
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 11 September 2019
Sec. Antimicrobials, Resistance and Chemotherapy
Volume 10 - 2019 | https://doi.org/10.3389/fmicb.2019.02131
This article is part of the Research Topic Fundamentals of, and Applications Based on, Quorum Sensing and Quorum Sensing Interference View all 21 articles
 F. Jerry Reen1,2
F. Jerry Reen1,2 José A. Gutiérrez-Barranquero1†
José A. Gutiérrez-Barranquero1† Ronan R. McCarthy1†
Ronan R. McCarthy1† David F. Woods1Sara Scarciglia1Claire Adams1Kristian Fog Nielsen3
David F. Woods1Sara Scarciglia1Claire Adams1Kristian Fog Nielsen3 Lone Gram3
Lone Gram3 Fergal O’Gara1,4,5*
Fergal O’Gara1,4,5*Despite the discovery of the first N-acyl homoserine lactone (AHL) based quorum sensing (QS) in the marine environment, relatively little is known about the abundance, nature and diversity of AHL QS systems in this diverse ecosystem. Establishing the prevalence and diversity of AHL QS systems and how they may influence population dynamics within the marine ecosystem, may give a greater insight into the evolution of AHLs as signaling molecules in this important and largely unexplored niche. Microbiome profiling of Stelletta normani and BD1268 sponge samples identified several potential QS active genera. Subsequent biosensor-based screening of a library of 650 marine sponge bacterial isolates identified 10 isolates that could activate at least one of three AHL biosensor strains. Each was further validated and profiled by Ultra-High Performance Liquid Chromatography Mass Spectrometry, with AHLs being detected in 8 out of 10 isolate extracts. Co-culture of QS active isolates with S. normani marine sponge samples led to the isolation of genera such as Pseudomonas and Paenibacillus, both of which were low abundance in the S. normani microbiome. Surprisingly however, addition of AHLs to isolates harvested following co-culture did not measurably affect either growth or biofilm of these strains. Addition of supernatants from QS active strains did however impact significantly on biofilm formation of the marine Bacillus sp. CH8a sporeforming strain suggesting a role for QS systems in moderating the microbe-microbe interaction in marine sponges. Genome sequencing and phylogenetic analysis of a QS positive Psychrobacter isolate identified several QS associated systems, although no classical QS synthase gene was identified. The stark contrast between the biodiverse sponge microbiome and the relatively limited diversity that was observed on standard culture media, even in the presence of QS active compounds, serves to underscore the extent of diversity that remains to be brought into culture.
The marine ecosystem is considered to be an underexplored resource for the study of bacterial interactions within eukaryotic hosts. Despite a number of well-studied examples of bacterial interactions within marine hosts such as the density dependent production of luminescence by Aliivibrio fischeri within the light organ of Euprymna scolopes, relatively little is known about the interactions that occur within marine microbial communities (Hmelo, 2017). This is particularly true in the case of the ancient invertebrate, the marine sponge. Marine sponges are sessile filter feeders that consume bacteria and other marine matter (Taylor et al., 2007). Bacteria can inhabit the mesophyll matrix of these invertebrates with almost 60% of the biomass of a marine sponge being comprised of bacterial endosymbionts (Wang, 2006). This symbiotic relationship is mutually beneficial whereby bacteria are provided with a sheltered nutrient rich environment and the marine sponges acquire limiting nutrients from the microflora (Mohamed et al., 2008; Blunt et al., 2009; Mayer et al., 2010, 2011). Within the dense polymicrobial environment of a marine sponge, bacteria can engage in a form of chemical communication termed quorum sensing (QS) (Taylor et al., 2004a; Diggle et al., 2007; Hmelo, 2017). Several classes of QS signaling system are known, with autoinducer peptides favored by gram positive bacteria while N-acyl homoserine lactones (AHLs) predominate within gram negative bacteria (Whiteley et al., 2017). AHLs are capable of activating an autoinducing transcriptional regulator which controls the transcription of target genes involved in a wide variety of cellular processes including the production of virulence determinants (Diggle et al., 2007). There are relatively few studies on the prevalence of functional AHL based QS systems within microorganisms inhabiting marine sponges (Taylor et al., 2004b; Mohamed et al., 2008; Cuadrado-Silva et al., 2013; Britstein et al., 2018). A number of studies have focused on the identification of homologs of genes associated with QS pathways (Zan et al., 2011). However, sequence based approaches provide limited information on the functionality of these homologous systems, which for the most part remains to be determined. This homology-based approach is also limited by the lack of nucleotide sequence homology among AHL synthases and AHL responsive transcriptional regulators (Steindler and Venturi, 2007). More recently, screening of marine sponges for AHL signals has revealed a rich diversity likely encoded by the microbial communities residing in those sponges (Britstein et al., 2018). Given the difficulties faced in bringing marine sponge biodiversity into culture, it is intriguing to speculate that these signals may play a role in moderating the dynamics of the microbial communities within which they operate.
To gain more insight into the relevance of AHL based QS systems within the microbiota inhabiting marine sponges, bacterial sponge isolates were screened for the production of AHLs using classical AHL reporter strains. A total of 10 QS producing isolates were identified and characterized for AHL production. Co-culture of QS positive isolates with marine sponge samples resulted in increased culturable plate diversity from these communities, although no new genera were identified. While addition of AHLs alone did not influence growth or biofilm in the marine sponge isolates, supernatants from several QS positive isolates suppressed biofilm formation in the marine sponge Bacillus sp. CH8a sporeforming strain. This suggests that the anti-biofilm activity of the QS active supernatants may be mediated downstream of intact QS signaling systems in the producing isolates. Genome sequencing of a QS positive Psychrobacter sp. isolate identified in this study revealed the presence of LuxR DNA binding domains. However, there was no evidence of a LuxR autoinducer domain or an AHL synthase domain in this or any other sequenced Psychrobacter genome. Further establishing the prevalence, structure and diversity of AHL based QS systems will give a better understanding of the role of AHL signaling in the marine ecosystem, potentially unlocking some of the natural biodiversity encoded therein.
Bacteria had previously been isolated from sponge genera including Hexactinellida, Stelletta, Lissodendoryx, Poecillastra, Inflatella. These sponges were collected using a remote operated vehicle on board the Celtic Explorer research vessel, 300 nautical miles off the west coast of Ireland as part of the marine biodiscovery cruise, May 2010. Sponge samples from the Amphilectus genus were collected in Gurraig Sound Kilkieran Bay, Galway (Kennedy et al., 2008). Whole sponge samples were rinsed with sterile artificial sea water (ASW, 3.33% (w/v) artificial sea salts, Instant Ocean) and immediately frozen at −80°C on board the ship until further processing. A sample of sponge tissue (1 g) was homogenized by grinding with a sterile porcelain pestle and mortar in 9 ml of sterile ASW. The sponge homogenate was subsequently serially diluted in ASW to 10–5 and 100 μl aliquots of the different dilutions were plated onto Marine agar (MA) (Difco, United Kingdom) and SYP-SW Agar (1% (w/v) starch, 0.4% (w/v) yeast extract, 0.2% (w/v) peptone and 3.33% (w/v) artificial sea salts, 1.5% (w/v) agar. Distinct morphologies were collected to form a library of approximately 650 isolates for subsequent screening and characterization.
As above, two independent samples of each sponge tissue (1 g) were homogenized by grinding with a sterile porcelain pestle and mortar in 10 ml of sterile PBS. DNA was extracted from 100 μl of these samples using the MoBio DNA extraction kit (MoBio) as per manufacturer’s instructions. The extracted gDNA was used as a template to amplify the v3-v5 region of the 16S rRNA gene, these amplicons were sequenced to 2 × 300 bp on a Next Gen Illumina MiSeq (V3) platform. Prior to the microbiome analysis, raw reads were demultiplexed based on inline-barcode sequences. The reads were processed using Minimum Entropy Decomposition (Eren et al., 2015). To assign taxonomic information to each Operational Taxonomic Unit (OTU), BLAST alignments of representative sequences to the NCBI database were performed. Further processing of OTUs was performed using the QIIME software package (version 1.8.01). Microbiome data has been uploaded on the NCBI Sequence Read Archive (SRA) database (BioProject No. PRJNA555824).
The following reporter strains were used to identify AHL production. Short chain AHLs were detected using the Serratia marcescens SP19 (Poulter et al., 2010). Upon production of short chain AHLs S. marcescens SP19 produces a red pigment, prodigiosin. S. marcescens SP19 is an AHL deficient mutant that only produces prodigiosin in response exogenous AHLs. Marine strains were cultured on Marine Broth (Difco, United Kingdom) supplemented with agar (1.5% w/v) for 72 h at 23°C. They were then overlaid with soft LB agar (0.1% agar) inoculated with S. marcescens SP19 at an OD600 nm of 0.5. Overlaid plates were incubated at 30°C overnight. QS was identified by prodigiosin production. As a positive control, 20 μM C4-HSL (Sigma, United Kingdom) was used. C4-C8 chain AHLs were detected using Chromobacterium violaceum CV026. C. violaceum CV026 produces a purple compound called violacein in a QS dependent manner. C. violaceum CV026 contains a transposon in its indigenous AHL synthase gene thus it only produces violacein in response to exogenous AHLs (McClean et al., 1997). Marine strains were cultured on Marine Agar (Difco, United Kingdom) for 72 h at 30°C. They were then overlaid with soft LB agar (0.1% agar) inoculated with C. violaceum CV026 at an OD600nm of 0.5. Overlaid plates were incubated at 30°C overnight. QS was identified by violacein production. 20 μM C8-HSL (Sigma, United Kingdom) was used as a positive control. The broad range Agrobacterium tumefaciens NTL4 biosensor was used for the detection of longer chain AHLs. It contains a plasmid pZLR4 carrying a traG:lacZ reporter fusion (Farrand et al., 1996; Yin et al., 2012). In response to exogenous AHLs the lacZ gene is transcribed resulting in the degradation of 5-bromo-4-chloro-3-indolyl-β-D-galactopyranoside in the media. The previously described plating protocol was observed except soft LB was supplemented with 50 μg/ml 5-bromo-4-chloro-3-indolyl-β-D-galactopyranoside. QS was identified by the breakdown of 5-bromo-4-chloro-3-indolyl-β-D-galactopyranoside resulting in a blue color. A concentration of 20 μM C10-HSL (Sigma, United Kingdom) was used as a positive control.
