 Amanda E. Price1†
Amanda E. Price1† Noelle C. Anastasio1,2†
Noelle C. Anastasio1,2† Sonja J. Stutz1
Sonja J. Stutz1 Jonathan D. Hommel1,2
Jonathan D. Hommel1,2 Kathryn A. Cunningham1,2*
Kathryn A. Cunningham1,2*- 1Center for Addiction Research, University of Texas Medical Branch, Galveston, TX, United States
- 2Department of Pharmacology and Toxicology, University of Texas Medical Branch, Galveston, TX, United States
Binge eating disorder (BED) is characterized by dysfunctional hedonic food intake and reward-related processes. Activation of the serotonin (5-HT) 5-HT2C receptor (5-HT2CR) suppresses both food intake and reward-related behaviors and is thus poised to regulate BED. This study assessed the effects of 5-HT2CR activation via the selective 5-HT2CR agonist WAY163909 on binge eating-related behaviors in adult male Sprague-Dawley rats. Low doses of WAY163909 (1.0, 2.0 mg/kg) suppressed high-fat food (HFF) binge intake, but not standard food non-binge intake. WAY163909 (1.0 mg/kg) also attenuated operant responding for self-administered HFF pellets on fixed and progressive ratio schedules of reinforcement, indicating that 5-HT2CR activation suppresses the reinforcing and motivational properties of HFF, respectively. These findings suggest that activation of the 5-HT2CR may be effective at suppressing binge eating in patients with BED via suppression of the reinforcing and motivational properties of HFF. This work supports future studies targeting the 5-HT2CR in the treatment of BED.
Introduction
Binge eating disorder (BED) is characterized by uncontrollable, recurrent episodes of excessive intake of food (American Psychiatric Association, 2013). Of note, food intake during binge episodes is often driven by hedonic rather than homeostatic mechanisms (i.e., food intake driven by wanting and liking factors but not necessary for energy balance; Finlayson et al., 2011; Dalton et al., 2013; Witt and Lowe, 2014). Patients with BED deem high-fat foods (HFFs) more rewarding than people without BED, which may motivate them to consume palatable food (Schebendach et al., 2013), and also exhibit disruptions in reward neurocircuitry (Kessler et al., 2016). Current treatments for BED are comprised of behavioral therapy, off-label use of pharmacotherapies, and lisdexamfetamine, the only clinically approved drug in the treatment of BED (Hutson et al., 2018). One avenue to identify novel treatment approaches in BED is to explore targets known to alter both food intake and reward-related behaviors. The serotonin (5-HT) 5-HT2C receptor (5-HT2CR) fulfills these criteria in that previous studies have demonstrated that 5-HT2CR activation suppresses feeding behavior (Hewitt et al., 2002; Bickerdike, 2003; Voigt and Fink, 2015) via promotion of satiety (for review, Higgins et al., 2017), which in part led to the development and subsequent FDA-approval of the weight loss drug lorcaserin (Belviq®), a first-in-class selective 5-HT2CR agonist (Fidler et al., 2011; O’Neil et al., 2012; Nigro et al., 2013; Aronne et al., 2014). In addition, preclinical studies demonstrate that 5-HT2CR activation also regulates the hedonic properties of rewarding substances such as food and drugs of abuse (for reviews, Higgins and Fletcher, 2003, 2015; Fletcher et al., 2010; Cunningham and Anastasio, 2014). Thus, suppression of the reinforcing and motivational properties of palatable food via 5-HT2CR activation may be one mechanism to decrease hedonic eating and ultimately binge intake.
The investigational compound WAY163909 is a high affinity, full efficacy 5-HT2CR agonist relative to the homologous 5-HT2AR and 5-HT2BR (Dunlop et al., 2005). WAY163909 suppresses food intake in Sprague-Dawley rats, obese Zucker rats, and diet-induced obese mice, effects that are completely reversed by a selective 5-HT2CR antagonist (Dunlop et al., 2005). Furthermore, WAY163909 decreases intake of both drug and sucrose reinforcers (Cunningham et al., 2011; Anastasio et al., 2014; Swinford-Jackson et al., 2016; Berro et al., 2017). To our knowledge, WAY163909 has not been previously tested for efficacy in preclinical binge eating models. Herein, we tested the hypothesis that WAY163909 would suppress binge intake of HFF in an intermittent access model at doses that do not interrupt standard food (SF) non-binge intake in adult male Sprague-Dawley rats. We further explored if WAY163909 alters hedonic eating via suppression of the reinforcing and motivational properties of HFF in two operant conditioning paradigms. These studies provide valuable insight into the potential therapeutic applicability of 5-HT2CR activation in BED.
Materials and Methods
Animals
Naïve male, outbred Sprague-Dawley rats (n = 42; Harlan, Houston, TX, United States) weighing 200–225 g at arrival were housed two per cage (except where noted below) under a 12-h light-dark cycle (lights on between 0600 and 1800 hours) with controlled temperature (21–23°C) and humidity (40–50%). Animals were acclimated for 7 days to the colony room prior to handling and experimental procedures. SF and water were available to rats ad libitum except during daily operant sessions and where noted below. All experiments were conducted in accordance with the NIH Guide for the Care and Use of Laboratory Animals (2011) and with the University of Texas Medical Branch Institutional Animal Care and Use Committee approval.
