 Katharina Röltgen
Katharina Röltgen Gerd Pluschke
Gerd Pluschke- 1Department of Pathology, Stanford School of Medicine, Stanford University, Stanford, CA, United States
- 2Medical Parasitology and Infection Biology, Swiss Tropical and Public Health Institute, Basel, Switzerland
- 3University of Basel, Basel, Switzerland
Buruli ulcer (BU) is a neglected, tropical infectious disease of the skin and the subcutaneous tissue caused by Mycobacterium ulcerans. This pathogen has emerged as a new species from a common ancestor with Mycobacterium marinum by acquisition of the virulence plasmid pMUM. The plasmid encodes enzymes required for the synthesis of the macrolide toxin mycolactone, which has cytotoxic and immunosuppressive activities. In advanced BU lesions, extracellular clusters of M. ulcerans reside in necrotic subcutaneous tissue and are protected from infiltrating leukocytes by the cytotoxic activity of secreted mycolactone. Several lines of evidence indicate that elements of the innate immune system eliminate in many cases the initial inoculum before bacterial clusters can form and that therefore exposure to M. ulcerans leads only in a minority of individuals to the characteristic chronic necrotizing BU lesions. It is assumed that phagocytes play a key role in early host defense against M. ulcerans. Antibodies against bacterial surface structures seem to have less potential to enhance innate immunity than TH1 cell responses. Precise innate and adaptive immune effector mechanisms leading to protective immunity are however unclear, complicating the development of effective vaccines, the most desired solution to control BU. The tuberculosis vaccine Mycobacterium bovis Bacillus Calmette–Guérin (BCG) has limited short-term protective activity against BU. Whether this effect is due to the broad antigenic cross-reactivity between M. bovis and M. ulcerans or is at least partly mediated by a non-specific enhanced responsiveness of innate immune cells to secondary stimulation, recently described as “trained immunity” or “innate immune memory” is unknown but has major implications for vaccine design. Current vaccine research and development activities are focusing on recombinant BCG, subunit vaccines with selected M. ulcerans proteins, and the neutralization of mycolactone.
Introduction
Innate immunity constitutes the first line of host defense against potentially pathogenic microbial invaders. It comprises physical and various chemical barriers, including antimicrobial proteins, to prevent entry into the host, as well as innate humoral (such as the alternative complement pathway) and cellular defense mechanisms that come into play if the epithelial barriers are breached. Innate immune cells, such as macrophages, neutrophils, and dendritic cells express pattern recognition receptors (PRRs), such as toll-like receptors (TLRs) that can detect so called pathogen-associated molecular patterns (PAMPs) common to many microorganisms, or damage-associated molecular patterns (DAMPs) of host molecules released by infected or dying cells. Upon recognition, these “sensor cells” can either act directly as effectors, phagocytosing and degrading the pathogens or indirectly, by producing inflammatory mediators, such as cytokines and chemokines that can attract and activate other immune cells. If the infection persists, phagocytes also connect the innate with the adaptive immune system by presenting antigens to antigen-specific T and B cells. Evidence has been accumulating in recent years that after infection or vaccination, innate immune cells display changes in their transcription programs and cell physiology, which may lead to transiently increased responsiveness upon secondary stimulation by microbial pathogens, a phenomenon termed “trained immunity” (Netea et al., 2011, 2016).
Pathogenic mycobacteria, including Mycobacterium tuberculosis and its near relative M. marinum (Stinear et al., 2008) have developed mechanisms to subvert the innate immune response. They can establish residence inside host macrophages and use host granulomas – organized immune cell aggregates, characterized by the presence of mature macrophages, that can contain but fail to eradicate infection foci – for their expansion and dissemination during the innate phases of infection (Ramakrishnan, 2013). M. marinum, which causes a granulomatous, tuberculosis-like disease in ectotherms (Tobin and Ramakrishnan, 2008), has gained popularity as a model organism for mycobacterial infections and has thus been extensively studied. Amongst different model systems, experimentally infected zebrafish (Danio rerio) embryos and early swimming larvae have become a powerful resource to study contributions of innate immune responses to combat mycobacterial infections (Davis et al., 2002). Due to the absence of an adaptive immune system at these early developmental stages, the zebrafish model has significantly advanced our understanding of innate host defense against mycobacterial infections.
In comparison, little is known on early interactions of the immune system with M. ulcerans, which causes the chronic, necrotizing skin disease Buruli ulcer (BU) and has emerged as a new species from a common ancestor with M. marinum by acquisition of a virulence plasmid (pMUM) and subsequent reductive evolution (Doig et al., 2012). Despite more than 98% overall nucleotide identity between the genomes of the two pathogens (Stinear et al., 2007), M. ulcerans has developed a markedly different strategy for immune evasion, primarily due to the pMUM-mediated ability to produce mycolactone, a diffusible cytotoxic and immunosuppressive macrolide toxin (George et al., 1999). While an early intra-macrophage growth phase of M. ulcerans has been postulated (Coutanceau et al., 2005; Silva et al., 2009), in advanced disease, M. ulcerans bacilli are predominantly found extracellularly in the necrotic core of BU lesions, that is devoid of living, infiltrating immune cells (Ruf et al., 2017). Infection with M. ulcerans can either be contained by the immune system as indicated by reports of spontaneous healing (Marion et al., 2016a; O’Brien et al., 2019) and of M. ulcerans-specific immune responses in exposed, but healthy individuals (Gooding et al., 2001; Diaz et al., 2006; Yeboah-Manu et al., 2012), or can lead to serious dermatologic manifestations and chronic necrotizing disease (Pluschke and Röltgen, 2019). Understanding of early immune mechanisms involved in the diverse outcome of infection with M. ulcerans is however incomplete. In this review article, we compare the pathogenesis of M. ulcerans and M. marinum infections and summarize current data on innate immune mechanisms against infection with M. ulcerans. Knowledge on correlates of protection against BU has important implications for the rational design of a vaccine – the ideal solution to control the disease as discussed at the end of this article.
Mycobacterium ulcerans Has Evolved From an M. marinum-Like Progenitor
Common Ancestry…
Buruli ulcer mainly affects inhabitants of rural, focal areas in West and Central Africa and yet the host range of M. ulcerans is broad (Röltgen and Pluschke, 2015). Apart from human BU lesions that most commonly involve the extremities, M. ulcerans has been isolated from lesions of other mammals in Australia (Fyfe et al., 2010) and from diseased fish and frogs around the world (Trott et al., 2004; Rhodes et al., 2005; Ranger et al., 2006; Stragier et al., 2008). Based on comparative genomic data, two major lineages of mycolactone-producing mycobacteria (MPM) have been distinguished, the classical lineage isolated from humans in Africa, Australia, and Papua New Guinea, and from other mammals and the ancestral lineage, which can be subdivided into at least two deep rooted sub-lineages; human disease isolates from Japanese patients (also designated Mycobacterium ulcerans subsp. shinshuense) and strains isolated from humans in the Americas and from ectotherms (Käser et al., 2007; Doig et al., 2012). Genomic data indicate that MPM have emerged only once through the acquisition of pMUM and therefore all MPM should be designated M. ulcerans (Yip et al., 2007; Pidot et al., 2010).
Proximity to aquatic habitats, and particularly activities within stagnant or slow flowing water bodies have been identified as a common risk factor for human BU in different geographical areas (Raghunathan et al., 2005; Kenu et al., 2014; N’krumah et al., 2017; Maman et al., 2018). Knowledge on transmission pathways and reservoirs of M. ulcerans is still fragmentary, but inoculation of the bacteria into the skin by postulated insect vectors or from environmental reservoirs via skin trauma is hypothesized. Association of M. ulcerans with aquatic environments has long been suspected due to its emergence from M. marinum, a ubiquitous pathogen of fish and other ectotherms. Occasional human M. marinum infections, which most commonly involve fingers and/or hands, are nowadays mainly connected with exposure to fish tanks, handling of fish, and boating/fishing-related activities (Johnson and Stout, 2015). Transmission of M. marinum is thought to occur through inoculation of the bacteria into the skin via cuts or lacerations (Petrini, 2006).
Mycobacterium ulcerans and M. marinum grow optimally at around 30°C and poorly at 37°C and above, which may at least in part explain their skin tropism and limited systemic dissemination. Both pathogens belong to the group of slow-growing mycobacteria, whereby the generation time in microbial culture medium of M. ulcerans (several days) is considerably longer than that of M. marinum (∼4–6 h) (Clark and Shepard, 1963; Marsollier et al., 2004). A mean incubation period of 4.5 months has been determined for M. ulcerans infections in a study on BU patients in Australia (Loftus et al., 2018). The incubation period of M. marinum is estimated to be ∼3 weeks, but can be up to several months long (Jernigan and Farr, 2000). Infections with both pathogens may resolve spontaneously by activities of the immune system but require long-term antibiotic treatment when established. Current WHO treatment recommendations for M. ulcerans infections comprise a combination therapy with rifampicin and clarithromycin (or streptomycin) for 8 weeks and surgery if indicated (World Health Organization, Global Buruli Ulcer Initiative, 2004; Phillips et al., 2020). There are no clinical trials to guide optimal management of M. marinum infections, but treatment with two active agents (clarithromycin/azithromycin, ethambutol, or rifampicin) for 3–4 months with adjunct surgical debridement for invasive infections has been reported (Griffith et al., 2007). Person to person transmission of M. ulcerans and M. marinum infection is considered unlikely. Thus, at first glance, characteristics of M. ulcerans and M. marinum and of the infections they cause seem very similar. Marked differences emerge however when comparing pathogenesis and host responses evoked by the two related mycobacterial species.
… but Vastly Different Pathogenesis
Infection with M. ulcerans initially produces subcutaneous nodules or papules with a necrotic core or less frequently plaques and edema with laterally extended destruction of subcutaneous tissue. As the early stages of the disease are often painless, patients tend to report late to health facilities, when severe skin and soft tissue destruction has started. In advanced stages of the infection, the epidermis overlying the necrotic deeper layers of the skin sloughs off, and chronic ulcers with undermined edges develop. Histopathologic hallmarks of BU lesions are a progressive contiguous coagulative necrosis of the deep dermis and subcutaneous fat tissue with clusters of acid-fast bacilli (AFB), but no viable infiltrating leukocytes in the core of necrotic areas. In patients with chronic non-healing BU, squamous cell carcinoma may develop (Evans et al., 1999). Human M. marinum disease, often referred to as “fish tank granuloma” is commonly limited to a single, nodular cutaneous lesion, but can progress to invasive disease such as tenosynovitis and less frequently arthritis and osteomyelitis (Aubry et al., 2002; Johnson and Stout, 2015). The histopathological spectrum of M. marinum infections is broad and depends on the course and stage of the disease (Travis et al., 1985; Sia et al., 2016). Features in skin biopsies range from poorly formed granulomas with loose infiltrates of epithelioid macrophages, scattered multinucleated giant cells, and lymphohistocytic dermal inflammation to well-formed granulomas with circumscribed, nodular macrophage infiltrates. Granulomas frequently contain a central necrotic core, often surrounded by mixed inflammation and granulation tissue (Sia et al., 2016). In deep soft tissue and synovial biopsies, moderately well-formed, non-caseating or necrotizing, suppurative granulomas with giant cells are seen (Beckman et al., 1985; Sia et al., 2016). Acute and chronic synovial inflammation characterized by a paucity of plasma cells, often accompanied by synovial hyperplasia, and fibrin exudation into the synovial space has been reported (Beckman et al., 1985; Travis et al., 1985; Sia et al., 2016). The normal synovial architecture may be replaced by extensive granulation tissue (Travis et al., 1985). In both skin and synovial samples, bacilli are not easily detected as they reside highly localized in necrotic, suppurative cores of the granuloma (Sia et al., 2016). Variation in pathogenicity may also be related to genetic differences, as M. marinum strains exhibit extensive genomic diversity (van der Sar et al., 2004; Broutin et al., 2012). Despite an overwhelming sequence similarity of 98% between M. ulcerans and M. marinum, explanations for differences in pathogenesis and immune defense can be found in the gene content of their genomes.
