 Marissa Sobolewski
Marissa Sobolewski Garima Singh2
Garima Singh2 Jay S. Schneider
Jay S. Schneider Deborah A. Cory-Slechta
Deborah A. Cory-Slechta- 1Department of Environmental Medicine, University of Rochester Medical Center, Rochester, NY, United States
- 2Department of Pathology, Anatomy and Cell Biology, Thomas Jefferson University, Philadelphia, PA, United States
While it is clear that behavioral experience modulates epigenetic profiles, it is less evident how the nature of that experience influences outcomes and whether epigenetic/genetic “biomarkers” could be extracted to classify different types of behavioral experience. To begin to address this question, male and female mice were subjected to either a Fixed Interval (FI) schedule of food reward, or a single episode of forced swim followed by restraint stress, or no explicit behavioral experience after which global expression levels of two activating (H3K9ac and H3K4me3) and two repressive (H3K9me2 and H3k27me3) post-translational histone modifications (PTHMs), were measured in hippocampus (HIPP) and frontal cortex (FC). The specific nature of the behavioral experience differentiated profiles of PTHMs in a sex- and brain region-dependent manner, with all 4 PTHMs changing in parallel in response to different behavioral experiences. These different behavioral experiences also modified the pattern of correlations of PTHMs both within and across FC and HIPP. Unexpectedly, highly robust correlations were found between global PTHM levels and behavioral performances, suggesting that global PTHMs may provide a higher-order pattern recognition function. Further efforts are needed to determine the generality of such findings and what characteristics of behavioral experience are critical for modulating PTHM responses.
Introduction
The importance of behavioral history to the nature of subsequent behavioral and brain function has long been recognized (Barrett, 1986; Pattij et al., 2004; Marchner and Preuschhof, 2018) and can include differential behavioral responses to subsequent drug treatments, or even the maintenance of such seemingly incongruous behavior as response-produced shock that can emerge following specific behavioral histories (Byrd, 1972). In children, intervention programs such as Head Start, have been shown to result in long-term benefits in school achievement, grade retention, special education placement, and social adjustment (Barnett, 1995). Parallels exist in animal models using environmental enrichment. Such enrichment is achieved in several different forms, both social and physical, with multiple reports showing its ability to produce beneficial effects (Hirase and Shinohara, 2014; Kelly, 2015; Kondo, 2017). Early adverse behavioral experiences also have persistent and profound damaging effects, both cognitive and emotional. Children living in poverty, for example, have reduced gray matter volume in the frontal and temporal cortex and in hippocampus (HIPP) (Brito and Noble, 2014; Hair et al., 2015; Noble et al., 2015). Early institutionalization or being raised in an orphanage, as deprivation-related adversities, resulted in developmental delays in social, behavioral and cognitive domains (Chugani et al., 2001; Johnson et al., 2006). Some studies have even suggested that such effects may be intergenerational (Galler and Rabinowitz, 2014). The long-lasting temporal influence of behavioral experience has invoked a variety of mechanistic explanations, most notably epigenetic mechanisms.
Indeed, behavioral experiences do influence brain epigenetic profiles (Szyf et al., 2016). Glucocorticoid receptor gene methylation was found to be higher in blood of adolescents that had experienced either childhood trauma or stressful life events (van der Knaap et al., 2014). Similar findings have emerged from studies of abused or neglected children (Guillemin et al., 2014; Weder et al., 2014; Romens et al., 2015; Turecki and Meaney, 2016). Living in poverty during childhood, with its adverse conditions, produces differences in blood profile DNA methylation profiles (Borghol et al., 2012). Early child abuse leads to greater DNA methylation of the hippocampal glucocorticoid receptor (McGowan et al., 2009). In animal models, maternal deprivation in non-human primates produced widespread changes in both blood and brain DNA methylation patterns (Provençal et al., 2012), as did hierarchical status in non-human primates (Tung et al., 2012). In rodents, studies of maternal separation and differences in maternal care alter hippocampal DNA methylation (Turecki and Meaney, 2016). Although much of the current focus has been related to the epigenetic consequences of adverse experiences, studies also show that positive reinforcement based learning experiences likewise alter epigenetic marks (Day et al., 2013). Thus, epigenetic marks, through dynamic control of gene transcription, essentially frame the parameters within which brain and behavioral function can occur. It has been suggested that the trajectory of behavioral experiences that vary in both intensity and quality, may further modify brain epigenetic marks, essentially assimilating an individual’s behavioral experience and reframing the boundaries of subsequent behavioral and brain function (Szyf and Bick, 2013). Such findings likely contribute to the repeated reports of epigenetic differences in monozygotic twins (Fraga et al., 2005; Nipa et al., 2009; Wong et al., 2010; Chiarella et al., 2015).
However, it is unknown how, and to what extent, different types of behavioral experiences influence the epigenome or what characteristics of those experiences are critical to subsequent epigenetic changes that shape the parameters of future brain and behavioral function. Comparisons of epigenetic profiles in response to differing behavioral experiences are limited. One study contrasted the impact of histone deacetylase (HDAC) inhibitors on bees that had been trained to discriminate between a rewarding odor stimulus (i.e., a positive conditioned stimulus; limonene) vs. an aversive odor stimulus (i.e., a negative conditioned stimulus; natural vanilla) as conditioned stimuli predictive of subsequent access to sucrose vs. saturated NaCl solutions, respectively. It was found that HDAC inhibitors impaired discrimination memory for the aversive stimulus, but did not impact the positive reward-based conditioning, despite the fact that both were forms of learning. Additional manipulations confirmed that these observations did not reflect differences in sucrose sensitivity or locomotor activity (Lockett et al., 2014).
Such studies demonstrate that differing characteristics of the behavioral experience influence subsequent epigenetic profiles, gene expression and cellular function. Understanding the controlling characteristics is critical to advancing behavioral epigenetics and to the use of behavioral experiences as intervention strategies to reverse or reprogram epigenetic changes induced by early adversity (Szyf et al., 2016). To evaluate the role of different types of behavioral experience, the current study examined differences in epigenetic profiles in mice that had been subjected to a fixed interval (FI) schedule of food reward which consisted of earning food rewards via a lever press response that were available after FIs of time, or to a single episode of forced swim followed by a single episode of restraint stress, or to no explicit behavioral experience (remaining in the home cage).
Gene transcription is controlled by a heritable triad of epigenetic mechanisms, including gene methylation, post-translational refinement by non-coding RNAs, chromatin remodeling and histone modifications. Previous work focused on determinations of gene and site-specific changes in epigenetic methylation patterns associated with behavioral experience has clearly advanced the understanding of the molecular mechanisms by which environmentally induced epigenetic changes influence gene expression and consequent function. However, such methods ignore the network-based function of brain. A recent study of in vivo epigenetic imaging of HDACs found them to be concurrently highly expressed in multiple brain regions in humans, with differences between gray and white matter as well as between cortical and subcortical gray matter (Wey et al., 2016). This is not surprising given that networked connectivity is a central feature of the brain and its operation is predicted by clustered gene expression hubs, with networked, connected regions of brain exhibiting similar transcriptional profiles (Fulcher and Fornito, 2016; Xu et al., 2016; Forest et al., 2017). This suggests that epigenetic influences on gene expression connected with specific behavioral experiences may likewise be clustered, and that the brain must ultimately integrate this aggregate information in responding to behavioral experiences (Greenspan, 2001; French and Pavlidis, 2011; Wolf et al., 2011; Ji et al., 2014; Hawrylycz et al., 2015; Richiardi et al., 2015; Fulcher and Fornito, 2016; Forest et al., 2017). Given the networked nature of brain function and the epigenetic consequences of behavioral experience, this study also sought to explore whether global PTHM levels could act as an integrated signal of epigenetic modifications, as they have previously been shown to predict behavioral differences (Bousiges et al., 2010; Nesbitt et al., 2014). Global post-translational histone modification (PTHM) levels [H3K9ac, H3K4me3 (activating marks) and H3K9me2 and H3K27me3 (repressive marks)] were measured in HIPP and frontal cortex (FC) following differential behavioral experience. As evidenced by distinctive patterns of correlations between PTHMs, both within and between brain regions, such data may suggest “biomarkers” or epigenetic/genetic signatures (Kubota, 2016) for subsequent classification of behavioral events and/or directions for future behavioral studies, including shifts in gene profiles and chromatin dynamics shaped by such changes in total histone modifications.
Materials and Methods
Animals
Adult C57Bl6 mice (Jackson Labs, Bar Harbor, ME, United States) that had been pair-housed by sex since weaning were randomly assigned at approximately 60 days of age to receive exposure to one of three behavioral experience conditions as described below. All three behavioral experience groups have no more than 1 pup/sex from any dam, so as to mitigate any litter specific effects within behavioral experience groups. Following the completion of all behavioral experience, FC and HIPP were extracted from mice for subsequent molecular analyses. Procedures used in this study had approval of the Institutional Animal Care and Use Committees at the University of Rochester, and all mice used in this study were treated humanely and with regard for alleviation of suffering.
