 Chen Kang1
Chen Kang1 Shaozhen He
Shaozhen He Hong Zhai
Hong Zhai Qingchang Liu
Qingchang Liu- 1Key Laboratory of Sweetpotato Biology and Biotechnology, Ministry of Agriculture, Beijing Key Laboratory of Crop Genetic Improvement, Laboratory of Crop Heterosis and Utilization, Ministry of Education, College of Agronomy and Biotechnology, China Agricultural University, Beijing, China
- 2College of Agronomy, Qingdao Agricultural University, Qingdao, China
Auxin response factors (ARFs) compose a family of transcription factors and have been found to play major roles in the process of plant growth and development. However, their roles in plant carotenoid biosynthesis and responses to abiotic stresses are rarely known to date. In the present study, we found that the IbARF5 gene from sweetpotato (Ipomoea batatas (L.) Lam.) line HVB-3 increased the contents of carotenoids and enhanced the tolerance to salt and drought in transgenic Arabidopsis. The transgenic Arabidopsis plants exhibited the increased abscisic acid (ABA) and proline contents and superoxide dismutase (SOD) activity and the decreased H2O2 content. Furthermore, it was found that IbARF5 positively regulated the genes associated with carotenoid and ABA biosynthesis and abiotic stress responses. These results suggest that IbARF5 is involved in carotenoid biosynthesis and salt and drought tolerance in transgenic Arabidopsis. This study provides a novel ARF gene for improving carotenoid contents and salt and drought tolerance of sweetpotato and other plants.
Introduction
In nature, more than 750 kinds of carotenoids are characterized structurally, which are widely found in bacteria, fungi, algae, and plants (Hirschberg, 2001; Takaichi, 2011). The biosynthesis pathway of carotenoids has been extensively studied in plants, and nearly all of the key genes have been isolated and characterized (Cunningham and Gantt, 1998; Fraser and Bramley, 2004; Colasuonno et al., 2017; Kang et al., 2018). Abiotic stresses, especially salt and drought, seriously affect the productivity and cultivation expansion of crop plants worldwide, accordingly, to develop their high tolerance to salt and drought is highly desirable (Zhu, 2002; Lindemose et al., 2013; Zhai et al., 2016; Li et al., 2017). As the precursor of abscisic acid (ABA), carotenoids have functional roles in development and environmental adaptation of plants (Schwartz et al., 2003; Nambara and Marion-Poll, 2005; Mehrotra et al., 2014; Li, 2015; Moreno et al., 2016). Thus, increasing the contents of carotenoids helps to enhance the adaptation of plants to harsh environments.
Auxin response factors (ARFs) constitute a family of plant specific transcription factors. A typical ARF protein contains a B3-DNA binding domain in the highly conserved N-terminal region (Ulmasov et al., 1997; Hagen and Guilfoyle, 2002; Mei et al., 2018). ARFs mediate responses to auxin and have been shown to be implicated in senescence (Ellis et al., 2005), hormone signaling (Li et al., 2006) and developmental programs (Krogan et al., 2012). In rice, OsARF1 was auxin-regulated and classified as a primary auxin responsive gene (Waller et al., 2002). In Arabidopsis, ARF2 mediated ABA response (Wang et al., 2011); MP/ARF5 regulated embryo and flower patterning and vascular differentiation (Hardtke and Berleth, 1998; Krogan et al., 2012); ARF6 and ARF8 promoted jasmonic acid production and flower maturation (Nagpal et al., 2005); NPH4/ARF7 and ARF19 controlled leaf expansion and lateral root growth (Okushima et al., 2005; Wilmoth et al., 2005). In tomato, SlARF2 regulated lateral root formation and flower senescence (Ren et al., 2017); ARF4 controlled sugar metabolism (Sagar et al., 2013); ARF10 increased chlorophyll and sugar accumulation during fruit development (Mei et al., 2018); ARF5 regulated fruit set and development (Liu et al., 2018). However, the roles of ARFs in plant carotenoid biosynthesis and abiotic stress responses are rarely known to date.
Sweetpotato (Ipomoea batatas (L.) Lam.) is an important food crop worldwide, which provides rich carbohydrates and carotenoids for human consumption (Teow et al., 2007; Zhai et al., 2016). This crop can also be used for bioenergy production on marginal lands due to its high adaption to harsh environments (Liu et al., 2014). Sweetpotato breeders are focusing on improving carotenoid contents and abiotic stresses tolerance of this crop. Kang et al. (2017) summarized the improvement of carotenoids by gene engineering in sweetpotato. Overexpression of the genes related to carotenoid biosynthesis have been shown to increase the contents of carotenoids and enhance the tolerance to abiotic stresses in sweetpotato (Kim et al., 2012, Kim et al., 2013b; Yu et al., 2013; Kim et al., 2014; Li et al., 2017; Kang et al., 2018). To date, ARFs have not been reported in sweetpotato. In this study, we found that the IbARF5 gene from storage roots of sweetpotato is involved in carotenoid biosynthesis and salt and drought tolerance in transgenic Arabidopsis.
Materials and Methods
Plant Materials
Sweetpotato line HVB-3 with high carotenoid content was employed to clone the IbARF5 gene in this study. The expressed sequence tag (EST) for IbARF5 was obtained from the transcriptome data of HVB-3 developed by Li et al. (2015). Arabidopsis wild type (Columbia-0, WT) was used for characterizing the IbARF5 gene.
Isolation and Sequence Analysis of IbARF5
Total RNA was extracted from freshly harvested storage roots of HVB-3 and transcribed into first-strand cDNA according to the method of Kang et al. (2018). The full-length cDNA of IbARF5 was amplified with specific primers (Supplementary Table S1) by rapid amplification of cDNA ends (RACE) method. Genomic DNA isolated from in vitro-grown plants of HVB-3 was used to amplify the genomic sequence of IbARF5. The IbARF5 cDNA was analyzed by an online BLAST1. The open-reading frame (ORF) Finder2 was used to predict the ORF of IbARF5. The DNAMAN software was applied to align the amino acid sequence of IbARF5 with those of ARF proteins from different plant species. The MEGA 7.0 software was employed to conduct the phylogenetic analysis with the neighbor-joining (NJ) method. Exon–intron structure was constructed using Splign tool3. The molecular weight and theoretical isoelectric point (pI) of IbARF5 were calculated at http://web.expasy.org/compute_pi/.
Subcellular Localization of IbARF5
The IbARF5 ORF amplified with specific primers (Supplementary Table S1) was ligated into pMDC83. pMDC83-IbARF5-GFP and pMDC83-GFP (as control) were transiently expressed in the onion epidermal cells with a GeneGun (HeliosTM, Biorad, United States). After co-cultivation on Murashige and Skoog (MS) medium (pH 5.8) at 28°C for 24 h, the onion cells were examined under a laser scanning confocal fluorescence microscope (Nikon Inc., Melville, NY, United States).
Transactivation Activity Assay of IbARF5 in Yeast
Transactivation activity of IbARF5 in yeast (Saccharomyces cerevisiae) was assayed as described by Jiang et al. (2014). The corresponding regions of IbARF5 were PCR-amplified using specific primers (Supplementary Table S1) and integrated into the yeast expression vector pGBKT7 (pBD). Expression vectors pBD-IbARF5, pBD-IbARF5-1, pBD-IbARF5-2, pBD-IbARF5-3, pGAL4 (as positive vector), and pBD (as negative vector) were transferred into the yeast strain AH109, respectively. The transactivation activity was determined as described in the yeast protocols handbook (PT3024-1; Clontech, Mountain View, CA, United States).
Expression Analysis of IbARF5 in Sweetpotato
Total RNA was isolated from storage root, stem, and leaf tissues of the 100-day-old HVB-3 and used to analyze the expression of IbARF5 by quantitative real-time PCR (qRT-PCR) with its specific primers (Supplementary Table S1). Ibactin (AY905538) was served as an internal control. Comparative CT method was employed to quantify the gene expression (Schmittgen and Livak, 2008).
After cultured on MS medium for 4 weeks, the HVB-3 plants were treated in liquid MS media containing H2O (as control), 200 mM NaCl, 20% PEG6000 and 100 μM ABA, respectively, and sampled at 0, 2, 4, 6, 12, 24, and 48 h after treatment for analyzing the expression of IbARF5.
Production of the Transgenic Arabidopsis Plants
The overexpression vector pC3301-121-IbARF5 was constructed through inserting 35S-IbARF5-NOS into pCAMBIA3301. The recombinant vector was introduced into the Agrobacterium tumefaciens strain GV3101. The dipping flower method was applied to transform Arabidopsis and putatively transgenic Arabidopsis seeds were sown on MS medium with 12.5 mg L-1 phosphinothricin (PPT) for selecting transgenic plants. Histochemical GUS assay (Jefferson et al., 1987) and PCR analysis were used to identify the transgenic Arabidopsis plants. Transgenic Arabidopsis was planted in pots with a soil, vermiculite and humus mixture (1:1:1, v/v/v) to obtain T3 seeds.
Measurement of Carotenoid Contents
Leaves (2-week-old) and seeds of the transgenic Arabidopsis plants were applied to extract α-carotene, lutein, β-carotene, β-cryptoxanthin, and zeaxanthin. High performance liquid chromatography (HPLC) system was used to determine their contents (Li et al., 2017).
Assay for Salt and Drought Tolerance
One-week-old in vitro-grown seedlings of transgenic Arabidopsis and WT were treated on MS media with 200 mM NaCl and 300 mM mannitol, respectively. After 2 weeks, their root length and fresh weight (FW) were investigated. Furthermore, the transgenic and WT seedlings were planted for 2 weeks in pots with a soil, vermiculite and humus mixture (1:1:1, v/v/v) and subsequently irrigated with a 33 mL of 300 mM NaCl solution for each pot once every 2 days for 2 weeks, or stressed by drought for 4 weeks followed by 2 days re-watering. The transgenic plants and WT grown for 6 weeks under normal condition were used as control. The proline and H2O2 contents and superoxide dismutase (SOD) activity in the transgenic plants and WT grown in pots for 4 weeks under normal condition, 1 week under 300 mM NaCl stress after 2 weeks of normal treatment, and 2 weeks under drought stress after 2 weeks of normal treatment, respectively, were determined with Assay Kits (Comin Biotechnology Co., Ltd. Suzhou, China). The ABA content was measured as described by Gao et al. (2011). Twenty-seven plants in three pots with nine plants per pot were treated for each line.
For ABA sensitivity assay, the transgenic and WT seeds were sown on MS media with 0, 0.5, and 1 μM ABA, respectively. After 1 week, their germination and cotyledon opening and greening rates were investigated. Fifty seeds of each line on a plate were analyzed.
