
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Pediatr. , 07 February 2020
Sec. Children and Health
Volume 8 - 2020 | https://doi.org/10.3389/fped.2020.00030
 Morika D. Williams1,2,3
Morika D. Williams1,2,3 B. Duncan X. Lascelles1,2,3,4,5*
B. Duncan X. Lascelles1,2,3,4,5*Modern health care has brought our society innumerable benefits but has also introduced the experience of pain very early in life. For example, it is now routine care for newborns to receive various injections or have blood drawn within 24 h of life. For infants who are sick or premature, the pain experiences inherent in the required medical care are frequent and often severe, with neonates requiring intensive care admission encountering approximately fourteen painful procedures daily in the hospital. Given that much of the world has seen a steady increase in preterm births for the last several decades, an ever-growing number of babies experience multiple painful events before even leaving the hospital. These noxious events occur during a critical period of neurodevelopment when the nervous system is very vulnerable due to immaturity and neuroplasticity. Here, we provide a narrative review of the literature pertaining to the idea that early life pain has significant long-term effects on neurosensory, cognition, behavior, pain processing, and health outcomes that persist into childhood and even adulthood. We refer to clinical and pre-clinical studies investigating how early life pain impacts acute pain later in life, focusing on animal model correlates that have been used to better understand this relationship. Current knowledge around the proposed underlying mechanisms responsible for the long-lasting consequences of neonatal pain, its neurobiological and behavioral effects, and its influence on later pain states are discussed. We conclude by highlighting that another important consequence of early life pain may be the impact it has on later chronic pain states—an area of research that has received little attention.
Newborn babies inevitably experience pain as a part of routine neonatal care, such as vitamin K injections and heel sticks to obtain blood for screening tests. These painful experiences are more frequent in infants who are sick or premature. In recent decades, much of the world has experienced a continued increase in preterm births with an estimated 15 million babies born prematurely each year globally (1). Preterm birth is defined as a baby born before 37 weeks of gestation and is further sub-categorized based on gestation age: extremely preterm (<28 weeks), very preterm (28– <32 weeks), and moderate to late preterm (32– <37 weeks) (2). A 2010 study of 184 countries found that the preterm birth rate, expressed as a percentage of livebirths, ranged between 5 and 18% and that this rate did not correlate directly to the economic development level of the country, e.g., northern European countries have a 5% rate whereas the United States is at 13% (3, 4). In the Unites States alone, the medical care, loss of productivity, maternal delivery, early intervention, and special education services are estimated to have cost $26.2 billion in 2005 (5). Furthermore, prematurity is the leading cause of death in children under the age of 5 years (1). Together, these factors make prematurity a modern global health crisis.
Though preterm births continue to increase, medical advancements in neonatal medicine have led to improved survival rates for preterm babies. This increase is associated with intense medical and surgical management over an extended hospital stay. Babies born at term (39–41 weeks) spend 4.9 days in the neonatal intensive care unit (NICU) (6). However, when adjusted for risk (i.e., birth weight, sex, small for gestational age status, ethnicity, fetal distress, and maternal stress), the average length of stay in the NICU for extremely preterm infants is 81 days (7). During this period, newborns may undergo multiple painful procedures each day for care and treatment. The demographics of the NICU is predominately premature newborns (24–36 weeks) when compared to term newborns (37–42 weeks)-−72.3 and 27.7% of NICU babies, respectively (8). Therefore, although preterm babies are at a higher risk for neonatal pain exposure, term newborns are not exempt. In fact, in the United States, ~460,000 neonates require NICU admission each year, and thus experience procedural, medical, or surgical pain (9).
The neonatal nervous system is very plastic because it is immature and undergoing major developmental changes; therefore, neonates may be vulnerable to neurodevelopmental changes from painful stimuli (10). Nociceptive input during early neurodevelopment is typically deleterious, increasing the risk of neurodevelopmental impairments, including short- and long-term physical and psychological disability (4, 11, 12), adverse changes in brain development and processing (13–15), and alterations in somatosensory function that lead to pain (16, 17). Such damage may have significant long-term effects that persist into childhood and even adulthood; a notion increasingly supported by the literature (17, 18).
The purpose of this report is to describe the current understanding of the relationship between early life pain and its implications on pain later in life, via a narrative review of clinical and pre-clinical studies. To this end, we highlight common painful procedures that occur early in life and the important consequences of these pain events to humans later in life. In each section we discuss the preclinical work that has advanced our understanding of these consequences and the potential mechanisms underlying them. We then review the animal model correlates used to investigate the effects of early life injury and discuss future directions for research.
The literature review was performed by searching the following databases, including PubMed, CAB Abstracts, and Google Scholar. Search was conducted using the following keywords: early life injury, early life pain, neonatal injury, neonatal pain, repetitive needle prick, heelstick, consequences, impairment, behavior, cognition, pain response, central nervous system, mechanisms, pain models, animal models, chronic pain, osteoarthritis, and inflammatory pain. For articles generated from this search, the abstracts were read, and then those relevant were downloaded and read in full. Those fitting in topic of relevance were human studies or rodent models of neonatal pain. Rat studies and behavioral studies were emphasized for the review. Studies investigating the effects of stress and other environmental factors were excluded from the review.
Newborns routinely experience invasive, painful procedures, such as blood collection, immunization, vitamin K injections, and/or circumcision. Preterm or sick term newborns in the NICU experience a much higher frequency of invasive and painful procedures (8, 18). On average, premature babies requiring NICU admission encounter about 14 painful procedures daily in the hospital (19, 20). These invasive procedures range from repetitive heel sticks to minimally invasive or open surgery (Table 1) (21–24).
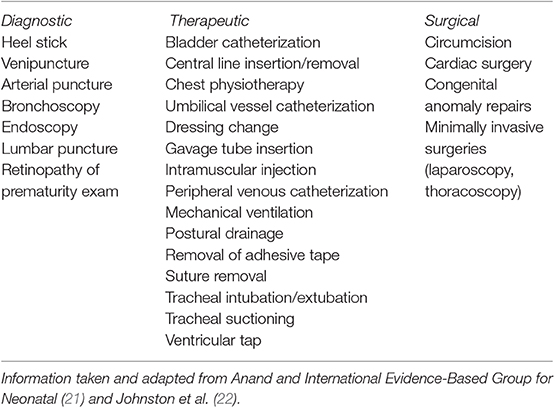
Table 1. Painful procedures commonly performed in the NICU.
One of the most frequently performed invasive procedures in the NICU is a heel stick, or heel lance/needle prick (8, 25). This procedure is a pinprick puncture into the heel of a newborn used to obtain blood samples for screening laboratory tests, glucose levels, general chemistries, complete blood counts, and toxicology screening. Generally, to obtain adequate amounts of blood for testing, the heel must be squeezed, which is more painful for newborns than venipunctures (26). Despite this fact and the recommendation to use less invasive alternatives (e.g., venipuncture, mechanical lances) or to incorporate non-pharmacological analgesia to minimize the pain (9), heel sticks continue to be common in practice.
Another painful procedure that may be required for newborns with significant respiratory compromise is endotracheal intubation, a procedure performed on both adults and infants that allows access to the airway to provide respiratory support via mechanical ventilation. Adult patients report mechanical ventilation to be a source of pain, discomfort, and depression (27). Because the procedure is known to be associated with pain (28, 29), the International Evidence-Based Group for Neonatal Pain recommends this procedure be performed ‘without the use of analgesia or sedation … only for resuscitation in the delivery room or for life-threatening situations associated with the unavailability of intravenous access’ (22). However, clinical use of pain control is still suboptimal (30) despite evidence that opioids reduce signs of pain during mechanical ventilation in newborns (31). In addition, there is concern around the long-term negative effects of opioids, such as neuromotor disability (32, 33). In the United States alone, mechanical ventilation is provided to ~35,000 preterm and 20,000 term neonates each year (34). Therefore, there is great concern that mechanical ventilation is a significant contributing factor to the early life pain experience.
To manage and correct life-threatening diseases or abnormalities, open and/or minimally invasive surgeries may be required. Although the exact number of surgeries performed on newborns is unknown, up to 33% of extremely preterm babies require surgery to treat congenital anomalies or manage complications (17, 35). Multiple surgeries may be needed, thus increasing the exposure to other painful procedures, such as venipunctures, endotracheal intubations, and anesthetic episodes—as well as the surgical trauma-associated and post-operative pain itself. As the procedural numbers rise, the likelihood of developing hypersensitivity and/or persistent pain at the site of damage (chronic post-surgical pain) increases within months following surgery (18, 36, 37). Chronic post-surgical pain is thought to be in large part due to a phenomenon called central sensitization, a manifestation of maladaptive responses of the spinal cord that lead to increases in excitability and reductions in inhibition resulting in enhanced pain responsiveness (38, 39).
Despite the high incidence of painful procedures performed in the NICU, most neonates receive no pharmacological analgesia (40–42) or receive an inadequate level of analgesic (43, 44). A combination of factors, such as the underestimation of pain or concerns for potential adverse effects, lead practitioners to withhold or inadequately dose these medications. However, recent studies exploring the long-term consequences of untreated, early life pain have shown that early life pain can have negative repercussions that can impact sensorimotor and cognitive development, behavior/mood, pain responses, pharmacological requirements, and health status seen as early as adolescence. We discuss these consequences below.
It is believed that early pain experiences shape the somatosensory scaffolding of later perceptual, cognitive, and social development (45). This belief is evidenced by a recent study that found an association between painful procedures in early life and a reduction in brain responses to non-noxious touch stimuli (45). Such risks of neurosensory impairment appear to be aggravated by prematurity. Sensorimotor impairments in children, such as impaired vision/blindness, impaired hearing/deafness, cerebral palsy, delayed development trajectory, and impaired intellect into childhood and adulthood have been attributed to painful medical procedures early in life (46). For example, neonatal surgery has led to major neurosensory disability with effects that persistent into childhood (8 years old), with males having a higher risk of disability (47).
Early exposure to painful procedures can negatively impact neurodevelopment, such as brain growth, which is directly related to cognition. For example, an inverse correlation between frequency of invasive procedures and amygdala and thalamus volumes was seen in 8 years old children born very preterm (48). Further, reduced brain volumes in children who survived very preterm birth and experienced early life pain were associated with poor cognitive outcomes, such as lower intelligence (IQ), language and attention deficits, poor visual-motor functions, and poor behavioral outcomes (48–51). These cognitive deficits have been shown to persist into early adolescence and young adulthood (19 years old) for those who are born premature (52–54). Overall, preterm boys appear to be at a greater risk for poor neurodevelopmental trajectories than their female counterparts (53, 55). In contrast, females with early pain seemed to be more vulnerable to brain development with slower growth of thalamic, basal ganglia, and total brain volumes (56). However, conflicting sex-differences may be attributed to varied study designs, outcome measures, environment, observer variation, or individual variation.
Rodent studies have found similar impairments to cognition and brain development caused by early life pain. In a rat model of neonatal pain, mimicking NICU care, long-term alterations in brain development were seen (57, 58). Rats exposed to acute repetitive needle prick stimulation (4 times per day for the first 2 weeks of life) suffered long-term memory impairment (59), while chronic repetitive needle prick stimulation (4 times per day, every other day, for the first 8 weeks of life) resulted in short term memory deficits (60). Similarly, neonatal rat inflammatory pain produced long-lasting alterations in hippocampal function that were more pronounced in middle-aged adult (P424–442) vs. adult (P144–158) rats, characterized by spatial memory impairment (61). Ranger et al. report that memory in adulthood was poorer for mice exposed to repetitive pain during the first week of life (62).
There is a strong association between exposure to painful procedures and altered behavioral development trajectories (63). Survivors of early repetitive pain may develop attention-deficit disorders, atypical behaviors, such as hypervigilance and exaggerated startled responses, and other forms of long-term stress-related psychosocial disabilities (64). Neonatal rodent models support these findings by showing that repetitive acute pain during the first week of life can lead to negatively altered locomotor activity (65), defensed withdrawal behaviors (57), anxiety/depressive behaviors (66), and reduced social behavior (67).
Internalizing behaviors are negative actions that are directed toward oneself manifesting as withdrawal, depression, and/or anxiety (68). Higher levels of internalizing behaviors are predictive of atypical social competence in children leading to greater social difficulties and lower peer acceptance (68, 69). These negative behaviors have been seen in children as early as 18 months old (70) and may persist throughout childhood and adulthood (54, 71, 72). Preterm children exposed to neonatal pain have a higher rate of internalizing behaviors than full term children (70, 73). For example, young adults born extremely preterm who underwent prior surgery had higher anxiety and pain catastrophizing than term controls (74). Internalized behaviors can also lead to the development of other serious health issues, such as drug addiction (75), alcoholism (76), or obesity (77).
