
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
PERSPECTIVE article
Front. Physiol. , 22 July 2020
Sec. Environmental, Aviation and Space Physiology
Volume 11 - 2020 | https://doi.org/10.3389/fphys.2020.00886
This article is part of the Research Topic Time Domains of Hypoxia Adaptation: Evolutionary Insights and Applications View all 24 articles
 Michael S. Tift1*
Michael S. Tift1* Rodrigo W. Alves de Souza2
Rodrigo W. Alves de Souza2 Janick Weber2
Janick Weber2 Erica C. Heinrich3
Erica C. Heinrich3 Francisco C. Villafuerte4
Francisco C. Villafuerte4 Atul Malhotra5
Atul Malhotra5 Leo E. Otterbein2
Leo E. Otterbein2 Tatum S. Simonson5
Tatum S. Simonson5Heme oxygenase (HO) enzymes catalyze heme into biliverdin, releasing carbon monoxide (CO) and iron into circulation. These byproducts of heme degradation can have potent cytoprotective effects in the face of stressors such as hypoxia and ischemia-reperfusion events. The potential for exogenous use of CO as a therapeutic agent has received increasing attention throughout the past few decades. Further, HO and CO are noted as putatively adaptive in diving mammals and certain high-altitude human populations that are frequently exposed to hypoxia and/or ischemia-reperfusion events, suggesting that HO and endogenous CO afford an evolutionary advantage for hypoxia tolerance and are critical in cell survival and injury avoidance. Our goal is to describe the importance of examining HO and CO in several systems, the physiological links, and the genetic factors that underlie variation in the HO/CO pathway. Finally, we emphasize the ways in which evolutionary perspectives may enhance our understanding of the HO/CO pathway in the context of diverse clinical settings.
In the late 1990s, a concept emerged suggesting that all cells possessed protective genes with the sole purpose to ensure survival (Platt and Nath, 1998). These cytoprotective genes played a role in the production of antioxidants, acute phase proteins, as well as regulating the effects of aging. A few were clearly master regulators, such that when not present, the cell or tissue was sensitive to stress. One of these was Hmox, which encodes the protein heme oxygenase (HO), the rate limiting enzyme responsible for the degradation of heme. HO catabolizes heme into biliverdin, which is subsequently converted to bilirubin by biliverdin reductase, releasing carbon monoxide (CO) and iron as byproducts. Paradoxically, CO was strictly considered a toxic pollutant. This notion stems from CO having a high affinity for the O2-binding site on hemoproteins. Therefore, elevated levels of CO can reduce O2 storage capacity and inhibit O2 transport, resulting in CO toxicity. However, recent studies have revealed the biological effects of the gas, leading to the acceptance of CO having therapeutic and cytoprotective effects in modulating inflammation, cell death, and proliferation (Motterlini and Otterbein, 2010).
Heme oxygenase genes are highly conserved and found in most living organisms (Wilks, 2002; Li and Stocker, 2009). The inducible HO-1 and constitutive HO-2 are ~40% identical in their amino acid sequence in humans (Cruse and Maines, 1988), with HO-1 observed at highest levels in the spleen, lung, and visceral adipose and HO-2 in the testes, heart, brain, and stomach (GTEx, V6). Stressors known to stimulate HO-1 expression include oxidative stress, hypoxia, cytokines, heavy metals, and bacterial endotoxins (Abraham, 2002). HO enzymes function as a crucial mechanism to recycle iron in the body through the degradation of heme (Wilks, 2002). The majority of extant vertebrates are known to utilize iron-containing hemoproteins (e.g., hemoglobin, myoglobin, neuroglobin, cytochromes, catalase, and peroxidase) as a mechanism to transport electrons in the respiratory chain for the production of ATP, to reduce oxidative stress and/or to increase the storage capacity and delivery of oxygen (O2) to cells (Hardison, 1996). Therefore, the ability to recycle iron stores in the body can offer an adaptive advantage if organisms find themselves iron-limited. The regulation of HO enzymes can also influence erythrocyte lifespan. Hmox1−/− mice rarely survive beyond the development and those that do have prolonged erythrocyte lifespans, reduced hemoglobin content, and a reduced erythrocyte size that leads to microcytic anemia (Fraser et al., 2015). Moreover, human patients with reduced erythrocyte lifespans display elevated blood and breath CO levels, suggesting increased HO activity (Strocchi et al., 1992).
Recent studies have shown that species and populations adapted to tolerate chronic hypoxia and/or ischemia/reperfusion (I/R) events have upregulated or modified the HO/CO pathway in a manner that could afford beneficial side effects (Herrera et al., 2008; Simonson et al., 2010; Tift et al., 2014). Here, we highlight some of those model species and populations and discuss how such modifications could provide an evolutionary advantage and how information from these studies may be used to learn more about the applications of the HO/CO pathway in the prevention and treatment of certain diseases (Goebel and Wollborn, 2020).
The late August Krogh stated that, “for such a large number of problems there will be some animal of choice, or a few such animals, on which it can be most conveniently studied” (Krogh, 1929). Model organisms naturally adapted for life in challenging conditions offer excellent opportunities to understand how extreme phenotypes afford protective advantages that can be directed to improve the treatment and/or the prevention of specific human pathologies (Carey et al., 2012; Carey, 2015). For example, some diving mammals experience dramatic hypoxemia (Meir et al., 2009) and I/R events (Zapol et al., 1979) during their repeated long-duration breath-holds and do not develop injuries from this lifestyle. Species that have adapted to live at high altitude or under other hypoxic conditions (e.g., burrowing) also avoid pathologies associated with exposure to chronic hypoxia (Logan et al., 2020). These natural models offer unique insights into the adaptive mechanisms underlying tolerance to hypoxia and/or I/R injury.
