 Dayu Zou
Dayu Zou Hongbin Liu
Hongbin Liu Meng Li
Meng Li- 1SZU-HKUST Joint Ph.D. Program in Marine Environmental Science, Shenzhen University, Shenzhen, China
- 2Shenzhen Key Laboratory of Marine Microbiome Engineering, Institute for Advanced Study, Shenzhen University, Shenzhen, China
- 3Department of Ocean Science, The Hong Kong University of Science and Technology, Hong Kong, China
- 4Hong Kong Branch of Southern Marine Science & Engineering Guangdong Laboratory, The Hong Kong University of Science and Technology, Hong Kong, China
Archaea are diverse and ubiquitous prokaryotes present in both extreme and moderate environments. Estuaries, serving as links between the land and ocean, harbor numerous microbes that are relatively highly active because of massive terrigenous input of nutrients. Archaea account for a considerable portion of the estuarine microbial community. They are diverse and play key roles in the estuarine biogeochemical cycles. Ammonia-oxidizing archaea (AOA) are an abundant aquatic archaeal group in estuaries, greatly contributing estuarine ammonia oxidation. Bathyarchaeota are abundant in sediments, and they may involve in sedimentary organic matter degradation, acetogenesis, and, potentially, methane metabolism, based on genomics. Other archaeal groups are also commonly detected in estuaries worldwide. They include Euryarchaeota, and members of the DPANN and Asgard archaea. Based on biodiversity surveys of the 16S rRNA gene and some functional genes, the distribution and abundance of estuarine archaea are driven by physicochemical factors, such as salinity and oxygen concentration. Currently, increasing amount of genomic information for estuarine archaea is becoming available because of the advances in sequencing technologies, especially for AOA and Bathyarchaeota, leading to a better understanding of their functions and environmental adaptations. Here, we summarized the current knowledge on the community composition and major archaeal groups in estuaries, focusing on AOA and Bathyarchaeota. We also highlighted the unique genomic features and potential adaptation strategies of estuarine archaea, pointing out major unknowns in the field and scope for future research.
Introduction
Archaea were proposed as the third domain of life by Woese and Fox (1977). The understanding of archaeal distribution, diversity and ecological functions has dramatically changed since then. Originally, archaea were thought to only inhabit extreme environments, and dwell under highly acidic, saline, and high-temperature conditions (Woese et al., 1978). Hence, for decades, they were considered to be obligate extremophiles. However, the discovery of mesophilic archaeal groups in temperate and oxygenated marine waters (DeLong, 1992; Fuhrman et al., 1992) overturned the previous view on the distribution of archaea. Subsequently, their presence was detected in both terrestrial and oceanic waters (DeLong, 1992; Fuhrman et al., 1992; Stein et al., 2002), soils and sediments (Bintrim et al., 1997; Walsh et al., 2005; Biddle et al., 2006; Teske and Sørensen, 2008), and under moderate conditions, which confirmed their ubiquity on a global scale (DeLong and Pace, 2001; Schleper et al., 2005).
The total cell number of bacteria and archaea exceeds 1.2 × 1030. These microorganisms mainly inhabit the deep oceanic subsurface (approximately 4 × 1029 cells), deep continental subsurface (approximately 3 × 1029 cells), soil (approximately 3 × 1029 cells) and open ocean (approximately 1 × 1029 cells) (Bar-On et al., 2018; Magnabosco et al., 2018; Flemming and Wuertz, 2019). Notably, archaea contribute a considerable proportion to the microbial biomass in moderate ecosystems (Moissl-Eichinger et al., 2018), accounting for approximately 40.0% and 12.8% prokaryotic cells in marginal regions and open ocean sites, respectively (Hoshino and Inagaki, 2019).
Although only few archaeal isolates have been cultured to date, cultivation-independent techniques, including metagenomics and single-cell genomics, have greatly broadened the understanding of archaeal diversity and distribution (Rinke et al., 2013). For a long time, Crenarchaeota (mainly consisting of thermophiles) and Euryarchaeota (mainly consisting of methanogens and halophiles) were the only defined archaeal phyla.
The number of proposed archaeal phyla has reached nearly 30 in recent years (Adam et al., 2017; Eme et al., 2017; Baker et al., 2020). Accordingly, several archaeal lineages have been proposed as new phyla based on phylogenetic analysis, including Thaumarchaeota (Brochier-Armanet et al., 2008), Korarchaeota (Barns et al., 1994; Elkins et al., 2008; Reigstad et al., 2010), and Aigarchaeota (Nunoura et al., 2011), which led to the establishment of TACK superphylum (Guy and Ettema, 2011) together with Crenarchaeota. Recently, metagenomic studies revealed new archaeal phyla belonging to the TACK superphylum, such as Verstraetearchaeota (Vanwonterghem et al., 2016), Geoarchaeota (Kozubal et al., 2013), Bathyarchaeota (Meng et al., 2014; Evans et al., 2015; He et al., 2016), Geothermarchaeota (Jungbluth et al., 2017), Marsarchaeota (Jay et al., 2018), and Nezharchaeota (Wang Y. et al., 2019). The TACK superphylum was proposed as the Proteoarchaeota (Petitjean et al., 2014) as the metabolic functions were highly diverse among different phyla. Members of the newly identified Asgard superphylum (i.e., Lokiarchaeota, Odinarchaeota, Thorarchaeota, Heimdallarchaeota, Helarchaeota, and Gerdarchaeota) were suggested to be the closest prokaryotes related to eukaryotes (Spang et al., 2015, 2018; Zaremba-Niedzwiedzka et al., 2017; Seitz et al., 2019; Cai et al., 2020). For example, members of Asgard archaea harbor genes encoding eukaryotic signature proteins previously believed to be specific to eukaryotes (Spang et al., 2015; Zaremba-Niedzwiedzka et al., 2017; Seitz et al., 2019; Cai et al., 2020). Further, the DPANN superphylum comprises Diapherotrites, Parvarchaeota, Aenigmarchaeota, Nanoarchaeota, Nanohaloarchaeota, and other archaeal groups with extremely small genomes. Some of its members have been hypothesized to have mutualistic or parasitic lifestyles (Baker et al., 2010; Rinke et al., 2013; Castelle et al., 2015; Eme and Doolittle, 2015; Dombrowski et al., 2019). Although most of the archaeal niches and metabolic functions remain unknown, valuable information on archaeal genomics, proteomics, and physiology is available to help us understand the vast diversity and ubiquitous archaea in different environments.
Estuaries act as connectors of the land and ocean, and hence, exhibit unique characteristics that are different from those of terrestrial and oceanic environments (McLusky and Elliott, 2004). Microbial activities in estuaries are stimulated by numerous nutrients carried by a river discharge (Milliman et al., 1985), that may favor the growth of estuarine microbes, including archaea. Previous reports related to estuarine archaea were only focused on limited lineages or genomes. Hence, a systematic research of estuarine archaea may offer a broader view of these microorganisms. Here, we review the community composition and distribution pattern of archaea in global estuarine ecosystems, focusing on the predominant aquatic and sedimentary archaeal groups (i.e., Thaumarchaeota and Bathyarchaeota, respectively), to better understand the diversity, ecological niches, as well as evolution and adaptation of archaea in estuarine environments.
Community Structure of Estuarine Archaea
Estuaries, accounting for only 0.4% of the global marine area, are considered to be one of the most productive ecosystems (Longhurst et al., 1995; Cloern et al., 2014) and are important sources of biogenetic emission of methane (Bange et al., 1994; Abril and Borges, 2005). They are also responsible for the delivery of terrigenous silicon, phosphorus, and nitrogen to the ocean, fueling a high primary production in coastal regions (Harrison et al., 2008). In estuaries, the continental freshwater runoff mixes with the coastal seawater. Distinct physicochemical gradients form along the mouth of the estuary as a result of many factors, such as river discharge, monsoons, and anthropogenic interferences (Vieira et al., 2007; Bernhard and Bollmann, 2010). These physicochemical gradients, e.g., decreasing organic compound and nitrogenous nutrient levels, or increasing salinity, and sulfate and chloride levels, may strongly affect the microbial community structure (Alla et al., 2006; Webster et al., 2014). In addition, terrestrial nutrients and microorganisms carried by a river discharge accumulate in estuaries, leading to a relatively high biodiversity and microbial activities therein, and subsequently influencing the biogeochemical and ecological processes of the estuarine ecosystems (Baird et al., 2004; Canfield and Thamdrup, 2009; Webster et al., 2014; Zhou et al., 2017; Liu et al., 2018b). On the other hand, because of the industrialization and urbanization along rivers, large quantities of varied continental inputs have resulted in severe environmental pollution, eutrophication, and oxygen deficit or hypoxia in estuaries (Diaz, 2001; Li et al., 2002; Huang et al., 2003; Howarth et al., 2011; Zheng et al., 2011; Lu et al., 2018). Therefore, a comprehensive understanding of the estuarine microbial community is critical for elucidating their ecological roles in the estuary.
Increasing numbers of archaeal groups are being recognized as vital players in estuarine biogeochemical cycles (Figure 1). These include: Thaumarchaeota, which comprise ammonia oxidizing archaea (AOA), one of the key players of ammonia oxidation in estuaries (Francis et al., 2005; Liu L. et al., 2011; Liu Z. et al., 2011; Zou et al., 2019, 2020b); Bathyarchaeota (formerly called Miscellaneous Crenarchaeotal Group, MCG), which are hypothesized to be important players in the benthic carbon cycle that, based on genomic analysis, may be able to utilize diverse organic substrates (Meng et al., 2014; Lazar et al., 2015, 2016; He et al., 2016; Zhou et al., 2018a); Euryarchaeota, e.g., most methanogens and anaerobic methanotrophs, are responsible for methane production and oxidation in estuaries, respectively (Oremland and Polcin, 1982; Serrano-Silva et al., 2014); and Asgard archaea, such as Thorarchaeota and Lokiarchaeota, which may participate in some biogeochemical cycles, as suggested by metagenomics (Seitz et al., 2016; Liu Y. et al., 2018; MacLeod et al., 2019; Cai et al., 2020). Recently, Liu et al. (2018b) explored the diversity and community structure of archaea in over 20 estuaries. Analysis of the numbers of operational taxonomic units (OTUs) highlighted the notion that the diversity of archaeal community is higher in low- and middle-latitude estuaries than in high-latitude estuaries. That is probably because the terrestrial and marine archaeal groups are continuously mixed in low- and middle-latitude estuarine regions, affected by high human activity (Xie et al., 2014; Liu et al., 2018b). Further, the archaeal community structure is co-influenced by many environmental factors, such as the latitude, salinity, dissolved oxygen levels, and nutrient conditions (Bernhard et al., 2010; Webster et al., 2014; Xie et al., 2014; Qin et al., 2017; Liu et al., 2018b).

Figure 1. Metabolic potentials of estuarine archaeal groups. Solid circles, metabolic pathways or functional genes identified in some genomes in the group. Empty circles, no reported pathways or functional genes in the genomes in the group. These results are re-summarized mainly based on Baker et al. (2020) and other previous papers, please see the main text and Supplementary Methods for details.
In this study, we have roughly revealed the archaeal composition in water columns and sediments of different estuaries summarized from published papers (Figure 2 and Supplementary Tables S1, S3 for detailed information). Generally, Bathyarchaeota dominate the sedimentary environment in most estuaries, while Thaumarchaeota and Euryarchaeota are predominant aquatic archaeal phyla. In addition, Woesearchaeota account for a high proportion of archaea in both aquatic and sedimentary environments in some estuaries, while Lokiarchaeota and Thorarchaeota are also observed in some estuaries. As shown in Figure 2, different estuaries harbor different archaeal communities, likely because of specific differences in the geographic location and physicochemical parameters. Liu et al. (2018b) suggested that Thaumarchaeota preferentially dwell in low-latitude estuarine environments, while Bathyarchaeota and Euryarchaeota are more abundant in mid- and high- latitude estuaries, respectively. The response and sensitivity of these archaeal groups to changes in the temperature and nutrient supply are different (Danovaro et al., 2016). This may be responsible for the variations in archaeal community at different latitudes. Intriguingly, although closely located along the west coastline of India, the archaeal communities in the Mandovi Estuary and Zuari Estuary are quite different (Singh et al., 2010; Khandeparker et al., 2017). The authors pointed out that the distinct microbial communities in these two estuaries might be affected by different localized interactions, however, more thorough investigations are needed to explain such differences. Similarly, a recent report revealed seasonal variations of archaeal communities in estuarine and coastal regions (Liu et al., 2020), highlighting the notion that spatiotemporal distribution of archaeal community is co-influenced by environmental parameters, including salinity and temperature. However, other than spatial and temporal variables, technical biases, such as sampling methods, library construction approaches, and different usage of primers and marker genes, might also impact the microbial community composition. Although it is difficult to measure the contribution of all variables, a precise and lucid understanding relies on considering different aspects as much as possible.
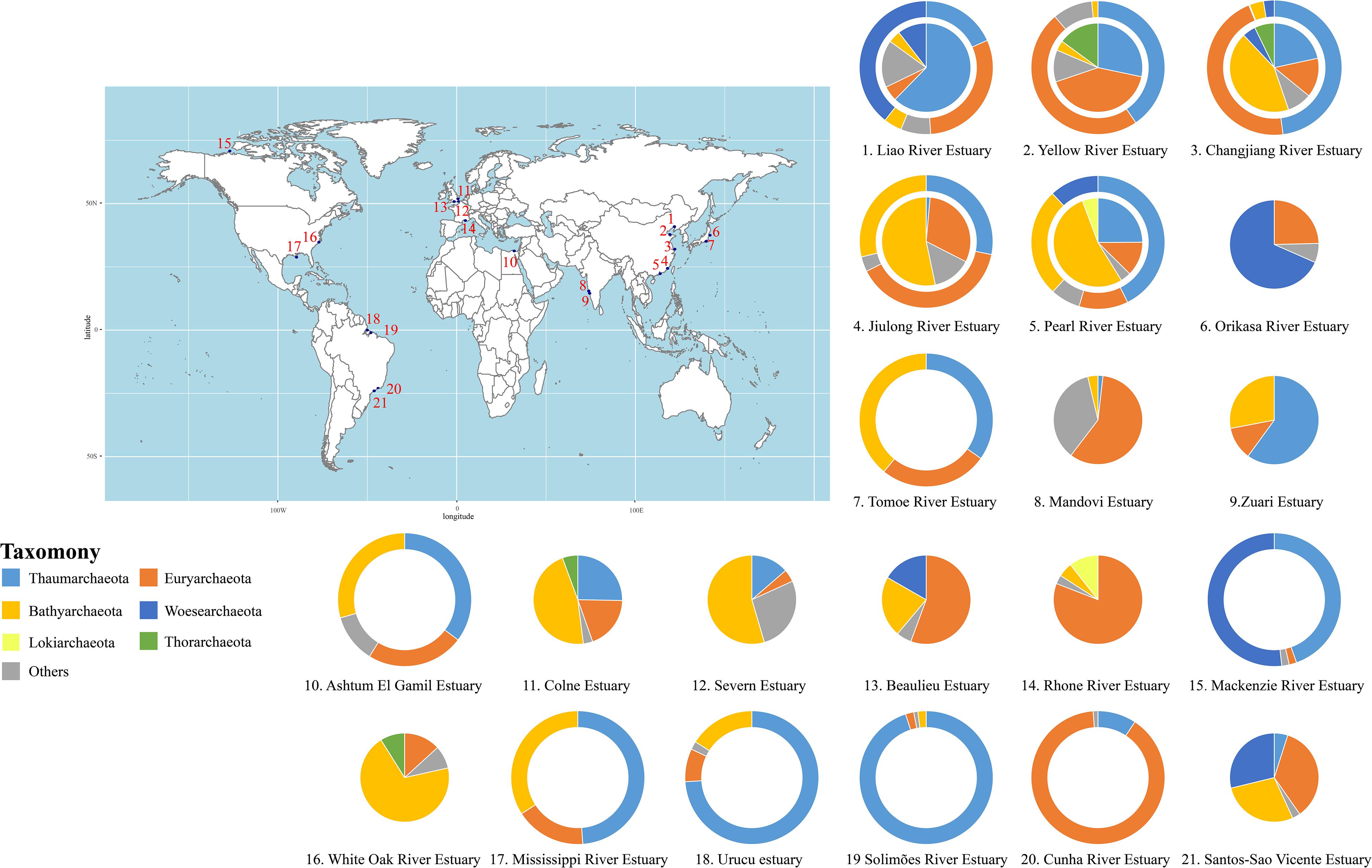
Figure 2. The archaeal community composition (on phylum level) in global estuaries. Outer rings, water samples; inner circles, sediment samples. Please see Supplementary Tables S1, S3 for details.
