 Laura Trovato1,2*
Laura Trovato1,2* Salvatore Oliveri1,2Maria Carmela Esposto3
Salvatore Oliveri1,2Maria Carmela Esposto3 Anna Prigitano3Luisa Romanò3
Anna Prigitano3Luisa Romanò3 Massimo Cogliati3
Massimo Cogliati3- 1U.O.C. Laboratory Analysis Unit, A.O.U. “Policlinico-Vittorio Emanuele”, Catania, Italy
- 2Department of Biomedical and Biotechnological Sciences, University of Catania, Catania, Italy
- 3Department of Biomedical Sciences for Health, Università degli Studi di Milano, Milan, Italy
This study investigated the presence of Cryptococcus neoformans and Cryptococcus gattii species complex isolates on olive trees growing in the Eastern part of Sicily (Italy) characterized by the presence of the volcano Etna and the ability of these fungal pathogens to sexually reproduce on medium containing volcanic soil. Samples from 124 olive trees were collected from 14 different sites around Mount Etna. Eighteen trees (14.5%) resulted colonized by C. neoformans VNI-αA isolates, one (0.8%) by VNIV-αD isolates, and two (1.6%) by C. gattii VGI-αB isolates. The ability of environmental and reference strains belonging to VNI, VNIV, and VGI molecular types to sexually reproduce on a medium containing volcanic soil was also tested. VNI and VNIV strains were able to produce filaments and basiodiospores more vigorously than on the control medium, whereas VGI strains were not fertile. In conclusion, the present study identified which C. neoformans and C. gattii species complex genotypes are circulating in Eastern Sicily and confirmed the ecological role of olive trees as environmental reservoir of these pathogens. It also showed that Cryptococcus is able to colonize and sexually reproduce in inhospitable environments such as the slopes of a volcano.
Introduction
Cryptococcosis is a pulmonary or disseminated infection caused by the encapsulated yeasts belonging to Cryptococcus neoformans and Cryptococcus gattii species complexes. Immunosuppression is the major predisposing condition involved in the development of cryptococcosis, which mainly affects HIV-infected patients, organ transplant recipients, patients with autoimmune diseases and those receiving corticosteroid or immunosuppressive therapies (Rajasingham et al., 2017; Henao-Martínez et al., 2018). Cryptococcosis is acquired from the environment by inhalation of basidiospores or small blastospores that reach the pulmonary alveoli and may frequently disseminate to meninges causing fatal meningitis or meningo-encephalitis (Kwon-Chung et al., 2014). Differences in biology, epidemiology, pathogenicity, clinical manifestations, and drug susceptibility have been observed between the two species complexes (Kwon-Chung and Varma, 2006; Lee et al., 2019). Infections caused by yeasts of the C. neoformans species complex, belonging to molecular types VNI, VNII, VNB, VNIII or VNIV, are primarily associated with immunosuppression (Kwon-Chung et al., 2014), whereas some yeasts of the C. gattii species complex (molecular types VGI and VGII) frequently infect immunocompetent hosts (Chen et al., 2014). Among each molecular type two mating type alleles can be distinguished, mating type a and mating type a, being the former more frequent than the latter (Heitman et al., 2011).
C. neoformans VNI is the most prevalent molecular type worldwide and has been recovered from bird excreta, soil, and trunk hollows of different tree species, suggesting that trees play a major role as reservoir in the environment (Costa et al., 2010; Chowdhary et al., 2012b; Colom et al., 2012; Cattana et al., 2014; Noguera et al., 2015; Cogliati et al., 2016a; Ellabib et al., 2016; Dou et al., 2017). In contrast, C. neoformans VNIV is mostly distributed in Europe, and less frequently in North and South America and Japan (Cogliati, 2013). VGI is the prevalent molecular type among the C. gattii species complex, which is mainly distributed in tropical and subtropical regions as well as in some geographical area with temperate climate such as the North Pacific coast of North America and Europe (Byrnes and Marr, 2011; Chowdhary et al., 2012a; Hagen et al., 2012; Springer et al., 2014; Cogliati et al., 2016a; Acheson et al., 2018). In addition, particular interest is recently addressed on C. gattii VGII which is causing outbreaks in British Columbia (Canada), North Pacific coast of United States, Brazil and Colombia (Lockhart et al., 2016; Acheson et al., 2018; Firacative et al., 2019; Maruyama et al., 2019).
A recent environmental survey carried out in Europe and the Mediterranean area showed the presence of both C. neoformans and C. gattii species complexes in the environment and that some trees, such as olive trees and carob trees, presented a rate of colonization higher than others (Cogliati et al., 2016a). Furthermore, a niche modeling study considering the European climatic conditions suggested that the potential areas of distribution of the two species complexes is often overlapping and that therefore, in Europe yeasts with different molecular types can frequently interact each with the other (Cogliati et al., 2017).
In Sicily, an island of Southern Italy, C. neoformans and C. gattii species complex isolates were previously recovered from Messina (Northeast Sicily) in several samples of bird excreta as well as in Eucalyptus camaldulensis, Prunus dulcis (almond), and Ceratonia siliqua (carob) (Criseo and Gallo, 1997; Pernice et al., 1998; Romeo et al., 2012). However, olive trees were not sampled during these environmental surveys. Eastern Sicily is characterized by the presence of Mount Etna, the largest volcano in Europe that reaches an altitude of 3350 m.a.s.l. Volcanic origin of soil, difference in altitudes, and exposure of the different sides of the mountain make unique the biotope of Mount Etna, which is very different from the rest of Sicily characterized by a Mediterranean climate.
The aim of this study, therefore, was to evaluate the distribution and molecular characterization of C. neoformans and C. gattii species complex isolates in Eastern Sicily, and the ability of these fungal pathogens to sexually reproduce on medium containing volcanic soil.
