 Nehal Adel Abdelsalam
Nehal Adel Abdelsalam Ahmed Tarek Ramadan
Ahmed Tarek Ramadan Marwa Tarek ElRakaiby
Marwa Tarek ElRakaiby Ramy Karam Aziz
Ramy Karam Aziz- 1The Center for Genome and Microbiome Research, Cairo University, Cairo, Egypt
- 2Department of Microbiology and Immunology, Faculty of Pharmacy, Cairo University, Cairo, Egypt
The harmful impact of xenobiotics on the environment and human health is being more widely recognized; yet, inter- and intraindividual genetic variations among humans modulate the extent of harm, mostly through modulating the outcome of xenobiotic metabolism and detoxification. As the Human Genome Project revealed that host genetic, epigenetic, and regulatory variations could not sufficiently explain the complexity of interindividual variability in xenobiotics metabolism, its sequel, the Human Microbiome Project, is investigating how this variability may be influenced by human-associated microbial communities. Xenobiotic-microbiome relationships are mutual and dynamic. Not only does the human microbiome have a direct metabolizing potential on xenobiotics, but it can also influence the expression of the host metabolizing genes and the activity of host enzymes. On the other hand, xenobiotics may alter the microbiome composition, leading to a state of dysbiosis, which is linked to multiple diseases and adverse health outcomes, including increased toxicity of some xenobiotics. Toxicomicrobiomics studies these mutual influences between the ever-changing microbiome cloud and xenobiotics of various origins, with emphasis on their fate and toxicity, as well the various classes of microbial xenobiotic-modifying enzymes. This review article discusses classic and recent findings in toxicomicrobiomics, with examples of interactions between gut, skin, urogenital, and oral microbiomes with pharmaceutical, food-derived, and environmental xenobiotics. The current state and future prospects of toxicomicrobiomic research are discussed, and the tools and strategies for performing such studies are thoroughly and critically compared.
Xenobiotics and Their Metabolism
Xenobiotics are chemical substances not normally present in the environment of living organisms. They are mostly regarded as synthetic substances, but the term may be more loosely used to include naturally occurring chemicals and endobiotics, when present in higher concentrations than their normal levels, or produced by certain organisms as a defense mechanism, such as the toxins produced by some fungi, bacteria, or even herbs (Soucek, 2011). From a metabolism viewpoint, they can be defined as chemicals that are extrinsic to the normal metabolism of a living organism (Koppel et al., 2017). Food with its variety of dietary compounds, environmental chemicals and pollutants, as well as medications are considered xenobiotics to the human body (Koppel et al., 2017).
Xenobiotic metabolism increases their water solubility, thus enhancing their elimination from the body (Clarke et al., 2019). Oral ingestion of xenobiotics passes them to the upper gastrointestinal tract, and those absorbed are translocated to the liver through the hepatic portal vein, a process commonly known as the first pass effect. The human liver, using its cytochrome P450 (CYP 450) family of enzymes, chemically transforms both endogenous and exogenous compounds (Nelson, 2005; Michalopoulos, 2007). Liver metabolism has three phases (Soucek, 2011): phase I (activation by oxidation, reduction, or hydrolysis), phase II (conjugation to polar moieties), and phase III (transport without chemical modification). Xenobiotics that are not metabolized and excreted accumulate in the body and may lead to chronic diseases and inflammation (Jain et al., 2005).
Poorly absorbed xenobiotics pass from the small intestine to the large intestine, where they are exposed to the gut microbial metabolizing niche. Metabolites released in the circulation are either excreted by the kidneys or return through biliary duct to the gut. The final fate of these metabolites is either excretion in the stool or reabsorption in the small intestine (Koppel et al., 2017).
Host Genetic Variability and Toxicogenomics
The efficiency of the host metabolizing enzymes varies from one individual to another according to the individual’s genetic makeup. Single nucleotide polymorphisms (SNPs) are among the reasons why individuals respond differently to xenobiotics and drugs (Soucek, 2011). Pharmacogenomics studies the impact of genetic discrepancies between individuals (usually SNPs) on their responses to drugs (Rizkallah et al., 2010; Saad et al., 2012). For example, genetic variants in uridine 5′-diphospho-glucuronosyltransferase (UGT) render some patients to slowly metabolize certain drugs such as NSAIDs and irinotecan due to lower glucuronidation rates (Lankisch et al., 2008; Stingl et al., 2014).
Likewise, toxicogenomics studies the impact of genetic discrepancies between individuals on their response to toxic substances. Philosophically speaking, toxicogenomics and pharmacogenomics are two manifestations of one phenomenon, given that all drugs can be regarded as poisons used at a non-toxic doses (Aziz, 2018). One may even go to an extreme and consider every chemical as a poison, following the classical principle of Paracelsus: “Sola dosis facit venenum” (Lt: only the dose makes the poison (quoted in Uppal et al., 2016)).
Although the host enzymes (notably liver enzymes) are variable and capable of metabolizing xenobiotics, their variability alone cannot explain the biotransformation of indigestible xenobiotics and dietary compounds. Human and microbial chemical transformations form a complex interchangeable network, in which they mutually affect one other (Soucek, 2011).
The Microbiome
The Human Microbiome: A Cloud Shrouded in Mystery
The microbiota is defined as all the microbial communities living on or in a particular biological system, formerly referred to as flora or microflora. The human body carries trillions of microbes with genes estimated to be at least 100 times more numerous than human coding genes. The term microbiome, disambiguated elsewhere (Saad et al., 2012), refers to a micro-ecosystem (micro.biome) or the summation of a microbiota and the genomes of its members (microbial metagenome). Sometimes it even describes the entire environment in which microbes reside (Marchesi and Ravel, 2015). Finally, microbiomics is defined as the field that uses “high throughput molecular techniques to study microbial communities” (Rajendhran and Gunasekaran, 2010).
A human’s microbiome, from one’s birth throughout his/her life, continuously evolves, impacting the body in both states of health and disease. Variations within a microbiome are not only developmental, but are also spatial, temporal, and are definitely affected by diet, hormones, stress, and even diurnal cycle. This continuous, multidimensional variability makes an individual’s microbiome akin to a cloud, with uncertainty of the exact composition and gene pool at any point of time and space (Elrakaiby et al., 2014).
The Human Microbiome: A Brief History
After the Human Genome Project (HGP) has come to a conclusion in 2003, researchers were baffled by its unexpected results (Relman and Falkow, 2001). Coding genes in the human genome were found to be much fewer than previously expected. The genetic, epigenetic, and regulatory variations in the human genome were not sufficient to explain the complexity of interindividual variability and its associated phenotypes. The human microbiome came then to the spotlight, with the promise that studying its composition, variability, and functional potential will allow the full understanding of the landscape of human phenotypic variations, and its effect on human health, immunity, and drug response.
In 2008, the NIH-funded Human Microbiome Project (HMP) was launched to explore intraindividual and interindividual variability influenced by the human microbiome (Turnbaugh et al., 2007). The project’s goals at launch were to (i) take advantage of the current high-throughput technologies to study the human microbiome by analyzing samples taken from multiple body sites (gastrointestinal tract, mouth, vagina, and skin) of more than 250 healthy volunteers; (ii) explore the microbiome’s role in health and disease by studying its variability under different medical conditions; (iii) construct a network that provides up-to-date data sources, new findings, standardized techniques, and technological approaches for the research community.
Currently, the HMP is in its second phase (the Integrative Human Microbiome Project or iHMP), characterized by focusing on selected health disorders (preterm birth, inflammatory bowel disease, and type 2 diabetes) to understand the microbiome’s role as well as the host involvement and interaction with its microbiome. With this second phase, a deluge of data, peer-reviewed articles, and news features with various degree of sophistication continue to tackle the role of the microbiome in almost every health condition.
In parallel to the rising excitement about microbiome research, which sometimes mixes established science with publicity, a handful of applications are revitalized, notably in the use of probiotics, functional foods, and nutraceuticals, and novel research areas are taking shape. Of these, two areas of research have expanded that relate to microbial interactions with chemicals to which the host is exposed: pharmacomicrobiomics is a relatively new field that investigates the interactions between human-associated microbes and drugs (Rizkallah et al., 2010) and toxicomicrobiomics is a branch of toxicology that extends toxicogenomics to exploring xenobiotic-microbiome interactions (Aziz et al., 2018).
The Microbiome Cloud in Health and Disease
Early data collected from the HMP (Turnbaugh et al., 2007) showed that samples from different body sites (oral, skin, gastrointestinal tract, and urogenital tract) contained unique microbial communities in the same individual. Surprisingly, researchers also found that if samples were drawn from two different individuals, one individual’s oral sample will look more similar to the other’s oral sample than his or her own skin sample (Turnbaugh et al., 2007).
Unlike the human genome, the gut microbiome is far from stable all through human life. In fact it varies all the time as it responds to multiple factors, including diet and the exposure to different xenobiotics (Clarke et al., 2019).
From one’s birth throughout his/her adulthood, the human microbiome continues to shift and change in an endless cycle influenced by numerous factors. At birth, newborns were shown to have the same microbes across all of their body parts; however, based on their method of delivery, the first acquired microbes vary greatly (Dominguez-Bello et al., 2010). Babies born through vaginal delivery have Lactobacillus spp. as their first microbes, along with a typical profile of their gut microbiota and a proper breast milk digestion support. Meanwhile, babies born through C-section have Staphylococcus spp. as their first microbes, along with an abnormal gut microbiota and a predisposition to a variety of food allergies and other disorders (e.g., asthma, atopic diseases, allergic rhinitis, and celiac disease).
Studies have also shown that breastfeeding plays a vital role in developing normal microbiome. A study conducted on 107 mother-newborn pairs in 2017 found that 30% of beneficial bacteria in a newborn’s intestinal tract are acquired by seeding during breastfeeding (Pannaraj et al., 2017). A prior study in 2016 demonstrated that early antibiotic use in newborns resulted in the loss of microbial diversity and early overabundance of resistance genes in the microbiome (Langdon et al., 2016).
As healthy infants grow up, microbial communities become more diverse across different body parts, until they resemble those of an adult. However, an adult’s microbiome cloud continues to change because of other factors, such as age, diet, genetic makeup, environmental factors, physiological, and psychological changes. As a person grows older, the microbiome tends to be stable because of reduced frequency of activities and motility, lower exposure to environmental factors, chronic diseases, etc. (Spor et al., 2011). Both long-term (Wu et al., 2011) and extreme short-term (David et al., 2014) changes in dietary habits drastically change the gut microbiota. The gut-brain microbiome communication axis is affected by alteration in the psychological and physiological state of a person (Grenham et al., 2011).
Excessive use of antibiotics in adulthood was found to result in irreversible changes in the human microbiome (Dethlefsen and Relman, 2011). All of the previously mentioned factors along with many more (environmental factors, daily habits, hygiene, etc.) contribute to the loss of microbial diversity shifting the microbial cloud to a state of dysbiosis.
In a healthy body state, human-associated microbes carry their beneficial roles by protecting the body against exogenous microbes and triggering non-specific immune responses against them (protective effect), producing essential metabolites required by the human body (productive effect), and aiding in the digestion and metabolism of food and xenobiotics (digestive and metabolic effect). The microbiota is said to be in a state of symbiosis or eubiosis. A diseased body state, on the other hand, is preceded by a shift towards microbial dysbiosis. In dysbiosis, the balance between different microbes is disturbed. Often, commensal or opportunistic microbes convert to pathogenic ones via horizontal gene transfer (HGT) or a break in organ barriers causes bacteria to infect sterile body parts or opportunistic microbes taking advantage of immunocompromised patients to cause diseases (Petersen and Round, 2014).
Several studies, over the past decade, have described associations between microbiome variations and various adverse health outcomes. The microbiome has been linked to the development of a diverse set of diseases ranging from minor ailments to inflammatory and degenerative diseases (e.g., obesity (Maruvada et al., 2017), inflammatory bowel diseases (Kostic et al., 2014; Moustafa et al., 2018), stage 4 hepatitis C (Aly et al., 2016), and various cancers (Elinav et al., 2019). With any such reported association, a cause-and-effect question is raised: do microbiome variations play a role in disease causation or is it just an association that could merely be used as a biomarker for the adverse health outcome (Maruvada et al., 2017; Rosato et al., 2018). For example, causation has been demonstrated in inflammatory bowel diseases (Moustafa et al., 2018) and colorectal cancer (Tjalsma et al., 2012; Scott et al., 2019).
In either case (correlation or causation), microbiome-driven biomarkers are valuable diagnostics. In the case of mere correlation, these biomarkers would be signs of exposure to a certain adverse health outcome, while, when causation is established, microbiome biomarkers are of predictive value (e.g., Fusobacterium nucleatum (Kostic et al., 2013; Rubinstein et al., 2013) and signature microbial metabolites (Vipperla and O’keefe, 2016) as predictors of colorectal cancer).