AOA in Estuaries
AOA Diversity Based on amoA Genes
Marine Group I (MG-I) archaea were first detected in the ocean water column (DeLong, 1992). MG-I are ubiquitous in the coastal water, and from shallow marine water to bathypelagic zones, accounting for a considerable fraction of the microbial community in those regions (Karner et al., 2001; Schattenhofer et al., 2009). The ability of archaeal ammonia oxidation was confirmed when the AOA Nitrosopumilus maritimus SCM1 (SCM1) was isolated (Könneke et al., 2005). Analyses of the archaeal amoA gene encoding the α-subunit of ammonia monooxygenase (AMO) revealed that archaea with an ammonia-oxidizing potential are ubiquitous in natural environments (Francis et al., 2005; Biller et al., 2012; Cao et al., 2013; Cheung et al., 2019). Members of chemolithoautotrophic AOA may be mixotrophic, as they harbor genes encoding extracellular peptidases and carbohydrate-active enzymes (Li et al., 2015; Zou et al., 2019). In addition, because of high affinity for ammonia, low oxygen demand, and tolerance of a wide salinity range, AOA are thought to have more prominent advantages for estuarine ammonia oxidation (Bollmann and Laanbroek, 2002; Martens-Habbena et al., 2009; Mosier et al., 2012b; Qin et al., 2017).
Based on the amoA genotypes as suggested by Alves et al. (2018), we summarized AOA community of different estuaries from published papers (Figure 3 and Supplementary Tables S2, S4 for detailed information). In aquatic samples, most estuarine AOA belong to lineage Nitrosopumilales (NP), while members of Nitrososphaerales (NS) are predominant in sediments, in agreement with previous observations (Alves et al., 2018). The genotypes NP-Gamma (SCM1-like) and NP-Epsilon (WCA) are more abundant in water columns than in sediments. According to Alves et al. (2018), these two groups are distributed in diverse environments, and are enriched in marine and estuarine regions, contributing to approximately 15% and 8% of all amoA sequences in global environments, respectively. On the other hand, the NS-Gamma and NS-Delta are more abundant in estuarine sediments, contributing to approximately 13% and 23% of all amoA sequences (Alves et al., 2018). The actual community composition varies among different estuaries, emphasizing the point that AOA distribution is environment-specific. Indeed, the approximate abundance of amoA genes in estuaries ranges from 1.0 × 103 to 1.7 × 108 copies/L and from 4.9 × 103 to 2.4 × 108 copies/g in water columns and sediments, respectively (Supplementary Table S2). Notably, the technical biases, such as different detection methods, PCR settings and primers, and analysis pipelines among these studies (Supplementary Tables S2, S4), may also have impacts on the results. Hence, they are not directly comparable to each other unless all these issues are carefully considered.

Figure 3. The AOA community composition (based on the amoA genotype) in global estuaries. Outer rings, water samples; inner circles, sediment samples. Please see Supplementary Tables S2, S4 for details.
Distribution Patterns of AOA
Ammonia-oxidizing archaea represent one of the most ubiquitous ammonia oxidizers on Earth, inhabiting both moderate and extreme environments, and also found as putative symbionts of marine invertebrates (Steger et al., 2008; Webster and Taylor, 2012; Erwin et al., 2014). Some AOA cultures grow at temperatures up to over 70°C (de la Torre et al., 2008; Daebeler et al., 2018), and in the range of pH values from 4.0 to 7.5 (Schleper and Nicol, 2010; Stieglmeier et al., 2014). However, the archaeal amoA genes have been detected at wider ranges of temperature (as low as 0.2°C and as high as 97°C) (Reigstad et al., 2008), pH (from 2.5 to 9.0), and salinity (from 0 to 38 psu) (Erguder et al., 2009), implying that AOA or amoA-encoding archaea (AEA) may be more widespread than current appreciated. Future cultivation or enrichment experiments may offer additional clues to better understand these microorganisms.
According to many amoA-based studies, the distribution and abundance of AOA or AEA are strongly associated with the environment. Hence, they can be generally categorized into marine and terrestrial groups (Francis et al., 2005; Biller et al., 2012; Pester et al., 2012; Cao et al., 2013; Yao et al., 2013). The distribution of the marine group is mainly related to the water depth, reflecting the response and adaptation of different ecotypes to different light exposures and ammonia concentrations (Sintes et al., 2013; Luo et al., 2014), while the terrestrial group is mainly partitioned by pH values (Gubry-Rangin et al., 2011).
Based on the archaeal amoA genotypes, a community shift of AOA or AEA along the estuary is commonly observed, with salinity one of the most important environmental factors influencing the distribution pattern (Dang et al., 2008; Mosier and Francis, 2008; Santoro et al., 2008; Bernhard et al., 2010; Zhang Y. et al., 2015). Salinity affects the ammonium adsorption in sediments (Boynton and Kemp, 1985), while the correlation between archaeal amoA gene abundance and salinity is different in different estuaries. For example, archaeal amoA genes are more abundant in low-salinity sediments of the San Francisco Bay than in high salinity regions (Mosier and Francis, 2008), but show positive correlation with salinity in the Elkhorn Slough Estuary (Caffrey et al., 2007). In addition, the abundance and transcriptional activity of archaeal amoA genes are high in estuarine regions with medium- and high-salinity (Zhang Y. et al., 2015). However, no significant correlations between the abundance of archaeal amoA genes and estuarine salinity were also reported (Santoro et al., 2008; Cao et al., 2011). The distinct seasonal changes of environmental variables in estuaries caused by river discharge may also influence the AOA community (Beman and Francis, 2006; Santoro et al., 2008; Cao et al., 2011).
The abundance and transcription of bacterial amoA genes are reduced with decreasing dissolved oxygen (DO), but DO changes do not significantly influence the diversity of archaeal amoA genes (Abell et al., 2011). The archaeal amoA genes are typically highly abundant in oceanic oxygen-minimum or -deficient zones (Molina et al., 2010; Pitcher et al., 2011; Peng et al., 2013). On the other hand, an increased relative abundance of archaeal amoA genes over bacterial amoA genes under low DO conditions in some estuaries was also reported (Santoro et al., 2008; Qin et al., 2017). According to a recent study, oxygen availability drives the Thaumarchaeota evolution (Ren et al., 2019), and is important for the expansion of AOA habitats from the terrestrial to hadopelagic ecosystems.
Light may also influence the AOA distribution. Archaeal AMO enzymes are more sensitive to photoinhibition than bacterial AMO enzymes (Merbt et al., 2012), which leads to AOA enrichment in the bottom water layers of estuaries (Zou et al., 2019). As another factor, temperature controls the diversity and distribution of AOA in the Westerschelde Estuary (Sahan and Muyzer, 2008). That is because variations in temperature may affect substrate availability, essential for the optimal microbial growth (Andersson et al., 2006). Seasonal variations of light and temperature are important factors controlling the distribution of different AOA species (Liu Q. et al., 2018). AOA are suggested as major players in the nitrogen cycle in low-nutrient environments (Erguder et al., 2009), such as the oligotrophic oceans (Francis et al., 2005). They are also reported as the major ammonia oxidizers in some eutrophic estuaries (Beman and Francis, 2006; Bernhard and Bollmann, 2010; Cao et al., 2011; Jin et al., 2011; Wang et al., 2014; Zou et al., 2019, 2020b) and lakes (Wu et al., 2010; Hou et al., 2013; Zhao et al., 2013; Bollmann et al., 2014). The impact of these environmental parameters, especially the salinity and ammonium concentration, on the distribution of AOA varies with species (Fukushima et al., 2012; Hatzenpichler, 2012; Stahl and de la Torre, 2012), which implies that AOA are well adapted to different environments.
Genomic Features of Estuarine AOA
Comparative analysis supports the notion that the genomic differences among AOA ecotypes are strongly associated with the habitat. For example, the gene kefA, encoding a K+ transporter, is rarely present in the terrestrial AOA group and is mainly harbored by marine AOA, which is interpreted as an adaptation to osmotic pressure in aquatic environments (Ren et al., 2019). Further, members of the shallow water group commonly encode the uvr system and pst systems, which are important for repairing ultraviolet light-induced DNA lesions and phosphate scavenging under phosphorus limitation, respectively; these genes are absent in members of the deep water group, consistent with the physicochemical conditions in deep oceans (Luo et al., 2014; Ren et al., 2019). Oxidative stress is more pronounced in epipelagic water than in deeper water layers, mainly because of a higher abundance of reactive oxygen species generated by many photochemical and photosynthetic processes (Diaz et al., 2013). Consequently, AOA from the epipelagic clade encode more genes related to superoxide dismutase than the bathypelagic clade (Luo et al., 2014). Further, genes related to signal transduction and regulation mechanisms are significantly enriched in the epipelagic clade compared with the mesopelagic clade (Rodriguez-Brito et al., 2006), which may be crucial for their adaptation to the changing upper oceanic environment.
Generally, based on the Cluster of Orthologous Groups (COG) categories, estuarine AOA have a higher proportion of genomic content related to transport and metabolism of amino acids, nucleotides, and lipids than marine AOA (Ngugi et al., 2015). Among many AOA isolates and enrichment cultures, the low-salinity type Candidatus Nitrosoarchaeum limnia (N. limnia) strain SFB1 enriched from estuarine sediments has several unique genomic features compared with other marine or soil AOA (Blainey et al., 2011; Mosier et al., 2012b). Growth experiments suggest that the N. limnia preferentially dwells in low-salinity environments, yet can grow in freshwater and under high-salinity conditions, which presumably is advantageous for adaptation to the estuarine environment, where the salinity typically fluctuates with the seasons (Mosier et al., 2012b). Genomic analysis revealed osmotic adaptation and niche differences between the halotolerant Nitrosopumilus-like RSA3 from the Red Sea Basin and other Nitrosopumilus-like AOA from epi- and mesopelagic oceans (Ngugi et al., 2015). Similarly, the N. limnia genome encodes many mechanosensitive channel proteins, which are necessary to protect microbes from hypoosmotic shock (Blainey et al., 2011). As confirmed by the electron microscopy, N. limnia cells are actively motile as they have flagella (Blainey et al., 2011). The genome of N. limnia, as well as other estuarine AOA enrichment cultures (strain BG20 and strain BD31) encode genes associated to flagellar biosynthesis and chemotaxis (Blainey et al., 2011; Mosier et al., 2012a).
The physicochemical parameters of estuaries change mainly as a result of river discharge, and it appears that high cell motility of estuarine AOA may be essential for their response to the variations in substrates and oxygen levels. Phosphate usually limits the growth of aquatic microbes in estuarine and coastal regions. Accordingly, estuarine AOA reportedly have many genes related to phosphate transport and regulation systems related to phosphate acquisition (Mosier et al., 2012b; Zou et al., 2019). Alternatively, they may utilize diverse types of phosphorus sources by using polyphosphate enzymes and phosphatases (Qin et al., 2020; Zou et al., 2020a). In eutrophic estuaries, such as the Pearl River Estuary and the Jiulong River Estuary, AOA genomes encode extra genes involved in heavy metal transport and regulation systems, and carbohydrate metabolisms, which may be important strategies to adapt the eutrophication and heavy metal pollution in these estuaries (Zou et al., 2019, 2020a).
Overall, estuarine AOA have evolved prominent coping strategies against osmotic pressure, eutrophication, and potential phosphorus limitation, and can sense and actively seek favorable microenvironments, perhaps via cell motility, thus forming a distinct ecotype in estuaries. However, only few published papers focused on the genomic and physiological features of estuarine AOA. Accordingly, more emphasiss should be placed on depicting their metabolic activity and contribution to nitrification.
Recent Findings and Research Hotspots Related to Thaumarchaeota
The origin and evolution of AOA is one of the research hotspots of recent years. The acquisition of a variant vacuolar-type (V-type) ATPase for AOA, via interphylum horizontal operon transfer, is tightly linked with their habitat expansion to acidic soils and to high-pressure hadopelagic ocean (Wang B. et al., 2019). In addition, the genotypic and gene content variability of marine AOA is driven by phosphorus and ammonia availabilities, and hydrostatic pressure (Qin et al., 2020). The acquisition of high-affinity ammonium transporter (amt) and high-affinity pst transporter for some marine AOA may play a crucial role in oligotrophic deep oceans (Qin et al., 2020). Therefore, variation in ATPase composition and structure, and genes related to ammonia and phosphorus transport may be important strategies for their adaptation to different environments.
Previous studies suggested that some members of Thaumarchaeota are non-AOA and are discovered in diverse environments (Rinke et al., 2013; Weber et al., 2015; Hua et al., 2018; Aylward and Santoro, 2020). Members of non-AOA represent the deepest lineages in the phylogenetic tree of Thaumarchaeota, forming the basal group (Group I.1c) (Beam et al., 2014; Hua et al., 2018). Terrestrial non-AOA are thought to be AOA ancestors. The acquisition of the aerobic ammonia oxidation, cobalamin, and biotin biosynthetic pathways for AOA, followed by their habitat expansion to marine environments, may be mainly driven by oxygen (Ren et al., 2019). Members of non-AOA can grow in both anaerobic and aerobic conditions, and oxygen may influence the diversity and structure of non-AOA community (Biggs-Weber et al., 2020). Since only few genomes and enrichment cultures for non-AOA Thaumarchaeota are available (Kato et al., 2019), most of their metabolic potentials remain underexplored. Hence, more detailed studies are needed to comprehensively describe their physiology.
