 Krishna C. Penumatsa1*
Krishna C. Penumatsa1* Ines Falcão-Pires2
Ines Falcão-Pires2 Sara Leite2
Sara Leite2 Adelino Leite-Moreira2Chinmayee D. Bhedi1Sabina Nasirova1Jing Ma3,4
Adelino Leite-Moreira2Chinmayee D. Bhedi1Sabina Nasirova1Jing Ma3,4 Roy L. Sutliff3,4
Roy L. Sutliff3,4 Barry L. Fanburg1*
Barry L. Fanburg1*- 1Pulmonary Critical Care and Sleep Division, Department of Medicine, Tufts Medical Center, Boston, MA, United States
- 2Faculty of Medicine of the University of Porto, Cardiovascular Research and Development Center, Porto, Portugal
- 3Department of Medicine, Atlanta Veterans Affairs and Emory University Medical Centers, Atlanta, GA, United States
- 4Department of Medicine, Emory University, Atlanta, GA, United States
Diastolic dysfunction of the heart and decreased compliance of the vasculature and lungs (i.e., increased organ tissue stiffness) are known features of obesity and the metabolic syndrome. Similarly, cardiac diastolic dysfunction is associated with aging. Elevation of the enzyme transglutaminase 2 (TG2) leads to protein cross-linking and enhanced collagen synthesis and participates as a candidate pathway for development of tissue stiffness. With these observations in mind we hypothesized that TG2 may be elevated in tissues of a rat model of obesity/metabolic syndrome (the ZSF 1 rat) and a mouse model of aging, i.e., the senescent SAMP8 mouse. In the experiments reported here, TG2 expression and activity were found for the first time to be spontaneously elevated in organs from both the ZSF1 rat and the SAMP8 mouse. These observations are consistent with a hypothesis that a TG2-related pathway may participate in the known tissue stiffness associated with cardiac diastolic dysfunction in these two rodent models. The potential TG2 pathway needs better correlation with physiologic dysfunction and may eventually provide novel therapeutic insights to improve tissue compliance.
Introduction
It is well known that the pathophysiology and progression of multiple human diseases are associated with tissue stiffness (or loss of compliance). Although such disease processes may also involve other non-contractile organs such as the kidney and liver, they are most evident in contractile tissues such as heart, lungs, and the vasculature. The stiffness is thought to be largely derived from changes in tissue extracellular matrix that consists of proteins such as fibronectin, collagen, and elastin. While stiffness of heart tissue may result in congestive heart failure, that of the lungs may produce respiratory failure and that of the circulation increases risk of hypertension.
The multifunctional enzyme transglutaminase 2 (TG2) is well known to post-translationally modify proteins by either creating cross-links between proteins at glycine and lysine residues (Greenberg et al., 1991; Griffin et al., 2002) or by the proteins with primary amine groups (Penumatsa and Fanburg, 2014; Muma and Mi, 2015; Lai and Lin, 2017). The food industry utilizes bacterial transglutaminase to enhance palatability of meats by a stiffening process (Kieliszek and Misiewicz, 2014). There has been a gradual accumulation of data that associates TG2 with synthesis and cross-linking of extracellular matrix proteins that produces tissue stiffness (Mosher and Schad, 1979; Orban et al., 2004). Highlighting the critical role of TG2 in tissue remodeling, recent studies by Santhanam et al. demonstrated that TG2 mediates ventricular (Oh et al., 2017) and aortic (Steppan et al., 2017) contractility, stiffness and function. Furthermore, the events of TG2 alterations are intricately interwoven with influences on transcription factors (Kumar and Mehta, 2012; Brown, 2013; Penumatsa et al., 2014), inflammation (Liu et al., 2019), oxidative stress (Lee et al., 2019), and fibrogenic signaling (Olsen et al., 2014; Penumatsa et al., 2017). Thus, TG2 is a potential target for disruption of tissue stiffness that occurs in diseases of the cardiovascular system and lungs.
In the present study, TG2 expression and activity were measured by immunoblotting in rodent models of obesity/metabolic syndrome and aging. Serotonylation of fibronectin (sFn), a technique previously developed by our laboratory (Liu et al., 2011), was used as a surrogate marker for increased TG2 transamidation activity. Our data indicate for the first-time that metabolic dysfunction and aging are associated with increased TG2 expression and activity in multiple tissues, including heart and lungs.
Materials and Methods
Animal Models
Male Wistar Kyoto (control; n = 6), ZSF1 lean (n = 6), and obese (n = 6) rats were obtained from Charles River (Barcelona, Spain) and fed with Purina Diet (#5008). By their 18th week of age, rats underwent metabolic, aerobic capacity, and echocardiographic evaluation. One to two weeks later, the rats were deeply anesthetized with 8% sevoflurane and immediately euthanized by exsanguination. Tissue samples were collected for molecular studies, as described below. All rat studies followed the recommendation of the Guide for the Care and Use of Laboratory Animals, published by the US National Institutes of Health (NIH Publication No. 85–23, Revised 2011) and were certified by the Portuguese Veterinary Governmental Authorities, approved by the Portuguese Foundation for Science and Technology and by the ethical committee of the institution. Only trained researchers, certified with a Laboratory Animal Science course according to the Federation of European Laboratory Animal Science Associations, performed rat handling and procedures. Aging mouse models, senescence-accelerated prone mice (SAMP8; n = 6) and senescence-accelerated resistant mice (SAMR1; control; n = 6) were purchased from Envigo (Formerly Harlan) (Indianapolis, IN, United States). All mouse studies were carried out using male 30-week-old mice fed ad libitum with irradiated 18% protein global rodent diet (Teklad 2918) (Reed et al., 2011). All mouse studies were performed in accordance with protocols approved by the Atlanta Veterans Affairs Institutional Animal Care and Use Committee.
Aerobic Capacity and Exercise Tolerance
On week before euthanasia, the rats were placed on a treadmill chamber coupled to a gas analyzer (LE8700C and LE405, Panlab Harvard Apparatus®). The gas flow was set at 700 mL.min–1 and treadmill was tilted at 10°. The adaptation was carried out at a speed of 15 cm.s–1 for 3 min. The maximum stress test started at a speed of 30 cm.s–1, with increments of 5 cm.s–1 every minute until the rats reached maximal aerobic capacity (VO2 max).
Metabolic Studies
After 12 h of fasting, rats underwent glycemic measurements (Freestyle-Mini) at baseline and after 15, 30, 60, 90, and 120 min of 1 g.Kg–1 glucose gavage (oral glucose tolerance test) or a 0.5 U.Kg–1 intraperitoneal insulin injection (insulin resistance test).
