 Jolanta Kowalska
Jolanta Kowalska Józef Tyburski2
Józef Tyburski2 Kinga Matysiak
Kinga Matysiak Eligio Malusá
Eligio Malusá- 1Institute of Plant Protection, Poznań, Poland
- 2Department of Agroecosystems, University of Warmia and Mazury in Olsztyn, Olsztyn, Poland
- 3Eurecat, Centre Tecnològic de Catalunya, Tarragona, Spain
- 4Research Institute of Horticulture, Skierniewice, Poland
Bioproducts, i.e., microbial based pesticides or fertilizers (biopesticides and biofertilizers), should be expected to play an ever-increasing role and application in agricultural practices world-wide in the effort to implement policies concerned with sustainable agriculture. However, several microbial strains have proven the capacity to augment plant productivity by enhancing crop nutrition and functioning as biopesticides, or vice-versa. This multifunctionality is an issue that is still not included as a concept and possibility in any legal provision regarding the placing on the market of bioproducts, and indicates difficulties in clearly classifying the purpose of their suitability. In this review, we overview the current understanding of the mechanisms in plant-microbe interactions underlining the dual function of microbial strains toward plant nutrition and protection. The prospects of market development for multifunctional bioproducts are then considered in view of the current regulatory approach in the European Union, in an effort that wants to stimulate a wider adoption of the new knowledge on the role played by microorganisms in crop production.
Introduction
Chemical, synthetic plant protection products (PPPs) and mineral fertilizers are criticized mainly for their potentially negative effects on human health (Bennekou, 2019) and/or the environment (Norse, 2005; Huang et al., 2017) including non-renewable resources depletion (Chojnacka et al., 2020), and overall negative impact on biodiversity (Mozumder and Berrens, 2007; Sánchez-Bayo and Wyckhuys, 2019). Because of these concerns, the application of sustainable crop production methods is required by consumers as well as by legal provisions (Marrone, 2019).
In this context, although microbial-based pesticides or fertilizers (hereafter biopesticides and biofertilizers, respectively) would not be expected to fully replace chemical pesticides and mineral fertilizers, they could play an ever-increasing role and application in agricultural practices world-wide (Alabouvette et al., 2012; Kurek and Ozimek, 2013).
The multifunctional use of bioinocula represents an issue that is emerging from several researches (Harman, 2011; Lopes et al., 2018), and could further support the development, marketing and application of microbial-based products. Yet, this prospective is neither included as a concept nor as a possibility in any legal provision dealing with the marketing of bioproducts, while it has started to be appraised in bioproducts for human consumption (Ma et al., 2019).
In this review we are summarizing recent findings on the multiple effects of microorganisms suitable as biofertilizers or biopesticides, in light of the intricated interactions between plants and microorganisms, in an effort to foster the discussion on new products that could find a better acceptance by farmers because of their multifunctional properties.
Plant-Microbial Inocula Interactions as the Basis for Multifunctional Bioproducts
The interaction between plants and beneficial fungi involves elicitors released by them which include several metabolites, including volatiles (Shoresh et al., 2010; Morath et al., 2012). These compounds function as signal transduction in plants, and as a result both the plant proteome and transcriptome are affected, as it has been observed with Trichoderma (Marra et al., 2006; Shoresh and Harman, 2008; Lorito et al., 2010; Lombardi et al., 2020) or arbuscular mycorrhizal fungi (Jung et al., 2012; Cameron et al., 2013; Rivero et al., 2015; Adolfsson et al., 2017). The effect of these modifications is translated into increased plant growth, particularly under stress, improved nutrient use efficiency, acquisition of a systemic resistance to diseases that goes beyond the commonly induced systemic and acquired resistances (Shoresh et al., 2010; Cameron et al., 2013). Qualitatively similar effects are induced in plants by rhizobacteria: the interactions involve different chemical compounds (Abriouel et al., 2011; Fickers, 2012; Lopes et al., 2015; Jasim et al., 2016) as well as priming (Brencic and Winans, 2005; van Wees et al., 2008). Mechanisms include induction of the plant innate immune response system (Jain et al., 2011) or acquired systemic resistance (Iavicoli et al., 2003; Choudhary and Johri, 2009), alteration of plant functional traits (Friesen et al., 2011) and prevention of pathogen settling (Bakker et al., 2012).
On the other hand, growth promotion in bacteria derives mainly from the synthesis of several plant growth hormones (Arkhipova et al., 2005; Xie et al., 2014; Radhakrishnan and Lee, 2016) or their indirect regulation through production of volatile organic compounds (Tahir et al., 2017; Rath et al., 2018) and 1-aminocyclopropane-1-carboxylate deaminase (Glick et al., 2007), as well as the solubilization or mineralization of mineral nutrients (Malusá et al., 2016). A key role in interaction between plants and microorganisms seems to be played by pattern recognition receptors (PRRs), localized in the plants’ plasma-membrane, which allow to recognize beneficial microbe/pathogen-associated molecular patterns (Boller and Felix, 2009; Zipfel, 2014; Trdá et al., 2015).
Nevertheless, the relation between plants and beneficial microorganisms inocula occurs within a wider framework of interactions, including those with the plant microbiome (Berg et al., 2017; Fadiji and Babalola, 2020) as well as with the soil physical, chemical and biological characteristics (Bardi and Malusá, 2012; Vimal et al., 2017), which all contribute to increase the complexity in developing sustainable management practices and agricultural products such as biofertilizers and biopesticides as well as for better exploiting their characteristics.
Biopesticides and Plant Growth Promotion
Several biopesticides have been developed to protect plants from pests since the mid-twentieth century (Copping and Menn, 2000; de Faria and Wraight, 2007) and among them several entomopathogenic fungi (e.g., Beauveria spp., Zimmermann, 2007) and bacteria (e.g., Bacillus thuringiensis, de Almeida Melo et al., 2016) are currently used in crop protection. However, recently published studies have provided evidence for the involvement of entomo- or myco-pathogenic microorganisms in promoting plant growth, thus opening new opportunities of their multifunctional use (Vega et al., 2009; Lacey et al., 2015; Table 1). Examples with entomopathogenic fungi include the significant increase in onion yields after Metarhizium anisopliae sprays (Maniania et al., 2003) or in growth of soybean seedlings (Khan et al., 2012) or maize plants (Liao et al., 2014) or cotton (Lopez and Sword, 2015) after soil inoculation with different entomopathogenic species. The mechanism of growth promotion is related to the transfer of nitrogen, also from the parasitized pest, which occurred in both leguminous and gramineous species (Behie et al., 2012). However, production of siderophores (Jirakkakul et al., 2015) or increased uptake of iron (Sánchez-Rodríguez et al., 2015) have also been demonstrated to occur in plants colonized with the entomopathogenic B. bassiana. The production of the auxin indole-3-acetic acid was likewise found to be associated to several Metarhizium and Beauveria strains (Liao et al., 2017). Nevertheless, as for the expression of the full efficacy in insect pests’ control, the ability of fungal entomopathogens to promote plant growth has resulted to depend on the inoculation method (Jaber and Enkerli, 2016, 2017) or the inoculation rate (García et al., 2011).
