 Wei Gao
Wei Gao Eunhee Cho
Eunhee Cho Yingying Liu
Yingying Liu Yuan Lu
Yuan Lu
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Pharmacol. , 29 May 2019
Sec. Translational Pharmacology
Volume 10 - 2019 | https://doi.org/10.3389/fphar.2019.00611
This article is part of the Research Topic Cell-Free Production Platforms for Developing Tools and Applications in Pharmacology View all 8 articles
Incorporation of unnatural amino acids (UNAAs) into proteins currently is an active biological research area for various fundamental and applied science. In this context, cell-free synthetic biology (CFSB) has been developed and recognized as a robust testing and biomanufacturing platform for highly efficient UNAA incorporation. It enables the orchestration of unnatural biological machinery toward an exclusive user-defined objective of unnatural protein synthesis. This review aims to overview the principles of cell-free unnatural protein synthesis (CFUPS) systems, their advantages, different UNAA incorporation approaches, and recent achievements. These have catalyzed cutting-edge research and diverse emerging applications. Especially, present challenges and future trends are focused and discussed. With the development of CFSB and the fusion with other advanced next-generation technologies, CFUPS systems would explicitly deliver their values for biopharmaceutical applications.
The incorporation of unnatural amino acids (UNAAs) in protein engineering has emerged as a key discipline in synthetic biology. The 20 standard amino acids (SAAs) on their own can no longer fulfill the growing demands from fundamental and applied science, due to their limited functional chemistries. Replacing the SAAs with UNAAs can provide proteins with novel physicochemical properties and biological functions. Moreover, the advances in cell-free protein synthesis (CFPS) systems have paved the way to accurate and efficient incorporation of UNAAs into proteins. Besides direct chemical reaction with the side chains of SAAs and the biological posttranslational modifications (PTMs) (Zhang et al., 2018), there has been an extensive research in making unnatural amino acids (UNAAs) with novel characteristics and incorporating them into proteins during translation (Liu and Schultz, 2010; Chin, 2017). Cohen and coworkers, for the first time in history, successfully incorporated SeMet into protein in E. coli (Munier and Cohen, 1959). Since then, chemists have created over 200 UNAAs with diverse characteristics (Gfeller et al., 2013), and different research groups have developed many approaches for effective UNAA incorporation in vivo or in vitro.
Site-specific incorporation of UNAAs featuring unique functional groups has been widely applied in fundamental science, including exploring protein properties (Liu and Schultz, 2010), characterizing molecular interactions (Yang et al., 2016), and mimicking eukaryotic PTMs (Brocker et al., 2014). Beyond these, the UNAA incorporation studies also show the growing influences on industrial research. It has opened the road to create novel biomaterials (Albayrak and Swartz, 2014), flexible bioconjugation (Lim and Kwon, 2016), unnatural enzymes (Ravikumar et al., 2015), and on-demand therapeutics (Si et al., 2016). In recent years, increasing attention has been paid on in vitro cell-free UNAA incorporation technology (Lee et al., 2016;Lu, 2019) to enhance the efficiency, achieve broader specificity, and increase fidelity through manipulating codon usage (Gan and Fan, 2017; Martin et al., 2018; Seki et al., 2018; Adachi et al., 2019).
In this review, we discuss the current knowledge, recent research achievements, and significant efforts carried out in the field of cell-free UNAA incorporation. The emerging frontiers, key challenges, and future perspectives are also further discussed.
Cell-free biosynthesis system aims to manipulate biological machinery to perform transcription and translation in vitro without using living cells. Its open nature provides excellent flexibility to design and engineer unnatural proteins with diverse UNAA incorporation. Conventional cell-based in vivo systems are facing many engineering challenges, including the UNAA transportation cross the cell membrane, the cytotoxicity of unnatural biological components, and low incorporation efficiency. In this context, the emerging cell-free unnatural protein synthesis (CFUPS) systems have been adopted to break through those restrictions.
Because of the open operation environment and no cell growth in cell-free systems, there is no need to concern issues such as the UNAA transportation and the cytotoxicity (Hong et al., 2014a). It is beneficial to the flexible regulation of reactions and versatile usage of different approaches for better UNAA incorporation (Des Soye et al., 2015). Compared with cell systems, because there is no complicated substance metabolism and all resources are used for the exclusive synthesis of target proteins, cell-free systems represent a faster method to make proteins with high yields in just a few hours from plasmids or linear DNAs (Carlson et al., 2012). One significant advantage of cell-free systems is the synthesis of proteins that are toxic and difficult to express in cell systems (Lu, 2017). Toxic biopharmaceutical proteins, such as anticancer drug onconase (Salehi et al., 2016), cancer therapeutic protein adenosine diphosphate (ADP)-ribosylases including pierisin-1 (Orth et al., 2011) and Pseudomonas exotoxin (Krinsky et al., 2016), and plasminogen activator urokinase (Yin and Swartz, 2004), have been successfully synthesized with high yields in cell-free systems. Difficult-to-express proteins typically include membrane proteins and protein complexes involving folding and assembly problems, which can be overcome in cell-free systems by directly manipulating the translation conditions. Aglycosylated monoclonal antibodies have been produced by optimizing the oxidizing environment in a Chinese hamster ovary (CHO) cell-free system (Martin et al., 2017). A drug target neuropeptide receptor Y2R has been successfully expressed in soluble form in E. coli cell-free system, and UNAA p-benzoyl-phenylalanine (BPa) was incorporated into it for structural studies (Kögler et al., 2019). The cell-free platform can also serve as a rapid screening pipelines for new drug candidates in a high-throughput manner due to its simple downstream processing without a cell lysis step (Jin and Hong, 2018). Recombinant malaria vaccines (Morita et al., 2017) and antibodies (Kanoi et al., 2017) have been screened by protein microarrays using E. coli or wheat germ cell-free systems. Therefore, CFUPS systems as good high-throughput screening platforms can speed up the design–build–test cycle for unnatural protein engineering (Mankowska et al., 2016). All these advantages have transformed the CFUPS system into a robust UNAA incorporation platform for the synthesis of unnatural proteins with great simplicity and flexibility.