Echocardiographic Evaluation
Anesthetized rats (3–4% sevoflurane) were placed on a heating pad and the ECG was monitored. A linear 15 MHz probe (Sequoia 15L8W) was used and a parasternal short axis view was performed to record M-mode as previously described (Hamdani et al., 2013). Aortic and mitral flow tracing velocity were recorded by pulsed-wave Doppler. Peak systolic tissue velocity and E’ were measured with tissue Doppler at the medial mitral annulus. Maximal left atrial dimensions were measured by 2D echocardiography in a four-chamber view. Acquisitions were done while transiently suspending mechanical ventilation and recordings from three consecutive heartbeats were averaged.
Western Blot Analysis
Tissue samples of both rats and mice were minced and homogenized in NP-40 lysis buffer (Boston Bio Products) containing protease and phosphatase inhibitor cocktails (MilliporeSigma). Tissue homogenates were centrifuged, and the protein content was determined using a Bradford Assay (Bio-Rad Laboratories). Reduced samples were then subjected to SDS-PAGE as described previously (Liu et al., 2011; Penumatsa et al., 2014). Western blotting analysis of total protein lysates were performed using the following primary antibodies: serotonin (5-HT; Sigma, Cat# S5545); transglutaminase 2 (TG2; Santa Cruz Biotechnology, Cat# sc-48387); β-actin (Cell Signaling Technology, Cat# 4970) and GAPDH (Cell Signaling Technology). Horseradish peroxidase-conjugated secondary antibodies (Santa Cruz Biotechnology) were used for immunoreactive protein detection. Serotonylation of fibronectin (sFN) represents TG2 transamidation activity (Liu et al., 2011). An ECL Western blotting substrate (Thermo Fisher Scientific) was used to detect the protein bands. Densitometry analysis was performed using Un-Scan-It gel analysis software (Silk Scientific, Orem, UT) as previously described (Penumatsa et al., 2014). The band intensity of the target protein was normalized against the β-Actin or GAPDH bands and the data was expressed as fold change in expression compared to respective controls.
Statistical Analysis
Data were expressed as mean ratios ± standard error of mean (SEM). Student’s t-test or One-way analysis of variance (ANOVA) followed by Tukey’s post hoc test (SigmaPlot 12.5 software; Systat Software, San Jose, CA, United States) were used to compare experimental groups. A p < 0.05 was considered statistically significant.
Results
TG2 Expression and Activity Are Increased in ZSF1 Obese Rats
Consistent with our previous studies (Hamdani et al., 2013), obese rats presented higher body, heart and peri-renal fat weight than lean ZSF1 and Wistar-Kyoto rats (Table 1). In addition, Obese ZSF1 rats showed hyperglycemia, impaired glucose tolerance and insulin resistance, consistent with the diagnosis of metabolic syndrome. Obese rats presented signs of congestion of the lungs, as assessed by increased weight and effort intolerance (lower values of VO2 max). Moreover, Table 1 displays other features that indicate a phenotype of HFpEF in ZSF1 obese rats (Leite et al., 2015a,b; van Dijk et al., 2016) and not in ZSF1 lean or Wistar Kyoto rats, such as normal left ventricular systolic function, as observed by similar cardiac index (CI) and ejection fraction (EF), and signs of diastolic dysfunction (increased E/e’ and left atrium area).
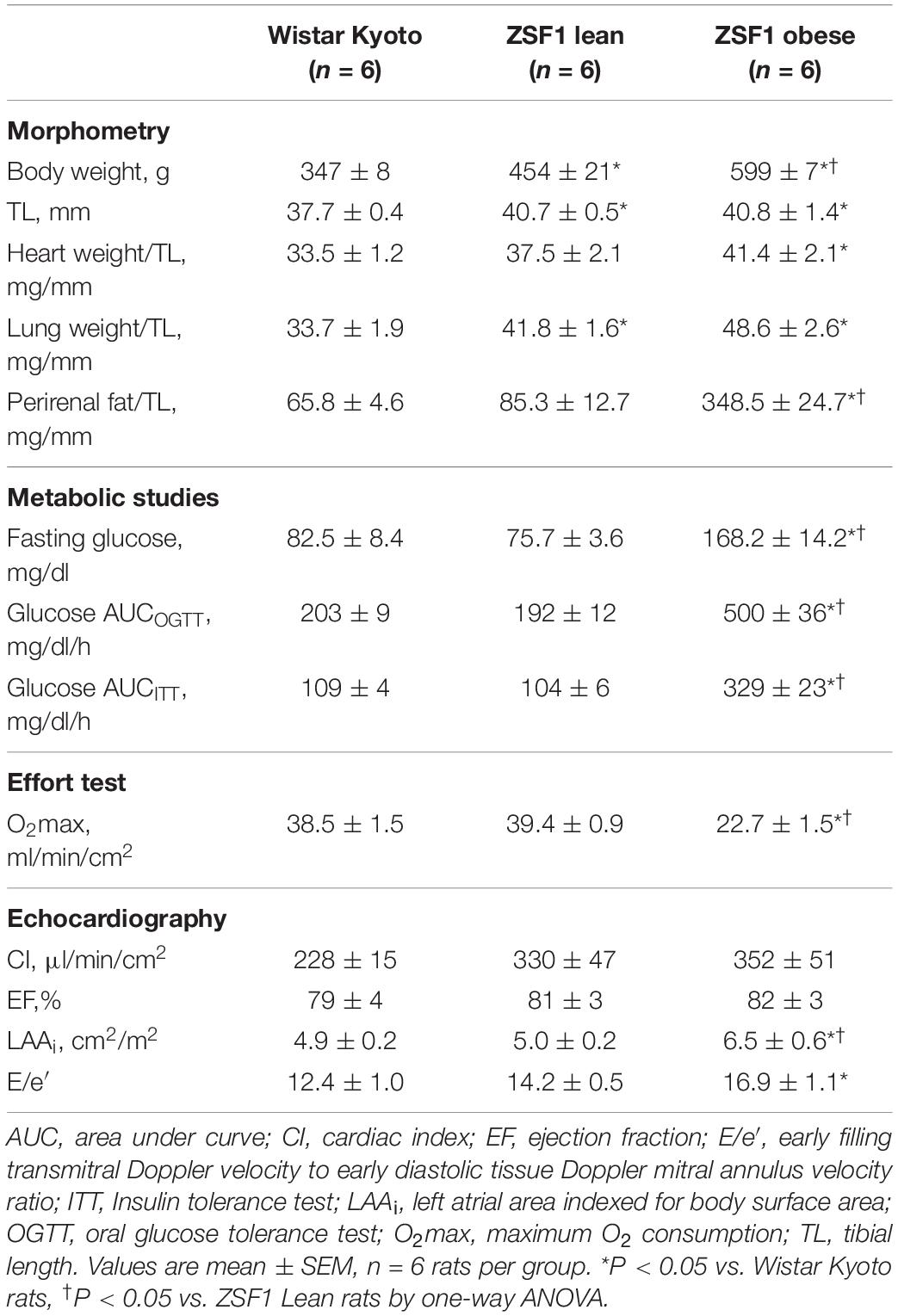
Table 1. Hemodynamic and metabolic changes in ZSF1 obese rats.