Materials and Methods
Environmental Sampling and Processing
In the present study olive trees growing at different altitude in urban and rural areas around Mount Etna were investigated. The environmental survey was carried out in October–November 2014 and April–May 2015. Samples were collected by rubbing the inner of hollows and fissures of olive trees with a sterile cotton-tipped swab moistened in a solution of sterile distilled water supplemented with chloramphenicol (10 mg/L), and processed and cultured on Niger seed agar as previously reported (Pham et al., 2014; Cogliati et al., 2016a). All brown colonies grown on the plates were isolated and species was identified by microscopy for assessing yeast morphology and capsule presence, and by testing urease activity, ability to grow at 37°C, and to assimilate myo-inositol as a carbon source. Isolates belonging to C. neoformans or C. gattii species complex were differentiated by inoculating a fresh colony onto L-canavanine-glycine-bromothymol blue (CGB) agar prepared as described elsewhere (Shadomy et al., 1987).
Molecular Typing
Genomic DNA was extracted as described by Viviani et al. (1997). Molecular type and mating type of all isolates was determined by four multiplex PCRs specific for both C. neoformans and C. gattii species complexes as previously described (Cogliati et al., 2000, 2015; Esposto et al., 2004; Feng et al., 2013). Molecular and mating types were assigned according to the standard nomenclature of the ISHAM Working Group for genotyping of C. neoformans and C. gattii (Meyer et al., 2009). Strains H99 (VNI-αA), JEC20 (VNIV-αD), JEC21 (VNIV-αD), IUM 96–2828 (VNII-αA), WM626 (VNII-αA), WM779 (VGIV-αC), NIH312 (VGIII-αB), NIH191 (VGIII-αC), WM201 (VGI-αB), and IUM 00–5363 (VGII-αB) were used as reference strains. Multi-locus sequence typing (MLST) according to the ISHAM scheme (Meyer et al., 2009) was performed for C. gattii species complex isolates. Sequence types (ST) were assigned comparing the MLST profile with those present in the C. gattii MLST database1.
Mating Assay
For the mating assay a volcanic soil medium (VSM) was prepared suspending 50 g of Etna volcanic soil in 1 L of distilled water. The suspension was boiled for at least 30 min and then filtered through gauze. Volume was adjusted with further distilled water and 20 g of agar were added. The medium was sterilized by autoclave and then poured in 90-mm Petri dishes. For sugar volcanic soil medium (SVSM), 0.5 g of glucose were added. Each plate containing VSM and SVSM was inoculated with a mix of two strains of opposite mating type as well as with the two strains independently to exclude haploid fruiting. Cultures were incubated at 25°C in the dark for at least 3–4 weeks and checked once a week for production of hyphae and basidiospores. One plate containing the control medium Murashige and Skoog agar (Sigma-Aldrich, Milano, Italy) was also inoculated as described above in order to test if the two strains were actually able to mate. Mating assays were performed as reported in Table 1. JEC20 (VNIV-αD), JEC21 (VNIV-αD), SPCCON11HO1-1 (VNIV-αA), and SPCCON11HO1-2 (VNI-αA) were used as reference tester strains, and strain ITBRF221HO1-5 (VGI-αB), isolated from Southern Italy (Montagna et al., 2018), was used for the mating assays with the VGI isolates from Sicily.
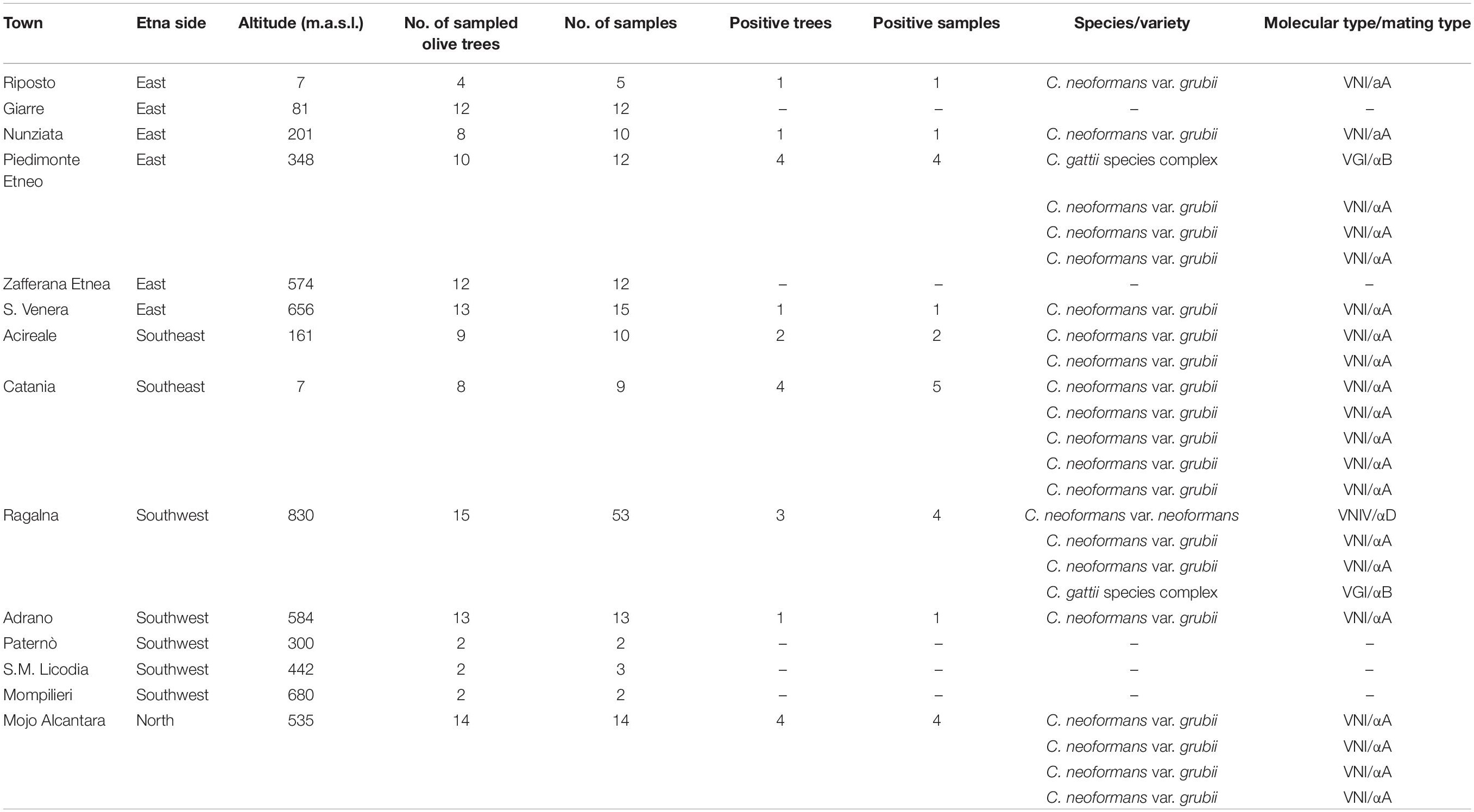
Table 1. Results of environmental samplings performed on the olive trees growing around Mount Etna.