Early life pain can be challenging to recognize due to variability in patients' responses toward pain and changes associated with age and subsequent painful experiences. Changes in facial expression and withdrawal reflex in the neonate are suggested to be most associated with nociception activity (78). However, facial grimacing after a heel stick was seen as early as 28 gestational weeks of age, becoming more recognizable with age. Recently, Green et al. demonstrate the emergence of discriminative facial expressions toward noxious and innocuous stimulation occurring in infants at ~33 weeks' gestation, coinciding with brain maturation (79). Some work has shown that the degree of grimacing was inversely related the number of invasive procedures (43). Additionally, a greater number of procedures during early life has been associated with lower behavioral pain responses (e.g., facial grimace, cry, state of arousal) and pain scores following subsequent pain exposure within 1 month of life (80–82). Although pain experiences may not produce noticeable behavioral changes, noxious stimulation (i.e., heel stick) in infants between 25 and 43 gestational weeks may be processed at the somatosensory cortical level (83). The presences of cortical activation, measured by increases in hemodynamic responses, without a facial motor response support the concept that pain is reflective of emotional responses requiring cortical processing (83, 84). Sex should also be considered when evaluating facial grimacing, as female preterm neonates showed more robust facial expressions than males in response to acute procedural pain (85).
Painful procedures during neonatal development have been shown to have later influences on sensitivity to noxious stimuli and pain. Negative hemodynamic effects, such as higher heart rates and lower oxygen saturations, following secondary noxious stimuli are reported in individuals with a history of neonatal pain (43, 80, 86). Males with a history of neonatal pain have more pronounced responses than females with respect to hemoglobin oxygenation following secondary venipuncture (87).
Procedural pain early in life appears to lead to alterations in neonatal sensory function resulting in hyposensitivity following acute pain as a neonate but hypersensitivity later in life (88). However, the directionality of changes in sensitivity later in life is inconsistent. Preterm or full term children between 10 and 12 years old who underwent neonatal surgery have shown generalized thermal hyposensitivity as well as mechanical and thermal hyposensitivity in areas around prior tissue damage (89, 90). In contrast, in preterm adolescents between 12 and 18 years old there was greater mechanical hypersensitivity compared to full term children, with females being consistently more sensitive than male counterparts (91). And, young adults (18–20 years old) who were extremely preterm and underwent prior neonatal surgery had higher pain intensity scores and reported moderate to severe persistent pain more than young adults who were term born (17, 74).
Investigations into whether painful early life experiences have a negative impact on pain responses following secondary acute injury later in life have also been conducted. Hypersensitivity and allodynia after secondary injuries have been demonstrated (88). Heightened pain responses (e.g., cried longer, higher pain scores) toward routine vaccinations given to 4–6 months old neonatally circumcised babies were seen when compared to uncircumcised babies (92, 93). Infants (4–21 weeks of age) requiring repeated surgery in areas of scaring, due to prior neonatal surgery, had higher post-operative pain scores and higher analgesic requirements than controls (94). Furthermore, higher pain intensity ratings during venipuncture at age 7.5 years were positively associated with a greater number of invasive procedures experienced in the neonatal period (95). Variations in pain responses (e.g., becoming hypersensitive or hyposensitive) with or without secondary injury are likely due to difficulties with controlling for: intensity/frequency of pain stimulus, time/age following painful event(s), and variability within study population, outcome measures, and other environmental influences.
Animal models of pain have aided in further exploration of these effects of early life pain as well as provided additional support for these clinical findings. After repetitive needle prick stimuli, neonatal rats had decreased thermal sensitivity during development (57) and adulthood (60). Similarly, adolescent rats demonstrated peripheral thermal hypoalgesia after experiencing repeated post-natal inflammatory peripheral pain (formalin) (96). In contrast, repeated neonatal hindpaw incision or needle prick has been shown to sensitize adult rats to secondary incisional injury resulting in increased incision-related hyperalgesia to cutaneous dynamic tactile (brush), pinch, and/or punctate (von Frey hair) stimuli (16, 97–100). For additional results, Walker et al. provides a list of long-term effects of neonatal injury demonstrated in laboratory models (101).
Early-life, pain-induced alterations can increase the likelihood of having subsequent poor health outcomes. Preterm babies, who are exposed to painful stimuli, are at a greater risk of developing non-communicable diseases (diabetes, hypertension) and other health conditions later in life (102), creating an intergenerational cycle of risk. Controlling for confounding factors, post-natal growth is reduced in infants that undergo repetitive procedural pain during early life (103), and infants who are smaller tend to have more cognitive problems than other children as previously discussed. Small infants often rapidly catch-up in growth during the first 2 years of life, but this ‘catch up’ is associated with the development of adiposity and impaired insulin sensitivity later in life (104). The development of these outcomes may lead to increased risk of obesity and metabolic disorders, such as type II diabetes in humans (105, 106) and in animals (107). Blood pressure in adults born premature may be higher than adults born full term, and thus prematurity exposes one to a higher risk of hypertension, cardiovascular disease, and stroke (108). People with these health conditions are predisposed to becoming chronic pain sufferers. There is a growing body of literature regarding the long-term health effects of early life injury, and a detailed, more exhaustive list can be found elsewhere (15, 109). Additionally, more clinical and research studies have been conducted to investigate the persistent ramifications of early life pain beyond infancy (67, 110–112).
Pain is a complex, multidimensional, and multisensory phenomenon involving many intact systems to produce the final emotional and sensory response. For the perception of pain, a noxious stimulus must be processed through the various levels of the neuroaxis, and arrive at higher brain centers. Nociception, or pain responses, can be inhibited at the level of the spinal cord through the descending inhibitory pain pathway. The descending pain circuit is composed of the periaqueductal gray matter in the upper brain stem, the locus coeruleus, the nucleus raphe magnus, and the nucleus reticularis gigantocellularis. Under normal conditions, the descending pain modulatory system controls the balance between facilitation and inhibition of nociception, maintaining a baseline of sensory processing (113). However, disruption in the system can lead to enhanced or facilitated nociception or promotion and maintenance of chronic pain (114).
The pain transmission system is further complicated by interactions between the central and peripheral nervous systems and the immune system—neuroimmune interactions. Neuroimmune interactions are responsible for: recruiting local neuronal elements for fine tuning the immune response; participating in the plasticity of synapses during development as well as in synaptic plasticity at adulthood; and engaging the rest of the body in the fight against infection from pathogenic microorganisms (115). This established bidirectional interaction between the neuronal and immune systems plays a crucial role in pain modulation (116–118).
During neonatal development newborns have a high degree of neuroplasticity and are very vulnerable to the modulating effects of noxious stimuli (88, 119–121), especially if repetitive in nature. Repetitive painful stimuli cause structural and functional reorganization of the nervous system at the level of the: periphery, spinal cord, and supraspinal pain processing; neuroendocrine function; and neurologic development (64, 82, 122). Human and animal studies have provided solid evidence that repetitive and prolonged pain early in life can alter subsequent pain processing (43, 123) and later pain sensitivity (59, 124). It remains to be determined if such changes also contribute to chronic pain. Specific underlying structural and functional reorganization that may be responsible for enhanced pain responses to subsequent pain as a result of early life pain are discussed here, with an emphasis on information gained from preclinical work in animal models.
It was once believed that neonates were unable to perceive pain due to the immaturity of the sensory nervous system (125). However, neuroanatomical and behavioral studies have provided strong evidence to the contrary (126–128). During the neonatal period, a considerable amount of maturation associated with pain transmission and modulation is taking place. Although the peripheral nervous system is mature and functional by 24 gestational weeks, there are substantial neuroanatomical shifts in the distribution of unmyelinated (C) and myelinated (A) ascending fibers occurring in the post-natal period (41). Early in life there is an abundance of myelinated Aδ sensory fibers, responsible for the initial perception of pain, and fewer low density C fibers, responsible for pain intensity (129). Pre-clinical non-human animal studies have further demonstrated the relative numbers of fiber types can be affected by early life pain, leading to hyperinnervation and subsequent peripheral sensitization and increased pain sensitivity later in life (130). Babies are more likely to have hypersensitivity due to imbalances between the numbers of afferent fibers and descending (negative) influences (124).
The spinal cord can undergo neuronal and synaptic changes due to inputs from the periphery, known as activity-dependent plasticity (131, 132). Spinal cord plasticity shapes its function during post-natal development and is continuously occurring throughout life (131, 133). Peripheral C-fiber activity drives activity-dependent cellular wind up, which initiates widespread changes in the function of the neuronal network in the spinal cord; this leads to clinical manifestations of spontaneous pain, abnormal sensitivity to noxious stimuli, or even innocuous stimuli, and referred pain that often follows injury to peripheral tissues (132). Changes induced by peripheral input depend on the intensity, duration, and the life stage at which the input occurs. Animal model research has been crucial to advance our understanding of the neuroanatomical and functional changes, and the effects on sensitivity to subsequent stimuli, produced by early life noxious stimuli. Neonatal rats exposed to a peripheral inflammatory stimulus in the hindpaw appear to have enhanced responsiveness to sensory stimulation due to altered axonal patterns in the substantia gelatinosa; these functional changes suggest that axons are sprouting into new areas of the dorsal horn (134). Furthermore, a single noxious input (CAR) at different ages between P0 and 8 in the neonatal rat has led to sustained thermal and mechanical hyperalgesia after re-inflammation (CFA at P40) in the ipsilateral, but not contralateral, hindpaw that is likely a result of local segmental circuitry involvement (135).
At the level of the spinal cord in rats, the balance between excitatory and inhibitory neurotransmitters shifts with maturity. In adults, substance P, calcitonin gene-related peptide, and glutamate are the neurotransmitters involved in excitatory pain transmission; γ-aminobutyric acid (GABA), norepinephrine, glycine, adenosine, endogenous cannabinoids, and opioid peptides are involved with inhibitory pain modulation (136). During early neurodevelopment (e.g., first post-natal week of rat life), GABA has an excitatory effect in the hippocampus caused by an inverted chloride gradient that induces depolarization (137). Similarly, there is a transient GABAergic excitatory effect in the immature dorsal horn; however, this shifts to inhibitory signaling by the end of the first post-natal week (138, 139).
Bremner et al. suggests that spinal GABAergic inhibitory transmission onto single dorsal horn cells “in vivo” is functional at P3 in rats and low chloride extrusion capacity does not impede the normal inhibitory functions of GABA (140). GABA activates voltage-gated sodium and calcium channels and potentiates the activity of N-methyl-D-aspartate (NMDA) receptors by reducing their voltage-dependent Mg2+ block (137). NMDA receptors, thought to be responsible for central sensitization (141), and activity-dependent tuning expand receptive fields in the dorsal horn until 42 gestational weeks, gradually reducing to adult size by 44 gestational weeks in humans (41). Together, increased expression of NMDA receptors and shifts in GABA function creates an excitatory neuronal environment during the most vulnerable period of a baby's life, when they may be exposed to multiple noxious insults. In an excitable neuronal environment, noxious and non-noxious stimuli may result in hypersensitivity, which is exacerbated with repeated stimuli (41).
In the mature nervous system, descending pathways are responsible for inhibiting noxious signals at the level of the spinal cord. Animal research has been crucial in understanding the maturation of descending modulatory systems, and therefore when the neonate is most vulnerable. During infancy the descending modulatory system is facilitatory, which is mediated by the mu-opioid receptor pathways in the rostroventral medulla (RVM) in the rat (142). As the RVM matures (e.g., P25–P40 in rat), the control over spinal nociceptive circuits transitions to descending inhibition (142). However, prior to P40, without full development of the descending inhibitory mechanisms, endogenously driven suppression of noxious stimuli from the periphery is not fully functional, thus the neonatal nervous system is more vulnerable to the effects of noxious stimuli (120, 121, 128, 143). Early noxious experiences can lead to permanent or long-lasting changes of the RVM circuits and other inhibitory mechanisms.
The vulnerability of the neonate to long term changes induced by noxious stimuli prior to maturation of the descending inhibitory pathway is highlighted by animal research. Neonatal incisional injury (P3) modifies descending pathways of spinal excitability from the RVM in early adulthood, producing acute hyperalgesia but long-term generalized hypoalgesia (P28) (144). In contrast, repetitive noxious stimulation (needle prick) in the neonatal rat (P0–14) has led to long-term thermal hypersensitivity (P56), likely due to alterations in the descending modulatory circuitry (59). Additionally, neonatal tissue inflammation (carrageenan; P3) has shown to lengthen thermal withdrawal latencies at the site of injury compared to controls; however, re-inflammation later in life (CFA; P50) exaggerates nocifensive responses, such as peripheral thermal hyperalgesia (145). These alterations in behavioral responses post-CFA re-injury are likely a result of enhanced RVM-evoked modulation of the pain reflexes, thus strengthened descending inhibition of the spinal cord (145).
Pruning, or the selection of active neuronal circuits, occurs throughout life; however, during the late second trimester and throughout the neonatal period, these processes and others that shape the neural architecture are particularly active in the human brain (146). In the neonatal period, there is rapid neuronal proliferation and differentiation including: maturation of oligodendrocytes; distribution and activation of microglia; differentiation, migration, and proliferation of cortical neurons; development of the subplate neurons, cerebral cortex, deep nuclear structures, and axons; formation of synaptic connections; and increase in cortical surface area and gyral formation (147). As discussed above, procedural pain in the NICU can have dramatic effects on brain development and function (13, 56).