It Is Written in the Genes…
Mycobacterium ulcerans has acquired both a virulence plasmid, encoding genes for the biosynthesis of the unique macrolide toxin mycolactone, and insertion sequence (IS) elements that have mediated extensive loss of DNA. Whereas the genome of M. marinum strain M encompasses a 6.6 Mb circular chromosome with 5424 coding sequences (CDS), and 65 pseudogenes (Stinear et al., 2008), the genome of the M. ulcerans classical lineage reference strain Agy99 is considerably smaller, comprising the virulence plasmid pMUM of 174 kb, and a 5.6 Mb circular chromosome with 4160 CDS, and 771 pseudogenes (Stinear et al., 2007). Production of the macrolide toxin mycolactone was likely the key factor enabling the evolution of M. ulcerans.
Mycolactone consists of a conserved 12-membered lactone ring with a C-linked upper side chain and a less conserved lower C5-O-linked polyunsaturated acyl side chain (George et al., 1999). The lower side chains of the mycolactone variants produced by different M. ulcerans sub-lineages are structurally diverse (Figure 1) and differ in toxic potency (Scherr et al., 2013). In addition to potent cytotoxic activity, mycolactone exhibits analgesic and immunosuppressive properties at sub-toxic concentrations (Phillips et al., 2009; Hall et al., 2014; Marion et al., 2014; Guenin-Macé et al., 2019). Low nanomolar concentrations of mycolactone cause in vitro apoptosis within 2–5 days in a wide range of mammalian cells (Bozzo et al., 2010; Scherr et al., 2013; Guenin-Mace et al., 2015; Ogbechi et al., 2015). The secreted toxin seems to diffuse passively through mammalian cell membranes and to promote Bim-dependent apoptosis through the Akt-FoxO3 axis, as demonstrated by the absence of necrotic BU lesions in M. ulcerans-infected Bim knock-out mice (Bieri et al., 2017). In contrast, necrotizing lesions with features of human BU are caused by injection of mycolactone or mycolactone-producing M. ulcerans into the skin of wild-type mice and other experimental animals (George et al., 2000; Oliveira et al., 2005; Sarfo et al., 2013). Among several distinct proposed molecular mechanisms for the mode of action of mycolactone, selective inhibition of the Sec61 translocon-mediated co-translational transport of secretory proteins into the ER seems to play the key role (Hall et al., 2014; Baron et al., 2016). The inhibition of protein translocation leads to pronounced proteomic changes and an integrated cellular stress response that ultimately seems to drive Bim-dependent apoptosis. In addition, downregulation of cytokines and chemokines at sub-toxic concentrations has strong immunosuppressive effects. The Wiskott–Aldrich syndrome protein (WASP) family has been proposed as another molecular target of mycolactone (Guenin-Macé et al., 2013). Binding of the toxin to WASP/N-WASP appears to lead to uncontrolled assembly of actin and defects in cell adhesion, which may obstruct innate cellular immune responses.

Figure 1. Structure of mycolactone variants. Mycolactone congeners were shown to be produced by M. ulcerans strains isolated from BU patients in Africa (mycolactone A/B), Australia (mycolactone C), and China (mycolactone D) and from fish and frogs of different geographical origin (mycolactone E and F).
One additional important difference in the gene content of the M. marinum and M. ulcerans genomes is the repertoire of ESX secretion systems and of PE/PPE proteins. The mycobacterial ESX loci are large gene clusters that encode a type VII secretory apparatus required for export of members of the 6-kDa early secreted antigenic target (ESAT-6) protein family together with other effector proteins across the complex cell envelope. Genes encoding ESAT-6 and the 10 kDa culture filtrate protein (CFP-10) are located directly adjacent to each other and are co-transcribed. The genome of the M. tuberculosis H37Rv strain contains 11 pairs of these esx tandem genes (Cole et al., 1998). M. marinum has 29 esx genes within five ESX loci (Stinear et al., 2008). The prototypical ESX-1 system is a major virulence determinant in M. tuberculosis and M. marinum, triggering granuloma formation as well as intercellular bacterial spread between macrophages (Volkman et al., 2004). The absence of a 9.5 kb genomic region across all Mycobacterium bovis Bacillus Calmette–Guérin (BCG) strains, termed Region of Difference 1 (RD1), located in the ESX-1 locus, is a major molecular determinant underlying BCG attenuation. M. marinum carries a partial duplication of the esx-1 gene cluster, resulting in more than one copy of several genes including esxA (ESAT-6) and esxB (CFP-10). On the other hand, M. ulcerans has retained only 13 esx genes and three intact ESX loci. Disruption of the ESX-1 locus in M. ulcerans classical lineage strains, which led to the abolishment of ESAT-6 and CFP-10 secretion (Huber et al., 2008), may contribute to the predominantly extracellular location of M. ulcerans bacilli (Stinear et al., 2007).
PE/PPE proteins, found mostly in slow-growing pathogenic mycobacteria, are characterized by conserved Pro-Glu (PE) and Pro-Pro-Glu (PPE) motifs at the N-termini. It has been reported that certain ESX secretion systems mediate the secretion of several PE/PPE proteins (Abdallah et al., 2009; Shah et al., 2015). The genome of the M. tuberculosis H37Rv strain contains 99 pe and 69 ppe genes, but this number can vary for different M. tuberculosis isolates (Fishbein et al., 2015). The M. marinum genome codes for 175 PE and 106 PPE proteins, whereas M. ulcerans has preserved only 70 intact pe and 46 ppe genes. The function of PE and PPE proteins is still enigmatic, but the limited data available suggest that they are important for mycobacterial virulence. Some of the PE proteins from M. marinum are thought to be involved in modulating the macrophage environment (Ramakrishnan et al., 2000; Tiwari et al., 2012; Fishbein et al., 2015). Compared to M. marinum, M. ulcerans has thus both gained a major virulence determinant – mycolactone and lost several other virulence factors mainly associated with the intracellular lifestyle of its ancestor. The following two paragraphs illustrate how these genomic differences are reflected in diverse interactions of M. marinum and M. ulcerans with the innate immune system.
Host-Mycobacterium Interactions – Insights Into Early Infection Events From a Zebrafish Model
In the past decades, M. marinum has become a model organism to study fundamental mechanisms of mycobacterial pathogen-host interactions. Zebrafish are naturally susceptible to M. marinum and upon infection develop organized granulomas similar to those caused by M. tuberculosis (Westerfield, 2000; Swaim et al., 2006). Zebrafish embryos and early swimming larvae are a powerful means to dissect innate immune responses to M. marinum, as at these early developmental stages, they rely solely on innate immune mechanisms mediated by macrophages and neutrophils, and lack the elements of adaptive immunity (Davis et al., 2002). In this paragraph, we describe how the genetic tractability and optical transparency of zebrafish embryos have enabled a variety of elegant experimental approaches to study early M. marinum infection events in vivo.
First Hours After Infection
Real-time imaging of zebrafish embryos revealed that after intravenous injection of M. marinum, blood macrophages immediately take up mycobacteria and extravasate into diverse tissues. Strikingly, recruitment of macrophages within 6 h after injection of M. marinum into the zebrafish hindbrain ventricle – an isolated cavity devoid of macrophages in the absence of pathogens – demonstrated redirection of normal migration and differentiation of embryonic macrophages (Davis et al., 2002). Both live and heat-killed M. marinum seem to be able to recruit phagocytes (heat-killed mycobacteria are subsequently readily degraded after phagocytosis), suggesting that cell wall lipids or heat-stable proteins stimulate migration (Clay et al., 2007). It has been postulated that M. marinum is able to evade microbicidal effects of TLR-activated macrophages by masking PAMPs with cell-surface associated lipids, and instead recruits permissive macrophages via an alternative pathway. In infected macrophages, the production of inflammatory cytokines, such as tumor necrosis factor (TNF)-α and interleukin (IL)-1β is upregulated (Cambier et al., 2014). Studies involving the depletion of macrophages in zebrafish embryos illustrated that macrophages are required to restrict proliferation of M. marinum and thus constitute only a suboptimal growth niche for the bacilli. They do however play an important role in the dissemination of mycobacteria into the tissues (Clay et al., 2007).
Migration Into Tissue and Granuloma Formation
After phagocytosing the M. marinum bacilli, macrophages migrate into deeper tissue, where they begin to form dynamic, granuloma-like aggregates, becoming visible three days after intravenous inoculation. Aggregated cells display euchromatic nuclei characteristic of activated macrophages and either tightly apposed cell membranes or indistinct cell boundaries, distinctive features of epithelioid cells and multinucleated giant cells in mature granulomas. M. marinum is found both intracellular, sequestered by the cellular aggregates, or extracellular, in necrotic (caseous) foci (Davis et al., 2002). TNF was found to be a key effector molecule required for the maintenance of granuloma integrity. Ablation of TNF signaling in mutant zebrafish embryos causes both accelerated intracellular bacterial growth and granuloma formation, followed by increased macrophage death and necrotic breakdown of granulomas with resultant exuberant growth of extracellular mycobacteria (Clay et al., 2008). Interestingly, granuloma formation is generally associated with accelerated bacterial proliferation, plateauing only after several weeks with the onset of adaptive immunity, indicating that nascent granulomas promote mycobacterial expansion (Volkman et al., 2004). Indeed, zebrafish studies have illustrated that granuloma-forming processes are mediated through mycobacterial virulence factors. By a mechanism requiring the mycobacterial RD1/ESX-1 secretion system, new macrophages are recruited to the granuloma via chemotactic signals, and phagocytose infected macrophages undergoing apoptosis, leading to rapid, iterative expansion of the number of infected macrophages and thereby to an increase in the bacterial burden (Davis and Ramakrishnan, 2009). The secretory protein ESAT-6 seems to play a central role in these chemotactic effects by inducing matrix metalloproteinase-9 (MMP-9) expression in epithelial cells surrounding the granuloma. In turn, MMP-9 is thought to enhance recruitment of macrophages to the growing granuloma. RD1/ESX-1-deficient M. marinum strains are still able to recruit macrophages to the infection site, but fail to elicit aggregation into granulomas and intercellular bacterial spread and thus presumably lack the ability to induce chemotactic signals required for the initiation of these processes (Volkman et al., 2004). Primary granulomas can seed new granulomas by the efflux of infected macrophages, constituting a means of disseminating infection (Davis and Ramakrishnan, 2009).
The Role of Neutrophils
Macrophages have been described as the predominant cell type phagocytosing M. marinum after microinjection into fluid-filled compartments, such as the blood or hindbrain ventricle. In contrast, M. marinum seems to evade contact with neutrophils at initial infection sites. Instead, neutrophils are subsequently recruited to the nascent granuloma in response to signals from dying infected macrophages within the granuloma. Neutrophils are able to phagocytose and rapidly kill the internalized mycobacteria through NADPH oxidase-dependent mechanisms (Yang et al., 2012). Interestingly, effective uptake of M. marinum by neutrophils has been observed after subcutaneous infection of zebrafish (Colucci-Guyon et al., 2011). Likewise, it was found in the zebrafish model of Escherichia coli infection that neutrophils efficiently engulf bacteria on tissue surfaces, but not in fluid environments (Le Guyader et al., 2008). The concept that the route of entry of the bacteria into the host may determine the role of neutrophils in infections may also be of relevance for human infectious diseases, particularly for the early phase of encounter with microbes (Colucci-Guyon et al., 2011).