In general, two types of CFUPS systems have been developed for effective UNAA incorporation. One is the extract-based PURE (protein synthesis using recombinant elements) system, which comprises a toolbox of transcriptional and translational components purified from E. coli (Shimizu et al., 2001; Shimizu et al., 2005). The other is the crude extract-based system; the crude cell extract is first prepared without purification, and then the system is supplemented with other essential components, including DNAs, RNA polymerase, energy-providing substrates, amino acids, nucleoside triphosphate (NTP), transfer ribonucleic acids (tRNAs), cofactors, and salts (Lu and Swartz, 2016) (Figure 1). The crude extracts have been made from many different prokaryotic and eukaryotic cells for unnatural protein synthesis. The most commonly used system is prokaryotic E. coli CFUPS system (Lu et al., 2013; Schinn et al., 2017; Martin et al., 2018; Gerrits et al., 2019). Eukaryotic CFUPS systems include wheat germ (plant) (Ogawa et al., 2016), Spodoptera frugiperda (insect) (Taki et al., 2006), and rabbit reticulocyte (mammalian) (Fahmi et al., 2007).
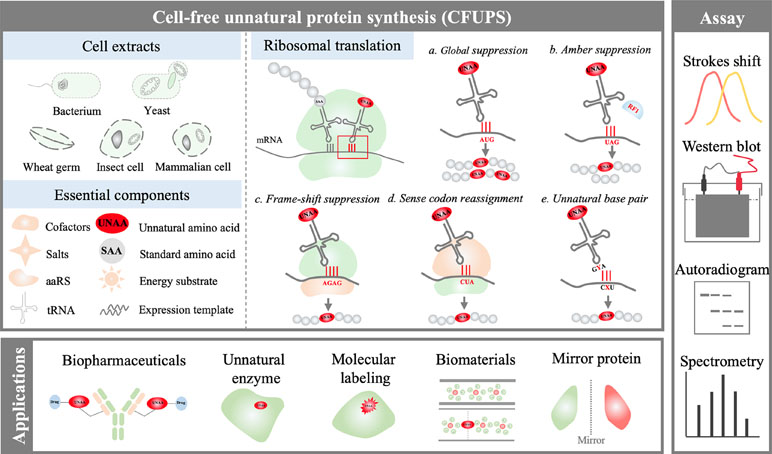
Figure 1 Overview of the cell-free unnatural protein synthesis (CFUPS) system.
In CFUPS systems, UNAAs are incorporated into proteins by transforming unnatural biological machinery, including codon, tRNA, aminoacyl-tRNA synthetase (aaRS), elongation factor (EF), release factor (RF), and ribosome (Des Soye et al., 2015). Because of defined components, the PURE CFUPS system is a potent fundamental tool for basic science study. However, the protein yield of the PURE system is 0.01–0.02 mg/mL and two to four times lower than E. coli crude extract-based cell-free system (Li et al., 2014), and the PURE system costs around 1,000 times more than the crude extract-based system on a protein-per-dollar basis (Hong et al., 2014a; Li et al., 2014). As a result, the crude extract-based CFUPS system is preferred for applied research.
Broadly defined, two UNAA incorporation methods have been developed in CFUPS systems (Figure 1). One is the global suppression method (GSM), which uses native biological machinery to incorporate UNAAs. The other method is the orthogonal translation system (OTS). In the OTS, the codon, tRNA, aaRS, EF, RF, and ribosome are reengineered by rational design or directed evolution (Neumann et al., 2010; Zeng et al., 2014). Compared with the GSM, the OTS is more widely used because of better site-specific UNAA incorporation ability.
In the GSM, the auxotrophic strains are prepared to make the extracts (Singh-Blom et al., 2014), or natural amino acids are depleted from the extracts by size-exclusion chromatographic approaches (Brodel et al., 2014). Afterward, amino acids including the desired UNAAs but without the SAAs to be globally replaced are supplied in the CFUPS reaction system. In this scheme, multiple identical UNAAs can be efficiently incorporated into a single protein (Merkel et al., 2010; Dumas et al., 2015). The lipase can still retain its enzymatic activity when up to 10% of sites are replaced with UNAAs (Merkel et al., 2010). The most used UNAAs in the GSM are methionine analogs, such as homopropargylglycine (HPG) and azidohomoalanine (AHA) (Lu et al., 2013; Lu and Swartz, 2016), to replace standard methionine. The GSM is compatible and can be combined with the OTS method for further genetic codon expansion. However, its apparent shortcoming is that the designed UNAAs must be similar to endogenous SAAs in structure and recognized by native biological machinery (Des Soye et al., 2015).
The difference of the OTS method with the GSM is the exogenous elements of the orthogonal translation pair, including orthogonal aaRS (o-aaRS), orthogonal tRNA (o-tRNA), and corresponding UNAA. The tyrosyl tRNA synthetase TyrRS/tRNACUA pair from Methanocaldococcus jannaschi and the pyrrolysyl tRNA synthetase PylRS/tRNACUA pair from Methanosarcina mazei and Methanosarcina barkeri are widely used (Des Soye et al., 2015; Lim and Kwon, 2016; Chin, 2017). Ideally, the actions of the exogenous orthogonal translation pairs should not be altered by endogenous native translation pairs (Hu et al., 2014). In the OTS method, searching for appropriate codon usage and corresponding efficient o-aaRS/tRNA pairs have been a challenge. Based on the variation in the codon usage, four different strategies have been developed in CFUPS systems, including the stop codon suppression, the frame-shift suppression, the sense codon reassignment, and unnatural base pair.
In the stop codon suppression strategy, as the name implies, three nonsense stop codons (UAG, UAA, UGA) are chosen to encode UNAAs (Bock et al., 1991; Hao et al., 2002; Xie and Schultz, 2006). Because the amber codon UAG is the least used in E. coli, it is most widely used in the UNAA incorporation (Nakamura et al., 2000; Xie and Schultz, 2005; Quast et al., 2015). A significant challenge is that the competition between RFs and suppressor tRNAs for interacting with stop codons causes truncated protein products. To solve this problem, researchers created the genetically engineered E. coli strain which lacks the release factor 1 (RF1) competing with the UAG codon (Johnson et al., 2011; Lajoie et al., 2013). In the RF1-deleted E. coli CFUPS system, the GFP yield increased 2.5 times (Hong et al., 2014b).
