 Priscila Gonçalves Ferreira1
Priscila Gonçalves Ferreira1 Blanka Tesla2
Blanka Tesla2 Elvira Cynthia Alves Horácio3,4
Elvira Cynthia Alves Horácio3,4 Laila Alves Nahum3,4,5
Laila Alves Nahum3,4,5 Melinda Ann Brindley2,6,7*
Melinda Ann Brindley2,6,7* Tiago Antônio de Oliveira Mendes1*
Tiago Antônio de Oliveira Mendes1* Courtney Cuinn Murdock2,7,8,9,10,11,12*
Courtney Cuinn Murdock2,7,8,9,10,11,12*- 1Department of Biochemistry and Molecular Biology, Universidade Federal de Viçosa, Viçosa, Brazil
- 2Department of Infectious Diseases, College of Veterinary Medicine, University of Georgia, Athens, GA, United States
- 3René Rachou Institute, Oswaldo Cruz Foundation, Belo Horizonte, Brazil
- 4Department of Genetics, Ecology and Evolution, Institute of Biological Sciences, Federal University of Minas Gerais, Belo Horizonte, Brazil
- 5Promove College of Technology, Belo Horizonte, Brazil
- 6Department of Population Health, College of Veterinary Medicine, University of Georgia, Athens, GA, United States
- 7Center for Vaccines and Immunology, University of Georgia, Athens, GA, United States
- 8Odum School of Ecology, University of Georgia, Athens, GA, United States
- 9Center for the Ecology of Infectious Diseases, University of Georgia, Athens, GA, United States
- 10Center for Emerging and Global Tropical Diseases, University of Georgia, Athens, GA, United States
- 11River Basin Center, University of Georgia, Athens, GA, United States
- 12Department of Entomology, College of Agriculture and Life Sciences, Cornell University, Ithaca, NY, United States
Vector-borne flaviviruses are emerging threats to human health. For successful transmission, the virus needs to efficiently enter mosquito cells and replicate within and escape several tissue barriers while mosquitoes elicit major transcriptional responses to flavivirus infection. This process will be affected not only by the specific mosquito-pathogen pairing but also by variation in key environmental variables such as temperature. Thus far, few studies have examined the molecular responses triggered by temperature and how these responses modify infection outcomes, despite substantial evidence showing strong relationships between temperature and transmission in a diversity of systems. To define the host transcriptional changes associated with temperature variation during the early infection process, we compared the transcriptome of mosquito midgut samples from mosquitoes exposed to Zika virus (ZIKV) and non-exposed mosquitoes housed at three different temperatures (20, 28, and 36°C). While the high-temperature samples did not show significant changes from those with standard rearing conditions (28°C) 48 h post-exposure, the transcriptome profile of mosquitoes housed at 20°C was dramatically different. The expression of genes most altered by the cooler temperature involved aspects of blood-meal digestion, ROS metabolism, and mosquito innate immunity. Further, we did not find significant differences in the viral RNA copy number between 24 and 48 h post-exposure at 20°C, suggesting that ZIKV replication is limited by cold-induced changes to the mosquito midgut environment. In ZIKV-exposed mosquitoes, vitellogenin, a lipid carrier protein, was most up-regulated at 20°C. Our results provide a deeper understanding of the temperature-triggered transcriptional changes in Aedes aegypti and can be used to further define the molecular mechanisms driven by environmental temperature variation.
Introduction
Over the past three decades, there have been significant advances in our understanding of the physiological and molecular interactions between pathogens and mosquito vectors (Bartholomay and Michel, 2018; Kumar et al., 2018; Simões et al., 2018). Research has provided substantial insights into the immune genes and pathways that shape resistance to vector-borne pathogens and has revealed many promising targets for genetic manipulation (Wilke and Marrelli, 2015; Yen et al., 2018; Shaw and Catteruccia, 2019). However, we are only just beginning to understand the complexity underlying mosquito–pathogen interactions. The response mosquitoes mount toward a given pathogen is a dynamic phenotype that is dependent upon both the specific mosquito–pathogen pairing (Lambrechts et al., 2013; Zouache et al., 2014; Duggal et al., 2015; Chouin-Carneiro et al., 2016) and the variation in key environmental factors (Okech et al., 2007; Alto et al., 2008; Parham and Michael, 2010; Parham et al., 2015; Cohen et al., 2016; Gloria-Soria et al., 2017; Murdock et al., 2017; Shragai et al., 2017; Siraj et al., 2018). Knowledge of how vectors respond to environmental variation is especially relevant for understanding how vector-borne pathogens emerge, defining the biological constraints on transmission, and anticipating the robustness of novel vector-control approaches (that manipulate mosquito physiological and immunological mechanisms to limit virus infection) in field settings.
Vector-borne flaviviruses are emerging threats to human health. Yellow fever virus (YFV), dengue virus (DENV), West Nile virus (WNV), and most recently, Zika virus (ZIKV) can be found throughout tropical and subtropical zones. For successful transmission, flaviviruses are taken up through the bite of a mosquito vector when it takes a blood meal from an infectious host. To complete infection within the mosquito vector, the virus needs to efficiently enter, replicate within, and escape several tissue barriers, primarily the midgut and salivary glands (Franz et al., 2015; Kumar et al., 2018). A number of studies have demonstrated that mosquitoes elicit major transcriptional changes in response to flavivirus infection, which could play an important role in limiting infection (Xi et al., 2008; Sim and Dimopoulos, 2010; Colpitts et al., 2011; Bonizzoni et al., 2012; Chauhan et al., 2012; Angleró-Rodríguez et al., 2017; Etebari et al., 2017; Saldaña et al., 2017a). These include differential regulation of genes involved in RNA interference (RNAi), classical immune pathways (e.g., JAK-STAT, Toll), production and transport of energy, and metabolism, as well as the production of non-coding RNAs (small and long) and microRNAs that could be involved in targeted gene regulation. Currently, it is unclear how conserved these responses to infection are across different mosquito–flavivirus combinations (Etebari et al., 2017) and how relevant environmental variation shapes the nature, magnitude, and timing of these responses (Murdock et al., 2012b, 2014; Adelman et al., 2013).
One of the major environmental variables that influence mosquito–pathogen interactions, as well as vector-borne transmission, is variation in environmental temperature. Mosquitoes are small ectothermic organisms, and many studies have already demonstrated that temperature can markedly affect diverse aspects of mosquito physiology, ecology, and pathogen replication. This, in turn, will shape the proportion of mosquitoes that become infected and infectious, the overall transmission potential of mosquito populations (Christofferson and Mores, 2016; Murdock et al., 2017; Tesla et al., 2018a; Mordecai et al., 2019), and how the distributions of mosquito vectors and vector-borne pathogens will shift in response to climate change (Siraj et al., 2014; Campbell et al., 2015; Ryan et al., 2015, 2018; Misslin et al., 2016). The extent to which temperature shapes transmission directly, through effects on pathogen biology, or indirectly, through effects on mosquito immunity and physiology, remains largely unexplored (Murdock et al., 2012a).
In previous work (Tesla et al., 2018a), we have demonstrated that ZIKV transmission by Aedes aegypti is optimized at a mean temperature of 29°C and has a thermal range from 22.7 to 34.7°C. Further, we observed constraints on ZIKV transmission at cool and warm temperatures to be mediated by different factors. To overcome the midgut barrier, a minimal viral load is necessary (Vazeille et al., 2019). Cool temperatures inhibited ZIKV transmission due to slow virus replication and escape from the midgut, resulting in a decrease in metrics of vector competence (e.g., the proportion of mosquitoes that became infected, disseminated infection, and became infectious) and lengthening the extrinsic incubation period (e.g., the interval of time before ZIKV can be detected in the saliva and mosquitoes become infectious [Tesla et al., 2018a; see Figures 1, 2)]. In contrast, increased mosquito mortality at hotter temperatures constrained transmission despite efficient ZIKV infection and rapid dissemination.
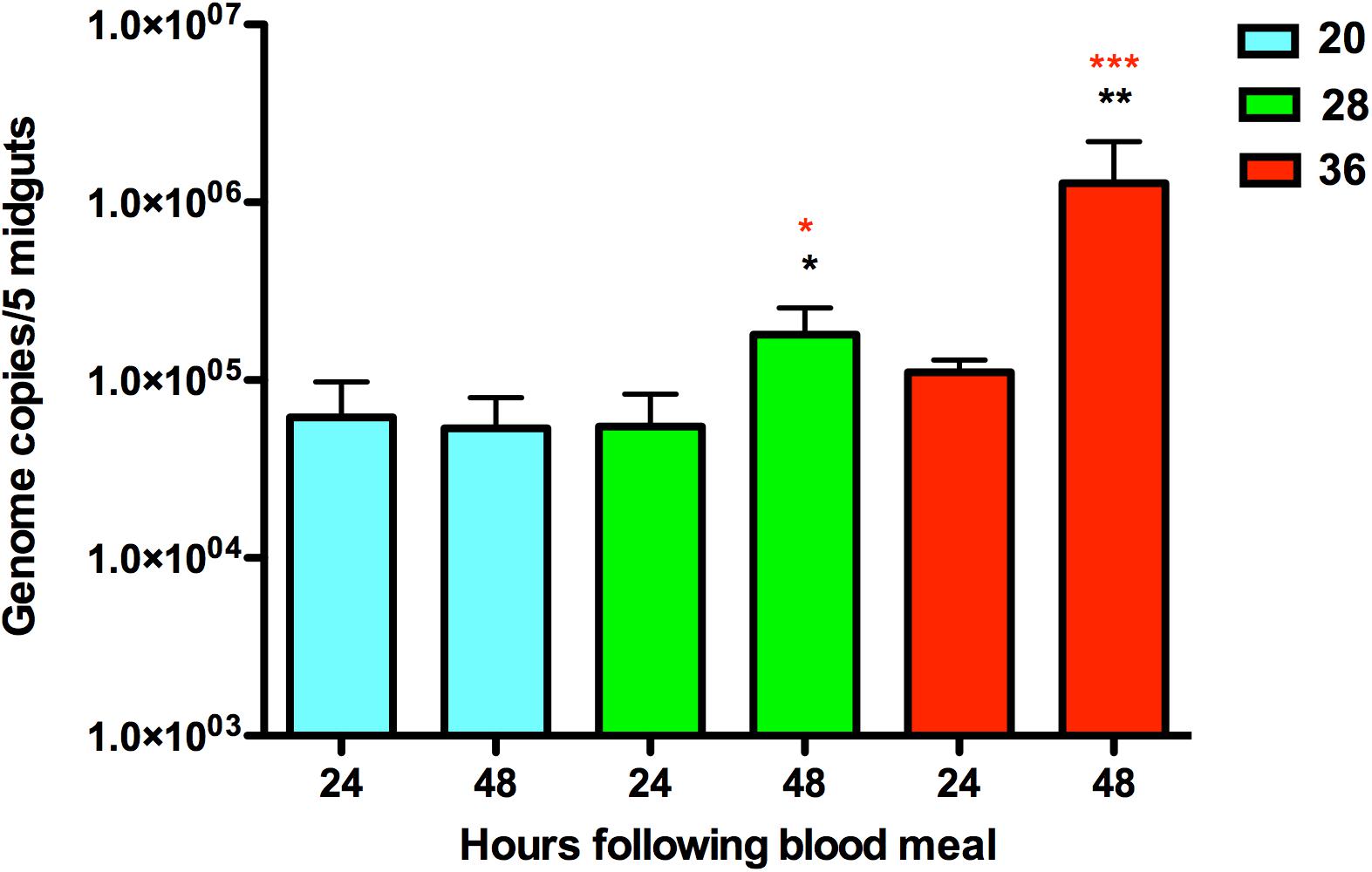
Figure 1. Quantification of ZIKV RNA in infected Aedes aegypti mosquitoes housed at 20, 28, and 36°C at two time points, 24 and 48 h. The average number of ZIKV genome copies present in pools of five midgut samples and standard error are shown. The y-axis is log-transformed. Black asterisks represent p-value < 0.05 calculated by T-test between the time points of 24 and 48 h post-infection for the same temperature. Red asterisks represent p-value < 0.05 calculated by ANOVA with Tukey correction for multiple hypotheses between the three different temperatures for the same time point after blood meal.
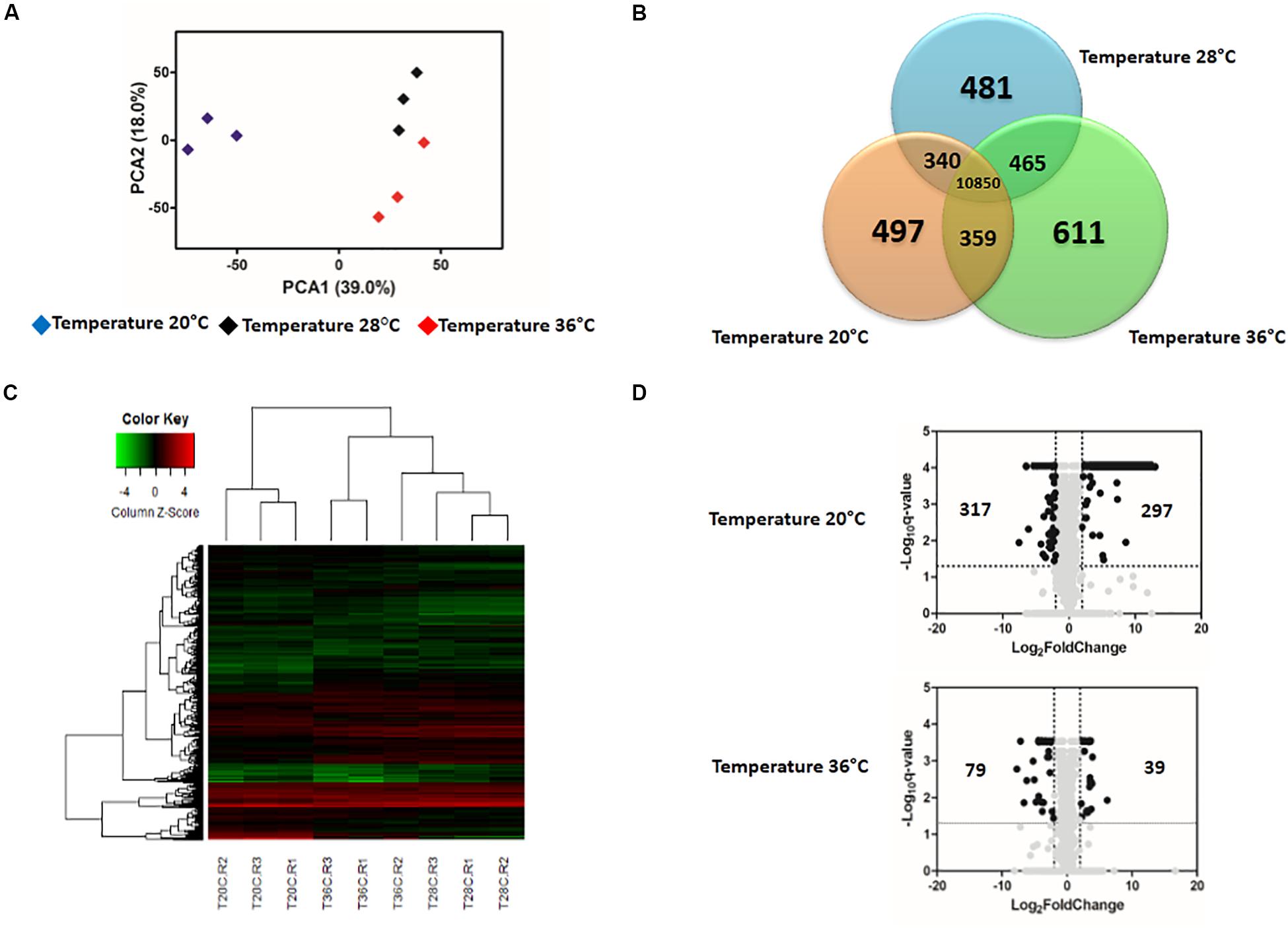
Figure 2. Effect of temperature in the expression profile of uninfected blood-fed Aedes aegypti. (A) Principal Component Analysis (PCA) plot showing the global gene expression profiles. (B) Venn Diagram reporting the number of specific and shared genes. (C) Heatmap plot showing local differences (naming scheme: TC20R1 = replicate 1 for the temperature 20°C). (D) Volcano plot representing the differential gene expression in RNAseq samples from Ae. aegypti exposed to three different constant temperatures (20, 28, and 36°C) at 48 h.