Some deep-diving mammals are known to experience levels of hypoxemia during dives that resemble arterial oxygen saturations of humans breathing ambient air on the summit of Mount Everest (Meir et al., 2009). These deep-divers also appear to be protected from repeated I/R events that occur in a majority of their tissues on a dive-to-dive basis as a result of the “dive response,” characterized by extreme bradycardia and peripheral ischemia to maintain mean arterial blood pressure during breath-holds (Halasz et al., 1974; Zapol et al., 1979; Allen and Vázquez-Medina, 2019). In 1959, Pugh reported levels of CO in the blood of deep-diving Weddell seals from Antarctica that resembled those of cigarette smokers (Pugh, 1959). Since then, Tift et al. (2014) have discovered that deep-diving northern elephant seals also maintain high levels of CO in the blood with carboxyhemoglobin (COHb) levels between 5 and 11% (Tift et al., 2014). While CO was shown to be high in the blood of two deep-diving pinnipeds, the distribution of CO concentrations in the blood of different diving species is still unknown.
The quantity of CO found in the blood of elephant seals is similar to that previously mentioned in laboratory studies, which demonstrate cytoprotective effects from exogenous CO delivery (Nakao et al., 2006). It is hypothesized that the elevated hemoprotein stores seen in deep-diving mammals are the source of the high CO levels, which could be used as a mechanism to reduce the development of tissue injuries from the chronic hypoxemia and I/R events they experience while diving (Tift and Ponganis, 2019). These high CO levels could indicate alterations in the erythrocyte lifespan and HO activity in specific tissues of these animals. Such levels of CO would likely increase the O2-binding affinity of hemoglobin, impacting O2-delivery mechanics. Considering the low arterial pO2 values routinely experienced during breath-holds (<15–20 mmHg), it is possible that a higher hemoglobin-O2 affinity due to moderate COHb levels could preserve blood O2 stores, increasing O2 availability later in the dive (Storz, 2016). Indeed, the Haldane effect suggests that mice exposed to trace amounts of CO will more efficiently acquire O2 and survive longer under conditions of severe hypoxia (Roughton and Darling, 1944). Endogenous CO also impacts mitochondrial O2 consumption and ATP production, which could contribute to hypometabolism during breath-holds and exposure to hypoxia (D’Amico et al., 2006).
Species adapted to live at high altitude face a different hypoxic challenge, in that the ambient environment has a reduced pO2. Certain pathologies may develop due to acute and chronic hypoxia exposure at altitude (e.g., pulmonary and cerebral edema, pulmonary hypertension, and excessive erythrocytosis). Considering hypoxia as a stressor that increases HO-1 activity (Shibahara et al., 2007), the HO/CO pathway is of interest when studying organisms at high altitude. When laboratory rats were brought from an elevation of 1,006 to 3,048 and 4,572 m, their COHb levels increased linearly with altitude from 0.68 to 1.16, and 1.68%, respectively (McGrath, 1992). Similarly, when healthy adult humans were brought from an altitude near sea level to 3,517 m, their COHb levels increased from 0.79 to 0.95% within 20 h (McGrath et al., 1993). When comparing neonatal sheep (lowland species) and llama (highland species) that underwent gestation at high altitude (3,600 m), only sheep developed pulmonary arterial hypertension (Herrera et al., 2008; Llanos et al., 2012). This was associated with reduced soluble guanylate cyclase, HO, and CO production despite increased nitric oxide synthase activity in the lungs of the sheep. In contrast, neonatal llamas avoided pulmonary hypertension and had an increase in pulmonary CO production and HO-1 expression with no change in nitric oxide levels (Herrera et al., 2008). This specific HO/CO protective effect has not been investigated in other species adapted for living with chronic hypoxia.
Humans of Tibetan ancestry exhibit a lower average hemoglobin concentration compared to sojourners and long-term highland residents in the Andes, where individuals exhibit varying degrees of polycythemia (Beall, 2007). Whether relatively lower hemoglobin provides a direct adaptive effect or is the side effect of other traits that provide an advantage remains an important question in the field (Storz, 2010; Simonson 2015). HMOX2 has been detected as a top selection candidate gene in ours and others’ studies of Tibetan adaptation and is hypothesized to play a role in regulating heme catabolism and CO production (Simonson et al., 2010; Yang et al., 2016). A study of 1,250 high-altitude native Tibetans found variants at this locus that are associated with decreased hemoglobin concentration in males, and in vitro analysis indicates that a derived intronic variant (rs4786504) is associated with increased HMOX2 expression (Yang et al., 2016). While it remains to be determined if regulatory variants at this locus lead to increased heme catabolism, such alterations could contribute to the lower hemoglobin concentrations seen in Tibetans living at altitude.
In contrast to Tibetans, adaptive HMOX genetic variants have not yet been identified in Andean or Ethiopian highlanders. However, HO could mitigate various challenges imposed by hypoxia at altitude (e.g., higher Hb-O2 affinity; Storz, 2016). Our recent examination in Andean adult males and females living at 4,350 m shows a positive relationship between endogenous CO in breath and blood with both hemoglobin concentration and altitude (Figure 1). These preliminary findings could reflect elevated erythrocyte destruction, reduced erythrocyte lifespan, and/or increased HO-1 or HO-2 activity. Genetic studies in lowlanders have shown that individuals with increased HO-1 expression are less prone to pathologies, such as diabetes, atherosclerosis, chronic obstructive pulmonary disease (COPD), and arthritis (Motterlini and Otterbein, 2010). Whether these levels contribute to the variability in pathologies associated with excessive erythrocytosis and chronic mountain sickness in Andeans remain to be determined.