The development of the frame-shift suppression strategy aims to address a major challenge that only one kind of UNAA can be encoded in the amber codon suppression strategy. The basic idea is creating an enlarged codon with four or five nucleotides (Neumann et al., 2010). By this approach, four-base or five-base codons have been successfully employed to the UNAA incorporation in prokaryotic E. coli CFUPS system (Hohsaka et al., 2001a; Hohsaka et al., 2001b) and eukaryotic CFUPS systems using rabbit reticulocyte lysates (Hohsaka et al., 2002). Because it is hard to seek effective orthogonal translation pairs, the frame-shift suppression technology develops very slowly in recent years.
The strategy using sense codons to encode UNAAs is defined as the sense codon reassignment. Theoretically, 30 to 40 codons are sufficient to encode genetic information of an organism (O’donoghue et al., 2013), and therefore, more than 20 codons could be reassigned to encode UNAAs (Krishnakumar and Ling, 2014). The main obstacle to this strategy is the competition between native tRNAs and artificial o-tRNAs (Cui et al., 2018).
A more advanced strategy is the unnatural base pair approach, which creates entirely new unnatural codons to encode UNAAs and possesses higher orthogonality than natural codons. The unnatural base pairs, such as Z-P (Chen et al., 2011; Leal et al., 2015), Ds-Px (Hirao et al., 2006), and NaM-5SICS (Malyshev et al., 2014; Zhang et al., 2017), have successfully realized the in vitro transcription and translation for unnatural protein synthesis in CFUPS systems. Further improvement will focus on engineering better DNA polymerases, RNA polymerases, EFs, and ribosomes to recognize the unnatural base pairs (Zhang et al., 2017; Dien et al., 2018).
Incorporation of UNAAs in CFUPS systems needs accurate assessment. The protein truncation can be initially analyzed by fluorescence measurement, sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), Western blot, or autoradiography (Lu et al., 2014; Lu et al., 2015). Mass spectrometry is generally used for accurate UNAA incorporation analysis (Gao et al., 2019). In some reports, the suppression efficiency is used for evaluating the UNAA incorporation, which is defined as the ratio of the active unnatural protein yield to the active native protein yield (Hong et al., 2014b).
Unnatural biopharmaceutical proteins have been attracting great attention in the biomedicine field. The incorporation of UNAAs provides a means for bioconjugation with other molecules to improve the potency (Murray and Baliga, 2013). The UNAA display on the virus-like particle (VLP) using the E. coli CFUPS system can regulate the immunogenicity of VLP (Lu et al., 2015; Ding et al., 2018). The flagellin as an effective immune stimulant can be orderly displayed on the VLPs by the click chemistry reaction of UNAAs with azide and alkyne groups incorporated into the proteins in CFUPS systems, which increases the activity of flagellin by 10 times (Lu et al., 2013; Lu and Swartz, 2016). The malaria vaccine candidate Pfs25 protein was synthesized with a C-terminal UNAA p-azidomethyl phenylalanine (pAMF), which was conjugated with dibenzocyclooctyne (DBCO) derivatized GPI for enhanced activity (Kapoor et al., 2018). In the development of antibody–drug conjugates (ADCs), CFUPS provides a more effective UNAA incorporation approach for stable linking between the antibody and the anticancer drug (Yin et al., 2012; Zimmerman et al., 2014; Currier et al., 2016). The UNAA Hco was incorporated into the target protein to make it sensitive to the inhibitors and small ligand molecules, which could be an attractive tool in the drug discovery (Ugwumba et al., 2011). The CFUPS system can be used as an effective solution for PTMs, such as phosphorylation, which was achieved by the incorporation of specially modified UNAA L-phosphoserine and is capable of producing active human mitogen-activated protein kinase kinase 1 (MEK1) kinase (Oza et al., 2015).
Design of mirror proteins or peptides made up of D-amino acids is another emerging research field for the CFUPS systems (Fujino et al., 2013). Introduction of D-amino acids into proteins or peptides could alter their functions. Many efforts have been made to devise a new ribosomal translation system that is compatible with the incorporation of D-amino acids (Katoh et al., 2017a; Katoh et al., 2017b). Up to 10 D-serine were successfully introduced into peptides, and macrocyclic peptides with five D-amino acids were also successfully expressed (Katoh et al., 2017b). Mirror protein or peptide can be used as a decoy for library screening to generate pharmacologically advantageous therapy (Zhao and Lu, 2014). Mirror peptide holds enhanced stability against protease digestion, and therefore it can circulate in the body for a longer time (Zhao and Lu, 2011). It could be available for oral administration due to the low efficiency of antigen presentation and poor immunogenicity (Welch et al., 2007).
CFUPS can be used as a powerful tool for stable labeling of proteins to study their structure, kinetic, and interaction characteristics (Gerrits et al., 2019). Incorporation of UNAAs was used for investigating the ligand-independent dimerization of functional human epidermal growth factor receptor (Quast et al., 2016). The CFUPS is also expanded to the biomaterials research field, such as protein polymers (Albayrak and Swartz, 2014) and biomimetic membranes (Wei et al., 2018).
Due to the advantages in terms of simplicity and flexibility over cell systems, the CFUPS systems have been well developed to customize proteins with novel structures and functions. Currently, it continues to incubate emerging applications in the biopharmaceuticals, enzymes, mirror proteins, molecular labeling, and biomaterials. However, with the increasing demand from fundamental and applied science, there are still many hurdles to overcome (Figure 2).