Neonatal pain has been associated with reduced brain microstructure and volumes in humans (56) and pigs (148). Proposed mechanisms responsible for reduced brain volume and structure due to early life pain include alterations mediated by excitotoxicity and disrupted axonal development. Persistent neonatal pain in the rat can result in neuronal death in cortical and subcortical areas due to excitoxicity (149). Additionally, microglial activation in the rat initiates downstream intracellular cascades (e.g., release of glutamate, proinflammatory cytokines) that can result in cell death via excitotoxicity (150). Human imaging studies (e.g., diffusion tensor imaging, MRI) suggest pain-associated impairment in axonal development as another mechanism responsible for abnormal brain development (13). For example, early procedural pain in the premature newborn likely contributed to the development of damaged subcortical neurons with secondary axonal changes in the white matter (13).
Regions of the brain that appear to be most affected by early life pain and noxious input are associated with or connected to the limbic system (e.g., hippocampus, amygdala, and thalamus) and basal ganglia (48). In studies conducted by Nuseir et al., lower levels of hippocampal neurotrophins were detected in injured neonatal rats as compared to control rats, and the authors suggested this as the underlying mechanism for memory impairment (59, 60). The thalamus is the relay center for sensory and motor signals ascending to the cerebral cortex. Human infants who experience pain have decreased thalamic volumes, accompanied by disruptions in thalamic metabolic growth and thalamocortical pathway maturation (14). Thalamocortical connections undergo rapid formation during gestation and early post-natal life making them particularly vulnerable to excitotoxicity (151). Interestingly, the rat thalamus is regulated by NMDA-dependent long-term potentiation, which is only active during the first week of life (152), when newborns are routinely exposed to painful procedures. Therefore, early pain can alter thalamic development due to NMDA-dependent mechanisms. Furthermore, NMDA and GABA receptors are also key components of synaptic plasticity in the amygdala of the rodent and are presumed responsible for neuronal responsiveness and structural reorganization (153). Early life insults in the piglet can alter the normal developmental expression of GABA receptors, thereby impacting neuronal cytoskeleton development and myelination (154), which may lead to downstream effects on learning and cognitive impairments. These changes within the limbic system appear to be conserved across species throughout life affecting lifelong memory and associated fear (153, 155, 156). Factors and mechanisms adversely influencing brain development are being elucidated by using animal models, but there is much more to understand about the cellular and molecular mechanisms.
The neonate's immune system is immature and undergoes rapid changes early in life (157). It is well-understood that the immune system plays a critical role in pain modulation due to the tightly woven interactions of the nervous system and immune system (116–118). Specific components of the immune system play an intimate role in modulating pain sensitivity (158). A key player in immune-related pain are microglia cells, which make up 15% of the total glial population in the central nervous system (CNS) (159). Microglia are present during the embryonic and early post-natal period during which they inhabit the spinal cord and brain (160). Animal research has shown that the development and architecture of the CNS is fine-tuned and shaped by microglia, where they also contribute to neuroplasticity and lifelong maintenance and protection of the CNS (161). Microglia respond to noxious stimulation by activating and proliferating, which contributes to maladaptive pain and central sensitization. Microglia reactivity is a complex multistage activation process, where microglia undergo a morphological and functional transformation process. Early life challenges, including tissue injury, lead to microglial activation that may result in the release of numerous signaling molecules that can have deleterious effects on the pathogenesis of pain, especially in the neonate (161, 162). For example, spinal microglial reactivity, measured by immunostaining of microglia-specific ionized calcium-binding adapter molecule 1 (Iba-1), was enhanced in rats exposed to a noxious insult (e.g., incision, inflammation) as a neonate (16, 88, 163–165).
With subsequent noxious experiences, activated microglia are primed to respond to injury more rapidly. Microglial priming appears to last for extended periods of time. As a result, microglia have gained attention as a suggested cellular mechanisms linking early life pain to altered pain processing and chronic pain in adulthood (161). Thus, microglia have become a major focus of investigative pre-clinical studies into pain processing pathways, neuroplasticity, and as potential treatment targets. However, the mechanisms driving microglial priming are unknown, and it is unknown how long microglial priming lasts. Interestingly, a murine study suggests that while male mice require microglial activation to develop pain hypersensitivity, females have an alternative route via the adaptive immune system (166).
The most commonly used clinically relevant models (we use ‘model’ to refer only to the induced pain state) of neonatal early life pain are the repetitive needle prick (RNP) (37, 57) and plantar incision (16, 100, 144), most often using a rat. These painful insults are performed at various time points during the early post-natal period (P0–P7) in the rat because this critical period corresponds with that of a preterm human infant (24–36 weeks gestation) (167). The RNP model involves repeated tissue-breaking injury to the rodent's hindpaw using a needle to mimic the heel stick frequently performed in human babies in the NICU (8). Usually the noxious stimulus is administered between 4 and 8 times per day over a defined period of time during the early neonatal period (P1–14).
The plantar incision model involves incising the skin and fascia of the mid-plantar aspect of the hindpaw and the underlying plantar muscles followed by routine closure. The plantar incision is a clinically relevant model used to mimic surgical injury and has been used in animals (e.g., rats, pig) of varying ages to investigate post-operative incisional pain and the efficacy of various analgesic treatments (168–173). In the context of neonatal early life pain, the incisional model has been used in rats between P3 and P17 (97, 144). Both paradigms result in reliable neonatal tissue-breaking injury and nociceptive input thus serving as clinically relevant models to better understand the mechanisms and consequences of early life pain.
Other models used to evaluate the long-term effects of neonatal injury include the induction of inflammation (e.g., carrageenan, CFA), full-thickness skin wounds, laparotomy, intramuscular injections, and peripheral nerve injury models (Tables 2, 3) (101). Alternative models have focused on the psychological implications of early life pain and prematurity, such as stress (67). The most commonly used species are rodents; however, there are reports of other species being used. For example, piglets who undergo tail docking (P3 or P63) have long-term peripheral mechanical hyperalgesia (tail stump) (212) and distinctive changes in gene expression up to 16 weeks following the procedure (213).
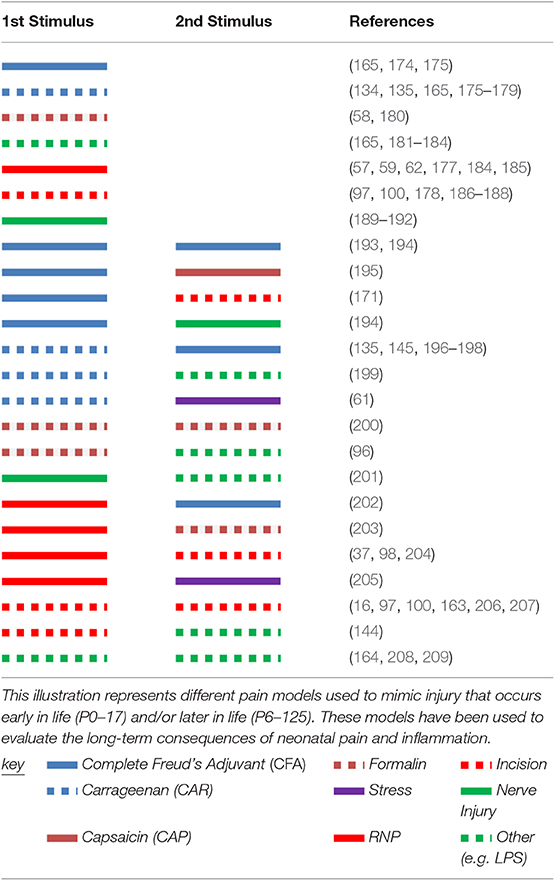
Table 2. Pain models used to evaluate long-term effects of neonatal pain with or without subsequent pain later in life.
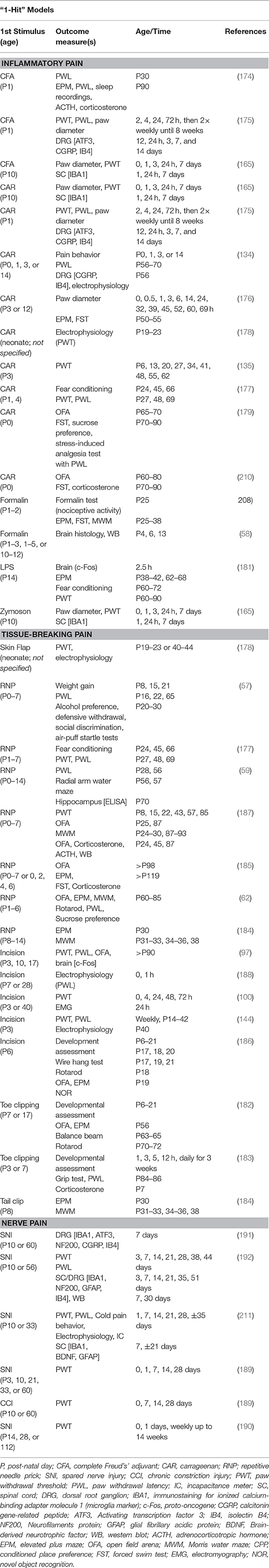
Table 3. Rodent models of neonatal pain and times of assessment.
Neonatal pain models (first-hit) have been superimposed with secondary (adult) painmodels (second-hit) to evaluate the effects of early life pain on subsequent pain later in life. Like one-hit neonatal pain models, two-hit models are primarily induced in the rat. Subsequent pain typically occurs between P25 and P60 and varies from inflammatory (e.g., CFA, CAR) to tissue-breaking (e.g., RNP, incision) in nature (Table 4). These time periods in the rat most likely resemble humans classified as young adults and middle-aged adults, respectively. Alone these models are used to evaluate the effects of early life pain on later acute pain (215–217). Two-hit models provide evidence that single or repetitive noxious stimulation during the early post-natal period can affect subsequent acute pain later in life in regards to mechanical and thermal sensitivity (37, 206), learning and memory (61, 200), and microglial activation (16, 163). However, the impact that early post-natal injury has on chronically painful conditions during adulthood has not been evaluated in animal models.
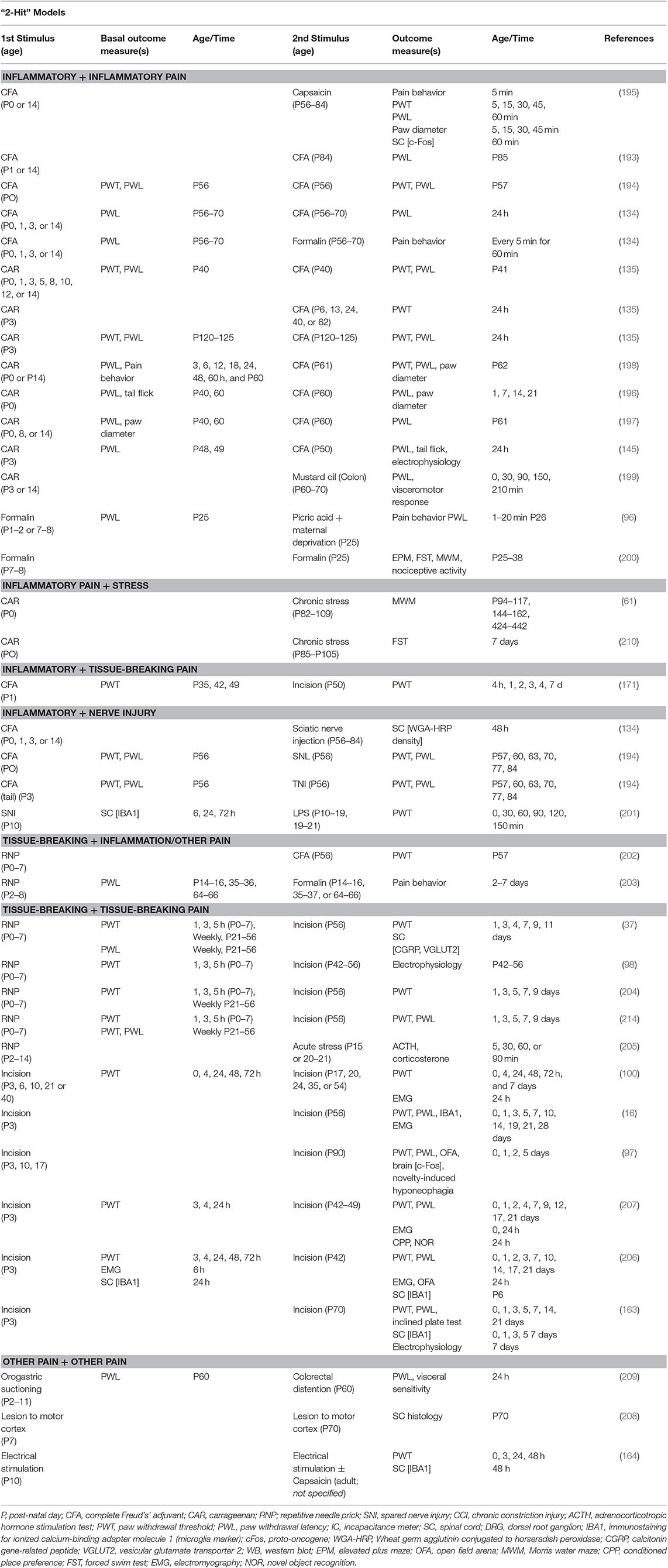
Table 4. Rodent models of neonatal pain with subsequent pain and time of assessments.
