
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
BRIEF RESEARCH REPORT article
Front. Neurol. , 18 December 2019
Sec. Movement Disorders
Volume 10 - 2019 | https://doi.org/10.3389/fneur.2019.01332
This article is part of the Research Topic Neurogenetics in neurology: from molecular neuroscience to precision medicine View all 13 articles
 Michaela Pogoda1†
Michaela Pogoda1† Franz-Joachim Hilke1,2†Ebba Lohmann3,4,5Marc Sturm1Florian Lenz1Jakob Matthes1Francesc Muyas1,6,7
Franz-Joachim Hilke1,2†Ebba Lohmann3,4,5Marc Sturm1Florian Lenz1Jakob Matthes1Francesc Muyas1,6,7 Stephan Ossowski1,6,7
Stephan Ossowski1,6,7 Alexander Hoischen8,9Ulrike Faust1Ilnaz Sepahi1
Alexander Hoischen8,9Ulrike Faust1Ilnaz Sepahi1 Nicolas Casadei1,10Sven Poths1
Nicolas Casadei1,10Sven Poths1 Olaf Riess1,10Christopher Schroeder1*
Olaf Riess1,10Christopher Schroeder1* Kathrin Grundmann1
Kathrin Grundmann1Background: This study's aim was to investigate a large cohort of dystonia patients for pathogenic and rare variants in the ATM gene, making use of a new, cost-efficient enrichment technology for NGS-based screening.
Methods: Single molecule Molecular Inversion Probes (smMIPs) were used for targeted enrichment and sequencing of all protein coding exons and exon-intron boundaries of the ATM gene in 373 dystonia patients and six positive controls with known ATM variants. Additionally, a rare-variant association study was performed.
Results: One patient (0.3%) was compound heterozygous and 21 others were carriers of variants of unknown significance (VUS) in the ATM gene. Although mutations in sporadic dystonia patients are not common, exclusion of pathogenic variants is crucial to recognize a potential tumor predisposition syndrome. SmMIPs produced similar results as routinely used NGS-based approaches.
Conclusion: Our results underline the importance of implementing ATM in the routine genetic testing of dystonia patients and confirm the reliability of smMIPs and their usability for germline screenings in rare neurodegenerative conditions.
Ataxia-telangiectasia (A-T) is a rare autosomal recessively inherited disease usually characterized by ataxia, neuro-motor impairment, ocular or cutaneous telangiectasia, high risk of malignancies and immunodeficiency (1–3). A-T patients are often extraordinarily sensitive to ionizing radiation, contraindicating radiation therapy for them as a standard therapy in case of cancer (4, 5). The disease is caused by mutations of the Ataxia telangiectasia mutated (ATM) gene located on chromosome 11q22-23 (6). ATM encodes the 350 kDa ATM protein, a nuclear serine/threonine-protein kinase which is crucial in the cellular response to DNA damage such as double-strand breaks (7, 8). Classic A-T is caused by biallelic truncating ATM mutations which lead to a total loss of ATM protein, resulting in an impaired cell cycle (9). Since variants in ATM are known to confer cancer risk in heterozygous carriers (10), and at the same time cause increased sensitivity of the patients to toxic effects of ionizing radiation (4, 5), identifying ATM mutation carriers can be highly relevant for adequate treatment and regular cancer control examinations.
Furthermore, recent reports describe non-classic forms of A-T or “variant A-T” (11, 12). In these cases, some residual ATM kinase function is maintained and the phenotype is highly variable, including incomplete or atypical phenotypes, e.g., ataxia plus extrapyramidal symptoms or choreoathetosis lacking the classical hallmarks, often masking the correct diagnosis (11, 13). Some of these ATM mutations manifest as pure generalized or focal dystonia (14).
However, the frequency of ATM mutations in different cohorts of dystonia patients is not well-described. Therefore, we tested a cohort of 373 dystonia patients for ATM alterations.