Expression Analysis of the Related Genes
Leaves (2-week-old) and seeds of the transgenic Arabidopsis plants and WT were applied to analyze the expression of the key genes in carotenoid biosynthesis. The leaves of the transgenic plants and WT potted for 4 weeks under normal condition, 1 week under 300 mM NaCl stress after 2 weeks of normal treatment, and 2 weeks under drought stress after 2 weeks of normal treatment, respectively, were used for analyzing the expression of the genes associated with ABA biosynthesis and abiotic stress responses. The specific primers of Atactin (NM112764) as internal control and the related genes were listed in Supplementary Table S1.
Statistical Analysis
All experiments were conducted with three biological replicates. Data presented as the mean ± SE were analyzed with Student’s t-test (two-tailed analysis) at P < 0.05 and P < 0.01.
Results
Cloning and Sequence Analysis of IbARF5
The 3757-bp cDNA of the IbARF5 gene contained a 2841-bp ORF encoding a 946-aa polypeptide with a molecular weight of 104.84 kDa and a predicted pI of 5.17. The IbARF5 protein shared a high sequence identity with ARF5 proteins in Nicotiana tabacum (XP_016465083.1, 74%), Capsicum annuum (XP_016568464.1, 72%), Sesamum indicum (XP_011083507.1, 72%), Solanum lycopersicum (NP_001234545.1, 72%), Solanum tuberosum (XP_006342026.1, 72%) and Vitis vinifera (XP_003634382.2, 68%). It contained one plant-specific B3-DNA binding domain, one Auxin_resp and one AUX_IAA (Supplementary Figure S1). Phylogenetic analysis showed that IbARF5 had a close relationship with that of N. tabacum (Figure 1A). The 4794-bp genomic DNA of IbARF5 contained 13 exons and 12 introns (Figure 1B).
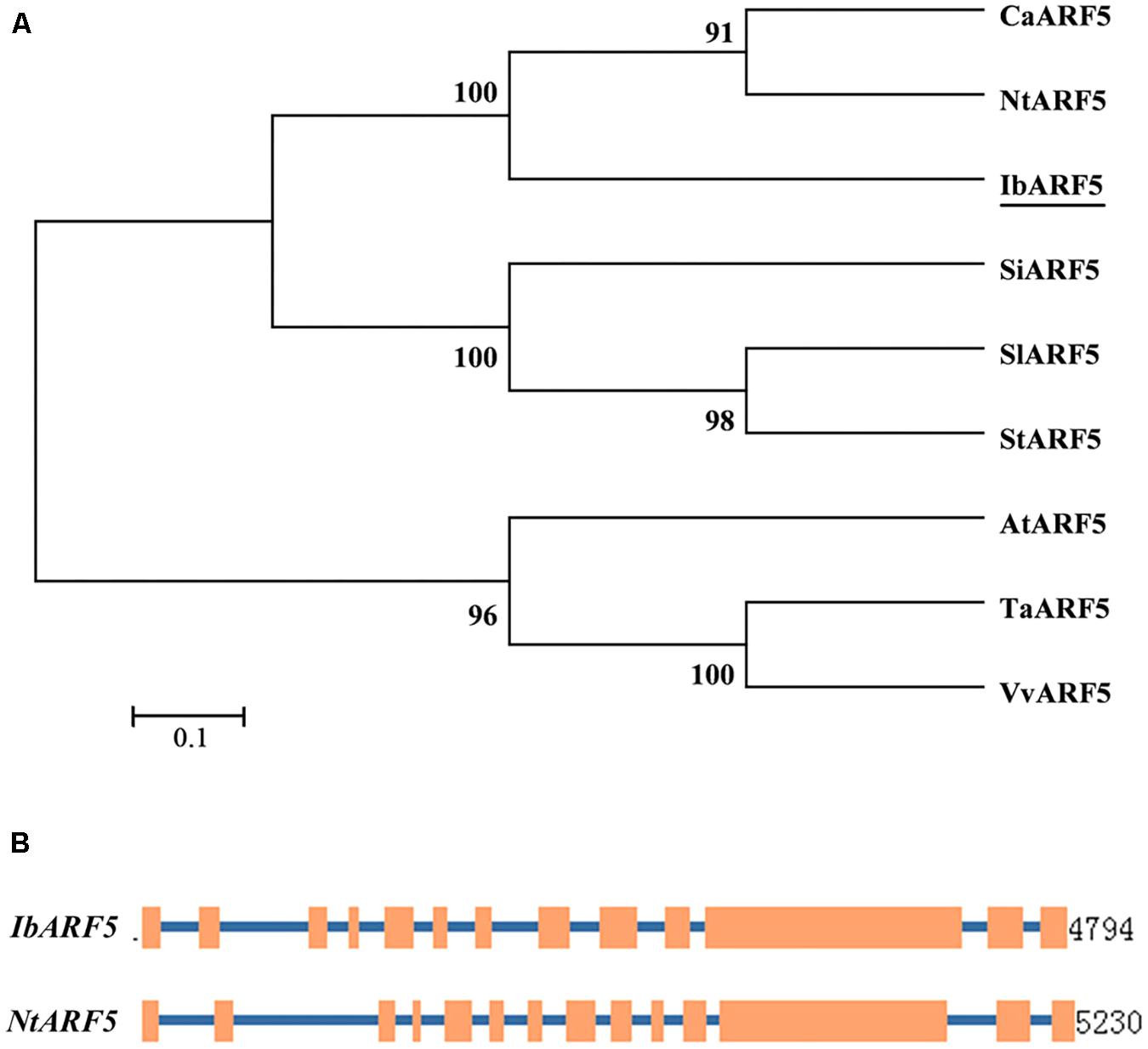
FIGURE 1. Phylogenetic tree of ARF5 in different plant species (A) and comparison of exon and intron constituents between IbARF5 and NtARF5 (B). Exons are represented by boxes and introns by lines.
IbARF5 Is Localized to Nuclei
The images from onion epidermal cells indicated that the green fluorescence emitted by IbARF5-GFP was exclusively distributed over the nuclei of the cells (Figure 2). These results showed that IbARF5 was localized to nuclei.
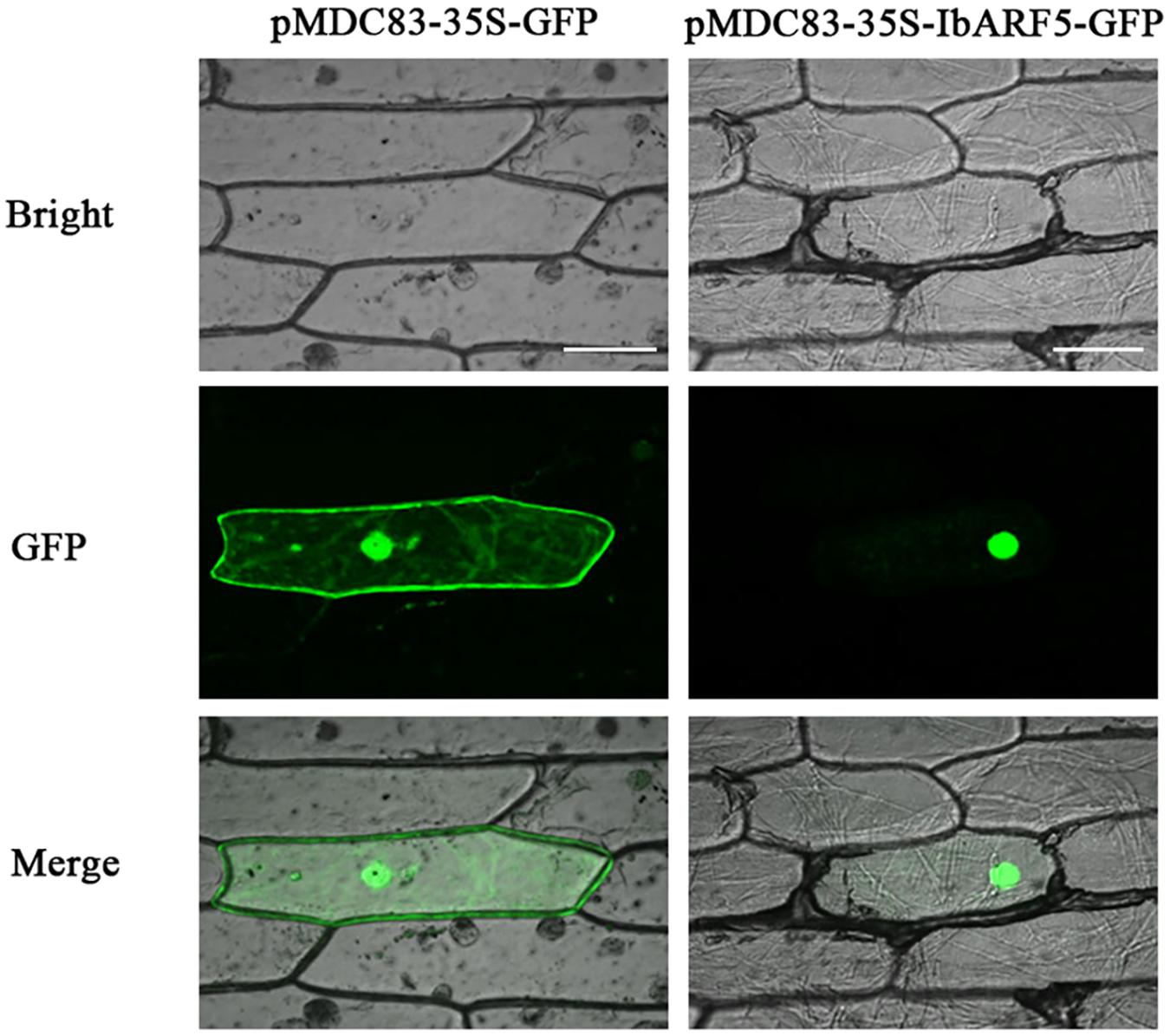
FIGURE 2. Subcellular localization of IbARF5 in onion leaf hypodermal cells. Confocal scanning microscopic images show localizations of IbARF5-GFP fusion proteins to nuclei in the right column vs. GFP as control in the left column. Bars = 20 μm.
IbARF5 Shows Transactivation Activity in Yeast
The yeast two-hybrid system was applied to identify a possible transactivation activity of IbARF5. The yeast cells harboring pBD-GAL4, pGBKT7-IbARF5 and pGBKT7-IbARF5-2, respectively, grew well on synthetic dropout (SD) plate without tryptophan and histidine and exhibited β-galactosidase activity, but the cells bearing pBD, pGBKT7-IbARF5-1, and pGBKT7-IbARF5-3, respectively, failed to grow (Figure 3A). These results demonstrated that IbARF5 might act as a transcription activator, and its transactivation activity was determined by the middle region, IbARF5-2 (Figure 3B).
