
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Oncol., 31 March 2020
Sec. Molecular and Cellular Oncology
Volume 10 - 2020 | https://doi.org/10.3389/fonc.2020.00413
This article is part of the Research TopicNeovascularization, Angiogenesis and Vasculogenic Mimicry in CancerView all 25 articles
 Erik Lizárraga-Verdugo1
Erik Lizárraga-Verdugo1 Melisa Avendaño-Félix1
Melisa Avendaño-Félix1 Mercedes Bermúdez1
Mercedes Bermúdez1 Rosalio Ramos-Payán1
Rosalio Ramos-Payán1 Carlos Pérez-Plasencia2
Carlos Pérez-Plasencia2 Maribel Aguilar-Medina1*
Maribel Aguilar-Medina1*Cancer stem cells (CSCs) are able to promote initiation, survival and maintenance of tumor growth and have been involved in gastrointestinal cancers (GICs) such as esophageal, gastric and colorectal. It is well known that blood supply facilitates cancer progression, recurrence, and metastasis. In this regard, tumor-induced angiogenesis begins with expression of pro-angiogenic molecules such as vascular endothelial growth factor (VEGF), which in turn lead to neovascularization and thus to tumor growth. Another pattern of blood supply is called vasculogenic mimicry (VM). It is a reminiscent of the embryonic vascular network and is carried out by CSCs that have the capability of transdifferentiate and form vascular-tube structures in absence of endothelial cells. In this review, we discuss the role of CSCs in angiogenesis and VM, since these mechanisms represent a source of tumor nutrition, oxygenation, metabolic interchange and facilitate metastasis. Identification of CSCs mechanisms involved in angiogenesis and VM could help to address therapeutics for GICs.
Gastrointestinal cancers (GICs) are among the most common malignancies worldwide that mainly include gastric, esophageal and colorectal cancers (1). Treatments for GICs commonly are chemotherapy, radiotherapy, surgery and most recently anti-angiogenic therapy. However, the efficiency of these treatments depends on multiple factors such as cancer staging and resistance to treatment and relapse, which are related to Cancer Stem Cells (CSCs) (2).
In normal and tumoral tissues, vasculature supply the nutrients and oxygen required to maintain homeostasis. Blood vessel formation in the embryo occurs by vasculogenesis, a process that involve de novo production of endothelial cells (ECs) (3). On the other hand, the process through which new blood vessels are formed by sprouting and splitting from pre-existing ones is called angiogenesis (4), which is an important cancer hallmark.
Self-renewal of CSCs and initiation of tumor is accompanied by the promotion of angiogenesis, through the secretion of proangiogenic factors such as Vascular Endothelial Growth Factor (VEGF) (5). However, angiogenesis is not the unique source of nutrients and oxygen for tumors (6), given that CSCs are able to transdifferentiate into endothelial-like cells enhancing neovascularization (7). This process, called vasculogenic mimicry (VM), is present in different types of cancers and is responsible of providing a sufficient blood supply to tumor tissues (8). Interestingly, CD133 positive glioma cells express that express VEGF are able to increase vascular density (9) and higher recruitment of endothelial progenitor cells (EPCs) is observed in tumors enriched with CSCs (10).
The aim of this review is to compile recent knowledge of gastrointestinal CSCs and their participation in VM and angiogenesis in order to understand the underlying mechanisms that lead to the development of more effective therapies.
Tumors are characterized by cell heterogeneity, according to CSCs theory, which hypothesizes that tumors are driven by a small cell subpopulation with stem cell properties, such as self-renewal and differentiation capacity (11, 12). Also, CSCs promote tumor initiation, growth and proliferation, leading to aberrant growth and slow cycle cell replacement, making them resistant to therapies (13) and are able to move outside of the primary site and metastasize (14).
CSCs were first isolated (CD34+CD38−) from Acute Myeloid Leukemia (AML) patient samples in late 90s. This small population, was capable to transfer AML from human patients to NOD/SCID mice (15). Since then, surface markers have been used to identify and isolate CSCs in several types of cancers, for instance, CD24, CD44, CD90, CD133, and CD166 for Gastrointestinal CSC, and it was demonstrated that they are generally tissue specific (Table 1) (2).
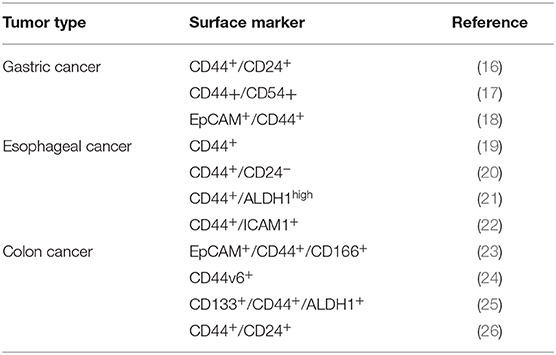
Table 1. Surface markers of gastrointestinal cancers stem cells.
Regarding to Esophageal Cancer Stem Cells (ECSCs), they were first isolated from Esophageal Squamous carcinoma cell line (ESCC) using colony morphology criteria (27). Nevertheless, isolation of ECSCs now is performed using CD44 and ALDH1 (19, 28).
CD44 was the first marker used to identify Gastric Cancer (GC) Stem Cells (GCSCs) (29). Moreover, the embryonic markers OCT-4, SOX2, NANOG and the surface maker CD133/Prom1 are highly expressed in GCSCs (30). Interestingly, CD44+/CD24+ GCSCs subpopulation has shown stem cell properties in vivo and in vitro (16). Also, EpCAM+/CD44+ phenotype present stem cell characteristics in GC tissues (18) Besides, isolated CD44+/CD54+ GCSCs from tumors and peripheral blood, are able to generate tumors both in vitro and in vivo (17). However, other molecules, such as, CD90, CD71, ABCB1, ABCG2, CD133, ALDH1, and Lgr5 are also considered as potential markers to GCSCs isolation (31–35).
Finally, Colorectal Cancer (CRC) Stem Cells (CRCSCs) were first isolated by CD133 expression, showing tumorigenic capabilities in mice (25, 36). Nevertheless, molecules such as EpCAM+/CD44+/CD166+, ALDH+, EphB2+, LGR5+, and CD44v6+ are commonly used to CRCSCs isolation from cell lines (23, 24, 37–39), despite these markers are shared with normal mesenchymal stem cells (MSCs). In this regard, it has been recently reported that Dclk1 discriminates between cancer and normal stem cells in the intestine (40).