We applied single molecule Molecular Inversion Probes (smMIPs) for targeted enrichment and sequencing of all protein coding exons and exon-intron boundaries of ATM. SmMIPs represent a cost-efficient and fast high-throughput technique to identify sequence variation in genes containing many exons (15). SmMIPs are oligonucleotide probes possessing two sequences complementary to defined genomic target regions (16–21). These complementary sequences are located at the 3′-end and at the 5′-end of the probe and hybridize to the single-stranded sample DNA upstream and downstream of the chosen target region of variable length. The gap between the complementary sequences is filled with the copy of the DNA target region by a polymerase, circularizing the probe in an additional ligation reaction.
The circular smMIP-target molecule can then be amplified in a PCR reaction and is, after a single library purification step, ready for sequencing. One main advantage of smMIPs is that probes can be designed in a modular way, tiling all relevant regions as closely as necessary, covering both DNA strands if desired (22). This is especially relevant for the avoidance of artifactual DNA sample damage, since artifacts usually only occur randomly in one strand (17, 23). Another asset of smMIPs is the usage of a unique single molecule (sm) molecular identifier (UMI = unique molecular identifier), a sequence of (in our case) 8 random bases in the probe that is individual for 48 ≈ 65,500 molecules. Since every smMIP molecule with an individual UMI-tag can only hybridize to one genomic DNA fragment, the UMI sequence can be used to retrace all originally different DNA molecules and to correct for PCR duplicates in the bioinformatical analysis. Thus, an accurate representation of the diversity of DNA molecules in the sample allows the sensitive detection of variants, even at low frequencies (17, 23, 24). Therefore, it is also most suitable for especially cost-efficient, reliable germline mutation screenings in large cohorts. An asset of the technology is that smMIPs can be designed in a customized way (22) and can be used for massively parallel resequencing of many thousands of target regions (25). In addition, most of the chemistry is independent of a specific supplier.
Informed consent was obtained from all patients. All samples were taken in accordance with the local Ethical Committee (# 847/2017BO). Genomic DNA was isolated from blood of 373 dystonia patients of Caucasian origin, who had been examined by specialists in movement disorders according to the current clinical criteria (26). Inclusion criteria were as follows: various degrees of dystonia as defined by published clinical criteria and a clinical course compatible with primary dystonia without features indicating secondary dystonia.
The whole smMIPs protocol was only slightly modified according to established protocols (beside the protocol, a detailed scheme of the methodology and workflow can be found in 15, 17, 21).
We designed smMIPs to screen the patients' DNA using the open source tool MIPGen (22). The design resulted in 190 smMIPs spanning all 62 protein coding exons of the ATM gene [transcript ENST00000278616, Ensemble (27)], covering both the sense and anti-sense strand of the DNA. Probes were synthesized by IDT (Integrated DNA Technologies; Coralville, USA). Upon arrival, all smMIPs were pooled in an equimolar manner. Six random samples were processed and sequenced to assess probe performance. Using the read depth of all individual probes, the pool was rebalanced twice, in order to improve uniformity of coverage of the target regions (17, 28) (see Supplementary Material for further information).
One hundred nanogram isolated DNA were used as input for the targeted capture of genomic regions of interest, and incubated for 21 h with smMIPs, using a ratio of 1:800 (DNA molecules:smMIPs), adding a polymerase and ligase for gap-filling and circularization. Subsequently, a 1-h exonuclease digestion was performed to remove any linear DNA. Thus, only circular smMIPs were amplified in a PCR using a high-fidelity polymerase. A small amount of each PCR product was categorized semi-quantitatively on an agarose gel, and the remaining PCR products were pooled accordingly to obtain equimolar representation of all samples. The pools were cleaned up for Illumina sequencing in one step using XP Ampure beads and the DNA concentration and fragment size was measured using a Qubit 2.0 device and an Agilent TapeStation 2200 to calculate the molarity of each pool. Samples were sequenced on a HiSeq2500 platform with 2 × 125 cycles (paired-end) and a target of 500 clusters per smMIP per sample.