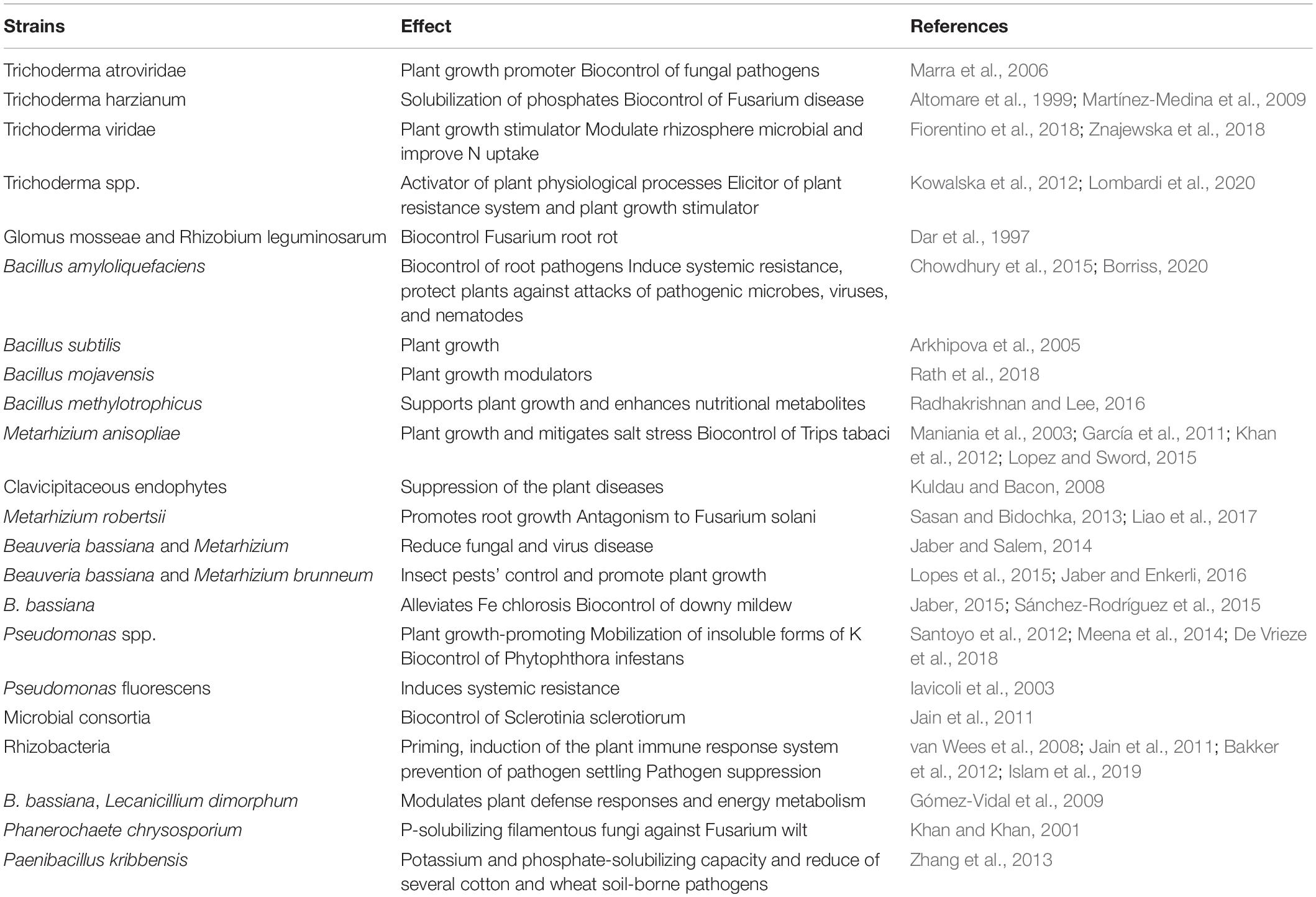
Table 1. Microbial strains showing plant protection and growth promotion effects.
Increased plant growth mediated by entomopathogenic fungi could result from the suppression of the plant diseases (Kuldau and Bacon, 2008; Jaber, 2015) or from a combination of reduced disease severity and more vigorous development of the plants as observed with Beauveria and Metarhizium strains and fungal or virus pathogens (Sasan and Bidochka, 2013; Jaber and Salem, 2014). In these cases, the mechanisms could derive from the capacity of entomopathogenic fungi to elicit the expression of photosynthesis- and energy metabolism-related proteins as well as plant defense responses (Gómez-Vidal et al., 2009).
Among pathogen biocontrol fungi, the dual effect of Trichoderma application has been observed in several studies. T. harzianum T-22 proved to solubilize in vitro insoluble rock phosphate likely by both chelation and reduction processes, since no release of organic acids nor acidification were observed (Altomare et al., 1999). Trichoderma-based products were shown to modulated rhizosphere microbial populations, improving nutrient uptake efficiency, yield, and nutritional quality of leafy vegetables (Fiorentino et al., 2018) or of strawberry plants (Lombardi et al., 2020). Dipping roots of strawberry cuttings in a suspension of T. asperellum prior to planting followed by foliage applications during the vegetation season stimulated plant growth (+ 24%) and health (Kowalska et al., 2012). The effect was reverberated on the control of Botrytis cinerea also on stored fruits, extending their shelf-life without symptoms of damage up to 7 days. Trichoderma spp. isolates significantly reduced the infection of germinating seeds and carrots seedlings by Pythium spp. and efficiently influenced the growth of the seedlings as compared to the standard chemical seed dressing (Sobolewski et al., 2013). Similarly, foliar application of T. asperellum increased seed yield and weight and improved lipid content of organic oilseed rape (Brassica napus L.) (Kowalska, 2014). The mechanism of these plant growth promoting effects could be explained by the growth stimulation, observed with a T. viridae strain, particularly of lateral roots and inhibition of the elongation of hypocotyls, resulting in about fourfold increase of dry biomass in comparison to the control (Znajewska et al., 2018).
Among the bacteria exploited for protection against pathogens, the Bacillus and Pseudomonas genera have common commercial use and frequently are exploited also for plant growth promotion (Santoyo et al., 2012). A rich literature exists on these microorganisms (e.g., Kumar et al., 2011; Chowdhury et al., 2015; Islam et al., 2019) thus the reader is advised to refer to it. However, it is interesting to note that the entomopathogenic B. thuringiensis, was able to in vitro solubilize low-soluble inorganic phosphate and simultaneously produce IAA when formulated in k-carrageenan (Vassilev et al., 2006). This formulation boosted plant growth and P-uptake when introduced into a soil–plant system, stimulating the establishment and development of the co-inoculated endomycorrhizal fungus Glomus deserticola (Vassilev and Vassileva, 2004).
Biocontrol Potential of Biofertilizers
Many plant growth promoters used for inoculation in cropping systems might serve as biocontrol microorganisms (Chowdhury et al., 2015; French, 2017; Table 1). The biocontrol potential of several P-solubilizers has been verified in several works of Vassilev and co-workers (Vassilev et al., 2006). Inoculation with G. intraradices significantly reduced the impact of the soil-borne pathogen F. oxysporum on tomato plants, paralleled by a significant decrease in the number of colony-forming units compared with the control treatment (Vassilev et al., 2008, 2009a). However, the further introduction of a filamentous fungus (A. niger) in the formulation, with different plant wastes and rock phosphate as microbial growing substrate, was more effective to control the pathogen. Similar results were achieved with Phanerochaete chrysosporium (Vassilev et al., 2009b) or in other field trials with P-solubilizing filamentous fungi against Fusarium wilt in tomato (Khan and Khan, 2001). In these cases, the biocontrol function was suggested to be based on production of hydrolytic enzymes or the competition for nutrients and space by the microbial P-solubilizers, as well as by hormones such as indole-3-acetic acid (IAA) and siderophores, being among the metabolites most frequently released by P-solubilizing microorganisms.
