 Thomas M. Drake
Thomas M. Drake Stephen R. Knight
Stephen R. Knight Ewen M. Harrison1
Ewen M. Harrison1 Kjetil Søreide
Kjetil Søreide- 1Department of Clinical Surgery, University of Edinburgh, Edinburgh, United Kingdom
- 2Department of Gastrointestinal Surgery, Stavanger University Hospital, Stavanger, Norway
- 3Department of Clinical Medicine, University of Bergen, Bergen, Norway
Precision medicine based upon molecular testing is heralded as a revolution in how cancer is prevented, diagnosed, and treated. Large efforts across the world aim to conduct comprehensive molecular profiling of disease to inform preclinical models, translational research studies and clinical trials. However, most studies have only been performed in patients from high-income countries. As the burden on non-communicable diseases increases, cancer will become a pressing burden across the world, disproportionately affecting low-middle income settings. There is emerging evidence that the molecular landscape of disease differs geographically and by genetic ancestry, which cannot be explained by environmental factors alone. There is a lack of good quality evidence that characterises the molecular landscape of cancers found in low-middle income countries. As cancer medicine becomes increasingly driven by molecular alterations in high-income settings, low-income settings may become left behind. Further efforts on an international scale must be made by researchers, funders, and policymakers to ensure cancer research addresses disease across the world, so models are not limited to subtypes of disease found in high-income countries. In this review, we discuss differences found in the molecular profiles of tumours worldwide and the implication this has for the future of global cancer care. Finally, we identify several barriers currently limiting progress in this field and innovative solutions, which may address these shortcomings.
Introduction
The incidence of cancer across the world is increasing, with the number of new cases set to rise by 70% in the next 20 years (1). This rise in incidence is accompanied by a sharp increase in cancer mortality, which disproportionately affects patients in low-middle income countries (2). As the global burden of communicable disease decreases with improvements in prevention, sanitation and treatment, non-communicable diseases such cancer will become a pressing burden. Whilst low and middle-income countries (LMICs) contend with barriers, such as delays in accessing healthcare, advanced disease at presentation, and limited access to treatment; research, and clinical practice in high income countries (HICs) is aimed toward developing treatment strategies tailored to individual patient characteristics and tumour biology.
Revolutionary polyomic (genomic, epigenomic, transcriptomic, proteomic, Figure 1) technologies have become established in clinical research and are increasingly commonplace in clinical practice in HICs. These technologies are set to revolutionise how research is performed and how therapies are selected—bringing precision medicine closer to reality than ever before. Indeed, the pharmaceutical industry are producing new agents designed to target specific molecular subtypes of disease. New prognostic modelling techniques, incorporating molecular data, clinical data and machine learning will provide more information to inform treatment choices for both patients and clinicians.
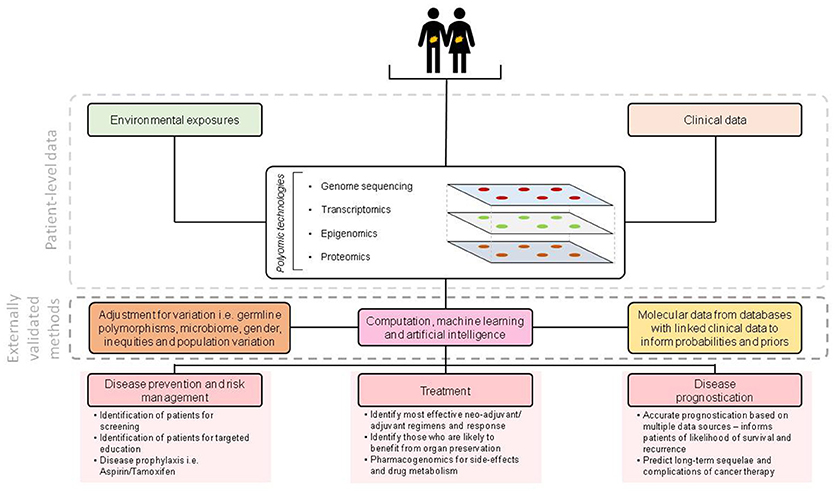
Figure 1. Overview of molecular technologies for enabling precision medicine.
These evolving capabilities are set to transform outcomes, however, there is little consideration given to how these technologies can improve cancer outcomes across the world and applicability to LMICs. Despite these remarkable advances, most research and clinical trials are conducted on populations within HICs, thus limiting global generalisability. There is clear evidence that basic cellular processes vary across different human populations (3, 4). Despite this, only 3% of genome-wide association studies have been performed in Africans, without considering sequencing studies (5, 6). Evidence in HICs from studies comparing individuals from different ancestries has found that despite controlling for socioeconomic factors and other environmental exposures, there is still a large disparity in cancer incidence and outcomes that remains to be addressed (7, 8). To prevent advances in technologies creating even greater disparity in cancer care across the world, work must now be expanded to include low and middle-income settings.
Cancer in Low and Middle-Income Settings
Across the spectrum of country development and geography, there are marked differences in the burden of cancer-related disease. Compared to well developed countries, LMICs have a higher age-standardised rate of gastric, oesophageal, bladder and liver cancers (Table 1). Although LMICs appear at present to have lower rates of lung, colorectal, pancreas and haematological malignancies, the 2016 Global Burden of Disease study has identified sustained rises between 1990 and 2016 of healthy years of life lost to these cancers (2).
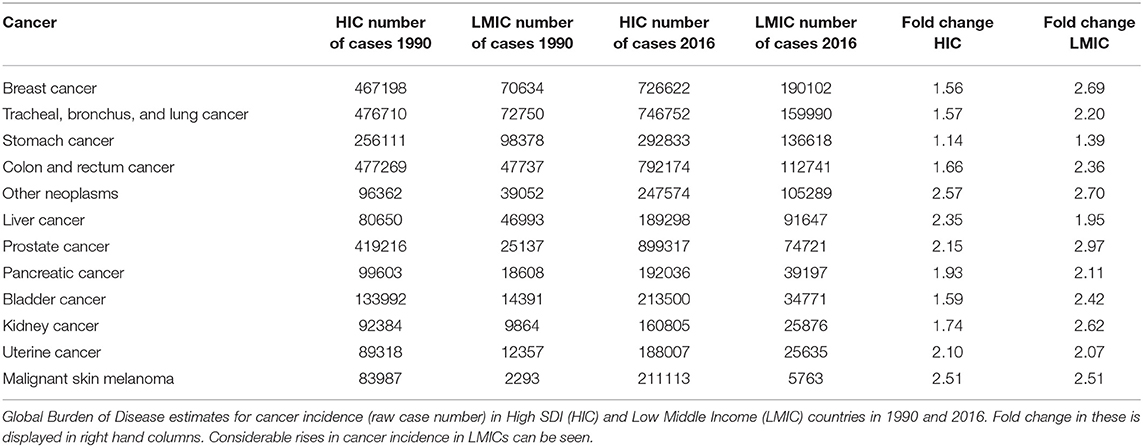
Table 1. Relative increases in cancer burden by income setting.
This socio-economic and geographical variation in causes of cancer across the world has implications for both clinical practice and future research. This difference in disease profile is multifactorial (Figure 2) and includes environmental factors, infectious agents (i.e., hepatitis B (HBV), hepatitis C (HCV), Human Immunodeficiency Virus (HIV), and Helicobacter pylori), occupational exposures and other lifestyle factors (smoking, alcohol use) amongst others. In addition to considerable differences in these risk factors, lower levels of resources for healthcare and education result in patients presenting to healthcare facilities with advanced disease in less developed countries.
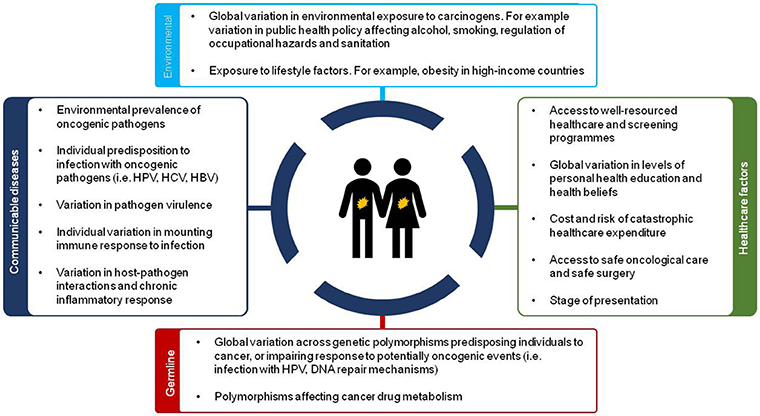
Figure 2. Overview of sources of disparity in future cancer care across the world.