Vascular niches are key for maintaining the stem phenotype, such as, self-renewal, undifferentiated state and dormancy in normal stem cells (41). In cancer context, neo-vascularization plays an important role during carcinogenesis and metastasis. This process was first described by Scherer in glioblastoma, where the cancer cells growth is possible by the proximity of surrounded blood vessels, now called “cancer vascular niche” (42). Normal stem cells and CSCs primordially growth in vascular niches, due to a perivascular microenvironment. However, cancer vascular niche is rich in abnormal blood vessels, connected and organized with each other in a different pattern from normal vessels (43, 44). These abnormalities are induced by hypoxia, low pH and high interstitial hostile fluid pressure, making a selection of hostile cells that can escape from the tumor through aberrant blood vessels to metastasize (45). Angiogenesis within the tumor mass harbors a variety of host-derived cells, regulated by monocytes Tie-2 expression, fibroblasts, ECs, as well as, innate and adaptive immune cells (46, 47).
Angiogenesis can be divided in two types: sprouting and intussusceptive (48–50). In the first one, ECs proliferate and sprout toward an angiogenic stimulator (e.g., VEGF), forming flat structures called filopodia, producing proteolytic enzymes to enhance angiogenic process (51). On the other hand, intussusceptive angiogenesis is independent of ECs, where an existing vessel is divided into two new vessels only by cellular reorganization (52). Interestingly, neovascularization is an important process to support tumor growth and metastasis; usually, tumors reach a size of ~2 mm in diameter when not fed by neovascularization (53). In this regard, CSCs are able to modify tumoral microenvironment by expressing angiogenic factors in order to enhance tumor neovascularization, contributing finally in their maintenance and proliferation (5).
Positive cells to Placental growth factor (PLGF), appear to be CSCs in esophageal cancer and have the capability to release PLGF, promoting cancer metastasis by the activation of MMP9 (54). Besides, CSCs that express PLGF are important due to the promotion (55) or inhibition of tumor angiogenesis depending on its interaction with VEGF (56).
Bone marrow mesenchymal stem cells (BM-MSCs) are implicated in the promotion of tumor angiogenesis in gastric cancer (GC) since SGC-7901 cells in both, in vitro and in vivo models, increases VEGF release from tumor cells by the activation ERK1/2 and p38 MAPK pathways, resulting in angiogenesis promotion (57). Moreover, gastric cancer-derived MSCs (GC-MSCs) are also able to promote angiogenesis when interact with BGC-823 and MKN-28 GC cell lines, inducing overexpression of pro-angiogenic factors, such as, VEGF, MIP-2, TGF-β1, IL-6, and IL-8 favoring tube formation (58).
Recently, the Leucine-rich repeat and immunoglobulin-like domain-containing Nogo receptor-interacting protein 2 (LINGO2) a novel gastric cancer stem cell-related marker has been associated with cancer progression (59). In this regard, gastric tumor tissues overexpressing LINGO2 shows elevated expression of the angiogenic marker pVEGFR2 and a blood vessel marker CD34, meanwhile the silencing of LINGO2 in Human Umbilical Vein Endothelial Cells (HUVEC) cells results in inhibition of tube formation, suggesting the involvement of positive-LINGO2 CSCs in angiogenesis (59).
CRCSCs are able to initiate vascularization via pericytes by growth promotion (5, 60). Thus, lack of pericytes recruitment impacts negatively in tumor size owing to poor vascular structure (61). This is also correlated to worst prognosis, due to leaky vessels that produces elevated local pressure, enhancing progression and metastasis. Nevertheless, higher vascular density has been associated with recurrence, metastasis and patient mortality (5, 62).
Co-cultivation of CRCSCs and SW620 cells enhances its stemness properties. Also, transplantation of SW48 and MSCs support angiogenesis in vivo (63). Additionally, conditioned media (CM) from SW480 cells pre-treated with CRCSCs CM enhances HUVEC tube formation and higher levels of VEGFA expression (63). Besides, BM-MSCs are able to induce angiogenesis, when treated with IFN-γ and TNF-α, by VEGF expression via the HIF-1α signaling pathway (64), meanwhile, IL-8 allows tumor angiogenesis (65).
Participation of CRCSCs in tumor neovascularization has been demonstrated in tumor tissues by CD31/CD133/Lgr5 co-expression (10). Besides, CRC cell lines HCT116 and HT29 spheroid-derived cells are able to co-act with endothelial progenitor cells (EPCs) in order to promote migration and tube formation by secreting VEGF. Meanwhile, EPCs also increases tumorigenesis of CRC cells through angiogenesis (10).
Little is known about cellular and molecular mechanistic features of CSCs roles in angiogenesis (Figure 1). For instance, Bone Morphogenic Protein 4 (BMP-4) plays a crucial role in angiogenesis by mediating vascular integrity. Besides, VEGF suppression is strongly regulated through BMP-9/ALK1. Conversely, TGFβ1/ALK5 pathway enhances angiogenesis by VEGF expression (66), being a critical signaling molecule for angiogenesis in CSCs (67). Moreover, VEGF-A/NRP-1 interaction promotes stemness properties in breast cancer (BC) cell lines by activation of Wnt/β-catenin pathway, since its inhibition relies in the attenuation of HUVEC-tube formation induced by co-culturing with extracts from Breast Cancer Stem Cells (BCSCs) (60). Moreover, glioblastoma stem-like cells (GSCs) produce VEGF-A, which is secreted in extracellular vesicles promoting permeability and angiogenesis in brain (68). Additionally, angiogenesis promotion can be stimulated by GSC-derived exosomes (GSC-EXs) trough miR-21/VEGF/VEGFR2 axis (69).