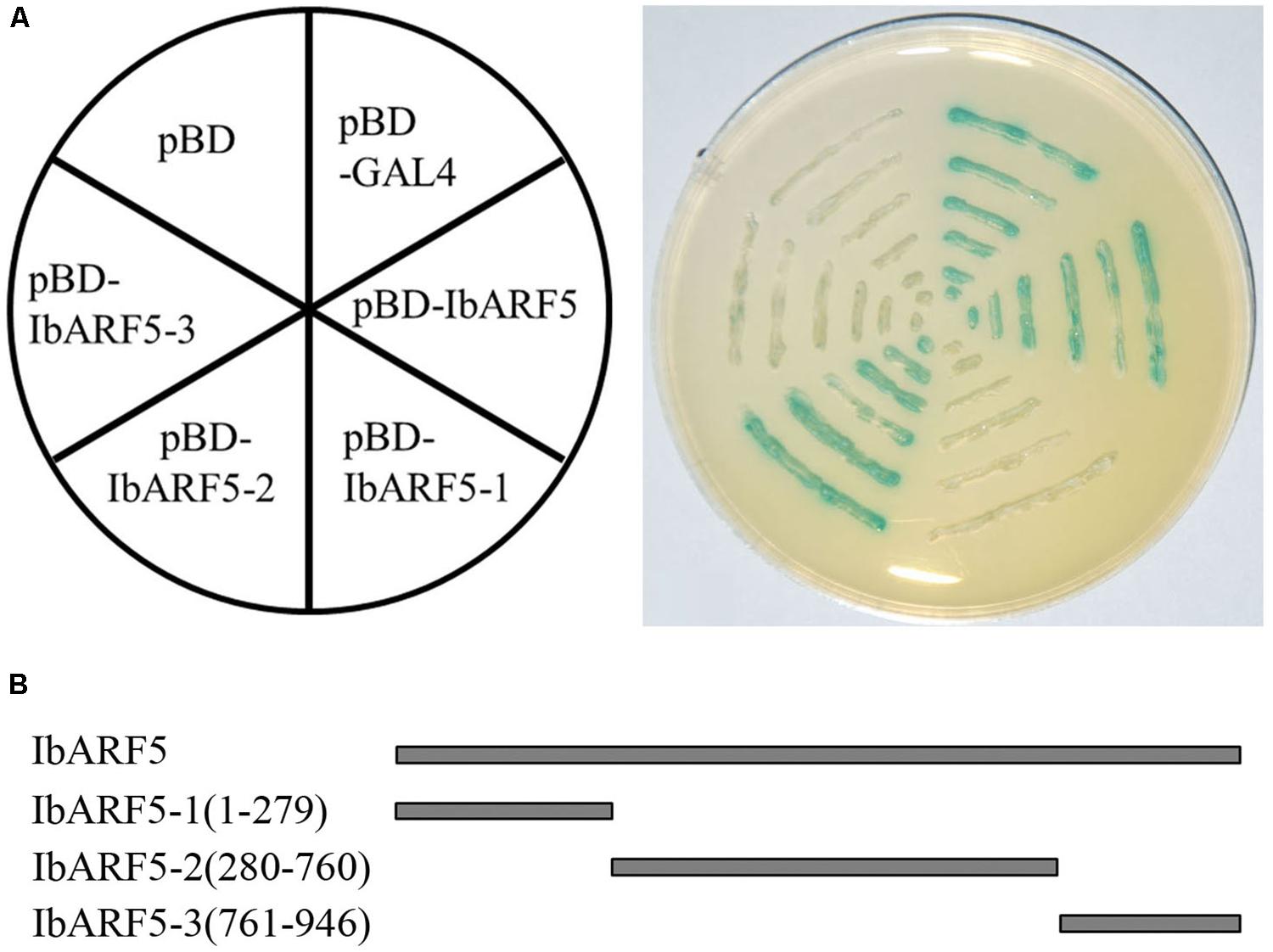
FIGURE 3. Transactivation activity of IbARF5 in the yeast. (A) the pBD-GAL4 vector as positive control; pBD-IbARF5; pBD-IbARF5-1; pBD-IbARF5-2; pBD-IbARF5-3; the empty pBD vector as negative control. The culture solution of the transformed yeast was drawn onto SD plate without tryptophan and histidine. (B) Different portions of IbARF5.
Expression Patterns of IbARF5 in Sweetpotato
Quantitative real-time PCR analysis revealed that IbARF5 exhibited higher expression level in the storage roots of HVB-3 than in its leaves and stems (Figure 4). Its expression in HVB-3 was strongly induced by NaCl, PEG6000 and ABA, and peaked (5.03-fold) at 4 h, (9.68-fold) at 24 h, and (12.18-fold) at 24 h, respectively (Figure 5).
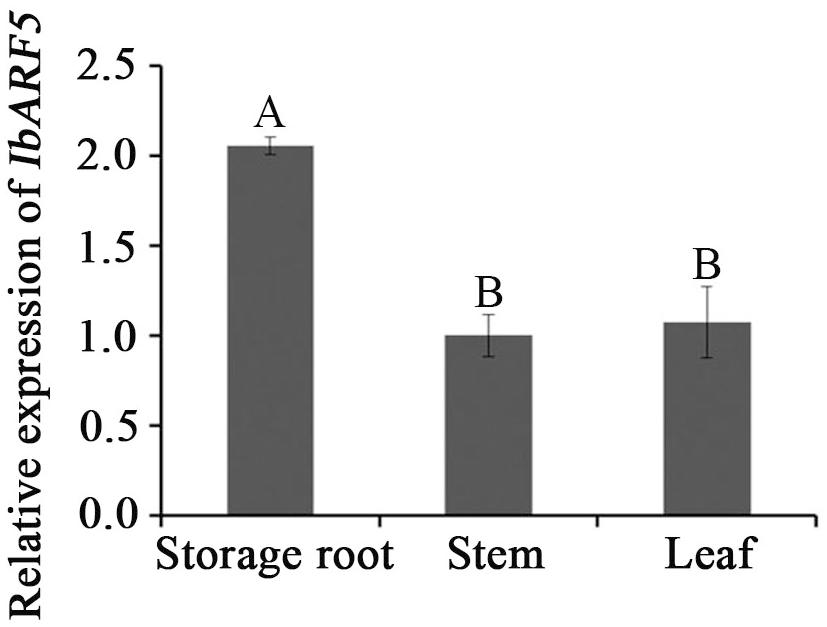
FIGURE 4. Expression analysis of IbARF5 in storage root, stem, and leaf tissues of HVB-3. Data are presented as means ± SE (n = 3). Different capital letters indicate a significant difference at P < 0.01 by Student’s t-test.
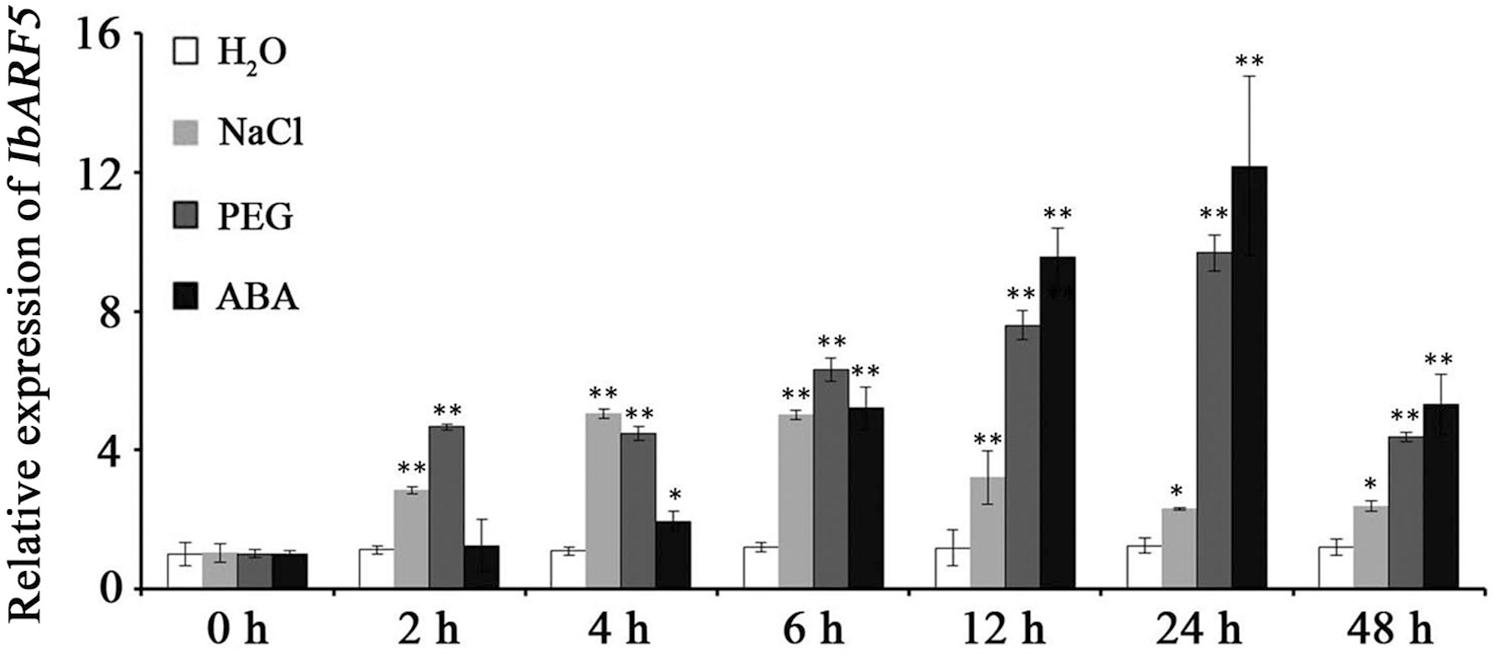
FIGURE 5. Expression analysis of IbARF5 in the in vitro-grown plants of HVB-3 after different times (h) in response to H2O, 200 mM NaCl, 20% PEG6000 and 100 μM ABA, respectively. Data are presented as means ± SE (n = 3). ∗ and ∗∗ indicate a significant difference from that of WT at P < 0.05 and P < 0.01, respectively, by Student’s t-test.
Production of the Transgenic Arabidopsis Plants
Putatively transgenic Arabidopsis seeds formed the plants on MS medium with 12.5 mg L-1 PPT. GUS assay and PCR analysis confirmed that 6 of the randomly sampled 60 plants were transgenic plants, named L1, L2, …, L6, respectively, from which T3 were generated. IbARF5 showed high expression levels in the transgenic Arabidopsis plants, especially L1, L4, L5, and L6 (Supplementary Figure S2).