Microbiome-Driven Xenobiotic Metabolism
With its immense diversity, the human microbiome has a powerful metabolizing capacity that even exceeds its host’s metabolic potential. The human gut microbiome, in particular, is capable of the biotransformation of xenobiotics starting from dietary compounds to pharmaceutical ingredients. The microbiome can change xenobiotics half-lives, their potential biological effect on the human body, and the rate and extent by which they reach the blood circulation or their biological targets/receptors (Koppel et al., 2017). Whether a xenobiotic is poorly absorbed and passes through the host’s small into large intestine, binds to one of the efflux proteins, or whether it is absorbed into the circulation, it will be exposed at some stage to the microbiota and its enzymes (Hall et al., 1999). The less a xenobiotic is absorbed or the more it is bound to proteins, the longer it resides in the gut and the more it is exposed to variable microbial enzymes (Hall et al., 1999). Microbial enzymes can activate prodrugs, inactivate drugs or increase or alleviate toxicity of others (Spanogiannopoulos et al., 2016; Kuntz and Gilbert, 2017; Wilson and Nicholson, 2017). The gut microbiome may also indirectly influence the host’s capacity to metabolize xenobiotics or drugs (Koppel et al., 2017).
The metabolic enzymatic capacity of the gut microbiota is clearly distinguished from the host’s metabolic capabilities to a degree that the microbial metabolism of xenobiotics is sometimes opposite to the host biotransformation. Host enzymes mainly carry out oxidation and conjugation, while microbial enzymatic reactions are mainly reduction and hydrolysis (Spanogiannopoulos et al., 2016; Wilson and Nicholson, 2017). Demethylation is another example of discrepancies between host and microbiome xenobiotic metabolism: whereas the host demethylation renders a xenobiotic more polar to be excreted outside the body, for microorganisms, demethylation generates carbon sources for further growth and division (Sutton et al., 1997; Kumano et al., 2016).
The microbial enzymatic repertoire in the human gut is complex and consists of divergent enzyme types, such as beta-glucosidases, beta-glucuronidases, aryl sulfatases, azoreductases and nitroreductases (Table 1). The human microbiome contributes to interindividual variations, not only because of the diversity of microbial types within an individual, but also because microbial enzymes differ from one species to another in sequence, activity, and abundance (Spanogiannopoulos et al., 2016; Wilson and Nicholson, 2017).

Table 1 Some gut bacterial enzyme families encoded in the microbiome and their impact on xenobiotics.
Hydrolysis, lyase-catalyzed bond breakage, reduction, and group transfer are some of the chemical reactions that gut microbial enzymes can catalyze (Koppel et al., 2017). Glycosidases, proteases, and sulfatases are among microbial hydrolytic enzymes. Glycosidases break the glycosidic bonds using water to release sugars (Kallemeijn et al., 2014). The xenobiotic molecule becomes less polar and thus acquires longer half-life in the human body which may increase it toxicity as seen in the non-steroidal anti-inflammatory drugs as diclofenac and ketoprofen (Saitta et al., 2014). Proteases break the bonds between amino acids in a polypeptide chain with different specificities (Tozaki et al., 1997). Phase II host metabolism results in sulfate ester which is broken down by sulfatases (Ulmer et al., 2014).
The bond cleavage, which neither requires oxygen nor water, is catalyzed by lyases. Microbial C–S beta-lyases break the carbon-sulfur bonds and the ammonia generated is the nutrient for bacterial growth (Rossol and Puhler, 1992). Reduction plays a role in opposing host metabolism mechanisms such as rendering xenobiotics less polar or drugs to either their inactive or active forms (Peppercorn and Goldman, 1972; Lavrijsen et al., 1995; Haiser et al., 2013). Methyl or acyl groups are transferred from or to xenobiotics by microbial transferase enzymes (Koppel et al., 2017). Acylation of 5-aminosalicylic acid, for example, leads to inactivation of the drug (Delomenie et al., 2001).
The presence or absence of a gut microbiota can have an indirect effect on regulating host metabolizing enzymes, either through competition of the microbial metabolites on host enzymes or through changing the expression of these enzymes. When gnotobiotic mice were studied in comparison to regular mice, the expression of liver enzyme genes, including active androstane receptor, was upregulated in germ-free mice (Bjorkholm et al., 2009). Expression levels of CYP3A (cytochrome P450 family 3 subfamily A) in mice decreased upon administration of ciprofloxacin antibiotic. This downregulation was attributed to the gut bacteria synthesizing lithocholic acid, which was hypothesized to affect CYP expression (Toda et al., 2009). In germ-free mice, transcription and translation of CYP3a11, the mouse homolog of human CYP3A4, were significantly lessened. However, gene expression of both CYP1a2 and CYP4a14 was elevated in germ-free mice (Selwyn et al., 2016).
Elemicin is an essential oil constituent incorporated into food and dietary supplements, which upon activation by CYP1A2 (the human homolog of mouse CYP1a2), is converted to more potential cytotoxic metabolite (Wang et al., 2019). CYP1A2 affects potent medications as theophylline, psychological disorders medications, warfarin, and other anticancer drugs (Wishart et al., 2018). CYP4a14 is the homolog of human CYP4A which is mainly responsible for arachidonic acid and fatty acids metabolism and may play a crucial role in prognosis of nonalcoholic fatty liver disease (Zhang et al., 2017b). Gene expression regulation of some of the host detoxifying enzymes such as glutathione peroxidases, sulfotransferase, epoxide hydrolases, and N-acteyltransferases was shown to be affected by the absence of gut bacteria (Meinl et al., 2009).
Gut microbiota-derived metabolites of steroid hormones, uremic solutes of dietary protein breakdown or herbal medicines, or bile acids may decrease CYP3A4 expression (Bjorkholm et al., 2009; Devlin et al., 2016). Low CYP3A4 results in less clearance of multiple CYP3A4-dependant medications, e.g., erythromycin and verapamil, affecting their action and adverse effects (Barnes et al., 2014).
Pharmaceutical Xenobiotics
The use of pharmaceutical products is undoubtedly on the rise over the past decades. According to the Global Use of Medicines report from the IQVIA Institute for human data science, the global market for pharmaceuticals reached $1.2 trillion in 2018, up $100 billion from 2017 (The Iqvia Institute, 2019). With the availability of pharmacies around every corner offering over-the-counter pharmaceutical products and the increasing number of patients with chronic illnesses, using such products becomes an integral part of our everyday lives. Effects of pharmaceuticals on the human body have long been studied, but with the importance of the microbiome lately being rediscovered, study results are accruing on the mutual interplay between pharmaceuticals and the microbiome.
As previously established, different body sites (oral, skin, gastrointestinal tract and urogenital tract) contain unique microbial communities (Turnbaugh et al., 2007), each of which interacts with pharmaceuticals in a different way (affecting and being affected by them). Some interactions may lead to an increase or a decrease in activity, while others result in increased toxicity of a pharmaceutical xenobiotic (Table 2), which fall within the scope of toxicomicrobiomics.

Table 2 Adverse effects/toxicities resulting from xenobiotic-microbiome interactions (pharmaceutical toxicomicrobiomic interactions).
Gut Microbiome
As defined earlier, the gut microbiota, which is the largest microbial community in the human body, contains trillions of microorganisms (bacteria, archaea and micro-eukaryotes), including more than 1,000 different species of bacteria (Bacteroidetes and Firmicutes being the two major phyla in most cases), with more than 3 million genes. The gut microbiota is characteristic in each individual, and only one third of it is shared between different people. Studies on the gut microbiota constitute more than 80% of the overall publications on human microbiome (Cani, 2018), which reflects both the importance of its role in human health and the booming research in that area.
A classic case of microbiome-driven modulation of drug pharmacology and toxicity is the reduction of the cardiac glycoside, digoxin, by some strains of Eggerthella lenta, resulting in an altered metabolism, concentration and toxicity (Haiser et al., 2013). This differential reduction explains earlier reports of variability in digoxin metabolism between North Americans and southern Indians, who had variable abundance of the microorganism (Mathan et al., 1989). Another classic case is that of acetaminophen (Table 2), an analgesic, antipyretic and weak anti-inflammatory drug. Acetaminophen’s variable toxicity among individuals is due to variability in the amounts of p-cresol, produced by some gut bacteria, and which competes with acetaminophen’s o-sulfonation in the liver (Clayton et al., 2009). Current guidelines recommend assessing the microbiome activity before the administration of acetaminophen.
Ranitidine, an H2 Blocker, was found to be differentially degraded depending on the presence of gut bacteria with ability to cleave an N-oxide bond (Basit and Lacey, 2001). Insulin, an anabolic hormone and a peptide drug, was found to be degraded by some proteolytic enzymes produced by the gut microbiota, rendering it ineffective (Tozaki et al., 1997). Metformin, an anti-diabetic drug, was found to be more effective orally than parenterally because of gut-based pharmacological effects (Napolitano et al., 2014).
Skin Microbiome
The human skin is the largest body organ and it plays an essential role in the body’s natural immunity together with its associated microbial communities. The skin microbiota has more than 19 major phyla (Grice et al., 2009) and defends the human body against pathogens. These defense mechanisms include releasing antimicrobial peptides, maintaining pH and competing with pathogens for available nutritional resources.
Owing to its direct contact with the environment, the skin microbiota continuously interacts with, and thus adapts to, environmental xenobiotics. Studies have recently unveiled the role of topical xenobiotics on skin microbiota. For example, a study conducted on infants revealed that emollients resulted in decreased skin pH and increased bacterial richness, diversity, and abundance of Streptococcus salivarius, which seemed to improve atopic dermatitis treatment (Glatz et al., 2018). Chitosan is a natural biopolymer with immunomodulatory and antimicrobial properties. The inclusion of chitosan in coating textiles may reduce atopic dermatitis severity by modulating skin staphylococcal communities (Lopes et al., 2015). Additionally, topical treatment with coal tar in atopic dermatitis was found to decrease the abundance of Staphylococcus aureus and increase that of Propionibacterium on the skin, shifting the microbiome into a healthier state (Smits et al., 2019).
Urogenital Microbiome
The female urinary system is sterile in its normal state as a result of the urea content, which inhibits microbial growth, the mechanical flushing of pathogens by urine, and the lactic acid and hydrogen peroxide secreted by vaginal lactobacilli.
Like with other body sites, the urogenital microbiota mutually interacts with pharmaceutical xenobiotics. For example, tenofovir, an antiviral used in the treatment of HIV, was found to be metabolically depleted in African women by Gardnerella vaginalis and other anaerobic bacteria (Klatt et al., 2017). Combined oral contraceptives were shown to be associated with increased vaginal colonization by healthy lactobacilli, and with reduced bacterial vaginosis-associated taxa in another study (Brooks et al., 2017). Contraceptive rings were found to decrease the severity of bacterial vaginosis in African women by promoting the growth of vaginal lactobacilli and decreasing the abundance of Gardnerella vaginalis and Atopobium vaginae (Crucitti et al., 2018).
Oral Microbiome
The interaction between the oral microbiome and xenobiotics has long been studied as the microbial community of the oral cavity is constantly subjected to habits of eating, drinking and smoking that affect and change the oral microbiome on a daily basis.
In a recent study, silver, titanium dioxide, and iron nanoparticles were found to have the lowest inhibitory and antibiofilm concentration against dental caries-causing bacteria like Streptococcus mutans and Streptococcus sanguinis, when compared to other nanoparticle-containing solutions, antibiotics, and chlorhexidine (Lavaee et al., 2016). Hyaluronic acid treatment in peri-implantitis was found to decrease the severity of the disease by decreasing the abundance of colonizing bacteria (Soriano-Lerma et al., 2019). The antibacterial and antibiofilm activity of abietic acid on Streptococcus mutans growth was recognized as a potential protective measure against dental caries (Ito et al., 2019b). Of note, silver nanoparticles have complex interactions with mucosal and intestinal microbiomes (reviewed in Bi et al., 2020).
Food Additives and Dietary Xenobiotics
Food is the source of energy and nutrients for the human body to grow, develop, and carry on everyday activities. The global human population is increasing over time, with incremental needs for ongoing food resources (Eshel et al., 2014). With this large demand, people have incorporated much more processed foods and artificial products into their diet to keep up with the rapid pace of their lifestyles. The enzymatic reserve of the gut microbiota has proven to play a pivotal role in metabolizing dietary compounds of natural, processed, or even artificial origins and making use of their metabolized products as a source of carbon or energy, not only for the microbiota itself, but also for the human host (Sharon et al., 2014).