Sequencing data were analyzed using an adapted in-house pipeline (available on https://github.com/imgag/megSAP, version 0.1-663-ged5a95d). Briefly, all sequences were identified by the UMI to allow later grouping and correction for PCR duplicates. Reads were aligned and mapped using paired-end reads (29), PCR duplicates were used to correct random PCR or sequencing errors labeling their base-quality to 0 to avoid calling falls-positive variants. BAM-files were generated, and variants called using the tool FreeBayes, version 1.1.0 (30). Variants were annotated and saved in GSvar-format.
For rare-variant association, a case-control study was performed (31). As a control cohort, vcf-files of the 1000 genomes project (32) were downloaded for 404 patients with European descent (populations Italian, Spanish, British, and Utah with European ancestry; Finnish not included). In order to obtain a representative and unbiased control cohort, we chose a public database for control data. Only target regions that were covered in both groups, case and control, were analyzed. All variants were subsequently filtered by the following criteria.
For filtering the variants, allele frequencies in the 1000 genomes database and in the ExAC database were necessitated to be below 1% and all variants needed to be reported <50 times in our in-house database. Synonymous and intronic variants (cut-off ± 8 bp) were excluded as well as variants that had been classified as benign or likely benign in our diagnostic in-house database. All variants were classified according to slightly modified guidelines (33, 34).
Statistical analyses were performed using JMP software version 13.0.0 (SAS Institute, Cary, NC). For rare-variant association, a Fisher's exact test was performed (α = 0.05).
The raw data supporting the conclusions of this manuscript will be made available by the authors, without undue reservation, to any qualified researcher. The data (vcf-files) of the control cohort were obtained from the 1000 genomes project (32) and are publicly available.
The median target region read depth for all samples (n = 373) was 218×, ranging from 34× for one sample to 840× in the sample with the highest coverage. The 20×-coverage of all target regions was 98.0%. Note that these figures only comprise condensed reads (no PCR duplicates) which were shown to be sufficient for allele frequencies typical in germline analyses (17).
Among 373 dystonia patients, we found 21 different variants in 22 of the dystonia patients (Table 1). Of these, 20 were of uncertain clinical significance (VUS) (33). Ten of them had high in silico prediction of pathogenicity [Combined Annotation-Dependent Depletion, CADD-Score > 20, (35, 36)]. One variant was classified as pathogenic (p. (Val2716Ala), class 5). There was no significant difference from the control group from the 1000 genomes project (p > 0.05 for both VUS or predicted pathogenic variants).
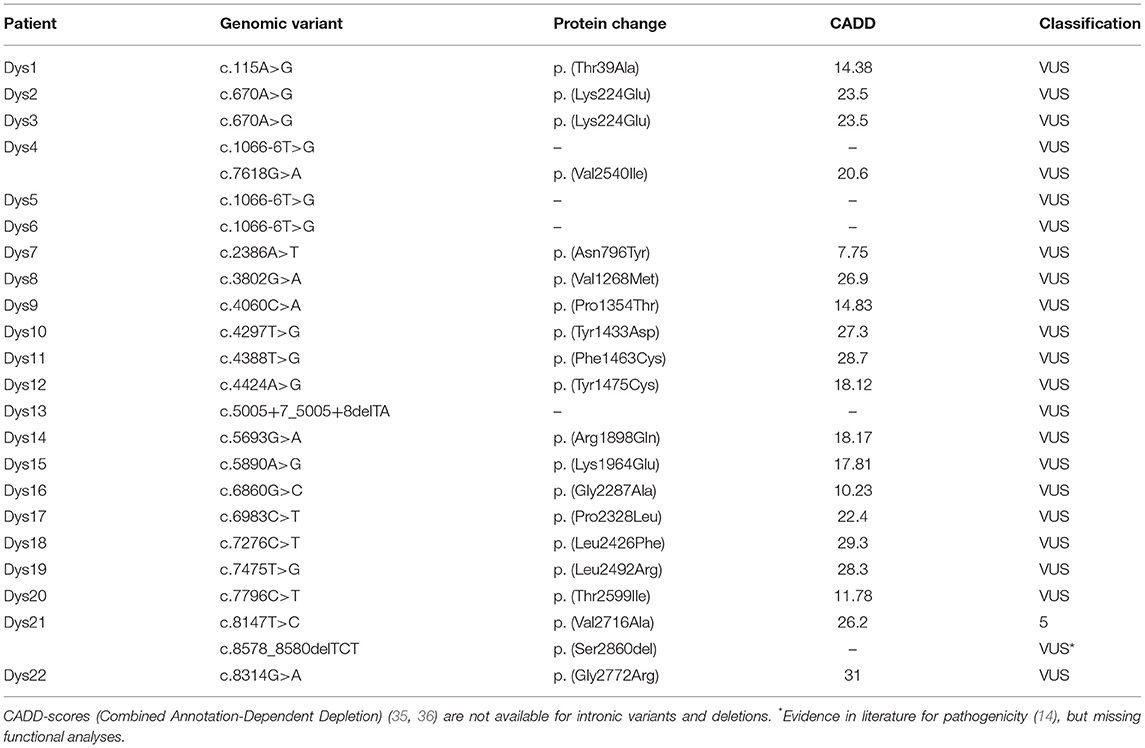
Table 1. Rare variants in the ATM gene among the 373 dystonia patients detected with smMIPs.