The observation that root colonization by AMF is not always associated to improved nutrition and increased vegetative biomass (Smith and Smith, 2011), has prompted to propose that improved stress tolerance is another major benefit of the symbiosis (Gianinazzi et al., 2010) and AM fungi are accepted as functioning in biocontrol (Johansson et al., 2004). Enhanced resistance of mycorrhizal plants to soil-borne pathogen attacks has been associated to the accumulation of phytoalexins, flavonoids, and isoflavonoids in AM-colonized root tissues (Ziedan et al., 2011; Jung et al., 2012). Interestingly, the bioprotective effect of Glomus mosseae against the soilborne pathogen Fusarium oxysporum f.sp. lycopersici was observed in different cultivars and genotypes which differed in their susceptibility to both the AMF and the pathogen (Steinkellner et al., 2012). Nevertheless, the response of phylogenetically diverse plants (i.e., tomato, soybean, and maize) to two mycorrhizal fungi – Funneliformis mosseae and Rhizophagus irregularis – depended on both the plant and the AMF species involved (Fernández et al., 2014). Although fungal pathogens reduce root colonization by AMF, the latter were shown to provide protection through increased enzymes activity, including those directly involved in the regulation of the symbioses. The biological protection of AMF has been also proved on plant parasitic nematodes, under greenhouse conditions: the population of Meloidogyne incognita or Pratilenchus penetrans was lowered by 45 and 87%, respectively, in mycorrhized roots in comparison to non-mycorrhized roots (Vos et al., 2012).
The effect of interactions between AMF and other agronomical practices shows how external factors can contribute to the expression of biocontrol potential. The interaction between AMF and different level of P availability was observed in the occurrence of Alternaria solani symptoms (Fritz et al., 2006). Mycorrhized tomato plants had significantly less A. solani symptoms than non-mycorrhizal plants, but increased P supply, which was paralleled to a reduction in mycorrhiza formation, led to a higher disease severity in mycorrhizal plants. On the other hand, individual co-inoculation of four Glomus species with T. harzianum affected the colony−forming capacity of the latter, but the combined inoculation – particularly with G. intraradices – resulted in a general synergistic effect on disease control (Martínez-Medina et al., 2009). The inoculation of bean plants with Glomus mosseae, besides decreasing propagule number of Fusarium solani in the rhizosphere, decreased pathogenic root rot by 34–77% (Dar et al., 1997). However, when co-inoculated with Rhizobium leguminosarum, the mycorrhized plants were more tolerant of the fungal root pathogen.
The induction of defense activity by AMF has been also proved in above ground tissues. Helicoverpa arimigera larvae feeding on leaves of tomato mycorrhized plants had a reduction of 62.3% in weight relative to non-inoculated plants, likely as a result of a priming effect related to jasmonate pathway (Song et al., 2013). Nevertheless, it could be speculated that the effect on above-ground herbivores derives also from reduced levels of metabolites connected to central catabolic and amino acid metabolism, particularly prominent in sink leaves, which prompted to suggest deteriorations rather than improvements in the nutritional value of colonized plants for higher trophic levels (Fester et al., 2011).
Several genera and species of bacteria (e.g., Pseudomonas or Bacillus) and fungi (e.g., Pennicillium or Aspergillus) ubiquitous in different soils are known to assist plants growth by mobilization of insoluble forms of K (Meena et al., 2014), with mechanisms similar to those found in P-solubilizers (Sheng and He, 2006). It is thus not unexpected that a strain of Paenibacillus kribbensis having potassium and phosphate-solubilizing capacity was also found to reduce the development of several cotton and wheat soil-borne pathogens in vitro (Zhang et al., 2013).
The potential function of plant-growth-promoting rhizobacteria in biocontrol has been long known and can be traced to bacterization studies with fluorescent pseudomonads beginning in the 1970s (Weller, 2007). Since then, a huge amount of studies has allowed to characterize the process of root colonization and the biotic and abiotic factors that are affecting it as well as the identification of genes and traits in bacterial fitness underlying the mechanisms of pathogen suppression (e.g., Labuschagne et al., 2010; Sayyed et al., 2013; Islam et al., 2019). However, notwithstanding this knowledge, the major difficulties and weakness in a broad use of PGPR strains in agricultural practices reside in formulation and registration of the bacteria for commercial use (Malusá et al., 2012; Bashan et al., 2014; Borriss, 2020).
Regulatory Future Perspectives of Multifunctional Bioproducts
A sustainable agriculture is a central pillar of the United Nations Sustainable Development Goals (United Nations, 2015), which can be pursued by the wide adoption of microbial-based products in agronomical practices. The regulatory and policy pressure posed by this international agreement could potentially transform the market of bioproducts into a key segment of the world economy. Such potential is confirmed by recent market analysis reports, which valued at about 10.2 billion USD by 2025 the global biopesticide market, with an annual growth rate of about 15% (Anonymous, 2019b) and projected 3.15 billion USD by the end of 2026 for the biofertilizers market, at an annual growth rate of about 11% (Anonymous, 2019a). However, it is intriguing that for biopesticides, the market value projected for 2025 was already expected to be reached by 2017 (Marrone, 2007).
Most microbial-based PPPs present on the market have been designed for annual crops (mainly legumes and cereals), but there is an increasing demand for these products in fruit and vegetable crops, particularly for organic production. On the other hand, even though biofertilizers would not fully replace mineral fertilizers (Adesemoye et al., 2009), their application, possibly in association with organic fertilizers or other carbon-based products (Saeid and Chojnacka, 2019), could substitute to a large extent mineral or synthetic inputs, having also a positive impact on plant protection strategies.
However, the legal framework regulating the production and marketing of bioproducts can pose a bottleneck to their wider adoption because it reflects the incomplete knowledge on microbial-based products as well as precautions in their safety assessment. Considering the current situation in the EU, known to have a well-developed regulatory framework on agricultural inputs, it emerges that the biopesticide registration process and data requirements are similar to those needed for chemical pesticides (Regulation Eu 1107/2019). Even though a legal provision (Parliament, 2009) has introduced in the EU a compulsory integrated pest management since 2014 for all crops, the unfamiliarity with biologically based pest management of the risk assessors and regulators has not fostered the modification of the authorization process, taking into consideration the peculiarities of the biopesticides mechanisms of action, as it had already been suggested by prominent researchers (Chandler et al., 2011). However, recently, a specific working group has been organized to this aim, and also the European Food Safety Agency has actively operated to find new assessment methods (Council of the European Union, 2019). Interestingly, similar bottlenecks have been hampering biopesticides’ development and use also in the Indian context, paralleled with a legislation aimed to support bioproducts for organic farming which resulted in an unfair competition from sub-standard or misbranded biopesticides (Keswani et al., 2019a).
In case of biofertilizers, the rules have been enacted patchily in the world (Malusá and Vassilev, 2014) and in the European Union only in 2019 a provision has been enacted, though limiting the marketing to just four kinds of biofertilizers: three related to N nutrition (based on symbiotic Rhizobium spp. and free-living Azotobacter spp. and Azospirillum spp.) and one for P nutrition (based on mycorrhizal fungi) (Regulation Eu 1107/2019, 2019). The limitation of species allowed to be marketed as biofertilizers contrasts with the plethora of genera and species recognized having positive effects on plant nutrition and available for commercial applications (Umesha et al., 2018). Furthermore, the EU Regulation allows only the drying or freeze-drying processes in the formulation of the product, which is also restrictive considering the technological possibilities in this respect (Bashan et al., 2014; Vassilev et al., 2020).