IbARF5 Increases Carotenoid Contents
The lutein and zeaxanthin contents were significantly increased, but α-carotene and β-carotene contents were not changed and β-cryptoxanthin was not detected in leaves of L1, L4, L5, and L6 (Table 1). In seeds of these four transgenic plants, the lutein, β-carotene, and zeaxanthin contents were significantly increased, but α-carotene was not detected and β-cryptoxanthin content was significantly increased only in L5 and L6 (Table 2). The total carotenoid contents in leaves and seeds were increased by 1.13–1.16 folds and 1.32–1.78 folds, respectively (Tables 1 and 2).
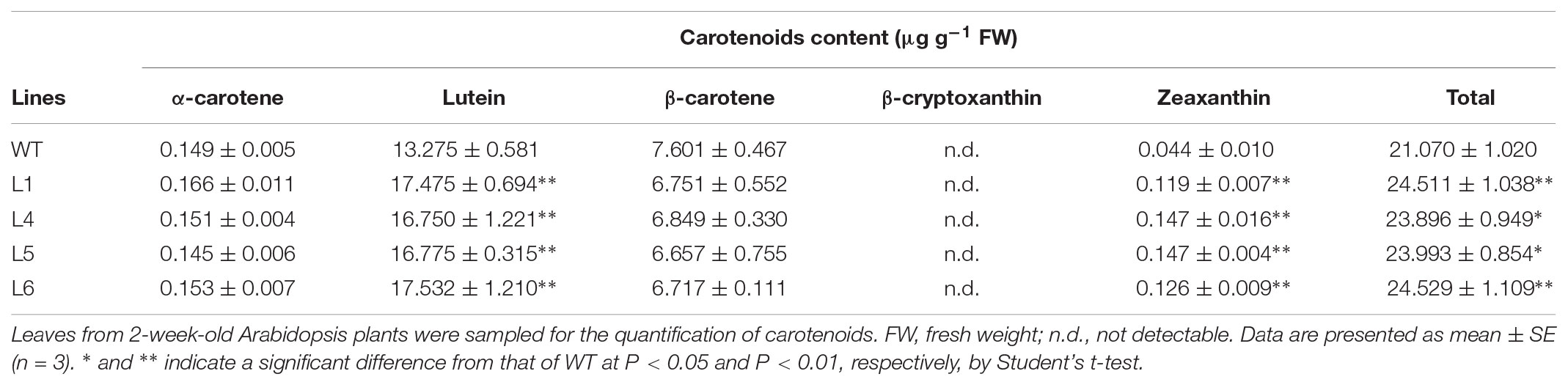
TABLE 1. Carotenoid contents in leaves of the IbARF5-overexpressing Arabidopsis plants.
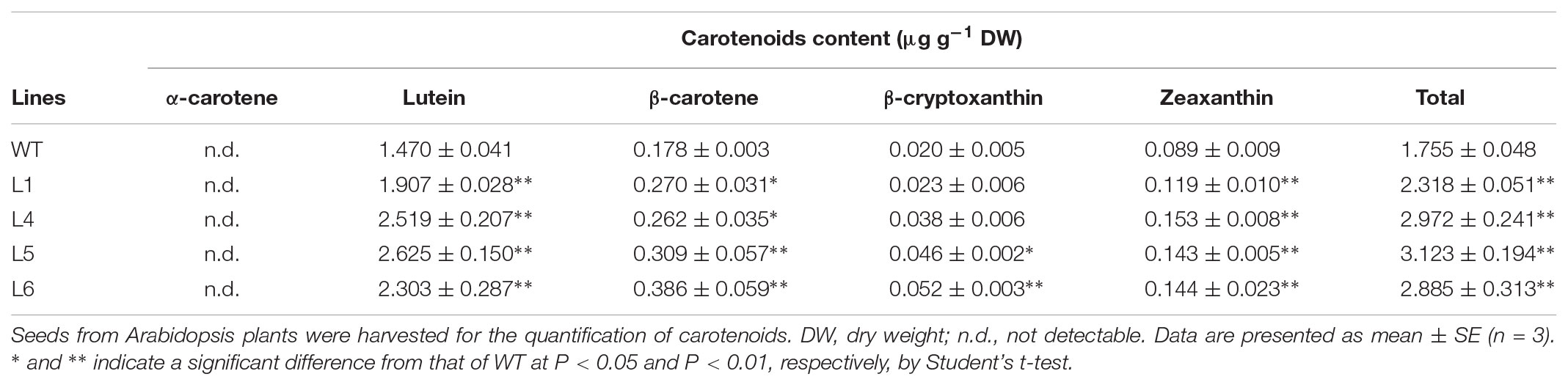
TABLE 2. Carotenoid contents in seeds of the IbARF5-overexpressing Arabidopsis plants.
IbARF5 Enhances Salt and Drought Tolerance
Four transgenic Arabidopsis plants, L1, L4, L5, and L6, and WT seedlings showed no significant differences in rooting and FW on MS medium without stresses (Figure 6). However, the transgenic plants exhibited good rooting and increased FW in contrast to WT on MS media with 200 mM NaCl and 300 mM mannitol, respectively (Figure 6).
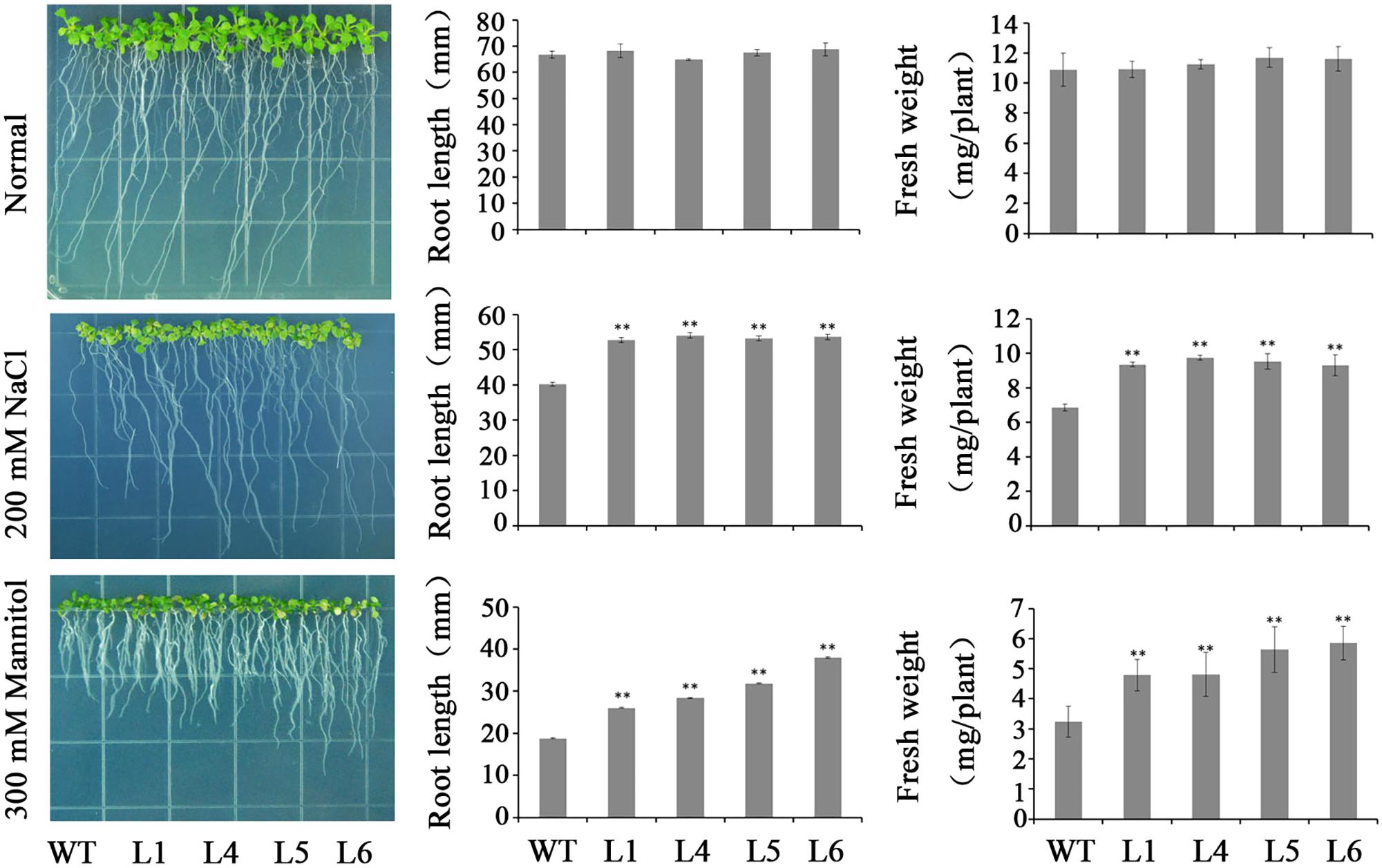
FIGURE 6. Responses of the transgenic Arabidopsis seedlings and WT cultured for 2 weeks on MS medium with 200 mM NaCl and 300 mM mannitol, respectively. Data are presented as mean ± SE (n = 3). ∗∗ indicates a significant difference from that of WT at P < 0.01 by Student’s t-test.
The transgenic plants and WT grown in pots showed similar growth trends under normal conditions (Figure 7A). Under NaCl and drought stresses, the transgenic plants showed good growth, while WT almost died (Figure 7A). Furthermore, it was found that the ABA and proline contents were increased, SOD activity was enhanced and H2O2 content was decreased in the transgenic plants (Figure 7B).