Chronic pain is a major global health concern affecting more than 1.5 billion people worldwide (218). Over the last decade, studies concerning the health status of adults with a history of varying degrees of prematurity, early life procedural pain, and/or early life surgery have been conducted to understand the effects of early life pain on diseases in adulthood (109, 219–221). Retrospective studies are very challenging to conduct because detailed medical records are not always accessible and/or there is a loss of patient retention due to geographic relocation or denied consent. These studies are further limited because of variability in details describing procedures performed including number of procedures, time of occurrence, and dose/route of drug administration in addition to differences in physician's and/or health facility's standard protocols, or lack thereof. Finally, early life painful events may not be captured during childhood or adult examinations because there is lack of patient recall, medical exam time constraints, or focused questions to gather pertinent information to current condition.
Regardless, untreated early life pain has been suggested to be a significant contributor to the development of chronic pain in children and adults (17, 97). It is speculated that this notion would be supported by animal models. However, two-hit models have focused on the effects of early pain on secondary acute pain in adult rats as previously discussed. Whether noxious stimuli during the neonatal period predisposes a patient (human or animal) to pain-associated with chronic painful conditions remains unknown. There are multiple animal models of chronic pain (222, 223) that could be coupled with an early life pain model to investigate whether there is an impact on chronic pain conditions. Well-documented chronic pain rodent models can be used to provide foundational data on pain responses and mechanistic pathways, and early life pain could be superimposed on this model to evaluate the influence of early life pain on chronic pain states. Indeed, this approach has been communicated in abstract form where we found that neonatal RNP pain appears to heighten reflexive and complex pain behaviors following chemical induction of osteoarthritis in adult rats, compared to osteoarthritis alone (224). However, there may be opportunities where using large animal models (e.g., dogs, pigs) would be preferred as these models are preexisting and better mimic human disease (225–228).
To date there is a complete lack of published clinical or basic science information on the effects of early life pain on persistently painful conditions in adults. Assessing ‘naturally occurring,’ or spontaneous, chronic pain conditions in animals with a history of early life pain could add to our understanding of the consequences of early life pain on later chronic pain (229). Companion animals commonly are surgically de-sexed, or sterilized (e.g., ovariohysterectomy or orchidectomy), at an early age (<6 months old), providing a natural model of early life surgery and pain. Such studies would be best performed prospectively, although one obvious shortcoming of such studies would be the removal of the influence (positive or negative) of sex hormones. To avoid hormonal confounders and the risk of missing the “critical period” of neurodevelopment other elective canine procedures may be used, such as tail docking (3–7 days old) or ear cropping (7–12 weeks old). Another clinically relevant example would be studying piglets who undergo painful procedures as a part of routine management practices (e.g., tail docking, castration, ear notching). Pigs commonly develop joint pain later in life and relevant outcome measures have been described. Interestingly, a piglet model has been developed to study the impact of NICU procedures (e.g., mechanical ventilation) in preterm infants (230), which could easily become a two-hit model.
Despite significant basic science work, chronic pain remains a significant public health crisis, where the prevalence of new cases has persistently increased in both adolescents (231) and adults (232–234). The influence of predisposing risk factors, such as age, gender, and obesity on chronic pain have been investigated for several decades, but there has been limited exploration of additional underlying risks, such as early life pain.
A growing body of evidence reports long-term consequences of early life pain in humans, and elegant animal studies are gradually shedding light on the neuroanatomical and functional mechanisms responsible for this (18, 88, 119, 235, 236). However, there is also the distinct possibility that early life pain may impact the degree and severity of chronically painful conditions that occur much later in life. To investigate this relationship, a series of systematic studies need to be conducted. First, a suitable two-hit model incorporating a painful procedure that consistently happens during the neonatal period and a disease that causes chronic pain in adulthood is needed. Second, an appropriate battery of outcome measures that are clinically relevant should be explored to evaluate changes to neurophysiology, psychology, and behavior. Finally, mechanisms responsible for long-term effects of early life pain should be determined using these models, and this will lay the foundation for the development of therapies to improve the quality of life of patients who suffer the consequences of early life pain experiences.
MW co-conceived the review, performed the literature searches and literature summaries, drafted and revised the manuscript, approved the final manuscript as submitted, and agreed to be accountable for all aspects of the work. BL co-conceived the review, assisted in the design of the content, made critical input and assisted in revisions, approved the final manuscript as submitted, and agreed to be accountable for all aspects of the work.
MW was supported by the National Institutes of Health (NIH) (T32 OD011130, 2017). BL salary was provided by NC State University.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
1. Howson CP, Kinney MV, Lawn JE. (editors). Born too Soon: the Global Action Report on Preterm Birth. Geneva: World Health Organization (2012). 126 p.
2. Dimes M, PMNCH, Children St, WHO. Born Too Soon: The Global Action Report on Preterm Birth. Geneva: World Health Organization (2012).
3. Blencowe H, Cousens S, Oestergaard MZ, Chou D, Moller AB, Narwal R, et al. National, regional, and worldwide estimates of preterm birth rates in the year 2010 with time trends since 1990 for selected countries: a systematic analysis and implications. Lancet. (2012) 379:2162–72. doi: 10.1016/S0140-6736(12)60820-4
4. Slattery MM, Morrison JJ. Preterm delivery. Lancet. (2002) 360:1489–97. doi: 10.1016/S0140-6736(02)11476-0
5. Behrman R, Butler A. Institute of Medicine (US) Committee on Understanding Premature Birth and Assuring Healthy Outcomes. Preterm Birth: Causes, Consequences, and Prevention. Washington, DC: National Academies Press (2007).
6. March of Dimes Perinatal Data Center: Special Care Nursery Admissions. White Plains, NY: National Perinatal Information System/Quality Analytic Services (2011).
7. Lee HC, Bennett MV, Schulman J, Gould JB. Accounting for variation in length of NICU stay for extremely low birth weight infants. J Perinatol. (2013) 33:872–6. doi: 10.1038/jp.2013.92
8. Carbajal R, Rousset A, Danan C, Coquery S, Nolent P, Ducrocq S, et al. Epidemiology and treatment of painful procedures in neonates in intensive care units. JAMA. (2008) 300:60–70. doi: 10.1001/jama.300.1.60
9. Hall RW, Anand KJ. Pain management in newborns. Clin Perinatol. (2014) 41:895–924. doi: 10.1016/j.clp.2014.08.010
10. Bower AJ. Plasticity in the adult and neonatal central nervous system. Br J Neurosurg. (1990) 4:253–64. doi: 10.3109/02688699008992734
11. Ochiai M, Kinjo T, Takahata Y, Iwayama M, Abe T, Ihara K, et al. Survival and neurodevelopmental outcome of preterm infants born at 22–24 weeks of gestational age. Neonatology. (2014) 105:79–84. doi: 10.1159/000355818
12. Saigal S, Doyle LW. An overview of mortality and sequelae of preterm birth from infancy to adulthood. Lancet. (2008) 371:261–9. doi: 10.1016/S0140-6736(08)60136-1
13. Brummelte S, Grunau RE, Chau V, Poskitt KJ, Brant R, Vinall J, et al. Procedural pain and brain development in premature newborns. Ann Neurol. (2012) 71:385–96. doi: 10.1002/ana.22267
14. Duerden EG, Grunau RE, Guo T, Foong J, Pearson A, Au-Young S, et al. Early procedural pain is associated with regionally-specific alterations in thalamic development in preterm neonates. J Neurosci. (2018) 38:878–86. doi: 10.1523/JNEUROSCI.0867-17.2017
15. Grunau RE. Neonatal pain in very preterm infants: long-term effects on brain, neurodevelopment and pain reactivity. Rambam Maimonides Med J. (2013) 4:e0025. doi: 10.5041/RMMJ.10132
16. Beggs S, Currie G, Salter MW, Fitzgerald M, Walker SM. Priming of adult pain responses by neonatal pain experience: maintenance by central neuroimmune activity. Brain. (2012) 135:404–17. doi: 10.1093/brain/awr288
17. Walker SM, Melbourne A, O'Reilly H, Beckmann J, Eaton-Rosen Z, Ourselin S, et al. Somatosensory function and pain in extremely preterm young adults from the UK EPICure cohort: sex-dependent differences and impact of neonatal surgery. Br J Anaesth. (2018) 121:623–35. doi: 10.1016/j.bja.2018.03.035
18. Grunau RE, Holsti L, Peters JW. Long-term consequences of pain in human neonates. Semin Fetal Neonatal Med. (2006) 11:268–75. doi: 10.1016/j.siny.2006.02.007
19. Johnston C, Barrington KJ, Taddio A, Carbajal R, Filion F. Pain in Canadian NICUs: have we improved over the past 12 years? Clin J Pain. (2011) 27:225–32. doi: 10.1097/AJP.0b013e3181fe14cf
20. Simons SH, van Dijk M, Anand KS, Roofthooft D, van Lingen RA, Tibboel D. Do we still hurt newborn babies? A prospective study of procedural pain and analgesia in neonates. Arch Pediatr Adolesc Med. (2003) 157:1058–64. doi: 10.1001/archpedi.157.11.1058
21. Johnston CC, Collinge JM, Henderson SJ, Anand KJ. A cross-sectional survey of pain and pharmacological analgesia in Canadian neonatal intensive care units. Clin J Pain. (1997) 13:308–12. doi: 10.1097/00002508-199712000-00008
22. Anand KJ, International Evidence-Based Group for Neonatal P. Consensus statement for the prevention and management of pain in the newborn. Arch Pediatr Adolesc Med. (2001) 155:173–80. doi: 10.1001/archpedi.155.2.173
23. Farmer D, Sitkin N, Lofberg K, Donkor P, Ozgediz D. Surgical interventions for congenital anomalies. In Debas HT, Donkor P, Gawande A, Jamison DT, Kruk ME, Mock CN, editors. Essential Surgery: Disease Control Priorities. The International Bank for Reconstruction and Development. Washington, DC: The World Bank (2015). p. 129–49. doi: 10.1596/978-1-4648-0346-8_ch8
24. Mitul AR, Sarin YK. Minimal access surgery in neonates. J Neonatal Surg. (2017) 6:59. doi: 10.21699/jns.v6i3.614
25. Courtois E, Droutman S, Magny JF, Merchaoui Z, Durrmeyer X, Roussel C, et al. Epidemiology and neonatal pain management of heelsticks in intensive care units: EPIPPAIN 2, a prospective observational study. Int J Nurs Stud. (2016) 59:79–88. doi: 10.1016/j.ijnurstu.2016.03.014
26. Shah VS, Ohlsson A. Venepuncture versus heel lance for blood sampling in term neonates. Cochrane Database Syst Rev. (2011) CD001452. doi: 10.1002/14651858.CD001452.pub4
27. Ma P, Liu J, Xi X, Du B, Yuan X, Lin H, et al. Practice of sedation and the perception of discomfort during mechanical ventilation in Chinese intensive care units. J Crit Care. (2010) 25:451–7. doi: 10.1016/j.jcrc.2009.11.006
28. Friesen RH, Honda AT, Thieme RE. Changes in anterior fontanel pressure in preterm neonates during tracheal intubation. Anesth Analg. (1987) 66:874–8. doi: 10.1213/00000539-198709000-00012
29. Gitto E, Aversa S, Salpietro CD, Barberi I, Arrigo T, Trimarchi G, et al. Pain in neonatal intensive care: role of melatonin as an analgesic antioxidant. J Pineal Res. (2012) 52:291–5. doi: 10.1111/j.1600-079X.2011.00941.x
31. Qiu J, Zhao L, Yang Y, Zhang JH, Feng Y, Cheng, R. Effects of fentanyl for pain control and neuroprotection in very preterm newborns on mechanical ventilation. J Matern Fetal Neonatal Med. (2019) 32:3734–40. doi: 10.1080/14767058.2018.1471593
32. Rozé JC, Denizot S, Carbajal R, Ancel PY, Kaminski M, Arnaud C, et al. Prolonged sedation and/or analgesia and 5-year neurodevelopment outcome in very preterm infants: results from the EPIPAGE cohort. Arch Pediatr Adolesc Med. (2008) 162:728–33. doi: 10.1001/archpedi.162.8.728
33. Zimmerman KO, Smith PB, Benjamin DK, Laughon M, Clark R, Traube C, et al. Sedation, analgesia, and paralysis during mechanical ventilation of premature infants. J Pediatr. (2017) 180:99–104.e1. doi: 10.1016/j.jpeds.2016.07.001
34. Anand KJ, Aranda JV, Berde CB, Buckman S, Capparelli EV, Carlo W, et al. Summary proceedings from the neonatal pain-control group. Pediatrics. (2006) 117:S9–22. doi: 10.1542/peds.2005-0620C
35. Doyle LW, Cheong JL, Burnett A, Roberts G, Lee KJ, Anderson PJ, et al. Biological and social influences on outcomes of extreme-preterm/low-birth weight adolescents. Pediatrics. (2015) 136:e1513–20. doi: 10.1542/peds.2015-2006
36. Fitzgerald M, Millard C, McIntosh N. Cutaneous hypersensitivity following peripheral tissue damage in newborn infants and its reversal with topical anaesthesia. Pain. (1989) 39:31–6. doi: 10.1016/0304-3959(89)90172-3
37. Knaepen L, Patijn J, van Kleef M, Mulder M, Tibboel D, Joosten EAJ. Neonatal repetitive needle pricking: Plasticity of the spinal nociceptive circuit and extended postoperative pain in later life. Dev Neurobiol. (2013) 73:85–97. doi: 10.1002/dneu.22047
38. Woolf CJ. Central sensitization: implications for the diagnosis and treatment of pain. Pain. (2011) 152:S2–15. doi: 10.1016/j.pain.2010.09.030
39. Woolf CJ, Thompson SW, King AE. Prolonged primary afferent induced alterations in dorsal horn neurones, an intracellular analysis in vivo and in vitro. J Physiol (Paris). (1988) 83:255–66.