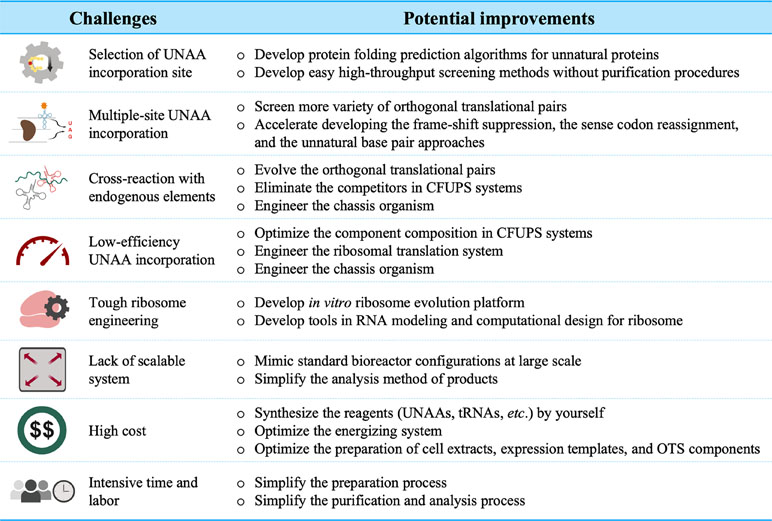
Figure 2 Major challenges in the CFUPS.
Multiple-site UNAA incorporation has been the major challenge. Because the UAA incorporation affects the protein expression and function, the selection of the incorporation sites is crucial, especially for multiple-site incorporation. Many prediction algorithms for the protein folding are based profoundly on homology, making them less useful when considering novel side-chain characteristics of UNAAs. As a consequence, high-throughput screening of incorporation sites has been attempted in CFUPS system (Schinn et al., 2017). Although the screening data offer some explanations on the effects of the incorporation sites, rational site prediction remains challenging. In the amber codon suppression approach, the elimination of the competitor RF1 significantly improves the incorporation of identical UNAAs at multiple sites (Martin et al., 2018; Seki et al., 2018; Adachi et al., 2019). However, incorporating different types of UNAAs at multiple sites is still a daunting task. To accomplish this task, more attention have been paid to the approaches such as the frame-shift suppression, the sense codon reassignment, and unnatural base pair, but recent progress is limited.
Low-efficiency problem always plagues further technology development. The UNAA incorporation efficiency is closely linked to the engineered orthogonal ribosomal translational system. More or less, exogenous UNAAs, o-aaRSs, and o-tRNAs interfere with endogenous SAAs, aaRSs, and tRNAs. The orthogonality needs to be further improved by rational design or directed evolution. However, the hardest part is evolving ribosomes (D’aquino et al., 2018). The reengineering of ribosomes could potentially improve the UNAA incorporation efficiency, increase the orthogonality, produce mirror-image proteins, and synthesize the proteins that are impossible with existing chemistry. Nevertheless, redesigning the ribosome is critical to protein synthesis. The in vitro ribosome evolution platform and computational modeling are required and developed to achieve the goals.
The scale-up and cost are two major concerns in industrial manufacturing. The lack of scalable systems hinders cell-free protein production at an adequate scale (Murray and Baliga, 2013). A 100-L level of cell-free system has been reached for the production of a human cytokine protein (Zawada et al., 2011). However, the scale-up of CFUPS has not been developed. Some applications are often limited by reagent expenses involving the synthesis of unnatural proteins. In E. coli CFUPS system, the total synthesis cost is USD 0.658 per 100 μg unnatural green fluorescent protein with unnatural amino acid para-Propargyloxyphenylalanine (pPaGFP) if using plasmid as the expression template and glucose in the energizing system (Shrestha et al., 2014). To further reduce the cost, efforts can focus on the energy regeneration system, the chassis organism, preparation of orthogonal translational system, and the UNAA synthesis. Apparently, the increase of the UNAA incorporation efficiency and the protein synthesis yield could dramatically decrease the production cost.
In the future development of CFUPS systems, overcoming the challenges presented above could deepen our understanding of living systems, promote the technological advance, expand the industrial applications, and realize significant economic benefits. To maximize the potential of CFUPS platform, it must be fused with other fast-developing technologies in biology, physics, chemistry, medicine, materials science, electronics, and computer science, so that it would open up pioneering and innovative research fields.
All authors listed have worked together to make the frame, compose the context, and revise the manuscript. WG, EC, and YL contributed to this paper equally.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
This work was supported by the National Natural Science Foundation of China (grant nos. 21706144 and 21878173) and the Beijing Natural Science Foundation (grant no. 2192023).

