 Manoj Kaushal
Manoj Kaushal- Plant Production and Plant Health, International Institute of Tropical Agriculture (IITA), Dar es Salaam, Tanzania
Salt stress threatens the achievement of sustainable global food security goals by inducing secondary stresses, such as osmotic, ionic, and oxidative stress, that are detrimental to plant growth and productivity. Various studies have reported the beneficial roles of microbes in ameliorating salt stress in plants. This review emphasizes salt tolerance and endurance mechanisms (STEM) in microbially inoculated (MI) plants that ensure plant growth and survival. Well-established STEM have been documented in MI plants and include conglomeration of osmolytes, antioxidant barricading, recuperating nutritional status, and ionic homeostasis. This is achieved via involvement of P solubilization, siderophore production, nitrogen fixation, selective ion absorption, volatile organic compound production, exopolysaccharide production, modifications to plant physiological processes (photosynthesis, transpiration, and stomatal conductance), and molecular alterations to alter various biochemical and physiological processes. Salt tolerance and endurance mechanism in MI plants ensures plant growth by improving nutrient uptake and maintaining ionic homeostasis, promoting superior water use efficiency and osmoprotection, enhancing photosynthetic efficiency, preserving cell ultrastructure, and reinforcing antioxidant metabolism. Molecular research in MI plants under salt stress conditions has found variations in the expression profiles of genes such as HKT1, NHX, and SOS1 (ion transporters), PIPs and TIPs (aquaporins), RBCS, RBCL (RuBisCo subunits), Lipoxygenase2 [jasmonic acid (JA) signaling], ABA (abscisic acid)-responsive gene, and APX, CAT, and POD (involved in antioxidant defense). Proteomic analysis in arbuscular mycorrhizal fungi-inoculated plants revealed upregulated expression of signal transduction proteins, including Ca2+ transporter ATPase, calcium-dependent protein kinase, calmodulin, and energy-related proteins (NADH dehydrogenase, iron-sulfur protein NADH dehydrogenase, cytochrome C oxidase, and ATP synthase). Future research should focus on the role of stress hormones, such as JA, salicylic acid, and brassinosteroids, in salt-stressed MI plants and how MI affects the cell wall, secondary metabolism, and signal transduction in host plants.
Introduction
Salinity or salt stress is a major threat to agricultural productivity and global food security. It can affect plant growth and development and thus reduce the biomass productivity of plants in arid and semiarid regions. Salt stress is detrimental to plant growth because it induces osmotic and ionic stress in plants, leading to reduced water uptake, transpiration, photosynthesis, and disrupted ionic homeostasis. Moreover, increased levels of reactive oxygen species (ROS) cause oxidative stress, which damages DNA, proteins, and membranes (Liu et al., 2016). Recent studies have confirmed that microbes can induce salt tolerance and endurance mechanisms (STEM) in plants (Table 1) to enable growth and development under harsh stress conditions (Porcel et al., 2016; Barnawal et al., 2017; Chen et al., 2017; Sapre et al., 2018; Yasin et al., 2018; Jia et al., 2019). The various functions of STEM mediating this process can be summarized as follows: (i) conglomeration of osmolytes to abate osmotic stress (Hajiboland et al., 2010; Talaat and Shawky, 2011; Evelin and Kapoor, 2014; Elhindi et al., 2017; Wu et al., 2017; Garg and Bharti, 2018; Hashem et al., 2018); (ii) antioxidant barricading to block oxidative stress (Bharti et al., 2016; Qin et al., 2016; Chang et al., 2018; Chu et al., 2019; Ye et al., 2019); (iii) recuperating nutritional status and ionic homeostasis through P solubilization, siderophore production, nitrogen fixation, ion transporter activity, and exopolysaccharide (EPS) production (Porcel et al., 2016; Elhindi et al., 2017; Zhou et al., 2017; Chang et al., 2018); (iv) physiological modifications in the plant (Barnawal et al., 2017; Chen et al., 2017; Elhindi et al., 2017; Hashem et al., 2018; Ren et al., 2018); and (v) molecular modification of stress-responsive gene expression (Barnawal et al., 2017; Yasin et al., 2018; El-Esawi et al., 2019; Jia et al., 2019). Plant growth-promoting rhizobacteria (PGPR) have been reported to have mitigative effects on the growth of pepper (Yasin et al., 2018), wheat (Bharti et al., 2016; Barnawal et al., 2017), soybean (Khan et al., 2019), oat (Sapre et al., 2018), Panax (Sukweenadhi et al., 2018), and maize (Chen et al., 2016) under salt stress conditions. Similarly, colonization by arbuscular mycorrhizal fungi (AMF) also ameliorated the effects of salt stress in wheat (Fileccia et al., 2017), rice (Porcel et al., 2016), watermelon (Ye et al., 2019), and cucumber (Hashem et al., 2018). The STEM exhibited by PGPR and AMF are illustrated in Figure 1.
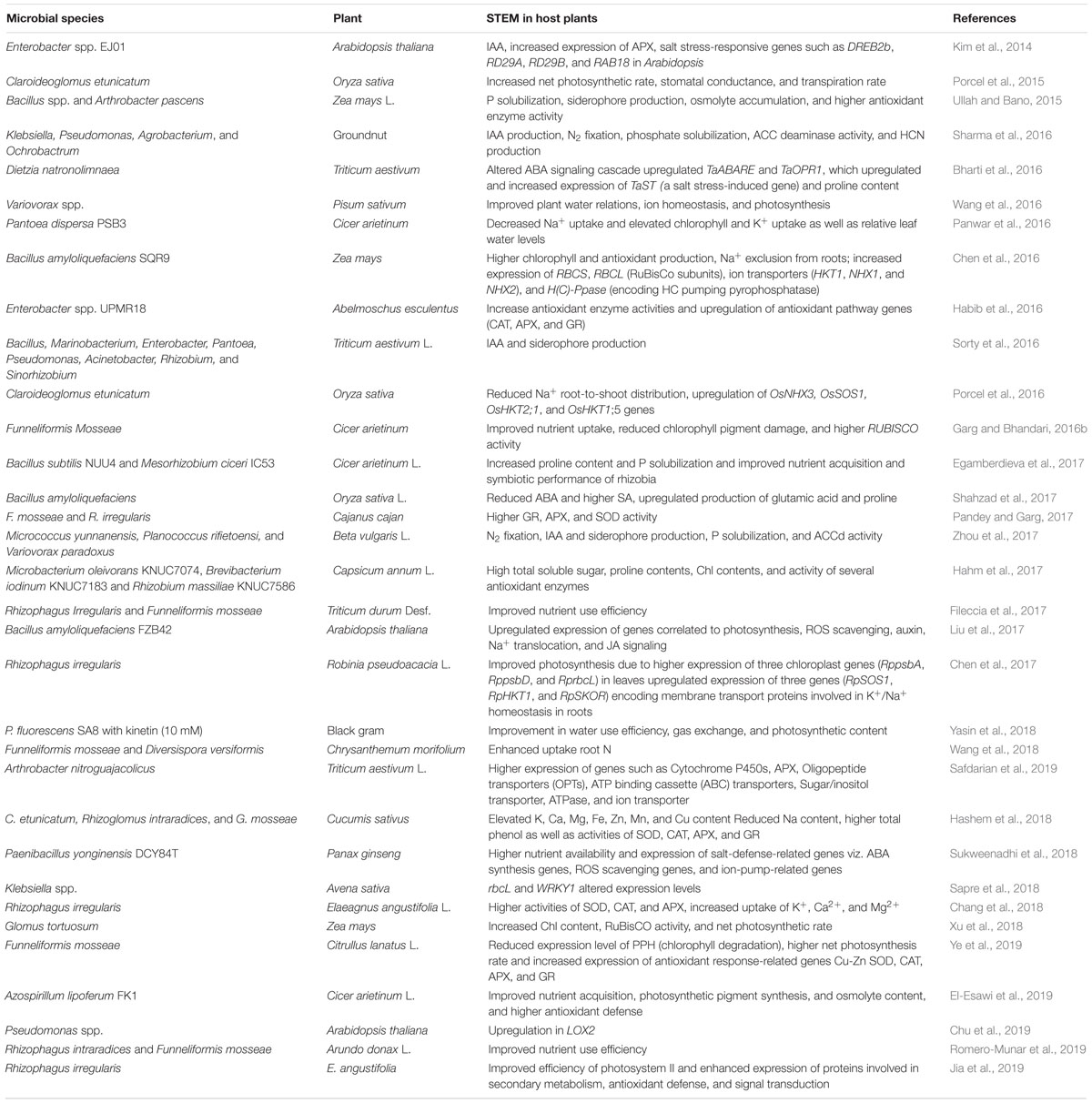
Table 1. STEM in various plant species under salt stress.
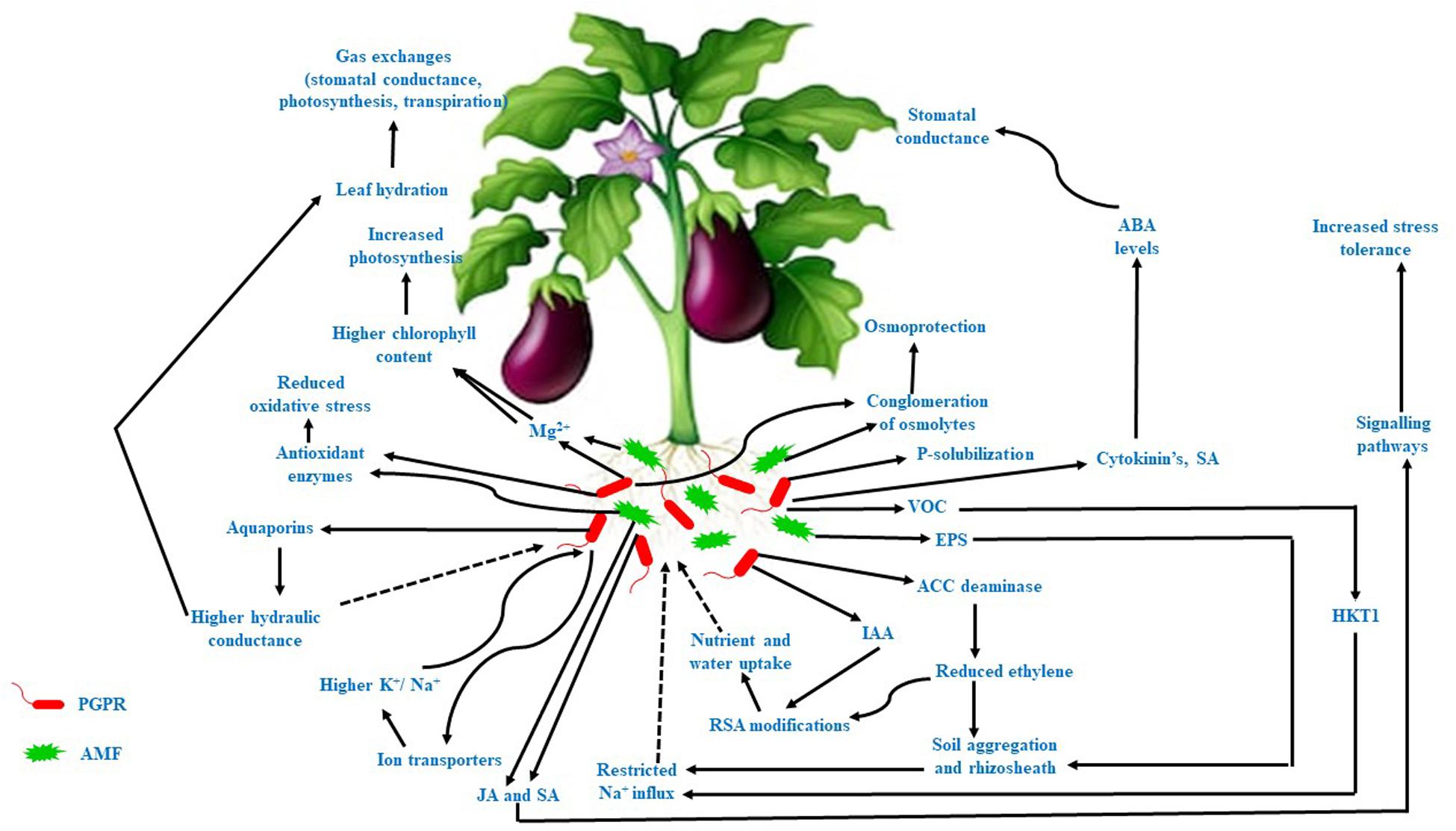
Figure 1. STEM exhibited by PGPR and AMF under salt stress.