Given that the ZSF1 obese rats show signs of metabolic dysfunction, we then assessed the TG2 expression and activity in multiple tissues. TG2 expression was significantly increased in the lungs (Figure 1A) of ZSF1 obese rats compared to control Wistar Kyoto rats. Additionally, we found increased TG2 transamidation activity as measured by sFn in lung tissues of both lean and obese ZSF1 rats compared to Wistar Kyoto rats (Figure 1A). Consistent with the observation that the ZSF1 obese rats show left ventricular diastolic dysfunction, both TG2 expression and activity were significantly increased in the left ventricles (Figure 1B). Consistent with the diabetic nephropathy observed in ZSF1 obese rats, TG2 levels are increased in kidneys (Figure 1C) and livers (Figure 1D) of this rat model.
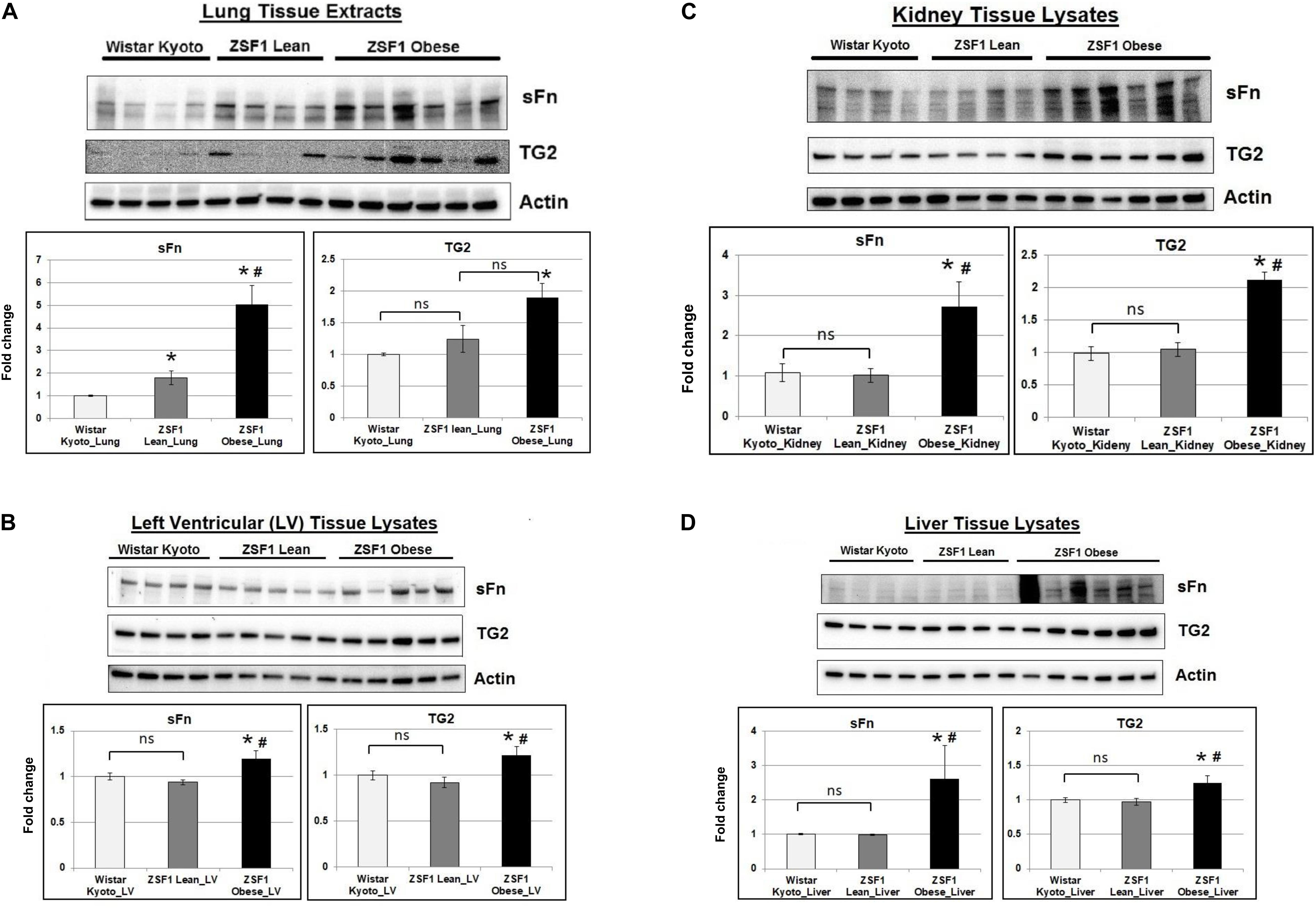
Figure 1. TG2 activity and expression are increased in a rat model of metabolic syndrome. Representative Western blot images showing serotonylated fibronectin (sFn) and transglutaminase 2 (TG2) expression levels in (A) lung, (B) cardiac left ventricular, (C) kidney, and (D) liver tissues of Wistar Kyoto, ZSF1 lean and ZSF1 obese rats. Beta-actin was used as a loading control. Bar graphs showing the normalized fold change differences compared to Wistar Kyoto rats. Data are expressed as mean ± SEM. n ≥ 4 rats per group. *p < 0.05 compared to Wistar Kyoto rats. #p < 0.05 compared to ZSF1 lean rats by one-way ANOVA. ns = not significant.
TG2 Expression and Activity Are Increased in SAMP8 Mice
The senescence-accelerated prone mouse (SAMP8) model of aging demonstrates several hallmarks of human cardiovascular pathology including aging-associated oxidative stress (Gan et al., 2012), metabolic dysfunction (Kurokawa et al., 1998), inflammation (Forman et al., 2011) and HFpEF (Reed et al., 2011). To determine the impact of aging on pro-fibrogenic phenotype, TG2 expression and activity were measured by Western blot analysis. We report here for the first-time that TG2 expression and activity are increased in lungs (Figure 2A) and left ventricles (Figure 2B) of senescent-prone (SAMP8) mice compared to control senescent-resistant (SAMR1) mice.