Food
Standard food available ad libitum and used in non-binge intake studies consisted of 25% protein, 58% carbohydrate, and 17% fat (by kcal; Teklad LM-485 Mouse/Rat Sterilizable Diet; Teklad Diets, Madison, WI, United States; 3.1 kcal/g). HFF employed for binge intake studies contained 20% protein, 35% carbohydrate, and 45% fat (by kcal; D12451, Research Diets, New Brunswick, NJ, United States; 4.73 kcal/g). HFF pellets used in operant assays consisted of 16% protein, 38% carbohydrate, and 46% fat (by kcal; BioServ product #F06162, 45 mg/pellet, Flemington, NJ, United States; 4.60 kcal/g).
Drugs
WAY163909 [(7b-R,10a-R)-1,2,3,4,8,9,10,10a-octahydro-7bH-cyclopenta[b][1,4] diazepino [6,7,1hi]indole] was a gift from Pfizer, Inc. (New York, NY, United States) and was dissolved in 0.9% NaCl (vehicle, VEH). WAY163909 was tested at a dose range (0–2 mg/kg) that did not alter total horizontal ambulation in a motor activity monitor (3 mg/kg WAY163909 has been shown to significantly suppress total horizontal ambulation) but dose-dependently suppressed operant responding for self-administered sucrose pellets (Cunningham et al., 2011). These effects are completely blocked following pretreatment with the selective 5-HT2CR antagonist SB242084 (Dunlop et al., 2005; Cunningham et al., 2011). All injections were administered intraperitoneally (i.p.) in a volume of 1 ml/kg.
HFF Binge Intake
Binge intake of HFF was assessed as described previously in a HFF intermittent access model (Benzon et al., 2014). Briefly, single-housed rats (n = 9) were acclimated to exclusive ad libitum access to HFF for 1 week to prevent food neophobia. Following this acclimation timeframe, rats were provided with exclusive ad libitum access to SF except during binge intake testing. The effects of 5-HT2CR activation on binge intake were determined after i.p. injection of vehicle or 0.5, 1, or 2 mg/kg WAY163909 15 min prior to the beginning of the dark cycle (1745 hours). At 1800 hours, SF was removed and 40 g of HFF was added to the home cage. HFF was removed 2 h later (2000 hours) and weighed to determine binge intake. Rats were then provided ad libitum access to SF. All rats received each of the four pharmacological treatments prior to HFF binge intake testing in a randomized manner with testing spaced at least 1 week apart. Previously published results using this paradigm demonstrate that rats receiving continuous access to HFF eat an average of 5 g of HFF in 2 h, whereas rats subjected to intermittent access to HFF eat an average of 7 g of HFF in 2 h (i.e., binge intake; Benzon et al., 2014). Thus, rats which consumed <5 g HFF in 2 h after vehicle pretreatment were excluded for not exhibiting HFF binge intake (n = 2). An additional rat was excluded as an outlier (i.e., intake greater than two standard deviations from the mean).
SF Non-binge Intake
The effect of 5-HT2CR activation on non-binge SF intake was also assessed in single-housed rats (n = 8) that were injected i.p. with vehicle or 0.5, 1, or 2 mg/kg WAY163909 15 min prior to the beginning of the dark cycle (1745 hours). At 1800 hours, all but 40 g of SF was removed from the home cage. At the end of 2 h (2000 hours), SF was removed and weighed to determine non-binge intake, and rats were allowed ad libitum access to SF. Rats received each of the four treatments prior to non-binge intake testing in a randomized manner at least 1 week apart.
Operant Conditioning for Self-Administration of HFF Pellets
Rats were trained to self-administer HFF pellets via an operant conditioning paradigm. Operant studies took place between 0900 and 1200 hours in standard operant chambers housed within a ventilated and sound-attenuated chamber and equipped with two retractable levers (Med Associates, Georgia, VT, United States). Operant studies consisted of 30-min sessions (5 days/week) during which rats were trained to lever press for a HFF pellet. Completion of the fixed ratio (FR) or progressive ratio (PR) schedule of reinforcement on the active lever resulted in delivery of the reinforcer (one HFF pellet); on the FR schedule, pellet delivery was paired with a discrete, flashing light. There were no scheduled consequences for lever presses on the inactive lever.
Effects of WAY163909 on FR Responding
Rats (n = 9) were SF restricted to 85–90% of free-feeding levels for the first 3 days of operant conditioning to facilitate acquisition of HFF self-administration and then provided with ad libitum access to SF while in the home cage for the remainder of the study. Rats were trained on an FR1 schedule of reinforcement for HFF pellets for 5 days, an FR3 schedule of reinforcement for 2 days, and moved to an FR5 schedule of reinforcement for the remainder of the study. The criterion for stable FR acquisition and responding (<25% variability in the number of HFF pellets earned over three consecutive FR5 training sessions) was achieved prior to initiation of test sessions. Once stable, rats underwent two consecutive days of testing in which they received vehicle on 1 day and 1 mg/kg WAY163909 the following day. After allowing at least 3 days for washout of drug and re-establishment of stability (Cunningham et al., 2011), rats underwent two additional consecutive days of testing in which they received vehicle on 1 day and 0.3 mg/kg WAY163909 the following day. All injections were administered i.p. 15 min prior to the beginning of the operant session. One rat was removed from the study for failure to achieve the stability criterion.