Conglomeration of Osmolytes and Water Homeostasis to Abate Osmotic Stress
In its initial phase, salt stress can be referred to as physiological drought because elevated ion levels during salt stress change the soil texture to reduce soil porosity and decrease water uptake. Osmolyte conglomeration is a major STEM that improves water uptake in microbially inoculated (MI) plants. This reduces the water potential by accumulating osmolytes, such as amino acids (proline), amines (e.g., glycinebetaine, polyamines), sugars, and organic acids (e.g., oxalate, malate). In addition to osmotic adjustment, these osmolytes are responsible for conserving membrane integrity, protein stability, and ROS scavenging to ultimately promote their positive effects on plant physiological functions, such as growth, photosynthesis, and crop yield, during salt stress (Zou et al., 2013). Proline and glycinebetaine enhance protein and membrane stabilization to impart osmoprotection to salt-stressed plants. However, contrasting results regarding proline production have been reported in MI and non-inoculated (NI) plants. Increased proline content in NI plants compared to AMF-inoculated (AI) plants can indicate higher stress conditions. AI plants show decreased proline content because microbial colonization helps the plant mitigate the stress. Some studies have suggested that proline accumulation is a salinity stress indicator rather than a consequence of mycorrhizal colonization (Sheng et al., 2011; Echeverria et al., 2013; Evelin et al., 2013; Evelin and Kapoor, 2014); however, other studies have found higher proline accumulations caused by AM colonization (Hajiboland et al., 2010; Talaat and Shawky, 2011; Garg and Baher, 2013; Elhindi et al., 2017; Hashem et al., 2018). Higher proline levels have been observed in PGPR-inoculated maize (Ullah and Bano, 2015), Gladiolus (Damodaran et al., 2014), Mentha (Bharti et al., 2014), Chrysanthemum (Wang et al., 2018), and Panax (Sukweenadhi et al., 2018). Increased proline levels in MI plants can be due to (i) upregulated expression P5CS, a gene involved in proline synthesis; (ii) increased efficiency of the enzymes P5CS and glutamate dehydrogenase (involved in glutamate synthesis) given that proline is synthesized from glutamate; and (iii) arrest of proline dehydrogenase (responsible for proline degradation) (Abo-Doma et al., 2016). In the nodules of AI-inoculated pigeon pea plants, reduced activity of trehalase (trehalose degrading enzyme) and increased activity of trehalose-6-P synthase and trehalose-6-phosphatase (enzymes involved in the biosynthesis of trehalose) led to higher trehalose levels (Garg and Pandey, 2016). Higher concentrations of acetic, malic, citric, oxalic, and fumaric acids were observed in AI maize plants compared to NI plants and led to enhanced salinity tolerance (Sheng et al., 2011). Arbuscular mycorrhizal fungi treatment alters polyamine levels in plants to impart stress tolerance (Evelin et al., 2013; Talaat and Shawky, 2013). Evelin et al. (2013) observed increased spermidine and spermine (Spd + Spm)/putrescine (Put) ratios in AI fenugreek plants compared to NI plants. Salinity tolerance in MI plants is advantageous and can correlate with their ability to join DNA, proteins, and phospholipids. Polyamine levels were modulated in response to mycorrhizal colonization in two cultivars of wheat (Sids 1 and Giza 168) under saline conditions. AM colonization led to higher putrescine but lower spermidine and spermine levels in Giza 168; however, in Sid 1, a reduction in putrescine and an increase in spermidine and spermine levels were reported (Talaat and Shawky, 2013). The accumulation of total soluble sugars (TSS), such as glucose, sucrose, and maltose, during salt stress in MI plants is another mode of STEM via osmotic adjustment. Conversion of starch into dextrins and maltose is accompanied by a- and b-amylases, respectively. Researchers have confirmed that enhanced salt stress tolerance in MI plants is due to higher TSS accumulation (Talaat and Shawky, 2011; Liu et al., 2016; Garg and Bharti, 2018; Zhu et al., 2018). Arbuscular mycorrhizal fungi inoculation modifies leaf sucrose and proline metabolism by regulating the enzymatic activities responsible for sucrose and proline metabolism to enhance osmotic tolerance in the host plant (Wu et al., 2017). Elevated TSS content can be caused by increased photosynthesis, amylase activity, and increased organic acid levels (Yu et al., 2015; Garg and Bharti, 2018; Zhu et al., 2018). Studies in MI chickpea plants have demonstrated that increased salt tolerance can be achieved by the synthesis of proline, glycinebetaine, and increasing TSS (Qurashi and Sabri, 2012; Upadhyay and Singh, 2015). Elevated glycinebetaine levels enhance salinity tolerance in rhizobacterially primed rice plants (Jha et al., 2011), AI wheat (Talaat and Shawky, 2011), and maize (Sheng et al., 2011). According to Rangel (2011), bacteria grown under glucose concentration have low cAMP levels, but when grown under carbon starvation, bacteria show higher cAMP levels. However, the converse is true for eukaryotes. This aspect needs to be addressed in relation to plant microbial crosstalk under salt stress conditions. Future research should focus on unraveling the molecular mechanisms underlying the role of microbes in promoting osmotic adjustment during salt stress.
Antioxidant Barricading to Caulk the Oxidative Stress
The hyperosmotic and hyperionic conditions present during salt stress disrupt cellular redox homeostasis by disrupting the equilibrium between the generation and elimination of ROS, leading to oxidative stress as a secondary stress. Reactive oxygen species target various biomolecules, including nucleic acids, proteins and fatty acids, to alter cellular function, cause DNA damage, reduce membrane fluidity, cause lipid peroxidation, and affect enzymatic activity. It is evident that ROS create oxidative stress; however, they are also involved in ethylene accumulation, auxin biosynthesis, and many signaling events (Kaushal, 2019). Thus, it is essential that an equilibrium is maintained between ROS generation and ROS scavenging systems to balance oxidative damage while managing endogenous signaling events. Plants are equipped with a robust antioxidant system consisting of enzymatic [superoxide dismutase (SOD), catalase (CAT), ascorbate peroxidase (APX), monodehydroascorbate reductase (MDHAR), and glutathione reductase (GR)] and non-enzymatic (cysteine, carotenoids, glutathione, tocopherols, and ascorbate) constituents. The induction of the antioxidative defense system has been shown to be another STEM activated in MI plants to abate salt stress (Talaat and Shawky, 2011; Li et al., 2012; Bharti et al., 2016; Chang et al., 2018). Higher antioxidant activities have been observed in AI tomato (Hajiboland et al., 2010; Latef and Chaoxing, 2011), Sesbania (Abd Allah et al., 2015), pigeon pea (Pandey and Garg, 2017), and Cucumis (Hashem et al., 2018) plants during salt stress. Wu et al. (2016), while investigating the impact of AMF colonization and salt stress on male and female Populus cathayana seedlings, observed significant increases in the activities of SOD and CAT in the roots of AI-colonized plants compared to those of NI plants; however, CAT activity was similar in the leaves of AI and NI plants. Three-way ANOVA revealed that the activities of SOD, POD, and CAT in roots were influenced by AMF × salt × sex, salt × sex, and AMF × sex, AMF × sex, and AMF × sex × salt, respectively. This demonstrated that the activities of different antioxidant enzymes were variably affected by the interactions between salt stress, gender, and AMF. A significant increase in the expression levels of genes related to the antioxidant response, such as Cu-Zn SOD, CAT, APX, and GR, was reported during salinity alkalinity stress and was further enhanced by AMF inoculation, thereby enabling host watermelon plants to cope with the stress (Ye et al., 2019). Higher root, stem, and leaf biomass was observed in AI seedlings of Elaeagnus angustifolia L. during salt stress, which was attributed to the increased activities of SOD, CAT, and APX in the leaves relative to those of NI plants (Chang et al., 2018). In addition to increased activity and expression levels of CAT, GPOX, APX, SOD, MDHAR, DHAR, and GR, AI Cicer arietinum plants also demonstrated enhanced levels of GSG, GSSH, and total glutathione (Garg and Bhandari, 2016a). Similar observations of STEM involving higher antioxidant barricading have been reported in rhizobacterially inoculated (RI) plants (Gururani et al., 2013; Kim et al., 2014; Tewari and Arora, 2014; Ullah and Bano, 2015; Bharti et al., 2016; Kaushal and Wani, 2016a; Singh and Jha, 2017). A significant increase in the specific activities of APX (1.4 times), SOD (2.4 times), and CAT (1.8 times) was observed in RI Solanum plants under salt stress, and antioxidant enzyme activity was positively correlated with the mRNA expression levels of the corresponding genes encoding these enzymes (Gururani et al., 2013). In a similar study, higher activities of APX and CAT and upregulation of antioxidant pathway genes (CAT, APX, and GR) were observed in Enterobacter spp. UPMR18-colonized okra plants, improving the physiological performance and salt tolerance of the plants (Habib et al., 2016). In Arabidopsis thaliana roots colonized with Burkholderia phytofirmans PsJNA, genes involved in ROS quenching (APX2) were significantly more transcribed, helping the plant to abate oxidative stress (Pinedo et al., 2015). In PGPR Dietzia natronolimnaea STR1-colonized wheat plants, the gene expression of certain antioxidant enzymes (APX, MnSOD, CAT, POD, GPX, and GR) was enhanced to alleviate salt stress (Bharti et al., 2016). Higher activities of antioxidant enzymes in MI plants can correlate with improved nutritional status (Cu, Mn, and Fe) because these enzymes are in fact metalloenzymes, and their activities are therefore governed by the presence of these nutrients (Kohler et al., 2009). Moreover, the activity of these enzymes also depends on the plant species, microbes, and stress timing. AI plants were able to abate the effect of salt stress by enhancing the activity of antioxidant enzymes, including SOD, CAT, and APX, and increasing ascorbic acid levels, which correlates with lower lipid peroxidation and electrolyte leakage. Zn, Cu, Mn, and Fe serve as co-factors for SOD isozymes, and their increased uptake in AI plants was able to boost SOD activity (Hashem et al., 2018).
Salinity Tolerance Through Recuperation of Nutritional Status and Ionic Homeostasis
Salinity can lead to altered nutritional status and ionic homeostasis in plants, hampering the plant’s productivity. Microbial colonization during stress improves the physiological performance of plants with an additional STEM involving enhanced nutrient uptake and selective ion absorption and translocation (Evelin et al., 2012; Bharti et al., 2014; Kang et al., 2014b; Tewari and Arora, 2014; Porcel et al., 2016; Hashem et al., 2018). The availability of nutrients such as P, N, Mn, and Fe is restricted in saline soils. Microbial inoculation simplifies the process of acquiring these nutrients for plants under salt stress conditions to promote plant health and productivity. Plant growth-promoting rhizobacteria strains act as phosphate-solubilizing rhizobacteria to increase the uptake and availability of P to plants (Prasad et al., 2015; Etesami, 2018). Salt tolerance was enhanced in PGPR-colonized Mentha (Bharti et al., 2014), wheat (Upadhyay and Singh, 2015), Chrysanthemum (Zhou et al., 2017), and groundnut (Shukla et al., 2012) plants due to enhanced phosphate nutrition. Improved phosphorus absorption was found in AI plants under mycorrhizal inoculation, even under salt stress conditions (Sharifi et al., 2007; Al-Khaliel, 2010; Bowles et al., 2016). It is postulated that phosphate is absorbed and converted to polyphosphate by the extraradical mycelium. Recent studies have demonstrated the involvement of AM aquaporins in the translocation of polyphosphate via mycorrhizal hyphae. Thus, adequate P uptake in AI plants helps selective ion absorption, limits toxic ions in vacuoles, and preserves membrane integrity (Evelin et al., 2012) to reverse the effects of salt stress. However, it has been suggested that enhanced growth in MI plants was caused by improved photosynthesis and WUE (water use efficiency) rather than by the increased mineral uptake (Ruiz-Lozano et al., 1996; Garg and Bhandari, 2016b; Chen et al., 2017). Moreover, Feng et al. (2002) reported that salt tolerance in AMF plants was conferred by increased soluble sugar accumulation rather than P levels. Rhizobacterial strains often secrete siderophores to cope with iron deficiency in plants surrounded by saline soil. Siderophores are high affinity low molecular weight Fe (III) chelators that scavenge Fe3+ to form an iron–siderophore complex that can be readily solubilized to increase iron availability to plants. Recent studies have reported enhanced salinity tolerance in rhizobacterially colonized plants resulting from siderophore production (Shukla et al., 2012; Sorty et al., 2016; Navarro-Torre et al., 2017; Zhou et al., 2017). Rhizobacterial strains that produce EPSs improve plant growth under salt stress through rhizosheath development around the plant roots, which limits the Na+ influx inside the stele (Ashraf et al., 2004). In wheat plants, EPS production by PGPR ameliorated salt stress via fusion of Na+ ions to EPS, leading to enhanced plant nutrition and growth. EPS adheres to soil particles to build macro aggregates that stabilize soil structure and ultimately improve its hydraulic water holding and cation exchange capacity (Upadhyay et al., 2011a, b).