40. Bellieni CV, Johnston CC. Analgesia, nil or placebo to babies, in trials that test new analgesic treatments for procedural pain. Acta Paediatr. (2016) 105:129–36. doi: 10.1111/apa.13210
41. Hatfield LA. Neonatal pain: what's age got to do with it? Surg Neurol Int. (2014) 5:S479–89. doi: 10.4103/2152-7806.144630
42. Roofthooft DW, Simons SH, Anand KJ, Tibboel D, van Dijk M. Eight years later, are we still hurting newborn infants? Neonatology. (2014) 105:218–26. doi: 10.1159/000357207
43. Johnston CC, Stevens BJ. Experience in a neonatal intensive care unit affects pain response. Pediatrics. (1996) 98:925–30.
44. Kaur H, Negi V, Sharma M, Mahajan G. Study of pain response in neonates during venipuncture with a view to analyse utility of topical anaesthetic agent for alleviating pain. Med J Armed Forces India. (2019) 75:140–5. doi: 10.1016/j.mjafi.2017.12.009
45. Maitre NL, Key AP, Chorna OD, Slaughter JC, Matusz PJ, Wallace MT, et al. The dual nature of early-life experience on somatosensory processing in the human infant brain. Curr Biol. (2017) 27:1048–54. doi: 10.1016/j.cub.2017.02.036
46. Burnett AC, Cheong JLY, Doyle LW. Biological and social influences on the neurodevelopmental outcomes of preterm infants. Clin Perinatol. (2018) 45:485–500. doi: 10.1016/j.clp.2018.05.005
47. Hunt RW, Hickey LM, Burnett AC, Anderson PJ, Cheong JLY, Doyle LW, et al. Early surgery and neurodevelopmental outcomes of children born extremely preterm. Arch Dis Child Fetal Neonatal Ed. (2018) 103:F227–32. doi: 10.1136/archdischild-2017-313161
48. Chau CMY, Ranger M, Bichin M, Park MTM, Amaral RSC, Chakravarty M, et al. Hippocampus, amygdala, and thalamus volumes in very preterm children at 8 years: neonatal pain and genetic variation. Front Behav Neurosci. (2019) 13:51. doi: 10.3389/fnbeh.2019.00051
49. Abernethy LJ, Cooke RW, Foulder-Hughes L. Caudate and hippocampal volumes, intelligence, and motor impairment in 7-year-old children who were born preterm. Pediatr Res. (2004) 55:884–93. doi: 10.1203/01.PDR.0000117843.21534.49
50. Allin M, Matsumoto H, Santhouse AM, Nosarti C, AlAsady MH, Stewart AL, et al. Cognitive and motor function and the size of the cerebellum in adolescents born very pre-term. Brain. (2001) 124:60–6. doi: 10.1093/brain/124.1.60
51. Ranger M, Zwicker JG, Chau CM, Park MT, Chakravarthy MM, Poskitt K, et al. Neonatal pain and infection relate to smaller cerebellum in very preterm children at school age. J Pediatr. (2015) 167:292–8.e1. doi: 10.1016/j.jpeds.2015.04.055
52. Jaekel J, Eryigit-Madzwamuse S, Wolke D. Preterm Toddlers' inhibitory control abilities predict attention regulation and academic achievement at age 8 years. J Pediatr. (2016) 169:87–92.e1. doi: 10.1016/j.jpeds.2015.10.029
53. Linsell L, Johnson S, Wolke D, O'Reilly H, Morris JK, Kurinczuk JJ, et al. Cognitive trajectories from infancy to early adulthood following birth before 26 weeks of gestation: a prospective, population-based cohort study. Arch Dis Child. (2018) 103:363–70. doi: 10.1136/archdischild-2017-313414
54. Olsen A, Dennis EL, Evensen KAI, Husby Hollund IM, Løhaugen GCC, Thompson PM, et al. Preterm birth leads to hyper-reactive cognitive control processing and poor white matter organization in adulthood. Neuroimage. (2018) 167:419–28. doi: 10.1016/j.neuroimage.2017.11.055
55. Skiöld B, Alexandrou G, Padilla N, Blennow M, Vollmer B, Adén U. Sex differences in outcome and associations with neonatal brain morphology in extremely preterm children. J Pediatr. (2014) 164:1012–8. doi: 10.1016/j.jpeds.2013.12.051
56. Schneider J, Duerden EG, Guo T, Ng K, Hagmann P, Bickle Graz M, et al. Procedural pain and oral glucose in preterm neonates: brain development and sex-specific effects. Pain. (2018) 159:515–25. doi: 10.1097/j.pain.0000000000001123
57. Anand KJS, Coskun V, Thrivikraman KV, Nemeroff CB, Plotksy PM. Long-term behavioral effects of repetitive pain in neonatal rat pups. Physiol Behav. (1999) 66:627–37. doi: 10.1016/S0031-9384(98)00338-2
58. Dührsen L, Simons SHP, Dzietko M, Genz K, Bendix I, Boos V, et al. Effects of repetitive exposure to pain and morphine treatment on the neonatal rat brain. Neonatology. (2013) 103:35–43. doi: 10.1159/000341769
59. Nuseir KQ, Alzoubi KH, Alhusban A, Bawaane A, Al-Azzani M, Khabour OF. Sucrose and naltrexone prevent increased pain sensitivity and impaired long-term memory induced by repetitive neonatal noxious stimulation: Role of BDNF and beta-endorphin. Physiol Behav. (2017) 179:213–9. doi: 10.1016/j.physbeh.2017.06.015
60. Nuseir KQ, Alzoubi KH, Alabwaini J, Khabour OF, Kassab MI. Sucrose-induced analgesia during early life modulates adulthood learning and memory formation. Physiol Behav. (2015) 145:84–90. doi: 10.1016/j.physbeh.2015.04.002
61. Henderson YO, Victoria NC, Inoue K, Murphy AZ, Parent MB. Early life inflammatory pain induces long-lasting deficits in hippocampal-dependent spatial memory in male and female rats. Neurobiol Learn Mem. (2015) 118:30–41. doi: 10.1016/j.nlm.2014.10.010
62. Ranger M, Tremblay S, Chau CMY, Holsti L, Grunau RE, Goldowitz D. Adverse behavioral changes in adult mice following neonatal repeated exposure to pain and sucrose. Front Psychol. (2018) 9:2394. doi: 10.3389/fpsyg.2018.02394
63. Ranger M, Grunau RE. Early repetitive pain in preterm infants in relation to the developing brain. Pain Manag. (2014) 4:57–67. doi: 10.2217/pmt.13.61
64. Hall RW, Anand KJS. Short- and long-term impact of neonatal pain and stress. Neoreviews. (2005) 6:e69–75. doi: 10.1542/neo.6-2-e69
65. Bhutta AT, Rovnaghi C, Simpson PM, Gossett JM, Scalzo FM, Anand KJ. Interactions of inflammatory pain and morphine in infant rats: long-term behavioral effects. Physiol Behav. (2001) 73:51–8. doi: 10.1016/S0031-9384(01)00432-2
66. Butkevich IP, Mikhailenko VA, Vershinina EA, Aloisi AM, Barr GA. Long-term effects of chronic buspirone during adolescence reduce the adverse influences of neonatal inflammatory pain and stress on adaptive behavior in adult male rats. Front Behav Neurosci. (2017) 11:11. doi: 10.3389/fnbeh.2017.00011
67. Burke NN, Finn DP, McGuire BE, Roche M. Psychological stress in early life as a predisposing factor for the development of chronic pain: clinical and preclinical evidence and neurobiological mechanisms. J Neurosci Res. (2017) 95:1257–70. doi: 10.1002/jnr.23802
68. Burt KB, Obradović J, Long JD, Masten AS. The interplay of social competence and psychopathology over 20 years: testing transactional and cascade models. Child Dev. (2008) 79:359–74. doi: 10.1111/j.1467-8624.2007.01130.x
69. Bauminger N, Solomon M, Rogers SJ. Externalizing and internalizing behaviors in ASD. Autism Res. (2010) 3:101–12. doi: 10.1002/aur.131
70. Vinall J, Miller SP, Synnes AR, Grunau RE. Parent behaviors moderate the relationship between neonatal pain and internalizing behaviors at 18 months corrected age in children born very prematurely. Pain. (2013) 154:1831–9. doi: 10.1016/j.pain.2013.05.050
71. Loe IM, Lee ES, Luna B, Feldman HM. Behavior problems of 9–16 year old preterm children: biological, sociodemographic, and intellectual contributions. Early Hum Dev. (2011) 87:247–52. doi: 10.1016/j.earlhumdev.2011.01.023
72. Schmidt LA, Miskovic V, Boyle M, Saigal S. Frontal electroencephalogram asymmetry, salivary cortisol, and internalizing behavior problems in young adults who were born at extremely low birth weight. Child Dev. (2010) 81:183–99. doi: 10.1111/j.1467-8624.2009.01388.x
73. Talge NM, Holzman C, Wang J, Lucia V, Gardiner J, Breslau N. Late-preterm birth and its association with cognitive and socioemotional outcomes at 6 years of age. Pediatrics. (2010) 126:1124–31. doi: 10.1542/peds.2010-1536
74. Walker SM, O'Reilly H, Beckmann J, Marlow N, Group ES. Conditioned pain modulation identifies altered sensitivity in extremely preterm young adult males and females. Br J Anaesth. (2018) 121:636–46. doi: 10.1016/j.bja.2018.05.066
75. Hussong AM, Jones DJ, Stein GL, Baucom DH, Boeding S. An internalizing pathway to alcohol use and disorder. Psychol Addict Behav. (2011) 25:390–404. doi: 10.1037/a0024519
76. Winters KC, Stinchfield RD, Latimer WW, Stone A. Internalizing and externalizing behaviors and their association with the treatment of adolescents with substance use disorder. J Subst Abuse Treat. (2008) 35:269–78. doi: 10.1016/j.jsat.2007.11.002
77. Pearl RL, White MA, Grilo CM. Weight bias internalization, depression, and self-reported health among overweight binge eating disorder patients. Obesity (Silver Spring). (2014) 22:E142–8. doi: 10.1002/oby.20617
78. Relland LM, Gehred A, Maitre NL. Behavioral and physiological signs for pain assessment in preterm and term neonates during a nociception-specific response: a systematic review. Pediatr Neurol. (2019) 90:13–23. doi: 10.1016/j.pediatrneurol.2018.10.001
79. Green G, Hartley C, Hoskin A, Duff E, Shriver A, Wilkinson D, et al. Behavioural discrimination of noxious stimuli in infants is dependent on brain maturation. Pain. (2019) 160:493–500. doi: 10.1097/j.pain.0000000000001425
80. Donia AE-S, Tolba OA. Effect of early procedural pain experience on subsequent pain responses among premature infants. Egypt Pediatr Assoc Gazette. (2016) 64:74–80. doi: 10.1016/j.epag.2016.03.002
81. Evans JC, McCartney EM, Lawhon G, Galloway J. Longitudinal comparison of preterm pain responses to repeated heelsticks. Pediatr Nurs. (2005) 31:216–21.