These temperature constraints on infection are likely regulated through different mechanisms. Temperature variation in general could profoundly impact arbovirus infection and replication early in infection due to shifts in the balance and dynamics of the midgut environment, the first host environment encountered. This environment is fairly complex, as arboviruses encounter oxidative and nitration stress associated with digestion of the blood meal (Luckhart et al., 1998; Graça-Souza et al., 2006; Xi et al., 2008), the presence [e.g., Wolbachia (Bian et al., 2010)] and proliferation of microbial flora (Xi et al., 2008; Carissimo et al., 2014; Hegde et al., 2015; Saraiva et al., 2016; Barletta et al., 2017; Saldaña et al., 2017b), and key immune factors (Campbell et al., 2008; Xi et al., 2008; Cirimotich et al., 2009; Sánchez-Vargas et al., 2009; Souza-Neto et al., 2009). To better understand the effects of temperature on the ZIKV-mosquito interaction, we used RNA sequencing to describe the transcriptional response of Ae. aegypti midguts to ZIKV during the early infection process at three different temperatures (20, 28, and 36°C) previously shown to impact ZIKV infection, dissemination, and transmission rates (Tesla et al., 2018a).
Materials and Methods
Ethics Statement
All mosquito infection work with ZIKV was approved by the University of Georgia, Athens Institutional Biosafety Committee (reference number 2015-0039).
Virus Production
Zika virus MEX1-44 was isolated from Ae. aegypti mosquitoes from Tapachula, Chiapas, Mexico in January 2016, kindly provided by the University of Texas Medical Branch Arbovirus Reference Collection. ZIKV stocks were propagated in Vero cells cultured in DMEM (Dulbeco’s Modified Eagle Medium), 5% fetal bovine serum (FBS) at 37°C and 5% CO2. Four days following inoculation, when cells showed visible cytopathic effect (>90%), supernatant containing virus was collected, cell debris was cleared by centrifugation (3,500 × g for 5 min), and virus was aliquoted and frozen at −80°C. The stock was titrated using standard plaque assays on Vero cells (Willard et al., 2017) and expressed in plaque-forming units per milliliter (PFU/mL).
Mosquito Husbandry
Aedes aegypti eggs collected in Chiapas, Mexico, were hatched in ddH2O under reduced pressure in a vacuum desiccator. Larvae were grown in trays, with 200 larvae in 1L ddH2O and four fish food pellets (Hikari Cichlid Cod Fish Pellets). Emerging adults were kept in rearing cages and fed with 10% sucrose solution ad libitum. Colonies were maintained on O-positive human blood (Interstate Blood Bank, male donors between 30 and 35 year). Both larvae and adults were maintained under the standard insectary conditions (27°C ± 0.5°C, 80% ± 5% relative humidity, and a 12:12 light: dark diurnal cycle) (Percival Scientific) that replicate rearing and experimental conditions used in Tesla et al. (2018a).
Experimental Mosquito Infection
Briefly, 3 to 4-day-old female mosquitoes (F5 generation; n = 600) were separated using a vacuum aspirator, transferred to an arthropod containment level three (ACL-3) facility at the University of Georgia and housed at 28°C ± 0.5°C, 80% ± 5% relative humidity, 12 h:12 h light:dark cycle (Percival Scientific). Mosquitoes were fed with a non-infectious or ZIKV-containing blood-meal (106 PFU/mL) after a 12-h period of starvation in a manner previously described (Tesla et al., 2018b). After the feed, engorged ZIKV-exposed (n = 120) and unexposed mosquitoes (n = 120) were randomly allocated across 12 paper cups (six with ZIKV-exposed and six with ZIKV-unexposed mosquitoes, n = 20 mosquitoes per cup). Forty engorged females that fed on either infectious blood or non-infectious blood were randomly distributed across one of three temperature treatments, 20°C, 28°C, and 36°C (±0.5°C) at 80% ± 5% relative humidity and with a 12 h:12 h light:dark cycle. Mosquitoes were provided 10% sucrose ad libitum throughout the duration of the experiment (48 h), and three full biological replicates were performed.
Viral Genome Quantification
Viral genomes were detected as previously described (Willard et al., 2019). Briefly, dissected mosquito midguts were collected in pools of five, and viral RNA was isolated using the Zymo Quick-RNA Viral Kit (Zymo, Irvine, CA, United States). Viral RNA samples were reverse-transcribed (RT) to cDNA (High Capacity RNA-to-cDNA Kit, Applied Biosystems, Foster City, CA, United States). The F5 generation was used to maintain genetic proximity with field populations of Ae. aegypti. To quantify the number of ZIKV genomes, we used the cDNA in a quantitative PCR (qPCR) reaction assay using TaqMan Gene Expression Master Mix (Applied Biosystems, ThermoFisher, Waltham, MA, United States), primers, and probes (F: ZIKV 1086, R: ZIKV 1162c, ZIKV 1107-FAM; TaqMan MGB Probe; Invitrogen Custom Primers) (Lanciotti et al., 2008). Each sample was analyzed in duplicate, and each plate contained a DNA plasmid standard curve (ZIKV molecular clone), no template, and no primer controls. ZIKV copy numbers were extrapolated from the generated standard curve using the Applied Biosystems protocol. The limit of detection was experimentally established to be 30 copies. Final copy numbers were adjusted by back-calculations to the total RNA and cDNA volume and expressed as copies per five-midgut pool. Outliers were identified by Grubbs test, implemented in Graphpad QuickCalcs1, and removed. The results were analyzed in GraphPad Prism 5.0. An unpaired T-test was applied to compare the two different times (24 and 48) post-infection for the same temperature. ANOVA with Tukey as the multiple hypothesis correction was used to compare the data from the three different temperatures (20, 28, and 36) at the same time points. A p-value below 0.05 was considered statistically significant.
Midgut Dissection and RNA Isolation
Mosquito responses to blood meal digestion, midgut microbial proliferation, and arbovirus midgut infection contribute significantly to viral bottlenecks during infection, and these processes could dynamically differ with temperature (Xi et al., 2008; Bonizzoni et al., 2011; Oliveira et al., 2011; Ramirez et al., 2012). Further, these responses likely occur early on during the infection process (<3 days post-infection) at optimal temperatures for ZIKV transmission (e.g., 29°C), as the proportion of mosquitoes with ZIKV-infected midguts plateaus by day three post-infection at temperatures above 28°C (Tesla et al., 2018a). To capture gene expression changes in the mosquito midgut early during the infection process, we isolated total RNA from midgut tissues. Fifteen ZIKV-exposed and 15 non-exposed mosquitoes were removed from each temperature at 24 and 48 h post-infection for RNA sequencing. The mosquitoes were killed with cold ethanol and washed in cold PBS containing 0.2% RNase inhibitor (Sigma-Aldrich) and 0.1% DEPC (Amresco). Midguts were carefully dissected and stored immediately in RNAlater (Invitrogen) at 4°C for 24 h, after which they were transferred to −80°C conditions. Total RNA was isolated from pools of 15 midguts using the Qiagen RNeasy Mini Kit as per the manufacturer’s instructions.
Library Preparation and Sequencing
Extracted RNA was sent to the University of Georgia Genomics Core for cDNA library preparation and RNA sequencing. The quality of the total RNA was analyzed using an Agilent Bioanalyzer RNA 6000 Pico Kit (Agilent Technologies). Poly(A) mRNA from total RNA was captured by magnetic oligo-dT beads and fragmented by heating (94°C, 6 min) in the presence of divalent cations (Mg2+) using the KAPA Stranded mRNA-Seq Kit for Illumina®. Fragmented poly(A) mRNA samples were converted to cDNA using random priming. After second-strand synthesis, an adenine residue was added to 3′-end of dscDNA fragments, and Illumina TruSeqLT adapter ligation to 3′-dAMP library fragments was performed. Adapter-ligated DNA was amplified by PCR using the KAPA Library Amplification Primer Mix. After creating the library of DNA templates, the fragment size distribution of these libraries was confirmed by electrophoresis and the library concentration was determined by qPCR. Sequencing was completed on an Illumina NextSeq 500 using the PE75bp (75 bp sequencing reads) settings on a High-Output 150-cycle kit using Illumina standard protocols for loading. Raw sequencing files were demultiplexed using the Illumina BaseSpace cloud platform demultiplexing pipeline. Four technical replicates were run per sample.
RNA Sequence Analysis in Response to Temperature and Zika Virus Exposure
The Tuxedo suite of tools was used to analyze the RNA-seq data (Trapnell et al., 2013). At each stage of the analysis pipeline, we were careful to identify and correct possible sources of bias in the study (Conesa et al., 2016). Read quality was assessed using FastQC (Andrews, 2010). Poor quality reads (quality score below 20), short reads (less than 25 bases), and adapter sequences (Bolger et al., 2014) were removed using Trimmomatic (v 0.36).
We aligned and mapped clean reads up to two different loci to the Ae. aegypti genome (NCBI ID: GCA_002204515.1) obtained from NCBI2 using TopHat (v 2.1.1) (Trapnell et al., 2009). We then performed differential gene expression analysis on RNAseq reads using Cuffdiff (Trapnell et al., 2013), which calculates expression levels in response to experimental treatments of interest (e.g., temperature and ZIKV-exposure) and determines whether observed variations in expression levels across groups are significantly different. The Ae. aegypti genome and its annotation file (NCBI ID: GCA_002204515.1) were used to run bias detection and determine the transcripts examined for differential expression, respectively. The relative expression levels were produced as fragments per kilobase of transcript per million fragments mapped (FPKM values), which normalized read count based on gene length and the total number of mapped reads. After quantification and normalization of expression values, differential expression analysis was carried out on the experimental data from 24 and 48 h following the blood meal. To characterize changes in the RNA transcriptome in response to temperature, we compared the RNA expression profiles of unexposed mosquitoes maintained at standard conditions and near the predicted thermal optimum for ZIKV transmission (28°C) to those of mosquitoes housed under cool (20°C) and hot (36°C) conditions. To determine whether temperature modified global RNA expression in ZIKV-exposed individuals, we compared whether the variation in expression profiles of ZIKV-exposed individuals across temperature treatments was similar to that of their unexposed counterparts. Finally, to evaluate the effects of ZIKV infection on global RNA expression, we compared RNA expression between unexposed and ZIKV-exposed mosquitoes within a given temperature treatment. The data quality assessment from the RNAseq analysis was performed using R package (v 3.4.4) (R Development Core Team, 2008) and plotted using GraphPad Prism (version 5.01 for Windows, GraphPad Software, San Diego, CA, United States)3. Perl scripts were written to extract specific information from RNAseq analysis output files necessary for each assessment of the results obtained. Principal-component analysis (PCA) was employed to examine the quality of replicates and the overall differences between samples and to determine the proximity among the experimental groups.
Gene Ontology (GO) Analysis
We employed BiNGO (v 3.0.3) (Maere et al., 2005) to determine which GO terms were significantly over-represented in a set of genes. First, the FASTA sequences of all differentially expressed genes were recovered using the protein database on the NCBI’s Batch Entrez4. The sequences were inputted into STRING (v 10.5) (Szklarczyk et al., 2017) to predict protein–protein interactions. GO functional annotations were provided by AgBase-Goanna (v 2.00) (Mccarthy et al., 2006), using the Ae. aegypti protein sequences in FASTA format from the STRING database as an input file. The similarity search for GO annotations was performed by Blastp using the UniProt Database with E-value: 10e–5, Matrix: BLOSUM62, and word size: 3. This file annotation was employed as a reference set for enrichment analysis. We used the default mode, employing Hypergeometric tests for assessing the enrichment of a GO term in the test set, and p-values were corrected by Benjamini and Hochberg False Discovery Rate (FDR) correction to control for type I error. Corrected p-values less than 0.05 were considered significant.
Results
From a total of 36 RNA samples that were sequenced using the Illumina platform, the number of clean paired reads varied between 2,063,917 and 3,028,305 for each library. All libraries resulted in a concordant pair alignment rate higher than 70%.
Cool Temperatures Restrict ZIKV Replication in the Mosquito Midgut
Viral RNA quantification of exposed mosquitoes revealed levels of ZIKV RNA in midgut samples from mosquitoes that imbibed a blood meal containing ZIKV (Figure 1). ZIKV replication was evident in mosquitoes housed at 28°C and 36°C, with increases in mosquito RNA copy number between 24 and 48 h post-feed (hpf). Further, the efficiency of ZIKV replication was maximized at the warmest temperature, with mosquitoes housed at 36°C having higher ZIKV RNA levels in their midguts than those housed at 28°C. Although the presence of ZIKV RNA was observed 24 and 48 h following the infectious blood meal in mosquitoes housed in the cool environment (20°C), we did not find significant increases in the viral copy number (Figure 1). This result suggests that cooler temperatures constrain ZIKV replication in the midgut.
The Effect of Temperature on Gene Expression in Unexposed Mosquitoes
Gene-expression patterns were influenced by both temperature and time post-blood feed. In general, the principal component analysis (PCA) plot showed a high degree of reproducibility among the replicate samples within each temperature treatment. At 24 hpf, temperature clearly distinguished gene expression profiles (Supplementary Figure S1A). However, by 48 hpf, mosquitoes housed at 28°C and 36°C had gene expression profiles that were more similar to each other than to mosquitoes housed at 20°C (Figure 2A). According to the Venn diagram, the vast majority of genes expressed in the midgut were similar for mosquitoes housed across the three temperature treatments (10,459 at 24 hpf, Supplementary Figure S1B; 10,850 at 48 hpf, Figure 2B). When concentrating on the differentially expressed genes, Euclidean distance heatmap analysis demonstrated that samples at 24 hpf were separated by temperature treatment in three distinct groups (Supplementary Figure S1C), while for samples at 48 hpf, 28 and 36°C samples cluster more closely with one another than with the 20°C (Figure 2C). This is the same profile as was seen in the PCA analysis, which employed all transcriptome data. Among these genes, 1665 differentially expressed genes (q-value < 0.05) were seen between mosquitoes held at 20 and 28°C for 24 h, and 3634 were seen by 48 h (Supplementary Table S1). In contrast, when comparing mosquitoes held at 36 and 28°C, we identified 1697 and 1695 differentially expressed genes after 24 and 48 h, respectively (Supplementary Table S2). In order to perform a more stringent analysis, we produced a volcano plot, which indicated a total of 70 and 297 upregulated genes [log2(Fold Change) >2 and q-value < 0.05] in mosquitoes housed at 20°C for 24 and 48 h, respectively, compared to those housed at 28°C. For downregulated genes [log2(Fold Change) <−2 and q-value < 0.05], we found 53 and 317 after 24 and 48 h, respectively (Supplementary Figure S1D and Figure 2D). We did not find as strong an effect of exposure time when mosquitoes were housed at 36°C, with only 19 and 39 genes up-regulated and 89 and 79 genes down-regulated after 24 and 48 h, respectively (Supplementary Figure S1D and Figure 2D).