Cooking or processing food may release undesirable xenobiotics that are not originally present in raw food such as polycyclic aromatic hydrocarbons, heterocyclic amines, and nitrosamines (Nogacka et al., 2019). The genotoxic consequences of the aforementioned xenobiotics are inextricable for both human and microbial genetic material (Nogacka et al., 2019). The gut microbiome may protect against the carcinogenic and genotoxic effects of these xenobiotics by biotransforming them to less toxic compounds or facilitating their excretion (Vanhaecke et al., 2008; Carmody and Turnbaugh, 2014). On the other hand, the gut microbiota may also transform heterocyclic amines, for example, into genotoxins, or may reverse the detoxification implied by host metabolism (Kassie et al., 2001). MeIQx (2-amino-3,8-dimethylimidazo[4,5-f]quinoxaline), one of the heterocyclic aromatic amines that are produced during red meat cooking, is believed to be associated with colorectal cancer. The gut microbiota is capable of transforming MelQx to MelQx-M1 which is of lower toxicity and mutagenicity to colon cells, thus it may be able to lessen the chances of cancer (Zhang et al., 2017a). The outcome of these diverse and sometimes contradictory effects will vary from person to person, and further studies in animal models and in vitro systems should address the effect of different microbial communities on a mixture of those chemicals.
On another front, the gut microbiome might be negatively affected by food additives, such as E171 or titanium dioxide. A recent study, conducted to observe the effect of E171 on the intactness of the colonic epithelium, showed that E171 altered the metabolites produced by the gut bacteria and helped forming a biofilm (Pinget et al., 2019). The imbalances in mucus layer thickness and epithelial permeability are involved in the development of colitis and colorectal cancer (Kim, 2014).
Non-caloric artificial sweeteners are currently consumed to give a sweet taste with much fewer or no calories. In one experiment, saccharin, sucralose, or aspartame were administered to mice in their drinking water. Consequently, mice developed marked glucose intolerance with distinct altered microbial composition (Suez et al., 2015). Xylitol, another food additive used to protect against caries, is capable of changing gut microbiota in mice, especially decreasing phylum Bacteroidetes, and enriching phylum Firmicutes and genus Prevotella (Uebanso et al., 2017). Benzoic acid is a preservative added to food to prevent bacterial and fungal growth. Dietary and supplement benzoic acid were proved to reduce Lactobacillus and E. coli in pig intestines (Mao et al., 2019). Emulsifiers, such as carboxymethylcellulose and polysorbate-80, caused microbial dysbiosis in mice and promoted low-grade inflammation which is one of the causes of metabolic syndrome (Chassaing et al., 2015).
Environmental Chemicals and Pollutants
Environmental pollutants and chemicals have become an integral part of the surroundings of people. It is becoming necessary to include the microbiome in considering the safety and toxicity of these compounds, as microbial enzymes can alter their half-life and dynamics (Koppel et al., 2017). Melamine is used as raw material for manufacturing adhesives, paints, tables, and engineered wood. Melamine traces were found in soil and wastewater close to melamine factories (Wang et al., 2014a). In 2008, melamine toxicity led to the death of children because of gut microbial transformation of melamine to cyanuric acid. The latter form is an insoluble complex that precipitates in the kidney and causes kidney stones and is biotransformed by Klebsiella terrigena, together with Pseudomonas spp. strain A (NRRL B-12227), Rhodococcus corallinus (NRRL B15444R), Klebsiella terrigena DRS-1 (ATCC 700372), Rhodococcus sp. strain Mel, Nocardioides sp. strain ATD6, and the novel bacterium CY1 or Melaminivora alkalimesophila gen. nov., sp. nov (Zheng et al., 2013; Wang et al., 2014b).
Halogenated compounds are another class of chemicals used in agriculture as pesticides, pharmaceuticals, bleaching and disinfecting agents. They possess a negative impact on the gut microbiota, changing the Firmicutes-to-Bacteroidetes ratio to a dysbiotic one, and hence exposing humans to obesity or immune system perturbations. The gut microbiome is capable of nitroreductive metabolism and reductive dehalogenation for organohalogens (Atashgahi et al., 2018). One of these compounds, 2,4-D (2,4-dichlorophenoxyacetic acid), is one of the most commonly used herbicides around the world (Peterson et al., 2017). It disrupts hormonal pathways in plants and eventually leads to plant death. Hence, it is used to control weeds in agriculture and at homes. In a study to observe the effect of 2,4-D on mouse gut microbiota, a state of dysbiosis was observed with an occupational-dose exposure in 2,4-D-treated mice. Dysbiosis was not only on the taxonomic level—in terms of alpha diversity and richness, but it was also strongly manifested on the microbial metabolic pathways and metabolites. Among the microbial metabolic pathways affected was the metabolism of urea, amino acids, and carbohydrates (Tu et al., 2019).
Polychlorinated biphenyls (PCBs), albeit banned in 1977 for their toxic effects, are still present in the environment (Centers for Disease Control and Prevention (CDC), 2009). Exposure to PCBs is mainly through ingestion and may lead to impairment in the neurological functions and behavior especially in children (Rude et al., 2019). In an animal study, two groups of mice were exposed to PCBs, a wild type group and another with a mutation in RyR1 and FMR1 genes. PCBs caused a state of dysbiosis, increased inflammatory state, and a more permeable gastrointestinal barrier in the offspring especially in the group with genetic mutations. The latter group also was more predisposed to develop defects in neurobehavioral development (Rude et al., 2019).
Interestingly, the microbiome composition of a pregnant mother varies upon her exposure to PCBs, which also leads to a variation in the gut bacterial community of her fetus (Laue et al., 2019). In that case, the fetal exposure to PCB shifted the children’s microbiome towards order Bacillales and family Propionibacteriaceae, a shift that lasted up to eight years after their birth (Laue et al., 2019).
Another example highlighting the combined power of environmental exposure, individual genetic makeup, and gut microbiota dysbiosis on human health is the effect of organophosphate pesticide chlorpyrifos (CPF). In a mouse model with different apolipoprotein E (APOE) genotypes, in which CPF was administered after birth, a distinct taxonomical pattern was observed after exposure. APOE genotype and CPF exposure diversely affected the types of short-chain fatty acids in the brain (Guardia-Escote et al., 2020). A correlation had previously been established between gut and brain short-chain fatty acids, with observed impact on brain functionality (Sun et al., 2016).
Glyphosate is as a weedicide used around the globe (Green, 2018). Whether alone or in combination, glyphosate significantly changes the gut microbial niche through shifting the abundance of its two main phyla, Bacteroidetes and Firmicutes. Consequently, it affects brain plasticity, maternal behavior, and parenting (Dechartres et al., 2019).
Disruptions in microbial processing of amino acids and carbohydrates affect the balanced state of the host amino acids and carbohydrate metabolism, and may contribute to metabolic diseases, such as type 2 diabetes mellitus or obesity (Flint et al., 2008; Neis et al., 2015). On the other hand, microbial urease activity has been associated with diseases of the gastrointestinal tract such as ulcers or stomach cancer (Mora and Arioli, 2014).
Arsenic is a naturally present metalloid that poses its toxic effects on human health especially in its inorganic form. It is a potential cancer-causing chemical for lungs, kidneys, liver, skin, and bladder. It was proved to negatively affect cardiovascular, neurological, and immunological functions (Naujokas et al., 2013). An investigation of the role of the gut microbiota in arsenic metabolism, conducted in mice, showed that gut dysbiosis enhanced the toxic effects of arsenic, one way through increasing arsenic load and the other through promoting one-carbon metabolism. However, a balanced gut microbiota absorbs arsenic and promotes its methylation (Chi et al., 2019). In addition, dysbiosis may alter the expressions of some genes involved in p35 signaling and many genes involved in hepatocellular carcinoma development (Chi et al., 2019). While arsenic by itself may disturb the gut microbial community (Lu et al., 2014), Bacteroides, Clostridium, Alistipes, Bilophila, and other bacterial genera are able to methylate arsenic and can survive exposure to it owing to multiple arsenic resistance genes encoded in their genome (Yin et al., 2015; Yu et al., 2016).
Cadmium and lead are inorganic heavy metals that are not degraded and tend to accumulate in an ecological system (Daisley et al., 2019). They enter the human body through ingestion of contaminated food, water, and even particles cleared by the respiratory system (Satarug et al., 2003). Pesticides, fertilizers, corrosion of lead-containing plumbing, mining, lead batteries, and burning of coal are different sources of lead and cadmium pollution (Wuana and Okieimen, 2011; Li et al., 2015; Daisley et al., 2019). Human exposure to cadmium may cause hypertension, diabetes, myocardial infarction, and impaired kidney function, and may thus imbalance minerals homeostasis (Satarug et al., 2010). Lead can lead to neurotoxicities by affecting neurotransmitters in the nervous system and interfering with calcium in mitochondria, hence increasing oxidative stress and mitochondrial death (Mason et al., 2014). Lactobacillus rhamnosus GR-1 was shown to minimize human exposure by binding or sequestering heavy metals and consequently improving their excretion in feces (Daisley et al., 2019). As a result, lactobacilli are considered a safe cost-effective solution to decrease exposure to heavy metals and their severely negative potential on human health (Daisley et al., 2019).
Azo compounds are widely used in pharmaceutical, textile, food, cosmetics production. Azodyes may cause mutations in the human DNA, allergic reactions and cancer (Gičević et al., 2020). The gut microbiota expresses azoreductase enzymes to metabolize azodyes. Among azo-reducing bacteria are Clostridium, Pseudomonas, Bacillus, Geobacillus, Lysinibacillus, Enterococcus, Eubacterium, and Escherichia (Misal and Gawai, 2018). Escherichia coli, Enterococcus faecalis, Enterococcus avium, and Bacillus cereus/thuriengensis have been recently isolated from human stool samples with variable azoreduction rates (Zahran et al., 2019).
Triclocarban (3,4,4,9-trichlorocarbanilide, TCC) is an antibacterial ingredient in personal care products such as toothpaste and hand soap (Halden et al., 2017). Eventually, it integrates into the environment through wastewater as a persistent non degradable organic pollutant (Petrie et al., 2015). TCC was recently shown to shift the gut microbial composition to an increased Firmicutes and Lactobacillus taxa, and decreased Bacteroidetes in low-dose TCC-exposed rats. On the microbial metabolic level, TCC increased short-chain fatty acids concentrations suggesting that TCC enhanced bacterial fermentation. These changes play a role in the synthesis of fatty acids by the liver either directly or indirectly (Poole et al., 2016).
Triclosan (5-chloro-2-(2,4-dichlorophenoxy)phenol, TCS) is another broad-spectrum antimicrobial added to personal use products including cosmetics and toys (Halden, 2014; Poole et al., 2016). In a recent study on mice, TCS affected beneficial gut microbiota such as Bifidobacterium. TCS treatment also caused colonic inflammation in conventionally raised mice, supporting the evidence that gut microbiota plays a role in TCS-caused inflammation (Yang et al., 2018).
Studying Toxicomicrobiomics
Tools and Techniques
The tools for studying toxicomicrobiomics, as well as pharmacomicrobiomics, are no different from the general tools used to decode the microbiome at different levels: its taxonomic and genomic composition, gene content, functional potential, actual expression of its genes (at the RNA or protein level), and finally its actual function as reflected by the combined microbiome-derived metabolites. As such, microbiome profiling by amplicon sequencing, shotgun microbiome sequencing or metagenomics, metatranscriptomics, metaproteomics, as well as metabolomics and metabonomics are all valid tools for the analysis of xenobiotic-microbiome interactions (Table 3).