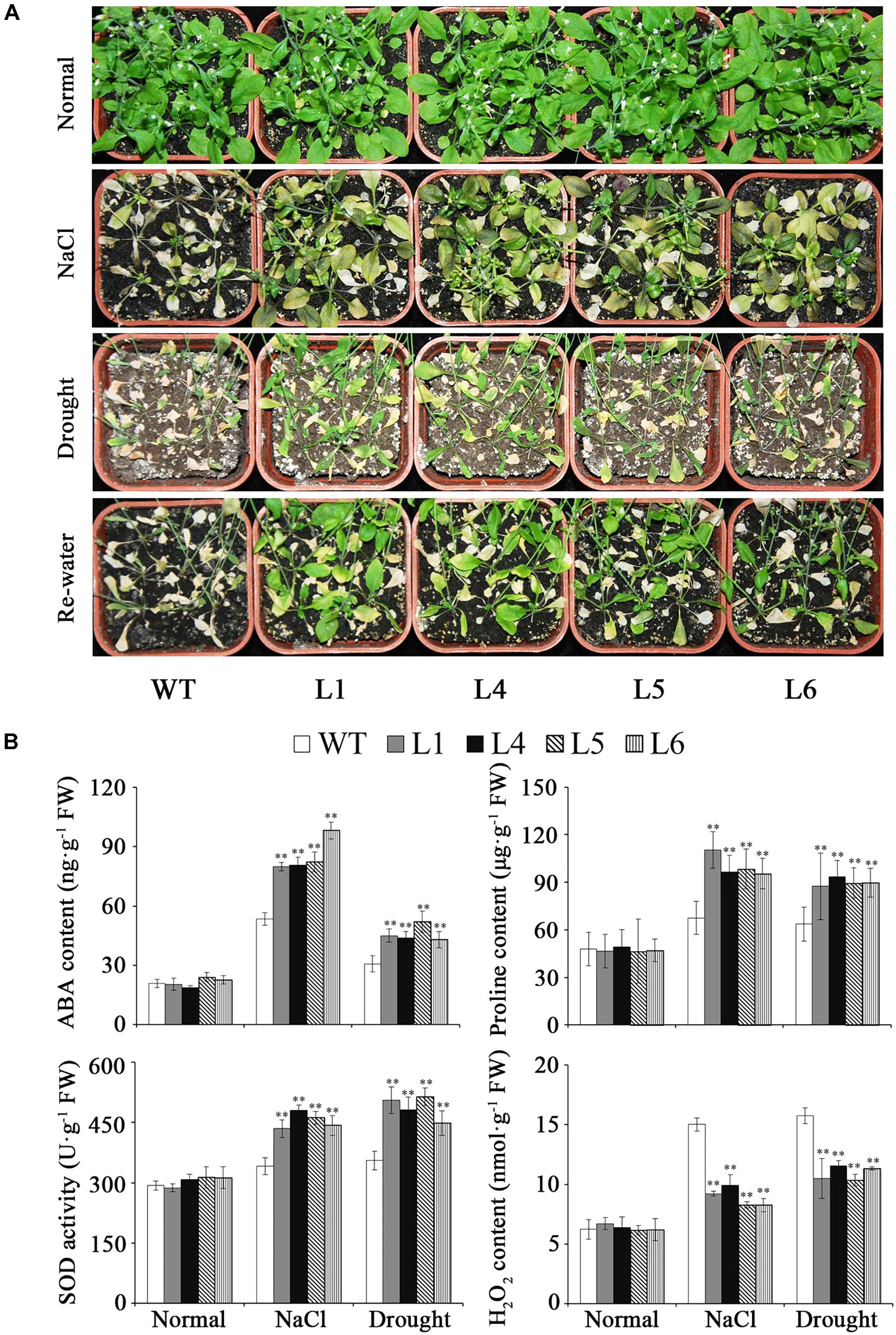
FIGURE 7. Responses of the transgenic Arabidopsis plants and WT grown in pots under salt and drought stresses. (A) Phenotypes of transgenic plants vs. WT grown for 6 weeks under normal condition, 2 weeks under 300 mM NaCl stress after 2 weeks of normal treatment, and 4 weeks under drought stress followed by 2 days re-watering after 2 weeks of normal treatment, respectively. (B) ABA, proline and H2O2 contents and SOD activity in the transgenic plants and WT grown for 4 weeks under normal condition, 1 week under 300 mM NaCl stress after 2 weeks of normal treatment, and 2 weeks under drought stress after 2 weeks of normal treatment, respectively. Data are presented as mean ± SE (n = 3). ∗∗ indicates a significant difference from that of WT at P < 0.01 by Student’s t-test.
No obvious differences in seed germination were observed between the transgenic plants and WT under normal condition (Figure 8). Under the treatment with different ABA concentrations, the germination of the transgenic seeds was more sensitive to ABA-elicited inhibition than that of WT though both germination rate and cotyledon opening and greening rate of the transgenic and WT seeds declined (Figure 8). These results demonstrated that IbARF5 might participate in the ABA signaling pathway.
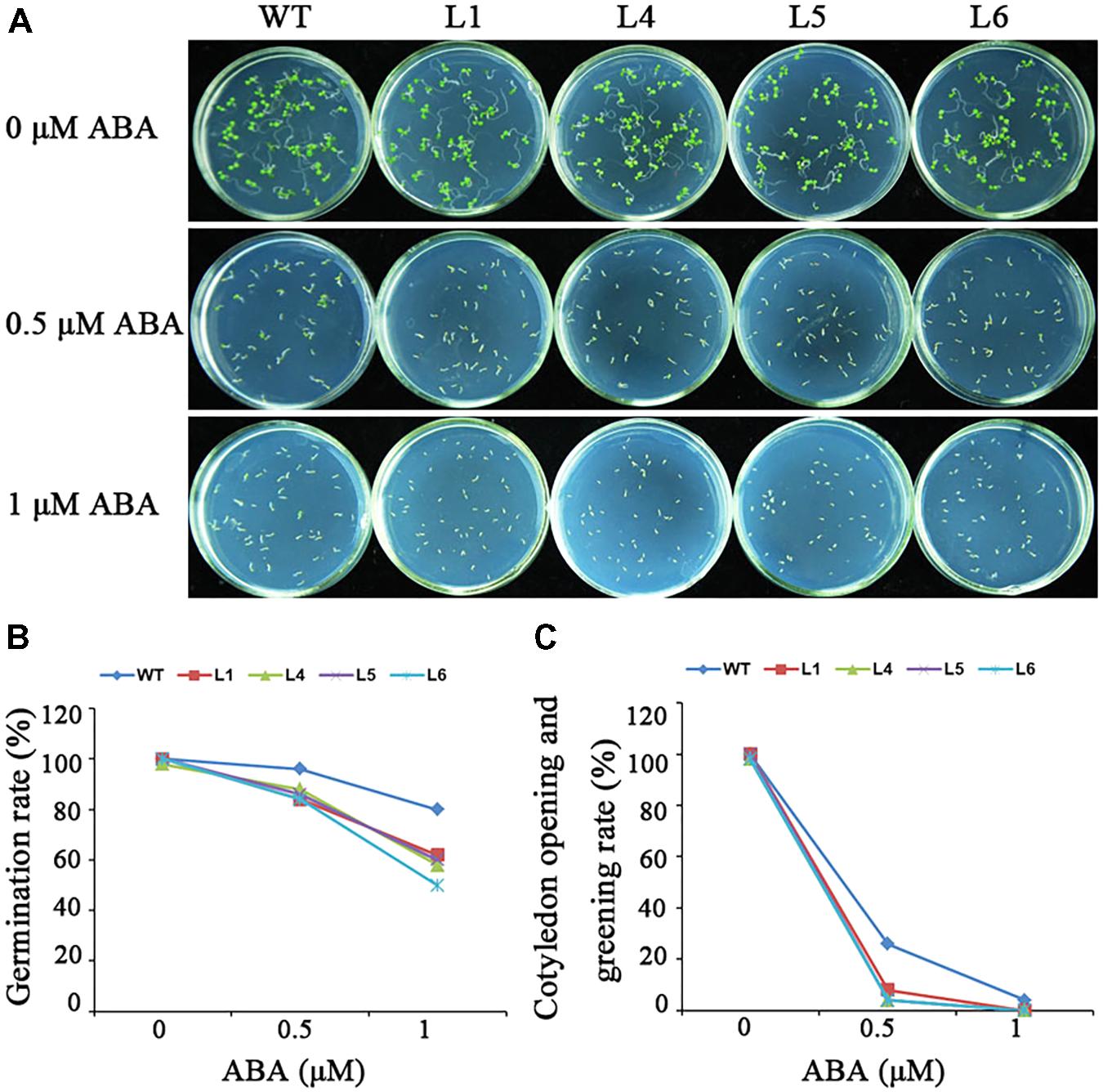
FIGURE 8. Responses of the transgenic Arabidopsis and WT seeds sown on MS medium with 0, 0.5, and 1 μM ABA for 1 week. (A) Growth vigor of the transgenic and WT seedlings. (B) Germination rates of the transgenic and WT seeds. (C) Cotyledon opening and greening rates of the transgenic and WT seeds.
IbARF5 Up-Regulates the Genes Involved in Carotenoid and ABA Biosynthesis and Abiotic Stress Responses
The genes encoding the key enzymes in carotenoid biosynthesis, eranylgeranyl pyrophosphate (GGPS), ζ-carotene desaturase (ZDS), phytoene synthase (PSY), 𝜀-carotene hydroxylase (𝜀-CHY), β-lycopene cyclase (β-LCY) and β-carotene hydroxylase (β-CHY) except for phytoene desaturase (PDS) and 𝜀-lycopene cyclase (𝜀-LCY) were systematically up-regulated in leaves of the transgenic Arabidopsis plants (Figure 9). GGPS, 𝜀-CHY, β-LCY, and β-CHY exhibited the increased expression levels, but ZDS, PSY, and 𝜀-LCY showed no changes in expression level and PDS was down-regulated in the transgenic seeds (Figure 9). Under NaCl and drought stresses, the genes encoding the key enzymes in ABA biosynthesis, zeaxanthin epoxidase (ZEP), 9-cis-epoxycarotenoid dioxygenase (NCED), and xanthoxin dehydrogenas (ABA2) were up-regulated, and abiotic stress-responsive genes encoding pyrroline-5-carboxylate synthase (P5CS), SOD, ascorbate peroxidase (APX), and dehydroascorbate reductase (DHAR) were also found to be up-regulated (Figure 10).