From Initial Granuloma Formation to Mature Granulomas
The view that granulomas are primarily host-beneficial protective structures, is thus challenged by the described findings in the zebrafish embryo model of M. marinum infection. Results demonstrate that pathogenic mycobacteria have developed mechanisms to harness nascent host granulomas for their dissemination and proliferation (Ramakrishnan, 2013). Pathologically, granulomas are defined as organized collection of differentiated macrophages with characteristic morphology, such as epithelioid histiocytes and giant cells (Figure 2A). In addition to macrophages, mature tuberculous granulomas in humans are populated by many other cell types, including neutrophils, dendritic cells, B and T cells, natural killer cells, fibroblasts, and epithelial cells (Figure 2B). The role of these cells in granulomatous infectious diseases is yet to be fully elucidated. Another characteristic of certain tuberculous granulomas is the presence of regions of acellular debris referred to as caseous necrosis. This limited central necrosis is also found in M. marinum infection; mycobacteria can be located both within macrophages and within the central caseous region. Microbial virulence factors may influence the cellular composition of granulomas and the role of these structures played in either the containment, persistence, or dissemination of infections.
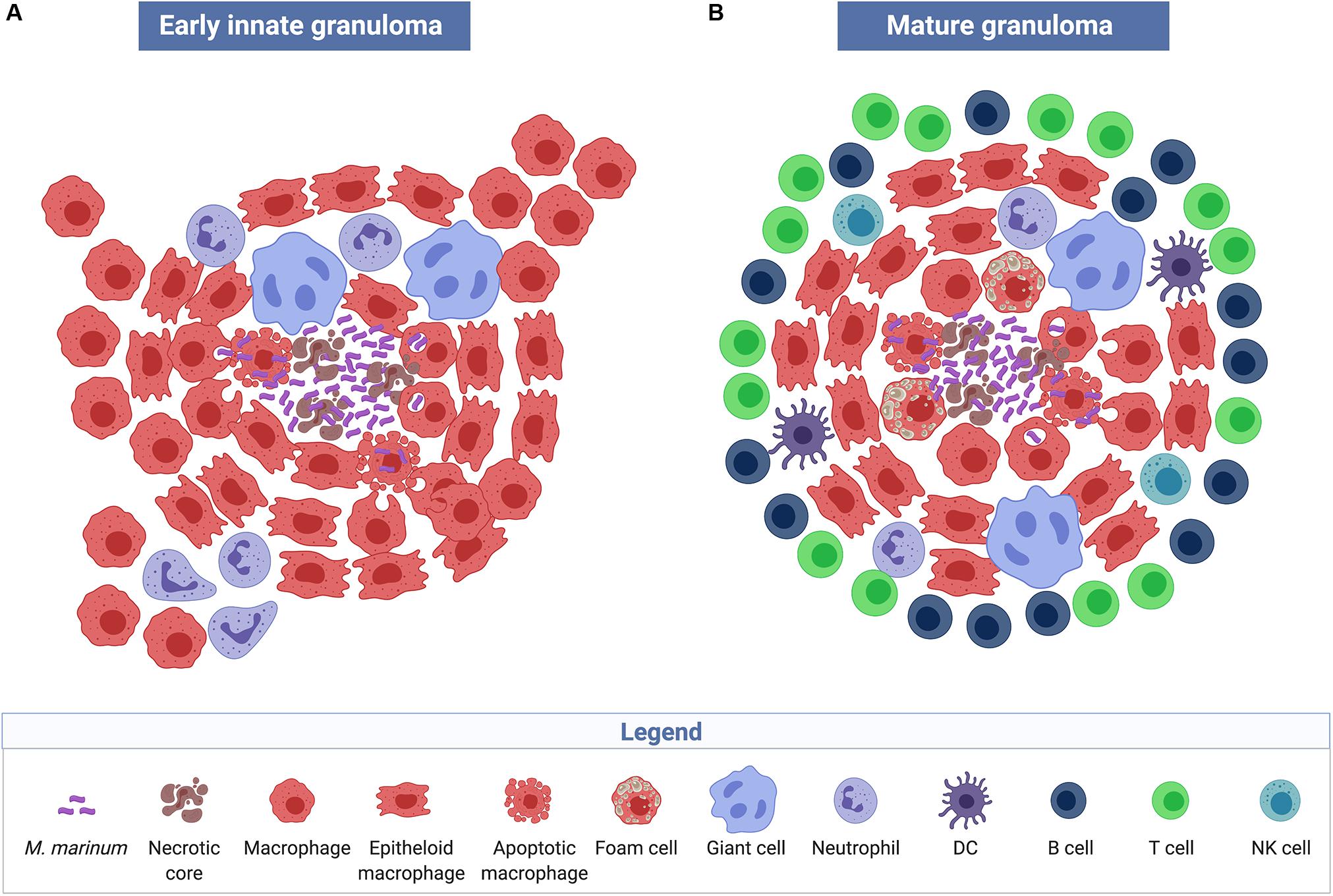
Figure 2. Structure and cellular composition of granulomas. Early innate M. marinum granulomas, as described in zebrafish embryos, are composed of compact, organized aggregates of macrophages that have either transformed into epithelioid cells with tightly interdigitated cell membranes or have fused to become multinucleated giant cells. Bacilli are found both extracellularly in the necrotic core of the granuloma and intracellularly within phagocytes (A). The characteristic mature tuberculous granuloma is populated by many other cell types such as neutrophils, dendritic cells, B and T cells, natural killer cells, and foam cells, which are macrophages with accumulated lipids (B) (created with biorender.com).
Innate Immune Mechanisms in M. ulcerans Infection
Entry Point: Skin
The site of M. ulcerans and M. marinum lesions is thought to also be the site of inoculation of the mycobacteria into the skin, as indicated by the vast majority of patients presenting with a single skin lesion on body parts that are likely exposed to the contaminated environment and/or, in the case of M. ulcerans, to potential vectors (Portaels et al., 1999; Johnson et al., 2007; Lavender et al., 2011). Definitive evidence for this assumption is however lacking. For both mycobacterial skin infections, the skin epithelium constitutes the first physical barrier. In healthy skin, stable homeostasis and barrier function are established by resident keratinocytes, dendritic cells, T cells, mast cells, fibroblasts, and macrophages together with the resident microbiome (Gallo and Nakatsuji, 2011). Contact with environmental reservoirs of M. ulcerans or M. marinum, and penetration of the bacilli into the subcutaneous tissue through trauma – in the case of M. ulcerans potentially including bites by contaminated insects acting as mechanical vectors (Wallace et al., 2017) – may be the most common mechanism of infection, although other mechanisms cannot be excluded. The outcome of an infection with both M. ulcerans and M. marinum may depend on the mode of transmission and the initial dose of inoculated bacteria. In a guinea pig infection model, it has been shown that BU can be produced by intra-dermal injection, but not through inoculation of M. ulcerans onto a superficial abrasion (Williamson et al., 2014). In advanced BU lesions, clusters of extracellular AFB are predominantly found in deep layers of the subcutaneous fat tissue (Ruf et al., 2016). This may reflect the location of the initial inoculum causing disease or the presence of a microenvironment in the necrotic tissue which is most favorable for the multiplication of M. ulcerans. In a case series of patients with M. marinum infection, boating or fishing were associated with invasive disease, whereas fish tank exposure was associated with cutaneous disease. This may be related to the mechanism of injury; boating and fishing injuries may involve deep puncture wounds from fish spines, fishhooks or other equipment, while fish tank injuries may involve more superficial exposures such as minor scrapes while cleaning or maintaining fish tanks (Johnson and Stout, 2015).
Once Inside the Host…
Once inside the host, the mycobacteria encounter innate immune cells expressing various PRRs. Based on a series of in vitro experiments it was postulated that keratinocyte TLRs may play a role in the innate immune response to M. ulcerans infections (Lee et al., 2009). The precise role (if any) of keratinocytes in the recognition of M. ulcerans and subsequent modulation of innate responses in the host is however unknown. If we assume that the mode of transmission of the mycobacteria allows for an encounter with these cells, keratinocyte TLRs, in response to sensing PAMPs expressed by microbes and DAMPs produced by the host, can initiate an inflammatory cascade including the release of inflammatory cytokines and host antimicrobial molecules. Early events in the innate immune response to skin injury include the recruitment of neutrophils via chemokines released by the keratinocytes. Later during the inflammatory cascade, macrophages are the predominant immune cell type (Coates et al., 2018). At this stage of the infection, the ability to produce mycolactone results in a fundamentally different interaction of the host with M. ulcerans as compared to M. marinum.
Insight Into Early Immune Defense Against M. ulcerans From Animal Models
To study immune responses setting in directly after infection with M. ulcerans, experimental infection models mostly involving mice (Mus musculus), but also other animals such as guinea pigs (Cavia porcellus) or pigs (Sus scrofa) have been developed (Bolz and Ruf, 2019). In the mouse model of BU, an immediate massive influx of neutrophils and to a lesser extent of monocytes/macrophages at the site of M. ulcerans intraperitoneal or intradermal injection is observed. One day after injection, bacilli are found within phagocytes and some of the inoculated bacilli are subsequently transported to the draining lymph nodes, where TH1-type cellular immune responses are initiated. Prompted by these findings, an early intracellular growth phase of M. ulcerans that induces inflammatory cellular responses has been postulated (Coutanceau et al., 2005; Oliveira et al., 2005; Torrado et al., 2007; Silva et al., 2009). As early as 24 h after injection of M. ulcerans into mouse footpads, lysis of infected phagocytes mediated by the production of mycolactone by M. ulcerans causes release of the bacteria into the extracellular space (Oliveira et al., 2005). In particular globi-like accumulations of bacilli released from phagocytes (Schütte et al., 2009; Ruf et al., 2011) may readily form a protective cloud of mycolactone and may represent starting points for the development of large extracellular clusters. During this second stage of the infection, bacteria can multiply predominantly extracellularly, as mycolactone appears to prevent infiltrating immune cells from reaching the mycobacteria (Ruf et al., 2017). M. ulcerans forms an extracellular matrix, which is rich in proteins, lipids and lipoglycans and is likely to play a role in the development of extracellular clusters (Marsollier et al., 2007). High concentrations of mycolactone in the lesion core cause apoptosis of both resident skin cells and infiltrating leukocytes. Chronic, necrotic lesions develop upon the invasion of the bacteria into healthy tissue and the progressive lateral destruction of subcutaneous tissue (Oliveira et al., 2005). Ischemia associated with vascular pathology may also contribute to the coagulative necrosis. In the infiltrate surrounding the necrotic core, intracellular bacilli can be detected, mainly in macrophages (Oliveira et al., 2005; Torrado et al., 2007). In the experimental mouse footpad infection model, dermal edema and footpad swelling become – dependent on the inoculation dose – evident a few weeks after infection. Necrosis extends to components of subcutaneous tissue, eventually resulting in extensive ulceration of the epidermis. That the continuous expansion of necrotic lesions is mainly mycolactone-mediated is illustrated in infections with mycolactone-negative M. ulcerans strains, which induce an initial acute neutrophilic response that gradually switches to a chronic mononuclear infiltrate devoid of massive necrosis (Oliveira et al., 2005).
The pig is widely used as a model in dermatological studies because pig and human skin share many morphological and physiological features. Pigs have been shown to develop single lesions at the site of injection of M. ulcerans into the skin, characterized by a central necrotic core containing large clumps of AFB surrounded by layers of neutrophilic debris, some intact neutrophils, and an outer belt of macrophages interspersed with T cells (Figures 3A–E). In contrast, lesions caused by mycolactone-deficient M. ulcerans strains present as multiple small central clusters of neutrophils and AFB with only limited necrosis, surrounded by a massive infiltration of macrophages interspersed with T-cells (Bolz et al., 2016b).
