 Quézia Silva Anders
Quézia Silva Anders Jaisa Klauss
Jaisa Klauss Livia Carla de Melo Rodrigues
Livia Carla de Melo Rodrigues Ester Miyuki Nakamura-Palacios
Ester Miyuki Nakamura-Palacios- 1Laboratory of Cognitive Sciences and Neuropsychopharmacology, Program of Post-Graduation in Physiological Sciences, Health Sciences Center, Federal University of Espírito Santo, Vitória, Brazil
- 2Laboratory of Neurotoxicology and Psychopharmacology, Program of Post-Graduation in Physiological Sciences, Federal University of Espírito Santo, Vitória, Brazil
FosB gene heterodimerizes with Jun family proteins to form activator protein 1 (AP-1) complexes that bind to AP-1 sites in responsive genes to regulate transcription in all cells. The genic expression of FosB seems to be modified after long time exposure to drugs of abuse and these changes may be involved in craving and addicted behavior. This study investigated the FosB mRNA expression in peripheral blood lymphocytes of drug addicted patients using real-time PCR approach. Thus, patients with crack-cocaine use disorder (CUD, n = 10), alcohol use disorder (AUD, n = 12), and healthy non-addicted subjects (CONT, n = 12) were assessed. FosB mRNA expression was reduced by 1.15-fold in CUD and 2.17-fold in AUD when compared to CONT. Hedge’s effect size gs over log FosB/Act was of 0.66 for CUD and of 0.30 for AUD when compared to controls. This study showed that FosB mRNA expression was detected in lymphocytes from peripheral blood for the first time, and it was less expressed in drug addicted patients. This molecular technique may constitute a potential peripheral marker for substance use disorder.
Introduction
The use of addictive drugs has been associated with alteration of gene expression in the brain that result in long-term changes on synapses and neural circuits and consequent neuroadaptive and behavioral changes such as tolerance and craving, which may underlie the development and maintenance of drug addiction (Nestler, 2008; Gajewski et al., 2016).
FosB gene heterodimerizes with Jun family proteins to form activator protein 1 (AP-1) complexes that bind to AP-1 sites in responsive genes to regulate transcription in all cells. There is evidence that the FosB genic expression can be modified after long-term drug exposure, increasing craving and aggravating the addictive behavior (Nestler, 2008; Gajewski et al., 2016).
According to Nestler (2012), drugs of abuse actives excitatory synapses increasing Ca2+ channels permeability into the neuron, triggering intracell targets to induce up or down regulation of genic expression. These alteration of genic expression in target genes like CREB, BDNF, and FosB would be involved in the development of a “state of addiction” (Nestler, 2013).
A truncated product of the FosB gene, the delta-FosB, gradually accumulates trough a course of repeated exposure to virtually all drugs of abuse and because of its unusual stability, its levels persist for weeks after drug cessation, mediating the sensitized responses to drug exposure (Nestler, 2008, 2013). Most studies focusing on molecular biology in drug addiction was conducted in rats (Robison and Nestler, 2012). There are no similar studies that has been done in the living brain of human drug addicts yet because of technical limitations. However, recently it has been suggested that the genic expression in peripheral blood lymphocytes may match to their expression state in the brain (Roozafzoon et al., 2010).
In this study, we analyzed FosB mRNA expression in peripheral blood lymphocytes (PBLs) of patients with crack-cocaine and alcohol use disorders in comparison to non-addicted subjects. To our knowledge this is the first study investigating the FosB gene expression in lymphocytes from peripheral blood as a potential marker for drug addiction condition.
Materials and Methods
Subjects
All subjects were informed about the purposes of the experiment by the principal investigator and signed a written consent before entering the study.
Ten patients, who met DSM V criteria for cocaine (crack) use disorder (CUD) and twelve alcohol use disorder patients (AUD) of both genders were successively recruited between October of 2017 and June of 2018 from a public hospital specialized in drug dependence treatment from Espírito Santo State, Brazil. They all received standard treatment provided by this hospital, consisting of psychosocial approaches – conducted by a professional team of psychologists, nurses, social workers, and physicians. They were not using pharmacological medications by the time the blood samples were collected for this study.
The control group was constituted by twelve healthy non-addicted and aged-matched subjects of both genders, recruited among workers from the University Hospital from Federal University of Espírito Santo and Hospital of Military Police of Espírito Santo. They were screened for drug use and included when were completely abstainer for alcohol and/or cocaine (Table 1).