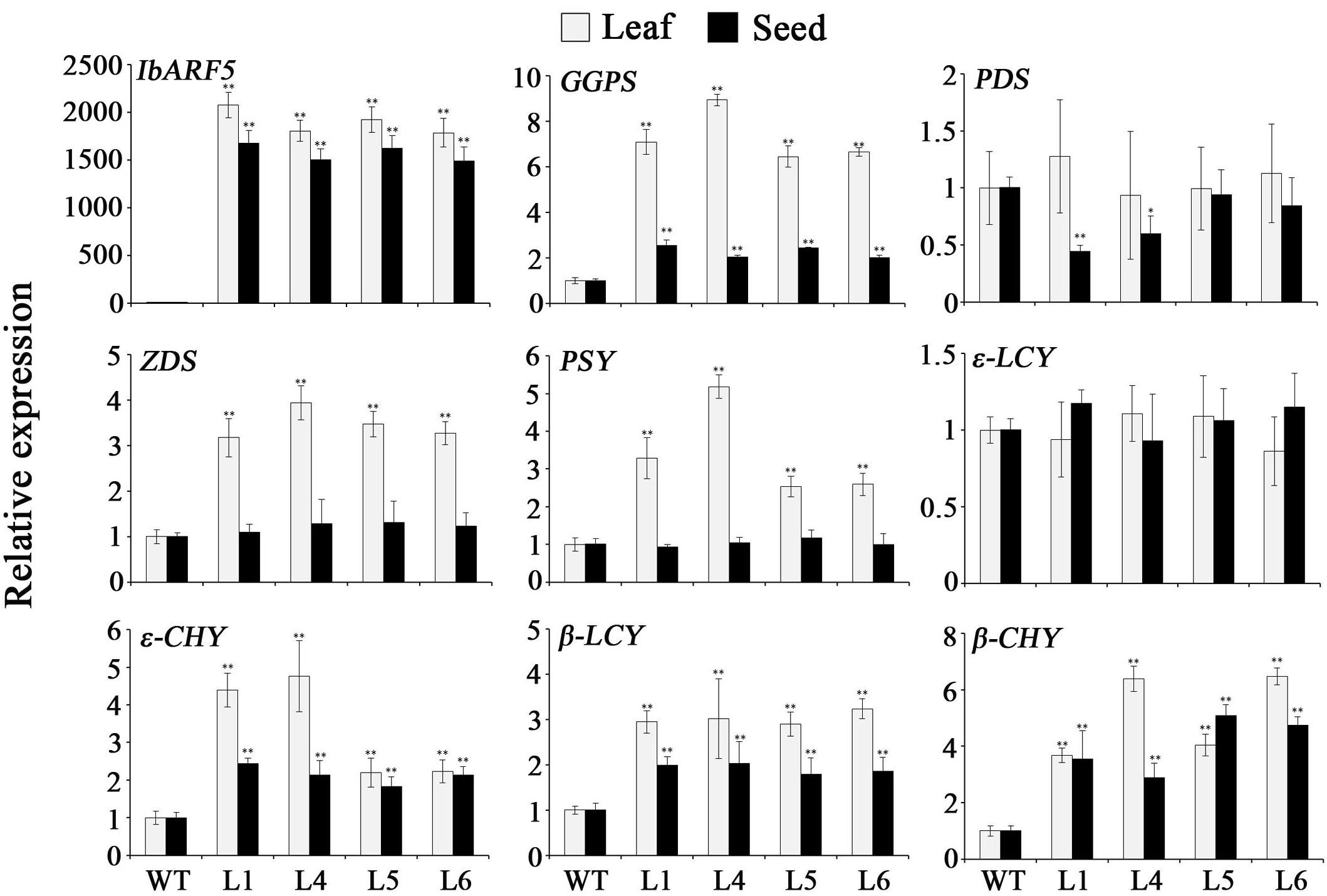
FIGURE 9. Transcript levels of IbARF5 and carotenoid biosynthesis genes in leaves and seeds of the transgenic Arabidopsis plants and WT. Data are presented as means ± SE (n = 3). ∗ and ∗∗ indicate a significant difference from that of WT at P < 0.05 and P < 0.01, respectively, by Student’s t-test.
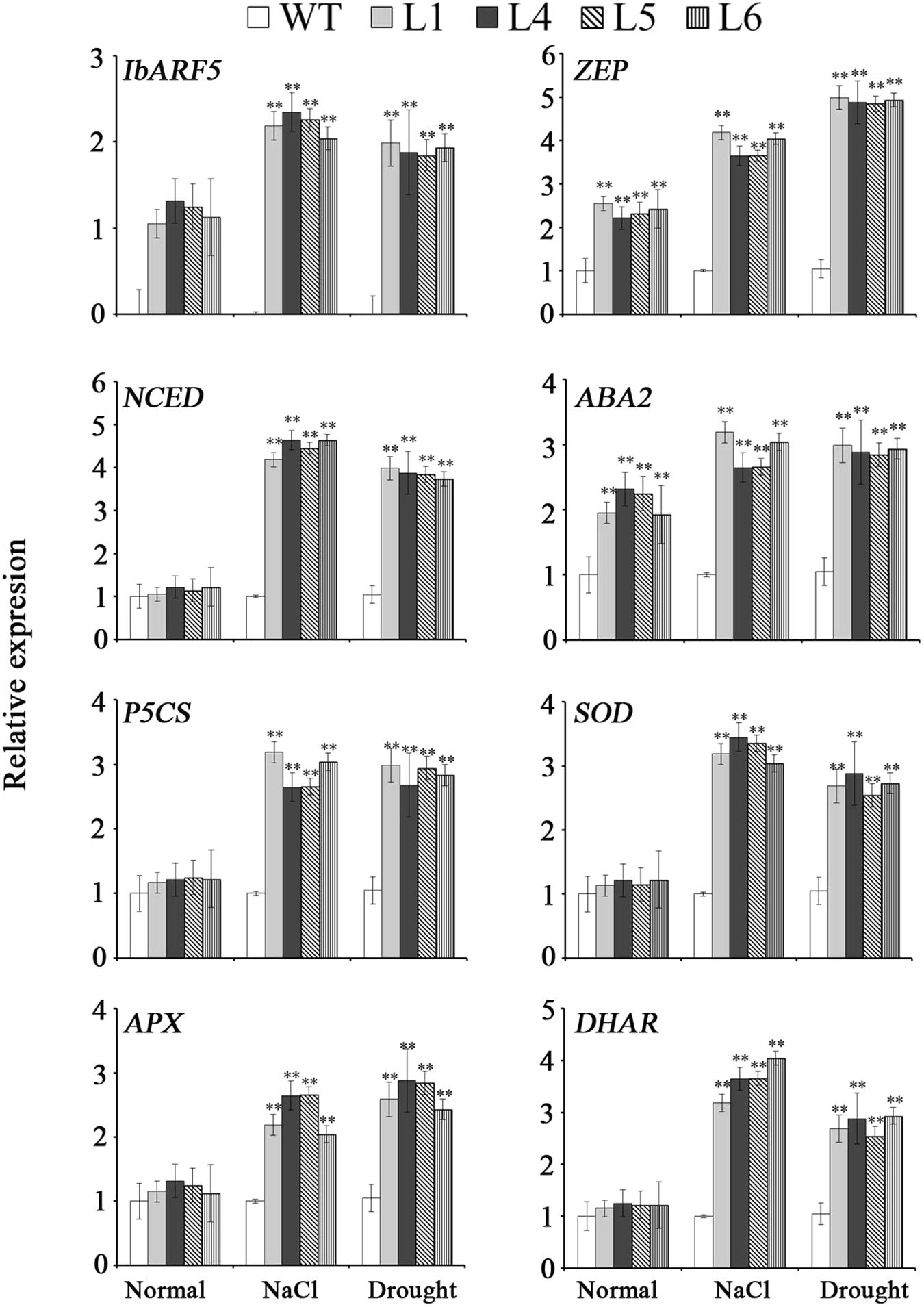
FIGURE 10. Transcript levels of salt and drought responsive genes in leaves of transgenic Arabidopsis plants and WT pot-grown for 4 weeks under normal condition, 1 week under 300 mM NaCl stress after 2 weeks of normal treatment, and 2 weeks under drought stress after 2 weeks of normal treatment, respectively. Data are presented as mean ± SE (n = 3). ∗∗ indicates a significant difference from that of WT at P < 0.01 by Student’s t-test.
Discussion
IbARF5 Increases Carotenoid Contents and Salt and Drought Tolerance
In plants, ARFs encode important transcription factors which regulate the expression of genes in response to auxin (Guilfoyle and Hagen, 2007). Several ARF transcription factor genes have been cloned from Arabidopsis, rice and tomato, and were found to play crucial roles in plant growth and developmental processes (Harper et al., 2000; Waller et al., 2002; Ellis et al., 2005; Okushima et al., 2005; Wilmoth et al., 2005; Wang et al., 2011; Ren et al., 2017; Liu et al., 2018). However, there is no report on the ARF transcription factors in improving carotenoid contents and abiotic stress tolerance in plants. Previous studies demonstrated that AtARF5 affected lateral organ development, primary root initiation, flower primordium initiation, and cotyledon development in Arabidopsis (Krogan et al., 2012) and SlARF5 controlled fruit set and development in tomato (Liu et al., 2018). In the present study, the IbARF5 gene was isolated from sweetpotato line HVB-3 with high carotenoid content. We found that its overexpression significantly increased the content of carotenoids and enhanced the tolerance to salt and drought in transgenic Arabidopsis (Tables 1, 2 and Figures 6, 7).
IbARF5 Up-Regulates the Genes Involved in Carotenoid Biosynthesis
It has been shown that carotenoid biosynthesis is mainly regulated at the transcript level of genes encoding the biosynthetic enzymes (Römer and Fraser, 2005; Sandmann et al., 2006; Li et al., 2017; Kang et al., 2018). In this study, we found that the key genes in carotenoid biosynthesis, GGPS, ZDS, PSY, 𝜀-CHY, β-LCY, and β-CHY in leaves and GGPS, 𝜀-CHY, β-LCY, and β-CHY in seeds of transgenic Arabidopsis were significantly up-regulated (Figure 9), which corresponded with the increase of carotenoid contents in transgenic Arabidopsis (Tables 1 and 2). These findings suggest that IbARF5 positively controls the expression of carotenoid biosynthetic genes, which resulted in the increased carotenoid contents in transgenic Arabidopsis. Overexpression of the Orange gene (IbOr) from sweetpotato increased carotenoid accumulation and abiotic stress tolerance in transgenic sweetpotato, potato, and alfalfa (Kim et al., 2013a; Goo et al., 2015; Wang et al., 2015). Furthermore, it was proved that similar to AtOr of Arabidopsis, IbOr directly interacted with PSY and increased carotenoid accumulation (Park et al., 2016; Kim et al., 2018). Therefore, the precise underlying mechanisms of IbARF5 in plant carotenoid accumulation need to be further investigated. In addition, we are developing the IbARF5-overexpressing sweetpotato plants for further analyzing its roles in carotenoid accumulation of the storage roots.
IbARF5 Up-Regulates the Genes Involved in ABA Biosynthesis
Carotenoids, especially β-branch carotenoids, serve as precursors for ABA biosynthesis and play a crucial role in plant tolerance and adaptation to abiotic stresses (Demmigadams and Adams, 2002; Xiong and Zhu, 2003; Sah et al., 2016). ABA regulates the expression of ABA-dependent stress-responsive genes and the increased level of ABA has been found to enhance the tolerance to salt and drought (Tuteja, 2007; Vishwakarma et al., 2017). It was reported that overexpression of IbMIPS1, IbZDS, and IbLCYB2 increased the level of ABA, which led to the enhanced tolerance to salt and drought in sweetpotato (Zhai et al., 2016; Li et al., 2017; Kang et al., 2018). In this study, the IbARF5-overexpressing Arabidopsis seeds showed the increased sensitivity to ABA in germination (Figure 8). The ABA biosynthetic genes IbZEP, IbNCED, and IbABA2 were up-regulated and ABA level was also significantly increased in transgenic Arabidopsis (Figures 7B and 10). These results suggest that overexpression of IbARF5 confers salt and drought tolerance by up-regulating the ABA biosynthetic genes and increasing ABA level in transgenic Arabidopsis.