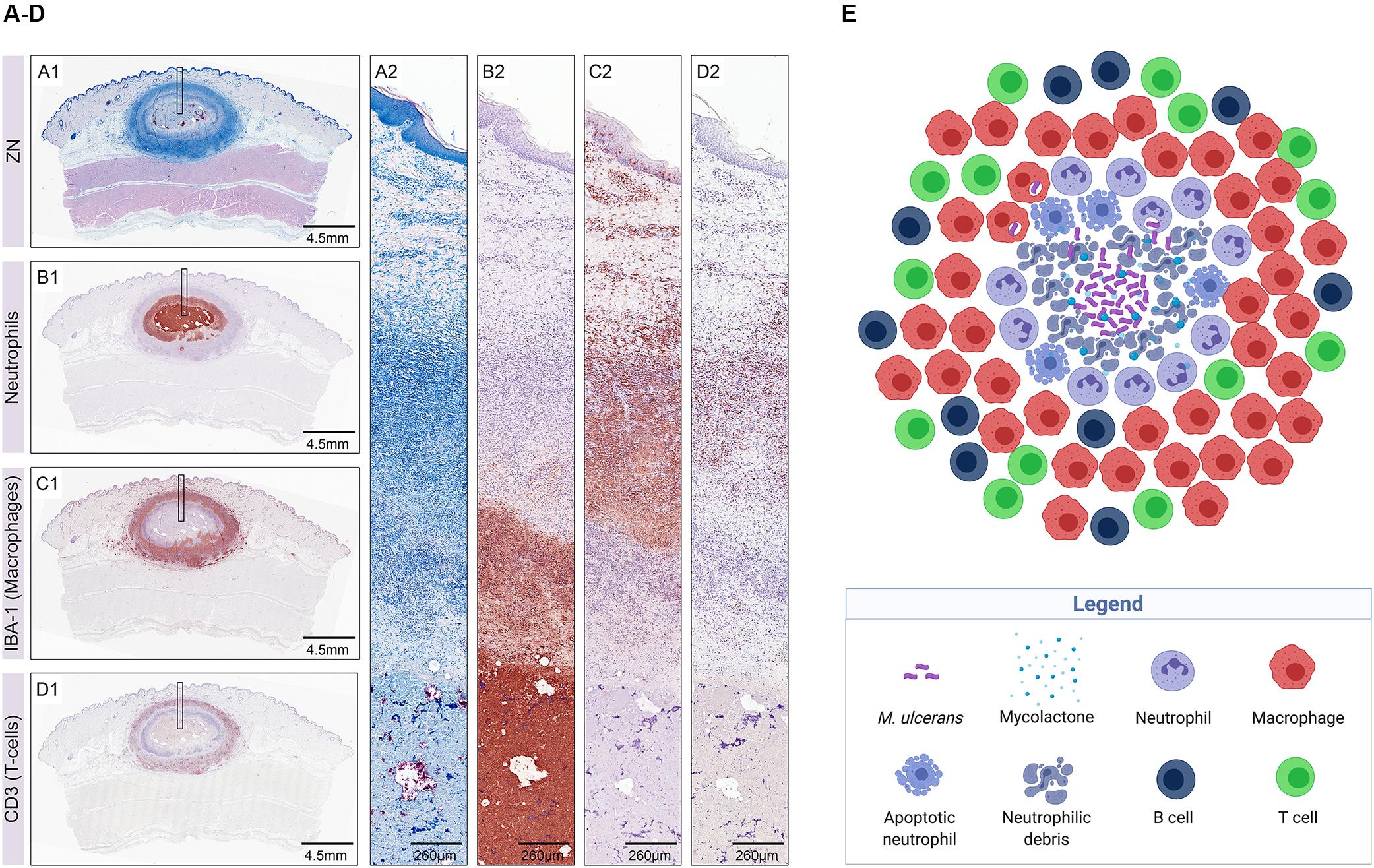
Figure 3. Early host immune response to M. ulcerans infection illustrated in histological sections of a nodular lesion 6 weeks after subcutaneous infection of pigs (A–D) (from Bolz et al., 2016b) and in a sketch depicting the cellular composition of the lesion (E) (created with biorender.com). Histopathological analysis of pig lesions shows a central necrotic core containing extracellular M. ulcerans bacilli (A1,A2; AFB stained with Ziehl–Neelsen staining) and neutrophilic debris (B1,B2). The necrotic core is surrounded by layers of neutrophils (B1,B2) and macrophages (C1,C2), heavily interspersed with T-cells (D1,D2).
Interestingly, both wild type and mutant M. ulcerans strains evoke the same sequential infiltration layers with neutrophils and neutrophilic debris in the necrotic lesion centers surrounded by macrophages and T cells (Bolz et al., 2016b), a composition which is in contrast to granulomatous lesions caused by M. marinum or M. tuberculosis, characterized by organized aggregates of mature macrophages.
Insight Into Early Immune Responses From the Study of Early Human BU Lesions
Buruli ulcer patients in rural, endemic areas of Africa typically report to health facilities at late stages of the disease and thus histopathological studies are mostly restricted to advanced ulcers. In a cohort of 12 BU patients from far north Queensland (Australia) presenting with early lesions, immunohistochemical analysis revealed an acellular, necrotic core containing the extracellularly multiplying AFB separated from intact tissue by a belt of infiltrating immune cells comprising clusters of CD20-positive B cells, CD3-positive T cells, neutrophils, and macrophages. Neutrophilic debris was found inside the lesion core and is indicative of a massive early neutrophil infiltration that was walled off by the cytotoxic actions of mycolactone (Figure 3E). Some AFB, located close to the infiltration belt, were detected intracellularly and may be signs of an expansion of the necrotic foci into healthy tissue (Ruf et al., 2017), a finding that was also reported in another study (Torrado et al., 2007). Although the pathogenesis of M. ulcerans infections in Australia and Africa seems comparable, it remains to be investigated if similar early immune responses are evoked in BU patients from Africa.
Immune Reconstitution After Antibiotic Therapy
Antibiotic treatment of BU patients results in a rapid onset of local cellular immune responses. Analysis of surgical specimens excised from patients after treatment with the standard antibiotic regimen for BU, revealed accumulation of infiltrating leukocytes in the BU lesions, presumably facilitated by decreasing concentrations of mycolactone associated with the suppression of the metabolic activity and finally with the killing of the bacilli. Already 4 weeks after start of antibiotic treatment AFB are primarily located within mononuclear phagocytes. Cellular infiltrates surrounding areas of coagulative necrosis display different levels of organization, including diffuse infiltrates present in all areas of connective and adipose tissue (Figure 4A) and, less frequently, organized epithelioid leukocyte accumulations located in deeper dermal tissue (Figure 4B) or dense lymphocyte aggregations in proximity to vessels (Figure 4C), reflecting a range of different functional activities required to clear the infection (Schütte et al., 2007).
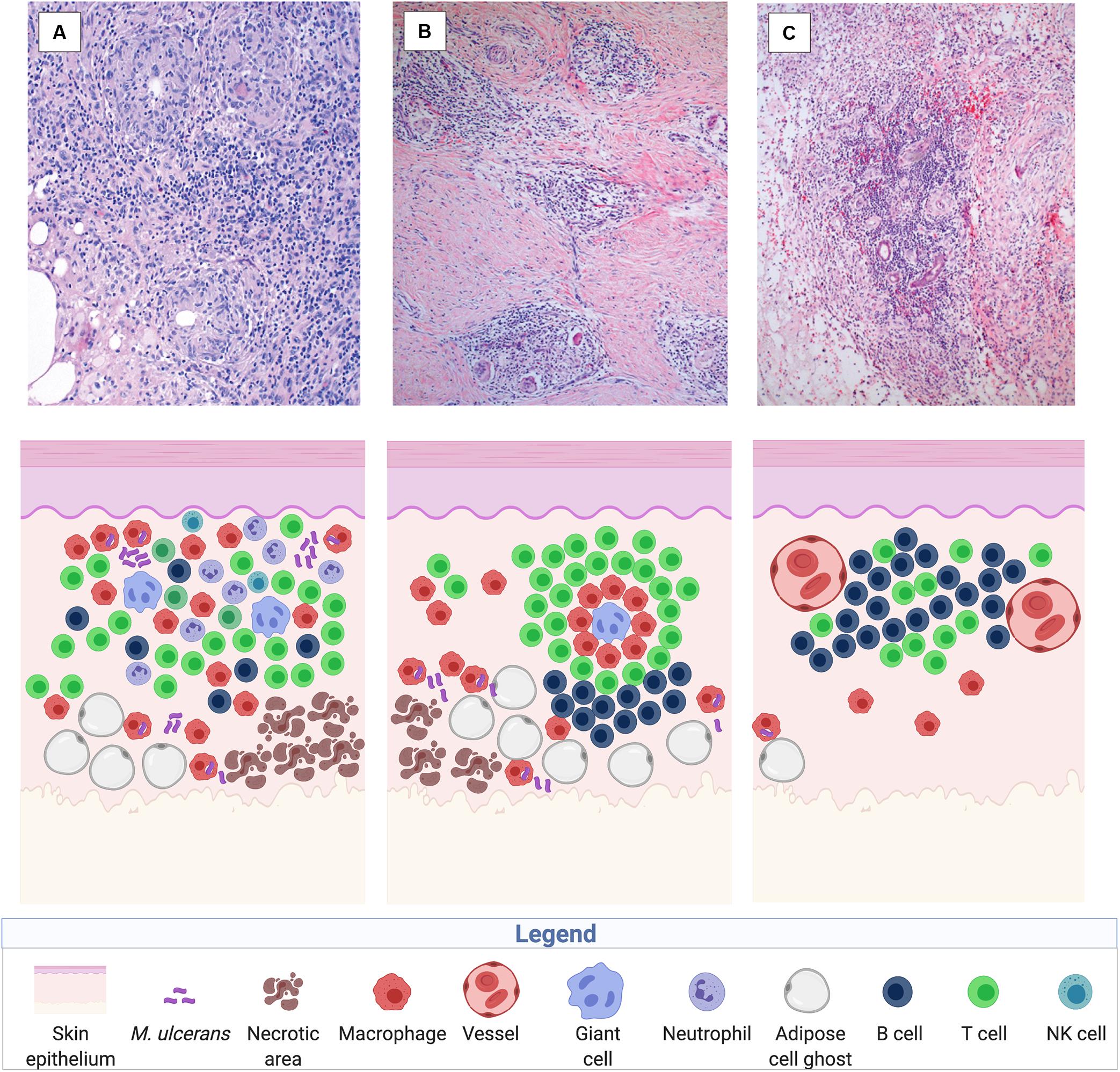
Figure 4. Immune reconstitution responses after chemotherapy illustrated in histological sections of a human BU lesion (from Schütte et al., 2007) and in a sketch of the participating immune components (created with biorender.com). Three main types of cellular infiltration have been observed in BU patients after treatment with the standard antibiotic regimen, including diffuse, heterogeneous cellular infiltration of the connective and adipose tissue (100× magnified) (A), granuloma-like structures in the connective tissue (40× magnified) (B), and follicle-like lymphocyte structures adjacent to vessels (40× magnified) (C).