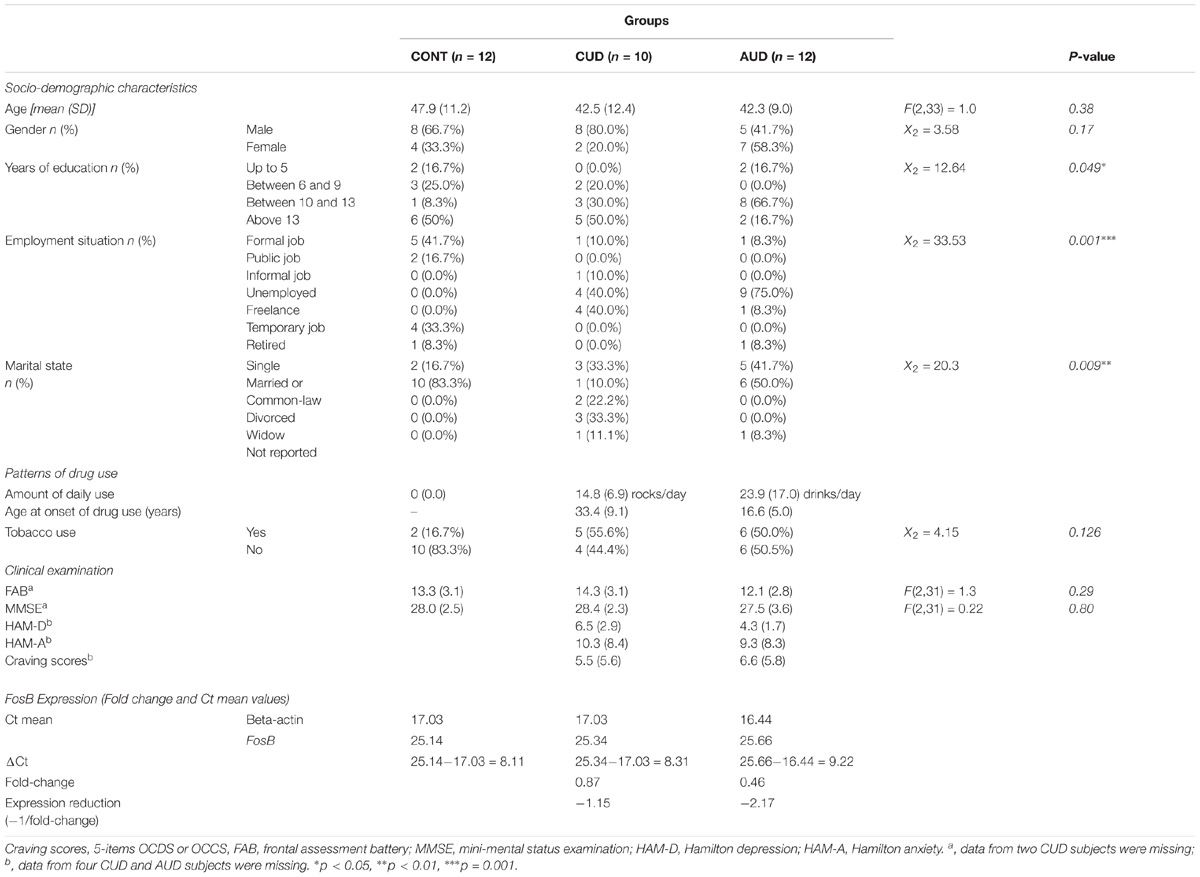
TABLE 1. Socio-demographic characteristics and FosB expression values of the healthy non-user controls (CONT, n = 12) subjects, crack-cocaine use disorder (CUD, n = 10), and alcohol use disorder (AUD, n = 12) patients.
The inclusion criteria for this study were: (1) male and female patients over the age of 18 years; (2) met criteria for crack-cocaine and alcohol dependence according to the ICD-10 Classification of Mental and Behavioral Disorders and the Diagnostic and Statistical Manual of Mental Disorders, fifth edition, as determined by clinical evaluation; (3) in stable clinical condition with no need for emergency care; (4) able to read, write, and speak Portuguese; and (5) no severe withdrawal signs or symptoms at baseline.