Some coastal AOA strains rely on the uptake and assimilation of organic carbon compounds and are considered as obligate mixotrophs (Qin et al., 2014). Although α-keto acids (e.g., pyruvate) enhance the activity of some AOA, an evidence of mixotrophy, α-keto acids also play an important role in H2O2 detoxification (Kim et al., 2016). Therefore, it is important to clarify whether AOA depend on organic compounds for energy required for growth, or whether these compounds are used in other metabolic pathways. Members of the Group I.1b Thaumarchaeota (Nitrososphaeraceae) lead a heterotrophic lifestyle in sludge, yet they encode the amoA gene (Mußmann et al., 2011). This indicates not all AOA are obligate autotrophic ammonia oxidizers under certain conditions. However, there is no clear evidence that these heterotrophic AOA cannot live autotrophically or are actually mixotrophs. Intriguingly, a group of heterotopic marine Thaumarchaeota, a sister group close to AOA rather than affiliated to the basal group, lacks the ability to oxidize ammonia and to engage in AOA-specific carbon-fixation (Aylward and Santoro, 2020; Reji and Francis, 2020). Based on metagenomics data, this group is widespread in marine environments, has smaller genome sizes than AOA, and encode the form III ribulose-bisphosphate carboxylase (RuBisCO) that potentially functions in nucleotide scavenging (Aylward and Santoro, 2020; Reji and Francis, 2020). The discovery of this group altered the knowledge of the metabolic diversity of Thaumarchaeota, and established solid connection between the basal non-AOA and AOA groups. Nevertheless, further physiological and biochemical evidence is necessary to confirm these inferred metabolic potentials.
Bathyarchaeota in Estuaries
Diversity of Bathyarchaeota Based on 16S rRNA Genes
Members of Bathyarchaeota were first described more than three decades ago (Barns et al., 1996). Thereafter, several close groups have been clustered into a single lineage, the MCG (Inagaki et al., 2003). The new archaeal phylum Bathyarchaeota was proposed recently to highlight its deep branching within the tree of Archaea (Meng et al., 2014). Although no pure cultures nor enrichment cultures of its members have yet been obtained, the phylum Bathyarchaeota is considered to be a cosmopolitan archaeal lineage with high phylogenetic diversity and abundance (Kubo et al., 2012; Lloyd et al., 2013; Meng et al., 2014; Zhou et al., 2018a). They are found in the terrestrial soil and marine sediments, from pelagic oligotrophic oceans to organic-rich coasts and estuaries (Kubo et al., 2012; Lloyd et al., 2013; Meng et al., 2014; Lazar et al., 2015; Yu et al., 2017; Pan et al., 2019), accounting for a considerable proportion of the microbial community composition in the freshwater and marine sediments (on average, 36 ± 22%) based on the 16S rRNA gene (Fillol et al., 2016). The 16S rRNA gene sequences of the most distant bathyarchaeotal members only share 76% similarity (Fry et al., 2008; Kubo et al., 2012). Consequently, this phylum was categorized into 25 subgroups (Zhou et al., 2018a). Because of river runoff, estuaries harbor many types of terrestrial and marine subgroups of Bathyarchaeota (Zhou et al., 2018a). The relatively high concentration of various organic compounds in sediments might be responsible for the high activity and abundance of Bathyarchaeota in these ecosystems (Lazar et al., 2016; Xiang et al., 2017).
Below, we discuss in detail the community composition of Bathyarchaeota in estuarine sediments based on the archaeal 16S rRNA gene analyses (Figure 4 and Supplementary Tables S1, S5 for detailed information), with the categorization suggested by Zhou et al. (2018a). Although the bathyarchaeotal community varies among estuaries, Bathy-6, -8, -15, and -17 are the predominant subgroups in estuarine sediments, accounting for approximately half of the local bathyarchaeotal community (Figure 4). Other major subgroups identified in estuarine sediments are Bathy-1, -3, -4, -12, -13, and -14, with the remaining community made up of rare subgroups. Xiang et al. (2017) identified several indicator subgroups in estuaries, including Bathy-3, -4, -13, and -16.
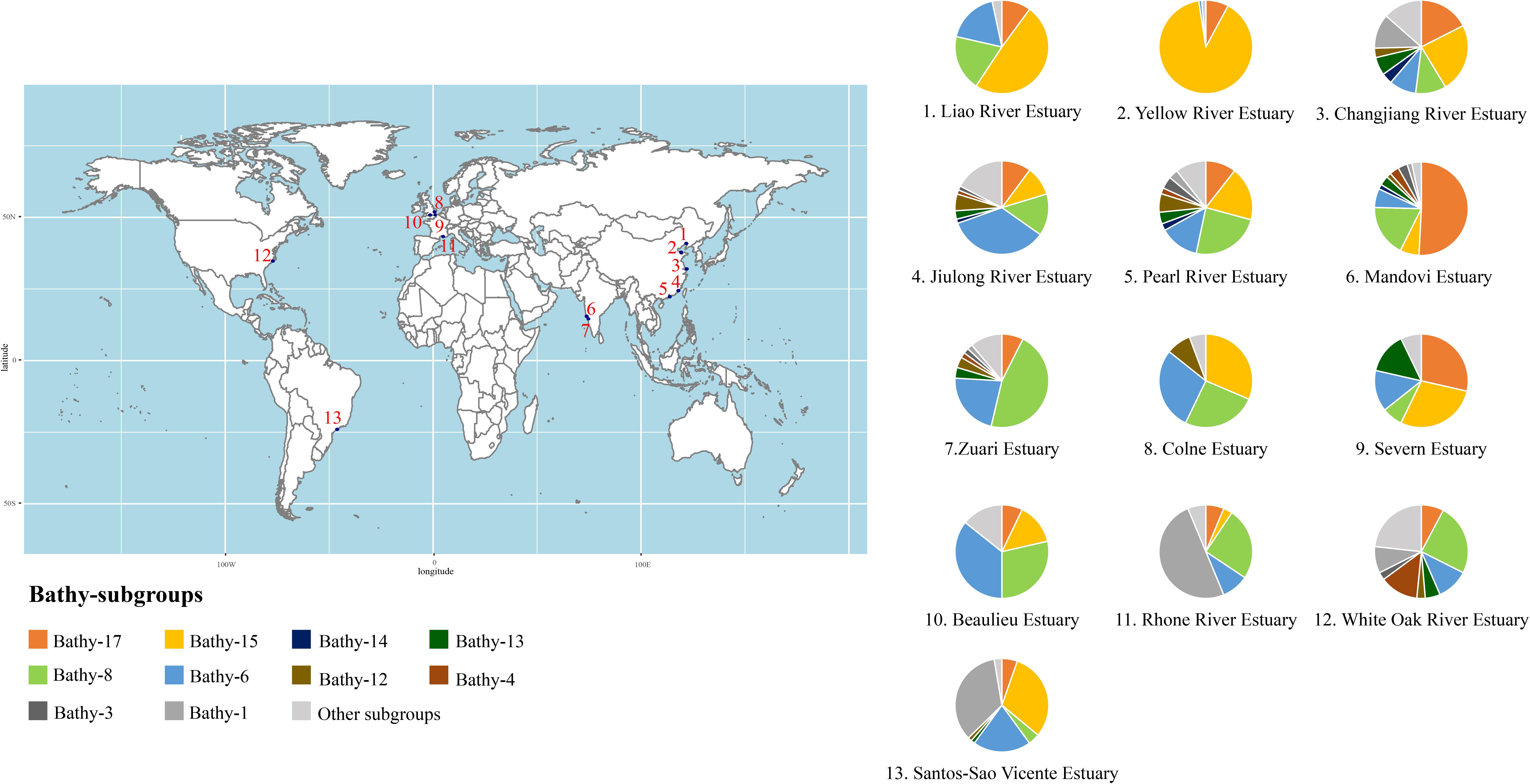
Figure 4. The bathyarchaeotal community composition (based on the 16S rRNA gene sequence) in global estuarine sediments. Please see Supplementary Tables S1, S5 for details.
Distribution Patterns of Bathyarchaeota
The abundance of Bathyarchaeota positively correlates with the concentration of total organic carbon in sediment cores in the South China Sea (Yu et al., 2017) and costal mangroves (Pan et al., 2019, 2020). Enrichment experiments also confirmed that Bathyarchaeota (Bathy-8) are able to grow heterotrophically on some organic carbons, such as lignin (Yu et al., 2018). Studies of the relationship between Bathyarchaeota and different environmental factors suggest that salinity is one of the most important influential factors (Fillol et al., 2016; Xiang et al., 2017). In one study, bathyarchaeotal community in global sediments was clearly separated into freshwater and saline groups based on principal coordinate analysis, and salinity was the best variable to explain the variance within the bathyarchaeotal community, as confirmed by permutational analysis of variance (Fillol et al., 2016). Further, Fillol et al. (2016) suggested that members of the bathyarchaeotal subgroups are bio-indicator lineages reflecting environments of different salinity (i.e., Bathy-1 and -8 are marine indicators and Bathy-5 and -11 are freshwater indicators), and the evolutionary progression of Bathyarchaeota largely occurred in the saline-to-freshwater direction. Because of the lack of pure or enriched cultures, these observations are mainly based on the 16S rRNA genes analysis. Hence, a detailed mechanistic proof of the influence of salinity on Bathyarchaeota should be obtained via genomics and, transcriptomic approaches, and in culture-based experiments.
Salinity and other environmental parameters are important for shaping the bathyarchaeotal community in costal and estuarine regions (Liu et al., 2014; Lazar et al., 2015; Yoshimura et al., 2018; Zhou et al., 2018b; Pan et al., 2019; Chen Y. et al., 2020; Zou et al., 2020b). The total abundance of Bathyarchaeota increases with the sediment depths in the Pearl River Estuary (Liu et al., 2014), and decreases with the decreasing reductive redox conditions in the sediment in the White Oak River (Lazar et al., 2015), but it remained stable in the surface sediments of the Pearl River Estuary (Zou et al., 2020b) and the Bay of Marennes-Oléron (Hélène et al., 2015). Salinity and ammonium levels are major factors influencing the distribution of Bathyarchaeota in surface sediments of the Pearl River Estuary (Zou et al., 2020b). On the other hand, the pH and oxygen levels, rather than salinity, are contributing factors shaping community structure of Bathyarchaeota community structure in costal mangroves and lakes (Yoshimura et al., 2018; Pan et al., 2019, 2020). Further, in the southern Yellow Sea and northern East China Sea, water depth, temperature, and salinity are the most influential factors determining the distribution of Bathyarchaeota in surface sediments (Chen Y. et al., 2020; Liu et al., 2020). Similarly, water depth is the major variable explaining the partitioned distribution pattern of Bathyarchaeota from the shallow marginal sea to the deep northern South China Sea (Zhou et al., 2018b). The prevalence of Bathyarchaeota in subsurface environments, and their relatively high abundance in costal and estuarine regions with high nutrient levels, suggests an anaerobic heterotrophic lifestyle and a possible ability to degrade organic compounds.
Although Bathyarchaeota are considered to be one of the most persistent and abundant core lineages of the sediment archaeal communities, the distribution of bathyarchaeotal subgroups is thought to be strongly associated with the environment, with a huge diversity within the lineage (Fillol et al., 2016). The Bathyarchaeota community is more complex and diverse in mangrove soils and estuary sediments than in other environments (Xiang et al., 2017). Studies suggest that Bathy-6 preferentially dwells in shallow suboxic sediments with low sulfide levels, while Bathy-8 persists in deeper and more reducing marine environments (Lazar et al., 2015; Zhou et al., 2018a). Even so, the predominant subgroup varies with the environment. For example, Bathy-6 predominates in the shallow water sediments of the northern South China Sea (Zhou et al., 2018b), inner surface sediments of the Pear River Estuary (Zou et al., 2020b) and the White Oak River Estuary (Lazar et al., 2015), as well as surface sediments of mangrove wetlands (Pan et al., 2019) and the karstic Lake Cisó (Fillol et al., 2015). Bathy-8 usually dominates in marine sediments and at greater depth (Lazar et al., 2016; Yu et al., 2017), while also dominating in the near-shore surface sediment samples in the northern East China Sea (Chen Y. et al., 2020). Bathy-15 and Bathy-17 are widely detected in freshwater and marine sediments (Fillol et al., 2015; Xiang et al., 2017), and Bathy-15 abundance is relatively stable with changing depth and redox conditions (Hélène et al., 2015; Yu et al., 2017). The environment-specific distribution of these subgroups underlines the distinct metabolic capacities, potential ecological functions and adaptation strategies of Bathyarchaeota in specific habitats.
Ecological Functions and Microbial Interactions of Estuarine Bathyarchaeota
Based on genomic analysis, Bathyarchaeota are important players in benthic C1 carbon compound cycling, potentially via methane metabolism (Evans et al., 2015) or acetogenesis (He et al., 2016), and they are genetically capable of degrading diverse organic compounds including detrital proteins, polymeric carbohydrates, fatty acids, and aromatic compounds (Meng et al., 2014; Seyler et al., 2014; Evans et al., 2015; Lazar et al., 2016). Their methylotrophic methanogenesis potential was first detected in bathyarchaeotal genome bins from the Surat Basin, which harbor key genes, such as the methyl-coenzyme reductase (MCR) and many methyltransferases, involved in utilizing a wide range of methylated compounds (Evans et al., 2015). Recently, a unique MCR type utilizing butane instead of methane was identified, clustering with the members of Bathyarchaeota, indicating a potential for butane oxidation (Laso-Pérez et al., 2016). Metagenomic analysis also revealed differences in metabolic capacities, substrate preferences, and ecological niches of bathyarchaeotal subgroups (Meng et al., 2014; He et al., 2016; Lazar et al., 2016), indicating metabolic flexibility of Bathyarchaeota.
Genomic analysis suggests that bin BA1 (Bathy-3) can utilize peptide and glucose and bin BA2 (Bathy-8) can degrade fatty-acids, while the lack of ATP-synthase may indicate the restricted substrate-level phosphorylation for energy (Evans et al., 2015). Bathy-6 genome form the White Oak River Estuarine sediments encodes genes involved in degradation of extracellular plant-derived mono- and polysaccharides, while Bathy-15 genome from the same environment encodes extracellular peptidases, underpinning a heterotrophic lifestyle (Lazar et al., 2016). In addition, bathyarchaeotal genomes (Bathy-1, -6, -15, and -17) from the White Oak River Estuary can generate acetyl-CoA autotrophically from CO2 and H2 through the Wood–Ljungdahl pathway, which might be involved in acetate generation (Lazar et al., 2016). Further, based on metagenomic and enzymatic analysis in the Guaymas Basin, He et al. (2016) suggested that members of Bathyarchaeota (Bathy-13, -16, -21, and -22) are acetogens and can utilize diverse organic substrates for fermentation. Genes related to dissimilatory nitrite reduction to ammonium were also identified in these genomes, suggesting a potential capacity for nitrite reduction (Lazar et al., 2016). Recently, diverse nitrogen metabolism related genes were identified in Bathyarchaeota genomes, for example the ammonium transporter (amt), hydroxylamine reductase (hcp), and nitrogenase iron protein (nifH) genes (Pan et al., 2020). Furthermore, potential urea production pathways, including the arginase (rocF) and agmatinase (speB) pathways, were identified in some subgroups (Bathy-6, -8, and -15) in mangrove sediments (Pan et al., 2020). In addition, some Bathyarchaeota may reduce S0 to sulfide using the hydrogenase/sulfur reductase (hydA), while members of Bathy-15 and -17 encode genes related to sulfate reduction and some Bathy-6 genomes harbor thiosulfate reduction genes (Pan et al., 2020). These findings highlight the important roles of Bathyarchaeota in benthic nitrogen and sulfur cycles, and underline their highly diverse metabolism.