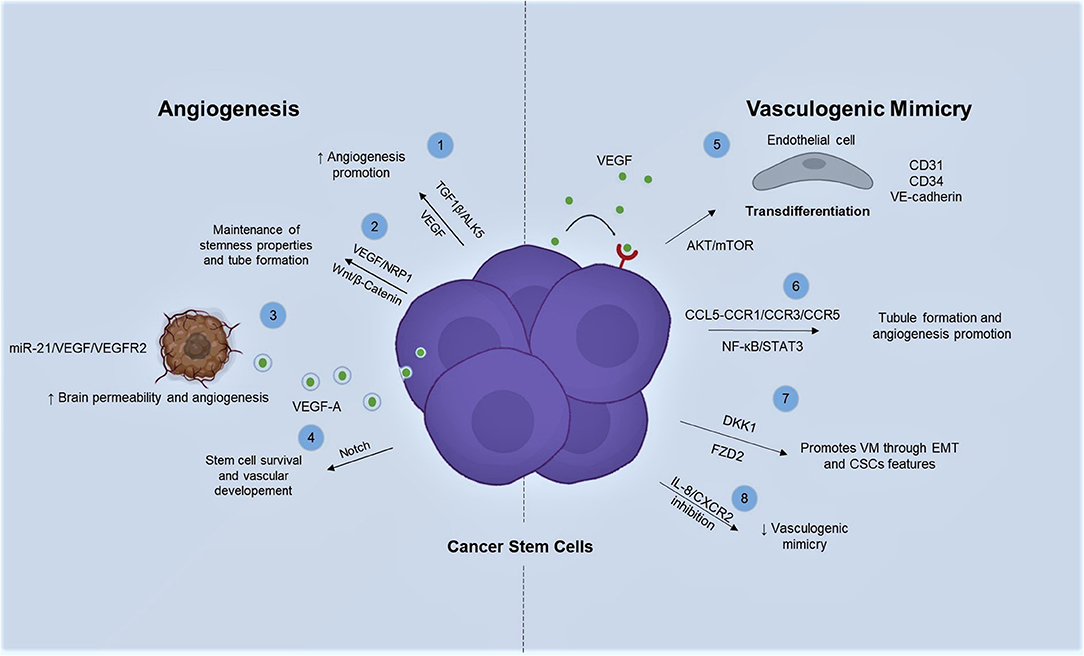
Figure 1. Angiogenic and vasculogenic mimicry promotion by CSCs is mainly triggered by VEGF among several types of cancer. There are different signaling pathways acting in order to promote and sustain neovascularization. 1. Angiogenesis promotion is leaded by TGFβ/ALK5 via VEGF expression in CSCs. 2. Wnt/β-catenin is activated by the interaction of VEGF-A/NRP-1 promoting tube formation. 3. CSCs are able to release VGEF-A by exosomes which in turn stimulates angiogenesis by miR-21/VEGF/VEGFR2 activation. 4. Notch signaling conserves stemness and vasculogenic markers in glioblastoma. 5. VEGFR2 through AKT/mTOR signaling pathway regulates transdifferentiation from poorly differentiated CRC cells into highly expressing CD31, CD34, and VE-cadherin ECs. 6. NF-κB/STAT3 pathway promotes tubule formation and angiogenesis on cancer stem-like cells via CCL5-CCR1/CCR3/CCR5. 7. VM can be influenced by DKK1 by EMT and CSCs behavior. 8. While FZD2 receptor can drive to EMT, thus enhancing stemness properties and VM capabilities.
Notch signaling pathway is also required for stem cell survival and vascular development and it is a crucial angiogenesis stimulator (70). Interestingly, inhibition of self-renewal capabilities and angiogenesis are orchestrated by Notch signaling repression in GSCs, as well as, reduction of vasculogenic markers, such as, CD105, CD31 and von Willebrand factor (vWF) (71).
The generation of vascular channels (VC) without ECs or fibroblasts was first identified in aggressive and metastatic melanoma in 1999, and was termed vasculogenic mimicry (6). In this specific case, the relationship between aggressive melanoma cells that co-expressed Vimentin and epithelial (keratin 8,18) intermediate filaments was particularly interesting, since these cells, where able to be aligned along the external walls of microvascular channels conducing red blood cells, without ECs (72).
Channels formed by VM are composed of a basement membrane and tumor cells that facilitate microcirculation plasma and blood supply from host normal vessels (73). VM can be classified in classical patterns in matrix type (6) and the tubular type (74). Besides, it has been described that VM is composed by matrix proteins such as Laminin, Heparan sulfate proteoglycan, and Collagens IV and VI (75).
VC network may be an independent angiogenesis mechanism for blood source, since angiogenesis inhibitors induce extracellular matrix-rich tubular network formation in vitro and are not able to suppress VM in several types of cancers, showing that VM works as an alternative mechanism for blood cells supply (76). Besides, VM is associated with tumor size, short overall survival (OS), high tumor grade, clinical staging, invasion and metastasis (77–79).
Interestingly, tumor cells associated to VM structures acquire an undifferentiated phenotype as well as ECs characteristics (80). Nowadays, CSCs have been involved in VC formation in cancer (81–87). For instance, in salivary adenoid cystic carcinoma (ACC) specimens CD133 is positively associated with VM formation. Besides, CD133+ ACC CSCs and xenograft tumors of nude mice injected with these cells show overexpression of VE-Cadherin and VM mediators (MMP-2, MMP-9) (86). Furthermore, an holoclone CD133+ isolated from MDA-MB-231 form VM and display MMP-2 and MMP-9 expression (87). In addition, VEGF-silenced cells, attenuate growth and promotes VM as adaptation mechanism associated to HIF-1α expression. Furthermore, enrichment of CD133+/CD271+ Melanoma CSCs is found in the perivascular niche in vivo (81).
It has been shown that epithelial–mesenchymal transition (EMT) cells present stem phenotype, showing a remarkable relationship between EMT and CSCs (88). For instances, Ginseng extract showed a negative effect on EMT, as well as, VM in ESCC lines (89). Besides, recombinant Endostatin (rh-Endo) protein combined with radiotherapy downregulates EMT characteristics and VC formation in ESCC through inactivation of AKT/GSK-3β signaling pathway (90).
In Gastric adenocarcinoma tissues, a positive relationship between CD133/Lgr5 expression and VC formations, microvessel density, tumor grade, lymph node metastasis and TNM staging has been shown (85). In the case of CRC, the upregulation of ZEB1 results in epithelial phenotype restoration, while, its silencing results in VM inhibition and VE-Cadherin and Flk-1 downregulation in HCT116 cell line (91).
CSCs and VM are involved in cell plasticity, which is the capability of an aberrant population to ECs transdifferentiation (Figure 1) (92). VEGF receptors regulate expression of specific marker for ECs, such as VE-Cadherin (93). In this regard, it has been described that primary and established sarcoma cell lines in contact with post-surgery fluids from Giant cell tumors of bone patients can enrich CD44/CD117 cell population and AKT/mTOR pathway activation. Moreover, it has been proved that prolonged stimulation results in transdifferentiation of tubule-like structures that express endothelial markers, such as, VE-Cadherin and CD31 (94). Additionally, CSCs switch on NF-κB and STAT3 signal pathways via CCL5-CCR1/CCR3/CCR5, stimulating endothelial differentiation and tubule formation (95).
It has been demonstrated that DKK1 enhances VM formation via EMT by developing CSC characteristics in not small cells lung carcinoma (NSCLC) (96). Besides, the Wnt signaling receptor FZD2 drives EMT process, enhancing stem-like properties and VM capacity in HCT116 cells (97). Interestingly, inhibition of IL-8/CXCR2 signaling by Transgelin results in suppression of VM with increased IL-8 levels due to IL-8 uptake inhibition in breast cancer stem cells (BCSCs) (98).
In CRC, the poorly differentiated cell line HCT116 expresses endothelial markers and form tube-like structure in vitro after endothelial-conditioned medium co-culture. In addition, under hypoxic conditions cells exhibit higher levels of VEGFR2/VEGFA, as well as, CD31, CD34 and VE-Cadherin overexpression (99).
Little is known about the role of CSCs promoting angiogenesis and VM. It has been shown that abnormal blood vessels are capable to obstruct immune response to the tumor, as wells as, the transportation and distribution of oxygen and chemotherapeutics. This hostile tumor microenvironment can also lead to selection of cells resistant to radiotherapy and chemotherapy (43). Altogether might suggest that anti-angiogenic drugs often induce tumor hypoxia, allowing CSCs to survive and propagate, thus driving tumor progression.