Statistics
Data were analyzed using the MedCalc Statistical Software version 17.9.2 (MedCalc Software bvba, Ostend, Belgium; 2017)2. Percentage of positive samples from North, East, Southeast, Southwest sides of Mount Etna as well as comparison of positive samples at different altitude were compared using the two-tailed χ2 test.
Results
The geographical distribution of environmental samplings is shown in Figure 1 and the results are reported in Table 1. One hundred and seventy-two samples were collected from 124 olive trees. A total of 21 isolates from 19 olive trees (15.3%) were identified as C. neoformans species complex whereas only two trees (1.6%), one growing on the East and the other on the Southwest side of Mount Etna, were colonized by C. gattii species complex isolates. Fourteen olive trees (14 samples) were sampled on the North, 59 (66 samples) on the East, 17 (19 samples) on the Southeast, and 34 (73 samples) on the Southwest side of Mount Etna, and the rate of positive trees was 28.6% (4/14), 11.9% (7/59), 35.3% (6/17), and 11.8% (4/34), respectively. Statistical analysis showed no significant difference between the four sides of Mount Etna, although the percentage of positive trees was higher in the North and the Southeast sides despite the number of specimens sampled was lower. Considering the altitude threshold value of 400 m.a.s.l., statistical analysis showed no significant difference of percentage of positive trees at different altitude, although it was higher at altitudes below 400 meters (22.6%, 12/53 vs. 12.7%, 9/71). Among the 21 isolates of the C. neoformans species complex, twenty were C. neoformans var. grubii belonging to molecular type VNI, mating type aA and one was C. neoformans var. neoformans, VNIV, mating type aD. The remaining two isolates were C. gattii, VGI, mating type aB. MLST analysis showed that both C. gattii VGI isolates belonged to ST197 which matched with the ST of other four isolates present in the MLST database: CCA320 and CCA321 environmental isolates from Barcelona (Spain), and CBS6290 and IHEM11792 clinical isolates from Kinshasa (Congo Republic). Results of mating assays showed that all tested strains were able to grow on both VSM and SVSM, although more vigorously on the latter one. The mating assay with the two VNIV reference tester strains (JEC21 × JEC20) produced filaments and basiodiospores on both VSM and SVSM, which showed to be a more suitable media compared to Murashige and Skoog agar (Figure 2). In addition, sporulation was also observed for the mating between the environmental VNIV isolate from Sicily (ITRU2HO2-1) and JEC20 (Figure 2). Among mating assays performed with VNI isolates only the one between the two tester strains (SPCCON11HO1-1 × SPCCON11HO1-2) resulted fertile on SVSM (Figure 2). None of the C. gattii mating assays resulted fertile after 4 weeks of incubation (Table 2).
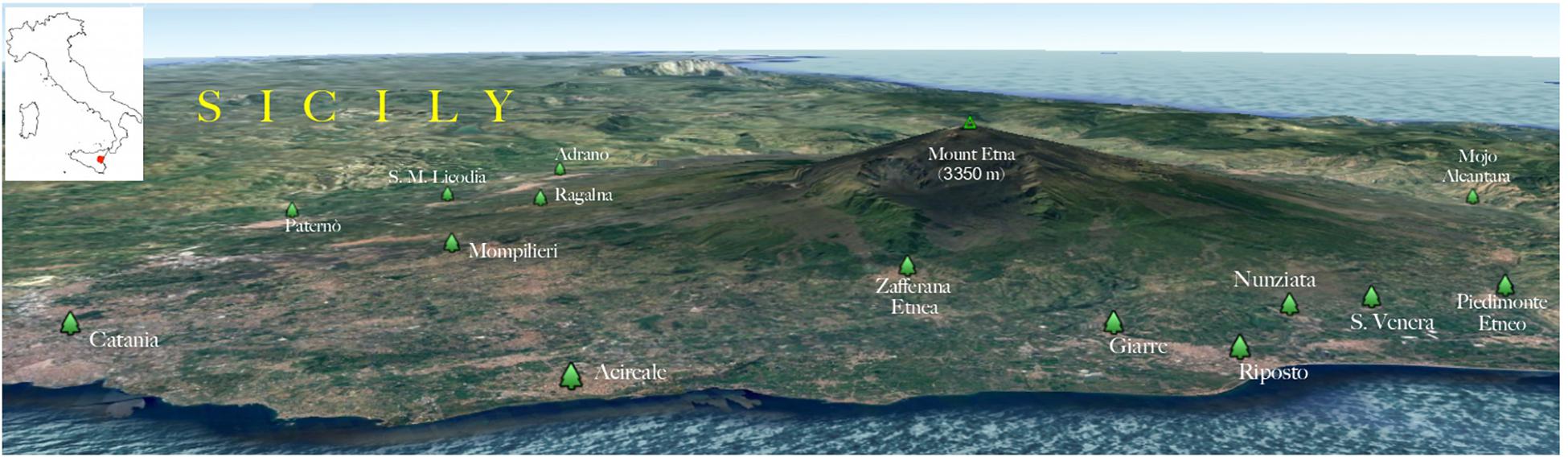
Figure 1. Three-dimensional map showing the geographical distribution of the environmental sampling sites. The insert shows where the geographical area is located in Italy (red area).
