
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
SYSTEMATIC REVIEW article
Front. Physiol. , 24 April 2019
Sec. Environmental, Aviation and Space Physiology
Volume 10 - 2019 | https://doi.org/10.3389/fphys.2019.00442
 Lucrezia Zuccarelli1,2
Lucrezia Zuccarelli1,2 Letizia Galasso3
Letizia Galasso3 Rachel Turner4
Rachel Turner4 Emily J. B. Coffey5
Emily J. B. Coffey5 Loredana Bessone2
Loredana Bessone2 Giacomo Strapazzon2,4*
Giacomo Strapazzon2,4*Background: Successful long-duration missions outside low-Earth orbit will depend on technical and physiological challenges under abnormal environmental conditions. Caves, characterized by absence of light, confinement, three-dimensional human movement and long-duration isolation, are identifiably one of the earliest examples of scientific enquiry into space analogs. However, little is known about the holistic human physiological response during cave exploration or prolonged habitation.
Objectives: The aim of our review was to conduct a systematic bibliographic research review of the effects of short and prolonged exposure to a cave environment on human physiology, with a view to extend the results to implications for human planetary exploration missions.
Methods: A systematic search was conducted following the structured PRISMA (Preferred Reporting Items for Systematic Reviews and Meta-Analyses) guidelines for electronic databases.
Results: The search retrieved 1,519 studies. There were 50 articles selected for further consideration, of which 31 met our inclusion criteria. Short-term cave exposure studies have investigated visual dysfunction, cardiovascular, endocrine-metabolic, immunologic-hematological and muscular responses in humans. Augmentations of heart rate, muscular damage, initial anticipatory stress reaction and inflammatory responses were reported during caving activity. Prolonged exposure studies mainly investigated whether biological rhythms persist or desist in the absence of standard environmental conditions. Changes were evident in estimated vs. actual rest-activity cycle periods and external desynchronization, body temperature, performance reaction time and heart rate cycles. All studies have shown a marked methodological heterogeneity and lack reproduction under controlled conditions.
Conclusions: This review facilitates a further comparison of the proposed physiological impact of a subterranean space analog environment, with existing knowledge in related disciplines pertaining to human operative preparation under challenging environmental conditions. This comprehensive overview should stimulate more reproducible research on this topic and offer the opportunity to advance study design and focus future human research in the cave environment on noteworthy, reproducible projects.
Long duration exploration and future human and robotic planetary surface missions are one of the long-term goals of space agencies (Robinson et al., 2007; Foing, 2016). Preparing for exploration-class missions to other planets requires a close replication of potential environmental and situational characteristics of extreme space conditions. Terrestrial space analogs offer a controlled environment for space research without leaving the planet (Pagel and Choukér, 2016). Since many years, isolation and confinement studies have been conducted in simulated scenarios such as the Mars500, Envihab and PlanHab facilities (Koch and Gerzer, 2008; Meigal and Fomina, 2016; Rittweger et al., 2016; Salvadego et al., 2018). In addition, real world scenarios based in extreme environmental conditions have been adapted to closely replicate the challenges of space exploration. What was previously the territory of only bold, enthusiastic explorers or honed ultra-endurance athletes has become the permanent training environment for the next generation of human elite space pioneers and clinicians. Simulated expeditions have been established in space analog locations around the globe, such as NASA Extreme Environment Mission Operations (NEEMO), Cooperative Adventure for Valuing and Exercising human behavior and performance Skills (ESA CAVES) and Antarctic programs in existing research stations (i.e., Concordia) (Lugg, 2005; Bessone et al., 2013; Anglin and Kring, 2016).
As a priority, better definition and investigation of physiological and psychosomatic effects of long-term permanence in space on the human body is required, as human performance directly impacts on safety and the mission outcome. In this regard, isolation and confinement specific to space habitat simulations have previously been identified as factors that heavily impact on the homeostatic regulation of human physiology, inclusive of various biological variables (Czeisler and Gooley, 2007), as well as behavioral health, cognitive and physical performance (Basner et al., 2014; Pagel and Choukér, 2016; Mogilever et al., 2018). Specifically, alteration of the biological rhythm (Pittendrigh, 1960) was one of the first topics, and is still of great interest to space agencies worldwide (Czeisler and Gooley, 2007; Robinson et al., 2007; Foing, 2016). Caves are one of the earliest scenarios where investigations specifying long-duration confined-isolation were conducted. As early as 1938, well before the launch of any Antarctic winter-over observational studies, Kleitman and his assistant spent 32 days in the Mammoth Cave, Kentucky, US, to specifically investigate how the absence of natural environmental time cues affects the circadian rhythm (Kleitman, 1963). Later isolation studies by Siffre and Montalbini attracted the interest of space agencies in the late 1970s (Siffre, 1988, 1990; Sonnenfeld et al., 1992). Despite this early interest and the inclusion of caves as an analog environment into the ESA astronaut training program since 2011 (Strapazzon et al., 2014; Mogilever et al., 2018), little data have been published in peer-reviewed journals since, and no systematic review exists of the work completed to date from a physiological or clinical perspective. International space agencies worldwide, motivated by a vision for long-term extra-terrestrial habitation and exploration, have expressed a need to better understand the potential effects of this complex cave space analog on the physiological functionality of trained astronauts. An analog which allows the direct investigation of individuals specifically trained to concurrently perform complex physical tasks and fulfill explorative responsibilities, while exposed to an enclosed, dark, atmospherically challenging, hot and humid environment.
The aim of this work is to systematically review studies investigating the effects of short and prolonged exposure to a cave environment on human physiology, to facilitate the greater understanding of all available primary research in this field and extend the results to the implications for human planetary exploration missions and space medicine considerations in the future.
A systematic review of the literature was conducted following the PRISMA (Preferred Reporting Items for Systematic Reviews and Meta-Analyses) Guidelines for Systematic Review (Liberati et al., 2009).
An extensive literature search was performed using recognized life science and biomedical electronic databases and by manually searching reference lists of those articles found, which specifically investigate human physiological responses to the cave environment. No language, publication date, or publication status restrictions were imposed. This search was applied to the following electronic databases: Medline (1966-Present), NASA Technical Reports Server (1915-Present), Google Scholar (all electronic resources-Present), Worldcat (1971-Present), OPAC (1831-Present), and PubMed (1997- Present). The last search was run on 24 November 2017.
Three members of the project team developed the search strategy (GS, LB, LZ). The search concepts included studies in the cave environment; the following search approach was used: 01. “cave/s” [All Fields], 02. “cave” [All Fields] AND “isolation” [All Fields] AND “human” [All Fields], 03. “free running isolation” [All Fields], 04. “potholing” [All Fields], 05. “potholer/s” [All Fields], 06. “caving” [All Fields], 07. “social isolation” [All Fields], 08. “subterranean” [All Fields] AND “isolation” [All Fields], 09. “underground environment” [All Fields], 10. “grotta/e” [All Fields], 11. “isolamento in grotta” [All Fields], 12. “isolamento spazio temporale” [All Fields], 13. “Montalbini” [author], 14. “grotte” [All Fields], 15. “sejours souterrain” [All Fields], 16. “Siffre” [author], 17. “cueva” [All Fields], 18. “aisolamento in cueva” [All Fields], 19. “permanencer bajo tierra” [All Fields], 20. “spelunka” [All Fields], and 21. “spelunking”. Additional manual reviews were performed in specialized speleological databases (see Supplementary Table 1), and forty experts (i.e., seven authors of scientific publications, twenty-one health care providers related to cave/ sports medicine, ten speleologists/ health care providers with direct experience in underground isolation; two journalists and librarians; see Supplementary Table 2) were surveyed/interviewed for additional information.
Animal studies in the cave environment were excluded from this review. Subjects of any age, gender, medical condition or caving experience were included. Full-length and abstract (i.e., when full-length was not available and the abstract contained original data) peer reviewed articles, as well as other relevant scholarly articles with original data, were considered eligible for inclusion. All retrieved records were screened by title and abstract by one review author (LZ). The review author (LZ) rated each citation using a ”relevant,“ ”irrelevant,“ or ”unsure“ designation. Only retrieved records that received a ”relevant“ or ”unsure“ classification were fully read. These selected articles were then classified into nine different categories (i.e., atmospheric science, cardiovascular function, emergency medicine, human factors, muscle/skeletal system, neuroscience, psychological aspects, radiation and respiratory function), in accordance with relevant topics for space medicine (Videbaek et al., 1993). Only records classified as ”physiological studies“ were included in the current review and two review authors (LZ, GS) independently assessed full-text records (i.e. each record was checked twice). The eligibility assessment was also performed independently, in a standardized manner by two different authors (LZ, GS). Disagreements between reviewers were resolved via discussion and all reasons for study exclusion were recorded.
The studies evaluated in this review were further categorized based on the duration of time spent in the cave environment. We classified studies based on cave exposure time: cave exposure up to 72 h as short and above 72 h as prolonged.
Data from included studies were summarized as follows (see Tables 1, 2): the publication details (e.g., publication year, format type, publication status); characteristics of participants (e.g., number, gender, age), and study specifics (e.g., experimental protocol, location of a study and measured parameters). The primary outcomes were defined as the effects of cave exposure on homeostatic control and physiological functions. For the short exposure studies, data about bone composition, cardiovascular, endocrine-metabolic, immunologic, muscular, and visual dysfunction were extracted. In addition, for prolonged exposure studies we extracted measurements related to the rest-activity cycle, thermoregulation, psychomotor function, cardiovascular adaptation, immunologic and hematological changes, bone composition, menstrual cycle, and renal system.