Diverse Outcome of Infection
The early immune response may in many cases be capable of clearing an initial M. ulcerans inoculum. Spontaneous healing of BU lesions and serological evidence of exposure of healthy individuals to M. ulcerans indicate that the host immune system can contain infections with the pathogen, although mechanisms conferring protection are not entirely clear. Progression of lesions is highly diverse and not all non-ulcerative lesions ulcerate (Capela et al., 2015). One factor for the diverse outcome of M. ulcerans infections may be the inoculation dose. In the pig model of M. ulcerans infection, lower inoculation doses led to limited tissue destruction and eventually to the clearance of the bacteria (Bolz et al., 2014). Healing of BU lesions without specific treatment has been described for nodules, small ulcers, and long-standing ulcerative lesions (Revill et al., 1973; Marion et al., 2016a; O’Brien et al., 2019). How frequently BU lesions heal spontaneously is however difficult to assess, as many patients with early stages of the disease, that are usually indolent and non-systemic, may not report to health facilities. In a series of 545 BU patients diagnosed at a BU treatment center in Benin, 5% of the cases healed without specific treatment (Marion et al., 2016a). Whereas subcutaneous injection of M. ulcerans into BALB/c or C57BL/6 mice eventually leads to irreversible ulceration and tissue necrosis, ulcerative lesions developing after infection of FVB/N mice with M. ulcerans healed spontaneously despite persistent bacterial load. The healing process in FVB/N mice was associated with an infiltration of predominantly mononuclear cells, such as macrophages, dendritic cells, and neutrophils at the site of infection and an accumulation of myeloid cells in the draining lymph nodes, suggesting an important role for innate cellular immune defense mechanisms in protection (Marion et al., 2016b). In contrast to FVB/N mice, experimentally infected guinea pigs and pigs are able to entirely clear the M. ulcerans bacilli during the process of spontaneous healing (Bolz et al., 2014, 2016b; Silva-Gomes et al., 2015).
Children living in BU endemic areas of Africa seem to gradually develop resistance against BU with age, as indicated by a decline in the risk to develop the disease after a peak at an age of 12–14 years (Debacker et al., 2004, 2006; Bratschi et al., 2013). An increase in risk to develop BU in the elderly may be related to a deterioration of the relevant immune defense mechanisms. Prevalence of human immunodeficiency virus (HIV) infection in BU patients is significantly higher than in the local control population (Johnson et al., 2008; Christinet et al., 2014), indicating that TH1 cell responses are important enhancers of the innate cellular immune defense against M. ulcerans infection. Moreover, skin lesions in BU-HIV co-infected patients tend to be more severe and more often multifocal (Johnson et al., 2002; Komenan et al., 2013). Case control studies aiming at the identification of host genetic factors relevant for susceptibility to BU have focused so far primarily on polymorphisms in genes known to be relevant for intracellular mycobacterial infections (Stienstra et al., 2006; Capela et al., 2016; Bibert et al., 2017). Significant associations observed with susceptibility to BU include polymorphisms affecting the promoter activity of the IFN-γ gene, the inducible nitric oxide synthase gene iNOS and the natural resistance-associated macrophage gene SLC11A1 (NRAMP1). These results for the primarily extracellular M. ulcerans support the view that macrophages are crucially important for the early containment of M. ulcerans infections. Diversity of the response to human infection may thus be influenced not only by factors like inoculation dose, age, and nutritional status, but also by a complex constellation of genetic factors.
Conclusion
Despite common ancestry and the high degree of genetic relatedness of M. marinum and M. ulcerans, the role of the innate host immune system in immune defense against the two pathogens seems to differ considerably. Macrophages are a key component in the innate immune response to M. marinum infection, where they inadvertently play a dual role, both containing mycobacterial growth and providing an environment where the bacilli can persist (Ramakrishnan, 2012). Continuous recruitment of macrophages (and other immune cells), at least partly dependent on mechanisms of the RD1/ESX-1 secretion system, leads to a characteristic localized inflammatory response and granuloma formation. On the contrary, phagocytes seem to be only transiently inhabited by M. ulcerans in very early stages of the infection (Coutanceau et al., 2005). Histopathological analyses of human BU lesions and of those of experimentally infected animals indicate that neutrophils are the main infiltrating cell type in early stages of M. ulcerans infection, later complemented by macrophages (Bolz et al., 2016b; Ruf et al., 2017). The early dominance of neutrophils may be explained by the route of M. ulcerans infection (Le Guyader et al., 2008; Colucci-Guyon et al., 2011). After a postulated short intra-phagocyte stage of M. ulcerans, production of mycolactone enables a second, extracellular growth phase. Bacterial clusters are formed, which produce a protective cloud of mycolactone, walling off and killing infiltrating immune cells. A progressive invasion of healthy tissue and potentially of phagocytes at the periphery of the lesions leads to the characteristic chronic, necrotic course of the disease (Ruf et al., 2017). In BU patients under chemotherapy, killing of the bacteria and consequently the decrease in mycolactone concentrations allows for an immune reconstitution reaction (Schütte et al., 2007).
Due to relatively unspecific first signs of M. ulcerans infections, the often indolent course of the disease, and the limited access of affected populations to medical care, patients commonly seek treatment primarily in advanced disease stages, leaving patients in many cases with permanent disabilities. This and the highly focal occurrence of the disease makes vaccination of inhabitants of endemic areas the desired solution for BU control, particularly because the limited understanding of risk factors for BU and reservoirs/vectors of M. ulcerans have so far hindered other preventive measures. Lack of knowledge on protective host immune responses has however also complicated the design of a vaccine for BU. Intriguingly, there is striking paucity of humoral immune responses upon infection with M. ulcerans, revealed after experimental M. ulcerans infection in the BU mouse model (Bieri et al., 2016; Bolz et al., 2016a), and corroborated by reports that sera of only a minority of BU patients contain M. ulcerans-specific antibodies (Yeboah-Manu et al., 2012). These findings may be explained by the cytotoxic and immunosuppressive actions of mycolactone on immune cells. This and evidence pointing toward an early intracellular phase of M. ulcerans call for a robust engagement of innate immune cells to boost cell-mediated immunity, eliminating the phagocytosed mycobacteria before larger intra- and later on extra-cellular toxin-producing bacterial clusters can form. Efficient activation of T cells orchestrated by antigen presenting cells is crucial to enable in turn activation (CD4 T cells) of host cells, such as macrophages to kill invading pathogens or direct killing (CD8 T cells) of infected host cells. The specific types of mycobacterial T cell antigens conferring protective immunity are yet to be determined. In addition to adaptive T cells activated upon mycobacterial peptide antigens presented by major histocompatibility complex (MHC) molecules, innate-like T cells with low antigen receptor diversity, which recognize lipid antigens presented by cluster of differentiation 1 (CD1), have been described (Beckman et al., 1994). T cells recognizing mycobacterial glycolipid antigens, which share some biological properties with both adaptive and innate-like T cells were shown to confer protection to tuberculosis in animal models (Larrouy-Maumus et al., 2017; James and Seshadri, 2020). In the past, strategies for the development of vaccines have mainly been empirical, with limited understanding of the underlying immune mechanisms, using killed or live attenuated forms of the pathogens. Live-attenuated vaccines such as BCG in the case of M. tuberculosis seem to be suitable for intracellular pathogens as they induce a broad range of immune responses including strong CD8 T cell responses. Interestingly, it has recently been reported that a prolonged increase in antimicrobial function of innate immune cells can itself contribute to protection from reinfection. In this context it was shown that BCG is capable of inducing non-specific cross-protection against microbes, a phenomenon that has been associated with a memory-like response in innate immune cells (Netea et al., 2011, 2016; Koeken et al., 2019). Such “trained innate immune cells” display functional and epigenetic reprogramming, leading to increased production of cytokines and chemokines, and improved phagocytotic and killing activities. Indeed, BCG was shown to offer a short-lived protective effect against BU in the first year after vaccination but limited to no protection thereafter (The Uganda Buruli Group, 1969; Smith et al., 1976). Whether this observed effect is due to mechanisms of trained immunity or is based on the broad antigenic cross-reactivity between M. bovis and M. ulcerans is however unknown. Similar results were found in the mouse model of experimental M. ulcerans infection, where BCG was shown to induce an immune response transiently containing proliferation of the bacilli but ultimately failing to prevent the typical BU pathology (Fraga et al., 2012). Differences in the effectiveness of BCG vaccination in different mouse strains has been reported (Converse et al., 2011). Genetically engineered BCG developed as a vehicle for BU vaccines offered marginally improved protection in the mouse model of M. ulcerans infection (Hart et al., 2015; Hart and Lee, 2016).
Considering that successful toxoid vaccines such as those against diphtheria and tetanus exist, targeting mycolactone itself may be a promising approach for the development of a BU vaccine. The presence of pre-existing neutralizing antibodies against mycolactone may physically block toxin interactions with host cells and thus aid cellular immune responses to the pathogen. Immunization of mice with a carrier protein conjugate of a non-toxic, synthetic mycolactone derivative has enabled for the first time the generation of antibody responses against the poorly immunogenic, cytotoxic mycolactone. Intriguingly, mycolactone-specific immune sera and mycolactone-specific mouse monoclonal antibodies showed toxin neutralizing activity, preventing mammalian cell apoptosis in an in vitro assay (Dangy et al., 2016). A more holistic approach targeting protective immune responses against both mycolactone and against other protein and potentially also glycolipid antigens may improve protective efficacy. To increase immunogenicity by activating appropriate elements of the innate immune system such a vaccine may need to be formulated with adjuvants stimulating various PRRs on innate immune cells (Tima et al., 2016).
Better understanding of the exact innate and adaptive immune mechanisms leading to protection from BU will help in the development of new strategies for effective vaccine design.
Author Contributions
GP was invited to writing the review. KR and GP drafted and revised the manuscript.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Abdallah, A. M., Verboom, T., Weerdenburg, E. M., Gey van Pittius, N. C., Mahasha, P. W., Jiménez, C., et al. (2009). PPE and PE_PGRS proteins of Mycobacterium marinum are transported via the type VII secretion system ESX-5. Mol. Microbiol. 73, 329–340. doi: 10.1111/j.1365-2958.2009.06783.x
Aubry, A., Chosidow, O., Caumes, E., Robert, J., and Cambau, E. (2002). Sixty-three cases of Mycobacterium marinum infection: clinical features, treatment, and antibiotic susceptibility of causative isolates. Arch. Intern. Med. 162, 1746–1752. doi: 10.1001/archinte.162.15.1746
Baron, L., Paatero, A. O., Morel, J.-D., Impens, F., Guenin-Macé, L., Saint-Auret, S., et al. (2016). Mycolactone subverts immunity by selectively blocking the Sec61 translocon. J. Exp. Med. 213, 2885–2896. doi: 10.1084/jem.20160662
Beckman, E. M., Porcelli, S. A., Morita, C. T., Behar, S. M., Furlong, S. T., and Brenner, M. B. (1994). Recognition of a lipid antigen by CD1-restricted alpha beta+ T cells. Nature 372, 691–694. doi: 10.1038/372691a0
Beckman, E. N., Pankey, G. A., and Mcfarland, G. B. (1985). The histopathology of Mycobacterium marinum synovitis. Am. J. Clin. Pathol. 83, 457–462. doi: 10.1093/ajcp/83.4.457
Bibert, S., Bratschi, M. W., Aboagye, S. Y., Collinet, E., Scherr, N., Yeboah-Manu, D., et al. (2017). Susceptibility to Mycobacterium ulcerans disease (Buruli ulcer) is associated with IFNG and iNOS gene polymorphisms. Front. Microbiol. 8:1903. doi: 10.3389/fmicb.2017.01903
Bieri, R., Bolz, M., Ruf, M.-T., and Pluschke, G. (2016). Interferon-γ is a crucial activator of early host immune defense against Mycobacterium ulcerans infection in mice. PLoS Negl. Trop. Dis. 10:e0004450. doi: 10.1371/journal.pntd.0004450
Bieri, R., Scherr, N., Ruf, M.-T., Dangy, J.-P., Gersbach, P., Gehringer, M., et al. (2017). The macrolide toxin mycolactone promotes bim-dependent apoptosis in Buruli ulcer through inhibition of mTOR. ACS Chem. Biol. 12, 1297–1307. doi: 10.1021/acschembio.7b00053
Bolz, M., Bénard, A., Dreyer, A. M., Kerber, S., Vettiger, A., Oehlmann, W., et al. (2016a). Vaccination with the surface proteins MUL_2232 and MUL_3720 of Mycobacterium ulcerans induces antibodies but fails to provide protection against Buruli ulcer. PLoS Negl. Trop. Dis. 10:e0004431. doi: 10.1371/journal.pntd.0004431
Bolz, M., Ruggli, N., Borel, N., Pluschke, G., and Ruf, M.-T. (2016b). Local cellular immune responses and pathogenesis of Buruli ulcer lesions in the experimental Mycobacterium ulcerans pig infection model. PLoS Negl. Trop. Dis. 10:e0004678. doi: 10.1371/journal.pntd.0004678
Bolz, M., and Ruf, M.-T. (2019). “Buruli ulcer in animals and experimental infection models,” in Buruli Ulcer: Mycobacterium ulcerans Disease, eds G. Pluschke and K. Röltgen (Cham: Springer International Publishing), 159–181. doi: 10.1007/978-3-030-11114-4_9
Bolz, M., Ruggli, N., Ruf, M.-T., Ricklin, M. E., Zimmer, G., and Pluschke, G. (2014). Experimental infection of the pig with Mycobacterium ulcerans: a novel model for studying the pathogenesis of Buruli ulcer disease. PLoS Negl. Trop. Dis. 8:e2968. doi: 10.1371/journal.pntd.0002968
Bozzo, C., Tiberio, R., Graziola, F., Pertusi, G., Valente, G., Colombo, E., et al. (2010). A Mycobacterium ulcerans toxin, mycolactone, induces apoptosis in primary human keratinocytes and in HaCaT cells. Microbes Infect. 12, 1258–1263. doi: 10.1016/j.micinf.2010.08.005
Bratschi, M. W., Bolz, M., Minyem, J. C., Grize, L., Wantong, F. G., Kerber, S., et al. (2013). Geographic distribution, age pattern and sites of lesions in a cohort of Buruli ulcer patients from the mapé basin of Cameroon. PLoS Negl. Trop. Dis. 7:e2252. doi: 10.1371/journal.pntd.0002252
Broutin, V., Bañuls, A.-L., Aubry, A., Keck, N., Choisy, M., Bernardet, J.-F., et al. (2012). Genetic diversity and population structure of Mycobacterium marinum: new insights into host and environmental specificities. J. Clin. Microbiol. 50, 3627–3634.
