
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol., 25 June 2019
Sec. Antimicrobials, Resistance and Chemotherapy
Volume 10 - 2019 | https://doi.org/10.3389/fmicb.2019.01394
This article is part of the Research Topic"One Health" Approach For Revealing Reservoirs And Transmission Of Antimicrobial ResistanceView all 27 articles
 Katia Suemi Gozi1
Katia Suemi Gozi1 Juliana Rodrigues Froes1
Juliana Rodrigues Froes1 Luana Perpetua Tobias Deus Ajude1
Luana Perpetua Tobias Deus Ajude1 Caroline Rodrigues da Silva1
Caroline Rodrigues da Silva1 Rafaela Speranza Baptista2
Rafaela Speranza Baptista2 Juliana Regina Peiró2
Juliana Regina Peiró2 Marcia Marinho2
Marcia Marinho2 Luiz Claudio Nogueira Mendes2
Luiz Claudio Nogueira Mendes2 Mara Corrêa Lelles Nogueira1
Mara Corrêa Lelles Nogueira1 Tiago Casella1,3*
Tiago Casella1,3*Antimicrobial resistance (AR) is a public health issue since it limits the choices to treat infections by Escherichia coli in humans and animals. In Brazil, the ovine meat market has grown in recent years, but studies about AR in sheep are still scarce. Thus, this study aims to investigate the presence of AR in E. coli isolated from lambs during feedlot. To this end, feces from 112 lambs with 2 months of age, after weaning, were collected on the first day of the animals in the feedlot (day 0), and on the last day before slaughtering (day 42). Isolates were selected in MacConkey agar supplemented with 4 mg/L of ceftiofur and identified by biochemical methods. Isolates were submitted to an antimicrobial susceptibility test by disc-diffusion and PCR to investigate genes for phylogenetic group, virulence determinants and resistance to the several antimicrobial classes tested. The genetic localization of the bla genes detected was elucidated by S1-PFGE followed by Southern blot-hybridizations. The isolates were typed by XbaI-PFGE and MLST methods. Seventy-eight E. coli were isolated from 8/112 (7.1%) animals on day 0, and from 55/112 (49.1%) animals on day 42. Since only fimH was present in almost all E. coli (97.4%) as a virulence gene, and also 88.5% belonged to phylogroups B1 or A, we consider that isolates represent intestinal commensal bacteria. The dendrogram separated the 78 non-virulent isolates in seven clusters, two of which comprised 50 E. coli belonging to ST/CC 1727/446 or ST 3994 recovered on day 42 commonly harboring the genotype blaCMY -2-aac(3)-IIa -tetA-sul1-sul2-floR-cmlA. Special attention should be given to the presence of blaCTX-M-15, a worldwide gene spread, and blaCTX-M-14, a hitherto undetected gene in Enterobacteriaceae from food-producing animals in Brazil. Importantly, E. coli lineages and plasmids carrying bla genes detected here have already been reported as sources of infection in humans either from animals, food, or the environment, which raises public health concerns. Hence, two types of commensal E. coli carrying important AR genes clearly prevailed during feedlot, but lambs are also reservoirs of bacteria carrying important AR genes such as blaCTX-M-14 and blaCTX-M-15, mostly related to antimicrobial treatment failure.
The use of antimicrobial agents in humans and animals can cause the emergence and dissemination of antimicrobial resistance (AR) in pathogens, which may compromise the effective treatment of infections in humans (Kaesbohrer et al., 2012). International public health agencies have reported the potential link and risks between the overuse or misuse of antimicrobials in veterinary practices and the emergence of human resistant pathogens, which encourage surveillance of AR and antimicrobial use worldwide (EFSA, 2011; WHO, 2017). Human exposure to AR bacteria through direct contact with animals, consumption and handling of contaminated food, and bacteria released into the environment may contribute to the spread of AR determinants (Kaesbohrer et al., 2012).
Infections caused by AR E. coli and their isolation from food-producing animals are increasing worldwide (EFSA, 2008, 2011; Kaesbohrer et al., 2012). This scenario is regarded as a consequence of the selective pressure exerted on the gastrointestinal tract (GIT) of the animals by the overuse of antimicrobials (Graham et al., 2017). During slaughtering, the carcass may be contaminated and AR commensal or pathogenic bacteria might reach humans through the food chain (Cyoia et al., 2019; Projahn et al., 2019). The relationship between AR strains isolated from humans and the food chain has been already reported (Belmar Campos et al., 2014). Therefore, the monitoring of commensal bacteria is important since it constitutes a reservoir of AR genes, which allows the tracking of emerging resistance in livestock and possible spread to animal-derived food and other zoonotic pathogens (EFSA, 2008; Kaesbohrer et al., 2012; Madec and Haenni, 2018).
The majority of studies about antimicrobial use and resistance in food-producing animals are carried out on cattle, chickens, and pigs, but in regard to other food-producing flocks, such as sheep, information is scarce. Little is known about AR in sheep in Brazil, despite the increased consumption of lamb meat (FAO, 2018). Therefore, this study aims to determine the distribution of AR E. coli in the fecal microbiota of feedlot lambs in Brazil.
A special feedlot comprising 140 lambs with 2 months of age, after weaning, coming from 35 different farms in the State of São Paulo, Southeastern Brazil was chosen for this study. Stool samples were collected weekly from the rectum of the animals for parasitological screening between September 14, 2016 and October 27, 2016, under the Ethics Committee approval number FOA00845-2017. Trimethoprim/sulfamethoxazole was used to prevent and to treat clinical manifestations of respiratory disease, and florfenicol was used to treat infectious keratoconjunctivitis. Stool samples from 112 lambs were collected immediately after the arrival of the sheep at the feedlot (day 0) and then on the day before the slaughtering of the animals (day 42) to further investigate the presence of AR E. coli.
About one gram of feces was diluted in 5 mL of sterile NaCl 0.9% and directly inoculated onto MacConkey agar (Oxoid) supplemented with 4 mg/L of ceftiofur (Lapisa). Following incubation at 37°C for 18–24 h, one of each of the different presumptive E. coli colonies (i.e., pinkish round colony due to lactose fermenting, dry to little mucous aspect, and characteristic odor) were selected for identification by biochemical essays using a commercial kit (NewProv) and further characterization described below.
Antimicrobial susceptibility testing was performed following the Clinical and Laboratory Standards Institute (CLSI, 2017) guidelines using the disc diffusion method. Bacterial susceptibility to 13 beta-lactam and non-beta-lactam antibiotics (Oxoid) of veterinary and human interest was tested: amoxicillin/clavulanate, ceftazidime, cefotaxime, ceftiofur, cefoxitin, ertapenem, amikacin, gentamicin, enrofloxacin, nalidixic acid, tetracycline, trimethoprim/sulfamethoxazole, florfenicol, and chloramphenicol. Parallel to the antimicrobial susceptibility test, the phenotypic test for production of extended-spectrum beta-lactamase was performed by the Modified Double Disc Synergy Test (Kaur et al., 2013). E. coli ATCC 25922 and Klebsiella pneumoniae ATCC 700603 were used as quality control strains.
Investigation of the main plasmidial genes associated with cephalosporins resistance (blaCTX-M and blaCMY), aminoglycosides resistance [aac(3)-Ia, aac(3)-IIa, acc(6′)-Ih, ant(2″)-Ia, aph(3′)-VI, aph(3′)-Ia and aac(6′)-Ib], quinolones resistance (qnrA, qnrB, qnrC, qnrD, qnrS, qepAB, and oqxAB), tetracycline resistance (tetA, tetB, and tetC), trimethoprim resistance (dfr Ia, dfr VII, and dfr XII), sulphas resistance (sul1 and sul2), and phenicols resistance (floR, cat and cmlA) was performed in each respective resistant strain by PCR according to previous protocols (Supplementary Table 1). Products of bla genes were sequenced using the corresponding primers in order to identify the variant detected and analyzed using BLAST1.
The following 20 virulence genes, which have been associated with Extraintestinal Pathogenic E. coli strains, were investigated by PCR as previously described (Supplementary Table 1): fimH, papEF, papG I, papG II, papG III, sfa/focDE, sfaS, focG, afa/draBC, nfaE, kpsMT K1, kpsMT K5, hlyA, cnf1, cdtB, sat, vat, fyuA, iutA, and iroN. Since sheep are known as an important source of Shiga-toxin producing-E. coli (STEC) (Vettorato et al., 2009), the stx1 and stx2 genes, as well as the aggR and the eae genes, associated with Enteroaggregative E. coli (EAEC) and Enteropathogenic E. coli (EPEC), respectively, were additionally searched by PCR according to previous protocols (Supplementary Table 1). E. coli isolates were also submitted to phylogenetic grouping for predicting of commensal or pathogenic isolates as previously described (Clermont et al., 2000; Doumith et al., 2012).
Replicon of the plasmids of the isolates was detected by PCR-based Replicon Typing scheme (Carattoli et al., 2005; Villa et al., 2010) using the PBRT 2.0 kit (DIATHEVA). S1 enzyme (Promega) was used for 45 min to linearize plasmids and results were visualized in Pulsed-Field Gel Electrophoresis (S1-PFGE) for 20 h with initial switch time = 1 s and final switch time = 30 s on an electric field of 6 V/cm. Southern blot-hybridization analysis on S1-PFGE gels was performed using adequate probes and the kits AmershamTM AlkPhos Direct Labeling Reagents and AmershamTM CDP-StarTM Detection Reagent (GE Healthcare).