Among the dystonia patients, one patient was found with two variants in the ATM gene: one was pathogenic and one a VUS with evidence for pathogenicity (p. (Val2617Ala), p. (Ser2860del)). Clinical examination had produced highly elevated AFP levels, familial aggregation of malignancies and a deterioration of the dystonic symptoms that were largely insensitive to treatment. This patient was recently published (14). In contrast, we found no biallelic pathogenic or likely pathogenic variants in the 1000 genomes control cohort.
All pathogenic variants reported for the six positive controls were also detected using smMIPs. The comparison of all variants showed no discrepancy between smMIPs and the diagnostic results.
Although we found no statistical association of ATM variants between dystonia patients and a control group, we found one patient with a pathogenic variant (p. (Val2617Ala)) and a VUS with evidence for pathogenicity (p. (Ser2860del)) among 373 dystonia patients. In accordance with other published data on the patient (14), our study confirms the efficiency of smMIPs as a diagnostic screening tool (15) and moreover the importance of identifying ATM as rare underlying cause in dystonia (37, 38). The genetic causes of dystonia, a rare disease, are very heterogeneous (39), and many genes have been identified in recent times. Among these genetic causes, ATM sequence variations are a crucial factor for patients regarding cancer susceptibility (10), disease progression and radiation toxicity (3, 4, 40, 41). Thus, we stress the importance of including ATM to the general screening for causes of dystonia.
Due to the usage of UMIs, smMIPs increase the confidence in variants without the necessity of a high coverage, because PCR duplicates can be used to correct for artifacts (17, 23). We successfully established smMIPs in a screening for ATM germline variants in dystonia patients. All pathogenic variants that had been detected with different diagnostic NGS approaches in six control patients (and one dystonia patient) were confirmed, validating smMIPs as a sensitive, competitive methodology. They provide a straight-forward wet lab protocol, with customizable chemistry, flexible probe design, sensitive variant detection, and cost-efficient sample processing, especially for high-throughput genetic testing (15, 21, 28, 42). Thus, they constitute a convenient tool for panel-based genetic testing.
Since ATM mutations confer a higher risk of developing cancer and radiation toxicity, it is crucial to detect ATM-variants as the underlying cause in any dystonic patient. In our study, we found smMIPs to be as sensitive as other NGS-based approaches while being highly cost-efficient and flexible.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation, to any qualified researcher. The data (vcf-files) of the control cohort were obtained from the 1000 genomes project and are publicly accessible here: http://www.internationalgenome.org/data#download.
All experimental protocols in this study were reviewed and approved by the Ethics Committee of the Eberhard Karls University of Tübingen and of the Medical Faculty of the University Hospital Tübingen. All patients gave written informed consent in accordance with the Declaration of Helsinki. The patients/participants provided their written informed consent to participate in this study.
The research project was conceptualized by OR, CS, F-JH, EL, and KG. Sample acquisition was accomplished by EL, KG, and UF. Methodology was implemented and supported by AH, F-JH, MP, NC, and SP. The computational analyses were developed and/or conducted by CS, MS, FL, SO, FM, and JM. Variant classification and (statistical) analyses were conducted by MP, IS, and SO. All authors revised and approved the final manuscript.