IbARF5 Up-Regulates Abiotic Stress-Responsive Genes and Changes Abiotic Stress-Associated Components
It is reported that the high level of ABA increases the transcript level of P5CS, which leads to more accumulation of proline under abiotic stresses (Sripinyowanich et al., 2013). Proline plays a pivotal role in maintaining osmotic balance, protecting integrity membrane and increasing reactive oxygen species (ROS) scavenging capacity, and its elevated level enhances salt and drought tolerance in plants (Yoshiba et al., 1997; Maggio et al., 2002; Neisiani et al., 2009; Gill and Tuteja, 2010; Kang et al., 2018). SOD as the first line of defense against ROS is induced by abiotic stresses to promote ROS scavenging (Wang et al., 2009). In the present study, P5CS, SOD, APX, and DHAR were up-regulated, proline level and SOD activity were increased and H2O2 content was decreased in transgenic Arabidopsis under salt and drought stresses (Figure 7B). Therefore, it is thought that the enhanced tolerance to salt and drought is due to up-regulation of abiotic stress-responsive genes and change of abiotic stress-associated components in transgenic Arabidopsis.
Conclusion
This study reveals, for the first time, that the IbARF5 gene from sweetpotato is involved in carotenoid biosynthesis and salt and drought tolerance of plants. Its overexpression increased the contents of carotenoids and conferred the tolerance to salt and drought by up-regulating the key genes involved in carotenoid and ABA biosynthesis and abiotic stress responses in transgenic Arabidopsis. This study provides a novel ARF gene for improving carotenoid contents and salt and drought tolerance of sweetpotato and other plants.
Author Contributions
QL and CK conceived and designed the experiments. CK and RL performed the experiments. CK and SH analyzed the data. QL, HZ, and NZ contributed reagents, materials, and analysis tools. QL and CK wrote the paper. All authors read and approved the final manuscript.
Funding
This work was supported by China Agriculture Research System (CARS-10, Sweetpotato) and National Natural Science Foundation of China (31461143017).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2018.01307/full#supplementary-material
FIGURE S1. Sequence alignment of IbARF5 with its homologs from other plants. Characteristic regions of ARF5 are indicated above the IbARF5 sequence.  ,Plant-specific B3-DNA binding domain;
,Plant-specific B3-DNA binding domain;  ,Auxin_resp;
,Auxin_resp;  ,AUX_IAA.
,AUX_IAA.
FIGURE S2. Expression analysis of IbARF5 in the transgenic Arabidopsis plants. The Arabidopsis actin gene was used as an internal control. Data are presented as means ± SE (n = 3). ∗∗ indicates a significant difference from that of WT at P < 0.01 by Student’s t-test.
TABLE S1. Primers used in this study.
Footnotes
- ^https://blast.ncbi.nlm.nih.gov/Blast.cgi
- ^https://www.ncbi.nlm.nih.gov/orffinder/
- ^https://www.ncbi.nlm.nih.gov/sutils/splign/splign.cgi?textpage=online&level=form
References
Colasuonno, P., Lozito, M. L., Marcotuli, I., Nigro, D., Giancaspro, A., Mangini, G., et al. (2017). The carotenoid biosynthetic and catabolic genes in wheat and their association with yellow pigments. BMC Genomics 18:122. doi: 10.1186/s12864-016-3395-6
Cunningham, F. X., and Gantt, E. (1998). Genes and enzymes of carotenoid biosynthesis in plants. Annu. Rev. Plant Physiol. Mol. Biol. 49, 557–583. doi: 10.1146/annurev.arplant.49.1.557
Demmigadams, B., and Adams, W. W. (2002). Antioxidants in photosynthesis and human nutrition. Science 298, 2149–2153. doi: 10.1126/science.1078002
Ellis, C. M., Nagpal, P., Young, J. C., Hagen, G., Guilfoyle, T. J., and Reed, J. W. (2005). Auxin response factor1 and auxin response factor2 regulate senescence and floral organ abscission in Arabidopsis thaliana. Development 132, 4563–4574. doi: 10.1242/dev.02012
Fraser, P. D., and Bramley, P. M. (2004). The biosynthesis and nutritional uses of carotenoids. Prog. Lipid Res. 43, 228–265. doi: 10.1016/j.plipres.2003.10.002
Gao, S., Yuan, L., Zhai, H., Liu, C. L., He, S. Z., and Liu, Q. C. (2011). Transgenic sweetpotato plants expressing an LOS5 gene are tolerant to salt stress. Plant Cell Tissue Organ Cult. 107, 205–213. doi: 10.1007/s11240-011-9971-1
Gill, S. S., and Tuteja, N. (2010). Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 48, 909–930. doi: 10.1016/j.plaphy.2010.08.016
Goo, Y., Han, E., Jeong, J. C., Kwak, S., Yu, J., Kim, Y., et al. (2015). Overexpression of the sweet potato IbOr gene results in the increased accumulation of carotenoid and confers tolerance to environmental stresses in transgenic potato. C. R. Biol. 338, 12–20. doi: 10.1016/j.crvi.2014.10.006
Guilfoyle, T. J., and Hagen, G. (2007). Auxin response factors. Curr. Opin. Plant Biol. 10, 453–460.
Hagen, G., and Guilfoyle, T. (2002). Auxin-responsive gene expression: genes, promoters and regulatory factors. Plant Mol. Biol. 49, 373–385.
Hardtke, C. S., and Berleth, T. (1998). The Arabidopsis gene MONOPTEROS encodes a transcription factor mediating embryo axis formation and vascular development. EMBO J. 17, 1405–1411. doi: 10.1093/emboj/17.5.1405
Harper, R. M., Stowe-Evans, E. L., Luesse, D. R., Muto, H., Tatematsu, K., Watahiki, M. K., et al. (2000). The NPH4 locus encodes the auxin response factor ARF7, a conditional regulator of differential growth in aerial Arabidopsis tissue. Plant Cell 12, 757–770. doi: 10.1105/tpc.12.5.757
Hirschberg, J. (2001). Carotenoid biosynthesis in flowering plants. Curr. Opin. Plant Biol. 4, 210–218. doi: 10.1016/S1369-5266(00)00163-1
Jefferson, R. A., Kavanagh, T. A., and Bevan, M. W. (1987). GUS fusions: β-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 6, 3901–3907.
Jiang, X., Zhang, C., Lü, P., Jiang, G., Liu, X., Dai, F., et al. (2014). RhNAC3, a stress-associated NAC transcription factor, has a role in dehydration tolerance through regulating osmotic stress-related genes in rose petals. Plant Biotechnol. J. 12, 38–48. doi: 10.1111/pbi.12114
Kang, C., Zhai, H., Xue, L. Y., Zhao, N., He, S. Z., and Liu, Q. C. (2018). A lycopene β-cyclase gene, IbLCYB2, enhances carotenoid contents and abiotic stress tolerance in transgenic sweetpotato. Plant Sci. 272, 243–254. doi: 10.1016/j.plantsci.2018.05.005
Kang, L., Park, S., Ji, C. Y., Kim, H. S., Lee, H., and Kwak, S. (2017). Metabolic engineering of carotenoids in transgenic sweetpotato. Breed. Sci. 67, 27–34. doi: 10.1270/jsbbs.16118
Kim, S. H., Ahn, Y. O., Ahn, M., Jeong, J. C., Lee, H., and Kwak, S. (2013a). Cloning and characterization of an Orange gene that increases carotenoid accumulation and salt stress tolerance in transgenic sweetpotato cultures. Plant Physiol. Biochem. 70, 445–454. doi: 10.1016/j.plaphy.2013.06.011
Kim, S. H., Kim, Y., Ahn, Y. O., Ahn, M., Jeong, J. C., Lee, H., et al. (2013b). Downregulation of the lycopene 𝜀-cyclase gene increases carotenoid synthesis via the β-branch-specific pathway and enhances salt-stress tolerance in sweetpotato transgenic calli. Physiol. Plant. 147, 432–442. doi: 10.1111/j.1399-3054.2012.01688.x
Kim, S. H., Ahn, Y. O., Ahn, M., Lee, H., and Kwak, S. (2012). Down-regulation of β-carotene hydroxylase increases β-carotene and total carotenoids enhancing salt stress tolerance in transgenic cultured cells of sweetpotato. Phytochemistry 74, 69–78. doi: 10.1016/j.phytochem.2011.11.003
Kim, S. H., Jeong, J. C., Park, S., Bae, J., Ahn, M., Lee, H., et al. (2014). Down-regulation of sweetpotato lycopene β-cyclase gene enhances tolerance to abiotic stress in transgenic calli. Mol. Biol. Rep. 41, 8137–8148. doi: 10.1007/s11033-014-3714-4
Kim, S. H., Ji, Y. C., Lee, C., Kim, S., Park, S., and Kwak, S. (2018). Orange: a target gene for regulating carotenoid homeostasis and increasing plant tolerance to environmental stress in marginal lands. J. Exp. Bot. 69, 3393–3400. doi: 10.1093/jxb/ery023
Krogan, N. T., Ckurshumova, W., Marcos, D., Caragea, A. E., and Berleth, T. (2012). Deletion of MP/ARF5 domains III and IV reveals a requirement for Aux/IAA regulation in Arabidopsis leaf vascular patterning. New Phytol. 194, 391–401. doi: 10.1111/j.1469-8137.2012.04064.x
Li, J., Dai, X., and Zhao, Y. (2006). A role for auxin response factor 19 in auxin and ethylene signaling in Arabidopsis. Plant Physiol. 140, 899–908. doi: 10.1104/pp.105.070987
Li, R. J., Kang, C., Song, X. J., Yu, L., Liu, D. G., He, S. Z., et al. (2017). A ζ-carotene desaturase gene, IbZDS, increases β-carotene and lutein contents and enhances salt tolerance in transgenic sweetpotato. Plant Sci. 262, 39–51. doi: 10.1016/j.plantsci.2017.05.014