Multifunctional bioproducts would also share with biopesticides and biofertilizers the issue of biosafety with respect to humans and the environment as, although only wild-type strains are being used for bioinocula development, the risk of pathogen spread cannot be completely excluded, thus requiring certain precautions (Keswani et al., 2019b).
In view of this situation, it appears quite difficult to expect that multifunctional bioproducts could soon be made available nor that manufacturers would advertise – not being it allowed by the legal provisions – either biocontrol or growth promotion features in bioproducts not registered for their respective purposes, even if the strains used could express them. The unlikeliness of a regulatory framework would also hamper the development of products based on microbial consortia that exhibit complementary and synergistic effects, through re-assembling strains with differing modes of action into small communities, thereby providing more consistent protection or growth promotion than with the application of single strains, which are now starting to gain attention as a possible strategy to widen the application of bioproducts (Reddy and Saravanan, 2013; Vassilev et al., 2015; De Vrieze et al., 2018). At the same time, the potential use of bioproducts for alleviating other abiotic stresses (Hassen et al., 2016; Rajendra Prasad et al., 2016), particularly relevant in the world-wide experienced climate change conditions, would also face difficulties due to lack of clear rules for their registration and marketing. The current regulatory framework in EU as well as that of other countries where bioproducts are highly promoted (see several articles in Singh et al., 2016) could be perceived as frustrating the researchers efforts in finding the best solutions to exploit microbial inocula, considering that plants (and animals) are no longer viewed as autonomous entities, but rather as “holobionts” (Bordenstein and Theis, 2015). Nevertheless, we believe that the research activity that is currently endeavored to better understand the biochemical and molecular mechanisms involved in plant–microbe–soil interactions, paralleled with their impact on the plant metabolomics and the interactions with endophytes, should also support the progress in manufacturing and the regulatory development, leading to the design and use of safe bioproducts with greater efficacy in enhancing the productivity of sustainable crops. To this aim, exploitation of endophytes (Fadiji and Babalola, 2020), or of pre-, pro-, and post-biotic approaches (Vassileva et al., 2020) as well as of the plants’ capacity to “Cry for Help,” i.e., recruit and subsequent assembly of protective specific microbiota (Bakker et al., 2018; Rodriguez and Durán, 2020), could represent possible research avenues to be explored.
Author Contributions
JK, JT, and EM designed and drafted the work. KM and BT contributed to the revision of the manuscript. All authors contributed to the article and approved the submitted version.
Funding
Part of this work was supported by the project EXCALIBUR (to EM) funded from the European Union’s Horizon 2020 Research and Innovation Program under grant agreement No. 817946.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Abriouel, H., Franz, C. M., Ben, Omar, N., and Galvez, A. (2011). Diversity and applications of Bacillus bacteriocins. FEMS Microbiol. Rev.. 35, 201–232.
Adesemoye, A. O., Torbert, H. A., and Kloepper, J. W. (2009). Plant growth-promoting rhizobacteria allow reduced application rates of chemical fertilizers. Microb. Ecol. 58, 921–929.
Adolfsson, L., Nziengui, H., Abreu, I. N., Šimura, J., Beebo, A., Herdean, A., et al. (2017). Enhanced secondary- and hormone metabolism in leaves of arbuscular mycorrhizal Medicago truncatula. Plant Physiol. 175, 392–411. doi: 10.1104/pp.16.01509
Alabouvette, C., Heilig, U., and Cordier, C. H. (2012). “Microbial control of plant diseases,” in Beneficial Microorganisms in Agriculture, Food and the Environment: Safety Assessment and Regulation, eds I. Sundh, A. Wilcks, M. Stanislaw Goettel, et al. (Wallingford: CAB International), 96–111. doi: 10.1079/9781845938109.0000
Altomare, C., Norvell, W. A., Bjorkman, T., and Harman, G. E. (1999). Solubilization of phosphates and micronutrients by the plant growth promoting and biocontrol fungus Trichoderma harzianum rifai 1295-22. Appl. Environ. Microbiol. 65, 2926–2933.
Anonymous (2019a). Biofertilizers Market by Product, Microorganism Type and Application, Crop Type: Global Opportunity Analysis and Industry Forecast, 2019-2026. Fortune Business Insights.
Anonymous (2019b). Biopesticides Market Size, Share and Industry Analysis by Product Type, Source, Mode of Application, Crops and Regional Forecast 2018 – 2025. Fortune Business Insights, 145.
Arkhipova, T. N., Veselov, S. U., Melentiev, A. I., Martynenko, E. V., and Kudoyarova, G. R. (2005). Ability of bacterium Bacillus subtilis to produce cytokinins and to influence the growth and endogenous hormone content of lettuce plants. Plant Soil 272, 201–209.
Bakker, M. G., Manter, D. K., Sheflin, A. M., Weir, T. L., and Vivanco, J. M. (2012). Harnessing the rhizosphere microbiome through plant breeding and agricultural management. Plant Soil 360, 1–13. doi: 10.1007/s11104-012-1361-x
Bakker, P. A. H. M., Pieterse, C. M. J., de Jonge, R., and Berendsen, R. L. (2018). The soil-borne legacy. Cell 172, 1178–1180. doi: 10.1016/j.cell.2018.02.024
Bardi, L., and Malusá, E. (2012). “Drought and nutritional stresses in plant: alleviating role of rhizospheric microorganisms,” in Abiotic Stress: New Research, eds N. Haryana and S. Punj (Hauppauge, NY: Nova Science Publishers, Inc.), 1–57.
Bashan, Y., de-Bashan, L. E., Prabhu, S. R., and Hernandez, J.-P. (2014). Advances in plant growth-promoting bacterial inoculant technology: formulations and practical perspectives (1998-2013). (A Marschner Review). Plant Soil 378, 1–33.
Behie, S. W., Zelisko, P. M., and Bidochka, M. J. (2012). Endophytic insect-parasitic fungi translocate nitrogen directly from insects to plants. Science 336, 1576–1577.
Bennekou, S. H. (2019). Moving towards a holistic approach for human health risk assessment – Is the current approach fit for purpose? EFSA J. 17:e170711. doi: 10.2903/j.efsa.2019.e170711
Berg, G., Köberl, M., Rybakova, D., Müller, H., Grosch, R., and Smalla, K. (2017). Plant microbial diversity is suggested as the key to future biocontrol and health trends. FEMS Microbiol. Ecol. 93:fix050. doi: 10.1093/femsec/fix050
Boller, T., and Felix, G. (2009). A renaissance of elicitors: perception of microbe-associated molecular patterns and danger signals by pattern-recognition receptors. Annu. Rev. Plant Biol. 60, 379–406. doi: 10.1146/annurev.arplant.57.032905.105346
Bordenstein, S. R., and Theis, K. R. (2015). Host biology in light of the microbiome: ten principles of holobionts and hologenomes. PLoS Biol. 13:e1002226. doi: 10.1371/journal.pbio.1002226
Borriss, R. (2020). “Phytostimulation and biocontrol by the plant-associated Bacillus amyloliquefaciens FZB42: An Update,” in Phyto-Microbiome in Stress Regulation. Environmental and Microbial Biotechnology, eds M. Kumar, V. Kumar, and R. Prasad (Singapore: Springer), doi: 10.1007/978-981-15-2576-6_1
Brencic, A., and Winans, S. C. (2005). Detection of and response to signals involved in host-microbe interactions by plant-associated bacteria. Microbiol. Mol. Biol. Rev. 69, 155–194. doi: 10.1128/MMBR.69.1.155-194.2005
Cameron, D. D., Neal, A. L., van Wees, S. C., and Ton, J. (2013). Mycorrhiza induced resistance: more than the sum of its parts? Trends Plant Sci. 18, 539–545. doi: 10.1016/j.tplants.2013.06.004
Chandler, D., Bailey, A. S., Tatchell, G. M., Davidson, G., Greaves, J., and Grant, W. P. (2011). The development, regulation and use of biopesticides for integrated pest management. Philos. Trans. R. Soc. Lond. Ser. B 366, 1987–1998. doi: 10.1098/rstb.2010.0390
Chojnacka, K., Moustakas, K., and Witek-Krowiak, A. (2020). Bio-based fertilizers: a practical approach towards circular economy. Bioresour. Technol. 295:122223. doi: 10.1016/j.biortech.2019.122223
Choudhary, D., and Johri, B. N. (2009). Interactions of Bacillus spp. and plants—with special reference to induced systemic resistance (ISR). Microbiol. Res. 164, 493–513.