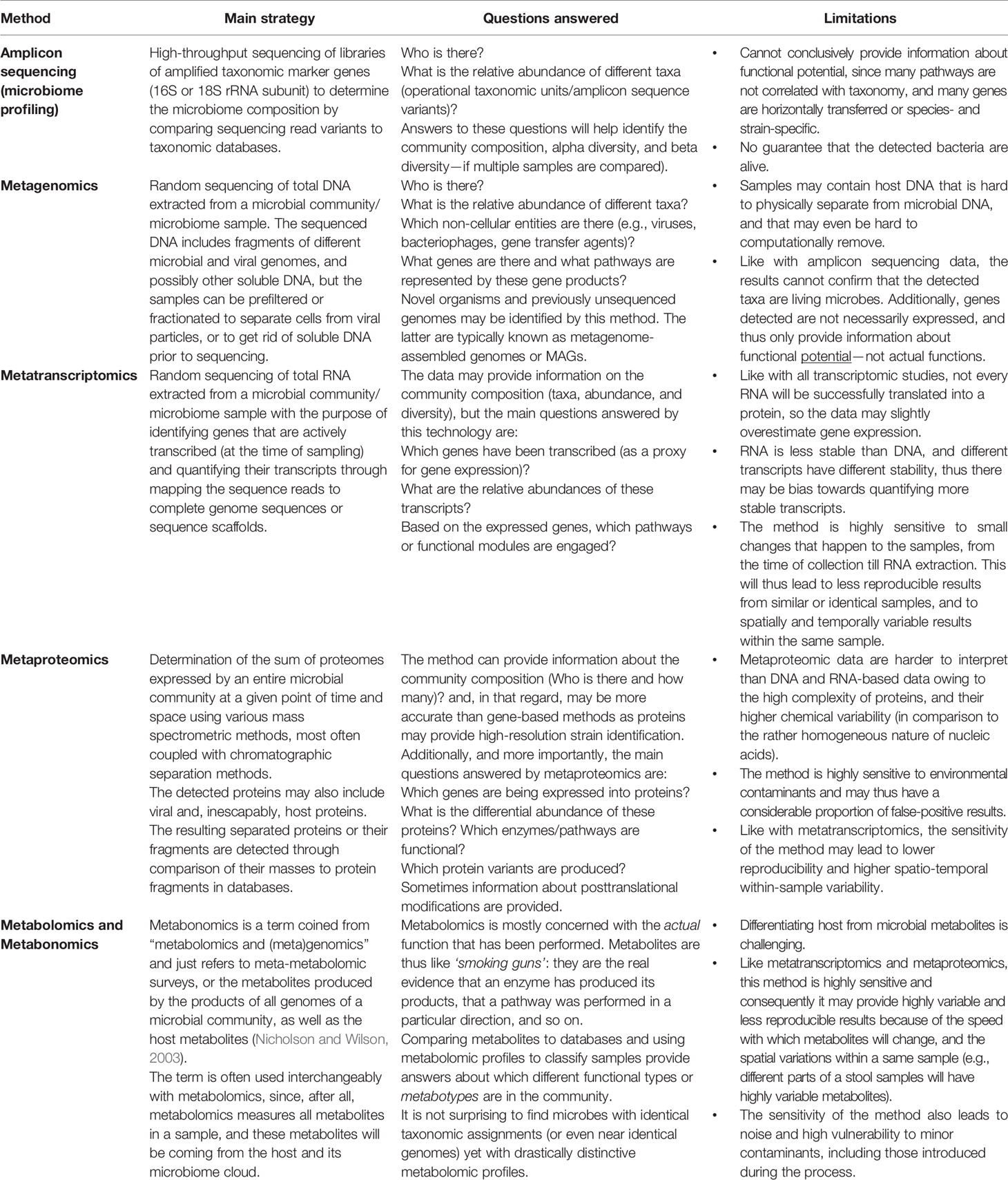
Table 3 Omics tools and technologies used in toxicomicrobiomic research.
Experimental Strategies for Discovery of Novel Xenobiotic-Microbiome Interactions
Using the above-mentioned ‘omics analysis technologies (Table 3), systematic studies need to be conducted to unearth new toxicomicrobiomic interactions. Strategies for these studies should benefit from environmental metagenomic approaches that seek to discover, for example, microbes and enzymes with biodegradation capabilities (Sayed et al., 2014; Bouhajja et al., 2016; Ngara and Zhang, 2018), or from pharmacomicrobiomic discovery methodologies (Bisanz et al., 2018). A remarkable example is the recent high-throughput systematic pharmacomicrobiomic screens to detect specific drug-bacterial interactions and separate them from drug-host interaction (e.g., Zimmermann et al., 2019a; Zimmermann et al., 2019b). Overall, experimental strategies, combining (wet) laboratory and computational (in silico approaches) may be classified into culture-based, sequencing-based, functional screening-based, and innovative hybrid strategies.
Culture-Independent Sequencing-Based Methods
● Amplicon sequencing: Using amplicon sequencing to determine the microbiome profiles has been coupled to different in silico function prediction tools [e.g., PICRUST (Langille et al., 2013) and FishTaco (Manor and Borenstein, 2017)], although functional prediction from taxonomic markers remains of limited reliability (Table 3, limitations). Once a set of taxa have been predicted as xenobiotic metabolizers, they can be suggested as biomarkers and as targets for further culture-based or hybrid approaches (which could be systems-based or reductionist studies focusing on a representative of each taxon, or combinations thereof).
● Shotgun metagenomics: Shotgun sequencing provides the benefit of sequence-based discovery of enzymes and pathways [e.g., biosynthetic or biodegradation operons (Mohamed et al., 2013; Ziko et al., 2019)]. The discovery of cardiac glycoside-degradation genetic locus was mostly based on shotgun and metatranscriptomic sequence analysis (Haiser et al., 2013). This strategy has the advantage of gene-level analysis, which allows not only gene and pathway identification, but also the ability to determine closely related gene variants. However, like all DNA-based methods, there is no guarantee that sequenced genes are ever expressed in the environment from which they were sampled, which may mislead researchers. For example, a novel detoxifying enzyme-coding gene, sequenced from the oral microbiome, may suggest that its encoded enzyme acts at a certain pH or salt concentration, when the actual enzyme may never be expressed in the oral cavity, and thus may have no bioremediation value.
● PCR libraries: Here, instead of sequencing all metagenomic DNA, the DNA is amplified with primers (often degenerate) to amplify members of gene families that encode the desired protein (e.g., enzyme, resistance factor, efflux pump). The generated PCR product is then extracted, used to build a library (usually a plasmid library), propagated, then sequenced. This strategy helps determining a large number of possible variants of the desired gene family within the microbial community. This approach was used, for example, to discover and characterize novel thermostable mercuric reductases (Maged et al., 2019). As with all DNA-based approaches, the discovery of any valuable gene with this strategy is no guarantee that the gene is actually expressed in its cell’s natural habitat.
Culture-Based Strategies
● Culturing the microbiome
1. Advances are continuously being made to optimize various aerobic and anaerobic culture methods and technologies to grow members of the human microbiota, with emphasis on those that have been long considered as yet-uncultured or uncultivable (Forster et al., 2019; Ito et al., 2019a).
2. Variations on this technology is the use of in vitro models for body sites or tissues, also considered as ex vivo models, e.g., fermentation chambers or reactors that simulate the human gut (Molly et al., 1993; Minekus et al., 1999; Takagi et al., 2016).
3. An innovative technological breakthrough, which is a variation of the same concept of simulating body sites, is the iChip, developed to isolate bacteria from their natural habitats (Nichols et al., 2010; Berdy et al., 2017). It is possible, in the future, to see similar in situ technologies to isolate bacteria from human stool or human mucus/body fluids. These iChips may eventually evolve into smaller miniaturized, nano-scale, or robotic devices.
● Systematic screens: These are simple yet labor-intensive high-throughput screens, in which different resident bacteria (usually directly isolated from human body sites: gut, skin, vagina) are co-cultured with a set of xenobiotics (drugs, heavy metals, environmental toxins), and the chemical changes/modifications to these xenobiotics are detected and quantified via chromatographic separation followed by spectroscopic methods. A typical example is the recent impressive work of Zimmerman and coworkers (Zimmermann et al., 2019a).
All culture-based methods offer great advantage as they allow actual experimentation and screening; however, this strategy remains biased as it enriches our knowledge about many already well-studied organisms. Culture-independent technologies were specifically developed to overcome this type of knowledge bias.
Function-Based (Functional Metagenomics Screening)
● In function-based metagenomic approaches, total DNA of environmental samples is extracted, fragmented, and expression libraries (in cosmids, fosmids, or other plasmids) are probed and screened for desired functions by positive or negative selection (Ngara and Zhang, 2018; Liu et al., 2019). These approaches offer the great advantage of actual expression of proteins, which allows testing their functionality, even if their producing microbes cannot be successfully cultured. For example, metagenomic libraries from soil, wastewater, river estuaries, or extreme environments (e.g., Red Sea brine pools (Mohamed et al., 2013; Sayed et al., 2014)) have been a good source for bioprospecting novel enzymes with ability to function at extreme conditions, or for the discovery of novel antibiotic or heavy-metal resistance genes via negative selection. This approach, which is well-established in applied and environmental microbiology, has been explored in screening the gut microbiome, for example, for resistance genes (Pehrsson et al., 2016) and biodegrading enzyme-coding genes (Cecchini et al., 2013). One limitation of this approach is the limited number of possible surrogate hosts (e.g., E. coli, Lactococcus, and yeast); however, anaerobic systems that are most relevant to the gut microbiota are being developed (e.g., in Bacteroides (Lam et al., 2018)).
Hybrid Methods (Sequence/Culture Based)
● Sequence-guided microbial augmentation: Recently a composition-guided approach has been adopted to better define the structure and function of microbial communities at polluted environments. In an innovative step to precision bioremediation, Redfern et al. (2019) combined amplicon-based microbiome analysis and chemical profiling of polluted sites to improve the biodegradation capabilities of targeted microbial consortia. Microbial communities of environmental samples collected from several locations with different contamination levels were taxonomically classified by high-throughput sequencing. This novel framework relied on establishing relationships between chemicals and taxa to select candidates for biostimulation, bioaugmentation and genetic bioaugmentation. Specifically, the biostimulation target, Geobacter, documented as pollutant degrader, was detected at high abundance at all the sampling sites. The bioaugmentation targets comprised a K-strategist, Mycobacterium, and an r-strategist, Thauera, to guarantee rapid and long-term bacterial adaptation. The genetic bioaugmentation target, Sphingomonas, possessed the ability to receive catabolic plasmids from augmented donors (Redfern et al., 2019).
● Reverse genomics: Another quite recent approach is the use of reverse genomics to identify surface antigenic markers for hard-to-culture bacteria, which is a reverse vaccinology-inspired approach (Cross et al., 2019). These in silico-identified specific antigens are then cloned and expressed in heterologous systems, used to generate antibodies, which are then purified and deployed to capture the desired bacteria in their natural microbiome habitat, even at a single-cell level. Such microbial enrichment makes culturing the rarest strains possible (Cross et al., 2019).
● Finally, a hybrid approach that combines high-throughput systematic screens, functional metagenomics, and high-throughput sequencing was developed and adopted to discover plant polymer-decomposing enzymes (Nyyssonen et al., 2013). This approach, which integrates several of the above-mentioned strategies, uses multiplexed fluorometric and colorimetric assays to screen metagenomic libraries, and only sequences DNA inserts with promising desired phenotypes. The approach can easily be adapted to human microbiome samples, notably fecal samples, to discover xenobiotic-metabolizing enzymes, notably from organisms that cannot be cultured yet.
Challenges and Future Aspects
Accruing reliable information about an individual’s microbiome may aid in preventing and treating certain diseases (Kosyakovsky, 2017). It may also lead to a decrease in the toxicity of some medications or the improvement in the efficacy of others (Haiser and Turnbaugh, 2012). A comprehensive “microbiome panel” may eventually be available as a routine laboratory screening to provide a microbiome profile of an individual and help defining the various options for a “healthy” microbiome. Once variants of healthy microbiomes and their biomarkers are defined, biomarkers for dysbiosis and disease susceptibility may also be defined. Consequently, xenobiotic-degrading bacterial taxa (or biomarkers thereof) may also be easily analyzed in the near future, and may be used in several therapeutic and toxicological decisions.
The highly disparate species and subspecies in the human microbiome interacting with every system in the body, including the immune system, raised the question about a core microbiome (Kuntz and Gilbert, 2017; Lloyd-Price et al., 2017). Core metabolic pathways are of more concern to comprehend the function and adaptation of the microbiome in a certain body location (Cardona et al., 2016). Researchers favor shotgun metagenomics to 16S rRNA gene profiling to analyze the whole genome of the microbial community (Cardona et al., 2016), which provides more information toward better deciphering of host-microbiome metabolic capabilities and interactions (Table 3). However, shotgun metagenomics is of higher cost and more complexity. In addition, achieving reliable personalized treatment based on pharmacomicrobiomics (and likewise toxicomicrobiomics) requires adopting standard reproducible approaches (Kashyap et al., 2017).
These challenges require multidisciplinary national and international collaborations to integrate big data, driven from multi-omics, to put pharmacomicrobiomics and toxicomicrobiomics into clinical applications. Microbiome studies should include populations with divergent geography and ethnicity (Lloyd-Price et al., 2017). Data and metadata of microbiome studies should be freely accessed over the time, according to peer-reviewed journal guidelines, to allow standardization and validation of bioinformatics and statistical analyses (Langille et al., 2018).
In this final section of the review, we propose ideas for practitioners, clinicians, and investigators in the field of microbiome research and its impact on human health.
Open-Access Curated Databases
There is a scarcity of databases of ready curated knowledge for extraction by practitioners and investigators which may cost valuable time, resulting in hindering of the research or deterring researchers from conducting any investigation in the field. For this reason, building databases with such curated knowledge or helping already established databases as a volunteer team member, crowdsourcer or crowdfunder is a must for the advancement of the field. Specifically, linking toxicology databases, chemical databases, and drug banks to pharmacogenomic and pharmacomicrobiomic data resources is becoming a pressing need
Predictive Models and Software Development
Building predictive models and developing software for simulating drug-microbiome interactions are essential as they will strongly reduce cost and researchers time. The only limitation to develop such models is the availability of curated knowledge, from which robust hypotheses can be driven and used for training and validating the model. Chemical similarity searches, if combined to gene/protein similarity searches, may allow the prediction of novel interactions between xenobiotic chemical moieties and different microbial enzymes.