82. Grunau RE, Oberlander TF, Whitfield MF, Fitzgerald C, Lee SK. Demographic and therapeutic determinants of pain reactivity in very low birth weight neonates at 32 weeks' postconceptional age. Pediatrics. (2001) 107:105–12. doi: 10.1542/peds.107.1.105
83. Slater R, Cantarella A, Franck L, Meek J, Fitzgerald M. How well do clinical pain assessment tools reflect pain in infants? PLoS Med. (2008) 5:e129. doi: 10.1371/journal.pmed.0050129
84. Slater R, Cantarella A, Gallella S, Worley A, Boyd S, Meek J, et al. Cortical pain responses in human infants. J Neurosci. (2006) 26:3662–6. doi: 10.1523/JNEUROSCI.0348-06.2006
85. Guinsburg R, de Araújo Peres C, Branco de Almeida MF, de Cássia Xavier Balda R, Cássia Berenguel R, Tonelotto J, et al. Differences in pain expression between male and female newborn infants. Pain. (2000) 85:127–33. doi: 10.1016/S0304-3959(99)00258-4
86. Gao H, Li M, Xu G, Li F, Zhou J, Zou Y, et al. Effect of non-nutritive sucking and sucrose alone and in combination for repeated procedural pain in preterm infants: a randomized controlled trial. Int J Nurs Stud. (2018) 83:25–33. doi: 10.1016/j.ijnurstu.2018.04.006
87. Bartocci M, Bergqvist LL, Lagercrantz H, Anand KJ. Pain activates cortical areas in the preterm newborn brain. Pain. (2006) 122:109–17. doi: 10.1016/j.pain.2006.01.015
88. Schwaller F, Fitzgerald M. The consequences of pain in early life: injury-induced plasticity in developing pain pathways. Eur J Neurosci. (2014) 39:344–52. doi: 10.1111/ejn.12414
89. Schmelzle-Lubiecki BM, Campbell KA, Howard RH, Franck L, Fitzgerald M. Long-term consequences of early infant injury and trauma upon somatosensory processing. Eur J Pain. (2007) 11:799–809. doi: 10.1016/j.ejpain.2006.12.009
90. Walker SM, Franck LS, Fitzgerald M, Myles J, Stocks J, Marlow N. Long-term impact of neonatal intensive care and surgery on somatosensory perception in children born extremely preterm. Pain. (2009) 141:79–87. doi: 10.1016/j.pain.2008.10.012
91. Buskila D, Neumann L, Zmora E, Feldman M, Bolotin A, Press J. Pain sensitivity in prematurely born adolescents. Arch Pediatr Adolesc Med. (2003) 157:1079–82. doi: 10.1001/archpedi.157.11.1079
92. Taddio A, Goldbach M, Ipp M, Stevens B, Koren G. Effect of neonatal circumcision on pain responses during vaccination in boys. Lancet. (1995) 345:291–2. doi: 10.1016/S0140-6736(95)90278-3
93. Taddio A, Katz J, Ilersich AL, Koren G. Effect of neonatal circumcision on pain response during subsequent routine vaccination. Lancet. (1997) 349:599–603. doi: 10.1016/S0140-6736(96)10316-0
94. Peters JW, Schouw R, Anand KJ, van Dijk M, Duivenvoorden HJ, Tibboel D. Does neonatal surgery lead to increased pain sensitivity in later childhood? Pain. (2005) 114:444–54. doi: 10.1016/j.pain.2005.01.014
95. Valeri BO, Ranger M, Chau CM, Cepeda IL, Synnes A, Linhares MB, et al. Neonatal invasive procedures predict pain intensity at school age in children born very preterm. Clin J Pain. (2016) 32:1086–93. doi: 10.1097/AJP.0000000000000353
96. Butkevich IP, Mikhailenko VA, Vershinina EA, Aloisi AM. Effects of neonatal pain, stress and their interrelation on pain sensitivity in later life in male rats. Chin J Physiol. (2016) 59:225–31. doi: 10.4077/CJP.2016.BAE412
97. Low LA, Fitzgerald M. Acute pain and a motivational pathway in adult rats: influence of early life pain experience. PLoS ONE. (2012) 7:e34316. doi: 10.1371/journal.pone.0034316
98. van den Hoogen NJ, Patijn J, Tibboel D, Joosten BA, Fitzgerald M, Kwok CHT. Repeated touch and needle-prick stimulation in the neonatal period increases the baseline mechanical sensitivity and postinjury hypersensitivity of adult spinal sensory neurons. Pain. (2018) 159:1166–75. doi: 10.1097/j.pain.0000000000001201
99. Walker SM, Beggs S, Baccei ML. Persistent changes in peripheral and spinal nociceptive processing after early tissue injury. Exp Neurol. (2016) 275:253–60. doi: 10.1016/j.expneurol.2015.06.020
100. Walker SM, Tochiki KK, Fitzgerald M. Hindpaw incision in early life increases the hyperalgesic response to repeat surgical injury: critical period and dependence on initial afferent activity. Pain. (2009) 147:99–106. doi: 10.1016/j.pain.2009.08.017
101. Walker SM. Biological and neurodevelopmental implications of neonatal pain. Clin Perinatol. (2013) 40:471–91. doi: 10.1016/j.clp.2013.05.002
102. Huang QT, Gao YF, Zhong M, Yu YH. Preterm birth and subsequent risk of acute childhood leukemia: a meta-analysis of observational studies. Cell Physiol Biochem. (2016) 39:1229–38. doi: 10.1159/000447828
103. Vinall J, Miller SP, Chau V, Brummelte S, Synnes AR, Grunau RE. Neonatal pain in relation to postnatal growth in infants born very preterm. Pain. (2012) 153:1374–81. doi: 10.1016/j.pain.2012.02.007
104. Ibáñez L, Ong K, Dunger DB, de Zegher F. Early development of adiposity and insulin resistance after catch-up weight gain in small-for-gestational-age children. J Clin Endocrinol Metab. (2006) 91:2153–8. doi: 10.1210/jc.2005-2778
105. Greer FR. Long-term adverse outcomes of low birth weight, increased somatic growth rates, and alterations of body composition in the premature infant: review of the evidence. J Pediatr Gastroenterol Nutr. (2007) 45:S147–51. doi: 10.1097/01.mpg.0000302961.01170.29
106. Ong KK, Ahmed ML, Emmett PM, Preece MA, Dunger DB. Association between postnatal catch-up growth and obesity in childhood: prospective cohort study. BMJ. (2000) 320:967–71. doi: 10.1136/bmj.320.7240.967
107. Alexeev EE, Lönnerdal B, Griffin IJ. Effects of postnatal growth restriction and subsequent catch-up growth on neurodevelopment and glucose homeostasis in rats. BMC Physiol. (2015) 15:3. doi: 10.1186/s12899-015-0017-5
108. Juonala M, Cheung MM, Sabin MA, Burgner D, Skilton MR, Kähönen M, et al. Effect of birth weight on life-course blood pressure levels among children born premature: the cardiovascular risk in Young Finns Study. J Hypertens. (2015) 33:1542–8. doi: 10.1097/HJH.0000000000000612
109. Raju TNK, Buist AS, Blaisdell CJ, Moxey-Mims M, Saigal S. Adults born preterm: a review of general health and system-specific outcomes. Acta Paediatr. (2017) 106:1409–37. doi: 10.1111/apa.13880
110. DiLorenzo M, Pillai Riddell R, Holsti L. Beyond acute pain: understanding chronic pain in infancy. Children (Basel). (2016) 3:26. doi: 10.3390/children3040026
111. Pillai Riddell RR, Stevens BJ, McKeever P, Gibbins S, Asztalos L, Katz J, et al. Chronic pain in hospitalized infants: health professionals' perspectives. J Pain. (2009) 10:1217–25. doi: 10.1016/j.jpain.2009.04.013
112. van Ganzewinkel CJ, Anand KJ, Kramer BW, Andriessen P. Chronic pain in the newborn: toward a definition. Clin J Pain. (2014) 30:970–7. doi: 10.1097/AJP.0000000000000056
113. Vanegas H, Schaible HG. Descending control of persistent pain: inhibitory or facilitatory? Brain Res Brain Res Rev. (2004) 46:295–309. doi: 10.1016/j.brainresrev.2004.07.004
114. Ossipov MH, Morimura K, Porreca F. Descending pain modulation and chronification of pain. Curr Opin Support Palliat Care. (2014) 8:143–51. doi: 10.1097/SPC.0000000000000055
115. Dantzer R. Neuroimmune interactions: from the brain to the immune system and vice versa. Physiol Rev. (2018) 98:477–504. doi: 10.1152/physrev.00039.2016
116. Grace PM, Hutchinson MR, Maier SF, Watkins LR. Pathological pain and the neuroimmune interface. Nat Rev Immunol. (2014) 14:217–31. doi: 10.1038/nri3621
117. Marchand F, Perretti M, McMahon SB. Role of the immune system in chronic pain. Nat Rev Neurosci. (2005) 6:521–32. doi: 10.1038/nrn1700
118. Ren K, Dubner R. Interactions between the immune and nervous systems in pain. Nat Med. (2010) 16:1267–76. doi: 10.1038/nm.2234
119. Beggs S. Long-term consequences of neonatal injury. Can J Psychiatry. (2015) 60:176–80. doi: 10.1177/070674371506000404
120. Beggs S, Torsney C, Drew LJ, Fitzgerald M. The postnatal reorganization of primary afferent input and dorsal horn cell receptive fields in the rat spinal cord is an activity-dependent process. Eur J Neurosci. (2002) 16:1249–58. doi: 10.1046/j.1460-9568.2002.02185.x
121. Fitzgerald M, Walker SM. Infant pain management: a developmental neurobiological approach. Nat Clin Pract Neurol. (2009) 5:35–50. doi: 10.1038/ncpneuro0984
122. Anand KJ, Scalzo FM. Can adverse neonatal experiences alter brain development and subsequent behavior? Biol Neonate. (2000) 77:69–82. doi: 10.1159/000014197
123. Grunau R. Early pain in preterm infants. A model of long-term effects. Clin Perinatol. (2002) 29:373–94, vii–viii. doi: 10.1016/S0095-5108(02)00012-X
124. Fitzgerald M. The development of nociceptive circuits. Nat Rev Neurosci. (2005) 6:507–20. doi: 10.1038/nrn1701
125. Rodkey EN, Pillai Riddell R. The infancy of infant pain research: the experimental origins of infant pain denial. J Pain. (2013) 14:338–50. doi: 10.1016/j.jpain.2012.12.017
126. Anand KJ. Clinical importance of pain and stress in preterm neonates. Biol Neonate. (1998) 73:1–9. doi: 10.1159/000013953
127. Fitzgerald M, Shaw A, MacIntosh N. Postnatal development of the cutaneous flexor reflex: comparative study of preterm infants and newborn rat pups. Dev Med Child Neurol. (1988) 30:520–6. doi: 10.1111/j.1469-8749.1988.tb04779.x
128. Goksan S, Hartley C, Emery F, Cockrill N, Poorun R, Moultrie F, et al. fMRI reveals neural activity overlap between adult and infant pain. Elife. (2015) 4:e6356. doi: 10.7554/eLife.06356
129. Steeds CE. The anatomy and physiology of pain. Surgery (Oxford). (2009) 27:507–11. doi: 10.1016/j.mpsur.2009.10.013
130. Zouikr I, Karshikoff B. Lifetime modulation of the pain system via neuroimmune and neuroendocrine interactions. Front Immunol. (2017) 8:276. doi: 10.3389/fimmu.2017.00276
131. Tahayori B, Koceja DM. Activity-dependent plasticity of spinal circuits in the developing and mature spinal cord. Neural Plast. (2012) 2012:964843. doi: 10.1155/2012/964843
132. Wolpaw JR, Tennissen AM. Activity-dependent spinal cord plasticity in health and disease. Annu Rev Neurosci. (2001) 24:807–43. doi: 10.1146/annurev.neuro.24.1.807
133. Smith CC, Paton JFR, Chakrabarty S, Ichiyama RM. Descending systems direct development of key spinal motor circuits. J Neurosci. (2017) 37:6372–87. doi: 10.1523/JNEUROSCI.0149-17.2017
134. Ruda MA, Ling QD, Hohmann AG, Peng YB, Tachibana T. Altered nociceptive neuronal circuits after neonatal peripheral inflammation. Science. (2000) 289:628–31. doi: 10.1126/science.289.5479.628
135. Ren K, Anseloni V, Zou SP, Wade EB, Novikova SI, Ennis M, et al. Characterization of basal and re-inflammation-associated long-term alteration in pain responsivity following short-lasting neonatal local inflammatory insult. Pain. (2004) 110:588–96. doi: 10.1016/j.pain.2004.04.006
136. Yam MF, Loh YC, Tan CS, Khadijah Adam S, Abdul Manan N, Basir R. General pathways of pain sensation and the major neurotransmitters involved in pain regulation. Int J Mol Sci. (2018) 19:E2164. doi: 10.3390/ijms19082164
137. Leinekugel X, Khalilov I, McLean H, Caillard O, Gaiarsa JL, Ben-Ari Y, et al. GABA is the principal fast-acting excitatory transmitter in the neonatal brain. Adv Neurol. (1999) 79:189–201.