Behavioral Experience
Fixed Interval (FI) Schedule Controlled Behavioral Experience
Fixed Interval schedule controlled behavioral experience was comprised of 30 behavioral test sessions (5 days per week, M-F) in which reinforcement (food reward; 20 mg food pellet; PJ Noyes) was programmed according to a FI 60 s schedule of reinforcement (Cory-Slechta et al., 1985, 1996, 1997). A prototypical behavioral pattern that is widespread across species emerges from this behavioral paradigm. FI test sessions were undertaken in operant chambers (Med Associates, Model ENV-307W) that were housed in cabinets that included sound-attenuation by white noise generators and fans for ventilation. The back wall of the chamber had three response levers configured horizontally, with a dual liquid dipper and food pellet dispenser delivery on the front (opposite) wall. The FI schedule requires a single lever press on the designated active lever (left lever) after the specified time interval (60 s) ends, which produces food delivery and starts the subsequent 60 s interval. Responses on any lever during the 60 s interval do not have any programmed consequences. FI test sessions were 30 min in duration. Standard performance measures of FI behavior were used were overall rate (defined as the total number of responses/total session time), the postreinforcement pause (PRP) time (defined as the time from the beginning of the 60 s interval to the occurrence of the first response), run rate (defined as the total number of responses/total time minus the PRP time) and interresponse time (IRT) or mean time between responses as previously described (Cory-Slechta et al., 1998).
Forced Swim-Restraint Stress (FS-RS) Behavioral Experience
Forced Swim-Restraint Stress (FS-RS) behavioral experience was comprised of an initial exposure to one 5 min session of forced swimming followed 1 week later by a single 30 min episode of restraint (immobilization) stress. For the forced swim exposure, mice were placed into a bucket of water that precluded escape, and that did not allow mice to use their tail to balance. The following week, a single 30 min immobilization restraint stress was carried out in which mice were placed in a restrictive plastic tube measuring 4.5″ in length and 1.1″ in width. Measures of forced swim included mean float duration, total float duration, mean duration swimming, and total duration swimming. There was no particular rationale for the order of these events, as both were considered highly salient.
No Behavioral Experience
No behavioral experience consisted of mice remaining in the home cages for the entire duration of behavioral experience afforded to the other groups.
Since the FI schedule of food reward requires food-motivation, all mice in all behavioral experience groups underwent caloric restriction beginning at approximately 50 days of age that maintained body weights at 90% of ad-lib weights; this continued for the duration of the experiment. Body weights were collected daily from M to F. Brains from all three experience groups were collected 1 week following completion of the FI schedule behavioral testing. For this purpose, consistent sections of FC and HIPP were dissected from fresh brain tissue. As FS-RS behavioral experience was completed before FI behavioral experience, a longer overall post-behavior experience time (approximately 1 week) elapsed for this group, resulting in some differences in overall timing between these experiences and global PTHM assessment. However, extending the duration of the FS-RS experience (i.e., repeated exposures) would also have led to habituation, and thus potentially diminished the salience of the differences in behavioral experience conditions (Babb et al., 2014; McInnis et al., 2015; Sadler and Bailey, 2016).
Measurement of Global Post-translational Histone Modifications Levels
A random subset of 5 of the 12 animals from each sex/behavioral experience group were used for these analyses. FC and HIPP were dissected from each of those 5 brain brains and homogenized for subsequent analyses. Thus, sample size for PTHM expression levels was n = 5 for each region/sex/behavioral treatment group and all PTHM expression level analyses utilized the homogenized tissue.
Histone Purification
Histone Purification An Active Motif Histone Purification mini kit (Cat. No. 40026) was used to purify histones via the protocol provided by the manufacturer, with some modifications. For this purpose, the dissected FC and HIPP collected at the termination of behavioral testing (five biological replicates per sex per behavioral experience group), all obtained from the left hemisphere were homogenized in 0.3 ml extraction buffer and incubated overnight at 4°C on a rotating platform. All steps were performed at 4°C. The crude histone extract was neutralized by adding 5× neutralization buffer prior to purifying the histone proteins. Purification was done using spin columns that were provided with the kit, which were then washed three times with 0.5 ml histone wash buffer. Elution was done in 0.05 ml elution buffer. The histone proteins were then precipitated overnight using perchloric acid to a final 4% concentration, after which they were pelleted by spinning at 14,000 rpm for 1 h at 4°C. Salts were eliminated by first washing twice with 0.5 ml 4% perchloric acid, then by acetone containing 0.2% HCl and finally with acetone. Pellets were then allowed to air dry, after which they were dissolved in 0.03 ml sterile water and snap frozen histone protein extract was stored at -80°C. Histone protein concentrations were measured according to manufacturer instructions using a Qubit Protein Assay Kit (Invitrogen, Cat No. Q33212).
Detection of Global Post-translational Histone Modifications Levels
Detection of global PTHMs levels The Active Motif Histone H3 PTM Multiplex Kit (Cat No. 33115) was used to measure global changes in PTHM levels. the Histone H3 Total Ab-conjugated beads (Cat No. 33116) were multiplexed with the Histone PTM Ab-conjugated beads (H3K9ac Cat No. 33117, H3K4me3 Cat No. 33121, H3K9me2 Cat No. 33119, and H3K27me3 Cat No. 33125) for normalization of Histone H3 levels between different samples. A total of 150 ng of purified histones were used for each assay. Data were collected using a FLEXMAP3D instrument and analyzed using xPONENT software (Luminex Corporation). Data were normalized to show the relative amount of each PTHM in different samples.
Statistical Analyses
Based on our prior repeated observations of sex-dependent differences in epigenetic marks (Cory-Slechta et al., 2004, 2010; Weston et al., 2014; Schneider et al., 2016), all statistical analyses were carried out separately by sex. For assessment of PTHM levels, one way analyses of variance (ANOVAs) were carried out with behavioral experience as a between groups factor (FI, FS-RS, no experience), followed by post hoc student’s t-test method tests as appropriate. Multivariate analyses were carried out to examine correlations of the global levels of PTHM marks within and across FC and HIPP. To determine whether the correlational structure changed significantly in response to behavioral experience conditions, chi square analyses were carried out across the three behavioral experience conditions, comparing the frequency of statistically significant vs. non-significant p-values. In addition, multivariate correlation analyses were carried out between measures of FI and of FS-RS behavioral performance with levels of global PTHMs. These were analyzed separately by sex and behavioral experience condition. For all analyses, a p ≤ 0.05 was considered statistically significant.
Results
Global PTHMs Differ in Response to Behavioral Experience: Sex and Brain-Region Dependence
Global PTHM levels changed in response to behavioral experience conditions and did so differentially by brain region for each sex as shown in Figure 1. Male-specific differences in global PTHM levels in FC in relation to behavioral experience conditions were found for H3K27me3 [F(2,14) = 4.43, p = 0.036] which reflected the higher levels of this mark in mice that had FI behavioral experience (p = 0.0124), compared to those with FS-RS behavioral experience, with a similar trend (p = 0.061) evident for H3K9ac in males. No significant differences were found for females. In HIPP, differences were particularly pronounced in females, with marked reductions (30–49%) in levels of all 4 PTHMs in groups that had either FI or FS-RS experiences [H3K9ac = F(2,14) = 7.75, p = 0.008; post hoc t-test p-values of 0.0061 and 0.0045, respectively, H3K4me3 = F(2,14) = 4.38, p = 0.04], with reductions of 46–49% (p = 0.02 and 0.028, respectively, from non-behavioral control); H3K9me2 = [F(2,14) = 8.55, p = 0.006; p = 0.003 and 0.005, respectively, H3K27me3 = F(2,14) = 9.12, p = 0.005]. In males, the trend in HIPP was for reduced levels (approximately 10–48%) in the FI behavioral experience groups relative to both non-behavioral and FS-RS behavioral experience groups for 3 of the 4 PTHMs examined, i.e., the opposite pattern of what was observed in FC. These trends in HIPP approached significance for H3K27me3 [F(2,14) = 3.81, p = 0.052] and for H3K9me2 [F(2,14) = 3.41, p = 0.067].
