
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
PERSPECTIVE article
Front. Phys. , 28 February 2020
Sec. Interdisciplinary Physics
Volume 8 - 2020 | https://doi.org/10.3389/fphy.2020.00031
This article is part of the Research Topic Complexity and Self-Organization View all 11 articles
 David Pastor-Escuredo1,2,3*
David Pastor-Escuredo1,2,3* Juan C. del Álamo2,3,4,5,6*
Juan C. del Álamo2,3,4,5,6*The growing availability of imaging data, calculation power, and algorithm sophistication are transforming the study of morphogenesis into a computation-driven discipline. In parallel, it is accepted that mechanics plays a role in many of the processes determining the cell fate map, providing further opportunities for modeling and simulation. We provide a perspective of this integrative field, discussing recent advances and outstanding challenges to understand the determination of the fate map. At the basis, high-resolution microscopy and image processing provide digital representations of embryos that facilitate quantifying their mechanics with computational methods. Moreover, innovations in in-vivo sensing and tissue manipulation can now characterize cell-scale processes to feed larger-scale representations. A variety of mechanical formalisms have been proposed to model cellular biophysics and its links with biochemical and genetic factors. However, there are still limitations derived from the dynamic nature of embryonic tissue and its spatio-temporal heterogeneity. Also, the increasing complexity and variety of implementations make it difficult to harmonize and cross-validate models. The solution to these challenges will likely require integrating novel in vivo measurements of embryonic biomechanics into the models. Machine Learning has great potential to classify spatio-temporally connected groups of cells with similar dynamics. Emerging Deep Learning architectures facilitate the discovery of causal links and are becoming transparent and interpretable. We anticipate these new tools will lead to multi-scale models with the necessary accuracy and flexibility to formulate hypotheses for in-vivo and in-silico testing. These methods have promising applications for tissue engineering, identification of therapeutic targets, and synthetic life.
Embryogenesis is a complicated ensemble of processes by which a single cell turns into a multi-cellular living organism. Through various developmental stages, the cell population proliferates while tissues develop, change their properties, differentiate, and gain their specific functionality [1]. During embryogenesis, biochemical, genetic, and epigenetic factors interact, forming a tangled network of processes with diverse physical length scales and time scales [2, 3]. Remarkably, the robustness and variability of these processes are balanced to make possible the reproducibility and diversity of living specimens [4].
Mechanics plays a central role in shaping the embryo [5, 6]. Gene expression gradients regulate tissue patterning and cellular properties, such as rheology, adhesion, and contractility [7]. At the same time, the embryonic cells sense mechanical cues from their microenvironment and convert them to biochemical signals, including gene expression [8, 9]. These cues are essential to guide morphogenesis but also tissue repair, given that immature cells can retain significant plasticity and reprogram in response to external forces [10, 11]. The cross-talk between biophysical and biochemical processes involves multiple mechanisms and molecules and occurs in multiple scales [12]. Besides, cells can follow complex trajectories within the developing embryo, thereby creating and being exposed to continuous changes in the microenvironment [13–16].
Researchers have been long interested in discovering mechanistic links between physical processes and gene expression that lead to cell fate determination [17–21]. Recent advances in microscopy, modeling, and computation have enabled quantifying 2D and 3D mechanical forces and rheological properties in multi-cellular colonies, including live developing embryos [22–28]. These methods provide local data in space and time, and analyzing them to unravel cell fate maps is challenging. High-resolution, long-term observation in two or three dimensions is desirable to consider the whole range of scales at which mechanics can impact cell fate. Still, it complicates the analysis further because it involves massive amounts of data. Furthermore, the statistical treatment of the data needs to accommodate the highly heterogeneous and time-evolving properties of developing tissues [29–31].
This perspective discusses current advances in computational methods for the characterization of mechanical processes during embryogenesis and how these processes influence cell fate. Sections Digital Reconstruction of Embryogenesis, In vivo Quantification of Forces and Mechanical Properties, and Computational Models in this perspective are organized according to key steps in the analysis of experimental data and relevant methodological approaches. Each section presents our view into key advances and outstanding challenges. Section Morphomechanical Domains in Developing Tissues: Follow the Cell, Not the Voxel proposes a paradigm to deal with the massive data produced by experimental techniques and construct a multi-scale representation of embryo dynamics. Finally, section Understanding Multi-Scale Embryonic Dynamics by Machine Learning presents problems at the intersection between morphogenesis and Machine Learning that has not been so far tackled by the community.
Progress in live microscopy and fluorescence reporters now allow high-resolution, time-lapse imaging of developing embryos in two and three dimensions [32–35]. Image analysis and computer vision methods can now create digital atlases of developing embryos (Figures 1A–F). These atlases contain spatio-temporal information about cell and tissue morphology, cell lineages, and functional patterns, such as gene expression or protein activity [36–42]. Moreover, novel visualization tools allow for systematically browsing these digital embryos (Figure 1), and integrating them into numerical simulations and machine learning algorithms [36, 42, 43].
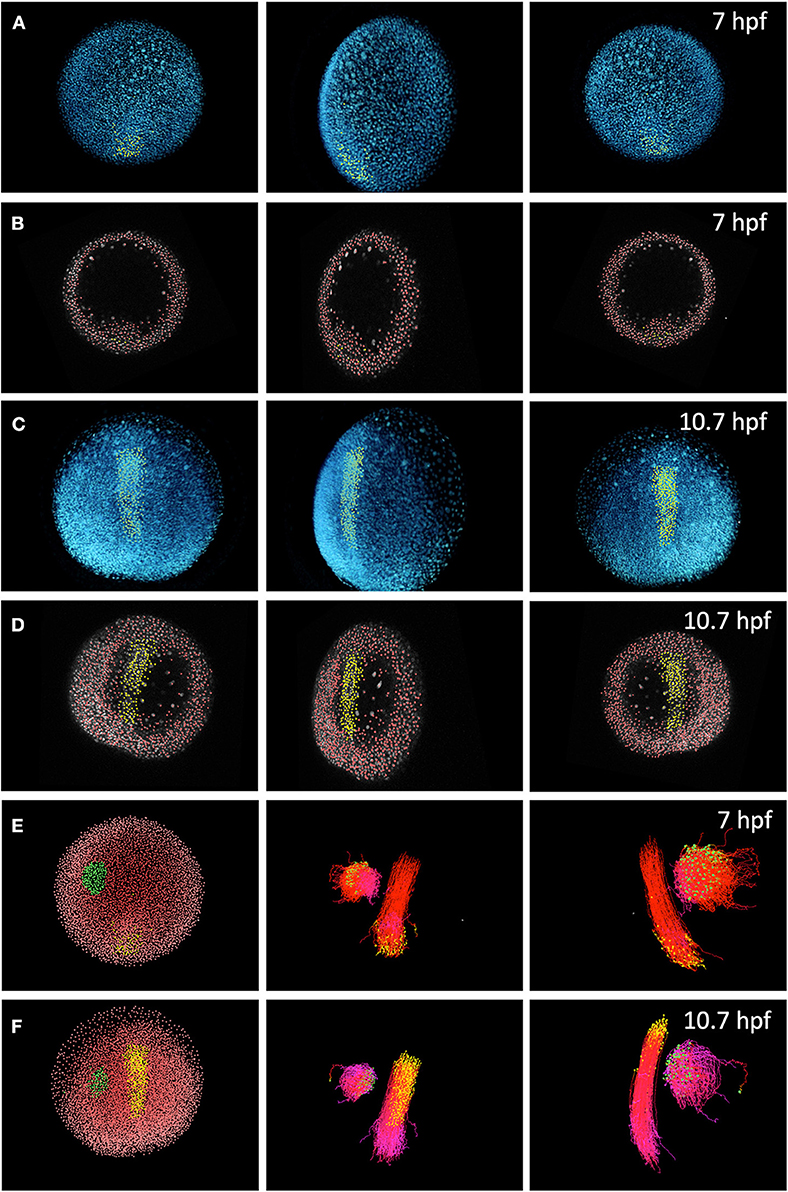
Figure 1. Digital reconstruction of cell trajectories in a zebrafish embryo. (A) Three-dimensional (3D) rendering of cell nuclei (blue) in raw data and selection of cells (yellow) in the dorsal line, performed at 10.7 h-post-fertilization and backtracked to 7 hpf. From left to right, each panel shows a different spatial orientation (animal pole, lateral and ventral). (B) Detected cell nuclei (red) and cell selection as in (A) (yellow), shown in a spatial slice over the orthoslice of the raw data channel (gray). Same time step and view angles as in (A). (C) 3D rendering of cell nuclei (blue) and selection of cells of the dorsal line at 10.7 hpf (yellow). (D) Cell detection (red) and cell selection (yellow) in the same slice as (B). (E) Left: Two cell selections (green and yellow) over nuclei detection domains at 7 hpf. Middle: forward tracking (yellow to red colormap indicates time advancement) of the two selected cell domains. Right: forward tracking of the two cell selections from a lateral point of view. (F) Cell selections (green and yellow) at 10.7 hpf. Cell backward tracking (yellow to purple colormap) in same view angles than (E). This dataset and the visualization tool Mov-IT are freely available from the BioEmergences open workflow http://bioemergences.iscpif.fr/bioemergences/openworkflow-index.php [36].