Effects of WAY163909 on PR Responding
Rats (n = 16) were SF restricted to 85–90% of free-feeding levels for the first 3 days of operant conditioning to facilitate acquisition of HFF self-administration and then provided with ad libitum access to SF while in the home cage for the remainder of the study. Rats were trained on an FR1 schedule of reinforcement for HFF pellets for 5 days, an FR3 schedule of reinforcement for 2 days, an FR5 schedule of reinforcement for 2 days, and then moved to a PR schedule of reinforcement for the remainder of the study. The PR schedule of reinforcement (1, 2, 4, 6, 9, 12, 15, 20, 25, 32, 40, 50, 62, 77, and 95) required rats to progressively increase the number of active lever presses needed to receive a single HFF reinforcer (Richardson and Roberts, 1996; McCue et al., 2017). PR sessions ended 10 min after the last reinforcer was received. The criterion for stable PR acquisition and responding (<25% variability in the number of HFF pellets earned over three consecutive PR training sessions) was achieved prior to initiation of test sessions. Following achievement of stability, rats were tested with 1 mg/kg of WAY163909, 0.3 mg/kg of WAY163909, and vehicle with at least 3 days between tests to allow for washout of drug and re-establishment of stability (Cunningham et al., 2011). All injections were administered i.p. 15 min prior to the beginning of the operant session. Four rats were excluded for not achieving stability and an additional rat was removed from the study due to an equipment malfunction.
Statistical Analyses
A repeated measures, one-way analysis of variance (ANOVA) was employed to assess the main effect of WAY163909 treatment on HFF binge intake, 2-h SF non-binge intake, and measures of operant responding for self-administration of HFF pellets (i.e., active and inactive lever presses, pellets earned, breakpoint, and latency to first reinforcer; Cunningham et al., 2011). Subsequent a priori comparisons to vehicle were analyzed using a one-tailed Dunnett’s procedure. A paired Student’s t-test was employed to assure consistent baseline responding between FR vehicle test days and between the days preceding the first and last PR tests. All statistical analyses were conducted with an experiment-wise error rate of α = 0.05 in SAS for Windows 9.4. Power analyses were completed using SPSS Statistics Version 25.
Results
HFF Binge and SF Non-binge Intake
The effect of the 5-HT2CR agonist WAY163909 on HFF binge intake was assessed in an intermittent HFF access model (n = 6). A repeated measures, one-way ANOVA revealed a main effect of treatment (F3,15 = 4.79; p = 0.0155). A priori comparisons demonstrated that 1 and 2 mg/kg of WAY163909 significantly suppressed binge intake compared to vehicle treatment (p < 0.05; Figure 1A). A power analysis determined an achieved power of 0.806 for this assessment. These data indicate that activation of 5-HT2CR signaling suppresses binge eating. The effect of WAY163909 on 2-h, non-binge SF intake was also assessed (n = 8). A repeated measures one-way ANOVA revealed no main effect of treatment (F3,21 = 1.12; p = 0.3642; Figure 1B). These results are consistent with published results indicating the ED50 for WAY163909-induced suppression of 2-h food intake in 24-h fasted Sprague-Dawley rats is 2.93 mg/kg (Dunlop et al., 2005). Together, these data indicate that lower doses of WAY163909 preferentially suppress HFF binge intake over non-binge intake of SF.
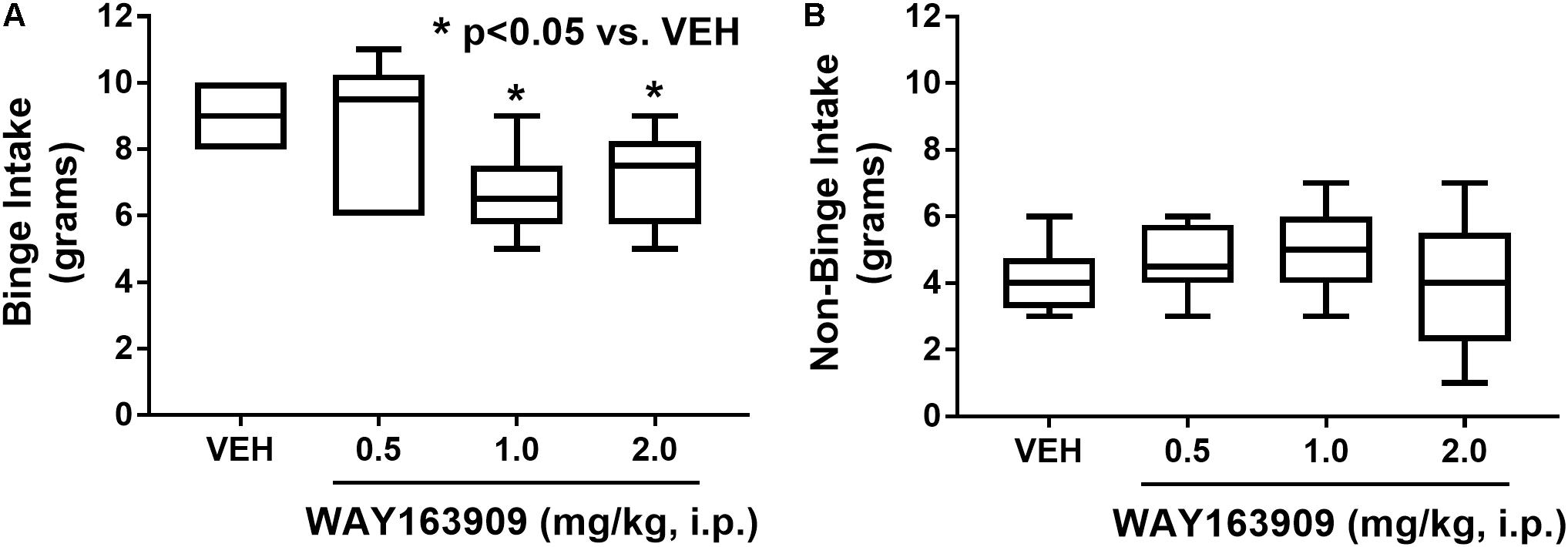
FIGURE 1. WAY163909 preferentially suppresses high-fat food (HFF) binge intake over standard food (SF) non-binge intake. (A) WAY163909 (1.0, 2.0 mg/kg) suppressed HFF binge intake in an intermittent access binge intake paradigm (n = 6). (B) WAY163909 did not alter SF non-binge intake (n = 8). ∗p < 0.05 vs. VEH (Dunnett’s test); boxes extend from the 25th to 75th percentiles, with whiskers representing minimum and maximum values and the line within the box representing the median value.