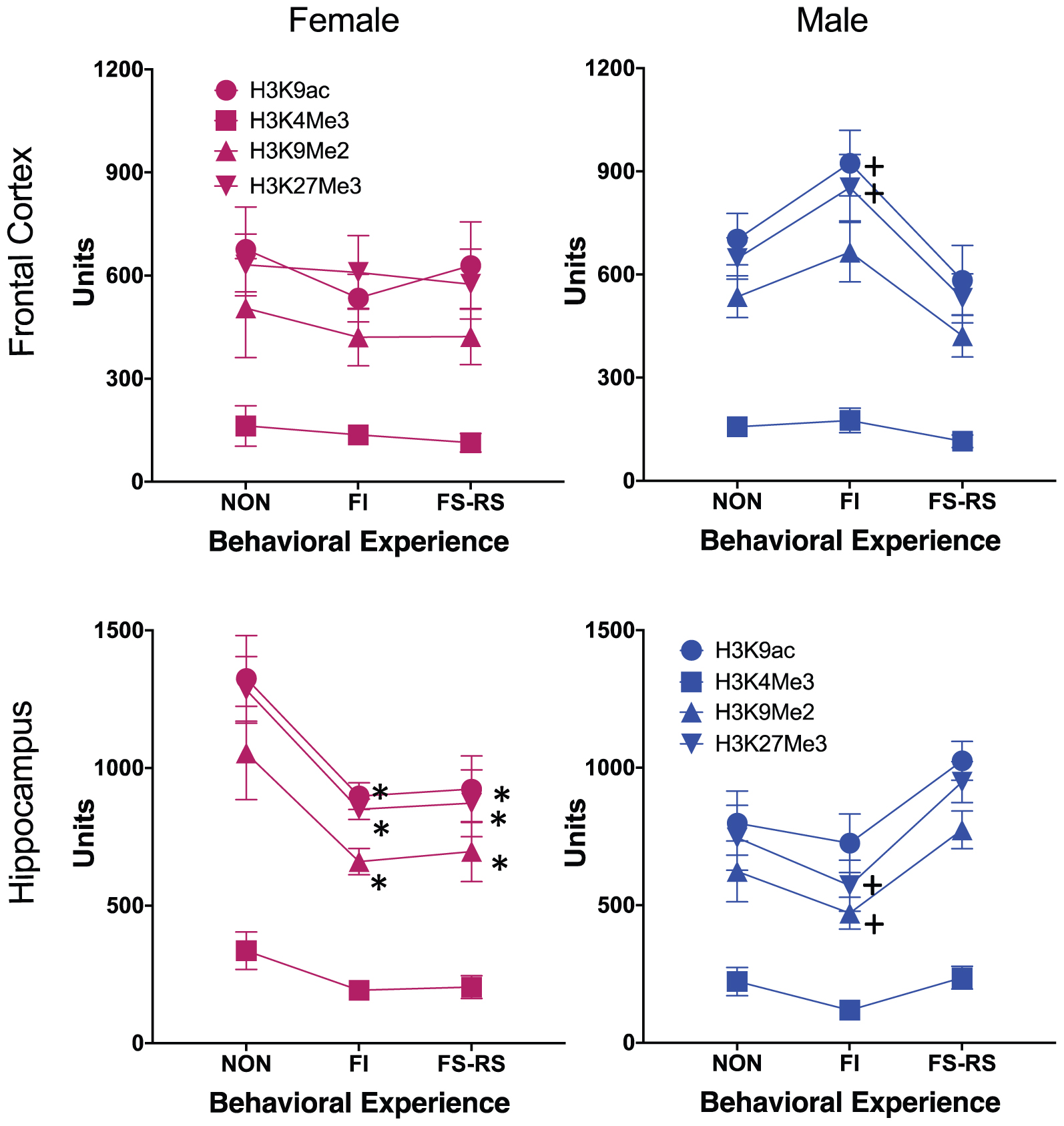
FIGURE 1. Group mean ± SE values for relative enrichment of the indicated global post-translational histone modification in frontal cortex and hippocampus (HIPP) of males and females in relation to the type of behavioral experience. Non, no behavioral experience group; POS, positive behavioral experience group; NEG, negative behavioral experience group (n = 12/group). ∗ significantly different from no behavior group; + significantly different from positive behavior group. + indicates significant difference from negative behavior group.
Correlations of Global PTHM Levels Within and Across Brain Regions in Relation to Behavioral Experience Condition
Figure 1 suggested that global levels of these 4 PTHM generally changed in parallel in response to a specific behavioral experience. Consequently, correlations of global PTHM levels within and across brain regions were examined in relation to behavioral experience conditions separately for each sex (Figure 2). Both within and across region correlations differed by behavioral experience for each sex.
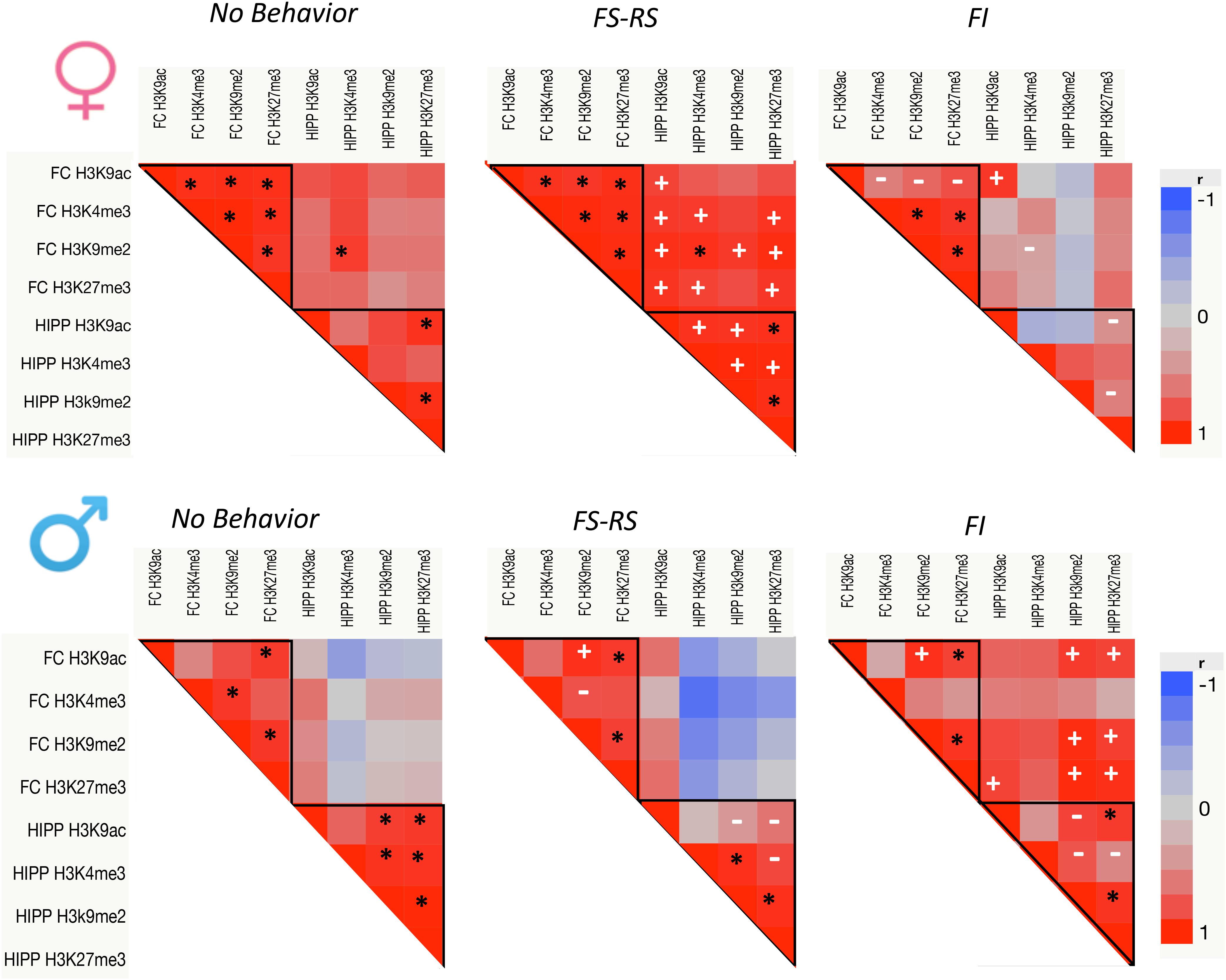
FIGURE 2. p-Value heat maps from multivariate correlation analyses across all PTHM levels in both FC and HIPP for each behavioral experience group of females and males (n = 5 per group). Black asterisks show significant correlations preserved across all three behavioral conditions; white plus marks show new correlations within the positive or negative behavioral experience groups; white dashes indicate significant correlations that were lost following either positive or negative behavioral experience relative to no behavior control.
For females (top row), under conditions of no behavioral experience, correlations of PTHMs largely occurred within a brain region, i.e., within FC and within HIPP regions, with only a single correlation across regions. Specifically, correlations between PTHMs were more pronounced within FC (r range = 0.92–0.995) with a more modest correlation profile within HIPP, and with a single inter FC-HIPP correlation. Following FS-RS experience, intra FC global PTHM correlations remained as in the no behavior conditions, while this experience significantly enhanced the number of within-HIPP PTHM level correlations (intra-FC correlations r range = 0.92–0.98 and intra-HIPP r range = 0.93–0.987). FS-RS experience also markedly increased the numbers of inter FC-HIPP correlations (r range = 0.865–0.974 for PTHMs). After FI behavioral experience, the number of within FC global PTHM correlations was reduced relative to no behavior control, particularly for H3K9ac, and within HIPP global PTHM correlations were eliminated. While the one inter FC-HIPP correlation seen under no behavioral experience conditions was also eliminated after FI experience, a new correlations between FC and HIPP global H3K9ac emerged. Chi-square analyses confirmed that the frequency of significant correlations of PTHM levels differed significantly in relation to behavioral experience (x2 = 48.98, p < 0.0001) with all three groups differing significantly from each other in frequency.
In males (bottom row) with no behavioral experience, global PTHM correlations were found within both HIPP and FC (r range = 0.887–0.996), but no inter FC-HIPP correlations occurred. After FS-RS behavioral experience, numbers of global PTHM level correlations were generally retained within FC, albeit in a different profile, while global PTHM correlations within HIPP were diminished relative to the no behavior condition, and no significant inter FC-HIPP correlations were found. After FI behavioral experience, within FC global PTHM correlation profiles were analogous to those seen after FS-RS experience, while within HIPP correlations were reduced. FI experience produced a marked increase in inter FC-HIPP correlations particularly between HIPP H3K9me2 and H3K27me3 with global levels of FC H3K9ac, H3K9me2 and H3K27me3. Chi-square analyses of global PTHM changes indicated a trend toward significant differences in the frequency of significant correlations by behavioral condition (x2 = 4.56, p < 0.102) which reflected the difference between the FI vs. RS-FS behavior groups.