Nevertheless, some inhibitors of VM are potential molecules to use in therapy of different types of cancers, such as LCS1269 that is capable of overcoming multidrug resistance for DNA-damaging agents in melanoma by VM inhibition (100). In addition, Hinokitiol, a tropolone-associated natural compound, has an important effect over EGFR expression and VM in BCSCs through proteasome-mediated EGFR degradation (101).
Molecules and signal pathways involved in angiogenesis and VM supported by CSCs are novel targets of cancer therapeutics. Nevertheless, information of GICs therapeutics in this matter is limited. Has been described that anti-CD133 has a great potential in treating CRC (96). Besides, targeting signaling pathways is possible, for instance, BBI-608 drug targeting STAT3 could be used for advanced CRC resistant to standard therapeutics or in mixture with Paclitaxel for advanced GC (2, 97). Moreover, Ginsenoside Rg3, a derived from ginseng, represses growth cells and CSCs properties in CRC cells, as well as, inhibits angiogenesis-related genes, suppressing vascularization in xenograft tumors (98).
Several authors suggest that interfering on growth and survival of tumoral ECs can be enough to inhibiting angiogenesis and CSCs self-renewal (99). In this regard, VEGF secreted by cancer cells is a well-recognized therapeutic target and several angiogenic inhibitors have been developed with the capability of also suppress self-renewal of CSCs leading to reduced tumor growth. It has been shown that, Bevacizumab expands survival time by targeting the perivascular niche by the inhibition of VEGF (102). Additionally, bevacizumab reduces metastatic niche formation in rectal carcinoma patients (103) and combined with an anti hepatoma-derived growth factor antibody prevents tumor relapse and progression in NSCLC by impairing CSCs (104). Conversely, the administration of Bevacizumab combined to Sunitinib (VEGF inhibitor) induces tumor hypoxia in BC cell lines resulting in the augment of CSCs population (105).
Recently, emerging evidence shows that tumors are heterogeneous, being constituted by multiple subpopulations such as CSCs that share self-renewal and differentiation characteristics with normal stem cells. Also, they are able to express specific surface markers that depend on the organ of origin. For instance, CD44, ALDH1, EpCAM, and Lrg5 are characteristics markers of gastrointestinal CSCs, in EC, GC, and CRC. Besides, vascular niches are important for maintaining tumor progression, since CSCs prefer a perivascular microenvironment, rich in blood vessels that often have an abnormal structure and is supported by hostile conditions such as, hypoxia, which in turn, enhances selection of more aggressive cells, able to invade and metastasize. In this regard, CSCs can be transdifferentiated into endothelial-like cells and pericytes, important lineages for maintenance of cancer vascular niche.
Some signaling pathways have been implicated in angiogenesis and VM. The most important molecules and pathways are VEGF/VEGFR2, Notch, BMP9/ALK1, PI3K/AKT/mTOR, NF-κB, and STAT3, that regulate different pivotal processes involved in angiogenesis promotion, such as permeability, endothelial and tubule-like transdifferentiation and promotion of endothelial markers expression, stem cell survival and vascular development.
Clinical relevance of angiogenesis in GICs is remarkable as poor pericyte coverage is correlated with worst prognosis due to leaky vessels that produce elevated local pressure and enhances progression and metastasis. Besides, a higher vascular density in the invasion front has been associated with recurrence, metastasis and patient mortality in CRC. Importantly, Dclk1 can discriminates between cancer and normal stem cells in the intestine.
CSCs are implicated in VM in different cancers, such as ACC, breast cancer and melanoma. In addition, there is a remarkable relationship between EMT and CSCs, due to EMT cells acquired stem phenotype. Importantly, GICs show that the use of drugs, certain proteins or radiotherapy that affect the EMT leads to inhibition of VM. Finally, clinical relevance of VM relies on its association with tumor size, short OS, high tumor grade, clinical staging, invasion and metastasis.
On this front, several drugs have been tested, for instance, Bevacizumab is able to expand survival time by targeting the perivascular niche by the inhibition of VEGF with effect on angiogenesis However, more studies are necessary in order to elucidate CSCs participation on VM and angiogenesis since this could help to address therapeutics for GICs.
EL-V, MA-F, MB, and MA-M conceived and designed the content of this review. EL-V, MA-F, MB, and RR-P wrote the paper. CP-P and MA-M contributed to the final version of the manuscript.
Consejo Nacional de Ciencia y Tecnología CONACYT, Mexico (Grant 290311).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The handling Editor declared a past co-authorship with several of the authors CP-P, RR-P.
The authors acknowledge CONACyT for MA-F (575985) and EL-V (304939) fellowships.
1. Grierson P, Lim K-H, Amin M. Immunotherapy in gastrointestinal cancers. J Gastrointest Oncol. (2017) 8:474–84. doi: 10.21037/jgo.2017.05.01
2. Taniguchi H, Moriya C, Igarashi H, Saitoh A, Yamamoto H, Adachi Y, et al. Cancer stem cells in human gastrointestinal cancer. Cancer Sci. (2016) 107:1556–62. doi: 10.1111/cas.13069
3. Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. (2011) 144:646–74. doi: 10.1016/j.cell.2011.02.013
4. Bergers G, Benjamin LE. Tumorigenesis and the angiogenic switch. Nat Rev Cancer. (2003) 3:401–10. doi: 10.1038/nrc1093