It is important to consider the context of cancer disease in LMICs can be somewhat different to developed settings and the implications this has. In this review we will explore how the molecular aetiology and epidemiology of cancer in less developed settings may differ and explore the impact this has for the future of clinical practice and research.
Molecular Divergence in Cancer Aetiology
Evidence suggests there is variation in both somatic, germline, and epigenetic alterations found across different human populations (3, 4). What is emerging suggests there are key genetic differences in some solid tumours when disease found in LMIC countries is compared to that in HICs (9–13).
In both African and middle-eastern countries, germline mutations in loci predisposing to breast, ovarian and colorectal cancer have been characterised in a small number of studies (14–19). In breast cancer for example, mutations in the BRCA1 and BRCA2 genes are more commonly found, with one Nigerian study identifying mutation rates of 7.9% for BRCA1 and 3.1% for BRCA2—far higher than in the cancer genome atlas (TCGA), where these are 1.3 and 1.5% respectively (16). Evidence from Tunisia and Morocco is concordant, with a higher rate of BRCA1/BRCA2 mutations, with some found to be novel, previously uncharacterised, of unknown clinical significance (14, 18, 20).
The same holds true for colorectal cancer, where there is little population-level data surrounding the prevalence of germline mutations in common cancer susceptibility genes. Germline mutations reported in the literature from African countries suggest that these variants are typically different from those found in HICs and the clinical significance of these mutations remains poorly characterised (7, 21–23).
Somatic and germline alterations across tumour types are not exclusively limited to low-middle income countries; racial disparity in the molecular composition of tumours also affects patients in HICs. Within the United States, for all cancers combined those of African-American ethnicity have been found to have mortality rates up to 25 percent higher than in Caucasian Americans (24). Several studies from TCGA and others have identified several key alterations in the somatic landscapes of tumours from African-Americans or Asian patients for renal, endometrial, breast, head and neck, colorectal, cervical and prostate cancers (25–29). Frequently, the differences in the mutational landscape of these tumours are in pivotal cancer driver genes, such as the von Hippel-Lindau (VHL) gene in clear cell renal cancer (26).
Furthermore, oncogenes are found to be differentially mutated in different ancestral groups, for example in endometrial cancer where PTEN mutations were found to predominate in those of Caucasian or Asian descent, whereas in African-Americans TP53 mutations are more common. These variations across ancestral groups extend to the nucleotide level, with mutational signatures having significantly divergent nucleotide signatures when compared to mutations found in Caucasian populations (27). Several initiatives are underway to address this, such as the American Association for Cancer Research (AACR) 2020 by 2020, which aims to sequence matched normal tissue and tumour for 2020 African-American cancer patients by 2020 (30). It is unclear whether the 2020 by 2020 initiative would utilise whole exome or whole genome sequencing. The latter would give a far richer pool of information on which to base further studies upon.
Globally, projects aimed at defining the human genome and cancer biology on a regional basis are beginning to emerge. For example, the International Cancer Genome Consortium (ICGC) is aimed at developing a comprehensive description of genomic, transcriptomic and epigenomic changes in 50 tumour types, with data from 16 countries already included (31). Recent publications have characterised the whole genomes of 560 breast cancers, demonstrating more than 90 mutated cancer genes were implicated in carcinogenesis, and that the mechanisms underlying most mutational signatures are presently unknown (32, 33). Unfortunately, LMICs (i.e., sub-Saharan Africa) are poorly represented within the project, as with many international genome collaborations, which limits conclusions and applicability on a truly global scale.
Pharmacogenomics
Genetic polymorphisms affecting the metabolism of chemotherapy drugs may also be different across different ancestral groups. Differences in frequencies of functional genetic variants in key drug response and metabolism genes may significantly influence drug response differences in different populations (34–37).
Evidence from LMICs across the world is sparse, however, studies examining ethnic groups within HICs has identified substantial differences in treatment response and toxicities across ethnic groups. Understanding how these polymorphisms affect treatment response and side effects is important if precision medicine strategies are to be successfully implemented worldwide. These polymorphisms found across different ethnic populations can be beneficial or harmful. For example, African-Americans are more likely to have variants of the DPYD and TYMS genes which predispose this group to haematological toxicities with 5-fluorouracil as compared to Caucasian-Americans (38). Conversely, with 5-fluorouracil, Caucasian-Americans are more likely to suffer diarrhoea, nausea, vomiting, and mucositis when compared with their African-American counterparts. A similar picture is true in the metabolism of Doxorubicin, where African-Americans are more likely to suffer cardiotoxicities than Caucasian-Americans. Polymorphisms found in those of African ancestry may lead to life-threatening toxicities, such as neutropaenia. A fall in neutrophil counts following chemotherapy is more commonly found in patients of African-American and Asian descent when compared with Europeans (39). This may be due to a constitutionally lower neutrophil count (in the absence of cancer therapy), which has been associated with the presence of the Duffy antigen/receptor chemokine gene (DARC) rs2814778 SNP in a study examining 261 healthy volunteers (40). Several small studies examining the cytochrome P450, have identified polymorphisms across ethnic groups (41). Diversity in alleles of P450 is greatest across the African continent, compared to in Europe, and Asia. Where the CYP2B6, CYP2C8, CYP2D6, CYP2D6, CYP2D6, CYP3A5, and CYP3A5 have greatest diversity. Drugs associated with varied metabolism in the presence of polymorphisms affecting these genes include cyclophosphamide (CYP2B6*6), paclitaxel (CYP2C8*2) and 5HT3 receptor antagonists (CYP2D6). Despite this, the clinical implications these polymorphisms have for cancer therapy in the context of LMICs are poorly characterised. The polymorphisms found across these populations should be considered in the context of the healthcare infrastructure available. If patients in LMICs have similar diversity in polymorphisms associated with drug metabolism, then consideration must be given to the risk of exposing these patients to serious chemotherapy toxicities. Work is currently underway to try and identify polymorphisms associated with the metabolism of drugs found on the WHO's essential medicines list, beginning with HIV, which could be extended into cancer therapeutics and provide useful information to those administering treatments in LMICs (34, 42). Whole-genome precision medicine approaches to pharmacogenomics at the individual patient level are likely to be some way off, however, a precision medicine approach to public health could have significant advantages. In some ways, this population level consideration of the genetic diversity within a given population has started to occur. For example, in Ethiopia studies have revealed that a high proportion of the population are rapid codeine metabolisers due to CYP2D6 polymorphisms, leading to rapid conversion of codeine to morphine and subsequent overdose at therapeutic doses (100).
Molecular Diversity Affecting Communicable Causes of Cancer
Communicable diseases contribute toward a considerable proportion of the cancer burden in LMICs. These are potentially preventable cancers, with infectious agents commonly arising from poor sanitation, vertical transmission (mother to child), horizontal transmission (person to person) and a lack of safe healthcare practices (i.e., needlestick injuries and reused sharps). Examples include infectious agents such as Hepatitis B Virus (HBV), Hepatitis C Virus (HCV), Human Immunodeficiency Virus (HIV), human papilloma virus (HPV), and Epstein-Barr virus (EBV), which are well known for their oncogenic potential. Prevention programmes over the past 20 years have increasingly begun to recognise this, and vaccination programmes aimed at preventing hepatitis B have been shown to be effective in reducing the incidence of hepatocellular carcinoma. Similar can be seen for HPV vaccination, where programmes have begun to be rolled out in an increasing number of LMIC settings (43). Despite these initiatives, little is known as to the molecular landscape of these organisms and subsequent host-pathogen interactions.
Hepatocellular Carcinoma (HCC)
The effects of viral hepatitis on the development of hepatocellular carcinoma is well characterised in HICs. With global vaccination programmes aimed at preventing HBV underway, we may observe a decrease in HBV associated HCC. Despite this, HCC is multifactorial and infection with HCV or other causes of cirrhosis typically contribute to the risk of HCC development. The somatic landscape of HCC has been well characterised in American, European, Chinese and Japanese populations; however, evidence is lacking on disease found in LMICs (44, 45). The practicalities HCC poses to obtain tumour samples in LMICs are challenging, primarily owing to the risks associated with liver surgery and very late stages of presentation.
The distribution of HBV and HCV across the world has substantial variation and drive HCC formation in separate manners. HBV has a higher prevalence in LMICs and is responsible for the majority (%) of virus-induced hepatocellular carcinoma, compared with HCV (%) (46, 47). Dysregulation of key cell-cycle proteins, including cyclin dependent kinase 2 and 4, upregulation of the RAS/MAPK/ERK pathways and maintenance of upregulated canonical Wnt signalling, on the background of chronic inflammation are believed to initiate and drive HCV-related hepatocellular carcinoma (HCC) (48). HBV integrates within the host genome, initiating HCC through the promotion of genomic instability (49, 50).