Correlations of Global PTHM Levels With Measures of Behavioral Experience
To further support the behavioral relevance of global PTHM measures, examination of correlations of global PTHM levels with measures of behavioral performance were carried out. Multiple significant correlations were found with very high r2-values. For example, in females (Figure 3), strong correlations between HIPP PTHM H3K9me2 occurred with multiple measures of performance on the FI schedule, particularly during the early sessions (period of learning of the FI schedule parameters). Specifically, higher overall rates and run rates on the FI schedule were associated with lower levels of HIPP H3K9me2, and correspondingly with shorter IRTs and PRP times. r2-values for these correlations were notable, and ranged from 0.72 to 0.97. Measures of FI performance in males (Figure 4) correlated with global PTHM levels in both FC (middle row) and HIPP (bottom row). Specifically, increasing levels of H3K9ac, H3K9me2, and H3K27me3 in both FC and HIPP were correlated with higher overall response rates on the FI schedule, with r2-values that ranged from 0.82 to 0.95.
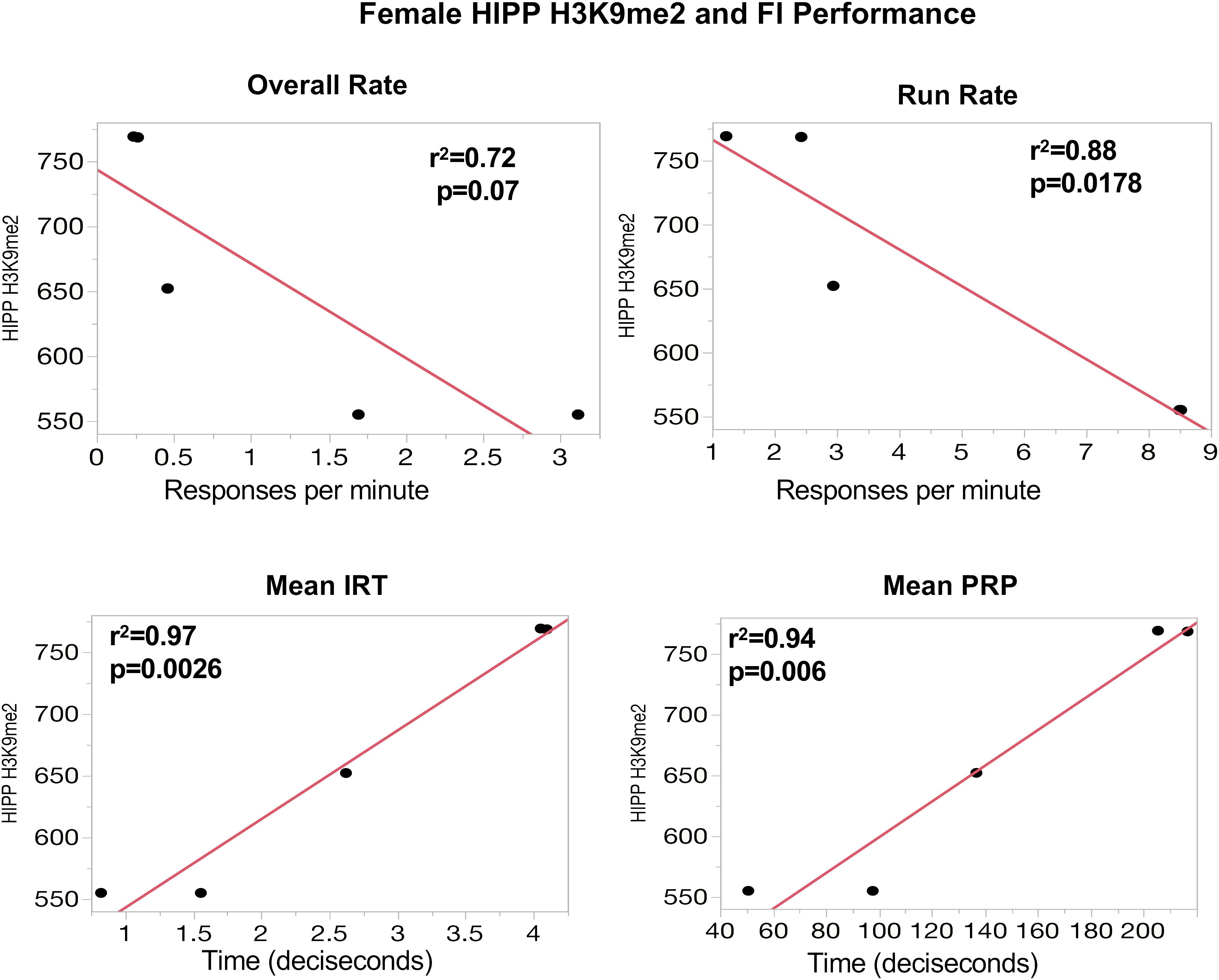
FIGURE 3. (Top) Correlations between measures of FI performance in session block 6–10 and global HIPP levels of H3K9me2 in females. The y-axis depicts levels of HIPP H3K9me2 expression and the x-axis depicts response rate values (responses/minute) for overall rate and run rate, or time (in deciseconds) for mean interresponse time (IRT) or postreinforcement pause (PRP) as indicated. The red line through each plot depicts the linear fit, and r2- and p-values are indicated for each plot.
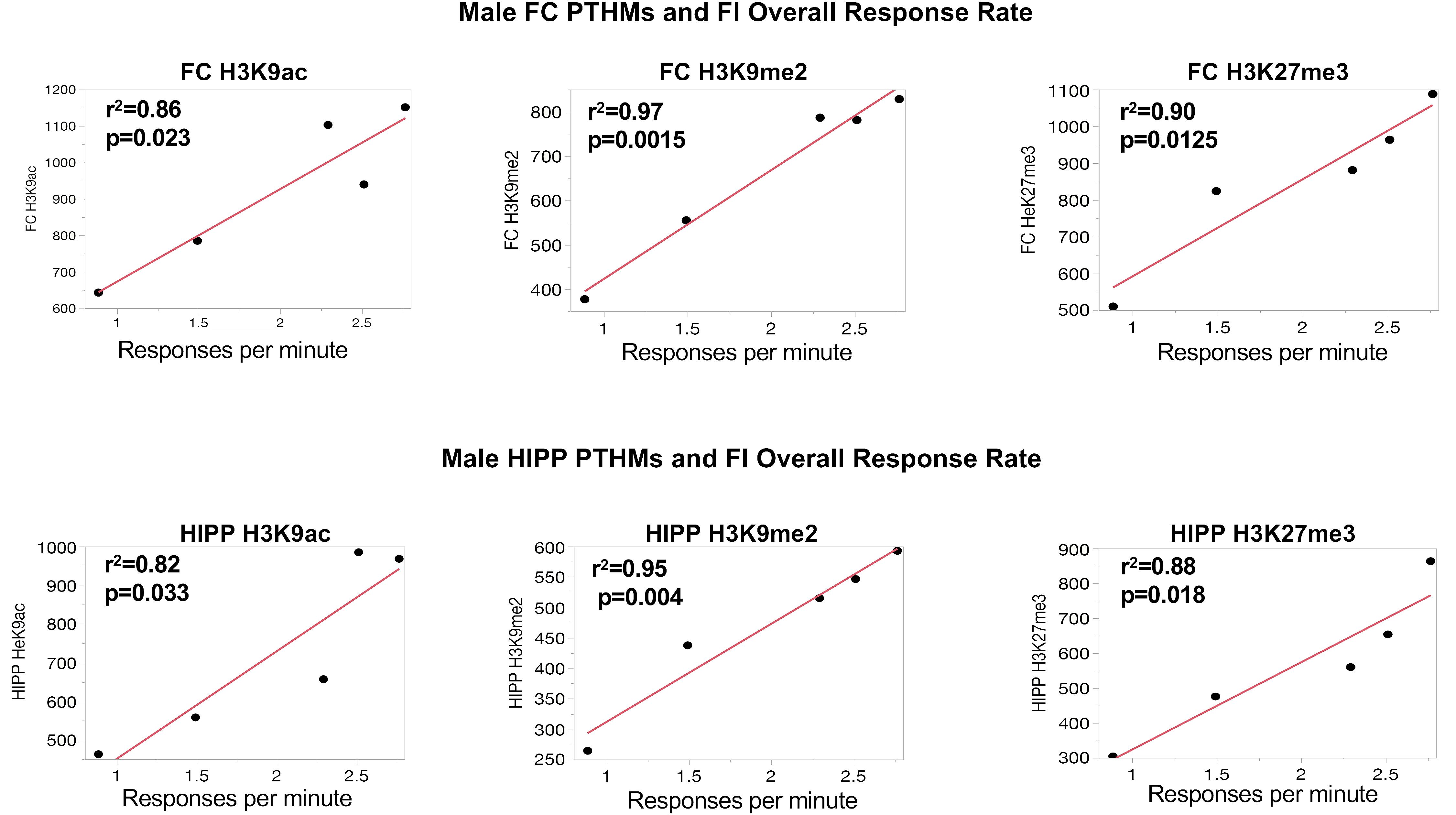
FIGURE 4. Correlations between measures of FI overall response rates and global levels of H3K9ac, H3K9me2, and H3K27me3 in FC and HIPP, respectively. The red line through each plot depicts the linear fit, and r2- and p-values are indicated for each plot.
