 Helge-André Erikstad
Helge-André Erikstad Ruben Michael Ceballos
Ruben Michael Ceballos Natalie Bennett Smestad
Natalie Bennett Smestad Nils-Kåre Birkeland
Nils-Kåre Birkeland- 1Department of Biological Sciences, University of Bergen, Bergen, Norway
- 2Department of Biological Sciences, University of Arkansas, Fayetteville, AR, United States
Thermoacidophilic methane-oxidizing Verrucomicrobia of the candidate genus Methylacidiphilum represent a bacterial taxon adapted to highly acidic (pH 1–4) and moderate temperature (∼65∘C) methane-containing geothermal environments. Their apparent ubiquity in acidic terrestrial volcanic areas makes them ideal model organisms to study prokaryotic biogeography. Three Methylacidiphilum species isolated from distantly-separated geothermal regions in Russia, New Zealand, and Italy were previously described. We have explored the intra-taxon phylogenetic patterns of these organisms based on comparative genome analyses and phenotypic comparisons with six new Verrucomicrobia methanotroph isolates from other globally-separated acidic geothermal locations. Comparison of rRNA and particulate methane monooxygenase (pmoCAB) operon sequences indicates a close phylogenetic relationship among the new isolates as well as with the previously characterized strains. All share similar cell morphology including the presence of extensive intracellular inclusion bodies and lack of intracellular membrane systems, which are typical for proteobacterial methanotrophs. However, genome sequence comparisons and concatenated MLST-based phylogenetic analyses separate the new isolates into three distinct species-level groups. Three recently processed isolates from the Azores (each from geographically-separate hot springs within the region) and a single isolate from Iceland are highly similar, sharing more than 88% in silico genome homology with each other as well as with the previous isolate, Methylacidiphilum fumariolicum strain SolV, from Italy. These appear to constitute a distinct European/Atlantic clade. However, two of the new isolates – one from the Yellowstone National Park (United States) and another from The Philippines – constitute separate and novel Methylacidiphilum species. There is no clear correlation between fatty acid profiles and geographic distance between origins, or any phylogenetic relationship. Serological analysis using antiserum raised against M. kamchatkense strain Kam1 revealed large differences in the degree of cross-reactivity with no correlation with other factors. However, the genetic distance between the strains does correlate to the distance between their geographic origins and suggests a global biogeographic pattern shaped by an isolation-by-distance mechanism. These results further confirm terrestrial geothermal springs as isolated islands featuring allopatric prokaryotic speciation.
Introduction
Microbial biogeography and mechanisms for speciation among microbes are still controversial and poorly understood topics. In general, prokaryotes are considered to be cosmopolitan. Specifically, it is thought that they exhibit global distribution without clear biogeographic boundaries. The long-standing Baas-Becking postulate, “Everything is everywhere, but the environment selects” (Baas-Becking, 1934), has been a cornerstone hypothesis for the evolution and diversification of prokaryotes for decades. A number of studies provide support for the cosmopolitan nature of prokaryotes. Identical microorganisms have been recovered from environments across the globe. Examples from marine (Massana et al., 2000), freshwater (Glöckner et al., 2000), and soil (Brewer et al., 2017) systems have been shown to be globally distributed and lack of any notable biogeographic structure. Furthermore, studies of bacterial pathogens and endospore formers also appear to demonstrate cosmopolitan distribution (Musser et al., 1990; Smith et al., 1991; Roberts and Cohan, 1995). Even prokaryotes recovered from deep subsurface environments, in some cases, have been shown to be virtually identical with isolates derived from widely-separated geographic regions. For example, an isolate of a thermophilic sulfate reducer belonging to the species Thermodesulfobacterium thermophilum sampled from hot water produced from deep oil wells in the North Sea is serologically indistinguishable from those found near the Caspian Sea (Christensen et al., 1992). A strain of the archaeal sulfate reducer, Archaeoglobus fulgidus, from a shallow hot vent in the Mediterranean Sea (Stetter, 1988), is almost indistinguishable from an isolate derived from an oil well in the North Sea (Beeder et al., 1994; Birkeland et al., 2017). Likewise, Thermosipho africanus strain TCF52B, isolated from a North Sea oil well (Dahle et al., 2008), is virtually identical with T. africanus strain Ob7T, isolated from a shallow hot vent in Djibouti, Africa as determined by genomic DNA-DNA hybridization (DDH) and 16S rRNA gene sequence analyses (Huber et al., 1989; Dahle et al., 2008). All three of these examples support the concept of a cosmopolitan distribution of prokaryotes and the idea that geographic distance plays a minor role in speciation and evolution in prokaryotic systems.