Bacterial DNA was typed by restriction with XbaI (Thermo Scientific) followed by a PFGE (XbaI-PFGE) for 22 h with initial switch time = 2.2 s and final switch time = 54.2 s, and 6 V/cm. The software BioNumericsTM version 7.6.3 (Applied Maths) was used for dendrogram construction and clustering based on the band-based Dice’s similarity coefficient and the unweighted pair group method using arithmetic averages. Isolates were considered to belong to the same cluster when the similarity coefficient was ≥90%.
Escherichia coli isolates were additionally submitted to Multilocus Sequence Typing according to the Achtman’s scheme2.
The bla genes sequences reported in this study have been deposited to GenBank under accession numbers MK896925 to MK896944 and MK917695 to MK917713.
Eight CTX-M-producing E. coli were isolated from eight animals on day 0, and 70 CTX-M- or CMY-2-producing E. coli were isolated from 55 lambs on day 42 (Figure 1 and Table 1). All 78 isolates presented resistance to at least one of the third-generation cephalosporins – 3GC tested (ceftazidime, cefotaxime, ceftiofur). The 53 CMY-2-producing E. coli presenting resistance to amoxicillin/clavulanic acid also presented resistance to the cephamycin cefoxitin (Figure 1). More than 80% of the isolates presented additional resistance to at least one of the phenicols tested (68, 87.2%), to tetracycline (66 isolates, 84.6%), to trimethoprim/sulfamethoxazole (65, 83.3%), and at least one of the aminoglycosides tested (64, 82.0%). Only seven isolates (9.0%) presented resistance to nalidixic acid and/or enrofloxacin, and all E. coli were susceptible to ertapenem (Table 1).

Figure 1. Dendrogram obtained from XbaI-PFGE typing of the 78 E. coli isolated. Dendrogram was constructed using Optimization 0% and Tolerance 1.5%. AMC, amoxicillin/clavulanate; CAZ, ceftazidime; CTX, cefotaxime; EFT, ceftiofur; FOX, cefoxitin; ETP, ertapenem; AK, amikacin; CN, gentamicin; ENR, enrofloxacin; NA, nalidixic acid; TE, tetracycline; STX, trimethoprim/sulfamethoxazole; FFC, florfenicol; C, chloramphenicol. Antimicrobial Susceptibility Profile squares: black, resistance; gray, intermediate resistance; white, susceptibility. Virulence genes squares: black, presence; white, absence. PhG, phylogenetic group. ST/CC, Sequence Type/Clonal Complex. ND, localization not detected. Isolation dates “Sep 14th, 2016” and “Oct 27th, 2016” refers to “day 0” and “day 42,” respectively.
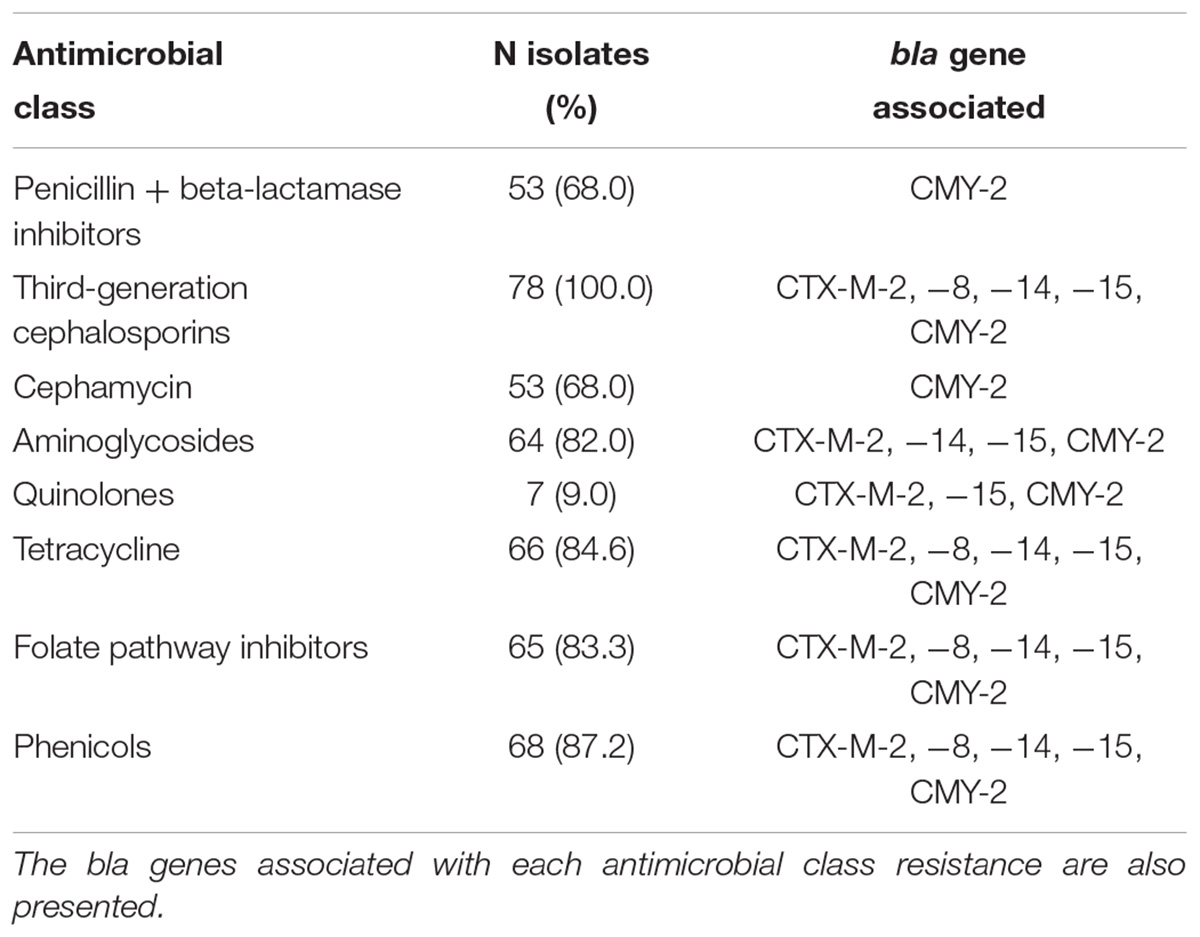
Table 1. Isolates presenting resistance to each antimicrobial class among the 78 E. coli from stools of sheep in Southeastern Brazil.
In total, 18 genes responsible for antimicrobial resistance were detected in this study, and all of the 78 isolates presented blaCMY -2 or blaCTX-M genes (Figure 1 and Table 2). The genes blaCTX-M-8, blaCTX-M-14, and blaCTX-M-15 were identified in the isolates from day 0 harbored by plasmids IncI1 of ∼97 kb or IncHI1 ∼194 kb for blaCTX-M-8, and plasmid FII of about 97 kb for blaCTX-M-15. We could not detect plasmids harboring blaCTX-M-14. The blaCTX-M-2, blaCTX-M-8, blaCTX-M-15, and blaCMY -2 were identified in isolates recovered on day 42 inserted into the chromosome in the case of blaCTX-M-2, and harbored by plasmids IncI1 of about 95 kb or 97 kb for blaCTX-M-8, plasmid IncHI2 of ∼335 kb for blaCTX-M-15, and plasmid IncA/C of ∼170 kb for all blaCMY -2 (Table 3). Regarding resistance to aminoglycosides, especially gentamicin, the aac(3)-IIa gene was detected in 60 isolates (76.9%) on days 0 and 42 of feedlot while the ant(2″)-Ia gene was detected only in five isolates (6.4%) on day 42. The qnrB gene was the only one detected as responsible for quinolone non-susceptibility, present in six isolates (7.7%) obtained on day 42. The tetA and tetB genes, responsible for tetracycline resistance, were detected in 54 (69.2%) and 10 (12.8%) isolates, respectively, on days 0 and 42 of feedlot. Concerning resistance to trimethoprim, the dfr VII gene was detected only on day 0 of feedlot in four isolates (5.1%), and dfr Ia and dfr XII were detected only on day 42 in 16 (20.5%) and 21 (26.9%) isolates, respectively. Resistance to sulphas was detected at both the first and last days of feedlot, with 61 isolates (78.2%) carrying sul1, and 65 (83.3%) carrying the sul2 gene. Lastly, in regard to phenicols resistance, the floR and the cmlA genes were detected in 65 (83.3%) and 67 (85.9%) isolates, respectively, while the cat gene was detected in only eight isolates (10.3%); all recovered on both days 0 and 42 of feedlot (Figure 1 and Table 2).