Salt stress impedes plant growth through elevated levels of Na+ and a lower K+/Na+ ratio. Bacillus-colonized Gladiolus plants displayed increased K+ uptake relative to Na+, reducing the Na+/K+ ratio under saline conditions (Damodaran et al., 2014). Rhizobacterially inoculated maize plants improved ionic balance by enhancing root K+ uptake and Na+ exclusion to confer salt tolerance (Rojas-Tapias et al., 2012). In an interesting study by Pinedo et al. (2015), PGPR B. phytofirmans PsJN-primed Arabidopsis plants were found to sustain salt stress conditions, and this correlated with altered expression of genes involved in ionic equilibrium, such as Arabidopsis K+ Transporter1 (KT1), High-Affinity K+ Transporter1 (HKT1), Sodium Hydrogen Exchanger2 (NHX2), and Arabidopsis Salt Overly Sensitive 1 (SOS1). Bacillus subtilis GB03-colonized Puccinellia tenuiflora displayed increased expression of PtHKT1 and PtSOS1 and downregulated expression of PtHKT2 genes in plant roots. Thus, limiting Na+ ion uptake in roots and their subsequent translocation reduced Na+ ion accumulation (Niu et al., 2016). Higher Na+ concentrations in rhizospheres provide strong competition against K+ ions, elevating the Na+/K+ ratio and causing higher stress by disrupting metabolic and physiological processes. Higher K+/Na+ ratios in AI plants is caused by the controlled translocation of Na+ ions to aboveground tissues of host plants and their accumulation in vacuoles. Salinity caused an escalation in the Na+ shoot to root ratio levels; however, this ratio was reduced in AMF seedlings (Evelin et al., 2012; Porcel et al., 2016). Additionally, it has been confirmed that enhanced K+ absorption and reduced Na+ transportation to shoot tissues lead to higher K+/Na+ ratios in AI plants during salt stress (Sharifi et al., 2007; Talaat and Shawky, 2011; Estrada et al., 2013a), which serve to preserve enzymatic processes and protein synthesis.
AMF acts as a primary barrier for absorption of ions during fungal nutrient uptake from soil or their transportation to host plants. This is attributed to the capability of AMF to retain these minerals in intraradical mycelium and vesicles via ionic accumulation in vacuoles (Mardukhi et al., 2011). This type of selective ion absorption (higher K+, Mg2+, and Ca2+ uptake; reduced Na+ uptake) leads to higher K+/Na+, Ca2+/Na+, and Mg2+/Na+ ratios in Rhizophagus intraradices. In addition, AI plants have increased Na+ content, up to a certain limit, which is then reduced at higher salinity levels, suggesting an AMF-induced buffering effect on Na+ uptake (Hammer and Rillig, 2011). AI-colonized wheat plants showed a significant increase in yield at various salinity levels that correlated to higher levels of N, P, and K, and reduced levels of Na+ in the leaves (Talaat and Shawky, 2013). Arbuscular mycorrhizal fungi improved K+ ion retention in maize plant tissues following upregulated expression of ZmAKT2 and ZmSKOR (Estrada et al., 2013b). A significant increase in K+ levels and decrease in Na+ levels were observed in AMF plants, suggesting selective uptake of K+ but not Na+ into the xylem of plant roots, which thus increases the K+/Na+ ratio under salt stress conditions to improve plant growth (Elhindi et al., 2017). Nitrogen assimilation in AI host plants is more efficient due to nitrate assimilation and higher enzyme production in the extraradical mycelia (Evelin et al., 2009; Kapoor et al., 2013). Enzymatic activities and protein synthesis were preserved in AI plants by increased nitrate reductase activity (caused by the elevated nitrate assimilation), increased K+ accumulation, and an improved K+/Na+ ratio (Talaat and Shawky, 2014). Wang et al. (2018) reported enhanced N uptake by plant roots in AI plants, which increased root length and root and shoot weight, to be the major mechanism underlying enhanced salt tolerance in Diversispora versiformis-colonized Chrysanthemum morifolium. Increased absorption of nutrients including Fe, K, Ca, Fe, Zn, and Mg, but restricted Na and Cl uptake in AI-colonized plants has been reported as a STEM to maintain ionic equilibrium and mitigate the effects of salt stress (Evelin et al., 2012; Kapoor et al., 2013). During salt stress, increased rhizospheric Na+ levels obstruct Ca2+ absorption and therefore disrupt the Ca2+:Na+ ratio of host plants, ultimately decreasing their hydraulic conductivity and disturbing Ca2+ signaling. Improving nutritional status is essential for conserving membrane integrity in AI plants. Mycorrhizal association increased Ca2+ and Mg2+ absorption by plant roots, even under soil salinity (Giri and Mukerji, 2004; Sharifi et al., 2007). Ca2+ levels were increased in Piriformospora indica -colonized barley plants, leading to the activation of signal transduction pathways to enhance stress tolerance in host plants (Alikhani et al., 2013). Given that Mg2+ is centrally located in the chlorophyll molecule, deceases in its uptake can reduce chlorophyll content and photosynthesis and eventually hamper plant growth. In AI host plants, an increase in Mg2+ uptake increases chlorophyll concentration to boost photosynthesis and plant performance while under stress conditions (Abdel Latef and Chaoxing, 2014). Enhanced salt tolerance in Rhizophagus irregularis-colonized E. angustifolia seedlings was correlated with increased K+, Ca2+, and Mg2+ uptake. Additionally, AM symbioses altered root architecture, and extraradical mycelia improved mineral uptake. K+ accumulation was increased in the roots and leaves of AI seedlings, leading to an enhanced K+/Na+ ratio in the plants and suggesting a STEM in plants (Chang et al., 2018).
Modifications in Plant Physiological Status
Microbially induced STEM includes phytohormonal modifications and alterations in other physiological processes, such as gas exchange, photosynthesis, and nutrient and water uptake. Various studies have established the role of microbes in alleviating the negative effects induced by salt stress on plant physiological performance (Porcel et al., 2015; Chen et al., 2017).
Phytohormonal Modulations
Microbes can promote plant growth during salt stress by altering the hormonal status of NI plants. Various phytohormones, including auxins, gibberellins, cytokinin, ethylene, ABA, JA, and SA, are involved in signaling events during plant–microbe interactions that can rescue plants under stress conditions. Numerous studies have reported roles for bacteria in phytohormonal modulations in response to salt stress, but there are relatively few studies related to AMF-mediated phytohormonal salt stress tolerance in host plants. Production of auxins, such as indole acetic acid (IAA), by PGPR strains has been well documented in RI plants and ensures plant survival during salt stress (Egamberdieva, 2009; Jha et al., 2012; Sharma et al., 2016; Sorty et al., 2016; Navarro-Torre et al., 2017). Rhizobacterially inoculated wheat plants showed elevated IAA levels in their rhizospheres compared to NI plants, which led to improved plant growth and survival under stress conditions (Tiwari et al., 2011). Higher root growth was reported in Pseudomonas chlororaphis TSAU13-primed wheat seedlings, tomato, and cucumber plants. IAA production by PGPR P. chlororaphis TSAU13 altered phytohormonal levels in plants and consequently enhanced stress tolerance compared to NI plants (Egamberdieva and Kucharova, 2009; Egamberdieva, 2012). Higher IAA levels in wheat plants inoculated with PGPR strains Arthrobacter protophormiae and D. natronolimnaea enabled host plants to survive salt stress (Barnawal et al., 2017). Chickpea plants co-inoculated with IAA-synthesizing rhizobacterial strains (B. subtilis NUU4 and Mesorhizobium ciceri IC53) exhibited increased root and shoot biomass, along with enhanced nodule formation relative to untreated plants and plants treated solely with M. ciceri IC53 (Egamberdieva et al., 2017). Similar observations of increased IAA levels have been reported in PGPR inoculated peanut (Sharma et al., 2016), barley (Cardinale et al., 2015), and wheat (Singh and Jha, 2016; Sorty et al., 2016). Plant growth-promoting rhizobacteria can modulate GA levels in RI plants (Kang et al., 2014a, b). Increased endogenous levels of gibberellins in Pseudomonas putida H-2-3 primed soybean plants (Kang et al., 2014b) and improved plant growth in salt stress conditions. Although cytokinin production is common in microbial strains and imparts stress tolerance to inoculated plants (Liu et al., 2013), very few studies have reported the role of cytokinin in MI plants. Higher proline content, in addition to increased shoot and root biomass, has been reported during salt stress in soybean plants inoculated with the cytokinin synthesizing rhizobacterial strains Arthrobacter, Azospirillum, and Bacillus (Naz et al., 2009). Accelerated ethylene synthesis above threshold values while under stress conditions restricts plant growth by negatively affecting root development and seed germination. However, rhizobacterial strains possessing ACCd can limit ethylene levels by cleaving the ethylene precursor, 1-aminocyclopropane-1-carboxylate (ACC), to produce ammonia and a-ketobutyrate. Various studies have reported plant growth promoted by ACCd-producing PGPR strains that also alleviates salt stress (Ali et al., 2014; Bharti et al., 2014). Nadeem et al. (2009) reported that ACCd-producing PGPR strains (Pseudomonas fluorescens, and Enterobacter spp.) improved the mineral status of maize plants, thereby helping them to counteract salt stress. Priming chickpea plants with ACCd-producing Pantoea dispersa PSB3 led to decreased Na+ uptake and elevated chlorophyll levels and relative leaf water levels, resulting in increased pod number and weight, biomass, and seed weight during salt stress (Panwar et al., 2016). Pea plants primed with ACCd-producing Variovorax paradoxus 5C-2 displayed root to shoot K+ ionic flow and Na+ ion root deposition, which led to elevated K+/Na+ ratios in the shoots. Moreover, a higher photosynthesis rate and decreased stomatal resistance caused the plant biomass to increase, thus enhancing stress tolerance (Wang et al., 2016). Enterobacter spp. UPMR18-treated okra plants displayed higher antioxidant enzyme activities and increased transcription of ROS pathway genes (Habib et al., 2016). Abscisic acid is a major stress phytohormone capable of alleviating abiotic stress by mediating the important physiological processes of stomatal opening and photosynthesis. Enhanced root and shoot growth were observed in rice plants treated with ABA-producing endophytic bacteria (Shahzad et al., 2017). D. natronolimnaea STR1-treated wheat plants showed enhanced salt tolerance via alteration of the ABA signaling cascade, which was confirmed by upregulation of TaABARE (ABA-responsive gene) and TaOPR1 (12-oxophytodienoate reductase 1) genes (Bharti et al., 2016). Treatment of maize plants with Bacillus amyloliquefaciens SQR9 conferred salt stress tolerance; treated plants counteracted increased ABA levels and exhibited enhanced chlorophyll levels, glutathione content, and K+/Na+ ratios (Chen et al., 2016). A significant increase in JA content and decrease in SA were recorded during salt stress in soybean plants (Kang et al., 2014b). Increased nutrient acquisition and salt stress tolerance were observed in maize plants upon inoculation with SA-synthesizing Serratia marcescens (Lavania and Nautiyal, 2013). Rice plants treated with B. amyloliquefaciens RWL-1 have been shown to have elevated endogenous SA levels and lower endogenous JA and ABA levels compared to plants treated with GA3 and water (Shahzad et al., 2016). Treatment with B. megaterium highlighted the role of JA-Ile turnover in the recovery of Arabidopsis plants from salt stress (Erice et al., 2017). Elevation of photosynthetic pigments and shoot biomass was reported in soybean plants under salt stress conditions in response to gibberellins produced by Aspergillus fumigatus (Khan et al., 2011). Decreased ABA production was found to regulate transpiration rate in cucumber plants colonized by AMF. However, JA and SA synthesis was increased upon AMF inoculation, which decreased oxidative damage to enhance salt stress tolerance (Hashem et al., 2018). In another study, higher levels of JA and its precursor, OPDA, were found in Digitaria eriantha colonized by R. irregularis under salt stress conditions, demonstrating the key role of JA in conferring salt tolerance to plants (Pedranzani et al., 2016). Strigolactones are the latest class of phytohormones found to be involved in adventitious root formation, reproductive development, and stress responses. A positive correlation was found between ABA and strigolactones in AMF-colonized Sesbania. Raised ABA levels caused higher H2O2 production, which led to increased SA synthesis and subsequently protected mycorrhizal plants from salt stress (Ren et al., 2018). Aroca et al. (2013) reported that strigolactone production was induced in AM-colonized lettuce plants under salt stress conditions and was correlated to ABA. More research studies targeting the role of AMF in altering phytohormonal levels in plants during salt stress are needed.