138. Baccei ML, Bardoni R, Fitzgerald M. Development of nociceptive synaptic inputs to the neonatal rat dorsal horn: glutamate release by capsaicin and menthol. J Physiol. (2003) 549:231–42. doi: 10.1113/jphysiol.2003.040451
139. Baccei ML, Fitzgerald M. Development of GABAergic and glycinergic transmission in the neonatal rat dorsal horn. J Neurosci. (2004) 24:4749–57. doi: 10.1523/JNEUROSCI.5211-03.2004
140. Bremner L, Fitzgerald M, Baccei M. Functional GABA(A)-receptor-mediated inhibition in the neonatal dorsal horn. J Neurophysiol. (2006) 95:3893–7. doi: 10.1152/jn.00123.2006
141. Latremoliere A, Woolf CJ. Central sensitization: a generator of pain hypersensitivity by central neural plasticity. J Pain. (2009) 10:895–926. doi: 10.1016/j.jpain.2009.06.012
142. Hathway GJ, Vega-Avelaira D, Fitzgerald M. A critical period in the supraspinal control of pain: opioid-dependent changes in brainstem rostroventral medulla function in preadolescence. Pain. (2012) 153:775–83. doi: 10.1016/j.pain.2011.11.011
143. Fitzgerald M, Millard C, MacIntosh N. Hyperalgesia in premature infants. Lancet. (1988) 1:292. doi: 10.1016/S0140-6736(88)90365-0
144. Walker SM, Fitzgerald M, Hathway GJ. Surgical injury in the neonatal rat alters the adult pattern of descending modulation from the rostroventral medulla. Anesthesiology. (2015) 122:1391–400. doi: 10.1097/ALN.0000000000000658
145. Zhang YH, Wang XM, Ennis M. Effects of neonatal inflammation on descending modulation from the rostroventromedial medulla. Brain Res Bull. (2010) 83:16–22. doi: 10.1016/j.brainresbull.2010.07.007
146. Huang H, Zhang J, Wakana S, Zhang W, Ren T, Richards LJ, et al. White and gray matter development in human fetal, newborn and pediatric brains. Neuroimage. (2006) 33:27–38. doi: 10.1016/j.neuroimage.2006.06.009
147. Volpe JJ. The encephalopathy of prematurity–brain injury and impaired brain development inextricably intertwined. Semin Pediatr Neurol. (2009) 16:167–78. doi: 10.1016/j.spen.2009.09.005
148. Plomgaard AM, Andersen AD, Petersen TH, van de Looij Y, Thymann T, Sangild PT, et al. Structural brain maturation differs between preterm and term piglets, whereas brain activity does not. Acta Paediatr. (2019) 108:637–44. doi: 10.1111/apa.14556
149. Anand KJ, Garg S, Rovnaghi CR, Narsinghani U, Bhutta AT, Hall RW. Ketamine reduces the cell death following inflammatory pain in newborn rat brain. Pediatr Res. (2007) 62:283–90. doi: 10.1203/PDR.0b013e3180986d2f
150. Gwak YS, Kang J, Unabia GC, Hulsebosch CE. Spatial and temporal activation of spinal glial cells: role of gliopathy in central neuropathic pain following spinal cord injury in rats. Exp Neurol. (2012) 234:362–72. doi: 10.1016/j.expneurol.2011.10.010
151. Verriotis M, Chang P, Fitzgerald M, Fabrizi L. The development of the nociceptive brain. Neuroscience. (2016) 338:207–19. doi: 10.1016/j.neuroscience.2016.07.026
152. Crair MC, Malenka RC. A critical period for long-term potentiation at thalamocortical synapses. Nature. (1995) 375:325–8. doi: 10.1038/375325a0
153. Pape HC, Stork O. Genes and mechanisms in the amygdala involved in the formation of fear memory. Ann N Y Acad Sci. (2003) 985:92–105. doi: 10.1111/j.1749-6632.2003.tb07074.x
154. Kalanjati VP, Wixey JA, Miller SM, Colditz PB, Bjorkman ST. GABAA receptor expression and white matter disruption in intrauterine growth restricted piglets. Int J Dev Neurosci. (2017) 59:1–9. doi: 10.1016/j.ijdevneu.2017.02.004
155. Janak PH, Tye KM. From circuits to behaviour in the amygdala. Nature. (2015) 517:284–92. doi: 10.1038/nature14188
156. McGaugh JL, Cahill L, Roozendaal B. Involvement of the amygdala in memory storage: interaction with other brain systems. Proc Natl Acad Sci USA. (1996) 93:13508–14. doi: 10.1073/pnas.93.24.13508
157. Goenka A, Kollmann TR. Development of immunity in early life. J Infect. (2015) 71:S112–20. doi: 10.1016/j.jinf.2015.04.027
158. Totsch SK, Sorge RE. Immune system involvement in specific pain conditions. Mol Pain. (2017) 13:1744806917724559. doi: 10.1177/1744806917724559
159. Lawson LJ, Perry VH, Dri P, Gordon S. Heterogeneity in the distribution and morphology of microglia in the normal adult mouse brain. Neuroscience. (1990) 39:151–70. doi: 10.1016/0306-4522(90)90229-W
160. Verney C, Monier A, Fallet-Bianco C, Gressens P. Early microglial colonization of the human forebrain and possible involvement in periventricular white-matter injury of preterm infants. J Anat. (2010) 217:436–48. doi: 10.1111/j.1469-7580.2010.01245.x
161. Burke NN, Fan CY, Trang T. Microglia in health and pain: impact of noxious early life events. Exp Physiol. (2016) 101:1003–21. doi: 10.1113/EP085714
162. Scholz J, Woolf CJ. The neuropathic pain triad: neurons, immune cells and glia. Nat Neurosci. (2007) 10:1361–8. doi: 10.1038/nn1992
163. Ding X, Liang YJ, Su L, Liao FF, Fang D, Tai J, et al. BDNF contributes to the neonatal incision-induced facilitation of spinal long-term potentiation and the exacerbation of incisional pain in adult rats. Neuropharmacology. (2018) 137:114–32. doi: 10.1016/j.neuropharm.2018.04.032
164. Hathway GJ, Vega-Avelaira D, Moss A, Ingram R, Fitzgerald M. Brief, low frequency stimulation of rat peripheral C-fibres evokes prolonged microglial-induced central sensitization in adults but not in neonates. Pain. (2009) 144:110–8. doi: 10.1016/j.pain.2009.03.022
165. Vega-Avelaira D, Ballesteros JJ, López-García JA. Inflammation-induced hyperalgesia and spinal microglia reactivity in neonatal rats. Eur J Pain. (2013) 17:1180–8. doi: 10.1002/j.1532-2149.2013.00308.x
166. Sorge RE, Mapplebeck JC, Rosen S, Beggs S, Taves S, Alexander JK, et al. Different immune cells mediate mechanical pain hypersensitivity in male and female mice. Nat Neurosci. (2015) 18:1081–3. doi: 10.1038/nn.4053
167. Kim JJ, Foy MR, Thompson RF. Behavioral stress modifies hippocampal plasticity through N-methyl-D-aspartate receptor activation. Proc Natl Acad Sci USA. (1996) 93:4750–3. doi: 10.1073/pnas.93.10.4750
168. Brennan TJ, Vandermeulen EP, Gebhart GF. Characterization of a rat model of incisional pain. Pain. (1996) 64:493–501. doi: 10.1016/0304-3959(95)01441-1
169. Brennan TJ, Zahn PK, Pogatzki-Zahn EM. Mechanisms of incisional pain. Anesthesiol Clin North Am. (2005) 23:1–20. doi: 10.1016/j.atc.2004.11.009
170. Castel D, Sabbag I, Meilin S. The effect of local/topical analgesics on incisional pain in a pig model. J Pain Res. (2019) 10:2169–75. doi: 10.2147/JPR.S144949
171. Chu YC, Chan KH, Tsou MY, Lin SM, Hsieh YC, Tao YX. Mechanical pain hypersensitivity after incisional surgery is enhanced in rats subjected to neonatal peripheral inflammation: effects of N-methyl-D-aspartate receptor antagonists. Anesthesiology. (2007) 106:1204–12. doi: 10.1097/01.anes.0000267604.40258.d1
172. Gilchrist HD, Allard BL, Simone DA. Enhanced withdrawal responses to heat and mechanical stimuli following intraplantar injection of capsaicin in rats. Pain. (1996) 67:179–88. doi: 10.1016/0304-3959(96)03104-1
173. Kang S, Wu C, Banik RK, Brennan TJ. Effect of capsaicin treatment on nociceptors in rat glabrous skin one day after plantar incision. Pain. (2010) 148:128–40. doi: 10.1016/j.pain.2009.10.031
174. Roizenblatt S, Andersen ML, Bignotto M, D'Almeida V, Martins PJ, Tufik S. Neonatal arthritis disturbs sleep and behaviour of adult rat offspring and their dams. Eur J Pain. (2010) 14:985–91. doi: 10.1016/j.ejpain.2010.03.008
175. Walker SM, Meredith-Middleton J, Cooke-Yarborough C, Fitzgerald M. Neonatal inflammation and primary afferent terminal plasticity in the rat dorsal horn. Pain. (2003) 105:185–95. doi: 10.1016/S0304-3959(03)00201-X
176. Anseloni VC, He F, Novikova SI, Turnbach Robbins M, Lidow IA, Ennis M, et al. Alterations in stress-associated behaviors and neurochemical markers in adult rats after neonatal short-lasting local inflammatory insult. Neuroscience. (2005) 131:635–45. doi: 10.1016/j.neuroscience.2004.11.039
177. Davis SM, Rice M, Rudlong J, Eaton V, King T, Burman MA. Neonatal pain and stress disrupts later-life pavlovian fear conditioning and sensory function in rats: evidence for a two-hit model. Dev Psychobiol. (2018) 60:520–33. doi: 10.1002/dev.21632
178. Torsney C, Fitzgerald M. Spinal dorsal horn cell receptive field size is increased in adult rats following neonatal hindpaw skin injury. J Physiol. (2003) 550:255–61. doi: 10.1113/jphysiol.2003.043661
179. Victoria NC, Inoue K, Young LJ, Murphy AZ. A single neonatal injury induces life-long deficits in response to stress. Dev Neurosci. (2013) 35:326–37. doi: 10.1159/000351121
180. Butkevich IP, Mikhailenko VA, Vershinina EA, Ulanova NA. Differences in adaptive behaviors of adolescent male and female rats exposed at birth to inflammatory pain or stress. J Evol Biochem Physiol. (2015) 51:305–15. doi: 10.1134/S0022093015040067
181. Doenni VM, Song CM, Hill MN, Pittman QJ. Early-life inflammation with LPS delays fear extinction in adult rodents. Brain Behav Immun. (2017) 63:176–85. doi: 10.1016/j.bbi.2016.11.022
182. Paluch LR, Lieggi CC, Dumont M, Monette S, Riedel ER, Lipman NS. Developmental and behavioral effects of toe clipping on neonatal and preweanling mice with and without vapocoolant anesthesia. J Am Assoc Lab Anim Sci. (2014) 53:132–40.