Genomic DNA from bacterial isolates was extracted using the MoBio UltraClean DNA extraction kit (MoBio, United States) following manufacturers’ guidelines. Strains were identified by PCR amplification of 16S rRNA genes which was carried out as described (Chan et al., 2010) using the universal primer pairs 27F (5′-AGAGTTTGATCMTGGCTCAG-3′) and 1525R (5′-AAGGAGGTGWTCCARCC-3′). PCR products were sequenced by MWG Eurofins, United Kingdom. Sequence identification was performed using BlastN and all sequences were submitted to the NCBI database (Accession numbers: MN209943-MN209952).
Extracts for thin layer chromatography (TLC) and ultra-high performance liquid chromatography-high resolution mass spectrometry (UHPLC-HRMS) were prepared from 200 ml cultures in Marine Broth (Difco, United Kingdom) that had been incubated for 72 h at 30°C and 180 rpm. Bacterial cells were pelleted by centrifugation at 4,000 rpm for 7 min and the supernatant was filter sterilized using a Nalgene® vacuum filtration system (0.2 μm, Sigma-Aldrich®, Germany). The supernatant was incubated with 1:1 volume of acidified ethyl acetate (1% formic acid) for 10 min at RT shaking at 180 rpm. The ethyl acetate phase was taken and dried using rotary evaporation. Residues were resuspended in 1 ml ethyl acetate and stored at −20°C. Ethyl acetate extracts were tested for QS activity by spotting 20 μL on C18 reverse phase TLC plates (20 cm × 20 cm TLC aluminum plates, Millipore, United Kingdom) with a methanol:water 7:3 (v/v) mobile phase. Once dried, TLC plates were overlaid with 25 ml of soft LB agar (0.1% agar) inoculated with the biosensor at an OD600nm of 0.5. Following confirmation of extract activity, extracts were dried using nitrogen evaporation and analyzed by UHPLC-HRMS for identification.
Samples were resuspended in approx. 150 μl 50:50 (vol/vol) acetonitrile (ACN)-water 50:50 (vol/vol) and 2 μl subsamples analyzed by UHPLC-HRMS. This was done on an Agilent Infinity 1290 UHPLC system (Agilent Technologies, Santa Clara, CA, United States) coupled to an Agilent 6550 QTOF MS operated in positive electrospray (ESI) mode, scanning m/z 50-1700. A lock mass solution of 10 μM Hexakis(2,2,3,3-tetrafluoropropoxy)phosphazene (Apollo Scientific Ltd., Cheshire, United Kingdom) dissolved in 95% acetonitrile and infused in the secondary ESI sprayer using an extra LC pump at a flow of 20 μl/min, and the [M + H]+ at m/z 922.0098 used as lock mass, resulting in a mass accuracy better than 3 ppm (deviation relative to theoretical m/z value).
An Agilent Poroshell 120 phenyl-hexyl column (2.1 × 150 mm, 2.7 μm), held at 60°C was used for separation. A linear gradient at 0.35 ml/min, consisting of water and ACN both buffered with 20 mM formic acid was started at 10% ACN and increased to 100% after 15 min, where it was held for 2 min. It was subsequently returned to 10% ACN in 0.1 min and maintained for 3 min (Kildgaard et al., 2014).
A reference standard mixture containing the following HSLs were included in the analytical sequence: C4, C6, C8, C10, C12, Oxo-C6, Oxo-C8, Oxo-C10, Oxo-C12, OH-C6, OH-C8, OH-C10, and OH-C12. Using the Agilent MassHunterQuant software, extracted ion chromatograms the [M + H]+ and [M + Na]+ ions ± 10 ppm were used for identification, along with the isotopic pattern (Kildgaard et al., 2014), and finally a retention time match ± 0.01 min.
Isolates with distinct morphologies that were cultured on QS-treated plates and identified by 16S rRNA sequencing were grown on marine agar for 72 h at 23°C. Cells were transferred into fresh marine broth, OD600 nm 0.05, in the presence and absence of 50 nM–10 μM of 3-oxo-C12 HSL, with DMSO as carrier control. Growth was measured spectrophotometrically at OD600 nm in honeycomb plates incubated at 23°C on a Bioscreen-C automated growth curves analysis system (Growth Curves USA).
Bacillus sp. CH8a was grown in marine broth (MB) at 23°C overnight with shaking (Phelan et al., 2012). In order to monitor the impact on biofilm formation, 500 μl of cell-free supernatant (CFS) from the QS strain 3-day cultures was added to 500 μl of test cultures at OD600 nm 0.1, grown in MB. As controls, 500 μl of fresh media was added to 500 μl of test cultures (media control) and 500 μl of the CFS of test cultures were added to 500 μl of corresponding test culture. In all cases, CFS was obtained by centrifugation at 8,000 rpm for 5 min followed by filtration through a 0.2 μm sterile filter. Bacillus sp. CH8a biofilms were incubated for 2 days at 23°C. Unattached cells were aspirated out of all wells which were washed once with 1 ml of sterile water. Attached cells were quantified using 0.1% (w/v) crystal violet. For marine isolate biofilm analysis, 3-oxo-C12-HSL was added at 10 μM to media and processed as described above.
A simple co-culture system was designed to expose marine sponge homogenate to signals and metabolites from actively growing QS active isolates. Costar Spin-X 0.22 μm cellulose acetate centrifuge tube filters (Corning) consisting of an upper chamber with a filter base and a lower receptacle chamber was used for this purpose. QS active isolates were grown in nutrient media in the top chamber, while marine sponge homogenate was incubated in the lower chamber. While the marine sponge homogenate was physically separate from the QS active isolate at all times, transfer of small molecular signals and metabolites between both chambers was possible. In order to establish the validity of the co-culture system for QS transfer between the upper and lower chambers, as well as the integrity of the filter system to prevent bacterial leakage, several test studies were performed. The upper chamber was removed, and 1 ml of LB broth was added to the lower chamber. The upper chamber was replaced and a known QS producing organism P. aeruginosa PA14 (500 μl) was added. The tubes were incubated at 37°C for 24 h after which time the top chamber was removed. Aliquots of the lower chamber were tested for (a) activation of AHL biosensor strains C. violaceum and S. marcescens and (b) contaminating bacterial growth.
Quorum sensing active isolates were grown in marine broth at 23°C for 72 h. Stelletta normani sponge sample (∼1 g) was homogenized in 10 ml of PBS. The upper chamber of the Costar Spin-X 0.22 μm cellulose acetate centrifuge tube filter (Corning) was removed, and 500 μl of sponge homogenate with 500 μl marine broth was added to the lower chamber of the filter tube. The filter was repositioned and 300 μl of QS active isolate in marine broth at OD600 nm 0.2 was added. Several controls were included: (i) 300 μl of marine broth was added to the top chamber to provide a baseline profile of bacteria that could be cultured under the standard media conditions used, (ii) each QS active isolate was prepared as above with no sponge sample in the lower chamber to control for leakage through the membrane, and (iii) media controls were included to control for inadvertent contamination (Figure 1). The tubes were incubated at 23°C with gentle shaking at 50 rpm for 72 h at which time the upper chambers were carefully removed. The contents of the lower chamber were mixed gently and serially diluted. Dilutions were plated on marine agar and incubated at 23°C for at least 72 h. Colony numbers and morphologies were profiled and distinct isolates were identified by 16S rDNA sequencing.