Improved Photosynthesis and Other Physiological Changes
Arbuscular mycorrhizal fungi-inoculated plants sustain higher chlorophyll and carotenoid levels through enhanced Mg2+ ion uptake (Evelin et al., 2012; Hashem et al., 2015), which is otherwise restricted by salt stress. Physiological changes of AI and RI plants also included a higher quantum yield of PSII and increased net photosynthetic rate (Talaat and Shawky, 2014; Chen et al., 2016, 2017; Hidri et al., 2016; Wang et al., 2016; Yasin et al., 2018) relative to NI or control plants. Glycinebetaine preserves the activities of RuBisCO and rubisco activase involved in CO2 fixation and secures PSII pigment-protein complexes (Talaat and Shawky, 2014) to confer salt stress tolerance to plants (Porcel et al., 2016; Hu et al., 2017). Higher RuBisCO activity has been reported in AMF plants (Garg and Bhandari, 2016b; Chen et al., 2017) and is correlated with increased RprbcL gene expression (Chen et al., 2017). AM symbiosis alleviates the physiological drought effects on photosynthesis by improving the water status of colonized plants through increases in their leaf area and stomatal conductance (Chen et al., 2017). Arbuscular mycorrhizal fungi can protect the photosystems by reducing non-photochemical quenching in inoculated plants relative to uninoculated ones and thus enhance their photosynthetic efficiency under salt stress conditions (Hu et al., 2017). Higher stomatal conductance and strong photosystem efficiency in AMF-colonized plants reduced photorespiration and lowered ROS production, which subsequently conferred salt stress tolerance (Ruiz-Sánchez et al., 2010). In AM rice plants under salt stress conditions, the enhanced quantum yield of photosystem II and reduced non-photochemical quenching-maintained photosynthesis, transpiration, and stomatal conductance led to increased biomass (Porcel et al., 2015). Ye et al. (2019) reported that AMF alleviated the decrease in the maximum photochemical efficiency of Photosystem II (Fv/Fm), photochemical quenching and increase in non-photochemical quenching (NPQ) observed in salt-stressed watermelon plants. Numerous studies have reported higher WUE, stomatal conductance, transpiration rate, and photosynthesis in AI plants exposed to salt stress compared to non-colonized plants (Sheng et al., 2008; Evelin et al., 2012; Elhindi et al., 2017). Wu et al. (2015) investigated the role of AMF in different sexes of P. cathayana during salt stress. Fv/Fm was elevated in males compared to females. Moreover, the improved efficiency of photosystem II and the antioxidant machinery in mycorrhizal seedlings alleviated the effects of salt stress. Increases in Fv/Fm, NPQ, and ETR were observed in the leaves of AI E. angustifolia compared to non-mycorrhizal plants under salt stress (Jia et al., 2019). Increased electrical conductivity was noticed in AMF-colonized plants and is caused by the reduced electrolyte permeability of root plasma membranes relative to non-mycorrhizal plant roots (Garg and Manchanda, 2008). Increased hydraulic conductance (Aroca et al., 2007), root system modifications (Campanelli et al., 2013; Alqarawi et al., 2014), and an improved water status were observed in AI plants under salt stress (Chen et al., 2017). Improved water status in AMF plants correlates with the expression of aquaporin genes (RpPIP1;1, RpPIP1;3, RpPIP2;1, RpTIP1;1, RpTIP1;3, RpTIP2;1) in leaves and roots of plants under salt stress; however, expression levels vary according to plant species, salinity levels, and location or tissue of expression (Chen et al., 2017). Higher WUE was observed in sweet basil plants colonized with Glomus deserticola compared to control plants under salt stress (Elhindi et al., 2017).
Significant increases in fresh and dry weight, plant height, and chlorophyll content were observed in PGPR-inoculated pepper plants relative to NI plants (Hahm et al., 2017). An increase in chlorophyll levels in PGPR-inoculated plants (Shukla et al., 2012; Bharti et al., 2014) and higher WUE of inoculated capsicum plants enabled them to survive under salt stress conditions (Yasin et al., 2018). Higher chlorophyll production, Na+ exclusion from roots, and antioxidant production enhanced salt stress tolerance in maize plants inoculated with B. amyloliquefaciens (Chen et al., 2016). Increased photosynthetic rates and decreased stomatal resistance enhanced the plant biomass of PGPR-inoculated pea plants (Wang et al., 2016). An interesting study by Chatterjee et al. (2018) found that treatment of rice plants with ACC deaminase-containing Brevibacterium linens RS16B decreased volatile organic compound (VOC) emissions and enhanced photosynthesis by reducing the availability of ACC and ACC oxidase activity, suggesting that research on volatile emissions during salt stress can reveal new insights related to stress severity and the initiation of secondary metabolism with stress progression. Ansari et al. (2019) demonstrated that PGPR inoculation increased root length, chlorophyll pigments, leaf number, relative water content (RWC), stomatal conductance (gs), and photosynthesis rate (Pn) in alfalfa plants under salt stress. Pseudomonas inoculation improved leaf number, chlorophyll pigments, nodule number, Na+ levels, and K+/Na+ ratios; however, Hartmannibacter inoculation improved carotenoid content, RWC, and K+ levels. Higher gs and chlorophyll pigments enhanced Pn in RI alfalfa plants. Inoculation with Bacillus megaterium strain A12 ameliorated salt stress in tomato plants by restoring redox homeostasis and photosynthesis to improve plant growth. Higher expression levels of the PBGD gene (encodes the enzyme needed for chlorophyll biosynthesis) enhanced chlorophyll content in tomato plants. In addition, reduction of ROS levels upregulated expression of the PsbA gene (encodes D1 protein that repairs stress damaged photosystem). Increased cytokinin production diminished the degradation of photosynthetic proteins and elevated the expression levels of genes related to photosystems under stress conditions (Akram et al., 2019).
Molecular Alterations
Multi-omics approaches, such as metagenomics, metatranscriptomics, and metaproteomics, can be utilized to enhance our understanding of plant behavior under stress conditions. However, it remains a challenge to integrate data from various “omics” tools, although this would be a step toward understanding the complex crosstalk between plants and microbes (Meena et al., 2017). STEM affecting various physiological and biochemical processes under salt stress involve variations in expression levels of genes, including ion transporters, aquaporins, and Δ1-pyrroline-5-carboxylate synthetase (P5CS), and variable levels of antioxidant defense enzymes, late embryogenesis abundant protein photosynthesis, antioxidant defense ionic homeostasis, and other signaling events. However, molecular alterations are categorized as a separate STEM to provide a better understanding, at transcriptional and proteomic levels, of how the plant response to salt stress is influenced.
Transcriptional Studies
Upregulation of SOS1, NADP-Me2 (NADP-malic enzyme), EREBP (ethylene-responsive element binding proteins), and SERK1 (somatic embryogenesis receptor-like kinase) and downregulation of GIG (glucose-insensitive growth) and SNF1 (serinethreonine protein kinase SAPK4) expression were observed in B. amyloliquefaciens SN13-inoculated rice plants exposed to salt stress. Modulated gene expression mitigated osmotic and ionic stress in plants to improve plant performance under stress conditions (Nautiyal et al., 2013). A significant increase in the transcriptional levels of AtRSA1 and AtWRKY8 and decrease in the expression of AtVQ9 (AtWRKY8 antagonist) in PGPR-treated Arabidopsis plants suggested enhanced plant performance under stress conditions resulting from microbial colonization. AtRSA1 forms a complex with the AtRITF1 transcription factor to regulate Na+ ion homeostasis and ROS detoxification during salt stress; AtWRKY8 and AtVQ9 are also involved in preserving ion homeostasis at reduced Na+/K+ ratios in the cytosol (Sukweenadhi et al., 2015). Upregulated expression of genes involved in stress responses [e.g., RAB18 (LEA), RD29B regulons of ABA-responsive elements], proline biosynthesis (e.g., P5CS1 and P5CS2), and MPK3 and MPK6 stress responses have also been reported (Kim et al., 2014). Upregulated expression of the TaCTR1 (Serine/Threonine protein kinase–ethylene responsive) and TaDREB2 (encodes a transcription factor enhancing abiotic stress tolerance in plants) genes was reported in wheat plants inoculated with PGPR strains (Kaushal and Wani, 2016b; Barnawal et al., 2017). B. subtilis GB03-inoculated P. tenuiflora plants showed less Na+ accumulation in response to upregulated expression of PtHKT1;5 (involved in the acquisition of Na+ from xylem) and SOS1 (role in Na+ efflux) and downregulation of PtHKT2;1 (involved in Na+ absorption in roots) to ameliorate salt stress in host plants (Niu et al., 2016). Improved physiological performance led to salt tolerance in B. amyloliquefaciens SQR9-treated maize plants and correlated to significant increases in the expression of RBCS and RBCL (RuBisCo subunits), ion transporters (HKT1, NHX1, and NHX2), and H(C)-Ppase (encoding HC pumping pyrophosphatase). However, the expression of NCED (encoding 9-cisepoxycarotenoid dioxygenase) was downregulated in RI seedlings (Chen et al., 2016). Increased expression of ionic transporters, such as TaNHX and TaHKT1, and the salt-induced stress gene TaST (reduces intracellular Na+ level, raises K+ content) was observed in PGPR-primed plants relative to NI plants. In addition, upregulated expression of the ABA-responsive gene (TaABARE), 12-oxophytodienoate reductase, or TaOPR1 (enhances antioxidant response) induced expression of TaMYB and TaWRKY, which led to expression of stress-related genes, including TaST. Plant tolerance to salt stress correlated with higher gene expression levels of CAT, APX, MnSOD, POD, GPX, and GR, which together modulated the antioxidant defense system to ultimately confer salt tolerance on inoculated host plants (Bharti et al., 2016). Volatile organic compounds generated by B. subtilis reduced gene expression of the high-affinity K+ transporter (HKT1) in the roots of inoculated Arabidopsis plants to limit Na+ uptake by the roots (Zhang et al., 2008). Higher transcriptional levels of genes involved in ABA signaling (RD29A and RD29B), ROS quenching (APX2), and detoxification (GLYI7) were reported in Burkholderia phytofirmans PsJN-colonized Arabidopsis plants. Additionally, expression of (LOX2) or Lipoxygenase2 (involved in JA biosynthesis) was downregulated; however, expression patterns of ion transport genes were varied between roots and shoots. Arabidopsis K+ Transporter 1 or AKT1 (plasma membrane transporter responsible for K+ uptake in roots) expression levels decreased in roots and rosettes after 24 h of salt stress. Sodium Hydrogen Exchanger 2 or NHX2 (a vacuolar antiporter engaged in ion compartmentalization) was upregulated in roots at all time points, but its expression varied at different points depending on salt stress and bacterial inoculation status. Salt Overly Sensitive 1 or SOS1 is another plasma membrane Na+/H+ antiporter (involved in Na+ removal from the cytoplasm). Bacterial inoculation caused upregulation of SOS1 expression in roots after 2 h, and expression was then downregulated after 24 and 72 h. However, in rosettes, increased expression levels were detected after 2 and 24 h. Bacterial inoculation downregulated the expression of High-Affinity K+ Transporter 1 or HKT 1 (sodium transporter) in roots under salt stress. Moreover, expression of HKT 1 was upregulated in the rosettes of non-stressed and colonized plants at 24 h but was downregulated at 24 and 72 h regardless of bacterial inoculation status (Pinedo et al., 2015). Arthrobacter woluwensis AK1-treated soybean plants exhibited upregulated expression of various genes, including GmLAX1 (auxin resistant 1), GmAKT2 (potassium channel), GmST1 (salt tolerance 1), and GmSALT3 (salt tolerance-related gene on chromosome 3) whereas downregulated expression of the ion transporter genes GmNHX1 (chloride channel gene) and GmCLC1 (Na+/H+ antiporter) was observed (Khan et al., 2019). A significant increase in the expression of stress related genes, such as CAPIP2 (aquaporin), stress related CaKR1, CaOSM1 (osmotin), and CAChi2 (Class II chitinase), was observed in PGPR inoculated capsicum plants, resulting in the modulation of various biochemical and physiological mechanisms to alleviate salt stress in plants (Yasin et al., 2018). In MI wheat plantlets, transcriptional studies found upregulated expression of P450s genes (CYP98A1, CYP734A5, CYP72A15, and CYP710A1) involved in redox reactions and stress responses, APX, and Nicotianamine synthase (NAS), which is responsible for iron absorption. Moreover, higher expression of oligopeptide transporters (membrane proteins able to transport different substrates), ATP binding cassette (ABC) transporters (proteins mediating energy-driven transport of various substrates), and HKT and NHX antiporters conferred salt stress resistance on inoculated plants (Safdarian et al., 2019). Increased expression of PIP genes was reported in AMF plants compared to non-AMF plants, which improved root water permeability and ameliorated the effects of salt stress conditions (Aroca et al., 2007; Jahromi et al., 2008). Mycorrhizal colonization boosted the expression levels of three chloroplast genes (RppsbA, RppsbD, and RprbcL) (encoding the larger subunit of rubisco) in leaves and genes involved in ion homeostasis (RpSOS1, RpHKT1, and RpSKOR). Higher expression of RpSOS1 and RpHKT1 decreased Na+ accumulation, while increased RpSKOR expression improved K+ accumulation in the leaves of mycorrhizal plants, leading to higher K+/Na+ ratios. Additionally, upregulated expression of RpPIP1;1 and RpPIP1;3 was observed in both leaves and roots; however, RpPIP2;1, and RpTIP1;1 were found to be more highly expressed in the roots of mycorrhizal plants under salt stress (Chen et al., 2017). Higher nitrogen uptake in salt-stressed mycorrhizal colonized wheat plants correlated with increased expression of NRT1.1 (involved in nitrate uptake in roots); however, the expression of ammonium transporters (AMT1.1 and AMT1.2) was unaffected. In addition, expression levels of genes related to drought stress (AQP1, AQP4, PIP1, DREB5, and DHN15.3) were lower compared to non-mycorrhizal wheat plants (Fileccia et al., 2017). Mycorrhizal colonization alleviated the reduced expression levels of RBCL (involved in photosynthesis) induced by salt stress; however, salinity raised expression levels of PPH (responsible for chlorophyll degradation), which was significantly increased after AMF colonization. Furthermore, genes involved in antioxidant defense responses (APX, Cu-Zn SOD, CAT, and GR) showed increased expression levels that were further enhanced by mycorrhizal treatment (Ye et al., 2019).