Microbiome Biomarkers as Future Non-Invasive Tools in Precision Medicine
As detailed above, the human microbiome is a rich source for discovery of biomarkers for diseases and adverse health outcomes. Using such biomarkers as non-invasive alternatives to biopsies and endoscopy, for example, would be a promising futuristic diagnostic (Ren et al., 2019) and predictive/preventive tool (Penalver Bernabe et al., 2018). For example, the human microbiota was found to release molecules and metabolites that affect the onset, progression, and treatment of cancer (Scott et al., 2019; Rao et al., 2020). True personalized cancer therapy may only be achieved through a full understanding of the effect of microbiome on chemotherapy (Table 2) and tumorigenesis, thus allowing proper patient stratification—based on specific biomarkers, microbiome types, and metabotypes (Rizkallah et al., 2010; Kuntz and Gilbert, 2017). Although inter-individual microbial diversity, emergence of novel and multiresistant microbial strains, and various microbial drug modification mechanisms may complicate or delay precision medicine, it remains the right choice for future theranotstics.
Clear Guidelines for Clinical Practice
Hopefully, the availability of online resources, biomarker discovery, development of assays, models, and software, and conducting more research will help practitioners and investigators to develop novel guidelines for clinical practice, and to implement guidelines for some of the well-established xenobiotic-microbe and xenobiotic-microbial enzyme interactions.
Author Contributions
RA conceived and conceptualized the article. All authors collected literature. NA and AR screened literature, made the final selection of reviewed articles, and drafted the article. ME reviewed toxicomicrobiomic strategies and tools. RA revised the draft and wrote the article in its final format. All authors read and approved the final format.
Funding
RA and ME are funded by the Egyptian Academy for Scientific Research and Technology (ASRT) JESOR program (project # 3046); however, the funders had no input in the content of this article.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Aly, A. M., Adel, A., El-Gendy, A. O., Essam, T. M., Aziz, R. K. (2016). Gut microbiome alterations in patients with stage 4 hepatitis C. Gut Pathog. 8, 42. doi: 10.1186/s13099-016-0124-2
Atashgahi, S., Shetty, S. A., Smidt, H., De Vos, W. M. (2018). Flux, impact, and fate of halogenated xenobiotic compounds in the gut. Front. Physiol. 9, 888. doi: 10.3389/fphys.2018.00888
Aziz, R. K., Hegazy, S. M., Yasser, R., Rizkallah, M. R., Elrakaiby, M. T. (2018). Drug pharmacomicrobiomics and toxicomicrobiomics: from scattered reports to systematic studies of drug-microbiome interactions. Expert Opin. Drug Metab. Toxicol. 14, 1043–1055. doi: 10.1080/17425255.2018.1530216
Aziz, R. K. (2018). Toxicomicrobiomics: narrowing the gap between environmental and medicinal toxicogenomics. Omics 22, 788–789. doi: 10.1089/omi.2018.0183
Barnes, K. J., Rowland, A., Polasek, T. M., Miners, J. O. (2014). Inhibition of human drug-metabolising cytochrome P450 and UDP-glucuronosyltransferase enzyme activities in vitro by uremic toxins. Eur. J. Clin. Pharmacol. 70, 1097–1106. doi: 10.1007/s00228-014-1709-7
Basit, A. W., Lacey, L. F. (2001). Colonic metabolism of ranitidine: implications for its delivery and absorption. Int. J. Pharm. 227, 157–165. doi: 10.1016/S0378-5173(01)00794-3
Basivireddy, J., Jacob, M., Ramamoorthy, P., Balasubramanian, K. A. (2005). Alterations in the intestinal glycocalyx and bacterial flora in response to oral indomethacin. Int. J. Biochem. Cell Biol. 37, 2321–2332. doi: 10.1016/j.biocel.2005.05.010
Berdy, B., Spoering, A. L., Ling, L. L., Epstein, S. S. (2017). In situ cultivation of previously uncultivable microorganisms using the ichip. Nat. Protoc. 12, 2232–2242. doi: 10.1038/nprot.2017.074
Bi, Y., Marcus, A. K., Robert, H., Krajmalnik-Brown, R., Rittmann, B. E., Westerhoff, P., et al. (2020). The complex puzzle of dietary silver nanoparticles, mucus and microbiota in the gut. J. Toxicol. Environ. Health B Crit. Rev. 23, 69–89. doi: 10.1080/10937404.2019.1710914
Bisanz, J. E., Spanogiannopoulos, P., Pieper, L. M., Bustion, A. E., Turnbaugh, P. J. (2018). How to determine the role of the microbiome in drug disposition. Drug Metab. Dispos. 46, 1588–1595. doi: 10.1124/dmd.118.083402
Bjorkholm, B., Bok, C. M., Lundin, A., Rafter, J., Hibberd, M. L., Pettersson, S. (2009). Intestinal microbiota regulate xenobiotic metabolism in the liver. PloS One 4, e6958. doi: 10.1371/journal.pone.0006958
Bouhajja, E., Agathos, S. N., George, I. F. (2016). Metagenomics: probing pollutant fate in natural and engineered ecosystems. Biotechnol. Adv. 34, 1413–1426. doi: 10.1016/j.biotechadv.2016.10.006
Brooks, J. P., Edwards, D. J., Blithe, D. L., Fettweis, J. M., Serrano, M. G., Sheth, N. U., et al. (2017). Effects of combined oral contraceptives, depot medroxyprogesterone acetate and the levonorgestrel-releasing intrauterine system on the vaginal microbiome. Contraception 95, 405–413. doi: 10.1016/j.contraception.2016.11.006
Cani, P. D. (2018). Human gut microbiome: hopes, threats and promises. Gut 67, 1716–1725. doi: 10.1136/gutjnl-2018-316723
Cardona, C., Weisenhorn, P., Henry, C., Gilbert, J. A. (2016). Network-based metabolic analysis and microbial community modeling. Curr. Opin. Microbiol. 31, 124–131. doi: 10.1016/j.mib.2016.03.008
Carmody, R. N., Turnbaugh, P. J. (2014). Host-microbial interactions in the metabolism of therapeutic and diet-derived xenobiotics. J. Clin. Invest. 124, 4173–4181. doi: 10.1172/JCI72335
Cecchini, D. A., Laville, E., Laguerre, S., Robe, P., Leclerc, M., Dore, J., et al. (2013). Functional metagenomics reveals novel pathways of prebiotic breakdown by human gut bacteria. PloS One 8, e72766. doi: 10.1371/journal.pone.0072766
Centers for Disease Control and Prevention (CDC) (2009). Fourth National Report on Human Exposure to Environmental Chemicals (Atlanta, Georgia: Department of Health and Human Services).
Chassaing, B., Koren, O., Goodrich, J. K., Poole, A. C., Srinivasan, S., Ley, R. E., et al. (2015). Dietary emulsifiers impact the mouse gut microbiota promoting colitis and metabolic syndrome. Nature 519, 92–96. doi: 10.1038/nature14232
Chi, L., Xue, J., Tu, P., Lai, Y., Ru, H., Lu, K. (2019). Gut microbiome disruption altered the biotransformation and liver toxicity of arsenic in mice. Arch. Toxicol. 93, 25–35. doi: 10.1007/s00204-018-2332-7
Clarke, G., Sandhu, K. V., Griffin, B. T., Dinan, T. G., Cryan, J. F., Hyland, N. P. (2019). Gut reactions: breaking down xenobiotic-microbiome interactions. Pharmacol. Rev. 71, 198–224. doi: 10.1124/pr.118.015768
Clayton, T. A., Baker, D., Lindon, J. C., Everett, J. R., Nicholson, J. K. (2009). Pharmacometabonomic identification of a significant host-microbiome metabolic interaction affecting human drug metabolism. Proc. Natl. Acad. Sci. U.S.A. 106, 14728–14733. doi: 10.1073/pnas.0904489106
Cross, K. L., Campbell, J. H., Balachandran, M., Campbell, A. G., Cooper, S. J., Griffen, A., et al. (2019). Targeted isolation and cultivation of uncultivated bacteria by reverse genomics. Nat. Biotechnol. 37, 1314–1321. doi: 10.1038/s41587-019-0260-6
Crucitti, T., Hardy, L., Van De Wijgert, J., Agaba, S., Buyze, J., Kestelyn, E., et al. (2018). Contraceptive rings promote vaginal lactobacilli in a high bacterial vaginosis prevalence population: A randomised, open-label longitudinal study in Rwandan women. PloS One 13, e0201003. doi: 10.1371/journal.pone.0201003
Dabek, M., Mccrae, S. I., Stevens, V. J., Duncan, S. H., Louis, P. (2008). Distribution of beta-glucosidase and beta-glucuronidase activity and of beta-glucuronidase gene gus in human colonic bacteria. FEMS Microbiol. Ecol. 66, 487–495. doi: 10.1111/j.1574-6941.2008.00520.x
Daisley, B. A., Monachese, M., Trinder, M., Bisanz, J. E., Chmiel, J. A., Burton, J. P., et al. (2019). Immobilization of cadmium and lead by Lactobacillus rhamnosus GR-1 mitigates apical-to-basolateral heavy metal translocation in a Caco-2 model of the intestinal epithelium. Gut Microbes 10, 321–333. doi: 10.1080/19490976.2018.1526581
David, L. A., Maurice, C. F., Carmody, R. N., Gootenberg, D. B., Button, J. E., Wolfe, B. E., et al. (2014). Diet rapidly and reproducibly alters the human gut microbiome. Nature 505, 559–563. doi: 10.1038/nature12820
Dechartres, J., Pawluski, J. L., Gueguen, M. M., Jablaoui, A., Maguin, E., Rhimi, M., et al. (2019). Glyphosate and glyphosate-based herbicide exposure during the peripartum period affects maternal brain plasticity, maternal behaviour and microbiome. J. Neuroendocrinol. 31, e12731. doi: 10.1111/jne.12731
Delomenie, C., Fouix, S., Longuemaux, S., Brahimi, N., Bizet, C., Picard, B., et al. (2001). Identification and functional characterization of arylamine N-acetyltransferases in eubacteria: evidence for highly selective acetylation of 5-aminosalicylic acid. J. Bacteriol. 183, 3417–3427. doi: 10.1128/JB.183.11.3417-3427.2001
Dethlefsen, L., Relman, D. A. (2011). Incomplete recovery and individualized responses of the human distal gut microbiota to repeated antibiotic perturbation. Proc. Natl. Acad. Sci. U.S.A. 108 Suppl 1, 4554–4561. doi: 10.1073/pnas.1000087107
Devlin, A. S., Marcobal, A., Dodd, D., Nayfach, S., Plummer, N., Meyer, T., et al. (2016). Modulation of a circulating uremic solute via rational genetic manipulation of the gut microbiota. Cell Host Microbe 20, 709–715. doi: 10.1016/j.chom.2016.10.021
Dominguez-Bello, M. G., Costello, E. K., Contreras, M., Magris, M., Hidalgo, G., Fierer, N., et al. (2010). Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc. Natl. Acad. Sci. U.S.A. 107, 11971–11975. doi: 10.1073/pnas.1002601107
Elinav, E., Garrett, W. S., Trinchieri, G., Wargo, J. (2019). The cancer microbiome. Nat. Rev. Cancer 19, 371–376. doi: 10.1038/s41568-019-0155-3
Elrakaiby, M., Dutilh, B. E., Rizkallah, M. R., Boleij, A., Cole, J. N., Aziz, R. K. (2014). Pharmacomicrobiomics: the impact of human microbiome variations on systems pharmacology and personalized therapeutics. OMICS 18, 402–414. doi: 10.1089/omi.2014.0018
Eshel, G., Shepon, A., Makov, T., Milo, R. (2014). Land, irrigation water, greenhouse gas, and reactive nitrogen burdens of meat, eggs, and dairy production in the United States. Proc. Natl. Acad. Sci. U.S.A. 111, 11996–12001. doi: 10.1073/pnas.1402183111
Flint, H. J., Bayer, E. A., Rincon, M. T., Lamed, R., White, B. A. (2008). Polysaccharide utilization by gut bacteria: potential for new insights from genomic analysis. Nat. Rev. Microbiol. 6, 121–131. doi: 10.1038/nrmicro1817
Forster, S. C., Kumar, N., Anonye, B. O., Almeida, A., Viciani, E., Stares, M. D., et al. (2019). A human gut bacterial genome and culture collection for improved metagenomic analyses. Nat. Biotechnol. 37, 186–192. doi: 10.1038/s41587-018-0009-7
Gičević, A., Hindija, L., Karačić, A. (2020). “Toxicity of Azo Dyes in Pharmaceutical Industry,” in CMBEBIH 2019. IFMBE Proceedings. Eds. Badnjevic, A., Škrbić, R., Gurbeta Pokvić, L. (Cham): Springer).