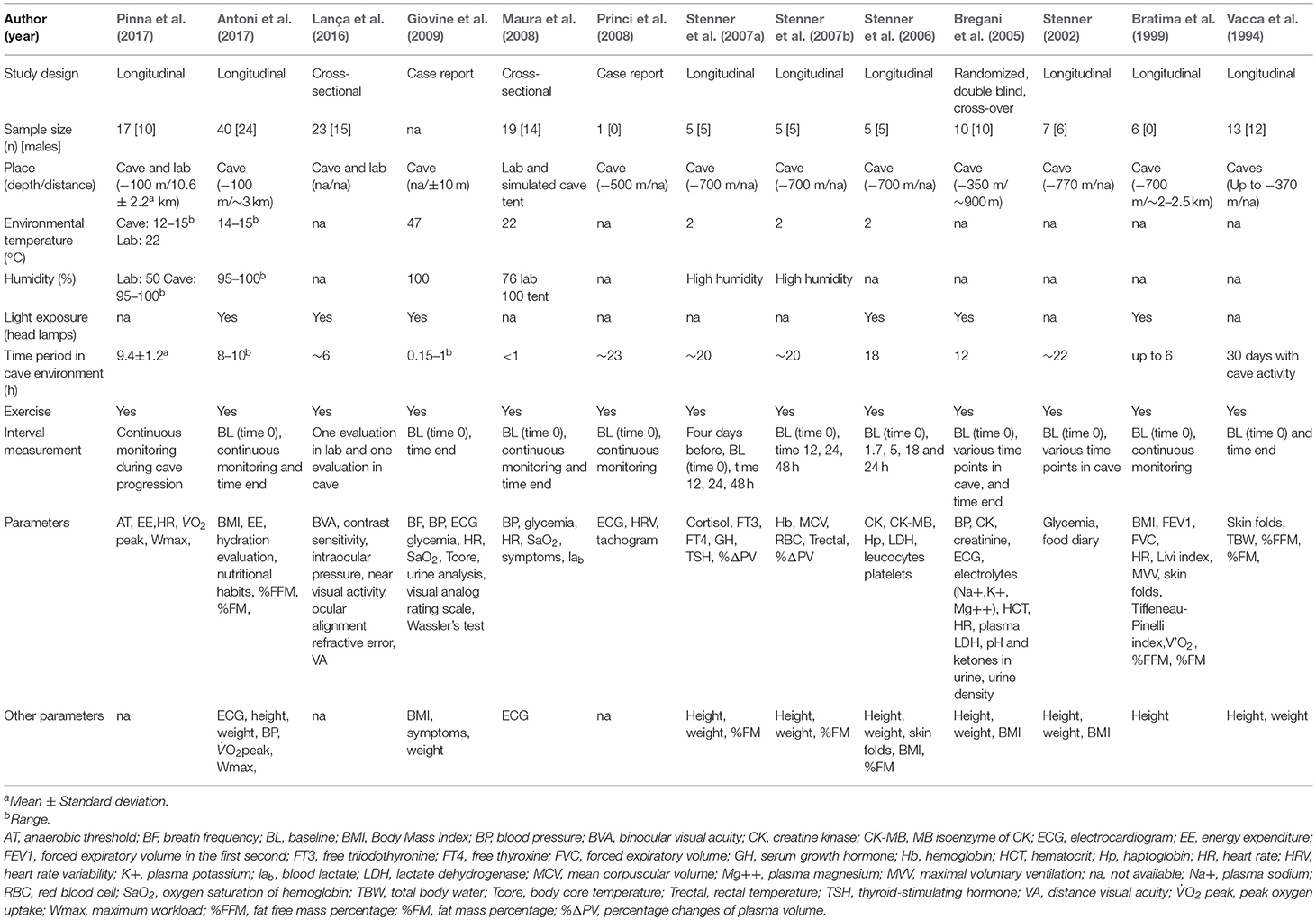
Table 1. Characteristics of short exposure studies.
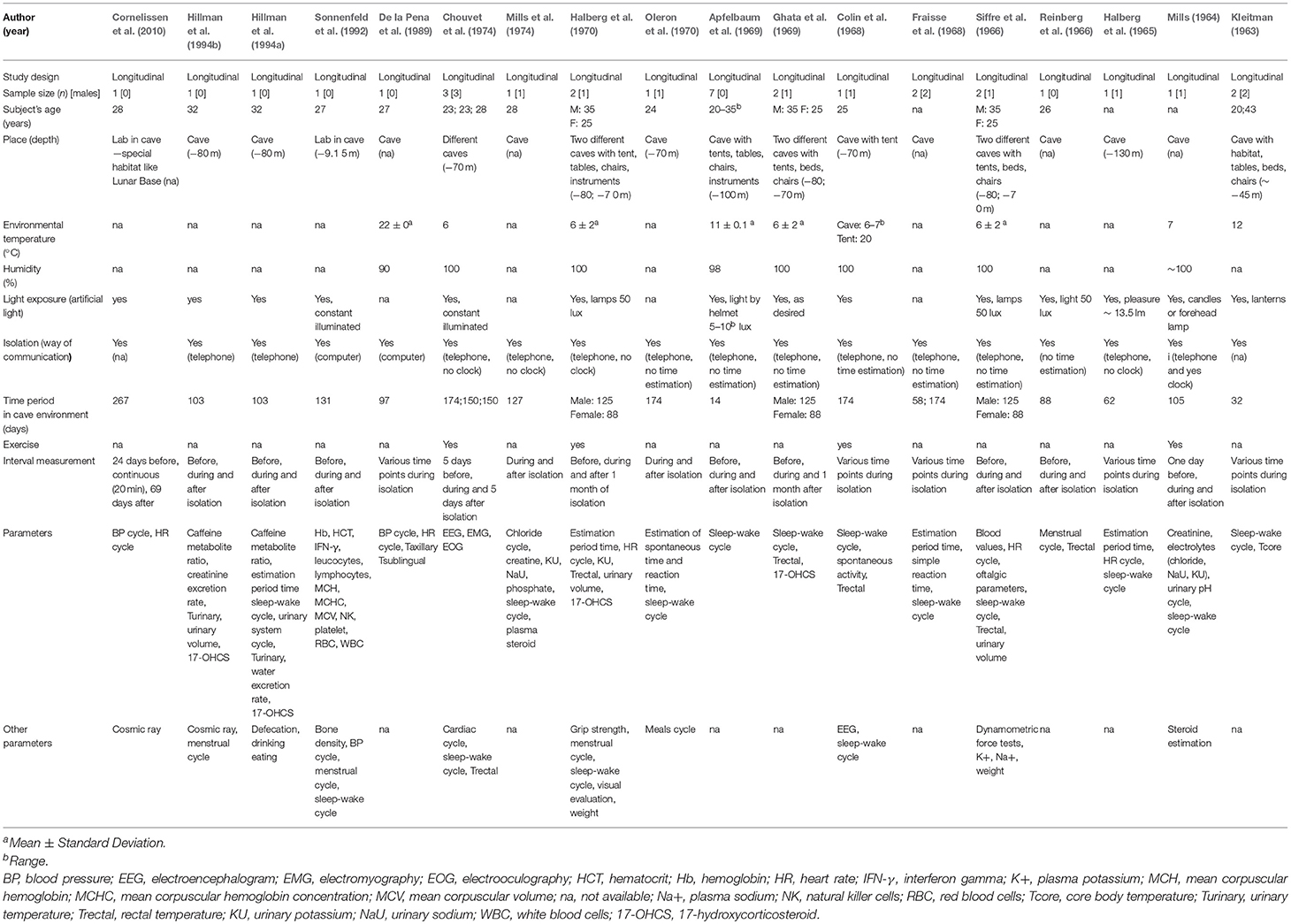
Table 2. Characteristics of prolonged exposure studies.
The systematic search (including manual review and expert suggestions) retrieved 1,519 articles (Figure 1). After the initial screening, 50 articles were retained; 31 of these were classified as ”physiological studies.“ A list of (and reasons for) the 19 excluded articles is recorded in Figure 1 and Supplementary Table 3.
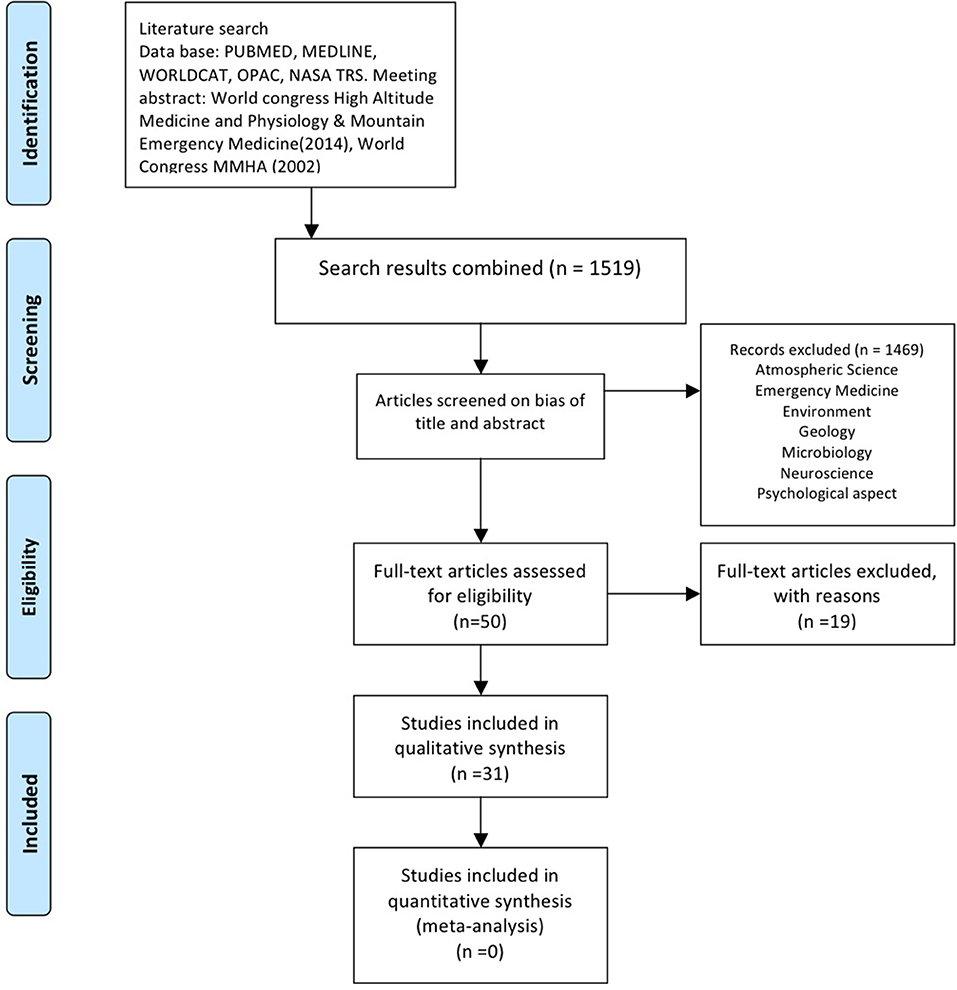
Figure 1. Search and screening strategy based on PRISMA guidelines.
Thirteen papers were categorized on the short exposure effects and 18 on prolonged exposure. The characteristics of those studies considered eligible for inclusion are summarized in Tables 1, 2, respectively.
All 31 studies included in the review contained original data and were published in either English, Italian or French. The studies differed in study design (e.g., time exposure, exercise, isolation and diet), cave environment characteristics (e.g., temperature, altitude and artificial light/environment exposure), monitored parameters, and measurement interval. The marked methodological heterogeneity across these studies and the limited number of papers prevented meta-analysis. Hence, we continue to describe these results qualitatively.
These 13 studies, all published between 1994 and 2017, include a total of 151 participants in different cave/lab simulation settings. Cave/lab simulation exercises were performed in all studies; exercise can be classified as cave progression (characterized by atypical, strenuous movement, often combining ascent, descent, and scrambling activity) in all studies apart from one, where an exercise stress test in the simulated cave-environment condition was completed (Maura et al., 2008). Anthropometric characteristics were measured in the majority of the studies (Vacca et al., 1994; Bratima et al., 1999; Stenner, 2002; Bregani et al., 2005; Stenner et al., 2006, 2007a,b; Maura et al., 2008; Antoni et al., 2017; Pinna et al., 2017). Overall, study subjects were healthy with heterogeneous anthropometric characteristics. Cardiovascular, endocrine-metabolic, immunologic-hematological, muscular responses, and visual dysfunction were investigated (Table 1). The corresponding trend analysis of the results for all studies can be found detailed in Table 3.