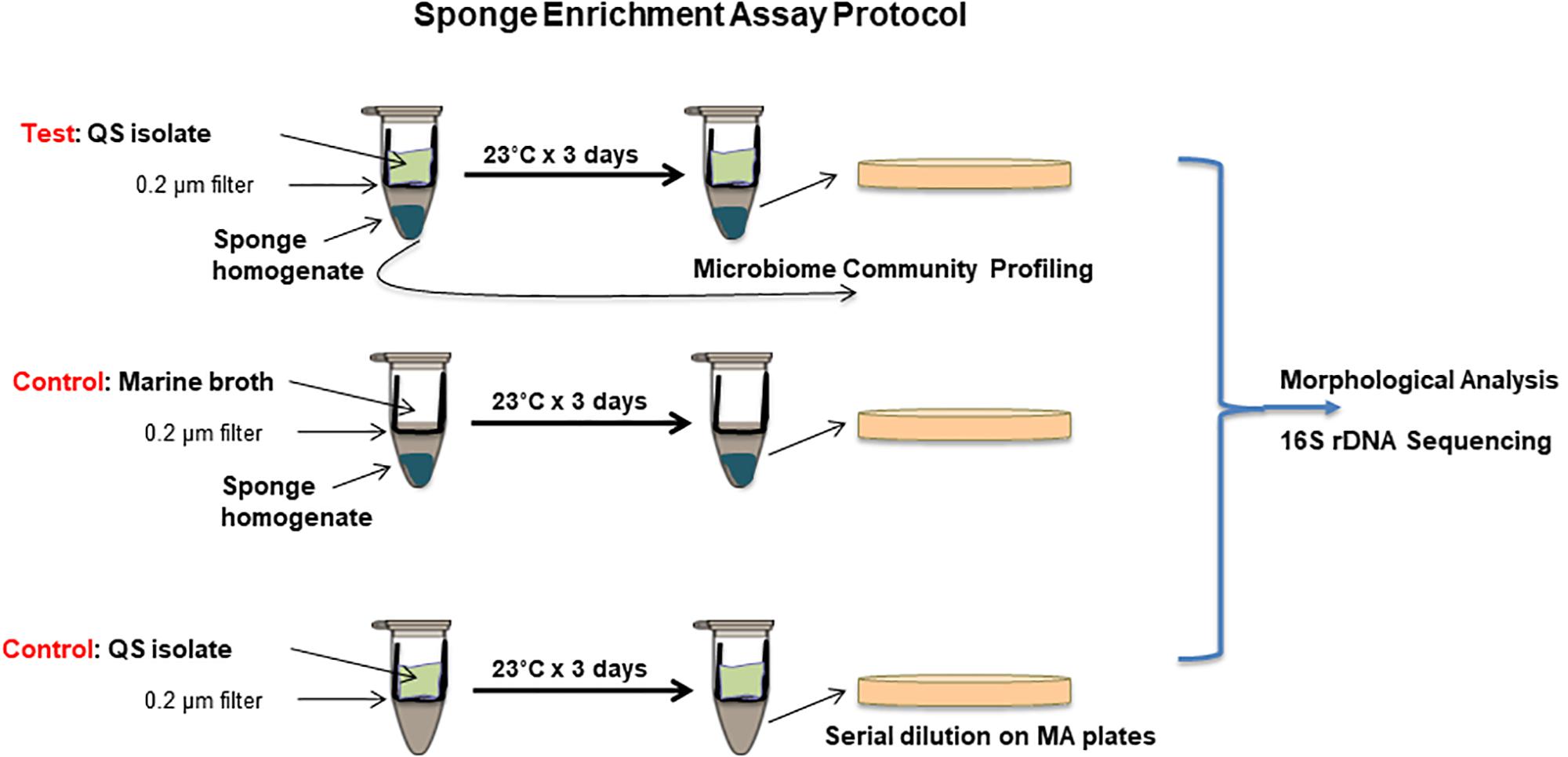
Figure 1. Co-culture sponge enrichment assay protocol incorporating QS and QQ samples with appropriate controls. QS or QQ producing isolates are placed in the top chamber with marine both media to facilitate growth. These are separated from the lower chamber containing sponge material by a 0.2 μm filter. QS or QQ signals can transition from the top chamber into the lower chamber by diffusion. Controls are included to ensure no microbial transfer between chambers. Isolation of culturable organisms is achieved on marine agar plates following 3 day incubation. Distinct morphologies are scored and selected for 16S rRNA sequencing.
Total DNA of Psychrobacter sp. 230 strain was obtained using the UltraClean microbial DNA isolation kit (Mo Bio Laboratories, Inc., Carlsbad, CA, United States) and was used for DNA library preparation using a TruSeq exome library prep kit. The draft genome sequencing project of Psychrobacter sp. 230 strain was performed by the Beijing Genomics Institute (BGI, China) using the Illumina HiSeq 4000 sequencing platform involving paired-end reads with a read length of 150 bp. The superfast FASTA/Q file manipulation tool, readfq.v5 (BGI, unpublished software), was used for quality trimming. This software removes the paired-end reads with a certain proportion of low-quality bases (default, 40%; parameter setting, 6 bp), reads with a certain proportion of Ns (ambiguous bases; default, 10%; parameter setting, 10 bp), reads with adapter contamination (default, 15 bp overlapped between adapter and reads), and duplicate sequences. Thus, the high-quality-filtered reads were all 150 bp long. From a total of 5.417.936 raw paired-end reads of 150 bp length, 3.329.182 high-quality reads were generated after processing with readfq.v5. Assembly was performed using SOAPdenovo 2.04 with default parameters. The sequencing depth provided 47.5 coverage of the genome. The draft genome assembly comprised 69 contigs with an N50 value of 183,949 grouped into 69 scaffolds with a total size of 3,290,930 bp and an overall GC content of 42.8%. The whole-genome shotgun project was deposited at NCBI under the accession number: NZ_SNVH00000000.1. The raw reads obtained after processing with readfq.v5 have been submitted to NCBI SRA under the accession number SRP216019.
To infer the phylogenetic history, nucleotide sequences were retrieved and downloaded from NCBI: GenBank2. Sequences were aligned using Clustal Omega and were cut to consistent lengths (Sievers et al., 2011). The evolutionary analysis was conducted in MEGA X using the Neighbor-Joining method (Saitou and Nei, 1987; Kumar et al., 2018). The clustering was tested using Bootstrapping with 1,000 replicates (Felsenstein, 1985). The Tajima–Nei method was used to calculate the evolutionary distances (Tajima and Nei, 1984). There was a total of 1,459 positions in the final dataset.
A range of QS active marine sponge microbial communities have been reported in recent years (Mohamed et al., 2008; Zan et al., 2012; Abbamondi et al., 2014; Britstein et al., 2018). The presence of QS systems in the sponge microbiota suggests a dynamic and ordered community that can respond to external cues and challenges. However, the extent to which QS producing bacteria colonize the marine sponge, and the role of QS within those microbial communities remains to be determined. Therefore, two distinct marine sponges were selected for microbial community profiling to establish the extent to which QS potential existed therein. Given the heterogeneity that exists in many clinical and ecological niches, with localized population profiles existing within relatively short distances of each other within a singular niche, two separate samples of each marine sponge species were selected. Homogenization and subsequent sequencing of two independently harvested triplicate samples revealed some interesting features of the respective microbiomes. Approximately 50% of the S. normani samples was identified only to the level of kingdom classification, suggesting a unique and unexplored bacterial diversity (Supplementary Table S1). The dominant phyla were Proteobacteria and Actinobacteria. At the genus level, Iamia [14.9% (±1.9)], Pseudoalteromonas [6.1% (±0.6)], Moraxellaceae [4.7% (±0.1)], and Nitrospira [2.5% (±0.6)] were the most abundant identifiable genera (Figure 2A). In contrast, the largest group identified for sponge BD1268 was the γ-Proteobacteria, representing between 85 and 94% of the OTUs in these samples. Sponge BD1268 was colonized by genera such as Candidatus Pelagibacter [12% (±4.5)], Thioprofundum [4.2% (±1.0)], and Thiohalophilus [2.4% (±0.4)]. Although the individual profiles remained relatively consistent from the perspective of the dominant phyla and families, there were differences in relative abundances between the microbiome profiles from the distinct sponge samples suggesting that heterogeneity of the population may exist within the sponge (Figure 2A). Diversity indexes (Shannon and Simpson) and OTU abundance was higher in the S. normani samples when compared with the respective samples from the BD1268 sponge (Figure 2B). As expected, independent samples clustered together based on sponge source (Supplementary Figure S1). Genera known to encode QS signaling systems (e.g., Pseudomonas, Halomonas, Psychrobacter) were present in the microbiomes of both sponges, although it was interesting to note that they were low in abundance when compared to the principal colonizers of the sponges. Therefore, notwithstanding the fact that genera previously shown to encode QS systems were present, the degree to which QS signaling pervades in these sponges remained to be determined.
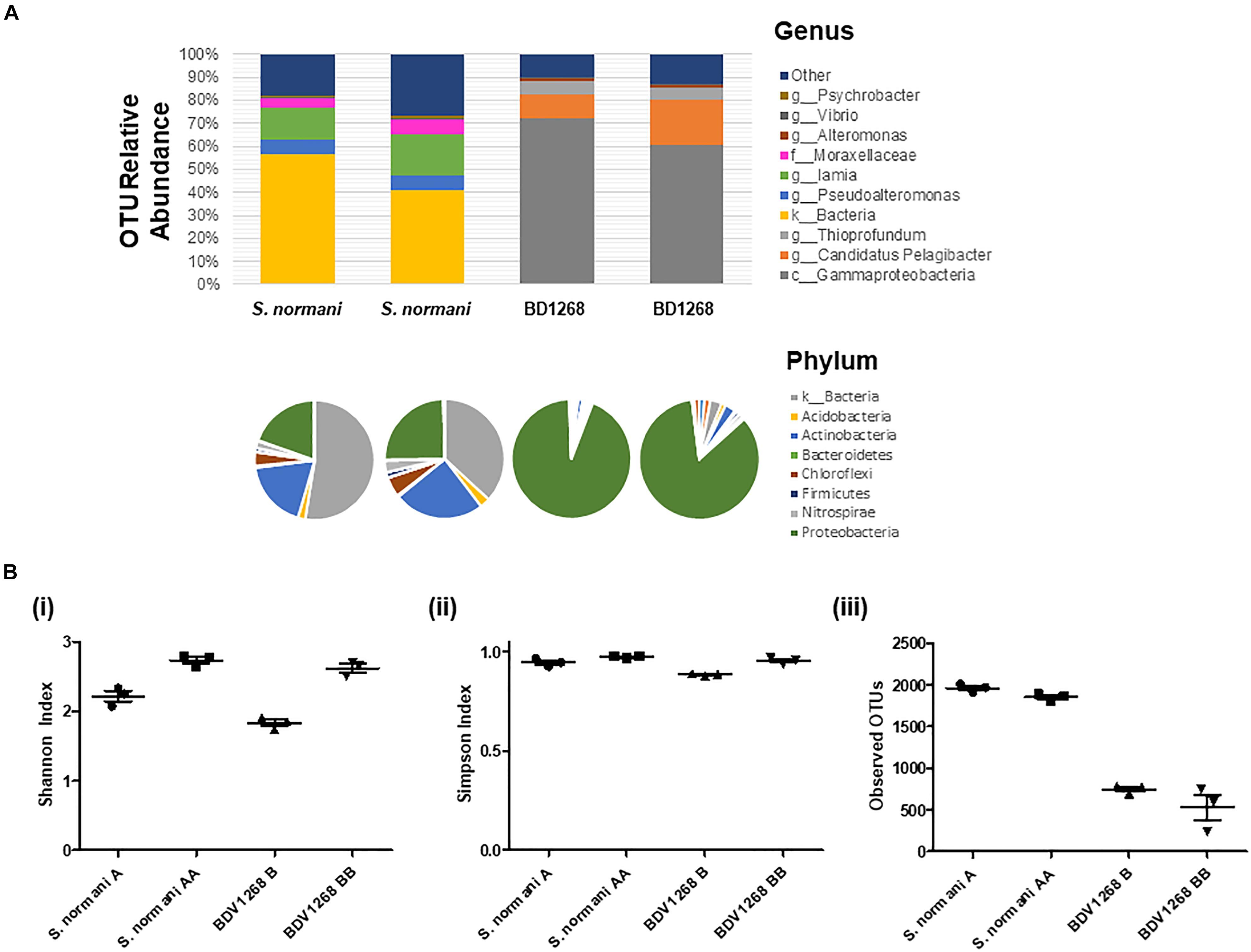
Figure 2. (A) Microbiome 16S DNA bacterial community profiling of sponge DNA. Stacked charts show relative abundance at the genus level. Phylum community profiles are presented in pie-chart format above the corresponding genus-level stacked chart. Comparison with culturable microbiota reveals considerable challenge in activating the “silent” majority of genera present. Two independent samples were taken for each sponge and the average of three independent DNA extraction sequenced replicates is presented for each sample. (B) Diversity and OTU analysis on microbiome data (i) Shannon diversity index (H = –Σpi ln pi), (ii) Simpson diversity index, and (iii) OTU abundance.