5. Garza Treviño EN, González PD, Valencia Salgado CI, Martinez Garza A. Effects of pericytes and colon cancer stem cells in the tumor microenvironment. Cancer Cell Int. (2019) 19:173. doi: 10.1186/s12935-019-0888-9
6. Maniotis AJ, Folberg R, Hess A, Seftor EA, Gardner LM, Pe'er J, et al. Vascular channel formation by human melanoma cells in vivo and in vitro: vasculogenic mimicry. Am J Pathol. (1999) 155:739–52. doi: 10.1016/S0002-9440(10)65173-5
7. Ricci-Vitiani L, Pallini R, Biffoni M, Todaro M, Invernici G, Cenci T, et al. Tumour vascularization via endothelial differentiation of glioblastoma stem-like cells. Nature. (2010) 468:824–8. doi: 10.1038/nature09557
8. Seftor RE, Seftor EA, Koshikawa N, Meltzer PS, Gardner LM, Bilban M, et al. Cooperative interactions of laminin 5 gamma2 chain, matrix metalloproteinase-2, and membrane type-1-matrix/metalloproteinase are required for mimicry of embryonic vasculogenesis by aggressive melanoma. Cancer Res. (2001) 61:6322–7. Available online at: https://cancerres.aacrjournals.org/content/61/17/6322
9. Bao S, Wu Q, Sathornsumetee S, Hao Y, Li Z, Hjelmeland AB, et al. Stem cell-like glioma cells promote tumor angiogenesis through vascular endothelial growth factor. Cancer Res. (2006) 66:7843–8. doi: 10.1158/0008-5472.CAN-06-1010
10. Wei B, Han XY, Qi CL, Zhang S, Zheng ZH, Huang Y, et al. Coaction of spheroid-derived stem-like cells and endothelial progenitor cells promotes development of colon cancer. PLoS ONE. (2012) 7:e39069. doi: 10.1371/journal.pone.0039069
12. Humphries A, Wright NA. Colonic crypt organization and tumorigenesis. Nat Rev Cancer. (2008) 8:415–24. doi: 10.1038/nrc2392
13. Kreso A, Dick JE. Evolution of the cancer stem cell model. Cell Stem Cell. (2014) 14:275–91. doi: 10.1016/j.stem.2014.02.006
14. Nandy SB, Lakshmanaswamy R. Cancer Stem Cells and Metastasis. Progr Mol Biol Transl Sci. (2017) 151:137–76. doi: 10.1016/bs.pmbts.2017.07.007
15. Bonnet D, Dick JE. Human acute myeloid leukemia is organized as a hierarchy that originates from a primitive hematopoietic cell. Nat Med. (1997) 3:730–7. doi: 10.1038/nm0797-730
16. Zhang C, Li C, He F, Cai Y, Yang H. Identification of CD44+CD24+ gastric cancer stem cells. J Cancer Res Clin Oncol. (2011) 137:1679–86. doi: 10.1007/s00432-011-1038-5
17. Chen T, Yang K, Yu J, Meng W, Yuan D, Bi F, et al. Identification and expansion of cancer stem cells in tumor tissues and peripheral blood derived from gastric adenocarcinoma patients. Cell Res. (2012) 22:248–58. doi: 10.1038/cr.2011.109
18. Han ME, Jeon TY, Hwang SH, Lee YS, Kim HJ, Shim HE, et al. Cancer spheres from gastric cancer patients provide an ideal model system for cancer stem cell research. Cell Mol Life Sci. (2011) 68:3589–605. doi: 10.1007/s00018-011-0672-z
19. Zhao J-S, Li W-J, Ge D, Zhang P-J, Li J-J, Lu C-L, et al. Tumor initiating cells in esophageal squamous cell carcinomas express high levels of CD44. PLoS ONE. (2011) 6:e21419. doi: 10.1371/journal.pone.0021419
20. Smit JK, Faber H, Niemantsverdriet M, Baanstra M, Bussink J, Hollema H, et al. Prediction of response to radiotherapy in the treatment of esophageal cancer using stem cell markers. Radiother Oncol. (2013) 107:434–41. doi: 10.1016/j.radonc.2013.03.027
21. Almanaa TN, Geusz ME, Jamasbi RJ. A new method for identifying stem-like cells in esophageal cancer cell lines. J Cancer. (2013) 4:536–48. doi: 10.7150/jca.6477
22. Tsai ST, Wang PJ, Liou NJ, Lin PS, Chen CH, Chang WC. ICAM1 is a potential cancer stem cell marker of esophageal squamous cell carcinoma. PLoS ONE. (2015) 10:e0142834. doi: 10.1371/journal.pone.0142834
23. Dalerba P, Dylla SJ, Park IK, Liu R, Wang X, Cho RW, et al. Phenotypic characterization of human colorectal cancer stem cells. Proc Natl Acad Sci USA. (2007) 104:10158–63. doi: 10.1073/pnas.0703478104
24. Todaro M, Gaggianesi M, Catalano V, Benfante A, Iovino F, Biffoni M, et al. CD44v6 is a marker of constitutive and reprogrammed cancer stem cells driving colon cancer metastasis. Cell Stem Cell. (2014) 14:342–56. doi: 10.1016/j.stem.2014.01.009
25. Ricci-Vitiani L, Lombardi DG, Pilozzi E, Biffoni M, Todaro M, Peschle C, et al. Identification and expansion of human colon-cancer-initiating cells. Nature. (2007) 445:111–5. doi: 10.1038/nature05384
26. Yeung TM, Gandhi SC, Wilding JL, Muschel R, Bodmer WF. Cancer stem cells from colorectal cancer-derivedstem-like cancer cells in melanoma cell lines. Proc Natl Acad Sci USA. (2010) 107:3722–7. doi: 10.1073/pnas.0915135107
27. Ban S, Ishikawa K-i, Kawai S, Koyama-Saegusa K, Ishikawa A, Shimada Y, et al. Potential in a single cancer cell to produce heterogeneous morphology, radiosensitivity and gene expression. J Radiat Res. (2005) 46:43–50. doi: 10.1269/jrr.46.43
28. Song S, Xie M, Scott AW, Jin J, Ma L, Dong X, et al. A novel YAP1 inhibitor targets CSC-enriched radiation-resistant cells and exerts strong antitumor activity in esophageal adenocarcinoma. Mol Cancer Ther. (2018) 17:443–54. doi: 10.1158/1535-7163.MCT-17-0560
29. Takaishi S, Okumura T, Tu S, Wang SS, Shibata W, Vigneshwaran R, et al. Identification of gastric cancer stem cells using the cell surface marker CD44. Stem Cells. (2009) 27:1006–20. doi: 10.1002/stem.30
30. Singh SR. Gastric cancer stem cells: a novel therapeutic target. Cancer Lett. (2013) 338:110–9. doi: 10.1016/j.canlet.2013.03.035
31. Katsuno Y, Ehata S, Yashiro M, Yanagihara K, Hirakawa K, Miyazono K. Coordinated expression of REG4 and aldehyde dehydrogenase 1 regulating tumourigenic capacity of diffuse-type gastric carcinoma-initiating cells is inhibited by TGF-beta. J Pathol. (2012) 228:391–404. doi: 10.1002/path.4020