Similarly, correlations between global PTHM levels and measures of FS-RS were observed, although of marginal significance. Figure 5 depicts correlations between FC PTHMs and FS-RS performance, with increases in total float duration times marginally correlated with reductions in levels of FC PTHMs H3K27me3, H3K4me3, and H3K9me2, with r2-values ranging from 0.69 to 0.74. In the case of males (Figure 6), a marginally significant correlation was observed in which increases in mean float duration were associated with reductions in HIPP PTHM H3K27me3 with an r2-value of 0.73; similar trends were evident for other PTHMs in both males and females.
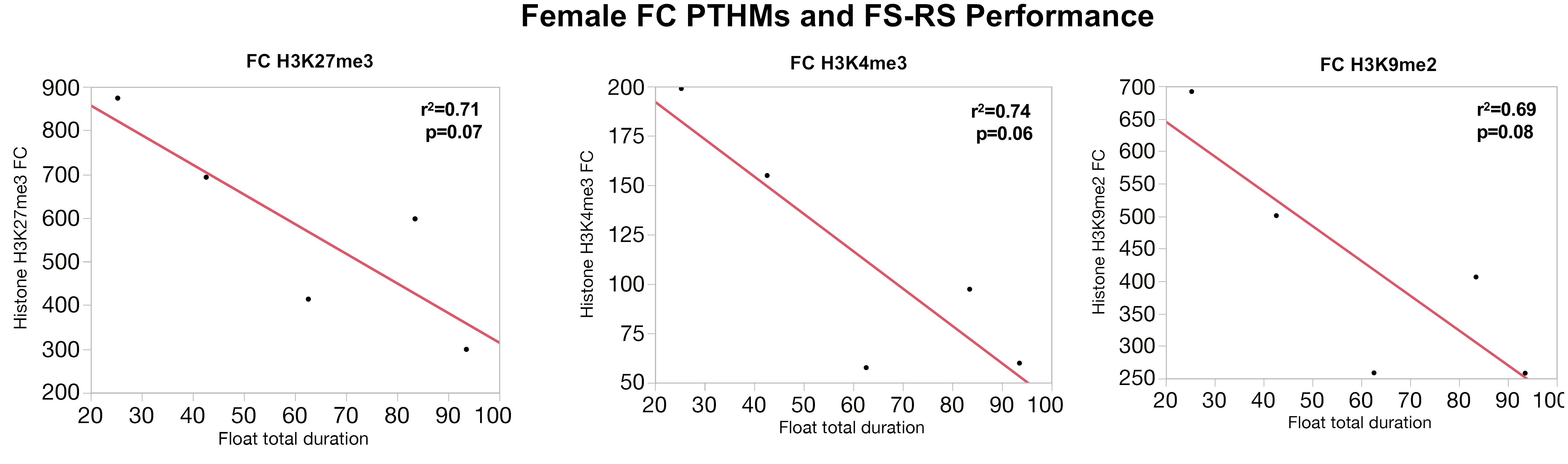
FIGURE 5. Correlations between measures of total duration floating in the FS paradigm and global levels of FC H3K27me3, H3K4me3, and H3K9me2 in females. The red line through each plot depicts the linear fit, and r2- and p-values are indicated for each plot.
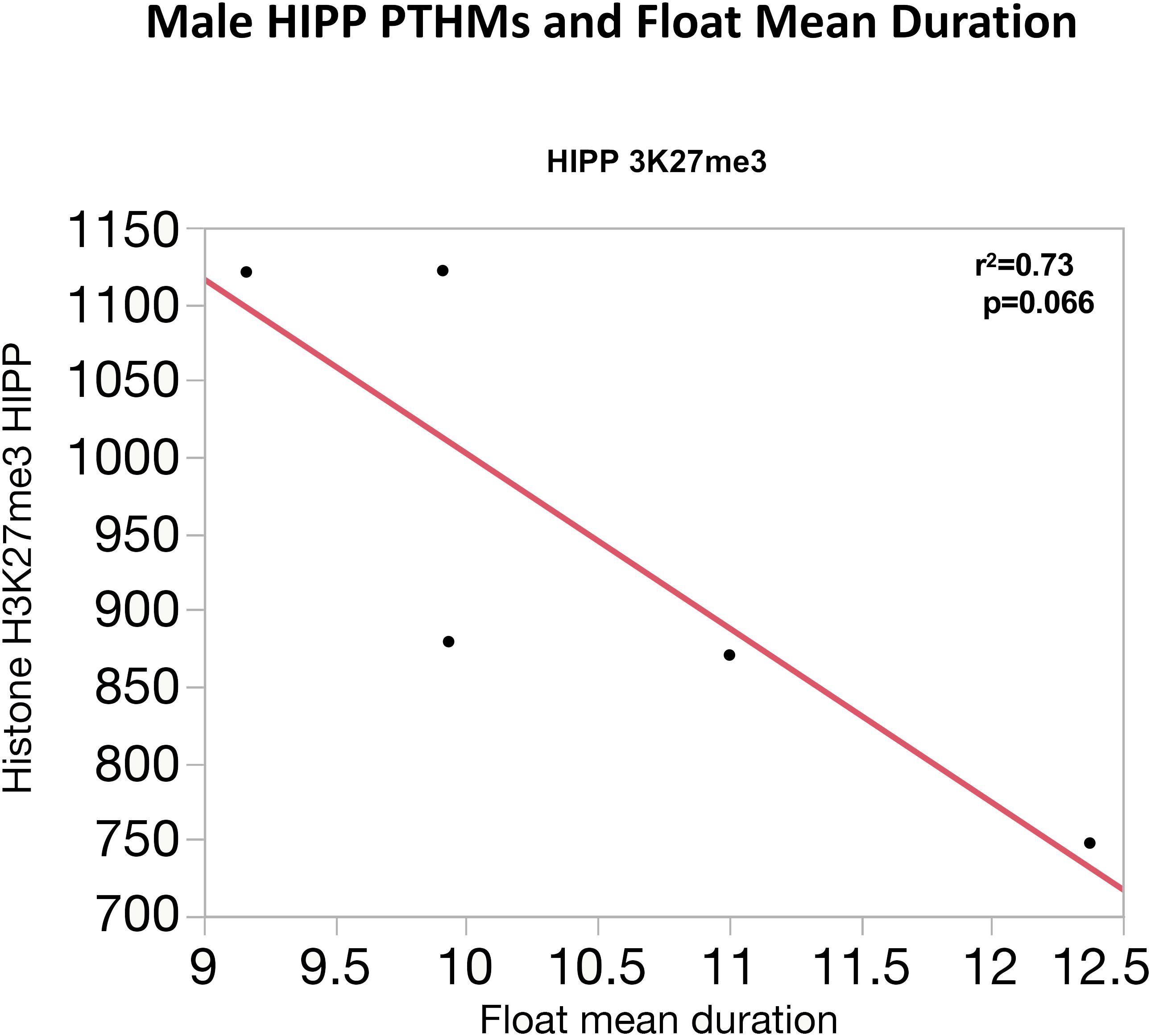
FIGURE 6. Correlation between mean floating duration time in the FS paradigm and global levels of HIPP H3K27me3 in males. The red line through each plot depicts the linear fit, and r2- and p-values are indicated for each plot.
Discussion
This study examined the influence of contrasting behavioral experiences, specifically food-rewarded learning performance on an FI schedule and single episodes of two different and salient stressors, forced swim followed by restraint stress, or to no explicit behavioral experience, on FC and hippocampal global levels of two global activating (H3K9ac and H3K4me3) and two global repressive (H3K9me2 and H3K27me3) PTHMs. Three overall findings from this study deserve mention, two of which were quite unexpected. First, the specific type of behavioral experience was shown to be a critical determinant of the subsequent global PTHM profiles, and these evidenced both brain region and sex differences. Second, these global PTHMs appeared to be changing in parallel in relation to the specific behavioral experience, i.e., with similar effects on levels of all 4 PTHMs in each behavioral condition. Third, even though these were global levels of histone modifications (and thus not indicative of changes at specific genes or sites on genes), they nevertheless showed robust correlations with measures of behavioral performance.
The results clearly demonstrate the importance of behavioral context and experience to subsequent PTHM modifications. For example, males showed trends toward increases in PTHM levels in FC, but decreases in HIPP after FI experience, whereas changes evident after FS-RS behavioral experience were not significant (Figure 1). In females, differences in global PTHM levels were restricted to HIPP, and comprised of equivalent reductions of PTHM levels in groups with either FI or FS-RS experience. Reports of direct comparisons in PTHM levels following differing behavioral experiences, and/or comparisons to a no-behavioral control group are limited to date. In one such study that did involve such comparisons in bees, systemic administration of HDAC inhibitors impaired olfactory discrimination memory for an aversive stimulus, but not for a rewarding odor, effects shown not to reflect differences in sucrose sensitivity or locomotor activity (Lockett et al., 2014). Although based on different outcome measures, both were measures of learning and show that the nature of the behavioral experience was important to the consequent epigenetic outcome. Another reported study (Bousiges et al., 2010) involved rats that had either undergone water maze learning that included a constant location visible platform for escape was possible or a water maze in which the platform was relocated for each trial. Findings showed increases in dorsal hippocampal global H2B and H4 acetylation levels in the hidden platform condition, but not the relocated platform condition., while an additional experiment revealed markedly increased H3 acetylation after the variable platform experience in comparison to levels obtained from rats that remained in their home cage.