To further define the effect of temperature on mosquito cellular and physiological responses, we sorted the top 20 up- or down-regulated genes at both the cool and hot temperatures, using mosquitoes maintained at 28°C as our standard (Table 1). The cool temperature produced a larger response, up-regulating genes hundreds of times compared to standard conditions, whereas the 36°C treatment had a more moderate effect. The transcript with the highest enrichment at 20°C was protein-G12 (1459-fold), whereas a serine protease easter was the most down-regulated (187-fold). For mosquitoes housed at 36°C, heat shock protein 70 (HSP 70) was the transcript most enriched (71-fold) relative to 28°C, while facilitated trehalose transporter Tret1 was the most down-regulated gene (212-fold). Surprisingly, both cool and warm temperature treatments induced some of the same genes to be differentially regulated. Six genes (protein G12, serine protease SP24D, and chymotrypsin-2) were up-regulated in mosquitoes housed at both 20°C and 36°C when compared to those at 28°C, while six genes (serine protease easter, facilitated trehalose transporter Tret1, venom protease, solute carrier family 2 facilitated glucose transporter member 3, and tryptase) were down-regulated.

Table 1. Top 20 up- and down-regulated genes of uninfected Aedes aegypti in response to low (20°C) and to high (36°C) temperature for 48 h post-blood-feeding (relative to standard rearing temperature of 28°C).
All differentially expressed genes in the midgut at 48 hpf were submitted for gene ontology (GO) analysis to identify cellular processes that were most perturbed in response to temperature treatment, as revealed by the transcriptome profile. GO analysis of the enriched and depleted transcripts from mosquitoes housed at 20°C revealed that 20 enriched GO terms were related to oxidation–reduction processes and 111 depleted GO terms were involved in gene expression, RNA processing, metabolic processes, and generation of energy (Figure 3). Mosquitoes housed at 36°C displayed up-regulated expression related to amine metabolism and cell redox homeostasis processes and down-regulated expression of genes associated with metabolic processes, cellular respiration, and energy derivation by oxidation of organic compounds (Figure 3).
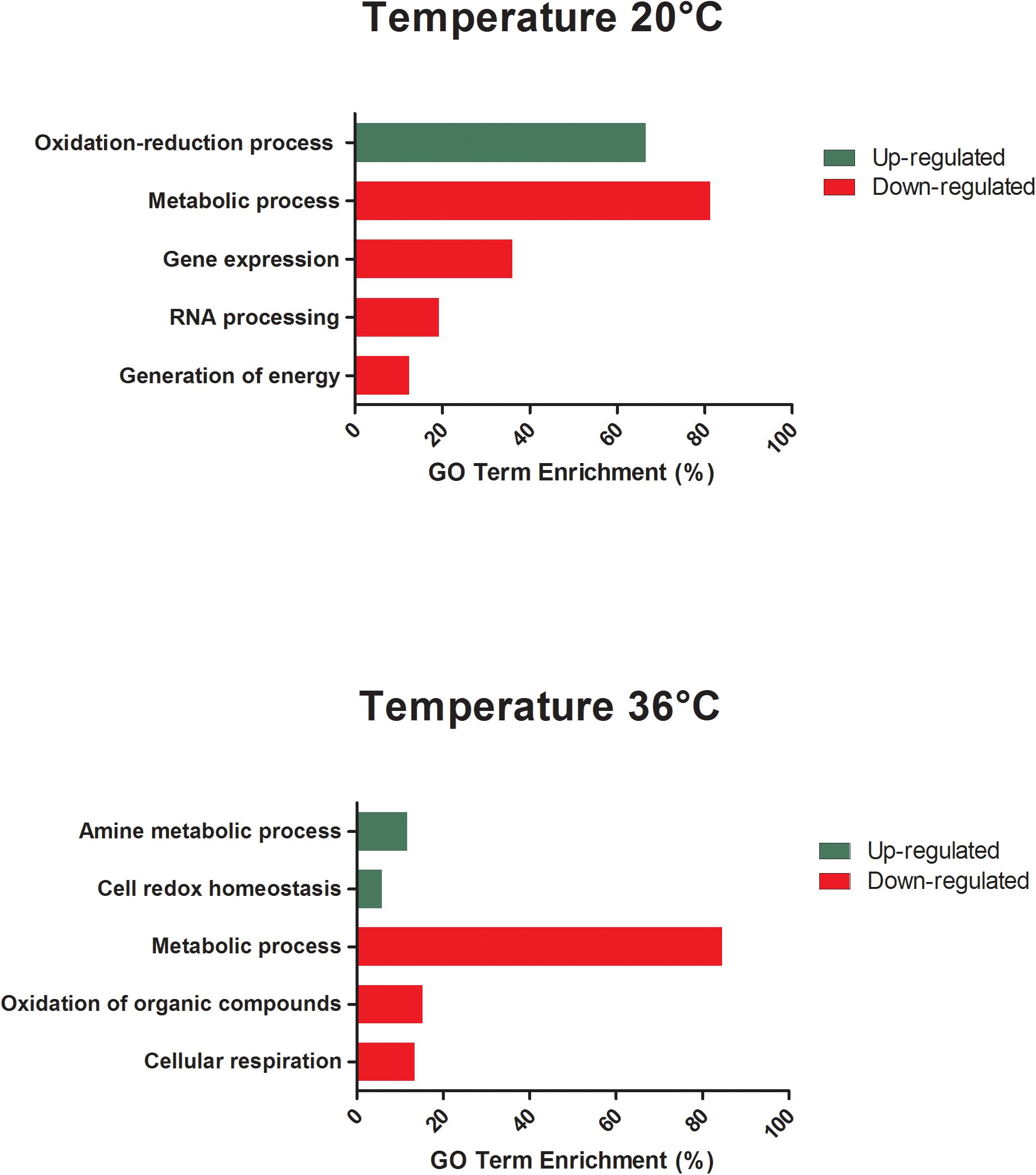
Figure 3. GO term enrichment analysis of differentially expressed genes of unexposed Aedes aegypti kept under the lowest (20°C) and the highest (36°C) temperature conditions in the category of biological processes for up- and down-regulated genes at 48 h.
The Effect of Temperature on Gene Expression in ZIKV-Exposed Mosquitoes
The gene expression profiles in ZIKV-exposed mosquitoes were also significantly affected by environmental temperature and time post-blood feed. In general, the effect of temperature on differential gene expression was similar to patterns observed in non-ZIKV-exposed blood-fed control mosquitoes outlined above. For example, PCA and heatmap analyses on differential gene expression from mosquito midguts at 24 hpf illustrated three distinct groups separated by temperature treatment (Supplementary Figures S2A,C). At 48 hpf, gene expression in mosquitoes housed at 20°C was more distinct than those housed at 28 and 36°C (Figures 4A,C). Further, Venn diagrams demonstrate that 1,416 and 10,786 genes were expressed across all temperatures at 24 hpf (Supplementary Figure S2B) and 48 hpf (Figure 4B), respectively, with the highest overlap in gene expression occurring in mosquitoes housed at 28°C and 36°C at both time points (Supplementary Figure S2B and Figure 4B). Finally, as seen in the absence of ZIKV infection, at 24 hpf, only 1669 and 1797 genes were differentially expressed in mosquitoes housed at 20 and 36°C, respectively, relative to those housed at 28°C. By 48 hpf, we observed a general increase in the number of genes differentially expressed at 20°C (3056, Supplementary Table S3) and a general decrease at 36°C (1518, Supplementary Table S4) relative to mosquitoes housed at 28°C. When we evaluated differential expression through a volcano plot, the profile was similar to that of mosquitoes that were not exposed to ZIKV (Supplementary Figure S2D and Figure 4D).
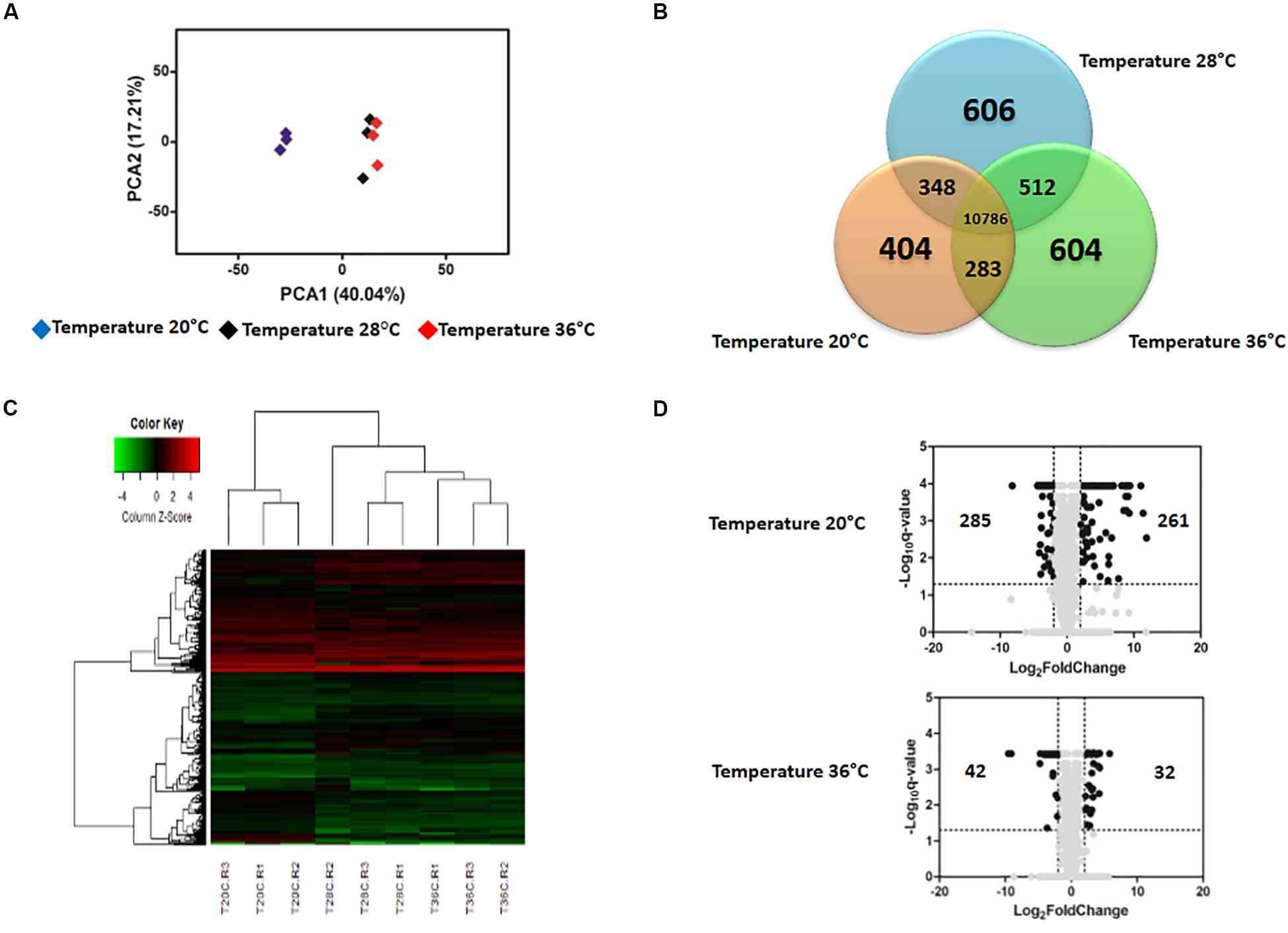
Figure 4. Effect of temperature on the expression profile of Zika-exposed Aedes aegypti. (A) Principal Component Analysis (PCA) plot showing the global gene expression profiles. (B) Venn Diagram reporting the number of specific and shared genes. (C) Heatmap plot showing local differences (naming scheme: TC20R1 = replicate 1 for the temperature 20°C). (D) Volcano plot representing the differential gene expression in RNAseq samples from infected Ae. aegypti exposed to three different constant temperatures (20, 28, and 36°C) at 48 h.
Several genes that strongly increased at 20°C remained among the top 20 differentially expressed after ZIKV exposure. Lysosomal alpha-mannosidase (XP_021703511.1), two vitellogenins (XP_001660818.2 and XP_001657509.1), phosphoenolpyruvate carboxykinase (XP_001647937.2), protein G12 isoforms (XP_021712126.1, XP_021701760.1, XP_001660827.1, XP_021701761.1, XP_001656377.1, and XP_001656375.1), beta-galactosidase (XP_021705369.1), alpha-N-acetylgalactosaminidase (XP_001650490.2), serine protease SP24D (XP_001659962.1), and chymotrypsin-2 (XP_021698904.1) are some of the genes that remained among the top 20 genes changed by cold temperature in the ZIKV-infection condition. We also saw that although glutamine synthetase (XP_001654186.1) and trypsin (XP_001663002.1) were not listed among the 20 most differentially expressed, they showed enrichment of 50- and 74-fold, respectively. Further, facilitated trehalose transporter Tret1, found to be the most down-regulated in unexposed mosquitoes kept at 36°C, was also the most down-regulated (25-fold) in ZIKV-exposed mosquitoes kept at the same temperature. Finally, serine protease SP24D (XP_001659962.1) and some G12 proteins (XP_021701760.1, XP_021701761.1, XP_001660827.1, and XP_001656377.1) that were enriched at both cool and warm temperatures remained enriched at these temperatures in ZIKV-exposed mosquitoes (Table 2).
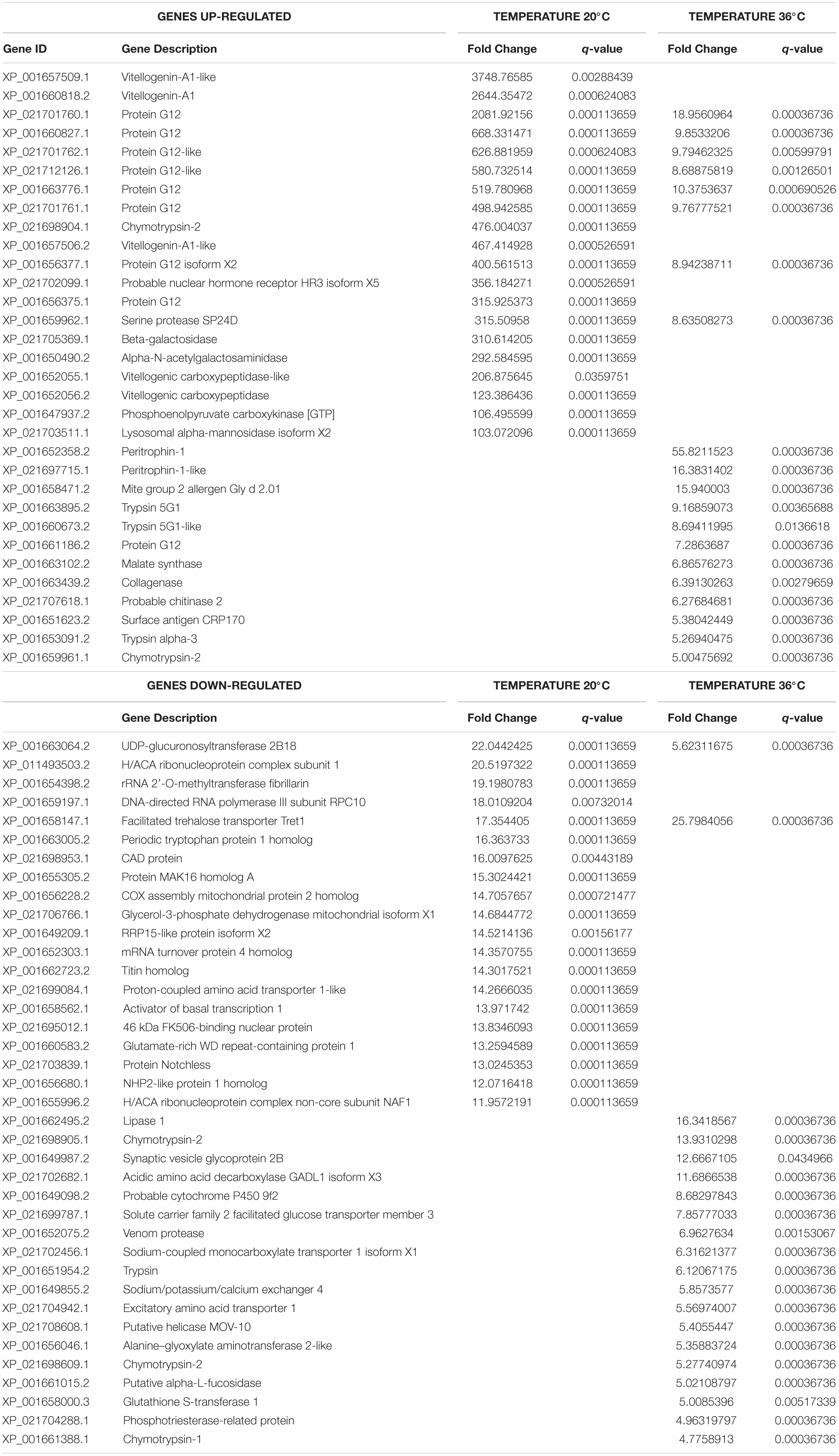
Table 2. Top 20 up- and down-regulated genes of Zika-exposed Aedes aegypti in response to low (20°C) and to high (36°C) temperature for 48 h post-blood-feeding relative to standard insectary conditions (28°C).