However, during the last 15 years, conspicuous patterns of biogeographic structure in global distribution of microbes have been reported. This has been shown for members of the thermophilic archaeal genus, Sulfolobus, both by comparison of selected chromosomal loci (Whitaker et al., 2003) and complete pan-genomes (Reno et al., 2009). These organisms are adapted to acidic terrestrial hot springs, and do not possess any spores or spore-like structures that can facilitate dispersion across large geographical distances. Isolates from similar hot springs as far apart as Kamchatka (Eastern Russia), Iceland, and the United States formed distinct phylogenetic clades, indicating evolution through local adaptation or random genetic drift with no evidence of gene flow between the isolated populations. Interestingly, the question of whether biogeographic structure exists in a common virus of the genus – namely, the Sulfolobus Spindle-shaped Virus – and the functional implications (e.g., infectivity) has been controversial (Snyder et al., 2007; Held and Whitaker, 2009; Ceballos et al., 2012). Terrestrial geothermal features represent environments with highly specific physicochemical conditions, often with extreme temperature and pH values. Dispersion of microorganisms between distantly located hot springs therefore seems highly unlikely, and such environments are thus regarded as isolated islands with strong geographic barriers. Select extremophiles from hypersaline lakes and mines (e.g., Nocardiopsis spp.) have also revealed a strong influence of spatial distance on the shaping of location-specific phylotypes of bacterial populations, especially across large spatial scales (He et al., 2015).
Three thermoacidophilic methanotrophs belonging to the phylum Verrucomicrobia, which have been given the provisional genus name Methylacidiphilum, have been isolated from acidic terrestrial geothermal environments in New Zealand, Italy, and Kamchatka, Russia (Dunfield et al., 2007; Pol et al., 2007; Islam et al., 2008). These organisms are extremely acidophilic (and moderately thermophilic) with optimal pH and temperatures for growth of 2.0–3.5 and 55–60°C, respectively (Op den Camp et al., 2009). Together with a mesophilic sister group, which is given the provisional genus name Methylacidimicrobium, these thermophilic methanotrophic Verrucomicrobia constitute a novel subdivision within the phylum Verrucomicrobia (Van Teeseling et al., 2014). In this study, we introduce six additional thermoacidophilic methanotrophic Verrucomicrobia isolates from acidic hot springs in: Iceland, Yellowstone National Park (United States), the Azores, and the Philippines. Together with previously described strains, a collection of nine isolates from closely located sampling sites from a few kilometers to 3000 km apart as well as from more distantly located sites (i.e., > 18,000 km apart) are available. Using genomic comparisons, our data demonstrate a clear correlation between the distance of the origin and relationship between strains, supporting a distance dependent speciation.