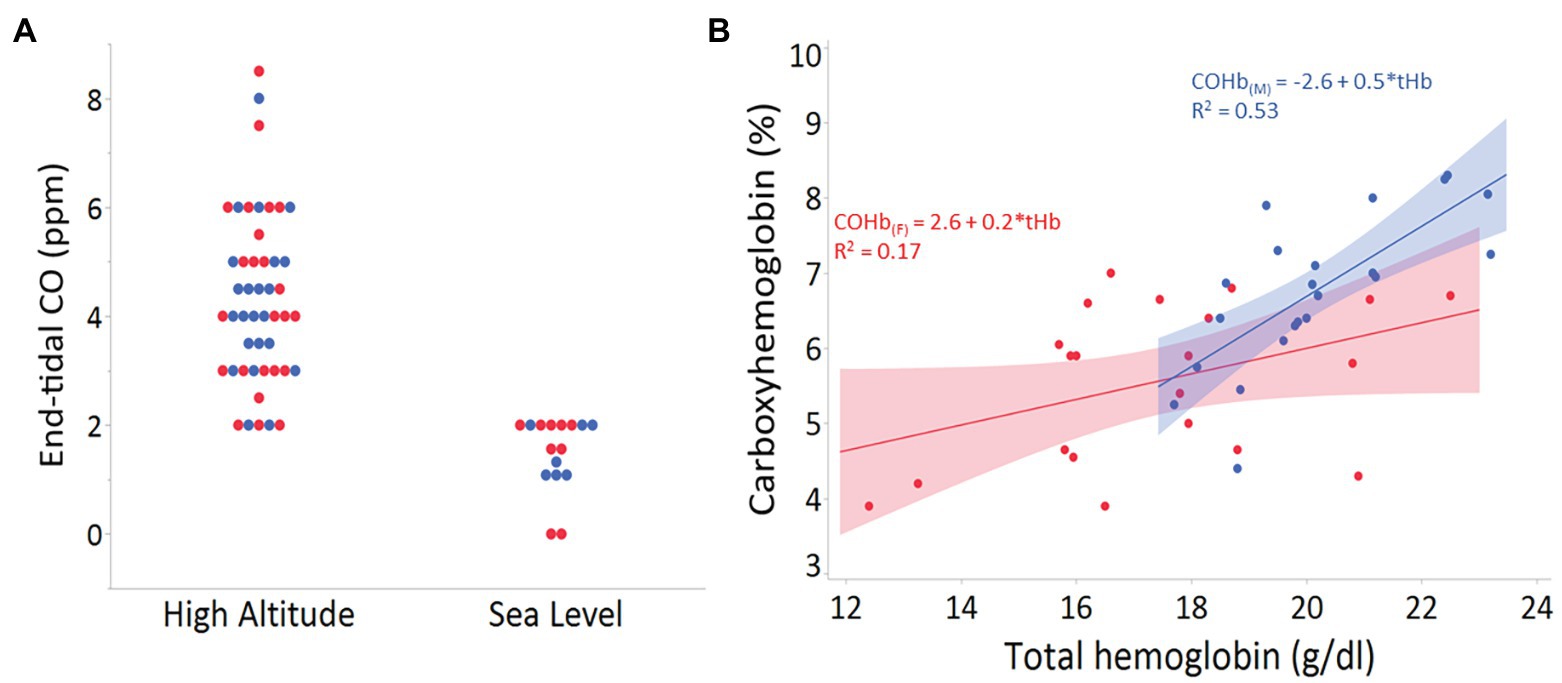
Figure 1. (A) End-tidal carbon monoxide (CO) levels (ppm) in 47 adults from Cerro de Pasco, Peru (high altitude), and 18 adults of mixed ancestry living in San Diego, California, USA (sea level). There was no statistical difference between males (blue) and females (red) from high or low altitude groups (p > 0.8 for both). (B) Relationship between total hemoglobin (g/dl) and the percent carboxyhemoglobin (COHb) in venous blood of adult males (n = 22; blue) and females (n = 21; red) from Cerro de Pasco, Peru (4,330 m).
Hypoxemia and I/R events are complications inherent to common disease states that often lead to a suite of downstream problems, including adverse cardiopulmonary effects, inflammation, and tissue death. Induction of a battery of stress response genes, such as HMOX1, respond metabolically to (1) degrade elevated heme released during tissue damage in part due to an increase in intracellular hemoproteins and (2) to generate bioactive products to further enhance cell and tissue recovery (Figure 2). Hypoxia and inflammation often occur in tandem during infection or I/R events, whereby the hypoxia-inducible factor (HIF) pathway interacts with NF-κB signaling to coordinate molecular responses, including regulation of HMOX1 (Lee et al., 1997; Rushworth and O’Connell, 2004).
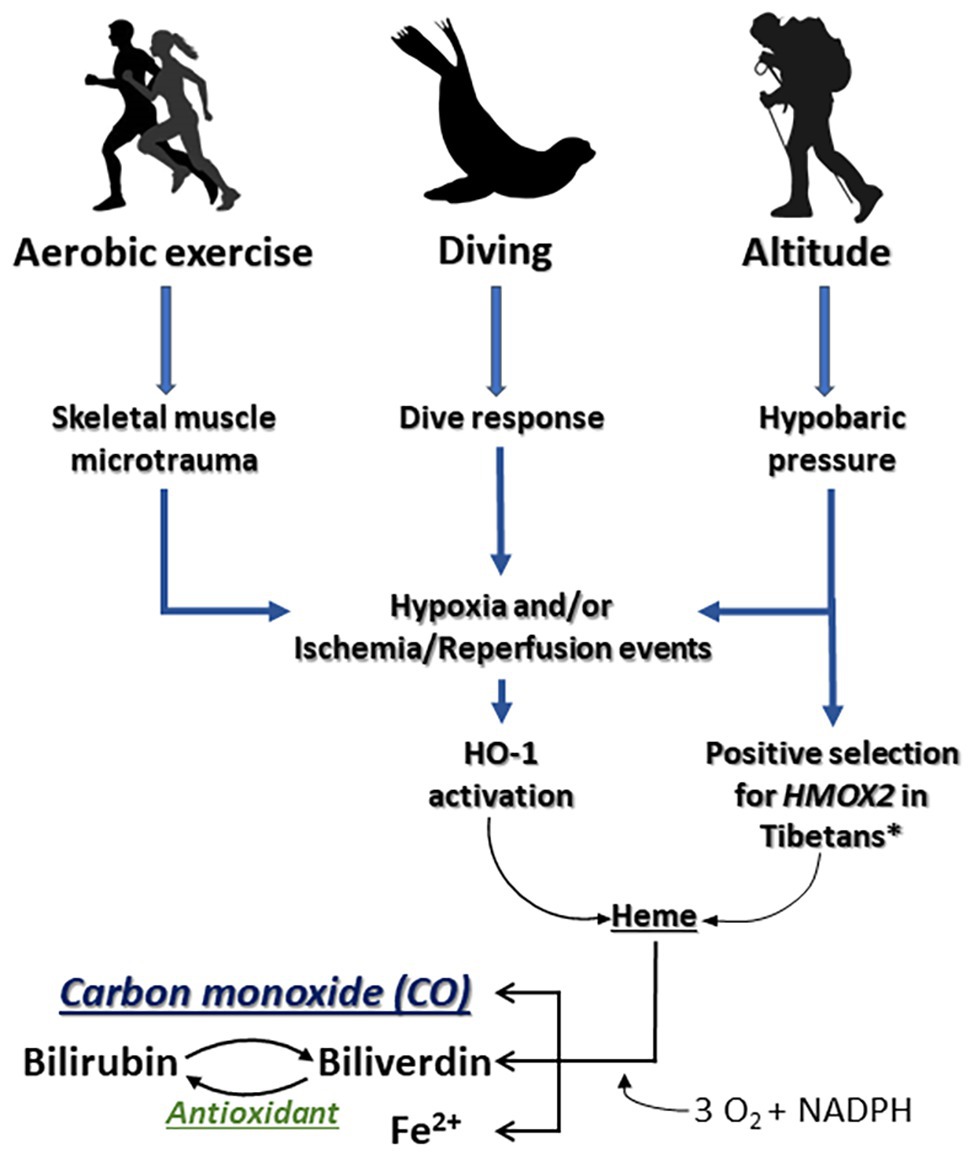
Figure 2. Impacts of aerobic exercise, breath-hold diving, and exposure to altitude on the HO/CO pathway. The stress response protein heme oxygenase-1 (HO-1) responds to intense aerobic exercise and/or skeletal muscle microtrauma by (1) degrading heme that is released during tissue injury as a danger associated molecular pattern and (2) generating bioactive products that contribute to cell and tissue recovery. *Simonson et al., 2010.
Why would evolution result in a system that increases CO in the height of hypoxia or an inflammatory sequelae and ongoing stress response that otherwise decreases tissue O2 availability? Perhaps permissive hypoxia, driven by several selective O2 sensors such as HIF-1α, HIF-2α, and/or mitochondrial oxidases, serves as a mechanism by which cells can dynamically adjust to optimize survival in different environments. In each instance, the availability of O2, which we define as that which is permissive or allowed, dictates specific cellular responses that benefit the needs of the tissue. While O2 is a requisite cofactor for HO-1 activity, HO-1 is also potently induced by hypoxia; in a somewhat paradoxical manner, this results in the generation of CO which then competes with O2 for heme binding sites (Figure 2). This observation has led to speculation that CO creates permissive hypoxia in tissues that serve to modulate cellular energetics and protection (D’Amico et al., 2006).