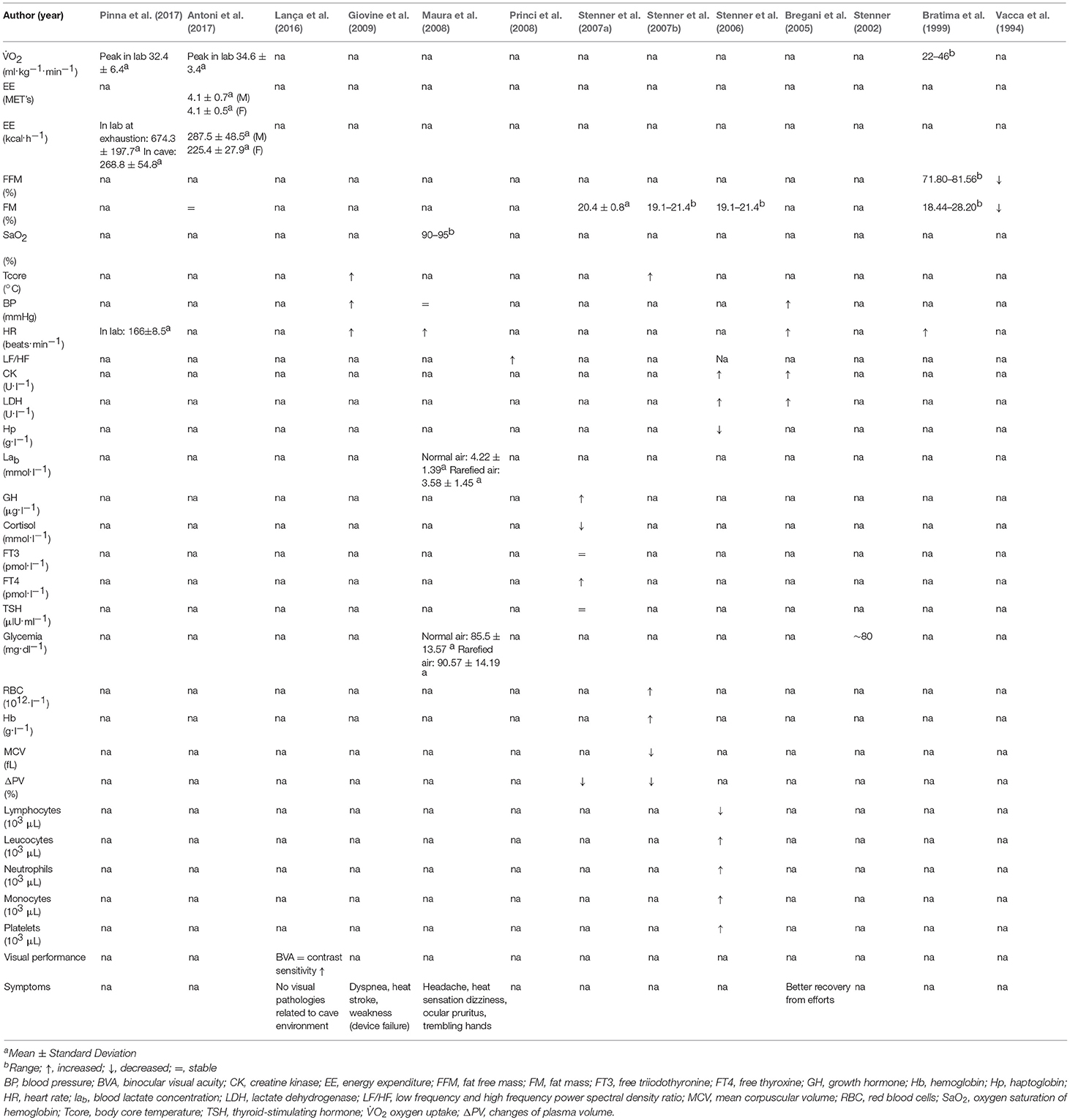
Table 3. Short exposure studies main results.
One study (Bratima et al., 1999) reported that heart rate (HR) increased more during cave traverse (ascent and scrambling activity) compared to descent into the cave. Other studies have not been able to substantiate these findings either pre or post, reporting only a trend for increased HR during cave exploration, without presenting sufficient data to that effect (Vacca et al., 1994; Giovine et al., 2009). Notably, a later study by Maura et al. (2008) used a unique scientific approach, directly studying the environmental factors that may be definitive in HR modification, in a specially built subterranean laboratory. This study demonstrated that in comparison to normoxic conditions, HR was on average increased by 10 beats.min−1 (p < 0.0002), when cavers performed a constant work rate exercise on a cycle ergometer at 75% of their maximum theoretical heart rate in an hypoxic (O2 ~15%) hypercapnic (CO2 ~1.9%) humid (100%) simulated cave atmosphere (Maura et al., 2008). These results were corroborated further in a later case report, detailing a cave exploration under similar conditions, which showed a relevant modification of the cardiac autonomic nervous system (ANS) activity through a reduction in heart rate variability (HRV) and a shift in the sympathovagal balance toward more sympathetic and less parasympathetic activity in a female elite potholer (Princi et al., 2008).
Physiological/psychological responses during cave progression have been characterized by indicative stressors considered able to alter the hormonal balance and metabolic homeostasis in humans. Previous work focused on hypothalamus-pituitary adrenocortical, hypothalamus-pituitary and hypothalamus-pituitary thyroid system responses to duration cave activity of ~20 h. Results demonstrated an evident elevation of serum cortisol prior to cave entry, even in elite cavers. This initial anticipatory stress reaction was reportedly reduced over time, followed by a noticeable increase in growth hormone (GH) production during prolonged explorative cave activity (Stenner et al., 2007a). Recently, Antoni et al. (2017) reported that total energy expenditure (TEE) was in the range 225–287 kcal.h−1 for women and men for a standardized cave progression (i.e., 10 h subterranean exploration), which is equivalent to an average metabolic equivalent of task (METs) of 4.1 METs. Energy intake during this cave exploration (determined by means of a self-administered dietary recall) was calculated to be ~100–120 kcal. h−1 (Antoni et al., 2017), this value was also supported by previous work in this field (Stenner, 2002). Bayesian statistical analysis estimated the effect of predictive variables on TEE, revealing that experienced cavers had a 5% lower TEE than those less skilled, plus female participants required a comparatively larger EE than men to perform the same task. Previous work has shown a significant decrease in fat mass estimated by skin folds and bio-impedance after repeated daily cave progression (Vacca et al., 1994), however as expected no change in fat mass was reported after only 10 h of cave exploration (Antoni et al., 2017). Glycemic values were shown to be in the normal range both during a 22 h continuous cave progression (Stenner, 2002) and during stress testing (≤ 1h) in a simulated cave environment (Maura et al., 2008).
A previous study demonstrated an initial increase in red blood cell (RBC) count and hemoglobin (Hb) concentration after only ~5 h of cave exploration, although this increase was in parallel with a concomitant reduction in plasma volume (PV) (Stenner et al., 2007b). Previous work from the same research group demonstrated that the number of leucocytes, neutrophils, monocytes, and platelets were significantly increased after cave progression, whereas lymphocytes showed a concomitant significant reduction (Stenner et al., 2006).
Two studies showed an increase in serum markers for muscle damage and metabolism during cave progression (Bregani et al., 2005; Stenner et al., 2006). Specifically, there was a clear increase in serum creatine kinase (CK) and lactate dehydrogenase (LDH) (Bregani et al., 2005; Stenner et al., 2006). Haptoglobin values were also significantly decreased (Stenner et al., 2006). The effect of specific nutritional supplementation of creatine and branched-chain amino acids administration after cave progression was investigated in a cross-over, randomized, double blind study (Bregani et al., 2005). Participants treated with creatine showed lower levels of CK and LDH after cave progression in relation to the control group. The LDH increase after exercise was more pronounced in controls, even if it was not significant; post cave exercise, CK values were increased significantly, but less so in supplemented participants.
No signs or symptoms of visual dysfunctions where reported post intermittent exposure to the cave environment in a recent study (Lança et al., 2016). Binocular visual acuity (BVA) was reported to be −0.05 ± 0.15 LogMAR (20/18) with a head light with a mean illuminance of 451.0 ± 305.7 lux. Significant improvements in contrast sensitivity were observed with 450 nm filters for 6 cycles per degree (cpd) and 18 cpd spatial frequencies.
The 18 studies included within this review that were related to prolonged exposure to the cave environment were published between 1963 and 2010. In total, all except one of these prolonged exposure studies exposed participants directly to the cave environment for >30 days (Table 2). Specifically, in Sonnenfeld et al. (1992) and Cornellissen at al. (2010), participants were accommodated within a laboratory built inside a cave, while in the others there was no such facility. Overall, in all 18 studies, participants were healthy and in a number of articles under review, the same participants were recruited repeatedly. In total, 20 participants were studied, with a good proportion of male and female (nine and 11, respectively).
Those studies inclusive of an isolation component mainly investigated whether biological rhythms persist or desist in the absence of standard environmental conditions (e.g., without information related to the 24 h clock). During underground isolation, participants were asked to follow a regular wake-sleep cycle, including customary meal times in the absence of any scheduling cues (Mills, 1964; Mills et al., 1974) (Table 2). Corresponding trend analysis of results for all studies can be found detailed in Table 4.
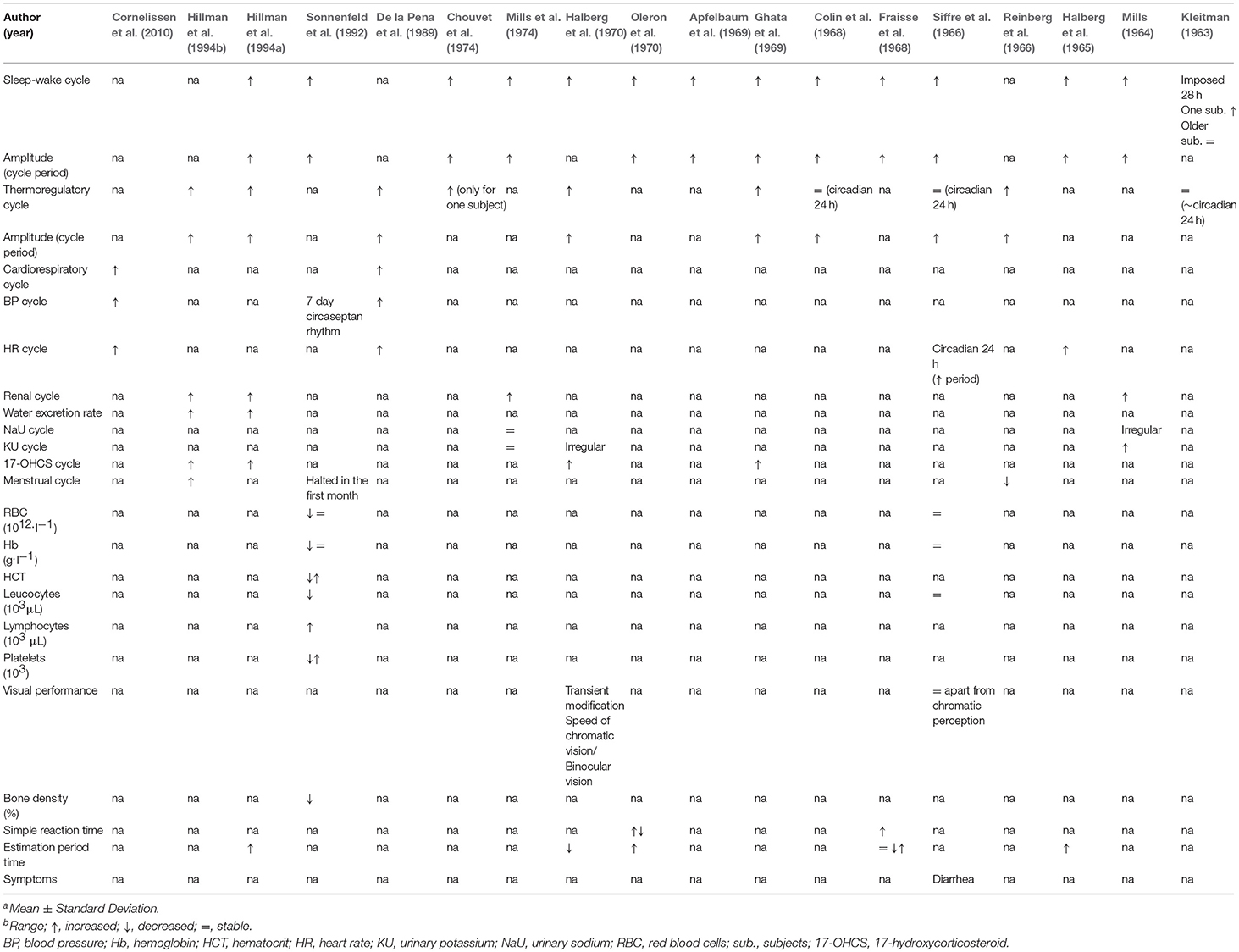
Table 4. Prolonged exposure studies main results.