Feng et al. (2019) compared genomic capacities of 10 bathyarchaeotal subgroups, and suggested that most Bathyarchaeota generally lead a lifestyle relying on both heterotrophic degradation and autotrophic carbon fixation. Recently, members of Bathy-6, -8, and -20 were found to encode potential anaerobic cobalamin biosynthesis pathways (Pan et al., 2020). Further, some Bathyarchaeota (Bathy-6) are potentially involved in light-sensing as they have genes for rhodopsin and porphyrin biosynthesis (Pan et al., 2020). In addition, a gene for the Form III RuBisCO and other genes related to the Calvin-Benson-Bassham cycle were identified in members of Bathyarchaeota (including Bathy-6, -8, -15, and -17), and their expression was supported by transcript analysis (Pan et al., 2020). This indicates that they might fix CO2 via multiple pathways, such as the Wood–Ljungdah pathway and Calvin-Benson-Bassham cycle. Although these findings expand the known metabolic potential of archaea and highlight the pivotal role of Bathyarchaeota in benthic carbon cycling, future experiments investigating substrate specificity of these proteins and analyses of the intermediate metabolites will help establish their actual functions.
Bathyarchaeota may closely interact with other microbes via the methanogenic and acetogenic processes. For example, other heterotrophic and acetoclastic microbes may feed on acetate generated by Bathyarchaeota (He et al., 2016; Lazar et al., 2016), while genomic inference suggests that Bathyarchaeota may have an anaerobic methane oxidizing capacity. Possible interactions between Bathyarchaeota, sulfate-reducing bacteria and anaerobic methane-oxidizing archaea were proposed (Evans et al., 2015; Zhou et al., 2018a). According to Xiang et al. (2017), Bathyarchaeota serve as “keystone species” in diverse environments, maintaining the stability and adaptability of the archaeal community. Potential symbiotic or synergistic relationships between Bathyarchaeota and Thermoprofundales (marine benthic group-D, MBG-D) (Zhou et al., 2019) were genetically inferred, as they share similar pathways, including acetogenesis and protein-degradation pathways (He et al., 2016; Lazar et al., 2016). However, most of these hypotheses are drawn based on limited genomic information. The detailed metabolic functions and interactions need further physiological exploration using more precise and rigorous experiments.
In summary, although no pure cultured strains of Bathyarchaeota have been established to date, the current knowledge of this diverse archaeal phylum in terms of their distribution patterns and metabolic functions is expanding, implying the significance of Bathyarchaeota in global biogeochemical cycling.
Further Research Prospects Related to Bathyarchaeota
Bathyarchaeota are highly diverse. Generally, each subgroup falls into a family or order level based on the 16S rRNA genes (Yarza et al., 2014), while the classification of subgroups varies in different studies. Phylogenetic trees based on 16S rRNA gene sequence and ribosomal proteins inferred from the available genomes of Bathyarchaeota from all subgroups share similar topology (Zhou et al., 2018a). This suggests that the current systematic nomenclature and classification of the 25 Bathyarchaeota subgroups is clear and definite. However, few 16S rRNA gene sequences remain ungrouped in the phylogenetic tree (Zhou et al., 2018a), making it difficult to predict their metabolic potentials, as representative genomes are lacking. Accordingly, the diversity of Bathyarchaeota deserves further exploration. Feng et al. (2019) reported that the deep-rooted Bathy-21 and -22 subgroups from hydrothermal environments might represent ancient types of Bathyarchaeota. In a recent study, several Bathyarchaeota genomes were retrieved from sediments of the Costa Rica margin subseafloor by metagenomics; they form a novel lineage in the phylogenetic tree, with suggestions that this lineage may couple methylotrophy to acetogenesis via the Wood–Ljungdahl pathway (Farag et al., 2020). We can expect a more comprehensive understanding of the diversity and evolution of Bathyarchaeota with the advances in sequencing technology and analysis approaches.
Previous studies also suggested that members of Bathyarchaeota grow on lignin (Yu et al., 2018), which offers valuable information for the enrichment of Bathyarchaeota. Similarly, the relative abundance of Bathyarchaeota increases upon the addition of humic acid and fulvic acid to paddy soils (Yi et al., 2019). Further, abundance of bathyarchaeotal 16S rRNA genes increases over 30-day incubation both, in biofilms supplemented with humic acids and in sediments supplemented with tryptophan (Compte-Port et al., 2020). The knowledge of the physiology, metabolic capacities, and adaptive strategies for Bathyarchaeota mainly based on genomic inference and is still rudimentary. Hence, successful enrichments or pure cultures, with the ensuing physiology and biochemistry experiments would accelerate research into these mysterious archaea, confirm the inferred genomic features, and allow improved understanding of their diversity and adaptation.
Other Active Archaeal Groups Inhabited in Estuaries
Although their abundance is relatively lower than that of the dominant Thaumarchaeota and Bathyarchaeota, other archaeal groups that inhabit in the estuaries may be also crucial players in the estuarine biochemical cycles, such as the methanogens (Jiang et al., 2011; Hu et al., 2016; Chen S. et al., 2020), Marine Group II (MGII) (Massana et al., 2000; Liu et al., 2014; Zhang C. L. et al., 2015; Xie et al., 2018), Thermoprofundales (Biddle et al., 2006; Lloyd et al., 2013; Zhou et al., 2019), and Asgard (Zaremba-Niedzwiedzka et al., 2017; MacLeod et al., 2019) and DPANN archaea (Rinke et al., 2013; Castelle et al., 2015).
Involvement of Diverse Euryarchaeota in Different Biogeochemical Cycles
The presence of MGII archaea in marine water was first reported over two decades ago. They are important planktonic archaea present in surface waters of both pelagic oceans, and in the coastal or estuarine environments, from the tropics to polar regions (Massana et al., 2000; Zhang C. L. et al., 2015). Currently, MGII archaea are proposed to be an order-level lineage namely, Candidatus Poseidoniales in the phylum Euryarchaeota, with 21 genera affiliated to two families, Candidatus Poseidonaceae fam. nov. (formerly subgroup MGIIa), and Candidatus Thalassarchaeaceae fam. nov. (formerly subgroup MGIIb) (Rinke et al., 2019). Genome inferred metabolic potential suggests a photoheterotrophic lifestyle for most MGII, and the capacity to degrade proteins, lipids, and other organic compounds, with varied cell motility among different genera (Rinke et al., 2019). The wide distribution of this photoheterotrophic archaeal plankton indicates their important role in carbon cycling, especially in surface waters.
Methanogenesis accounts for a large portion of global methane emission. This process is mainly performed by diverse methanogenic archaea utilizing H2/CO2, methyl compounds, or acetate to anaerobically generate methane (Thauer et al., 2008). Early studies suggested that methanogens belong to six Euryarchaeota genera, as class I methanogens (Methanococcales, Methanopyrales, and Methanobacteriales) and class II methanogens (Methanosarcinales, Methanomicrobiales, and Methanocellales) (Bapteste et al., 2005; Adam et al., 2017). Recently, several new methanogenic archaeal groups within Euryarchaeota were identified based on the mcrA gene (encoding α-subunit of the methyl-coenzyme M reductase), including Methanomassiliicoccales (formerly RC-III) (Borrel et al., 2014), Methanofastidiosa (formerly WSA2) (Nobu et al., 2016) and Methanonatronarchaeia (Sorokin et al., 2017). Further, archaeal mcrA genes have been identified in different archaeal phyla, such as Bathyarchaeota (Evans et al., 2015), Verstraetearchaeota (Vanwonterghem et al., 2016), Geoarchaeota (Wang Y. et al., 2019), and Nezhaarchaeota (Wang Y. et al., 2019), indicating a vast diversity of potential methanogenic archaea.
Methanogenic communities have been described in diverse soil and sedimentary habitats, such as paddies, wetlands, lakes, estuaries, and geothermal or hydrothermal environments (Thauer et al., 2008; Serrano-Silva et al., 2014; Wen et al., 2017). The structure and distribution of methanogens are driven by a series of physicochemical parameters in estuaries. For example, pH strongly influenced the activity of methanogens that use acetate or H2 (Kim et al., 2004; Kotsyurbenko et al., 2007), and increasing salinity reportedly to inhibits hydrogenotrophic methanogens, while enhancing acetoclastic methanogenesis (Liu et al., 2016). Hence, the community structure of estuarine methanogens usually exhibits seasonal or spatial variation (Chen S. et al., 2020; Zhang et al., 2020). Among the different habitats, the highest richness of methanogenic linages is observed in estuary sediments (Wen et al., 2017), suggesting higher diversity of methanogens in estuaries than in other habitats.
In addition to Bathyarchaeota, Thermoprofundales (formally MBG-D archaea) is another important sedimentary archaeal group ubiquitously distributed in marine subsurface ecosystems, and considerably contributing to the benthic biogeochemical cycles (Biddle et al., 2006; Lloyd et al., 2013). According to their unique phylogenetic position and metabolic potentials, MBG-D archaea were recently proposed as a new order Thermoprofundales within the class Thermoplasmata in the phylum Euryarchaeota, with 16 subgroups (Zhou et al., 2019). MBG-D archaea are most abundant in marine sediments and coastal regions, such as mangroves and estuaries, and subgroup distribution is strongly associated with specific environments, illustrating niche-specific adaptation (Zhou et al., 2019). Metagenomics provide insight into the metabolic potential and ecological functions of MBG-D archaea. Such potential includes exogenous protein mineralization, acetate and ethanol through generation via fermentation, and autotrophic growth linked to the Wood–Ljungdahl pathway (Lloyd et al., 2013; Lazar et al., 2017; Zhou et al., 2019). Microbial co-occurrence analyses indicate close interactions of MBG-D archaea with Bathyarchaeota, Lokiarchaeota, and anaerobic methanotrophic archaea in diverse environments (Zhou et al., 2019), suggesting potential synergistic or syntrophic relationships between these archaeal groups, and highlighting the important role of MBG-D archaea in benthic ecosystems.
The Ubiquitous and Diverse Asgard Archaea
Asgard archaea are considered as the bridge between eukaryotes and prokaryotes because they are phylogenetically close to eukaryotic cells and encode many genes related to eukaryotic signature proteins (Zaremba-Niedzwiedzka et al., 2017). Asgard archaea have been commonly described in a wide range of habitats, from freshwater to marine environments, such as anaerobic marine, estuarine and limnic sediments, and pelagic waters (MacLeod et al., 2019). They are also more abundant in methane-rich or hydrothermal environments than in terrestrial soil or freshwater environments Among Asgard archaea, Lokiarchaeota and Thorarchaeota are observed in diverse environments (Zaremba-Niedzwiedzka et al., 2017; Liu Y. et al., 2018; MacLeod et al., 2019; Cai et al., 2020), Odinarchaeota are most abundant in geothermal environments, and Heimdallarchaeota are enriched in marine sediments. Genomic explorations reveal that Asgard archaea might be important players in the nitrogen and sulfur cycles (MacLeod et al., 2019). Different lineages have diverse metabolic potentials. For example, Lokiarchaeota are proposed to utilize halogenated organic matter and are hydrogen-dependent (Sousa et al., 2016; Manoharan et al., 2019); Thorarchaeota are thought to be mixotrophy and may be capable of acetogenesis (Seitz et al., 2016; Liu Y. et al., 2018); Heimdallarchaeota have the potential to sense light (Pushkarev et al., 2018; Bulzu et al., 2019); Helarchaeota possibly oxidize hydrocarbon in anaerobic environments (Seitz et al., 2019); and Gerdarchaeota may use both organic and inorganic carbon (Cai et al., 2020). Furthermore, Asgard archaea are proposed to play key roles in diverse environments via microbe−microbe interactions, such as signaling, metabolite exchange, and other biotic or physicochemical activities (MacLeod et al., 2019; Imachi et al., 2020).
The Ubiquitous and Diverse DPANN Archaea
The discovery of the ultra-small ecto-symbiotic archaeon Nanoarchaeum equitans has changed our understanding of archaea. It is understood to be a parasitic ectosymbiont of Ignicoccus and other Crenarchaeota, and belongs to a separate new archaeal phylum Nanoarchaeota (Huber et al., 2002; Waters et al., 2003). Additional lineages closely related to Nanoarchaeota have since been identified via single-cell and metagenomic approaches. This led to the recognition of the superphylum DPANN, as most of these microbes have small genomes and limited metabolic capacity (Rinke et al., 2013; Castelle et al., 2015).
Woesearchaeota (formerly Deep-sea Hydrothermal Vent Euryarchaeota Group 6, DHVEG-6) are one of the most ubiquitous and abundant archaeal phyla in the superphylum DPANN (Teske and Sørensen, 2008; Castelle et al., 2015). Their habitats span terrestrial waters and soils (Castelle et al., 2015), costal and estuarine regions (Long et al., 2016; Liu et al., 2018b), marine environments (Ding et al., 2017), and even the human microbiome (Koskinen et al., 2017). Based on phylogenetic analysis, Liu et al. (2018a) proposed 26 subgroups for this highly diverse phylum, and suggested oxygen might as a significant factor driving the distribution and evolution of Woesearchaeota. Low- and mid-latitude estuaries harbor a higher diversity and abundance of Woesearchaeota than high-latitude estuaries, and the community structure is mainly partitioned by oxygen, salinity, and temperature (Liu et al., 2018a). An anaerobic syntrophic lifestyle with obvious metabolic deficiencies has been suggested for members of Woesearchaeota, highlighting the requirement for metabolic complementarity with other microbes (Castelle et al., 2015; Liu et al., 2018a). Genomic characteristics suggest that Woesearchaeota may be able to generate acetate and hydrogen (Liu et al., 2018a). Network and metabolic modeling imply a possible syntrophic relationship between Woesearchaeota and some methanogens. For example, Woesearchaeota might support the growth of some H2/CO2-using and acetate-using methanogens to compete with hydrogenotrophic and acetotrophic methanogens, in exchange receiving amino-acids and other compounds (Liu et al., 2018a). Deciphering the functional importance and ecological role of the vastly diverse DPANN archaea is crucial for improved understanding of the evolution of symbiosis in the archaeal domain of life.
Summary and Perspectives
Archaea are currently understood to be ubiquitously distributed in diverse environments, and are recognized as important players in the global biogeochemical cycles. Advances in culture-independent technologies facilitate the research into most archaeal groups, which expands our understanding of their diversity, distribution, metabolic potentials, and ecological niches. Estuaries usually harbor a highly diverse active microbial community, largely because of the mixing of terrestrial and oceanic species via river runoff. The community composition and distribution patterns of archaea in different estuaries are widely studied. Thaumarchaeota, Bathyarchaeota, and Euryarchaeota are the most abundant estuarine archaeal phyla. Among them, Thaumarchaeota and Bathyarchaeota are more abundant in water columns and sediments, respectively, while Euryarchaeota inhabit both aquatic and sedimentary environments. Many environmental factors drive the archaeal distribution in estuaries, such as geographic location, salinity, and oxygen levels, while the response to these factors varies among the different archaeal groups. Although the knowledge of estuarine archaea is increasing, most studies are mainly based on the analysis of archaeal 16S rRNA gene or other functional genes. Hence, a more detailed view of the metabolic features and environmental adaptations of estuarine archaea is needed. The future research focus may include:
(1) Detailing genomic characteristics of estuarine archaea using metagenomic approaches and single cell sequencing technologies to better delineate their adaptation strategies.