The three-dimensional in-vivo imaging of whole embryos has challenges associated with image resolution, quality, and artifacts (e.g., anisotropic point spread function). Besides, photobleaching and phototoxicity make it challenging to extend imaging over intervals long enough to capture relevant morphogenetic processes. Multi-view light-sheet microscopy (LSM) [35, 44, 45] and view fusion algorithms [46, 47] allow for 3D imaging large embryos with cellular isotropic resolution. Recently, advances proposing adaptive optics and lattice LSM with ultrathin light-sheet excitation featured, promising sub-cellular resolution during long-term observation [48].
Reconstructing the multi-scale dynamics of embryogenesis requires not only long-term imaging with sub-cellular spatial resolution but also sub-minute temporal resolution. An established approach to achieve these joint demands is to record images of several embryos within the same cohort with different temporal resolutions and to register the resulting images spatially onto a common template [49, 50]. The projected growth in computing power of microscopy systems (e.g., by embedded GPU computing) makes it possible to envision enhanced microscopes with real-time adaptive multi-scale observation [51, 52].
Image processing workflows must be able to handle the massive amounts of complex data resulting from microscopy modalities to provide a quantification of structures, motion, and hierarchy [3, 53]. Intensity-based methods, such as optical flow or image registration produce continuous velocity fields [53–55] that can leverage the powerful modeling and descriptive tools of continuum and statistical mechanics [56, 57]. On the other hand, tracking the motion and divisions of single cells yields discrete cell lineages, which presents apparent advantages [4, 36, 40].
Motion estimation is critical because determining cell fate involves reconstructing 3D cell trajectories across the various developmental stages, imposing quasi-error-free requirements (Figures 1E,F). Deep Learning tools, such as Convolutional Neural Networks can help to improve the performance under challenging conditions, such as deep-tissue segmentation provided tagged training data [58, 59]. Interactive annotation tools for correction and validation are still a suitable approach to generate reliable expert-driven data [36, 42, 43, 60] and potentially allow crowdsourced results [61]. Beyond image data repositories, sharing detailed experiment metadata through systematic frameworks (e.g., based on ontologies) can provide a “Big Data” substrate for machine learning to optimize pipelines.
Digital reconstruction of morphogenesis already provides structured data, but embedding biophysical formalisms is invaluable to decipher multi-scale dynamics. The study of biophysics in single cells is not new: the measurement of the rheological properties of cells, their internal stresses and the forces they exert has received significant attention in the past two decades [12, 23, 62]. The requirement of non-invasiveness, three-dimensionality, and the need for calibrated sensors that sensitive enough to resolve minute forces and deformations make this task particularly challenging in live embryos.
Laser ablation was one of the pioneering methods to quantify embryonic mechanics in vivo. This technique produces a localized cut in a tissue, which allows for estimating tissue tensions by letting the ablated region relax to a stress-free configuration [63–65]. This technique is still widely used but it is disruptive. A non-invasive alternative is to use fluorescence reporters to measure acto-myosin activity as a surrogate metric of force generation. Still, both methods rely on independent measurements of tissue rheology [66, 67]. Molecular sensors based on fluorescence resonance energy transfer (FRET) also provide a minimally invasive means of measuring forces in vivo [68]. This modality is very attractive since it probes the tension born by specific molecules. However, it requires careful calibration, does not provide vector or tensor data, and needs a different sensor to measure the tension born by each molecule. It is undoubtable that these approaches will continue to shed light on numerous embryogenic processes. Even so, their critical examination has kindled the search for easy-to-calibrate quantifications of the strains, stresses and material properties inside live tissues.
Because in vitro assays allow for careful control of experimental parameters, they have experienced significant progress in the past 20 years, thus offering valuable lessons for the development of in vivo techniques. In particular, there is a diversity of force microscopy methods that exploit the linear properties and high deformability of hydrogelsto provide sensitive, calibrated strain-stress sensors. Cells are cultured on these hydrogels, the deformation caused by the cells on the hydrogel is measured, typically by tracking the motion of fiduciary markers (e.g., fluorescent microspheres), and the traction forces exerted by the cells are recovered from the measured deformations [69–71]. Monolayer Stress Microscopy is an extension of traction microscopy that quantifies the collective distribution of intracellular stress in thin confluent cell cultures [72]. A similar approach was proposed to estimate ventral furrow invagination in Drosophila although in that case the stress-free configuration was not known [73]. Of note, traction forces can be highly three-dimensional even when the cells are plated on flat hydrogels [74], leading to significant bending and additional intracellular stress in cell monolayers [75]. Quantifying the forces involved in epithelial bending and invagination could offer new biomechanical insights about the morphogenesis of tissues and organs.
In live developing embryos, it is now feasible to measure strains (and strain rates, Figure 2) at the cellular level by tracking the morphological changes of segmented cells [55, 76]. Tissue-level strain fields can be derived from cell tracking and optical flow methods (Figures 2A–C) [57, 77]. By combining the cell-level and tissue-level strain quantifications it is possible to infer tissue rearrangements, such as cell deformation and cell intercalation [55, 60, 77, 78]. These metrics can be overlaid with functional data, such as gene expression and acto-myosin activity, to provide a correlation-based understanding of tissue dynamics [53, 77, 79]. Moreover, continuum strain fields enable the quantification of internal stresses based on a prescribed mechanical model for the embryo. These formulations are very advantageous—they allow for writing sets of equations that can be solved analytically or numerically to fully characterize the mechanical state of the tissue [80]. A mechanical formalism that has been applied to developing embryos with notable success relies on enforcing static equilibrium of forces between intracellular pressure and cortical tension. This formulation leads to a geometrical problem for cell shapes that can be closed by analyzing experimental images [81–86]. However, it must be recalled that embryonic tissue properties are heterogeneous, highly non-linear and time-evolving, which makes it challenging to develop mechanical formalisms that are uniformly valid across different regions of space, instants of time, and genetic and pharmacological manipulations. Furthermore, a significant challenge is to establish the stress-free reference state to properly quantify visco-elastic forces.
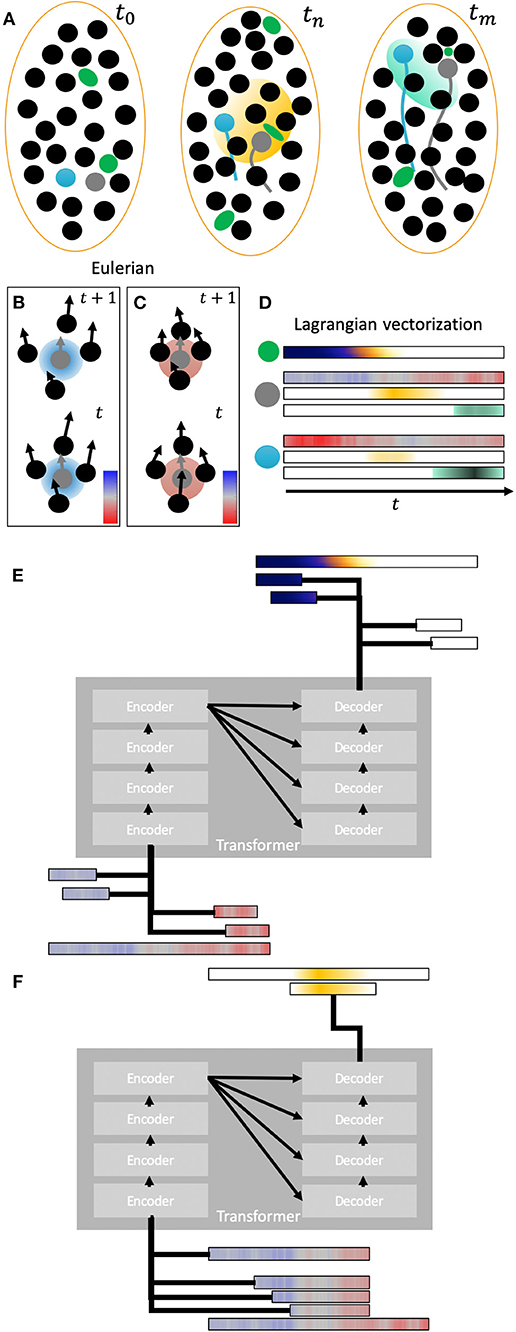
Figure 2. Lagrangian representation of kinematics and genetic expression. (A) Three schematic snapshots of different time intervals of migrating cell nuclei within an area. The trajectories of two cells (gray and blue) are highlighted. In the second time step a gene expression pattern in yellow is shown affecting several cells. In the last time step another different gene expression pattern is represented in cyan affecting another set of cells. Green blobs represent mechanical sensors that sense local deformation. (B) Two snapshots showing the relative displacements of neighbor cells with respect of a reference cell (gray). These relative displacements are translated into a kinematic descriptor of relative area change rate that representation expansion (blue) and compression (red). The relative displacements in this schematic panel imply a local expansion (blue value) around the reference cell between timesteps t and t + 1 as shown by the average increasing distance between the cell nuclei. (C) same schematic than in (B) for a compressive case with cells getting closer to the reference cell (red value). (D) Lagrangian vectorization of compression/expansion descriptor [same colormap than (B,C)] and gene expression along time for the two reference cells. The data sensed with the mechanical probe is also vectorized in a Lagrangian representation with colormap dark blue to white. Gene expression is vectorized along the reference cell trajectories. (E) Schematic of a transformer (encoder-decoder) architecture trained to infer local forces from deformation measurements (input). The input is segmented into different temporal frames as subvectors. (F) Schema of a transformer architecture trained to infer mechanical factors (input) involved in the appearance of expression patterns at different temporal scales.