Materials and Methods
Sampling, Isolation, and Cultivation
Sampling was done by collecting a slurry of water and top layer of sediment (to < 3 cm depth) from shallow (5–20 cm deep) sites of acidic hot springs in The Azores (Furnas, Ribeira Grande and Furnas das Lagos), Iceland (Krysuvik), The Philippines (Makiling mud spring) and Yellowstone National Park (the Norris geyser basin), United States. Characteristics and geographic coordinates of the springs are provided in Table 1, including time of sampling and strain designations. The chemical composition of spring waters is given in Supplementary Table S1. The distance between sampling sites were calculated using Google Maps1. This is based on shortest distance between points defined by latitude:longitude coordinates. Inoculation and enrichment was done using a 10-fold dilution of a standard methanotrophic ammonium-based mineral medium (Whittenbury et al., 1970) adjusted to pH 3.5 and prepared as previously described (Islam et al., 2008; Erikstad et al., 2012). Enrichment cultures were prepared by inoculation of 1 ml sample slurry to 10 ml medium in 120 ml serum flasks which subsequently were closed with a butyl rubber cap and an aluminum crimp seal. Methane was added by a syringe to 20% headspace concentration. Flasks were incubated at 55°C with shaking at 150 rpm. Growth was monitored by phase contrast microscopy, using a Nikon, Eclipse E400 microscope. CH4 consumption was analyzed in a Hewlet Packard 6890 gas chromatograph equipped with a 1.83 m Haysep R column (80/100 mesh) and a thermal conductivity detector (150°C). After verification of growth and methane consumption, isolation was done by two times dilution to extinction. All isolates were identified as methanotrophic Verrucomicrobia by PCR and sequencing of their 16S rRNA genes as earlier described (Islam et al., 2008). All subsequent cultivations were performed in the medium and under conditions as described above. Since 2015 the medium has been supplemented with 300 nM CeCl3 as it was shown that lanthanides strongly enhance growth of these microorganisms (Pol et al., 2014).
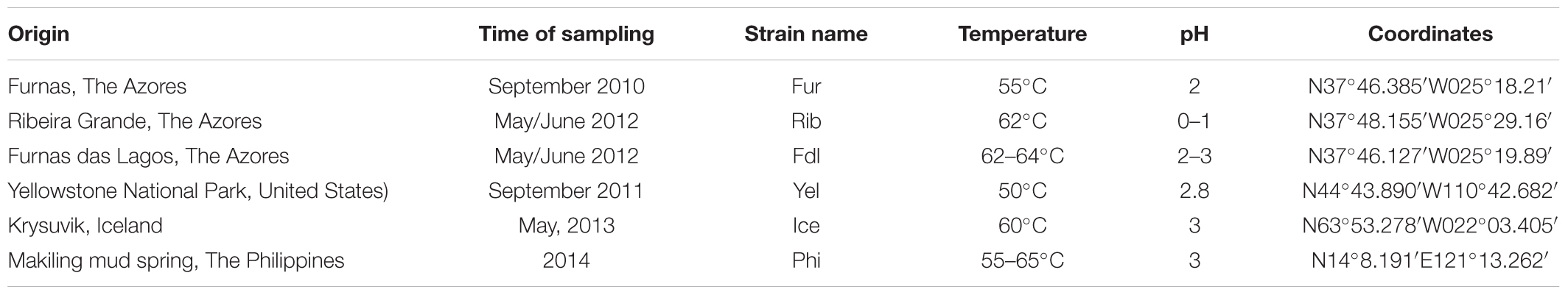
Table 1. Overview of the characteristics and location of the sampling sites from which the novel methanotrophic isolates were recovered.
Electron Microscopy
Transmission electron microscopy was performed as described previously (Islam et al., 2008). Briefly, harvested cells were fixed in 2.5% glutaraldehyde at 4°C overnight followed by 3 washes in 0.25% NaCl for 15 min each before post-fixation with 1% OsO4 for 60 min. Then, cells were washed and dehydrated in increasing concentrations of ethanol before being embedded in Agar 100 resin. Thin sections were prepared using a Reichert Ultracut ultramicrotome and stained with lead and uranyl before being examined with a Jeol JEM-1230 electron microscope.
For scanning electron microscopy, 2 ml samples of six to 7 days old cultures were removed from serum bottles with a 10 ml BD syringe with a Luer-LokTM tip. Polycarbonate membrane filters (Millipore IsoporeTM, 0.2 μm pore size) were inserted into 13 mm Millipore Swinnex filter holders by the use of forceps, and then mounted onto the syringes containing the cultures. The cultures were filtrated through the filters, which were subsequently washed with cacodylate buffer (pH 7.4) before being fixed in 6% glutaraldehyde in cacodylate buffer for 2 h. The filters were then washed with 0.5 ml cacodylate buffer 10 × 5 min and distilled H2O for 5 × 5 min. Dehydration of the cells were then performed by step-wise treatment with 50, 75, 96% and finally 2 × 100% ethanol. Filters were incubated for 15 min in each step and then transferred to a Quorum Technologies Critical point dryer CPD7501. The filters were first flushed with 100% liquid CO2 three times using a pressure between 700 and 900 psi at -20°C to 4°C. A critical point was then reached by increasing the pressure to 1300–1500 psi and the temperature to 35–37°C. The filters were then mounted onto aluminum stubs with double sided carbon tape and coated with a 10 nm thick layer of gold palladium in a sputter coater (Jeol JFC-2300HR High resolution fine coater). The samples were then examined in a Jeol JSM-7400F scanning electron microscope.