Abe, M., Ohno, S., Yokogawa, T., Nakanishi, T., Arisaka, F., Hosoya, T., et al. (2007). Detection of structural changes in a cofactor binding protein by using a wheat germ cell-free protein synthesis system coupled with unnatural amino acid probing. Proteins 67, 643–652. doi: 10.1002/prot.21341
Adachi, J., Katsura, K., Seki, E., Takemoto, C., Shirouzu, M., Terada, T., et al. (2019). Cell-free protein synthesis using S30 extracts from Escherichia coli RFzero strains for efficient incorporation of non-natural amino acids into proteins. Int. J. Mol. Sci. 20, 492. doi: 10.3390/ijms20030492
Albayrak, C., Swartz, J. R. (2014). Direct polymerization of proteins. ACS Synth. Biol. 3, 353–362. doi: 10.1021/sb400116x
Bock, A., Forchhammer, K., Heider, J., Baron, C. (1991). Selenoprotein synthesis: an expansion of the genetic code. Trends Biochem. Sci. 16, 463–467. doi: 10.1016/0968-0004(91)90180-4
Brocker, M. J., Ho, J. M., Church, G. M., Soll, D., O’donoghue, P. (2014). Recoding the genetic code with selenocysteine. Angew. Chem. Int. Ed. Engl. 53, 319–323. doi: 10.1002/anie.201308584
Brodel, A. K., Sonnabend, A., Kubick, S. (2014). Cell-free protein expression based on extracts from CHO cells. Biotechnol. Bioeng. 111, 25–36. doi: 10.1002/bit.25013
Carlson, E. D., Gan, R., Hodgman, C. E., Jewett, M. C. (2012). Cell-free protein synthesis: applications come of age. Biotechnol. Adv. 30, 1185–1194. doi: 10.1016/j.biotechadv.2011.09.016
Chen, F., Yang, Z., Yan, M., Alvarado, J. B., Wang, G., Benner, S. A. (2011). Recognition of an expanded genetic alphabet by type-II restriction endonucleases and their application to analyze polymerase fidelity. Nucleic Acids Res. 39, 3949–3961. doi: 10.1093/nar/gkq1274
Chin, J. W. (2017). Expanding and reprogramming the genetic code. Nature 550, 53–60. doi: 10.1038/nature24031
Cui, Z., Wu, Y., Mureev, S., Alexandrov, K. (2018). Oligonucleotide-mediated tRNA sequestration enables one-pot sense codon reassignment in vitro. Nucleic Acids Res. 46, 6387–6400. doi: 10.1093/nar/gky365
Currier, N. V., Ackerman, S. E., Kintzing, J. R., Chen, R., Filsinger Interrante, M., Steiner, A., et al. (2016). Targeted drug delivery with an integrin-binding Knottin–Fc–MMAF conjugate produced by cell-free protein synthesis. Mol. Cancer Ther. 15, 1291–1300. doi: 10.1158/1535-7163.MCT-15-0881
D’aquino, A. E., Kim, D. S., Jewett, M. C. (2018). Engineered ribosomes for basic science and synthetic biology. Annu. Rev. Chem. Biomol. Eng. 9, 311–340. doi: 10.1146/annurev-chembioeng-060817-084129
Des Soye, B. J., Patel, J. R., Isaacs, F. J., Jewett, M. C. (2015). Repurposing the translation apparatus for synthetic biology. Curr. Opin. Chem. Biol. 28, 83–90. doi: 10.1016/j.cbpa.2015.06.008
Dien, V. T., Morris, S. E., Karadeema, R. J., Romesberg, F. E. (2018). Expansion of the genetic code via expansion of the genetic alphabet. Curr. Opin. Chem. Biol. 46, 196–202. doi: 10.1016/j.cbpa.2018.08.009
Ding, X., Liu, D., Booth, G., Gao, W., Lu, Y. (2018). Virus-like particle engineering: from rational design to versatile applications. Biotechnol. J. 13, e1700324. doi: 10.1002/biot.201700324
Dumas, A., Lercher, L., Spicer, C. D., Davis, B. G. (2015). Designing logical codon reassignment—expanding the chemistry in biology. Chem. Sci. 6, 50–69. doi: 10.1039/C4SC01534G
Fahmi, N. E., Dedkova, L., Wang, B., Golovine, S., Hecht, S. M. (2007). Site-specific incorporation of glycosylated serine and tyrosine derivatives into proteins. J. Am. Chem. Soc. 129, 3586–3597. doi: 10.1021/ja067466n
Fujino, T., Goto, Y., Suga, H., Murakami, H. (2013). Reevaluation of the D-amino acid compatibility with the elongation event in translation. J. Am. Chem. Soc. 135, 1830–1837. doi: 10.1021/ja309570x
Gan, Q., Fan, C. (2017). Increasing the fidelity of noncanonical amino acid incorporation in cell-free protein synthesis. Biochim biophys acta. General subjects 1861, 3047–3052. doi: 10.1016/j.bbagen.2016.12.002
Gao, W., Bu, N., Lu, Y. (2019). Efficient incorporation of unnatural amino acids into proteins with a robust cell-free system. Methods Protoc. 2, 16. doi: 10.3390/mps2010016
Gerrits, M., Budisa, N., Merk, H. (2019). Site-specific chemoselective pyrrolysine analogues incorporation using the cell-free protein synthesis system. ACS Synth. Biol. 8, 381–390. doi: 10.1021/acssynbio.8b00421
Gfeller, D., Michielin, O., Zoete, V. (2013). SwissSidechain: a molecular and structural database of non-natural sidechains. Nucleic Acids Res. 41, E327–E332. doi: 10.1093/nar/gks991
Hao, B., Gong, W., Ferguson, T. K., James, C. M., Krzycki, J. A., Chan, M. K. (2002). A new UAG-encoded residue in the structure of a methanogen methyltransferase. Science 296, 1462–1466. doi: 10.1126/science.1069556
Hirao, I., Kimoto, M., Mitsui, T., Fujiwara, T., Kawai, R., Sato, A., et al. (2006). An unnatural hydrophobic base pair system: site-specific incorporation of nucleotide analogs into DNA and RNA. Nat. Methods 3, 729–735. doi: 10.1038/nmeth915
Hohsaka, T., Ashizuka, Y., Murakami, H., Sisido, M. (2001a). Five-base codons for incorporation of nonnatural amino acids into proteins. Nucleic Acids Res. 29, 3646–3651. doi: 10.1093/nar/29.17.3646