The alterations observed in individual rest-activity cycles affected all physiological variables considered in the studies under review. A rest-activity cycle persisted in the absence of any environmental synchronizer or deliberate scheduling, however it appeared desynchronized from an exact 24 h period in all the 14 studies included in review where it was investigated (Kleitman, 1963; Mills, 1964; Halberg et al., 1965, 1970; Siffre et al., 1966; Colin et al., 1968; Fraisse et al., 1968; Apfelbaum et al., 1969; Ghata et al., 1969; Oleron et al., 1970; Chouvet et al., 1974; Mills et al., 1974; Sonnenfeld et al., 1992; Hillman et al., 1994a).
Colin et al. (1968) observed that at the beginning of their 6-month isolation case study, both the actual and estimated rest-activity cycles of their single male participant was near circadian (25 h and 27 h, respectively). After only 10 days of isolation, the participant reached values of 45 h referring to the time between two awakenings and 50 h referring to the estimated period of activity. The estimated period of activity progressively increased (bicircadian), while the wakening-to-wakening cycles returned to circadian (Colin et al., 1968). Later, a further study with a similarly long duration isolation (Chouvet et al., 1974) also showed similar discrepancies between the actual and the estimated wakening-to-wakening cycles, reporting data from polygraphic recordings [i.e., electroencephalographic activities (EEG) and electromyography (EMG)] in three participants with variable latencies and several variations in sleep observations. All participants reached a bicircadian rhythm (34 h of wakefulness followed by 14 h of sleep), which they subjectively considered to be a 24 h cycle. However, only one participant was able to adapt completely to this new rhythm and maintain it >2 months. The use of an external Zeitgeber (i.e., 500 W lamp) was effective in reducing the relative variability of this rhythm. Paradoxical sleep (PS; also known as rapid eye movement or REM sleep) remained unchanged while the sleep-waking rhythm was disrupted in all, but PS onset latencies in the two participants unable to maintain a bicircadian rhythm were shortened (Chouvet et al., 1974). Hillman et al. (1994a), showed that a female participant at the start of 103 days of isolation demonstrated an immediate change in rest-activity cycle to ~25 h period. In the middle of the span, this period reportedly shortened slightly but still remained longer than 24 h. In addition, the participant showed a dissociation between observed and estimated daily duration, averaging ~58 h per subjective ”day.“ Differences between the perceived time elapsed and the actual time elapsed (calendar days) was found also by Halberg et al. (1970), indicating that the two participants both perceived the day to be longer than normal, over the course of the isolation period (88–128 days).
Similar trends were also previously observed in long duration isolation studies (Fraisse et al., 1968; Mills et al., 1974), where different male participants also reported dissociation between observed and estimated cycle duration. More specifically, Mills et al. (1974) demonstrated in a male participant isolated for a total of 4 months (127 days) an extended estimated day length adopted by the subject, with an estimated activity period ranging from ~19 to 66 h (Mills et al., 1974). Previously, Mills demonstrated that the sleep-activity cycle was considered externally desynchronized. However, the internal synchronization reportedly persisted into a rhythmic process, with a period of ~24.5 h (Mills, 1964). Finally, Halberg et al. (1965) showed that a 2-month isolation in a cave environment, despite the desynchronization with respect to local time, an internal synchronization of the rhythm functions was maintained (Halberg et al., 1965). Overall an internal rhythm was maintained, without the influence of environmental synchronizers, with different period not based on the 24 h (and for this reason is called free-running).
The circadian rhythm of the body temperature was investigated in 10 studies (Kleitman, 1963; Reinberg et al., 1966; Siffre et al., 1966; Colin et al., 1968; Ghata et al., 1969; Halberg et al., 1970; Chouvet et al., 1974; De la Pena et al., 1989; Hillman et al., 1994a,b). The first underground study investigating the rhythm of body temperature, Kleitman (1963), revealed a persistence of a circadian rhythm for a month in a participant living on a forced-desynchrony protocol (i.e., non 24 h sleep/wake cycle, with no regular light stimulus). In contrast to the work by Kleitman (1963), Colin et al. (1968) compared the rhythms of body temperature and spontaneous activity for a 6-month isolation period, observing maintenance of a circadian rhythm of the rectal temperature throughout the duration of isolation with a progressive lengthening. However, in an earlier study, over the course of ~3 months subterranean isolation rectal temperature decreased with circadian desynchronization compared to local time following a circatrigintan rhythm (i.e., a biologic variation or rhythm with a frequency of 1 cycle in 30 ± 5 days) (Reinberg et al., 1966). There was a similar observation in later study, in which an internal circasemiseptan rhythm desynchronization (relating to biologic variations or rhythms with a frequency of 1 cycle in 3.5 ± 1 days) was observed in urinary-temperature (Hillman et al., 1994b).
Several studies reported data related to psychomotor tests performed by subjects during underground isolation (Halberg et al., 1965, 1970; Fraisse et al., 1968; Oleron et al., 1970; Hillman et al., 1994a). Estimation of a short interval of time (i.e., number of seconds the subject took to count from 1 to 120 s with the goal of evaluating a duration of 2 min) showed a progressive increase within a 2-month cave isolation (Halberg et al., 1965). Oleron et al. (1970) and Fraisse et al. (1968), reporting results from one subject, showed an accelerated spontaneous time at the end of the isolation period. During these studies, the participant was required to tap as regularly as possible on a telegraphic key with their middle finger only. The number of taps per 10 s was calculated, plus the interval duration between two taps on average. Moreover, the simple reaction time (i.e., subject was asked to press as quickly as possible on a telegraph key at the appearance of a light signal) was reportedly increased during a prolonged stay in the cave environment.
HR values and spectrum analysis were covered by four of the studies under review (Halberg et al., 1965; Siffre et al., 1966; De la Pena et al., 1989; Cornelissen et al., 2010). In all of these studies, the HR cycle was desynchronized from environmental synchronizers (i.e., alternation of light-dark cycle, seasons, and social routine), which help to maintain a normal circadian rhythm of ~24 h. HR spectrum analysis during a free-running isolation experience (Siffre et al., 1966) showed that the circadian rhythm persisted with a slight lengthening (24.8 and 24.6 h for the male and female subject, respectively). The three other studies supported this proposition for a persistent circadian drive, regardless of external desynchronization (Halberg et al., 1965; De la Pena et al., 1989; Cornelissen et al., 2010). Equally, similar results were found for blood pressure (BP) spectrum analysis (De la Pena et al., 1989; Cornelissen et al., 2010).
Only two studies investigated the immunologic and hematological responses due to underground isolation (Siffre et al., 1966; Sonnenfeld et al., 1992). Sonnenfeld et al. (1992) demonstrated in a female a decrease in leukocytes and concomitant increase in both lymphocytes and monocytes, including natural killer T-cells, during a prolonged isolation (131 days). After an initial decrease in RBC, Hb, platelets and hematocrit (HCT) after two weeks, RBC and Hb values reportedly normalized, whereas HCT and platelets increased over and after the isolation period (4 months). However, these results are in direct conflict with the observations in two subjects of Siffre et al. (1966), which demonstrated no changes in RBC, Hb, and leucocytes after an isolation period of either 2 or 4 months.
One study reported an overall bone loss of 2% (up to 8.2% in trabecular bone) after 131 days of cave isolation in a woman with a vitamin D-depleted diet (Sonnenfeld et al., 1992).
Changes in the menstrual cycle have been studied in two different women during long isolation in a cave (Reinberg et al., 1966; Hillman et al., 1994b). The menstrual cycle was slightly reduced (from 29 to 26 days) during the isolation period of 88 days (Reinberg et al., 1966). Opposite results, with a lengthening and desynchronization of the menstrual cycle (from 25 to 27–32 days) were also observed in a similar 3-month isolation study (Hillman et al., 1994b).
Seven studies investigated parameters of renal function during cave isolation (Mills, 1964; Siffre et al., 1966; Ghata et al., 1969; Halberg et al., 1970; Mills et al., 1974; Hillman et al., 1994a,b). During a long permanence in a cave (≤ 4 months), the biological rhythm of urinary excretion of 17-Hydroxycorticosteroids persisted with a desynchronization from circadian rhythm (24 h) (Ghata et al., 1969). Equally, after a similar prolonged isolation study (3 months), the excretion of potassium was also shown to be irregular (Halberg et al., 1970). Mills (1964) in their early work supported this finding, demonstrating a desynchronized rhythm of excretion in both potassium and particularly chloride. Urinary pH was roughly correlated with rate of potassium excretion and creatinine excretion pattern was irregular, without definitive conclusions (Mills, 1964). However, in a later publication they concluded the period of circadian excretion of potassium, sodium and chloride remained similar during cave isolation (127 days), demonstrating a potential variability in methodology, or simply a lack of reproducibility of specific study conditions (Mills et al., 1974). Finally, subsequent studies continued to further demonstrate an internal circasemiseptan desynchronization in water excretion rate, intermicturition interval and caffeine metabolite ratio over a 3-month isolation period (Hillman et al., 1994a,b).