A recent approach for the in-vivo characterization of embryonic mechanics, without prior assumptions, consists of injecting microdroplets or hydrogel microspheres of size comparable to one cell, and that can act as calibrated sensors and/or actuators (Figure 2) [87]. After appropriate functionalization by surface coating, these sensors can be made biocompatible and are internalized by the embryo, thereby minimizing the invasiveness of the method. Incompressible fluorescent oil-droplets allow for quantifying anisotropic stresses [88], whereas hydrogel droplets with characterized compressibility allow for quantifying isotropic ones [89]. Moreover, ferrofluid droplets can be act as active sensors to measure the local tissue rheology [90]. An additional feature of these sensors is that they move with their neighboring cells during development, thus providing valuable information about the temporal evolution of mechanical stresses and tissue rheology. Their limitations stem from reduced sampling ability, given by the limited number of sensors that can be used per embryo, and the current lack of scalable computational frameworks to relate the measurements with cell fate determination. Even so, it is reasonable to expect that emerging innovations will simplify the implementation of these techniques, enabling their widespread application.
Computational models with explanatory and predictive power can infer causal links and contribute to the mechanistic understanding of embryogenesis. These models allow researchers to observe processes, reverse engineer mechanisms, and test hypotheses with much looser constraints than pharmacological or genetic manipulations. Many biological problems involving collective cell-cell and cell-matrix interactions have been simulated using discrete, continuum, and hybrid physical models [91, 92]. Discrete agent-based models initially considered cellular movements within a lattice to investigate multicellular interactions [93]. Lattice-free agent-based models consider continuous movements of each agent. A common approach is to conceptualize cells as objects with fixed geometry and biophysical properties, whose trajectories are dictated by the balance of forces exerted by their neighbors and the environment [94]. Subcellular resolution can be achieved through agent-based models in which each agent is deformable and occupies several nodes [95]. The cellular Potts model (CPM) is an energy-based stochastic model, typically defined on a lattice that can have subcellular resolution, that is particularly well-suited to deal with large deformations and multi-scale phenomena [96]. These features make the CPM well-suited to simulate collective cell dynamics in a diversity of scenarios, including morphogenesis [97]. While they are mostly phenomenological, these models are a promising, computationally efficient approach to study how meso-scale multicellular phenomena emerge from the self-organization of sub-cellular and cellular processes.
The cellular Potts model was initially applied to quantify epithelial dynamics including the rearrangements of different cells [98]. Subsequently, the CPM has provided insight about how cortical tension and cell adhesions drive cell sorting and tissue organization [99, 100]. More recently, agent-based models have proven useful to integrate mechanical cues with gene expression. Epithelial and mesenchymal tectonics were simulated together with gene regulatory network dynamics to recapitulate the dynamics of early zebrafish development [26]. Deformable agent-based models are a promising approach to quantify mechanotransduction, the heterogeneity of embryonic tissues, and their impact in larger-scale developmental processes [92, 101].
Vertex models bridge the discrete and continuum descriptions [102]. In these models, each cell is approximated by a polygon in 2D or a polyhedron in 3D, and the tissue measurements are sampled at the junction of three or more cells [102, 103]. Vertex models provide more information on cell interfaces than agent-based models permitting the analysis of topological changes in the cell environment [104, 105]. Curved cell geometries can be resolved with finite-elements [106, 107], and the biophysical interaction between the membrane and the cytoplasmic fluid can also be incorporated using immersed boundary methods [108]. Vertex models have been widely applied to study the mechanics of epithelia, which are represented as manifolds that can fold or invaginate [109–114]. These models have made contributions to our understanding various tissue behaviors: growth [115–117], cell division and packing [118], planar polarity [119] and the formation of compartments [120]. Dynamic cellular finite-element models have been also proposed for individual and collective cell movements and mechanics [121].
As stated above, continuum models can adapt mechanical theories, such as hydrodynamics and statistical mechanics to live matter [122], taking advantage of a massive body of knowledge and powerful tools from applied mathematics and computation, such as stability theory, perturbation methods, and computational fluid dynamics. In addition to providing a means to relate measurements of strain fields to internal stresses [78, 123], these models are well-suited to perform predictive simulations large-scale embryo dynamics. The widely studied formation of the ventral furrow in Drosophila [124] is a good example of a process governed by hydrodynamics [56]. Most continuum models are limited by their inherent coarse-grained, but fusion between these models and agent-based models could help resolve the contribution individual cells to tissue behavior [125].
Although microscopy experiments provide increasingly rich data about embryonic development, the data is obtained in a form that makes it difficult to extract the relationships between cellular and subcellular dynamics, large-scale biomechanical phenomena, and cell fate maps. The root for this difficulty can be illustrated using the analogy between the cell trajectories and a flow; observation through the microscopy imposes a perspective in a fixed reference frame as an external observer of embryogenesis (i.e., Eulerian frame). However, a perspective as an internal observer that records data along the trajectory of each cell would be more suitable (i.e., Lagrangian frame). The Lagrangian framework allows for computing deformation rates and finite deformations over arbitrarily long time intervals [57]. It also helps discover Lagrangian coherent structures [126, 127] formed by cells that experience similar histories of mechanical cues, and which potentially organize the embryogenic flow (Figure 2D).
The Lagrangian trajectories of embryonic cells can be obtained by single-cell tracking or by approximating their motion as a continuous flow [14, 15, 53]. Moreover, in the Lagrangian framework, descriptors related to morphology, mechanics, genetics, etc. can be expressed in terms of the cell trajectories at specific time intervals. The usefulness of this approach depends on whether it can identify true morphomechanical fields. That is, if it finds connected domains of cells with a similar history of cues, if these domains are reproducible across several specimens, and if they can be related to the fate maps. We previously showed that machine learning does identify morphomechanical fields by classifying cell populations with similar Lagrangian cues either via clustering or with training data [57]. Comparison of cohorts can be either performed using a canonical embryo as reference or computing a statistical average of morphomechanical fields. This is a different approach from statistical spatial atlases frequently used to align information within a cohort [49, 50]. However, several fundamental questions and methodological obstacles remain unanswered. In particular, the sensitivity of the automatic classification of morphomechanical fields to intra-phenotypic variability, and its usefulness in establishing inter-phenotype differences need to be addressed in more detail. In particular, automating these analyses for cohorts of embryos requires systematic scanning across entire embryos to compensate for the different development rates of each embryo and its phenotype variability. Then, through the spatio-temporal registration of fields [54], it could be possible to infer robust phenotyping structures and assess the impact of dynamics variability into morphological configuration of tissues and organs.
Biological systems are often defined as networks of discrete elements or biochemical processes, which serve as a conceptual framework to glean mechanistic insight about their organization [128, 129]. Framing embryogenesis using this paradigm involves identifying morphogenetic events and fields [130], which can be diverse in nature, duration, and length-scale. Based on image data, one can define morphogenetic events as spatio-temporal spots of statistically abnormal behavior given a reference window. They may comprise subcellular or mesoscopic regions and a variable number of time frames and can be encapsulated by applying spatio-temporal connectivity [131]. When these fields are backtracked, they become unwound in time and space, allowing the discovery of intersections with past events and/or environmental cues. Likewise, forward tracking of events can reveal cascade effects that propagate into one or more morphogenetic fields. The structured representation of digital embryos as spatio-temporally connected fields is a form of dimensionality reduction that fits machine learning-driven approaches.
Owing to recent advances in machine learning methods, computers can now perform human-like reasoning in tasks, such as conversation or gaming [132–134]. Deep learning (DL) architectures, such as Feed-Forward Networks, Recurrent Neural Networks (RNN) and Convolutional Neural Networks (CNN) can be applied to analyze interactions in the networks of elements employed in systems biology [135]. Consequently, the applications of DL to biomedicine are quickly becoming ubiquitous [136–139]. The analysis of high-throughput genomics data to study genetic variations in regulatory networks is no exception [140, 141]. A main barrier toward adopting DL in developmental biology has been its black-box nature, which does not easily permit inferring mechanisms or causal relationships, and makes it challenging to manipulate models to test hypotheses. Most ongoing efforts to mitigate these limitations can be encompassed under the epistemological paradigm of the Visible Neural Network [142]. One approach toward VNN is to represent the nodes in the network as “visible” data-driven models. This approach has been used to relate cell genotypes and phenotypes based on cell ontologies [143]. An alternate approach is to build the nodes in the network using explicit models based on theoretical or semi-empirical laws [144]. Both approaches allow for manipulating the inner machinery of the DL architecture, thereby facilitating hypothesis testing, the inference of causal relationships, and elucidating mechanisms. Furthermore, coupling DL model-driven architectures with multi-level structured training data can help reduce the amount of inputs, simplify the architecture and facilitate its interpretation [145]. Exhaustive simulations running on cloud technologies [146, 147] can leverage computational models and feed machine learning workflows to create multiple hypothesis to be tested in-vivo. In the case of embryo development, most theoretical and computational models are coarse grained and, thus, better suited to represent meso-scale and large-scale phenomena (see section Computational Models). Consequently, it could be beneficial to develop hybrid approaches in which cell-scale phenomena are modeled with DL. This type of bottom-up methodology has shown great potential to improve the prediction of chaotic deterministic systems, such as turbulent flow [148], but it should be noted that, epistemologically, it constitutes a transparent network of opaque nodes. Given that multiple relationships among genetic and biophysical processes evolve dynamically in space and time during morphogenesis, RNNs are a suitable approach to treat experimental data sequences. Several architectures of RNN have been proposed to improve training and solve the vanishing gradient problem through time [149]. LSTM comprise memory cells to infer long-term dependencies in sequences [150–152]. Gated Recurrent Units are another RNN architecture that addresses the long-term memory problem and outperforms LSTM in some applications [153, 154]. Sets of LSTM can be combined to design an encoder-decoder that approaches the problem as a conversion of the input sequence into an intermediate fixed-length sequence (encoder) that can be further classified (decoder) [153, 155]. Recent advances in sequence analysis have been based on the idea of attention [156–161]. Attention architectures deal with long inputs by focusing on relevant frames of the sequence, eliminating the restriction of a fixed-length intermediate sequence, and leveraging intermediate states of the encoder as additional input to the decoder. Attention also provides clarity of the input-output relationships [156] and has shown promising results in end-to-end entailment of complex data sequences [162]. The transformer, an architecture without recurrence that relies on feed-forward layers and attention, has been proposed to exploit the potential of attention while allowing for massive parallelization [161, 163].