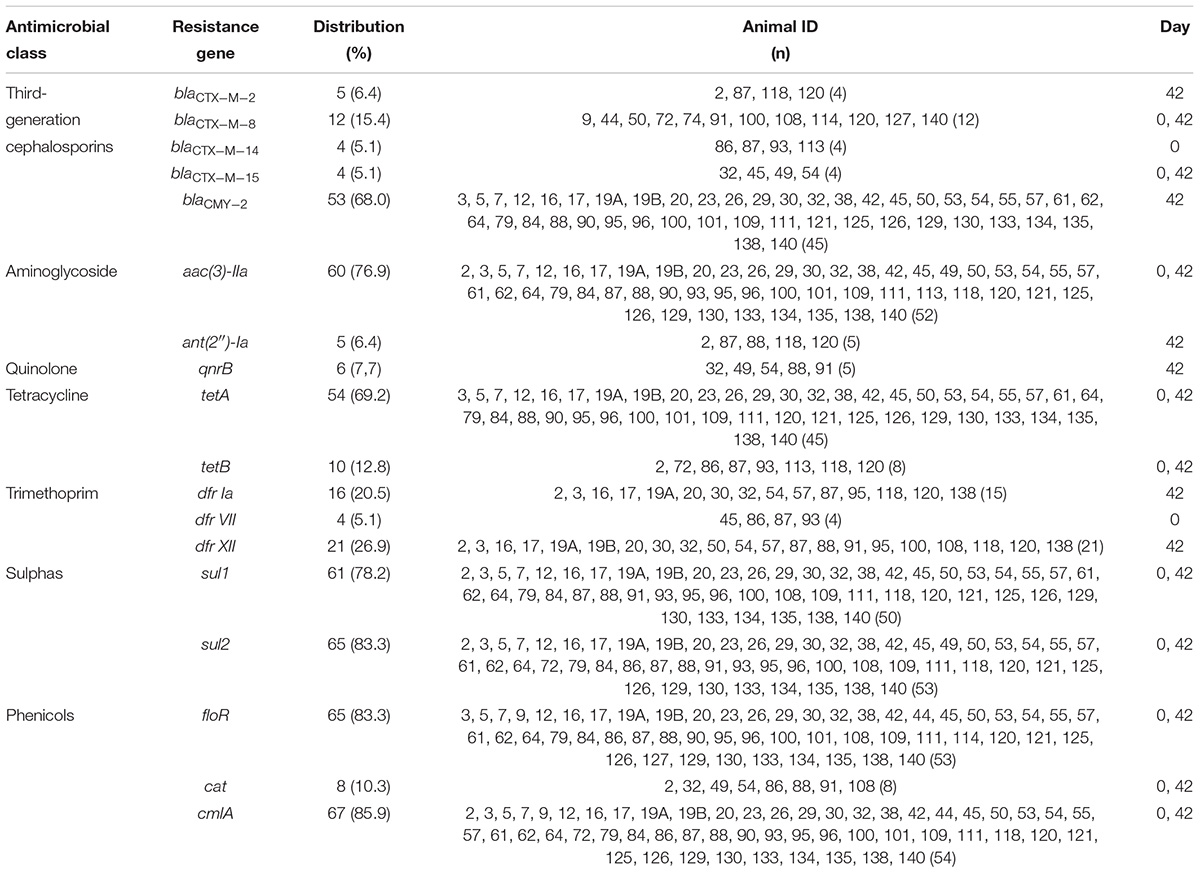
Table 2. Antimicrobial resistance genes distribution among the 78 E. coli from stools of sheep in Southeastern Brazil, according to the animals (ID) and the day of feedlot they were detected.
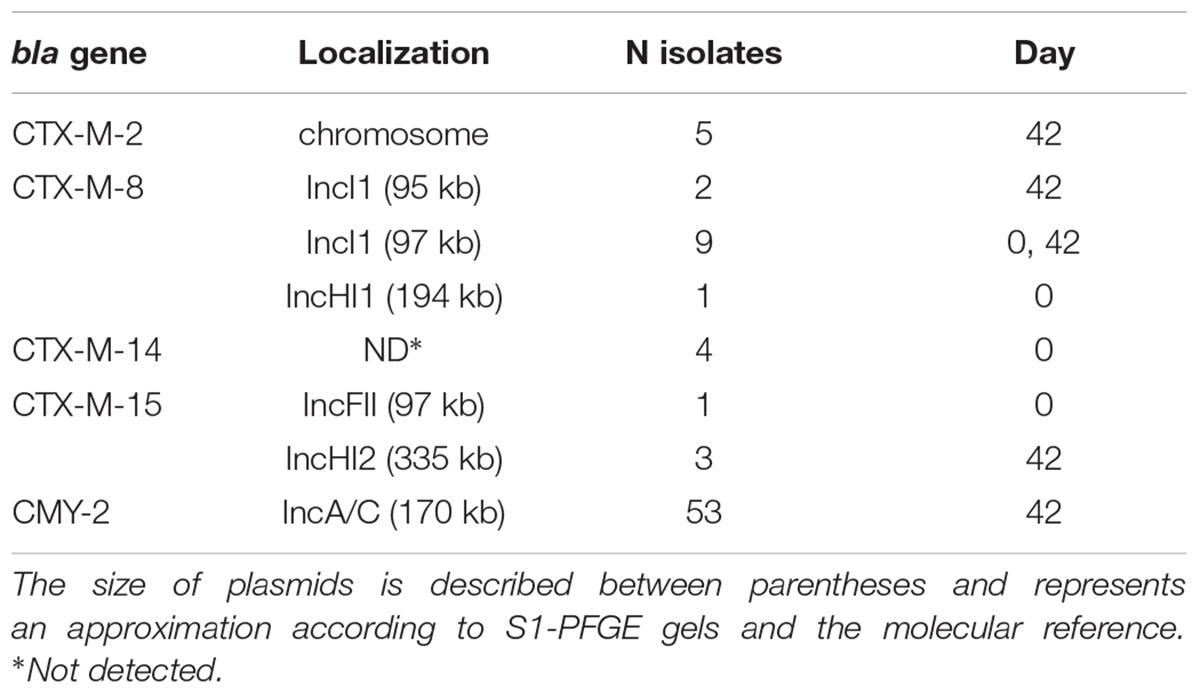
Table 3. Genetic localization of bla genes detected in the 78 E. coli isolates.
Four virulence genes were detected, but only fimH was present in the majority (76 isolates, 97.4%). The papEF was detected in 3 isolates (3.8%), and papG II and fyuA in 2 (2.6%), as presented in Figure 1. Furthermore, five genotypes concerning virulence were detected, including the absence of any gene, the presence of only fimH or a combination of it and the other genes detected (Table 4). No genes predictive of STEC, EAEC or EPEC were detected. Forty-eight (61.5%) E. coli belonged to phylogroup B1, 21 (27.0%) to phylogroup A, 5 (6.4%) to phylogroup D, and 4 (5.1%) to phylogroup B2. Phylogroup D was related only to the blaCTX-M-2 gene (Table 4).
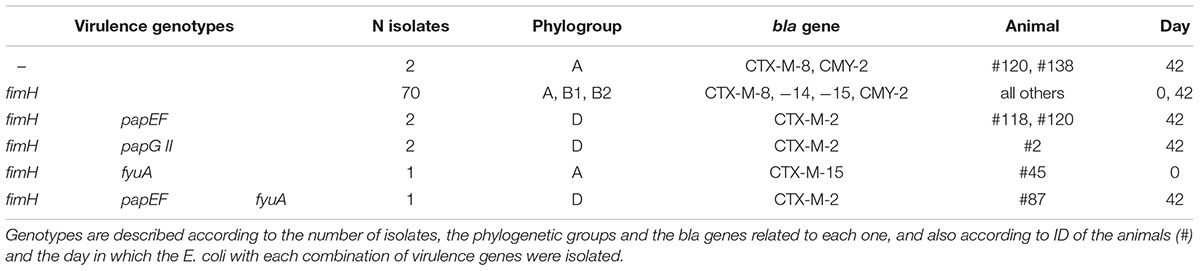
Table 4. Genotypes detected concerning virulence genes in the 78 E. coli isolates from stools of sheep in Southeastern Brazil.
The XbaI-PFGE typing distinguished the 78 isolates in seven major clusters. Two lambs (animals #45 and #87) presented AR E. coli (isolates 0.45E1 and 42.45E1, and 0.87E1 and 42.87E1, respectively) on both days 0 and 42, but the strains are not similar by XbaI-PFGE and neither carry the same AR genes (Figure 1). Typing with the MLST scheme revealed 10 Sequence Types (ST) of E. coli in the studied feedlot, and one new allele profile in seven blaCTX-M-8-carrying isolates (0.44E1, 42.91E1, 42.108E1, 42.114E1, 42.120E2, 42.127E1, and 42.140E1) recovered on the first and last days of feedlot. The ST 1727 Clonal Complex (CC) 446 was predominant and present only on day 42 in 52.6% of the total isolates regarding just blaCMY -2-carrying E. coli from phylogenetic group B1, followed by the ST 3994, the new combination of MLST alleles, ST/CC 58/155, ST 6618, ST 1585, and the ST/CC 410/23, ST 457, ST 962, ST 1642, ST 5204 (Table 5).