Operant Responding for Self-Administration of HFF Pellets
The effect of WAY163909 on the reinforcing value of HFF was assessed using FR responding for HFF pellets in freely fed rats (n = 8). A paired Student’s t-test between the first and second vehicle test indicated there were no significant differences in active lever presses (p = 0.934), inactive lever presses (p = 0.621), pellets earned (p = 0.605), or latency to first reinforcer (p = 0.089); thus, the average vehicle response was used as control for analysis of the WAY163909 dose–response relationship. A repeated measures, one-way ANOVA revealed a main effect of treatment on active lever presses (F2,14 = 47.54; p < 0.0001) and pellets earned (F2,14 = 42.08; p < 0.0001), but not for inactive lever presses (F2,14 = 0.34; p = 0.7185) or latency to first reinforcer (F2,14 = 1.39; p = 0.2819). A priori comparisons indicated that 1 mg/kg of WAY163909 significantly suppressed active lever presses (p < 0.05) and pellets earned (p < 0.05) compared to vehicle (Figure 2). These data indicate that activation of the 5-HT2CR suppresses the reinforcing value of HFF, a finding which coalesces with previously published results demonstrating that WAY163909 also dose-dependently suppresses operant responding for self-administration of sucrose pellets (Cunningham et al., 2011).
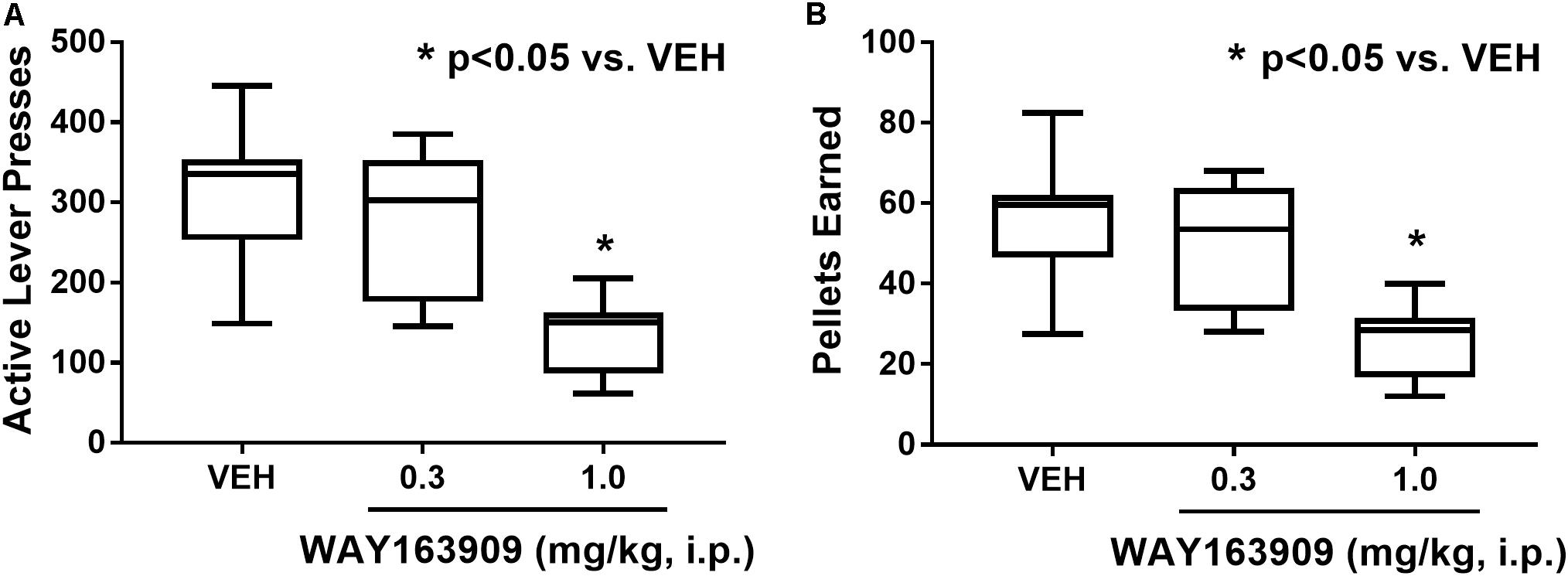
FIGURE 2. WAY163909 suppresses the reinforcing efficacy of high-fat food (HFF). WAY163909 (1.0 mg/kg) suppressed (A) active lever presses and (B) pellets earned during self-administration of HFF pellets on a fixed ratio schedule of reinforcement (n = 8). ∗p < 0.05 vs. VEH (Dunnett’s test); boxes extend from the 25th to 75th percentiles, with whiskers representing minimum and maximum values and the line within the box representing the median value.
The effect of WAY163909 on the motivational value of HFF was assessed using PR responding for HFF pellets in freely fed rats (n = 11). A paired Student’s t-test was used to assess consistent baseline responding between the days preceding the first and last test sessions; analyses indicated there was no difference in active lever presses (p = 0.819) or pellets earned (p = 0.714). A repeated measures one-way ANOVA revealed a main effect of treatment on active lever presses (F2,20 = 9.12; p = 0.0015) and breakpoint (F2,20 = 19.11; p < 0.0001), but not inactive lever presses (F2,20 = 2.59; p = 0.0998) or latency to first reinforcer (F2,20 = 1.81; p = 0.1888). A priori comparisons indicated that 1 mg/kg of WAY163909 significantly suppressed active lever presses (p < 0.05) and breakpoint (p < 0.05) compared to vehicle treatment (Figure 3). These data indicate that 5-HT2CR activation suppresses the motivational value of HFF.