Space medicine is akin to sports medicine in that it is commonly described as dealing with normal physiology in abnormal conditions. Despite this reality, knowledge surrounding the provision of successful physiological preparation in subterranean space analog scenarios remains limited. A current proposition for Martian caves and lava tubes as potential locations for sheltered habitation on Mars, have highlighted the need to better understand both the environmental impact this may infer on pioneering space cohorts and inform the type of training necessary to enable further human space exploration. Previous human studies investigating the effect of the unique cave environment on homeostatic physiology fall into two categories: either short or prolonged exposure. Short-term cave exposure studies have shown an augmentation of heart rate, muscular damage, initial anticipatory stress reaction, and inflammatory responses during cave progression. The cave environment seems to lead to supra-elevated energy expenditure, despite the moderate intensity implied, especially if continued for several hours. Prolonged exposure studies mainly investigated whether biological rhythms persist or desist in the absence of standard environmental conditions. Changes were evident in estimated vs. actual rest-activity cycle periods and external desynchronization, body temperature, performance reaction time, and heart rate cycles. Studies have shown a variability related to the different study environments and design (e.g., time period in cave environment, exercise protocol, control of the influence of environmental characteristics). The establishment of the cave environment as a space analog has further focused these studies on both the acute response to cave progression, as well as adaptive mechanisms involved when mid and long duration exposure to altered atmospheric composition, artificial light, isolation, confinement and challenging combinations of heat and high humidity were imposed. As a result, the environmental effect of cave progression, inclusive of alterations in the circadian rhythm, have been identified as important for informing practical considerations when defining implications for astronaut expeditionary training courses in subterranean space analogs. Nevertheless, through systematic review, we have identified knowledge gaps and areas that would benefit from additional research efforts, such as developing specific criteria for the collection of scientific data specifically related to clinical grade physiological monitoring in challenging subterranean environments. This approach could prove popular throughout organizations where highly trained human operatives are exposed to complex, dangerous working conditions, often in remote locations, void of immediate clinical care and evacuation.
Cave exploration is an atypical, strenuous, three-dimensional human movement specific to cave progression, with some similarities with mountaineering (i.e., a combination of ascending, descending and scrambling). The cave studies selected for review herein include a number of cave progressions (see Table 3), which typically alternate between aerobic and anaerobic exercise models in individuals who specialized in caving. Antoni et al. estimated an EE during cave progression for up to 10 h with a prevalence of scrambling in agreement with previous work by Bratima et al., which detailed a prevalence of ascending and descending activity during the traverse (Bratima et al., 1999; Antoni et al., 2017). Nevertheless, similar to a number of case studies including long duration cave progression, there is a lack of detailed analysis of the traverse obstacles or segmentation of the cave into different logistical categories. Current study results pertaining to estimated EE (as well as METs) have shown that a standardized cave traverse can be considered to elicit an energy expenditure equivalent to moderate physical activity (Ainsworth et al., 2000): comparable to modest intensity activities such as fishing, hunting and visiting natural environments recreationally (Ainsworth et al., 2000; Elliott et al., 2015). Despite this, it should be kept in mind that cave explorations are usually carried out for prolonged periods of time, under atypical atmospheric and temperature conditions. This type of prolonged physical effort, despite the moderate intensity implied, could lead to supra-elevated energy expenditure, which should be considered during space exploration analog training models. Previous research regarding Extravehicular Activity (EVA) during Shuttle and ISS missions, as well as in Lunar analogs, seem to be characterized by a very low to moderate metabolic demand and several hours of activity (Johnston et al., 1975; Downs et al., 2018). The estimated EE of 4.1 METs reported during caving activity can be compared to the amount of oxygen required for 28 Apollo EVA (i.e., mean O2 = 10–14 ml.kg−1.min−1) (Johnston et al., 1975). Moreover, “Martian” surface traverse (i.e., 1,500 m track covered with rocks and pebbles) and hill climbing, movements typically present during a cave progression, were recently used as tests able to simulate crewmember mission tasks (Sutterfield et al., 2019). In order to further knowledge on the physical ‘cost’ of exploratory movement in these subterranean environments, more specific studies are required. Mapping atypical movement patterns in such a subterranean environment is challenging. Accurate data collection in these challenging environments still remains predominantly a technological challenge, but it is an area where large gains could be made once movement patterns and profiling of such atypical, specialist movement are better characterized. However, even with fundamental assessment, many studies have postulated an inadequate restoration of lost calories and fluid in relation to the level and duration of movement recorded in the cave environment (Vacca et al., 1994; Bratima et al., 1999; Stenner, 2002; Stenner et al., 2007b; Antoni et al., 2017). Moreover, water losses during cave activity may also lead to acute hematological modification (i.e., increase in red blood cells and hemoglobin) as hypothesized by Stenner et al. (2007b).
Only studies in high altitude mountaineering scenarios have focused on energy expenditure over a long duration (Westerterp et al., 1992; Hoyt et al., 1994; Pulfrey and Jones, 1996; Reynolds et al., 1999; Miller et al., 2013). However, despite similarities in movement patterns, high altitude ascent does not represent a comparable model (or subsequent physiological impact) to those environmental conditions encountered in caves, especially in relation to humidity, constant temperature, and levels of hypercapnia. In order to further delineate the combined effect of multiple environmental factors found in a cave, Maura et al. (2008) tried to determine the physiological effect of an atmosphere similar to a possible cave environment at high altitude (O2~15%, CO2~1.9%, humidity 100%) during which participants completed a stress test. The given hypothesis, that when completing strenuous activity under hypoxic, hypercapnic and humid conditions, for the same given exercise an additional physiological burden and symptomology is evident and should be pre-empted in future training and analog assessments (Maura et al., 2008).
Equally, previous research related to endocrine and immune-hematological changes not only showed the potential for an anticipatory stress reaction to the cave environment, based on increased cortisol levels (Stenner et al., 2007a), but also suggested that there may be a compensatory response to both exercise and environment [evidenced by decreased cortisol levels and restoration of GH levels 24 h post cave exit (Stenner et al., 2006, 2007a,b)]. Due to the pulsatile nature of GH levels in the blood, conventional measurements of serum GH are highly variable dependent on a number of environmental stressors. GH values are usually increased with stress, exercise and low blood glucose values (Godfrey et al., 2003). Given the challenging terrain and climate intrinsic to the cave environment, caving may prove to be a strong stimulus for GH secretion. The concentration of GH in the blood can increase with time for given work intensity and can increase 10-fold during prolonged moderate exercise (Saugy et al., 2006). Therefore, long periods of potentially cyclical aerobic-anaerobic caving activity, normally almost without rest, could elicit similar increases in GH. Plus, traversing specific near vertical ascents could be considered comparable to a short duration intensive exercise (with accumulation of lactate at 70% O2 max for a short term period such as 10–20 min), which has previously been proven to increase GH by 5/10-fold (Felsing et al., 1992). Further long duration study of endocrinological factors, plus potential hormonal compensation, while completing exploratory tasks in the cave environment, would further add to both understanding of appetite regulation and quantification of the level of physiological stress induced during both short and prolonged active exposures. Finally, cave progression can include prolonged, multimodal, challenging traverse activity with intensive moments that can induce skeletal muscle damage and intravascular hemolysis also accompanied by the activation of inflammatory cells, as shown by the increased levels of CK and LDH, decreased level of haptoglobin and the significant increase in leucocytes due to neutrophila and monocytosis (Bregani et al., 2005; Stenner et al., 2006). These responses seem to be related both to the intensity of the exercise, pre-determined fitness/training level, overall fluid losses and the level of physiological strain induced by the specific atmospheric environmental conditions of the cave environment (Stenner et al., 2007b), and could be reduced by a specific diet. Specifically, supplementation of creatine seems to mitigate muscular damage, which typically occurs during cave progression (Bregani et al., 2005).
During underground cave isolation, inclusive of the absence of direct interpersonal relations and external time cues, changes in the circadian system frequency are demonstrated (i.e., rest-activity cycle, rectal temperature, urine volume. and excretion of potassium) (Kleitman, 1963; Mills, 1964; Halberg et al., 1965, 1970; Reinberg et al., 1966; Siffre et al., 1966; Colin et al., 1968; Fraisse et al., 1968; Apfelbaum et al., 1969; Ghata et al., 1969; Oleron et al., 1970; Chouvet et al., 1974; Mills et al., 1974; De la Pena et al., 1989; Sonnenfeld et al., 1992; Hillman et al., 1994a,b). Consequently, a desynchronization of the biological rhythm from the environmental cycle is evident. External desynchronization is evident when a biologic rhythm becomes desynchronized from an environmental cycle. On the contrary, internal de-synchronization is described when two or more biologic rhythms in the same entity become desynchronized from each other (Halberg, 1968).
Interestingly, desynchronization with respect to local time has been observed in participants isolated in caves without time cues: in some cases individual rhythms showed external circadian de-synchronization while maintaining reasonably fixed time relations among themselves, i.e., a continuation of internal circadian synchronization (Colin et al., 1968; Mills et al., 1974; De la Pena et al., 1989; Hillman et al., 1994b; Cornelissen et al., 2010). For example, when referring to the rest-activity cycle of subject isolated in a cave for several months, a free-running circadian component was reported (i.e., independent of any rhythm in habit or environment). Nevertheless, even in the absence of natural light (or other external cues) as a synchronizer (i.e., an environmental or operative periodicity that determines the temporal placement of a given biologic rhythm by impelling the rhythm to assume synchronization), the circadian rhythm of many physiological parameters is actually maintained at the beginning of a given cave isolation. However, with a subsequent increase in the duration of cave occupation desynchronization occurs, especially related to the duration of circadian rhythm (Halberg et al., 1965). Changes were evident not only in estimated vs. actual rest-activity cycle periods and external desynchronization, but also related to body temperature, performance reaction time and heart rate cycles. Conversely, changes related to the immunological and hematological cycle, as well as the menstrual cycle and renal secretory circadian function, were reported with uncertainties.
During a forced desynchrony protocol in which light was externally controlled, Kleitman (1963) observed a persistent of a circadian rhythm of body temperature. In agreement, but under different free of time cues isolation conditions inside a laboratory, Czeisler et al. (1999) reported that the intrinsic period of the core body temperature rhythm does not appear to have been dependent on the length of the imposed sleep-wake cycle (i.e., 20 h and 28 h forced desynchronization). In fact, the temperature period estimates are nearly equivalent under both forced-desynchrony protocols (average value of 24.28 h both under 20 h and 28 h), independent of the imposed rest-activity cycle. It seems that, despite the desynchronization with respect to local time, an internal synchronization of the rhythm function was maintained due to the presence of the human circadian pacemaker.
Finally, during early cave research, study protocol usually involved the majority of participants to be hospitalized post cave isolation (Mills, 1964; Halberg et al., 1970). In doing so, they were exposed to a new influence of alternate synchronizers. Full external circadian resynchronization of all physiologic functions occurred after a return to the social routine, inclusive of the alternation of day and night, normal meal times, plus small cues such as variations in noise from hospital and urban activity (Halberg et al., 1970).