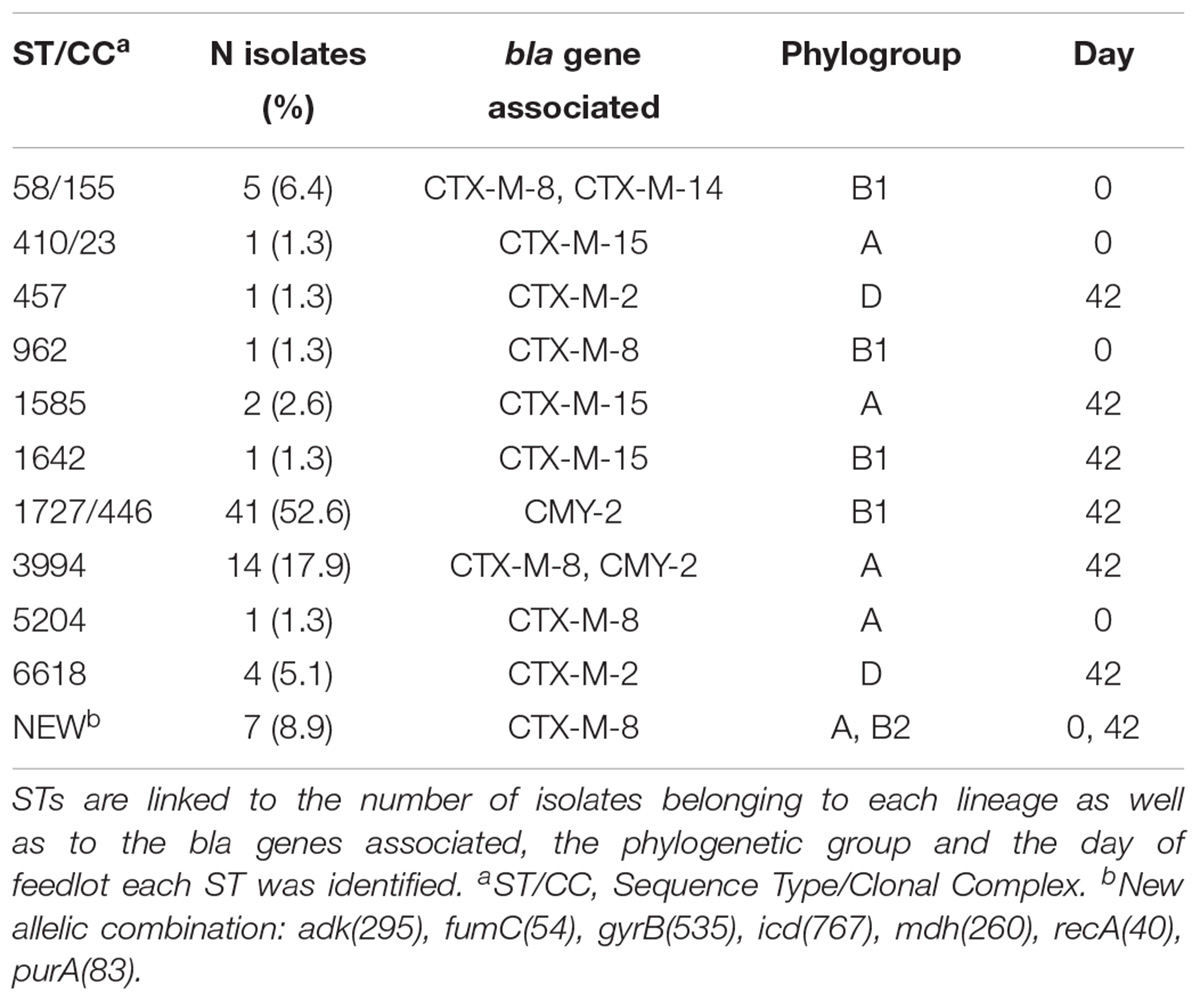
Table 5. Sequence Types and Clonal Complexes detected for the E. coli isolated from sheep in Southeastern Brazil.
Seventy-four (94.9%) E. coli isolates presented a multidrug-resistant antibiotype (MRAb) according to the antimicrobial susceptibility test, with the exception of the isolate 0.74E1, recovered on day 0 from animal #74, and the isolates 42.120E2, 42.127E1, 42.140E1, obtained on day 42 from animals #120, #127 and #140. Interestingly, all the non-MRAb E. coli were associated with blaCTX-M-8 gene (Figure 1). The high percentage of MRAb isolates illustrates the potential for spread of AR bacteria through a flock. Studies have already reported that the resistance rate to some antimicrobials rises during cattle or pig feedlot because of antimicrobial usage (Benedict et al., 2015; Gibbons et al., 2016; Weinroth et al., 2018). However, a Canadian study surveyed sheep flocks over a 1-year period and found no significant difference between the initial and the final visits (Scott et al., 2012), which is in disagreement with this study.
No isolate presented resistance to ertapenem (Table 1), which could be related to the fact that carbapenems are not approved for use in animals (OIE, 2018). The detection of AR E. coli in 55 animals after feedlot, in comparison to eight animals on day 0, indicates a selection pressure acting on the flock. Beta-lactams, florfenicol, macrolides, quinolones, tetracycline, and trimethoprim/sulfamethoxazole are administered in sheep (OIE, 2018). In fact, some animals included in this study received florfenicol or trimethoprim/sulfamethoxazole, and this could explain the presence of the AR E. coli because of direct or co-selection of resistance determinants in the GIT of the animals (Collignon et al., 2016; Makita et al., 2016; Knudsen et al., 2018).
The set of genes codifying beta-lactamase enzymes carried by the E. coli isolated on the 2 days of analysis was diverse. On day 0, 7.1% (8/112 animals) of the sampled lambs presented E. coli harboring some blaCTX-M-variant. However, after 42 days of feedlot the majority of E. coli isolated (53/78, 68.0% of the total) harbored the blaCMY -2 gene, comprising essentially the two great clusters of the dendrogram and the ST/CC 1727/446 and ST 3994 (Figure 1). Besides, blaCMY -2, the blaCTX-M-2 gene was detected only on day 42, while blaCTX-M-14 was detected in isolates recovered only on day 0 (Table 2). It seems that the first two genes entered into the flock during feedlot by some external factor such as surrounding animals, insects, or the environment (Blaak et al., 2015; Huijbers et al., 2015; Solà-Ginés et al., 2015), and the latter disappeared during feedlot perhaps because of competition between the blaCTX-M-14-carrying E. coli and other more successful strains, possibly the blaCMY -2-carrying E. coli. On the other hand, blaCTX-M-8 and blaCTX-M-15 were present on the first day of feedlot and persisted until the end (Table 2), which is clearly not linked to the maintenance of isolates into the feedlot, since the CTX-M-8- and the CTX-M-15-producing E. coli isolated on days 0 and 42 are not clonally related by PFGE or MLST (Figure 1). However, the majority of blaCTX-M-8 detected in isolates from day 42 are harbored by IncI1 plasmids of ∼97 kb, the same as two detected on day 0, which illustrates the maintenance and spreading of that plasmid through the feedlot. On the other hand, the blaCTX-M-15 gene identified in three E. coli recovered on day 42 probably entered the feedlot at some point since they are harbored by plasmid IncHI2 of ∼335 kb, differently from the blaCTX-M-15 harbored by an IncFII of ∼97 kb on day 0 (Table 3). Remarkably, some animals (2, 16, 19A, 19B, 20, 32, 50, 54, 57, 100, 111, 120, 140) carried more than one CMY-2 or CTX-M-producing E. coli on day 42, which are also present in other animals (Figure 1 and Table 2), which demonstrates the exchanging of commensal GIT bacteria among animals in the feedlot.
The use of a 3GC to enrich medium for recovery of E. coli from the feces of broilers induced a positivity of 99% of the samples containing blaCMY -2- and/or blaCTX-M-isolates (Verrette et al., 2019), which could be the explanation for the high percentage of such E. coli in our study. The blaCMY -2 gene has been reported as frequent in E. coli isolates causing urinary tract infections in Brazil (Rocha D. A. C. et al., 2016), and CMY-2- and CTX-M-producing E. coli were already isolated from poultry and buffalo in the country (Aizawa et al., 2014; Casella et al., 2018; Hoepers et al., 2018) but never in sheep. Apart from the prevalence of isolates presenting the blaCMY -2 gene, the occurrence of blaCTX-M-14- and blaCTX-M-15-carrying E. coli in this study is remarkable. Those genes are the dominant blaCTX-M variants in most regions worldwide, concerning isolates from human infections and food-producing or companion animals (Zhao and Hu, 2013; Bevan et al., 2017; Chong et al., 2018; Dandachi et al., 2018). This means that the studied lambs represent a potential source of hard-to-treat infections caused by E. coli or at least a reservoir of important AR genes that could reach human pathogens. The blaCTX-M-8 gene was the second most detected in the studied population after blaCMY -2, present on both first and last days of feedlot (Table 2). CTX-M-8 was firstly identified in Brazil (Bonnet et al., 2000) and is still frequent in isolates from food-producing animals and meat in the country (Fernandes et al., 2016; Ferreira et al., 2016). However, it is thought to have a relatively low prevalence in other territories and is supposed to be transmitted by travelers or contaminated food (Dhanji et al., 2010; Egervärn et al., 2014; Eller et al., 2014).