The study was funded by the Exzellenzinitiative Zukunftskonzept Universität Tübingen, project ID D.27.13047.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Illumina sequencing was performed by the DFG NGS Competence Center Tübingen (NCCT, Tübingen, Germany). We would also like to acknowledge all patients and their families who contributed to this study, as well as the clinicians and the technical staff. We acknowledge support by the Deutsche Forschungsgemeinschaft and the Open Access Publishing Fund of the University of Tübingen. This manuscript has been part of an unpublished Master Thesis (43).
2. Gatti RA, Painter RB. Ataxia-telangiectasia. Luxembourg: Springer Science & Business Media (2013).
3. Gilad S, Chessa L, Khosravi R, Russell P, Galanty Y, Piane M, et al. Genotype-phenotype relationships in ataxia-telangiectasia and variants. Am J Hum Genet. (1998) 62:551–61. doi: 10.1086/301755
4. Abadir R, Hakami N. Ataxia telangiectasia with cancer. An indication for reduced radiotherapy and chemotherapy doses. Brit J Radiol. (1983) 56:343–5. doi: 10.1259/0007-1285-56-665-343
5. Gatti RA. Ataxia-telangiectasia. Dermatol Clin. (1995) 13:1–6. doi: 10.1016/S0733-8635(18)30100-1
6. Savitsky K, Bar-Shira A, Gilad S, Rotman G, Ziv Y, Vanagaite L, et al. A single ataxia telangiectasia gene with a product similar to PI-3 kinase. Science. (1995) 268:1749–53. doi: 10.1126/science.7792600
7. Gutiérrez-Enríquez S, Fernet M, Dörk T, Bremer M, Lauge A, Stoppa-Lyonnet D, et al. Functional consequences of ATM sequence variants for chromosomal radiosensitivity. Genes Chromosomes Cancer. (2004) 40:109–19. doi: 10.1002/gcc.20025
8. Savitsky K, Sfez S, Tagle DA, Ziv Y, Sartiel A, Collins FS, et al. The complete sequence of the coding region of the ATM gene reveals similarity to cell cycle regulators in different species. Hum Mol Genet. (1995) 4:2025–32. doi: 10.1093/hmg/4.11.2025
9. McKinnon PJ. ATM and the molecular pathogenesis of ataxia telangiectasia. Annu Rev Pathol. (2012) 7:303–21. doi: 10.1146/annurev-pathol-011811-132509
10. Renwick A, Thompson D, Seal S, Kelly P, Chagtai T, Ahmed M, et al. ATM mutations that cause ataxia-telangiectasia are breast cancer susceptibility alleles. Nat Genet. (2006) 38:873–5. doi: 10.1038/ng1837
11. Lohmann E, Krüger S, Hauser A-K, Hanagasi H, Guven G, Erginel-Unaltuna N, et al. Clinical variability in ataxia–telangiectasia. J Neurol. (2015) 262:1724–7. doi: 10.1007/s00415-015-7762-z
12. Méneret A, Ahmar-Beaugendre Y, Rieunier G, Mahlaoui N, Gaymard B, Apartis E, et al. The pleiotropic movement disorders phenotype of adult ataxia-telangiectasia. Neurology. (2014) 83:1087–95. doi: 10.1212/WNL.0000000000000794
13. McConville CM, Stankovic T, Byrd PJ, McGuire GM, Yao Q-Y, Lennox GG, et al. Mutations associated with variant phenotypes in ataxia-telangiectasia. Am J Hum Genet. (1996) 59:320.