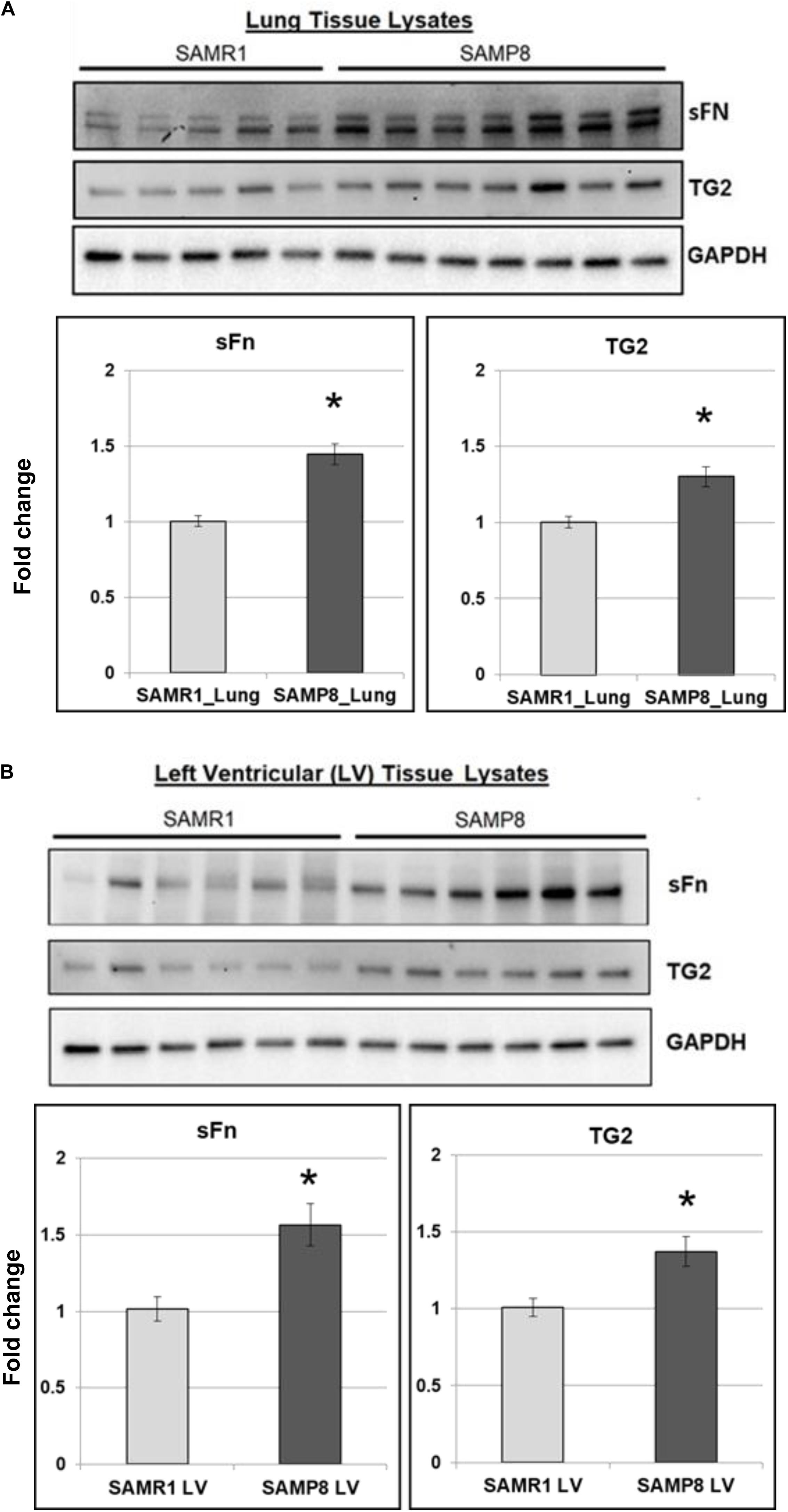
Figure 2. TG2 activity and expression and fibrogenic markers are increased in mouse model of aging. Representative Western blot images showing serotonylated fibronectin (sFn) and transglutaminase 2 (TG2) expression levels in (A) lungs and (B) left ventricles of senescence-resistant (SAMR1) and senescence-accelerated (SAMP8) mice. GAPDH was used as a loading control. Bar graphs showing the normalized fold change differences compared to SAMR1 mice. Data are expressed as mean ± SEM. n ≥ 5 mice per group. *p < 0.05 compared to SAMR1 mice by t-test.
Discussion
Our present studies establish that expression and activity of TG2, a protein cross- linking enzyme with fibrogenic properties, are increased in multiple organs from rodent models of both metabolic dysfunction and senescence. Additionally, we have reported previously that hypoxia (Penumatsa et al., 2017) and hyperglycemia (Bhedi et al., 2020) stimulate expression and activity of TG2 in cardiac and pulmonary vascular fibroblasts in culture. Although we have not yet specifically related TG2 activity to tissue compliance, our present in vivo observations with rodent models of metabolic syndrome (ZSF1 obese rats) and aging (SAMP8 mice) are consistent with participation of TG2 in cardiac and lung matrix changes that are associated with tissue stiffness in multiple cardiovascular and pulmonary diseases mediated by these conditions.
TG2 in Pressure Overload Induced Cardiopulmonary Remodeling
The multifunctional enzyme TG2 is now receiving attention for its potential role in vascular (Bakker et al., 2006; Penumatsa et al., 2014; Steppan et al., 2014), cardiac (Kashima et al., 1997; Small et al., 1999; Wang et al., 2018), and pulmonary (DiRaimondo et al., 2014; Olsen et al., 2014) diseases. Our previous studies have identified that TG2 activity is largely controlled by physiologic events associated with increased cardiac pressure overload models, in particular those of chronic hypoxia (DiRaimondo et al., 2014; Penumatsa et al., 2017) and transverse aortic constriction (Bhedi et al., 2020). Studies by others have largely supported this concept (Baandrup et al., 2011; Shinde et al., 2018; Wang et al., 2018). A particular phenotype of cardiac disease characterized by increased stiffness or impaired relaxation is that of diastolic dysfunction. When combined with signs and symptoms of heart failure and a normal systolic function, diastolic dysfunction leads to heart failure with preserved ejection fraction (HFpEF) (Kass et al., 2004; Piccini et al., 2004; Reed et al., 2011). In addition, a possible common denominator of these events is that TG2 activity is under regulation by glycolysis and perhaps specifically the terminal glycolytic enzyme phosphokinase M2 (Bhedi et al., 2020).
TG2 in Hyperglycemia Mediated Cardiopulmonary Remodeling
There is now general recognition that the pathology of diabetes mellitus (DM) includes remodeling of extracellular tissue and intercellular matrices with the formation of fibrosis and collagen alterations that promote tissue stiffness (Ban and Twigg, 2008; Law et al., 2012). Although fibrosis may be present in any organ of the body (Gilbert and Cooper, 1999; Patil et al., 2011), its involvement of the cardiovascular system is particularly prevalent in DM. While there may be more subtle processes involving the pulmonary arterial stiffness and cardiac right ventricle in association with pulmonary hypertension (PH) in DM (Grinnan et al., 2016; Whitaker et al., 2018), metabolic dysfunction has been identified in particular (Irwin et al., 2014; Lai et al., 2019). Up to 30% of patients with HFpEF have DM (Piccini et al., 2004). The basic mechanisms by which the intercellular matrix alteration occurs in DM are receiving increased attention. Appealing theories regarding mechanisms include direct effects of hyperglycemia on fibroblasts to promote their transition to myofibroblasts and subsequent fibrosis; glycation of proteins to form advanced glycation end products (AGEs) that alter the tissue matrices; and increased oxidative stress and inflammation (Asbun and Villarreal, 2006; Ban and Twigg, 2008). Studies have demonstrated associations between hyperglycemia (Skill et al., 2004; Bhedi et al., 2020), reactive oxygen species (Lee et al., 2019) and inflammatory (Liu et al., 2019) responses and TG2 regulation. Of interest, TG2 -mediated protein serotonylation has been found to be modulated in pancreatic beta cells (Paulmann et al., 2009).