(2) Dissecting the relationship and dependency between certain types of estuarine archaea and environmental factors in enrichment experiments.
(3) Conducting modeling and long-term monitoring analyses to predict and verify the influence of seasonal changes on the estuarine archaeal community.
(4) Determining the ecological importance of archaea in estuaries using various geochemical approaches to quantitatively analyze the roles of archaea in biogeochemical cycling.
(5) Cultivating and isolating estuarine archaea to verify their inferred metabolic capacities, thus providing more accurate and detailed understanding for their ecological niche.
Author Contributions
DZ and ML conceived the study. DZ wrote the manuscript with the help from all co-authors. All authors contributed to the article and approved the submitted version.
Funding
This work was financially supported by the National Natural Science Foundation of China (Grant Nos. 91851105 and 31970105); the National Science and Technology Fundamental Resources Investigation Program of China (Grant No. 2019FY100700); the Shenzhen Science and Technology Program (Grant Nos. JCYJ20170818091727570 and KQTD20180412181334790); the Key Project of Department of Education of Guangdong Province (Grant No. 2017KZDXM071); the CAS Interdisciplinary Innovation Team (Grant No. JCTD-2018-16); and the Post-graduate Studentship of SZU-HKUST Joint Ph.D. Program in Marine Environmental Science.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We want to give our sincere thanks to the editor, all reviewers, and Dr. Joanna Mackie for their detailed advices and suggestions.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2020.02060/full#supplementary-material
References
Abell, G. C. J., Banks, J., Ross, D. J., Keane, J. P., Robert, S. S., Revill, A. T., et al. (2011). Effects of estuarine sediment hypoxia on nitrogen fluxes and ammonia oxidizer gene transcription. FEMS Microbiol. Ecol. 75, 111–122. doi: 10.1111/j.1574-6941.2010.00988.x
Abril, G., and Borges, A. V. (2005). “Carbon dioxide and methane emissions from estuaries,” in Greenhouse Gas Emissions—Fluxes and Processes, eds A. Tremblay, L. Varfalvy, C. Roehm, and M. Garneau (Berlin: Springer), 187–207. doi: 10.1007/3-540-26643-7_7
Adam, P. S., Borrel, G., Brochier-Armanet, C., and Gribaldo, S. (2017). The growing tree of Archaea: new perspectives on their diversity, evolution and ecology. ISME J. 11, 2407. doi: 10.1038/ismej.2017.122
Alla, A. A., Mouneyrac, C., Durou, C., Moukrim, A., and Pellerin, J. (2006). Tolerance and biomarkers as useful tools for assessing environmental quality in the Oued Souss estuary (Bay of Agadir, Morocco). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 143, 23–29. doi: 10.1016/j.cbpc.2005.11.015
Alves, R. J. E., Minh, B. Q., Urich, T., von Haeseler, A., and Schleper, C. (2018). Unifying the global phylogeny and environmental distribution of ammonia-oxidising archaea based on amoA genes. Nat. Commun. 9:1517.
Andersson, M. G., van Rijswijk, P., and Middelburg, J. J. (2006). Uptake of dissolved inorganic nitrogen, urea and amino acids in the Scheldt estuary: comparison of organic carbon and nitrogen uptake. Aqu. Microb. Ecol. 44, 303–315. doi: 10.3354/ame044303
Aylward, F. O., and Santoro, A. E. (2020). Heterotrophic Thaumarchaea with small genomes are widespread in the dark ocean. mSystems 5, e00415–e00420.
Baird, D., Christian, R. R., Peterson, C. H., and Johnson, G. A. (2004). Consequences of hypoxia on estuarine ecosystem function: energy diversion from consumers to microbes. Ecol. Appl. 14, 805–822. doi: 10.1890/02-5094
Baker, B. J., Comolli, L. R., Dick, G. J., Hauser, L. J., Hyatt, D., Dill, B. D., et al. (2010). Enigmatic, ultrasmall, uncultivated Archaea. Proc. Natl. Acad. Sci. U.S.A. 107, 8806–8811. doi: 10.1073/pnas.0914470107
Baker, B. J., De Anda, V., Seitz, K. W., Dombrowski, N., Santoro, A. E., and Lloyd, K. G. (2020). Diversity, ecology and evolution of Archaea. Nat. Microbiol. 5, 1–14.
Bange, H. W., Bartell, U. H., Rapsomanikis, S., and Andreae, M. O. (1994). Methane in the Baltic and North Seas and a reassessment of the marine emissions of methane. Global Biogeochem. Cycles 8, 465–480. doi: 10.1029/94GB02181
Bapteste, É, Brochier, C., and Boucher, Y. (2005). Higher-level classification of the Archaea: evolution of methanogenesis and methanogens. Archaea 1, 353–363. doi: 10.1155/2005/859728
Barns, S. M., Delwiche, C. F., Palmer, J. D., and Pace, N. R. (1996). Perspectives on archaeal diversity, thermophily and monophyly from environmental rRNA sequences. Proc. Natl. Acad. Sci. U.S.A. 93, 9188–9193. doi: 10.1073/pnas.93.17.9188
Barns, S. M., Fundyga, R. E., Jeffries, M. W., and Pace, N. R. (1994). Remarkable archaeal diversity detected in a Yellowstone National Park hot spring environment. Proc. Natl. Acad. Sci. U.S.A. 91, 1609–1613. doi: 10.1073/pnas.91.5.1609
Bar-On, Y. M., Phillips, R., and Milo, R. (2018). The biomass distribution on Earth. Proc. Natl. Acad. Sci. U.S.A. 115, 6506–6511.
Beam, J. P., Jay, Z. J., Kozubal, M. A., and Inskeep, W. P. (2014). Niche specialization of novel Thaumarchaeota to oxic and hypoxic acidic geothermal springs of Yellowstone National Park. ISME J. 8, 938–951. doi: 10.1038/ismej.2013.193
Beman, J. M., and Francis, C. A. (2006). Diversity of ammonia-oxidizing archaea and bacteria in the sediments of a hypernutrified subtropical estuary: Bahia del Tobari. Mexico. Appl. Environ. Microbiol. 72, 7767–7777. doi: 10.1128/aem.00946-06
Bernhard, A. E., and Bollmann, A. (2010). Estuarine nitrifiers: new players, patterns and processes. Estuar. Coast. Shelf Sci. 88, 1–11. doi: 10.1016/j.ecss.2010.01.023
Bernhard, A. E., Landry, Z. C., Blevins, A., José, R., Giblin, A. E., and Stahl, D. A. (2010). Abundance of ammonia-oxidizing archaea and bacteria along an estuarine salinity gradient in relation to potential nitrification rates. Appl. Environ. Microbiol. 76, 1285–1289. doi: 10.1128/aem.02018-09
Biddle, J. F., Lipp, J. S., Lever, M. A., Lloyd, K. G., Sørensen, K. B., Anderson, R., et al. (2006). Heterotrophic Archaea dominate sedimentary subsurface ecosystems off Peru. Proc. Natl. Acad. Sci. U.S.A. 103, 3846–3851. doi: 10.1073/pnas.0600035103
Biggs-Weber, E., Aigle, A., Prosser, J. I., and Gubry-Rangin, C. (2020). Oxygen preference of deeply-rooted mesophilic thaumarchaeota in forest soil. Soil Biol. Biochem. 148, 107848. doi: 10.1016/j.soilbio.2020.107848
Biller, S. J., Mosier, A. C., Wells, G. F., and Francis, C. A. (2012). Global biodiversity of aquatic ammonia-oxidizing archaea is partitioned by habitat. Front. Microbiol. 3:252. doi: 10.3389/fmicb.2012.00252
Bintrim, S. B., Donohue, T. J., Handelsman, J., Roberts, G. P., and Goodman, R. M. (1997). Molecular phylogeny of Archaea from soil. Proc. Natl. Acad. Sci. U.S.A. 94, 277–282. doi: 10.1073/pnas.94.1.277
Blainey, P. C., Mosier, A. C., Potanina, A., Francis, C. A., and Quake, S. R. (2011). Genome of a low-salinity ammonia-oxidizing archaeon determined by single-cell and metagenomic analysis. PLoS One 6:e16626. doi: 10.1371/journal.pone.0016626
Bollmann, A., Bullerjahn, G. S., and McKay, R. M. (2014). Abundance and diversity of ammonia-oxidizing archaea and bacteria in sediments of trophic end members of the Laurentian Great Lakes, Erie and Superior. PLoS One 9:e97068. doi: 10.1371/journal.pone.0097068
Bollmann, A., and Laanbroek, H. J. (2002). Influence of oxygen partial pressure and salinity on the community composition of ammonia-oxidizing bacteria in the Schelde estuary. Aqu. Microb. Ecol. 28, 239–247. doi: 10.3354/ame028239
Borrel, G., Parisot, N., Harris, H. M., Peyretaillade, E., Gaci, N., Tottey, W., et al. (2014). Comparative genomics highlights the unique biology of Methanomassiliicoccales, a Thermoplasmatales-related seventh order of methanogenic archaea that encodes pyrrolysine. BMC Genomics 15:679. doi: 10.1186/1471-2164-15-679
Boynton, W. R., and Kemp, W. M. (1985). Nutrient regeneration and oxygen consumption by sediments along an estuarine salinity gradient. Mar. Ecol. Prog. Ser. Oldendorf 23, 45–55. doi: 10.3354/meps023045
Brochier-Armanet, C., Boussau, B., Gribaldo, S., and Forterre, P. (2008). Mesophilic Crenarchaeota: proposal for a third archaeal phylum, the Thaumarchaeota. Nat. Rev. Microbiol. 6, 245–252. doi: 10.1038/nrmicro1852
Bulzu, P. A., Andrei, A. Ş, Salcher, M. M., Mehrshad, M., Inoue, K., Kandori, H., et al. (2019). Casting light on Asgardarchaeota metabolism in a sunlit microoxic niche. Nat. Microbiol. 4, 1129–1137. doi: 10.1038/s41564-019-0404-y
Caffrey, J. M., Bano, N., Kalanetra, K., and Hollibaugh, J. T. (2007). Ammonia oxidation and ammonia-oxidizing bacteria and archaea from estuaries with differing histories of hypoxia. ISME J. 1, 660–662. doi: 10.1038/ismej.2007.79
Cai, M., Liu, Y., Yin, X., Zhou, Z., Friedrich, M. W., Richter-Heitmann, T., et al. (2020). Diverse Asgard archaea including the novel phylum Gerdarchaeota participate in organic matter degradation. Scie. China Life Sci. 63, 886–897. doi: 10.1007/s11427-020-1679-1
Canfield, D. E., and Thamdrup, B. (2009). Towards a consistent classification scheme for geochemical environments, or, why we wish the term ‘suboxic’would go away. Geobiology 7, 385–392. doi: 10.1111/j.1472-4669.2009.00214.x
Cao, H., Auguet, J. C., and Gu, J. D. (2013). Global ecological pattern of ammonia-oxidizing archaea. PLoS One 8:e52853. doi: 10.1371/journal.pone.0052853
Cao, H., Hong, Y., Li, M., and Gu, J. D. (2011). Diversity and abundance of ammonia-oxidizing prokaryotes in sediments from the coastal Pearl River estuary to the South China Sea. Antonie Van Leeuwenhoek 100, 545–556. doi: 10.1007/s10482-011-9610-1
Castelle, C. J., Wrighton, K. C., Thomas, B. C., Hug, L. A., Brown, C. T., Wilkins, M. J., et al. (2015). Genomic expansion of domain archaea highlights roles for organisms from new phyla in anaerobic carbon cycling. Curr. Biol. 25, 690–701. doi: 10.1016/j.cub.2015.01.014
Chen, S., Wang, P., Liu, H., Xie, W., Wan, X. S., Kao, S. J., et al. (2020). Population dynamics of methanogens and methanotrophs along the salinity gradient in Pearl River Estuary: implications for methane metabolism. Appl. Microbiol. Biotechnol. 104, 1331–1346. doi: 10.1007/s00253-019-10221-6
Chen, Y., Li, S., Yu, Z., Chen, Y., Mi, T., and Zhen, Y. (2020). Characteristics of the Bathyarchaeota community in surface sediments from the southern Yellow Sea and northern East China sea. Estuar. Coast. Shelf Sci. 235:106595. doi: 10.1016/j.ecss.2020.106595
Cheung, S., Mak, W., Xia, X., Lu, Y., Cheung, Y., and Liu, H. (2019). Overlooked genetic diversity of ammonia oxidizing archaea lineages in the global oceans. J. Geophys. Res. Biogeosci. 124, 1799–1811. doi: 10.1029/2018jg004636
Cloern, J. E., Foster, S. Q., and Kleckner, A. E. (2014). Phytoplankton primary production in the world’s estuarine-coastal ecosystems. Biogeosciences 11, 2477–2501. doi: 10.5194/bg-11-2477-2014
Compte-Port, S., Fillol, M., Gich, F., and Borrego, C. M. (2020). Metabolic versatility of freshwater sedimentary archaea feeding on different organic carbon sources. PLoS One 15:e0231238. doi: 10.1371/journal.pone.0231238
Daebeler, A., Herbold, C. W., Vierheilig, J., Sedlacek, C. J., Pjevac, P., Albertsen, M., et al. (2018). Cultivation and genomic analysis of “Candidatus Nitrosocaldus islandicus,” an obligately thermophilic, ammonia-oxidizing Thaumarchaeon from a hot spring biofilm in Graendalur Valley, Iceland. Front. Microbiol. 9:193. doi: 10.3389/fmicb.2018.00193
Dang, H., Zhang, X., Sun, J., Li, T., Zhang, Z., and Yang, G. (2008). Diversity and spatial distribution of sediment ammonia-oxidizing crenarchaeota in response to estuarine and environmental gradients in the Changjiang Estuary and East China Sea. Microbiology 154, 2084–2095. doi: 10.1099/mic.0.2007/013581-0
Danovaro, R., Molari, M., Corinaldesi, C., and Dell’Anno, A. (2016). Macroecological drivers of archaea and bacteria in benthic deep-sea ecosystems. Sci. Adv. 2:e1500961. doi: 10.1126/sciadv.1500961
de la Torre, J. R., Walker, C. B., Ingalls, A. E., Könneke, M., and Stahl, D. A. (2008). Cultivation of a thermophilic ammonia oxidizing archaeon synthesizing crenarchaeol. Environ. Microbiol. 10, 810–818. doi: 10.1111/j.1462-2920.2007.01506.x
DeLong, E. F. (1992). Archaea in coastal marine environments. Proc. Natl. Acad. Sci. U.S.A. 89, 5685–5689.