Both genes aac(3)-IIa and ant(2″)-Ia codify resistance to gentamicin, and are present in plasmids (Ramirez and Tolmasky, 2010; Norris and Serpersu, 2013; Cox et al., 2015). In this study, aac(3)-IIa clearly predominated in relation to ant(2″)-Ia (Table 2). Notably, both genes reported here are clearly related to E. coli associated with infections (Miró et al., 2013; Fernández-Martínez et al., 2015). Resistance to phenicols was detected on the first and last days of feedlot, with floR and cmlA present in higher frequencies than the cat gene (Figure 1 and Table 2). A Portuguese study found only cmlA in E. coli isolated from sheep (Ramos et al., 2013), and a Brazilian study carried out with Salmonella Typhimurium isolated from humans and food revealed floR associated with food isolates and the cat gene associated with human Salmonella (Almeida et al., 2018). Furthermore, cmlA has already been reported in E. coli from chicken meat in the country (Casella et al., 2017a). Sixty-five isolates (83.3%) presented resistance to trimethoprim/sulfamethoxazole, but more than 60% of the E. coli presented at least one of the sul genes screened while 33.3% presented some dfr gene. Both sul1 and sul2 have been detected in E. coli isolated from sheep in Portugal (Ramos et al., 2013), and those genes have already been reported in E. coli isolated from clinical specimens (Oliveira-Pinto et al., 2017), chicken meat (Casella et al., 2015) and even surface water (Canal et al., 2016) in Brazil, but once again we know nothing about the subject in sheep. Resistance to tetracycline was detected during the entire feedlot stay of the lambs, with tetA and tetB detected on days 0 and 42, with considerable predominance of the first (Figure 1 and Table 2). Interestingly, the isolate 42.120E1 carried tetA and tetB, which is unexpected since both express the same tetracycline efflux mechanism (Thaker et al., 2010). tetA and tetB have already been detected in high frequencies in E. coli isolated from sheep (Ramos et al., 2013). The rising in the content of genes codifying resistance to tetracycline has been observed during bovine feedlot (Weinroth et al., 2018), but to our knowledge, there is no report of such an event concerning resistance to other antimicrobial classes in general, as observed in this study. In fact, the use of tetracyclines and trimethoprim/sulphonamides in sheep has already been reported as presenting a significant association with tetracycline resistance (Scott et al., 2012), and the tetA gene was positively associated with blaCMY -2 after ceftiofur followed chlortetracycline treatment in cattle (Kanwar et al., 2013), which is in agreement with our study. The qnrB gene was detected in six isolates recovered only on day 42, with all but one presenting intermediate resistance to the quinolones (Figure 1 and Table 2). A Chinese study reported qnrB as low-frequency among the genes detected in E. coli recovered from swine (Liu et al., 2018), and a recent study conducted in Brazil showed E. coli isolates carrying qnrB associated with the genes blaCTX-M-2 and blaCMY -2 in poultry (Ferreira et al., 2019). In our study, resistance to quinolones had little importance as a disseminated mechanism through the feedlot. Therefore, the presence of such genes codifying resistance to different antimicrobial classes in commensal isolates of food-producing animals as lambs raises public health concerns. The occurrence of MRAb E. coli in the studied lambs may be caused by the presence of animals and insects carrying these bacteria in the surroundings of the feedlot or even the environment (Blaak et al., 2015; Huijbers et al., 2015; Chong et al., 2018). Since we have collected feces from 112/140 flock animals, another possibility is a lamb not sampled as the source of that E. coli. Indeed, blaCMY -2-floR-tetA-sul2-harboring plasmids have already been identified in food-producing animals (Fernández-Alarcón et al., 2011) and could represent a similarity found in this study.
Regarding virulence genes, most isolates presented only fimH (Table 4), which is related to adhesion and is necessary for GIT colonization (Waksman and Hultgren, 2009). The absence of other virulence genes is not surprising, considering that the E. coli were isolated from feces of healthy animals and represent the GIT microbiota of the lambs. Instead of a known source of STEC strains in Brazil (Vettorato et al., 2009), sheep studied here did not present any evidence of carrying diarrheagenic E. coli (DEC). Nevertheless, all isolates were primarily selected from stools with the 3GC ceftiofur, which could represent a bias in the absence of STEC, EAEC or EPEC strains since such DEC could be present but do not carry genes for 3GC-resistance. The majority of the isolates (61.5%) belong to the phylogenetic group B1, 27.0% were classified as A, and 11.5% belong to phylogroups B2 or D (Figure 1 and Table 4). These results are in agreement with another study (Ramos et al., 2013), in which 61.1% of E. coli isolated from sheep were classified as phylogroup B1, 31.5% were phylogroup A, and 7.4% as phylogroups B2 or D. Traditionally, phylogenetic groups A and B1 are associated with commensal E. coli, while B2 and D with pathogenic isolates (Clermont et al., 2000), which is also in concordance with the few virulence genes detected.
Although the CMY-2-producers were distributed in different clusters according to XbaI-PFGE and belong to two different lineages according to MLST, the IncA/C plasmid of about 170 kb was confirmed as responsible for blaCMY -2 mobilization. This fact also illustrates the dissemination of that plasmid through the feedlot, which was indeed related to blaCMY -2 mobilization in food-producing animals and meat before, suggesting spread of the plasmid worldwide and in Brazil (Guo et al., 2014; Casella et al., 2017b; Dame-Korevaar et al., 2017). blaCTX-M-8 was carried by an IncI1 plasmid of ∼97 kb in isolates recovered on days 0 and 42, which seems to be responsible for the maintenance of that gene in the feedlot during the period analyzed. blaCTX-M-8-IncI1 plasmids have already been reported in E. coli isolated from humans, wastewater, food-producing animals and meat, and appear to be more responsible for the mobilization of that gene in several countries, including Brazil (Ferreira et al., 2014b; Dropa et al., 2016; Norizuki et al., 2017; Casella et al., 2018; Dantas Palmeira et al., 2018). The blaCTX-M-15 gene was carried by very different plasmids on the first and last days of feedlot (Table 3), which means that the blaCTX-M-15-IncFII present on day 0 probably disappeared and the blaCTX-M-15-IncHI2 entered the feedlot at any time point during the period. Since both plasmids are carried by extremely different E. coli, according to XbaI-PFGE and MLST methodologies (Figure 1), it seems that the change on plasmids responsible for blaCTX-M-15 mobilization was due to the disappearance and entry of respective strains into the feedlot, contrary to what happened to the blaCTX-M-8-IncI1 plasmids of about 97 kb mentioned above. IncHI2 plasmids have also been reported as responsible for mobilization of blaCTX-M-15 in several Enterobacteriaceae species isolated from humans or animals (Kariuki et al., 2015; Haenni et al., 2016) and have been detected in 3/4 of the CTX-M-15-producing E. coli in this study. The CTX-M-2-producers identified in this study seem to carry the blaCTX-M-2 inserted into the chromosome. This is not a rare event nowadays and is plausible since it has already been reported in E. coli isolated from chickens and chicken meat in Brazil (Ferreira et al., 2014a; Casella et al., 2018). In addition to that, blaCTX-M-2-carrying E. coli were isolated just on day 42 and were clonally related by XbaI-PFGE and MLST, with the exception of isolate 42.2E2 (Figure 1). Finally, we could not detect the plasmid linked to blaCTX-M-14, and this gene has already been described inserted into the chromosome (Hamamoto et al., 2016; Hamamoto and Hirai, 2019), which could be the explanation for the present isolates. Further studies are required to elucidate this subject.
XbaI-PFGE typing grouped most of the blaCMY -2-carrying E. coli in the two major clusters, composed of 37 and 13 E. coli that carry essentially blaCMY -2-aac(3)-IIa-tetA-sul1-sul2-floR-cmlA, with exceptions, belonging to phylogroups B1-ST/CC 1727/446 or A-ST 3994, respectively (Figure 1). Strains belonging to the later cluster additionally carry the dfr Ia and dfr XII genes. This finding indicates that two strains have spread among animals throughout the feedlot, but all harboring the same blaCMY -2-carrying plasmid as mentioned above. Interestingly, two lineages were detected carrying different bla genes, such as ST/CC 58/155 presenting blaCTX-M-8 or blaCTX-M-14 and ST 3994 presenting blaCTX-M-8 or blaCMY -2, and both groups have a considerable relationship within isolates (Figure 1). E. coli ST/CC 58/155 has already been reported harboring blaCTX-M-14 and others from clinical specimens and healthy people in several countries (Gerhold et al., 2016; Kawamura et al., 2017). In Brazil, this lineage has already been reported carrying blaCTX-M-8 or blaCMY -2 in dogs, and the blaCTX-M-8 gene was also harbored by an IncI1 plasmid (Melo et al., 2018), as in this study. Furthermore, the same Brazilian study showed an E. coli phylogroup D-ST 457 isolated from a diseased dog carrying the blaCTX-M-2 inserted in the chromosome, the same as the only CTX-M-2-producing isolate ST 457 in this study, which demonstrates the presence of that clone in different animals in the country. Contrary to the clonality described above regarding ST/CC 58/155, the E. coli ST/CC 1727/446 isolated in this study carry only blaCMY -2, but isolates were not clonally related according to XbaI-PFGE typing (Figure 1). This could represent micro-evolution occurring in the E. coli strains in the feedlot during the period of 42 days. The new combination of alleles (new ST) found in seven related blaCTX-M-8-carrying isolates was the unique lineage recovered on days 0 and 42, which means that the clone remained in the studied feedlot lambs carrying the same blaCTX-M-8-IncI1 plasmid (Figure 1 and Table 5).