Human sleep and circadian rhythms have evolved by way of adaptation to an environment characterized by a 24 h light-dark cycle and a gravitational force (Dijk et al., 2001). However, there are situations, similar to cave isolation, in which human beings are exposed to a non 24 h cycle. For example, during spaceflight astronauts are removed from the main synchronizer of the circadian system, the 24 h alternation of day-light and darkness (Monk et al., 1998). A free-run due to the absence of a 24 h light/dark cycle and increased sleep disturbances in astronauts occur more frequently during spaceflight than on the ground. Such disturbances may be caused also by exogenous factors (i.e., noise and uncomfortable temperature), but changes in the physiological basis for the regulation of sleep and the circadian clock may result specifically in sleep discomfort (Gundel et al., 1997). In fact, during spaceflight the crewmembers have reported a reduction in subjective sleep quality (Dijk et al., 2001), plus a reduction in sleep efficiency and a long sleep onset latency (Gundel et al., 1997). Decline in sleep quality may lead to a decrease in exercise performance and cognitive function (Fullagar et al., 2015), deterioration of proper immune system function (Lorton et al., 2006), and may even negatively impact upon individual probability of developing a form of cardiovascular disease (Reitz and Martino, 2015). Similarly, space flight can reportedly impact on body temperature regulation, i.e. body temperature may be increased during the imposed ”night cycle“ in comparison to normal, which may be an indication of reduced circadian amplitude (Gundel et al., 1997; Fortney et al., 1998; Polyakov et al., 2001). Moreover, spaceflight induces a pro-inflammatory response, which has also been shown to play an important role in down regulating body temperature (Luheshi et al., 1999; Crucian et al., 2014). Circadian phase, as assessed by the trough in body temperature, is delayed in astronauts during spaceflight in which they were only exposed to artificial light: the delay relative to ground-based measurements amounts to more than 2 h (Gundel et al., 1997). Spaceflight could generate circadian misalignment, which has been linked to adverse health consequences, such as impaired glucose regulation and metabolism, increased cardiovascular risk, and higher prevalence of cancers. This proposed circadian misalignment may also contribute to the use of medication (Flynn-Evans et al., 2016). One alternative countermeasure to facilitate circadian alignment is an appropriately timed exposure to both light and darkness: in fact, the circadian system is sensitive to even low levels of light (Basner et al., 2013). Specifically, the alteration of room lighting with artificial simulation of day and night may be sufficient to entrain circadian systems to a 24 h day during the preflight (Mrosovsky, 1988; Boivin et al., 1996). Aside from spaceflight, circadian disruption is also evident in terrestrial studies (Basner et al., 2013), including over-winter polar expeditions (Arendt, 2012).
Microgravity is also known to affect cardiovascular, pulmonary, neuro-vestibular, and musculoskeletal systems (Baker et al., 2008). Terrestrial partial gravity simulation models seek to simulate reduced gravity and the consequential impact it has on human physiology in relation to the proposed Lunar or Martian environment. The main problem remains that there is a lack of physiological data collected in ”real“ partial gravity scenarios and it is challenging to validate current partial gravity simulation models (Richter et al., 2017). However, in the literature to date, there are no indications that gravity directly influenced circadian rhythms and sleep in astronauts. However, in other species an effect of gravity on the circadian system is discussed (Ferraro et al., 1995). Despite this major limitation to identify space analogs suitable to directly inform extraterrestrial programs, the cave space analog has proven itself in review highly compatible to the current space environment, inclusive of EVA and isolation considerations.
In the cave environment there is usually a strong reduction in sunlight exposure requiring consequential adaptation to artificial light, as well as other external cues leading to augmentation of perceived vs. actual time. In addition, true isolation can be achieved, as communication can be hard to establish and prolonged confinement can be simulated under challenging conditions for days to weeks, or even months. Since 2011 ESA has organized an astronaut expeditionary training course in a space analog environment called CAVES (Cooperative Adventure for Valuing and Exercising human behavior and performance Skills) (Bessone et al., 2013; Strapazzon et al., 2014). CAVES is a scientific exploration expedition, taking place in an underground environment, to prepare international astronauts to become effective long-duration spaceflight crewmembers. Isolated cave environment allows the participants to develop autonomous operational skills in a multi-cultural team, while being challenged by an intense scientific and technological program in a surrounding that naturally recreates space-like stressors. In the 5 editions currently completed, 28 astronauts from 6 different space agencies have taken part in the training, plus more than 30 members of the scientific and organization staff stayed with them during the 7-day underground period.
A cave environment draws many parallels to current conditions on the International Space Station (ISS), inclusive the chance to replicate cues to simulate the 16 h day/night cycle, plus proposed environmental conditions for long duration interplanetary travel. Furthermore, artificial habitats will most probably be hypobaric, hyperoxic and hypercapnic in future, mainly due to the technical limitations of space travel (Norcross et al., 2013). Equally, the increase in demand for Extravehicular Activities (EVA), means increased time in the pressurized suits, which can cause thermal discomfort and are difficult to maneuver. In addition, analogous movement challenges include: the restrictive nature of cave passages, the lack of common reference points, necessity for three dimensional movement patterns, plus low visibility and perpetual darkness: all of which can reduce orientation capabilities, causing confusion and a potentially hostile perception of the environment, similar to that experienced during EVA maneuvers, or similar condition that could be found in lunar and extra-terrestrial cave (sites that could be chosen for exploration or for setting human habitat due to partial protection from cosmic radiation and stable environmental conditions compared to planetary surface). The scope of this review does not focus solely on the short effects' response, but also to prolonged effects of this atypical, subterranean environment and the potential for further development and understanding of one of the most challenging space analogs to date. This unique cave environment and the current space-agencies—led CAVES and PANGAEA (Planetary ANalogue Geological and Astrobiological Exercise for Astronauts) programs provide a unique opportunity to build upon subterranean experimental models employed previously and clearly identified here in review (Bessone et al., 2013; Strapazzon et al., 2014; Sauro et al., 2018). A more robust scientific approach, inclusive of increased numbers of trained personnel as participants could greatly improve not only the quality of research, but also the application of the results for practical considerations for future space missions. Specifically, piloting technological advances in medical monitoring equipment for real-time measurement of core temperature, cardiovascular health and energy expenditure, whilst completing specific subterranean routes and different operative conditions would help to inform estimations related to physiological strain induced under underground challenging conditions.
In review, structured cave research has previously lacked robust, reproducible, scientific data and is mainly limited to case studies. The reality of individual variation in response to physiological strain induced by challenging environmental conditions, even in a potentially homogeneous population such as cavers or astronauts, must be carefully considered when looking to inform practical recommendations for future space analog development. This current review identifies the need for further studies dedicated to investigating the cave environment as a space analog for exploratory missions. Precisely controlled, reproducible studies (both in the lab and cave environment) should be adopted in future, incorporating larger samples sizes, as well as a deliberate unification of protocol design, measurement validity, techniques employed, and standardized reporting. Equally, better specification and characterization of specific subterranean traverse routes and subterranean characteristics, in particular the characterization of particular obstacles, should be made to improve the level of scientific reproducibility and enable robust comparisons in terms of physiological exertion (i.e., TEE or O2) with alternate analogs and EVA specific tasks. A major limitation includes the pronounced methodological heterogeneity across those studies which met our selection criteria, plus the overall dearth of scientific papers which focused on this topic with sufficient clarity and thus prevented completion of a meta-analysis. It is clear that the reductive approach previously employed has led to highly fragmented understanding of current knowledge and observations gathered in the subterranean environment related to the physiological impact of a long-term exploratory cave isolation model. If these limitations were to be addressed, the cave space analog could provide the larger community an opportunity to collect novel data sets, which better inform a specific physiological strain index for subterranean activities and help delineate the physiological requirement to operate effectively in this unique environment, for the benefit of future exploratory programs.
Through the systematic review, we have identified the current status of knowledge, potential gaps and areas within sports and aerospace medicine literature pertaining to the impact of subterranean exploration on human physiology that would clearly benefit from additional research efforts. The aim in the future will be to better inform operational concerns for future training and deployment in cave analogs, as well as develop specific criteria for reproducible scientific data collection related to physiological monitoring in these complex environments. Despite growing expectation from different space agencies for advancements in data collection and collation of such data, the subterranean environment has consistently proven to be one of the most challenging. Consequently, the development of a specific physiological strain index and associated protocol development for explorative caving activities and prolonged permanence is yet to be fully defined. Since previous publications focusing on caving physiology are scarce, this complex space analog calls for more specific research to better determine the physiological basis of the cave traverse, in a variety of settings and for multiple purposes.
Topics highlighted within this review for further scientific investigation include: (i) cardiovascular considerations: cardiovascular response to both short and long duration cave exploration, with specific focus on the cardiac autonomic nervous system, heart rate variability, and potential augmentation of the sympatho-vagal balance; (ii) environmental cost: the metabolic impact of multiple challenging environmental factors (e.g., heat, humidity, hypoxia, hypercapnia), on subsequent estimations of TEE for a given activity and aspects of the potential performance deficit over long-duration exploratory scenarios; (iii) basic endocrinology: previous data on anticipatory and acute stress responses to cave exploration currently lacks translation to a prolonged model of cave isolation, mainly due to the logistical challenge of long duration isolation of large groups with relevant expertise for sufficient comparison; (iv) hydration and hematological values: previous work has identified potential hypo-osmolal dehydration in the case of prolonged cave traverse activity (Stenner et al., 2007b). Levels of hydration and subtle changes in plasma volume should be better determined when estimating both changes in body composition and hematological changes in muscle damage proteins and immunological markers, (v) desynchronization: reasonable inference exists that during specific subterranean isolation protocols there is a further requirement to closely assess self-reported rest activity cycles, renal output, menstruation and fluctuations in core body temperature, to better identify whether prolonged exposure to the cave environment, or specific ambient conditions, are responsible for further circadian desynchronization.
LB and GS developed the concept for this review. LB, GS, and LZ designed the study. Data collection and analysis were completed by LZ and LG. LZ, GS, LG, RT, EC, and LB interpreted the results. Creation of figures was completed by LZ and GS. LZ, GS, and LG wrote the article and the final draft was edited by RT, EC, and LB. All authors approved the final manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors acknowledge Victor De Maria for his contribution to this bibliographic research. We also show appreciation to Michel Siffre and Maurizio Montalbini for the outstanding dedication to the cave environment and to Michel Siffre for his help in retrieving material for this paper. The authors also acknowledge Nicolette B. Mogilever for her valuable help. The authors thank the Department of Innovation, Research and University of the Autonomous Province of Bozen/Bolzano for covering the Open Access publication costs.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphys.2019.00442/full#supplementary-material
ANS, Autonomic Nervous System; BP, Blood Pressure; BVA, Binocular Visual Acuity; CK, Serum Creatine Kinase; cpd, Cycles per degree; EE, Energy Expenditure; EEG, Electroencephalogram; EMG, Electromyography; ESA, European Space Agency; ESA CAVES, Cooperative Adventure for Valuing and Exercising human behavior and performance Skills; EVA, Extravehicular Activities; GH, Growth hormone; Hb, Hemoglobin; HCT, Hematocrit; HR, Heart Rate; HRV, Heart Rate Variability; LDH, Lactate Dehydrogenase; METs, Metabolic Equivalent of Task; NASA, National Aeronautics and Space Administration; NASA NEEMO, NASA Extreme Environment Mission Operations; PANGAEA, Planetary ANalogue Geological and Astrobiological Exercise for Astronauts; PRISMA, Preferred Reporting Items for Systematic Reviews and Meta-Analyses; PS, Paradoxical Sleep; PV, Plasma Volume; RBC, Red Blood Cell; TEE, Total Energy Expenditure.