DeLong, E. F., and Pace, N. R. (2001). Environmental diversity of bacteria and archaea. Syst. Biol. 50, 470–478. doi: 10.1080/10635150118513
Diaz, J. M., Hansel, C. M., Voelker, B. M., Mendes, C. M., Andeer, P. F., and Zhang, T. (2013). Widespread production of extracellular superoxide by heterotrophic bacteria. Science 340, 1223–1226. doi: 10.1126/science.1237331
Diaz, R. J. (2001). Overview of hypoxia around the world. J. Environ. Qual. 30, 275–281. doi: 10.2134/jeq2001.302275x
Ding, J., Zhang, Y., Wang, H., Jian, H., Leng, H., and Xiao, X. (2017). Microbial community structure of deep-sea hydrothermal vents on the ultraslow spreading Southwest Indian ridge. Front. Microbiol. 8:1012. doi: 10.3389/fmicb.2017.01012
Dombrowski, N., Lee, J. H., Williams, T. A., Offre, P., and Spang, A. (2019). Genomic diversity, lifestyles and evolutionary origins of DPANN archaea. FEMS Microbiol. Lett. 366:fnz008.
Elkins, J. G., Podar, M., Graham, D. E., Makarova, K. S., Wolf, Y., Randau, L., et al. (2008). A korarchaeal genome reveals insights into the evolution of the Archaea. Proc. Natl. Acad. Sci. U.S.A. 105, 8102–8107. doi: 10.1073/pnas.0801980105
Eme, L., and Doolittle, W. F. (2015). Microbial diversity: a bonanza of phyla. Curr. Biol. 25, R227–R230.
Eme, L., Spang, A., Lombard, J., Stairs, C. W., and Ettema, T. J. (2017). Archaea and the origin of eukaryotes. Nat. Rev. Microbiol. 15, 711–723. doi: 10.1038/nrmicro.2017.133
Erguder, T. H., Boon, N., Wittebolle, L., Marzorati, M., and Verstraete, W. (2009). Environmental factors shaping the ecological niches of ammonia-oxidizing archaea. FEMS Microbiol. Rev. 33, 855–869. doi: 10.1111/j.1574-6976.2009.00179.x
Erwin, P. M., Pineda, M. C., Webster, N., Turon, X., and Lopez-Legentil, S. (2014). Down under the tunic: bacterial biodiversity hotspots and widespread ammonia-oxidizing archaea in coral reef ascidians. ISME J. 8, 575–588. doi: 10.1038/ismej.2013.188
Evans, P. N., Parks, D. H., Chadwick, G. L., Robbins, S. J., Orphan, V. J., Golding, S. D., et al. (2015). Methane metabolism in the archaeal phylum Bathyarchaeota revealed by genome-centric metagenomics. Science 350, 434–438. doi: 10.1126/science.aac7745
Farag, I. F., Biddle, J. F., Zhao, R., Martino, A. J., House, C. H., and León-Zayas, R. I. (2020). Metabolic potentials of archaeal lineages resolved from metagenomes of deep Costa Rica sediments. ISME J. 14, 1345–1358. doi: 10.1038/s41396-020-0615-5
Feng, X., Wang, Y., Zubin, R., and Wang, F. (2019). Core metabolic features and hot origin of Bathyarchaeota. Engineering 5, 498–504. doi: 10.1016/j.eng.2019.01.011
Fillol, M., Auguet, J. C., Casamayor, E. O., and Borrego, C. M. (2016). Insights in the ecology and evolutionary history of the Miscellaneous Crenarchaeotic Group lineage. ISME J. 10, 665–677. doi: 10.1038/ismej.2015.143
Fillol, M., Sànchez-Melsió, A., Gich, F., and Borrego, C. M. (2015). Diversity of Miscellaneous Crenarchaeotic Group archaea in freshwater karstic lakes and their segregation between planktonic and sediment habitats. FEMS Microbiol. Ecol. 91:fiv020. doi: 10.1093/femsec/fiv020
Flemming, H. C., and Wuertz, S. (2019). Bacteria and archaea on earth and their abundance in biofilms. Nat. Rev. Microbiol. 17, 247–260. doi: 10.1038/s41579-019-0158-9
Francis, C. A., Roberts, K. J., Beman, J. M., Santoro, A. E., and Oakley, B. B. (2005). Ubiquity and diversity of ammonia-oxidizing archaea in water columns and sediments of the ocean. Proc. Natl. Acad. Sci. U.S.A. 102, 14683–14688. doi: 10.1073/pnas.0506625102
Fry, J. C., Parkes, R. J., Cragg, B. A., Weightman, A. J., and Webster, G. (2008). Prokaryotic biodiversity and activity in the deep subseafloor biosphere. FEMS Microbiol. Ecol. 66, 181–196. doi: 10.1111/j.1574-6941.2008.00566.x
Fuhrman, J. A., McCallum, K., and Davis, A. A. (1992). Novel major archaebacterial group from marine plankton. Nature 356, 148–149. doi: 10.1038/356148a0
Fukushima, T., Wu, Y. J., and Whang, L. M. (2012). The influence of salinity and ammonium levels on amoA mRNA expression of ammonia-oxidizing prokaryotes. Water Sci. Technol. 65, 2228–2235. doi: 10.2166/wst.2012.142
Gubry-Rangin, C., Hai, B., Quince, C., Engel, M., Thomson, B. C., James, P., et al. (2011). Niche specialization of terrestrial archaeal ammonia oxidizers. Proc. Natl. Acad. Sci. U.S.A. 108, 21206–21211. doi: 10.1073/pnas.1109000108
Guy, L., and Ettema, T. J. (2011). The archaeal ‘TACK’superphylum and the origin of eukaryotes. Trends Microbiol. 19, 580–587. doi: 10.1016/j.tim.2011.09.002
Harrison, P. J., Yin, K., Lee, J. H. W., Gan, J., and Liu, H. (2008). Physical–biological coupling in the Pearl River Estuary. Cont. Shelf Res. 28, 1405–1415. doi: 10.1016/j.csr.2007.02.011
Hatzenpichler, R. (2012). Diversity, physiology, and niche differentiation of ammonia-oxidizing archaea. Appl. Environ. Microbiol. 78, 7501–7510. doi: 10.1128/aem.01960-12
He, Y., Li, M., Perumal, V., Feng, X., Fang, J., Xie, J., et al. (2016). Genomic and enzymatic evidence for acetogenesis among multiple lineages of the archaeal phylum Bathyarchaeota widespread in marine sediments. Nat. Microbiol. 1:16035.
Hélène, A., Mylène, H., Christine, D., and Céline, L. (2015). “C3 group: a rare but active Thaumarchaeal group in intertidal muddy sediment,” in Proceedings of the Aquatic Sciences Meeting on Association for the Sciences of Limnology & Oceanography, Granada.
Hoshino, T., and Inagaki, F. (2019). Abundance and distribution of Archaea in the subseafloor sedimentary biosphere. ISME J. 13, 227–231. doi: 10.1038/s41396-018-0253-3
Hou, J., Song, C., Cao, X., and Zhou, Y. (2013). Shifts between ammonia-oxidizing bacteria and archaea in relation to nitrification potential across trophic gradients in two large Chinese lakes (Lake Taihu and Lake Chaohu). Water Res. 47, 2285–2296. doi: 10.1016/j.watres.2013.01.042
Howarth, R., Chan, F., Conley, D. J., Garnier, J., Doney, S. C., Marino, R., et al. (2011). Coupled biogeochemical cycles: eutrophication and hypoxia in temperate estuaries and coastal marine ecosystems. Front. Ecol. Environ. 9:18–26. doi: 10.1890/100008
Hu, A., Wang, H., Li, J., Liu, J., Chen, N., and Yu, C. P. (2016). Archaeal community in a human-disturbed watershed in southeast China: diversity, distribution, and responses to environmental changes. Appl. Microbiol. Biotechnol. 100, 4685–4698. doi: 10.1007/s00253-016-7318-x
Hua, Z. S., Qu, Y. N., Zhu, Q., Zhou, E. M., Qi, Y. L., Yin, Y. R., et al. (2018). Genomic inference of the metabolism and evolution of the archaeal phylum Aigarchaeota. Nat. Commun. 9:2832.
Huang, X. P., Huang, L. M., and Yue, W. Z. (2003). The characteristics of nutrients and eutrophication in the Pearl River estuary, South China. Mar. Pollut. Bull. 47, 30–36. doi: 10.1016/S0025-326X(02)00474-5
Huber, H., Hohn, M. J., Rachel, R., Fuchs, T., Wimmer, V. C., and Stetter, K. O. (2002). A new phylum of Archaea represented by a nanosized hyperthermophilic symbiont. Nature 417, 63–67. doi: 10.1038/417063a
Imachi, H., Nobu, M. K., Nakahara, N., Morono, Y., Ogawara, M., Takaki, Y., et al. (2020). Isolation of an archaeon at the prokaryote–eukaryote interface. Nature 577, 519–525.
Inagaki, F., Suzuki, M., Takai, K., Oida, H., Sakamoto, T., Aoki, K., et al. (2003). Microbial communities associated with geological horizons in coastal subseafloor sediments from the Sea of Okhotsk. Appl. Environ. Microbiol. 69, 7224–7235. doi: 10.1128/aem.69.12.7224-7235.2003
Jay, Z. J., Beam, J. P., Dlakić, M., Rusch, D. B., Kozubal, M. A., and Inskeep, W. P. (2018). Marsarchaeota are an aerobic archaeal lineage abundant in geothermal iron oxide microbial mats. Nat. Microbiol. 3, 732–740. doi: 10.1038/s41564-018-0163-1
Jiang, L., Zheng, Y., Chen, J., Xiao, X., and Wang, F. (2011). Stratification of Archaeal communities in shallow sediments of the Pearl River Estuary. Southern China. Antonie Van Leeuwenhoek 99, 739–751. doi: 10.1007/s10482-011-9548-3
Jin, T., Zhang, T., Ye, L., Lee, O. O., Wong, Y. H., and Qian, P. Y. (2011). Diversity and quantity of ammonia-oxidizing Archaea and Bacteria in sediment of the Pearl River Estuary, China. Appl. Microbiol. Biotechnol. 90, 1137–1145. doi: 10.1007/s00253-011-3107-8
Jungbluth, S. P., Amend, J. P., and Rappé, M. S. (2017). Metagenome sequencing and 98 microbial genomes from Juan de Fuca Ridge flank subsurface fluids. Sci. Data 4, 1–11.
Karner, M. B., DeLong, E. F., and Karl, D. M. (2001). Archaeal dominance in the mesopelagic zone of the Pacific Ocean. Nature 409, 507–510. doi: 10.1038/35054051
Kato, S., Itoh, T., Yuki, M., Nagamori, M., Ohnishi, M., Uematsu, K., et al. (2019). Isolation and characterization of a thermophilic sulfur-and iron-reducing thaumarchaeote from a terrestrial acidic hot spring. ISME J. 13, 2465–2474. doi: 10.1038/s41396-019-0447-3
Khandeparker, L., Kuchi, N., Kale, D., and Anil, A. C. (2017). Microbial community structure of surface sediments from a tropical estuarine environment using next generation sequencing. Ecol. Indic. 74, 172–181. doi: 10.1016/j.ecolind.2016.11.023
Kim, I. S., Hwang, M. H., Jang, N. J., Hyun, S. H., and Lee, S. T. (2004). Effect of low pH on the activity of hydrogen utilizing methanogen in bio-hydrogen process. Int. J. Hydro. Energy 29, 1133–1140.
Kim, J. G., Park, S. J., Damsté, J. S. S., Schouten, S., Rijpstra, W. I. C., Jung, M. Y., et al. (2016). Hydrogen peroxide detoxification is a key mechanism for growth of ammonia-oxidizing archaea. Proc. Natl. Acad. Sci. U.S.A. 113, 7888–7893. doi: 10.1073/pnas.1605501113
Könneke, M., Bernhard, A. E., José, R., Walker, C. B., Waterbury, J. B., and Stahl, D. A. (2005). Isolation of an autotrophic ammonia-oxidizing marine archaeon. Nature 437, 543–546. doi: 10.1038/nature03911
Koskinen, K., Pausan, M. R., Perras, A. K., Beck, M., Bang, C., Mora, M., et al. (2017). First insights into the diverse human archaeome: specific detection of archaea in the gastrointestinal tract, lung, and nose and on skin. mBio 8:e00824-17.
Kotsyurbenko, O. R., Friedrich, M. W., Simankova, M. V., Nozhevnikova, A. N., Golyshin, P. N., Timmis, K. N., et al. (2007). Shift from acetoclastic to H2-dependent methanogenesis in a West Siberian peat bog at low pH values and isolation of an acidophilic Methanobacterium strain. Appl. Environ. Microbiol. 73, 2344–2348. doi: 10.1128/AEM.02413-06
Kozubal, M. A., Romine, M., deM Jennings, R., Jay, Z. J., Tringe, S. G., Rusch, D. B., et al. (2013). Geoarchaeota: a new candidate phylum in the Archaea from high-temperature acidic iron mats in Yellowstone National Park. ISME J. 7, 622–634. doi: 10.1038/ismej.2012.132
Kubo, K., Lloyd, K. G., Biddle, J. F., Amann, R., Teske, A., and Knittel, K. (2012). Archaea of the Miscellaneous Crenarchaeotal Group are abundant, diverse and widespread in marine sediments. ISME J. 6, 1949–1965. doi: 10.1038/ismej.2012.37
Laso-Pérez, R., Wegener, G., Knittel, K., Widdel, F., Harding, K. J., Krukenberg, V., et al. (2016). Thermophilic archaea activate butane via alkyl-coenzyme M formation. Nature 539, 396–401. doi: 10.1038/nature20152
Lazar, C. S., Baker, B. J., Seitz, K., Hyde, A. S., Dick, G. J., Hinrichs, K. U., et al. (2016). Genomic evidence for distinct carbon substrate preferences and ecological niches of Bathyarchaeota in estuarine sediments. Environ. Microbiol. 18, 1200–1211. doi: 10.1111/1462-2920.13142
Lazar, C. S., Baker, B. J., Seitz, K. W., and Teske, A. P. (2017). Genomic reconstruction of multiple lineages of uncultured benthic archaea suggests distinct biogeochemical roles and ecological niches. ISME J. 11, 1118–1129. doi: 10.1038/ismej.2016.189
Lazar, C. S., Biddle, J. F., Meador, T. B., Blair, N., Hinrichs, K. U., and Teske, A. P. (2015). Environmental controls on intragroup diversity of the uncultured benthic archaea of the miscellaneous Crenarchaeotal group lineage naturally enriched in anoxic sediments of the White Oak River estuary (North Carolina, USA). Environ. Microbiol. 17, 2228–2238. doi: 10.1111/1462-2920.12659
Li, D., Zhang, J., Huang, D., Wu, Y., and Liang, J. (2002). Oxygen depletion off the Changjiang (Yangtze River) estuary. Sci. China Ser. D Earth Sci. 45, 1137–1146.
Li, M., Baker, B. J., Anantharaman, K., Jain, S., Breier, J. A., and Dick, G. J. (2015). Genomic and transcriptomic evidence for scavenging of diverse organic compounds by widespread deep-sea archaea. Nat. Commun. 6:8933.
