
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Pediatr., 14 May 2020
Sec. Pediatric Infectious Diseases
Volume 8 - 2020 | https://doi.org/10.3389/fped.2020.00244
 Massimiliano Bergallo1,2
Massimiliano Bergallo1,2 Luca Marozio3Giovanni Botta4Annalisa Tancredi3Valentina Daprà1,2Ilaria Galliano1,2Paola Montanari1,2Alessandra Coscia5Chiara Benedetto3
Luca Marozio3Giovanni Botta4Annalisa Tancredi3Valentina Daprà1,2Ilaria Galliano1,2Paola Montanari1,2Alessandra Coscia5Chiara Benedetto3 Pier Angelo Tovo2*
Pier Angelo Tovo2*Background: Placenta shows high transcription levels of human endogenous retroviruses (HERVs) that are overexpressed during embryonic and fetal development.
Methods: In order to gather further information on the degree of HERV activation in maternal and fetal tissues we assessed the transcription levels of pol genes of HERV-H, -K, and -W in PBMCs of newborns and their mothers as well as in chorion (fetal part) and decidua basalis (maternal part) of the placenta using a real time PCR assay.
Results: Transcripts of pol genes of the three HERV families were significantly higher in mononuclear cells from cord blood than from maternal blood and in the fetal part than in the maternal part of the placenta.
Conclusions: The HERV over-expressions in cells and tissues of the offspring are further clues that they play pivotal physiologic roles during early life events and suggest that HERV-driven abnormalities of pregnancy and fetal development may derive mostly from the conceptus, not from the mother.
Human endogenous retroviruses (HERVs) derive from ancestral infections of somatic cells with subsequent integration into germ line of primates more than 25 millions years ago. During evolution, mutations, recombinations, insertions and/or deletions have rendered their sequences unable to produce infectious virions. However, HERVs maintain their retroviral structure with gag, pol, and env genes, flanked by two non-coding long terminal repeats, that function as control regions (1). Most HERV sequences are inactive, but some encode proteins of viral origin. HERV proviruses can modulate the expression of adjacent cellular genes and their transcripts, through viral reverse transcriptase and integrase, may generate novel insertions into the DNA (2).
HERVs represent 8% of our genome, but their functions remain poorly defined. They have been associated with a large array of diseases, such as cancers, autoimmune diseases and neurologic disorders. On the other hand, their massive and prolonged persistence within our DNA suggests that they also have some beneficial roles. For instance, accumulating evidence highlights the importance of HERV elements in placentation (3–6), in the early phases of embryogenesis (7), in fetal development and in maintaining stemness of pluripotent cells (8).
The human placenta is composed of various cell types, that include extravillous and villous cytotrophoblasts. The latter can further be differentiated into an overlaying structure, referred to as the syncytiotrophoblast. This regulates the proper maternal-fetal exchange of nutrients and the production of important hormones, such as human chorionic gonadotropin and human placental lactogen whose production is regulated by retroviral elements (9). The placenta actively expresses a number of HERV genes. Among these, two envelope proteins, termed syncytin-1 (3) and syncytin-2 (4), are involved in normal placenta formation, drive the fusion between villous cytotrophoblast and the syncytiotrophoblast layer (5, 6), and may influence the feto-maternal tolerance (10). Genes similar to syncytins and with comparable functional properties are expressed in the placenta of various eutherian mammals. This suggests a convergent evolutionary process and represents a typical example how the host has co-opted retroviral elements to his own survival (11).
Several retroviral groups have been identified (2). Among these HERV-H, HERV-K, and HERV-W are those most widely studied. We previously observed that pol genes of these three retroviral groups are overexpressed in peripheral blood mononuclear cells (PBMCs) from cord blood, particularly in preterm newborns in inversed correlation with gestational age (12). The aims of the current project were to assess whether this enhanced HERV activation is present at birth also in the mother. Furthermore, since the activation of HERV-pol genes in the placenta has not been investigated and the localization of their transcripts in different placental cell populations is unkown, we evaluated the expression of pol genes of HERV-H, -K, and -W in both the maternal and fetal components of placenta.