Proteomics Studies
Microbial colonization under salt stress leads to up- and downregulated expression of various proteins. Proteomic analysis can reveal the protein profile of inoculated plants to decode the STEM. Detected proteins can be utilized for genetic transformation to boost salt tolerance in crops. Molecular studies performed by Alikhani et al. (2013) revealed that proteome trends in P. indica-inoculated barley plants were different from those of NI plants under salt stress. The abundance of the protein peroxiredoxins-2E-2 (component of antioxidant defense system) was increased in both PGPR-colonized and NI plants in the presence of 300 mM NaCl; however, the increase in protein level was greater in P. indica-primed plants. Higher expression levels of RBCS (small chain of RUBISCO) were reported in P. indica-colonized plants compared to NI counterparts. Increased expression levels of xyloglucan endotransglycosylase or XET (involved in cell wall biosynthesis) and tubulin-folding cofactor A (involved in cell wall division) were observed in inoculated plants; however, there was no change in expression levels in inoculated plants (with 300 mM NaCl). Furthermore, the expression of papain-like cysteine proteases (cell signaling pathways) was reduced in NI plants, whereas its expression levels were increased in inoculated plants, enhancing salt stress tolerance. Proteomic analysis in AI E. angustifolia seedlings showed interesting results. Increased expression of a core protein of PS II (D1 precursor processing PSB27) involved in stabilizing the PSII reaction center was found in chloroplasts, and mitochondria showed increased levels of various energy-related proteins, such as NADH dehydrogenase, iron-sulfur protein NADH dehydrogenase, cytochrome C oxidase, and ATP synthase, to provide energy for cellular activities. Phosphoribosyl transferase or APT (enzyme in tryptophan synthesis) was also upregulated in response to AMF colonization. Upregulation of four peptidyl prolyl cis-trans isomerases (peptidyl-prolyl cis-trans isomerase (FKBP12), peptidyl-prolyl cis-trans isomerase (CYP18-1), FKBP-type peptidyl-prolyl cis-trans isomerase 5 isoform 1, and peptidyl-prolyl cis-trans isomerase (FKBP62), along with four molecular chaperones, prefoldin subunit 1, prefoldin subunit 2, heat shock 70 kDa, and partial and small hsp 17.3 kDa, was observed in mycorrhizal seedlings to enable correct protein folding under salt stress. In this study, upregulated expression of proteins involved in signal transduction, such as G proteins, plasma membrane Ca2+ transporter ATPase (PMCA), calcium-dependent protein kinases (CDPKs), and calmodulin (CaM), enhanced Ca2+ signaling. Thus, AMF colonization enhanced expression of proteins involved in secondary metabolism, antioxidant defense, and signal transduction to lead to increased salt tolerance in E. augustifolia seedlings (Jia et al., 2019).
Conclusion and Recommendations
Microbially inoculated plants induce STEM to counteract salt stress and enhance plant productivity. STEM can promote nutrient uptake, enhanced WUE and photosynthesis, preservation of ionic homeostasis and osmoprotection, and efficient antioxidant metabolism. In recent years, various studies have reported that plant–microbe interactions can develop STEM in host plants; however, some aspects of this phenomenon remain poorly understood. Future studies should investigate the role of phytohormonal crosstalk (e.g., BR, JA, and strigolactones) in MI plants to understand their role in eliciting stress signals during salt stress. Metabolomic studies should aim to understand the STEM underlying the secondary metabolism in salt-stressed MI plants. There is also a need for studies investigating the nutritional uptake of sulfur in MI plants under salt stress, given its involvement with glutathione and cysteine (involved in ABA synthesis). Moreover, as the cell wall is the first line of defense during salt stress, subsequent studies should target biochemical and molecular alterations relating to the cell wall of MI plants. Insights into the plant immune system triggered in response to microbial partners while under stress conditions should also be addressed to harness plant microbial interactions for agricultural benefits.
Author Contributions
MK reviewed, wrote, and revised the manuscript.
Conflict of Interest
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Abd Allah, E. F., Hashem, A., Alqarawi, A. A., Bahkali, A. H., and Alwhibi, M. S. (2015). Enhancing growth performance and systemic acquired resistance of medicinal plant Sesbania sesban (L.) Merr using arbuscular mycorrhizal fungi under salt stress. Saudi J. Biol. Sci. 22, 274–283. doi: 10.1016/j.sjbs.2015.03.004
Abdel Latef, A. A., and Chaoxing, H. (2014). Does the inoculation with Glomus mosseae improve salt tolerance in pepper plants? J. Plant Growth Regul. 33, 644–653. doi: 10.1007/s00344-014-9414-4
Abo-Doma, A., Edrees, S., and Abdel-Aziz, S. H. (2016). The effect of mycorrhiza growth and expression of some genes in barley. Egypt J. Genet. Cytol. 40, 301–313. doi: 10.21608/ejgc.2011.10794
Akram, W., Aslam, H., Ahmad, S. R., Anjum, T., Yasin, N. A., Khan, W. U., et al. (2019). Bacillus megaterium strain A12 ameliorates salinity stress in tomato plants through multiple mechanisms. J. Plant Inter. 14, 506–518. doi: 10.1080/17429145.2019.1662497
Ali, S., Charles, T. C., and Glick, B. R. (2014). Amelioration of high salinity stress damage by plant growth promoting bacterial endophytes that contain ACC deaminase. Plant Physiol. Biochem. 80, 160–167.
Alikhani, M., Khatabi, B., Sepehri, M., Nekouei, M. K., Mardi, M., and Salekdeh, G. H. (2013). A proteomics approach to study the molecular basis of enhanced salt tolerance in barley (Hordeum vulgare L.) conferred by the root mutualistic fungus Piriformospora indica. Mol Biosyst. 9, 1498–1510. doi: 10.1039/c3mb70069k
Al-Khaliel, A. S. (2010). Effect of salinity stress on mycorrhizal association and growth response of peanut infected by Glomus mosseae. Plant Soil Environ. 56, 318–324.
Alqarawi, A. A., Abd-Allah, E. F., and Hashem, A. (2014). Alleviation of salt-induced adverse impact via mycorrhizal fungi in Ephedra aphylla Forssk. J. Plant Interact. 9, 802–810.
Ansari, M., Shekari, F., Mohammadi, M. H., Juhos, K., Vegvari, G., and Biro, B. (2019). Salt-tolerant plant growth-promoting bacteria enhanced salinity tolerance of salt-tolerant alfalfa (Medicago sativa L.) cultivars at high salinity. Acta Physiol. Plant 41:195. doi: 10.1007/s11738-019-2988-5
Aroca, R., Porcel, R., and Ruiz-Lozano, J. M. (2007). How does arbuscular mycorrhizal symbiosis regulate root hydraulic properties and plasma membrane aquaporins in Phaseolus vulgaris under drought, cold or salinity stresses? New Phytol. 173, 808–816.
Aroca, R., Ruiz-Lozano, J. M., Zamarreno, A. M., Paz, J. A., Garcia-Mina, J. M., Pozo, M. J., et al. (2013). Arbuscular mycorrhizal symbiosis influences strigolactone production under salinity and alleviates salt stress in lettuce plants. J. Plant Physiol. 170, 47–55. doi: 10.1016/j.jplph.2012.08.020
Ashraf, M., Hasnain, S., Berge, O., and Mahmood, T. (2004). Inoculating wheat seedling with exopolysaccharide-producing bacteria restricts sodium uptake and stimulates plant growth under salt stress. Biol. Fert. Soils 40, 157–162.
Barnawal, D., Bharti, N., Pandey, S. S., Pandey, A., Chanotiya, C. S., and Kalra, A. (2017). Plant growth promoting rhizobacteria enhances wheat salt and drought stress tolerance by altering endogenous phytohormone levels and TaCTR1/TaDREB2 expression. Physiol. Plant. 161, 502–514. doi: 10.1111/ppl.12614
Bharti, N., Barnawal, D., Awasthi, A., Yadav, A., and Kalra, A. (2014). Plant growth promoting rhizobacteria alleviate salinity induced negative effects on growth, oil content and physiological status in Mentha arvensis. Acta Physiol. Plant. 36, 45–60.