Liu, J., Yang, H., Zhao, M., and Zhang, X. H. (2014). Spatial distribution patterns of benthic microbial communities along the Pearl Estuary, China. Syst. Appl. Microbiol. 37, 578–589. doi: 10.1016/j.syapm.2014.10.005
Liu, J., Zhu, S., Liu, X., Yao, P., Ge, T., and Zhang, X. H. (2020). Spatiotemporal dynamics of the archaeal community in coastal sediments: assembly process and co-occurrence relationship. ISME J. 14, 1463–1478. doi: 10.1038/s41396-020-0621-7
Liu, L., Yang, J., and Zhang, Y. (2011). Genetic diversity patterns of microbial communities in a subtropical riverine ecosystem (Jiulong River, southeast China). Hydrobiologia 678, 113–125. doi: 10.1007/s10750-011-0834-x
Liu, Q., Tolar, B. B., Ross, M. J., Cheek, J. B., Sweeney, C. M., Wallsgrove, N. J., et al. (2018). Light and temperature control the seasonal distribution of thaumarchaeota in the South Atlantic bight. ISME J. 12, 1473–1485. doi: 10.1038/s41396-018-0066-4
Liu, X., Li, M., Castelle, C. J., Probst, A. J., Zhou, Z., Pan, J., et al. (2018a). Insights into the ecology, evolution, and metabolism of the widespread Woesearchaeotal lineages. Microbiome 6:102.
Liu, X., Pan, J., Liu, Y., Li, M., and Gu, J. D. (2018b). Diversity and distribution of Archaea in global estuarine ecosystems. Sci. Total Environ. 637, 349–358. doi: 10.1016/j.scitotenv.2018.05.016
Liu, Y., Zhou, Z., Pan, J., Baker, B. J., Gu, J. D., and Li, M. (2018). Comparative genomic inference suggests mixotrophic lifestyle for Thorarchaeota. ISME J. 12, 1021–1031. doi: 10.1038/s41396-018-0060-x
Liu, Y., Priscu, J. C., Xiong, J., Conrad, R., Vick-Majors, T., Chu, H., et al. (2016). Salinity drives archaeal distribution patterns in high altitude lake sediments on the Tibetan Plateau. FEMS Microbiol. Ecol. 92:fiw033.
Liu, Z., Huang, S., Sun, G., Xu, Z., and Xu, M. (2011). Diversity and abundance of ammonia-oxidizing archaea in the Dongjiang River, China. Microbiol. Res. 166, 337–345. doi: 10.1016/j.micres.2010.08.002
Lloyd, K. G., Schreiber, L., Petersen, D. G., Kjeldsen, K. U., Lever, M. A., Steen, A. D., et al. (2013). Predominant archaea in marine sediments degrade detrital proteins. Nature 496, 215–218. doi: 10.1038/nature12033
Long, Y., Yi, H., Chen, S., Zhang, Z., Cui, K., Bing, Y., et al. (2016). Influences of plant type on bacterial and archaeal communities in constructed wetland treating polluted river water. Environ. Sci. Pollut. Res. 23, 19570–19579. doi: 10.1007/s11356-016-7166-3
Longhurst, A., Sathyendranath, S., Platt, T., and Caverhill, C. (1995). An estimate of global primary production in the ocean from satellite radiometer data. J. Plankt. Res. 17, 1245–1271. doi: 10.1093/plankt/17.6.1245
Lu, Z., Gan, J., Dai, M., Liu, H., and Zhao, X. (2018). Joint effects of extrinsic biophysical fluxes and intrinsic hydrodynamics on the formation of hypoxia west off the Pearl River estuary. J. Geophys. Res. Oceans 123, 6241–6259. doi: 10.1029/2018jc014199
Luo, H., Tolar, B. B., Swan, B. K., Zhang, C. L., Stepanauskas, R., Moran, M. A., et al. (2014). Single-cell genomics shedding light on marine Thaumarchaeota diversification. ISME J. 8, 732–736. doi: 10.1038/ismej.2013.202
MacLeod, F., Kindler, G. S., Wong, H. L., Chen, R., and Burns, B. P. (2019). Asgard archaea: diversity, function, and evolutionary implications in a range of microbiomes. AIMS Microbiol. 5, 48–61. doi: 10.3934/microbiol.2019.1.48
Magnabosco, C., Lin, L. H., Dong, H., Bomberg, M., Ghiorse, W., Stan-Lotter, H., et al. (2018). The biomass and biodiversity of the continental subsurface. Nat. Geosci. 11, 707–717. doi: 10.1038/s41561-018-0221-6
Manoharan, L., Kozlowski, J. A., Murdoch, R. W., Löffler, F. E., Sousa, F. L., and Schleper, C. (2019). Metagenomes from coastal marine sediments give insights into the ecological role and cellular features of Loki-and Thorarchaeota. mBio 10:e02039-19.
Martens-Habbena, W., Berube, P. M., Urakawa, H., José, R., and Stahl, D. A. (2009). Ammonia oxidation kinetics determine niche separation of nitrifying Archaea and Bacteria. Nature 461, 976–979. doi: 10.1038/nature08465
Massana, R., DeLong, E. F., and Pedrós-Alió, C. (2000). A few cosmopolitan phylotypes dominate planktonic archaeal assemblages in widely different oceanic provinces. Appl. Environ. Microbiol. 66, 1777–1787. doi: 10.1128/aem.66.5.1777-1787.2000
McLusky, D. S., and Elliott, M. (2004). The Estuarine Ecosystem: Ecology, Threats and Management, Third Edn. Oxford: OUP. 216.
Meng, J., Xu, J., Qin, D., He, Y., Xiao, X., and Wang, F. (2014). Genetic and functional properties of uncultivated MCG archaea assessed by metagenome and gene expression analyses. ISME J. 8, 650–659. doi: 10.1038/ismej.2013.174
Merbt, S. N., Stahl, D. A., Casamayor, E. O., Martí, E., Nicol, G. W., and Prosser, J. I. (2012). Differential photoinhibition of bacterial and archaeal ammonia oxidation. FEMS Microbiol. Lett. 327, 41–46. doi: 10.1111/j.1574-6968.2011.02457.x
Milliman, J. D., Huang-Ting, S., Zuo-Sheng, Y., and Mead, R. H. (1985). Transport and deposition of river sediment in the Changjiang estuary and adjacent continental shelf. Cont. Shelf Res. 4, 37–45. doi: 10.1016/0278-4343(85)90020-2
Moissl-Eichinger, C., Pausan, M., Taffner, J., Berg, G., Bang, C., and Schmitz, R. A. (2018). Archaea are interactive components of complex microbiomes. Trends Microbiol. 26, 70–85. doi: 10.1016/j.tim.2017.07.004
Molina, V., Belmar, L., and Ulloa, O. (2010). High diversity of ammonia-oxidizing archaea in permanent and seasonal oxygen-deficient waters of the eastern South Pacific. Environ. Microbiol. 12, 2450–2465. doi: 10.1111/j.1462-2920.2010.02218.x
Mosier, A. C., Allen, E. E., Kim, M., Ferriera, S., and Francis, C. A. (2012a). Genome sequence of “Candidatus Nitrosopumilus salaria” BD31, an ammonia-oxidizing archaeon from the San Francisco Bay estuary. J. Bacteriol. 194, 2121–2122. doi: 10.1128/jb.00013-12
Mosier, A. C., Lund, M. B., and Francis, C. A. (2012b). Ecophysiology of an ammonia-oxidizing archaeon adapted to low-salinity habitats. Microb. Ecol. 64, 955–963. doi: 10.1007/s00248-012-0075-1
Mosier, A. C., and Francis, C. A. (2008). Relative abundance and diversity of ammonia-oxidizing archaea and bacteria in the San Francisco Bay estuary. Environ. Microbiol. 10, 3002–3016. doi: 10.1111/j.1462-2920.2008.01764.x
Mußmann, M., Brito, I., Pitcher, A., Damsté, J. S. S., Hatzenpichler, R., Richter, A., et al. (2011). Thaumarchaeotes abundant in refinery nitrifying sludges express amoA but are not obligate autotrophic ammonia oxidizers. Proc. Natl. Acad. Sci. U.S.A. 108, 16771–16776. doi: 10.1073/pnas.1106427108
Ngugi, D. K., Blom, J., Alam, I., Rashid, M., Ba-Alawi, W., Zhang, G., et al. (2015). Comparative genomics reveals adaptations of a halotolerant thaumarchaeon in the interfaces of brine pools in the Red Sea. ISME J. 9, 396–411. doi: 10.1038/ismej.2014.137
Nobu, M. K., Narihiro, T., Kuroda, K., Mei, R., and Liu, W. T. (2016). Chasing the elusive Euryarchaeota class WSA2: genomes reveal a uniquely fastidious methyl-reducing methanogen. ISME J. 10, 2478–2487. doi: 10.1038/ismej.2016.33
Nunoura, T., Takaki, Y., Kakuta, J., Nishi, S., Sugahara, J., and Kazama, H., et al. (2011). Insights into the evolution of Archaea and eukaryotic protein modifier systems revealed by the genome of a novel archaeal group. Nucl. Acids Res. 39, 3204–3223. doi: 10.1093/nar/gkq1228
Oremland, R. S., and Polcin, S. (1982). Methanogenesis and sulfate reduction: competitive and noncompetitive substrates in estuarine sediments. Appl. Environ. Microbiol. 44, 1270–1276. doi: 10.1128/aem.44.6.1270-1276.1982
Pan, J., Chen, Y., Wang, Y., Zhou, Z., and Li, M. (2019). Vertical distribution of bathyarchaeotal communities in mangrove wetlands suggests distinct Niche preference of Bathyarchaeota subgroup 6. Microb. Ecol. 77, 417–428. doi: 10.1007/s00248-018-1309-7
Pan, J., Zhou, Z., Béjà, O., Cai, M., Yang, Y., Liu, Y., et al. (2020). Genomic and transcriptomic evidence of light-sensing, porphyrin biosynthesis, Calvin-Benson-Bassham cycle, and urea production in Bathyarchaeota. Microbiome 8:43.
Peng, X., Jayakumar, A., and Ward, B. B. (2013). Community composition of ammonia-oxidizing archaea from surface and anoxic depths of oceanic oxygen minimum zones. Front. Microbiol. 4:177. doi: 10.3389/fmicb.2013.00177
Pester, M., Rattei, T., Flechl, S., Gröngröft, A., Richter, A., Overmann, J., et al. (2012). amoA-based consensus phylogeny of ammonia-oxidizing archaea and deep sequencing of amoA genes from soils of four different geographic regions. Environ. Microbiol. 14, 525–539. doi: 10.1111/j.1462-2920.2011.02666.x
Petitjean, C., Deschamps, P., López-García, P., and Moreira, D. (2014). Rooting the domain archaea by phylogenomic analysis supports the foundation of the new kingdom Proteoarchaeota. Genome Biol. Evol. 7, 191–204. doi: 10.1093/gbe/evu274
Pitcher, A., Villanueva, L., Hopmans, E. C., Schouten, S., Reichart, G. J., and Damsté, J. S. S. (2011). Niche segregation of ammonia-oxidizing archaea and anammox bacteria in the Arabian Sea oxygen minimum zone. ISME J. 5, 1896–1904. doi: 10.1038/ismej.2011.60
Pushkarev, A., Inoue, K., Larom, S., Flores-Uribe, J., Singh, M., Konno, M., et al. (2018). A distinct abundant group of microbial rhodopsins discovered using functional metagenomics. Nature 558, 595–599. doi: 10.1038/s41586-018-0225-9
Qin, W., Amin, S. A., Martens-Habbena, W., Walker, C. B., Urakawa, H., Devol, A. H., et al. (2014). Marine ammonia-oxidizing archaeal isolates display obligate mixotrophy and wide ecotypic variation. Proc. Natl. Acad. Sci. U.S.A. 111, 12504–12509. doi: 10.1073/pnas.1324115111
Qin, W., Meinhardt, K. A., Moffett, J. W., Devol, A. H., Virginia Armbrust, E., Ingalls, A. E., et al. (2017). Influence of oxygen availability on the activities of ammonia-oxidizing archaea. Environ. Microbiol. Rep. 9, 250–256. doi: 10.1111/1758-2229.12525
Qin, W., Zheng, Y., Zhao, F., Wang, Y., Urakawa, H., and Martens-Habbena, W., et al. (2020). Alternative strategies of nutrient acquisition and energy conservation map to the biogeography of marine ammonia-oxidizing archaea. ISME J. 1–15.
Reigstad, L. J., Jorgensen, S. L., and Schleper, C. (2010). Diversity and abundance of Korarchaeota in terrestrial hot springs of Iceland and Kamchatka. ISME J. 4, 346–356. doi: 10.1038/ismej.2009.126
Reigstad, L. J., Richter, A., Daims, H., Urich, T., Schwark, L., and Schleper, C. (2008). Nitrification in terrestrial hot springs of Iceland and Kamchatka. FEMS Microbiol. Ecol. 64, 167–174. doi: 10.1111/j.1574-6941.2008.00466.x
Reji, L., and Francis, C. A. (2020). Metagenome-assembled genomes reveal unique metabolic adaptations of a basal marine Thaumarchaeota lineage. ISME J. 14, 2105–2115. doi: 10.1038/s41396-020-0675-6
Ren, M., Feng, X., Huang, Y., Wang, H., Hu, Z., Clingenpeel, S., et al. (2019). Phylogenomics suggests oxygen availability as a driving force in Thaumarchaeota evolution. ISME J. 13, 2150–2161. doi: 10.1038/s41396-019-0418-8
Rinke, C., Rubino, F., Messer, L. F., Youssef, N., Parks, D. H., Chuvochina, M., et al. (2019). A phylogenomic and ecological analysis of the globally abundant Marine Group II archaea (Ca. Poseidoniales ord. nov.). ISME J. 13, 663–675. doi: 10.1038/s41396-018-0282-y
Rinke, C., Schwientek, P., Sczyrba, A., Ivanova, N. N., Anderson, I. J., Cheng, J. F., et al. (2013). Insights into the phylogeny and coding potential of microbial dark matter. Nature 499, 431–437. doi: 10.1038/nature12352
Rodriguez-Brito, B., Rohwer, F., and Edwards, R. A. (2006). An application of statistics to comparative metagenomics. BMC Bioinform. 7:162. doi: 10.1186/1471-2105-7-162
Sahan, E., and Muyzer, G. (2008). Diversity and spatio-temporal distribution of ammonia-oxidizing Archaea and Bacteria in sediments of the Westerschelde estuary. FEMS Microbiol. Ecol. 64, 175–186. doi: 10.1111/j.1574-6941.2008.00462.x
Santoro, A. E., Francis, C. A., De Sieyes, N. R., and Boehm, A. B. (2008). Shifts in the relative abundance of ammonia-oxidizing bacteria and archaea across physicochemical gradients in a subterranean estuary. Environ. Microbiol. 10, 1068–1079. doi: 10.1111/j.1462-2920.2007.01547.x
Schattenhofer, M., Fuchs, B. M., Amann, R., Zubkov, M. V., Tarran, G. A., and Pernthaler, J. (2009). Latitudinal distribution of prokaryotic picoplankton populations in the Atlantic Ocean. Environ. Microbiol. 11, 2078–2093. doi: 10.1111/j.1462-2920.2009.01929.x
Schleper, C., Jurgens, G., and Jonuscheit, M. (2005). Genomic studies of uncultivated archaea. Nat. Rev. Microbiol. 3, 479–488. doi: 10.1038/nrmicro1159
Schleper, C., and Nicol, G. W. (2010). Ammonia-oxidising archaea–physiology, ecology and evolution. Adv. Microb. Physiol. 57, 1–41. doi: 10.1016/b978-0-12-381045-8.00001-1
Seitz, K. W., Dombrowski, N., Eme, L., Spang, A., Lombard, J., Sieber, J. R., et al. (2019). Asgard archaea capable of anaerobic hydrocarbon cycling. Nat. Commun. 10, 1–11.