Despite these similarities, we did note some key differences between the top 20 genes most differentially expressed in unexposed (Table 1) and ZIKV-exposed mosquitoes (Table 2), with the greatest change in expression reflected in mosquitoes housed at 20°C and 36°C at 48 h relative to those housed at 28°C. In unexposed mosquitoes kept at 20°C, vitellogenin-A1-like (XP_001657509.1) and vitellogenin-A1 (XP_001660818.2) were up-regulated, yet ZIKV exposure amplified the enrichment from 138- and 68-fold to 3748- and 2644-fold (Tables 1, 2). Surprisingly, in ZIKV-exposed mosquitoes, we did not detect a large up-regulation of Hsp70 at 36°C like we did in the unexposed population.
The GO analysis of differentially expressed genes at 48 hpf in ZIKV-exposed mosquitoes demonstrated distinct effects of cool and warm temperatures on cellular and metabolic function relative to mosquitoes housed at 28°C (Figure 5). Further, the functions of these differentially expressed genes were, in part, different from those in unexposed mosquitoes housed at these temperatures (Figure 3). For example, when maintained at 20°C, oxidative-reduction processes were no longer enriched, as was seen in unexposed mosquitoes. Instead, ZIKV-exposed mosquitoes housed at 20°C had significant enrichment of the Toll signaling pathway, a known anti-dengue pathway in Ae. aegypti (Tchankouo-Nguetcheu et al., 2010). Additionally, endosome transport, Ras protein signal transduction, actin cytoskeleton organization, epithelial tube morphogenesis, pH reduction, and proteolysis were enriched. In contrast, nuclear transport, regulation of viral reproduction, “de novo” protein folding, generation of energy, gene expression, RNA processing, and metabolic processes were down-regulated in addition to the expression associated with gene expression, RNA processing, and metabolic processes observed in unexposed counterparts at this temperature (Figure 5). ZIKV-exposed mosquitoes housed at 36°C no longer had significant enrichment in genes associated with cellular amine processes and had significant depletion in genes associated with hexose and phosphate metabolic processes and with the generation of precursor metabolites and energy (Figure 5).
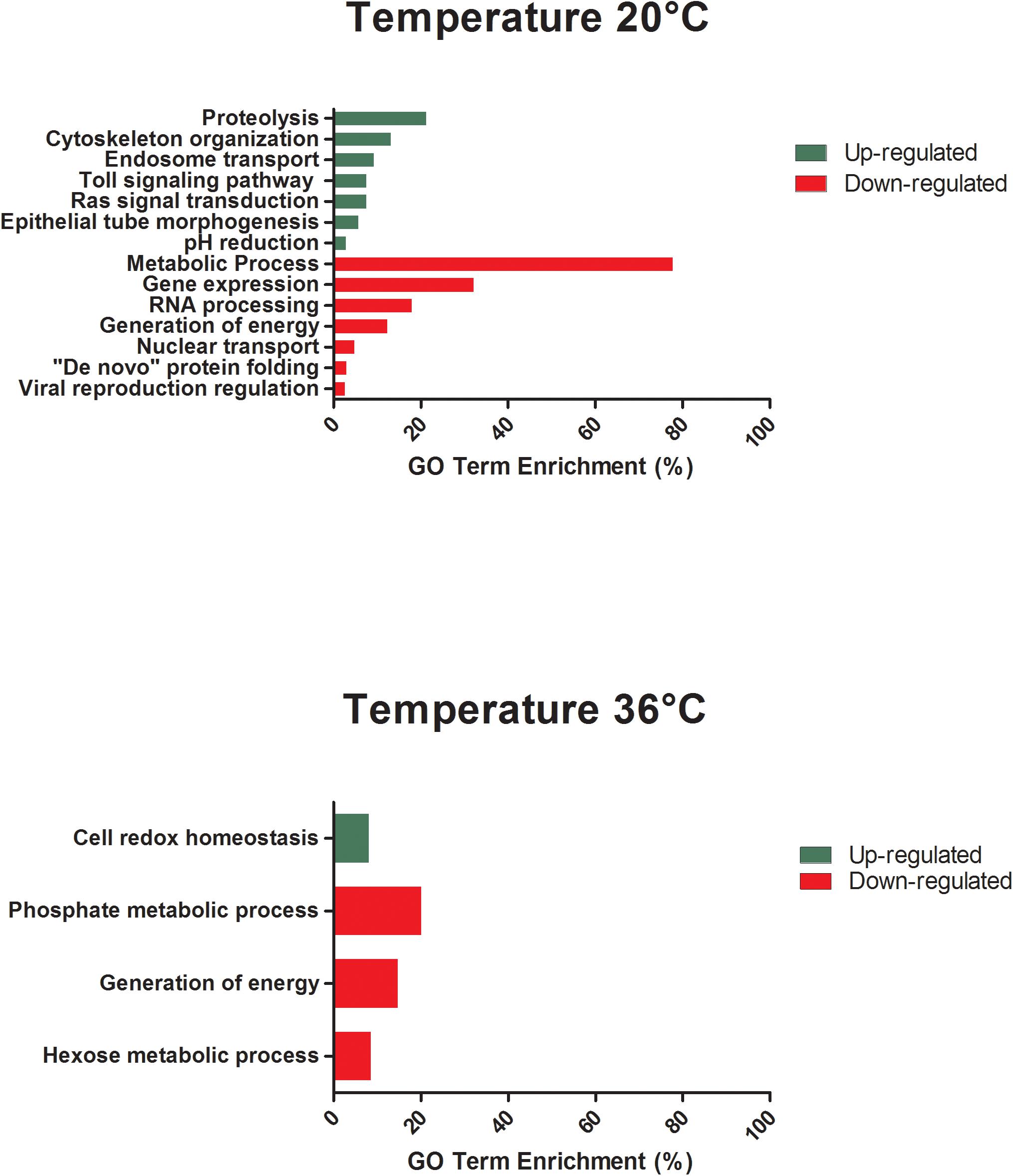
Figure 5. GO term enrichment analysis of differentially expressed genes of Aedes aegypti exposed to Zika virus under the lowest (20°C) and in the highest (36°C) temperature conditions in the category of biological processes for up- and down-regulated genes at 48 h.
Effects of Temperature on Oxidative Stress and Innate Immune Mechanisms
To further explore the effects of temperature on uninfected and ZIKV-exposed mosquitoes, we highlight differences in those genes involved in managing oxidative stress, innate immunity, and apoptosis at both 20 and 36°C compared to 28°C at 48 hpf (Supplementary Tables S5, S6 and Supplementary Figures S3, S4). Virus infection has previously been shown to be modified by complex responses related to detoxification of the blood meal, metabolism, immunity, and apoptosis in some systems (Sanders et al., 2005; Girard et al., 2010; Tchankouo-Nguetcheu et al., 2010; Colpitts et al., 2011; Wang et al., 2012; Neill et al., 2015; Eng et al., 2016). Further, these responses need not respond equivalently to temperature variation, as shown in previous work, which demonstrated that mosquito immune responses differed qualitatively and quantitatively across a range of environmental temperatures (Murdock et al., 2012b). The majority of genes involved in managing oxidative stress, innate immunity, and apoptosis exhibited qualitatively different patterns in gene expression in response to cool and warm temperatures in uninfected and ZIKV-exposed mosquitoes. However, while significant, the majority of these differences were very subtle (<2.0-fold; Supplementary Figures S3, S4). We did observe components of the melanization and Toll pathways to be modestly expressed (>2.0 fold; Supplementary Figures S3, S4) in response to temperature. An isoform of phenoloxidase (XP_021699380.1), a c-type lectin (XP_001661643.1), and the Toll receptor 6 (TLR6) (XP_021712805.1) were significantly enriched in both uninfected and ZIKV-exposed mosquitoes housed at 20°C.
Mosquito Responses to ZIKV Infection Are Remarkably Modulated by Environmental Temperature
To investigate the effects of ZIKV-exposure on global gene-expression patterns from mosquito midguts early on in the infection process, we compared gene expression between ZIKV-exposed and unexposed mosquitoes within each temperature treatment. The PCA plots demonstrate that ZIKV exposure does not alter mosquito transcription at 24 hpf (Supplementary Figure S5) within a given temperature treatment. However, 48-hpf ZIKV-exposed and unexposed samples under the cold temperature condition were distributed in two distinct groups, although one of the ZIKV-exposed biological replicates was relatively close to the control group (Figure 6A). Also, the overall number of differentially expressed genes between ZIKV-exposed and unexposed mosquitoes varied across temperature treatments. For example, at 24 hpf, we observed a total of 225, 154, and 161 genes differentially expressed between ZIKV-exposed and unexposed mosquitoes at 20°C, 28°C, and 36°C, respectively (Supplementary Tables S7–S9). We identified only two proteins – a sodium/potassium/calcium exchanger 4 (XP_001649855.2) and an uncharacterized protein (XP_001654261.2) – that were up-regulated in ZIKV-exposed mosquitoes at all temperatures (Supplementary Figure S6A) and one protein – chymotrypsin-2 (XP_021698609.1) that was down-regulated (Supplementary Figure S6B). At 48 hpf, we observed more genes to be differentially expressed (1188 genes) in mosquitoes housed at 20°C between ZIKV-exposed and unexposed mosquitoes, versus only 180 and 50 genes at 28 and 36°C, respectively (Supplementary Tables S7–S9). Only one uncharacterized protein (XP_001658660.2) was up-regulated in the ZIKV-exposed mosquitoes at all temperatures (Figures 7A,B) at this sampling time point. These results indicate that while the physiological responses of mosquito midguts to ZIKV exposure early in the infection process are similar within a given temperature treatment, these responses are significantly distinct across different environmental temperatures.
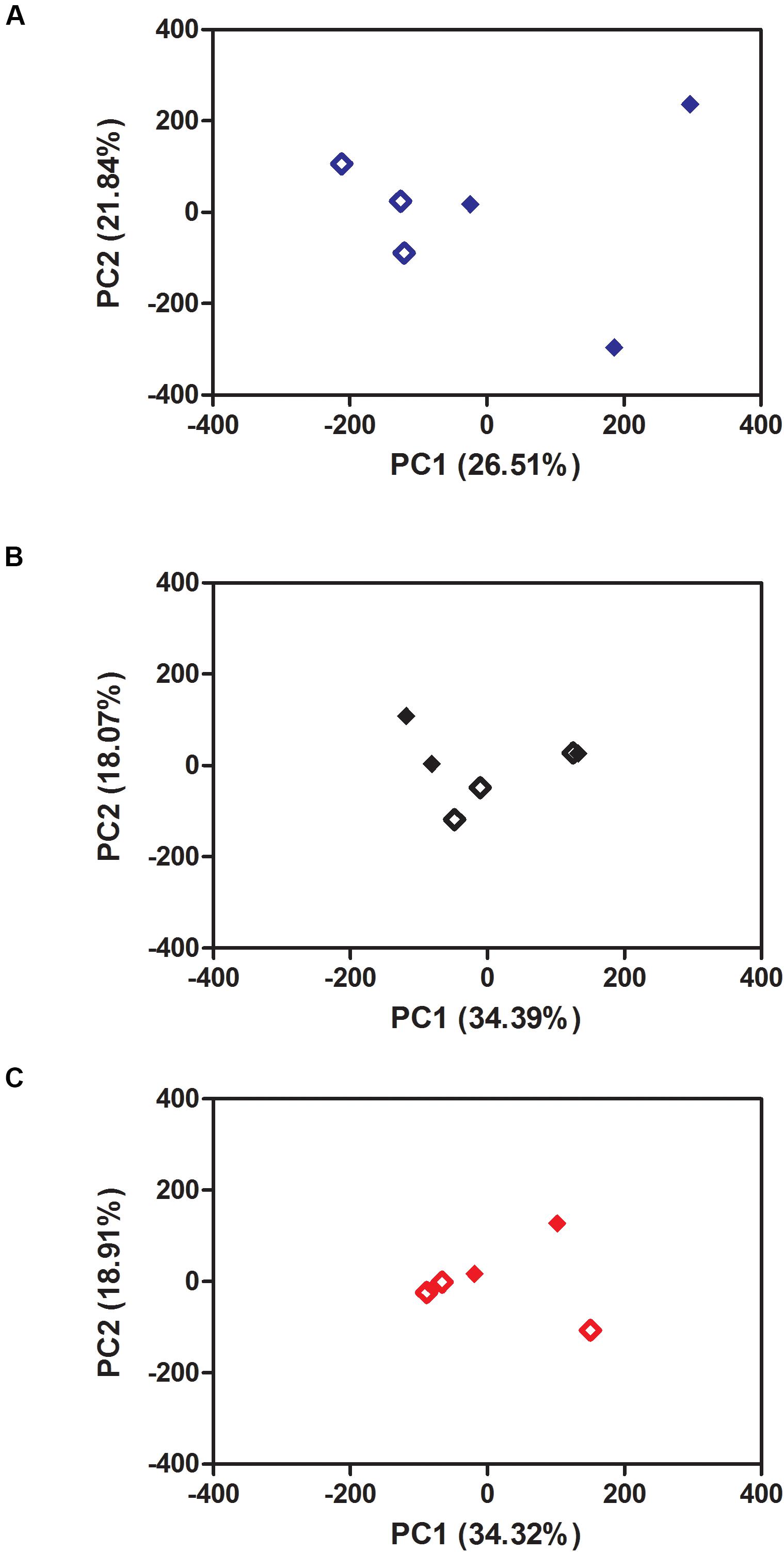
Figure 6. Principal Component Analysis (PCA) of the effect of Zika exposure on Aedes aegypti gene expression at three different constant temperatures: (A) 20°C, (B) 28°C, (C) 36°C at 48 h. Open diamond: non-exposed mosquitoes. Closed diamond: Zika-exposed mosquitoes.