Furthermore, exclusion criteria included: (1) a condition of intoxication or withdrawal due to a substance other than crack-cocaine and alcohol, (2) unstable mental or medical disorder or substance abuse or addiction other than crack-cocaine and alcohol dependence, except nicotine and/or caffeine; (3) diagnosis of epilepsy, convulsions, or delirium tremens during abstinence from crack-cocaine and alcohol.
This study was approved by the Brazilian Institutional Review Board of the Federal University of Espírito Santo (CAAE 19403713.6.0000.5060), Brazil. It was conducted in strict adherence to the Declaration of Helsinki and is in accordance with the ethical standards of the Committee on Human Experimentation of the Federal University of Espírito Santo, Vitória, Brazil.
Experimental Protocol
Peripheral blood samples (5 ml) were collected from the cubital vein in tubes containing ethylenediaminetetraacetic acid (EDTA). The interval between this blood collection and the isolation of lymphocytes was of the maximum of 3 h. Total RNA was extracted from lymphocytes using the QIAamp Blood Mini Kit® (Qiagen, Germany), and the amount and purity of RNA was determined by spectrophotometry. Aliquots of RNA were subjected to reverse transcribed for complementary DNA (cDNA) using RT2 First Strand Kit® (Qiagen, Germany) according to the manufacturer’s protocol in a final volume of 20 μl.
The primers used for amplification of FosB and beta-actin genes in real-time PCR reaction were purchased from Qiagen company primer bank.
Real-time quantitative PCR (RT-PCR) was performed using the ABI PRISM 7500 Sequence Detection Systems® (Applied Biosystems, United States) in combination with SYBR green detection (Qiagen, Germany). The reactions were optimized in a 10 ml reaction volume containing 2 μl cDNA, 5 μl RT2 SYBR Green ROX FAST Mastermix® (Qiagen, Germany), 0.4 μl beta-actin (NM_001101.3, Qiagen, Germany) and FosB (NM_006732.2, Qiagen, Germany) and 2.6 μl H2O. The general PCR condition profile was: Taq polymerase activation at 95°C for 10 min, followed by 40 cycles of denaturing at 95°C for 15 s, annealing at 60°C for 1 min, and extension at 95°C for 15 s. After amplification, a melting curve was acquired to determine the optimal PCR conditions.
Data Analysis
The Ct mean for FosB was subtracted from Ct mean for beta-actin for group control, CUD and AUD, yielding ΔCt. Then the ΔCt from control group was subtracted from ΔCt of CUD and AUD, yielding a ΔΔCt values. Fold-change was found using the formula:
2−ΔCtAddicted/2−ΔCtControl and the values expression reduction was found according with Schmittgen and Livak (2008) proceeding the division of −1/calculated fold-change.
Statistical Analysis
ANCOVA was used to compare quantitative data among groups considering age as covariate and the proportion of Ct values of FosB over Ct values of beta-actin (FosB/Act) from each subject converted in logarithm scale (Yamada et al., 2013). SPSS Statistics Base 24.0 (SPSS Inc., United States) and GraphPad Prism 7.0 (GraphPad Software Inc., United States) were employed for statistical analysis and graphic presentations, and the P-value of less than 0.05 was considered statistically significant.
Effect sizes were calculated using Cohen’s d and corrected by Hedges’s gs for between-group comparisons of Log FosB/Act values (Lakens, 2013).
Results
We used RT-PCR to measure mRNA expression levels in human PBLs of CUD and AUD patients in comparison with non-addicted controls.
These groups were well-paired by age and gender, but other socio-demographic characteristics was unequal (Table 1). Schooling was found different among groups (p < 0.05) possibly because of higher proportion of middle school degree in AUD patients when compared to higher school degree in control and CUD patients; employment situation was different among groups (p = 0.001) as larger proportion of CUD and AUD patients was unemployed and/or working as freelancers and smaller proportion of them was formally employed; and marital state was also different among groups (p < 0.01) as higher proportion was married or living in common law in control and AUD groups, whereas CUD patients were mostly single or widow (Table 1). These differences could be expected to be seen in crack-cocaine and alcohol addicted population due to important behavioral and social consequences of these substance use disorders.