A key issue is how to pre-train [163, 164] and train these architectures with the data streams of morphogenesis. For instance, contextual bidirectional pre-training might facilitate characterizing strain-stress relationships given past and future tissue states (Figures 2E,F), in order to generate stress maps. Also, entailment of morphogenetic cues and mechanical events with fate map determination could be possible using the input defined by the profiles of cell trajectories, labeled according to a given morphogenetic field or a mature organ. In this regard, the scalability of biological domain tagging could introduce bottlenecks in the generation of training sets, particularly when considering the inherent variability of biological data. These tasks may require using several input vectors at the same time requiring extending current speech-oriented DL architectures [165, 166].
In this perspective, we have critically surveyed recent advances in computational methods for the characterization of embryogenesis, focusing on how to integrate data from biophysical measurements or models into cell fate maps. The ongoing surge in research efforts to quantify the biophysics of morphogenesis is leading to important methodological contributions and new insights about how genetics unfold into phenotypes. Despite these advances, the mechanistic description of morphogenesis remains challenging, given the dynamic and multi-scale nature of the underlying processes and the notable plasticity of immature cells. Thus, new methods are required to understand the interplay of physics, genetics, and epigenetics, leading to cell fate map determination. State-of-the-art imaging systems, image analyses, and computer vision methods are enabling the digital curation of multi-dimensional, high-resolution atlases of developing embryos. These data need to be structured in a systematic way to ensure experimental reproducibility and compatibility of different databases, which are necessary for statistically significant comparisons of large cohorts. In this sense, we posit that data analysis would benefit from a Lagrangian representation based on cell trajectories containing the cumulative histories of the spatio-temporal events and environmental cues cells experience along their paths. This representation integrates spatial information into temporal sequences allowing for multi-scale discovery of morphomechanical fields.
Computational models offer a powerful toolbox to assimilate and explain experimental data, as well as to test new hypotheses. As these models grow in sophistication, they are beginning to predict and decipher the dynamics of developing embryos, based on multi-scale biophysical formalisms that can tackle spatio-temporal heterogeneity and complex mechanobiological interplays. These formalisms are benefitting from novel, minimally-invasive experimental approaches to measure the evolving mechanical properties of live embryos. However, the increasing diversity of models makes it difficult to identify, harmonize, and cross-validate a set of laws that govern the dynamics of morphogenesis. The lack of long-term maintenance of many open-source modeling codes makes this task additionally challenging.
In parallel, machine learning is quickly gaining traction as an alternative to classic model-driven computation to leverage intensive experimentation machine learning and causality inference tools [167, 168] can help test the completeness of models. In particular, these tools can elucidate morphomechanical domains formed by cells with similar dynamics, and link the formation of these domains with upstream biomechanical events. Deep learning (DL) architectures are becoming transparent and interpretable by nesting data-driven or model-driven visible nodes, and have been proven useful to discover causal links in other biological processes. For a holistic approach, DL is suitable to analyze spatio-temporal profiles, seek for events, discover patterns and identify dynamic entities. Multi-scale comparison of cohorts with model-driven DL architectures can be the basis to discover “missing data,” factors and critical spatio-temporal processes regulating phenotype configuration. Overall, the methodologies and approaches here discussed will have valuable practical applications for tissue engineering, stem cell research, genetics and behavior of diseases, drug studies, and synthetic life.
The dataset used for visualization in Figure 1 and the free version of the visualization tool Mov-IT are freely available from the BioEmergences open workflow http://bioemergences.iscpif.fr/bioemergences/openworkflow-index.php. Data and tool are described in Faure et al. [36].
A dataset of a developing wild-type zebrafish embryo was presented in Figure 1. This dataset was produced by the BioEmergences lab (bioemergences.eu) as described in Faure et al. [36].
DP-E conceived the work, made the figures, and co-wrote the manuscript. JÁ advised for this work and co-wrote the manuscript.
This work was supported by NIH grants 1 R01 HD092216-01A1, NIH 1R01HL128630, 1R01HL130840, NIH 2R01 GM084227, and NSF grant NSF CBET – 1706436/1706571.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank BioEmergences Lab-CNRS and Nadine Peyriéras for the joint work on computational developmental biology that inspired this work. We also thank Nicole Gorfinkiel for discussions on tissue mechanics. We thank the Biomedical Image Technologies Lab-UPM, Andres Santos, María Jesús Ledesma-Carbayo and Jose M. Goicolea for their collaboration on previous work.
1. Keller R. Physical biology returns to morphogenesis. Science. (2012) 338:201–3. doi: 10.1126/science.1230718
2. Davidson L, von Dassow M, Zhou J. Multi-scale mechanics from molecules to morphogenesis. Int J Biochem Cell Biol. (2009) 41:2147–62. doi: 10.1016/j.biocel.2009.04.015
3. Blanchard GB, Adams RJ. Measuring the multi-scale integration of mechanical forces during morphogenesis. Curr Opin Genet Dev. (2011) 21:653–63. doi: 10.1016/j.gde.2011.08.008
4. Gilmour D, Rembold M, Leptin M. From morphogen to morphogenesis and back. Nature. (2017) 541:311–20. doi: 10.1038/nature21348
5. Heisenberg C-P, Bellaïche Y. Forces in tissue morphogenesis and patterning. Cell. (2013) 153:948–62. doi: 10.1016/j.cell.2013.05.008
6. Lecuit T, Lenne P-F, Munro E. Force generation, transmission, and integration during cell and tissue morphogenesis. Annu Rev Cell Dev Biol. (2011) 27:157–84. doi: 10.1146/annurev-cellbio-100109-104027
7. Heller E, Fuchs E. Tissue patterning and cellular mechanics. J Cell Biol. (2015) 211:219–31. doi: 10.1083/jcb.201506106
8. Wozniak MA, Chen CS. Mechanotransduction in development: a growing role for contractility. Nat Rev Mol Cell Biol. (2009) 10:34–43. doi: 10.1038/nrm2592
9. Mammoto T, Ingber DE. Mechanical control of tissue and organ development. Development. (2010) 137:1407–20. doi: 10.1242/dev.024166
10. Zhang R, Han P, Yang H, Ouyang K, Lee D, Lin Y-F, et al. In vivo cardiac reprogramming contributes to zebrafish heart regeneration. Nature. (2013) 498:497–501. doi: 10.1038/nature12322
11. Gálvez-Santisteban M, Chen D, Zhang R, Serrano R, Nguyen C, Zhao L, et al. Hemodynamic-mediated endocardial signaling controls in vivo myocardial reprogramming. Elife. (2019) 8:e44816. doi: 10.7554/eLife.44816
12. Roca-Cusachs P, Conte V, Trepat X. Quantifying forces in cell biology. Nat Cell Biol. (2017) 19:742–51. doi: 10.1038/ncb3564
13. Kwan KM, Otsuna H, Kidokoro H, Carney KR, Saijoh Y, Chien C-B. A complex choreography of cell movements shapes the vertebrate eye. Development. (2012) 139:359–72. doi: 10.1242/dev.071407
14. Briggs JA, Weinreb C, Wagner DE, Megason S, Peshkin L, Kirschner MW, et al. The dynamics of gene expression in vertebrate embryogenesis at single-cell resolution. Science. (2018) 360:eaar5780. doi: 10.1126/science.aar5780
15. Wagner DE, Weinreb C, Collins ZM, Briggs JA, Megason SG, Klein AM. Single-cell mapping of gene expression landscapes and lineage in the zebrafish embryo. Science. (2018) 360:981–7. doi: 10.1126/science.aar4362
16. Farrell JA, Wang Y, Riesenfeld SJ, Shekhar K, Regev A, and Schier AF. Single-cell reconstruction of developmental trajectories during zebrafish embryogenesis. Science. (2018) 360:eaar3131. doi: 10.1126/science.aar3131
17. Turing AM. The chemical basis of morphogenesis. Bull Math Biol. (1990) 52:153–97. doi: 10.1007/BF02459572
18. Kimmel CB, Warga RM, Schilling TF. Origin and organization of the zebrafish fate map. Development. (1990) 108:581–94.