The effects of HO-1 and CO on tissue protection have been clearly demonstrated in models of I/R injury and many other tissue ischemic pathologies (Wegiel et al., 2014). Due to space constraints, we highlight a few of the seminal findings. Induction of HO-1 or exposure to CO offer dose-dependent anti-inflammatory and cytoprotective effects (Otterbein et al., 2016). The cytoprotective effects of CO were first demonstrated in acute lung injury, and these findings rapidly expanded through the work of numerous laboratories to most models of acute organ injury (Hopper et al., 2018). Administration of exogenous CO or higher HO-1 activity increases the expression of anti-inflammatory cytokines and reduces expression of multiple pro-inflammatory cytokines during acute pulmonary inflammation (Konrad et al., 2016; Zhang et al., 2019). In particular, CO inhibited LPS-induced pro-inflammatory signaling by downregulating NADPH oxidase-dependent reactive oxygen species (ROS) production in macrophages, thereby inhibiting toll-like receptor (TLR) signaling (Nakahira et al., 2006). Additionally, CO can inhibit tumor necrosis factor-induced apoptosis via a p38 MAPK-dependent mechanism (Ryter et al., 2007). These beneficial effects may also result in part from CO stabilization of HIF-1α (Chin et al., 2007). This is important considering the impact of pulmonary function in hypoxia-induced diseases at high altitude (Selland et al., 1993) and the consistent hypoxemia and lung collapse seen during deep-dives in marine mammals (McDonald and Ponganis, 2012).
CO is also hypothesized to play a role in the control of breathing via peripheral O2 sensing in the carotid body, where HO-2 is expressed (Prabhakar, 2012). In the presence of hypoxia, decreased HO-2 activity in glomus cells may reduce CO generation and, since CO inhibits cystathionine-γ-lyase production of the excitatory gasotransmitter H2S, decreased CO generation may result in increased sensory activity of the glomus cells. This is intriguing given that the gene HMOX2 is under evolutionary selection in Tibetan populations, who maintain elevated hypoxic ventilatory responses compared to other high-altitude groups (Beall, 2007).
Organ transplant has been perhaps the most well studied when considering translational potential of the HO/CO pathway. The role of HO-1/CO in the kidney was first described in models of I/R injury, where induction of HO-1 or exposure to CO protected the kidney (Schaaf-Lafontaine and Courtoy, 1986; Blydt-Hansen et al., 2003). This large body of work culminated in the initiation of multiple clinical trials, where exogenous CO has safely been administered to transplant patients1. The mechanism by which CO imparts its salutary effects remain incompletely understood, but likely targets include hemoproteins such as mitochondria oxidases and therein the regulation of bioenergetics, ROS formation, and consumption of O2 (Schallner and Otterbein, 2015; Otterbein et al., 2016). The beneficial effects of HO-1 and CO in organ injury result from preserving mitochondrial health and would dovetail with the observations observed in diving mammals and populations that live at altitude. It is intriguing to speculate that one mechanism for the cytoprotective effects of CO, in instances of tissue hypoxia, may be that CO permits displacement and redistribution of O2 within intracellular stores. The activity of HO-1 is known to increase during hypoxia, increasing CO production, which can alter cellular metabolism; thus, another potential mechanism of CO that plays a role in hypoxia tolerance is slowing or shifting oxidative metabolism to glycolysis through permissive use of O2 (Figure 2).