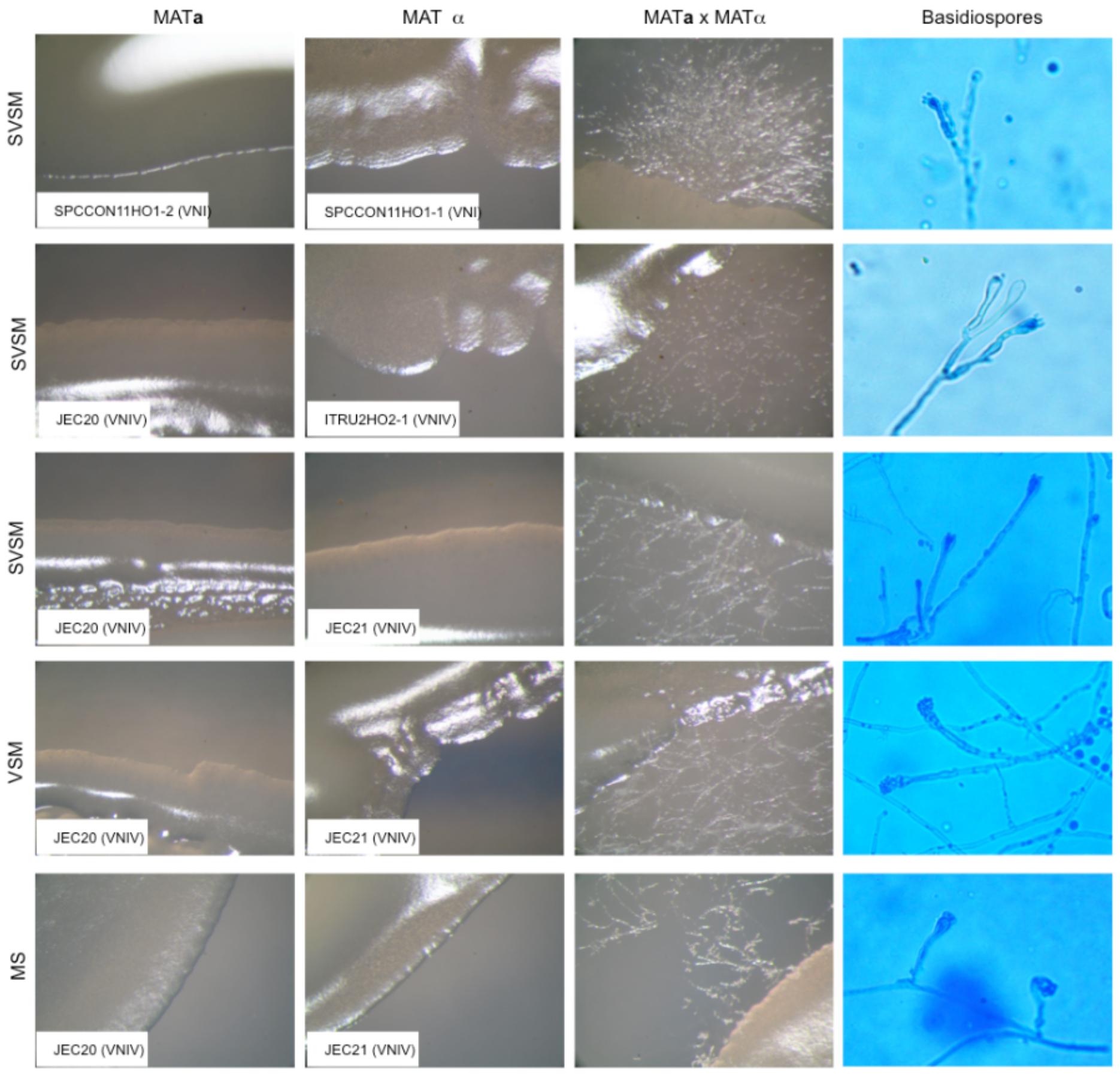
Figure 2. Results of fertile mating type assays. MATa, MATα, and MATa × MATα cultures were observed at 50× magnification with a stereomicroscope (Zeiss, Essingen, Germany). Basidiospores were observed at 400× magnification with a light microscope Axioskope 40 (Zeiss). SVSM = Sugar volcanic soil medium; VSM = Volcanic soil medium; MS = Murashige and Skoog agar.