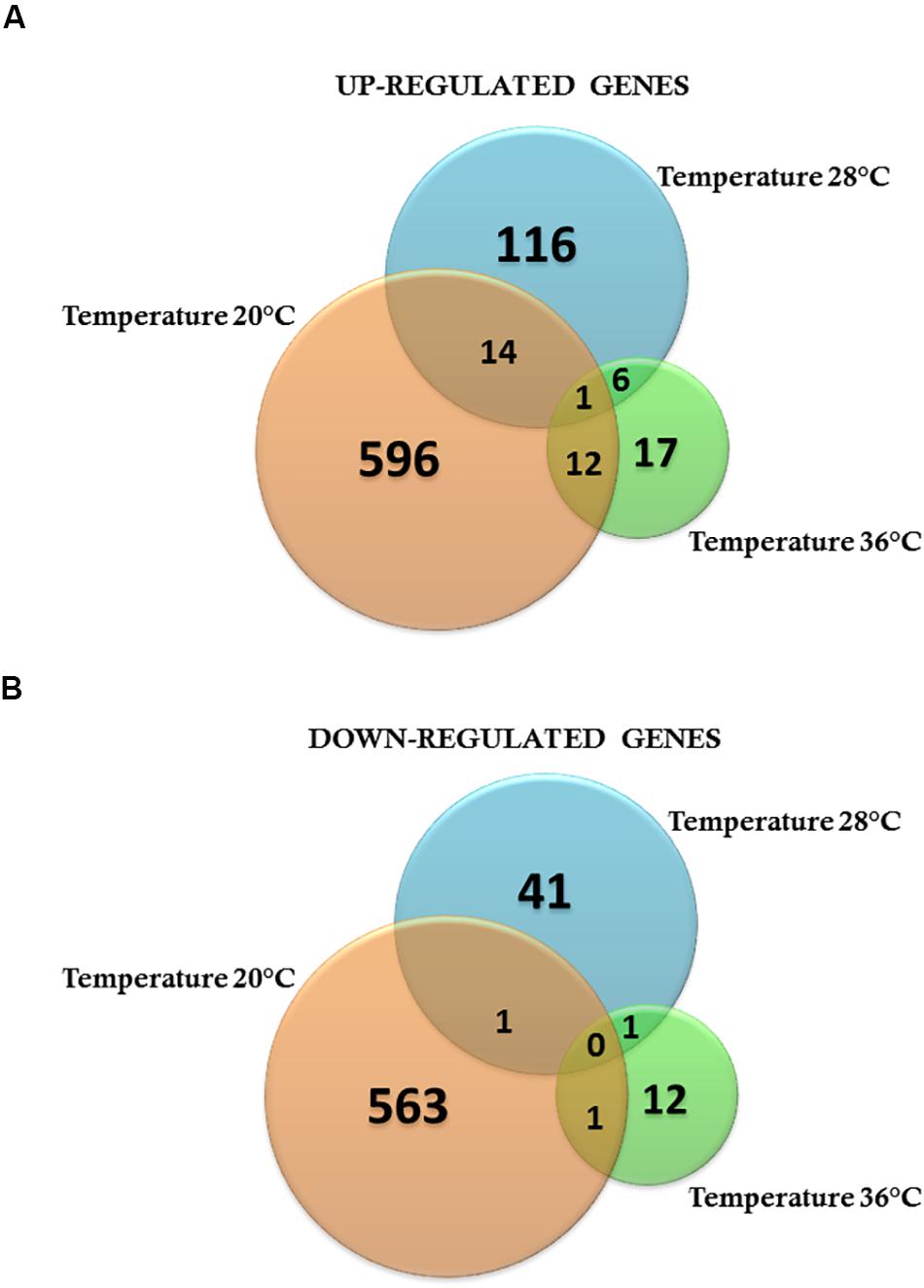
Figure 7. Venn diagram representing the number of specific differentially expressed genes for Zika-infected Aedes aegypti exposed to three different constant temperatures (20, 28, and 36°C). (A) Up-regulated genes. (B) Down-regulated genes at 48 h.
When concentrating on the top 10 most differentially expressed genes between ZIKV-exposed and unexposed mosquitoes at each temperature treatment at 48 hpf (Table 3), only at 20°C did we observe genes with altered expression (enrichment or depletion) of 10-fold or more. GO analysis demonstrated that functions associated with these genes included metallopeptidases (angiotensin-converting enzyme and vitellogenic carboxypeptidase), ammonia/nitrogen metabolism (argininosuccinate lyase and alanine aminotransferase 1), and iron ion binding (a member of the cytochrome P450 family, 4g15).

Table 3. Top 10 up- and down-regulated genes of Zika-exposed Aedes aegypti relative to uninfected mosquitoes kept at low (20°C), standard (28°C), and high (36°C) temperatures 48 h post-blood-feeding.
Interestingly, two vitellogenins (XP_001660818.2 and XP_001657509.1) that were strongly up-regulated in ZIKV-exposed and unexposed mosquitoes housed at 20°C relative to those housed at 28°C were the genes most enriched by ZIKV exposure at the cold temperature (Table 3). The expression of these genes did not change in response to ZIKV exposure at 28°C (Supplementary Table S8) and 36°C (Supplementary Table S9), suggesting that cold stress alters the midgut vitellogenin expression and may be more significant during a viral infection. ZIKV exposure induced a depletion of beta-1,3-glucan-binding protein (GNBP), which binds to β−1,3-glucan and lipopolysaccharide on the surface of pathogens (Dimopoulos et al., 1997), when mosquitoes were maintained at 20°C. Finally, among the most down-regulated genes in ZIKV-exposed mosquitoes at 28°C, solute carrier family 22 (XP_001656519.2), synaptic vesicle glycoprotein (XP_001651077.2), and vitellogenic carboxypeptidase (XP_001652056.2) (Table 3) were also among the most down-regulated genes in unexposed mosquitoes housed at 36°C relative to 28°C (Table 1).
Discussion
The dynamics and distribution of vector-borne diseases depend on the interplay between the pathogen, the mosquito, and the environment. Temperature is a strong driver of vector-borne disease transmission (Kilpatrick et al., 2008; Lambrechts et al., 2011; Carrington et al., 2013a, b; Mordecai et al., 2013, 2017; Johnson et al., 2015; Shocket et al., 2018; Tesla et al., 2018a). Despite the strong effects of temperature on mosquito-borne pathogens, little is known about the underlying mechanisms involved (Adelman et al., 2013). In this study, RNA sequencing of Ae. aegypti midguts unexposed or exposed to ZIKV, taken early in the infection process, revealed different transcriptional responses to variation in environmental temperature, with ZIKV infection modifying these responses to temperature.
Previously, we found that temperature significantly affected the efficiency of the establishment of ZIKV infection in Ae. aegypti midguts, with cool temperatures limiting ZIKV transmission primarily due to poor midgut infection, slow replication, and poor dissemination, while high mosquito mortality at warmer temperatures inhibited ZIKV transmission despite efficient ZIKV infection (Tesla et al., 2018a). Similar to our previous study, we were unable to detect ZIKV replication when mosquitoes were housed in cool conditions (Figure 1), which is similar to findings for other flavivirus systems in which infection rates were measured across different constant temperatures (Watts et al., 1987; Kilpatrick et al., 2008; Johansson et al., 2012; Xiao et al., 2014). There was no significant difference in the viral RNA levels quantified from the three temperatures at 24 hpf, reflecting the initial concentration of viral RNA ingested in the blood meal (Figure 1). Therefore, we can confirm that while all treatment groups obtained ZIKV in the blood meal, only the mosquitoes housed at standard (28°C) or warm (36°C) temperatures were actively replicating virus by 48 hpf.
We found that variation in temperature elicited strong expression responses in unexposed and ZIKV-exposed mosquitoes. We observed that mosquitoes housed at a cold temperature (20°C) had more genes differentially expressed at 48 hpf relative to mosquitoes housed in standard (28°C) and warm environments (36°C), which exhibited more similar patterns in gene expression (Figures 2A, 4A). This is not entirely surprising, as metabolic theory predicts that low body temperatures will inevitably depress the rates of biochemical reactions (Angilletta et al., 2010). Our data support this hypothesis, as many of the genes drastically altered at 20°C participate in blood meal digestion, peritrophic membrane (PM) formation, metabolism, and managing oxidative stress associated with the breakdown of hemoglobin into heme, a cytotoxic product (Tables 1, 2). Phosphoenolpyruvate carboxykinase and trypsin are upregulated in Ae. aegypti midgut during the first few hours after ingestion of a blood meal (Sanders et al., 2003) but were extraordinarily up-regulated in mosquitoes 48 hpf when housed in a cool environment in our study. Additionally, protein G12, which has previously been associated with blood meal digestion and nitrile-specific detoxification (Morlais et al., 2003; Fischer et al., 2008; Bonizzoni et al., 2011), was one of the most enriched transcripts. Further, two digestive proteases involved in glycoside hydrolysis, beta-galactosidase and alpha-N-acetylgalactosaminidase (Santamaría et al., 2015), were highly induced at 48 hpf. Glutamine synthetase, an enzyme that contributes to PM formation, was also highly induced, further demonstrating that cool temperatures delay blood meal digestion (Supplementary Tables S1, S3). Although the PM is semi-permeable, it is thought to form a barrier and protect the midgut from pathogens [e.g., viruses (Wang and Granados, 2000), bacteria (Kuraishi et al., 2011; Jin et al., 2019), malaria (Rodgers et al., 2017), and protozoa (Weiss et al., 2014)] and other harmful substances present in the insect gut after a blood meal (Wang et al., 2004; Shibata et al., 2015).
We also observed several genes involved in the innate immune response to be modestly upregulated in response to 20°C relative to their levels under warmer temperature conditions in both unexposed and ZIKV-exposed mosquitoes. Melanization is a major effector mechanism of the mosquito immune response and has been implicated in the defense against a diversity of pathogens [e.g., bacteria (Hillyer et al., 2003a, b), malaria (Kumar et al., 2003; Jaramillo-Gutierrez et al., 2009), filarial worms (Christensen et al., 2005; Huang et al., 2005), and viruses (Rodriguez-Andres et al., 2012)]. Phenoloxidase, a key enzyme in the melanization pathway, was up-regulated in mosquito midguts at 20°C (Supplementary Figures S3B, S4B). Studies in both butterflies and Anopheles stephensi demonstrated that phenoloxidase activity was higher at cool temperatures and becomes less efficient at warmer temperatures (Suwanchaichinda and Paskewitz, 1998; Murdock et al., 2012b). The production of melanin is also essential for other physiological processes such as cold acclimatization in insects (Crill et al., 1996; Kutch et al., 2014), the formation of the hard protective layer around eggs, and wound healing (Lai et al., 2009). Our data also reveal that c-type lectin, reported to participate in the activation of the melanization cascade (Yu and Kanost, 2000; Christensen et al., 2005), was also up-regulated. Therefore, our results suggest that cold stress triggers numerous molecular changes in the mosquito, including modest changes in the levels of important immune effectors that could have important consequences for arboviral infection.
Contrary to what we observed at the cool temperature, exposure to hot conditions (36°C) does not trigger pronounced up- or down-regulation of genes relative to standard conditions (28°C). The heat shock protein 70 (HSP 70) transcript was most enriched in response to the hot environment (Table 1). The upregulation of HSP 70 is associated with reduced lifespans in other insect systems (Feder and Krebs, 1998; Feder, 1999), which may explain the rapid mosquito mortality we observed at this temperature in previous work (Tesla et al., 2018a). HSP70 has also been suggested to facilitate arbovirus infection in mosquitoes in terms of viral entry, viral RNA synthesis, and virion production (Kuadkitkan et al., 2010; Taguwa et al., 2015).
Zika virus exposure induced very modest effects when comparing ZIKV-exposed and unexposed mosquitoes reared at 28 and 36°C, while those that experienced the cool temperature exhibited a larger alteration in gene expression at 48 hpf (Figure 6 and Supplementary Figure S5). This may not be entirely surprising, as ZIKV-induced transcriptional changes under standard rearing conditions (28°C) have previously been shown to be subtle 48 h post-infection (Murdock et al., 2012a) and ZIKV was only observed to be actively replicating at 28 and 36°C. When concentrating on differentially expressed genes between ZIKV-exposed and unexposed mosquitoes at each temperature treatment (Table 3), only at 20°C do we observe changes of 10-fold or more. Therefore, the presence of ZIKV in the blood meal did alter the response of mosquitoes to temperature variation, with the most pronounced differences occurring in mosquitoes housed at the cool temperature. In particular, the midguts of ZIKV-exposed mosquitoes experienced enhanced signal transduction processes, pH modification, midgut epithelial morphogenesis, and Toll pathway activation relative to ZIKV-exposed mosquitoes at 28°C (Figure 5), and this pattern was qualitatively different to a similar comparison in unexposed mosquitoes (Figure 3). These changes could be reflective of patterns observed in other studies demonstrating that mosquitoes infected with blood-borne pathogens actively modify ROS metabolism in midgut cells to control levels of hydrogen peroxide (H2O2), which in turn is an important modulator of downstream innate immune responses (e.g., Toll pathway), antimicrobial peptide production, and pathogen infection (Molina-Cruz et al., 2008; Herrera-Ortiz et al., 2011; Oliveira et al., 2011, 2012). Furthermore, the presence and abundance of particular microbial flora [e.g., Wolbachia (Pan et al., 2012)] and the proliferation of the midgut flora due to ingestion of the blood meal (Carissimo et al., 2014; Saraiva et al., 2016; Barletta et al., 2017) can also trigger ROS production, with temperature variation modifying these effects.
Additionally, vitellogenin proteins (Vg) were highly upregulated (>3000 fold) when ZIKV-exposed mosquitoes were housed at cool temperatures relative to ZIKV-exposed mosquitoes at 28°C (Table 2) and unexposed mosquitoes at 20°C (Table 3). Vg is a precursor egg-yolk protein but may also function by shielding cells from the negative effects of inflammation and infection (Corona et al., 2007; Azevedo et al., 2011). Work with honey bees suggests that Vg-incubated insect cells display enhanced tolerance against H2O2-induced oxidative stress (Fluri et al., 1977; Seehuus et al., 2006). Vg also binds to dying cells, suggesting that it may play a role in recognizing damaged cells and shielding healthy cells from toxic by-products (Havukainen et al., 2013). Both caspase dronc [an inhibitor caspase (Cooper et al., 2007)] and an effector caspase (Bryant et al., 2008), which are important components of the apoptotic pathway controlling mechanisms of cell death, were enriched in mosquitoes housed at 20°C (Supplementary Figures S3E, S4E). Thus, elevated Vg expression may combat cellular damage resulting from elevated heme toxicity and oxidative stress (Havukainen et al., 2013) associated with the delayed breakdown of hemoglobin in the blood meal (Bottino-Rojas et al., 2015) in mosquitoes housed at cool temperatures. There may also be a direct effect of Vg on ZIKV, as it has been associated with antiviral effects in some fish species (Garcia et al., 2010). Entry of ZIKV into mammalian cells is associated with apoptotic mimicry, with viral lipids interacting with phosphatidylserine receptors on cells gaining entry in a pathway similar to clearance of apoptotic cells (Hamel et al., 2015). Honey bee Vg binds to dying cells through interactions with lipids (Havukainen et al., 2013). Although the receptors ZIKV uses to interact and enter mosquito cells have not been identified, if Vg protein coats viral particles, it may impede normal cellular interaction and entry. Alternatively, ZIKV infection could be limited at the cooler temperature if infected mosquitoes balance ROS metabolism toward a higher state of oxidative stress, as shown in other systems (Molina-Cruz et al., 2008; Herrera-Ortiz et al., 2011; Pan et al., 2012; Bottino-Rojas et al., 2015; Wong et al., 2015), facilitating downstream innate immune mechanisms and virus killing. Whether overexpressions of Vg lipoproteins have direct effects on ZIKV infection or reflect a response to buffer ZIKV-exposed mosquitoes to a higher state of oxidative stress in the midgut remain open questions that will be explored in future experiments.