14. Kuhm C, Gallenmüller C, Dörk T, Menzel M, Biskup S, Klopstock T. Novel ATM mutation in a German patient presenting as generalized dystonia without classical signs of ataxia-telangiectasia. J Neurol. (2015) 262:768–70. doi: 10.1007/s00415-015-7636-4
15. Neveling K, Mensenkamp AR, Derks R, Kwint M, Ouchene H, Steehouwer M, et al. BRCA testing by single-molecule molecular inversion probes. Clin Chem. (2017) 63:503–12. doi: 10.1373/clinchem.2016.263897
16. Nilsson M, Malmgren H, Samiotaki M, Kwiatkowski M, Chowdhary BP, Landegren U. Padlock probes: circularizing oligonucleotides for localized DNA detection. Science. (1994) 265:2085–8. doi: 10.1126/science.7522346
17. Eijkelenboom A, Kamping EJ, Kastner-van Raaij AW, Hendriks-Cornelissen SJ, Neveling K, Kuiper RP, et al. Reliable next-generation sequencing of formalin-fixed, paraffin-embedded tissue using single molecule tags. J Mol Diagn. (2016) 18:851–63. doi: 10.1016/j.jmoldx.2016.06.010
18. Cantsilieris S, Stessman HA, Shendure J, Eichler EE. Targeted capture and high-throughput sequencing using molecular inversion probes (MIPs). Genotyping. (2017) 1492:95–106. doi: 10.1007/978-1-4939-6442-0_6
19. Hardenbol P, Banér J, Jain M, Nilsson M, Namsaraev EA, Karlin-Neumann GA, et al. Multiplexed genotyping with sequence-tagged molecular inversion probes. Nat Biotechnol. (2003) 21:673. doi: 10.1038/nbt821
20. Umbarger MA, Kennedy CJ, Saunders P, Breton B, Chennagiri N, Emhoff J, et al. Next-generation carrier screening. Genet Med. (2014) 16:132. doi: 10.1038/gim.2013.83
21. O'Roak BJ, Vives L, Fu W, Egertson JD, Stanaway IB, Phelps IG, et al. Multiplex targeted sequencing identifies recurrently mutated genes in autism spectrum disorders. Science. (2012) 338:1619–22. doi: 10.1126/science.1227764
22. Boyle EA, O'Roak BJ, Martin BK, Kumar A, Shendure J. MIPgen: optimized modeling and design of molecular inversion probes for targeted resequencing. Bioinformatics. (2014) 30:2670–2. doi: 10.1093/bioinformatics/btu353
23. Hiatt JB, Pritchard CC, Salipante SJ, O'Roak BJ, Shendure J. Single molecule molecular inversion probes for targeted, high-accuracy detection of low-frequency variation. Genome Res. (2013) 23:843–54. doi: 10.1101/gr.147686.112
24. Shen P, Wang W, Krishnakumar S, Palm C, Chi A-K, Enns GM, et al. High-quality DNA sequence capture of 524 disease candidate genes. Proc Natl Acad Sci USA. (2011) 108:6549–54. doi: 10.1073/pnas.1018981108
25. Turner EH, Lee C, Ng SB, Nickerson DA, Shendure J. Massively parallel exon capture and library-free resequencing across 16 genomes. Nat Methods. (2009) 6:315. doi: 10.1038/nmeth.f.248
26. Fahn S, Eldridge R. Definition of dystonia and classification of the dystonic states. Adv Neurol. (1976) 14:1–5.
27. Aken BL, Ayling S, Barrell D, Clarke L, Curwen V, Fairley S, et al. The Ensembl gene annotation system. Database. (2016) 2016:baw093. doi: 10.1093/database/baw093
28. Porreca GJ, Zhang K, Li JB, Xie B, Austin D, Vassallo SL, et al. Multiplex amplification of large sets of human exons. Nat Methods. (2007) 4:931. doi: 10.1038/nmeth1110
29. Li H, Durbin R. Fast and accurate short read alignment with Burrows–Wheeler transform. Bioinformatics. (2009) 25:1754–60. doi: 10.1093/bioinformatics/btp324
30. Garrison E, Marth G. Haplotype-based variant detection from short-read sequencing. arXiv preprint. (2012) arXiv arXiv:1207.3907.