Glatz, M., Jo, J. H., Kennedy, E. A., Polley, E. C., Segre, J. A., Simpson, E. L., et al. (2018). Emollient use alters skin barrier and microbes in infants at risk for developing atopic dermatitis. PloS One 13, e0192443. doi: 10.1371/journal.pone.0192443
Green, J. M. (2018). The rise and future of glyphosate and glyphosate-resistant crops. Pest. Manage. Sci. 74, 1035–1039. doi: 10.1002/ps.4462
Grenham, S., Clarke, G., Cryan, J. F., Dinan, T. G. (2011). Brain-gut-microbe communication in health and disease. Front. Physiol. 2, 94. doi: 10.3389/fphys.2011.00094
Grice, E. A., Kong, H. H., Conlan, S., Deming, C. B., Davis, J., Young, A. C., et al. (2009). Topographical and temporal diversity of the human skin microbiome. Science 324, 1190–1192. doi: 10.1126/science.1171700
Guardia-Escote, L., Basaure, P., Biosca-Brull, J., Cabre, M., Blanco, J., Perez-Fernandez, C., et al. (2020). APOE genotype and postnatal chlorpyrifos exposure modulate gut microbiota and cerebral short-chain fatty acids in preweaning mice. Food. Chem. Toxicol. 135, 110872. doi: 10.1016/j.fct.2019.110872
Haiser, H. J., Turnbaugh, P. J. (2012). Is it time for a metagenomic basis of therapeutics? Science 336, 1253–1255. doi: 10.1126/science.1224396
Haiser, H. J., Gootenberg, D. B., Chatman, K., Sirasani, G., Balskus, E. P., Turnbaugh, P. J. (2013). Predicting and manipulating cardiac drug inactivation by the human gut bacterium Eggerthella lenta. Science 341, 295–298. doi: 10.1126/science.1235872
Halden, R. U., Lindeman, A. E., Aiello, A. E., Andrews, D., Arnold, W. A., Fair, P., et al. (2017). The Florence Statement on triclosan and triclocarban. Environ. Health Perspect. 125, 064501. doi: 10.1289/EHP1788
Halden, R. U. (2014). On the need and speed of regulating triclosan and triclocarban in the United States. Environ. Sci. Technol. 48, 3603–3611. doi: 10.1021/es500495p
Hall, S. D., Thummel, K. E., Watkins, P. B., Lown, K. S., Benet, L. Z., Paine, M. F., et al. (1999). Molecular and physical mechanisms of first-pass extraction. Drug Metab. Dispos. 27, 161–166.
Ito, T., Sekizuka, T., Kishi, N., Yamashita, A., Kuroda, M. (2019a). Conventional culture methods with commercially available media unveil the presence of novel culturable bacteria. Gut Microbes 10, 77–91. doi: 10.1080/19490976.2018.1491265
Ito, Y., Ito, T., Yamashiro, K., Mineshiba, F., Hirai, K., Omori, K., et al. (2019b). Antimicrobial and antibiofilm effects of abietic acid on cariogenic Streptococcus mutans. Odontology. 108, 57–65. doi: 10.1007/s10266-019-00456-0
Jain, R. K., Kapur, M., Labana, S., Lal, B., Sarma, P. M., Bhattacharya, D., et al. (2005). Microbial diversity: Application of microorganisms for the biodegradation of xenobiotics. Curr. Sci. 89, 101–112.
Kallemeijn, W. W., Witte, M. D., Wennekes, T., Aerts, J. M. (2014). Mechanism-based inhibitors of glycosidases: design and applications. Adv. Carbohydr. Chem. Biochem. 71, 297–338. doi: 10.1016/B978-0-12-800128-8.00004-2
Kashyap, P. C., Chia, N., Nelson, H., Segal, E., Elinav, E. (2017). Microbiome at the frontier of personalized medicine. Mayo Clin. Proc. 92, 1855–1864. doi: 10.1016/j.mayocp.2017.10.004
Kassie, F., Rabot, S., Kundi, M., Chabicovsky, M., Qin, H. M., Knasmuller, S. (2001). Intestinal microflora plays a crucial role in the genotoxicity of the cooked food mutagen 2-amino-3-methylimidazo [4,5-f]quinoline. Carcinogenesis 22, 1721–1725. doi: 10.1093/carcin/22.10.1721
Kim, J. M. (2014). Antimicrobial proteins in intestine and inflammatory bowel diseases. Intest. Res. 12, 20–33. doi: 10.5217/ir.2014.12.1.20
Klatt, N. R., Cheu, R., Birse, K., Zevin, A. S., Perner, M., Noel-Romas, L., et al. (2017). Vaginal bacteria modify HIV tenofovir microbicide efficacy in African women. Science 356, 938–945. doi: 10.1126/science.aai9383
Koeth, R. A., Wang, Z., Levison, B. S., Buffa, J. A., Org, E., Sheehy, B. T., et al. (2013). Intestinal microbiota metabolism of L-carnitine, a nutrient in red meat, promotes atherosclerosis. Nat. Med. 19, 576–585. doi: 10.1038/nm.3145
Koppel, N., Maini Rekdal, V., Balskus, E. P. (2017). Chemical transformation of xenobiotics by the human gut microbiota. Science 356 (6344), 1–11. doi: 10.1126/science.aag2770
Kostic, A. D., Chun, E., Robertson, L., Glickman, J. N., Gallini, C. A., Michaud, M., et al. (2013). Fusobacterium nucleatum potentiates intestinal tumorigenesis and modulates the tumor-immune microenvironment. Cell Host Microbe 14, 207–215. doi: 10.1016/j.chom.2013.07.007
Kostic, A. D., Xavier, R. J., Gevers, D. (2014). The microbiome in inflammatory bowel disease: current status and the future ahead. Gastroenterology 146, 1489–1499. doi: 10.1053/j.gastro.2014.02.009
Kosyakovsky, L. B. (2017). The emerging role of the microbiome in precision medicine: An overview. UBCMJ 9, 2.
Kumano, T., Fujiki, E., Hashimoto, Y., Kobayashi, M. (2016). Discovery of a sesamin-metabolizing microorganism and a new enzyme. Proc. Natl. Acad. Sci. U.S.A. 113, 9087–9092. doi: 10.1073/pnas.1605050113
Kuntz, T. M., Gilbert, J. A. (2017). Introducing the microbiome into precision medicine. Trends Pharmacol. Sci. 38, 81–91. doi: 10.1016/j.tips.2016.10.001
Lam, K. N., Martens, E. C., Charles, T. C. (2018). Developing a Bacteroides system for function-based screening of DNA from the human gut microbiome. mSystems 3, e00195-17. doi: 10.1128/mSystems.00195-17
Langdon, A., Crook, N., Dantas, G. (2016). The effects of antibiotics on the microbiome throughout development and alternative approaches for therapeutic modulation. Genome Med. 8, 39. doi: 10.1186/s13073-016-0294-z
Langille, M. G., Zaneveld, J., Caporaso, J. G., Mcdonald, D., Knights, D., Reyes, J. A., et al. (2013). Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 31, 814–821. doi: 10.1038/nbt.2676
Langille, M. G. I., Ravel, J., Fricke, W. F. (2018). “Available upon request”: not good enough for microbiome data! Microbiome 6, 8. doi: 10.1186/s40168-017-0394-z
Lankisch, T. O., Schulz, C., Zwingers, T., Erichsen, T. J., Manns, M. P., Heinemann, V., et al. (2008). Gilbert’s Syndrome and irinotecan toxicity: combination with UDP-glucuronosyltransferase 1A7 variants increases risk. Cancer Epidemiol. Biomarkers Prev. 17, 695–701. doi: 10.1158/1055-9965.EPI-07-2517
Laue, H. E., Brennan, K. J. M., Gillet, V., Abdelouahab, N., Coull, B. A., Weisskopf, M. G., et al. (2019). Associations of prenatal exposure to polybrominated diphenyl ethers and polychlorinated biphenyls with long-term gut microbiome structure: a pilot study. Environ. Epidemiol. 3 (1), e039. doi: 10.1097/EE9.0000000000000039
Lavaee, F., Faez, K., Faez, K., Hadi, N., Modaresi, F. (2016). Antimicrobial and antibiofilm activity of silver, titanium dioxide and iron nano particles. Am. J. Dent. 29, 315–320.
Lavrijsen, K., Van Dyck, D., Van Houdt, J., Hendrickx, J., Monbaliu, J., Woestenborghs, R., et al. (1995). Reduction of the prodrug loperamide oxide to its active drug loperamide in the gut of rats, dogs, and humans. Drug Metab. Dispos. 23, 354–362.
Li, P., Lin, C., Cheng, H., Duan, X., Lei, K. (2015). Contamination and health risks of soil heavy metals around a lead/zinc smelter in southwestern China. Ecotoxicol. Environ. Saf. 113, 391–399. doi: 10.1016/j.ecoenv.2014.12.025
Liu, N., Li, H., Chevrette, M. G., Zhang, L., Cao, L., Zhou, H., et al. (2019). Functional metagenomics reveals abundant polysaccharide-degrading gene clusters and cellobiose utilization pathways within gut microbiota of a wood-feeding higher termite. ISME J. 13, 104–117. doi: 10.1038/s41396-018-0255-1
Lloyd-Price, J., Mahurkar, A., Rahnavard, G., Crabtree, J., Orvis, J., Hall, A. B., et al. (2017). Strains, functions and dynamics in the expanded Human Microbiome Project. Nature 550, 61–66. doi: 10.1038/nature23889
Lopes, C., Soares, J., Tavaria, F., Duarte, A., Correia, O., Sokhatska, O., et al. (2015). Chitosan-coated textiles may improve atopic dermatitis severity by modulating skin staphylococcal profile: a randomized controlled trial. PloS One 10, e0142844. doi: 10.1371/journal.pone.0142844
Lu, K., Abo, R. P., Schlieper, K. A., Graffam, M. E., Levine, S., Wishnok, J. S., et al. (2014). Arsenic exposure perturbs the gut microbiome and its metabolic profile in mice: an integrated metagenomics and metabolomics analysis. Environ. Health Perspect. 122, 284–291. doi: 10.1289/ehp.1307429
Maged, M., El Hosseiny, A., Saadeldin, M. K., Aziz, R. K., Ramadan, E. (2019). Thermal stability of a mercuric reductase from the Red Sea Atlantis II hot brine environment as analyzed by site-directed mutagenesis. Appl. Environ. Microbiol. 85 (3), e02387-18, 1–12. doi: 10.1128/AEM.02387-18
Manor, O., Borenstein, E. (2017). Systematic characterization and analysis of the taxonomic drivers of functional shifts in the human microbiome. Cell Host Microbe 21, 254–267. doi: 10.1016/j.chom.2016.12.014
Mao, X., Yang, Q., Chen, D., Yu, B., He, J. (2019). Benzoic acid used as food and feed additives can regulate gut functions. Biomed. Res. Int. 2019, 5721585. doi: 10.1155/2019/5721585
Marchesi, J. R., Ravel, J. (2015). The vocabulary of microbiome research: a proposal. Microbiome 3, 31. doi: 10.1186/s40168-015-0094-5
Maruvada, P., Leone, V., Kaplan, L. M., Chang, E. B. (2017). The human microbiome and obesity: moving beyond associations. Cell Host Microbe 22, 589–599. doi: 10.1016/j.chom.2017.10.005
Mason, L. H., Harp, J. P., Han, D. Y. (2014). Pb neurotoxicity: neuropsychological effects of lead toxicity. Biomed. Res. Int. 2014, 840547. doi: 10.1155/2014/840547
Mathan, V. I., Wiederman, J., Dobkin, J. F., Lindenbaum, J. (1989). Geographic differences in digoxin inactivation, a metabolic activity of the human anaerobic gut flora. Gut 30, 971–977. doi: 10.1136/gut.30.7.971
Meinl, W., Sczesny, S., Brigelius-Flohe, R., Blaut, M., Glatt, H. (2009). Impact of gut microbiota on intestinal and hepatic levels of phase 2 xenobiotic-metabolizing enzymes in the rat. Drug Metab. Dispos. 37, 1179–1186. doi: 10.1124/dmd.108.025916
Michalopoulos, G. K. (2007). Liver regeneration. J. Cell. Physiol. 213, 286–300. doi: 10.1002/jcp.21172
Minekus, M., Smeets-Peeters, M., Bernalier, A., Marol-Bonnin, S., Havenaar, R., Marteau, P., et al. (1999). A computer-controlled system to simulate conditions of the large intestine with peristaltic mixing, water absorption and absorption of fermentation products. Appl. Microbiol. Biotechnol. 53, 108–114. doi: 10.1007/s002530051622
Misal, S. A., Gawai, K. R. (2018). Azoreductase: a key player of xenobiotic metabolism. Bioresour. Bioprocess. 5, 17. doi: 10.1186/s40643-018-0206-8
Mohamed, Y. M., Ghazy, M. A., Sayed, A., Ouf, A., El-Dorry, H., Siam, R. (2013). Isolation and characterization of a heavy metal-resistant, thermophilic esterase from a Red Sea brine pool. Sci. Rep. 3, 3358. doi: 10.1038/srep03358