Cambier, C. J., Takaki, K. K., Larson, R. P., Hernandez, R. E., Tobin, D. M., Urdahl, K. B., et al. (2014). Mycobacteria manipulate macrophage recruitment through coordinated use of membrane lipids. Nature 505, 218–222. doi: 10.1038/nature12799
Capela, C., Dossou, A. D., Silva-Gomes, R., Sopoh, G. E., Makoutode, M., Menino, J. F., et al. (2016). Genetic variation in autophagy-related genes influences the risk and phenotype of Buruli ulcer. PLoS Negl. Trop. Dis. 10:e0004671. doi: 10.1371/journal.pntd.0004671
Capela, C., Sopoh, G. E., Houezo, J. G., Fiodessihoué, R., Dossou, A. D., Costa, P., et al. (2015). Clinical epidemiology of Buruli ulcer from Benin (2005-2013): effect of time-delay to diagnosis on clinical forms and severe phenotypes. PLoS Negl. Trop. Dis. 9:e0004005. doi: 10.1371/journal.pntd.0004005
Christinet, V., Comte, E., Ciaffi, L., Odermatt, P., Serafini, M., Antierens, A., et al. (2014). Impact of human immunodeficiency virus on the severity of Buruli ulcer disease: results of a retrospective study in Cameroon. Open Forum Infect. Dis. 1:ofu021. doi: 10.1093/ofid/ofu021
Clark, H. F., and Shepard, C. C. (1963). Effect of environmental temperatures on infection with Mycobacterium marinum (BALNEI) of mice and a number of poikilothermic species. J. Bacteriol. 86, 1057–1069.
Clay, H., Davis, J. M., Beery, D., Huttenlocher, A., Lyons, S., and Ramakrishnan, L. (2007). Dichotomous role of the macrophage in early Mycobacterium marinum infection of the zebrafish. Cell Host Microbe 2, 29–39. doi: 10.1016/j.chom.2007.06.004
Clay, H., Volkman, H. E., and Ramakrishnan, L. (2008). Tumor necrosis factor signaling mediates resistance to mycobacteria by inhibiting bacterial growth and macrophage death. Immunity 29, 283–294. doi: 10.1016/j.immuni.2008.06.011
Coates, M., Blanchard, S., and MacLeod, A. S. (2018). Innate antimicrobial immunity in the skin: a protective barrier against bacteria, viruses, and fungi. PLoS Pathog. 14:e1007353. doi: 10.1371/journal.ppat.1007353
Cole, S. T., Brosch, R., Parkhill, J., Garnier, T., Churcher, C., Harris, D., et al. (1998). Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence. Nature 393, 537–544. doi: 10.1038/31159
Colucci-Guyon, E., Tinevez, J.-Y., Renshaw, S. A., and Herbomel, P. (2011). Strategies of professional phagocytes in vivo: unlike macrophages, neutrophils engulf only surface-associated microbes. J. Cell Sci. 124, 3053–3059. doi: 10.1242/jcs.082792
Converse, P. J., Almeida, D. V., Nuermberger, E. L., and Grosset, J. H. (2011). BCG-mediated protection against Mycobacterium ulcerans infection in the mouse. PLoS Negl. Trop. Dis. 5:e985. doi: 10.1371/journal.pntd.0000985
Coutanceau, E., Marsollier, L., Brosch, R., Perret, E., Goossens, P., Tanguy, M., et al. (2005). Modulation of the host immune response by a transient intracellular stage of Mycobacterium ulcerans: the contribution of endogenous mycolactone toxin. Cell. Microbiol. 7, 1187–1196. doi: 10.1111/j.1462-5822.2005.00546.x
Dangy, J.-P., Scherr, N., Gersbach, P., Hug, M. N., Bieri, R., Bomio, C., et al. (2016). Antibody-mediated neutralization of the exotoxin mycolactone, the main virulence factor produced by Mycobacterium ulcerans. PLoS Negl. Trop. Dis. 10:e0004808. doi: 10.1371/journal.pntd.0004808
Davis, J. M., Clay, H., Lewis, J. L., Ghori, N., Herbomel, P., and Ramakrishnan, L. (2002). Real-time visualization of mycobacterium-macrophage interactions leading to initiation of granuloma formation in zebrafish embryos. Immunity 17, 693–702.
Davis, J. M., and Ramakrishnan, L. (2009). The role of the granuloma in expansion and dissemination of early tuberculous infection. Cell 136, 37–49. doi: 10.1016/j.cell.2008.11.014
Debacker, M., Aguiar, J., Steunou, C., Zinsou, C., Meyers, W. M., Scott, J. T., et al. (2004). Mycobacterium ulcerans disease: role of age and gender in incidence and morbidity. Trop. Med. Int. Health 9, 1297–1304. doi: 10.1111/j.1365-3156.2004.01339.x
Debacker, M., Portaels, F., Aguiar, J., Steunou, C., Zinsou, C., Meyers, W., et al. (2006). Risk factors for Buruli ulcer, Benin. Emerg. Infect. Dis. 12, 1325–1331. doi: 10.3201/eid1209.050598
Diaz, D., Döbeli, H., Yeboah-Manu, D., Mensah-Quainoo, E., Friedlein, A., Soder, N., et al. (2006). Use of the immunodominant 18-kiloDalton small heat shock protein as a serological marker for exposure to Mycobacterium ulcerans. Clin. Vaccine Immunol. 13, 1314–1321.
Doig, K. D., Holt, K. E., Fyfe, J. A. M., Lavender, C. J., Eddyani, M., Portaels, F., et al. (2012). On the origin of Mycobacterium ulcerans, the causative agent of Buruli ulcer. BMC Genomics 13:258. doi: 10.1186/1471-2164-13-258
Evans, M. R., Etuaful, S. N., Amofah, G., Adjei, O., Lucas, S., and Wansbrough-Jones, M. H. (1999). Squamous cell carcinoma secondary to Buruli ulcer. Trans. R. Soc. Trop. Med. Hyg. 93, 63–64. doi: 10.1016/s0035-9203(99)90181-5
Fishbein, S., van Wyk, N., Warren, R. M., and Sampson, S. L. (2015). Phylogeny to function: PE/PPE protein evolution and impact on Mycobacterium tuberculosis pathogenicity. Mol. Microbiol. 96, 901–916. doi: 10.1111/mmi.12981
Fraga, A. G., Martins, T. G., Torrado, E., Huygen, K., Portaels, F., Silva, M. T., et al. (2012). Cellular immunity confers transient protection in experimental Buruli ulcer following BCG or mycolactone-negative Mycobacterium ulcerans vaccination. PLoS One 7:e33406. doi: 10.1371/journal.pone.0033406
Fyfe, J. A. M., Lavender, C. J., Handasyde, K. A., Legione, A. R., O’Brien, C. R., Stinear, T. P., et al. (2010). A major role for mammals in the ecology of Mycobacterium ulcerans. PLoS Negl. Trop. Dis. 4:e791. doi: 10.1371/journal.pntd.0000791
Gallo, R. L., and Nakatsuji, T. (2011). Microbial symbiosis with the innate immune defense system of the skin. J. Invest. Dermatol. 131, 1974–1980. doi: 10.1038/jid.2011.182
George, K. M., Chatterjee, D., Gunawardana, G., Welty, D., Hayman, J., Lee, R., et al. (1999). Mycolactone: a polyketide toxin from Mycobacterium ulcerans required for virulence. Science 283, 854–857. doi: 10.1126/science.283.5403.854
George, K. M., Pascopella, L., Welty, D. M., and Small, P. L. (2000). A Mycobacterium ulcerans toxin, mycolactone, causes apoptosis in guinea pig ulcers and tissue culture cells. Infect. Immun. 68, 877–883. doi: 10.1128/iai.68.2.877-883.2000
Gooding, T. M., Johnson, P. D., Campbell, D. E., Hayman, J. A., Hartland, E. L., Kemp, A. S., et al. (2001). Immune response to infection with Mycobacterium ulcerans. Infect. Immun. 69, 1704–1707. doi: 10.1128/IAI.69.3.1704-1707.2001
Griffith, D. E., Aksamit, T., Brown-Elliott, B. A., Catanzaro, A., Daley, C., Gordin, F., et al. (2007). An official ATS/IDSA statement: diagnosis, treatment, and prevention of nontuberculous mycobacterial diseases. Am. J. Respir. Crit. Care Med. 175, 367–416. doi: 10.1164/rccm.200604-571ST
Guenin-Mace, L., Baron, L., Chany, A.-C., Tresse, C., Saint-Auret, S., Jonsson, F., et al. (2015). Shaping mycolactone for therapeutic use against inflammatory disorders. Sci. Transl. Med. 7:289ra85. doi: 10.1126/scitranslmed.aab0458
Guenin-Macé, L., Ruf, M.-T., Pluschke, G., and Demangel, C. (2019). “Mycolactone: more than just a cytotoxin,” in Buruli Ulcer: Mycobacterium ulcerans Disease, eds G. Pluschke and K. Röltgen (Cham: Springer International Publishing), 117–134. doi: 10.1007/978-3-030-11114-4_7
Guenin-Macé, L., Veyron-Churlet, R., Thoulouze, M.-I., Romet-Lemonne, G., Hong, H., Leadlay, P. F., et al. (2013). Mycolactone activation of Wiskott-Aldrich syndrome proteins underpins Buruli ulcer formation. J. Clin. Invest. 123, 1501–1512. doi: 10.1172/JCI66576
Hall, B. S., Hill, K., McKenna, M., Ogbechi, J., High, S., Willis, A. E., et al. (2014). The pathogenic mechanism of the Mycobacterium ulcerans virulence factor, mycolactone, depends on blockade of protein translocation into the ER. PLoS Pathog. 10:e1004061. doi: 10.1371/journal.ppat.1004061
Hart, B. E., Hale, L. P., and Lee, S. (2015). Recombinant BCG expressing Mycobacterium ulcerans Ag85A imparts enhanced protection against experimental Buruli ulcer. PLoS Negl. Trop. Dis. 9:e0004046. doi: 10.1371/journal.pntd.0004046
Hart, B. E., and Lee, S. (2016). Overexpression of a Mycobacterium ulcerans Ag85B-EsxH fusion protein in recombinant BCG improves experimental Buruli ulcer vaccine efficacy. PLoS Negl. Trop. Dis. 10:e0005229. doi: 10.1371/journal.pntd.0005229
Huber, C. A., Ruf, M.-T., Pluschke, G., and Käser, M. (2008). Independent loss of immunogenic proteins in Mycobacterium ulcerans suggests immune evasion. Clin. Vaccine Immunol. 15, 598–606.