Bharti, N., Pandey, S. S., Barnawal, D., Patel, V. K., and Kalra, A. (2016). Plant growth promoting rhizobacteria Dietzia natronolimnaea modulates the expression of stress responsive genes providing protection of wheat from salinity stress. Sci. Rep. 6:34768. doi: 10.1038/srep34768
Bowles, T. M., Barrios-Masias, F. H., Carlisle, E. A., Cavagnaro, T. R., and Jackson, L. E. (2016). Effects of arbuscular mycorrhizae on tomato yield, nutrient uptake, water relations, and soil carbon dynamics under deficit irrigation in field conditions. Sci. Total Environ. 566, 1223–1234. doi: 10.1016/j.scitotenv.2016.05.178
Campanelli, A., Ruta, C., De Mastro, G., and Morone-Fortunato, I. (2013). The role of arbuscular mycorrhizal fungi in alleviating salt stress in Medicago sativa L. var. icon. Symbiosis 59, 65–76. doi: 10.1007/s13199-012-0191-1
Cardinale, M., Ratering, S., Suarez, C., Montoya, A. M. Z., Geissler-Plaum, R., and Schnell, S. (2015). Paradox of plant growth promotion potential of rhizobacteria and their actual promotion effect on growth of barley (Hordeum vulgare L.) under salt stress. Microbiol. Res. 181, 22–32. doi: 10.1016/j.micres.2015.08.002
Chang, W., Sui, X., Fan, X. X., Jia, T. T., and Song, F. Q. (2018). Arbuscular mycorrhizal symbiosis modulates antioxidant response and ion distribution in salt-stressed Elaeagnus angustifolia seedlings. Front. Microbiol. 9:652. doi: 10.3389/fmicb.2018.00652
Chatterjee, P., Kanagendran, A., Samaddar, S., Pazouki, L., Sa, T. M., and Niinemets, Ü (2018). Inoculation of Brevibacterium linens RS16 in Oryza sativa genotypes enhanced salinity resistance: impacts on photosynthetic traits and foliar volatile emissions. Sci. Total Environ. 645, 721–732. doi: 10.1016/j.scitotenv.2018.07.187
Chen, J., Zhang, H., Zhang, X., and Tang, M. (2017). Arbuscular mycorrhizal symbiosis alleviates salt stress in black locust through improved photosynthesis, water status, and K+/Na+ homeostasis. Front. Plant Sci. 8:1739. doi: 10.3389/fpls.2017.01739
Chen, L., Liu, Y., Wu, G., Veronican Njeri, K., Shen, Q., Zhang, N., et al. (2016). Induced maize salt tolerance by rhizosphere inoculation of Bacillus amyloliquefaciens SQR9. Physiol. Plant. 158, 34–44.
Chu, T. N., Tran, B. T. H., Van Bui, L., and Hoang, M. T. T. (2019). Plant growth-promoting rhizobacterium Pseudomonas PS01 induces salt tolerance in Arabidopsis thaliana. BMC Res. Notes 12:11. doi: 10.1186/s13104-019-4046-1
Damodaran, T., Rai, R. B., Jha, S. K., Kannan, R., Pandey, B. K., Sah, V., et al. (2014). Rhizosphere and endophytic bacteria for induction of salt tolerance in Gladiolus grown in sodic soils. J. Plant Inter. 9, 577–584.
Echeverria, M., Sannazarro, A. I., Ruiz, O. A., and Menendez, A. B. (2013). Modulatory effects of Mesorhizobium tianshanense and Glomus intraradices on plant proline and polyamine levels during early plant response of Lotus tenuis to salinity. Plant Soil 364, 69–79. doi: 10.1007/s11104-012-1312-6
Egamberdieva, D. (2009). Alleviation of salt stress by plant growth regulators and IAA producing bacteria in wheat. Acta Physiol. Plant. 31, 861–864. doi: 10.1007/s11738-009-0297-0
Egamberdieva, D. (2012). Pseudomonas chlororaphis: a salt tolerant bacterial inoculant for plant growth stimulation under saline soil conditions. Acta Physiol. Plant. 34, 751–756. doi: 10.1007/s11738-011-0875-9
Egamberdieva, D., and Kucharova, Z. (2009). Selection for root colonising bacteria stimulating wheat growth in saline soils. Biol. Fertil. Soils 45, 563–571. doi: 10.1007/s00374-009-0366-y
Egamberdieva, D., Wirth, S., Shurigin, V., Hashem, A., and Abd_Allah, E. F. (2017). Endophytic bacteria improve plant growth, symbiotic performance of chickpea (Cicer arietinum L.) and induce suppression of root rot caused by Fusarium solani under salt stress. Front. Microbiol. 8:1887. doi: 10.3389/fmicb.2017.01887
El-Esawi, M. A., Al-Ghamdi, A. A., Ali, H. M., and Alayafi, A. A. (2019). Azospirillum lipoferum FK1 confers improved salt tolerance in chickpea (Cicer arietinum L.) by modulating osmolytes, antioxidant machinery and stress related genes expression. Environ. Exp. Bot. 159, 55–65.
Elhindi, K. M., El-Din, A. S., and Elgorban, A. M. (2017). The impact of arbuscular mycorrhizal fungi in mitigating salt-induced adverse effects in sweet basil (Ocimum basilicum L). Saudi J. Biol. Sci. 24, 170–179.
Erice, G., Ruíz-Lozano, J. M., Zamarreño, A. M., García-Mina, J. M., and Aroca, R. (2017). Transcriptomic analysis reveals the importance of JA-Ile turnover in the response of Arabidopsis plants to plant growth promoting rhizobacteria and salinity. Environ. Exp. Bot. 143, 10–19. doi: 10.1016/j.envexpbot.2017.08.006
Estrada, B., Aroca, R., Barea, J. M., and Ruiz-Lozano, J. M. (2013a). Native arbuscular mycorrhizal fungi isolated from a saline habitat improved maize antioxidant systems and plant tolerance to salinity. Plant Sci. 20, 42–51. doi: 10.1016/j.plantsci.2012.11.009
Estrada, B., Aroca, R., Maathuis, F. J., Barea, J. M., and Ruiz-Lozano, J. M. (2013b). Arbuscular mycorrhizal fungi native from a Mediterranean saline area enhance maize tolerance to salinity through improved ion homeostasis. Plant Cell Environ. 36, 1771–1782. doi: 10.1111/pce.12082
Etesami, H. (2018). Can interaction between silicon and plant growth promoting rhizobacteria benefit in alleviating abiotic and biotic stresses in crop plants? Agric. Ecosyst. Environ. 253, 98–112. doi: 10.1016/j.agee.2017.11.007
Evelin, H., Giri, B., and Kapoor, R. (2012). Contribution of Glomus intraradices inoculation to nutrient acquisition and mitigation of ionic imbalance in NaCl stressed Trigonella foenum-graecum. Mycorrhiza 22, 203–217. doi: 10.1007/s00572-011-0392-0
Evelin, H., Giri, B., and Kapoor, R. (2013). Ultrastructural evidence for AMF mediated salt stress mitigation in Trigonella foenum-graecum. Mycorrhiza 23, 71–86. doi: 10.1007/s00572-012-0449-8
Evelin, H., and Kapoor, R. (2014). Arbuscular mycorrhizal symbiosis modulates antioxidant response in salt-stressed Trigonella foenum-graecum plants. Mycorrhiza 24, 197–208. doi: 10.1007/s00572-013-0529-4
Evelin, H., Kapoor, R., and Giri, B. (2009). Arbuscular mycorrhizal fungi in alleviation of salt stress: a review. Ann. Bot. 104, 1263–1280. doi: 10.1093/aob/mcp251
Feng, G., Zhang, F. S., Li, X. L., Tian, C. Y., Tang, C., and Rengel, Z. (2002). Improved tolerance of maize plants to salt stress by arbuscular mycorrhiza is related to higher accumulation of soluble sugars in roots. Mycorrhiza 12, 185–190. doi: 10.1007/s00572-002-0170-0
Fileccia, V., Ruisi, P., Ingraffia, R., Giambalvo, D., Frenda, A. S., and Martinelli, F. (2017). Arbuscular mycorrhizal symbiosis mitigates the negative effects of salinity on durum wheat. PLoS One 12:e0184158.
Garg, N., and Baher, N. (2013). Role of arbuscular mycorrhizal symbiosis in proline biosynthesis and metabolism of Cicer arietinum L. (chickpea) genotypes under salt stress. J. Plant Growth Regul. 32, 767–778. doi: 10.1007/s00344-013-9346-4
Garg, N., and Bhandari, P. (2016a). Interactive effects of silicon and arbuscular mycorrhiza in modulating ascorbate-glutathione cycle and antioxidant scavenging capacity in differentially salt-tolerant Cicer arietinum L. genotypes subjected to long-term salinity. Protoplasma 253, 1325–1345. doi: 10.1007/s00709-015-0892-4
Garg, N., and Bhandari, P. (2016b). Silicon nutrition and mycorrhizal inoculations improve growth, nutrient status, K+/Na+ ratio and yield of Cicer arietinum L. genotypes under salinity stress. Plant Growth Regul. 78, 371–387. doi: 10.1007/s10725-015-0099-x
Garg, N., and Bharti, A. (2018). Salicylic acid improves arbuscular mycorrhizal symbiosis, and chickpea growth and yield by modulating carbohydrate metabolism under salt stress. Mycorrhiza 28, 727–746. doi: 10.1007/s00572-018-0856-6
Garg, N., and Manchanda, G. (2008). Effect of arbuscular mycorrhizal inoculation of salt-induced nodule senescence in Cajanus cajan (pigeon pea). J. Plant Growth Regul. 27, 115–124.
Garg, N., and Pandey, R. (2016). High effectiveness of exotic arbuscular mycorrhizal fungi is reflected in improved rhizobial symbiosis and trehalose turnover in Cajanus cajan genotypes grown under salinity stress. Fungal Ecol. 21, 57–67. doi: 10.1016/j.funeco.2016.04.001
Giri, B., and Mukerji, K. G. (2004). Mycorrhizal inoculant alleviates salt stress in Sesbania aegyptiaca and Sesbania grandiflora under field conditions: evidence for reduced sodium and improved magnesium uptake. Mycorrhiza 14, 307–312.
Gururani, M. A., Upadhyaya, C. P., Baskar, V., Venkatesh, J., Nookaraju, A., and Park, S. W. (2013). Plant growth-promoting rhizobacteria enhance abiotic stress tolerance in Solanum tuberosum through inducing changes in the expression of ROS-scavenging enzymes and improved photosynthetic performance. J. Plant Growth Regul. 32, 245–258. doi: 10.1007/s00344-012-9292-6
Habib, S. H., Kausar, H., and Saud, H. M. (2016). Plant growth-promoting rhizobacteria enhance salinity stress tolerance in okra through ROS-scavenging enzymes. Biomed. Res. Int. 2016:6284547. doi: 10.1155/2016/628454
Hahm, M. S., Son, J. S., Hwang, Y. J., Kwon, D. K., and Ghim, S. Y. (2017). Alleviation of salt stress in pepper (Capsicum annum L.) plants by plant growth-promoting rhizobacteria. J. Microbiol. Biotechnol. 27, 1790–1797. doi: 10.4014/jmb.1609.09042
Hajiboland, R., Aliasgharzadeh, N., Laiegh, S. F., and Poschenrieder, C. (2010). Colonization with arbuscular mycorrhizal fungi improves salinity tolerance of tomato (Solanum lycopersicum L.) plants. Plant Soil 331, 313–327. doi: 10.1007/s11104-009-0255-z
Hammer, E. C., and Rillig, M. C. (2011). The influence of different stresses on glomalin levels in an arbuscular mycorrhizal fungus-salinity increases glomalin content. PLoS One 6:e28426. doi: 10.1371/journal.pone.0028426
Hashem, A., Abd-Allah, E. F., Alqarawi, A. A., Aldubise, A., and Egamberdieva, D. (2015). Arbuscular mycorrhizal fungi enhances salinity tolerance of Panicum turgidum Forssk by altering photosynthetic and antioxidant pathways. J. Plant Interact. 10, 230–242. doi: 10.1080/17429145.2015.1052025
Hashem, A., Alqarawi, A. A., Radhakrishnan, R., Al-Arjani, A. B. F., Aldehaish, H. A., Egamberdieva, D., et al. (2018). Arbuscular mycorrhizal fungi regulate the oxidative system, hormones and ionic equilibrium to trigger salt stress tolerance in Cucumis sativus L. Saudi J. Biol. Sci. 25, 1102–1114.