19. Woo K, Fraser SE. Order and coherence in the fate map of the zebrafish nervous system. Development. (1995) 121:2595–609.
20. Sako K, Pradhan SJ, Barone V, Inglés-Prieto Á, Müller P, Ruprecht V, et al. Optogenetic control of nodal signaling reveals a temporal pattern of nodal signaling regulating cell fate specification during gastrulation. Cell Rep. (2016) 16:866–77. doi: 10.1016/j.celrep.2016.06.036
21. Chan CJ, Heisenberg C-P, Hiiragi T. Coordination of morphogenesis and cell-fate specification in development. Curr Biol. (2017) 27:R1024–35. doi: 10.1016/j.cub.2017.07.010
22. Keller PJ. Imaging morphogenesis: technological advances and biological insights. Science. (2013) 340:1234168. doi: 10.1126/science.1234168
23. Polacheck WJ, Chen CS. Measuring cell-generated forces: a guide to the available tools. Nat Methods. (2016) 13:415–23. doi: 10.1038/nmeth.3834
24. Villoutreix P, Delile J, Rizzi B, Duloquin L, Savy T, Bourgine P, et al. An integrated modelling framework from cells to organism based on a cohort of digital embryos. Sci Rep. (2016) 6:37438. doi: 10.1038/srep37438
25. Sharpe J. Computer modeling in developmental biology: growing today, essential tomorrow. Development. (2017) 144:4214–25. doi: 10.1242/dev.151274
26. Delile J, Herrmann M, Peyriéras N, Doursat R. A cell-based computational model of early embryogenesis coupling mechanical behaviour and gene regulation. Nat Commun. (2017) 8:13929. doi: 10.1038/ncomms13929
27. Yeh Y-T, Serrano R, François J, Chiu J-J, Li Y-SJ, Del Álamo JC, et al. Three-dimensional forces exerted by leukocytes and vascular endothelial cells dynamically facilitate diapedesis. Proc Natl Acad Sci USA. (2018) 115:133–8. doi: 10.1073/pnas.1717489115
28. Latorre E, Kale S, Casares L, Gómez-González M, Uroz M, Valon L, et al. Active superelasticity in three-dimensional epithelia of controlled shape. Nature. (2018) 563:203–8. doi: 10.1038/s41586-018-0671-4
29. Forgacs G, Foty RA, Shafrir Y, Steinberg MS. Viscoelastic properties of living embryonic tissues: a quantitative study. Biophys J. (1998) 74:2227–34. doi: 10.1016/S0006-3495(98)77932-9
30. Marmottant P, Mgharbel A, Käfer J, Audren B, Rieu J-P, Vial J-C, et al. The role of fluctuations and stress on the effective viscosity of cell aggregates. Proc Natl Acad Sci USA. (2009) 106:17271–5. doi: 10.1073/pnas.0902085106
31. Wu P-H, Aroush DR-B, Asnacios A, Chen W-C, Dokukin ME, Doss BL, et al. A comparison of methods to assess cell mechanical properties. Nat Methods. (2018) 15:491–8. doi: 10.1038/s41592-018-0015-1
32. Olivier N, Luengo-Oroz MA, Duloquin L, Faure E, Savy T, Veilleux I, et al. Cell lineage reconstruction of early zebrafish embryos using label-free nonlinear microscopy. Science. (2010) 329:967–71. doi: 10.1126/science.1189428
33. Supatto W, Truong TV, Débarre D, Beaurepaire E. Advances in multiphoton microscopy for imaging embryos. Curr Opin Genet Dev. (2011) 21:538–48. doi: 10.1016/j.gde.2011.08.003
34. Gao L, Shao L, Chen B-C, Betzig E. 3D live fluorescence imaging of cellular dynamics using Bessel beam plane illumination microscopy. Nat Protoc. (2014) 9:1083–1101. doi: 10.1038/nprot.2014.087
35. Wolff C, Tinevez J-Y, Pietzsch T, Stamataki E, Harich B, Guignard L, et al. Multi-view light-sheet imaging and tracking with the MaMuT software reveals the cell lineage of a direct developing arthropod limb. Elife. (2018) 7:e34410. doi: 10.7554/eLife.34410
36. Faure E, Savy T, Rizzi B, Melani C, Stašová O, Fabrèges D, et al. A workflow to process 3D+ time microscopy images of developing organisms and reconstruct their cell lineage. Nat Commun. (2016) 7:8674. doi: 10.1038/ncomms9674
37. Amat F, Lemon W, Mossing DP, McDole K, Wan Y, Branson K, et al. Fast, accurate reconstruction of cell lineages from large-scale fluorescence microscopy data. Nat Methods. (2014) 11:951–8. doi: 10.1038/nmeth.3036
38. Tinevez J-Y, Pietzsch T, Rueden C. MaMuT. Github (2018). Available online at: https://github.com/fiji/MaMuT (accessed September 1, 2019).
39. Stegmaier J, Amat F, Lemon WC, McDole K, Wan Y, Teodoro G, et al. Real-time three-dimensional cell segmentation in large-scale microscopy data of developing embryos. Dev Cell. (2016) 36:225–40. doi: 10.1016/j.devcel.2015.12.028
40. Ulman V, Maška M, Magnusson KE, Ronneberger O, Haubold C, Harder N, et al. An objective comparison of cell-tracking algorithms. Nat Methods. (2017) 14:1141–52. doi: 10.1038/nmeth.4473
41. Dufour AC, Jonker AH, Olivo-Marin J-C. Deciphering tissue morphodynamics using bioimage informatics. Philos Trans R Soc B Biol Sci. (2017) 372:20150512. doi: 10.1098/rstb.2015.0512
42. Schott B, Traub M, Schlagenhauf C, Takamiya M, Antritter T, Bartschat A, et al. EmbryoMiner: a new framework for interactive knowledge discovery in large-scale cell tracking data of developing embryos. PLoS Comput Biol. (2018) 14:e1006128. doi: 10.1371/journal.pcbi.1006128
43. Leggio B, Laussu J, Carlier A, Godin C, Lemaire P, Faure E. MorphoNet: an interactive online morphological browser to explore complex multi-scale data. Nat Commun. (2019) 10:2812. doi: 10.1038/s41467-019-10668-1
44. Tomer R, Khairy K, Amat F, Keller PJ. Quantitative high-speed imaging of entire developing embryos with simultaneous multiview light-sheet microscopy. Nat Methods. (2012) 9:755–63. doi: 10.1038/nmeth.2062
45. Wu Y, Chandris P, Winter PW, Kim EY, Jaumouillé V, Kumar A, et al. Simultaneous multiview capture and fusion improves spatial resolution in wide-field and light-sheet microscopy. Optica. (2016) 3:897–910. doi: 10.1364/OPTICA.3.000897
46. Rubio-Guivernau JL, Gurchenkov V, Luengo-Oroz MA, Duloquin L, Bourgine P, Santos A, et al. Wavelet-based image fusion in multi-view three-dimensional microscopy. Bioinformatics. (2011) 28:238–45. doi: 10.1093/bioinformatics/btr609
47. Schmied C, Stamataki E, Tomancak P. Open-source solutions for SPIMage processing. Methods Cell Biol. (2014) 505–29. doi: 10.1016/B978-0-12-420138-5.00027-6
48. Liu T-L, Upadhyayula S, Milkie DE, Singh V, Wang K, Swinburne IA, et al. Observing the cell in its native state: imaging subcellular dynamics in multicellular organisms. Science. (2018) 360:eaaq1392. doi: 10.1126/science.aaq1392
49. Ronneberger O, Liu K, Rath M, Rueβ D, Mueller T, Skibbe H, et al. ViBE-Z: a framework for 3D virtual colocalization analysis in zebrafish larval brains. Nat Methods. (2012) 9:735–42. doi: 10.1038/nmeth.2076
50. Castro-González C, Luengo-Oroz MA, Duloquin L, Savy T, Rizzi B, Desnoulez S, et al. A digital framework to build, visualize and analyze a gene expression atlas with cellular resolution in zebrafish early embryogenesis. PLoS Comput Biol. (2014) 10:e1003670. doi: 10.1371/journal.pcbi.1003670
51. Power RM, Huisken J. A guide to light-sheet fluorescence microscopy for multiscale imaging. Nat Methods. (2017) 14:360–73. doi: 10.1038/nmeth.4224
52. Ovečka M, von Wangenheim D, Tomančák P, Šamajová O, Komis G, Šamaj J. Multiscale imaging of plant development by light-sheet fluorescence microscopy. Nature Plants. (2018) 4:639–50. doi: 10.1038/s41477-018-0238-2
53. Pastor Escuredo D. Methods for the Analysis of Multi-Scale Cell Dynamics From Fluorescence Microscopy Images. Madrid: Universidad Politécnica de Madrid, ETSIT (2015).