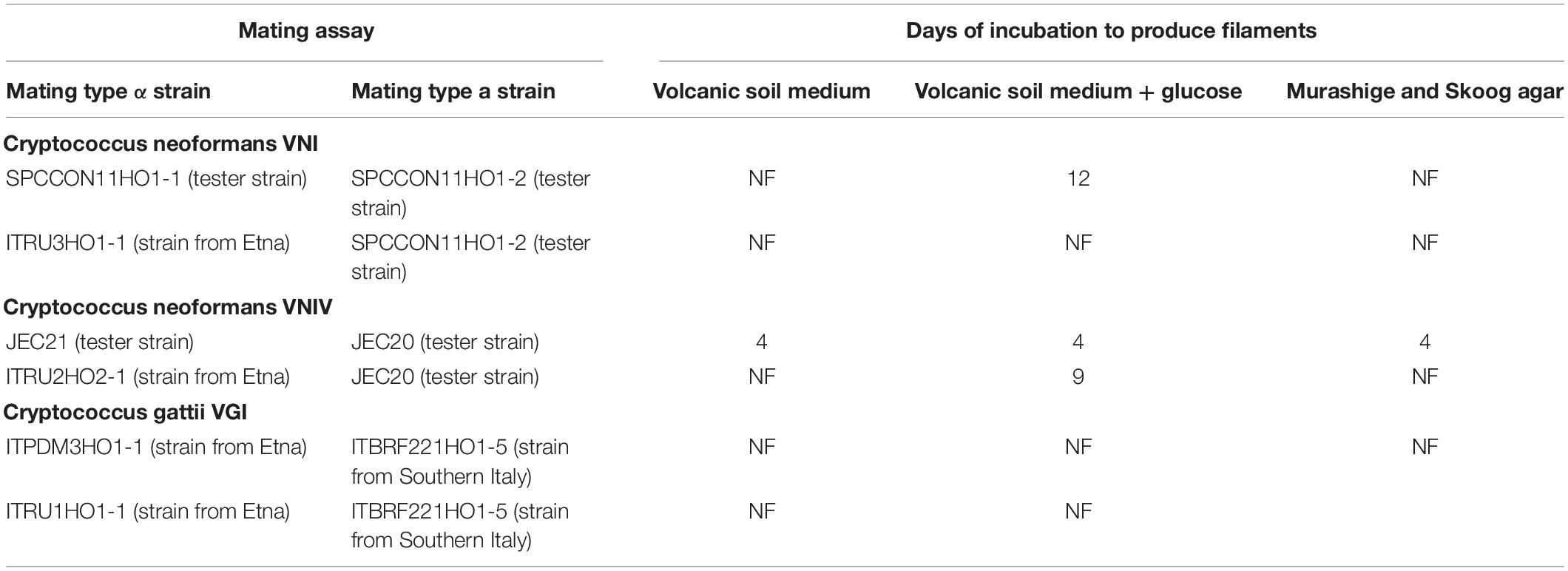
Table 2. Mating assays performed in the present study.
Discussion
This study confirms the presence of C. neoformans and C. gattii species complex on olive trees growing in Eastern Sicily on the four sides of Mount Etna with a high prevalence on North and Southeast sides. The percentage of positive olive trees was 15.3 and 1.6% for C. neoformans and C. gattii species complex, respectively, confirming the important role of these trees as an ecological niche for both species complexes. A recent environmental survey carried out in Apulia (another region of Southern Italy) reported a percentage of positive olive trees (18%) similar to that found in our study (Montagna et al., 2018). Since Sicily is the third Italian region for the production of olive oil, with more than 150,000 hectares of its territory (about 6%) dedicated to olive tree cultivation, this region represents an area with high suitability for the survival of Cryptococcus as also confirmed by a recent niche modeling study (Cogliati et al., 2017). VNI was the prevalent molecular type among isolates collected in the present study as observed in other regions of Southern Italy (Romeo et al., 2012; Cogliati et al., 2016a; Montagna et al., 2018), confirming the ubiquitous presence of this fungal pathogen. The unique isolate with molecular type VNIV was found in an olive tree on the Southwest side of Etna and in particular at an altitude of 820 m.a.s.l. This could reflect the ability of VNIV isolates to tolerate lower temperature better than VNI and C. gattii VGI isolates as previously reported (Cogliati et al., 2017). Other authors reported the isolation of this yeast in the nostrils of squirrels living in the Appenine area of Calabria (Iatta et al., 2015). All together these data confirm that VNIV isolates are able to survive in the European areas with a sub-continental climate (Cogliati et al., 2017). Both C. gattii species complex isolates belonged to VGI molecular type and ST197 genotype. This genotype was previously reported to be genetically related to several other European genotypes forming a Mediterranean cluster (Hagen et al., 2012). Therefore, the isolates recovered in Sicily during this study are probably endemic of the Mediterranean area and have been present in the environment since long time.