Li, R. J., Zhai, H., Kang, C., Liu, D. G., He, S. Z., and Liu, Q. C. (2015). De novo transcriptome sequencing of the orange-fleshed sweet potato and analysis of differentially expressed genes related to carotenoid biosynthesis. Int. J. Genomics 2015:843802. doi: 10.1155/2015/843802
Li, T. (2015). Recent advances in understanding carotenoid-derived signaling molecules in regulating plant growth and development. Front. Plant Sci. 6:790. doi: 10.3389/fpls.2015.00790
Lindemose, S., O’Shea, C., Jensen, M., and Skriver, K. (2013). Structure, function and networks of transcription factors involved in abiotic stress responses. Int. J. Mol. Sci. 14, 5842–5878. doi: 10.3390/ijms14035842
Liu, D. G., He, S. Z., Zhai, H., Wang, L. J., Zhao, Y., Wang, B., et al. (2014). Overexpression of IbP5CR enhances salt tolerance in transgenic sweetpotato. Plant Cell Tissue Organ Cult. 117, 1–16. doi: 10.1007/s11240-013-0415-y
Liu, S. Y., Zhang, Y. W., Feng, Q. S., Qin, L., Pan, C. T., Lamin-Samu, A. T., et al. (2018). Tomato auxin response factor 5 regulates fruit set and development via the mediation of auxin and gibberellin signaling. Sci. Rep. 8:2971. doi: 10.1038/s41598-018-21315-y
Maggio, A., Miyazaki, S., Veronese, P., Fujita, T., and Ibeas, J. I. (2002). Does proline accumulation play an active role in stress-induced growth reduction? Plant J. 31, 699–712. doi: 10.1046/j.1365-313X.2002.01389.x
Mehrotra, R., Bhalothia, P., Bansal, P., Basantani, M. K., Bharti, V., and Mehrotra, S. (2014). Abscisic acid and abiotic stress tolerance-different tiers of regulation. J. Plant Physiol. 171, 486–496. doi: 10.1016/j.jplph.2013.12.007
Mei, L. Y., Yuan, Y. J., Wu, M. B., Gong, Z. H., et al. (2018). SlARF10, an auxin response factor, is required for chlorophyll and sugar accumulation during tomato fruit development. bioRxiv [Preprint]. doi: 10.1101/253237
Moreno, J. C., Cerda, A., Simpson, K., Lopez-Diaz, I., Carrera, E., Handford, M., et al. (2016). Increased Nicotiana tabacum fitness through positive regulation of carotenoid, gibberellin and chlorophyll pathways promoted by Daucus carota lycopene β-cyclase (Dclcyb1) expression. J. Exp. Bot. 67, 2325–2338. doi: 10.1093/jxb/erw037
Nagpal, P., Ellis, C. M., Weber, H., Ploense, S. E., Barkawi, L. S., Guilfoyle, T. J., et al. (2005). Auxin response factors ARF6 and ARF8 promote jasmonic acid production and flower maturation. Development 132, 4107–4118. doi: 10.1242/dev.01955
Nambara, E., and Marion-Poll, A. (2005). Abscisic acid biosynthesis and catabolism. Annu. Rev. Plant Biol. 56, 165–185. doi: 10.1146/annurev.arplant.56.032604.144046
Neisiani, F. F., Sanavy, S. A. M. M., Ghanati, F., and Dolatabadian, A. (2009). Effect of foliar application of pyridoxine on antioxidant enzyme activity, proline accumulation and lipid peroxidation of Maize (Zea mays L.) under water deficit. Not. Bot. Horti Agrobot. Cluj Napoca 37, 116–121.
Okushima, Y., Overvoorde, P. J., Arima, K., Alonso, J. M., Chan, A., Chang, C., et al. (2005). Functional genomic analysis of the AUXIN RESPONSE FACTOR gene family members in Arabidopsis thaliana: unique and overlapping functions of ARF7 and ARF19. Plant Cell 17, 444–463. doi: 10.1105/tpc.104.028316
Park, S., Kim, H. S., Jung, Y. J., Kim, S. H., Ji, C. Y., Wang, Z., et al. (2016). Orange protein has a role in phytoene synthase stabilization in sweetpotato. Sci. Rep. 6:33563. doi: 10.1038/srep33563
Ren, Z., Liu, R., Gu, W., and Dong, X. (2017). The Solanum lycopersicum auxin response factor SlARF2 participates in regulating lateral root formation and flower organ senescence. Plant Sci. 256, 103–111. doi: 10.1016/j.plantsci.2016.12.008
Römer, S., and Fraser, P. D. (2005). Recent advances in carotenoid biosynthesis, regulation and manipulation. Planta 221, 305–308. doi: 10.1007/s00425-005-1533-5
Sagar, M., Chervin, C., Mila, I., Hao, Y., Roustan, J. P., Benichou, M., et al. (2013). SlARF4, an auxin response factor involved in the control of sugar metabolism during tomato fruit development. Plant Physiol. 161, 1362–1374. doi: 10.1104/pp.113.213843
Sah, S. K., Reddy, K. R., and Li, J. X. (2016). Abscisic acid and abiotic stress tolerance in crop plants. Front. Plant Sci. 7:571. doi: 10.3389/fpls.2016.00571
Sandmann, G., Mer, S. R., and Fraser, P. D. (2006). Understanding carotenoid metabolism as a necessity for genetic engineering of crop plants. Metab. Eng. 8, 291–302. doi: 10.1016/j.ymben.2006.01.005
Schmittgen, T. D., and Livak, K. J. (2008). Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 3, 1101–1108. doi: 10.1038/nprot.2008.73
Schwartz, S. H., Qin, X., and Zeevaart, J. A. (2003). Elucidation of the indirect pathway of abscisic acid biosynthesis by mutants, genes, and enzymes. Plant Physiol. 131, 1591–1601. doi: 10.1104/pp.102.017921
Sripinyowanich, S., Klomsakul, P., Boonburapong, B., Bangyeekhun, T., Asami, T., Gu, H., et al. (2013). Exogenous ABA induces salt tolerance in indica rice (Oryza sativa L.): the role of OsP5CS1 and OsP5CR gene expression during salt stress. Environ. Exp. Bot. 86, 94–105. doi: 10.1016/j.envexpbot.2010.01.009
Takaichi, S. (2011). Carotenoids in algae distributions biosyntheses and functions. Mar. Drugs 9, 1101–1118. doi: 10.3390/md9061101
Teow, C. C., Truong, V., McFeeters, R. F., Thompson, R. L., Pecota, K. V., and Yencho, G. C. (2007). Antioxidant activities, phenolic and β-carotene contents of sweet potato genotypes with varying flesh colours. Food Chem. 103, 829–838. doi: 10.1016/j.foodchem.2006.09.033
Ulmasov, T., Hagen, G., and Guilfoyle, T. J. (1997). ARF1, a transcription factor that binds to auxin response elements. Science 276, 1865–1868. doi: 10.1126/science.276.5320.1865
Vishwakarma, K., Upadhyay, N., Kumar, N., Yadav, G., Singh, J., Mishra, R. K., et al. (2017). Abscisic acid signaling and abiotic stress tolerance in plants: a review on current knowledge and future prospects. Front. Plant Sci. 8:161. doi: 10.3389/fpls.2017.00161
Waller, F., Furuya, M., and Nick, P. (2002). OsARF1, an auxin response factor from rice, is auxin-regulated and classifies as a primary auxin responsive gene. Plant Mol. Biol. 50, 415–425. doi: 10.1023/A:1019818110761
Wang, L., Hua, D., He, J., Duan, Y., Chen, Z., Hong, X., et al. (2011). Auxin Response Factor 2 (ARF2) and its regulated homeodomain gene HB33 mediate abscisic acid response in Arabidopsis. PLoS Genet. 7:e1002172. doi: 10.1371/journal.pgen.1002172
Wang, W. B., Kim, Y. H., Lee, H. S., Kim, K. Y., and Deng, X. (2009). Analysis of antioxidant enzyme activity during germination of alfalfa under salt and drought stresses. Plant Physiol. Biochem. 47, 570–577. doi: 10.1016/j.plaphy.2009.02.009
Wang, Z., Ke, Q., Kim, M. D., Kim, S. H., Ji, C. Y., Jeong, J. C., et al. (2015). Transgenic alfalfa plants expressing the sweetpotato Orange gene exhibit enhanced abiotic stress tolerance. PLoS One 10:e0126050. doi: 10.1371/journal.pone.0126050
Wilmoth, J. C., Wang, S., Tiwari, S. B., Joshi, A. D., Hagen, G., Guilfoyle, T. J., et al. (2005). NPH4/ARF7 and ARF19 promote leaf expansion and auxin-induced lateral root formation. Plant J. 43, 118–130. doi: 10.1111/j.1365-313X.2005.02432.x
Xiong, L. M., and Zhu, J. K. (2003). Regulation of abscisic acid biosynthesis. Plant Physiol. 133, 29–36. doi: 10.1104/pp.103.025395
Yoshiba, Y., Kiyosue, T., Nakashima, K., Yamaguchishinozaki, K., and Shinozaki, K. (1997). Regulation of levels of proline as an osmolyte in plants under water stress. Plant Cell Physiol. 38, 1095–1102. doi: 10.1093/oxfordjournals.pcp.a029093
Yu, L., Zhai, H., Chen, W., He, S. Z., and Liu, Q. C. (2013). Cloning and functional analysis of lycopene ?-cyclase (IbLCYe) gene from sweetpotato, Ipomoea batatas (L.)lam. J. Integr. Agr. 12, 773–780. doi: 10.1016/S2095-3119(13)60299-3
Zhai, H., Wang, F. B., Si, Z. Z., Huo, J. X., Xing, L., An, Y. Y., et al. (2016). A myo-inositol-1-phosphate synthase gene, IbMIPS1, enhances salt and drought tolerance and stem nematode resistance in transgenic sweet potato. Plant Biotechnol. J. 14, 592–602. doi: 10.1111/pbi.12402
Keywords: sweetpotato, IbARF5, Arabidopsis, carotenoid content, salt and drought tolerance
Citation: Kang C, He S, Zhai H, Li R, Zhao N and Liu Q (2018) A Sweetpotato Auxin Response Factor Gene (IbARF5) Is Involved in Carotenoid Biosynthesis and Salt and Drought Tolerance in Transgenic Arabidopsis. Front. Plant Sci. 9:1307. doi: 10.3389/fpls.2018.01307
Received: 25 June 2018; Accepted: 17 August 2018;
Published: 11 September 2018.
Edited by:
Ruth Grene, Virginia Tech, United StatesReviewed by:
Kazuo Nakashima, Japan International Research Center for Agricultural Sciences, JapanSang-Soo Kwak, Korea Research Institute of Bioscience and Biotechnology (KRIBB), South Korea
Copyright © 2018 Kang, He, Zhai, Li, Zhao and Liu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Qingchang Liu, bGl1cWNAY2F1LmVkdS5jbg==