PBMCs were collected after an uneventful, at term pregnancy and vaginal delivery from parturients and from cord blood of newborns without any clinical or laboratory abnormalities.
Placental specimens were obtained from uneventful, at term (>37 weeks of gestation) pregnancies after vaginal or Cesarean delivery. Placenta tissues were washed with Hank's solution to remove contaminating blood. The decidua basalis (i.e., the maternal part of the placenta) and the chorion (i.e., the fetal part of the placenta, represented mainly by syncytiotrophoblast) were macroscopically identified and separated by an expert pathologist (GB) and confirmed by a microscopic examination. Small samples (~5–10 mg wet weight) of each part of the placental tissue were placed in dry tubes and immediately processed for RNA extraction.
Total RNA was extracted from chorion, decidua basalis, and PBMCs using the automated extractor Maxwell (Promega, Madison, WI) following the RNA Blood Kit protocol without modification. This kit provides treatment with DNase during the RNA extraction process. Four hundred nanograms of total RNA was reverse-transcribed with 2 μl of buffer 10X, 4.8 μl of MgCl2 25 mM, 2 μl ImpromII (Promega), 1 μl of RNase inhibitor 20U/l, 0.4 μl random hexamers 250 μM (Promega), 2 μl mix dNTPs 100 mM (Promega), and dd-water in a final volume of 20 μl. The reaction mix was carried out in a GeneAmp PCR system 9,700 Thermal Cycle (Applied Biosystems, Foster City, CA, USA) under the following conditions: 5 min at 25°C, 60 min at 42°C and 15 min at 70°C for the inactivation of enzyme; the cDNAs were stored at −80° until use. For control of genomic DNA contamination we amplified directly RNA extracts without reverse transcription.
Relative quantification of mRNA expression of HERV-H, -K, and -W was achieved by means of PCR real time Taqman amplification and normalization to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) using the ABI PRISM 7,500 real time system (Life technologies, Texas, USA). GAPDH was chosen as reference gene being the most stable among 9 reference genes (13) and already used in our previous studies (12, 14, 15). Forty nanograms of cDNA were amplified in a 20 μl total volume reaction containing Go-Taq mastermix probe (Promega), 500 nmol of specific primers and 200 nmol of specific probes. The following primers and probes for pol genes were used: HERV-K primers (KPOLF-5′-CCACTGTAGAGCCTCCTAAACCC-3′) (KPOLR-5′-TTGGTAGCGGCCACTGATTT-3′) and probe (KPOLP-6FAM-CCCACACCGGTTTTTCTGTTTTCCAAGTTAA-TAMRA) as reported by Schaban et al. (16); for HERV-H primers (HPOLF-5′- TGGACTGTGCTGCCGCAA-3′) (HPOLR-5′-GAAGSTCATCAATATATTGAATAAGGTGAGA-3′) and probe (HPOLP-6FAM- TTCAGGGACAGCCCTCGTTACTTCAGCCAAGCTC-TAMRA; HERV-W primers (WPOLF-5′-ACMTGGAYKRTYTTRCCCCAA-3′) (WPOLR-5′- GTAAATCATCCACMTAYYGAAGGAYMA-3′) and probe (WPOLP-6FAM-TYAGGGATAGCCCYCATCTRTTTGGYCAGGCA-TAMRA); GAPDH primers (GAPDHF-5′-CCAAGGTCATCCATGACAAC-3′) (GAPDHR-5′- GTGGCAGTGATGGCATGGAC-3′) and probe (GAPDH-6FAM- TGGTATCGTGGAAGGA-3′ MGB). The established assays use probes and primers designed by Primer Express TM software version 3.0 (Applied Biosystems, Foster City, USA). Basic Local Alignment Search Tool (BLAST) analysis confirmed no cross-reaction between HERV primers.