Different genotypes of the Hepatitis C virus are known to lead to higher risk of HCC (51). Classically, the type 1b genotype has been associated with the highest risk of HCC formation. The global prevalence of HCV genotypes and variation worldwide has been studied in detail. Modelling studies demonstrate a high level of variation across the world in genotype, even within continents (46). Type 1 HCV predominates worldwide, however in central and west Africa, type 4 is more commonly found. Little data is available on why this variation exists and how this may reflect in disparity in HCC rates worldwide (46). In particular, study of genotype 4 and how this type mediates HCC formation in LMICs is missing. Host-pathogen interactions are known to play a crucial role in the clearance of these viruses and hence the subsequent risk of developing virus associated HCC. It is well known that patients of African ancestry have lower viral clearance rates than Caucasians (52). Several genome-wide association studies performed on patients of African descent have identified polymorphisms in alleles near class II Human Leukocyte Antigens on chromosome 6, the IL28B gene and other SNPs (53–56). This evidence draws upon a limited number of participants from LMICs and does not study the subsequent likelihood of HCC development.
The interaction between environmental and genetic factors may have a significant influence on the risk of HCC. The study of these interactions is limited in LMICs, however there are some examples of where this has been proven to be successful. Environmental exposures such as Aflatoxin B1 (found in certain grains and funghi), alcohol use, obesity, amongst others are known to contribute (57, 58). The interaction between Aflatoxin B1 and host genetics is particularly interesting: Aflatoxin B1 exposure is associated with HCC expressing more p53 mutations than in unexposed patients and may lead to greater genomic instability. Aflatoxin B1 has synergy with HBV, promoting HCC formation (59). Strategies to identify ways to abrogate DNA damage exerted by Aflatoxin B1 are under investigation and other environmental exposures are currently under study. Nevertheless, owing to little data on the molecular landscape of HCC in LMICs, it is unclear whether these research findings will have benefit for these patient groups. Similar epidemiological association studies have been done for other cancers, however, the data which provides the basis for disease models and translation to clinical practice in LMICs is lacking.
Cervical Cancer
Cervical cancer disproportionately affects women in LMICs, with the highest incidence found in sub-Saharan Africa (1, 60). Cervical cancer deaths are higher in LMICs, with 9 out of 10 deaths from cervical cancer worldwide occurring in LMIC settings. At present radiotherapy and surgery are the mainstays of cervical cancer management. The cause for this disparity in cancer incidence across the world is poorly understood and is multifactorial. Availability of radiotherapy in LMICs is also known to be extremely limited (61).
A proportion of cervical cancer cases are preventable, through early identification of dysplastic disease and immunisation against HPV. There are close connections with HIV too, with women who are HIV at a higher risk of developing cervical cancer. The prevalence of HPV is higher in women in LMICs than in HICs (62). There is a particularly high prevalence in Africa and Oceania, with higher exposure at a younger age. This in part accounts for the higher burden of cervical cancer in LMICs and the epidemiological burden of HPV associated cervical cancer is well described. In HIC populations genetic associations have been associated with the development of cervical cancer and are relatively well characterised (63, 64). Host-pathogen interactions between HPV and the immune system are also relatively well-characterised with respect to disease found in HICs. In HICs, genes such as TGF-β, those governing toll-like-receptors (TLRs), MHC genes and expression of cytokines, intricately linked with immune responses are associated with effective clearance of HPV (65–67). The somatic landscape of these tumours in LMICs is poorly characterised.
Evidence in LMICs suggests polymorphisms on the TYMS and RPS19 genes are associated with high-risk infection in Nigerian women, but overall in LMICs evidence is lacking on factors influencing effective HPV clearance (68). Several studies of women in HICs of African ancestry have identified that women of African-American descent despite having a similar prevalence of HPV, take longer to clear the virus (69). This delayed clearance, combined with known polymorphisms affecting genes responsible for immune response may go some way to explaining the higher rates of cervical cancer in LMICs (70). Further evidence is emerging that the distribution of HPV genotypes differs across separate geographical locations, with HPV 16 predominating worldwide, but HPV 58 and 31 more commonly found across Africa and East Asia. HPV 58 has been associated with increased risk of cervical cancer (71). Despite this work, there are scarce data from studies into virus genotype and genome-wide factors that may drive cervical cancer in LMICs.
Barriers to Global Research in Precision Medicine
Genetic variation through germline polymorphisms and somatic mutations associated with cancers in LMIC populations, suggests there may be opportunity for implementing global strategies for more effective individualised treatment and better prognostication. In HICs, molecular testing is already being used to target therapies to specific alterations in tumours. Examples of this include the use of trastuzumab following HER2 testing in breast cancer, use of endocrine therapy in breast cancer and guiding EGFR targeted therapies in colorectal cancer using KRAS testing. In the case of colorectal cancer, both type and location of KRAS mutations are known to predict response to EGFR inhibition in colorectal cancer (72). There is some limited evidence to suggest EGFR/KRAS mutations occur in a similar pattern and distribution in LMICs as compared to HICs (22, 73, 74). However, in African-American populations within HICs, there is an increased frequency of mutations found in genes implicated in EGFR signalling, which may correspond with more aggressive disease (75, 76). Despite this, no clinical trials have aimed to include patients within these settings or sequence tumours to identify predictors of response. Building evidence to support precision medicine at a global level is important, both in terms of providing effective cancer therapies and being able to decide whether targeted therapies are cost effective in LMICs.
Building a sustainable workforce of clinicians, laboratory medicine and scientific leaders in this field is key. At present, in LMICs, implementing effective national programmes for precision cancer therapy and prevention following similar models to examples within HICs is unlikely to be feasible. A lack of trained laboratory medicine workforce, instruments, transportation, finances, and evidence to support the applicability of clinical response are all key factors (77–79). Access to pathology and laboratory medicine services in their current format is a major issue, with some LMICs having no workforce at all (77, 78). To support the implementation of precision medicine approaches to therapies, there is a pressing need to change this and ensure the emerging workforce have the skills to support the transition to precision medicine in these settings. Alongside the development of cancer therapies and research models, delivery of increasingly complex therapies requires major improvements in healthcare infrastructure and resource and it is optimistic to say that workforce development alone will enable this. Establishing infrastructure to support translational research in LMIC settings has challenges. The use of regional biobanks may provide a method to collect tissue samples now, for processing later, when infrastructure is in place, or even to demonstrate feasibility of systematic tissue collection—derisking investments that would otherwise have been spent on building an entire sample handling and sequencing pipeline. Biobanks, however, require large amounts of energy resources to ensure samples are frozen, with some biobanks using liquid nitrogen to freeze samples. Liquid nitrogen transport is difficult in HICs, let alone securing a reliable supply in LMICs. Electricity supplies in some LMIC locations is sporadic too, which would be required to operate freezer systems. Other preservative solutions could be used to preserve nucleic acids in tissues at room temperature as a stop-gap solution, or as a means of extending the time available to transport specimens to a central repository. Transporting biospecimens and tissue is subject to tight United Nations control, thus making international efforts more difficult for countries without existing expertise to contribute. Furthermore, times of epidemic and regional spread of disease has implications for whether it would be safe to transport biospecimens across the world. Further issues surrounding the logistics remain and policy makers should identify solutions to this as a priority.
Global Cancer Trials
Conducting high-quality cancer trials is challenging in high-income settings and even more so in locations with little clinical trial infrastructure (80). Considerable methodological challenges exist around patient stratification by treatment response. Biomarker identification and sample handling must be robust and timely, together with the requirement for high levels of patient follow-up. In all countries, undertaking follow-up, and transporting biological specimens is challenging. These challenges are amplified in LMIC settings, with many countries lacking postal address systems, patient records and the infrastructure to process clinical specimens. Furthermore, a supply of clinical triallists is short in LMIC settings. Training and exchange programmes with HIC partners may help provide solutions in the sohrt term, however, long term infrastructure building must be given priority.
One example of where this is changing is Rwanda, where electronic patient record systems are being introduced (81). Integrating clinical systems into research in such settings would enable efficient research to be undertaken. In HICs, registry-based trials provide an efficient means of producing follow-up data and this approach could be emulated in LMICs where electronic records exist. A further consideration to any future trials of precision in LMIC settings is whether this can be continued after the trial concludes, should an intervention prove effective. For this reason, details on sequelae of interventions should be collected to ensure health-care systems can handle any treatment related harms. Non-governmental organisations such as the World Health Organisation and other non-governmental organisations (NGOs) often perform scoping studies in LMIC countries, but few cancer specific trials have been iniated. These NGOs should consider whether building research infrastructure in medicine is a sufficient priority to enable tailored solutions to be led by LMIC investigators independently.