The current findings also show clear brain region and sex-dependent differences in epigenetic profiles following behavioral experience, confirming many previous reports (McCarthy and Nugent, 2015). For example, exposure to subchronic variable stress in rodents resulted in significant sex-specific differences in Dnmt3a gene expression levels in nucleus accumbens (Hodes et al., 2015). Genome-wide assessment of H3K4me3 in adult mice revealed differences in its expression by sex in 248 genes and loci (Shen et al., 2015). Prenatal stress differentially altered gene expression and epigenetic regulation in HIPP and FC of rats in a sex-dependent manner (Van den Hove et al., 2013). Further, levels of histone modifications were found to differ by sex in neonatal mouse brain (Tsai et al., 2009). Thus, the sex differences observed here both confirm and extend those findings.
With respect to brain region differences, studies to date have often focused on single brain regions. Yet, regionally specific responses in histone marks to behavioral experience are increasingly being identified. For example, in an object recognition task, PTHM level profiles and activity over time differed significantly in HIPP and prefrontal cortex in mice, as did expression of the learning-related gene zif268 (Graff et al., 2012). Furthermore, in a report consistent with our findings, levels of histones H2B, H3, and H4 acetylation were found to differ both within region and across regions (HIPP and dorsal striatum) in mice trained in spatial vs. cued water maze learning (Dagnas and Mons, 2013).
Remarkably, global PTHM levels appeared to be changing in parallel in relation to behavioral experience, i.e., with generally similar effects on levels of all 4 PTHMs within a brain region in each behavioral condition. Furthermore, the significance of those changes was clearly demonstrated by their very robust correlations with measures of behavioral performance. Such findings demonstrate that both total levels of histone modifications, in addition to studies that suggest that effects are gene/loci specific (Tsankova et al., 2004; Park et al., 2017), are needed to understand the complex suite of epigenetic shifts occurring following behavioral experience. It seems intuitive that the understanding behavioral function, which involves the integrated and immediate activity of networks of regions within the brain, requires an iterative research approach of integrative/higher-order and gene-specific measures. Studies show brain network connectivity to be a central feature of its operation that can actually be predicted by clustered gene expression profiles across regions (Greenspan, 2001; French and Pavlidis, 2011; Wolf et al., 2011; Ji et al., 2014; Whitney et al., 2014; Hawrylycz et al., 2015; Richiardi et al., 2015; Fulcher and Fornito, 2016; Xu et al., 2016; Forest et al., 2017). These gene clusters may be best represented by a global or total measurement of histone modifications across the genome (Nesbitt et al., 2014). Epigenetic changes, even while differing at local sites, nevertheless occur simultaneously in multiple brain regions, as shown by in vivo imaging of HDACs in humans (Wey et al., 2016). In fact, our findings of robust correlations of behavioral performance with measures of global PTHMs are similar to a previous report of correlations of locomotor activity with global levels of hippocampal acetylation which differentiated animals exhibiting low vs. high levels of activity. Here too correlations were observed with global PTHMs, albeit not generally as robust as those shown here, likely reflecting the fact that brains were obtained 4 months after behavioral assessment (Nesbitt et al., 2014). One implication of our findings would be that such global histone marks could serve as “biomarkers” or indicators (Kubota, 2016) of past behavioral experience which could provide a far more cost-effective approach to delineating the full spectrum of individual PTHMs by gene, region and cell type (Henikoff and Shilatifard, 2011).
One question raised by these findings is what role such global changes could play in brain. Nesbitt et al. (2014) suggested that such global changes may represent a potential mechanism to regulate cellular plasticity in brain, and further, that reports identifying gene-specific changes in chromatin marks could be due to primary or upstream changes in total levels of chromatin marks that then facilitate the progression or trajectory of gene expression programs. Extending this idea, a higher order level of control is suggested, i.e., a type of “pattern recognition” by the brain controlling gene expression, as ultimately the brain must integrate more localized epigenetic changes across systems/networks. Evidence for importance of global histone marks in predicting multiple types of cancer, as well as its progression and response to chemotherapy is already well established (Ebrahimi et al., 2013; Karczmarski et al., 2014; Ellinger et al., 2016; Zhao et al., 2016; Ngollo et al., 2017).
The current findings also raise questions as to what specific characteristics/processes of behavioral experiences differentiate their epigenetic consequences, and whether similar behavioral domains (e.g., learning of any kind) produce corresponding profiles of epigenetic changes. To date, studies of behavioral epigenetics have often tended to focus on single behaviors (e.g., learning, drug abuse, or stress) and frequently examine that behavior using a very limited range of paradigms. For example, studies of epigenetic coding of learning/memory have primarily relied on the use of fear conditioning (Day and Sweatt, 2011; Sweatt, 2016). This raises the question of whether reported epigenetic changes in such studies are purely “learning”-related, i.e., generalizable across different learning paradigms, or reflect specific components of the paradigm used, e.g., shock-based learning. Would positive reinforcement-based learning paradigms produce the same epigenetic profiles as negative reinforcement based learning paradigms? One limitation of our study was the differences in timing between the end of behavioral testing and consequent determination of PTHMs, which was longer in the FS-RS as compared to the FI behavioral experience group by 1 week, providing an opportunity for more PTHM changes in that condition post-testing. This could account for the lesser evidence for correlations between global PTHM and FS-RS experience as compared to FI-based correlations. Even with this limitation, however, overall behavioral experience conditions of the groups were markedly different and still evidenced in differences in global PTHM levels and in correlations of these global PTHMs with behavior. Further, correlations between global acetylation levels and locomotor activity survived a 4 months post-behavioral assessment time frame in the study by Nesbitt et al. (2014).
These global differences in PTHM profiles would seem to be consistent with the potential for differential types of networked activity following FI vs. FS-RS vs. no behavioral experience. While it might be asserted that some within-animal correlations would certainly be expected, this seems an unlikely explanation for the patterns observed here for at least two reasons. First, it would not explain why all PTHMs were not uniformly or close to uniformly correlated. Nor would it explain the fact that marked differences in the PTHM correlation profiles occurred in response to different types of behavioral experience. Future studies of gene specific genome wide and transcriptomic effects, using ChiP-sequencing and RNA sequencing methods, of these behavioral experiences will be required to fully elucidate the influence of these behaviors on the neural connectome at a gene-specific level. Further, while recognizing that correlational analyses does not recapitulate direct molecular mechanistic function or indicate causation, advancing the understanding of behavioral epigenetics will also require systematic assessment of network-based outcomes to answer the questions related to how brain ultimately integrates the localized molecular changes to particular patterns of behavioral experiences.
Author Contributions
DC-S, JS, and MS conceived the experiments. MS and DC-S oversaw the behavioral assessments and all statistical analyses and GS and JS contributed in histone determinations. DC-S wrote the manuscript with input from JS, MS, and GS.
Funding
This work was supported in part by R01 ES021534 (Cory-Slechta; Schneider, JS multi-PIs) and by P30 ES001247 (Cory-Slechta, P.I.).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Babb, J. A., Masini, C. V., Day, H. E., and Campeau, S. (2014). Habituation of hypothalamic-pituitary-adrenocortical axis hormones to repeated homotypic stress and subsequent heterotypic stressor exposure in male and female rats. Stress 17, 224–234. doi: 10.3109/10253890.2014.905534
Barnett, W. S. (1995). Long-term effects of early childhood programs on cognitive and school outcomes. Future Child 5, 25–50. doi: 10.2307/1602366
Barrett, J. E. (1986). “Behavioral history: residual influences on subsequent behavior and drug effects,” in Developmental Behavioral Pharmacology, ed. T. Thompson (Hillsdale, NJ: Lawrence Erlbaum Associates), 99–114.
Borghol, N., Suderman, M., Mcardle, W., Racine, A., Hallett, M., Pembrey, M., et al. (2012). Associations with early-life socio-economic position in adult DNA methylation. Int. J. Epidemiol. 41, 62–74. doi: 10.1093/ije/dyr147
Bousiges, O., Vasconcelos, A. P. D., Neidl, R., Cosquer, B., Herbeaux, K., Panteleeva, I., et al. (2010). Spatial memory consolidation is associated with induction of several lysine-acetyltransferase (histone acetyltransferase) expression levels and H2B/H4 acetylation-dependent transcriptional events in the rat hippocampus. Neuropsychopharmacology 35, 2521–2537. doi: 10.1038/npp.2010.117
Brito, N. H., and Noble, K. G. (2014). Socioeconomic status and structural brain development. Front. Neurosci. 8:276. doi: 10.3389/fnins.2014.00276
Byrd, L. D. (1972). Responding in the squirrel monkey under second-order schedules of shock delivery. J. Exp. Anal. Behav. 18, 155–167. doi: 10.1901/jeab.1972.18-155
Chiarella, J., Tremblay, R. E., Szyf, M., Provençal, N., and Booij, L. (2015). Impact of early environment on children’s mental health: lessons from DNA methylation studies with monozygotic twins. Twin Res. Hum. Genet. 18, 623–634. doi: 10.1017/thg.2015.84
Chugani, H. T., Behen, M. E., Muzik, O., Juhász, C., Nagy, F., and Chugani, D. C. (2001). Local brain functional activity following early deprivation: a study of postinstitutionalized romanian orphans. Neuroimage 14, 1290–1301. doi: 10.1006/nimg.2001.091712
Cory-Slechta, D. A., O’mara, D. J., and Brockel, B. J. (1998). Nucleus accumbens dopaminergic mediation of fixed interval schedule-controlled behavior and its modulation by low-level lead exposure. J. Pharm. Exp. Ther. 286, 794–805.