The amplifications were run in a 96-well plate at 95°C for 10 min, followed by 40 cycles at 95°C for 15 s and at 60°C for 1 min. Furthermore, in order to confirm that there was no DNA genomic contamination, control PCR was performed with RNA before reverse transcription using the same primers and probes described above. Each sample was run in triplicate. Relative quantification of target gene expression was performed with the ΔCt method. Using 40 ng of cDNA in amplification we obtained Ct value from 26 to 30.4. These Ct values correspond to a good performance of real time PCR. Since we measured Ct for every target in all the samples tested, we argued that our methods were suitable for HERV detection and quantification. Results were expressed as 1/ΔCt.
Mann-Whitney test was used to compare the transcriptional levels of pol genes of every HERV group in chorion vs. decidua basalis as well as in PBMCs from cord blood vs. maternal blood. Statistical analyses were done using the Prism software (GraphPad Software, La Jolla, CA). In all analyses, p < 0.05 was taken to be statistically significant.
PBMCs were collected after vaginal delivery from 35 women and from cord blood of 47 newborns (13 males, 34 females). The gestational age was comparable between the maternal and the neonatal population (median gestational age 281 days, range 270–292 vs. 277 days, range 260–295, respectively). Placental tissue was collected from 57 women at delivery (24 vaginal deliveries, 33 Cesarean sections).
The pol genes of HERV-H, HERV-K, and HERV-W were always transcriptionally active in PBMCs from mothers and newborns as well as in the chorion and in the decidua basalis.
As detailed in Figure 1 the number of transcripts of pol genes of HERV-H, HERV-K, and HERV-W were significantly higher in chorion than in decidua basalis. In particular, the HERV-H-pol values (mean +/– SD) were: 0.26 +/– 0.14 in chorion vs. 0.24 +/– 0.04 in decidua basalis, p = 0.0167; the HERV-K-pol values were: 0.23 +/– 0.19 in chorion vs. 0.19 +/– 0.02 in decidua basalis, p = 0.0459; and the HERV-W-pol values were: 0.87 +/– 2.55 in chorion vs. 0.79 +/– 1.03 in decidua, p = 0.0419, with the largest range of values (including a few negative findings). The preferential HERV expression in the fetal part of placenta was not influenced by mode of delivery (data not shown).
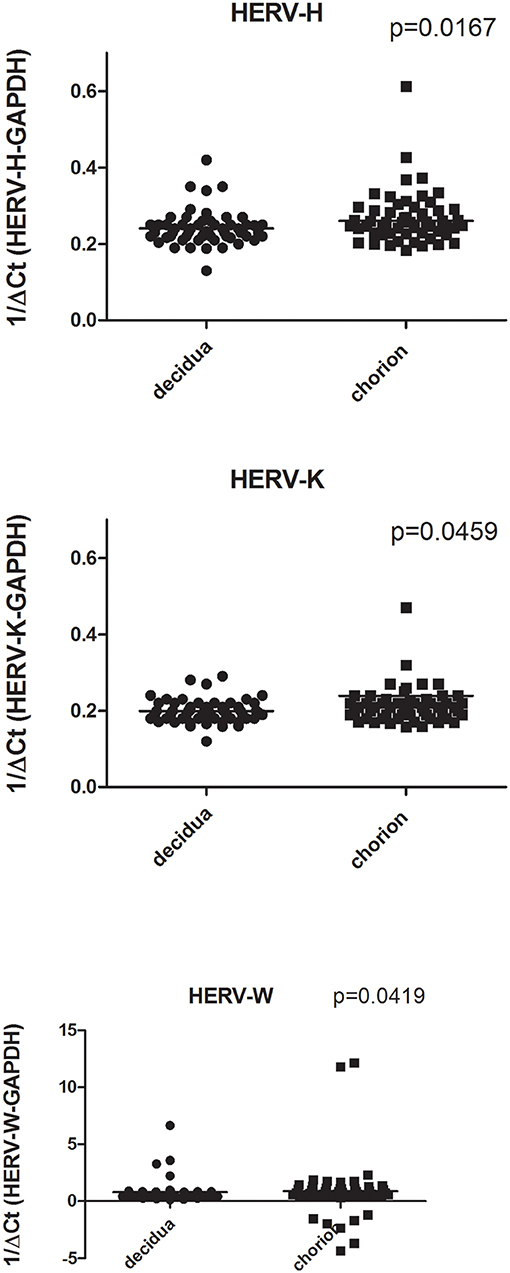
Figure 1. Transcription levels of pol genes of HERV-H, HERV-K, and HERV-W in decidua basalis and chorion from placentae of uneventful, at term pregnancies. Data are represented as black dot. Relative pol levels were assessed by real-time PCR and represented by 1/ΔCt. Statistical significance was calculated by Mann-Whitney test.