We have proposed that TG2 is an important mediator of an alteration in intercellular matrix protein cross-linking that occurs in DM. We have tested a mouse model of type 2 diabetes, the Akita mouse, for expression and activity of TG2 in multiple organs and have identified an elevation of TG2 expression and activity in all organs studied as compared to controls (Bhedi et al., 2020). These observations further led us to examine the response of fibroblasts in culture to elevated glucose. We reported that in addition to stimulation of TG2, glucose elevates glycolysis, synthesis of TGFβ1 and cellular collagen content (Bhedi et al., 2020). In these studies we found that elevated synthesis of TGFβ1, type 1 collagen and α-smooth muscle actin are blocked by inhibition of TG2 or glycolysis (Bhedi et al., 2020). These observations suggest that enhanced glycolysis and TG2 production induced by hyperglycemia may lead to enhanced tissue stiffness either as a result of fibrosis or alteration of cardiac contractile mechanisms influenced by TG2.
TG2 in Obesity/Metabolic Syndrome Mediated Cardiopulmonary Remodeling
It is well known that alterations in tissue matrix reflected as fibrosis or other alterations resulting in increased tissue stiffness occur in obesity and specifically in obesity associated with metabolic syndrome (Cavalera et al., 2014; Aroor et al., 2019). These features of the disease manifest as HFpEF and loss of pulmonary and systemic arterial compliance (Lai et al., 2019; Leite et al., 2019). Although animal models of the metabolic syndrome, such as the one used in the present study, have been identified as having increased tissue stiffness, the levels of TG2 activity in tissues of these animal models have never been evaluated. The present study was designed to address this deficiency. We have utilized ZSF1 rats with obesity, diabetes and hypertension as a model of the metabolic syndrome. This model has been used previously by Hamdani et al. for studies in which they demonstrated the presence of left ventricular diastolic dysfunction (Hamdani et al., 2013). With this model, we have now found for the first-time elevation of both TG2 expression and activity in multiple tissues including heart, kidney and liver of ZSF1 obese rats compared to ZSF1 lean and control ones. These results suggest a more generalized elevation of tissue TG2 in these animals, perhaps attributed to a hyperglycemia and/or an inflammatory phenotype. Interestingly, TG2 was elevated in lungs of both lean and obese ZSF1 rats compared control rats. Given that both lean and obese ZSF1 rats have previously shown elevated systemic blood pressure (Griffin et al., 2007), further studies are needed to assess the pulmonary circulation in these rat models.
TG2 in Aging Mediated Cardiopulmonary Remodeling
Congestive heart failure is often associated with cardiac fibrosis and HFpEF (Wong et al., 1989; Biernacka and Frangogiannis, 2011; Loffredo et al., 2014; Diez-Villanueva and Alfonso, 2016). Clinical prevalence of HFpEF increases with aging (Fleg and Strait, 2012; Strait and Lakatta, 2012; Loffredo et al., 2014). Aging-associated fibrosis also occurs in other organs, including lungs (Schafer et al., 2017; Sicard et al., 2018); the prevalence of chronic lung diseases and idiopathic pulmonary fibrosis in the elderly may reflect this occurrence (Budinger et al., 2017). It has been previously reported that TG2 mediates cardiac ventricular (Oh et al., 2017) and systemic arterial (Armstrong et al., 2018) stiffness in rodent models of aging. Similar to that of the obesity/metabolic syndrome, it has been proposed that the aging process may be under the regulation of enhanced glycolysis with reduced mitochondrial function (James et al., 2015; Feng et al., 2016; Cho et al., 2017; Jang et al., 2018). Of note, altered glucose metabolism (Costantino et al., 2016; Cho et al., 2017; Zank et al., 2018) and inflammation (including elevation of cytokines) (Maggio et al., 2006; Forman et al., 2011) have been recognized to be driving metabolic forces in aging-associated tissue dysfunction.
Our current findings extend previous observations indicating an elevation of TG2 expression and activity in hearts and lungs of mouse models that experience glycolysis-inducing events such as exposure to hypoxia and transverse aortic banding which imposes a pressure overload on the left ventricle (Bhedi et al., 2020). Further studies are needed to determine the mechanisms that promote increased TG2 in these animal models and to relate these findings to the physiologic process of decreased tissue compliance. Since small molecule inhibitors of TG2 are currently available (Kim, 2018) and anti-TG2 monoclonal antibodies are being developed, these novel therapeutics may prove to be useful in modifying tissue stiffness occurring with alterations in tissue pressure-overload, diabetes, obesity/metabolic syndrome or aging.
Data Availability Statement
All datasets generated for this study are included in the article/supplementary material.
Ethics Statement
The animal studies were reviewed and approved by the Portuguese Foundation for Science and Technology or Atlanta VA IACUC.
Author Contributions
KP, IF-P, RS, and BF conceived and designed the experiments and wrote the manuscript. KP, IF-P, SL, AL-M, CB, SN, JM, and RS performed the experiments. KP, IF-P, SL, AL-M, CB, and SN analyzed the data. All authors contributed to the final manuscript.
Funding
This research was supported by the funding from an American Heart Association Career Development Award 18CDA34140005 (KP), Tupper Research Fund at Tufts Medical Center (KP), the National Institutes of Health AG064064 (KP), HL107713 (BF), and HL070892 (RS), the Fundo Europeu de Desenvolvimento Regional (FEDER) through Compete 2020 – Programa Operacional Competitividade e Internacionalização (POCI) and the project NETDIAMOND (POCI-01-0145-FEDER-016385), which was supported by European Structural and Investment Funds, Lisbon’s regional operational program 2020.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Dulce Fontoura for support with the rat model maintenance.