Finally, in ZIKV-exposed mosquitoes at 36°C, we observed peritrophin, one of the components of the peritrophic matrix, to be highly up-regulated relative to ZIKV-exposed mosquitoes housed at 28°C (Table 2) and unexposed mosquitoes housed at 36°C (Table 3). Although PM formation is highly induced 3–24 hpf, peritrophin can undergo positive modulation by pathogens in other vector-borne disease systems (e.g., Le. major) (Coutinho-Abreu et al., 2013). While we cannot confirm whether HSP70 or modulation of peritrophin play a role in ZIKV infection, these could be potential mechanisms explaining why we detect higher viral RNA levels at 36°C in this study (Figure 1) and efficient viral dissemination and salivary gland invasion in previous work (Tesla et al., 2018a).
Although we estimated the effects of mean constant temperatures on the immune-physiological profiles of Ae. aegypti in response to ZIKV infection to maintain experimental tractability, mosquitoes and their pathogens live in a variable world where temperatures fluctuate daily and seasonally (easily encompassing the range of temperatures explored here). Both mosquito immunity and virus transmission have previously been shown to differ in fluctuating environments relative to constant temperature environments (Lambrechts et al., 2011; Carrington et al., 2013a, b; Murdock et al., 2013). There is also a substantial body of work demonstrating carry-over effects of environmental variation in the larval environment on adult mosquito phenotypes, fitness, and metrics of transmission (Muturi and Alto, 2011; Muturi et al., 2011a, b; Alto and Bettinardi, 2013; Buckner et al., 2016; Evans et al., 2018). While outside the scope of this study, future work should investigate how both fluctuating environmental conditions and the larval environment modify the physiological and immunological responses of adult mosquitoes to arbovirus infection and temperature variation in the adult environment. Finally, while there is currently limited evidence demonstrating that mosquitoes select microhabitats to optimize internal body temperatures for metabolic demands (Blanford et al., 2009), as in other ectothermic organisms (Huey, 1991; Hertz et al., 1994; Gvoždík, 2002; Dillon et al., 2009), if this did occur in the field, it would modify the range of field-relevant temperatures mosquitoes effectively experience.
In this study, we demonstrate profound effects of temperature on ZIKV viral replication and the transcriptional responses of mosquitoes. Temperature variation may alter the ZIKV infection process either through modifying the response of mosquitoes to ZIKV infection, altering the efficiencies of viral-specific processes, or, more likely, both. Our study focused on midgut responses early in the infection process. However, disentangling these effects will require the sampling of other immunological tissues and at later time points where high levels of ZIKV RNA can be detected. While further work is needed to determine the precise mechanisms at play, our results indicate that temperature shifts the balance and dynamics of the midgut environment, which could result in direct and indirect consequences for the ZIKV-infection process. These results do reinforce the assertion that the conventional approach of studying the mechanisms underpinning mosquito–pathogen interactions under a narrow set of laboratory conditions or across canonical innate immune pathways is likely missing important biological complexity. To move forward, we need to begin framing our mechanistic understanding of this dynamic phenotype in the ecologically variable world in which mosquitoes and pathogens associate. This study represents a key advance toward this objective.
Data Availability Statement
The fast raw data were deposited in the NCBI SRA database under accession number PRJNA615972. This Sequence Read Archive (SRA) submission will be released on 2020–10–20 or upon publication, whichever is first.
Ethics Statement
All mosquito infection work with ZIKV was approved by the University of Georgia, Athens Institutional Biosafety Committee (reference number 2015-0039).
Author Contributions
BT performed the experiments. PF and TO analyzed the RNAseq data. CM, MB, TO, and LN designed the experiments, acquired the funding, and supervised the project. All authors wrote the manuscript.
Funding
This work was supported by co-funding from Brazilian Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG) (APQ-04765-16), the University of Georgia’s Office for Research (OVPR, grant 302070035), and an NSF-RAPID award 1640780. The funders had no role in study design, data collection and interpretation, and decision to submit the work for publication.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank the University of Texas Medical Branch Arbovirus Reference Collection for providing the virus and Dr. Americo Rodriguez from the Instituto Nacional de Salud Publica for providing mosquito eggs. We also thank the Universidade Federal de Viçosa and the University of Georgia for the use of the facilities in which this work was carried out. The bioinformatics analyses were performed in the UFV Cluster.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2020.00901/full#supplementary-material
Footnotes
- ^ https://www.graphpad.com/quickcalcs/Grubbs1.cfm
- ^ https://www.ncbi.nlm.nih.gov/
- ^ www.graphpad.com
- ^ https://www.ncbi.nlm.nih.gov/sites/batchentrez
References
Adelman, Z. N., Anderson, M. A. E., Wiley, M. R., Murreddu, M. G., Samuel, G. H., Morazzani, E. M., et al. (2013). Cooler temperatures destabilize RNA interference and increase susceptibility of disease vector mosquitoes to viral infection. PLoS Negl. Trop. Dis. 7:e2239. doi: 10.1371/journal.pntd.0002239
Alto, B. W., and Bettinardi, D. (2013). Temperature and dengue virus infection in mosquitoes: independent effects on the immature and adult stages. Am. J. Trop. Med. Hyg. 88, 497–505. doi: 10.4269/ajtmh.12-0421
Alto, B. W., Lounibos, L. P., Mores, C. N., and Reiskind, M. H. (2008). Larval competition alters susceptibility of adult Aedes mosquitoes to dengue infection. Proc. R. Soc. B Biol. Sci. 275, 463–471. doi: 10.1098/rspb.2007.1497
Andrews, S. (2010). FastQC: A Quality Control Tool for High Throughput Sequence Data. Available online at: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed March 11, 2018).
Angilletta, M. J., Huey, R. B., and Frazier, M. R. (2010). Thermodynamic effects on organismal performance: is hotter better? Physiol. Biochem. Zool. 83, 197–206. doi: 10.1086/648567
Angleró-Rodríguez, Y. I., MacLeod, H. J., Kang, S., Carlson, J. S., and Dimopoulos, N. J. G. (2017). Aedes aegypti molecular responses to Zika Virus: modulation of infection by the toll and Jak/Stat immune pathways and virus host factors. Front. Microbiol. 8:2050. doi: 10.3389/fmicb.2017.02050
Azevedo, D. O., Zanuncio, J. C., Delabie, J. H. C., and Serrão, J. E. (2011). Temporal variation of vitellogenin synthesis in Ectatomma tuberculatum (Formicidae: Ectatomminae) workers. J. Insect Physiol. 57, 972–977. doi: 10.1016/j.jinsphys.2011.04.015
Barletta, A. B. F., Nascimento-Silva, M. C. L., Talyuli, O. A. C., Oliveira, J. H. M., Pereira, L. O. R., Oliveira, P. L., et al. (2017). Microbiota activates IMD pathway and limits Sindbis infection in Aedes aegypti. Parasit. Vectors 10:103. doi: 10.1186/s13071-017-2040-2049
Bartholomay, L. C., and Michel, K. (2018). Mosquito immunobiology: the intersection of vector health and vector competence. Annu. Rev. Entomol. 63, 145–167. doi: 10.1146/annurev-ento-010715-023530
Bian, G., Xu, Y., Lu, P., Xie, Y., and Xi, Z. (2010). The endosymbiotic bacterium Wolbachia induces resistance to dengue virus in Aedes aegypti. PLoS Pathog. 6:e833. doi: 10.1371/journal.ppat.1000833
Blanford, S., Read, A. F., and Thomas, M. B. (2009). Thermal behaviour of Anopheles stephensi in response to infection with malaria and fungal entomopathogens. Malar. J. 8, 1–9. doi: 10.1186/1475-2875-8-72
Bolger, A. M., Lohse, M., and Usadel, B. (2014). Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 30, 2114–2120. doi: 10.1093/bioinformatics/btu170
Bonizzoni, M., Dunn, W. A., Campbell, C. L., Olson, K. E., Dimon, M. T., Marinotti, O., et al. (2011). RNA-seq analyses of blood-induced changes in gene expression in the mosquito vector species, Aedes aegypti. BMC Genomics 12:82. doi: 10.1186/1471-2164-12-82
Bonizzoni, M., Dunn, W. A., Campbell, C. L., Olson, K. E., Marinotti, O., and James, A. A. (2012). Complex modulation of the Aedes aegypti transcriptome in response to dengue virus infection. PLoS One 7:e50512. doi: 10.1371/journal.pone.0050512
Bottino-Rojas, V., Talyuli, O. A. C., Jupatanakul, N., Sim, S., Dimopoulos, G., Venancio, T. M., et al. (2015). Heme signaling impacts global gene expression, immunity and dengue virus infectivity in Aedes aegypti. PLoS One 10:e0135985. doi: 10.1371/journal.pone.0135985
Bryant, B., Blair, C. D., Olson, K. E., and Clem, R. J. (2008). Annotation and expression profiling of apoptosis-related genes in the yellow fever mosquito Aedes aegypti. Insect Biochem. Mol. Biol. 38, 331–345. doi: 10.1016/j.ibmb.2007.11.012
Buckner, E. A., Alto, B. W., and Lounibos, L. P. (2016). Larval temperature-food effects on adult mosquito infection and vertical transmission of dengue-1 virus. J. Med. Entomol. 53, 91–98. doi: 10.1093/jme/tjv145
Campbell, C. L., Keene, K. M., Brackney, D. E., Olson, K. E., Blair, C. D., Wilusz, J., et al. (2008). Aedes aegypti uses RNA interference in defense against Sindbis virus infection. BMC Microbiol. 8:47. doi: 10.1186/1471-2180-8-47
Campbell, L. P., Luther, C., Moo-Llanes, D., Ramsey, J. M., Danis-Lozano, R., and Peterson, A. T. (2015). Climate change influences on global distributions of dengue and chikungunya virus vectors. Philos. Trans. R. Soc. B Biol. Sci. 370:20140135. doi: 10.1098/rstb.2014.0135
Carissimo, G., Pondeville, E., McFarlane, M., Dietrich, I., Mitri, C., Bischoff, E., et al. (2014). Antiviral immunity of Anopheles gambiae is highly compartmentalized, with distinct roles for RNA interference and gut microbiota. Proc. Natl. Acad. Sci. U.S.A. 112, E176–E185. doi: 10.1073/pnas.1412984112
Carrington, L. B., Armijos, M. V., Lambrechts, L., and Scott, T. W. (2013a). Fluctuations at a low mean temperature accelerate dengue virus transmission by Aedes aegypti. PLoS Negl. Trop. Dis. 7:e2190. doi: 10.1371/journal.pntd.0002190
Carrington, L. B., Seifert, S. N., Armijos, M. V., Lambrechts, L., and Scott, T. W. (2013b). Reduction of Aedes aegypti vector competence for dengue virus under large temperature fluctuations. Am. J. Trop. Med. Hyg. 88, 689–697. doi: 10.4269/ajtmh.12-0488
Chauhan, C., Behura, S. K., DeBruyn, B., Lovin, D. D., Harker, B. W., Gomez-Machorro, C., et al. (2012). Comparative expression profiles of midgut genes in dengue virus refractory and susceptible Aedes aegypti across critical period for virus infection. PLoS One 7:e47350. doi: 10.1371/journal.pone.0047350
Chouin-Carneiro, T., Vega-Rua, A., Vazeille, M., Yebakima, A., Girod, R., Goindin, D., et al. (2016). Differential susceptibilities of Aedes aegypti and Aedes albopictus from the Americas to Zika Virus. PLoS Negl. Trop. Dis. 10:1–11. doi: 10.1371/journal.pntd.0004543
Christensen, B. M., Li, J., Chen, C. C., and Nappi, A. J. (2005). Melanization immune responses in mosquito vectors. Trends Parasitol. 21, 192–199. doi: 10.1016/j.pt.2005.02.007
Christofferson, R. C., and Mores, C. N. (2016). Potential for extrinsic incubation temperature to alter interplay between transmission potential and mortality of dengue-infected Aedes aegypti. Environ. Health Insights 10, 119–123. doi: 10.4137/ehi.s38345
Cirimotich, C. M., Scott, J. C., Phillips, A. T., Geiss, B. J., and Olson, K. E. (2009). Suppression of RNA interference increases alphavirus replication and virus-associated mortality in Aedes aegypti mosquitoes. BMC Microbiol. 9:49. doi: 10.1186/1471-2180-9-49
Cohen, J. M., Civitello, D. J., Brace, A. J., Feichtinger, E. M., Ortega, C. N., Richardson, J. C., et al. (2016). Spatial scale modulates the strength of ecological processes driving disease distributions. Proc. Natl. Acad. Sci. U.S.A. 113, E3359–E3364. doi: 10.1073/pnas.1521657113
Colpitts, T. M., Cox, J., Vanlandingham, D. L., Feitosa, F. M., Cheng, G., Kurscheid, S., et al. (2011). Alterations in the Aedes aegypti transcriptome during infection with west nile, dengue and yellow fever viruses. PLoS Pathog. 7:e1002189. doi: 10.1371/journal.ppat.1002189
Conesa, A., Madrigal, P., Tarazona, S., Gomez-Cabrero, D., Cervera, A., McPherson, A., et al. (2016). A survey of best practices for RNA-seq data analysis. Genome Biol. 17:13. doi: 10.1186/s13059-016-0881-888
Cooper, D. M., Thi, E. P., Chamberlain, C. M., Pio, F., and Lowenberger, C. (2007). Aedes Dronc: a novel ecdysone-inducible caspase in the yellow fever mosquito. Aedes aegypti. Insect Mol. Biol. 16, 563–572. doi: 10.1111/j.1365-2583.2007.00758.x
Corona, M., Velarde, R. A., Remolina, S., Moran-Lauter, A., Wang, Y., Hughes, K. A., et al. (2007). Vitellogenin, juvenile hormone, insulin signaling, and queen honey bee longevity. Proc. Natl. Acad. Sci. U.S.A. 104, 7128–7133. doi: 10.1073/pnas.0701909104
Coutinho-Abreu, I. V., Sharma, N. K., Robles-Murguia, M., and Ramalho-Ortigao, M. (2013). Characterization of Phlebotomus papatasi Peritrophins, and the Role of PpPer1 in Leishmania major Survival in its Natural Vector. PLoS Negl. Trop. Dis. 7:e2132. doi: 10.1371/journal.pntd.0002132
Crill, W. D., Huey, R. B., and Gilchrist, G. W. (1996). Within- and Between-Generation Effects of Temperature on the Morphology and Physiology of Drosophila melanogaster. Evolution. 50, 1205–1218. doi: 10.2307/2410661
Dillon, M. E., Wang, G., Garrity, P. A., and Huey, R. B. (2009). Review: thermal preference in Drosophila. J. Therm. Biol. 34, 109–119. doi: 10.1016/j.jtherbio.2008.11.007
Dimopoulos, G., Richman, A., Müller, H. M., and Kafatos, F. C. (1997). Molecular immune responses of the mosquito Anopheles gambiae to bacteria and malaria parasites. Proc. Natl. Acad. Sci. U.S.A. 94, 11508–11513. doi: 10.1073/pnas.94.21.11508
Duggal, N. K., Reisen, W. K., Fang, Y., Newman, R. M., Yang, X., Ebel, G. D., et al. (2015). Genotype-specific variation in West Nile virus dispersal in California. Virology 485, 79–85. doi: 10.1016/j.virol.2015.07.004
Eng, M. W., van Zuylen, M. N., and Severson, D. W. (2016). Apoptosis-related genes control autophagy and influence DENV-2 infection in the mosquito vector, Aedes aegypti. Insect Biochem. Mol. Biol. 76, 70–83. doi: 10.1016/j.ibmb.2016.07.004
Etebari, K., Hegde, S., Saldaña, M. A., Widen, S. G., Wood, T. G., Asgari, S., et al. (2017). Global transcriptome analysis of Aedes aegypti mosquitoes in response to Zika Virus Infection. mSphere 2:e00456-17. doi: 10.1128/mSphere.00456-417
Evans, M. V., Shiau, J. C., Solano, N., Brindley, M. A., Drake, J. M., and Murdock, C. C. (2018). Carry-over effects of urban larval environments on the transmission potential of dengue-2 virus. Parasit. Vectors 11:426. doi: 10.1186/s13071-018-3013-3013
Feder, M. E. (1999). Engineering candidate genes in studies of adaptation: the heat-shock protein Hsp70 in Drosophila melanogaster. Am. Nat. 154, S55–S66.