As detailed in Figure 2 the expression levels of pol genes of HERV-H, HERV-K, and HERV-W were significantly higher in PBMCs from newborns than from parturients. In particular, the HERV-H-pol values (mean +/– SD) were: 0.26 +/– 0.03 in newborns vs. 0.16 +/– 0.02 in parturients, p < 0.0001; the HERV-K-pol values were: 0.22 +/– 0.04 in newborns vs. 0.17 +/– 0.02 in parturients, p < 0.0001; and the HERV-W-pol values were: 0.66 +/– 0.2 in newborns vs. 0.28 +/– 0.04 in parturients, p < 0.0001. No significant differences were found in newborns between males and females (data not shown).
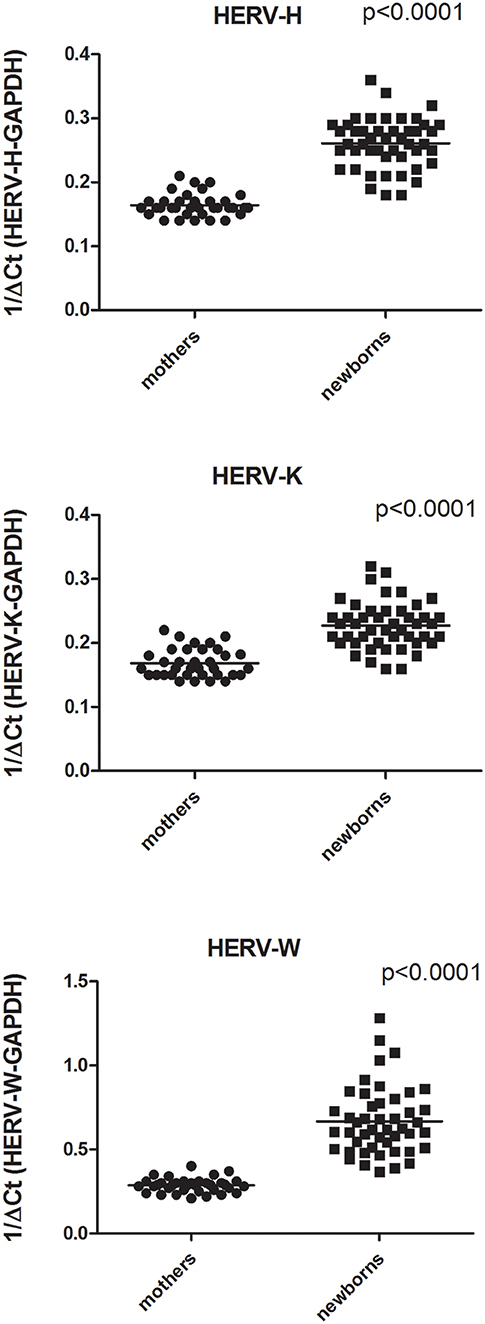
Figure 2. Transcription levels of pol genes of HERV-H, HERV-K, and HERV-W in mononuclear cells from peripheral blood of parturients and from cord blood of newborns collected after an at term pregnancy and vaginal delivery. Data are represented as black dot. Relative pol levels were assessed by real-time PCR and represented.