31. Lee S, Abecasis GR, Boehnke M, Lin X. Rare-variant association analysis: study designs and statistical tests. Am J Hum Genet. (2014) 95:5–23. doi: 10.1016/j.ajhg.2014.06.009
32. Genomes Project Consortium. A global reference for human genetic variation. Nature. (2015) 526:68–74. doi: 10.1038/nature15393
33. Plon SE, Eccles DM, Easton D, Foulkes WD, Genuardi M, Greenblatt MS, et al. Sequence variant classification and reporting: recommendations for improving the interpretation of cancer susceptibility genetic test results. Hum Mutat. (2008) 29:1282–91. doi: 10.1002/humu.20880
34. Spurdle AB, Healey S, Devereau A, Hogervorst FB, Monteiro AN, Nathanson KL, et al. ENIGMA—Evidence-based network for the interpretation of germline mutant alleles: an international initiative to evaluate risk and clinical significance associated with sequence variation in BRCA1 and BRCA2 genes. Hum Mutat. (2012) 33:2–7. doi: 10.1002/humu.21628
35. Kircher M, Witten DM, Jain P, O'roak BJ, Cooper GM, Shendure J. A general framework for estimating the relative pathogenicity of human genetic variants. Nat Genet. (2014) 46:310–5. doi: 10.1038/ng.2892
36. Velde KJ, Kuiper J, Thompson BA, Plazzer JP, Valkenhoef G, Haan M, et al. Evaluation of CADD scores in curated mismatch repair gene variants yields a model for clinical validation and prioritization. Hum Mutat. (2015) 36:712–9. doi: 10.1002/humu.22798
37. Bodensteiner JB, Goldblum RM, Goldman AS. Progressive dystonia masking ataxia in ataxia-telangiectasia. Arch Neurol. (1980) 37:464–5. doi: 10.1001/archneur.1980.00500560094020
38. Thompson S, Iyer A, Byrd P, Taylor M, Spinty S. Dopa-Responsive dystonia and chorea as a presenting feature in ataxia-telangiectasia. Mov Disord Clin Pract. (2014) 1:249–51. doi: 10.1002/mdc3.12048
39. Lohmann K, Klein C. Genetics of dystonia: what's known? What's new? What's next? Mov Disord. (2013) 28:899–905. doi: 10.1002/mds.25536
40. Barlow C, Eckhaus MA, Schäffer AA, Wynshaw-Boris A. Atm haploinsufficiency results in increased sensitivity to sublethal doses of ionizing radiation in mice. Nat Genet. (1999) 21:359. doi: 10.1038/7684
41. Verhagen MM, Last JI, Hogervorst FB, Smeets DF, Roeleveld N, Verheijen F, et al. Presence of ATM protein and residual kinase activity correlates with the phenotype in ataxia-telangiectasia: a genotype–phenotype study. Hum Mutat. (2012) 33:561–71. doi: 10.1002/humu.22016
42. Niedzicka M, Fijarczyk A, Dudek K, Stuglik M, Babik W. Molecular Inversion Probes for targeted resequencing in non-model organisms. Sci Rep. (2016) 6:24051. doi: 10.1038/srep24051
Keywords: dystonia, ATM, NGS, ataxia-telangiectasia, MIPs
Citation: Pogoda M, Hilke F-J, Lohmann E, Sturm M, Lenz F, Matthes J, Muyas F, Ossowski S, Hoischen A, Faust U, Sepahi I, Casadei N, Poths S, Riess O, Schroeder C and Grundmann K (2019) Single Molecule Molecular Inversion Probes for High Throughput Germline Screenings in Dystonia. Front. Neurol. 10:1332. doi: 10.3389/fneur.2019.01332
Received: 05 June 2019; Accepted: 02 December 2019;
Published: 18 December 2019.
Edited by:
Matthew James Farrer, University of Florida, United StatesReviewed by:
Pedro Chana, Universidad de Santiago de Chile, ChileCopyright © 2019 Pogoda, Hilke, Lohmann, Sturm, Lenz, Matthes, Muyas, Ossowski, Hoischen, Faust, Sepahi, Casadei, Poths, Riess, Schroeder and Grundmann. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Christopher Schroeder, Y2hyaXN0b3BoZXIuc2Nocm9lZGVyQG1lZC51bmktdHVlYmluZ2VuLmRl
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.