The HO/CO pathway has also been the focus of many reports in models of cardiovascular disease, including heart failure and cardiac arrest (Shih et al., 2011). Cardiac muscle cells deficient in HO-1 accumulate lethal amounts of ROS, and mice that survive with embryonic HO-1 deletion exhibit many deleterious effects (Kapturczak et al., 2004). Furthermore, Hmox1−/− mice are highly susceptible to I/R injury and, after hypoxia, these animals show evidence of right ventricular infarction (Yoshida et al., 2001; Liu et al., 2005). During reperfusion in the heart, CO administration can decrease infarct size, reduce apoptosis, and increase inotropy. Although those effects were examined mostly in a cardiovascular system, HO-1 also affects other cell types, including skeletal muscle and the physiologic response to exercise (Kozakowska et al., 2018).
Skeletal muscle comprises almost 40% of total body mass in humans, exhibiting major metabolic activity by contributing up to 30% of the resting metabolic rate in adults (Zurlo et al., 1990). The tissue can respond to numerous environmental and physiological challenges (e.g., hypoxia and I/R events) by changing its phenotypic profile and is one commonality across diving animals and individuals living at altitude (Pette and Staron, 2001; Spangenburg et al., 2008; Pabst et al., 2016). Skeletal muscle contraction during exhaustive exercise generates ROS that can promote oxidative damage to myofibers (Reid, 2001), and it has been suggested that HO-1 can protect against exercise-induced injury (Saxena et al., 2010). HO-1 is normally expressed at very low levels in skeletal muscle, but increases dramatically with exhaustive exercise (Pilegaard et al., 2000). The discovery that CO induces mitochondrial biogenesis through specific signaling pathways has raised the possibility that it contributes to the resolution of skeletal muscle injury during and after episodes of oxidative stress, such as exercise or I/R events (Suliman et al., 2007). Intermittent CO breathing after a single exercise test led to an increase in mitochondrial oxidative stress markers and mitochondrial fusion protein expression indicative of mitochondrial biogenesis in skeletal muscle.
From a clinical perspective, based on high disease prevalence and consequences, there is intense interest in both intermittent hypoxemia (as seen in obstructive sleep apnea-OSA) and sustained hypoxemia (as seen in COPD or at high altitude; Soler et al., 2015; Benjafield et al., 2019). The combination of these stimuli (sustained plus intermittent hypoxemia) occurs in some clinical settings, including overlap syndrome (OSA plus COPD), or in OSA patients at high altitude (Marin et al., 2010). The deleterious effects of hypoxemia are well established (Drager et al., 2015; Umeda et al., 2020), although some literature supports a potential protective role of ischemic preconditioning with more mild levels of hypoxemia (Sánchez-de-la-Torre et al., 2018). Regarding CO, some data support its role as a biomarker in OSA (Kobayashi et al., 2008), although its biological impact in these patients has been debated (Owens et al., 2008). Further translational research is certainly encouraged to apply knowledge regarding hypoxia and CO from comparative biology to the clinical setting.
The concept that evolutionary mechanisms have optimized the biology and physiology of HO and CO is relevant in the context of hypoxia adaptation. Examples can be observed across species and under varying physiologic and pathophysiologic conditions ranging from diving mammals to individuals living at altitude, as well as physical exercise and in response to tissue injury. Increasing our understanding of the natural activity and regulation of the HO/CO pathway associated with injury avoidance and maintenance of cellular health will contribute valuable knowledge regarding the benefits of tissue-specific, moderate-level CO exposure.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The studies involving human participants were reviewed and approved by UCSD IRB #171772. The patients/participants provided their written informed consent to participate in this study.
All authors contributed to the article and approved the submitted version.
This work was supported by grants from the National Science Foundation (1927616), Department of Defense (W81XWH-16-0464), National Institutes of Health (R01DK119202, R01HL145470), the UCSD Center for Physiological Genomics of Low Oxygen (CPGLO), and American Heart Association (#19CDA34760244).
LO is a Scientific Advisor for Hillhurst Biopharmaceuticals. AM is funded by NHLBI. He reports income from Merck and Livanova related to medical education. ResMed provided a philanthropic donation to UC San Diego.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Abraham, N. G. (ed.) (2002). Heme oxygenase in biology and medicine. (New York: Springer Science & Business Media). Available at: https://www.springer.com/gp/book/9780306472640#aboutBook
Allen, K. N., and Vázquez-Medina, J. P. (2019). Natural tolerance to ischemia and hypoxemia in diving mammals: a review. Front. Physiol. 10:1199. doi: 10.3389/fphys.2019.01199
Beall, C. M. (2007). Two routes to functional adaptation: tibetan and Andean high-altitude natives. Proc. Natl. Acad. Sci. U. S. A. 104, 8655–8660. doi: 10.1073/pnas.0701985104
Benjafield, A. V., Ayas, N. T., Eastwood, P. R., Heinzer, R., Ip, M. S. M., Morrell, M. J., et al. (2019). Estimation of the global prevalence and burden of obstructive sleep apnoea: a literature-based analysis. Lancet Respir. Med. 7, 687–698. doi: 10.1016/S2213-2600(19)30198-5
Blydt-Hansen, T. D., Katori, M., Lassman, C., Ke, B., Coito, A. J., Iyer, S., et al. (2003). Gene transfer-induced local heme oxygenase-1 overexpression protects rat kidney transplants from ischemia/reperfusion injury. J. Am. Soc. Nephrol. 14, 745–754. doi: 10.1097/01.asn.0000050760.87113.25
Carey, H. V. (2015). Lessons learned from comparative and evolutionary physiology. Physiology 30, 80–81. doi: 10.1152/physiol.00003.2015
Carey, H. V., Martin, S. L., Horwitz, B. A., Yan, L., Bailey, S. M., Podrabsky, J., et al. (2012). Elucidating nature’s solutions to heart, lung, and blood diseases and sleep disorders. Circ. Res. 110, 915–921. doi: 10.1161/CIRCRESAHA.111.255398
Chin, B. Y., Jiang, G., Wegiel, B., Wang, H. J., MacDonald, T., Zhang, X. C., et al. (2007). Hypoxia-inducible factor 1α stabilization by carbon monoxide results in cytoprotective preconditioning. Proc. Natl. Acad. Sci. U. S. A. 104, 5109–5114. doi: 10.1073/pnas.0609611104
Cruse, I., and Maines, M. D. (1988). Evidence suggesting that the two forms of heme oxygenase are products of different genes. J. Biol. Chem. 263, 3348–3353.