In conclusion, feedlot lambs act as reservoirs of commensal multidrug-resistant E. coli, and those AR genes or bacteria can reach humans through the food chain. The presence of blaCTX-M-14 and blaCTX-M-15 deserves special attention since they are the genes most related to human infections worldwide. To the best of our knowledge, this is the first report of blaCTX-M-14 in Enterobacteriaceae isolated from food-producing animals in Brazil. Additionally, E. coli ST lineages and plasmids harboring the bla genes detected have already been identified in humans, animals, meat and the environment, which demonstrates the concern for their dissemination and for public health. Further studies are needed in order to determine the reasons for the success of the blaCMY -2-aac(3)-IIa-tetA-sul1-sul2-floR-cmlA-carrying E. coli in the studied feedlot. To the best of our knowledge, this is the first study reporting such a broad characterization of antimicrobial resistant E. coli isolated from sheep.
The datasets generated for this study can be found in GenBank, MK896925 to MK896944, and MK917695 to MK917713.
Ethics Committee approval number FOA00845-2017. Universidade Estadual Paulista (UNESP) “Júlio de Mesquita Filho,” campus de Araçatuba, Faculdade de Medicina Veterinária.
TC, MN, and LM designed the study. KG, JF, LDA, and CS performed all the laboratorial experiments. RB handled with the animals and their stools. TC, MN, LM, JP, and MM wrote and revised the whole manuscript.
This work was supported by the São Paulo Research Foundation (FAPESP) (Grant Nos. 2018/02691-4, 2018/16343-8, and 2018/16147-4); the Brazilian Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES); and the Brazilian National Council for Scientific and Technological Development (CNPq) (Grant No. 121464/2018-0).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We are very grateful to Luciana Moran Conceição and Valéria D’Artibale Fraga for the technical support. We also thank Nathalia Paschoalli, a professional translator for English language editing.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2019.01394/full#supplementary-material
Aizawa, J., Neuwirt, N., Barbato, L., Neves, P. R., Leigue, L., Padilha, J., et al. (2014). Identification of fluoroquinolone-resistant extended-spectrum β-lactamase (CTX-M-8)-producing Escherichia coli ST224, ST2179 and ST2308 in buffalo (Bubalus bubalis). J. Antimicrob. Chemother. 69, 2866–2869. doi: 10.1093/jac/dku218
Almeida, F., Seribelli, A. A., Cazentini Medeiros, M. I., dos Rodrigues, D. P., De MelloVarani, A., Luo, Y., et al. (2018). Phylogenetic and antimicrobial resistance gene analysis of Salmonella Typhimurium strains isolated in Brazil by whole genome sequencing. PLoS One 13:e0201882. doi: 10.1371/journal.pone.0201882
Belmar Campos, C., Fenner, I., Wiese, N., Lensing, C., Christner, M., Rohde, H., et al. (2014). Prevalence and genotypes of extended spectrum beta-lactamases in Enterobacteriaceae isolated from human stool and chicken meat in Hamburg, Germany. Int. J. Med. Microbiol. 304, 678–684. doi: 10.1016/j.ijmm.2014.04.012
Benedict, K. M., Gow, S. P., McAllister, T. A., Booker, C. W., Hannon, S. J., Checkley, S. L., et al. (2015). Antimicrobial resistance in Escherichia coli recovered from feedlot cattle and associations with antimicrobial use. PLoS One 10:e0143995. doi: 10.1371/journal.pone.0143995
Bevan, E. R., Jones, A. M., and Hawkey, P. M. (2017). Global epidemiology of CTX-M β-lactamases: temporal and geographical shifts in genotype. J. Antimicrob. Chemother. 72, 2145–2155. doi: 10.1093/jac/dkx146
Blaak, H., Van Hoek, A. H., Hamidjaja, R. A., Van Der Plaats, R. Q., Kerkhof-De Heer, L., De Roda Husman, A. M., et al. (2015). Distribution, numbers, and diversity of ESBL-producing E. coli in the poultry farm environment. PLoS One 10:e0135402. doi: 10.1371/journal.pone.0135402
Bonnet, R., Sampaio, J. L. M., Labia, R., De Champs, C., Sirot, D., Chanal, C., et al. (2000). A novel CTX-M β-lactamase (CTX-M-8) in cefotaxime-resistant Enterobacteriaceae isolated in Brazil. Antimicrob. Agents Chemother. 44, 1936–1942. doi: 10.1128/AAC.44.7.1936-1942.2000
Canal, N., Meneghetti, K. L., De Almeida, C. P., Da Rosa Bastos, M., Otton, L. M., and Corção, G. (2016). Characterization of the variable region in the class 1 integron of antimicrobial-resistant Escherichia coli isolated from surface water. Brazilian J. Microbiol. 47, 337–344. doi: 10.1016/j.bjm.2016.01.015
Carattoli, A., Bertini, A., Villa, L., Falbo, V., Hopkins, K. L., and Threlfall, E. J. (2005). Identification of plasmids by PCR-based replicon typing. J. Microbiol. Methods 63, 219–228. doi: 10.1016/j.mimet.2005.03.018
Casella, T., Cerdeira, L. T., Fernandes, M. R., Souza, T. A., Haenni, M., Madec, J.-Y., et al. (2017a). Draft genome sequence of a CTX-M-15-producing Escherichia coli ST345 from commercial chicken meat in Brazil. J. Glob. Antimicrob. Resist. 9, 124–125. doi: 10.1016/j.jgar.2017.04.002
Casella, T., Nogueira, M. C. L., Saras, E., Haenni, M., and Madec, J. Y. (2017b). High prevalence of ESBLs in retail chicken meat despite reduced use of antimicrobials in chicken production, France. Int. J. Food Microbiol. 257, 271–275. doi: 10.1016/j.ijfoodmicro.2017.07.005
Casella, T., Haenni, M., Madela, N. K., de Andrade, L. K., Pradela, L. K., de Andrade, L. N., et al. (2018). Extended-spectrum cephalosporin-resistant Escherichia coli isolated from chickens and chicken meat in Brazil is associated with rare and complex resistance plasmids and pandemic ST lineages. J. Antimicrob. Chemother. 73, 3293–3297. doi: 10.1093/jac/dky335
Casella, T., Rodríguez, M. M., Takahashi, J. T., Ghiglione, B., Dropa, M., Assunção, E., et al. (2015). Detection of blaCTX-M-type genes in complex class 1 integrons carried by Enterobacteriaceae isolated from retail chicken meat in Brazil. Int. J. Food Microbiol. 197, 88–91. doi: 10.1016/j.ijfoodmicro.2014.12.001
Chong, Y., Shimoda, S., and Shimono, N. (2018). Current epidemiology, genetic evolution and clinical impact of extended-spectrum β-lactamase-producing Escherichia coli and Klebsiella pneumoniae. Infect. Genet. Evol. 61, 185–188. doi: 10.1016/j.meegid.2018.04.005
Clermont, O., Bonacorsi, S., and Bingen, E. (2000). Rapid and Simple Determination of the Escherichia coli Phylogenetic Group. Appl. Environ. Microbiol. 66, 4.