Feder, M. E., and Krebs, R. A. (1998). Natural and genetic engineering of the heat-shock protein Hsp70 in drosophila melanogaster: consequences for thermotolerance. Am. Zool. 38, 503–517.
Fischer, H. M., Wheat, C. W., Heckel, D. G., and Vogel, H. (2008). Evolutionary origins of a novel host plant detoxification gene in butterflies. Mol. Biol. Evol. 25, 809–820. doi: 10.1093/molbev/msn014
Fluri, P., Wille, H., Gerig, L., and Liischer, M. (1977). Juvenile hormone, vitellogenin and haemocyte composition in winter worker honeybees (Apis mellifera). Experientia 33, 1240–1241. doi: 10.1007/BF01922354
Franz, A. W. E., Kantor, A. M., Passarelli, A. L., and Clem, R. J. (2015). Tissue barriers to arbovirus infection in mosquitoes. Viruses 7, 3741–3767. doi: 10.3390/v7072795
Garcia, J., Munro, E. S., Monte, M. M., Fourrier, M. C. S., Whitelaw, J., Smail, D. A., et al. (2010). Atlantic salmon (Salmo salar L.) serum vitellogenin neutralises infectivity of infectious pancreatic necrosis virus (IPNV). Fish Shellfish Immunol. 29, 293–297. doi: 10.1016/j.fsi.2010.04.010
Girard, Y. A., Mayhew, G. F., Fuchs, J. F., Li, H., Schneider, B. S., McGee, C. E., et al. (2010). Transcriptome Changes in Culex quinquefasciatus (Diptera: Culicidae) salivary glands during west nile virus infection. J. Med. Entomol. 47, 421–435. doi: 10.1603/me09249
Gloria-Soria, A., Armstrong, P. M., Turner, P. E., and Turner, P. E. (2017). Infection rate of Aedes aegypti mosquitoes with dengue virus depends on the interaction between temperature and mosquito genotype. Proc. R. Soc. B Biol. Sci. 284:20171506. doi: 10.1098/rspb.2017.1506
Graça-Souza, A. V., Maya-Monteiro, C., Paiva-Silva, G. O., Braz, G. R. C., Paes, M. C., Sorgine, M. H. F., et al. (2006). Adaptations against heme toxicity in blood-feeding arthropods. Insect Biochem. Mol. Biol. 36, 322–335. doi: 10.1016/j.ibmb.2006.01.009
Gvoždík, L. (2002). To heat or to save time? Thermoregulation in the lizard Zootoca vivipara (Squamata: Lacertidae) in different thermal environments along an altitudinal gradient. Can. J. Zool. 80, 479–492. doi: 10.1139/z02-015
Hamel, R., Dejarnac, O., Wichit, S., Ekchariyawat, P., Neyret, A., Luplertlop, N., et al. (2015). Biology of Zika virus infection in human skin cells. J. Virol. 89, 8880–8896. doi: 10.1128/jvi.00354-315
Havukainen, H., Münch, D., Baumann, A., Zhong, S., Halskau, Ø, Krogsgaard, M., et al. (2013). Vitellogenin recognizes cell damage through membrane binding and shields living cells from reactive oxygen species. J. Biol. Chem. 288, 28369–28381. doi: 10.1074/jbc.M113.465021
Hegde, S., Rasgon, J. L., and Hughes, G. L. (2015). The microbiome modulates arbovirus transmission in mosquitoes. Curr. Opin. Virol. 15, 97–102. doi: 10.1016/j.coviro.2015.08.011
Herrera-Ortiz, A., Martínez-Barnetche, J., Smit, N., Rodriguez, M. H., and Lanz-Mendoza, H. (2011). The effect of nitric oxide and hydrogen peroxide in the activation of the systemic immune response of Anopheles albimanus infected with Plasmodium berghei. Dev. Comp. Immunol. 35, 44–50. doi: 10.1016/j.dci.2010.08.004
Hertz, P. E., Fleishman, L. J., and Armsby, C. (1994). The influence of light intensity and temperature on microhabitat selection in two anolis lizards. Funct. Ecol. 8, 720–729. doi: 10.2307/2390231
Hillyer, J. F., Schmidt, S. L., and Christensen, B. M. (2003a). Hemocyte-mediated phagocytosis and melanization in the mosquito Armigeres subalbatus following immune challenge by bacteria. Cell Tissue Res. 313, 117–127. doi: 10.1007/s00441-003-0744-y
Hillyer, J. F., Schmidt, S. L., and Christensen, B. M. (2003b). Rapid phagocytosis and melanization of bacteria and plasmodium sporozoites by hemocytes of the mosquito Aedes aegypti. J. Parasitol. 89, 62–69.
Huang, C. Y., Christensen, B. M., and Chen, C. C. (2005). Role of dopachrome conversion enzyme in the melanization of filarial worms in mosquitoes. Insect Mol. Biol. 14, 675–682. doi: 10.1111/j.1365-2583.2005.00597.x
Jaramillo-Gutierrez, G., Rodrigues, J., Ndikuyeze, G., Povelones, M., Molina-Cruz, A., and Barillas-Mury, C. (2009). Mosquito immune responses and compatibility between Plasmodium parasites and anopheline mosquitoes. BMC Microbiol. 9:154. doi: 10.1186/1471-2180-9-154
Jin, M., Liao, C., Chakrabarty, S., Wu, K., and Xiao, Y. (2019). Comparative proteomics of peritrophic matrix provides an insight into its role in Cry1Ac resistance of cotton bollworm helicoverpa armigera. Toxins 11:92. doi: 10.3390/toxins11020092
Johansson, M. A., Arana-Vizcarrondo, N., Biggerstaff, B. J., Gallagher, N., Marano, N., and Staples, J. E. (2012). Assessing the risk of international spread of yellow fever virus: a mathematical analysis of an urban outbreak in Asunción, 2008. Am. J. Trop. Med. Hyg. 86, 349–358. doi: 10.4269/ajtmh.2012.11-0432
Johnson, L. R., Ben-Horin, T., Lafferty, K. D., McNally, A., Mordecai, E., Paaijmans, K. P., et al. (2015). Understanding uncertainty in temperature effects on vector-borne disease: a Bayesian approach. Ecology 96, 203–213. doi: 10.1890/13-1964.1
Kilpatrick, A. M., Meola, M. A., Moudy, R. M., and Kramer, L. D. (2008). Temperature, viral genetics, and the transmission of West Nile virus by Culex pipiens mosquitoes. PLoS Pathog. 4:e1000092. doi: 10.1371/journal.ppat.1000092
Kuadkitkan, A., Wikan, N., Fongsaran, C., and Smith, D. R. (2010). Identification and characterization of prohibitin as a receptor protein mediating DENV-2 entry into insect cells. Virology 406, 149–161. doi: 10.1016/j.virol.2010.07.015
Kumar, A., Srivastava, P., Sirisena, P. D. N. N., Dubey, S. K., Kumar, R., Shrinet, J., et al. (2018). Mosquito innate immunity. Insects 9:E95. doi: 10.3390/insects9030095
Kumar, S., Christophides, G. K., Cantera, R., Charles, B., Han, Y. S., Meister, S., et al. (2003). The role of reactive oxygen species on Plasmodium melanotic encapsulation in Anopheles gambiae. Proc. Natl. Acad. Sci. U.S.A. 100, 14139–14144. doi: 10.1073/pnas.2036262100
Kuraishi, T., Binggeli, O., Opota, O., Buchon, N., and Lemaitre, B. (2011). Genetic evidence for a protective role of the peritrophic matrix against intestinal bacterial infection in Drosophila melanogaster. Proc. Natl. Acad. Sci. U.S.A. 108, 15966–15971. doi: 10.1073/pnas.1105994108
Kutch, I. C., Sevgili, H., Wittman, T., and Fedorka, K. M. (2014). Thermoregulatory strategy may shape immune investment in Drosophila melanogaster. J. Exp. Biol. 217, 3664–3669. doi: 10.1242/jeb.106294
Lai, S.-C., Chen, C.-C., and Hou, R. F. (2009). Immunolocalization of prophenoloxidase in the process of wound healing in the mosquito Armigeres subalbatus (Diptera: Culicidae). J. Med. Entomol. 39, 266–274. doi: 10.1603/0022-2585-39.2.266
Lambrechts, L., Paaijmans, K. P., Fansiri, T., Carrington, L. B., Kramer, L. D., Thomas, M. B., et al. (2011). Impact of daily temperature fluctuations on dengue virus transmission by Aedes aegypti. Proc. Natl. Acad. Sci. U.S.A. 108, 7460–7465. doi: 10.1073/pnas.1101377108
Lambrechts, L., Quillery, E., Noël, V., Richardson, J. H., Jarman, R. G., Scott, T. W., et al. (2013). Specificity of resistance to dengue virus isolates is associated with genotypes of the mosquito antiviral gene Dicer-2. Proc. R. Soc. B Biol. Sci. 280:20122437. doi: 10.1098/rspb.2012.2437
Lanciotti, R. S., Kosoy, O. L., Laven, J. J., Velez, J. O., Lambert, A. J., Johnson, A. J., et al. (2008). Genetic and serologic properties of Zika virus associated with an epidemic. Yap State, Micronesia, 2007. Emerg. Infect. Dis. 14, 1232–1239. doi: 10.3201/eid1408.080287
Luckhart, S., Vodovotz, Y., Cui, L., and Rosenberg, R. (1998). The mosquito Anopheles stephensi limits malaria parasite development with inducible synthesis of nitric oxide. Proc. Natl. Acad. Sci. U.S.A. 95, 5700–5705.
Maere, S., Heymans, K., and Kuiper, M. (2005). BiNGO: a Cytoscape plugin to assess overrepresentation of Gene Ontology categories in Biological Networks. Bioinformatics 21, 3448–3449. doi: 10.1093/bioinformatics/bti551
Mccarthy, F. M., Wang, N., Magee, G. B., Nanduri, B., Lawrence, M. L., Camon, E. B., et al. (2006). AgBase: a functional genomics resource for agriculture. BMC Genomics 7:229. doi: 10.1186/1471-2164-7-229
Misslin, R., Telle, O., Daudé, E., Vaguet, A., and Paul, R. E. (2016). Urban climate versus global climate change—what makes the difference for dengue? Ann. N. Y. Acad. Sci. 1382, 56–72. doi: 10.1111/nyas.13084
Molina-Cruz, A., DeJong, R. J., Charles, B., Gupta, L., Kumar, S., Jaramillo-Gutierrez, G., et al. (2008). Reactive oxygen species modulate Anopheles gambiae immunity against bacteria and Plasmodium. J. Biol. Chem. 283, 3217–3223. doi: 10.1074/jbc.M705873200
Mordecai, E. A., Caldwell, J. M., Grossman, M. K., Lippi, C. A., Johnson, L. R., Neira, M., et al. (2019). Thermal biology of mosquito-borne disease. Ecol. Lett. 22, 1690–1708. doi: 10.1111/ele.13335
Mordecai, E. A., Cohen, J. M., Evans, M. V., Gudapati, P., Johnson, L. R., Lippi, C. A., et al. (2017). Detecting the impact of temperature on transmission of Zika, dengue, and chikungunya using mechanistic models. PLoS Negl. Trop. Dis. 11:e0005568. doi: 10.1371/journal.pntd.0005568
Mordecai, E. A., Paaijmans, K. P., Johnson, L. R., Balzer, C., Ben-Horin, T., de Moor, E., et al. (2013). Optimal temperature for malaria transmission is dramatically lower than previously predicted. Ecol. Lett. 16, 22–30. doi: 10.1111/ele.12015
Morlais, I., Mori, A., Schneider, J. R., and Severson, D. W. (2003). A targeted approach to the identification of candidate genes determining susceptibility to Plasmodium gallinaceum in Aedes aegypti. Mol. Genet. Genomics 269, 753–764. doi: 10.1007/s00438-003-0882-887
Murdock, C. C., Blanford, S., Hughes, G. L., Rasgon, J. L., and Thomas, M. B. (2014). Temperature alters Plasmodium blocking by Wolbachia. Sci. Rep. 4:3932. doi: 10.1038/srep03932
Murdock, C. C., Cox-foster, D., Read, A. F., and Thomas, M. B. (2012a). Rethinking vector immunology: the role of environmental temperature in shaping resistance. Nat. Rev. Microbiol. 10, 869–876. doi: 10.1038/nrmicro2900
Murdock, C. C., Paaijmans, K. P., Bells, A. S., King, J. G., Hillyer, J. F., Read, A. F., et al. (2012b). Complex effects of temperature on mosquito immune function. Proc. R. Soc. B Biol. Sci. 279, 3357–3366. doi: 10.1098/rspb.2012.0638
Murdock, C. C., Evans, M. V., McClanahan, T. D., Miazgowicz, K. L., and Tesla, B. (2017). Fine-scale variation in microclimate across an urban landscape shapes variation in mosquito population dynamics and the potential of Aedes albopictus to transmit arboviral disease. PLoS Negl. Trop. Dis. 11:e5640. doi: 10.1371/journal.pntd.0005640
Murdock, C. C., Moller-Jacobs, L. L., and Thomas, M. B. (2013). Complex environmental drivers of immunity and resistance in malaria mosquitoes. Proc. R. Soc. B 280:20132030. doi: 10.1098/rspb.2013.2030
Muturi, E. J., and Alto, B. W. (2011). Larval environmental temperature and insecticide exposure alter Aedes aegypti competence for arboviruses. Vector-Borne Zoonotic Dis. 11, 1157–1163. doi: 10.1089/vbz.2010.0209
Muturi, E. J., Kim, C. H., Alto, B. W., Berenbaum, M. R., and Schuler, M. A. (2011a). Larval environmental stress alters Aedes aegypti competence for Sindbis virus. Trop. Med. Int. Heal. 16, 955–964. doi: 10.1111/j.1365-3156.2011.02796.x
Muturi, E. J., Lampman, R., Costanzo, K., and Alto, B. W. (2011b). Effect of temperature and insecticide stress on life-history traits of Culex restuans and Aedes albopictus (Diptera: Culicidae). J. Med. Entomol. 48, 243–250. doi: 10.1603/me10017
Neill, K. O., Huang, N., Unis, D., and Clem, R. J. (2015). Rapid selection against arbovirus-induced apoptosis during infection of a mosquito vector. Proc. Natl. Acad. Sci. U.S.A. 112, E1152–E1161. doi: 10.1073/pnas.1424469112
Okech, B. A., Gouagna, L. C., Yan, G., Githure, J. I., and Beier, J. C. (2007). Larval habitats of Anopheles gambiae s.s. (Diptera: Culicidae) influences vector competence to Plasmodium falciparum parasites. Malar. J. 6:50. doi: 10.1186/1475-2875-6-50
Oliveira, G., de, A., Lieberman, J., and Barillas-Mury, C. (2012). Epithelial Nitration by a Peroxidase/NOX5 System Mediates Mosquito Antiplasmodial Immunity. Science 335, 856–859. doi: 10.1126/science.1209678
Oliveira, J. H. M., Gonçalves, R. L. S., Lara, F. A., Dias, F. A., Gandara, A. C. P., Menna-Barreto, R. F. S., et al. (2011). Blood meal-derived heme decreases ROS levels in the midgut of Aedes aegypti and allows proliferation of intestinal microbiota. PLoS Pathog. 7:e1001320. doi: 10.1371/journal.ppat.1001320