References
Armstrong, D. M. F., Sikka, G., Armstrong, A. D. C., Saad, K. R., Freitas, W. R., Berkowitz, D. E., et al. (2018). Knockdown of transglutaminase-2 prevents early age-induced vascular changes in mice1. Acta Cir. Bras. 33, 991–999. doi: 10.1590/s0102-865020180110000006
Aroor, A. R., Habibi, J., Nistala, R., Ramirez-Perez, F. I., Martinez-Lemus, L. A., Jaffe, I. Z., et al. (2019). Diet-induced obesity promotes kidney endothelial stiffening and fibrosis dependent on the endothelial mineralocorticoid receptor. Hypertension 73, 849–858. doi: 10.1161/hypertensionaha.118.12198
Asbun, J., and Villarreal, F. J. (2006). The pathogenesis of myocardial fibrosis in the setting of diabetic cardiomyopathy. J. Am. Coll. Cardiol. 47, 693–700. doi: 10.1016/j.jacc.2005.09.050
Baandrup, J. D., Markvardsen, L. H., Peters, C. D., Schou, U. K., Jensen, J. L., Magnusson, N. E., et al. (2011). Pressure load: the main factor for altered gene expression in right ventricular hypertrophy in chronic hypoxic rats. PLoS One 6:e15859. doi: 10.1371/journal.pone.0015859
Bakker, E. N., Pistea, A., Spaan, J. A., Rolf, T., de Vries, C. J., van Rooijen, N., et al. (2006). Flow-dependent remodeling of small arteries in mice deficient for tissue-type transglutaminase: possible compensation by macrophage-derived factor XIII. Circ. Res. 99, 86–92. doi: 10.1161/01.res.0000229657.83816.a7
Ban, C. R., and Twigg, S. M. (2008). Fibrosis in diabetes complications: pathogenic mechanisms and circulating and urinary markers. Vasc. Health Risk Manag. 4, 575–596. doi: 10.2147/vhrm.s1991
Bhedi, C. D., Nasirova, S., Toksoz, D., Warburton, R. R., Morine, K. J., Kapur, N. K., et al. (2020). Glycolysis regulated transglutaminase 2 activation in cardiopulmonary fibrogenic remodeling. FASEB J. 34, 930–944. doi: 10.1096/fj.201902155r
Brown, K. D. (2013). Transglutaminase 2 and NF-kappaB: an odd couple that shapes breast cancer phenotype. Breast Cancer Res. Treat. 137, 329–336. doi: 10.1007/s10549-012-2351-7
Budinger, G. R. S., Kohanski, R. A., Gan, W., Kobor, M. S., Amaral, L. A., Armanios, M., et al. (2017). The intersection of aging biology and the pathobiology of lung diseases: a joint NHLBI/NIA workshop. J. Gerontol. A Biol. Sci. Med. Sci. 72, 1492–1500. doi: 10.1093/gerona/glx090
Cavalera, M., Wang, J., and Frangogiannis, N. G. (2014). Obesity, metabolic dysfunction, and cardiac fibrosis: pathophysiological pathways, molecular mechanisms, and therapeutic opportunities. Transl. Res. 164, 323–335. doi: 10.1016/j.trsl.2014.05.001
Cho, S. J., Moon, J. S., Lee, C. M., Choi, A. M., and Stout-Delgado, H. W. (2017). Glucose transporter 1-dependent glycolysis is increased during aging-related lung fibrosis, and phloretin inhibits lung fibrosis. Am. J. Respir. Cell Mol. Biol. 56, 521–531. doi: 10.1165/rcmb.2016-0225oc
Costantino, S., Paneni, F., and Cosentino, F. (2016). Ageing, metabolism and cardiovascular disease. J. Physiol. 594, 2061–2073. doi: 10.1113/jp270538
Diez-Villanueva, P., and Alfonso, F. (2016). Heart failure in the elderly. J. Geriatr. Cardiol. 13, 115–117.
DiRaimondo, T. R., Klock, C., Warburton, R., Herrera, Z., Penumatsa, K., Toksoz, D., et al. (2014). Elevated transglutaminase 2 activity is associated with hypoxia-induced experimental pulmonary hypertension in mice. ACS Chem. Biol. 9, 266–275. doi: 10.1021/cb4006408
Feng, Z., Hanson, R. W., Berger, N. A., and Trubitsyn, A. (2016). Reprogramming of energy metabolism as a driver of aging. Oncotarget 7, 15410–15420. doi: 10.18632/oncotarget.7645
Fleg, J. L., and Strait, J. (2012). Age-associated changes in cardiovascular structure and function: a fertile milieu for future disease. Heart Fail. Rev. 17, 545–554. doi: 10.1007/s10741-011-9270-2
Forman, K., Vara, E., Garcia, C., Kireev, R., Cuesta, S., Escames, G., et al. (2011). Effect of a combined treatment with growth hormone and melatonin in the cardiological aging on male SAMP8 mice. J. Gerontol. A Biol. Sci. Med. Sci. 66, 823–834. doi: 10.1093/gerona/glr083
Gan, W., Nie, B., Shi, F., Xu, X. M., Qian, J. C., Takagi, Y., et al. (2012). Age-dependent increases in the oxidative damage of DNA, RNA, and their metabolites in normal and senescence-accelerated mice analyzed by LC-MS/MS: urinary 8-oxoguanosine as a novel biomarker of aging. Free Radic. Biol. Med. 52, 1700–1707. doi: 10.1016/j.freeradbiomed.2012.02.016
Gilbert, R. E., and Cooper, M. E. (1999). The tubulointerstitium in progressive diabetic kidney disease: more than an aftermath of glomerular injury? Kidney Int. 56, 1627–1637. doi: 10.1046/j.1523-1755.1999.00721.x
Greenberg, C. S., Birckbichler, P. J., and Rice, R. H. (1991). Transglutaminases: multifunctional cross-linking enzymes that stabilize tissues. FASEB J. 5, 3071–3077. doi: 10.1096/fasebj.5.15.1683845
Griffin, K. A., Abu-Naser, M., Abu-Amarah, I., Picken, M., Williamson, G. A., and Bidani, A. K. (2007). Dynamic blood pressure load and nephropathy in the ZSF1 (fa/fa cp) model of type 2 diabetes. Am. J. Physiol. Renal Physiol. 293, F1605–F1613.
Griffin, M., Casadio, R., and Bergamini, C. M. (2002). Transglutaminases: nature’s biological glues. Biochem. J. 368(Pt 2), 377–396. doi: 10.1042/BJ20021234
Grinnan, D., Farr, G., Fox, A., and Sweeney, L. (2016). The role of hyperglycemia and insulin resistance in the development and progression of pulmonary arterial hypertension. J. Diabetes Res. 2016:2481659.
Hamdani, N., Franssen, C., Lourenco, A., Falcao-Pires, I., Fontoura, D., Leite, S., et al. (2013). Myocardial titin hypophosphorylation importantly contributes to heart failure with preserved ejection fraction in a rat metabolic risk model. Circ. Heart Fail. 6, 1239–1249. doi: 10.1161/circheartfailure.113.000539
Irwin, D. C., Garat, C. V., Crossno, J. T. Jr., MacLean, P. S., Sullivan, T. M., Erickson, P. F., et al. (2014). Obesity-related pulmonary arterial hypertension in rats correlates with increased circulating inflammatory cytokines and lipids and with oxidant damage in the arterial wall but not with hypoxia. Pulm. Circ. 4, 638–653. doi: 10.1086/678510
James, E. L., Michalek, R. D., Pitiyage, G. N., de Castro, A. M., Vignola, K. S., Jones, J., et al. (2015). Senescent human fibroblasts show increased glycolysis and redox homeostasis with extracellular metabolomes that overlap with those of irreparable DNA damage, aging, and disease. J. Proteome Res. 14, 1854–1871. doi: 10.1021/pr501221g
Jang, J. Y., Blum, A., Liu, J., and Finkel, T. (2018). The role of mitochondria in aging. J. Clin. Invest. 128, 3662–3670.