The collection of morphologically diverse bacterial isolates from the S. normani sponge described above had previously been reported (Gutierrez-Barranquero et al., 2017). Together with this, cultivation of bacteria from sponge tissue derived from other sponge families such as Hexactinellida, Lissodendoryx, Poecillastra, Inflatella, and Amphilectus had yielded a collection of bacteria from the Proteobacteria, Actinobacteria, Bacteroidetes and Firmicutes groups (Gutierrez-Barranquero et al., 2017). Within these groups a wide range of different families were represented. The most abundant families included Pseudoalteromonadaceae, Flavobacteriaceae, and Moraxellaceae. To establish the prevalence of AHL based QS within the sponge microbiome, over 650 sponge isolated bacteria were screened for AHL production. In total 10 AHL producing candidates were identified, representing approximately 0.015% of the culture collection (Supplementary Figure S2 and Table 1). In order to identify the AHL positive isolates to a species level, the full length 16S rRNA (1,400 bp) sequence was determined. Among the isolates identified with AHL activity were a number of known AHL producers these included members of the Halomonas, Psychrobacter, Vibrio, Pseudoaltermonas, and Pseudomonas genera (Bruhn et al., 2005; Huang et al., 2009; Tahrioui et al., 2011; Ma et al., 2016). Despite the fact that the culture collection was populated by bacterial isolates harvested from six sponges, only four of these (Stelletta, Hexactinellida, Inflatella, and Lissodendoryx) yielded QS active isolates. The detection of QS active genera such as Pseudomonas and Halomonas from within the S. normani strains, despite the low abundance evident in the microbiome profile (Supplementary Table S1), is an important finding and suggests potentially a temporal role for QS active strains within communities. All QS active isolates were further validated by TLC analysis and soft agar biosensor overlay prior to selection for UHPLC-HRMS analysis and structural characterization (Supplementary Figure S2).

Table 1. AHL profile of marine sponge QS active bacterial isolates.
To identify the AHLs being produced by each of the QS positive candidates UHPLC-HRMS was performed (Supplementary Figure S2). Ethyl acetate extracts were prepared for all isolates that had tested positive for AHL. All extracts were tested to ensure activation of respective AHL biosensors. Based on UHPLC-HRMS analysis, AHLs were identified in the majority of isolates (Table 1). The most abundant AHLs being produced by these isolates were 3-oxo-C10-HSL and 3-oxo-C12-HSL. Both AHLs, in addition to 3-OH-C10 HSL, were identified in extracts from Psychrobacter sp. and Pseudoalteromonas sp., the latter also producing trace amounts of 3-OH-C6 HSL. Only one isolate produced detectable amounts of C4-HSL, with the same Pseudomonas sp. isolate also producing trace amounts of C6-HSL. In some cases, no AHL traces were identified, notwithstanding the ability of the isolates to activate the respective biosensors.
The marine sponges profiled in this study appear to sustain a significant network of QS systems which may be important in modulating the population in response to external cues. Previous studies have shown that the ratio of QS and QQ strains can change dramatically in response to environmental conditions (Tan et al., 2015). Therefore, we investigated whether the secretome of the QS active isolates could impact on the profile of culturable bacteria that could be obtained from marine sponges under the standard media conditions used. Filter based microtubes were used to establish a co-culture system whereby the QS active isolate was added to a well with a porous membrane through which secreted QS signals can be transferred to the chamber below containing S. normani sponge homogenate (Figure 3A). In three independent studies, distinct colony morphologies were observed on plates with QS treated samples when compared to untreated controls (Figure 3B). The fidelity of all controls was maintained throughout the experiments with no leakage or inadvertent contamination observed.
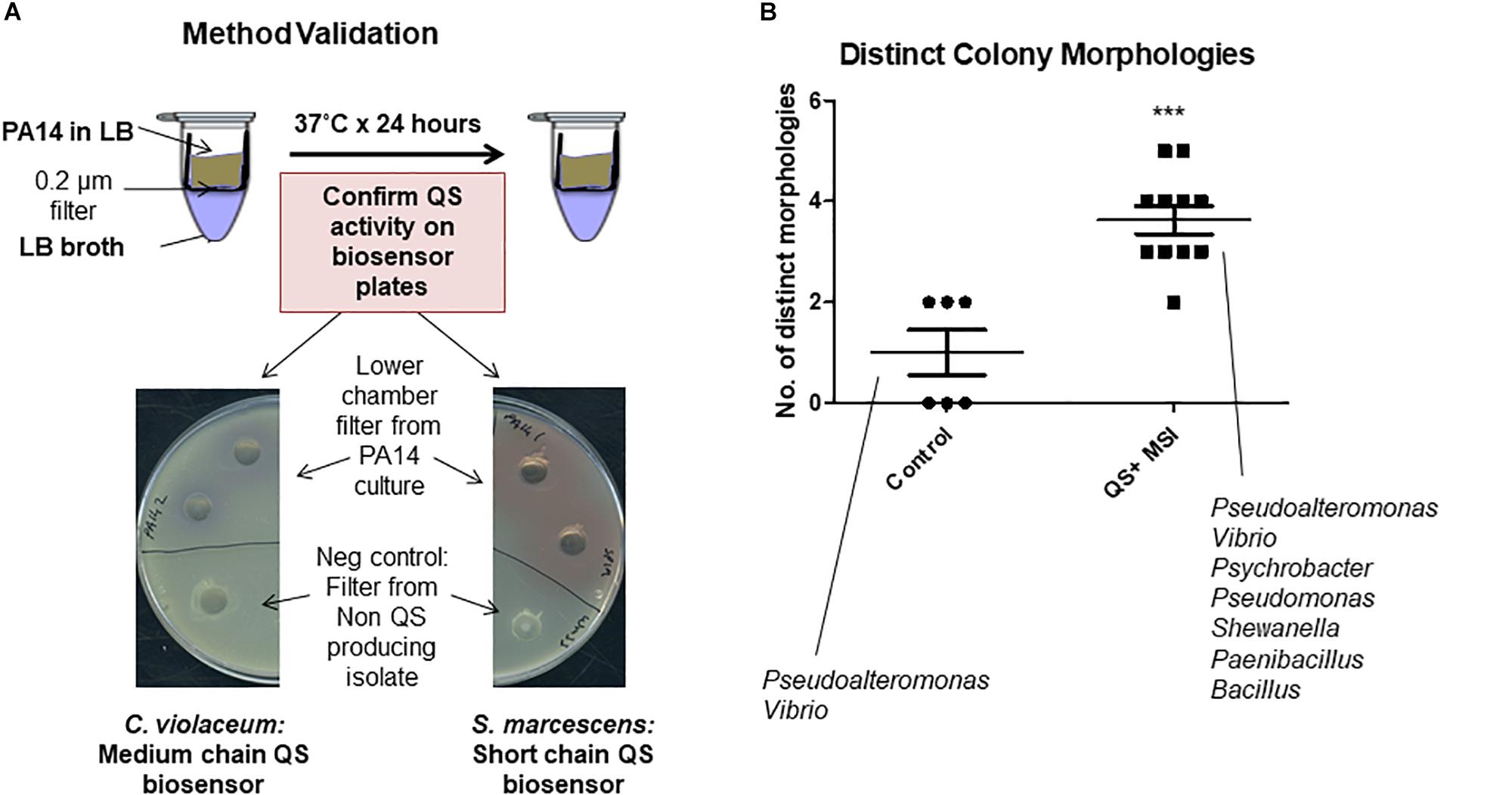
Figure 3. Co-culture sponge enrichment analysis. (A) Proof of concept using QS producing strains. Transfer of the QS active compounds confirmed by validation on Biosensor seeded plates. (B) Outcome of QS-mediated sponge enrichment assays measuring recoverable morphologies which were subsequently 16S rRNA-typed. Each datapoint refers to the number of distinct species recovered after the designated treatment. The species listed represent recoverable isolates identified in the study. Data presented represents at least three independent assays encompassing distinct S. normani sponge preparations. Statistical analysis was performed by Student’s t-test. ∗∗∗p ≤ 0.001.