Pan, X., Zhou, G., Wu, J., Bian, G., Lu, P., Raikhel, A. S., et al. (2012). Wolbachia induces reactive oxygen species (ROS)-dependent activation of the Toll pathway to control dengue virus in the mosquito Aedes aegypti. Proc. Natl. Acad. Sci. U.S.A. 109, E23–E31. doi: 10.1073/pnas.1116932108
Parham, P. E., and Michael, E. (2010). Modeling the effects of weather and climate change on malaria transmission. Environ. Health Perspect. 118, 620–626. doi: 10.1289/ehp.0901256
Parham, P. E., Waldock, J., Christophides, G. K., Hemming, D., Agusto, F., Evans, K. J., et al. (2015). Climate, environmental and socio-economic change: weighing up the balance in vector-borne disease transmission. Philos. Trans. R. Soc. B Biol. Sci. 370:20130551. doi: 10.1098/rstb.2013.0551
Ramirez, J. L., Souza-Neto, J., Cosme, R. T., Rovira, J., Ortiz, A., Pascale, J. M., et al. (2012). Reciprocal tripartite interactions between the Aedes aegypti midgut microbiota, innate immune system and dengue virus influences vector competence. PLoS Negl. Trop. Dis. 6:e1561. doi: 10.1371/journal.pntd.0001561
Rodgers, F. H., Gendrin, M., Wyer, C. A. S., and Christophides, G. K. (2017). Microbiota-induced peritrophic matrix regulates midgut homeostasis and prevents systemic infection of malaria vector mosquitoes. PLoS Pathog. 13:e1006391. doi: 10.1371/journal.ppat.1006391
Rodriguez-Andres, J., Rani, S., Varjak, M., Chase-Topping, M. E., Beck, M. H., Ferguson, M. C., et al. (2012). Phenoloxidase activity acts as a mosquito innate immune response against infection with semliki Forest Virus. PLoS Pathog. 8:e1002977. doi: 10.1371/journal.ppat.1002977
Ryan, S. J., Carlson, C. J., Mordecai, E. A., and Johnson, L. R. (2018). Global expansion and redistribution of Aedes-borne virus transmission risk with climate change. PLoS Negl. Trop. Dis. 13:e7213. doi: 10.1371/journal.pntd.0007213
Ryan, S. J., Mcnally, A., Johnson, L. R., Mordecai, E. A., Ben-Horin, T., Paaijmans, K., et al. (2015). Mapping physiological suitability limits for malaria in Africa under climate change. Vector-Borne Zoonotic Dis. 15, 718–725. doi: 10.1089/vbz.2015.1822
Saldaña, M. A., Etebari, K., Hart, C. E., Widen, S. G., Wood, T. G., Thangamani, S., et al. (2017a). Zika virus alters the microRNA expression profile and elicits an RNAi response in Aedes aegypti mosquitoes. PLoS Negl. Trop. Dis. 11:e0005760. doi: 10.1371/journal.pntd.0005760
Saldaña, M. A., Hegde, S., and Hughes, G. L. (2017b). Microbial control of arthropod-borne disease. Mem. Inst. Oswaldo Cruz 112, 81–93. doi: 10.1590/0074-02760160373
Sánchez-Vargas, I., Scott, J. C., Poole-Smith, B. K., Franz, A. W. E., Barbosa-Solomieu, V., Wilusz, J., et al. (2009). Dengue virus type 2 infections of Aedes aegypti are modulated by the mosquito’s RNA interference pathway. PLoS Pathog. 5:e1000299. doi: 10.1371/journal.ppat.1000299
Sanders, H. R., Evans, A. M., Ross, L. S., and Gill, S. S. (2003). Blood meal induces global changes in midgut gene expression in the disease vector, Aedes aegypti. Insect Biochem. Mol. Biol. 33, 1105–1122. doi: 10.1016/S0965-1748(03)00124-3
Sanders, H. R., Foy, B. D., Evans, A. M., Ross, L. S., Beaty, B. J., Olson, K. E., et al. (2005). Sindbis virus induces transport processes and alters expression of innate immunity pathway genes in the midgut of the disease vector, Aedes aegypti. Insect Biochem. Mol. Biol. 35, 1293–1307. doi: 10.1016/j.ibmb.2005.07.006
Santamaría, M. E., González-Cabrera, J., Martínez, M., Grbic, V., Castañera, P., Díaz, I., et al. (2015). Digestive proteases in bodies and faeces of the two-spotted spider mite, Tetranychus urticae. J. Insect Physiol. 78, 69–77. doi: 10.1016/j.jinsphys.2015.05.002
Saraiva, R. G., Kang, S., Simões, M. L., Angleró-Rodríguez, Y. I., and Dimopoulos, G. (2016). Mosquito gut antiparasitic and antiviral immunity. Dev. Comp. Immunol. 64, 53–64. doi: 10.1016/j.dci.2016.01.015
Seehuus, S.-C., Norberg, K., Gimsa, U., Krekling, T., and Amdam, G. V. (2006). Reproductive protein protects functionally sterile honey bee workers from oxidative stress. PNAS 103, 962–967. doi: 10.1073/pnas.0502681103
Shaw, W. R., and Catteruccia, F. (2019). Vector biology meets disease control: using basic research to fight vector-borne diseases. Nat. Microbiol. 4, 20–34. doi: 10.1038/s41564-018-0214-7
Shibata, T., Maki, K., Hadano, J., Fujikawa, T., Kitazaki, K., Koshiba, T., et al. (2015). Crosslinking of a peritrophic matrix protein protects gut epithelia from bacterial exotoxins. PLoS Pathog. 11:e5244. doi: 10.1371/journal.ppat.1005244
Shocket, M. S., Ryan, S. J., and Mordecai, E. A. (2018). Temperature explains broad patterns of Ross River virus transmission. eLife 7:e37762. doi: 10.7554/eLife.37762
Shragai, T., Tesla, B., Murdock, C., and Harrington, L. C. (2017). Zika and chikungunya: mosquito-borne viruses in a changing world. Ann. N. Y. Acad. Sci. 1399, 61–77. doi: 10.1111/nyas.13306
Sim, S., and Dimopoulos, G. (2010). Dengue virus inhibits immune responses in Aedes aegypti cells. PLoS One 5:e10678. doi: 10.1371/journal.pone.0010678
Simões, M. L., Caragata, E. P., and Dimopoulos, G. (2018). Diverse host and restriction factors regulate mosquito–pathogen interactions. Trends Parasitol. 34, 603–616. doi: 10.1016/j.pt.2018.04.011
Siraj, A. S., Rodriguez-Barraquer, I., Barker, C. M., Tejedor-Garavito, N., Harding, D., Lorton, C., et al. (2018). Spatiotemporal incidence of Zika and associated environmental drivers for the 2015-2016 epidemic in Colombia Amir. Sci. Data. 5:180073. doi: 10.1038/sdata.2018.73
Siraj, A. S., Santos-Vega, M., Bouma, M. J., Yadeta, D., Ruiz Carrascal, D., and Pascual, M. (2014). Altitudinal changes in malaria incidence in highlands of Ethiopia and Colombia. Science 343, 1154–1158. doi: 10.1126/science.1244325
Souza-Neto, J. A., Sim, S., and Dimopoulos, G. (2009). An evolutionary conserved function of the JAK-STAT pathway in anti-dengue defense. Proc. Natl. Acad. Sci. U.S.A. 106, 17841–17846. doi: 10.1073/pnas.0905006106
Suwanchaichinda, C., and Paskewitz, S. M. (1998). Effects of larval nutrition, adult body size, and adult temperature on the ability of Anopheles gambiae (Diptera: Culicidae) to Melanize Sephadex beads. J. Med. Entomol. 35, 157–161.
Szklarczyk, D., Morris, J. H., Cook, H., Kuhn, M., Wyder, S., Simonovic, M., et al. (2017). The STRING database in 2017: quality-controlled protein – protein association networks, made broadly accessible. Nucleic Acids Res. 45, 362–368. doi: 10.1093/nar/gkw937
Taguwa, S., Maringer, K., Li, X., Bernal-Rubio, D., Rauch, J. N., Gestwicki, J. E., et al. (2015). Defining Hsp70 subnetworks in dengue virus replication reveals key vulnerability in flavivirus infection. Cell 163, 1108–1123. doi: 10.1016/j.cell.2015.10.046
Tchankouo-Nguetcheu, S., Khun, H., Pincet, L., Roux, P., Bahut, M., Huerre, M., et al. (2010). Differential protein modulation in midguts of Aedes aegypti infected with chikungunya and dengue 2 viruses. PLoS One 5:e13149. doi: 10.1371/journal.pone.0013149
R Development Core Team (2008). R: A Language and Environment for Statistical Computing. Vienna: R Foundation Statics Computing, doi: 10.1007/978-3-540-74686-7
Tesla, B., Demakovsky, L. R., Mordecai, E. A., Ryan, S. J., Bonds, M. H., Ngonghala, C. N., et al. (2018a). Temperature drives Zika virus transmission: evidence from empirical and mathematical models. Proc. R. Soc. B Biol. Sci. 285:20180795. doi: 10.1098/rspb.2018.0795
Tesla, B., Demakovsky, L. R., Packiam, H. S., Mordecai, E. A., Rodríguez, A. D., Bonds, M. H., et al. (2018b). Estimating the effects of variation in viremia on mosquito susceptibility, infectiousness, and R0 of Zika in Aedes aegypti. PLoS Negl. Trop. Dis. 12:e0006733. doi: 10.1371/journal.pntd.0006733
Trapnell, C., Pachter, L., and Salzberg, S. L. (2009). TopHat: discovering splice junctions with RNA-Seq. Bioinformatics 25, 1105–1111. doi: 10.1093/bioinformatics/btp120
Trapnell, C., Roberts, A., Goff, L., Pertea, G., Kim, D., Kelley, D. R., et al. (2013). Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 7, 562–578. doi: 10.1038/nprot.2012.016
Vazeille, M., Madec, Y., Mousson, L., Bellone, R., Barré-Cardi, H., Sousa, C. A., et al. (2019). Zika virus threshold determines transmission by European Aedes albopictus mosquitoes. Emerg. Microbes Infect. 8, 1668–1678. doi: 10.1080/22221751.2019.1689797
Wang, H., Gort, T., Boyle, D. L., and Clem, R. J. (2012). Effects of manipulating apoptosis on sindbis virus infection of Aedes aegypti mosquitoes. J. Virol. 86, 6546–6554. doi: 10.1128/jvi.00125-12
Wang, P., and Granados, R. R. (2000). Calcofluor disrupts the midgut defense system in insects. Insect Biochem. Mol. Biol. 30, 135–143. doi: 10.1016/S0965-1748(99)00108-3
Wang, P., Li, G., and Granados, R. R. (2004). Identification of two new peritrophic membrane proteins from larval Trichoplusia ni: structural characteristics and their functions in the protease rich insect gut. Insect Biochem. Mol. Biol. 34, 215–227. doi: 10.1016/j.ibmb.2003.10.001
Watts, D. M., Burke, D. S., Harrison, B. A., Whitmire, R. E., and Nisalak, A. (1987). Effect of Temperature on the Vector Efficiency of Aedes aegypti for Dengue 2 Virus. Am. J. Trop. Med. Hyg. 36, 143–152.
Weiss, B. L., Savage, A. F., Griffith, B. C., Wu, Y., and Aksoy, S. (2014). The peritrophic matrix mediates differential infection outcomes in the tsetse fly gut following challenge with commensal, pathogenic, and parasitic microbes. J. Immunol. 193, 773–782. doi: 10.4049/jimmunol.1400163
Wilke, A. B. B., and Marrelli, M. T. (2015). Paratransgenesis: a promising new strategy for mosquito vector control. Parasit. Vectors 8:342. doi: 10.1186/s13071-015-0959-2
Willard, K. A., Demakovsky, L., Tesla, B., Goodfellow, F. T., Stice, S. L., Murdock, C. C., et al. (2017). Zika virus exhibits lineage-specific phenotypes in cell culture, in Aedes aegypti mosquitoes, and in an embryo model. Viruses 9:E383. doi: 10.3390/v9120383
Willard, K. A., Elling, C. L., Stice, S. L., and Brindley, M. A. (2019). The oxysterol 7-ketocholesterol reduces zika virus titers in vero cells and human neurons. Viruses 11, 1–17. doi: 10.3390/v11010020
Wong, Z. S., Brownlie, J. C., and Johnson, K. N. (2015). Oxidative stress correlates with Wolbachia-mediated antiviral protection in Wolbachia-Drosophila associations. Appl. Environ. Microbiol. 81, 3001–3005. doi: 10.1128/AEM.03847-14
Xi, Z., Ramirez, J. L., and Dimopoulos, G. (2008). The Aedes aegypti toll pathway controls dengue virus infection. PLoS Pathog. 4:e1000098. doi: 10.1371/journal.ppat.1000098
Xiao, F. Z., Zhang, Y., Deng, Y. Q., He, S., Xie, H. G., Zhou, X. N., et al. (2014). The effect of temperature on the extrinsic incubation period and infection rate of dengue virus serotype 2 infection in Aedes albopictus. Arch. Virol. 159, 3053–3057. doi: 10.1007/s00705-014-2051-1
Yen, P.-S., James, A., Li, J.-C., Chen, C.-H., and Failloux, A.-B. (2018). Synthetic miRNAs induce dual arboviral-resistance phenotypes in the vector mosquito Aedes aegypti. Commun. Biol. 1:11. doi: 10.1038/s42003-017-0011-5
Yu, X. Q., and Kanost, M. R. (2000). Immulectin-2, a lipopolysaccharide-specific lectin from an insect, Manduca sexta, is induced in response to Gram-negative bacteria. J. Biol. Chem. 275, 37373–37381. doi: 10.1074/jbc.M003021200
Zouache, K., Fontaine, A., Vega-Rua, A., Mousson, L., Thiberge, J. M., Lourenco-De-Oliveira, R., et al. (2014). Three-way interactions between mosquito population, viral strain and temperature underlying chikungunya virus transmission potential. Proc. R. Soc. B Biol. Sci. 281:20141078. doi: 10.1098/rspb.2014.1078
Keywords: temperature, Aedes aegypti, Zika virus, RNA-seq, transcriptome, immune response
Citation: Ferreira PG, Tesla B, Horácio ECA, Nahum LA, Brindley MA, de Oliveira Mendes TA and Murdock CC (2020) Temperature Dramatically Shapes Mosquito Gene Expression With Consequences for Mosquito–Zika Virus Interactions. Front. Microbiol. 11:901. doi: 10.3389/fmicb.2020.00901
Received: 27 September 2019; Accepted: 16 April 2020;
Published: 12 June 2020.
Edited by:
Declan C. Schroeder, University of Minnesota, United StatesReviewed by:
Tadahisa Teramoto, Georgetown University, United StatesAnna-Bella Failloux, Institut Pasteur, France
Copyright © 2020 Ferreira, Tesla, Horácio, Nahum, Brindley, de Oliveira Mendes and Murdock. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Melinda Ann Brindley, bWJyaW5kbGVAdWdhLmVkdQ==; Tiago Antônio de Oliveira Mendes, dGlhZ29hb21lbmRlc0B1ZnYuYnI=; Courtney Cuinn Murdock, Y2NtMjU2QGNvcm5lbGwuZWR1