Chowdhury, S. P., Hartmann, A., Gao, X., and Borriss, R. (2015). Biocontrol mechanism by root-associated Bacillus amyloliquefaciens FZB42 – a review. Front. Microbiol. 6:780. doi: 10.3389/fmicb.2015.00780
Copping, L. G., and Menn, J. J. (2000). Biopesticides: a review of their action, applications and efficacy. Pest Manage. Sci. 56, 651–676.
Council of the European Union (2019). Progress Report on the Implementation Plan to Increase the Availability of Low-Risk Plant Protection Products and Accelerate Implementation of Integrated Pest Management in Member States. Available online at: https://data.consilium.europa.eu/doc/document/ST-10238-2019-INIT/en/pdf.
Dar, G. H., Zargar, M. Y., and Beigh, G. M. (1997). Biocontrol of fusarium root rot in the common bean (Phaseolus vulgaris L.) by using symbiotic Glomus mosseae and Rhizobium leguminosarum. Microb. Ecol. 34, 74–80.
de Almeida Melo, A. M., Soccol, V. T., and Soccol, C. R. (2016). Bacillus thuringiensis: mechanism of action, resistance, and new applications: a review. Crit. Rev. Biotechnol. 36, 317–326. doi: 10.3109/07388551.2014.960793
de Faria, M. R., and Wraight, S. P. (2007). Mycoinsecticides and mycoacaricides: a comprehensive list with worldwide coverage and international classification of formulation types. Biol. Control. 43, 237–256.
De Vrieze, M., Germanier, F., Vuille, N., and Weisskopf, L. (2018). Combining different potato-associated Pseudomonas strains for improved biocontrol of Phytophthora infestans. Front. Microbiol. 9:2573. doi: 10.3389/fmicb.2018.02573
Fadiji, A. E., and Babalola, O. O. (2020). Elucidating mechanisms of endophytes used in plant protection and other bioactivities with multifunctional prospects. Front. Bioeng. Biotechnol. 8:467. doi: 10.3389/fbioe.2020.00467
Fernández, I., Merlos, M., López-Ráez, J. A., Martínez-Medina, A., Ferrol, N., Azcón, C., et al. (2014). Defense related phytohormones regulation in arbuscular mycorrhizal symbioses depends on the partner genotypes. J. Chem. Ecol. 40, 791–803.
Fester, T., Fetzer, I., Buchert, S., Lucas, R., Rillig, M. C., and Hartig, C. (2011). Towards a systemic metabolic signature of the arbuscular mycorrhizal interaction. Oecologia 167, 913–924.
Fickers, P. (2012). Antibiotic compounds from Bacillus: why are they so amazing? Am. J. Biochem. Biotechnol. 8, 38–43.
Fiorentino, N., Ventorino, V., Woo, S. L., Pepe, O., De Rosa, A., Gioia, L., et al. (2018). Trichoderma-based biostimulants modulate rhizosphere microbial populations and improve N uptake efficiency, yield, and nutritional quality of leafy vegetables. Front. Plant Sci. 9:743. doi: 10.3389/fpls.2018.00743
French, K. E. (2017). Engineering mycorrhizal symbioses to alter plant metabolism and improve crop health. Front. Microbiol. 8:1403. doi: 10.3389/fmicb.2017.01403
Friesen, M. L., Porter, S. S., Stark, S. C., von Wettberg, E. J., Sachs, J. L., and Martinez-Romero, E. (2011). Microbially mediated plant functional traits. Annu. Rev. Ecol. Evol. Systemat. 42, 23–46.
Fritz, M., Jakobsen, I., Lyngkjær, M. F., Thordal-Christensen, H., and Pons-Kühnemann, J. (2006). Arbuscular mycorrhiza reduce ssusceptibility of tomatoto Alternaria solani. Mycorrhiza 16, 413–419.
García, J. E., Posadas, J. B., Perticari, A., and Lecuona, R. E. (2011). Metarhizium anisopliae (Metschnikoff) Sorokin promotes growth and has endophytic activity in tomato plants. Adv. Biol. Res. 5, 22–27.
Gianinazzi, S., Gollotte, A., Gollotte, A., Binet, M. N., Van Tuinen, D., Redecker, D., et al. (2010). Agroecology: the key role of arbuscular mycorrhizas in ecosystem services. Mycorrhiza 20, 519–530.
Glick, B. R., Todorovic, B., Czarny, J., Cheng, Z., Duan, J., and McConkey, B. (2007). Promotion of plant growth by bacterial ACC deaminase. Crit. Rev. Plant Sci. 26, 227–242.
Gómez-Vidal, S., Salinas, J., Tena, M., and Lopez-Llorca, L. V. (2009). Proteomic analysis of date palm (Phoenix dactylifera L.) responses to endophytic colonization by entomopathogenic fungi. Electrophoresis 30, 2996–3005.
Harman, G. E. (2011). Multifunctional fungal plant symbionts: new tools to enhance plant growth and productivity. New Phytol. 189, 647–649.
Hassen, A. I., Bopape, F. L., and Sanger, L. K. (2016). “Microbial inoculants as agents of growth promotion and abiotic stress tolerance in plants,” in Microbial Inoculants in Sustainable Agricultural Productivity, eds D. Singh, H. Singh, and R. Prabha (New Delhi: Springer), 23–36. doi: 10.1007/978-81-322-2647-5_2
Huang, J., Xu, C.-C., Ridoutt, B. G., Wang, X.-C., and Ren, P.-A. (2017). Nitrogen and phosphorus losses and eutrophication potential associated with fertilizer application to cropland in China. J. Clean. Product. 159, 171–179. doi: 10.1016/j.jclepro.2017.05.008
Iavicoli, A., Boutet, E., Buchala, A., and Métraux, J. P. (2003). Induced systemic resistance in Arabidopsis thaliana in response to root inoculation with Pseudomonas fluorescens CHA0. Mol. Plant-Microbe Interact. 16, 851–858.
Islam, M. d. T, Rahman, M. M., Pandey, P., Boehme, M. H., and Haesaert, G. (2019). Bacilli and Agrobiotechnology: Phytostimulation and Biocontrol, Vol. 2. Berlin: Springer, doi: 10.1007/978-3-030-15175-1
Jaber, L. R. (2015). Grapevine leaf tissue colonization by the fungal entomopathogen Beauveria bassiana s.l. and its effect against downy mildew. BioControl. 60, 103–112.
Jaber, L. R., and Enkerli, J. (2016). Effect of seed treatment duration on growth and colonization of Vicia faba by endophytic Beauveria bassiana and Metarhizium brunneum. Biol. Control 103, 187–195.
Jaber, L. R., and Enkerli, J. (2017). Fungal entomopathogens as endophytes: can they promote plant growth? Biocontrol Sci. Technol. 27, 28–41.