Hohsaka, T., Ashizuka, Y., Taira, H., Murakami, H., Sisido, M. (2001b). Incorporation of nonnatural amino acids into proteins by using various four-base codons in an Escherichia coli in vitro translation system. Biochemistry 40, 11060–11064. doi: 10.1021/bi0108204
Hohsaka, T., Fukushima, M., Sisido, M. (2002). Nonnatural mutagenesis in E. coli and rabbit reticulocyte lysates by using four-base codons. Nucleic Acids Res. Suppl., 2, 201–202. doi: 10.1093/nass/2.1.201
Hong, S. H., Kwon, Y. C., Jewett, M. C. (2014a). Non-standard amino acid incorporation into proteins using Escherichia coli cell-free protein synthesis. Front. Chem. 2, 34. doi: 10.3389/fchem.2014.00034
Hong, S. H., Ntai, I., Haimovich, A. D., Kelleher, N. L., Isaacs, F. J., Jewett, M. C. (2014b). Cell-free protein synthesis from a release factor 1 deficient Escherichia coli activates efficient and multiple site-specific nonstandard amino acid incorporation. ACS Synth. Biol. 3, 398–409. doi: 10.1021/sb400140t
Hu, C., Chan, S. I., Sawyer, E. B., Yu, Y., Wang, J. (2014). Metalloprotein design using genetic code expansion. Chem. Soc. Rev. 43, 6498–6510. doi: 10.1039/C4CS00018H
Jin, X., Hong, S. H. (2018). Cell-free protein synthesis for producing ‘difficult-to-express’ proteins. Biochem. Eng. J. 138, 156–164. doi: 10.1016/j.bej.2018.07.013
Johnson, D. B. F., Xu, J. F., Shen, Z. X., Takimoto, J. K., Schultz, M. D., Schmitz, R. J., et al. (2011). RF1 knockout allows ribosomal incorporation of unnatural amino acids at multiple sites. Nat. Chem. Biol. 7, 779–786. doi: 10.1038/nchembio.657
Kanoi, B. N., Takashima, E., Morita, M., White, M. T., Palacpac, N. M. Q., Ntege, E. H., et al. (2017). Antibody profiles to wheat germ cell-free system synthesized Plasmodium falciparum proteins correlate with protection from symptomatic malaria in Uganda. Vaccine 35, 873–881. doi: 10.1016/j.vaccine.2017.01.001
Kapoor, N., Vanjak, I., Rozzelle, J., Berges, A., Chan, W., Yin, G., et al. (2018). Malaria derived glycosylphosphatidylinositol anchor enhances anti-Pfs25 functional antibodies that block malaria transmission. Biochemistry 57, 516–519. doi: 10.1021/acs.biochem.7b01099
Katoh, T., Iwane, Y., Suga, H. (2017a). Logical engineering of D-arm and T-stem of tRNA that enhances d-amino acid incorporation. Nucleic Acids Res. 45, 12601–12610. doi: 10.1093/nar/gkx1129
Katoh, T., Tajima, K., Suga, H. (2017b). Consecutive elongation of D-amino acids in translation. Cell Chem Biol 24, 46–54. doi: 10.1016/j.chembiol.2016.11.012
Kögler, L. M., Stichel, J., Kaiser, A., Beck-Sickinger, A. G. (2019). Cell-free expression and photo-crosslinking of the human neuropeptide Y2 receptor. Front. Pharmacol. 10, 1–17. doi: 10.3389/fphar.2019.00176
Krinsky, N., Kaduri, M., Shainsky-Roitman, J., Goldfeder, M., Ivanir, E., Benhar, I., et al. (2016). A simple and rapid method for preparing a cell-free bacterial lysate for protein synthesis. PLoS One 11, 1–13. doi: 10.1371/journal.pone.0165137
Krishnakumar, R., Ling, J. (2014). Experimental challenges of sense codon reassignment: an innovative approach to genetic code expansion. FEBS Lett. 588, 383–388. doi: 10.1016/j.febslet.2013.11.039
Lajoie, M. J., Rovner, A. J., Goodman, D. B., Aerni, H. R., Haimovich, A. D., Kuznetsov, G., et al. (2013). Genomically recoded organisms expand biological functions. Science 342, 357–360. doi: 10.1126/science.1241459
Leal, N. A., Kim, H., Hoshika, S., Kim, M. J., Carrigan, M. A., Benner, S. A. (2015). Transcription, reverse transcription, and analysis of RNA containing artificial genetic components. ACS Synth. Biol. 4, 407–413. doi: 10.1021/sb500268n
Lee, K.-H., Catherine, C., Kim, D.-M. (2016). Enhanced production of unnatural amino acid-containing proteins in a cell-free protein synthesis system. J. Ind. Eng. Chem. 37, 90–94. doi: 10.1016/j.jiec.2016.03.008
Li, J., Gu, L., Aach, J., Church, G. M. (2014). Improved cell-free RNA and protein synthesis system. PLoS One 9, e106232. doi: 10.1371/journal.pone.0106232
Lim, S. I., Kwon, I. (2016). Bioconjugation of therapeutic proteins and enzymes using the expanded set of genetically encoded amino acids. Crit. Rev. Biotechnol. 36, 803–815. doi: 10.3109/07388551.2015.1048504
Liu, C. C., Schultz, P. G. (2010). Adding new chemistries to the genetic code. Annu. Rev. Biochem. 79, 413–444. doi: 10.1146/annurev.biochem.052308.105824
Lu, Y. (2017). Cell-free synthetic biology: engineering in an open world. Synth. Syst. Biotechnol. 2, 23–27. doi: 10.1016/j.synbio.2017.02.003
Lu, Y. (2019). “Chapter 2—advances in cell-free biosynthetic technology,” in Current developments in biotechnology and bioengineering. Eds. Singh, S. P., Pandey, A., Du, G., Kumar, S.. (Cambridge, MA: Elsevier), 23–45. doi: 10.1016/B978-0-444-64085-7.00002-2
Lu, Y., Chan, W., Ko, B. Y., Vanlang, C. C., Swartz, J. R. (2015). Assessing sequence plasticity of a virus-like nanoparticle by evolution toward a versatile scaffold for vaccines and drug delivery. Proc. Natl. Acad. Sci. U. S. A. 112, 12360–12365. doi: 10.1073/pnas.1510533112
Lu, Y., Swartz, J. R. (2016). Functional properties of flagellin as a stimulator of innate immunity. Sci. Rep. 6, 18379. doi: 10.1038/srep18379