The results of this study document a significantly higher expression of pol genes of all the three HERV groups in the chorion (the fetal part of the placenta) than in the decidua basalis (maternal part). Our findings on HERV-pol genes are consistent with other studies, focused on HERV-env genes, demonstrating that their transcripts are mainly localized in villous cytotrophoblast and/or syncytiotrophoblast (5, 6, 17, 18). Notably, the expression of endogenous retroviruses occurs mostly in the fetal part of the placenta not only in invasive placental types with syncytiotrophoblast formation, such as the hemochorial placenta of human beings, but also in epitheliochorial placentas, such as in horses (19), where the syncytiotrophoblast is absent and fetal trophoblast cells are simply apposed to the intact maternal epithelium. This suggests that the preferential fetal expression of HERVs is an important feature for the placental structure and functions that has been preserved over the phylogenesis.
Present results for the first time also show that at birth the transcriptional levels of pol genes of HERV-H, HERV-K, and HERV-W are significantly higher in PBMCs from cord blood than from maternal blood. Therefore, our findings evidence that during pregnancy there is an enhanced HERV transcription that typically involves cells and tissues of fetal origin, whereas cells and tissues of maternal origin are less interested by the phenomenon. Several findings support this consideration. HERV-K is efficiently transcribed during human embryogenesis beginning at the 8-cell stage and continuing until the blastocyst outgrowths (3). Besides the aforementioned syncytins, recently a new tamed retroviral envelope protein produced from the fetus and then shed extracellularly into the maternal blood has been identified (8). We found that HERV-pol transcripts are significantly higher in PBMCs from premature than in full-term neonates in inversed correlation with gestational age, while postnatally they decrease with no further variations throughout the first decade of life (12). Also in rabbits the placenta-specific expression of endogenous retroviruses shows a 4-fold decrease with the progression of gestational age (20). It must be underlined that HERVs are efficiently transcribed in germ cells and pluripotent stem cells (2, 8) where they contribute to maintain their pluripotency circuit and cell fate reprogramming (21). Cord blood is reach of CD34+ hematopoietic and progenitor stem cells particularly in preterm newborns (22). The high proportion of stem cells in cord blood might thus explain, at least in part, why HERV transcription is higher in neonatal than in maternal PBMCs.
Taken together all these findings suggest that HERVs play crucial physiologic roles in the first life events, starting from the preimplanted blastocyst and continuing from placentation till birth through the placental tissues and the fetal development and differentiation. Indeed the primers and probes we used encompass several HERV-pol sequences, with the consequent limit that their localization in specific loci could not be identified. PBMCs from mothers and newborns and placental tissues were obtained from partly different populations. Furthermore, we did not assess their protein-coding capacity, though this does not undermine their potential importance, since also in placenta HERVs can act as non-coding regulatory elements able to promote or enhance the activity of neighboring cellular genes (10). In general, the emerging contribution of HERVs to a variety of ontogenetic processes may explain why a substantial portion of the genome is occupied, from millions of years, by retroviruses.
The underlying mechanisms responsible for the increased HERV transcription in the placenta and during the intrauterine life remain to be fully elucidated. HERV expression is regulated by epigenetic mechanisms, such as the methylation processes of DNA (23). The placental tissues are characterized by DNA hypometylation that triggers HERV transcription (24) and decreases with the progression of pregnancy (25). In contrast, pathologic placentae are characterized by epigenetic hypermethylation of HERV promoters (26). Notably, both the DNA hypomethylation and the HERV overexpression are inversely correlated with gestational age (12, 27). Furthermore, sexual hormones contribute to the regulation of HERVs (28), therefore, the peculiar hormonal patterns occurring throughout pregnancy might represent typical temporary factors conditioning HERV activation.
The present high placental and fetal HERV transcripts were detected in physiological conditions. Alterations of HERV activity might however result in failures or abnormalities of pregnancy or in disturbaces of offspring development. For instance, pre-eclampsia (PE) is a typical pregnancy disorder associated with a defect in placentation that causes important maternal and neonatal morbidity and mortality. Syncytin-1 and −2 transcripts are reduced in placental tissues of women with PE (29, 30). Dysregulations of HERV gene levels were found in placentae of patients with intrauterine growth restriction (31). Furthermore, maternal smoking, typically associated with impaired fetal development, has a significant negative impact on the physiologic overexpression of HERVs in newborns (15).