32. Jiang J, Zhang Y, Chuai S, Wang Z, Zheng D, Xu F, et al. Trastuzumab (herceptin) targets gastric cancer stem cells characterized by CD90 phenotype. Oncogene. (2012) 31:671–82. doi: 10.1038/onc.2011.282
33. Ohkuma M, Haraguchi N, Ishii H, Mimori K, Tanaka F, Kim HM, et al. Absence of CD71 transferrin receptor characterizes human gastric adenosquamous carcinoma stem cells. Ann Surg Oncol. (2012) 19:1357–64. doi: 10.1245/s10434-011-1739-7
34. Jiang Y, He Y, Li H, Li HN, Zhang L, Hu W, et al. Expressions of putative cancer stem cell markers ABCB1, ABCG2, and CD133 are correlated with the degree of differentiation of gastric cancer. Gastric Cancer. (2012) 15:440–50. doi: 10.1007/s10120-012-0140-y
35. Barker N, Huch M, Kujala P, van de Wetering M, Snippert HJ, van Es JH, et al. Lgr5 (+ve) stem cells drive self-renewal in the stomach and build long-lived gastric units in vitro. Cell Stem Cell. (2010) 6:25–36. doi: 10.1016/j.stem.2009.11.013
36. O'Brien CA, Pollett A, Gallinger S, Dick JE. A human colon cancer cell capable of initiating tumour growth in immunodeficient mice. Nature. (2007) 445:106–10. doi: 10.1038/nature05372
37. Huang EH, Hynes MJ, Zhang T, Ginestier C, Dontu G, Appelman H, et al. Aldehyde dehydrogenase 1 is a marker for normal and malignant human colonic stem cells (SC) and tracks SC overpopulation during colon tumorigenesis. Cancer Res. (2009) 69:3382–9. doi: 10.1158/0008-5472.CAN-08-4418
38. Kemper K, Versloot M, Cameron K, Colak S, de Sousa e Melo F, de Jong JH, et al. Mutations in the Ras-Raf Axis underlie the prognostic value of CD133 in colorectal cancer. Clin Cancer Res. (2012) 18:3132–41. doi: 10.1158/1078-0432.CCR-11-3066
39. Jung P, Sato T, Merlos-Suárez A, Barriga FM, Iglesias M, Rossell D, et al. Isolation and in vitro expansion of human colonic stem cells. Nat Med. (2011) 17:1225. doi: 10.1038/nm.2470
40. Nakanishi Y, Seno H, Fukuoka A, Ueo T, Yamaga Y, Maruno T, et al. Dclk1 distinguishes between tumor and normal stem cells in the intestine. Nat Genet. (2012) 45:98. doi: 10.1038/ng.2481
41. Vissapragada R, Contreras MA, da Silva CG, Kumar VA, Ochoa A, Vasudevan A, et al. Bidirectional crosstalk between periventricular endothelial cells and neural progenitor cells promotes the formation of a neurovascular unit. Brain Res. (2014) 1565:8–17. doi: 10.1016/j.brainres.2014.03.018
42. Scherer HJ. The forms of growth in gliomas and their practical significance. Brain. (1940) 63:1–35. doi: 10.1093/brain/63.1.1
43. Carmeliet P, Jain RK. Principles and mechanisms of vessel normalization for cancer and other angiogenic diseases. Nat Rev Drug Discov. (2011) 10:417–27. doi: 10.1038/nrd3455
44. Ping YF, Zhang X, Bian XW. Cancer stem cells and their vascular niche: Do they benefit from each other? Cancer Lett. (2016) 380:561–7. doi: 10.1016/j.canlet.2015.05.010
45. Jain RK. Normalization of tumor vasculature: an emerging concept in antiangiogenic therapy. Science. (2005) 307:58–62. doi: 10.1126/science.1104819
46. De Palma M, Venneri MA, Galli R, Sergi Sergi L, Politi LS, Sampaolesi M, et al. Tie2 identifies a hematopoietic lineage of proangiogenic monocytes required for tumor vessel formation and a mesenchymal population of pericyte progenitors. Cancer Cell. (2005) 8:211–26. doi: 10.1016/j.ccr.2005.08.002
47. Chouaib S, Kieda C, Benlalam H, Noman MZ, Mami-Chouaib F, Ruegg C. Endothelial cells as key determinants of the tumor microenvironment: interaction with tumor cells, extracellular matrix and immune killer cells. Crit Rev Immunol. (2010) 30:529–45. doi: 10.1615/CritRevImmunol.v30.i6.30
48. Azad T, Ghahremani M, Yang X. The role of YAP and TAZ in angiogenesis and vascular mimicry. Cells. (2019) 8:407. doi: 10.3390/cells8050407
49. Hillen F, Griffioen AW. Tumour vascularization: sprouting angiogenesis and beyond. Cancer Metastasis Rev. (2007) 26:489–502. doi: 10.1007/s10555-007-9094-7
50. Djonov V, Baum O, Burri PH. Vascular remodeling by intussusceptive angiogenesis. Cell Tissue Res. (2003) 314:107–17. doi: 10.1007/s00441-003-0784-3
51. Lamalice L, Le Boeuf F, Huot J. Endothelial cell migration during angiogenesis. Circ Res. (2007) 100:782–94. doi: 10.1161/01.RES.0000259593.07661.1e
52. Caduff JH, Fischer LC, Burri PH. Scanning electron microscope study of the developing microvasculature in the postnatal rat lung. Anat Rec. (1986) 216:154–64. doi: 10.1002/ar.1092160207
53. Zetter BR. Angiogenesis and tumor metastasis. Ann Rev Med. (1998) 49:407–24. doi: 10.1146/annurev.med.49.1.407
54. Chen Y, Jiang T, Mao A, Xu J. Esophageal cancer stem cells express PLGF to increase cancer invasion through MMP9 activation. Tumour Biol. (2014) 35:12749–55. doi: 10.1007/s13277-014-2601-x
55. Carmeliet P, Moons L, Luttun A, Vincenti V, Compernolle V, De Mol M, et al. Synergism between vascular endothelial growth factor and placental growth factor contributes to angiogenesis and plasma extravasation in pathological conditions. Nat Med. (2001) 7:575–83. doi: 10.1038/87904
56. Eriksson A, Cao R, Pawliuk R, Berg SM, Tsang M, Zhou D, et al. Placenta growth factor-1 antagonizes VEGF-induced angiogenesis and tumor growth by the formation of functionally inactive PlGF-1/VEGF heterodimers. Cancer Cell. (2002) 1:99–108. doi: 10.1016/S1535-6108(02)00028-4
57. Zhu W, Huang L, Li Y, Zhang X, Gu J, Yan Y, et al. Exosomes derived from human bone marrow mesenchymal stem cells promote tumor growth in vivo. Cancer Lett. (2012) 315:28–37. doi: 10.1016/j.canlet.2011.10.002
58. Li W, Zhou Y, Yang J, Zhang X, Zhang H, Zhang T, et al. Gastric cancer-derived mesenchymal stem cells prompt gastric cancer progression through secretion of interleukin-8. J Exp Clin Cancer Res. (2015) 34:52. doi: 10.1186/s13046-015-0172-3