In other areas of medicine (such as malaria, HCV and HIV) have successfully delivered clinical trials that integrate molecular or genotypic testing to enable molecular determinants of disease response to be identified in LMICs (82). Malaria is a good example, where trials often collect data on genotypes and information on microsatellites of malaria parasites from DNA isolated from blood films or spots. The markers used can be tested in a polymerase chain reaction (PCR) assay by research staff at that centre. Malaria trials have used these to investigate markers of treatment resistance and response (83, 84). Similarly, for trials in antiretroviral therapy for HIV and HCV, there are several studies which utilise commercial molecular testing kits at the centre level for identifying genotypes found in disease (85). However, this approach is challenging to adopt and thought must be given to the sustainability of testing when it is attempted to adopt trial findings into routine clinical practice.
Capacity Building for Precision Medicine
The most notable effort currently underway in LMICs to build evidence and crucially capacity for genomic sequencing is the Human Heredity and Health in Africa initiative (H3Africa) (86). This project is a collaboration of African clinicians, scientists and bioinformaticians who conduct large-scale sequencing and genetic association studies. So far, they have largely focussed on communicable disease such as trypanosomiasis, stroke and other neurological diseases affecting patients across the continent. Relevant to the field of oncology, some work has been undertaken into HPV infection in Nigeria (68), with the women included in this study demonstrating similar genetic susceptibility to infection as other populations. This group are set to expand into the field of breast cancer, which will provide useful data to study the genomic epidemiology of the disease on the African continent. The MRC Centre for Genomics and Global Health in the Gambia has also conducted successful genome wide and sequencing techniques in other disease areas, including in P. falciparum (87, 88).
There have been several success stories in LMICs, with some African countries delivering exciting genetic epidemiology studies. The Nigerian Breast Cancer Study, led by the University of Idaban has produced several large studies, underpinning the understanding of breast cancer genetics in Nigerian women (89). Through collaborative support from the University of Chicago, this group has gone on to publish multiple genetic epidemiology studies, and have even attempted randomised clinical trials (90). Building partnerships between institutions with experience in polyomics may help foster knowledge exchange and promote the implementation of best practice.
Local Lead and Oversight of Research Projects
Any solution must be led and maintained locally, rather than researchers from HICs taking data from local populations. It must also maintain practicality and clinical relevance to the local patient population. The H3Africa initiative has recently published guidelines for researchers wishing to do research in the African continent which are aimed at empowering local scientists and ensuring benefit for African patients (5). Developing home-grown expertise in polyomic technologies in LMICs would also have economic benefits in addition to building the global laboratory medicine workforce of the future. In fact, LMICs may be at some advantage in building competitive platforms for precision medicine as they can adopt new technologies without having to make mistakes or go through the intermediate steps of technological evolution that have been observed in high-income settings.
Disruptive Near-Future Technologies
New technologies may provide innovative solutions to some of the barriers to precision medicine. Genome sequencing over the past 15 years has fallen by 100,000-fold, yet with the cost of sequencing an entire human genome around the region of $1000, this is still prohibitively expensive for patients and healthcare systems (91). Furthermore, the sample handling and analysis pipelines required to support genome sequencing is logistically challenging. Establishing infrastructure and identifying expert staff to deal with sequencing data and ensure appropriate quality control is a large barrier. Large quantities of computational resource are required and may work as a centralised resource across, if shared across multiple LMICs. Whilst a centralised model may be a more cost and resource effective model, other challenges, including specimen transportation, and political stability could pose barriers to such a model. Furthermore, models of research and practice must be sensitive to resources of countries involved (Figure 3). Examples such as nanopore sequencing devices may provide an answer to these logistical challenges, with handheld and smartphone based sequencing devices available which reduce requirements for library preparation (92, 93). This has the potential to enable targeted sequencing of patients, at a far lower cost. With other multiplexing approaches, including developments in nucleotide barcoding, many samples could be rapidly processed at a very low cost.
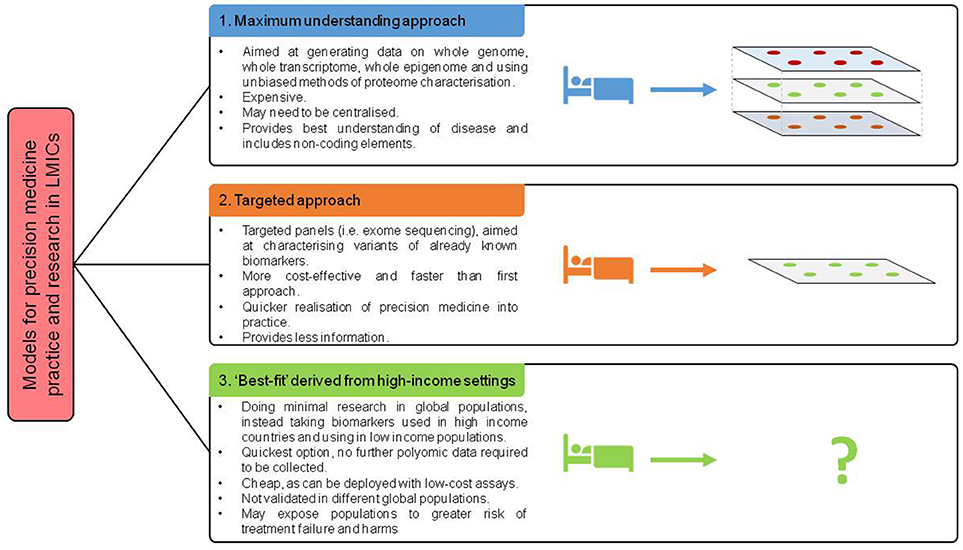
Figure 3. Proposed models of future deployment of precision medicine across world.
Targeted sequencing and mutation detection assays (PCR or ligation based methods, Table 2) offer a good alternative, but require that there is data to support that these assays bring clinical benefit, often derived from sequencing data. New, more portable approaches to molecular testing are being developed, which may be useful in resource-limited settings. Paper-based oligonucleotide based ligation assays, originally developed to detect HIV genotypes and monitor resistance, could be adapted for use in cancer therapy (94, 95). Paper-based immunochromatography assays, popularised for detection of HIV, could also be adapted for the detection of cancer specific ligands at the point of care. Nevertheless, these approaches lack the ability to resolve molecular alterations at the nucleotide level and rely on these already being known. The potential impact for these assays is great, particularly in the context of cancer diagnosis in limited resource settings, where other common diagnostic tests such as endoscopy or imaging maybe unavailable.
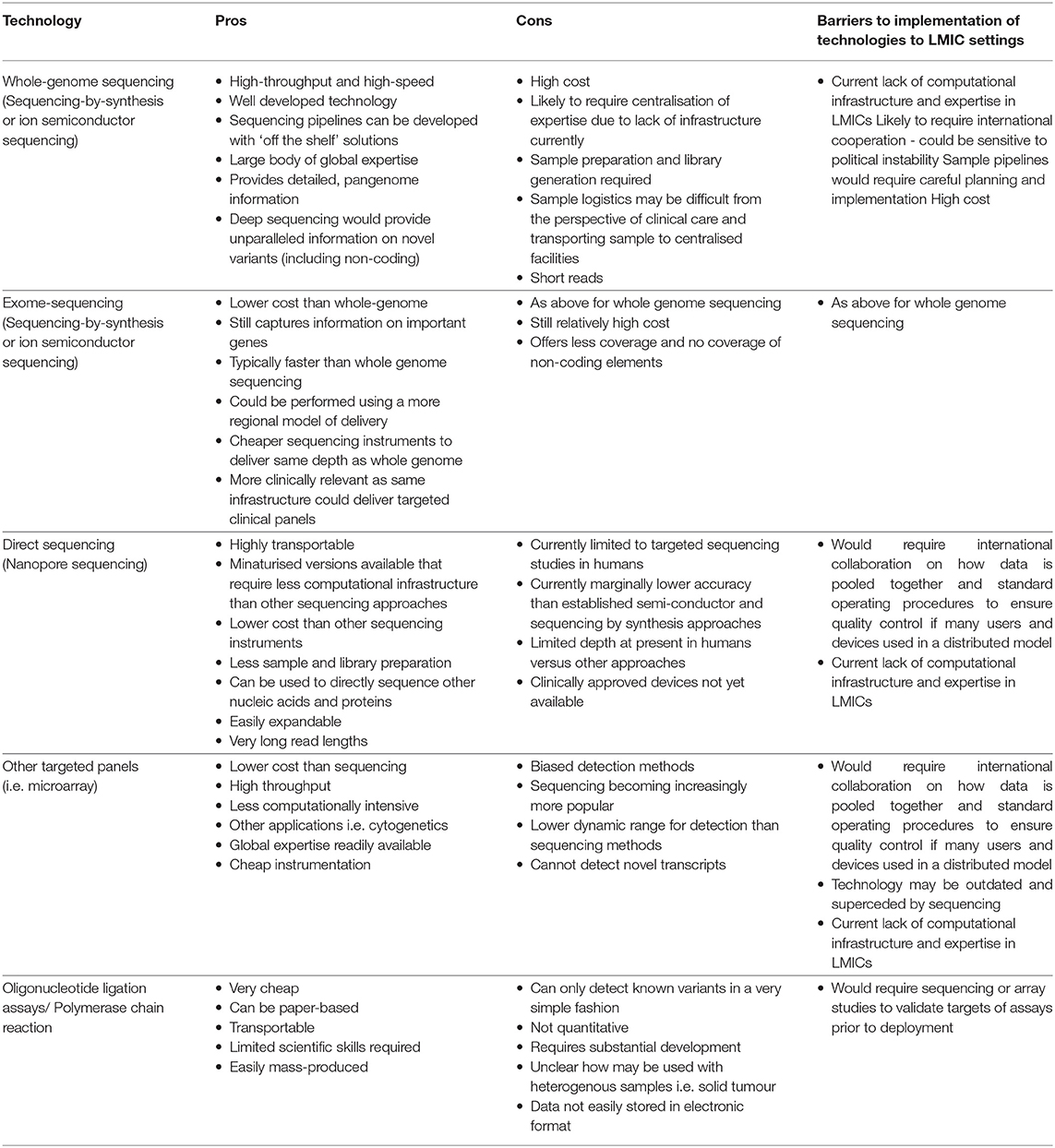
Table 2. Possible enabling technologies for global precision medicine.