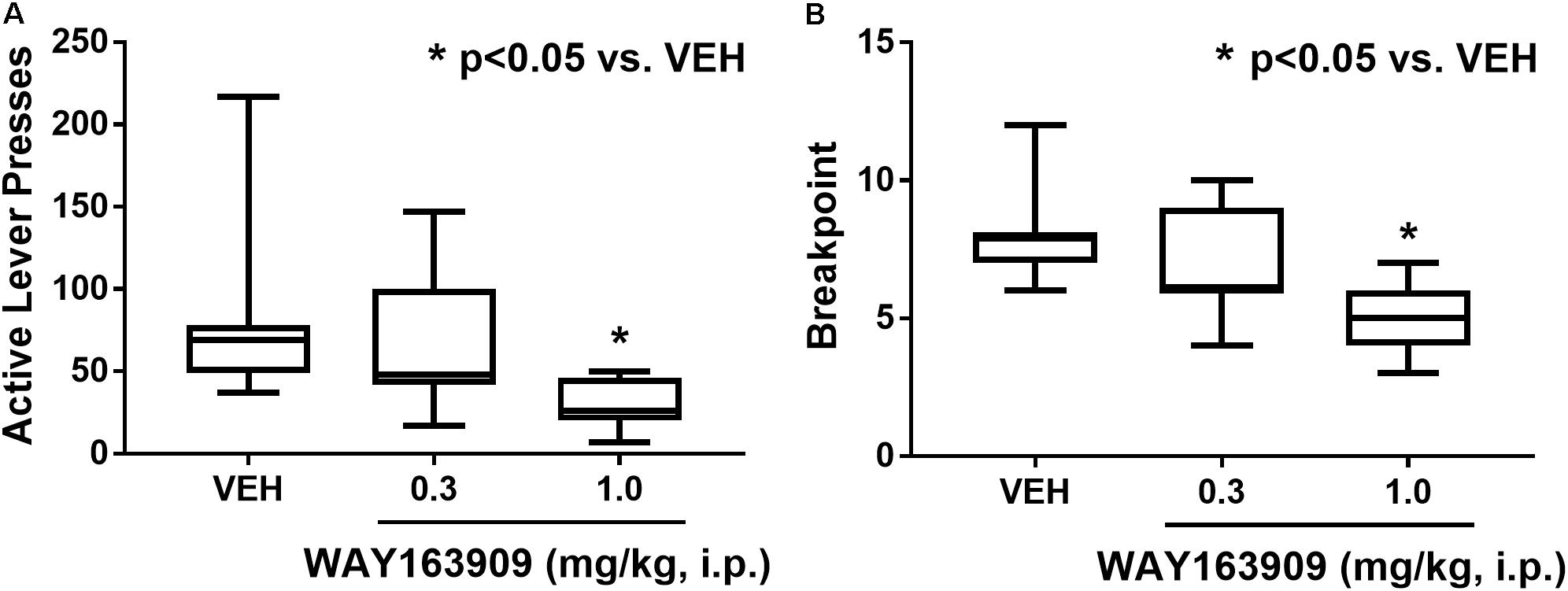
FIGURE 3. WAY163909 suppresses the motivational value of high-fat food (HFF). WAY163909 (1.0 mg/kg) suppressed (A) active lever presses and (B) breakpoint during self-administration of HFF pellets on a progressive ratio schedule of reinforcement (n = 11). ∗p < 0.05 vs. VEH (Dunnett’s test); boxes extend from the 25th to 75th percentiles, with whiskers representing minimum and maximum values and the line within the box representing the median value.
Discussion
Food intake can be described as homeostatic (intake necessary to maintain energy balance) or hedonic (intake driven by reward-related factors; Lutter and Nestler, 2009). Hedonic processes are postulated as an important component of binge eating episodes seen in disorders such as BED (Finlayson et al., 2011). To model binge eating in rodents, we gave rats ad libitum access to standard chow along with intermittent access to a highly palatable food (Corwin and Wojnicki, 2006; Corwin et al., 2011; Benzon et al., 2014). The palatable food used in this study was a HFF chow, which is nutritionally representative of foods that patients with BED may eat in excess during a binge episode (Corwin et al., 2011). Administration of 1 mg/kg of WAY163909 significantly suppressed HFF binge intake in rodents, suggesting that 5-HT2CR activation may be a viable therapeutic approach to suppress binge eating episodes in patients with BED. This same dose of WAY163909 did not alter non-binge intake, consistent with previous literature that higher doses of WAY163909 (i.e., 3–10 mg/kg) are required to suppress 2-h SF intake in 24-h fasted male Sprague-Dawley rats (Dunlop et al., 2005). This suggests that 5-HT2CR activation preferentially suppresses hedonic intake of food at lower doses but attenuates both hedonic and homeostatic intake of food at higher doses. We then demonstrated that 1 mg/kg of WAY163909 suppressed both FR and PR responding, suggesting that 5-HT2CR activation attenuates the reinforcing efficacy and motivational properties of HFF, respectively. These findings are congruent with literature demonstrating 5-HT2CR activation decreases intake of other types of palatable food, such as those high in carbohydrates (Martin et al., 1998; Fletcher et al., 2010; Higgins et al., 2013b).