Alcohol use disorder and CUD patients used high amount of alcohol drinks and rocks of crack-cocaine per day, respectively (Table 1). More than half of them were tobacco users but not significantly different from control group. No between group differences were found regarding cognitive performance (FAB and MMSE) (Table 1, see Supplementary Material for clinical measurements description). Symptoms of depression in the HAM-D scored into normal range and of anxiety (HAM-A) indicated mild symptoms in both AUD and CUD patients. Craving scores (5-items of OCDS or OCCS) were mild to moderate in both substance use disorder. Mean scores of these clinical measurements are similar to those found in our previous studies in these specific substance use disorders (Klauss et al., 2014; Batista et al., 2015).
Mean Log FosB/Act values from both CUD and AUD patients were slightly larger from the mean found in the control group (Figure 1). However, no statistically significant difference was found in the ANCOVA having age as covariate [F(2,30) = 1.002, p = 0.379, η2 = 0.063]. But, Hedge’s effect size gs was of 0.66 for CUD and of 0.30 for AUD when Log FosB/Act was individually compared to controls.
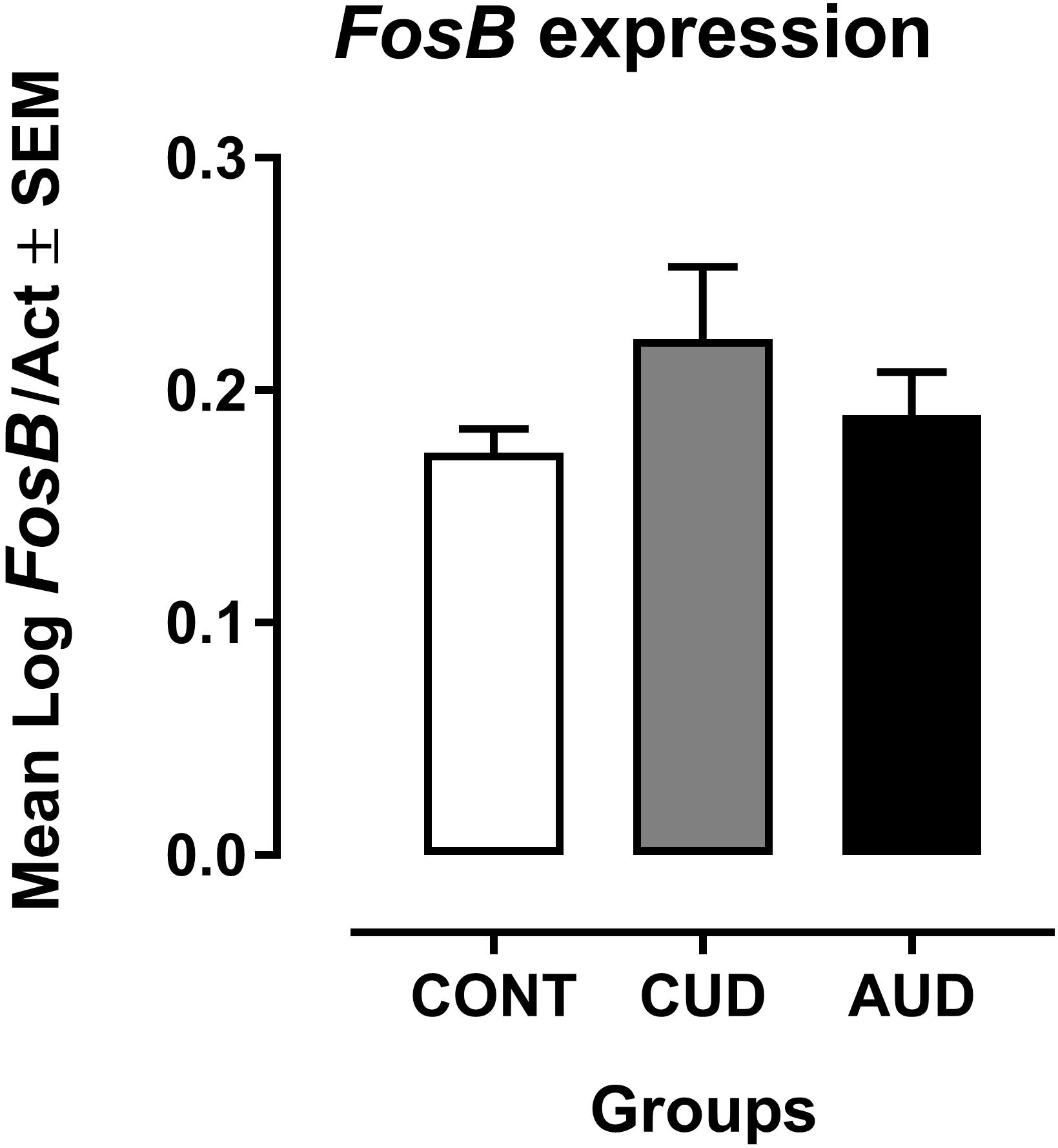
FIGURE 1. Mean Log FosB/Act values from both CUD (n = 10) and AUD (n = 12) patients compared from the mean of the control group (CONT, n = 12). Larger Ct values reflect later threshold cycle, that is, the quantitative cycle in which the target was captured is delayed, meaning that the target is less expressed.