Genome Sequence Analysis and Phylogeny
Genomic DNA was extracted with the use of a GenEluteTM Bacterial Genomic DNA kit following the manufacturer’s instructions and sequenced at GATC Biotech, Germany2 using Illumina paired-end technology. A total of 11,700 to more than 28,000 reads accounting for 1.5–3.6 Giga bases were obtained. Assembly by CLC Genomics Workbench 8.5.1 resulted in 2.3–2.4 Mega bases of unique sequence data distributed into 58–113 contigs. Annotation was performed using RAST3 and the NCBI Prokaryotic Annotation Pipeline4. The whole-genome shotgun projects have been deposited at DDBJ/EMBL/GenBank under the accession numbers LXJS00000000 (Strain Fur), LXNL00000000 (Strain Fdl), LXNK00000000, LXOX00000000 (Strain Ice), LXQC00000000 (Strain Phi) and LXQB00000000 (Strain Yel). Pairwise genome sequence similarities were calculated using the in-silico Genome-To-Genome Distance Calculator available at DSMZ5. Contigs obtained from the de novo assembly were ordered and aligned using the M. fumariolicum SolV genome (Anvar et al., 2014) as template by the use of MAUVE version 2.4.06 (Darling et al., 2004).
The evolutionary history of Methylacidiphilum strains was inferred using sequence concatemers of the genes gyrA, gyrB, ftsZ, rho, dnaG, rpoE, and groEL, by using the Maximum Likelihood method based on the Tamura-Nei model (Tamura and Nei, 1993). Reference genes were extracted from the complete genome sequences of M. fumariolicum SolV (Anvar et al., 2014), Methylacidiphilum infernorum strain V4 (Hou et al., 2008) and the draft sequence of Methylacidiphilum kamchatkense strain Kam1 (Erikstad and Birkeland, 2015). The mesophilic methanotrophic verrucomicrobial isolates, Methylacidimicrobium fagopyrum 3C (Van Teeseling et al., 2014; BioProject PRJNA165235) and Methylacidimicrobium sp. LP2A (Op den Camp et al., 2009; Acc. no. JAFS01000000), were also included. Initial tree(s) for the heuristic search were obtained automatically by applying Neighbor-Join and BioNJ algorithms to a matrix of pairwise distances estimated using the Maximum Composite Likelihood approach, and then selecting the topology with superior log likelihood value. All positions containing gaps and missing data were eliminated. The analysis was conducted in MEGA7 (Kumar et al., 2016).
From DNA sequence alignments of select genes (e.g., pmoAB2), identity matrices were constructed to illustrate pairwise comparisons of sequence similarities between different strains. In addition, a geographic distance matrix was constructed, which denotes physical separation (km) between sites from which each isolate was derived. Using a “genetic distance” matrix with a geographic distance matrix, a Mantel test was employed to compute correlation between the two distance matrices. The resulting test statistic, rM, is defined as:
where and n is the number of rows/columns (i.e., 9) in each of the distance matrices (Mantel, 1967). Possible values for rM from -1 to +1 indicate the presence of either a negative or positive correlation (or absence of correlation if rM = 0) for genetic distance versus geographic distance. To test for potential correlation between genetic divergence and tectonic plate sympatry, the Spearman correlation method was employed (Dodge, 2008). The Spearman rank correlation (rs) is defined by:
where cov(rggenetic, rgtectonic) is the covariance of the rank variables and σrggenetic and σrgtectonic are the standard deviations of the rank variables.