Activation of the 5-HT2CR is postulated to primarily suppress feeding by increasing production of α-melanocyte stimulating hormone which acts on melanocortin 4 receptors in the paraventricular nucleus of the hypothalamus to promote satiety (Heisler et al., 2002; Xu et al., 2008; Lam et al., 2008). However, 5-HT2CR activation-induced suppression of operant responding for food in self-administration studies suggests that the 5-HT2CR regulates intake of food in additional ways since total food intake in these paradigms is often not enough to promote satiety (Higgins et al., 2013a). Previous studies have also indicated that 5-HT2CR agonists suppress intake of palatable food in non-food deprived rats (Rowland et al., 2008; Canal et al., 2014). A recent study demonstrated that the clinically approved 5-HT2CR agonist lorcaserin suppressed both binge-like eating and hunger-driven feeding in wild-type mice (Xu et al., 2017). Interestingly, lorcaserin-induced suppression of binge-like eating in mice is dependent upon 5-HT2CR expression on dopaminergic neurons in the ventral tegmental area, cells highly implicated in reward-related behaviors (Xu et al., 2017). In mice with selective knockout of 5-HT2CR in dopaminergic neurons, lorcaserin is unable to suppress binge-like eating, suggesting that the 5-HT2CR is mediating its effects on hedonic feeding through mechanisms beyond hypothalamus-dependent promotion of satiety (Xu et al., 2017). Indeed, lorcaserin also suppresses PR responding for chocolate pellets via activation of 5-HT2CR in the ventral tegmental area (Valencia-Torres et al., 2017), further supporting a role for mesolimbic 5-HT2CR in the control of palatable food intake. To our knowledge, the role of the 5-HT2CR in hypothalamic neurons has not been explored in binge eating behavior, but future studies should assess if the 5-HT2CR in hypothalamic subregions contributes to WAY163909-mediated suppression of binge eating because of the well-established role of this population in mediating food intake.
Elevations in body weight are seen in approximately 70% of patients with BED (Kessler et al., 2013). Thus, a medication that effectively suppresses both binge episodes and overall food intake at therapeutic doses would be highly beneficial in the treatment of comorbid BED and obesity. The FDA-approved selective 5-HT2CR agonist lorcaserin may be of benefit for this population of patients. Interestingly, clinical trials for lorcaserin showed that while, on average, treatment produced modest effects (about 3% weight loss when accounting for the effects of placebo), lorcaserin treatment resulted in 5% or even 10% body weight loss in certain subpopulations, a phenomenon occurring at twice the frequency in the lorcaserin group compared to the placebo group (Aronne et al., 2014). Our findings combined with previous literature suggest that 5-HT2CR activation suppresses hedonic intake of food in addition to homeostatic intake of food; thus, lorcaserin may show higher efficacy in patients seeking weight loss treatment with comorbid BED compared to patients without BED. To our knowledge, a behavioral subtyping of the individuals who exhibit the greatest weight loss upon lorcaserin treatment has not yet been reported. The present study suggests that analyses assessing the efficacy of lorcaserin in different behavioral subtypes of obesity are warranted.
This study supports 5-HT2CR activation as a novel therapeutic target to suppress hedonic food intake in patients with BED. Future studies should assess the viability of repurposing the weight loss drug, lorcaserin, in the treatment of BED. These findings, in combination with the larger body of literature surrounding the role of 5-HT2CR in food intake, suggest that 5-HT2CR activation may be especially helpful in the treatment of comorbid BED and obesity.
Author Contributions
NA, JH, and KC planned the experiments. NA and SS completed the experiments. AP and NA analyzed the data and interpreted the results. AP drafted the manuscript. All authors edited and approved the manuscript.
Funding
This work was supported by NIDA grants T32 DA007287 (AP), F30 DA042617 (AP), P50 DA033935 (KC, NA), and K05 DA020087 (KC), the Klarman Family Foundation, and the Center for Addiction Research at UTMB. WAY163909 was a generous gift from Pfizer, Inc.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
American Psychiatric Association (2013). Feeding and Eating Disorders. Washington, DC: American Psychiatric Publishing.