In general, the preferential HERV expression in cells and tissues of the offspring rather than of the mother suggests that, in a number of instances, abnormalities of pregnancy and of fetal development may result from HERV-driven alterations deriving mostly from the conceptus, even if targeted by maternal factors, such as environmental and hormonal factors. For instance, we looked at possible abnormal HERV activation in blood samples from women with recurrent miscarriages, but we did not find any alterations (unpublished data). In contrast, enhanced HERV expression is associated with development and invasion of several cancers, e.g., the up-regulation of syncytin-1 promotes the malignant transformation of hydatidiform moles (32) and the spread of endometrial carcinoma (33), and all malignancies of placental origin (including those of late appearance) derive from cells of the conceptus, not of the mother.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation, to any qualified researcher.
The subjects included were asked to sign an informed consent. The study was approved by the local Ethical Committee (Comitato Etico Interaziendale AOU Città della Salute e della Scienza di Torino—AO Ordine Mauriziano di Torino—ASL Città di Torino. Protocol no. 0067257–2018). This study was carried out in accordance with the recommendations of the Declaration of Helsinki, and all parents gave informed written consent. The protocol was approved.
MB, LM, and PT contributed in conceptualizing the study design. MB and PT prepared the first draft of the manuscript. LM, AT, CB, and AC enrolled the study populations, obtained their informed consent, were responsible for ethical approval, and collected placentae and PBMCs. GB separated placental tissues. MB, VD, IG, and PM were involved in processing samples, data collection, and data analysis. All authors approved the final manuscript.
This study was supported by a grant RILO 60% University of Turin.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
1. Johnson WE. Origins and evolutionary consequences of ancient endogenous retroviruses. Nat Rev Microbiol. (2019) 17:355–70. doi: 10.1038/s41579-019-0189-2
2. Grandi N, Tramontano E. Human endogenous retroviruses are ancient acquired elements still shaping innate immune responses. Front Immunol. (2018) 9:2039. doi: 10.3389/fimmu.2018.02039
3. Mi S, Lee X, Li X, Veldman GM, Finnerty H, Racie L, et al. Syncytin is a captive retroviral envelope protein involved in human placental morphogenesis. Nature. (2000) 403:785–9. doi: 10.1038/35001608
4. Blaise LS, de Parseval N, Benit L, Heidmann T. Genomewide screening for fusogenic human endogenous retrovirus envelopes identifies syncytin 2, a gene conserved on primate evolution. Proc Natl Acad Sci USA. (2003) 100:13013–18. doi: 10.1073/pnas.2132646100
5. Malassine A, Blaise S, Handschuh K, Lalucque H, Dupressoir A, Evain-Brion D, et al. Expression of the fusogenic herv-frd env glycoprotein (syncytin 2) in human placenta is restricted to villous cytotrophoblastic cells. Placenta. (2007) 28:185–91. doi: 10.1016/j.placenta.2006.03.001
6. Vargas A, Moreau J, Landry S, LeBellego F, Toufaily C, Rassart E, et al. Syncytin-2 plays an important role in the fusion of human trophoblast cells. J Mol Biol. (2009) 392:301–18. doi: 10.1016/j.jmb.2009.07.025
7. Grow EJ, Flynn RA, Chavez SL, Bayless NL, Wossidlo M, Wesche DJ, et al. Intrinsic retroviral reactivation in human preimplantation embryos and pluripotent cells. Nature. (2015) 52:221–5. doi: 10.1038/nature14308