Lu, Y., Welsh, J. P., Chan, W., Swartz, J. R. (2013). Escherichia coli-based cell free production of flagellin and ordered flagellin display on virus-like particles. Biotechnol. Bioeng. 110, 2073–2085. doi: 10.1002/bit.24903
Lu, Y., Welsh, J. P., Swartz, J. R. (2014). Production and stabilization of the trimeric influenza hemagglutinin stem domain for potentially broadly protective influenza vaccines. Proc. Natl. Acad. Sci. U. S. A. 111, 125–130. doi: 10.1073/pnas.1308701110
Malyshev, D. A., Dhami, K., Lavergne, T., Chen, T., Dai, N., Foster, J. M., et al. (2014). A semi-synthetic organism with an expanded genetic alphabet. Nature 509, 385–388. doi: 10.1038/nature13314
Mankowska, S. A., Gatti-Lafranconi, P., Chodorge, M., Sridharan, S., Minter, R. R., Hollfelder, F. (2016). A shorter route to antibody binders via quantitative in vitro bead-display screening and consensus analysis. Sci. Rep. 6, 1–11. doi: 10.1038/srep36391
Martin, R. W., Des Soye, B. J., Kwon, Y. C., Kay, J., Davis, R. G., Thomas, P. M., et al. (2018). Cell-free protein synthesis from genomically recoded bacteria enables multisite incorporation of noncanonical amino acids. Nat. Commun. 9, 1203. doi: 10.1038/s41467-018-03469-5
Martin, R. W., Majewska, N. I., Chen, C. X., Albanetti, T. E., Jimenez, R. B. C., Schmelzer, A. E., et al. (2017). Development of a CHO-based cell-free platform for synthesis of active monoclonal antibodies. ACS Synth. Biol. 6, 1370–1379. doi: 10.1021/acssynbio.7b00001
Merkel, L., Schauer, M., Antranikian, G., Budisa, N. (2010). Parallel incorporation of different fluorinated amino acids: on the way to “teflon” proteins. Chembiochem 11, 1505–1507. doi: 10.1002/cbic.201000295
Mikami, S., Kobayashi, T., Yokoyama, S., Imataka, H. (2006). A hybridoma-based in vitro translation system that efficiently synthesizes glycoproteins. J. Biotechnol. 127, 65–78. doi: 10.1016/j.jbiotec.2006.06.018
Morita, M., Takashima, E., Ito, D., Miura, K., Thongkukiatkul, A., Diouf, A., et al. (2017). Immunoscreening of Plasmodium falciparum proteins expressed in a wheat germ cell-free system reveals a novel malaria vaccine candidate. Sci. Rep. 7, 46086. doi: 10.1038/srep46086
Munier, R., Cohen, G. N. (1959). Incorporation of structural analogues of amino acids into bacterial proteins during their synthesis in vivo. Biochim. Biophys. Acta 31, 378–391. doi: 10.1016/0006-3002(59)90011-3
Murray, C. J., Baliga, R. (2013). Cell-free translation of peptides and proteins: from high throughput screening to clinical production. Curr. Opin. Chem. Biol. 17, 420–426. doi: 10.1016/j.cbpa.2013.02.014
Nakamura, Y., Gojobori, T., Ikemura, T. (2000). Codon usage tabulated from international DNA sequence databases: status for the year 2000. Nucleic Acids Res. 28, 292. doi: 10.1093/nar/28.1.292
Neumann, H., Wang, K., Davis, L., Garcia-Alai, M., Chin, J. W. (2010). Encoding multiple unnatural amino acids via evolution of a quadruplet-decoding ribosome. Nature 464, 441–444. doi: 10.1038/nature08817
O’donoghue, P., Ling, J., Wang, Y. S., Soll, D. (2013). Upgrading protein synthesis for synthetic biology. Nat. Chem. Biol. 9, 594–598. doi: 10.1038/nchembio.1339
Ogawa, A., Namba, Y., Gakumasawa, M. (2016). Rational optimization of amber suppressor tRNAs toward efficient incorporation of a non-natural amino acid into protein in a eukaryotic wheat germ extract. Org. Biomol. Chem. 14, 2671–2678. doi: 10.1039/C5OB02533H
Orth, J. H. C., Schorch, B. R., Boundy, S., Ffrench-Constant, R., Kubick, S., Aktories, K. (2011). Cell-free synthesis and characterization of a novel cytotoxic pierisin-like protein from the cabbage butterfly Pieris rapae. Toxicon 57, 199–207. doi: 10.1016/j.toxicon.2010.11.011
Oza, J. P., Aerni, H. R., Pirman, N. L., Barber, K. W., Ter Haar, C. M., Rogulina, S., et al. (2015). Robust production of recombinant phosphoproteins using cell-free protein synthesis. Nat. Commun. 6, 1–7. doi: 10.1038/ncomms9168
Quast, R. B., Ballion, B., Stech, M., Sonnabend, A., Varga, B. R., Wustenhagen, D. A., et al. (2016). Cell-free synthesis of functional human epidermal growth factor receptor: investigation of ligand-independent dimerization in Sf21 microsomal membranes using non-canonical amino acids. Sci. Rep. 6, 34048. doi: 10.1038/srep34048
Quast, R. B., Mrusek, D., Hoffmeister, C., Sonnabend, A., Kubick, S. (2015). Cotranslational incorporation of non-standard amino acids using cell-free protein synthesis. FEBS Lett. 589, 1703–1712. doi: 10.1016/j.febslet.2015.04.041
Ravikumar, Y., Nadarajan, S. P., Yoo, T. H., Lee, C. S., Yun, H. (2015). Unnatural amino acid mutagenesis-based enzyme engineering. Trends Biotechnol. 33, 462–70. doi: 10.1016/j.tibtech.2015.05.002
Salehi, A. S. M., Smith, M. T., Bennett, A. M., Williams, J. B., Pitt, W. G., Bundy, B. C. (2016). Cell-free protein synthesis of a cytotoxic cancer therapeutic: onconase production and a just-add-water cell-free system. Biotechnol. J. 11, 274–281. doi: 10.1002/biot.201500237
Schinn, S. M., Bradley, W., Groesbeck, A., Wu, J. C., Broadbent, A., Bundy, B. C. (2017). Rapid in vitro screening for the location-dependent effects of unnatural amino acids on protein expression and activity. Biotechnol. Bioeng. 114, 2412–2417. doi: 10.1002/bit.26305