Promising developments are on the horizon for the diagnosis and monitoring of disease, such as liquid biopsy and measurement of circulating nucleic acids (circulating tumour DNA/ RNA) (96). At present, these technologies rely of measurement of small quantities of nucleic acids, circulating tumour cells or other targets in peripheral blood via sequencing and subsequent computational processing (97). Although a reliable and universal test is yet to be developed based on this methodology, the development of such a test has the potential to improve early diagnosis and reduce reliance on other expensive methods of diagnostic testing in LMICs. However, given the lack of workforce, infrastructure and expertise available in LMICs at present, new technologies may further exacerbate global inequalities. Similarly, for other biomarkers of cancer which could be utilised as substrates to bind ligands to deliver cancer therapies with (such as fluorescence imaging (98), chemotherapeutics, or radiotherapeutics), there is little data to support whether what works in HICs can be translated straight into LMICs safely. Indeed, at present there is very little data that underpins the patient pathway or outcomes after cancer treatment in LMICs.
Collaborative approaches are crucial to ensuring future success (80, 99). Several partnerships between HICs and LMICs have begun to collect prospective, high quality evidence and establish clinical research networks. One such example is the GlobalSurg collaborative, a network of over 5,000 surgeons throughout the world who deliver large multicentre prospective studies (100). Recently, this network has established several trials units in LMIC settings that will deliver cancer trials and test whether new devices being concurrently developed in HICs can be utilised in LMICs. Building sustainable capacity concurrently with new developments will enable local economies to thrive and patients in LMICs to receive cutting edge care. Working with LMIC partners to facilitate global translation should be a priority when developing potentially disruptive technologies for both researchers and industry.
Summary
Funders, scientists, genome consortia, scientific journals and policy makers have an important role to play in a drive to ensure cancer research is generalisable across the world and will benefit patients in LMICs. Many major journals now mandate that polyomic data is deposited in databases for future use by the scientific community and other interested partners. This has been highly successful, with databases such as RNAcentral containing 13 million sequences and the ICGC Project available to the worldwide scientific community (31, 101). In addition, collaboratives such as the 1,000 genome project, containing 2,500 human genomes and representing 26 populations, demonstrate the growing potential to sequence genomes on a global scale (102). However, most data currently deposited is from HICs and more work is required to increase the availability of data from LMICs. Funders have begun to recognise this, and recent discussions have begun to focus on building precision medicine at a population level in LMICs (5, 103). At present, this initial funding is focussed on precision medicine for communicable disease, despite the rising burden of non-communicable disease.
Precision medicine clearly offers numerous advantages for patients and recent efforts to characterise the landscape of cancers using polyomic technologies is changing research and practice. There is, however, a lack of evidence, data and clear strategy on how this will be used to benefit patients across the world, particularly in LMICs. Emerging evidence suggests there are differences at the molecular level between cancer in HICs and that found in LMICs. To ensure inequity in cancer care between high and LMIC settings is not worsened, steps must be taken to improve the mechanistic understanding of cancer at a global level.
Summary Points
• There is known variation in basic cellular processes such as DNA methylation, epigenomic alterations and frequencies of polymorphisms across different human populations. These are likely to affect cancer risk, disease behaviour and treatment response.
• LMIC populations are under-represented in large genome wide association studies and sequencing studies. In an era where genomic technologies are driving drug development and targeted therapies, this may result in global inequities for cancer therapy.
• To prevent cancer inequities worsening further, funders, researchers and scientists should aim to include patients from LMICs in international studies. This would ensure that emerging consensus molecular subtypes are representative of disease worldwide.
• New technologies present exciting opportunities to improve cancer care and the representation of LMIC countries in cancer research. Further work should be done to ensure LMIC representation and identify novel ways of implementing cost-effective approaches to precision medicine or precision public health within LMIC settings.
Author Contributions
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Ferlay J, Soerjomataram I, Ervik M, Dikshit R, Eser S, Mathers C, et al. GLOBOCAN 2012 v1.0, Cancer Incidence and Mortality Worldwide: IARC CancerBase No. 11 [Internet]. Lyon: International Agency for Research on Cancer (2013) Available online at: http://globocan.iarc.fr (Accessed March 2, 2018).
2. Wang H, Naghavi M, Allen C, Barber RM, Carter A, Casey DC, et al. Global, regional, and national life expectancy, all-cause mortality, and cause-specific mortality for 249 causes of death, 1980–2015: a systematic analysis for the Global Burden of Disease Study 2015. Lancet (2016) 388:1459–544. doi: 10.1016/S0140-6736(16)31012-1
3. Kader F, Ghai M. DNA methylation-based variation between human populations. Mol Genet Genomics (2017) 292:5–35. doi: 10.1007/s00438-016-1264-2
4. Song M-A, Brasky TM, Marian C, Weng DY, Taslim C, Dumitrescu RG, et al. Racial differences in genome-wide methylation profiling and gene expression in breast tissues from healthy women. Epigenetics (2015) 10:1177–87. doi: 10.1080/15592294.2015.1121362
6. Tan DSW, Mok TSK, Rebbeck TR. Cancer genomics: diversity and disparity across ethnicity and geography. J Clin Oncol. (2016) 34:91–101. doi: 10.1200/JCO.2015.62.0096
7. Jovov B, Araujo-Perez F, Sigel CS, Stratford JK, McCoy AN, Yeh JJ, Keku T. Differential gene expression between African American and European American colorectal cancer patients. PLoS ONE (2012) 7: e30168. doi: 10.1371/journal.pone.0030168
8. Paulucci DJ, Sfakianos JP, Skanderup AJ, Kan K, Tsao CK, Galsky MD, et al. Genomic differences between black and white patients implicate a distinct immune response to papillary renal cell carcinoma. Oncotarget (2017) 8:5196–5205. doi: 10.18632/oncotarget.14122
9. Saffroy R, Morère JF, Bosselut N, Innominato PF, Hamelin J, Trédaniel J, et al. Impact of country of birth on genetic testing of metastatic lung adenocarcinomas in France: African women exhibit a mutational spectrum more similar to Asians than to Caucasians. Oncotarget (2017) 8:50792–803. doi: 10.18632/oncotarget.15132
10. Pollard JA, Burns PA, Hughes TA, Mukherjee G, Jones JL, Ho-Yen C, et al. Differential expression of microRNAs in breast cancers from four different ethnicities. (2018)S24. doi: 10.1159/000488456
11. Hansen MEB, Hunt SC, Stone RC, Horvath K, Herbig U, Ranciaro A, et al. Shorter telomere length in Europeans than in Africans due to polygenetic adaptation. Hum Mol Genet. (2016) 25:2324–30. doi: 10.1093/hmg/ddw070
12. Zhang Y, Zeng X, Lu H, Ji H, Zhao E, Li Y. Association between 8q24 (rs13281615 and rs6983267) polymorphism and breast cancer susceptibility: a meta-analysis involving 117,355 subjects. Oncotarget (2016) 7:68002–11. doi: 10.18632/oncotarget.12009
13. Amador MAT, Cavalcante GC, Santos NPC, Gusmão L, Guerreiro JF, Ribeiro-Dos-Santos Â, et al. Distribution of allelic and genotypic frequencies of IL1A, IL4, NFKB1 and PAR1 variants in Native American, African, European and Brazilian populations. BMC Res Notes. (2016) 9:101. doi: 10.1186/s13104-016-1906-9