Pseudoalteromonas and Vibrio were both cultured from the S. normani sponge homogenate in the absence of any treatments. Upon co-culture with QS active isolates, Pseudomonas, Paenibacillus and Psychrobacter were isolated in independent experiments. This was in addition to Pseudoalteromonas and Vibrio as seen on the control plates. Co-culture with QS-active isolates 12 and 411 resulted in isolation of Pseudomonas while 214 and 211 led to isolation of Paenibacillus. Psychrobacter was isolated upon co-culture with all 211, 214, and 411. No representative from these genera was observed on the untreated control plates (Figure 3). All three genera were represented at a very low relative abundance within the microbiome from the untreated S. normani sponge samples (Pseudomonas [0.003% (±0.003), Psychrobacter [0.6% (±0.1)], and Paenibacillus only evident at Order level of Bacillales [0.02% (±0.05)]) suggesting a change in community structure (Supplementary Table S1). Together, these data suggest that cell-cell communication and QS can impact the dynamics of population growth within microbiomes and influence the culture-readiness of genera in response to external cues. It was notable that the impact of QS active supernatants was not restricted to gram negative organisms. However, the vast majority of microbiome constituents were not represented in the culturable diversity on the plates. Representatives of e.g., Iamia, Nitrospira, Caldilinea, or Gaiella were not observed on either control or test plates. Therefore, modification of the media to cater for additional supplemental requirements, or indeed to reduce nutritional richness, may be required to capture these unculturable components.
In order to understand how QS signaling might influence species dynamics within the marine sponge, we investigated whether marine isolates obtained following co-culture with QS positive marine sponge isolates responded to exogenous AHLs. As the most frequently identified AHL in this study, 3-oxo-C12 HSL was selected to assess its influence on two key QS associated phenotypes i.e., biofilm formation and growth. Somewhat surprisingly, neither was affected upon addition of 3-oxo-C12 HSL when compared to untreated samples (Figure 4). Addition of either 10 or 50 μM 3-oxo-C12 HSL did not impact on growth of test strains, either in exponential or stationary phase, as determined over 72 h (Figure 4 and Supplementary Figure S3). Similarly, although the test isolates formed biofilms in multi-well plates, addition of either 10 or 50 μM 3-oxo-C12 HSL did not significantly alter total attached biomass (Figure 4).
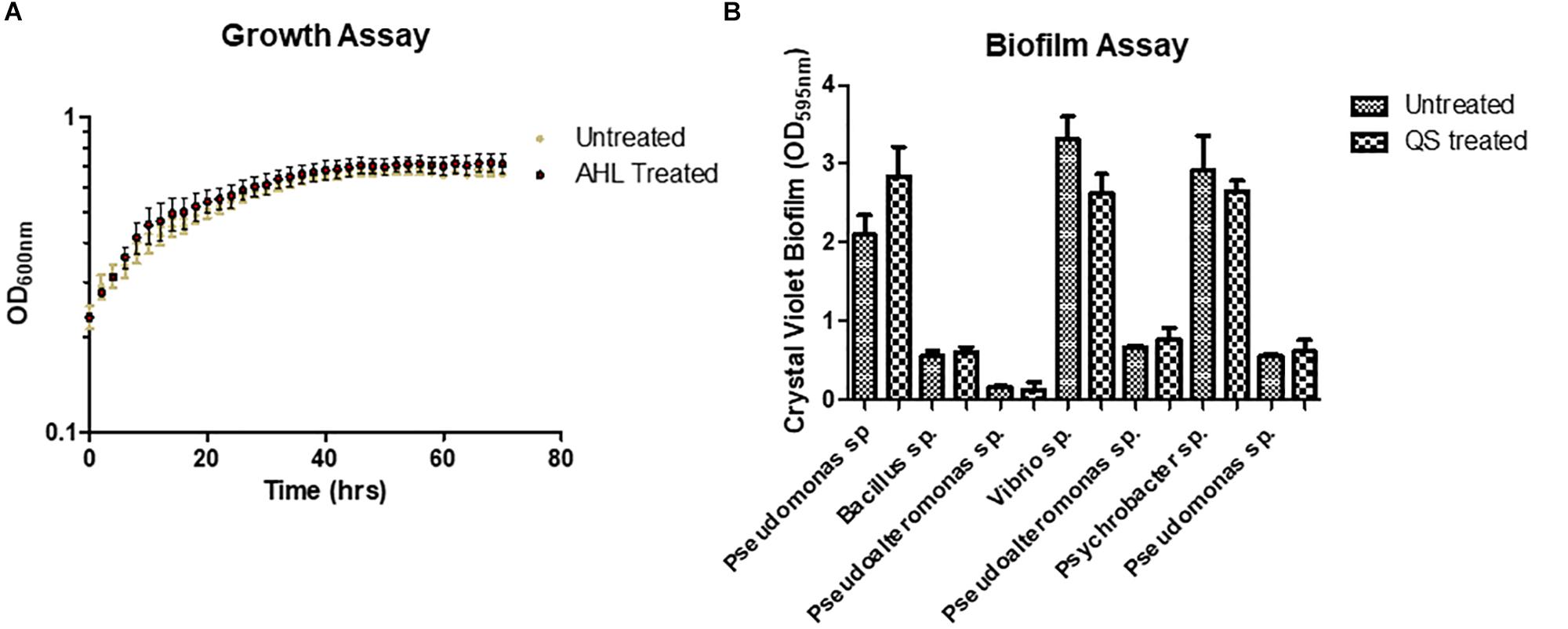
Figure 4. Addition of purified 3-oxo-C12-HSL did not influence (A) growth of a Pseudomonas sp. isolate or (B) biofilm formation of marine sponge isolates recovered in this study following co-culture.
The ability of QS to control a range of virulence related phenotypes is well established. In addition to controlling biofilm formation and toxin secretion in important pathogens, QS is also known to elicit an antagonistic response toward co-colonizing organisms in producing strains. To test if this applied to the marine sponge QS active isolates, biofilm formation in the marine sponge sporefomer Bacillus sp. CH8a was investigated in the presence and absence of extracts from QS positive isolates. Addition of extracts from several of the isolates led to a significant reduction in biofilm formation by CH8a when compared with the untreated control (Figure 5). This suggests that QS active strains would likely produce compound(s) that would moderate the behavior of co-existing microbes within the sponge microbiome.
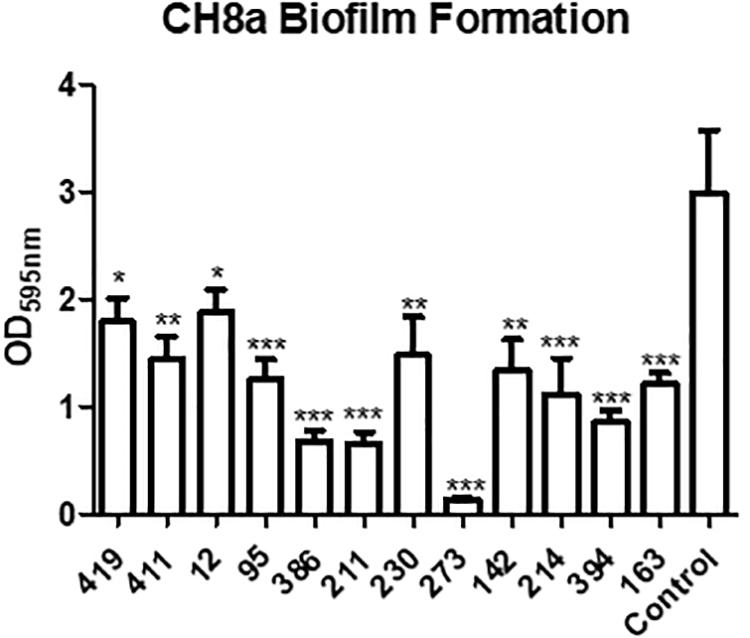
Figure 5. Modulation of Bacillus sp. CH8a biofilm formation by extracts from QS active isolates. Data presented is the average of at least three independent biological replicates. Statistical analysis was performed by one-way ANOVA with post hoc Bonferroni corrective testing (∗p ≤ 0.05, ∗∗p ≤ 0.005, ∗∗∗p ≤ 0.001).
Suppression of biofilm and activation of QS biosensors suggest that QS signaling is a highly networked and evolved system in the marine sponge niche. However, very little is known about the factors involved in newly emerging QS positive species. Psychrobacter sp. have only recently been shown to be QS positive, although the molecular mechanism underpinning this activity remains to be elucidated. To assess if any proteins encoded within Psychrobacter genomes could function as LuxR homologs the available Psychrobacter genomes were investigated via the Pfam Domain Search Tool to identify proteins with specific domains associated with LuxR type transcriptional regulators, i.e., the autoinducer binding domain (PFAM03472) or the LuxR DNA binding domain (PFAM00196). A total of 70 proteins were identified in 38 genomes that possessed a LuxR type DNA binding domain (Supplementary Table S1). However, neither an autoinducer binding domain nor a classical autoinducer synthase domain (PFAM00765) were identified in any of the available genome sequences. This raised two possibilities; (i) a potential novel AHL synthase with a low sequence homology to known AHL synthases may be functionally active within the Psychrobacter genus, or (ii) marine Psychrobacter sp. encode novel regions of DNA carrying the capacity for AHL production. Several reports exist in the literature of QS active strains where the corresponding genetic systems encoding that activity remain to be identified. The prevalence of horizontal gene transfer, and the phenotypic and genotypic heterogeneity that exists within communities is such that interrogating model genomes can be limited when searching for a particular functionality that may be strain specific. We considered it important to investigate the QS signaling potential that is encoded in the genome of the Psychrobacter species, particularly as we had a QS active strain with which to interrogate.
Whole genome sequencing of the Psychrobacter sp. 230 isolate from this study revealed a genome encoding 2908 genes (Supplementary Figure S4). The draft genome assembly comprised 69 contigs with an N50 value of 183,949 grouped into 69 scaffolds with a total size of 3,290,930 bp and an overall GC content of 42.8% (Table 2). The Psychrobacter sp. 230 genome was searched for Lux domains and three putative LuxR domain proteins were identified. BLASTX sequence searches and SMART domain analysis suggested that these three proteins were transcriptional response regulator proteins, with no evidence of autoinducer domains. A LysE family homoserine(lactone) translocation protein was also encoded in the genome, as was a dienelactone hydrolase family protein. However, the absence of a LuxI-like synthase gene in the Psychrobacter genome indicates that the molecular mechanism through which AHL based QS is performed in this isolate remains to be ascertained. Cluster based analysis revealed that this Pscyhrobacter sp. grouped with three other Psychrobacter sp. isolates and distinct from other members of the genus suggesting it may be a genetic outlier within Psychrobacter (Figure 6). Previously, Ma et al. (2016) identified a Psychrobacter sp. isolate from mangrove with QS activity that clustered with Psychrobacter sp. isolates from deep sea sediments of the east Pacific Ocean. Therefore, a functional approach may be warranted to uncover the molecular basis of QS signaling in this species.