8. Heidmann O, Béguin A, Paternina J, Berthier R, Deloger M, Bawa O, et al. HEMO, an ancestral endogenous retroviral envelope protein shed in the blood of pregnant women and expressed in pluripotent stem cells and tumors. Proc Natl Acad Sci USA. (2017) 114:E6642–51. doi: 10.1073/pnas.1702204114
9. Dunn-Fletcher CE, Muglia LM, Pavlicev M, Wolf G, Sun MA, Hu YC, et al. Anthropoid primate–specific retroviral element THE1B controls expression of CRH in placenta and alters gestation length. PLoS Biol. (2108) 16:e2006337. doi: 10.1371/journal.pbio.2006337
10. Lokossou AG, Toudic C, Barbeau B. Implication of human endogenous retrovirus envelope proteins in placental functions. Viruses. (2014) 6:4609–27. doi: 10.3390/v6114609
11. Chuong EB. The placenta goes viral: retroviruses control gene expression in pregnancy. PLoS Biol. (2018) 16:e3000028. doi: 10.1371/journal.pbio.3000028
12. Bergallo M, Galliano I, Pirra A, Daprà V, Licciardi F, Montanari P, et al. Transcriptional activity of human endogenous retroviruses is higher at birth in inversed correlation with gestational age. Infect Genet Evol. (2019) 68:273–9. doi: 10.1016/j.meegid.2018.12.018
13. Garson JA, Usher L, Al-Chalabi A, Huggett J, Day EF, McCormick AL. Quantitative analysis of human endogenous retrovirus-K transcripts in postmortem premotor cortex fails to confirm elevated expression of HERV-K RNA in amyotrophic lateral sclerosis. Acta Neuropathol Commun. (2019) 7:45. doi: 10.1186/s40478-019-0698-2
14. Bergallo M, Montanari P, Mareschi K, Merlino C, Berger M, Bini I, et al. Expression of the pol gene of human endogenous retroviruses HERV-K and -W in leukemia patients. Arch Virol. (2017) 162:3639–44. doi: 10.1007/s00705-017-3526-7
15. Bergallo M, Galliano I, Daprà V, Pirra A, Montanari P, Pavan M, et al. Transcriptional activity of human endogenous retroviruses in response to prenatal exposure of maternal cigarette smoking. Am J Perinatol. (2019) 36:1060–5. doi: 10.1055/s-0038-1675768
16. Schanab O, Humer J, Gleiss A, Mikula M, Sturlan S, Grunt S, et al. Expression of human endogenous retrovirus K is stimulated by ultraviolet radiation in melanoma. Pigment Cell Melanoma Res. (2011) 24:656–65. doi: 10.1111/j.1755-148X.2011.00860.x
17. Kudaka W, Oda T, Jinno Y, Yoshimi N, Aoki Y. Cellular localization of placenta-specific human endogenous retrovirus (HERV) transcripts and their possible implication in pregnancy-induced hypertension. Placenta. (2008) 29:282–9. doi: 10.1016/j.placenta.2007.11.009
18. Kammerer U, Germeyer A, Stengel S, Kapp M, Denner J. Human endogenous retrovirus K (HERV-K) is expressed in villous and extravillous cytotrophoblast cells of the human placenta. J Reprod Immunol. (2011) 91:1–8. doi: 10.1016/j.jri.2011.06.102
19. Stefanetti V, Marenzoni ML, Passamonti M, Cappelli K, Garcia-Etxebarria K, Coletti MS. High expression of endogenous retroviral envelope gene in the equine fetal part of the placenta. PLoS ONE. (2016) 11:e0155603. doi: 10.1371/journal.pone.0155603
20. Heidmann O, Vernochet C, Dupressoir A, Heidmann T. Identification of an endogenous retroviral envelope gene with fusogenic activity and placenta-specific expression in the rabbit: a new “syncytin” in a third order of mammals. Retrovirology. (2009) 6:107. doi: 10.1186/1742-4690-6-107
21. Yin Y, Zhou L, Yuan S. Enigma of retrotransposon biology in mammalian early embryos and embryonic stem cells. Stem Cells Int. (2018) 2018:6239245. doi: 10.1155/2018/6239245