14. Tazzite A, Jouhadi H, Nadifi S, Aretini P, Falaschi E, Collavoli A, et al. BRCA1 and BRCA2 germline mutations in Moroccan breast/ovarian cancer families: novel mutations and unclassified variants. Gynecol Oncol. (2012) 125:687–92. doi: 10.1016/j.ygyno.2012.03.007
15. Zhang J, Fackenthal JD, Huo D, Zheng Y, Olopade OI. Searching for large genomic rearrangements of the BRCA1 gene in a Nigerian population. Breast Cancer Res Treat (2010) 124:573–7. doi: 10.1007/s10549-010-1006-9
16. Fackenthal JD, Zhang J, Zhang B, Zheng Y, Hagos F, Burrill DR, et al. High prevalence of BRCA1 and BRCA2 mutations in unselected Nigerian breast cancer patients. Int J Cancer (2012) 131:1114–23. doi: 10.1002/ijc.27326
17. Cherbal F, Salhi N, Bakour R, Adane S, Boualga K, Maillet P. BRCA1 and BRCA2 unclassified variants and missense polymorphisms in Algerian breast/ovarian cancer families. Dis Markers (2012) 32:343–53. doi: 10.3233/DMA-2012-0893
18. Riahi A, Kharrat M, Lariani I, Chaabouni-Bouhamed H. High-resolution melting (HRM) assay for the detection of recurrent BRCA1/BRCA2 germline mutations in Tunisian breast/ovarian cancer families. Fam Cancer (2014) 13:603–9. doi: 10.1007/s10689-014-9740-5
19. Valarmathi MT, Sawhney M, Deo SS V., Shukla NK, Das SN. Novel germline mutations in theBRCA1 andBRCA2 genes in Indian breast and breast-ovarian cancer families. Hum Mutat. (2004) 23:205. doi: 10.1002/humu.9213
20. Koboldt DC, Fulton RS, McLellan MD, Schmidt H, Kalicki-Veizer J, McMichael JF, et al. Comprehensive molecular portraits of human breast tumours. Nature (2012) 490:61–70. doi: 10.1038/nature11412
21. The Cancer Genome Atlas. Comprehensive molecular characterization of human colon and rectal cancer. Nature (2012) 487:330–7. doi: 10.1038/nature11252
22. Raskin L, Dakubo JCB, Palaski N, Greenson JK, Gruber SB. Distinct molecular features of colorectal cancer in ghana. Cancer Epidemiol. (2013) 37:556–61. doi: 10.1016/j.canep.2013.07.007
23. Abdel-Malak C, Darwish H, Elsaid A, El-Tarapely F, Elshazli R. Association of APC I1307K and E1317Q polymorphisms with colorectal cancer among Egyptian subjects. Fam Cancer (2016) 15:49–56. doi: 10.1007/s10689-015-9834-8
24. National Cancer Institute DCCPS Surveillance Research Program. Surveillance, Epidemiology, and End Results (SEER). SEER*Stat Database Mortality—All COD, Total US (1990-2004)—Linked To Cty Attrib US, 1969-2004 Counties (2004) Available online at: http://www.seer.cancer.gov (Accessed April 2, 2018).
25. Ramakodi MP, Kulathinal RJ, Chung Y, Serebriiskii I, Liu JC, Ragin CC. Ancestral-derived effects on the mutational landscape of laryngeal cancer. Genomics (2016) 107:76–82. doi: 10.1016/j.ygeno.2015.12.004
26. Krishnan B, Rose TL, Kardos J, Milowsky MI, Kim WY. Intrinsic genomic differences between african American and white patients with clear cell renal cell carcinoma. JAMA Oncol. (2016) 2:664–7. doi: 10.1001/jamaoncol.2016.0005
27. Guttery DS, Blighe K, Polymeros K, Paul Symonds R, Macip S, Moss EL. Racial differences in endometrial cancer molecular portraits in The Cancer Genome Atlas. Oncotarget (2018) 9:17093–103. doi: 10.18632/oncotarget.24907
28. Spratt DE, Chan T, Waldron L, Speers C, Feng FY, Ogunwobi O, Osborne JR. Racial/ethnic disparities in genomic sequencing. JAMA Oncol. (2016) 2:1070–4. doi: 10.1001/jamaoncol.2016.1854
29. Zhang W, Edwards A, Flemington EK, Zhang K. Racial disparities in patient survival and tumor mutation burden, and the association between tumor mutation burden and cancer incidence rate. Sci Rep. (2017) 7:1–9. doi: 10.1038/s41598-017-13091-y
30. Jayaraman K. 2020 African American Genomes. Nat Publ Gr. (2018) 36:380–1. doi: 10.1038/nbt0518-380b
31. International Cancer Genome Consortium. Available online at: http://icgc.org/content/icgc-home-0#about (Accessed May 28, 2018).
32. Nik-Zainal S, Alexandrov LB, Wedge DC, Van Loo P, Greenman CD, Raine K, et al. Mutational processes molding the genomes of 21 breast cancers. Cell (2012) 149:979–93. doi: 10.1016/j.cell.2012.04.024
33. Nik-Zainal S, Davies H, Staaf J, Ramakrishna M, Glodzik D, Zou X, et al. Landscape of somatic mutations in 560 breast cancer whole genome sequences. Nature (2016) 534:47–54. doi: 10.1038/nature17676
34. Aminkeng F, Ross CJD, Rassekh SR, Brunham LR, Sistonen J, Dube M-P, et al. Higher frequency of genetic variants conferring increased risk for ADRs for commonly used drugs treating cancer, AIDS and tuberculosis in persons of African descent. Pharmacogenomics J. (2014) 14:160–70. doi: 10.1038/tpj.2013.13
35. Bu R, Gutiérrez MI, Al-Rasheed M, Belgaumi A, Bhatia K. Variable drug metabolism genes in Arab population. Pharmacogenomics J. (2004) 4:260–6. doi: 10.1038/sj.tpj.6500251
36. de Jong FA, Marsh S, Mathijssen RHJ, King C, Verweij J, Sparreboom A, Mcleod HL. ABCG2 pharmacogenetics: ethnic differences in allele frequency and assessment of influence on irinotecan disposition abcg2 pharmacogenetics: ethnic differences in Allele Frequency and Assessment of Influence on Irinotecan Disposition. (2004) 10:5889–94. doi: 10.1158/1078-0432.CCR-04-0144
37. Dandara C, Swart M, Mpeta B, Wonkam A, Masimirembwa C. Cytochrome P450 pharmacogenetics in African populations: implications for public health. Expert Opin Drug Metab Toxicol. (2014) 10:769–85. doi: 10.1517/17425255.2014.894020
38. O'Donnell PH, Dolan ME. Cancer pharmacoethnicity: ethnic differences in susceptibility to the effects of chemotherapy. Clin Cancer Res. (2009) 15:4806–14. doi: 10.1158/1078-0432.CCR-09-0344
39. Sekine I, Yamamoto N, Nishio K, Saijo N. Emerging ethnic differences in lung cancer therapy. Br J Cancer (2008) 99:1757–62. doi: 10.1038/sj.bjc.6604721
40. Grann VR, Bowman N, Joseph C, Wei Y, Horwitz MS, Jacobson JS, et al. Neutropenia in 6 ethnic groups from the Caribbean and the U.S. Cancer (2008) 113:854–60. doi: 10.1002/cncr.23614
41. Warnich L, Drögemöller BI, Pepper MS, Dandara C, Wright GEB. Pharmacogenomic research in south africa: lessons learned and future opportunities in the rainbow nation. Curr Pharmacogenomics Person Med. (2011) 9:191–207. doi: 10.2174/187569211796957575
42. Wadelius M, Alfirevic A. Pharmacogenomics and personalized medicine: the plunge into next-generation sequencing. Genome Med. (2011) 3:78. doi: 10.1186/gm294
43. Bruni L, Diaz M, Barrionuevo-Rosas L, Herrero R, Bray F, Bosch FX, et al. Global estimates of human papillomavirus vaccination coverage by region and income level: a pooled analysis. Lancet Glob Heal. (2016) 4:e453–63. doi: 10.1016/S2214-109X(16)30099-7
44. Ally A, Balasundaram M, Carlsen R, Chuah E, Clarke A, Dhalla N, et al. Comprehensive and integrative genomic characterization of hepatocellular carcinoma. Cell (2017) 169:1327–41.e23. doi: 10.1016/j.cell.2017.05.046
45. Yao S, Johnson C, Hu Q, Yan L, Liu B, Ambrosone CB, Wang J, Liu S. Differences in somatic mutation landscape of hepatocellular carcinoma in Asian American and European American populations. Oncotarget (2016) 7:40491–9. doi: 10.18632/oncotarget.9636
46. Blach S, Zeuzem S, Manns M, Altraif I, Duberg A-S, Muljono DH, et al. Global prevalence and genotype distribution of hepatitis C virus infection in 2015: a modelling study. Lancet Gastroenterol Hepatol. (2017) 2:161–76. doi: 10.1016/S2468-1253(16)30181-9
47. Razavi-Shearer D, Gamkrelidze I, Nguyen MH, Chen D-S, Van Damme P, Abbas Z, et al. Global prevalence, treatment, and prevention of hepatitis B virus infection in 2016: a modelling study. Lancet Gastroenterol Hepatol. (2018) 3:383–403. doi: 10.1016/S2468-1253(18)30056-6
48. Zhang Q. Activation of the Ras/Raf/MEK pathway facilitates hepatitis C virus replication via attenuation of the interferon-JAK-STAT Pathway. J Virol. (2012) 86:1544–54. doi: 10.1128/JVI.00688-11
49. Neuveut C, Wei Y, Buendia MA. Mechanisms of HBV-related hepatocarcinogenesis. J Hepatol. (2010) 52:594–604. doi: 10.1016/j.jhep.2009.10.033
50. Benn J, Schneider RJ. Hepatitis B virus HBx protein deregulates cell cycle checkpoint controls. Proc Natl Acad Sci USA. (1995) 92:11215–9.