Molly, K., Vande Woestyne, M., Verstraete, W. (1993). Development of a 5-step multi-chamber reactor as a simulation of the human intestinal microbial ecosystem. Appl. Microbiol. Biotechnol. 39, 254–258. doi: 10.1007/BF00228615
Mora, D., Arioli, S. (2014). Microbial urease in health and disease. PloS Pathog. 10, e1004472. doi: 10.1371/journal.ppat.1004472
Moustafa, A., Li, W., Anderson, E. L., Wong, E. H. M., Dulai, P. S., Sandborn, W. J., et al. (2018). Genetic risk, dysbiosis, and treatment stratification using host genome and gut microbiome in inflammatory bowel disease. Clin. Transl. Gastroenterol. 9, e132. doi: 10.1038/ctg.2017.58
Nakayama, H., Kinouchi, T., Kataoka, K., Akimoto, S., Matsuda, Y., Ohnishi, Y. (1997). Intestinal anaerobic bacteria hydrolyse sorivudine, producing the high blood concentration of 5-(E)-(2-bromovinyl)uracil that increases the level and toxicity of 5-fluorouracil. Pharmacogenetics 7, 35–43. doi: 10.1097/00008571-199702000-00005
Napolitano, A., Miller, S., Nicholls, A. W., Baker, D., Van Horn, S., Thomas, E., et al. (2014). Novel gut-based pharmacology of metformin in patients with type 2 diabetes mellitus. PloS One 9, e100778. doi: 10.1371/journal.pone.0100778
Naujokas, M. F., Anderson, B., Ahsan, H., Aposhian, H. V., Graziano, J. H., Thompson, C., et al. (2013). The broad scope of health effects from chronic arsenic exposure: update on a worldwide public health problem. Environ. Health Perspect. 121, 295–302. doi: 10.1289/ehp.1205875
Neis, E. P., Dejong, C. H., Rensen, S. S. (2015). The role of microbial amino acid metabolism in host metabolism. Nutrients 7, 2930–2946. doi: 10.3390/nu7042930
Nelson, D. R. (2005). Book Review: Cytochrome P450: Structure, Mechanism, and Biochemistry, 3rd ed Edited by Paul R. Ortiz de Montellano (University of California, San Francisco). J. Am. Chem. Soc 127, 12147–12148. doi: 10.1021/ja041050x
Ngara, T. R., Zhang, H. (2018). Recent advances in function-based metagenomic screening. Genomics Proteomics Bioinf. 16, 405–415. doi: 10.1016/j.gpb.2018.01.002
Nichols, D., Cahoon, N., Trakhtenberg, E. M., Pham, L., Mehta, A., Belanger, A., et al. (2010). Use of ichip for high-throughput in situ cultivation of “uncultivable” microbial species. Appl. Environ. Microbiol. 76, 2445–2450. doi: 10.1128/AEM.01754-09
Nicholson, J. K., Wilson, I. D. (2003). Opinion: understanding ‘global’ systems biology: metabonomics and the continuum of metabolism. Nat. Rev. Drug Discovery 2, 668–676. doi: 10.1038/nrd1157
Nogacka, A. M., Gomez-Martin, M., Suarez, A., Gonzalez-Bernardo, O., De Los Reyes-Gavilan, C. G., Gonzalez, S. (2019). Xenobiotics formed during food processing: Their relation with the intestinal microbiota and colorectal cancer. Int. J. Mol. Sci. 20 (8), 2051. doi: 10.3390/ijms20082051
Nyyssonen, M., Tran, H. M., Karaoz, U., Weihe, C., Hadi, M. Z., Martiny, J. B., et al. (2013). Coupled high-throughput functional screening and next generation sequencing for identification of plant polymer decomposing enzymes in metagenomic libraries. Front. Microbiol. 4, 282. doi: 10.3389/fmicb.2013.00282
Pannaraj, P. S., Li, F., Cerini, C., Bender, J. M., Yang, S., Rollie, A., et al. (2017). Association between breast milk bacterial communities and establishment and development of the infant gut microbiome. JAMA Pediatr. 171, 647–654. doi: 10.1001/jamapediatrics.2017.0378
Pehrsson, E. C., Tsukayama, P., Patel, S., Mejia-Bautista, M., Sosa-Soto, G., Navarrete, K. M., et al. (2016). Interconnected microbiomes and resistomes in low-income human habitats. Nature 533, 212–216. doi: 10.1038/nature17672
Pellock, S. J., Walton, W. G., Ervin, S. M., Torres-Rivera, D., Creekmore, B. C., Bergan, G., et al. (2019). Discovery and characterization of FMN-binding beta-glucuronidases in the human gut microbiome. J. Mol. Biol. 431, 970–980. doi: 10.1016/j.jmb.2019.01.013
Penalver Bernabe, B., Cralle, L., Gilbert, J. A. (2018). Systems biology of the human microbiome. Curr. Opin. Biotechnol. 51, 146–153. doi: 10.1016/j.copbio.2018.01.018
Peppercorn, M. A., Goldman, P. (1972). The role of intestinal bacteria in the metabolism of salicylazosulfapyridine. J. Pharmacol. Exp. Ther. 181, 555–562.
Petersen, C., Round, J. L. (2014). Defining dysbiosis and its influence on host immunity and disease. Cell Microbiol. 16, 1024–1033. doi: 10.1111/cmi.12308
Peterson, M. A., Mcmaster, S. A., Riechers, D. E., Skelton, J., Stahlman, P. W. (2017). 2,4-D past, present, and future: a review. Weed Technol. 30, 303–345. doi: 10.1614/WT-D-15-00131.1
Petrie, B., Barden, R., Kasprzyk-Hordern, B. (2015). A review on emerging contaminants in wastewaters and the environment: current knowledge, understudied areas and recommendations for future monitoring. Water Res. 72, 3–27. doi: 10.1016/j.watres.2014.08.053
Pinget, G., Tan, J., Janac, B., Kaakoush, N. O., Angelatos, A. S., O’sullivan, J., et al. (2019). Impact of the food additive titanium dioxide (E171) on gut microbiota-host interaction. Front. Nutr. 6, 57. doi: 10.3389/fnut.2019.00057
Poole, A. C., Pischel, L., Ley, C., Suh, G., Goodrich, J. K., Haggerty, T. D., et al. (2016). Crossover control study of the effect of personal care products containing triclosan on the microbiome. mSphere 1 (3), e00056-15. doi: 10.1128/mSphere.00056-15
Rafii, F., Sutherland, J. B., Hansen, E. B., Jr., Cerniglia, C. E. (1997). Reduction of nitrazepam by Clostridium leptum, a nitroreductase-producing bacterium isolated from the human intestinal tract. Clin. Infect. Dis. 25 Suppl 2, S121–S122. doi: 10.1086/516204
Rajendhran, J., Gunasekaran, P. (2010). Human microbiomics. Indian J. Microbiol. 50, 109–112. doi: 10.1007/s12088-010-0034-9
Rao, B. C., Lou, J. M., Wang, W. J., Li, A., Cui, G. Y., Yu, Z., et al. (2020). Humanmicrobiome is a diagnostic biomarker in hepatocellular carcinoma. Hepatob.Pancreat. Dis. Int.. doi: 10.1016/j.hbpd.2020.01.003
Redfern, L. K., Gardner, C. M., Hodzic, E., Ferguson, P. L., Hsu-Kim, H., Gunsch, C. K. (2019). A new framework for approaching precision bioremediation of PAH contaminated soils. J. Hazard. Mater. 378, 120859. doi: 10.1016/j.jhazmat.2019.120859
Relman, D. A., Falkow, S. (2001). The meaning and impact of the human genome sequence for microbiology. Trends Microbiol. 9, 206–208. doi: 10.1016/S0966-842X(01)02041-8
Ren, Z., Li, A., Jiang, J., Zhou, L., Yu, Z., Lu, H., et al. (2019). Gut microbiome analysis as a tool towards targeted non-invasive biomarkers for early hepatocellular carcinoma. Gut 68, 1014–1023. doi: 10.1136/gutjnl-2017-315084
Rigby, R. J., Carr, J., Orgel, K., King, S. L., Lund, P. K., Dekaney, C. M. (2016). Intestinal bacteria are necessary for doxorubicin-induced intestinal damage but not for doxorubicin-induced apoptosis. Gut Microbes 7, 414–423. doi: 10.1080/19490976.2016.1215806
Rizkallah, M. R., Saad, R., Aziz, R. K. (2010). The Human Microbiome Project, personalized medicine and the birth of pharmacomicrobiomics. Curr. Pharmacogenomics Person. Med. 8, 182–193. doi: 10.2174/187569210792246326
Rosato, A., Tenori, L., Cascante, M., De Atauri Carulla, P. R., Santos, M. D., Saccenti, E. (2018). From correlation to causation: analysis of metabolomics data using systems biology approaches. Metabolomics 14, 37. doi: 10.1007/s11306-018-1335-y
Rossol, I., Puhler, A. (1992). The Corynebacterium glutamicum aecD gene encodes a C-S lyase with alpha, beta-elimination activity that degrades aminoethylcysteine. J. Bacteriol. 174, 2968–2977. doi: 10.1128/JB.174.9.2968-2977.1992
Rubinstein, M. R., Wang, X., Liu, W., Hao, Y., Cai, G., Han, Y. W. (2013). Fusobacterium nucleatum promotes colorectal carcinogenesis by modulating E-cadherin/beta-catenin signaling via its FadA adhesin. Cell Host Microbe 14, 195–206. doi: 10.1016/j.chom.2013.07.012
Rude, K. M., Pusceddu, M. M., Keogh, C. E., Sladek, J. A., Rabasa, G., Miller, E. N., et al. (2019). Developmental exposure to polychlorinated biphenyls (PCBs) in the maternal diet causes host-microbe defects in weanling offspring mice. Environ. Pollut. 253, 708–721. doi: 10.1016/j.envpol.2019.07.066
Saad, R., Rizkallah, M. R., Aziz, R. K. (2012). Gut Pharmacomicrobiomics: the tip of an iceberg of complex interactions between drugs and gut-associated microbes. Gut Pathog. 4, 16. doi: 10.1186/1757-4749-4-16
Saitta, K. S., Zhang, C., Lee, K. K., Fujimoto, K., Redinbo, M. R., Boelsterli, U. A. (2014). Bacterial beta-glucuronidase inhibition protects mice against enteropathy induced by indomethacin, ketoprofen or diclofenac: mode of action and pharmacokinetics. Xenobiotica 44, 28–35. doi: 10.3109/00498254.2013.811314
Satarug, S., Baker, J. R., Urbenjapol, S., Haswell-Elkins, M., Reilly, P. E., Williams, D. J., et al. (2003). A global perspective on cadmium pollution and toxicity in non-occupationally exposed population. Toxicol. Lett. 137, 65–83. doi: 10.1016/S0378-4274(02)00381-8
Satarug, S., Garrett, S. H., Sens, M. A., Sens, D. A. (2010). Cadmium, environmental exposure, and health outcomes. Environ. Health Perspect. 118, 182–190. doi: 10.1289/ehp.0901234
Sayed, A., Ghazy, M. A., Ferreira, A. J., Setubal, J. C., Chambergo, F. S., Ouf, A., et al. (2014). A novel mercuric reductase from the unique deep brine environment of Atlantis II in the Red Sea. J. Biol. Chem. 289, 1675–1687. doi: 10.1074/jbc.M113.493429
Scott, A. J., Alexander, J. L., Merrifield, C. A., Cunningham, D., Jobin, C., Brown, R., et al. (2019). International Cancer Microbiome Consortium consensus statement on the role of the human microbiome in carcinogenesis. Gut 68, 1624–1632. doi: 10.1136/gutjnl-2019-318556
Selwyn, F. P., Cheng, S. L., Klaassen, C. D., Cui, J. Y. (2016). Regulation of hepatic drug-metabolizing enzymes in germ-free mice by conventionalization and probiotics. Drug Metab. Dispos. 44, 262–274. doi: 10.1124/dmd.115.067504
Sharon, G., Garg, N., Debelius, J., Knight, R., Dorrestein, P. C., Mazmanian, S. K. (2014). Specialized metabolites from the microbiome in health and disease. Cell Metab. 20, 719–730. doi: 10.1016/j.cmet.2014.10.016
Shen, S., Lim, G., You, Z., Ding, W., Huang, P., Ran, C., et al. (2017). Gut microbiota is critical for the induction of chemotherapy-induced pain. Nat. Neurosci. 20, 1213–1216. doi: 10.1038/nn.4606
Smits, J. P. H., Ederveen, T. H. A., Rikken, G., Van Den Brink, N. J. M., Van Vlijmen-Willems, I., Boekhorst, J., et al. (2019). Targeting the cutaneous microbiota in atopic dermatitis by coal tar via AHR-dependent induction of antimicrobial peptides. J. Invest. Dermatol. 139 (9), S251. doi: 10.1016/j.jid.2019.07.215
Soriano-Lerma, A., Magan-Fernandez, A., Gijon, J., Sanchez-Fernandez, E., Soriano, M., Garcia-Salcedo, J. A., et al. (2019). Short-term effects of hyaluronic acid on the subgingival microbiome in peri-implantitis: A randomized controlled clinical trial. J. Periodontol. (0), 1–12. doi: 10.1002/JPER.19-0184
Soucek, P. (2011). “Xenobiotics,” in Encyclopedia of Cancer. Ed. Schwab, M. (Berlin, Heidelberg: Springer Berlin Heidelberg), 3964–3967.