Seitz, K. W., Lazar, C. S., Hinrichs, K. U., Teske, A. P., and Baker, B. J. (2016). Genomic reconstruction of a novel, deeply branched sediment archaeal phylum with pathways for acetogenesis and sulfur reduction. ISME J. 10, 1696–1705. doi: 10.1038/ismej.2015.233
Serrano-Silva, N., Sarria-Guzmán, Y., Dendooven, L., and Luna-Guido, M. (2014). Methanogenesis and methanotrophy in soil: a review. Pedosphere 24, 291–307. doi: 10.1016/s1002-0160(14)60016-3
Seyler, L. M., McGuinness, L. M., and Kerkhof, L. J. (2014). Crenarchaeal heterotrophy in salt marsh sediments. ISME J. 8, 1534–1543. doi: 10.1038/ismej.2014.15
Singh, S. K., Verma, P., Ramaiah, N., Chandrashekar, A. A., and Shouche, Y. S. (2010). Phylogenetic diversity of archaeal 16S rRNA and ammonia monooxygenase genes from tropical estuarine sediments on the central west coast of India. Res. Microbiol. 161, 177–186. doi: 10.1016/j.resmic.2010.01.008
Sintes, E., Bergauer, K., De Corte, D., Yokokawa, T., and Herndl, G. J. (2013). Archaeal amoA gene diversity points to distinct biogeography of ammonia-oxidizing Crenarchaeota in the ocean. Environ. Microbiol. 15, 1647–1658. doi: 10.1111/j.1462-2920.2012.02801.x
Sorokin, D. Y., Makarova, K. S., Abbas, B., Ferrer, M., Golyshin, P. N., Galinski, E. A., et al. (2017). Discovery of extremely halophilic, methyl-reducing euryarchaea provides insights into the evolutionary origin of methanogenesis. Nat. Microbiol. 2:17081.
Sousa, F. L., Neukirchen, S., Allen, J. F., Lane, N., and Martin, W. F. (2016). Lokiarchaeon is hydrogen dependent. Nat. Microbiol. 1:16034.
Spang, A., Eme, L., Saw, J. H., Caceres, E. F., Zaremba-Niedzwiedzka, K., Lombard, J., et al. (2018). Asgard archaea are the closest prokaryotic relatives of eukaryotes. PLoS Genet. 14:e1007080. doi: 10.1371/journal.pgen.1007080
Spang, A., Saw, J. H., Jørgensen, S. L., Zaremba-Niedzwiedzka, K., Martijn, J., Lind, A. E., et al. (2015). Complex archaea that bridge the gap between prokaryotes and eukaryotes. Nature 521, 173–179. doi: 10.1038/nature14447
Stahl, D. A., and de la Torre, J. R. (2012). Physiology and diversity of ammonia-oxidizing archaea. Annu. Rev. Microbiol. 66, 83–101. doi: 10.1146/annurev-micro-092611-150128
Steger, D., Ettinger-Epstein, P., Whalan, S., Hentschel, U., De Nys, R., Wagner, M., et al. (2008). Diversity and mode of transmission of ammonia-oxidizing archaea in marine sponges. Environ. Microbiol. 10, 1087–1094. doi: 10.1111/j.1462-2920.2007.01515.x
Stein, L. Y., Jones, G., Alexander, B., Elmund, K., Wright-Jones, C., and Nealson, K. H. (2002). Intriguing microbial diversity associated with metal-rich particles from a freshwater reservoir. FEMS Microbiol. Ecol. 42, 431–440. doi: 10.1111/j.1574-6941.2002.tb01032.x
Stieglmeier, M., Alves, R. J., and Schleper, C. (2014). “The phylum thaumarchaeota,” in The Prokaryotes, eds E. Rosenberg, E. F. DeLong, S. Lory, E. Stackebrandt, and F. Thompson (Berlin: Springer), 347–362. doi: 10.1007/978-3-642-38954-2_338
Teske, A., and Sørensen, K. B. (2008). Uncultured archaea in deep marine subsurface sediments: have we caught them all? ISME J. 2, 3–18. doi: 10.1038/ismej.2007.90
Thauer, R. K., Kaster, A. K., Seedorf, H., Buckel, W., and Hedderich, R. (2008). Methanogenic archaea: ecologically relevant differences in energy conservation. Nat. Rev. Microbiol. 6, 579–591. doi: 10.1038/nrmicro1931
Vanwonterghem, I., Evans, P. N., Parks, D. H., Jensen, P. D., Woodcroft, B. J., Hugenholtz, P., et al. (2016). Methylotrophic methanogenesis discovered in the archaeal phylum Verstraetearchaeota. Nat. Microbiol. 1:16170.
Vieira, R. P., Clementino, M. M., Cardoso, A. M., Oliveira, D. N., Albano, R. M., Gonzalez, A. M., et al. (2007). Archaeal communities in a tropical estuarine ecosystem: Guanabara Bay, Brazil. Microb. Ecol. 54, 460–468. doi: 10.1007/s00248-007-9261-y
Walsh, D. A., Papke, R. T., and Doolittle, W. F. (2005). Archaeal diversity along a soil salinity gradient prone to disturbance. Environ. Microbiol. 7, 1655–1666. doi: 10.1111/j.1462-2920.2005.00864.x
Wang, B., Qin, W., Ren, Y., Zhou, X., Jung, M. Y., Han, P., et al. (2019). Expansion of Thaumarchaeota habitat range is correlated with horizontal transfer of ATPase operons. ISME J. 13, 3067–3079. doi: 10.1038/s41396-019-0493-x
Wang, Y., Wegener, G., Hou, J., Wang, F., and Xiao, X. (2019). Expanding anaerobic alkane metabolism in the domain of Archaea. Nat. Microbiol. 4, 595–602. doi: 10.1038/s41564-019-0364-2
Wang, Z., Wang, Z., Huang, C., and Pei, Y. (2014). Vertical distribution of ammonia-oxidizing archaea (AOA) in the hyporheic zone of a eutrophic river in North China. World J. Microbiol. Biotechnol. 30, 1335–1346. doi: 10.1007/s11274-013-1559-y
Waters, E., Hohn, M. J., Ahel, I., Graham, D. E., Adams, M. D., Barnstead, M., et al. (2003). The genome of Nanoarchaeum equitans: insights into early archaeal evolution and derived parasitism. Proc. Natl. Acad. Sci. U.S.A. 100, 12984–12988. doi: 10.1073/pnas.1735403100
Weber, E. B., Lehtovirta-Morley, L. E., Prosser, J. I., and Gubry-Rangin, C. (2015). Ammonia oxidation is not required for growth of Group 1.1 c soil Thaumarchaeota. FEMS Microbiol. Ecol. 91:fiv001.
Webster, G., O’Sullivan, L. A., Meng, Y., Williams, A. S., Sass, A. M., Watkins, A. J., et al. (2014). Archaeal community diversity and abundance changes along a natural salinity gradient in estuarine sediments. FEMS Microbiol. Ecol. 91, 1–18. doi: 10.1093/femsec/fiu025
Webster, N. S., and Taylor, M. W. (2012). Marine sponges and their microbial symbionts: love and other relationships. Environ. Microbiol. 14, 335–346. doi: 10.1111/j.1462-2920.2011.02460.x
Wen, X., Yang, S., Horn, F., Winkel, M., Wagner, D., and Liebner, S. (2017). Global biogeographic analysis of methanogenic archaea identifies community-shaping environmental factors of natural environments. Front. Microbiol. 8:1339. doi: 10.3389/fmicb.2017.01339
Woese, C. R., and Fox, G. E. (1977). Phylogenetic structure of the prokaryotic domain: the primary kingdoms. Proc. Natl. Acad. Sci. U.S.A. 74, 5088–5090. doi: 10.1073/pnas.74.11.5088
Wu, Y., Xiang, Y., Wang, J., Zhong, J., He, J., and Wu, Q. L. (2010). Heterogeneity of archaeal and bacterial ammonia-oxidizing communities in Lake Taihu, China. Environ. Microbiol. Rep. 2, 569–576. doi: 10.1111/j.1758-2229.2010.00146.x
Xiang, X., Wang, R., Wang, H., Gong, L., Man, B., and Xu, Y. (2017). Distribution of Bathyarchaeota communities across different terrestrial settings and their potential ecological functions. Sci. Rep. 7:45028.
Xie, W., Luo, H., Murugapiran, S. K., Dodsworth, J. A., Chen, S., Sun, Y., et al. (2018). Localized high abundance of Marine Group II archaea in the subtropical Pearl River Estuary: implications for their niche adaptation. Environ. Microbiol. 20, 734–754. doi: 10.1111/1462-2920.14004
Xie, W., Zhang, C., Zhou, X., and Wang, P. (2014). Salinity-dominated change in community structure and ecological function of Archaea from the lower Pearl River to coastal South China Sea. Appl. Microbiol. Biotechnol. 98, 7971–7982. doi: 10.1007/s00253-014-5838-9
Yao, H., Campbell, C. D., Chapman, S. J., Freitag, T. E., Nicol, G. W., and Singh, B. K. (2013). Multi-factorial drivers of ammonia oxidizer communities: evidence from a national soil survey. Environ. Microbiol. 15, 2545–2556. doi: 10.1111/1462-2920.12141
Yarza, P., Yilmaz, P., Pruesse, E., Glöckner, F. O., Ludwig, W., Schleifer, K. H., et al. (2014). Uniting the classification of cultured and uncultured bacteria and archaea using 16S rRNA gene sequences. Nat. Rev. Microbiol. 12, 635–645. doi: 10.1038/nrmicro3330
Yi, X. Y., Yang, Y. P., Yuan, H. Y., Chen, Z., Duan, G. L., and Zhu, Y. G. (2019). Coupling metabolisms of arsenic and iron with humic substances through microorganisms in paddy soil. J. Hazard. Mater. 373, 591–599. doi: 10.1016/j.jhazmat.2019.03.113
Yoshimura, K. M., York, J., and Biddle, J. F. (2018). Impacts of salinity and oxygen on particle-associated microbial communities in the Broadkill River, Lewes DE. Front. Mar. Sci. 5:100. doi: 10.3389/fmars.2018.00100
Yu, T., Liang, Q., Niu, M., and Wang, F. (2017). High occurrence of Bathyarchaeota (MCG) in the deep-sea sediments of South China Sea quantified using newly designed PCR primers. Environ. Microbiol. Rep. 9, 374–382. doi: 10.1111/1758-2229.12539
Yu, T., Wu, W., Liang, W., Lever, M. A., Hinrichs, K. U., and Wang, F. (2018). Growth of sedimentary Bathyarchaeota on lignin as an energy source. Proc. Natl. Acad. Sci. U.S.A. 115, 6022–6027. doi: 10.1073/pnas.1718854115
Zaremba-Niedzwiedzka, K., Caceres, E. F., Saw, J. H., Bäckström, D., Juzokaite, L., Vancaester, E., et al. (2017). Asgard archaea illuminate the origin of eukaryotic cellular complexity. Nature 541, 353–358. doi: 10.1038/nature21031
Zhang, C. J., Chen, Y. L., Pan, J., Wang, Y. M., and Li, M. (2020). Spatial and seasonal variation of methanogenic community in a river-bay system in South China. Appl. Microbiol. Biotechnol. 104, 4593–4603. doi: 10.1007/s00253-020-10613-z
Zhang, C. L., Xie, W., Martin-Cuadrado, A. B., and Rodriguez-Valera, F. (2015). Marine Group II Archaea, potentially important players in the global ocean carbon cycle. Front. Microbiol. 6:1108. doi: 10.3389/fmicb.2015.01108
Zhang, Y., Chen, L., Dai, T., Tian, J., and Wen, D. (2015). The influence of salinity on the abundance, transcriptional activity, and diversity of AOA and AOB in an estuarine sediment: a microcosm study. Appl. Microbiol. Biotechnol. 99, 9825–9833. doi: 10.1007/s00253-015-6804-x
Zhao, D., Zeng, J., Wan, W., Liang, H., Huang, R., and Wu, Q. L. (2013). Vertical distribution of ammonia-oxidizing archaea and bacteria in sediments of a eutrophic lake. Curr. Microbiol. 67, 327–332. doi: 10.1007/s00284-013-0369-7
Zheng, S., Qiu, X., Chen, B., Yu, X., Liu, Z., Zhong, G., et al. (2011). Antibiotics pollution in Jiulong River estuary: source, distribution and bacterial resistance. Chemosphere 84, 1677–1685. doi: 10.1016/j.chemosphere.2011.04.076
Zhou, Z., Liu, Y., Lloyd, K. G., Pan, J., Yang, Y., Gu, J. D., et al. (2019). Genomic and transcriptomic insights into the ecology and metabolism of benthic archaeal cosmopolitan, Thermoprofundales (MBG-D archaea). ISME J. 13, 885–901. doi: 10.1038/s41396-018-0321-8
Zhou, Z., Meng, H., Liu, Y., Gu, J. D., and Li, M. (2017). Stratified bacterial and archaeal community in mangrove and intertidal wetland mudflats revealed by high throughput 16S rRNA gene sequencing. Front. Microbiol. 8:2148. doi: 10.3389/fmicb.2017.02148
Zhou, Z., Pan, J., Wang, F., Gu, J. D., and Li, M. (2018a). Bathyarchaeota: globally distributed metabolic generalists in anoxic environments. FEMS Microbiol. Rev. 42, 639–655. doi: 10.1093/femsre/fuy023
Zhou, Z., Zhang, G. X., Xu, Y. B., and Gu, J. D. (2018b). Successive transitory distribution of Thaumarchaeota and partitioned distribution of Bathyarchaeota from the Pearl River estuary to the northern South China Sea. Appl. Microbiol. Biotechnol. 102, 8035–8048. doi: 10.1007/s00253-018-9147-6
Zou, D., Li, Y., Kao, S. J., Liu, H., and Li, M. (2019). Genomic adaptation to eutrophication of ammonia-oxidizing archaea in the Pearl River estuary. Environ. Microbiol. 21, 2320–2332. doi: 10.1111/1462-2920.14613
Zou, D., Wan, R., Han, L., Xu, M. N., Liu, Y., and Liu, H., et al. (2020a). Genomic characteristics of a novel species of ammonia-oxidizing Archaea from the Jiulong River Estuary. Appl. Environ. Microbiol. 86. doi: 10.1128/AEM.00736-20
Keywords: Thaumarchaeota, Bathyarchaeota, estuarine archaea, distribution, environmental adaptation
Citation: Zou D, Liu H and Li M (2020) Community, Distribution, and Ecological Roles of Estuarine Archaea. Front. Microbiol. 11:2060. doi: 10.3389/fmicb.2020.02060
Received: 15 May 2020; Accepted: 05 August 2020;
Published: 28 August 2020.
Edited by:
Chuanlun Zhang, Southern University of Science and Technology, ChinaReviewed by:
Roland Hatzenpichler, Montana State University, United StatesJong-Geol Kim, Chungbuk National University, South Korea
Shengwei Hou, University of Southern California, United States
Copyright © 2020 Zou, Liu and Li. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Meng Li, bGltZW5nODQ4QHN6dS5lZHUuY24=