D’Amico, G., Lam, F., Hagen, T., and Moncada, S. (2006). Inhibition of cellular respiration by endogenously produced carbon monoxide. J. Cell Sci. 119, 2291–2298. doi: 10.1242/jcs.02914
Drager, L. F., Polotsky, V. Y., O’Donnell, C. P., Cravo, S. L., Lorenzi-Filho, G., and Machado, B. H. (2015). Translational approaches to understanding metabolic dysfunction and cardiovascular consequences of obstructive sleep apnea. Am. J. Physiol. Heart Circ. Physiol. 309, H1101–H1111. doi: 10.1152/ajpheart.00094.2015
Fraser, S. T., Midwinter, R. G., Coupland, L. A., Kong, S., Berger, B. S., Yeo, J. H., et al. (2015). Heme oxygenase-1 deficiency alters erythroblastic island formation, steady-state erythropoiesis and red blood cell lifespan in mice. Haematologica 100, 601–610. doi: 10.3324/haematol.2014.116368
Goebel, U., and Wollborn, J. (2020). Carbon monoxide in intensive care medicine-time to start the therapeutic application?! Intensive Care Med. Exp. 8:2. doi: 10.1186/s40635-020-0292-8
Halasz, N. A., Elsner, R., Garvie, R. S., and Grotke, G. T. (1974). Renal recovery from ischemia: a comparative study of harbor seal and dog kidneys. Am. J. Phys. 227, 1331–1335. doi: 10.1152/ajplegacy.1974.227.6.1331
Hardison, R. C. (1996). A brief history of hemoglobins: plant, animal, protist, and bacteria. Proc. Natl. Acad. Sci. U. S. A. 93, 5675–5679. doi: 10.1073/pnas.93.12.5675
Herrera, E. A., Reyes, R. V., Giussani, D. A., Riquelme, R. A., Sanhueza, E. M., Ebensperger, G., et al. (2008). Carbon monoxide: a novel pulmonary artery vasodilator in neonatal llamas of the Andean altiplano. Cardiovasc. Res. 77, 197–201. doi: 10.1093/cvr/cvm013
Hopper, C. P., Meinel, L., Steiger, C., and Otterbein, L. E. (2018). Where is the clinical breakthrough of heme oxygenase-1/carbon monoxide therapeutics? Curr. Pharm. Des. 24, 2264–2282. doi: 10.2174/1381612824666180723161811
Kapturczak, M. H., Wasserfall, C., Brusko, T., Campbell-Thompson, M., Ellis, T. M., Atkinson, M. A., et al. (2004). Heme oxygenase-1 modulates early inflammatory responses: evidence from the heme oxygenase-1-deficient mouse. Am. J. Pathol. 165, 1045–1053. doi: 10.1016/S0002-9440(10)63365-2
Kobayashi, M., Miyazawa, N., Takeno, M., Murakami, S., Kirino, Y., Okouchi, A., et al. (2008). Circulating carbon monoxide level is elevated after sleep in patients with obstructive sleep apnea. Chest 134, 904–910. doi: 10.1378/chest.07-2904
Konrad, F. M., Knausberg, U., Höne, R., Ngamsri, K. -C., and Reutershan, J. (2016). Tissue heme oxygenase-1 exerts anti-inflammatory effects on LPS-induced pulmonary inflammation. Mucosal Immunol. 9, 98–111. doi: 10.1038/mi.2015.39
Kozakowska, M., Pietraszek-Gremplewicz, K., Ciesla, M., Seczynska, M., Bronisz-Budzynska, I., Podkalicka, P., et al. (2018). Lack of heme oxygenase-1 induces inflammatory reaction and proliferation of muscle satellite cells after cardiotoxin-induced skeletal muscle injury. Am. J. Pathol. 188, 491–506. doi: 10.1016/j.ajpath.2017.10.017
Lee, P. J., Jiang, B. H., Chin, B. Y., Iyer, N. V., Alam, J., Semenza, G. L., et al. (1997). Hypoxia-inducible factor-1 mediates transcriptional activation of the heme oxygenase-1 gene in response to hypoxia. J. Biol. Chem. 272, 5375–5381. doi: 10.1074/jbc.272.9.5375
Li, C., and Stocker, R. (2009). Heme oxygenase and iron: from bacteria to humans. Redox Rep. 14, 95–101. doi: 10.1179/135100009x392584
Liu, X., Wei, J., Peng, D. H., Layne, M. D., and Yet, S. -F. (2005). Absence of heme oxygenase-1 exacerbates myocardial ischemia/reperfusion injury in diabetic mice. Diabetes 54, 778–784. doi: 10.2337/diabetes.54.3.778
Llanos, A. J., Ebensperger, G., Herrera, E. A., Reyes, R. V., Cabello, G., Díaz, M., et al. (2012). The heme oxygenase-carbon monoxide system in the regulation of cardiorespiratory function at high altitude. Respir. Physiol. Neurobiol. 184, 186–191. doi: 10.1016/j.resp.2012.05.003
Logan, S. M., Szereszewski, K. E., Bennett, N. C., Hart, D. W., van Jaarsveld, B., Pamenter, M. E., et al. (2020). The brains of six African mole-rat species show divergent responses to hypoxia. J. Exp. Biol. 223:jeb215905. doi: 10.1242/jeb.215905
Marin, J. M., Soriano, J. B., Carrizo, S. J., Boldova, A., and Celli, B. R. (2010). Outcomes in patients with chronic obstructive pulmonary disease and obstructive sleep apnea: the overlap syndrome. Am. J. Respir. Crit. Care Med. 182, 325–331. doi: 10.1164/rccm.200912-1869OC