59. Jo JH, Park SB, Park S, Lee HS, Kim C, Jung DE, et al. Novel gastric cancer stem cell-related marker LINGO2 is associated with cancer cell phenotype and patient outcome. Int J Mol Sci. (2019) 20:E555. doi: 10.3390/ijms20030555
60. Shangguan W, Fan C, Chen X, Lu R, Liu Y, Li Y, et al. Endothelium originated from colorectal cancer stem cells constitute cancer blood vessels. Cancer Sci. (2017) 108:1357–67. doi: 10.1111/cas.13262
61. Shenoy AK, Jin Y, Luo H, Tang M, Pampo C, Shao R, et al. Epithelial-to-mesenchymal transition confers pericyte properties on cancer cells. J Clin Invest. (2016) 126:4174–86. doi: 10.1172/JCI86623
62. Ribeiro AL, Okamoto OK. Combined effects of pericytes in the tumor microenvironment. Stem Cells Int. (2015) 2015:868475. doi: 10.1155/2015/868475
63. Zhang X, Hu F, Li G, Li G, Yang X, Liu L, et al. Human colorectal cancer-derived mesenchymal stem cells promote colorectal cancer progression through IL-6/JAK2/STAT3 signaling. Cell Death Dis. (2018) 9:25. doi: 10.1038/s41419-017-0176-3
64. Liu Y, Han Z-P, Zhang S-S, Jing Y-Y, Bu X-X, Wang C-Y, et al. Effects of inflammatory factors on mesenchymal stem cells and their role in the promotion of tumor angiogenesis in colon cancer. J Biol Chem. (2011) 286:25007–15. doi: 10.1074/jbc.M110.213108
65. Wang J, Wang Y, Wang S, Cai J, Shi J, Sui X, et al. Bone marrow-derived mesenchymal stem cell-secreted IL-8 promotes the angiogenesis and growth of colorectal cancer. Oncotarget. (2015) 6:42825–37. doi: 10.18632/oncotarget.5739
66. Shao ES, Lin L, Yao Y, Boström KI, Dc W, Bostro KI. Regulated by the activin-like kinase receptors 1 and 5 in endothelial cells expression of vascular endothelial growth factor is coordinately regulated by the activin-like kinase receptors 1 and 5 in endothelial cells. Blood. (2013) 114:2197–206. doi: 10.1182/blood-2009-01-199166
67. Mercurio AM. VEGF/neuropilin signaling in cancer stem cells. Int J Mol Sci. (2019) 20:490. doi: 10.3390/ijms20030490
68. Treps L, Perret R, Edmond S, Ricard D, Gavard J. Glioblastoma stem-like cells secrete the pro-angiogenic VEGF-A factor in extracellular vesicles. J Extracell Vesicles. (2017) 6:1359479. doi: 10.1080/20013078.2017.1359479
69. Sun X, Ma X, Wang J, Zhao Y, Wang Y, Bihl JC, et al. Glioma stem cells-derived exosomes promote the angiogenic ability of endothelial cells through miR-21/VEGF signal. Oncotarget. (2017) 8:36137–48. doi: 10.18632/oncotarget.16661
70. Gridley T. Notch signaling in vascular development and physiology. Development. (2007) 134:2709–18. doi: 10.1242/dev.004184
71. Hovinga KE, Shimizu F, Wang R, Panagiotakos G, Van Der Heijden M, Moayedpardazi H, et al. Inhibition of notch signaling in glioblastoma targets cancer stem cells via an endothelial cell intermediate. Stem Cells. (2010) 28:1019–29. doi: 10.1002/stem.429
72. Folberg R, Hendrix MJ, Maniotis AJ. Vasculogenic mimicry and tumor angiogenesis. Am J Pathol. (2000) 156:361–81. doi: 10.1016/S0002-9440(10)64739-6
73. Sun B, Zhang D, Zhao N, Zhao X. Epithelial-to-endothelial transition and cancer stem cells: two cornerstones of vasculogenic mimicry in malignant tumors. Oncotarget. (2017) 8:30502–10. doi: 10.18632/oncotarget.8461
74. El Hallani S, Boisselier B, Peglion F, Rousseau A, Colin C, Idbaih A, et al. A new alternative mechanism in glioblastoma vascularization: tubular vasculogenic mimicry. Brain. (2010) 133(Pt 4):973–82. doi: 10.1093/brain/awq044
75. Folberg R, Maniotis AJ. Vasculogenic mimicry. Acta Pathol Microbiol Immunol Scand. (2004) 112:508–25. doi: 10.1111/j.1600-0463.2004.apm11207-0810.x
76. van der Schaft DW, Seftor RE, Seftor EA, Hess AR, Gruman LM, Kirschmann DA, et al. Effects of angiogenesis inhibitors on vascular network formation by human endothelial and melanoma cells. J Natl Cancer Inst. (2004) 96:1473–7. doi: 10.1093/jnci/djh267
77. Sun B, Zhang S, Zhao X, Zhang W, Hao X. Vasculogenic mimicry is associated with poor survival in patients with mesothelial sarcomas and alveolar rhabdomyosarcomas. Int J Oncol. (2004) 25:1609–14. doi: 10.3892/ijo.25.6.1609
78. Sun B, Zhang S, Zhang D, Du J, Guo H, Zhao X, et al. Vasculogenic mimicry is associated with high tumor grade, invasion and metastasis, and short survival in patients with hepatocellular carcinoma. Oncol Rep. (2006) 16:693–8. doi: 10.3892/or.16.4.693
79. Zhang D, Sun B, Zhao X, Ma Y, Ji R, Gu Q, et al. Twist1 expression induced by sunitinib accelerates tumor cell vasculogenic mimicry by increasing the population of CD133+ cells in triple-negative breast cancer. Mol Cancer. (2014) 13:207. doi: 10.1186/1476-4598-13-207
80. Hendrix MJ, Seftor EA, Hess AR, Seftor RE. Vasculogenic mimicry and tumour-cell plasticity: lessons from melanoma. Nat Rev Cancer. (2003) 3:411–21. doi: 10.1038/nrc1092
81. Schnegg CI, Yang MH, Ghosh SK, Hsu MY. Induction of vasculogenic mimicry overrides VEGF-a silencing and enriches stem-like cancer cells in melanoma. Cancer Res. (2015) 75:1682–90. doi: 10.1158/0008-5472.CAN-14-1855
82. Zhao X, Sun B. Long noncoding RNA n339260 promotes vasculogenic mimicry and cancer stem cell development in hepatocellular carcinoma. Cancer Sci. (2018) 109:3197–208. doi: 10.1111/cas.13740
83. Liu Y, Mei L, Yu Q, Xu C, Qiu Y, Yang Y, et al. Multifunctional tandem peptide modified paclitaxel-loaded liposomes for the treatment of vasculogenic mimicry and cancer stem cells in malignant glioma. ACS Appl Mater Interfaces. (2015) 7:16792–801. doi: 10.1021/acsami.5b04596
84. Wang H, Huang B, Li BM, Cao KY, Mo CQ, Jiang SJ, et al. ZEB1-mediated vasculogenic mimicry formation associates with epithelial-mesenchymal transition and cancer stem cell phenotypes in prostate cancer. J Cell Mol Med. (2018) 22:3768–3781. doi: 10.1111/jcmm.13637