54. Ledesma-Carbayo MJ, Kybic J, Desco M, Santos A, Suhling M, Hunziker P, et al. Spatio-temporal nonrigid registration for ultrasound cardiac motion estimation. IEEE Trans Med Imaging. (2005) 24:1113–26. doi: 10.1109/TMI.2005.852050
55. Blanchard GB, Kabla AJ, Schultz NL, Butler LC, Sanson B, Gorfinkiel N, et al. Tissue tectonics: morphogenetic strain rates, cell shape change and intercalation. Nat Methods. (2009) 6:458–64. doi: 10.1038/nmeth.1327
56. He B, Doubrovinski K, Polyakov O, Wieschaus E. Apical constriction drives tissue-scale hydrodynamic flow to mediate cell elongation. Nature. (2014) 508:392–6. doi: 10.1038/nature13070
57. Pastor-Escuredo D, Lombardot B, Savy T, Boyreau A, Goicolea JM, Santos A, et al. Kinematic analysis of cell lineage reveals coherent and robust mechanical deformation patterns in zebrafish gastrulation. bioRxiv 054353. doi: 10.1101/054353
58. Stegmaier J, Spina TV, Falcão AX, Bartschat A, Mikut R, Meyerowitz E, et al. Cell segmentation in 3D confocal images using supervoxel merge-forests with CNN-based hypothesis selection. In: 2018 IEEE 15th International Symposium on Biomedical Imaging (ISBI 2018). Washington, DC: IEEE (2018). p. 382–386.
59. Caicedo JC, Roth J, Goodman A, Becker T, Karhohs KW, Broisin M, et al. Evaluation of deep learning strategies for nucleus segmentation in fluorescence images. J Quant Cell Sci. 95:952–65. doi: 10.1002/cyto.a.23863
60. Etournay R, Merkel M, Popović M, Brandl H, Dye NA, Aigouy B, et al. TissueMiner: a multiscale analysis toolkit to quantify how cellular processes create tissue dynamics. Elife. (2016) 5:e14334. doi: 10.7554/eLife.14334.033
61. Linares M, Postigo M, Cuadrado D, Ortiz-Ruiz A, Gil-Casanova S, Vladimirov A, et al. Collaborative intelligence and gamification for on-line malaria species differentiation. Malar J. (2019) 18:21. doi: 10.1186/s12936-019-2662-9
62. Sugimura K, Lenne P-F, Graner F. Measuring forces and stresses in situ in living tissues. Development. (2016) 143:186–96. doi: 10.1242/dev.119776
63. Rauzi M, Verant P, Lecuit T, Lenne P-F. Nature and anisotropy of cortical forces orienting Drosophila tissue morphogenesis. Nat Cell Biol. (2008) 10:1401–10. doi: 10.1038/ncb1798
64. Hutson MS, Veldhuis J, Ma X, Lynch HE, Cranston PG, Brodland GW. Combining laser microsurgery and finite element modeling to assess cell-level epithelial mechanics. Biophys J. (2009) 97:3075–85. doi: 10.1016/j.bpj.2009.09.034
65. Behrndt M, Salbreux G, Campinho P, Hauschild R, Oswald F, Roensch J, et al. Forces driving epithelial spreading in zebrafish gastrulation. Science. (2012) 338:257–60. doi: 10.1126/science.1224143
66. Levayer R, Lecuit T. Biomechanical regulation of contractility: spatial control and dynamics. Trends Cell Biol. (2012) 22:61–81. doi: 10.1016/j.tcb.2011.10.001
67. Machado PF, Duque J, Étienne J, Martinez-Arias A, Blanchard GB, Gorfinkiel N. Emergent material properties of developing epithelial tissues. BMC Biol. (2015) 13:98. doi: 10.1186/s12915-015-0200-y
68. Jurchenko C, Salaita KS. Lighting up the force: investigating mechanisms of mechanotransduction using fluorescent tension probes. Mol Cell Biol. (2015) 35:2570–82. doi: 10.1128/MCB.00195-15
69. Dembo M, Wang Y-L. Stresses at the cell-to-substrate interface during locomotion of fibroblasts. Biophys J. (1999) 76:2307–16. doi: 10.1016/S0006-3495(99)77386-8
70. Butler JP, Tolic-Nørrelykke IM, Fabry B, Fredberg JJ. Traction fields, moments, and strain energy that cells exert on their surroundings. Am J Physiol Cell Physiol. (2002) 282:C595–605. doi: 10.1152/ajpcell.00270.2001
71. Del Alamo JC, Meili R, Alonso-Latorre B, Rodríguez-Rodríguez J, Aliseda A, Firtel RA, et al. Spatio-temporal analysis of eukaryotic cell motility by improved force cytometry. Proc Natl Acad Sci. (2007) 104:13343–8. doi: 10.1073/pnas.0705815104
72. Trepat X, Wasserman MR, Angelini TE, Millet E, Weitz DA, Butler JP, et al. Physical forces during collective cell migration. Nat Phys. (2009) 5:426–30. doi: 10.1038/nphys1269
73. Brodland GW, Conte V, Cranston PG, Veldhuis J, Narasimhan S, Hutson MS, et al. Video force microscopy reveals the mechanics of ventral furrow invagination in Drosophila. Proc Natl Acad Sci USA. (2010) 107:22111–6. doi: 10.1073/pnas.1006591107
74. Hur SS, Zhao Y, Li Y-S, Botvinick E, Chien S. Live cells exert 3-dimensional traction forces on their substrata. Cell Mol Bioeng. (2009) 2:425–36. doi: 10.1007/s12195-009-0082-6
75. Serrano R, Aung A, Varghese S, del Álamo JC. Three-dimensional monolayer stress microscopy. Biophys J. (2016) 110:330a. doi: 10.1016/j.bpj.2015.11.1776
76. Heller D, Hoppe A, Restrepo S, Gatti L, Tournier AL, Tapon N, et al. EpiTools: an open-source image analysis toolkit for quantifying epithelial growth dynamics. Dev Cell. (2016) 36:103–16. doi: 10.1016/j.devcel.2015.12.012
77. Bosveld F, Bonnet I, Guirao B, Tlili S, Wang Z, Petitalot A, et al. Mechanical control of morphogenesis by Fat/Dachsous/Four-jointed planar cell polarity pathway. Science. (2012) 336:724–7. doi: 10.1126/science.1221071
78. Blanchard GB. Taking the strain: quantifying the contributions of all cell behaviours to changes in epithelial shape. Philos Trans R Soc B Biol Sci. (2017) 372:20150513. doi: 10.1098/rstb.2015.0513
79. Tetley RJ, Blanchard GB, Fletcher AG, Adams RJ, Sanson B. Unipolar distributions of junctional Myosin II identify cell stripe boundaries that drive cell intercalation throughout Drosophila axis extension. Elife. (2016) 5:e12094. doi: 10.7554/eLife.12094
80. Tlili S, Gay C, Graner F, Marcq P, Molino F, Saramito P. Colloquium: mechanical formalisms for tissue dynamics. Eur Phys J E. (2015) 38:33. doi: 10.1140/epje/i2015-15033-4
81. Chiou KK, Hufnagel L, Shraiman BI. Mechanical stress inference for two dimensional cell arrays. PLoS Comput Biol. (2012) 8:e1002512. doi: 10.1371/journal.pcbi.1002512
82. Ishihara S, Sugimura K. Bayesian inference of force dynamics during morphogenesis. J Theor Biol. (2012) 313:201–11. doi: 10.1016/j.jtbi.2012.08.017
83. Ishihara S, Sugimura K, Cox S, Bonnet I, Bellaiche Y, Graner F. Comparative study of non-invasive force and stress inference methods in tissue. Eur Phys J E. (2013) 36:45. doi: 10.1140/epje/i2013-13045-8
84. Brodland GW, Veldhuis JH, Kim S, Perrone M, Mashburn D, Hutson MS. CellFIT: a cellular force-inference toolkit using curvilinear cell boundaries. PLoS ONE. (2014) 9:e99116. doi: 10.1371/journal.pone.0099116
85. Guirao B, Rigaud SU, Bosveld F, Bailles A, Lopez-Gay J, Ishihara S, et al. Unified quantitative characterization of epithelial tissue development. Elife. (2015) 4:e08519. doi: 10.7554/eLife.08519
86. Veldhuis JH, Ehsandar A, Maître J-L, Hiiragi T, Cox S, Brodland GW. Inferring cellular forces from image stacks. Philos Trans R Soc B Biol Sci. (2017) 372:20160261. doi: 10.1098/rstb.2016.0261
87. Mongera A, Rowghanian P, Gustafson HJ, Shelton E, Kealhofer DA, Carn EK, et al. A fluid-to-solid jamming transition underlies vertebrate body axis elongation. Nature. (2018) 561:401–5. doi: 10.1038/s41586-018-0479-2
88. Campàs O, Mammoto T, Hasso S, Sperling RA, O'connell D, Bischof AG, et al. Quantifying cell-generated mechanical forces within living embryonic tissues. Nat Methods. (2014) 11:183–9. doi: 10.1038/nmeth.2761
89. Dolega M, Delarue M, Ingremeau F, Prost J, Delon A, Cappello G. Cell-like pressure sensors reveal increase of mechanical stress towards the core of multicellular spheroids under compression. Nat Commun. (2017) 8:14056. doi: 10.1038/ncomms14056
90. Serwane F, Mongera A, Rowghanian P, Kealhofer DA, Lucio AA, Hockenbery ZM, et al. In vivo quantification of spatially varying mechanical properties in developing tissues. Nat Methods. (2017) 14:181–6. doi: 10.1038/nmeth.4101
91. Byrne H, Drasdo D. Individual-based and continuum models of growing cell populations: a comparison. J Math Biol. (2009) 58:657. doi: 10.1007/s00285-008-0212-0
92. Van Liedekerke P. (2019). Quantitative Modeling of Cell and Tissue Mechanics With Agent-Based Models. Paris: Inria Paris, Sobonne Université.