McDonald, B. I., and Ponganis, P. J. (2012). Lung collapse in the diving sea lion: hold the nitrogen and save the oxygen. Biol. Lett. 8, 1047–1049. doi: 10.1098/rsbl.2012.0743
McGrath, J. J. (1992). Effects of altitude on endogenous carboxyhemoglobin levels. J. Toxicol. Environ. Health 35, 127–133. doi: 10.1080/15287399209531601
McGrath, J. J., Schreck, R. M., and Lee, P. S. (1993). Carboxyhemoglobin levels in humans: effects of altitude. Inhal. Toxicol. 5, 241–249. doi: 10.3109/08958379309034504
Meir, J. U., Champagne, C. D., Costa, D. P., Williams, C. L., and Ponganis, P. J. (2009). Extreme hypoxemic tolerance and blood oxygen depletion in diving elephant seals. Am. J. Phys. Regul. Integr. Comp. Phys. 297, R927–R939. doi: 10.1152/ajpregu.00247.2009
Motterlini, R., and Otterbein, L. E. (2010). The therapeutic potential of carbon monoxide. Nat. Rev. Drug Discov. 9, 728–743. doi: 10.1038/nrd3228
Nakahira, K., Kim, H. P., Geng, X. H., Nakao, A., Wang, X., Murase, N., et al. (2006). Carbon monoxide differentially inhibits TLR signaling pathways by regulating ROS induced trafficking of TLRs to lipid rafts. J. Exp. Med. 203, 2377–2389. doi: 10.1084/jem.20060845
Nakao, A., Toyokawa, H., Tsung, A., Nalesnik, M. A., Stolz, D. B., Kohmoto, J., et al. (2006). Ex vivo application of carbon monoxide in University of Wisconsin solution to prevent intestinal cold ischemia/reperfusion injury. Am. J. Transplant. 6, 2243–2255. doi: 10.1111/j.1600-6143.2006.01465.x
Otterbein, L. E., Foresti, R., and Motterlini, R. (2016). Heme oxygenase-1 and carbon monoxide in the heart: the balancing act between danger signaling and pro-survival. Circ. Res. 118, 1940–1959. doi: 10.1161/CIRCRESAHA.116.306588
Owens, R. L., Yim-Yeh, S., and Malhotra, A. (2008). Carbon monoxide poisoning, or carbon monoxide protection? Chest 134, 895–896. doi: 10.1378/chest.08-1728
Pabst, D. A., McLellan, W. A., and Rommel, S. A. (2016). How to build a deep diver: the extreme morphology of mesoplodonts. Integr. Comp. Biol. 56, 1337–1348. doi: 10.1093/icb/icw126
Pette, D., and Staron, R. S. (2001). Transitions of muscle fiber phenotypic profiles. Histochem. Cell Biol. 115, 359–372. doi: 10.1007/s004180100268
Pilegaard, H., Ordway, G. A., Saltin, B., and Darrell Neufer, P. (2000). Transcriptional regulation of gene expression in human skeletal muscle during recovery from exercise. Am. J. Physiol. Endocrinol. Metab. 279, E806–E814. doi: 10.1152/ajpendo.2000.279.4.E806
Platt, J. L., and Nath, K. A. (1998). Heme oxygenase: protective gene or Trojan horse. Nat. Med. 4, 1364–1365. doi: 10.1038/3947
Prabhakar, N. R. (2012). Carbon monoxide (CO) and hydrogen sulfide (H(2)S) in hypoxic sensing by the carotid body. Respir. Physiol. Neurobiol. 184, 165–169. doi: 10.1016/j.resp.2012.05.022
Pugh, L. G. (1959). Carbon monoxide content of the blood and other observations on Weddell seals. Nature 183, 74–76. doi: 10.1038/183074a0
Reid, M. B. (2001). Nitric oxide, reactive oxygen species, and skeletal muscle contraction. Med. Sci. Sports Exerc. 33, 371–376. doi: 10.1097/00005768-200103000-00006
Roughton, F. J. W., and Darling, R. C. (1944). The effect of carbon monoxide on the oxyhemoglobin dissociation curve. Am. J. Phys. 141, 737–737. doi: 10.1152/ajplegacy.1944.141.5.737-s
Rushworth, S. A., and O’Connell, M. A. (2004). Haem oxygenase-1 in inflammation. Biochem. Soc. Trans. 32, 1093–1094. doi: 10.1042/BST0321093
Ryter, S. W., Kim, H. P., Nakahira, K., Zuckerbraun, B. S., Morse, D., and Choi, A. M. K. (2007). Protective functions of heme oxygenase-1 and carbon monoxide in the respiratory system. Antioxid. Redox Signal. 9, 2157–2173. doi: 10.1089/ars.2007.1811
Sánchez-de-la-Torre, A., Soler, X., Barbé, F., Florés, M., Maisel, A., Malhotra, A., et al. (2018). Cardiac troponin values in patients with acute coronary syndrome and sleep apnea: a pilot study. Chest 153, 329–338. doi: 10.1016/j.chest.2017.06.046
Saxena, S., Shukla, D., Saxena, S., Khan, Y. A., Singh, M., Bansal, A., et al. (2010). Hypoxia preconditioning by cobalt chloride enhances endurance performance and protects skeletal muscles from exercise-induced oxidative damage in rats. Acta Physiol. 200, 249–263. doi: 10.1111/j.1748-1716.2010.02136.x
Schaaf-Lafontaine, N., and Courtoy, R. (1986). Transfer into the mouse of mast cells differentiated in vitro. C. R. Seances Soc. Biol. Fil. 180, 234–242.