Cory-Slechta, D. A., Pazmino, R., and Bare, C. (1997). The critical role of the nucleus accumbens dopamine systems in the mediation of fixed interval schedule-controlled operant behavior. Brain Res. 764, 253–256. doi: 10.1016/S0006-8993(97)00591-X
Cory-Slechta, D. A., Pokora, M. J., and Preston, R. A. (1996). The effects of dopamine agonists on fixed interval schedule-controlled behavior are selectively altered by low level lead exposure. Neurotoxicol. Teratol. 18, 565–575. doi: 10.1016/0892-0362(96)00082-7
Cory-Slechta, D. A., Stern, S., Weston, D., Allen, J. L., and Liu, S. (2010). Enhanced learning deficits in female rats following lifetime Pb exposure combined with prenatal stress. Toxicol. Sci. 117, 427–438. doi: 10.1093/toxsci/kfq221
Cory-Slechta, D. A., Virgolini, M. B., Thiruchelvam, M., Weston, D. D., and Bauter, M. R. (2004). Maternal stress modulates the effects of developmental lead exposure. Environ. Health Perspect. 112, 717–730. doi: 10.1289/ehp.6481
Cory-Slechta, D. A., Weiss, B., and Cox, C. (1985). Performance and exposure indices of rats exposed to low concentrations of lead. Toxicol. Appl. Pharmacol. 78, 291–299. doi: 10.1016/0041-008X(85)90292-3
Dagnas, M., and Mons, N. (2013). Region- and age-specific patterns of histone acetylation related to spatial and cued learning in the water maze. Hippocampus 23, 581–591. doi: 10.1002/hipo.22116
Day, J. J., Childs, D., Guzman-Karlsson, M. C., Kibe, M., Moulden, J., Song, E., et al. (2013). DNA methylation regulates associative reward learning. Nat. Neurosci. 16, 1445–1452. doi: 10.1038/nn.3504
Day, J. J., and Sweatt, J. D. (2011). Epigenetic mechanisms in cognition. Neuron 70, 813–829. doi: 10.1016/j.neuron.2011.05.019
Ebrahimi, A., Schittenhelm, J., Honegger, J., and Schluesener, H. (2013). Prognostic relevance of global histone 3 lysine 9 acetylation in ependymal tumors. J. Neurosurg. 119, 1424–1431. doi: 10.3171/2013.9.JNS13511
Ellinger, J., Schneider, A. C., Bachmann, A., Kristiansen, G., Muller, S. C., and Rogenhofer, S. (2016). Evaluation of global histone acetylation levels in bladder cancer patients. Anticancer Res. 36, 3961–3964.
Forest, M., Iturria-Medina, Y., Goldman, J. S., Kleinman, C. L., Lovato, A., Oros Klein, K., et al. (2017). Gene networks show associations with seed region connectivity. Hum. Brain Mapp. 38, 3126–3140. doi: 10.1002/hbm.23579
Fraga, M. F., Ballestar, E., Paz, M. F., Ropero, S., Setien, F., Ballestar, M. L., et al. (2005). Epigenetic differences arise during the lifetime of monozygotic twins. Proc. Natl. Acad. Sci. U.S.A. 102, 10604–10609. doi: 10.1073/pnas.0500398102
French, L., and Pavlidis, P. (2011). Relationships between gene expression and brain wiring in the adult rodent brain. PLoS Comput. Biol. 7:e1001049. doi: 10.1371/journal.pcbi.1001049
Fulcher, B. D., and Fornito, A. (2016). A transcriptional signature of hub connectivity in the mouse connectome. Proc. Natl. Acad. Sci. U.S.A. 113, 1435–1440. doi: 10.1073/pnas.1513302113
Galler, J., and Rabinowitz, D. G. (2014). “Chapter seven - the intergenerational effects of early adversity,” in Progress in Molecular Biology and Translational Science, eds A. Schahram and L. Farah (Cambridge, MA: Academic Press), 177–198.
Graff, J., Woldemichael, B. T., Berchtold, D., Dewarrat, G., and Mansuy, I. M. (2012). Dynamic histone marks in the hippocampus and cortex facilitate memory consolidation. Nat. Commun. 3:991. doi: 10.1038/ncomms1997
Guillemin, C., Provencal, N., Suderman, M., Cote, S. M., Vitaro, F., Hallett, M., et al. (2014). DNA methylation signature of childhood chronic physical aggression in T cells of both men and women. PLoS One 9:e86822. doi: 10.1371/journal.pone.0086822
Hair, N. L., Hanson, J. L., Wolfe, B. L., and Pollak, S. D. (2015). Association of child poverty, brain development, and academic achievement. JAMA Pediatr. 169, 822–829. doi: 10.1001/jamapediatrics.2015.1475
Hawrylycz, M., Miller, J. A., Menon, V., Feng, D., Dolbeare, T., Guillozet-Bongaarts, A. L., et al. (2015). Canonical genetic signatures of the adult human brain. Nat. Neurosci. 18, 1832–1844. doi: 10.1038/nn.4171
Henikoff, S., and Shilatifard, A. (2011). Histone modification: cause or cog? Trends Genet. 27, 389–396. doi: 10.1016/j.tig.2011.06.006
Hirase, H., and Shinohara, Y. (2014). Transformation of cortical and hippocampal neural circuit by environmental enrichment. Neuroscience 280, 282–298. doi: 10.1016/j.neuroscience.2014.09.031
Hodes, G. E., Pfau, M. L., Purushothaman, I., Ahn, H. F., Golden, S. A., Christoffel, D. J., et al. (2015). Sex differences in nucleus accumbens transcriptome profiles associated with susceptibility versus resilience to subchronic variable stress. J. Neurosci. 35, 16362–16376. doi: 10.1523/JNEUROSCI.1392-15.2015
Ji, S., Fakhry, A., and Deng, H. (2014). Integrative analysis of the connectivity and gene expression atlases in the mouse brain. Neuroimage 84, 245–253. doi: 10.1016/j.neuroimage.2013.08.049
Johnson, R., Browne, K., and Hamilton-Giachritsis, C. (2006). Young children in institutional care at risk of harm. Trauma Violence Abuse 7, 34–60. doi: 10.1177/1524838005283696
Karczmarski, J., Rubel, T., Paziewska, A., Mikula, M., Bujko, M., Kober, P., et al. (2014). Histone H3 lysine 27 acetylation is altered in colon cancer. Clin. Proteomics 11:24. doi: 10.1186/1559-0275-11-24
Kelly, ÁM. (2015). “Non-pharmacological approaches to cognitive enhancement,” in Cognitive Enhancement, eds K. M. Kantak and J. G. Wettstein (Cham: Springer), 417–439.
Kondo, M. (2017). Molecular mechanisms of experience-dependent structural and functional plasticity in the brain. Anat. Sci. Int. 92, 1–17. doi: 10.1007/s12565-016-0358-6
Kubota, T. (2016). Epigenetic alterations induced by environmental stress associated with metabolic and neurodevelopmental disorders. Environ. Epigenetics 2, dvw017–dvw017. doi: 10.1093/eep/dvw017
Lockett, G., Wilkes, F., Helliwell, P., and Maleszka, R. (2014). Contrasting effects of histone deacetylase inhibitors on reward and aversive olfactory memories in the honey bee. Insects 5, 377–398. doi: 10.3390/insects5020377
Marchner, J. R., and Preuschhof, C. (2018). Reward history but not search history explains value-driven attentional capture. Atten. Percept. Psychophys. doi: 10.3758/s13414-018-1513-z [Epub ahead of print].