Collignon, P. C., Conly, J. M., Andremont, A., McEwen, S. A., Aidara-Kane, A., Griffin, P. M., et al. (2016). World health organization ranking of antimicrobials according to their importance in human medicine: a critical step for developing risk management strategies to control antimicrobial resistance from food animal production. Clin. Infect. Dis. 63, 1087–1093. doi: 10.1093/cid/ciw475
Cox, G., Stogios, P. J., Savchenko, A., and Wright, G. D. (2015). Structural and molecular basis for resistance to aminoglycoside antibiotics by the Adenylyltransferase ANT(2′′)-Ia. MBio 6, 1–9. doi: 10.1128/mbio.02180-2114
Cyoia, P. S., Koga, V. L., Nishio, E. K., Houle, S., Dozois, C. M., de Brito, K. C. T., et al. (2019). Distribution of ExPEC virulence factors, blaCTX-M, fosA3, and mcr-1 in Escherichia coli isolated from commercialized chicken carcasses. Front. Microbiol. 9:3254. doi: 10.3389/fmicb.2018.03254
Dame-Korevaar, A., Fischer, E. A. J., Stegeman, A., Mevius, D., van Essen-Zandbergen, A., Velkers, F., et al. (2017). Dynamics of CMY-2 producing E. coli in a broiler parent flock. Vet. Microbiol. 203, 211–214. doi: 10.1016/j.vetmic.2017.03.024
Dandachi, I., Chabou, S., Daoud, Z., and Rolain, J. M. (2018). Prevalence and emergence of extended-spectrum cephalosporin-, carbapenem- and colistin-resistant gram negative bacteria of animal origin in the Mediterranean basin. Front. Microbiol. 9:2299. doi: 10.3389/fmicb.2018.02299
Dantas Palmeira, J., Ferreira, H., Madec, J. Y., and Haenni, M. (2018). Pandemic Escherichia coli ST648 isolate harbouring fosA3 and blaCTX-M-8 on an IncI1/ST113 plasmid: a new successful combination for the spread of fosfomycin resistance? J. Glob. Antimicrob. Resist. 15, 254–255. doi: 10.1016/j.jgar.2018.10.025
Dhanji, H., Murphy, N. M., Doumith, M., Durmus, S., Lee, S. S., Hope, R., et al. (2010). Cephalosporin resistance mechanisms in Escherichia coli isolated from raw chicken imported into the UK. J. Antimicrob. Chemother. 65, 2534–2537. doi: 10.1093/jac/dkq376
Doumith, M., Day, M. J., Hope, R., Wain, J., and Woodford, N. (2012). Improved multiplex PCR strategy for rapid assignment of the four major Escherichia coli phylogenetic groups. J. Clin. Microbiol. 50, 3108–3110. doi: 10.1128/JCM.01468-1412
Dropa, M., Lincopan, N., Balsalobre, L. C., Oliveira, D. E., Moura, R. A., Fernandes, M. R., et al. (2016). Genetic background of novel sequence types of CTX-M-8- and CTX-M-15-producing Escherichia coli and Klebsiella pneumoniae from public wastewater treatment plants in São Paulo, Brazil. Environ. Sci. Pollut. Res. Int. 23, 4953–4958. doi: 10.1007/s11356-016-6079-6075
EFSA (2008). Force on Zoonoses Data Collection including guidance for harmonized monitoring and reporting of antimicrobial resistance in commensal Escherichia coli and Enterococcus spp. in food animals. EFSA J. 141, 1–44. doi: 10.2903/j.efsa.2008.141r
EFSA (2011). Scientific Opinion on the public health risks of bacterial strains producing extended-spectrum β-lactamases and/or AmpC β-lactamases in food and food-producing animals. EFSA J. 9, 2322. doi: 10.2903/j.efsa.2011.2322
Egervärn, M., Borjesson, S., Byfors, S., Finn, M., Kaipe, C., Englund, S., et al. (2014). Escherichia coli with extended-spectrum beta-lactamases or transferable AmpC beta-lactamases and Salmonella on meat imported into Sweden. Int. J. Food Microbiol. 171, 8–14. doi: 10.1016/j.ijfoodmicro.2013.11.005
Eller, C., Leistner, R., Guerra, B., Fischer, J., Wendt, C., Rabsch, W., et al. (2014). Emergence of extended-spectrum β-lactamase (ESBL) CTX-M-8 in Germany. J. Antimicrob. Chemother. 69, 562–564. doi: 10.1093/jac/dkt387
FAO (2018). FAOSTAT. Food Agric. Organ. United Nations - Stat. Div. Available at: http://www.fao.org/faostat/en/#data/QL (accessed January 18, 2019).
Fernandes, S. A., Camargo, C. H., Francisco, G. R., Bueno, M. F. C., Garcia, D. O., Doi, Y., et al. (2016). Prevalence of Extended-Spectrum β-Lactamases CTX-M-8 and CTX-M-2-Producing Salmonella Serotypes from Clinical and Nonhuman Isolates in Brazil. Microb. Drug Resist. 23, 580–589. doi: 10.1089/mdr.2016.0085
Fernández-Alarcón, C., Singer, R. S., and Johnson, T. J. (2011). Comparative genomics of multidrug resistance-encoding incA/C plasmids from commensal and pathogenic Escherichia coli from multiple animal sources. PLoS One 6:e23415. doi: 10.1371/journal.pone.0023415
Fernández-Martínez, M., Miró, E., Ortega, A., Bou, G., González-López, J. J., Oliver, A., et al. (2015). Molecular identification of aminoglycoside-modifying enzymes in clinical isolates of Escherichia coli resistant to amoxicillin/clavulanic acid isolated in Spain. Int. J. Antimicrob. Agents 46, 157–163. doi: 10.1016/j.ijantimicag.2015.03.008
Ferreira, J. C., Penha Filho, R. A. C., Andrade, L. N., Berchieri, A. Jr., and Darini, A. L. (2016). Evaluation and characterization of plasmids carrying CTX-M genes in a non-clonal population of multidrug-resistant Enterobacteriaceae isolated from poultry in Brazil. Diagn. Microbiol. Infect. Dis. 85, 444–448. doi: 10.1016/j.diagmicrobio.2016.05.011
Ferreira, J. C., Penha Filho, R. A. C., Andrade, L. N., Berchieri, A., and Darini, A. L. C. (2014a). Detection of chromosomal blaCTX-M-2 in diverse Escherichia coli isolates from healthy broiler chickens. Clin. Microbiol. Infect. 20, O623–O626. doi: 10.1111/1469-0691.12531
Ferreira, J. C., Penha Filho, R. A. C., Andrade, L. N., Berchieri, A. Jr., and Darini, A. L. (2014b). IncI1/ST113 and IncI1/ST114 conjugative plasmids carrying blaCTX-M-8 in Escherichia coli isolated from poultry in Brazil. Diagn. Microbiol. Infect. Dis. 80, 304–306. doi: 10.1016/j.diagmicrobio.2014.09.012
Ferreira, J. C., Penha Filho, R. A. C., Andrade, L. N., and Darini, A. L. C. (2019). Evaluation of heavy metal tolerance genes in plasmids harbored in multidrug-resistant and isolated from poultry in Brazil. Diagn. Microbiol. Infect. Dis doi: 10.1016/j.diagmicrobio.2019.01.019 [Epub ahead of print].
Gerhold, G., Schulze, M. H., Gross, U., and Bohne, W. (2016). Multilocus sequence typing and CTX-M characterization of ESBL-producing E. coli?: a prospective single-centre study in Lower Saxony, Germany. Epidemiol. Infect. 144, 3300–3304. doi: 10.1017/s0950268816001412
Gibbons, J. F., Boland, F., Egan, J., Fanning, S., Markey, B. K., and Leonard, F. C. (2016). Antimicrobial resistance of faecal Escherichia coli isolates from pig farms with different durations of in-feed antimicrobial use. Zoonoses Public Health 63, 241–250. doi: 10.1111/zph.12225
Graham, J. P., Eisenberg, J. N. S., Trueba, G., Zhang, L., and Johnson, T. J. (2017). Small-scale food animal production and antimicrobial resistance: mountain, molehill, or something in-between? Environ. Health Perspect. 125, 1–5. doi: 10.1289/EHP2116
Guo, Y. F., Zhang, W. H., Ren, S. Q., Yang, L., Lü, D. H., Zeng, Z. L., et al. (2014). IncA/C plasmid-mediated spread of CMY-2 in multidrug-resistant Escherichia coli from food animals in China. PLoS One 9:e96738. doi: 10.1371/journal.pone.0096738
Haenni, M., Saras, E., Ponsin, C., Dahmen, S., Petitjean, M., Hocquet, D., et al. (2016). High prevalence of international ESBL CTX-M-15-producing Enterobacter cloacae ST114 clone in animals. J. Antimicrob. Chemother 71, 1497–1500. doi: 10.1093/jac/dkw006
Hamamoto, K., and Hirai, I. (2019). Characterisation of chromosomally-located bla CTX-M and its surrounding sequence in CTX-M-type extended-spectrum β-lactamase-producing Escherichia coli isolates. J. Glob. Antimicrob. Resist. 17, 53–57. doi: 10.1016/j.jgar.2018.11.006
Hamamoto, K., Ueda, S., Toyosato, T., Yamamoto, Y., and Hirai, I. (2016). High Prevalence of Chromosomal bla CTX-M-14 in Escherichia coli Isolates Possessing bla CTX-M-14. Antimicrob. Agents Chemother. 60, 2582–2584. doi: 10.1128/AAC.00108-116
Hoepers, P. G., Silva, P. L., Rossi, D. A., Valadares Júnior, E. C., Ferreira, B. C., Zuffo, J. P., et al. (2018). The association between extended spectrum beta-lactamase (ESBL) and ampicillin C (AmpC) beta-lactamase genes with multidrug resistance in Escherichia coli isolates recovered from turkeys in Brazil. Br. Poult. Sci. 59, 396–401. doi: 10.1080/00071668.2018.1468070