Schallner, N., and Otterbein, L. E. (2015). Friend or foe? Carbon monoxide and the mitochondria. Front. Physiol. 6:17. doi: 10.3389/fphys.2015.00017
Selland, M. A., Stelzner, T. J., Stevens, T., Mazzeo, R. S., McCullough, R. E., and Reeves, J. T. (1993). Pulmonary function and hypoxic ventilatory response in subjects susceptible to high-altitude pulmonary edema. Chest 103, 111–116. doi: 10.1378/chest.103.1.111
Shibahara, S., Han, F., Li, B., and Takeda, K. (2007). Hypoxia and heme oxygenases: oxygen sensing and regulation of expression. Antioxid. Redox Signal. 9, 2209–2225. doi: 10.1089/ars.2007.1784
Shih, R. -H., Cheng, S. -E., Hsiao, L. -D., Kou, Y. R., and Yang, C. -M. (2011). Cigarette smoke extract upregulates heme oxygenase-1 via PKC/NADPH oxidase/ROS/PDGFR/PI3K/Akt pathway in mouse brain endothelial cells. J. Neuroinflammation 8:104. doi: 10.1186/1742-2094-8-104
Simonson, T. S. (2015). Altitude adaptation: a glimpse through various lenses. High Alt. Med. Biol. 16, 125–137. doi: 10.1089/ham.2015.0033
Simonson, T. S., Yang, Y., Huff, C. D., Yun, H., Qin, G., Witherspoon, D. J., et al. (2010). Genetic evidence for high-altitude adaptation in Tibet. Science 329, 72–75. doi: 10.1126/science.1189406
Soler, X., Gaio, E., Powell, F. L., Ramsdell, J. W., Loredo, J. S., Malhotra, A., et al. (2015). High prevalence of obstructive sleep apnea in patients with moderate to severe chronic obstructive pulmonary disease. Ann. Am. Thorac. Soc. 12, 1219–1225. doi: 10.1513/AnnalsATS.201407-336OC
Spangenburg, E. E., Le Roith, D., Ward, C. W., and Bodine, S. C. (2008). A functional insulin-like growth factor receptor is not necessary for load-induced skeletal muscle hypertrophy. J. Physiol. 586, 283–291. doi: 10.1113/jphysiol.2007.141507
Storz, J. F. (2010). Evolution. Genes for high altitudes. Science 329, 40–41. doi: 10.1126/science.1192481
Storz, J. F. (2016). Hemoglobin-oxygen affinity in high-altitude vertebrates: is there evidence for an adaptive trend? J. Exp. Biol. 219, 3190–3203. doi: 10.1242/jeb.127134
Strocchi, A., Schwartz, S., Ellefson, M., Engel, R. R., Medina, A., and Levitt, M. D. (1992). A simple carbon monoxide breath test to estimate erythrocyte turnover. J. Lab. Clin. Med. 120, 392–399.
Suliman, H. B., Carraway, M. S., Ali, A. S., Reynolds, C. M., Welty-Wolf, K. E., and Piantadosi, C. A. (2007). The CO/HO system reverses inhibition of mitochondrial biogenesis and prevents murine doxorubicin cardiomyopathy. J. Clin. Invest. 117, 3730–3741. doi: 10.1172/JCI32967
Tift, M. S., and Ponganis, P. J. (2019). Time domains of hypoxia adaptation-elephant seals stand out among divers. Front. Physiol. 10:677. doi: 10.3389/fphys.2019.00677
Tift, M. S., Ponganis, P. J., and Crocker, D. E. (2014). Elevated carboxyhemoglobin in a marine mammal, the northern elephant seal. J. Exp. Biol. 217, 1752–1757. doi: 10.1242/jeb.100677
Umeda, A., Miyagawa, K., Mochida, A., Takeda, H., Takeda, K., Okada, Y., et al. (2020). Intermittent hypoxia, energy expenditure, and visceral adipocyte recovery. Respir. Physiol. Neurobiol. 273:103332. doi: 10.1016/j.resp.2019.103332
Wegiel, B., Nemeth, Z., Correa-Costa, M., Bulmer, A. C., and Otterbein, L. E. (2014). Heme oxygenase-1: a metabolic Nike. Antioxid. Redox Signal. 20, 1709–1722. doi: 10.1089/ars.2013.5667
Wilks, A. (2002). Heme oxygenase: evolution, structure, and mechanism. Antioxid. Redox Signal. 4, 603–614. doi: 10.1089/15230860260220102
Yang, D., Peng, Y., Ouzhuluobu, Bianbazhuoma, Cui, C., Bianba, et al. (2016). HMOX2 functions as a modifier gene for high-altitude adaptation in Tibetans. Hum. Mutat. 37, 216–223. doi: 10.1002/humu.22935
Yoshida, T., Maulik, N., Ho, Y. S., Alam, J., and Das, D. K. (2001). H(mox-1) constitutes an adaptive response to effect antioxidant cardioprotection: a study with transgenic mice heterozygous for targeted disruption of the heme oxygenase-1 gene. Circulation 103, 1695–1701. doi: 10.1161/01.CIR.103.12.1695
Zapol, W. M., Liggins, G. C., Schneider, R. C., Qvist, J., Snider, M. T., Creasy, R. K., et al. (1979). Regional blood flow during simulated diving in the conscious Weddell seal. J. Appl. Physiol. 47, 968–973. doi: 10.1152/jappl.1979.47.5.968
Zhang, R., Pan, K., Hao, Y., Yip, C., and Ko, W. (2019). Anti-inflammatory action of HO-1/CO in human bronchial epithelium in response to cationic polypeptide challenge. Mol. Immunol. 105, 205–212. doi: 10.1016/j.molimm.2018.12.002
Keywords: carbon monoxide, heme oxygenase, hypoxia, altitude, diving, evolution, cytoprotection
Citation: Tift MS, Alves de Souza RW, Weber J, Heinrich EC, Villafuerte FC, Malhotra A, Otterbein LE and Simonson TS (2020) Adaptive Potential of the Heme Oxygenase/Carbon Monoxide Pathway During Hypoxia. Front. Physiol. 11:886. doi: 10.3389/fphys.2020.00886
Edited by:
Dieter Blottner, Charité – Universitätsmedizin Berlin, GermanyReviewed by:
Enrico M. Camporesi, USF Health, United StatesCopyright © 2020 Tift, Alves de Souza, Weber, Heinrich, Villafuerte, Malhotra, Otterbein and Simonson. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Michael S. Tift, dGlmdG1AdW5jdy5lZHU=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.