Hidri, R., Barea, J. M., Mahmoud, O. M. B., Abdelly, C., and Azcón, R. (2016). Impact of microbial inoculation on biomass accumulation by Sulla carnosa provenances, and in regulating nutrition, physiological and antioxidant activities of this species under non-saline and saline conditions. J. Plant Physiol. 201, 28–41. doi: 10.1016/j.jplph.2016.06.013
Hu, W., Zhang, H., Chen, H., and Tang, M. (2017). Arbuscular mycorrhizas influence Lycium barbarum tolerance of water stress in a hot environment. Mycorrhiza 27, 451–463. doi: 10.1007/s00572-017-0765-0
Jahromi, F., Aroca, R., Porcel, R., and Ruiz-Lozano, J. M. (2008). Influence of salinity on the in vitro development of Glomus intraradices and on the in vivo physiological and molecular responses of mycorrhizal lettuce plants. Microb. Ecol. 55, 45–53. doi: 10.1007/s00248-007-9249-7
Jha, B., Gontia, I., and Hartmann, A. (2012). The roots of the halophyte Salicornia brachiata are a source of new halotolerant diazotrophic bacteria with plant growth-promoting potential. Plant Soil 356, 265–277. doi: 10.1007/s11104-011-0877-9
Jha, Y., Subramanian, R. B., and Patel, S. (2011). Combination of endophytic and rhizospheric plant growth promoting rhizobacteria in Oryza sativa shows higher accumulation of osmoprotectant against saline stress. Acta Physiol. Plant. 33, 797–802.
Jia, T., Wang, J., Chang, W., Fan, X., Sui, X., and Song, F. (2019). Proteomics analysis of E. angustifolia seedlings inoculated with arbuscular mycorrhizal fungi under salt stress. Int. J. Mol. Sci. 20:788. doi: 10.3390/ijms20030788
Kang, S. M., Khan, A. L., Waqas, M., You, Y. H., Kim, J. H., Kim, J. G., et al. (2014a). Plant growth-promoting rhizobacteria reduce adverse effects of salinity and osmotic stress by regulating phytohormones and antioxidants in Cucumis sativus. J. Plant Inter. 9, 673–682.
Kang, S. M., Radhakrishnan, R., Khan, A. L., Kim, M.-J., Park, J.-M., Kim, B.-R., et al. (2014b). Gibberellin secreting rhizobacterium Pseudomonas putida H-2-3 modulates the hormonal and stress physiology of soybean to improve the plant growth under saline and drought conditions. Plant Physiol. Biochem. 84, 115–124.
Kapoor, R., Evelin, H., Mathur, P., and Giri, B. (2013). “Arbuscular mycorrhiza: approaches for abiotic stress tolerance in crop plants for sustainable agriculture,” in Plant Acclimation to Environmental Stress, eds N. Tuteja and S. S. Gill (Berlin: Springer), 359–401.
Kaushal, M. (2019). Microbes in cahoots with plants: MIST to hit the jackpot of agricultural productivity during drought. Int. J. Mol. Sci. 20:1769. doi: 10.3390/ijms20071769
Kaushal, M., and Wani, S. P. (2016a). Plant growth promoting rhizobacteria: drought stress alleviators to ameliorate crop production in drylands. Ann. Microbiol. 66, 35–42.
Kaushal, M., and Wani, S. P. (2016b). Rhizobacterial-plant interactions: strategies ensuring plant growth promotion under drought and salinity stress. Agric. Ecosyst. Environ. 231, 68–78.
Khan, A. L., Hamayun, M., Kim, Y. H., Kang, S. M., and Lee, I. J. (2011). Ameliorative symbiosis of endophyte (Penicillium funiculosum LHL06) under salt stress elevated plant growth of Glycine max L. Plant Physiol. Biochem. 49, 852–861. doi: 10.1016/j.plaphy.2011.03.005
Khan, M. A., Asaf, S., Khan, A. L., Jan, R., Kang, S. M., Kim, K. M., et al. (2019). Rhizobacteria AK1 remediates the toxic effects of salinity stress via regulation of endogenous phytohormones and gene expression in soybean. Biochem. J. 476, 2393–2409. doi: 10.1042/BCJ20190435
Kim, K., Jang, Y. J., Lee, S. M., Oh, B. T., Chae, J. C., and Lee, K. J. (2014). Alleviation of salt stress by Enterobacter sp. EJ01 in tomato and Arabidopsis is accompanied by up-regulation of conserved salinity responsive factors in plants. Mol. Cells 37, 109–117. doi: 10.14348/molcells.2014.2239
Kohler, J., Hernández, J. A., Caravaca, F., and Roldán, A. (2009). Induction of antioxidant enzymes is involved in the greater effectiveness of a PGPR versus AM fungi with respect to increasing the tolerance of lettuce to severe salt stress. Environ. Exp. Bot. 65, 245–252. doi: 10.1016/j.envexpbot.2008.09.008
Latef, A. A. H. A., and Chaoxing, H. (2011) Effect of arbuscular mycorrhizal fungi on growth, mineral nutrition, antioxidant enzymes activity and fruit yield of tomato grown under salinity stress. Sci. Hortic. 127, 228–233. doi: 10.1016/j scienta.2010.09.020.
Lavania, M., and Nautiyal, C. (2013). Solubilization of tricalcium phosphate by temperature and salt tolerant Serratia marcescens NBRI1213 isolated from alkaline soils. Afr. J. Microbiol. Res. 7, 4403–4413.
Li, T., Run-Jin, L., Xin-Hua, H., and Bao-Shan, W. (2012). Enhancement of superoxide dismutase and catalase activities and salt tolerance of euhalophyte Suaeda salsa L. by mycorrhizal fungus Glomus mosseae. Pedosphere 22, 217–224. doi: 10.1016/S1002-0160(12)60008-3
Liu, B., Zhao, S., Tan, F., Zhao, H., Wang, D., Si, H., et al. (2017). Changes in ROS production and antioxidant capacity during tuber sprouting in potato. Food Chem. 237, 205–213. doi: 10.1016/j.foodchem.2017.05.107
Liu, F., Xing, S., Ma, H., Du, Z., and Ma, B. (2013). Cytokinin-producing, plant growth promoting rhizobacteria that confer resistance to drought stress in Platycladus orientalis container seedlings. Appl. Microbiol. Biotechnol. 97, 9155–9164.
Liu, S., Guo, X., Feng, G., Maimaitiaili, B., Fan, J., and He, X. (2016). Indigenous arbuscular mycorrhizal fungi can alleviate salt stress and promote growth of cotton and maize in saline fields. Plant Soil 398, 195–206. doi: 10.1007/s11104-015-2656-5
Mardukhi, B., Rejali, F., Daei, G., Ardakani, M. R., Malakouti, M. J., and Miransari, M. (2011). Arbuscular mycorrhizas enhance nutrient uptake in different wheat genotypes at high salinity levels under field and greenhouse conditions. C. R. Biol. 334, 564–571. doi: 10.1016/j.crvi.2011.05.001
Meena, K. K., Sorty, A. M., Bitla, U. M., Choudhary, K., Gupta, P., Pareek, A., et al. (2017). Abiotic stress responses and microbe-mediated mitigation in plants: the omics strategies. Front. Plant Sci. 8:172. doi: 10.3389/fpls.2017.00172
Nadeem, S. M., Zahir, Z. A., Naveed, M., and Arshad, M. (2009). Rhizobacteria containing ACC-deaminase confer salt tolerance in maize grown on salt-affected fields. Can. J. Microbiol. 55, 1302–1309.
Nautiyal, C. S., Srivastava, S., Chauhan, P. S., Seem, K., Mishra, A., and Sopory, S. K. (2013). Plant growth-promoting bacteria Bacillus amyloliquefaciens NBRISN13 modulates gene expression profile of leaf and rhizosphere community in rice during salt stress. Plant Physiol. Biochem. 66, 1–9.
Navarro-Torre, S., Barcia-Piedras, J. M., Mateos-Naranjo, E., Redondo-Gómez, S., Camacho, M., Caviedes, M. A., et al. (2017). Assessing the role of endophytic bacteria in the halophyte Arthrocnemum macrostachyum salt tolerance. Plant Biol. 19, 249–256. doi: 10.1111/plb.12521
Naz, I., Bano, A., and Ul-Hassan, T. (2009). Isolation of phytohormones producing plant growth promoting rhizobacteria from weeds growing in Khewra salt range, Pakistan and their implication in providing salt tolerance to Glycine max L. Afr. J. Biotechnol. 8, 5762–5766.
Niu, S. Q., Li, H. R., Pare, P. W., Aziz, M., Wang, S. M., Shi, H. Z., et al. (2016). Induced growth promotion and higher salt tolerance in the halophyte grass Puccinellia tenuiflora by beneficial rhizobacteria. Plant Soil 407, 217–230. doi: 10.1007/s11104-015-2767-z
Pandey, R., and Garg, N. (2017). High effectiveness of Rhizophagus irregularis is linked to superior modulation of antioxidant defence mechanisms in Cajanus cajan (L.) Millsp. genotypes grown under salinity stress. Mycorrhiza 27, 669–682. doi: 10.1007/s00572-017-0778-8
Panwar, M., Tewari, R., Gulati, A., and Nayyar, H. (2016). Indigenous salt-tolerant rhizobacterium Pantoea dispersa (PSB3) reduces sodium uptake and mitigates the effects of salt stress on growth and yield of chickpea. Acta Physiol. Plant. 38:278. doi: 10.1007/s11738-016-2284-6
Pedranzani, H., Rodríguez-Rivera, M., Gutiérrez, M., Porcel, R., Hause, B., and Ruiz-Lozano, J. M. (2016). Arbuscular mycorrhizal symbiosis regulates physiology and performance of Digitaria eriantha plants subjected to abiotic stresses by modulating antioxidant and jasmonate levels. Mycorrhiza 26, 141–152. doi: 10.1007/s00572-015-0653-4
Pinedo, I., Ledger, T., Greve, M., and Poupin, M. J. (2015). Burkholderia phytofirmans PsJN induces long-term metabolic and transcriptional changes involved in Arabidopsis thaliana salt tolerance. Front. Plant Sci. 6:466. doi: 10.3389/fpls.2015.00466
Porcel, R., Aroca, R., Azcon, R., and Ruiz-Lozano, J. M. (2016). Regulation of cation transporter genes by the arbuscular mycorrhizal symbiosis in rice plants subjected to salinity suggests improved salt tolerance due to reduced NaCl root-to-shoot distribution. Mycorrhiza 26, 673–684. doi: 10.1007/s00572-016-0704-5
Porcel, R., Redondo-Gómez, S., Mateos-Naranjo, E., Aroca, R., Garcia, R., and Ruiz-Lozano, J. M. (2015). Arbuscular mycorrhizal symbiosis ameliorates the optimum quantum yield of photosystem II and reduces non-photochemical quenching in rice plants subjected to salt stress. J. Plant Physiol. 185, 75–83. doi: 10.1016/j.jplph.2015.07.006
Prasad, R., Kumar, M., and Varma, A. (2015). “Role of PGPR in soil fertility and plant health,” in Plant Growth Promoting Rhizobacteria and Medicinal Plants, eds D. Egamberdeiva, S. Shrivastava, and A. Varma (Switzerland: Springer International Publishing), 247–260.
Qin, Y., Druzhinina, I. S., Pan, X., and Yuan, Z. (2016). Microbially mediated plant salt tolerance and microbiome-based solutions for saline agriculture. Biotechnol. Adv. 34, 1245–1259. doi: 10.1016/j.biotechadv.2016.08.005
Qurashi, A. W., and Sabri, A. N. (2012). Bacterial exopolysaccharide and biofilm formation stimulate chickpea growth and soil aggregation under salt stress. Braz. J. Microbiol. 43, 1183–1191. doi: 10.1590/S1517-838220120003000046
Rangel, D. E. (2011). Stress induced cross-protection against environmental challenges on prokaryotic and eukaryotic microbes. World J. Microbiol. Biotechnol. 27, 1281–1296. doi: 10.1007/s11274-010-0584-3
Ren, C. G., Kong, C. C., and Xie, Z. H. (2018). Role of abscisic acid in strigolactone induced salt stress tolerance in arbuscular mycorrhizal Sesbania cannabina seedlings. BMC Plant Biol. 18:74. doi: 10.1186/s12870-018-1292-7
Rojas-Tapias, D., Moreno-Galvan, A., Pardo-Diaz, S., Obando, M., Rivera, D., and Bonilla, R. (2012). Effect of inoculation with plant growth-promoting bacteria (PGPB) on amelioration of saline stress in maize (Zea mays). Appl. Soil Ecol. 61, 264–272.