93. Van Liedekerke P, Palm M, Jagiella N, Drasdo D. Simulating tissue mechanics with agent-based models: concepts, perspectives and some novel results. Comput Part Mech. (2015) 2:401–44. doi: 10.1007/s40571-015-0082-3
94. Drasdo D, Höhme S. A single-cell-based model of tumor growth in vitro: monolayers and spheroids. Phys Biol. (2005) 2:133–47. doi: 10.1088/1478-3975/2/3/001
95. Van Liedekerke P, Neitsch J, Johann T, Warmt E, Grosser S, Valverde IG, et al. Quantifying the mechanics and growth of cells and tissues in 3D using high resolution computational models. bioRxiv 470559. doi: 10.1101/470559
96. Swat MH, Thomas GL, Belmonte JM, Shirinifard A, Hmeljak D, Glazier JA. Multi-scale modeling of tissues using CompuCell3D. Methods Cell Biol. (2012) 325–66. doi: 10.1016/B978-0-12-388403-9.00013-8
97. Voss-Böhme A. Multi-scale modeling in morphogenesis: a critical analysis of the cellular Potts model. PLoS ONE. (2012) 7:e42852. doi: 10.1371/journal.pone.0042852
98. Graner F, Glazier JA. Simulation of biological cell sorting using a two-dimensional extended Potts model. Phys Rev Lett. (1992) 69:2013. doi: 10.1103/PhysRevLett.69.2013
99. Käfer J, Hayashi T, Marée AF, Carthew RW, Graner F. Cell adhesion and cortex contractility determine cell patterning in the Drosophilaretina. Proc Natl Acad Sci USA. (2007) 104:18549–54. doi: 10.1073/pnas.0704235104
100. Krieg M, Arboleda-Estudillo Y, Puech P-H, Käfer J, Graner F, Müller D, et al. Tensile forces govern germ-layer organization in zebrafish. Nat Cell Biol. (2008) 10:429–36. doi: 10.1038/ncb1705
101. Fletcher AG, Cooper F, Baker RE. Mechanocellular models of epithelial morphogenesis. Philos Trans R Soc B Biol Sci. (2017) 372:20150519. doi: 10.1098/rstb.2015.0519
102. Alt S, Ganguly P, Salbreux G. Vertex models: from cell mechanics to tissue morphogenesis. Philos Trans R Soc B Biol Sci. (2017) 372:20150520. doi: 10.1098/rstb.2015.0520
103. Fletcher AG, Osterfield M, Baker RE, Shvartsman SY. Vertex models of epithelial morphogenesis. Biophys J. (2014) 106:2291–304. doi: 10.1016/j.bpj.2013.11.4498
104. Nagai T, Honda H. A dynamic cell model for the formation of epithelial tissues. Philos Mag B. (2001) 81:699–719. doi: 10.1080/13642810108205772
105. Farhadifar R, Röper J-C, Aigouy B, Eaton S, Jülicher F. The influence of cell mechanics, cell-cell interactions, and proliferation on epithelial packing. Curr Biol. (2007) 17:2095–104. doi: 10.1016/j.cub.2007.11.049
106. Ishimoto Y, Morishita Y. Bubbly vertex dynamics: a dynamical and geometrical model for epithelial tissues with curved cell shapes. Phys Rev E. (2014) 90:052711. doi: 10.1103/PhysRevE.90.052711
107. Perrone MC, Veldhuis JH, Brodland GW. Non-straight cell edges are important to invasion and engulfment as demonstrated by cell mechanics model. Biomech Model Mechanobiol. (2016) 15:405–18. doi: 10.1007/s10237-015-0697-6
108. Tanaka S, Sichau D, Iber D. LBIBCell: a cell-based simulation environment for morphogenetic problems. Bioinformatics. (2015) 31:2340–7. doi: 10.1093/bioinformatics/btv147
109. Brezavšček AH, Rauzi M, Leptin M, Ziherl P. A model of epithelial invagination driven by collective mechanics of identical cells. Biophys J. (2012) 103:1069–77. doi: 10.1016/j.bpj.2012.07.018
110. Okuda S, Inoue Y, Eiraku M, Sasai Y, Adachi T. Apical contractility in growing epithelium supports robust maintenance of smooth curvatures against cell-division-induced mechanical disturbance. J Biomech. (2013) 46:1705–13. doi: 10.1016/j.jbiomech.2013.03.035
111. Murisic N, Hakim V, Kevrekidis IG, Shvartsman SY, Audoly B. From discrete to continuum models of three-dimensional deformations in epithelial sheets. Biophys J. (2015) 109:154–63. doi: 10.1016/j.bpj.2015.05.019
112. Monier B, Gettings M, Gay G, Mangeat T, Schott S, Guarner A, et al. Apico-basal forces exerted by apoptotic cells drive epithelium folding. Nature. (2015) 518:245–8. doi: 10.1038/nature14152
113. Rauzi M, Krzic U, Saunders TE, Krajnc M, Ziherl P, Hufnagel L, et al. Embryo-scale tissue mechanics during Drosophila gastrulation movements. Nat Commun. (2015) 6:8677. doi: 10.1038/ncomms9677
114. Misra M, Audoly B, Kevrekidis IG, Shvartsman SY. Shape transformations of epithelial shells. Biophys J. (2016) 110:1670–8. doi: 10.1016/j.bpj.2016.03.009
115. Hufnagel L, Teleman AA, Rouault H, Cohen SM, Shraiman BI. On the mechanism of wing size determination in fly development. Proc Natl Acad Sci USA. (2007) 104:3835–40. doi: 10.1073/pnas.0607134104
116. Aegerter-Wilmsen T, Smith AC, Christen AJ, Aegerter CM, Hafen E, Basler K. Exploring the effects of mechanical feedback on epithelial topology. Development. (2010) 137:499–506. doi: 10.1242/dev.041731
117. Li Y, Naveed H, Kachalo S, Xu LX, Liang J. Mechanisms of regulating cell topology in proliferating epithelia: impact of division plane, mechanical forces, and cell memory. PLoS ONE. (2012) 7:e43108. doi: 10.1371/journal.pone.0043108
118. Marinari E, Mehonic A, Curran S, Gale J, Duke T, Baum B. Live-cell delamination counterbalances epithelial growth to limit tissue overcrowding. Nature. (2012) 484:542–45. doi: 10.1038/nature10984
119. Aigouy B, Farhadifar R, Staple DB, Sagner A, Röper J-C, Jülicher F, et al. Cell flow reorients the axis of planar polarity in the wing epithelium of Drosophila. Cell. (2010) 142:773–86. doi: 10.1016/j.cell.2010.07.042
120. Landsberg KP, Farhadifar R, Ranft J, Umetsu D, Widmann TJ, Bittig T, et al. Increased cell bond tension governs cell sorting at the Drosophila anteroposterior compartment boundary. Curr Biol. (2009) 19:1950–5. doi: 10.1016/j.cub.2009.10.021
121. Zhao J, Cao Y, DiPietro LA, Liang J. Dynamic cellular finite-element method for modelling large-scale cell migration and proliferation under the control of mechanical and biochemical cues: a study of re-epithelialization. J R Soc Interface. (2017) 14. doi: 10.1098/rsif.2016.0959
122. Marchetti MC, Joanny J-F, Ramaswamy S, Liverpool TB, Prost J, Rao M, et al. Hydrodynamics of soft active matter. Rev Mod Phys. (2013) 85:1143. doi: 10.1103/RevModPhys.85.1143
123. Blanchard GB, Fletcher AG, Schumacher LJ. The devil is in the mesoscale: mechanical and behavioural heterogeneity in collective cell movement. Semin Cell Dev Biol. (2018) 93:46–54. doi: 10.1016/j.semcdb.2018.06.003
124. Conte V, Muñoz JJ, Miodownik M. A 3D finite element model of ventral furrow invagination in the Drosophila melanogaster embryo. J Mech Behav Biomed Mater. (2008) 1:188–98. doi: 10.1016/j.jmbbm.2007.10.002
125. González-Valverde I, García-Aznar JM. Mechanical modeling of collective cell migration: an agent-based and continuum material approach. Comput Methods Appl Mech Eng. (2018) 337:246–62. doi: 10.1016/j.cma.2018.03.036
126. Shadden SC, Lekien F, Marsden JE. Definition and properties of Lagrangian coherent structures from finite-time Lyapunov exponents in two-dimensional aperiodic flows. Physica D. (2005) 212:271–304. doi: 10.1016/j.physd.2005.10.007
127. Haller G. Lagrangian coherent structures. Annu Rev Fluid Mech. (2015) 47:137–62. doi: 10.1146/annurev-fluid-010313-141322
128. Kitano H. Systems biology: a brief overview. Science. (2002) 295:1662–4. doi: 10.1126/science.1069492
129. Biasuz K, Leggio B, Faure E, Lemaire P. The “computable egg”: myth or useful concept? Curr Opin Syst Biol. (2018) 11:91–7. doi: 10.1016/j.coisb.2018.09.003
130. Vecchi D, Hernández I. The epistemological resilience of the concept of morphogenetic field. In: Towards a Theory of Development. Oxford, UK: Oxford University Press (2014). p. 79.