Kashima, K., Yokoyama, S., Daa, T., Nakayama, I., and Iwaki, T. (1997). Immunohistochemical study on tissue transglutaminase and copper-zinc superoxide dismutase in human myocardium: its relevance to apoptosis detected by the nick end labelling method. Virchows Arch. 430, 333–338. doi: 10.1007/bf01092757
Kass, D. A., Bronzwaer, J. G., and Paulus, W. J. (2004). What mechanisms underlie diastolic dysfunction in heart failure? Circ. Res. 94, 1533–1542. doi: 10.1161/01.res.0000129254.25507.d6
Kieliszek, M., and Misiewicz, A. (2014). Microbial transglutaminase and its application in the food industry. A review. Folia Microbiol. 59, 241–250. doi: 10.1007/s12223-013-0287-x
Kim, S. Y. (2018). New insights into development of transglutaminase 2 inhibitors as pharmaceutical lead compounds. Med. Sci. 6:87. doi: 10.3390/medsci6040087
Kumar, S., and Mehta, K. (2012). Tissue transglutaminase constitutively activates HIF-1alpha promoter and nuclear factor-kappaB via a non-canonical pathway. PLoS One 7:e49321. doi: 10.1371/journal.pone.0049321
Kurokawa, T., Ozaki, N., and Ishibashi, S. (1998). Difference between senescence-accelerated prone and resistant mice in response to insulin in the heart. Mech. Ageing Dev. 102, 25–32. doi: 10.1016/s0047-6374(98)00009-8
Lai, T. S., and Lin, C. J. (2017). Greenberg CS. Role of tissue transglutaminase-2 (TG2)-mediated aminylation in biological processes. Amino Acids 49, 501–515. doi: 10.1007/s00726-016-2270-8
Lai, Y. C., Wang, L., and Gladwin, M. T. (2019). Insights into the pulmonary vascular complications of heart failure with preserved ejection fraction. J. Physiol. 597, 1143–1156. doi: 10.1113/jp275858
Law, B., Fowlkes, V., Goldsmith, J. G., Carver, W., and Goldsmith, E. C. (2012). Diabetes-induced alterations in the extracellular matrix and their impact on myocardial function. Microsc. Microanal. 18, 22–34. doi: 10.1017/s1431927611012256
Lee, J. Y., Lee, Y. J., Jeon, H. Y., Han, E. T., Park, W. S., Hong, S. H., et al. (2019). The vicious cycle between transglutaminase 2 and reactive oxygen species in hyperglycemic memory-induced endothelial dysfunction. FASEB J. 33, 12655–12667. doi: 10.1096/fj.201901358rr
Leite, S., Cerqueira, R. J., Ibarrola, J., Fontoura, D., Fernandez-Celis, A., Zannad, F., et al. (2019). Arterial remodeling and dysfunction in the ZSF1 rat model of heart failure with preserved ejection fraction. Circ. Heart Fail. 12:e005596.
Leite, S., Oliveira-Pinto, J., Tavares-Silva, M., Abdellatif, M., Fontoura, D., Falcao-Pires, I., et al. (2015a). Echocardiography and invasive hemodynamics during stress testing for diagnosis of heart failure with preserved ejection fraction: an experimental study. Am. J. Physiol. Heart Circ. Physiol. 308, H1556–H1563.
Leite, S., Rodrigues, S., Tavares-Silva, M., Oliveira-Pinto, J., Alaa, M., Abdellatif, M., et al. (2015b). Afterload-induced diastolic dysfunction contributes to high filing pressures in experimental heart failure with preserved ejection fraction. Am. J. Physiol. Heart Circ. Physiol. 309, H1648–H1654.
Liu, C., Luo, R., Wang, W., Peng, Z., Johnson, G. V. W., Kellems, R. E., et al. (2019). Tissue transglutaminase-mediated AT1 receptor sensitization underlies pro-inflammatory cytokine LIGHT-induced hypertension. Am. J. Hypertens. 32, 476–485. doi: 10.1093/ajh/hpz018
Liu, Y., Wei, L., Laskin, D. L., and Fanburg, B. L. (2011). Role of protein transamidation in serotonin-induced proliferation and migration of pulmonary artery smooth muscle cells. Am. J. Respir. Cell Mol. Biol. 44, 548–555. doi: 10.1165/rcmb.2010-0078oc
Loffredo, F. S., Nikolova, A. P., Pancoast, J. R., and Lee, R. T. (2014). Heart failure with preserved ejection fraction: molecular pathways of the aging myocardium. Circ. Res. 115, 97–107. doi: 10.1161/circresaha.115.302929
Maggio, M., Guralnik, J. M., Longo, D. L., and Ferrucci, L. (2006). Interleukin-6 in aging and chronic disease: a magnificent pathway. J. Gerontol. A Biol. Sci. Med. Sci. 61, 575–584. doi: 10.1093/gerona/61.6.575
Mosher, D. F., and Schad, P. E. (1979). Cross-linking of fibronectin to collagen by blood coagulation Factor XIIIa. J. Clin. Invest. 64, 781–787. doi: 10.1172/JCI109524
Muma, N. A., and Mi, Z. (2015). Serotonylation and transamidation of other monoamines. ACS Chem. Neurosci. 6, 961–969. doi: 10.1021/cn500329r
Oh, Y. J., Pau, V. C., Steppan, J., Sikka, G., Bead, V. R., Nyhan, D., et al. (2017). Role of tissue transglutaminase in age-associated ventricular stiffness. Amino Acids 49, 695–704. doi: 10.1007/s00726-016-2295-z
Olsen, K. C., Epa, A. P., Kulkarni, A. A., Kottmann, R. M., McCarthy, C. E., Johnson, G. V., et al. (2014). Inhibition of transglutaminase 2, a novel target for pulmonary fibrosis, by two small electrophilic molecules. Am. J. Respir. Cell Mol. Biol. 50, 737–747. doi: 10.1165/rcmb.2013-0092OC
Orban, J. M., Wilson, L. B., Kofroth, J. A., El-Kurdi, M. S., Maul, T. M., and Vorp, D. A. (2004). Crosslinking of collagen gels by transglutaminase. J. Biomed. Mater. Res. A 68, 756–762. doi: 10.1002/jbm.a.20110
Patil, V. C., Patil, H. V., Shah, K. B., Vasani, J. D., and Shetty, P. (2011). Diastolic dysfunction in asymptomatic type 2 diabetes mellitus with normal systolic function. J. Cardiovasc. Dis. Res. 2, 213–222. doi: 10.4103/0975-3583.89805
Paulmann, N., Grohmann, M., Voigt, J. P., Bert, B., Vowinckel, J., Bader, M., et al. (2009). Intracellular serotonin modulates insulin secretion from pancreatic beta-cells by protein serotonylation. PLoS Biol. 7:e1000229. doi: 10.1371/journal.pbio.1000229
Penumatsa, K. C., and Fanburg, B. L. (2014). Transglutaminase 2-mediated serotonylation in pulmonary hypertension. Am. J. Physiol. Lung Cell. Mol. Physiol. 306, L309–L315. doi: 10.1152/ajplung.00321.2013
Penumatsa, K. C., Toksoz, D., Warburton, R. R., Hilmer, A. J., Liu, T., Khosla, C., et al. (2014). Role of hypoxia-induced transglutaminase 2 in pulmonary artery smooth muscle cell proliferation. Am. J. Physiol. Lung Cell. Mol. Physiol. 307, L576–L585.