Romero-Munar, A., Baraza, E., Gulías, J., and Cabot, C. (2019). Arbuscular mycorrhizal fungi confer salt tolerance in giant reed (Arundo donax L.) plants grown under low phosphorus by reducing leaf NaCl concentration and improving phosphorus use efficiency. Front. Plant Sci. 10:843. doi: 10.3389/fpls.2019.00843
Ruiz-Lozano, J. M., Azcon, R., and Gomez, M. (1996). Alleviation of salt stress by arbuscular mycorrhizal Glomus species in Lactuca sativa plants. Physiol. Plantarum 98, 767–772.
Ruiz-Sánchez, M., Aroca, R., Muñoz, Y., Polón, R., and Ruiz-Lozano, J. M. (2010). The arbuscular mycorrhizal symbiosis enhances the photosynthetic efficiency and the antioxidative response of rice plants subjected to drought stress. J. Plant Physiol. 167, 862–869. doi: 10.1016/j.jplph.2010.01.018
Safdarian, M., Askari, H., Shariati, V., and Nematzadeh, G. (2019). Transcriptional responses of wheat roots inoculated with Arthrobacter nitroguajacolicus to salt stress. Sci. Rep. 9:1792. doi: 10.1038/s41598-018-38398-2
Sapre, S., Gontia-Mishra, I., and Tiwari, S. (2018). Klebsiella sp. confers enhanced tolerance to salinity and plant growth promotion in oat seedlings (Avena sativa). Microbiol. Res. 206, 25–32. doi: 10.1016/j.micres.2017.09.009
Shahzad, R., Khan, A. L., Bilal, S., Waqas, M., Kang, S. M., and Lee, I. J. (2017). Inoculation of abscisic acid-producing endophytic bacteria enhances salinity stress tolerance in Oryza sativa. Environ. Exp. Bot. 136, 68–77. doi: 10.1016/j.envexpbot.2017.01.010
Shahzad, R., Waqas, M., Khan, A. L., Asaf, S., Khan, M. A., Kang, S. M., et al. (2016). Seed-borne endophytic Bacillus amyloliquefaciens RWL-1 produces gibberellins and regulates endogenous phytohormones of Oryza sativa. Plant Physiol. Biochem. 106, 236–243. doi: 10.1016/j.plaphy.2016.05.006
Sharifi, M., Ghorbanli, M., and Ebrahimzadeh, H. (2007). Improved growth of salinity-stressed soybean after inoculation with pre-treated mycorrhizal fungi. J. Plant Physiol. 164, 1144–1151.
Sharma, S., Kulkarni, J., and Jha, B. (2016). Halotolerant rhizobacteria promote growth and enhance salinity tolerance in peanut. Front. Microbiol. 7:1600. doi: 10.3389/fmicb.2016.01600
Sheng, M., Tang, M., Chen, H., Yang, B., Zhang, F., and Huang, Y. (2008). Influence of arbuscular mycorrhizae on photosynthesis and water status of maize plants under salt stress. Mycorrhiza 18, 287–296. doi: 10.1007/s00572-008-0180-7
Sheng, M., Tang, M., Zhang, F., and Huang, Y. (2011). Influence of arbuscular mycorrhiza on organic solutes in maize leaves under salt stress. Mycorrhiza 21, 423–430. doi: 10.1007/s00572-010-0353-z
Shukla, P. S., Agarwal, P. K., and Jha, B. (2012). Improved salinity tolerance of Arachis hypogaea (L.) by the interaction of halotolerant plant growth- promoting rhizobacteria. J. Plant Growth Regul. 31, 195–206. doi: 10.1007/s00344-011-9231-y
Singh, R. P., and Jha, P. N. (2016). Alleviation of salinity-induced damage on wheat plant by an ACC deaminase-producing halophilic bacterium Serratia sp SL-12 isolated from a salt lake. Symbiosis 69, 101–111. doi: 10.1007/s13199-016-0387-x
Singh, R. P., and Jha, P. N. (2017). The PGPR Stenotrophomonas maltophilia SBP-9 augments resistance against biotic and abiotic stress in wheat plants. Front. Microbiol. 8:1945. doi: 10.3389/fmicb.2017.01945
Sorty, A. M., Meena, K. K., Choudhary, K., Bitla, U. M., Minhas, P. S., and Krishnani, K. K. (2016). Effect of plant growth promoting bacteria associated with halophytic weed (Psoralea corylifolia L) on germination and seedling growth of wheat under saline conditions. Appl. Biochem. Biotechnol. 180, 872–882. doi: 10.1007/s12010-016-2139-z
Sukweenadhi, J., Balusamy, S. R., Kim, Y. J., Lee, C. H., Kim, Y. J., Koh, S. C., et al. (2018). A growth-promoting bacteria, Paenibacillus yonginensis DCY84T enhanced salt stress tolerance by activating defense-related systems in Panax ginseng. Front. Plant Sci. 9:813. doi: 10.3389/fpls.2018.00813
Sukweenadhi, J., Kim, Y. J., Choi, E. S., Koh, S. C., Lee, S. W., Kim, Y. J., et al. (2015). Paenibacillus yonginensis DCY84T induces changes in Arabidopsis thaliana gene expression against aluminum, drought, and salt stress. Microbiol. Res. 172, 7–15.
Talaat, N. B., and Shawky, B. T. (2011). Influence of arbuscular mycorrhizae on yield, nutrients, organic solutes, and antioxidant enzymes of two wheat cultivars under salt stress. J. Plant Nutr. Soil Sci. 174, 283–291.
Talaat, N. B., and Shawky, B. T. (2013). Modulation of nutrient acquisition and polyamine pool in salt-stressed wheat (Triticum aestivum L.) plants inoculated with arbuscular mycorrhizal fungi. Acta Physiol. Plant. 35, 2601–2610. doi: 10.1007/s11738-013-1295-9
Talaat, N. B., and Shawky, B. T. (2014). Protective effects of arbuscular mycorrhizal fungi on wheat (Triticum aestivum L.) plants exposed to salinity. Environ. Exp. Bot. 98, 20–31. doi: 10.1016/j.envexpbot.2013.10.005
Tewari, S., and Arora, N. K. (2014). Multifunctional exopolysaccharides from Pseudomonas aeruginosa PF23 involved in plant growth stimulation, biocontrol and stress amelioration in sunflower under saline conditions. Curr. Microbiol. 69, 484–494.
Tiwari, S., Singh, P., Tiwari, R., Meena, K. K., Yandigeri, M., Singh, D. P., et al. (2011). Salt-tolerant rhizobacteria-mediated induced tolerance in wheat (Triticum aestivum) and chemical diversity in rhizosphere enhance plant growth. Biol. Fertil. Soils 47:907. doi: 10.1007/s00374-011-0598-5
Ullah, S., and Bano, A. (2015). Isolation of plant-growth-promoting rhizobacteria from rhizospheric soil of halophytes and their impact on maize (Zea mays L.) under induced soil salinity. Can. J. Microbiol. 61, 307–313. doi: 10.1139/cjm-2014-0668
Upadhyay, S. K., and Singh, D. P. (2015). Effect of salt-tolerant plant growth promoting rhizobacteria on wheat plants and soil health in a saline environment. Plant Biol. 17, 288–293. doi: 10.1111/plb.12173
Upadhyay, S. K., Singh, J. S., Saxena, A. K., and Singh, D. P. (2011a). Impact of PGPR inoculation on growth and antioxidant status of wheat under saline conditions. Plant Biol. 14, 605–611.
Upadhyay, S. K., Singh, J. S., and Singh, D. P. (2011b). Exopolysaccharide-Producing plant growth-promoting rhizobacteria under salinity condition. Pedosphere 21, 214–222.
Wang, Q., Dodd, I. C., Belimov, A. A., and Jiang, F. (2016). Rhizosphere bacteria containing 1-aminocyclopropane-1-carboxylate deaminase increase growth and photosynthesis of pea plants under salt stress by limiting Na+ accumulation. Funct. Plant Biol. 43, 161–172.
Wang, Y., Wang, M., Li, Y., Wu, A., and Huang, J. (2018). Effects of arbuscular mycorrhizal fungi on growth and nitrogen uptake of Chrysanthemum morifolium under salt stress. PLoS One 13:e0196408. doi: 10.1371/journal.pone.0196408
Wu, H. H., Zou, Y. N., Rahman, M. M., Ni, Q. D., and Wu, Q. S. (2017). Mycorrhizas alter sucrose and proline metabolism in trifoliate orange exposed to drought stress. Sci. Rep. 7:42389. doi: 10.1038/srep42389
Wu, N., Li, Z., Liu, H., and Tang, M. (2015). Influence of arbuscular mycorrhiza on photosynthesis and water status of Populus cathayana Rehder males and females under salt stress. Acta Physiol. Plant. 37, 183–197. doi: 10.1007/s11738-015-1932-6
Wu, N., Li, Z., Wu, F., and Tang, M. (2016). Comparative photochemistry activity and antioxidant responses in male and female Populuscathayana cuttings inoculated with arbuscular mycorrhizal fungi under salt. Sci. Rep. 6:37663. doi: 10.1038/srep37663
Xu, H., Lu, Y., and Tong, S. (2018). Effects of arbuscular mycorrhizal fungi on photosynthesis and chlorophyll fluorescence of maize seedlings under salt stress. Emir. J. Food Agric. 30, 199–204. doi: 10.9755/ejfa.2018.v30.i3.1642
Yasin, N. A., Akram, W., Khan, W. U., Ahmad, S. R., Ahmad, A., and Ali, A. (2018). Halotolerant plant-growth promoting rhizobacteria modulate gene expression and osmolyte production to improve salinity tolerance and growth in Capsicum annum L. Environ Sci. Pollut. Res. 25, 23236–23250. doi: 10.1007/s11356-018-2381-8
Ye, L., Zhao, X., Bao, E., Cao, K., and Zou, Z. (2019). Effects of arbuscular mycorrhizal fungi on watermelon growth, elemental uptake, antioxidant, and photosystem II activities and stress-response gene expressions under salinity-alkalinity stresses. Front. Plant Sci. 10:863. doi: 10.3389/fpls.2019.00863
Yu, J., Sun, L., Fan, N., Yang, Z., and Huang, B. (2015). Physiological factors involved in positive effects of elevated carbon dioxide concentration on Bermudagrass tolerance to salinity stress. Environ. Exp. Bot. 115, 20–27. doi: 10.1016/j.envexpbot.2015.02.003
Zhang, H., Kim, M. S., Sun, Y., Dowd, S. E., Shi, H., and Paré, P. W. (2008). Soil bacteria confer plant salt tolerance by tissue-specific regulation of the sodium transporter HKT1. Mol. Plant-Microbe Interact. 21, 737–744. doi: 10.1094/MPMI-21-6-0737
Zhou, C., Zhu, L., Xie, Y., Li, F., Xiao, X., Ma, Z., et al. (2017). Bacillus licheniformis SA03 confers increased saline-alkaline tolerance in Chrysanthemum plants by induction of abscisic acid accumulation. Front. Plant Sci. 8:1143. doi: 10.3389/fpls.2017.01143
Zhu, X., Song, F., Liu, S., Liu, F., and Li, X. (2018). Arbuscular mycorrhiza enhances nutrient accumulation in wheat exposed to elevated CO2 and soil salinity. J. Plant Nutr. Soil Sci. 81, 836–846. doi: 10.1002/jpln.201700575
Zou, Y. N., Wu, Q. S., Huang, Y. M., Ni, Q. D., and He, X. H. (2013). Mycorrhizal mediated lower proline accumulation in Poncirus trifoliata under water deficit derives from the integration of inhibition of proline synthesis with increase of proline degradation. PLoS One 8:e80568. doi: 10.1371/journal.pone.0080568
Keywords: salt stress, microbes, ion transporters, signal transduction, aquaporins, photosynthesis
Citation: Kaushal M (2020) Insights Into Microbially Induced Salt Tolerance and Endurance Mechanisms (STEM) in Plants. Front. Microbiol. 11:1518. doi: 10.3389/fmicb.2020.01518
Received: 28 January 2020; Accepted: 11 June 2020;
Published: 26 August 2020.
Edited by:
Naveen Kumar Arora, Babasaheb Bhimrao Ambedkar University, IndiaReviewed by:
Suprasanna Penna, Bhabha Atomic Research Centre (BARC), IndiaJitendra Mishra, Babasaheb Bhimrao Ambedkar University, India
Copyright © 2020 Kaushal. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Manoj Kaushal, a2F1c2hhbC5tYmdAZ21haWwuY29t; TS5LYXVzaGFsQGNnaWFyLm9yZw==