Anastasio, N. C., Liu, S., Maili, L., Swinford, S. E., Lane, S. D., Fox, R. G., et al. (2014). Variation within the serotonin (5-HT) 5-HT2C receptor system aligns with vulnerability to cocaine cue reactivity. Transl. Psychiatry 4:e369. doi: 10.1038/tp.2013.131
Aronne, L., Shanahan, W., Fain, R., Glicklich, A., Soliman, W., Li, Y., et al. (2014). Safety and efficacy of lorcaserin: a combined analysis of the BLOOM and BLOSSOM trials. Postgrad. Med. 126, 7–18. doi: 10.3810/pgm.2014.10.2817
Benzon, C. R., Johnson, S. B., McCue, D. L., Li, D., Green, T. A., and Hommel, J. D. (2014). Neuromedin U receptor 2 knockdown in the paraventricular nucleus modifies behavioral responses to obesogenic high-fat food and leads to increased body weight. Neuroscience 258, 270–279. doi: 10.1016/j.neuroscience.2013.11.023
Berro, L. F., Perez Diaz, M., Maltbie, E., and Howell, L. L. (2017). Effects of the serotonin 2C receptor agonist way163909 on the abuse-related effects and mesolimbic dopamine neurochemistry induced by abused stimulants in rhesus monkeys. Psychopharmacology 234, 2607–2617. doi: 10.1007/s00213-017-4653-2
Bickerdike, M. J. (2003). 5-HT2C receptor agonists as potential drugs for the treatment of obesity. Curr. Top. Med. Chem. 3, 885–897. doi: 10.2174/1568026033452249
Canal, C. E., Morgan, D., Felsing, D., Kondabolu, K., Rowland, N. E., Robertson, K. L., et al. (2014). A novel aminotetralin-type serotonin (5-HT) 2C receptor-specific agonist and 5-HT2A competitive antagonist/5-HT2B inverse agonist with preclinical efficacy for psychoses. J. Pharmacol. Exp. Ther. 349, 310–318. doi: 10.1124/jpet.113.212373
Corwin, R. L., Avena, N. M., and Boggiano, M. M. (2011). Feeding and reward: perspectives from three rat models of binge eating. Physiol. Behav. 104, 87–97. doi: 10.1016/j.physbeh.2011.04.041
Corwin, R. L., and Wojnicki, F. H. (2006). Binge eating in rats with limited access to vegetable shortening. Curr. Protoc. Neurosci. 36, 9.23B.1–9.23B.11. doi: 10.1002/0471142301.ns0923bs36
Cunningham, K. A., and Anastasio, N. C. (2014). Serotonin at the nexus of impulsivity and cue reactivity in cocaine addiction. Neuropharmacology 76(Pt B), 460–478. doi: 10.1016/j.neuropharm.2013.06.030
Cunningham, K. A., Fox, R. G., Anastasio, N. C., Bubar, M. J., Stutz, S. J., Moeller, F. G., et al. (2011). Selective serotonin 5-HT2C receptor activation suppresses the reinforcing efficacy of cocaine and sucrose but differentially affects the incentive-salience value of cocaine- vs. sucrose-associated cues. Neuropharmacology 61, 513–523. doi: 10.1016/j.neuropharm.2011.04.034
Dalton, M., Blundell, J., and Finlayson, G. (2013). Effect of BMI and binge eating on food reward and energy intake: further evidence for a binge eating subtype of obesity. Obes. Facts 6, 348–359. doi: 10.1159/000354599
Dunlop, J., Sabb, A. L., Mazandarani, H., Zhang, J., Kalgaonker, S., Shukhina, E., et al. (2005). Way-163909 ((7bR,10aR)-1,2,3,4,8,9,10,10a-octahydro-7bH-cyclopenta-[b][1,4]diazepino[ 6,7,1hi]indole): a novel 5-HT2C receptor selective agonist with anorectic activity. J. Pharmacol. Exp. Ther. 313, 862–869. doi: 10.1124/jpet.104.075382
Fidler, M. C., Sanchez, M., Raether, B., Weissman, N. J., Smith, S. R., Shanahan, W. R., et al. (2011). A one-year randomized trial of lorcaserin for weight loss in obese and overweight adults: the BLOSSOM trial. J. Clin. Endocrinol. Metab. 96, 3067–3077. doi: 10.1210/jc.2011-1256
Finlayson, G., Arlotti, A., Dalton, M., King, N., and Blundell, J. E. (2011). Implicit wanting and explicit liking are markers for trait binge eating. A susceptible phenotype for overeating. Appetite 57, 722–728. doi: 10.1016/j.appet.2011.08.012
Fletcher, P. J., Sinyard, J., and Higgins, G. A. (2010). Genetic and pharmacological evidence that 5-HT2C receptor activation, but not inhibition, affects motivation to feed under a progressive ratio schedule of reinforcement. Pharmacol. Biochem. Behav. 97, 170–178. doi: 10.1016/j.pbb.2010.07.002
Heisler, L. K., Cowley, M. A., Tecott, L. H., Fan, W., Low, M. J., Smart, J. L., et al. (2002). Activation of central melanocortin pathways by fenfluramine. Science 297, 609–611. doi: 10.1126/science.1072327
Hewitt, K. N., Lee, M. D., Dourish, C. T., and Clifton, P. G. (2002). Serotonin 2C receptor agonists and the behavioural satiety sequence in mice. Pharmacol. Biochem. Behav. 71, 691–700. doi: 10.1016/S0091-3057(01)00709-2
Higgins, G. A., and Fletcher, P. J. (2003). Serotonin and drug reward: focus on 5-HT(2C) receptors. Eur. J. Pharmacol. 480, 151–162. doi: 10.1016/j.ejphar.2003.08.102
Higgins, G. A., and Fletcher, P. J. (2015). Therapeutic potential of 5-HT2C receptor agonists for addictive disorders. ACS Chem. Neurosci. 6, 1071–1088. doi: 10.1021/acschemneuro.5b00025
Higgins, G. A., Sellers, E. M., and Fletcher, P. J. (2013a). From obesity to substance abuse: therapeutic opportunities for 5-HT2C receptor agonists. Trends Pharmacol. Sci. 34, 560–570. doi: 10.1016/j.tips.2013.08.001
Higgins, G. A., Silenieks, L. B., Lau, W., de Lannoy, I., Lee, D. K., Izhakova, J., et al. (2013b). Evaluation of chemically diverse 5-HT2C receptor agonists on behaviours motivated by food and nicotine and on side effect profiles. Psychopharmacology 226, 475–490. doi: 10.1007/s00213-012-2919-2
Higgins, G. A., Silenieks, L. B., Patrick, A., De Lannoy, I. A. M., Fletcher, P. J., Parker, L. A., et al. (2017). Studies to examine potential tolerability differences between the 5-HT2C receptor selective agonists lorcaserin and CP-809101. ACS Chem. Neurosci. 8, 1074–1084. doi: 10.1021/acschemneuro.6b00444
Hutson, P. H., Balodis, I. M., and Potenza, M. N. (2018). Binge-eating disorder: clinical and therapeutic advances. Pharmacol. Ther. 182, 15–27. doi: 10.1016/j.pharmthera.2017.08.002
Kessler, R. C., Berglund, P. A., Chiu, W. T., Deitz, A. C., Hudson, J. I., Shahly, V., et al. (2013). The prevalence and correlates of binge eating disorder in the World Health Organization World Mental Health Surveys. Biol. Psychiatry 73, 904–914. doi: 10.1016/j.biopsych.2012.11.020
Kessler, R. M., Hutson, P. H., Herman, B. K., and Potenza, M. N. (2016). The neurobiological basis of binge-eating disorder. Neurosci. Biobehav. Rev. 63, 223–238. doi: 10.1016/j.neubiorev.2016.01.013
Lam, D. D., Przydzial, M. J., Ridley, S. H., Yeo, G. S. H., Rochford, J. J., O’Rahilly, S., et al. (2008). Serotonin 5-HT2C receptor agonist promotes hypophagia via downstream activation of melanocortin 4 receptors. Endocrinology 149, 1323–1328. doi: 10.1210/en.2007-1321
Lutter, M., and Nestler, E. J. (2009). Homeostatic and hedonic signals interact in the regulation of food intake. J. Nutr. 139, 629–632. doi: 10.3945/jn.108.097618
Martin, J. R., Bos, M., Jenck, F., Moreau, J. L., Mutel, V., Sleight, A. J., et al. (1998). 5-HT2C receptor agonists: pharmacological characteristics and therapeutic potential. J. Pharmacol. Exp. Ther. 286, 913–924.