The mating type a was the only mating type identified among all isolates. Although mating type a isolates were not recovered during this study, future surveys could be able to detect them considering that such isolates have been already found in Southern Italy (Viviani et al., 2001; Cogliati et al., 2016a; Montagna et al., 2018). Mating assays showed that C. neoformans and C. gattii species complex strains are able to grow on media containing volcanic soil with and without addition of glucose. This means that volcanic soil is a suitable substrate for the growth of these fungi and therefore, that the peculiar characteristics of the soil on Mount Etna do not limit their survival. In addition, our results showed that C. neoformans species complex strains belonging to VNI and VNIV molecular types are able to sexually reproduce on medium containing volcanic soil, and that one environmental VNIV isolate recovered from Mount Etna was fertile. This is in agreement with the recent findings reporting that VNIV population is more recombinant than VNI population and thus sexual reproduction occurs more frequently in the former than in the latter (Cogliati et al., 2016b, 2019). In contrast, none of the tested C. gattii strains was able to mate probably due to the fact that VGI strains are the least fertile strains among C. gattii species complex and the absence of adequate tester strains (Springer et al., 2017). Several studies report the isolation of C. neoformans species complex strains from soil samples (Machado et al., 1993; Montagna et al., 2003, 2018; Cogliati, 2013; Escandón and Castañeda, 2015; Cogliati et al., 2016a) and, recently, it was also showed its ability to mate and produce basidiospores on media prepared with different soil substrates (Cogliati, 2018). However, at present, similar studies on media containing volcanic soil have not yet been carried out. The ability of this yeast to grow on the volcanic soil confirms that C. neoformans species complex strains are able to colonize a wide range of microhabitats including those more inhospitable. Volcanic soil of Mount Etna is a sub-acid substrate (pH 6.8), rich of microelements such as iron and copper, with a medium content of potassium, phosphorus and magnesium, but poor of nitrogen and calcium (D’Antone et al., 2017). These characteristics are very similar to those recommended to induce filamentation and sporulation of cryptococcal yeasts (Hsueh et al., 2011) and therefore, volcanic soil of Mount Etna results a suitable substrate for sexual reproduction of this fungal pathogens. Furthermore, the ability of cryptococcal yeasts to grow and produce blastospores and basidiospores on the volcanic soil represents a potential source of infection since soil aerosols could transfer small cells and spores in pulmonary alveoli of humans and animals causing the onset of the infection. In conclusion, this study identified some of C. neoformans and C. gattii species complex genotypes present in Eastern Sicily and confirmed the ecological role of olive trees as environmental reservoir of these pathogens. It also showed that Cryptococcus is able to colonize and sexually reproduce in inhospitable environments such as the slopes of a volcano.
Data Availability Statement
All datasets generated for this study are included in the manuscript/supplementary files.
Author Contributions
LT, SO, and MC designed the study. LT and SO performed the environmental samplings. MC, ME, AP, and LR performed the molecular typing and mating assay. LT, SO, and MC analyzed the data. LT and MC wrote the manuscript. All authors read and approved the final version of the manuscript.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We wish to thank the Scientific Bureau of the University of Catania for language support.
Footnotes
References
Acheson, E. S., Galanis, E., Bartlett, K., Mak, S., and Klinkenberg, B. (2018). Searching for clues for eighteen years: deciphering the ecological determinants of Cryptococcus gattii on Vancouver Island, British Columbia. Med. Mycol. 56, 129–144. doi: 10.1093/mmy/myx037
Byrnes, E. J. III, and Marr, K. A. (2011). The outbreak of Cryptococcus gattii in western north america: epidemiology and clinical issues. Curr. Infect. Dis. Rep. 13, 256–261. doi: 10.1007/s11908-011-0181-0
Cattana, M. E., Sosa Mde, L., Fernández, M., Rojas, F., Mangiaterra, M., and Giusiano, G. (2014). Native trees of the northeast argentine: natural hosts of the Cryptococcus neoformans-Cryptococcus gattii species complex. Rev. Iberoam. Micol. 31, 188–192. doi: 10.1016/j.riam.2013.06.005
Chen, S. C., Meyer, W., and Sorrell, T. C. (2014). Cryptococcus gattii infections. Clin. Microbiol. Rev. 27, 980–1024. doi: 10.1128/CMR.00126-13
Chowdhary, A., Randhawa, H. S., Boekhout, T., Hagen, F., Klaassen, C. H., and Meis, J. F. (2012a). Temperate climate niche for Cryptococcus gattii in Northern Europe. Emerg. Infect. Dis. 18, 172–174. doi: 10.3201/eid1801.111190
Chowdhary, A., Rhandhawa, H. S., Prakash, A., and Meis, J. F. (2012b). Environmental prevalence of Cryptococcus neoformans and Cryptococcus gattii in India: an update. Crit. Rev. Microbiol. 38, 1–16. doi: 10.3109/1040841X.2011.606426