It must be noted that in gene expression, larger Ct values reflect later threshold cycle, that is, the quantitative cycle in which the target was captured is delayed, meaning that the target is less expressed (Schmittgen and Livak, 2008).
In fact, when considering the fold-change analysis, the FosB mRNA was less expressed by 1.15-fold in the CUD patient and by 2.17-fold in the AUD patients when normalized to controls (Table 1).
Discussion
According to “peripheral marker hypothesis,” changes in genetics expression in the brain are reflected in peripheral blood lymphocytes (Roozafzoon et al., 2010). Basing on this theory, our study may constitute the first to demonstre the FosB mRNA expression in lymphocytes from peripheral blood, and to show that it is possibly altered in drug addicted patients.
Hedge’s gs of 0.66 for CUD and of 0.30 for AUD indicate, respectively, medium and small effect sizes according to Cohen (1992) in between-groups comparisons, suggesting that although no significant differences were found in the statistical analysis, effects sizes over controls could be meaningful.
Previous studies in rodents indicated that chronic exposure to drugs modulates brain reward regions through the increased of a FosB isoform (delta-FosB) directly measured in reward circuits (Vialou et al., 2012). We found that expression levels of FosB mRNA was decreased in peripheral blood of CUD and AUD patients. This reduced FosB mRNA expression may reflect the long-term exposition to the drugs as there is experimental evidence that the degree of FosB gene induction partially desensitizes with repeated exposure to amphetamine (Alibhai et al., 2007). Besides, Renthal et al. (2008) demonstrated that accumulation of delta-FosB after chronic amphetamine exposure desensitizes c-fos mRNA induction to a subsequent drug dose.
A recent post-mortem study showed that the expression of FosB isoforms was downregulated in the hippocampus of addicted patients (Gajewski et al., 2016). Differently, we are here measuring in vivo FosB gene expression in PBLs.
Levels of FosB mRNA expression in PBLs may represent a peripheral marker for molecular changes in substance use disorders and could be useful to follow the after effects of drug dependence treatment, especially those that potentially modify synaptic plasticity such as non-invasive brain stimulation (Cirillo et al., 2017), which has shown to decrease craving and relapses to the drug use (Klauss et al., 2014; Batista et al., 2015). This possibility needs to be carefully investigated.
There are limitations that must be considered. The high complexity and cost of the method with limited budget, and the restricted inclusion and exclusion criteria, have limited the number of subjects included in our samples. We have collected 36 samples from non-addicted controls, 27 from AUD and 17 from CUD patients, but included in the analysis only technically adequate samples. Here we explored the expression of only one gene, but other gene expressions such as of BDNF and dopamine receptors are under collection or processing and will be published in the near future.
Patients included in this study were recruited from our major clinical trial registered in ClinicalTrials.gov1,2
In summary, FosB mRNA expression was detected in lymphocytes from peripheral blood and showed to be mildly reduced in drug addicted patients. Thus, with a great caution because these are preliminary data, it may be suggested that this molecular technique could constitute a potential peripheral marker to measure changes in substance use disorders.
Author Contributions
All authors have read and approved the manuscript for submission; have made a substantial contribution to the conception, design, gathering, analysis and/or interpretation of data and a contribution to the writing and intellectual content of the article; and acknowledge that they have exercised due care in ensuring the integrity of the work.