Table 2. Genome data for Psychrobacter sp. 230 marine sponge isolate.
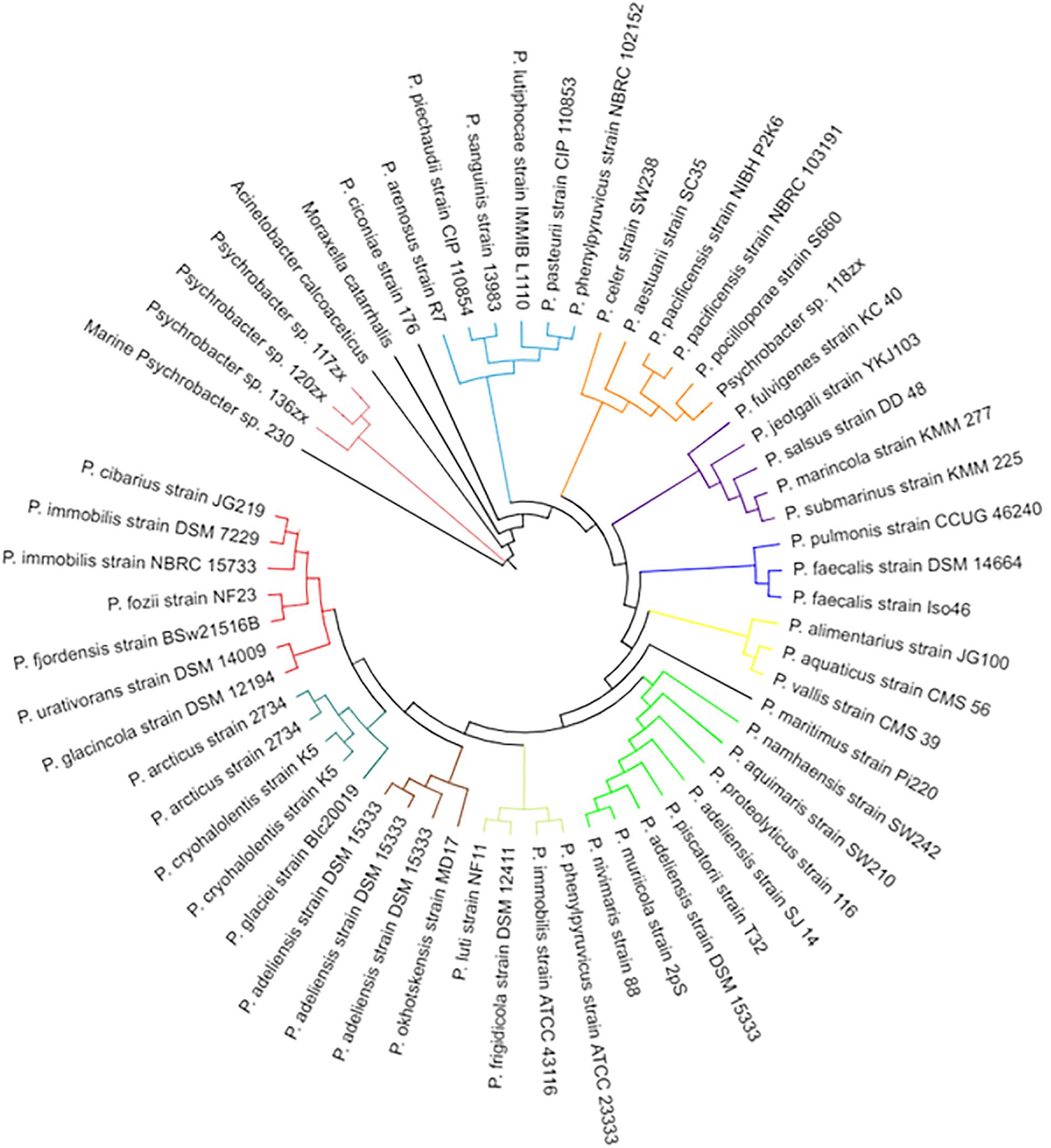
Figure 6. Phylogenetic tree of Psychrobacter species including the marine Psychrobacter sp. 230 isolate identified in this study. Species names are listed on the outer edge of each branch and an arbitrary color based system is used to distinguish the various clades in the tree.
In this study, 650 marine sponge bacterial isolates were screened for the ability to produce AHLs. A total of 10 isolates were identified that were capable of activating AHL biosensor reporter strains. Mass spectrometry revealed that several of the isolates produced the same or similar AHLs (OC10–OC12 HSL). The capacity for AHL based signaling in the marine ecosystem has previously been reported. AHL signaling in marine snow was first described by Gram et al. (2002), with species of Roseobacter shown to be QS active. More recently, Pantoea ananatis has been reported to produce a spectrum of AHL signals in marine snow, governing extracellular enzyme production in producing strains (Jatt et al., 2015). Since the first description of AHL based QS in A. fischeri species (Nealson and Hastings, 1979), where the LuxIR paradigm system was first identified, AHL signals have been found in a broad diversity of marine isolates (Hmelo, 2017). Rasch et al. (2007) described AHL production in Aeromonas salmonicida isolates, while a number of studies profiled members of the Vibrionaceae for AHL production (Yang et al., 2011; Purohit et al., 2013). The diversity of AHL signals that are encoded in the marine ecosystem has been highlighted by a recent study reporting AHLs with long (up to 19 carbons) and poly-hydroxylated acyl side chains (Doberva et al., 2017). At the same time, studies reporting QS inhibition or quenching in the marine environment have also received considerable attention in recent years (Romero et al., 2012; Gutierrez-Barranquero et al., 2017; Ma et al., 2018). Primarily produced by microbial species, host derived quorum quenching (QQ) has also been described (Weiland-Brauer et al., 2019). Elucidating and profiling the extent of QS signaling within these environments is a key step in understanding the functional role played by QS in the host-microbe interaction.
Interspecies communication within the microbial communities of the marine sponge may offer a competitive advantage through cross-genus coordinated behavior. If several species all produce the same or similar AHLs, then they potentially could adopt community like behaviors more rapidly than species that are not part of this interspecies signaling network. This could arise from the activity threshold being reached more rapidly if several different species produce the same signaling molecule. Of course, conservation within receptor systems would also be an integral factor in moderating these responses. The prevalence of orphan LuxR receptor systems in sequenced microbial genomes highlights the complexity of signaling interactions that remain to be identified and understood (Patankar and Gonzalez, 2009). Adopting community-like behaviors through QS systems may offer a distinct competitive advantage as bacteria can attach to a form a biofilm like structure within the environment of the sponge. Community-based small molecular interactions may also be important with respect to intracellular sponge symbionts, such as the recently reported Candidatus Endohaliclona renieramycinifaciens intracellular interaction with Haliclona (Tianero et al., 2019).
The prevalence and diversity of AHLs being produced by the sponge bacterial isolates identified in this study suggests that mechanisms to inhibit these systems may also exist within the sponge microenvironment. The identification of novel compounds that are capable of inhibiting AHL based QS systems is one of the key areas of focus in the development of next generation antimicrobials. Previously, we have reported on the profiling of a subset of this collection of marine sponge isolates for quorum sensing inhibitory (QSI) or QQ activity. A total of 18/440 culturable isolates were found to encode QSI, being able to supress AHL signaling is an isolate dependent manner (Gutierrez-Barranquero et al., 2017). It was interesting to note in that study that several species possessed dual QS and QSI activities. In this current study the finding that Psychrobacter sp. isolates from the same sponge collection were also capable of QS activity suggests that community level moderation of group behavior is a highly evolved trait in the marine ecosystem. Tan et al. (2015) previously showed how the dynamics of QS and QSI/QQ producing organisms can fluctuate in response to changes in environmental conditions. It is noticeable in this regard that two species cultured from QS treated sponge homogenate, Pseudomonas and Paenibacillus, are themselves known to possess AHL signaling systems (Ma et al., 2016). Understanding the interplay between QS and QSI/QQ in the marine sponge ecosystem and the role of QQ in moderating community behavior will underpin advances in marine ecology and beyond.
The dynamics of AHL production in marine microbial communities is seen as a mechanism to enhance culturability of rare genera, many of which encode valuable biosynthetic gene clusters for natural products such as antibiotics and anti-cancer drugs (Reen et al., 2015). While co-culture with QS positive isolates did alter the profile of culturable bacteria isolated from marine sponge homogenates, they failed to introduce new genera into culture. This of course could be due to limitations in the culture conditions, including the general nature of the media used which is more conducive to the culture of fast-growing bacteria. Dilution based methods and modification of the growth conditions with regard to media, temperature, and time may provide the optimum conditions for culture of QS dependent organisms (Rygaard et al., 2017).