51. Bruno S, Silini E, Crosignani A, Borzio F, Leandro G, Bono F, et al. Hepatitis C virus genotypes and risk of hepatocellular carcinoma in cirrhosis: a prospective study. Hepatology (1997) 25:754–8. doi: 10.1002/hep.510250344
52. Thomas DL, Astemborski J, Rai RM, Anania FA, Schaeffer M, Galai N, et al. The natural history of hepatitis C virus infection. JAMA (2000) 284:450–6. doi: 10.1001/jama.284.4.450
53. Thio CL, Carrington M, Marti D, O'Brien SJ, Vlahov D, Nelson KE, et al. Class II HLA alleles and hepatitis B virus persistence in African Americans. J Infect Dis. (1999) 179:1004–6. doi: 10.1086/314684
54. Mosbruger TL, Duggal P, Goedert JJ, Kirk GD, Hoots WK, Tobler LH, et al. Large scale candidate gene analysis of spontaneous clearance of hepatitis C Virus. J Infect Dis. (2010) 201:1371–80. doi: 10.1086/651606
55. Duggal P, Thio CL, Wojcik GL, Goedert JJ, Mangia A, Latanich R, et al. Genome-wide association study of spontaneous resolution of hepatitis C virus infection: data from multiple cohorts. Ann Intern Med. (2013) 158:235–45. doi: 10.7326/0003-4819-158-4-201302190-00003
56. Thomas DL, Thio CL, Martin MP, Qi Y, Ge D, Ohuigin C, et al. Genetic variation in IL28B and spontaneous clearance of hepatitis C virus. Nature (2009) 461:798–801. doi: 10.1038/nature08463
57. Hamid AS, Tesfamariam SG, Zhang Y, Zhang ZG. Aflatoxin B1-induced hepatocellular carcinoma in developing countries: geographical distribution, mechanism of action and prevention (Review). Oncol Lett. (2013) 5:1087–92. doi: 10.3892/ol.2013.1169
58. Herbst DA, Reddy KR. Risk factors for hepatocellular carcinoma. Clin Liver Dis. (2012) 1:180–2. doi: 10.1002/cld.111
59. Kew MC. Synergistic interaction between aflatoxin B1 and hepatitis B virus in hepatocarcinogenesis. Liver Int. (2003) 23:405–9. doi: 10.1111/j.1478-3231.2003.00869.x
60. GLOBOCAN Cancer Fact Sheets: Cervical Cancer. Available online at: http://globocan.iarc.fr/old/FactSheets/cancers/cervix-new.asp (Accessed May 29, 2018).
61. Zubizarreta EH, Fidarova E, Healy B, Rosenblatt E. Need for radiotherapy in low and middle income countries - the silent crisis continues. Clin Oncol. (2015) 27:107–14. doi: 10.1016/j.clon.2014.10.006
62. Human, Papillomavirus and Related Diseases Report WORLD,. Available online at: http://www.hpvcentre.net/statistics/reports/XWX.pdf (Accessed May 29, 2018).
63. de Sanjosé S, Diaz M, Castellsagué X, Clifford G, Bruni L, Muñoz N, et al. Worldwide prevalence and genotype distribution of cervical human papillomavirus DNA in women with normal cytology: a meta-analysis. Lancet Infect Dis. (2007) 7:453–9. doi: 10.1016/S1473-3099(07)70158-5
64. Crosbie EJ, Einstein MH, Franceschi S, Kitchener HC. Human papillomavirus and cervical cancer. Lancet (2013) 382:889–99. doi: 10.1016/S0140-6736(13)60022-7
65. Hardikar S, Johnson LG, Malkki M, Petersdorf EW, Galloway DA, Schwartz SM, et al. A population-based case-control study of genetic variation in cytokine genes associated with risk of cervical and vulvar cancers. Gynecol Oncol. (2015) 139:90–6. doi: 10.1016/j.ygyno.2015.07.110
66. Levovitz C, Chen D, Ivansson E, Gyllensten U, Finnigan JP, Alshawish S, et al. Tgf β receptor 1: An immune susceptibility gene in hpv-associated cancer. Cancer Res. (2014) 74:6833–44. doi: 10.1158/0008-5472.CAN-14-0602-T
67. Leo PJ, Madeleine MM, Wang S, Schwartz SM, Newell F, Pettersson-Kymmer U, et al. Defining the genetic susceptibility to cervical neoplasia—A genome-wide association study. PLOS Genet. (2017) 13:e1006866. doi: 10.1371/journal.pgen.1006866
68. Famooto A, Almujtaba M, Dareng E, Akarolo-Anthony S, Ogbonna C, Offiong R, et al. RPS19 and TYMS SNPs and prevalent high risk human papilloma virus infection in nigerian women. PLoS ONE (2013) 8:19–24. doi: 10.1371/journal.pone.0066930
69. Banister CE, Messersmith AR, Cai B, Spiryda LB, Glover SH, Pirisi L, et al. Disparity in the persistence of high-risk human papillomavirus genotypes between African American and European American Women of College Age. J Infect Dis. (2015) 211:100–8. doi: 10.1093/infdis/jiu394
70. Sudenga SL, Wiener HW, Shendre A, Wilson CM, Tang J, Shrestha S. Variants in interleukin family of cytokines genes influence clearance of high risk HPV in HIV-1 coinfected African–American adolescents. Hum Immunol. (2013) 74:1696–700. doi: 10.1016/J.HUMIMM.2013.08.010
71. Chan PKS, Lam CW, Cheung T-H, Li WWH, Lo KWK, Chan MYM, et al. Association of Human Papillomavirus type 58 variant with the risk of cervical cancer. JNCI J Natl Cancer Inst. (2002) 94:1249–53. doi: 10.1093/jnci/94.16.1249
72. Bertotti A, Papp E, Jones S, Adleff V, Anagnostou V, Lupo B, et al. The genomic landscape of response to EGFR blockade in colorectal cancer. Nature (2015) 526:263–7. doi: 10.1038/nature14969
73. Abdulkareem FB, Sanni LA, Richman SD, Chambers P, Hemmings G, Grabsch H, et al. KRAS and BRAF mutations in Nigerian colorectal cancers. West Afr J Med. (2012) 31:198–203.