Funding
EN-P was recipient of a researcher fellowship from Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) (proc. 304374/2014-8) and was also funded by this agency (proc. 466650/2014-0). JK and QA were recipient of graduate student fellowship from Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphar.2018.01205/full#supplementary-material
Footnotes
References
Alibhai, I. N., Green, T. A., Potashkin, J. A., and Nestler, E. J. (2007). Regulation of fosB and (fosB mRNA expression: in vivo and in vitro studies. Brain Res. 1143, 22–33. doi: 10.1016/j.brainres.2007.01.069
Batista, E. K., Klauss, J., Fregni, F., Nitsche, M. A., and Nakamura-Palacios, E. M. (2015). A Randomized placebo-controlled trial of targeted prefrontal cortex modulation with bilateral tDCS in patients with crack-cocaine dependence. Int. J. Neuropsychopharmacol. 18, 1–11. doi: 10.1093/ijnp/pyv066
Cirillo, G., Di Pino, G., Capone, F., Ranieri, F., Florio, L., Todisco, V., et al. (2017). Neurobiological after-effects of non-invasive brain stimulation. Brain Stimul. 10, 1–18. doi: 10.1016/j.brs.2016.11.009
Gajewski, P. A., Turecki, G., and Robison, A. J. (2016). Differential expression of fosb proteins and potential target genes in select brain regions of addiction and depression patients. PLoS One 11:355. doi: 10.1371/journal.pone.0160355
Klauss, J., Penido Pinheiro, L. C., Silva Merlo, B. L., Correia Santos, G. D. E., Fregni, F., Nitsche, M. A., et al. (2014). A randomized controlled trial of targeted prefrontal cortex modulation with tDCS in patients with alcohol dependence. Int. J. Neuropsychopharmacol. 17, 1793–1803. doi: 10.1017/S1461145714000984
Lakens, D. (2013). Calculating and reporting effect sizes to facilitate cumulative science: a practical primer for t-tests and ANOVAs. Front. Psychol. 4:863. doi: 10.3389/fpsyg.2013.00863
Nestler, E. J. (2008). Transcriptional mechanisms of addiction: role of DeltaFosB. Philos. Trans. R. Soc. Lond. B Biol. Sci. 363, 3245–3255. doi: 10.1098/rstb.2008.0067
Nestler, E. J. (2012). Transcriptional mechanisms of drug addiction. Clin. Psychopharmacol. Neurosci. 10, 136–143. doi: 10.9758/cpn.2012.10.3.136
Renthal, W., Carle, T. L., Maze, I., Covington, H. E. III, Truong, H.-T., Alibhai, I., et al. (2008). (FosB mediates epigenetic desensitization of the c-fos gene after chronic amphetamine exposure. J. Neurosci. 28, 7344–7349. doi: 10.1523/JNEUROSCI.1043-08.2008
Robison, A. J., and Nestler, E. J. (2012). Transcriptional and epigenetic mechanisms of addiction. Nat. Rev. Neurosci. 12, 623–637. doi: 10.1038/nrn3111
Roozafzoon, R., Goodarzi, A., Vousooghi, N., Sedaghati, M., Yaghmaei, P., and Mohammad-Reza, Z. (2010). Expression of NMDA receptor subunits in human peripheral blood lymphocytes in opioid addiction. J. Pharmacol. 638, 29–32. doi: 10.1016/j.ejphar.2010.04.017
Schmittgen, T. D., and Livak, K. J. (2008). Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 3, 1101–1108. doi: 10.1038/nprot.2008.73
Vialou, V., Feng, J., Robison, A. J., Ku, S. M., Ferguson, D., Scobie, K., et al. (2012). Serum response factor and cAMP response element binding protein are both required for cocaine induction of ΔFosB. J. Neurosci. 32, 7577–7584. doi: 10.1523/JNEUROSCI.1381-12.2012
Keywords: FosB, mRNA expression, lymphocyte, crack-cocaine use disorder, alcohol use disorder
Citation: Anders QS, Klauss J, Rodrigues LCM and Nakamura-Palacios EM (2018) FosB mRNA Expression in Peripheral Blood Lymphocytes in Drug Addicted Patients. Front. Pharmacol. 9:1205. doi: 10.3389/fphar.2018.01205
Received: 29 June 2018; Accepted: 02 October 2018;
Published: 24 October 2018.
Edited by:
Juan J. Canales, University of Tasmania, AustraliaReviewed by:
Fabio Cardoso Cruz, Federal University of São Paulo, BrazilMarcelo Tadeu Marin, Universidade Estadual Paulista Júlio de Mesquita Filho (UNESP), Brazil
Copyright © 2018 Anders, Klauss, Rodrigues and Nakamura-Palacios. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ester Miyuki Nakamura-Palacios, ZW1ucGFsYWNpb3NAZ21haWwuY29t