McCarthy, M. M., and Nugent, B. M. (2015). At the frontier of epigenetics of brain sex differences. Front. Behav. Neurosci. 9:221. doi: 10.3389/fnbeh.2015.00221
McGowan, P. O., Sasaki, A., D’alessio, A. C., Dymov, S., Labonte, B., Szyf, M., et al. (2009). Epigenetic regulation of the glucocorticoid receptor in human brain associates with childhood abuse. Nat. Neurosci. 12, 342–348. doi: 10.1038/nn.2270
McInnis, C. M., Wang, D., Gianferante, D., Hanlin, L., Chen, X., Thoma, M. V., et al. (2015). Response and habituation of pro- and anti-inflammatory gene expression to repeated acute stress. Brain Behav. Immun. 46, 237–248. doi: 10.1016/j.bbi.2015.02.006
Nesbitt, A. M., Mccurdy, R. D., Bryant, S. M., and Alter, M. D. (2014). Total levels of hippocampal histone acetylation predict normal variability in mouse behavior. PLoS One 9:e94224. doi: 10.1371/journal.pone.0094224
Ngollo, M., Lebert, A., Daures, M., Judes, G., Rifai, K., Dubois, L., et al. (2017). Global analysis of H3K27me3 as an epigenetic marker in prostate cancer progression. BMC Cancer 17:261. doi: 10.1186/s12885-017-3256-y
Nipa, H. F., Gottesman, I. I., and Wong, A. H. C. (2009). Not really identical: Epigenetic differences in monozygotic twins and implications for twin studies in psychiatry. Am. J. Med. Genet. C Semin. Med. Genet. 151C, 136–141. doi: 10.1002/ajmg.c.30206
Noble, K. G., Houston, S. M., Brito, N. H., Bartsch, H., Kan, E., Kuperman, J. M., et al. (2015). Family income, parental education and brain structure in children and adolescents. Nat. Neurosci. 18, 773–778. doi: 10.1038/nn.3983
Park, S. W., Lee, J. G., Seo, M. K., Ly, N. N., Lee, C. H., Cho, H. Y., et al. (2017). Epigenetic modification of glucocorticoid receptor promoter I7 in maternally separated and restraint-stressed rats. Neurosci. Lett. 650, 38–44. doi: 10.1016/j.neulet.2017.04.024
Pattij, T., Broersen, L. M., Peter, S., and Olivier, B. (2004). Impulsive-like behavior in differential-reinforcement-of-low-rate 36 s responding in mice depends on training history. Neurosci. Lett. 354, 169–171. doi: 10.1016/j.neulet.2003.10.004
Provençal, N., Suderman, M. J., Guillemin, C., Massart, R., Ruggiero, A., Wang, D., et al. (2012). The signature of maternal rearing in the methylome in rhesus macaque prefrontal cortex and T cells. J. Neurosci. 32, 15626–15642. doi: 10.1523/JNEUROSCI.1470-12.2012
Richiardi, J., Altmann, A., Milazzo, A. C., Chang, C., Chakravarty, M. M., Banaschewski, T., et al. (2015). BRAIN NETWORKS. Correlated gene expression supports synchronous activity in brain networks. Science 348, 1241–1244. doi: 10.1126/science.1255905
Romens, S. E., Mcdonald, J., Svaren, J., and Pollak, S. D. (2015). Associations between early life stress and gene methylation in children. Child Dev. 86, 303–309. doi: 10.1111/cdev.12270
Sadler, A. M., and Bailey, S. J. (2016). Repeated daily restraint stress induces adaptive behavioural changes in both adult and juvenile mice. Physiol. Behav. 167, 313–323. doi: 10.1016/j.physbeh.2016.09.014
Schneider, J. S., Anderson, D. W., Kidd, S. K., Sobolewski, M., and Cory-Slechta, D. A. (2016). Sex-dependent effects of lead and prenatal stress on post-translational histone modifications in frontal cortex and hippocampus in the early postnatal brain. Neurotoxicology 54, 65–71. doi: 10.1016/j.neuro.2016.03.016
Shen, E. Y., Ahern, T. H., Cheung, I., Straubhaar, J., Dincer, A., Houston, I., et al. (2015). Epigenetics and sex differences in the brain: a genome-wide comparison of histone-3 lysine-4 trimethylation (H3K4me3) in male and female mice. Exp. Neurol. 268, 21–29. doi: 10.1016/j.expneurol.2014.08.006
Sweatt, J. D. (2016). Neural plasticity and behavior – sixty years of conceptual advances. J. Neurochem. 139, 179–199. doi: 10.1111/jnc.13580
Szyf, M., and Bick, J. (2013). DNA methylation: a mechanism for embedding early life experiences in the genome. Child Dev. 84, 49–57. doi: 10.1111/j.1467-8624.2012.01793.x
Szyf, M., Tang, Y.-Y., Hill, K. G., and Musci, R. (2016). The dynamic epigenome and its implications for behavioral interventions: a role for epigenetics to inform disorder prevention and health promotion. Transl. Behav. Med. 6, 55–62. doi: 10.1007/s13142-016-0387-7
Tsai, H.-W., Grant, P. A., and Rissman, E. F. (2009). Sex differences in histone modifications in the neonatal mouse brain. Epigenetics 4, 47–53. doi: 10.4161/epi.4.1.7288
Tsankova, N. M., Kumar, A., and Nestler, E. J. (2004). Histone modifications at gene promoter regions in rat hippocampus after acute and chronic electroconvulsive seizures. J. Neurosci. 24, 5603–5610. doi: 10.1523/JNEUROSCI.0589-04.2004
Tung, J., Barreiro, L. B., Johnson, Z. P., Hansen, K. D., Michopoulos, V., Toufexis, D., et al. (2012). Social environment is associated with gene regulatory variation in the rhesus macaque immune system. Proc. Natl. Acad. Sci. U.S.A. 109, 6490–6495. doi: 10.1073/pnas.1202734109
Turecki, G., and Meaney, M. J. (2016). Effects of the social environment and stress on glucocorticoid receptor gene methylation: a systematic review. Biol. Psychiatry 79, 87–96. doi: 10.1016/j.biopsych.2014.11.022
Van den Hove, D. L. A., Kenis, G., Brass, A., Opstelten, R., Rutten, B. P. F., Bruschettini, M., et al. (2013). Vulnerability versus resilience to prenatal stress in male and female rats; Implications from gene expression profiles in the hippocampus and frontal cortex. Eur. Neuropsychopharmacol. 23, 1226–1246. doi: 10.1016/j.euroneuro.2012.09.011
van der Knaap, L. J., Riese, H., Hudziak, J. J., Verbiest, M. M., Verhulst, F. C., Oldehinkel, A. J., et al. (2014). Glucocorticoid receptor gene (NR3C1) methylation following stressful events between birth and adolescence. The TRAILS study. Transl. Psychiatry 4:e381. doi: 10.1038/tp.2014.22
Weder, N., Zhang, H., Jensen, K., Yang, B. Z., Simen, A., Jackowski, A., et al. (2014). Child abuse, depression, and methylation in genes involved with stress, neural plasticity, and brain circuitry. J. Am. Acad. Child Adolesc. Psychiatry 53, 417.e–424.e. doi: 10.1016/j.jaac.2013.12.025
Weston, H. I., Weston, D. D., Allen, J. L., and Cory-Slechta, D. A. (2014). Sex-dependent impacts of low-level lead exposure and prenatal stress on impulsive choice behavior and associated biochemical and neurochemical manifestations. Neurotoxicology 44, 169–183. doi: 10.1016/j.neuro.2014.06.013
Wey, H.-Y., Gilbert, T. M., Zürcher, N. R., She, A., Bhanot, A., Taillon, B. D., et al. (2016). Insights into neuroepigenetics through human histone deacetylase PET imaging. Sci. Transl. Med. 8, ra106–ra351. doi: 10.1126/scitranslmed.aaf7551
Whitney, O., Pfenning, A. R., Howard, J. T., Blatti, C. A., Liu, F., Ward, J. M., et al. (2014). Core and region-enriched networks of behaviorally regulated genes and the singing genome. Science 346:1256780. doi: 10.1126/science.1256780
Wolf, L., Goldberg, C., Manor, N., Sharan, R., and Ruppin, E. (2011). Gene expression in the rodent brain is associated with its regional connectivity. PLoS Comput. Biol. 7:e1002040. doi: 10.1371/journal.pcbi.1002040
Wong, C. C. Y., Caspi, A., Williams, B., Craig, I. W., Houts, R., Ambler, A., et al. (2010). A longitudinal study of epigenetic variation in twins. Epigenetics 5, 516–526. doi: 10.4161/epi.5.6.12226
Xu, J., Potenza, M. N., Calhoun, V. D., Zhang, R., Yip, S. W., Wall, J. T., et al. (2016). Large-scale functional network overlap is a general property of brain functional organization: reconciling inconsistent fMRI findings from general-linear-model-based analyses. Neurosci. Biobehav. Rev. 71, 83–100. doi: 10.1016/j.neubiorev.2016.08.035
Keywords: behavior, FI schedule, forced swim, restraint stress, global histone modification, frontal cortex, hippocampus, epigenetics
Citation: Sobolewski M, Singh G, Schneider JS and Cory-Slechta DA (2018) Different Behavioral Experiences Produce Distinctive Parallel Changes in, and Correlate With, Frontal Cortex and Hippocampal Global Post-translational Histone Levels. Front. Integr. Neurosci. 12:29. doi: 10.3389/fnint.2018.00029
Received: 08 May 2018; Accepted: 29 June 2018;
Published: 19 July 2018.
Edited by:
Martín Cammarota, Federal University of Rio Grande do Norte, BrazilReviewed by:
Gregg Stanwood, Florida State University, United StatesD. Caroline Blanchard, University of Hawaii at Manoa, United States
Copyright © 2018 Sobolewski, Singh, Schneider and Cory-Slechta. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marissa Sobolewski, bWFyaXNzYV9zb2JvbGV3c2tpQHVybWMucm9jaGVzdGVyLmVkdQ==