James, C. A., and Seshadri, C. (2020). T cell responses to mycobacterial glycolipids: on the spectrum of “innateness.” Front. Immunol. 11:170. doi: 10.3389/fimmu.2020.00170
Jernigan, J. A., and Farr, B. M. (2000). Incubation period and sources of exposure for cutaneous Mycobacterium marinum infection: case report and review of the literature. Clin. Infect. Dis. 31, 439–443. doi: 10.1086/313972
Johnson, M. G., and Stout, J. E. (2015). Twenty-eight cases of Mycobacterium marinum infection: retrospective case series and literature review. Infection 43, 655–662.
Johnson, P. D. R., Azuolas, J., Lavender, C. J., Wishart, E., Stinear, T. P., Hayman, J. A., et al. (2007). Mycobacterium ulcerans in mosquitoes captured during outbreak of Buruli ulcer, southeastern Australia. Emerg. Infect. Dis. 13, 1653–1660. doi: 10.3201/eid1311.061369
Johnson, R. C., Ifebe, D., Hans-Moevi, A., Kestens, L., Houessou, R., Guédénon, A., et al. (2002). Disseminated Mycobacterium ulcerans disease in an HIV-positive patient: a case study. AIDS 16, 1704–1705.
Johnson, R. C., Nackers, F., Glynn, J. R., de Biurrun Bakedano, E., Zinsou, C., Aguiar, J., et al. (2008). Association of HIV infection and Mycobacterium ulcerans disease in Benin. AIDS 22, 901–903. doi: 10.1097/QAD.0b013e3282f7690a
Käser, M., Rondini, S., Naegeli, M., Stinear, T., Portaels, F., Certa, U., et al. (2007). Evolution of two distinct phylogenetic lineages of the emerging human pathogen Mycobacterium ulcerans. BMC Evol. Biol. 7:177. doi: 10.1186/1471-2148-7-177
Kenu, E., Nyarko, K. M., Seefeld, L., Ganu, V., Käser, M., Lartey, M., et al. (2014). Risk factors for Buruli ulcer in Ghana-a case control study in the suhum-kraboa-coaltar and akuapem South districts of the eastern region. PLoS Negl. Trop. Dis. 8:e3279. doi: 10.1371/journal.pntd.0003279
Koeken, V. A. C. M., Verrall, A. J., Netea, M. G., Hill, P. C., and van Crevel, R. (2019). Trained innate immunity and resistance to Mycobacterium tuberculosis infection. Clin. Microbiol. Infect. 25, 1468–1472. doi: 10.1016/j.cmi.2019.02.015
Komenan, K., Elidjé, E. J., Ildevert, G. P., Yao, K. I., Kanga, K., Kouamé, K. A., et al. (2013). Multifocal Buruli ulcer associated with secondary infection in HIV positive patient. Case Rep. Med. 2013:348628. doi: 10.1155/2013/348628
Phillips, R. O., Robert, J., Abass, K. M., Thompson, W., Sarfo, F. S., Wilson, T., (2020). Rifampicin and clarithromycin (extended release) versus rifampicin and streptomycin for limited Buruli ulcer lesions: a randomised, open-label, non-inferiority phase 3 trial. Lancet 395, 1259–1267. doi: 10.1016/S0140-6736(20)30047-7
Larrouy-Maumus, G., Layre, E., Clark, S., Prandi, J., Rayner, E., Lepore, M., et al. (2017). Protective efficacy of a lipid antigen vaccine in a guinea pig model of tuberculosis. Vaccine 35, 1395–1402. doi: 10.1016/j.vaccine.2017.01.079
Lavender, C. J., Fyfe, J. A. M., Azuolas, J., Brown, K., Evans, R. N., Ray, L. R., et al. (2011). Risk of Buruli ulcer and detection of Mycobacterium ulcerans in mosquitoes in southeastern Australia. PLoS Negl. Trop. Dis. 5:e1305. doi: 10.1371/journal.pntd.0001305
Le Guyader, D., Redd, M. J., Colucci-Guyon, E., Murayama, E., Kissa, K., Briolat, V., et al. (2008). Origins and unconventional behavior of neutrophils in developing zebrafish. Blood 111, 132–141.
Lee, H.-M., Shin, D.-M., Choi, D.-K., Lee, Z.-W., Kim, K.-H., Yuk, J.-M., et al. (2009). Innate immune responses to Mycobacterium ulcerans via toll-like receptors and dectin-1 in human keratinocytes. Cell. Microbiol. 11, 678–692. doi: 10.1111/j.1462-5822.2009.01285.x
Loftus, M. J., Trubiano, J. A., Tay, E. L., Lavender, C. J., Globan, M., Fyfe, J. A. M., et al. (2018). The incubation period of Buruli ulcer (Mycobacterium ulcerans infection) in Victoria, Australia - remains similar despite changing geographic distribution of disease. PLoS Negl. Trop. Dis. 12:e0006323. doi: 10.1371/journal.pntd.0006323
Maman, I., Tchacondo, T., Kere, A. B., Piten, E., Beissner, M., Kobara, Y., et al. (2018). Risk factors for Mycobacterium ulcerans infection (Buruli ulcer) in Togo - a case-control study in Zio and Yoto districts of the maritime region. BMC Infect. Dis. 18:48. doi: 10.1186/s12879-018-2958-3
Marion, E., Chauty, A., Kempf, M., Le Corre, Y., Delneste, Y., Croue, A., et al. (2016a). Clinical features of spontaneous partial healing during Mycobacterium ulcerans infection. Open Forum Infect. Dis. 3:ofw013. doi: 10.1093/ofid/ofw013
Marion, E., Jarry, U., Cano, C., Savary, C., Beauvillain, C., Robbe-Saule, M., et al. (2016b). FVB/N mice spontaneously heal ulcerative lesions induced by Mycobacterium ulcerans and switch M. ulcerans into a low mycolactone producer. J. Immunol. 196, 2690–2698. doi: 10.4049/jimmunol.1502194
Marion, E., Song, O.-R., Christophe, T., Babonneau, J., Fenistein, D., Eyer, J., et al. (2014). Mycobacterial toxin induces analgesia in Buruli ulcer by targeting the angiotensin pathways. Cell 157, 1565–1576. doi: 10.1016/j.cell.2014.04.040
Marsollier, L., Brodin, P., Jackson, M., Korduláková, J., Tafelmeyer, P., Carbonnelle, E., et al. (2007). Impact of Mycobacterium ulcerans biofilm on transmissibility to ecological niches and Buruli ulcer pathogenesis. PLoS Pathog. 3:e62. doi: 10.1371/journal.ppat.0030062
Marsollier, L., Stinear, T., Aubry, J., Saint André, J. P., Robert, R., Legras, P., et al. (2004). Aquatic plants stimulate the growth of and biofilm formation by Mycobacterium ulcerans in axenic culture and harbor these bacteria in the environment. Appl. Environ. Microbiol. 70, 1097–1103. doi: 10.1128/aem.70.2.1097-1103.2004
Netea, M. G., Joosten, L. A. B., Latz, E., Mills, K. H. G., Natoli, G., Stunnenberg, H. G., et al. (2016). Trained immunity: a program of innate immune memory in health and disease. Science 352:aaf1098. doi: 10.1126/science.aaf1098
Netea, M. G., Quintin, J., and van der Meer, J. W. M. (2011). Trained immunity: a memory for innate host defense. Cell Host Microbe 9, 355–361. doi: 10.1016/j.chom.2011.04.006
N’krumah, R. T. A. S., Koné, B., Cissé, G., Tanner, M., Utzinger, J., Pluschke, G., et al. (2017). Characteristics and epidemiological profile of Buruli ulcer in the district of Tiassalé, south Côte d’Ivoire. Acta Trop. 175, 138–144. doi: 10.1016/j.actatropica.2016.12.023
O’Brien, D. P., Murrie, A., Meggyesy, P., Priestley, J., Rajcoomar, A., and Athan, E. (2019). Spontaneous healing of Mycobacterium ulcerans disease in Australian patients. PLoS Negl. Trop. Dis. 13:e0007178. doi: 10.1371/journal.pntd.0007178
Ogbechi, J., Ruf, M.-T., Hall, B. S., Bodman-Smith, K., Vogel, M., Wu, H.-L., et al. (2015). Mycolactone-dependent depletion of endothelial cell thrombomodulin is strongly associated with fibrin deposition in Buruli ulcer lesions. PLoS Pathog. 11:e1005011. doi: 10.1371/journal.ppat.1005011
Oliveira, M. S., Fraga, A. G., Torrado, E., Castro, A. G., Pereira, J. P., Filho, A. L., et al. (2005). Infection with Mycobacterium ulcerans induces persistent inflammatory responses in mice. Infect. Immun. 73, 6299–6310. doi: 10.1128/IAI.73.10.6299-6310.2005
Petrini, B. (2006). Mycobacterium marinum: ubiquitous agent of waterborne granulomatous skin infections. Eur. J. Clin. Microbiol. Infect. Dis. 25, 609–613.
Phillips, R., Sarfo, F. S., Guenin-Macé, L., Decalf, J., Wansbrough-Jones, M., Albert, M. L., et al. (2009). Immunosuppressive signature of cutaneous Mycobacterium ulcerans infection in the peripheral blood of patients with Buruli ulcer disease. J. Infect. Dis. 200, 1675–1684. doi: 10.1086/646615
Pidot, S. J., Asiedu, K., Käser, M., Fyfe, J. A. M., and Stinear, T. P. (2010). Mycobacterium ulcerans and other mycolactone-producing mycobacteria should be considered a single species. PLoS Negl. Trop. Dis. 4:e663. doi: 10.1371/journal.pntd.0000663
Pluschke, G., and Röltgen, K. (eds). (2019). Buruli Ulcer: Mycobacterium ulcerans Disease. Cham: Springer International Publishing.
Portaels, F., Elsen, P., Guimaraes-Peres, A., Fonteyne, P.-A., and Meyers, W. M. (1999). Insects in the transmission of Mycobacterium ulcerans infection. Lancet 353:986.
Raghunathan, P. L., Whitney, E. A. S., Asamoa, K., Stienstra, Y., Taylor, T. H., Amofah, G. K., et al. (2005). Risk factors for Buruli ulcer disease (Mycobacterium ulcerans infection): results from a case-control study in Ghana. Clin. Infect. Dis. 40, 1445–1453. doi: 10.1086/429623
Ramakrishnan, L. (2012). Revisiting the role of the granuloma in tuberculosis. Nat. Rev. Immunol. 12, 352–366. doi: 10.1038/nri3211
Ramakrishnan, L. (2013). “Looking within the zebrafish to understand the tuberculous granuloma,” in The New Paradigm of Immunity to Tuberculosis Advances in Experimental Medicine and Biology, ed. M. Divangahi (New York, NY: Springer), 251–266. doi: 10.1007/978-1-4614-6111-1_13
Ramakrishnan, L., Federspiel, N. A., and Falkow, S. (2000). Granuloma-specific expression of mycobacterium virulence proteins from the glycine-rich PE-PGRS family. Science 288, 1436–1439. doi: 10.1126/science.288.5470.1436
Ranger, B. S., Mahrous, E. A., Mosi, L., Adusumilli, S., Lee, R. E., Colorni, A., et al. (2006). Globally distributed mycobacterial fish pathogens produce a novel plasmid-encoded toxic macrolide, mycolactone F. Infect. Immun. 74, 6037–6045.