Penumatsa, K. C., Toksoz, D., Warburton, R. R., Kharnaf, M., Preston, I. R., Kapur, N. K., et al. (2017). Transglutaminase 2 in pulmonary and cardiac tissue remodeling in experimental pulmonary hypertension. Am. J. Physiol. Lung Cell. Mol. Physiol. 313, L752–L762.
Piccini, J. P., Klein, L., Gheorghiade, M., and Bonow, R. O. (2004). New insights into diastolic heart failure: role of diabetes mellitus. Am. J. Med. 116(Suppl. 5A), 64S–75S.
Reed, A. L., Tanaka, A., Sorescu, D., Liu, H., Jeong, E. M., Sturdy, M., et al. (2011). Diastolic dysfunction is associated with cardiac fibrosis in the senescence-accelerated mouse. Am. J. Physiol. Heart Circ. Physiol. 301, H824–H831.
Schafer, M. J., White, T. A., Iijima, K., Haak, A. J., Ligresti, G., Atkinson, E. J., et al. (2017). Cellular senescence mediates fibrotic pulmonary disease. Nat. Commun. 8:14532.
Shinde, A. V., Su, Y., Palanski, B. A., Fujikura, K., Garcia, M. J., and Frangogiannis, N. G. (2018). Pharmacologic inhibition of the enzymatic effects of tissue transglutaminase reduces cardiac fibrosis and attenuates cardiomyocyte hypertrophy following pressure overload. J. Mol. Cell. Cardiol. 117, 36–48. doi: 10.1016/j.yjmcc.2018.02.016
Sicard, D., Haak, A. J., Choi, K. M., Craig, A. R., Fredenburgh, L. E., and Tschumperlin, D. J. (2018). Aging and anatomical variations in lung tissue stiffness. Am. J. Physiol. Lung Cell. Mol. Physiol. 314, L946–L955.
Skill, N. J., Johnson, T. S., Coutts, I. G., Saint, R. E., Fisher, M., Huang, L., et al. (2004). Inhibition of transglutaminase activity reduces extracellular matrix accumulation induced by high glucose levels in proximal tubular epithelial cells. J. Biol. Chem. 279, 47754–47762. doi: 10.1074/jbc.m402698200
Small, K., Feng, J. F., Lorenz, J., Donnelly, E. T., Yu, A., Im, M. J., et al. (1999). Cardiac specific overexpression of transglutaminase II (G(h)) results in a unique hypertrophy phenotype independent of phospholipase C activation. J. Biol. Chem. 274, 21291–21296. doi: 10.1074/jbc.274.30.21291
Steppan, J., Bergman, Y., Viegas, K., Armstrong, D., Tan, S., Wang, H., et al. (2017). Tissue transglutaminase modulates vascular stiffness and function through crosslinking-dependent and crosslinking-independent functions. J. Am. Heart Assoc. 6:e004161.
Steppan, J., Sikka, G., Jandu, S., Barodka, V., Halushka, M. K., Flavahan, N. A., et al. (2014). Exercise, vascular stiffness, and tissue transglutaminase. J. Am. Heart Assoc. 3:e000599.
Strait, J. B., and Lakatta, E. G. (2012). Aging-associated cardiovascular changes and their relationship to heart failure. Heart Fail. Clin. 8, 143–164. doi: 10.1016/j.hfc.2011.08.011
van Dijk, C. G., Oosterhuis, N. R., Xu, Y. J., Brandt, M., Paulus, W. J., van Heerebeek, L., et al. (2016). Distinct endothelial cell responses in the heart and kidney microvasculature characterize the progression of heart failure with preserved ejection fraction in the obese ZSF1 rat with cardiorenal metabolic syndrome. Circ. Heart Fail. 9:e002760.
Wang, Z., Stuckey, D. J., Murdoch, C. E., Camelliti, P., Lip, G. Y. H., and Griffin, M. (2018). Cardiac fibrosis can be attenuated by blocking the activity of transglutaminase 2 using a selective small-molecule inhibitor. Cell Death Dis. 9:613.
Whitaker, M. E., Nair, V., Sinari, S., Dherange, P. A., Natarajan, B., Trutter, L., et al. (2018). Diabetes mellitus associates with increased right ventricular afterload and remodeling in pulmonary arterial hypertension. Am. J. Med. 131, 702.e7–702.e13.
Wong, W. F., Gold, S., Fukuyama, O., and Blanchette, P. L. (1989). Diastolic dysfunction in elderly patients with congestive heart failure. Am. J. Cardiol. 63, 1526–1528. doi: 10.1016/0002-9149(89)90022-2
Keywords: transglutaminase 2, heart, lung, obesity, metabolic syndrome, aging, tissue stiffness, diastolic dysfunction
Citation: Penumatsa KC, Falcão-Pires I, Leite S, Leite-Moreira A, Bhedi CD, Nasirova S, Ma J, Sutliff RL and Fanburg BL (2020) Increased Transglutaminase 2 Expression and Activity in Rodent Models of Obesity/Metabolic Syndrome and Aging. Front. Physiol. 11:560019. doi: 10.3389/fphys.2020.560019
Received: 11 May 2020; Accepted: 17 August 2020;
Published: 15 September 2020.
Edited by:
James Todd Pearson, National Cerebral and Cardiovascular Center, JapanReviewed by:
Stephanie W. Watts, Michigan State University, United StatesRiccardo Ientile, University of Messina, Italy
Copyright © 2020 Penumatsa, Falcão-Pires, Leite, Leite-Moreira, Bhedi, Nasirova, Ma, Sutliff and Fanburg. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Krishna C. Penumatsa, a3BlbnVtYXRzYUB0dWZ0c21lZGljYWxjZW50ZXIub3Jn; Barry L. Fanburg, YmZhbmJ1cmdAdHVmdHNtZWRpY2FsY2VudGVyLm9yZw==