Cogliati, M. (2013). Global molecular epidemiology of Cryptococcus neoformans and gattii: an atlas of the molecular types. Scientifica 2013:675213. doi: 10.1155/2013/675213
Cogliati, M. (2018). Soil enables mating and sporulation of Cryptococcus neoformans complex. Int. J. Fungal Genet. Biol. 1, 1–4.
Cogliati, M., Allaria, M., Tortorano, A. M., and Viviani, M. A. (2000). Genotyping Cryptococcus neoformans var. neoformans with specific primers designed from PCR-fingerprinting bands sequenced using a modified PCR-based strategy. Med. Mycol. 38, 97–103. doi: 10.1080/mmy.38.2.97.103
Cogliati, M., D’Amicis, R., and Tortorano, A. M. (2015). Cryptococcus gattii sero-mating type allelic pattern determined by multiplex PCR. Clin. Microbiol. Infect. 21:190.e1-4. doi: 10.1016/j.cmi.2014.09.003
Cogliati, M., D’Amicis, R., Zani, A., Montagna, M. T., Caggiano, G., De Giglio, O., et al. (2016a). Environmental distribution of Cryptococcus neoformans and C. gattii around the Mediterranean basin. FEMS Yeast Res. 16:fow045. doi: 10.1093/femsyr/fow045
Cogliati, M., Zani, A., Rickerts, V., McCormick, I., Desnos-Ollivier, M., Velegraki, A., et al. (2016b). Multilocus sequence typing analysis reveals that Cryptococcus neoformans var. neoformans is a recombinant population. Fungal Genet. Biol. 87, 22–29. doi: 10.1016/j.fgb.2016.01.003
Cogliati, M., Desnos-Ollivier, M., McCormick-Smith, I., Rickerts, V., Ferreira-Paim, K., Meyer, W., et al. (2019). Genotypes and population genetics of Cryptococcus neoformans and Cryptococcus gattii species complexes in Europe and the mediterranean area. Fungal Genet. Biol. 129, 16–29. doi: 10.1016/j.fgb.2019.04.001
Cogliati, M., Puccianti, E., Montagna, M. T., De Donno, A., Susever, S., Ergin, C., et al. (2017). Fundamental niche prediction of the pathogenic yeasts Cryptococcus neoformans and Cryptococcus gattii in Europe. Environ. Microbiol. 19, 4318–4325. doi: 10.1111/14622920.13915
Colom, M. F., Hagen, F., Gonzalez, A., Mellado, A., Morera, N., Linares, C., et al. (2012). Ceratonia siliqua (carob) trees as natural habitat and source of infection by Cryptococcus gattii in the Mediterranean environment. Med. Mycol. 50, 67–73. doi: 10.3109/13693786.2011.574239
Costa, A. K., Sidrim, J. J., Cordeiro, R. A., Brilhante, R. S., Monteiro, A. J., and Rocha, M. F. (2010). Urban pigeons (Columba livia) as a potential source of pathogenic yeasts: a focus on antifungal susceptibility of Cryptococcus strains in Northeast Brazil. Mycopathologia 169, 207–213. doi: 10.1007/s11046-009-9245-1
Criseo, G., and Gallo, M. (1997). Serotyping of Cryptococcus neoformans isolates from environmental and clinical sources in extreme southern Italy (Calabria and Sicily, central Mediterranean area). Mycoses 40, 95–100. doi: 10.1111/j.1439-0507.1997.tb00194.x
D’Antone, C., Punturo, R., and Vaccaro, C. (2017). Rare earth elements distribution in grapevine varieties grown on volcanic soils: an example from Mount Etna (Sicily, Italy). Environ. Assess. 189:160. doi: 10.1007/s10661-017-5878-6
Dou, H., Wang, H., Xie, S., Chen, X., Xu, Z., and Xu, Y. (2017). Molecular characterization of Cryptococcus neoformans isolated from the environment in Beijing. China Med. Mycol. 55, 737–747. doi: 10.1093/mmy/myx026
Ellabib, M. S., Aboshkiwa, M. A., Husien, W. M., D’Amicis, R., and Cogliati, M. (2016). Isolation, identification and molecular typing of Cryptococcus neoformans from pigeon droppings and other environmental sources in Tripoli. Libya. Mycopathologia 181, 603–608. doi: 10.1007/s11046-016-9996-4
Escandón, P., and Castañeda, E. (2015). Long-term survival of Cryptococcus neoformans and Cryptococcus gattii in stored environmental samples from Colombia. Rev. Iberoam. Micol. 32, 197–199. doi: 10.1016/j.riam.2014.06.007
Esposto, M. C., Cogliati, M., Tortorano, A. M., and Viviani, M. A. (2004). Determination of Cryptococcus neoformans var. neoformans mating type by multiplex PCR. Clin. Microbiol. Infect. 10, 1092–1094. doi: 10.1111/j.1469-0691.2004.00972.x
Feng, X., Fu, X., Ling, B., Wang, L., Liao, W., Pan, W., et al. (2013). Rapid differentiation of cryptic species within Cryptococcus gattii by a duplex PCR assay. J. Clin. Microbiol. 51, 3110–3112. doi: 10.1128/JCM.01455-13
Firacative, C., Torres, G., Meyer, W., and Escandón, P. (2019). Clonal dispersal of Cryptococcus gattii VGII in an endemic region of cryptococcosis in Colombia. J. Fungi 5:E32. doi: 10.3390/jof5020032
Hagen, F., Colom, M. F., Swinne, D., Tintelnot, K., Iatta, R., Montagna, M. T., et al. (2012). Autochthonous and dormant Cryptococcus gattii infections in Europe. Emerg. Infect. Dis. 18, 1618–1624. doi: 10.3201/eid1810.120068
Heitman, J., Kozel, T. R., Kwon-Chung, K. J., Perfect, J. R., and Casadevall, A. (2011). Cryptococcus: from Human Pathogen to Model Yeast. Washington, DC: ASM Press.
Henao-Martínez, A. F., Chastain, D. B., and Franco-Paredes, C. (2018). Treatment of cryptococcosis in non-HIV immunocompromised patients. Curr. Opin. Infect. Dis. 31, 278–285. doi: 10.1097/QCO.0000000000000458
Hsueh, Y. P., Lin, X., Kwon-Chung, K. J., and Heitman, J. (2011). “Sexual reproduction of Cryptococcus,” in Cryptococcus: From Human Pathogen to Model Yeast, eds J. Heitman, T. Kozel, K. J. Kwon-Chung, J. Perfect, and A. Casadevall, (Washington, DC: American Society for Microbiology Press).