McCue, D. L., Kasper, J. M., and Hommel, J. D. (2017). Regulation of motivation for food by neuromedin U in the paraventricular nucleus and the dorsal raphe nucleus. Int. J. Obes. 41, 120–128. doi: 10.1038/ijo.2016.178
Nigro, S. C., Luon, D., and Baker, W. L. (2013). Lorcaserin: a novel serotonin 2C agonist for the treatment of obesity. Curr. Med. Res. Opin. 29, 839–848. doi: 10.1185/03007995.2013.794776
O’Neil, P. M., Smith, S. R., Weissman, N. J., Fidler, M. C., Sanchez, M., Zhang, J., et al. (2012). Randomized placebo-controlled clinical trial of lorcaserin for weight loss in type 2 diabetes mellitus: the BLOOM-DM study. Obesity 20, 1426–1436. doi: 10.1038/oby.2012.66
Richardson, N. R., and Roberts, D. C. (1996). Progressive ratio schedules in drug self-administration studies in rats: a method to evaluate reinforcing efficacy. J. Neurosci. Methods 66, 1–11. doi: 10.1016/0165-0270(95)00153-0
Rowland, N. E., Crump, E. M., Nguyen, N., Robertson, K., Sun, Z., and Baooth, R. G. (2008). Effect of (-)-trans-PAT, a novel 5-HT2C receptor agonist, on intake of palatable food in mice. Pharmacol. Biochem. Behav. 91, 176–180. doi: 10.1016/j.pbb.2008.07.004
Schebendach, J., Broft, A., Foltin, R. W., and Walsh, B. T. (2013). Can the reinforcing value of food be measured in bulimia nervosa? Appetite 62, 70–75. doi: 10.1016/j.appet.2012.11.009
Swinford-Jackson, S. E., Anastasio, N. C., Fox, R. G., Stutz, S. J., and Cunningham, K. A. (2016). Incubation of cocaine cue reactivity associates with neuroadaptations in the cortical serotonin (5-HT) 5-HT2C receptor (5-HT2CR) system. Neuroscience 324, 50–61. doi: 10.1016/j.neuroscience.2016.02.052
Valencia-Torres, L., Olarte-Sanchez, C. M., Lyons, D. J., Georgescu, T., Greenwald-Yarnell, M., Myers, M. G. Jr., et al. (2017). Activation of ventral tegmental area 5-HT2C receptors reduces incentive motivation. Neuropsychopharmacology 42, 1511–1521. doi: 10.1038/npp.2016.264
Voigt, J. P., and Fink, H. (2015). Serotonin controlling feeding and satiety. Behav. Brain Res. 277, 14–31. doi: 10.1016/j.bbr.2014.08.065
Witt, A. A., and Lowe, M. R. (2014). Hedonic hunger and binge eating among women with eating disorders. Int. J. Eat. Disord. 47, 273–280. doi: 10.1002/eat.22171
Xu, P., He, Y., Cao, X., Valencia-Torres, L., Yan, X., Saito, K., et al. (2017). Activation of serotonin 2C receptors in dopamine neurons inhibits binge-like eating in mice. Biol. Psychiatry 81, 737–747. doi: 10.1016/j.biopsych.2016.06.005
Keywords: serotonin, 5-HT2C receptor, binge eating, high-fat food, motivation
Citation: Price AE, Anastasio NC, Stutz SJ, Hommel JD and Cunningham KA (2018) Serotonin 5-HT2C Receptor Activation Suppresses Binge Intake and the Reinforcing and Motivational Properties of High-Fat Food. Front. Pharmacol. 9:821. doi: 10.3389/fphar.2018.00821
Received: 25 May 2018; Accepted: 09 July 2018;
Published: 27 July 2018.
Edited by:
Stella Dracheva, Icahn School of Medicine at Mount Sinai, United StatesReviewed by:
Anthony Isles, Cardiff University, United KingdomClinton Canal, Mercer University, United States
Copyright © 2018 Price, Anastasio, Stutz, Hommel and Cunningham. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kathryn A. Cunningham, a2N1bm5pbmdAdXRtYi5lZHU=
†These authors have contributed equally to this work.