Huijbers, P. M. C., Blaak, H., De Jong, M. C. M., Graat, E. A. M., Vandenbroucke-Grauls, C. M. J. E., and De Roda Husman, A. M. (2015). Role of the environment in the transmission of antimicrobial resistance to humans: a review. Environ. Sci. Technol. 49, 11993–12004. doi: 10.1021/acs.est.5b02566
Kaesbohrer, A., Schroeter, A., Tenhagen, B. A., Alt, K., Guerra, B., and Appel, B. (2012). Emerging antimicrobial resistance in commensal escherichia coli with public health relevance. Zoonoses Public Health 59, 158–165. doi: 10.1111/j.1863-2378.2011.01451.x
Kanwar, N., Scott, H. M., Norby, B., Loneragan, G. H., Vinasco, J., McGowan, M., et al. (2013). Effects of ceftiofur and chlortetracycline treatment strategies on antimicrobial susceptibility and on tet(A), tet(B), and blaCMY-2 resistance genes among E. coli isolated from the feces of feedlot cattle. PLoS One 8:e80575. doi: 10.1371/journal.pone.0080575
Kariuki, S., Okoro, C., Kiiru, J., Njoroge, S., Omuse, G., Langridge, G., et al. (2015). Ceftriaxone-resistant Salmonella enterica serotype typhimurium sequence type 313 from Kenyan patients is associated with the blaCTX-M-15 gene on a novel IncHI2 plasmid. Antimicrob. Agents Chemother. 59, 3133–3139. doi: 10.1128/AAC.00078-15
Kaur, J., Chopra, S., Sheevani, and Mahajan, G. (2013). Modified double disc synergy test to detect ESBL production in urinary isolates of Escherichia coli and Klebsiella pneumoniae. J. Clin. Diagnostic Res. 7, 229–233. doi: 10.7860/JCDR/2013/4619.2734
Kawamura, K., Nagano, N., Suzuki, M., Wachino, J., Kimura, K., and Arakawa, Y. (2017). ESBL-producing Escherichia coli and its rapid rise among healthy people. Food Saf. 5, 122–150. doi: 10.14252/foodsafetyfscj.2017011
Knudsen, P. K., Gammelsrud, K. W., Alfsnes, K., Steinbakk, M., Abrahamsen, T. G., Müller, F., et al. (2018). Transfer of a bla CTX-M-1-carrying plasmid between different Escherichia coli strains within the human gut explored by whole genome sequencing analyses. Sci. Rep. 8, 1–10. doi: 10.1038/s41598-017-18659-18652
Liu, X., Liu, H., Wang, L., Peng, Q., Li, Y., Zhou, H., et al. (2018). Molecular characterization of extended-spectrum β-lactamase-producing multidrug resistant Escherichia coli from swine in Northwest China. Front. Microbiol 9:1756. doi: 10.3389/fmicb.2018.01756
Madec, J.-Y., and Haenni, M. (2018). Antimicrobial resistance plasmid reservoir in food and food-producing animals. Plasmid 99, 72–81. doi: 10.1016/j.plasmid.2018.09.001
Makita, K., Goto, M., Ozawa, M., Kawanishi, M., Koike, R., Asai, T., et al. (2016). Multivariable analysis of the association between antimicrobial use and antimicrobial resistance in Escherichia coli Isolated from apparently healthy pigs in Japan. Microb. Drug Resist. 22, 28–39. doi: 10.1089/mdr.2014.0311
Melo, L. C., Oresco, C., Leigue, L., Netto, H. M., Melville, P. A., Benites, N. R., et al. (2018). Prevalence and molecular features of ESBL/pAmpC-producing Enterobacteriaceae in healthy and diseased companion animals in Brazil. Vet. Microbiol. 221, 59–66. doi: 10.1016/j.vetmic.2018.05.017
Miró, E., Grünbaum, F., Gómez, L., Rivera, A., Mirelis, B., Coll, P., et al. (2013). Characterization of aminoglycoside-modifying enzymes in Enterobacteriaceae clinical strains and characterization of the plasmids implicated in their diffusion. Microb. Drug Resist. 19, 94–99. doi: 10.1089/mdr.2012.0125
Norizuki, C., Wachino, J., Suzuki, M., Kawamura, K., Nagano, N., Kimura, K., et al. (2017). Specific bla CTX-M-8 /IncI1 plasmid transfer among genetically diverse Escherichia coli Isolates between humans and chickens. Antimicrob. Agents Chemother. 61, e663–e617. doi: 10.1128/AAC.00663-617
Norris, A. L., and Serpersu, E. H. (2013). Ligand promiscuity through the eyes of the aminoglycoside N3 acetyltransferase IIa. Protein Sci. 22, 916–928. doi: 10.1002/pro.2273
Oliveira-Pinto, C., Diamantino, C., Oliveira, P. L., Reis, M. P., Costa, P. S., Paiva, M. C., et al. (2017). Occurrence and characterization of class 1 integrons in Escherichia coli from healthy individuals and those with urinary infection. J. Med. Microbiol. 66, 577–583. doi: 10.1099/jmm.0.000468
Projahn, M., von Tippelskirch, P., Semmler, T., Guenther, S., Alter, T., and Roesler, U. (2019). Contamination of chicken meat with extended-spectrum beta-lactamase producing- Klebsiella pneumoniae and Escherichia coli during scalding and defeathering of broiler carcasses. Food Microbiol. 77, 185–191. doi: 10.1016/j.fm.2018.09.010
Ramirez, M. S., and Tolmasky, M. E. (2010). Aminoglycoside modifying enzymes. Drug Resist. Updat. 13, 151–171. doi: 10.1016/j.drup.2010.08.003
Ramos, S., Silva, N., Caniça, M., Capelo-Martinez, J. L., Brito, F., Igrejas, G., et al. (2013). High prevalence of antimicrobial-resistant Escherichia coli from animals at slaughter: a food safety risk. J. Sci. Food Agric. 93, 517–526. doi: 10.1002/jsfa.5814
Rocha, D. A. C., Campos, J. C., Passadore, L. F., Sampaio, S. C. F., Nicodemo, A. C., and Sampaio, J. L. M. (2016). Frequency of Plasmid-Mediated AmpC β-Lactamases in Escherichia coli isolates from urine samples in São Paulo, Brazil. Microb. Drug Resist. 22, 321–327. doi: 10.1089/mdr.2015.0190
Scott, L., Menzies, P., Reid-Smith, R. J., Avery, B. P., McEwen, S. A., Moon, C. S., et al. (2012). Antimicrobial resistance in fecal generic Escherichia coli and Salmonella spp. obtained from Ontario sheep flocks and associations between antimicrobial use and resistance. Can. J. Vet. Res. 76, 109–119.
Solà-Ginés, M., González-López, J. J., Cameron-Veas, K., Piedra-Carrasco, N., Cerdà-Cuéllar, M., and Migura-Garcia, L. (2015). Houseflies (Musca domestica) as vectors for extended-spectrum β-lactamase-producing Escherichia coli on Spanish broiler farms. Appl. Environ. Microbiol. 81, 3604–3611. doi: 10.1128/aem.04252-4214
Thaker, M., Spanogiannopoulos, P., and Wright, G. D. (2010). The tetracycline resistome. Cell. Mol. Life Sci. 67, 419–431. doi: 10.1007/s00018-009-0172-176
Verrette, L., Fairbrother, J. M., and Boulianne, M. (2019). Effect of ceftiofur cessation and substitution with lincomycin-spectinomycin on extended-spectrum-beta-lactamase/AmpC genes and multidrug resistance in Escherichia coli from a Canadian broiler production pyramid. Appl. Environ. Microbiol. [Epub ahead of print].
Vettorato, M. P., De Castro, A. F. P., Cergole-Novella, M. C., Camargo, F. L. L., Irino, K., and Guth, B. E. C. (2009). Shiga toxin-producing Escherichia coli and atypical enteropathogenic Escherichia coli strains isolated from healthy sheep of different populations in São Paulo, Brazil. Lett. Appl. Microbiol. 49, 53–59. doi: 10.1111/j.1472-765X.2009.02617.x
Villa, L., García-Fernandez, A., Fortini, D., and Carattoli, A. (2010). Replicon sequence typing of IncF plasmids carrying virulence and resistance determinants. J. Antimicrob. Chemother. 65, 2518–2529. doi: 10.1093/jac/dkq347
Waksman, G., and Hultgren, S. J. (2009). Structural biology of the chaperone-usher pathway of pilus biogenesis. Nat. Rev. Microbiol. 7, 765–774. doi: 10.1038/nrmicro2220
Weinroth, M. D., Scott, H. M., Norby, B., Loneragan, G. H., Noyes, N. R., Rovira, P., et al. (2018). Effects of ceftiofur and chlortetracycline on the resistomes of feedlot cattle. Appl. Environ. Microbiol. 84, 1–12. doi: 10.1128/AEM.00610-618
WHO (2017). Integrated Surveillance of Antimicrobial Resistance in Foodborne Bacteria: Application of a One Health Approach. Geneva: World Health Organization.
Keywords: Escherichia coli, sheep, multidrug resistance, cephalosporin, aminoglycoside, tetracycline, trimethoprim/sulfamethoxazole, phenicols
Citation: Gozi KS, Froes JR, Deus Ajude LPT, da Silva CR, Baptista RS, Peiró JR, Marinho M, Mendes LCN, Nogueira MCL and Casella T (2019) Dissemination of Multidrug-Resistant Commensal Escherichia coli in Feedlot Lambs in Southeastern Brazil. Front. Microbiol. 10:1394. doi: 10.3389/fmicb.2019.01394
Received: 12 February 2019; Accepted: 04 June 2019;
Published: 25 June 2019.
Edited by:
Ziad Daoud, University of Balamand, LebanonReviewed by:
Mueen Aslam, Agriculture and Agri-Food Canada, CanadaCopyright © 2019 Gozi, Froes, Deus Ajude, da Silva, Baptista, Peiró, Marinho, Mendes, Nogueira and Casella. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tiago Casella, dGlhZ29fY2FzZWxsYUB5YWhvby5jb20uYnI=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.