Jaber, L. R., and Salem, N. M. (2014). Endophytic colonisation of squash by the fungal entomopathogen Beauveria bassiana (Ascomycota: Hypocreales) for managing Zucchini yellow mosaic virus in cucurbits. Biocontrol Sci. Technol. 24, 1096–1109.
Jain, A., Singh, S., Sarma, B. K., and Singh, H. B. (2011). Microbial consortium mediated reprogramming of defense network in pea to enhance tolerance against Sclerotinia sclerotiorum. J. Appl. Microbiol. 12, 537–550.
Jasim, B., Sreelakshmi, K. S., Mathew, J., and Radhakrishnan, E. K. (2016). Surfactin, iturin, and fengycin biosynthesis by endophytic Bacillus sp. from Bacopa monnieri. Microb. Ecol. 72, 106–119.
Jirakkakul, J., Cheevadhanarak, S., Punya, J., Chutrakul, C., Senachak, J., Buajarern, T., et al. (2015). Tenellin acts as an iron chelator to prevent iron-generated reactive oxygen species toxicity in the entomopathogenic fungus Beauveria bassiana. FEMS Microbiol. Lett. 362, 1–8.
Johansson, J., Paul, L. R., and Finlay, R. D. (2004). Microbial interactions in the mycorrhizosphere and their significance for sustainable agriculture. FEMS Microbiol. Ecol. 1, 1–13.
Jung, S. C., Martínez-Medina, A., López-Ráez, J. A., and Pozo, M. J. (2012). Mycorrhiza-induced resistance and priming of plant defenses. J. Chem. Ecol. 38, 651–664.
Keswani, C., Dilnashin, H., Birla, H., and Singh, S. P. (2019a). Re-addressing the commercialization and regulatory hurdles for biopesticides in India. Rhizosphere 11:100155. doi: 10.1016/j.rhisph.2019.100155
Keswani, C., Prakash, O., Bharti, N., Vílchez, J. I., Sansinenea, E., and Lally, et al. (2019b). Re-addressing the biosafety issues of plant growth promoting rhizobacteria. Sci. Total Environ. 690, 841–852. doi: 10.1016/j.scitotenv.2019.07.046
Khan, A. L., Hamayun, M., Khan, S. A., Kang, S.-M., Shinwari, Z. K., Kamran, M., et al. (2012). Pure culture of Metarhizium anisopliae LHL07 reprograms soybean to higher growth and mitigates salt stress. World J. Microbiol. Biotechnol. 28, 1483–1494.
Khan, M. R., and Khan, S. M. (2001). Biomanagement of Fusarium wilt of tomato by the soil application of certain phosphate-solubilizing microorganisms. Int. J. Pest Manage. 47, 227–231.
Kowalska, J. (2014). Organically grown Brassica napus – use of border strips and Trichoderma. Acta Agric. Scand. Sect. B – Soil Plant Sci. 64, 529–536. doi: 10.1080/09064710.2014.929730
Kowalska, J., Remlein-Starosta, D., and Drożdżyński, D. (2012). Using of Trichoderma asperellum in organic strawberry production. Progr. Plant Protect. 52, 351–353.
Kuldau, G., and Bacon, C. (2008). Clavicipitaceous endophytes: their ability to enhance resistance of grasses to multiple stresses. Biological Control. 46, 57–71.
Kumar, A., Prakash, A., and Johri, B. N. (2011). “Bacillus as PGPR in crop ecosystem,” in Bacteria in Agrobiology: Crop Ecosystems, ed. D. Maheshwari (Berlin: Springer), doi: 10.1007/978-3-642-18357-7_2
Kurek, E., and Ozimek, E. (2013). “The use of microbiological soil activity to increase availability of phosphorous and potassium for plants,” in Soil Microbial Activity and Its Fertility, eds J. Tyburski, B. Godden, and M. J. Pennickx (Olsztyn: University of Warmia and Mazury in Olsztyn), 119–136.
Labuschagne, N., Pretorius, T., and Idris, A. H. (2010). “Plant growth promoting rhizobacteria as biocontrol agents against soil-borne plant diseases,” in Plant Growth and Health Promoting Bacteria. Microbiology Monographs, Vol. 18, ed. D. Maheshwari (Berlin: Springer), doi: 10.1007/978-3-642-13612-2_9
Lacey, L. A., Grzywacz, D., Shapiro-Ilan, D., Frutos, R., Brownbridge, M., and Goettel, M. S. (2015). Insect pathogens as biological control agents: back to the future. J. Inverteb. Pathol. 132, 1–41. doi: 10.1016/j.jip.2015.07.009
Liao, X., Lovett, B., Fang, W., and St. Leger, R. J. (2017). Metarhizium robertsii produces indole-3-acetic acid, which promotes root growth in Arabidopsis and enhances virulence to insects. Microbiology 163, 980–991.
Liao, X., O’Brien, T. R., Fang, W., and St. Leger, R. J. (2014). The plant beneficial effects of Metarhizium species correlate with their association with roots. Appl. Microbiol. Biotechnol. 98, 7089–7096.
Lombardi, N., Caira, S., Troise, A. D., Scaloni, A., Vitaglione, P., Vinale, F., et al. (2020). Trichoderma applications on strawberry plants modulate the physiological processes positively affecting fruit production and quality. Front. Microbioogyl. 11:1364. doi: 10.3389/fmicb.2020.01364
Lopes, R., Tsui, S., Gonçalves, P., and de Queiroz, M. V. (2018). A look into a multifunctional toolbox: endophytic Bacillus species provide broad and underexploited benefits for plants. World J. Microbiol. Biotechnol. 34:94.
Lopes, R. B. M., Costa, L. E., Vanetti, M. C., de Araujo, E. F., and de Queiroz, M. V. (2015). Endophytic bacteria isolated from common bean (Phaseolus vulgaris) exhibiting high variability showed antimicrobial activity and quorum sensing inhibition. Curr. Microbiol. 71, 509–516.
Lopez, D. C., and Sword, G. A. (2015). The endophytic fungal entomopathogens Beauveria bassiana and Purpureocillium lilacinum enhance the growth of cultivated cotton (Gossypium hirsutum) and negatively affect survival of the cotton bollworm (Helicoverpa zea). Biol. Control. 89, 53–60.
Lorito, M., Woo, S. L., Harman, G. E., and Monte, E. (2010). Translational research on Trichoderma: from ‘omics to the field. Annu. Rev. Phytopathol. 48, 395–417.
Ma, W., Mao, Q., Xia, W., Dong, G., Yu, C., and Jiang, F. (2019). Gut microbiota shapes the efficiency of cancer therapy. Front. Microbiol. 10:1050. doi: 10.3389/fmicb.2019.01050
Malusá, E., Pinzari, F., and Canfora, L. (2016). “Efficacy of biofertilizers: challenges to improve crop production,” in Microbial Inoculants in Sustainable Agricultural Productivity: Functional Applications, Vol. 2, eds D. Singh, P. Singh, H. Bahadur, and R. Ratna (Berlin: Springer), 17–40. doi: 10.1007/978-81-322-2644-4_2
Malusá, E., Sas-Paszt, L., and Ciesielska, J. (2012). Technologies for beneficial microorganisms inocula used as biofertilizers. Sci. World J. 2012:491206. doi: 10.1100/2012/491206
Malusá, E., and Vassilev, N. (2014). A contribution to set a legal framework for biofertilizers. Appl. Microbiol. Biotechnol. 98, 6599–6607. doi: 10.1007/s00253-014-5828-y
Maniania, N. K., Sithanantham, S., Ekesi, S., Ampong-Nyarko, K., Baumgärtner, J., Löhr, B., et al. (2003). A field trial of the entomogenous fungus Metarhizium anisopliae for control of onion thrips, Thrips tabaci. Crop Protect. 22, 553–559.