Ainsworth, B. E., Haskell, W. L., Whitt, M. C., Irwin, M. L., Swartz, A. M., Strath, S. J., et al. (2000). Compendium of physical activities: an update of activity codes and MET intensities. Med. Sci. Sports Exerc. 32(9 Suppl.1), S498–S504. doi: 10.1097/00005768-200009001-00009
Anglin, K. M., and Kring, J. P. (2016). Lessons from a space analog on adaptation for long-duration exploration missions. Aerosp. Med. Hum. Perform. 87, 406–410. doi: 10.3357/AMHP.4489.2016
Antoni, G., Marini, E., Curreli, N., Tuveri, V., Comandini, O., Cabras, S., et al. (2017). Energy expenditure in caving. PLoS ONE 12:e0170853. doi: 10.1371/journal.pone.0170853
Apfelbaum, M., Reinberg, A., Nillus, P., and Halberg, F. (1969). Rythmes circadiens de l'alternance veille sommeil pendant l'isolement souterrain de sept jeunes femmes. Presse Méd. 77, 879–882.
Arendt, J. (2012). Biological rhythms during residence in polar regions. Chronobiol. Int. 29, 379–394. doi: 10.3109/07420528.2012.668997
Baker, E. S., Barratt, M. R., and Wear, M. L. (2008). “Human response to space flight,” in Principles of Clinical Medicine for Space Flights, eds M. R. Barratt and S. L. Pool (New York, NY: Springer), 27–57. doi: 10.1007/978-0-387-68164-1_2
Basner, M., Dinges, D. F., Mollicone, D., Ecker, A., Jones, C. W., Hyder, E. C., et al. (2013). Mars 520-d mission simulation reveals protracted crew hypokinesis and alternations of sleep duration and timing. Proc. Natl. Acad. Sci. U.S.A. 110, 2635–2640. doi: 10.1073/pnas.1212646110
Basner, M., Dinges, D. F., Mollicone, D. J., Savelev, I., Ecker, A. J., Di Antonio, A., et al. (2014). Psychological and behavioral changes during confinement in a 520-day simulated interplanetary mission to mars. PLoS ONE 9:e93298. doi: 10.1371/journal.pone.0093298
Bessone, L., Beblo-Vranesevic, K., Cossu, Q. A., De Waele, J., Leuko, S., Marcia, P., et al. (2013). “ESA CAVES: Training astronauts for space exploration,” in Proceedings of the 16th International Congress of Speleology. (Brno),321–317.
Boivin, D. B., Duffy, J. F., Kronauer, R. E., and Czeisler, C. A. (1996). Dose-response relationships for resetting of human circadian clock by light. Nature 379, 540–542. doi: 10.1038/379540a0
Bratima, F., Del Bello, G., and Stenner, E. (1999). Valutazione Funzionale di Atlete Praticanti la Speleologia. Grotte, Attività e Riflessioni Della Commissione E. Boegan ”PROGRESSIONE 40.“. Available online at: http://www.boegan.it/2009/11/valutazione-funzionale-di-atlete-praticanti-la-speleologia/
Bregani, E. R., Aliberti, S., and Guariglia, A. (2005). Creatine combined with branched-chain amino acids supplement in speleological practice: a scientific controlled trial. Med. Sport. 58, 233–239.
Chouvet, G., Mouret, J., Coindet, J., Siffre, M., and Jouvet, M. (1974). Periodicite bicircadienne du cycle veille-sommeil dans des conditions hors du temps. Etude polygraphique. Electroencephalogr. Clin. Neurophysiol. 37, 367–380. doi: 10.1016/0013-4694(74)90112-6
Colin, J., Timbal, J., Boutelier, C., Houdas, Y., and Siffre, M. (1968). Rhythm of the rectal temperature during a 6-month free-running experiment. J. Appl. Physiol. 25, 170–176. doi: 10.1152/jappl.1968.25.2.170
Cornelissen, G., Halberg, F., Siegelova, J., and Galvagno, A. (2010). “The moon's image in prolonged human isolation,” in Noninvasive Methods in Caridology, eds F. Halberg, T. Kenner, B. Fiser, and J. Siegelova (Brno: Masaryk University), 69–74.
Crucian, B. E., Zwart, S. R., Mehta, S., Uchakin, P., Quiriarte, H. D., Pierson, D., et al. (2014). Plasma cytokine concentrations indicate that in vivo hormonal regulation of immunity is altered during long-duration spaceflight. J. Interferon Cytokine Res. 34, 778–786. doi: 10.1089/jir.2013.0129
Czeisler, C. A., Duffy, J. F., Shanahan, T. L., Brown, E. N., Mitchell, J. F., Rimmer, D. W., et al. (1999). Stability, precision and near-24-hour period of the human circadian pacemaker. Science 284, 2177–2181. doi: 10.1126/science.284.5423.2177
Czeisler, C. A., and Gooley, J. J. (2007). Sleep and circadian rhythms in humans. Cold Spring Harb. Symp. Quant. Biol. 72, 579–597. doi: 10.1101/sqb.2007.72.064
De la Pena, S. S., Halberg, F., Galvagno, A., Montalbini, M., Follini, S., Wu, J., et al. (1989). “Circadian and circaseptan (about-7-day) free-running physiologic rhythms of a woman in social isolation,” in Second Annual IEEE Symposium Computer-Based Medical Systems (Minneapolis, MN), 273–278.
Dijk, D. J., Neri, D. F., Wyatt, J. K., Ronda, J. M., Riel, E., Ritz-De Cecco, A., et al. (2001). Sleep performance, circadian rhythms, and light-dark cycles during two space shuttle flights. Am. J. Physiol. Regul. Integr. Comp. Physiol. 281, R1647–R1164. doi: 10.1152/ajpregu.2001.281.5.R1647
Downs, M., Norcross, J. R., English, K., et al. (2018). “ISS crewmember aerobic capacity compared with metabolic cost of simulated exploration extravehicular activity (abstract),” in Proceedings of the AsMA Annual Scientific Meeting (Dallas, TX).
Elliott, L. R., White, M. P., Taylor, A. H., and Herbert, S. (2015). Energy expenditure on recreational visits to different natural environments. Soc. Sci. Med. 139, 53–60. doi: 10.1016/j.socscimed.2015.06.038
Felsing, N. E., Brasel, J. A., and Cooper, D. M. (1992). Effect of low and high intensity exercise on circulating growth hormone in men. J. Clin. Endocrinol. Metab. 75, 157–162. doi: 10.1210/jcem.75.1.1619005
Ferraro, J. S., Sulzman, F. M., and Dorsett, J. A. (1995). Alterations in growth rate associated with normally persisting circadian rhythm during spaceflight. Aviat. Space Environ. Med. 66, 1079–1085.
Flynn-Evans, E. E., Barger, L. K., Kubey, A. A., Sullivan, J. P., and Czeisler, C. A. (2016). Circadian misalignment affects sleep and medication use before and during spaceflight. NPJ Microgravity 2:15019. doi: 10.1038/npjmgrav.2015.19
Foing, B. (2016). “Towards a moon village: vision and opportunities,” in EGU General Assembly Conference (Vienna).
Fortney, S. M., Mikhaylov, V., Lee, S. M., Kobzev, Y, Gonzalez, R. R., and Greenleaf, J. E. (1998). Body temperature and thermoregulation during submaximal exercise after 115-day spaceflight. Aviat. Space Environ. Med. 69, 137–141.
Fraisse, P., Siffre, M., Oléron, G., and Zuili, N. (1968). “Le rythme veille-sommeil et l'estimation du temps,” in Cycles Biologiques et Psychiatrie, Simposium Bel-Air III (Paris: Masson and Cle), 257–265.
Fullagar, H. H., Skorski, S, Duffield, R., Hammes, D., Coutts, A. J., and Meyer, T. (2015). Sleep and athletic performance: the effects of sleep loss on exercise performance, and physiological and cognitive responses to exercise. Sports Med. 45, 161–186. doi: 10.1007/s40279-014-0260-0
Ghata, J., Halberg, F., Reinberg, A., and Siffre, M. (1969). Rythmes circadiens désynchronisés du cycle social (17-hydroxycorticostéroïdes, température rectale, veille-sommeil) chez deux sujets adultes sains. Ann. Endocrinol. 30, 245–260.
Giovine, G., Giovanni, B., De Vivo, A., Lomastro, F., Casagrande, G., Davila, A., et al. (2009). “The Naica caves and human physiology,” in Proceeding of the 15th International Congress of Speleology (Kerrville, TX), 1980–1984.
Godfrey, R. J., Madgwick, Z., and Whyte, G. P. (2003). The exercise-induced growth hormone response in athletes. Sports Med. 33, 599–613. doi: 10.2165/00007256-200333080-00005
Gundel, A., Polyakov, V. V., and Zulley, J. (1997). The alteration of human sleep and circadian rhythms during spaceflight. J. Sleep Res. 6, 1–8. doi: 10.1046/j.1365-2869.1997.00028.x
Halberg, F. (1968). “Physiologic considerations underlying rhythmometry, with special reference to emotional illness,” in Cycles Biologiques et Psychiatrie, Symposium Bel-Air III, 73–126.
Halberg, F., Reinberg, A., Haus, E., Ghata, J., and Siffre, M. (1970). Human biological rhythms during and after several months of isolation underground in natural caves. Nat. Speleol. Soc. Bull. 32, 89–115.
Halberg, F., Siffre, M., Engeli, M., Hillman, D., and Reinberg, A. (1965). Etude en libre-cours des rythmes circadiens du pouls de lalternance veille-sommeil et de lestimation du temps pendant les deux mois de sejour souterrain dun homme adulte jeune. C.R. Acad. Sci. 260:1259.
Hillman, D. C., Siffre, M., Milano, G., and Halberg, F. (1994a). Free-running psycho-physiologic circadians and three-month pattern in a woman isolated in a cave. New Trends Exp. Clin. Psychiatry. 10, 127–133.
Hillman, D. C., Siffre, M., Milano, G., and Halberg, F. (1994b). Urinary about-84-hour (circasemiseptan) variations of a woman isolated in a cave and cosmic ray effects. New Trends Exp. Clin. Psychiatry. 10, 173–178.
Hoyt, R. W., Jones, T. E., Baker-Fulco, C. J., Schoeller, D. A., Schoene, R. B., Schwartz, R. S., et al. (1994). Doubly labeled water measurement of human energy expenditure during exercise at high altitude. Am. J. Physiol. 266, R966–R971. doi: 10.1152/ajpregu.1994.266.3.R966
Johnston, R. S., Dietlein, L. F., and Berry, C. A. (1975). Biomedical Results of Apollo. For Sale by the Supt. of Docs. U.S. Govt. Print. Off . Washington, DC: Scientific and Technical Information Office, National Aeronautics and Space Administration, 592.
Koch, B., and Gerzer, R. (2008). A research facility for habitation questions to be built at the German Aerospace Center in Cologne: future challenges of Space medicine. Hippokratia. 12, 91–96.
Lança, C. C., Fernandes, N., Monteiro, A., and Almeida, A. (2016). Lighting conditions and optical filters effects on visual performance of speleologists exposed to cave environments. Int. J. Speleol. 45, 27–33. doi: 10.5038/1827-806X.45.1.1933
Liberati, A., Altman, D. G., Tetzlaff, J., Mulrow, C., Gøtzsche, P. C., Ioannidis, J. P., et al. (2009). The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate health care interventions: explanation and elaboration. BMJ 339:b2700. doi: 10.1136/bmj.b2700
Lorton, D., Lubahn, C. L., Estus, C., Millar, B. A., Carter, J. L., Wood, C. A., et al. (2006). Bidirectional communication between the brain and the immune system: implications for physiological sleep and disorders with disrupted sleep. Neuroimmunomodulation 13, 357–374. doi: 10.1159/000104864
Lugg, D. J. (2005). Behavioral health in Antarctica: implications for long-duration space missions. Aviat. Space Environ. Med. 76, B74–B77.