131. Luengo-Oroz MA, Pastor-Escuredo D, Castro-Gonzalez C, Faure E, Savy T, Lombardot B, et al. $3 D+ t $ morphological processing: applications to embryogenesis image analysis. IEEE Trans Image Proc. (2012) 21:3518–30. doi: 10.1109/TIP.2012.2197007
132. Hinton G, Deng L, Yu D, Dahl G, Mohamed AR, Jaitly N, et al. Deep neural networks for acoustic modeling in speech recognition. IEEE Signal Proc Mag. (2012) 29:82–97. doi: 10.1109/MSP.2012.2205597
133. Sainath TN, Mohamed AR, Kingsbury B, Ramabhadran B. Deep convolutional neural networks for LVCSR. In: 2013 IEEE International Conference on Acoustics, Speech and Signal Processing. Vancouver, BC: IEEE (2013). p. 8614–8.
134. Silver D, Huang A, Maddison CJ, Guez A, Sifre L, Van Den Driessche G, et al. Mastering the game of Go with deep neural networks and tree search. Nature. (2016) 529:484–9. doi: 10.1038/nature16961
135. LeCun Y, Bengio Y, Hinton G. Deep learning. Nature. (2015) 521:436–44. doi: 10.1038/nature14539
136. Mamoshina P, Vieira A, Putin E, Zhavoronkov A. Applications of deep learning in biomedicine. Mol Pharm. (2016) 13:1445–54. doi: 10.1021/acs.molpharmaceut.5b00982
137. Min S, Lee B, Yoon S. Deep learning in bioinformatics. Brief Bioinformatics. (2017) 18:851–69. doi: 10.1093/bib/bbw068
138. Wainberg M, Merico D, Delong A, Frey BJ. Deep learning in biomedicine. Nat Biotechnol. (2018) 36:829–38. doi: 10.1038/nbt.4233
139. Zitnik M, Nguyen F, Wang B, Leskovec J, Goldenberg A, Hoffman MM. Machine learning for integrating data in biology and medicine: Principles, practice, and opportunities. Inform Fus. (2019) 50:71–91. doi: 10.1016/j.inffus.2018.09.012
140. Angermueller C, Pärnamaa T, Parts L, Stegle O. Deep learning for computational biology. Mol Syst Biol. (2016) 12:878. doi: 10.15252/msb.20156651
141. Eraslan G, Avsec Ž, Gagneur J, Theis FJ. Deep learning: new computational modelling techniques for genomics. Nat Rev Genet. 20:389–403. doi: 10.1038/s41576-019-0122-6
142. Michael KY, Ma J, Fisher J, Kreisberg JF, Raphael BJ, Ideker T. Visible machine learning for biomedicine. Cell. (2018) 173:1562–5. doi: 10.1016/j.cell.2018.05.056
143. Ma J, Yu MK, Fong S, Ono K, Sage E, Demchak B, et al. Using deep learning to model the hierarchical structure and function of a cell. Nat Methods. (2018) 15:290–8. doi: 10.1038/nmeth.4627
144. Gazestani VH, Lewis NE. From genotype to phenotype: Augmenting deep learning with networks and systems biology. Curr Opin Syst Biol. (2019) 15:68–73. doi: 10.1016/j.coisb.2019.04.001
145. Wang D, Liu S, Warrell J, Won H, Shi X, Navarro FC, et al. (2018). Comprehensive functional genomic resource and integrative model for the human brain. Science. 362:eaat8464. doi: 10.1126/science.aat8464
146. Coulier A, Hellander A. Orchestral: a lightweight framework for parallel simulations of cell-cell communication. In: 2018 IEEE 14th International Conference on e-Science (e-Science). Amsterdam: IEEE (2018), 168–176.
147. Ghaffarizadeh A, Heiland R, Friedman SH, Mumenthaler SM, Macklin P. PhysiCell: an open source physics-based cell simulator for 3-D multicellular systems. PLoS Comput Biol. (2018) 14:e1005991. doi: 10.1371/journal.pcbi.1005991
148. Tracey BD, Duraisamy K, and Alonso JJ. A machine learning strategy to assist turbulence model development. In: 53rd AIAA Aerospace Sciences Meeting. Kissimmee, FL (2015). p. 1287.
149. Jozefowicz R, Zaremba W, Sutskever I. An empirical exploration of recurrent network architectures. In: International Conference on Machine Learning. Lille (2015). p. 2342–50.
150. Hochreiter S, Schmidhuber J. Long short-term memory. Neural Comput. (1997) 9:1735–80. doi: 10.1162/neco.1997.9.8.1735
151. Sak H, Senior A, Beaufays F. Long short-term memory recurrent neural network architectures for large scale acoustic modeling. In: Fifteenth Annual Conference of the International Speech Communication Association. Singapore (2014).
152. Malhotra P, Vig L, Shroff G, and Agarwal P. Long short term memory networks for anomaly detection in time series. In: Proceedings: Presses universitaires de Louvain. Louvain (2015). p. 89.
153. Cho K, Van Merriënboer B, Gulcehre C, Bahdanau D, Bougares F, Schwenk H, et al. Learning phrase representations using RNN encoder-decoder for statistical machine translation. arXiv [Preprint] arXiv:1406.1078 (2014). doi: 10.3115/v1/D14-1179
154. Chung J, Gulcehre C, Cho K, and Bengio Y. Empirical evaluation of gated recurrent neural networks on sequence modeling. arXiv [Preprint] arXiv:1412.3555 (2014).
155. Sutskever I, Vinyals O, Le QV. Sequence to sequence learning with neural networks. In: Advances in Neural Information Processing Systems. Montréal, QC (2014). p. 3104–12.
156. Bahdanau D, Cho K, and Bengio Y. Neural machine translation by jointly learning to align and translate. arXiv [Preprint] arXiv:1409.0473 (2014).
157. Xu K, Ba J, Kiros R, Cho K, Courville A, Salakhudinov R, et al. Show, attend and tell: neural image caption generation with visual attention. In: International Conference on Machine Learning. Lille (2015). p. 2048–57.
158. Hermann KM, Kocisky T, Grefenstette E, Espeholt L, Kay W, Suleyman M, et al. Teaching machines to read and comprehend. In: Advances in Neural Information Processing Systems. Montréal, QC (2015). p. 1693–1701.
159. Luong M T, Pham H, Manning CD. Effective approaches to attention-based neural machine translation. arXiv [Preprint] arXiv:1508.04025 (2015). doi: 10.18653/v1/D15-1166
160. You Q, Jin H, Wang Z, Fang C, and Luo J. Image captioning with semantic attention. In: Proceedings of the IEEE Conference on Computer Vision and Pattern Recognition. Las Vegas, NV (2016). p. 4651–9.
161. Vaswani A, Shazeer N, Parmar N, Uszkoreit J, Jones L, Gomez AN, et al. Attention is all you need. In: Advances in Neural Information Processing Systems. Long Beach, CA (2017). p. 5998–6008.
162. Rocktäschel T, Grefenstette E, Hermann KM, Kočiský T, Blunsom P. Reasoning about entailment with neural attention. arXiv [Preprint] arXiv:1509.06664 (2015).
163. Devlin J, Chang MW, Lee K, Toutanova K. Bert: pre-training of deep bidirectional transformers for language understanding. arXiv [Preprint] arXiv:1810.04805 (2018).
164. Dai AM, Le QV. Semi-supervised sequence learning. In: Advances in Neural Information Processing systems. Montréal, QC (2015). p. 3079–87.
165. Chorowski JK, Bahdanau D, Serdyuk D, Cho K, Bengio Y. Attention-based models for speech recognition. In: Advances in Neural Information Processing systems. Montréal, QC (2015). p. 577–85.
166. Rush AM, Chopra S, Weston J. A neural attention model for abstractive sentence summarization. arXiv [Preprint] arXiv:1509.00685 (2015). doi: 10.18653/v1/D15-1044
Keywords: morphogenesis, cell mechanics, multi-scale modeling, morphomechanical fields, deep learning, cell fate map, fluorescence microscopy, digital embryo
Citation: Pastor-Escuredo D and del Álamo JC (2020) How Computation Is Helping Unravel the Dynamics of Morphogenesis. Front. Phys. 8:31. doi: 10.3389/fphy.2020.00031
Received: 31 October 2019; Accepted: 04 February 2020;
Published: 28 February 2020.
Edited by:
Carlos Gershenson, National Autonomous University of Mexico, MexicoReviewed by:
Ignazio Licata, Institute for Scientific Methodology (ISEM), ItalyCopyright © 2020 Pastor-Escuredo and del Álamo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: David Pastor-Escuredo, ZGF2aWRAbGlmZWRsYWIub3Jn; Juan C. del Álamo, amNAdWNzZC5lZHU=; anVhbmNhckB1dy5lZHU=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.