The absence of a LuxIR system in the QS positive Pychrobacter sp. 230 isolate would suggest that a hidden diversity to the molecular mechanisms underpinning QS signaling remains to be elucidated. This is consistent with previous reports of AinS and LuxM family autoinducer synthase enoding genes, quite distinct from their LuxI counterparts (Venturi and Subramoni, 2009). Recently, a new LuxIR based system termed TswIR has been identified in an uncultured symbiont from the Red Sea Sponge Theonella swinhoei (Britstein et al., 2016). The synthase protein TswI (COG3916) was annotated as both an autoinducer synthase and a GNAT acetyltransferase activity and while GNAT acetyltransferase proteins were identified in the Psychrobacter genomes, no members of the COG3916 family were found. Furthermore, the recent finding that LuxIR homologs can synthesize and respond to non-acyl HSL signals, serves to underscore the hidden complexity in these systems (Ahlgren et al., 2011). Two orphan Photorhabdus LuxR proteins, PluR and PauR, sense alpha-pyrones and dialkylresorcinols, respectively (Brameyer and Heermann, 2015). It is possible that other examples of non-AHL LuxR interactions may be uncovered in the future, something that would add greatly to the complexity of the signaling interactions as currently understood. The absence of homologs of these proteins in the Psychrobacter sp. 230 genome may necessitate a functional approach in order to elucidate the molecular mechanism through which AHL signaling is established in this and other marine genera.
The datasets generated for this study can be found in the NCBI Database accession nos: MN209943-MN209952, NZ_SNVH00000000.1, SRP216019 and PRJNA555824.
FR and FO’G conceived the study. FR, JG-B, CA, DW, RM, SS, and KN performed the experimental analysis. FR wrote the manuscript with inputs from all the authors. FR and FO’G finalized the manuscript for submission.
FR and FO’G acknowledge support from Enterprise Ireland (CF-2017-0757-P) and the Health Research Board/Irish Thoracic Society (MRCG-2018-6). This research was also supported in part by grants awarded to FO’G by the European Commission (EU2020-634486-2015), Science Foundation Ireland (SSPC-2, 15/TIDA/2977), the Irish Research Council for Science, Engineering and Technology (GOIPG/2014/647), the Cystic Fibrosis Foundation, United States (OG1710), and the Health Research Board/Irish Thoracic Society (MRCG-2014-6). KN is grateful to Agilent technologies for the Thought Leader Donation of the UHPLC-QTOF system.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors thank Iwona Kozak and Niall Dunphy for excellent technical assistance and Jamie Deery for generation of the PCA plots.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2019.02131/full#supplementary-material
FIGURE S1 | PCA bi-plot cluster analysis of microbiome samples performed using R (v. 3.5.2). Visualization was performed using the ggplot and ggfortify packages. Samples from both sponges form separate clusters reflecting their distinct microbial community profiles.
FIGURE S2 | (A) Screen of Marine Sponge Isolates using QS-Biosensor Strains. (B) QS positive isolates were grown in culture flasks to confirm biosensor activation. Species level identification was achieved by 16S rRNA sequencing and subsequent BLAST analysis. (C) Extracts were validated by TLC overlay and subsequently sent for UHPLC-HRMS analysis and classification.
FIGURE S3 | (A–F) Growth profiling of isolates harvested following co-culture with QS active strains in the presence of 10 μM 3-oxo-C12-HSL or DMSO carrier control. Data presented is the average (±SEM) of two independent biological replicates with five technical replicates in each experiment performed on the BioScreen-C.
FIGURE S4 | Genome representation of the newly sequenced Psychrobacter sp. 230 isolate identified as a 3-oxo-C12-HSL producer in this study. COG functional categories are presented on the outer ring, while forward and reverse strand gene annotations are presented in the inner rings in red and blue, respectively.
Abbamondi, G. R., De Rosa, S., Iodice, C., and Tommonaro, G. (2014). Cyclic dipeptides produced by marine sponge-associated bacteria as quorum sensing signals. Nat. Prod. Comm. 9, 229–232.
Ahlgren, N. A., Harwood, C. S., Schaefer, A. L., Giraud, E., and Greenberg, E. P. (2011). Aryl-homoserine lactone quorum sensing in stem-nodulating photosynthetic bradyrhizobia. Proc. Natl. Acad. Sci. U.S.A. 108, 7183–7188. doi: 10.1073/pnas.1103821108
Blunt, J. W., Copp, B. R., Hu, W. P., Munro, M. H., Northcote, P. T., and Prinsep, M. R. (2009). Marine natural products. Nat. Prod. Rep. 26, 170–244. doi: 10.1039/b805113p
Brameyer, S., and Heermann, R. (2015). Specificity of signal-binding via Non-AHL LuxR-type receptors. PLoS One 10:e0124093. doi: 10.1371/journal.pone.0124093
Britstein, M., Devescovi, G., Handley, K. M., Malik, A., Haber, M., Saurav, K., et al. (2016). A new N-Acyl homoserine lactone synthase in an uncultured symbiont of the red sea sponge Theonella swinhoei. Appl. Environ. Microbiol. 82, 1274–1285. doi: 10.1128/AEM.03111-15
Britstein, M., Saurav, K., Teta, R., Sala, G. D., Bar-Shalom, R., Stoppelli, N., et al. (2018). Identification and chemical characterization of N-acyl-homoserine lactone quorum sensing signals across sponge species and time. FEMS Microbiol. Ecol. 94:fix182. doi: 10.1093/femsec/fix182
Bruhn, J. B., Dalsgaard, I., Nielsen, K. F., Buchholtz, C., Larsen, J. L., and Gram, L. (2005). Quorum sensing signal molecules (acylated homoserine lactones) in gram-negative fish pathogenic bacteria. Dis. Aquat. Organ. 65, 43–52. doi: 10.3354/dao065043
Chan, K. G., Wong, C. S., Yin, W. F., Sam, C. K., and Koh, C. L. (2010). Rapid degradation of N-3-oxo-acylhomoserine lactones by a Bacillus cereus isolate from Malaysian rainforest soil. Antonie Van Leeuwenhoek 98, 299–305. doi: 10.1007/s10482-010-9438-0
Cuadrado-Silva, C. T., Castellanos, L., Arevalo-Ferro, C., and Osorno, O. E. (2013). Detection of quorum sensing systems of bacteria isolated from fouled marine organisms. Biochem. Syst. Ecol. 46, 101–107. doi: 10.1016/j.bse.2012.09.010
Doberva, M., Stien, D., Sorres, J., Hue, N., Sanchez-Ferandin, S., Eparvier, V., et al. (2017). Large diversity and original structures of acyl-homoserine lactones in strain MOLA 401, a marine rhodobacteraceae bacterium. Front. Microbiol. 8:1152. doi: 10.3389/fmicb.2017.01152
Eren, A. M., Morrison, H. G., Lescault, P. J., Reveillaud, J., Vineis, J. H., and Sogin, M. L. (2015). Minimum entropy decomposition: unsupervised oligotyping for sensitive partitioning of high-throughput marker gene sequences. ISME J. 9, 968–979. doi: 10.1038/ismej.2014.195
Farrand, S. K., Hwang, I., and Cook, D. M. (1996). The tra region of the nopaline-type Ti plasmid is a chimera with elements related to the transfer systems of RSF1010, RP4, and F. J. Bacteriol. 178, 4233–4247. doi: 10.1128/jb.178.14.4233-4247.1996
Felsenstein, J. (1985). Confidence-limits on phylogenies - an approach using the bootstrap. Evolution 39, 783–791. doi: 10.1111/j.1558-5646.1985.tb00420.x
Gram, L., Grossart, H. P., Schlingloff, A., and Kiorboe, T. (2002). Possible quorum sensing in marine snow bacteria: production of acylated homoserine lactones by Roseobacter strains isolated from marine snow. Appl. Environ. Microbiol. 68, 4111–4116. doi: 10.1128/aem.68.8.4111-4116.2002
Gutierrez-Barranquero, J. A., Reen, F. J., Parages, M. L., McCarthy, R., Dobson, A. D. W., and O’Gara, F. (2017). Disruption of N-acyl-homoserine lactone-specific signalling and virulence in clinical pathogens by marine sponge bacteria. Microb. Biotechnol. 12, 1049–1063. doi: 10.1111/1751-7915.12867
Hmelo, L. R. (2017). Quorum sensing in marine microbial environments. Annu. Rev. Mar. Sci. 9, 257–281. doi: 10.1146/annurev-marine-010816-060656
Huang, Y. L., Ki, J. S., Lee, O. O., and Qian, P. Y. (2009). Evidence for the dynamics of Acyl homoserine lactone and AHL-producing bacteria during subtidal biofilm formation. ISME J. 3, 296–304. doi: 10.1038/ismej.2008.105
Jatt, A. N., Tang, K., Liu, J., Zhang, Z., and Zhang, X. H. (2015). Quorum sensing in marine snow and its possible influence on production of extracellular hydrolytic enzymes in marine snow bacterium Pantoea ananatis B9. FEMS Microbiol. Ecol. 91, 1–13. doi: 10.1093/femsec/fiu030
Kennedy, J., Codling, C. E., Jones, B. V., Dobson, A. D. W., and Marchesi, J. R. (2008). Diversity of microbes associated with the marine sponge, Haliclona simulans, isolated from Irish waters and identification of polyketide synthase genes from the sponge metagenome. Environ. Microbiol. 10, 1888–1902. doi: 10.1111/j.1462-2920.2008.01614.x
Kildgaard, S., Mansson, M., Dosen, I., Klitgaard, A., Frisvad, J. C., Larsen, T. O., et al. (2014). Accurate dereplication of bioactive secondary metabolites from marine-derived fungi by UHPLC-DAD-QTOFMS and a MS/HRMS library. Mar. Drugs 12, 3681–3705. doi: 10.3390/md12063681
Kumar, S., Stecher, G., Li, M., Knyaz, C., and Tamura, K. (2018). MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 35, 1547–1549. doi: 10.1093/molbev/msy096
Ma, Z. P., Lao, Y. M., Jin, H., Lin, G. H., Cai, Z. H., and Zhou, J. (2016). Diverse profiles of AI-1 type quorum sensing molecules in cultivable bacteria from the Mangrove (Kandelia obovata) rhizosphere environment. Front. Microbiol. 7:1957. doi: 10.3389/fmicb.2016.01957
Ma, Z. P., Song, Y., Cai, Z. H., Lin, Z. J., Lin, G. H., Wang, Y., et al. (2018). Anti-quorum sensing activities of selected coral symbiotic bacterial extracts from the South China Sea. Front. Cell. Infect. Microbiol. 8:144. doi: 10.3389/fcimb.2018.00144
Mayer, A. M. S., Glaser, K. B., Cuevas, C., Jacobs, R. S., Kem, W., Little, R. D., et al. (2010). The odyssey of marine pharmaceuticals: a current pipeline perspective. Trends Pharmacol. Sci. 31, 255–265. doi: 10.1016/j.tips.2010.02.005
Mayer, A. M. S., Rodriguez, A. D., Berlinck, R. G. S., and Fusetani, N. (2011). Marine pharmacology in 2007-8: marine compounds with antibacterial, anticoagulant, antifungal, anti-inflammatory, antimalarial, antiprotozoal, antituberculosis, and antiviral activities; affecting the immune and nervous system, and other miscellaneous mechanisms of action. Compar. Biochem. Physiol. C-Toxicol. Pharmacol. 153, 191–222. doi: 10.1016/j.cbpc.2010.08.008
McClean, K. H., Winson, M. K., Fish, L., Taylor, A., Chhabra, S. R., Camara, M., et al. (1997). Quorum sensing and Chromobacterium violaceum: exploitation of violacein production and inhibition for the detection of N-acylhomoserine lactones. Microbiology 143, 3703–3711. doi: 10.1099/00221287-143-12-3703
Mohamed, N. M., Cicirelli, E. M., Kan, J., Chen, F., Fuqua, C., and Hill, R. T. (2008). Diversity and quorum-sensing signal production of Proteobacteria associated with marine sponges. Environ. Microbiol. 10, 75–86. doi: 10.1111/j.1462-2920.2007.01431.x
Nealson, K. H., and Hastings, J. W. (1979). Bacterial bioluminescence: its control and ecological significance. Microbiol. Rev. 43, 496–518.