Marra, R., Ambrosino, P., Carbone, V., Vinale, F., Woo, S. L., Ruocco, M., et al. (2006). Study of the three-way interaction between Trichoderma atroviride, plant and fungal pathogens using a proteome approach. Curr. Genet. 50, 307–321.
Marrone, P. G. (2007). Barriers to adoption of biological control agents and biological pesticides. CAB Reviews: Perspectives in Agriculture, Veterinary Science, Nutrition and Natural Resources 2. doi: 10.1079/PAVSNNR20072051
Marrone, P. G. (2019). Pesticidal natural products – status and future potential. Pest Manage. Sci. 75, 2325–2340.
Martínez-Medina, A., Pascual, J. A., Lloret, E., and Roldán, A. (2009). Interactions between arbuscular mycorrhizal fungi and Trichoderma harzianum and their effects on Fusarium wilt in melon plants grown in seedling nurseries. J. Sci. Food Agric. 89, 1843–1850. doi: 10.1002/jsfa.3660
Meena, V. S., Maurya, B. R., and Verma, J. P. (2014). Does a rhizospheric microorganism enhance K+availability inagricultural soils? Microbiol. Res. 169, 337–347. doi: 10.1016/j.micres.2013.09.003
Morath, S. U., Hung, R., and Bennett, J. W. (2012). Fungal volatile organic compounds: a review with emphasis on their biotechnological potential. Fungal Biol. Rev. 26, 73–83.
Mozumder, P., and Berrens, R. P. (2007). Inorganic fertilizer use and biodiversity risk: an empirical investigation. Ecol. Econ. 62, 538–543. doi: 10.1016/j.ecolecon.2006.07.016
Norse, D. (2005). Non-point pollution from crop production: global, regional and national issues. Pedosphere 15, 1–10.
Parliament, E. (2009). Directive 2009/128/EC of the european parliament and of the council of 21 October 2009 establishing a framework for community action to achieve the sustainable use of pesticides. Offi. J. Eur. Union L 39, 71–86.
Radhakrishnan, R., and Lee, I. J. (2016). Gibberellins producing Bacillus methylotrophicus KE2 supports plant growth and enhances nutritional metabolites and food values of lettuce. Plant Physiol. Biochem. 109, 181–189.
Rajendra Prasad, S., Kamble, U. R., Sripathy, K. V., Udaya Bhaskar, K., and Singh, D. P. (2016). “Seed bio-priming for biotic and abiotic stress management,” in Microbial Inoculants in Sustainable Agricultural Productivity, eds D. Singh, H. Singh, and R. Prabha (New Delhi: Springer), 211–228. doi: 10.1007/978-81-322-2647-5_12
Rath, M., Mitchell, T. R., and Gold, S. E. (2018). Volatiles produced by Bacillus mojavensis RRC101 act as plant growth modulators and are strongly culture-dependent. Microbiol. Res. 208, 76–84.
Reddy, C. H. A., and Saravanan, R. S. (2013). Polymicrobial multi-functional approach for enhancement of crop productivity. Adv. Appl. Microbiol. 82, 53–113.
Regulation Eu 2019/1009. (2019). Regulation (EU) 2019/1009 of the European Parliament and of the Council of 5 June 2019 laying down rules on the making available on the market of EU fertilising products and amending Regulations (EC) No 1069/2009 and (EC) No 1107/2009 and repealing Regulation (EC) No 2003/2003. Offi. J. Eur. Union 62, 1–114.
Regulation, E. U. (1107/2009). Regulation (EC) No 1107/2009 of the European Parliament and of the Council of 21 October 2009 concerning the placing of plant protection products on the market. OJL 309, 1–50.
Rivero, J., Gamir, J., Aroca, R., Pozo, M. J., and Flors, V. (2015). Metabolic transition in mycorrhizal tomato roots. Front. Microbiol. 6:598. doi: 10.3389/fmicb.2015.00598
Rodriguez, R., and Durán, P. (2020). Natural holobiome engineering by using native extreme microbiome to counteract the climate change effects. Front. Bioeng. Biotechnol. 8:568. doi: 10.3389/fbioe.2020.00568
Saeid, A., and Chojnacka, K. (2019). “Fertlizers: need for: organic farming new strategies,” in Global Perspectives and Methods, eds S. Chandran and U. M. R. Sabu Thomas (Sawston: Woodhead Publishing), 91–116.
Sánchez-Bayo, F., and Wyckhuys, K. A. G. (2019). Worldwide decline of the entomofauna: a review of its drivers. Biol. Conserv. 232, 8–27. doi: 10.1016/j.biocon.2019.01.020
Sánchez-Rodríguez, A. R., Del Campillo, M. C., and Quesada-Moraga, E. (2015). Beauveria bassiana: an entomopathogenic fungus alleviates Fe chlorosis symptoms in plants grown on calcareous substrates. Sci. Horticult. 197, 193–202.
Santoyo, G., del Carmen, Orozco-Mosqueda, M., and Govindappa, M. (2012). Mechanisms of biocontrol and plant growth-promoting activity in soil bacterial species of Bacillus and Pseudomonas: a review. Biocontrol Sci. Technol. 22, 855–872.
Sasan, R. K., and Bidochka, M. J. (2013). Antagonism of the endophytic insect pathogenic fungus Metarhizium robertsii against the bean plant pathogen Fusarium solani f.sp. phaseoli. Can. J. Plant Pathol. 35, 288–293.
Sayyed, R. Z., Chincholkar, S. B., Reddy, M. S., Gangurde, N. S., and Patel, P. R. (2013). “Siderophore producing PGPR for crop nutrition and phytopathogen suppression,” in Bacteria in Agrobiology: Disease Management, ed. D. Maheshwari (Berlin: Springer), doi: 10.1007/978-3-642-33639-3_17
Sheng, X. F., and He, L. Y. (2006). Solubilization of potassium-bearing minerals by a wild type strain of Bacillus edaphicus and its mutants and increased potassium uptake by wheat. Can. J. Microbiol. 52, 66–72.
Shoresh, M., and Harman, G. E. (2008). The molecular basis of maize responses to Trichoderma harzianum T22 inoculation: a proteomic approach. Plant Physiol. 147, 2147–2163.
Shoresh, M., Mastouri, F., and Harman, G. E. (2010). Induced systemic resistance and plant responses to fungal biocontrol agents. Annu. Rev. Phytopathol. 48, 21–43.
Singh, H. B, Sarma, B. K., and Keswani, C. (2016). Agriculturally Important Microorganisms: Commercialization and Regulatory Requirements in Asia. Singapore:pringer, 336
Smith, S. E., and Smith, F. A. (2011). Roles of arbuscular mycorrhizas in plant nutrition and growth: new paradigms from cellular to ecosystem scales. Annu. Rev. Plant Biol. 62, 227–250.
Sobolewski, J., Gidelska, A., Szczech, M., and Robak, J. (2013). Trichoderma spp. jako zaprawa nasienna przeciwko zgorzelom siewek roślin warzywnych. Progr. Plant Protect. 53, 340–344.
Song, Y. Y., Ye, M., Li, C. Y., Wang, R. L., Wei, X. C., Luo, S. M., et al. (2013). Priming of anti-herbivore defense in tomato by arbuscular mycorrhizal fungus and involvement of the jasmonate pathway. J. Chem. Ecol. 39, 1036–1044.