22. Wisgrill L, Schuller S, Bammer M, Berger A, Pollak A, Radke TF, et al. Hematopoietic stem cells in neonates: Any differences between very preterm and term neonates? PLoS ONE. (2014) 9:e106717. doi: 10.1371/journal.pone.0106717
23. Groh S, Schotta G. Silencing of endogenous retroviruses by heterochromatin. Cell Mol Life Sci. (2017) 74:2055–65. doi: 10.1007/s00018-017-2454-8
24. Schroeder DI, Jayashankar K, Douglas KC, Thirkill TL, York D, Dickinson PJ, et al. Early developmental and evolutionary origins of gene body DNA methylation patterns in mammalian placentas. PLoS Genet. (2015) 11:e1005442. doi: 10.1371/journal.pgen.1005442
25. Novakovic B, Yuen RK, Gordon L, Penaherrera MS, Sharkey A, Moffett A, et al. Evidence for widespread changes in promoter methylation profile in human placenta in response to increasing gestational age and environmental/stochastic factors. BMC Genomics. (2011) 12:529. doi: 10.1186/1471-2164-12-529
26. Ruebner M, Strissel PL, Ekici AB, Stiegler E, Dammer U, Goecke TW, et al. Reduced syncytin-1 expression levels in placental syndromes correlates with epigenetic hypermethylation of the ERVW-1 promoter region. PLoS ONE. (2013) 8:e56145. doi: 10.1371/journal.pone.0056145
27. Knight AK, Craij JM, Theda C, Bækvad-Hansen M, Bybjerg-Grauholm J, Hansen CS, et al. An epigenetic clock for gestational age at birth based on blood methylation data. Genome Biol. (2016) 17:206. doi: 10.1186/s13059-016-1068-z
28. Nguyen TD, Davis J, Eugenio RA, Liu Y. Female sex hormones activate human endogenous retrovirus type K through the OCT4 transcription factor in T47D breast cancer cells. AIDS Res Hum Retroviruses. (2019) 35:348–56. doi: 10.1089/aid.2018.0173
29. Vargas A, Toufaily C, LeBellego F, Rassart E, Lafond J, Barbeau B. Reduced expression of both syncytin 1 and syncytin 2 correlates with severity of preeclampsia. Reprod Sci. (2011) 18:1085–91. doi: 10.1177/1933719111404608
30. Zhuang XW, Li J, Brost BC, Xia XY, Chen HB, Wang CX, et al. Decreased expression and altered methylation of syncytin-1 gene in human placentas associated with preeclampsia. Curr Pharm Des. (2014) 20:1796–802. doi: 10.2174/13816128113199990541
31. Ruebner M, Strissel PL, Langbein M, Pahbusch F, Watcher DL, Faschingbauer F, et al. Impaired cell fusion and differentiation in the placentae from patients with intrauterine growth restriction correlate with reduced levels of HERV envelope genes. J Mol Med. (2010) 88:1143–56. doi: 10.1007/s00109-010-0656-8
32. Bolze PA, Patrier S, Cheynet V, Oriol G, Massardier J, Hajri T, et al. Expression patterns of ERVWE1/Syncytin-1 and other placentally expressed human endogenous retroviruses along the malignant transformation process of hydatidiform moles. Placenta. (2016) 39:116–24. doi: 10.1016/j.placenta.2016.01.011
Keywords: HERV human endogenous retroviruses, PCR, placenta, cord blood, mRNA expression
Citation: Bergallo M, Marozio L, Botta G, Tancredi A, Daprà V, Galliano I, Montanari P, Coscia A, Benedetto C and Tovo PA (2020) Human Endogenous Retroviruses Are Preferentially Expressed in Mononuclear Cells From Cord Blood Than From Maternal Blood and in the Fetal Part of Placenta. Front. Pediatr. 8:244. doi: 10.3389/fped.2020.00244
Received: 23 October 2019; Accepted: 20 April 2020;
Published: 14 May 2020.
Edited by:
Enzo Tramontano, University of Cagliari, ItalyReviewed by:
Hervé Perron, Geneuro-Innovation, FranceCopyright © 2020 Bergallo, Marozio, Botta, Tancredi, Daprà, Galliano, Montanari, Coscia, Benedetto and Tovo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Pier Angelo Tovo, cGllcmFuZ2Vsby50b3ZvQHVuaXRvLml0
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.