Luheshi, G. N., Gardner, J. D., Rushforth, D. A., Loudon, A. S., and Rothwell, N. J. (1999). Leptin actions on food intake and body temperature are mediated by IL-1. Proc. Natl. Acad. Sci. U.S.A. 96, 7047–7052. doi: 10.1073/pnas.96.12.7047
Maura, I. D. Y. I., Mora, J. E. I., Cornet, J. B., Rincón, J. A. G., Zabala, D. D., and Cardona, R.S. (2008). Adaptation to the rarefied air of abysses and caves. A laboratory study. Apunts Medicina de l' Esport (English Edition). 43, 135–140.
Meigal, A., and Fomina, E. (2016). Electromyographic evaluation of countermeasures during the terrestrial simulation of interplanetary spaceflight in Mars500 project. Pathophysiology 23, 11–18. doi: 10.1016/j.pathophys.2015.10.001
Miller, A. D., Taylor, B. J., and Johnson, B. D. (2013). Energy expenditure and intensity levels during a 6170-m summit in the Karakoram mountains. Wilderness Environ. Med. 24, 337–344. doi: 10.1016/j.wem.2013.06.009
Mills, J. N. (1964). Circadian rhythms during and after three months in solitude underground. J. Physiol. 174, 217–231. doi: 10.1113/jphysiol.1964.sp007483
Mills, J. N., Minors, D. S., and Waterhouse, J. M. (1974). The circadian rhythms of human subjects without timepieces or indication of the alternation of day and night. J. Physiol. 240, 567–594. doi: 10.1113/jphysiol.1974.sp010623
Mogilever, N., Zuccarelli, L., Burles, F., Iaria, G., Strapazzon, G., Bessone, L., and Coffey, E. B. (2018). Expedition cognition: a review and prospective of subterranean neuroscience with spaceflight applications. Front. Hum. Neurosci. 12:407. doi: 10.3389/fnhum.2018.00407
Monk, T. H., Buysse, D. J., Billy, B. D., Kennedy, K. S., and Willrich, L. M. (1998). Sleep and circadian rhythms in four orbiting astronauts. J. Biol. Rhythms 13, 188–201. doi: 10.1177/074873098129000039
Mrosovsky, N. (1988). Phase response curves for social entrainment. J. Comp. Phys. A 162, 35–46. doi: 10.1007/BF01342701
Norcross, J., Norsk, P., Law, J., Arias, D., Conkin, J., Perchonok, M., et al. (2013). Effects of the 8 psia/32% O2 Atmosphere on the Human in the Spaceflight Environment. Hanover, MD: National Aeronautics and Space Administration Center for AeroSpace Information.
Oleron, G., Fraisse, P., Siffre, M., and Zuili, N. (1970). Les variations circadiennes du temps de reaction et du tempo spontane au cours dune experience ≪hors du temps≫. Annee Psychol. 70, 347–356. doi: 10.3406/psy.1970.27900
Pagel, J. I., and Choukér, A. (2016). Effects of isolation and confinement on humans-implications for manned space explorations. J. Appl. Physiol. 120, 1449–1457. doi: 10.1152/japplphysiol.00928.2015
Pinna, V., Magnani, S., Sainas, G., Ghiani, G., Vanni, S., Olla, S., et al. (2017). Physical capacity and energy expenditure of cavers. Front. Physiol. 8:1067. doi: 10.3389/fphys.2017.01067
Pittendrigh, C. S. (1960). Circadian rhythms and the circadian organization of living systems. Cold Spring Harb. Symp. Quant. Biol. 25, 159–184. doi: 10.1101/SQB.1960.025.01.015
Polyakov, V. V., Lacota, N. G., and Gundel, A. (2001). Human thermohomeostasis onboard “Mir” and in simulated microgravity studies. Acta Astonaut. 49, 137–143. doi: 10.1016/S0094-5765(01)00091-1
Princi, T., Stenner, E., Biasioli, B., Peterec, D., and Accardo, A. (2008). “The heart rate variability in a female élite potholer at rest and during the cave exploration: a case report,” in 5th European Symposium on Biomedical Engineering (ESMBE 2008) (Chania).
Pulfrey, S. M., and Jones, P. J. (1996). Energy expenditure and requirement while climbing above 6,000 m. J. Appl. Physiol. 81, 1306–1311. doi: 10.1152/jappl.1996.81.3.1306
Reinberg, A., Halberg, F., Ghata, J., and Siffre, M. (1966). Spectre thermique (rythmes de la temperature rectale) d'une femme adulte avant, pendant et apres son isolement souterrain de trois mois. C.R. Acad. Sci. 262, 782–785.
Reitz, C. J., and Martino, T. A. (2015). Disruption of circadian rhythms and sleep on critical illness and the impact on cardiovascular events. Curr. Pharm. Des. 21, 3505–3511. doi: 10.2174/1381612821666150706105926
Reynolds, R. D., Lickteig, J. A., Deuster, P. A., Howard, M. P., Conway, J. M., Pietersma, A., et al. (1999). Energy metabolism increases and regional body fat decreases while regional muscle mass is spared in humans climbing Mt. Everest. J. Nutr. 129, 1307–1314. doi: 10.1093/jn/129.7.1307
Richter, C., Braunstein, B., Winnard, A., Nasser, M., and Weber, T. (2017). Human biomechanical and cardiopulmonary responses to partial gravity- a systematic review. Front. Physiol. 8:583. doi: 10.3389/fphys.2017.00583
Rittweger, J., Debevec, T., Frings-Meuthen, P., Lau, P., Mittag, U., Ganse, B., et al. (2016). On the combined effects of normobaric hypoxia and bed rest upon bone and mineral metabolism: results from the PlanHab study. Bone 91, 130–138. doi: 10.1016/j.bone.2016.07.013
Robinson, J. A., Thumm, T. L., and Thomas, D. A. (2007). NASA utilization of the international space station and the vision for space exploration. Acta Astronaut. 61, 176–184. doi: 10.1016/j.actaastro.2007.01.019
Salvadego, D., Keramidas, M. E., Kölegård, R., Brocca, L., Lazzer, S., Mavelli, I., et al. (2018). PlanHab*: hypoxia does not worsen the impairment of skeletal muscle oxidative function induced by bed rest alone. J. Physiol. 596, 3341–3355. doi: 10.1113/JP275605
Saugy, M., Robinson, N., Saudan, C., Baume, N., Avois, L., and Mangin, P. (2006). Human growth hormone doping in sport. Br. J. Sports Med. 40(Suppl. 1), i35–i39. doi: 10.1136/bjsm.2006.027573
Sauro, F., Massironi, M., Pozzobon, R., Hiesinger, H., Mangold, N., Frias, J. M., et al. (2018). “Training astronauts for field geology: The ESA PANGAEA Training and PANGAEA-extension testing analogue,” in Lunar and Planetary Science Conference 49 (The Woodlands, TX).
Siffre, M. (1988). “Ryhtmes biologiques, sommeil et vigilance en confinement prolonge,” in ESA, Proceedings of the Colloquium on Space and Sea (SEE N 88-26016 19-51) (Marseille), 53–68.
Siffre, M. (1990). “Some problems of desynchronisation of sleep-wakefulness and circadian rhythm s for long duration spaceflights,” in ESA, Proceedings of the Space e Sea Colloquium (Paris).
Siffre, M., Reinberg, A., Halberg, F., Ghata, J., Perdriel, G., and Slind, R. (1966). L'isolement souterrain prolongé. Étude de deux sujets adultes sains avant, pendant et apres cet isolement. Presse Méd. 74, 915–919.
Sonnenfeld, G., Measel, J., Loken, M. R., Degioanni, J., Follini, S., Galvagno, A., et al. (1992). Effects of isolation on interferon production and hematological and immunological parameters. J. Interferon. Res. 12, 75–81. doi: 10.1089/jir.1992.12.75
Stenner, E. (2002). Valori Della Glicemia Rilevati Durante le Esplorazioni Dell'abisso Laricetto. Grotte, Attività e Riflessioni Della Commissione E. Boegan ”PROGRESSIONE 46“. Available online at: http://www.boegan.it/2009/11/glicemia-negli-speleologi/
Stenner, E., Gianoli, E., Biasioli, B., Piccinini, C., Delbello, G., and Bussani, A (2006). Muscular damage and intravascular haemolysis during an 18 hour subterranean exploration in a cave of 700 m depth. Br. J. Sports Med. 40, 235–238. doi: 10.1136/bjsm.2005.021402
Stenner, E., Gianoli, E., Piccinini, C., Biasioli, B., Bussani, A., and Delbello, G. (2007a). Hormonal responses to a long duration exploration in a cave of 700 m depth. Eur. J. Appl. Physiol. 100, 71–78. doi: 10.1007/s00421-007-0408-9
Stenner, E., Piccinini, C., Biasioli, B., Gianoli, E., Bussani, A., and Delbello, G. (2007b). Red cell and core temperature in spelunking. Int. J. Sports Med. 28, 727–231. doi: 10.1055/s-2007-964978
Strapazzon, G., Pilo, L., Bessone, L., and Barratt, M. R. (2014). CAVES as an environment for astronaut training. Wilderness Environ. Med. 225, 244–245. doi: 10.1016/j.wem.2013.12.003
Sutterfield, S. L., Alexander, A. M., Hammer, S. M., Didier, K. D., Caldwell, J. T., Barstow, T. J., et al. (2019). Prediction of planetary mission task performance for long-duration spaceflight. Med. Sci. Sports Exerc. doi: 10.1249/MSS.0000000000001980. [Epub ahead of print].
Vacca, U., Tuveri, V., and Irsara, S. (1994). Analisi Della Composizione Corporea di Individui Impegnati in Attività Speleologica. Grotte, Attività e Riflessioni Della Commissione E. Boegan ”PROGRESSIONE 30“. Available online at: http://www.boegan.it/1993/01/relazione-medica-tepuy-93/
Videbaek, H. R., Johansen, L. B., and Norsk, P. (1993). Human Anatomy and Physiology for the European Astronaut Team. Copenhagen: Damec Research Inc.
Keywords: space analog, spaceflight, extreme environments, circadian rhythms, human physiology, cave
Citation: Zuccarelli L, Galasso L, Turner R, Coffey EJB, Bessone L and Strapazzon G (2019) Human Physiology During Exposure to the Cave Environment: A Systematic Review With Implications for Aerospace Medicine. Front. Physiol. 10:442. doi: 10.3389/fphys.2019.00442
Received: 14 December 2018; Accepted: 01 April 2019;
Published: 24 April 2019.
Edited by:
Jörn Rittweger, German Aerospace Center, GermanyReviewed by:
Ana Diaz Artiles, Texas A&M University, United StatesCopyright © 2019 Zuccarelli, Galasso, Turner, Coffey, Bessone and Strapazzon. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Giacomo Strapazzon, Z2lhY29tby5zdHJhcGF6em9uQGV1cmFjLmVkdQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.