183. Schaefer DC, Asner IN, Seifert B, Bürki K, Cinelli P. Analysis of physiological and behavioural parameters in mice after toe clipping as newborns. Lab Anim. (2010) 44:7–13. doi: 10.1258/la.2009.009020
184. Schellinck HM, Stanford L, Darrah M. Repetitive acute pain in infancy increases anxiety but does not alter spatial learning ability in juvenile mice. Behav Brain Res. (2003) 142:157–65. doi: 10.1016/S0166-4328(02)00406-0
185. Page GG, Blakely WP, Kim M. The impact of early repeated pain experiences on stress responsiveness and emotionality at maturity in rats. Brain Behav Immun. (2005) 19:78–87. doi: 10.1016/j.bbi.2004.05.002
186. Catré D, Lopes MF, Cabrita AS. Lasting developmental effects of neonatal fentanyl exposure in preweanling rats. Anesthesiol Res Pract. (2012) 2012:180124. doi: 10.1155/2012/180124
187. Chen M, Xia D, Min C, Zhao X, Chen Y, Liu L, et al. Neonatal repetitive pain in rats leads to impaired spatial learning and dysregulated hypothalamic-pituitary-adrenal axis function in later life. Sci Rep. (2016) 6:39159. doi: 10.1038/srep39159
188. Ririe DG, Bremner LR, Fitzgerald M. Comparison of the immediate effects of surgical incision on dorsal horn neuronal receptive field size and responses during postnatal development. Anesthesiology. (2008) 109:698–706. doi: 10.1097/ALN.0b013e3181870a32
189. Howard RF, Walker SM, Mota PM, Fitzgerald M. The ontogeny of neuropathic pain: postnatal onset of mechanical allodynia in rat spared nerve injury (SNI) and chronic constriction injury (CCI) models. Pain. (2005) 115:382–9. doi: 10.1016/j.pain.2005.03.016
190. Ririe DG, Eisenach JC. Age-dependent responses to nerve injury-induced mechanical allodynia. Anesthesiology. (2006) 104:344–50. doi: 10.1097/00000542-200602000-00021
191. Vega-Avelaira D, Géranton SM, Fitzgerald M. Differential regulation of immune responses and macrophage/neuron interactions in the dorsal root ganglion in young and adult rats following nerve injury. Mol Pain. (2009) 5:70. doi: 10.1186/1744-8069-5-70
192. Vega-Avelaira D, McKelvey R, Hathway G, Fitzgerald M. The emergence of adolescent onset pain hypersensitivity following neonatal nerve injury. Mol Pain. (2012) 30:30. doi: 10.1186/1744-8069-8-30
193. Blom JM, Benatti C, Alboni S, Capone G, Ferraguti C, Brunello N, et al. Early postnatal chronic inflammation produces long-term changes in pain behavior and N-methyl-D-aspartate receptor subtype gene expression in the central nervous system of adult mice. J Neurosci Res. (2006) 84:1789–98. doi: 10.1002/jnr.21077
194. Lim EJ, Back SK, Kim MA, Li C, Lee J, Jeong KY, et al. Long-lasting neonatal inflammation enhances pain responses to subsequent inflammation, but not peripheral nerve injury in adult rats. Int J Dev Neurosci. (2009) 27:215–22. doi: 10.1016/j.ijdevneu.2009.01.005
195. Hohmann AG, Neely MH, Pina J, Nackley AG. Neonatal chronic hind paw inflammation alters sensitization to intradermal capsaicin in adult rats: a behavioral and immunocytochemical study. J Pain. (2005) 6:798–808. doi: 10.1016/j.jpain.2005.07.009
196. Laprairie JL, Johns ME, Murphy AZ. Preemptive morphine analgesia attenuates the long-term consequences of neonatal inflammation in male and female rats. Pediatr Res. (2008) 64:625–30. doi: 10.1203/PDR.0b013e31818702d4
197. LaPrairie JL, Murphy AZ. Female rats are more vulnerable to the long-term consequences of neonatal inflammatory injury. Pain. (2007) 132:S124–33. doi: 10.1016/j.pain.2007.08.010
198. Lidow MS, Song ZM, Ren K. Long-term effects of short-lasting early local inflammatory insult. Neuroreport. (2001) 12:399–403. doi: 10.1097/00001756-200102120-00042
199. Wang G, Ji Y, Lidow MS, Traub RJ. Neonatal hind paw injury alters processing of visceral and somatic nociceptive stimuli in the adult rat. J Pain. (2004) 5:440–9. doi: 10.1016/j.jpain.2004.07.003
200. Mikhailenko VA, Butkevich IP, Vershinina EA, Ulanova NA. Long-term changes in adaptive behavior of rats after neonatal inflammatory pain. Zh Evol Biokhim Fiziol. (2015) 51:108–14. doi: 10.1134/S0022093015020052
201. Moss A, Beggs S, Vega-Avelaira D, Costigan M, Hathway GJ, Salter MW, et al. Spinal microglia and neuropathic pain in young rats. Pain. (2007) 128:215–24. doi: 10.1016/j.pain.2006.09.018
202. Knaepen L, Patijn J, Tibboel D, Joosten EA. Sex differences in inflammatory mechanical hypersensitivity in later life of rats exposed to repetitive needle pricking as neonates. Neurosci Lett. (2012) 516:285–9. doi: 10.1016/j.neulet.2012.04.012
203. Johnston CC, Walker CD. The effects of exposure to repeated minor pain during the neonatal period on formalin pain behaviour and thermal withdrawal latencies. Pain Res Manag. (2003) 8:213–7. doi: 10.1155/2003/305409
204. van den Hoogen NJ, Tibboel D, Honig WM, Hermes D, Patijn J, Joosten EA. Neonatal paracetamol treatment reduces long-term nociceptive behaviour after neonatal procedural pain in rats. Eur J Pain. (2016) 20:1309–18. doi: 10.1002/ejp.855
205. Walker CD, Kudreikis K, Sherrard A, Johnston CC. Repeated neonatal pain influences maternal behavior, but not stress responsiveness in rat offspring. Dev Brain Res. (2003) 140:253–61. doi: 10.1016/S0165-3806(02)00611-9
206. Moriarty O, Tu Y, Sengar AS, Salter MW, Beggs S, Walker SM. Priming of adult incision response by early life injury: neonatal microglial inhibition has persistent but sexually dimorphic effects in adult rats. J Neurosci. (2019) 39:3081–93. doi: 10.1523/JNEUROSCI.1786-18.2019
207. Moriarty O, Harrington L, Beggs S, Walker S. Opioid analgesia and the somatosensory memory of neonatal surgical injury in the adult rat. Br J Anaesth. (2018) 121:314–24. doi: 10.1016/j.bja.2017.11.111
208. Gibson CL, Arnott GA, Clowry GJ. Plasticity in the rat spinal cord seen in response to lesions to the motor cortex during development but not to lesions in maturity. Exp Neurol. (2000) 166:422–34. doi: 10.1006/exnr.2000.7511
209. Smith C, Nordstrom E, Sengupta JN, Miranda A. Neonatal gastric suctioning results in chronic visceral and somatic hyperalgesia: role of corticotropin releasing factor. Neurogastroenterol Motil. (2007) 19:692–9. doi: 10.1111/j.1365-2982.2007.00949.x
210. Victoria NC, Karom MC, Murphy AZ. Analgesia for early-life pain prevents deficits in adult anxiety and stress in rats. Dev Neurosci. (2015) 37:1–13. doi: 10.1159/000366273
211. McKelvey R, Berta T, Old E, Ji RR, Fitzgerald M. Neuropathic pain is constitutively suppressed in early life by anti-inflammatory neuroimmune regulation. J Neurosci. (2015) 35:457–66. doi: 10.1523/JNEUROSCI.2315-14.2015
212. Giminiani PD, Edwards SA, Malcolm EM, Leach MC, Herskin MS, Sandercock DA. Characterization of short- and long-term mechanical sensitisation following surgical tail amputation in pigs. Sci Rep. (2017) 7:1–9. doi: 10.1038/s41598-017-05404-y
213. Sandercock DA, Barnett MW, Coe JE, Downing AC, Nirmal AJ, Di Giminiani P, et al. Transcriptomics analysis of porcine caudal dorsal root ganglia in tail amputated pigs shows long-term effects on many pain-associated genes. Front Vet Sci. (2019) 6:314. doi: 10.3389/fvets.2019.00314
214. van den Hoogen NJ, Patijn J, Tibboel D, Joosten EA. Repetitive noxious stimuli during early development affect acute and long-term mechanical sensitivity in rats. Pediatr Res. (2019) 87:26–31. doi: 10.1038/s41390-019-0420-x
215. Fehrenbacher JC, Vasko MR, Duarte DB. Models of inflammation: Carrageenan- or complete Freund's Adjuvant (CFA)-induced edema and hypersensitivity in the rat. Curr Protoc Pharmacol. (2012) Chapter 5:Unit5.4. doi: 10.1002/0471141755.ph0504s56
216. McKeon GP, Pacharinsak C, Long CT, Howard AM, Jampachaisri K, Yeoman DC, et al. Analgesic effects of tramadol, tramadol–gabapentin, and buprenorphine in an incisional model of pain in rats (Rattus norvegicus). J Am Assoc Lab Anim Sci. (2011) 50:192–7.
217. Peters CM, Hayashida K, Suto T, Houle TT, Aschenbrenner CA, Martin TJ, et al. Individual differences in acute pain-induced endogenous analgesia predict time to resolution of postoperative pain in the rat. Anesthesiology. (2015) 122:895–907. doi: 10.1097/ALN.0000000000000593
218. Institute of Medicine Committee on Advancing Pain Research CE. The National Academies Collection: Reports funded by National Institutes of Health. Relieving Pain in America: A Blueprint for Transforming Prevention, Care, Education, and Research. Washington, DC: National Academies Press; National Academy of Sciences (2011).
219. Jones GT. Psychosocial vulnerability and early life adversity as risk factors for central sensitivity syndromes. Curr Rheumatol Rev. (2016) 12:140–53. doi: 10.2174/1573397112666151231113438
220. Littlejohn C, Pang D, Power C, Macfarlane GJ, Jones GT. Is there an association between preterm birth or low birthweight and chronic widespread pain? Results from the 1958 Birth Cohort Study. Eur J Pain. (2012) 16:134–9. doi: 10.1016/j.ejpain.2011.05.015
221. Pyhala R, Wolford E, Kautiainen H, Andersson S, Bartmann P, Baumann N, et al. Self-reported mental health problems among adults born preterm: a meta-analysis. Pediatrics. (2017) 139:e20162690. doi: 10.1542/peds.2016-2690
222. Burma NE, Leduc-Pessah H, Fan CY, Trang T. Animal models of chronic pain: advances and challenges for clinical translation. J Neurosci Res. (2017) 95:1242–56. doi: 10.1002/jnr.23768
223. Gregory NS, Harris AL, Robinson CR, Dougherty PM, Fuchs PN, Sluka KA. An overview of animal models of pain: disease models and outcome measures. J Pain. (2013) 14:1255–69. doi: 10.1016/j.jpain.2013.06.008
224. Williams M, Meyers R, Lascelles B. PS19 long-term impacts of early life injury on monoiodoacetate induced osteoarthritis in adult rats. J Am Assoc Lab Anim Sci. (2018) 57:610. Available online at: https://www.ingentaconnect.com/contentone/aalas/jaalas/2018/00000057/00000005/art00013
225. Cope PJ, Ourradi K, Li Y, Sharif M. Models of osteoarthritis: the good, the bad and the promising. Osteoarthritis Cartilage. (2019) 27:230–9. doi: 10.1016/j.joca.2018.09.016
226. Knazovicky D, Helgeson ES, Case B, Thomson A, Gruen ME, Maixner W, et al. X. Replicate effects and test-retest reliability of quantitative sensory threshold testing in dogs with and without chronic pain. Vet Anaesth Analg. (2017) 44:615–24. doi: 10.1016/j.vaa.2016.08.008
227. Uilenreef J, van der Staay FJ, Meijer E. A monosodium iodoacetate osteoarthritis lameness model in growing pigs. Animals (Basel). (2019) 9:405. doi: 10.3390/ani9070405
228. Wilkes D, Li G, Angeles CF, Patterson JT, Huang LY. A large animal neuropathic pain model in sheep: a strategy for improving the predictability of preclinical models for therapeutic development. J Pain Res. (2012) 5:415–24. doi: 10.2147/JPR.S34977
229. Lascelles BDX, Brown DC, Maixner W, Mogil JS. Spontaneous painful disease in companion animals can facilitate the development of chronic pain therapies for humans. Osteoarthritis Cartilage. (2018) 26:175–83. doi: 10.1016/j.joca.2017.11.011
230. Eiby YA, Wright LL, Kalanjati VP, Miller SM, Bjorkman ST, Keates HL, et al. A pig model of the preterm neonate: anthropometric and physiological characteristics. PLoS ONE. (2013) 8:e68763. doi: 10.1371/journal.pone.0068763
231. Gobina I, Villberg J, Välimaa R, Tynjälä J, Whitehead R, Cosma A, et al. Prevalence of self-reported chronic pain among adolescents: evidence from 42 countries and regions. Eur J Pain. (2019) 23:316–26. doi: 10.1002/ejp.1306
232. Kennedy J, Roll JM, Schraudner T, Murphy S, McPherson S. Prevalence of persistent pain in the U.S. adult population: new data from the 2010 national health interview survey. J Pain. (2014) 15:979–84. doi: 10.1016/j.jpain.2014.05.009
233. Nahin RL. Estimates of pain prevalence and severity in adults: United States, 2012. J Pain. (2015) 16:769–80. doi: 10.1016/j.jpain.2015.05.002
234. Pitcher MH, Von Korff M, Bushnell MC, Porter L. Prevalence and profile of high-impact chronic pain in the United States. J Pain. (2019) 20:146–60. doi: 10.1016/j.jpain.2018.07.006
235. Victoria NC, Murphy AZ. Exposure to early life pain: long term consequences and contributing mechanisms. Curr Opin Behav Sci. (2016) 7:61–8. doi: 10.1016/j.cobeha.2015.11.015
Keywords: neonatal pain, early life, consequences, neurobiology, mechanisms, animal models, chronic pain
Citation: Williams MD and Lascelles BDX (2020) Early Neonatal Pain—A Review of Clinical and Experimental Implications on Painful Conditions Later in Life. Front. Pediatr. 8:30. doi: 10.3389/fped.2020.00030
Received: 18 November 2019; Accepted: 20 January 2020;
Published: 07 February 2020.
Edited by:
Udo Rolle, University Hospital Frankfurt, GermanyReviewed by:
Burak Tander, Acibadem University, TurkeyCopyright © 2020 Williams and Lascelles. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: B. Duncan X. Lascelles, ZHhsYXNjZWxAbmNzdS5lZHU=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.