Spanogiannopoulos, P., Bess, E. N., Carmody, R. N., Turnbaugh, P. J. (2016). The microbial pharmacists within us: a metagenomic view of xenobiotic metabolism. Nat. Rev. Microbiol. 14, 273–287. doi: 10.1038/nrmicro.2016.17
Spatz, M., Smith, D. W., Mcdaniel, E. G., Laqueur, G. L. (1967). Role of intestinal microorganisms in determining cycasin toxicity. Proc. Soc Exp. Biol. Med. 124, 691–697. doi: 10.3181/00379727-124-31826
Spor, A., Koren, O., Ley, R. (2011). Unravelling the effects of the environment and host genotype on the gut microbiome. Nat. Rev. Microbiol. 9, 279–290. doi: 10.1038/nrmicro2540
Stingl, J. C., Bartels, H., Viviani, R., Lehmann, M. L., Brockmoller, J. (2014). Relevance of UDP-glucuronosyltransferase polymorphisms for drug dosing: A quantitative systematic review. Pharmacol. Ther. 141, 92–116. doi: 10.1016/j.pharmthera.2013.09.002
Stringer, A. M., Gibson, R. J., Logan, R. M., Bowen, J. M., Yeoh, A. S., Hamilton, J., et al. (2009). Gastrointestinal microflora and mucins may play a critical role in the development of 5-Fluorouracil-induced gastrointestinal mucositis. Exp. Biol. Med. (Maywood) 234, 430–441. doi: 10.3181/0810-RM-301
Suez, J., Korem, T., Zilberman-Schapira, G., Segal, E., Elinav, E. (2015). Non-caloric artificial sweeteners and the microbiome: findings and challenges. Gut Microbes 6, 149–155. doi: 10.1080/19490976.2015.1017700
Sun, J., Ling, Z., Wang, F., Chen, W., Li, H., Jin, J., et al. (2016). Clostridium butyricum pretreatment attenuates cerebral ischemia/reperfusion injury in mice via anti-oxidation and anti-apoptosis. Neurosci. Lett. 613, 30–35. doi: 10.1016/j.neulet.2015.12.047
Sutton, D., Butler, A. M., Nadin, L., Murray, M. (1997). Role of CYP3A4 in human hepatic diltiazem N-demethylation: inhibition of CYP3A4 activity by oxidized diltiazem metabolites. J. Pharmacol. Exp. Ther. 282, 294–300.
Swanson, H. I. (2015). Drug metabolism by the host and gut microbiota: A partnership or rivalry? Drug Metab. Dispos. 43, 1499–1504. doi: 10.1124/dmd.115.065714
Takagi, R., Sasaki, K., Sasaki, D., Fukuda, I., Tanaka, K., Yoshida, K., et al. (2016). A single-batch fermentation system to simulate human colonic microbiota for high-throughput evaluation of prebiotics. PloS One 11, e0160533. doi: 10.1371/journal.pone.0160533
Takasuna, K., Hagiwara, T., Hirohashi, M., Kato, M., Nomura, M., Nagai, E., et al. (1996). Involvement of beta-glucuronidase in intestinal microflora in the intestinal toxicity of the antitumor camptothecin derivative irinotecan hydrochloride (CPT-11) in rats. Cancer Res. 56, 3752–3757.
Tjalsma, H., Boleij, A., Marchesi, J. R., Dutilh, B. E. (2012). A bacterial driver-passenger model for colorectal cancer: beyond the usual suspects. Nat. Rev. Microbiol. 10, 575–582. doi: 10.1038/nrmicro2819
Toda, T., Ohi, K., Kudo, T., Yoshida, T., Ikarashi, N., Ito, K., et al. (2009). Ciprofloxacin suppresses Cyp3a in mouse liver by reducing lithocholic acid-producing intestinal flora. Drug Metab. Pharmacokinet. 24, 201–208. doi: 10.2133/dmpk.24.201
Tozaki, H., Emi, Y., Horisaka, E., Fujita, T., Yamamoto, A., Muranishi, S. (1997). Degradation of insulin and calcitonin and their protection by various protease inhibitors in rat caecal contents: implications in peptide delivery to the colon. J. Pharm. Pharmacol. 49, 164–168. doi: 10.1111/j.2042-7158.1997.tb06773.x
Tu, P., Gao, B., Chi, L., Lai, Y., Bian, X., Ru, H., et al. (2019). Subchronic low-dose 2,4-D exposure changed plasma acylcarnitine levels and induced gut microbiome perturbations in mice. Sci. Rep. 9, 4363. doi: 10.1038/s41598-019-40776-3
Turnbaugh, P. J., Ley, R. E., Hamady, M., Fraser-Liggett, C. M., Knight, R., Gordon, J. I. (2007). The Human Microbiome Project. Nature 449, 804–810. doi: 10.1038/nature06244
Uebanso, T., Kano, S., Yoshimoto, A., Naito, C., Shimohata, T., Mawatari, K., et al. (2017). Effects of consuming xylitol on gut microbiota and lipid metabolism in mice. Nutrients 9 (7), 756 doi: 10.3390/nu9070756
Ulmer, J. E., Vilen, E. M., Namburi, R. B., Benjdia, A., Beneteau, J., Malleron, A., et al. (2014). Characterization of glycosaminoglycan (GAG) sulfatases from the human gut symbiont Bacteroides thetaiotaomicron reveals the first GAG-specific bacterial endosulfatase. J. Biol. Chem. 289, 24289–24303. doi: 10.1074/jbc.M114.573303
Uppal, K., Walker, D. I., Liu, K., Li, S., Go, Y. M., Jones, D. P. (2016). Computational metabolomics: A framework for the million metabolome. Chem. Res. Toxicol. 29, 1956–1975. doi: 10.1021/acs.chemrestox.6b00179
Vanhaecke, L., Vercruysse, F., Boon, N., Verstraete, W., Cleenwerck, I., De Wachter, M., et al. (2008). Isolation and characterization of human intestinal bacteria capable of transforming the dietary carcinogen 2-amino-1-methyl-6-phenylimidazo[4,5-b]pyridine. Appl. Environ. Microbiol. 74, 1469–1477. doi: 10.1128/AEM.02064-07
Vipperla, K., O’keefe, S. J. (2016). Diet, microbiota, and dysbiosis: a ‘recipe’ for colorectal cancer. Food Funct. 7, 1731–1740. doi: 10.1039/C5FO01276G
Wang, H., Geng, C., Li, J., Hu, A., Yu, C. P. (2014a). Characterization of a novel melamine-degrading bacterium isolated from a melamine-manufacturing factory in China. Appl. Microbiol. Biotechnol. 98, 3287–3293. doi: 10.1007/s00253-013-5363-2
Wang, H., Li, J., Hu, A., Qin, D., Xu, H., Yu, C. P. (2014b). Melaminivora alkalimesophila gen. nov., sp. nov., a melamine-degrading betaproteobacterium isolated from a melamine-producing factory. Int. J. Syst. Evol. Microbiol. 64, 1938–1944. doi: 10.1099/ijs.0.055103-0
Wang, Y. K., Yang, X. N., Zhu, X., Xiao, X. R., Yang, X. W., Qin, H. B., et al. (2019). Role of metabolic activation in elemicin-induced cellular toxicity. J. Agric. Food Chem. 67, 8243–8252. doi: 10.1021/acs.jafc.9b02137
Wilson, I. D., Nicholson, J. K. (2017). Gut microbiome interactions with drug metabolism, efficacy, and toxicity. Transl. Res. 179, 204–222. doi: 10.1016/j.trsl.2016.08.002
Wishart, D. S., Feunang, Y. D., Guo, A. C., Lo, E. J., Marcu, A., Grant, J. R., et al. (2018). DrugBank 5.0: a major update to the DrugBank database for 2018. Nucleic Acids Res. 46, D1074–d1082. doi: 10.1093/nar/gkx1037
Wu, G. D., Chen, J., Hoffmann, C., Bittinger, K., Chen, Y. Y., Keilbaugh, S. A., et al. (2011). Linking long-term dietary patterns with gut microbial enterotypes. Science 334, 105–108. doi: 10.1126/science.1208344
Wuana, R. A., Okieimen, F. E. (2011). Heavy metals in contaminated soils: A review of sources, chemistry, risks and best available strategies for remediation. ISRN Ecol. 2011, 20. doi: 10.5402/2011/402647
Yang, H., Wang, W., Romano, K. A., Gu, M., Sanidad, K. Z., Kim, D., et al. (2018). A common antimicrobial additive increases colonic inflammation and colitis-associated colon tumorigenesis in mice. Sci. Transl. Med. 10. (443), eaan4116 doi: 10.1126/scitranslmed.aan4116
Yin, N., Zhang, Z., Cai, X., Du, H., Sun, G., Cui, Y. (2015). In vitro method to assess soil arsenic metabolism by human gut microbiota: arsenic speciation and distribution. Environ. Sci. Technol. 49, 10675–10681. doi: 10.1021/acs.est.5b03046
Yu, H., Wu, B., Zhang, X. X., Liu, S., Yu, J., Cheng, S., et al. (2016). Arsenic metabolism and toxicity influenced by ferric iron in simulated gastrointestinal tract and the roles of gut microbiota. Environ. Sci. Technol. 50, 7189–7197. doi: 10.1021/acs.est.6b01533
Yu, C., Zhou, B., Xia, X., Chen, S., Deng, Y., Wang, Y., et al. (2019). Prevotella copri is associated with carboplatin-induced gut toxicity. Cell. Death Dis. 10, 714. doi: 10.1038/s41419-019-1963-9
Zahran, S. A., Ali-Tammam, M., Hashem, A. M., Aziz, R. K., Ali, A. E. (2019). Azoreductase activity of dye-decolorizing bacteria isolated from the human gut microbiota. Sci. Rep. 9, 5508. doi: 10.1038/s41598-019-41894-8
Zhang, J., Empl, M. T., Schwab, C., Fekry, M. I., Engels, C., Schneider, M., et al. (2017a). Gut microbial transformation of the dietary imidazoquinoxaline mutagen MelQx reduces its cytotoxic and mutagenic potency. Toxicol. Sci. 159, 266–276. doi: 10.1093/toxsci/kfx132
Zhang, X., Li, S., Zhou, Y., Su, W., Ruan, X., Wang, B., et al. (2017b). Ablation of cytochrome P450 omega-hydroxylase 4A14 gene attenuates hepatic steatosis and fibrosis. Proc. Natl. Acad. Sci. U.S.A. 114, 3181–3185. doi: 10.1073/pnas.1700172114
Zheng, X., Zhao, A., Xie, G., Chi, Y., Zhao, L., Li, H., et al. (2013). Melamine-induced renal toxicity is mediated by the gut microbiota. Sci. Transl. Med. 5, 172ra122. doi: 10.1126/scitranslmed.3005114
Ziko, L., Saqr, A. A., Ouf, A., Gimpel, M., Aziz, R. K., Neubauer, P., et al. (2019). Antibacterial and anticancer activities of orphan biosynthetic gene clusters from Atlantis II Red Sea brine pool. Microb. Cell Fact. 18, 56. doi: 10.1186/s12934-019-1103-3
Zimmermann, M., Zimmermann-Kogadeeva, M., Wegmann, R., Goodman, A. L. (2019a). Mapping human microbiome drug metabolism by gut bacteria and their genes. Nature 570, 462–467. doi: 10.1038/s41586-019-1291-3
Keywords: microbiota, microbiome, pharmacomicrobiomics, metabolism, secondary metabolism, cytochromes
Citation: Abdelsalam NA, Ramadan AT, ElRakaiby MT and Aziz RK (2020) Toxicomicrobiomics: The Human Microbiome vs. Pharmaceutical, Dietary, and Environmental Xenobiotics. Front. Pharmacol. 11:390. doi: 10.3389/fphar.2020.00390
Received: 01 November 2019; Accepted: 16 March 2020;
Published: 16 April 2020.
Edited by:
Khare Sangeeta, National Center for Toxicological Research (FDA), United StatesReviewed by:
Alberto Mantovani, Higher Institute of Health (ISS), ItalyAlbert P. Li, In Vitro ADMET Laboratories, United States
Copyright © 2020 Abdelsalam, Ramadan, ElRakaiby and Aziz. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ramy Karam Aziz, cmFteS5heml6QGdtYWlsLmNvbQ==
†These authors have contributed equally to this work