85. Zhou L, Yu L, Feng ZZ, Gong XM, Cheng ZN, Yao N, et al. Aberrant expression of markers of cancer stem cells in gastric adenocarcinoma and their relationship to vasculogenic mimicry. Asian Pac J Cancer Prev. (2015) 16:4177–83. doi: 10.7314/APJCP.2015.16.10.4177
86. Wang SS, Gao XL, Liu X, Gao SY, Fan YL, Jiang YP, et al. CD133+ cancer stem-like cells promote migration and invasion of salivary adenoid cystic carcinoma by inducing vasculogenic mimicry formation. Oncotarget. (2016) 7:29051–62. doi: 10.18632/oncotarget.8665
87. Liu TJ, Sun BC, Zhao XL, Zhao XM, Sun T, Gu Q, et al. CD133+ cells with cancer stem cell characteristics associates with vasculogenic mimicry in triple-negative breast cancer. Oncogene. (2013) 32:544–53. doi: 10.1038/onc.2012.85
88. Radisky DC, LaBarge MA. Epithelial-mesenchymal transition and the stem cell phenotype. Cell Stem Cell. (2008) 2:511–2. doi: 10.1016/j.stem.2008.05.007
89. Shi Q, Diao Y, Jin F, Ding Z. Anti-metastatic effects of Aidi on human esophageal squamous cell carcinoma by inhibiting epithelial-mesenchymal transition and angiogenesis. Mol Med Rep. (2018) 18:131–8. doi: 10.3892/mmr.2018.8976
90. Chen X, Zhang H, Zhu H, Yang X, Yang Y, Yang Y, et al. Endostatin combined with radiotherapy suppresses vasculogenic mimicry formation through inhibition of epithelial-mesenchymal transition in esophageal cancer. Tumour Biol. (2016) 37:4679–88. doi: 10.1007/s13277-015-4284-3
91. Liu Z, Sun B, Qi L, Li H, Gao J, Leng X. Zinc finger E-box binding homeobox 1 promotes vasculogenic mimicry in colorectal cancer through induction of epithelial-to-mesenchymal transition. Cancer Sci. (2012) 103:813–20. doi: 10.1111/j.1349-7006.2011.02199.x
92. Angelini G, Pisani M, Mobbili G, Marini M, Gasbarri C. Neutral liposomes containing crown ether-lipids as potentialDNA vectors. Biochim Biophys Acta. (2013) 1828:2506–12. doi: 10.1016/j.bbamem.2013.06.003
93. Breier G, Grosser M, Rezaei M. Endothelial cadherins in cancer. Cell Tissue Res. (2014) 355:523–7. doi: 10.1007/s00441-014-1851-7
94. Fazioli F, Colella G, Miceli R, Di Salvatore MG, Gallo M, Boccella S, et al. Post-surgery fluids promote transition of cancer stem cell-to-endothelial and AKT/mTOR activity, contributing to relapse of giant cell tumors of bone. Oncotarget. (2017) 8:85040–53. doi: 10.18632/oncotarget.18783
95. Tang S, Xiang T, Huang S, Zhou J, Wang Z, Xie R, et al. Ovarian cancer stem-like cells differentiate into endothelial cells and participate in tumor angiogenesis through autocrine CCL5 signaling. Cancer Lett. (2016) 376:137–47. doi: 10.1016/j.canlet.2016.03.034
96. Bach P, Abel T, Hoffmann C, Gal Z, Braun G, Voelker I, et al. Specific elimination of CD133+ tumor cells with targeted oncolytic measles virus. Cancer Res. (2013) 73:865–74. doi: 10.1158/0008-5472.CAN-12-2221
97. Chen Z, Han ZC. STAT3: a critical transcription activator in angiogenesis. Med Res Rev. (2008) 28:185–200. doi: 10.1002/med.20101
98. Tang YC, Zhang Y, Zhou J, Zhi Q, Wu MY, Gong FR, et al. Ginsenoside Rg3 targets cancer stem cells and tumor angiogenesis to inhibit colorectal cancer progression in vivo. Int J Oncol. (2018) 52:127–38. doi: 10.3892/ijo.2017.4183
99. Gu JW, Rizzo P, Pannuti A, Golde T, Osborne B, Miele L. Notch signals in the endothelium and cancer “stem-like” cells: opportunities for cancer therapy. Vasc Cell. (2012) 4:7. doi: 10.1186/2045-824X-4-7
100. Vartanian A, Baryshnikova M, Burova O, Afanasyeva D, Misyurin V, Belysmall a CA, et al. Inhibitor of vasculogenic mimicry restores sensitivity of resistant melanoma cells to DNA-damaging agents. Melanoma Res. (2017) 27:8–16. doi: 10.1097/CMR.0000000000000308
101. Tu D-G, Yu Y, Lee C-H, Kuo Y-L, Lu Y-C, Tu C-W, et al. Hinokitiol inhibits vasculogenic mimicry activity of breast cancer stem/progenitor cells through proteasome-mediated degradation of epidermal growth factor receptor. Oncol Lett. (2016) 11:2934–40. doi: 10.3892/ol.2016.4300
102. Barone A, Sengupta R, Warrington NM, Smith E, Wen PY, Brekken RA, et al. Combined VEGF and CXCR4 antagonism targets the GBM stem cell population and synergistically improves survival in an intracranial mouse model of glioblastoma. Oncotarget. (2014) 5:9811–22. doi: 10.18632/oncotarget.2443
103. Willett CG, Boucher Y, di Tomaso E, Duda DG, Munn LL, Tong RT, et al. Direct evidence that the VEGF-specific antibody bevacizumab has antivascular effects in human rectal cancer. Nat Med. (2004) 10:145–7. doi: 10.1038/nm988
104. Zhao J, Ma MZ, Ren H, Liu Z, Edelman MJ, Pan H, et al. Anti-HDGF targets cancer and cancer stromal stem cells resistant to chemotherapy. Clin Cancer Res. (2013) 19:3567–76. doi: 10.1158/1078-0432.CCR-12-3478
Keywords: CSCs, esophageal, gastric, colorectal cancer, angiogenesis, vasculogenic mimicry
Citation: Lizárraga-Verdugo E, Avendaño-Félix M, Bermúdez M, Ramos-Payán R, Pérez-Plasencia C and Aguilar-Medina M (2020) Cancer Stem Cells and Its Role in Angiogenesis and Vasculogenic Mimicry in Gastrointestinal Cancers. Front. Oncol. 10:413. doi: 10.3389/fonc.2020.00413
Received: 22 August 2019; Accepted: 10 March 2020;
Published: 31 March 2020.
Edited by:
Laurence A. Marchat, National Polytechnic Institute of Mexico, MexicoReviewed by:
Ali Syed Arbab, Augusta University, United StatesCopyright © 2020 Lizárraga-Verdugo, Avendaño-Félix, Bermúdez, Ramos-Payán, Pérez-Plasencia and Aguilar-Medina. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Maribel Aguilar-Medina, bWFyaWJlbGFndWlsYXJAdWFzLmVkdS5teA==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.