Steinkellner, S., Hage-Ahmed, K., Garcia-Garrido, J. M., Illana, A., Ocampo, J. A., and Vierheilig, H. (2012). A comparison of wild-type, old and modern tomato cultivars in the interaction with the arbuscular mycorrhizal fungus Glomusmosseae and the tomato pathogen Fusarium oxysporum f. sp. lycopersici. Mycorrhiza 22, 189–194. doi: 10.1007/s00572-011-0393-z
Tahir, H. A., Gu, Q., Wu, H., Raza, W., Hanif, A., Wu, L., et al. (2017). Plant growth promotion by volatile organic compounds produced by Bacillus subtilis SYST2. Front. Microbiol. 8:171. doi: 10.3389/fmicb.2017.00171
Trdá, L., Boutrot, F., Claverie, J., Brulé, D., Dorey, S., and Poinssot, B. (2015). Perception of pathogenic or beneficial bacteria and their evasion of host immunity: pattern recognition receptors in the frontline. Front. Plant Sci. 6:219. doi: 10.3389/fpls.2015.00219
Umesha, S., Singh, P. K., and Singh, R. P. (2018). “Microbial biotechnology and sustainable agriculture,” in Biotechnology for Sustainable Agriculture, Emerging Approaches and Strategies, eds R. L. Singh and S. Monda (Chichester: Woodhead Publishing), 185–205.
United Nations (2015). Transforming Our World: The 2030 Agenda for Sustainable Development. A/RES/70/1 21/10/2015. New York, NY: United Nations.
van Wees, S. C. M., van der Ent, S., and Pieterse, C. M. J. (2008). Plant immune responses triggered by beneficial microbes. Curr. Opin. Plant Biol. 11, 443–448.
Vassilev, N., Nikolaeva, I., Jurado, E., Reyes, A., Fenice, M., and Vassileva, M. (2008). “Antagonistic effect of microbially-treated mixture of agro-industrial wastes and inorganic insoluble phosphate to Fusarium wilt disease,” in Progress in Environmental Microbiology, ed. M. B. Kim (Hauppauge: Nova Publishers), 223–234.
Vassilev, N., Requena, A., Nieto, L., Nikolaeva, I., and Vassileva, M. (2009a). Production of manganese peroxidase by Phanerochaete chrysosporium grown on medium containing agro-wastes/rock phosphate and biocontrol properties of the final product. Ind. Crops Products 30, 28–32.
Vassilev, N., Someus, E., Serrano, M., Bravo, V., Garcia, R. M., Reyes, A., et al. (2009b). “Novel approaches in phosphate-fertilizer production based on wastes derived from rock phosphate mining and food processing industry,” in Industrial Waste: Environmental Impact, Disposal and Treatment, ed. J. P. Samuelson (New York, NY: Nova SciPubl), 387–391.
Vassilev, N., and Vassileva, M. (2004). “Multifunctional properties of a plant growth promoting bacterium entrapped in k-carrageenan,” in XII International Workshop on Bioencapsulation, eds J. L. Pedraz, G. Orive, and D. Poncelet (Vitoria: Univ Pais Vasco), 162–166.
Vassilev, N., Vassileva, M., Lopez, A., Martos, V., Reyes, A., Maksimovic, I., et al. (2015). Unexploited potential of some biotechnological techniques for biofertilizer production and formulation. Appl. Microbiol. Biotechnol. 99, 4983–4996.
Vassilev, N., Vassileva, M., Martos, V., Garcia, Del Moral, L. F., Kowalska, J., et al. (2020). Formulation of microbial inoculants by encapsulation in natural polysaccharides: focus on beneficial properties of carrier additives and derivates. Front. Plant Sci. 11:270. doi: 10.3389/fpls.2020.00270
Vassilev, N., Vassileva, M., and Nikolaeva, I. (2006). Simultaneous P-solubilizing and biocontrol activity of microorganisms: potentials and future trends. Appl. Microbiol. Biotechnol. 71, 137–144. doi: 10.1007/s00253-006-0380-z
Vassileva, M., Flor-Peregrin, E., Malusá, E., and Vassilev, N. (2020). Towards better understanding of the interactions and efficient application of plant beneficial prebiotics, probiotics, postbiotics and synbiotics. Front. Plant Sci. 11:553774. doi: 10.3389/fpls.2020.01068
Vega, F. E., Goettel, M. S., Blackwell, M., Chandler, D., Jackson, M. A., Keller, S., et al. (2009). Fungal entomopathogens: new insights on their ecology. Fungal Ecol. 2, 149–159. doi: 10.1016/j.funeco.2009.05.001
Vimal, S. R., Singh, J. S., Arora, N. K., and Singh, S. (2017). Soil-Plant-Microbe interactions in stressed agriculture management: a review. Pedosphere 27, 177–192. doi: 10.1016/S1002-0160(17)60309-6
Vos, C. M., Tesfahun, A. N., Panis, B., De Waele, D., and Elsen, A. (2012). Arbuscular mycorrhizal fungi induce systemic resistance in tomato against the sedentary nematode Meloidogyne incognita and the migratory nematode Pratylenchus penetrans. Appl. Soil Ecol. 61, 1–6. doi: 10.1016/j.apsoil.2012.04.007
Weller, D. M. (2007). Pseudomonas biocontrol agents of soilborne pathogens: looking back over 30 years. Phytopathology 97, 250–256.
Xie, S. S., Wu, H. J., Zang, H. Y., Wu, L. M., Zhu, Q. Q., and Gao, X. W. (2014). Plant growth promotion by spermidine-producing Bacillus subtilis OKB105. Mol. Plant Microbe Interact. J. 27, 655–663.
Zhang, A., Zhao, G., Gao, T., Wang, W., Li, J., Zhang, S., et al. (2013). Solubilization of insoluble potassium and phosphate by Paenibacillus kribensis CX-7: a soil microorganism with biological control potential. Afr. J. Microbiol. Res. 7, 41–47. doi: 10.5897/AJMR12.1485
Ziedan, E. S., Elewa, I., Mostafa, M., and Sahab, A. (2011). Application of mycorrhizae for controlling root diseases of sesame. J. Plant Prot. Res. 51, 355–361.
Zimmermann, G. (2007). Review on safety of the entomopathogenic fungi Beauveria bassiana and Beauveria brongniartii. Biocontrol Sci. Technol. 17, 533–596.
Zipfel, C. (2014). Plant pattern-recognition receptors. Trends Immunol. 35, 345–351. doi: 10.1016/j.it.2014.05.004
Keywords: biofertilizers, biopesticides, microbial consortia, multifunctional bioproducts, plant growth-promoting microorganisms, endophytes
Citation: Kowalska J, Tyburski J, Matysiak K, Tylkowski B and Malusá E (2020) Field Exploitation of Multiple Functions of Beneficial Microorganisms for Plant Nutrition and Protection: Real Possibility or Just a Hope? Front. Microbiol. 11:1904. doi: 10.3389/fmicb.2020.01904
Received: 30 May 2020; Accepted: 20 July 2020;
Published: 05 August 2020.
Edited by:
Giorgio Gambino, Institute for Sustainable Plant Protection (CNR), ItalyReviewed by:
Chetan Keswani, Banaras Hindu University, IndiaShengjun Xu, Research Center for Eco-Environmental Sciences (CAS), China
Copyright © 2020 Kowalska, Tyburski, Matysiak, Tylkowski and Malusá. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jolanta Kowalska, ai5rb3dhbHNrYUBpb3JwaWIucG96bmFuLnBs