Iatta, R., Immediato, D., Puttilli, M. R., Danesi, P., Passantino, G., Parisi, A., et al. (2015). Cryptococcus neoformans in the respiratory tract of squirrels, Callosciurus finlaysonii (Rodentia, Sciuridae). Med. Mycol. 53, 666–673. doi: 10.1093/mmy/myv045
Kwon-Chung, K. J., Fraser, J. A., Doering, T. L., Wang, Z., Janbon, G., Idnurm, A., et al. (2014). Cryptococcus neoformans and Cryptococcus gattii, the etiologic agents of cryptococcosis. Cold Spring Harb. Perspect. Med. 4:a019760. doi: 10.1101/cshperspect.a019760
Kwon-Chung, K. J., and Varma, A. (2006). Do major species concepts support one, two or more species within Cryptococcus neoformans? FEMS Yeast Res. 6, 574–587. doi: 10.1111/j.1567-1364.2006.00088.x
Lee, G. A., Arthur, I., Merritt, A., and Leung, M. (2019). Molecular types of Cryptococcus neoformans and Cryptococcus gattii in Western Australia and correlation with antifungal susceptibility. Med. Mycol. 57, 1004–1010. doi: 10.1093/mmy/myy161
Lockhart, S. R., McCotter, O. Z., and Chiller, T. M. (2016). Emerging fungal infections in the Pacific Northwest: the unrecognized burden and geographic range of Cryptococcus gattii and Coccidioides immitis. Microbiol. Spectr. 4:EI10–EI0016. doi: 10.1128/microbiolspec.EI10-0016-2016
Machado, C. C., Amaral, A. A., and Severo, L. C. (1993). Cryptococcus neoformans var. neoformans isolated from soil. Rev Inst. Med. Trop. Sao. Paulo. 35, 77–79. doi: 10.1590/s0036-46651993000100011
Maruyama, F. H., de Paula, D. A. J., Menezes, I. G., Favalessa, O. C., Hahn, R. C., de Almeida, A. D. B. P. F., et al. (2019). Genetic diversity of the Cryptococcus gattii species complex in Mato Grosso State, Brazil. Mycopathologia 184, 45–51. doi: 10.1007/s11046-018-0313-2
Meyer, W., Aanensen, D. M., Boekhout, T., Cogliati, M., Diaz, M. R., Esposto, M. C., et al. (2009). Consensus multilocus sequence typing scheme for Cryptococcus neoformans and Cryptococcus gattii. Med. Mycol. 47, 561–570. doi: 10.1080/13693780902953886
Montagna, M. T., De Donno, A., Caggiano, G., Serio, F., De Giglio, O., Bagordo, F., et al. (2018). Molecular characterization of Cryptococcus neoformans and Cryptococcus gattii from environmental sources and genetic comparison with clinical isolates in Apulia, Italy. Environ. Res. 160, 347–352. doi: 10.1016/j.envres.2017.09.032
Montagna, M. T., Santacroce, M. P., Caggiano, G., Tatò, D., and Ajello, L. (2003). Cavernicolous habitats harbouring Cryptococcus neoformans: results of a speleological survey in Apulia, Italy, 1999-2000. Med. Mycol. 41, 451–455. doi: 10.1080/13693780310001602785
Noguera, M. C., Escandón, P., and Castañeda, E. (2015). Cryptococcosis in Atlántico, Colombia: an approximation of the prevalence of this mycosis and the distribution of the etiological agent in the environment. Rev. Soc. Bras. Med. Trop. 48, 580–586. doi: 10.1590/0037-8682-0178-2015
Pernice, I., Lo Passo, C., Criseo, G., Pernice, A., and Todaro-Luck, F. (1998). Molecular subtyping of clinical and environmental strains of Cryptococcus neoformans variety neoformans serotype A isolated from southern Italy. Mycoses 41, 117–124. doi: 10.1111/j.1439-0507.1998.tb00312.x
Pham, C. D., Ahn, S., Turner, L. A., Wohrle, R., and Lockhart, S. R. (2014). Development and validation of benomyl birdseed agar for the isolation of Cryptococcus neoformans and Cryptococcus gattii from environmental samples. Med. Mycol. 52, 417–421. doi: 10.1093/mmy/myt028
Rajasingham, R., Smith, R. M., Park, B. J., Jarvis, J. N., Govender, N. P., Chiller, T. M., et al. (2017). Global burden of disease of HIV-associated cryptococcal meningitis: an updated analysis. Lancet Infect. Dis. 17, 873–881. doi: 10.1016/S1473-3099(17)30243-8
Romeo, O., Scordino, F., Chillemi, V., and Criseo, G. (2012). Cryptococcus neoformans/Cryptococcus gattii species complex in southern Italy: an overview on the environmental di?usion of serotypes, genotypes and mating-types. Mycopathologia 174, 283–291. doi: 10.1007/s11046-012-9547-6
Shadomy, H. J., Wood-Helie, S., Shadomy, S., Dismukes, W. E., and Chau, R. Y. (1987). Biochemical serogrouping of clinical isolates of Cryptococcus neoformans. Diagn. Microbiol. Infect. Dis. 6, 131–138. doi: 10.1016/0732-8893(87)90097-6
Springer, D. J., Billmyre, R. B., Filler, E. E., Voelz, K., Pursall, R., Mieczkowski, P. A., et al. (2014). Cryptococcus gattii VGIII isolates causing infections in HIV/AIDS patients in Southern California: identification of the local environmental source as arboreal. PLoS Pathog. 10:e1004285. doi: 10.1371/journal.ppat.1004285
Springer, D. J., Mohan, R., and Heitman, J. (2017). Plants promote mating and dispersal of the human pathogenic fungus Cryptococcus. PLoS One 12:e0171695. doi: 10.1371/journal.pone.0171695
Viviani, M. A., Esposto, M. C., Cogliati, M., Montagna, M. T., and Wickes, B. L. (2001). Isolation of a Cryptococcus neoformans serotype A MATa strain from the Italian environment. Med. Mycol. 39, 383–386. doi: 10.1080/mmy.39.5.383.386
Keywords: Cryptococcus neoformans complex, Cryptococcus gattii complex, environment, molecular typing, mating assay
Citation: Trovato L, Oliveri S, Esposto MC, Prigitano A, Romanò L and Cogliati M (2019) Cryptococcus neoformans and Cryptococcus gattii Species Complex Isolates on the Slopes of Mount Etna, SICILY, Italy. Front. Microbiol. 10:2390. doi: 10.3389/fmicb.2019.02390
Received: 27 June 2019; Accepted: 02 October 2019;
Published: 18 October 2019.
Edited by:
Orazio Romeo, University of Messina, ItalyReviewed by:
Bodo Wanke, National Institute of Infectology Evandro Chagas (INI), BrazilMarilene Henning Vainstein, Federal University of Rio Grande do Sul, Brazil
Copyright © 2019 Trovato, Oliveri, Esposto, Prigitano, Romanò and Cogliati. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Laura Trovato, bHRyb3ZhdG9AdW5pY3QuaXQ=