Patankar, A. V., and Gonzalez, J. E. (2009). Orphan LuxR regulators of quorum sensing. FEMS Microbiol. Rev. 33, 739–756. doi: 10.1111/j.1574-6976.2009.00163.x
Phelan, R. W., O’Halloran, J. A., Kennedy, J., Morrissey, J. P., Dobson, A. D., O’Gara, F., et al. (2012). Diversity and bioactive potential of endospore-forming bacteria cultured from the marine sponge Haliclona simulans. J. Appl. Microbiol. 112, 65–78. doi: 10.1111/j.1365-2672.2011.05173.x
Poulter, S., Carlton, T. M., Su, X. B., Spring, D. R., and Salmond, G. P. C. (2010). Engineering of new prodigiosin-based biosensors of Serratia for facile detection of short-chain N-acyl homoserine lactone quorum-sensing molecules. Environ. Microbiol. Rep. 2, 322–328. doi: 10.1111/j.1758-2229.2010.00140.x
Purohit, A. A., Johansen, J. A., Hansen, H., Leiros, H. K., Kashulin, A., Karlsen, C., et al. (2013). Presence of acyl-homoserine lactones in 57 members of the Vibrionaceae family. J. Appl. Microbiol. 115, 835–847. doi: 10.1111/jam.12264
Rasch, M., Kastbjerg, V. G., Bruhn, J. B., Dalsgaard, I., Givskov, M., and Gram, L. (2007). Quorum sensing signals are produced by Aeromonas salmonicida and quorum sensing inhibitors can reduce production of a potential virulence factor. Dis. Aquat. Organ. 78, 105–113. doi: 10.3354/dao01865
Reen, F. J., Romano, S., Dobson, A. D., and O’Gara, F. (2015). The sound of silence: activating silent biosynthetic gene clusters in marine microorganisms. Mar. Drugs 13, 4754–4783. doi: 10.3390/md13084754
Romero, M., Martin-Cuadrado, A. B., and Otero, A. (2012). Determination of whether quorum quenching is a common activity in marine bacteria by analysis of cultivable bacteria and metagenomic sequences. Appl. Environ. Microbiol. 78, 6345–6348. doi: 10.1128/AEM.01266-12
Rygaard, A. M., Thogersen, M. S., Nielsen, K. F., Gram, L., and Bentzon-Tilia, M. (2017). Effects of gelling agent and extracellular signaling molecules on the culturability of marine bacteria. Appl. Environ. Microbiol. 83:e00243-17. doi: 10.1128/AEM.00243-17
Saitou, N., and Nei, M. (1987). The neighbor-joining method - a new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 4, 406–425.
Sievers, F., Wilm, A., Dineen, D., Gibson, T. J., Karplus, K., Li, W., et al. (2011). Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 7:539. doi: 10.1038/msb.2011.75
Steindler, L., and Venturi, V. (2007). Detection of quorum-sensing N-acyl homoserine lactone signal molecules by bacterial biosensors. FEMS Microbiol. Lett. 266, 1–9. doi: 10.1111/j.1574-6968.2006.00501.x
Tahrioui, A., Quesada, E., and Llamas, I. (2011). The hanR/hanI quorum-sensing system of Halomonas anticariensis, a moderately halophilic bacterium. Microbiology 157, 3378–3387. doi: 10.1099/mic.0.052167-0
Tajima, F., and Nei, M. (1984). Estimation of evolutionary distance between nucleotide-sequences. Mol. Biol. Evol. 1, 269–285.
Tan, C. H., Koh, K. S., Xie, C., Zhang, J., Tan, X. H., Lee, G. P., et al. (2015). Community quorum sensing signalling and quenching: microbial granular biofilm assembly. NPJ Biofilms Microbiomes 1:15006. doi: 10.1038/npjbiofilms.2015.6
Taylor, M. W., Radax, R., Steger, D., and Wagner, M. (2007). Sponge-associated microorganisms: evolution, ecology, and biotechnological potential. Microbiol. Mol. Biol. Rev. 71, 295–347. doi: 10.1128/mmbr.00040-06
Taylor, M. W., Schupp, P. J., Baillie, H. J., Charlton, T. S., de Nys, R., Kjelleberg, S., et al. (2004a). Evidence for acyl homoserine lactone signal production in bacteria associated with marine sponges. Appl. Environ. Microbiol. 70, 4387–4389. doi: 10.1128/aem.70.7.4387-4389.2004
Taylor, M. W., Schupp, P. J., Dahllof, I., Kjelleberg, S., and Steinberg, P. D. (2004b). Host specificity in marine sponge-associated bacteria, and potential implications for marine microbial diversity. Environ. Microbiol. 6, 121–130. doi: 10.1046/j.1462-2920.2003.00545.x
Tianero, M. D., Balaich, J. N., and Donia, M. S. (2019). Localized production of defence chemicals by intracellular symbionts of Haliclona sponges. Nat. Microbiol. 4, 1149–1159. doi: 10.1038/s41564-019-0415-8
Venturi, V., and Subramoni, S. (2009). Future research trends in the major chemical language of bacteria. HFSP J. 3, 105–116. doi: 10.2976/1.3065673
Wang, G. (2006). Diversity and biotechnological potential of the sponge-associated microbial consortia. J. Ind. Microbiol. Biotechnol. 33, 545–551. doi: 10.1007/s10295-006-0123-2
Weiland-Brauer, N., Fischer, M. A., Pinnow, N., and Schmitz, R. A. (2019). Potential role of host-derived quorum quenching in modulating bacterial colonization in the moon jellyfish Aurelia aurita. Sci. Rep. 9:34. doi: 10.1038/s41598-018-37321-z
Whiteley, M., Diggle, S. P., and Greenberg, E. P. (2017). Progress in and promise of bacterial quorum sensing research. Nature 551, 313–320. doi: 10.1038/nature24624
Yang, Q., Han, Y., and Zhang, X. H. (2011). Detection of quorum sensing signal molecules in the family Vibrionaceae. J. Appl. Microbiol. 110, 1438–1448. doi: 10.1111/j.1365-2672.2011.04998.x
Yin, W. F., Purmal, K., Chin, S., Chan, X. Y., and Chan, K. G. (2012). Long chain N-acyl homoserine lactone production by Enterobacter sp. isolated from human tongue surfaces. Sensors 12, 14307–14314. doi: 10.3390/s121114307
Zan, J., Cicirelli, E. M., Mohamed, N. M., Sibhatu, H., Kroll, S., Choi, O., et al. (2012). A complex LuxR-LuxI type quorum sensing network in a roseobacterial marine sponge symbiont activates flagellar motility and inhibits biofilm formation. Mol. Microbiol. 85, 916–933. doi: 10.1111/j.1365-2958.2012.08149.x
Keywords: quorum sensing (QS), microbiome, marine sponge-associated bacteria, cell–cell communication, acyl homoserine lactone (AHL)
Citation: Reen FJ, Gutiérrez-Barranquero JA, McCarthy RR, Woods DF, Scarciglia S, Adams C, Fog Nielsen K, Gram L and O’Gara F (2019) Quorum Sensing Signaling Alters Virulence Potential and Population Dynamics in Complex Microbiome-Host Interactomes. Front. Microbiol. 10:2131. doi: 10.3389/fmicb.2019.02131
Received: 20 May 2019; Accepted: 29 August 2019;
Published: 11 September 2019.
Edited by:
Ana Maria Otero, University of Santiago de Compostela, SpainReviewed by:
Jürgen Tomasch, Helmholtz Center for Infection Research, Helmholtz Association of German Research Centers (HZ), GermanyCopyright © 2019 Reen, Gutiérrez-Barranquero, McCarthy, Woods, Scarciglia, Adams, Fog Nielsen, Gram and O’Gara. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Fergal O’Gara, Zi5vZ2FyYUB1Y2MuaWU=
†Present address: José A. Gutiérrez-Barranquero, Facultad de Ciencias, Departamento de Microbiología, Instituto de Hortofruticultura Subtropical y Mediterránea La Mayora (IHSM-UMA-CSIC), Universidad de Málaga, Málaga, Spain Ronan R. McCarthy, Department of Life Sciences, Brunel University London, London, United Kingdom
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.