Seki, E., Yanagisawa, T., Yokoyama, S. (2018). “Cell-free protein synthesis for multiple site-specific incorporation of noncanonical amino acids using cell extracts from RF-1 deletion E. coli strains,” in Noncanonical amino acids: methods and protocols. Ed. Lemke, E. A. (New York, NY: Springer New York), 49–65. doi: 10.1007/978-1-4939-7574-7_3
Shimizu, Y., Inoue, A., Tomari, Y., Suzuki, T., Yokogawa, T., Nishikawa, K., et al. (2001). Cell-free translation reconstituted with purified components. Nat. Biotechnol. 19, 751–755. doi: 10.1038/90802
Shimizu, Y., Kanamori, T., Ueda, T. (2005). Protein synthesis by pure translation systems. Methods 36, 299–304. doi: 10.1016/j.ymeth.2005.04.006
Shrestha, P., Smith, M. T., Bundy, B. C. (2014). Cell-free unnatural amino acid incorporation with alternative energy systems and linear expression templates. N. Biotechnol. 31, 28–34. doi: 10.1016/j.nbt.2013.09.002
Si, L., Xu, H., Zhou, X., Zhang, Z., Tian, Z., Wang, Y., et al. (2016). Generation of influenza A viruses as live but replication-incompetent virus vaccines. Science 354, 1170–1173. doi: 10.1126/science.aah5869
Singh-Blom, A., Hughes, R. A., Ellington, A. D. (2014). An amino acid depleted cell-free protein synthesis system for the incorporation of non-canonical amino acid analogs into proteins. J. Biotechnol. 178, 12–22. doi: 10.1016/j.jbiotec.2014.02.009
Taki, M., Tokuda, Y., Ohtsuki, T., Sisido, M. (2006). Design of carrier tRNAs and selection of four-base codons for efficient incorporation of various nonnatural amino acids into proteins in Spodoptera frugiperda 21 (Sf21) insect cell-free translation system. J. Biosci. Bioeng. 102, 511–517. doi: 10.1263/jbb.102.511
Ugwumba, I. N., Ozawa, K., De La Cruz, L., Xu, Z. Q., Herlt, A. J., Hadler, K. S., et al. (2011). Using a genetically encoded fluorescent amino acid as a site-specific probe to detect binding of low-molecular-weight compounds. Assay Drug Dev. Technol. 9, 50–57. doi: 10.1089/adt.2010.0306
Wei, P., Zhuang, B., Yu, D., Sharipova, A., Cai, J., Huang, L., et al. (2018). Cell-free expression of unnatural amino acid incorporated aquaporin SS9 with improved separation performance in biomimetic membranes. Biomed. Res. Int. 2018, 3560894. doi: 10.1155/2018/3560894
Welch, B. D., Vandemark, A. P., Heroux, A., Hill, C. P., Kay, M. S. (2007). Potent D-peptide inhibitors of HIV-1 entry. Proc. Natl. Acad. Sci. U. S. A. 104, 16828–16833. doi: 10.1073/pnas.0708109104
Xie, J., Schultz, P. G. (2005). An expanding genetic code. Methods 36, 227–238. doi: 10.1016/j.ymeth.2005.04.010
Xie, J., Schultz, P. G. (2006). A chemical toolkit for proteins—an expanded genetic code. Nat. Rev. Mol. Cell Biol. 7, 775–782. doi: 10.1038/nrm2005
Yang, Y., Song, H., Chen, P. R. (2016). Genetically encoded photocrosslinkers for identifying and mapping protein-protein interactions in living cells. IUBMB Life 68, 879–886. doi: 10.1002/iub.1560
Yin, G., Garces, E. D., Yang, J., Zhang, J., Tran, C., Steiner, A. R., et al. (2012). Aglycosylated antibodies and antibody fragments produced in a scalable in vitro transcription-translation system. MAbs 4, 217–225. doi: 10.4161/mabs.4.2.19202
Yin, G., Swartz, J. R. (2004). Enhancing multiple disulfide bonded protein folding in a cell-free system. Biotechnol. Bioeng. 86, 188–195. doi: 10.1002/bit.10827
Zawada, J. F., Yin, G., Steiner, A. R., Yang, J., Naresh, A., Roy, S. M., et al. (2011). Microscale to manufacturing scale-up of cell-free cytokine production—a new approach for shortening protein production development timelines. Biotechnol. Bioeng. 108, 1570–1578. doi: 10.1002/bit.23103
Zeng, Y., Wang, W., Liu, W. S. R. (2014). Towards reassigning the rare AGG codon in Escherichia coli. Chembiochem 15, 1750–1754. doi: 10.1002/cbic.201400075
Zhang, Y., Park, K. Y., Suazo, K. F., Distefano, M. D. (2018). Recent progress in enzymatic protein labelling techniques and their applications. Chem. Soc. Rev. 47, 9106–9136. doi: 10.1039/C8CS00537K
Zhang, Y., Ptacin, J. L., Fischer, E. C., Aerni, H. R., Caffaro, C. E., San Jose, K., et al. (2017). A semi-synthetic organism that stores and retrieves increased genetic information. Nature 551, 644–647. doi: 10.1038/nature24659
Zhao, L., Lu, W. (2011). D-peptide-based drug discovery aided by chemical protein synthesis. Isr. J. Chem. 51, 868–875. doi: 10.1002/ijch.201100085
Zhao, L., Lu, W. (2014). Mirror image proteins. Curr. Opin. Chem. Biol. 22, 56–61. doi: 10.1016/j.cbpa.2014.09.019
Keywords: cell-free protein synthesis, cell-free synthetic biology, unnatural amino acid, unnatural protein, global suppression, orthogonal translation system
Citation: Gao W, Cho E, Liu Y and Lu Y (2019) Advances and Challenges in Cell-Free Incorporation of Unnatural Amino Acids Into Proteins. Front. Pharmacol. 10:611. doi: 10.3389/fphar.2019.00611
Received: 22 February 2019; Accepted: 15 May 2019;
Published: 29 May 2019.
Edited by:
Frank Bernhard, Goethe-Universität Frankfurt am Main, GermanyReviewed by:
Martin Siemann-Herzberg, University of Stuttgart, GermanyCopyright © 2019 Gao, Cho, Liu and Lu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yuan Lu, eXVhbmx1QHRzaW5naHVhLmVkdS5jbg==
†These authors have contributed equally to this work.
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.