74. Yoon HH, Shi Q, Alberts SR, Goldberg RM, Thibodeau SN, Sargent DJ, et al. Racial differences in BRAF/KRAS mutation rates and survival in stage III colon cancer patients. J Natl Cancer Inst. (2015) 107:1–10. doi: 10.1093/jnci/djv186
75. Ashktorab H, Daremipouran M, Devaney J, Varma S, Rahi H, Lee E, et al. Identification of novel mutations by exome sequencing in African American colorectal cancer patients. Cancer (2015) 121:34–42. doi: 10.1002/cncr.28922
76. Staudacher JJ, Yazici C, Bul V, Zeidan J, Khalid A, Xia Y, et al. Increased frequency of kras mutations in african americans compared with caucasians in sporadic colorectal cancer. Clin Transl Gastroenterol. (2017) 8:e124. doi: 10.1038/ctg.2017.48
77. Horton S, Sullivan R, Flanigan J, Fleming KA, Kuti MA, Looi LM, et al. Delivering modern, high-quality, affordable pathology and laboratory medicine to low-income and middle-income countries: a call to action. Lancet (2018) 391:1953–64. doi: 10.1016/S0140-6736(18)30460-4
78. Wilson ML, Fleming KA, Kuti MA, Looi LM, Lago N, Ru K. Access to pathology and laboratory medicine services: a crucial gap. Lancet (2018) 391:1927–38. doi: 10.1016/S0140-6736(18)30458-6
79. Sayed S, Cherniak W, Lawler M, Tan SY, El Sadr W, Wolf N, et al. Improving pathology and laboratory medicine in low-income and middle-income countries: roadmap to solutions. Lancet (2018) 391:1939–52. doi: 10.1016/S0140-6736(18)30459-8
80. Soreide K, Alderson D, Bergenfelz A, Beynon J, Connor S, Deckelbaum DL, et al. Strategies to improve clinical research in surgery through international collaboration. Lancet (2013) 382:1140–51. doi: 10.1016/S0140-6736(13)61455-5
81. Amoroso CL, Akimana B, Wise B, Fraser HSF. Using electronic medical records for HIV care in rural Rwanda. Stud Health Technol Inform. (2010) 160:337–41. doi: 10.3233/978-1-60750-588-4-337
82. Opi DH, Swann O, Macharia A, Uyoga S, Band G, Ndila CM, et al. Two complement receptor one alleles have opposing associations with cerebral malaria and interact with α+thalassaemia. Elife (2018) 7:1–27. doi: 10.7554/eLife.31579
83. Arinaitwe E, Sandison TG, Wanzira H, Kakuru A, Homsy J, Kalamya J, et al. Artemether lumefantrine versus dihydroartemisinin piperaquine for falciparum malaria: a longitudinal, randomized trial in young ugandan children. Clin Infect Dis. (2009) 49:1629–37. doi: 10.1086/647946
84. Yeka A, Dorsey G, Kamya MR, Talisuna A, Lugemwa M, Rwakimari JB, et al. Artemether-lumefantrine versus dihydroartemisinin-piperaquine for treating uncomplicated malaria: a randomized trial to guide policy in Uganda. PLoS ONE (2008) 3:e2390. doi: 10.1371/journal.pone.0002390
85. Gregson J, Kaleebu P, Marconi VC, van Vuuren C, Ndembi N, Hamers RL, et al. Occult HIV-1 drug resistance to thymidine analogues following failure of first-line tenofovir combined with a cytosine analogue and nevirapine or efavirenz in sub Saharan Africa: a retrospective multi-centre cohort study. Lancet Infect Dis. (2017) 17:296–304. doi: 10.1016/S1473-3099(16)30469-8
86. Mulder NJ, Adebiyi E, Adebiyi M, Adeyemi S, Ahmed A, Ahmed R, et al. Development of bioinformatics infrastructure for genomics research. Glob Heart (2017) 12:91–8. doi: 10.1016/j.gheart.2017.01.005
87. Reid AJ, Talman AM, Bennett HM, Gomes AR, Sanders MJ, Illingworth CJR, et al. Single-cell RNA-seq reveals hidden transcriptional variation in malaria parasites. Elife (2018) 7:e33105. doi: 10.7554/eLife.33105
88. Jallow M, Teo YY, Small KS, Rockett KA, Deloukas P, Clark TG, et al. Genome-wide and fine-resolution association analysis of malaria in West Africa. Nat Genet. (2009) 41:657–65. doi: 10.1038/ng.388
89. Oguntunde PE, Adejumo AO, Okagbue HI. Breast cancer patients in Nigeria: data exploration approach. Data Br. (2017) 15:47–57. doi: 10.1016/j.dib.2017.08.038
90. Arowolo OA, Njiaju UO, Ogundiran TO, Abidoye O, Lawal OO, Obajimi M, et al. Neo-adjuvant capecitabine chemotherapy in women with newly diagnosed locally advanced breast cancer in a resource-poor setting (Nigeria): efficacy and safety in a phase II feasibility study. Breast J. (2013) 19:470–7. doi: 10.1111/tbj.12149
91. The, Cost of Sequencing a Human Genome - National Human Genome Research Institute (NHGRI),. Available online at: https://www.genome.gov/27565109/the-cost-of-sequencing-a-human-genome/ (Accessed May 29, 2018).
92. Jain M, Koren S, Miga KH, Quick J, Rand AC, Sasani TA, et al. Nanopore sequencing and assembly of a human genome with ultra-long reads. Nat Biotechnol. (2018) 36:338–45. doi: 10.1038/nbt.4060
93. Suzuki A, Suzuki M, Mizushima-Sugano J, Frith MC, Makałowski W, Kohno T, et al. Sequencing and phasing cancer mutations in lung cancers using a long-read portable sequencer. DNA Res. (2017) 24:585–96. doi: 10.1093/dnares/dsx027
94. Natoli ME, Rohrman BA, De Santiago C, van Zyl GU, Richards-Kortum RR. Paper-based detection of HIV-1 drug resistance using isothermal amplification and an oligonucleotide ligation assay. Anal Biochem. (2018) 544:64–71. doi: 10.1016/j.ab.2017.12.008
95. Jauset-Rubio M, Svobodová M, Mairal T, McNeil C, Keegan N, Saeed A, et al. Ultrasensitive, rapid and inexpensive detection of DNA using paper based lateral flow assay. Sci Rep. (2016) 6:1–10. doi: 10.1038/srep37732
96. Nordgård O, Tjensvoll K, Gilje B, Søreide K. Circulating tumour cells and DNA as liquid biopsies in gastrointestinal cancer. Br J Surg. (2018) 105:e110–20. doi: 10.1002/bjs.10782
97. Cohen AJD, Li L, Wang Y, Thorburn C, Danilova L, Douville C, et al. Detection and localization of surgically resectable cancers with a multi- analyte blood test. Science (2018) 3247:1–10. doi: 10.1126/science.aar3247
98. Seth S, Akram AR, McCool P, Westerfeld J, Wilson D, McLaughlin S, et al. Assessing the utility of autofluorescence-based pulmonary optical endomicroscopy to predict the malignant potential of solitary pulmonary nodules in humans. Sci Rep. (2016) 6:1–10. doi: 10.1038/srep31372
99. Hyman SE. The importance of true collaboration in efforts to increase diversity in genetic analyses. Curr Biol. (2018) 28:R598. doi: 10.1016/j.cub.2018.04.039
100. Bhangu A, Ademuyiwa AO, Aguilera ML, Alexander P, Al-Saqqa SW, Borda-Luque G, et al. Surgical site infection after gastrointestinal surgery in high-income, middle-income, and low-income countries: a prospective, international, multicentre cohort study. Lancet Infect Dis. (2018) 18:516–525. doi: 10.1016/S1473-3099(18)30101-4
101. RNAcentral. Available online at: http://rnacentral.org/about-us (Accessed May 29, 2018).
102. Altshuler DL, Durbin RM, Abecasis GR, Bentley DR, Chakravarti A, Clark AG, et al. A map of human genome variation from population-scale sequencing. Nature (2010) 467:1061–73. doi: 10.1038/nature09534
103. African, Academy of Sciences|Alliance for Accelerating Excellence in Science in Africa (AESA) | AESA,. Available online at: https://aasciences.ac.ke/programmes/easa/alliance-for-accelerating-excellence-in-science-in-africa-aesa/ (Accessed May 29, 2018).
Keywords: cancer, surgery, oncology, genomics, low-income, global health, global surgery, precision medicine
Citation: Drake TM, Knight SR, Harrison EM and Søreide K (2018) Global Inequities in Precision Medicine and Molecular Cancer Research. Front. Oncol. 8:346. doi: 10.3389/fonc.2018.00346
Received: 31 May 2018; Accepted: 07 August 2018;
Published: 04 September 2018.
Edited by:
Dana Hashim, Cancer Registry of Norway, NorwayReviewed by:
Eiko Saito, The University of Tokyo, JapanUmberto Malapelle, Università degli Studi di Napoli Federico II, Italy
Copyright © 2018 Drake, Knight, Harrison and Søreide. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Thomas M. Drake, dC5kcmFrZUBlZC5hYy51aw==
Kjetil Søreide, a3NvcmVpZGVAbWFjLmNvbQ==