Revill, W. D., Morrow, R. H., Pike, M. C., and Ateng, J. (1973). A controlled trial of the treatment of Mycobacterium ulcerans infection with clofazimine. Lancet 2, 873–877.
Rhodes, M. W., Kator, H., McNabb, A., Deshayes, C., Reyrat, J.-M., Brown-Elliott, B. A., et al. (2005). Mycobacterium pseudoshottsii sp. nov., a slowly growing chromogenic species isolated from Chesapeake Bay striped bass (Morone saxatilis). Int. J. Syst. Evol. Microbiol. 55, 1139–1147.
Röltgen, K., and Pluschke, G. (2015). Mycobacterium ulcerans disease (Buruli ulcer): potential reservoirs and vectors. Curr. Clin. Microbiol. Rep. 2, 35–43.
Ruf, M.-T., Bolz, M., Vogel, M., Bayi, P. F., Bratschi, M. W., Sopho, G. E., et al. (2016). Spatial distribution of Mycobacterium ulcerans in Buruli ulcer lesions: implications for laboratory diagnosis. PLoS Negl. Trop. Dis. 10:e0004767. doi: 10.1371/journal.pntd.0004767
Ruf, M.-T., Chauty, A., Adeye, A., Ardant, M.-F., Koussemou, H., Johnson, R. C., et al. (2011). Secondary Buruli ulcer skin lesions emerging several months after completion of chemotherapy: paradoxical reaction or evidence for immune protection? PLoS Negl. Trop. Dis. 5:e1252. doi: 10.1371/journal.pntd.0001252
Ruf, M.-T., Steffen, C., Bolz, M., Schmid, P., and Pluschke, G. (2017). Infiltrating leukocytes surround early Buruli ulcer lesions, but are unable to reach the mycolactone producing mycobacteria. Virulence 8, 1918–1926. doi: 10.1080/21505594.2017.1370530
Sarfo, F. S., Converse, P. J., Almeida, D. V., Zhang, J., Robinson, C., Wansbrough-Jones, M., et al. (2013). Microbiological, histological, immunological, and toxin response to antibiotic treatment in the mouse model of Mycobacterium ulcerans disease. PLoS Negl. Trop. Dis. 7:e2101. doi: 10.1371/journal.pntd.0002101
Scherr, N., Gersbach, P., Dangy, J.-P., Bomio, C., Li, J., Altmann, K.-H., et al. (2013). Structure-activity relationship studies on the macrolide exotoxin mycolactone of Mycobacterium ulcerans. PLoS Negl. Trop. Dis. 7:e2143. doi: 10.1371/journal.pntd.0002143
Schütte, D., Um-Boock, A., Mensah-Quainoo, E., Itin, P., Schmid, P., and Pluschke, G. (2007). Development of highly organized lymphoid structures in Buruli ulcer lesions after treatment with Rifampicin and streptomycin. PLoS Negl. Trop. Dis. 1:e2. doi: 10.1371/journal.pntd.0000002
Schütte, D., Umboock, A., and Pluschke, G. (2009). Phagocytosis of Mycobacterium ulcerans in the course of Rifampicin and streptomycin chemotherapy in Buruli ulcer lesions. Br. J. Dermatol. 160, 273–283. doi: 10.1111/j.1365-2133.2008.08879.x
Shah, S., Cannon, J. R., Fenselau, C., and Briken, V. (2015). A duplicated ESAT-6 region of ESX-5 is involved in protein export and virulence of mycobacteria. Infect. Immun. 83, 4349–4361.
Sia, T. Y., Taimur, S., Blau, D. M., Lambe, J., Ackelsberg, J., Yacisin, K., et al. (2016). Clinical and pathological evaluation of Mycobacterium marinum group skin infections associated with fish markets in New York city. Clin. Infect. Dis. 62, 590–595. doi: 10.1093/cid/civ937
Silva, M. T., Portaels, F., and Pedrosa, J. (2009). Pathogenetic mechanisms of the intracellular parasite Mycobacterium ulcerans leading to Buruli ulcer. Lancet Infect. Dis. 9, 699–710.
Silva-Gomes, R., Marcq, E., Trigo, G., Gonçalves, C. M., Longatto-Filho, A., Castro, A. G., et al. (2015). Spontaneous healing of Mycobacterium ulcerans lesions in the guinea pig model. PLoS Negl. Trop. Dis. 9:e0004265. doi: 10.1371/journal.pntd.0004265
Smith, P. G., Revill, W. D., Lukwago, E., and Rykushin, Y. P. (1976). The protective effect of BCG against Mycobacterium ulcerans disease: a controlled trial in an endemic area of Uganda. Trans. R. Soc. Trop. Med. Hyg. 70, 449–457. doi: 10.1016/0035-9203(76)90128-0
Stienstra, Y., Werf, T. S., van der, Oosterom, E., Nolte, I. M., Graaf, W. T. A., et al. (2006). Susceptibility to Buruli ulcer is associated with the SLC11A1 (NRAMP1) D543N polymorphism. Genes Immun. 7, 185–189. doi: 10.1038/sj.gene.6364281
Stinear, T. P., Seemann, T., Harrison, P. F., Jenkin, G. A., Davies, J. K., Johnson, P. D. R., et al. (2008). Insights from the complete genome sequence of Mycobacterium marinum on the evolution of Mycobacterium tuberculosis. Genome Res. 18, 729–741. doi: 10.1101/gr.075069.107
Stinear, T. P., Seemann, T., Pidot, S., Frigui, W., Reysset, G., Garnier, T., et al. (2007). Reductive evolution and niche adaptation inferred from the genome of Mycobacterium ulcerans, the causative agent of Buruli ulcer. Genome Res. 17, 192–200. doi: 10.1101/gr.5942807
Stragier, P., Hermans, K., Stinear, T., and Portaels, F. (2008). First report of a mycolactone-producing mycobacterium infection in fish agriculture in Belgium. FEMS Microbiol. Lett. 286, 93–95. doi: 10.1111/j.1574-6968.2008.01264.x
Swaim, L. E., Connolly, L. E., Volkman, H. E., Humbert, O., Born, D. E., and Ramakrishnan, L. (2006). Mycobacterium marinum infection of adult zebrafish causes caseating granulomatous tuberculosis and is moderated by adaptive immunity. Infect. Immun. 74, 6108–6117.
The Uganda Buruli Group (1969). BCG vaccination against Mycobacterium ulcerans infection (Buruli ulcer): first results of a trial in Uganda. Lancet 293, 111–115.
Tima, H. G., Huygen, K., and Romano, M. (2016). Innate signaling by mycobacterial cell wall components and relevance for development of adjuvants for subunit vaccines. Expert Rev. Vaccines 15, 1409–1420. doi: 10.1080/14760584.2016.1187067
Tiwari, B. M., Kannan, N., Vemu, L., and Raghunand, T. R. (2012). The Mycobacterium tuberculosis PE proteins Rv0285 and Rv1386 modulate innate immunity and mediate bacillary survival in macrophages. PLoS One 7:e51686. doi: 10.1371/journal.pone.0051686
Tobin, D. M., and Ramakrishnan, L. (2008). Comparative pathogenesis of Mycobacterium marinum and Mycobacterium tuberculosis. Cell. Microbiol. 10, 1027–1039. doi: 10.1111/j.1462-5822.2008.01133.x
Torrado, E., Fraga, A. G., Castro, A. G., Stragier, P., Meyers, W. M., Portaels, F., et al. (2007). Evidence for an intramacrophage growth phase of Mycobacterium ulcerans. Infect. Immun. 75, 977–987.
Travis, W. D., Travis, L. B., Roberts, G. D., Su, D. W., and Weiland, L. W. (1985). The histopathologic spectrum in Mycobacterium marinum infection. Arch. Pathol. Lab. Med. 109, 1109–1113.
Trott, K. A., Stacy, B. A., Lifland, B. D., Diggs, H. E., Harland, R. M., Khokha, M. K., et al. (2004). Characterization of a Mycobacterium ulcerans-like infection in a colony of African tropical clawed frogs (Xenopus tropicalis). Comp. Med. 54, 309–317.
van der Sar, A. M., Abdallah, A. M., Sparrius, M., Reinders, E., Vandenbroucke-Grauls, C. M. J. E., and Bitter, W. (2004). Mycobacterium marinum strains can be divided into two distinct types based on genetic diversity and virulence. Infect. Immun. 72, 6306–6312. doi: 10.1128/IAI.72.11.6306-6312.2004
Volkman, H. E., Clay, H., Beery, D., Chang, J. C. W., Sherman, D. R., and Ramakrishnan, L. (2004). Tuberculous granuloma formation is enhanced by a mycobacterium virulence determinant. PLoS Biol. 2:e367. doi: 10.1371/journal.pbio.0020367
Wallace, J. R., Mangas, K. M., Porter, J. L., Marcsisin, R., Pidot, S. J., Howden, B., et al. (2017). Mycobacterium ulcerans low infectious dose and mechanical transmission support insect bites and puncturing injuries in the spread of Buruli ulcer. PLoS Negl. Trop. Dis. 11:e0005553. doi: 10.1371/journal.pntd.0005553
Westerfield, M. (2000). The Zebrafish Book: A Guide for the Laboratory Use of Zebrafish (Danio rerio). Eugene, OR: University of Oregon Press.
Williamson, H. R., Mosi, L., Donnell, R., Aqqad, M., Merritt, R. W., and Small, P. L. C. (2014). Mycobacterium ulcerans fails to infect through skin abrasions in a guinea pig infection model: implications for transmission. PLoS Negl. Trop. Dis. 8:e2770. doi: 10.1371/journal.pntd.0002770
World Health Organization, Global Buruli Ulcer Initiative (2004). Provisional Guidance on the Role of Specific Antibiotics in the Management of Mycobacterium ulcerans Disease (Buruli ulcer). Geneva: World Health Organization.
Yang, C.-T., Cambier, C. J., Davis, J. M., Hall, C. J., Crosier, P. S., and Ramakrishnan, L. (2012). Neutrophils exert protection in the early tuberculous granuloma by oxidative killing of mycobacteria phagocytosed from infected macrophages. Cell Host Microbe 12, 301–312. doi: 10.1016/j.chom.2012.07.009
Yeboah-Manu, D., Röltgen, K., Opare, W., Asan-Ampah, K., Quenin-Fosu, K., Asante-Poku, A., et al. (2012). Sero-epidemiology as a tool to screen populations for exposure to Mycobacterium ulcerans. PLoS Negl. Trop. Dis. 6:e1460. doi: 10.1371/journal.pntd.0001460
Keywords: Mycobacterium ulcerans, skin neglected tropical disease, mycolactone, Mycobacterium marinum, zebrafish
Citation: Röltgen K and Pluschke G (2020) Buruli ulcer: The Efficacy of Innate Immune Defense May Be a Key Determinant for the Outcome of Infection With Mycobacterium ulcerans. Front. Microbiol. 11:1018. doi: 10.3389/fmicb.2020.01018
Received: 14 January 2020; Accepted: 24 April 2020;
Published: 25 May 2020.
Edited by:
Malcolm Scott Duthie, HDT Bio Corp, United StatesReviewed by:
António Gil Castro, University of Minho, PortugalMark H. Wansbrough-Jones, St George’s, University of London, United Kingdom
Paul Converse, Johns Hopkins University, United States
Copyright © 2020 Röltgen and Pluschke. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Katharina Röltgen, roeltgen@stanford.edu