Fatty Acid Analysis
Analysis of the cellular fatty acid composition was determined by Leibniz Institute DSMZ-German collection of Microorganisms and Cell Cultures7. Fatty acid methyl esters obtained from freeze-dried cell pellets by saponification, methylation and extraction using minor modifications of a method previously described (Miller, 1982; Kuykendall et al., 1988). The fatty acid methyl ester mixtures were separated using Sherlock Microbial Identification System (MIS) (MIDI, Microbial ID, Newark, DE 19711, United States) which consisted of an Agilent model 6890N gas chromatograph fitted with a 5% phenyl-methyl silicone capillary column (0.2 mm × 25 m), a flame ionization detector, Agilent model 7683A automatic sampler, and a HP-computer with MIDI database (Hewlett-Packard Co., Palo Alto, CA, United States). Peaks are automatically integrated and fatty acid names and percentages calculated by the MIS Standard Software (Microbial ID). The gas chromatographic parameters were as follows: carrier gas, ultra-high-purity hydrogen; column head pressure 60 kPa; injection volume 2 μl; column split ratio, 100:1; septum purge 5 ml/min; column temperature, 170–270°C at 5°C/min; injection port temperature, 240°C; and detector temperature, 300°C.
Serology
Cells from an actively growing culture of Kam1 were harvested by centrifugation and washed twice in phosphate buffered saline (PBS), pH 7, containing 3.7% formaldehyde, and washed twice in PBS. The final resuspension contained 5 × 109 cells per ml and was shipped on ice to Biogenes (Germany)8 for production of polyclonal antibodies in rabbits through intramuscular injections. The rabbits were first injected with 1 ml antigen followed by weekly injections with 0.5 ml antigen for 3 weeks. Two months later, a boost of 1 ml was injected followed by final bleeding 1 week later. Pre-immunserum was collected prior to starting the immunization programme.
ELISA was performed on freeze-dried pellets of our bacterial isolates. The 96- well immunoplate (Nunc MaxiSorpTM) was coated with 150 μl of the antigen solution overnight at 4°C (10 μg/ml in PBS- buffer). The antigens were not applied to the outermost wells. Skimmed milk powder (5 μg/ml) was used as a negative control. All the following washing steps was done with x3 PBS-Tween buffer (PBS-T), pH 7.4 (200 μL to each well). After coating, the wells were washed and subsequently blocked with a 3% skimmed milk powder blocking solution in PBS-T for 1 h at room temperature (200 μL to each well) and followed by additional washes. The primary antiserum was initially diluted 1:400 in PBS-T followed by consecutive to-fold dilutions per well (200 μL to each well in total). The negative control serum was initially diluted 1:100 in PBS-T otherwise the same dilution pattern as for the primary antiserum. After incubating with primary antiserum at room temperature for 2 h, the plate was washed and added 50 μl pr well of 1:3000 diluted “goat anti-rabbit” Ig conjugate (diluted in PBS-T), followed by another incubation at room temperature for 1 h. Fifty microliter of peroxidase substrate solution, o-Phenyleneidamine (OPD), was added to each well. After 6 min, 50 μl of 2.5N H2SO4 was added each well to stop the substrate reaction. Titertek Multiscan spectrophotometric microplate reader was used to read the results at OD492 nm.
Results
Isolation and Initial Characterization
The six methanotrophic isolates presented here, belonging to the phylum Verrucomicrobia, were isolated from geothermal environments across the globe (Figure 1 and Table 1). In addition, M. kamchatkense strain Kam1, which was previously isolated by our lab (Islam et al., 2008), was included in this study. Three of the new isolates originate from three different locations on the San Miguel Island in The Azores. The isolates are designated according to their respective sampling sites: Furnas (Fur), Ribeira Grande (Rib), and Furnas Das Lagos (Fdl). Temperatures at sample sites located in the Azores ranged from 55 to 64°C and pH from 0 to 3. The isolate from Yellowstone National Park (Yel) was collected in the Norris geyser basin from a site at 50°C and pH 2.8. We collected the Iceland isolate (Ice), from a spring at 60°C and pH 3, which was located in the Seltun area in the Krysuvik geothermal field. The isolate from the Philippines was recovered from the Mount Makiling mud spring in the Laguna province on the island of Luzon. This sampling site was between 55 and 65°C and pH 3.
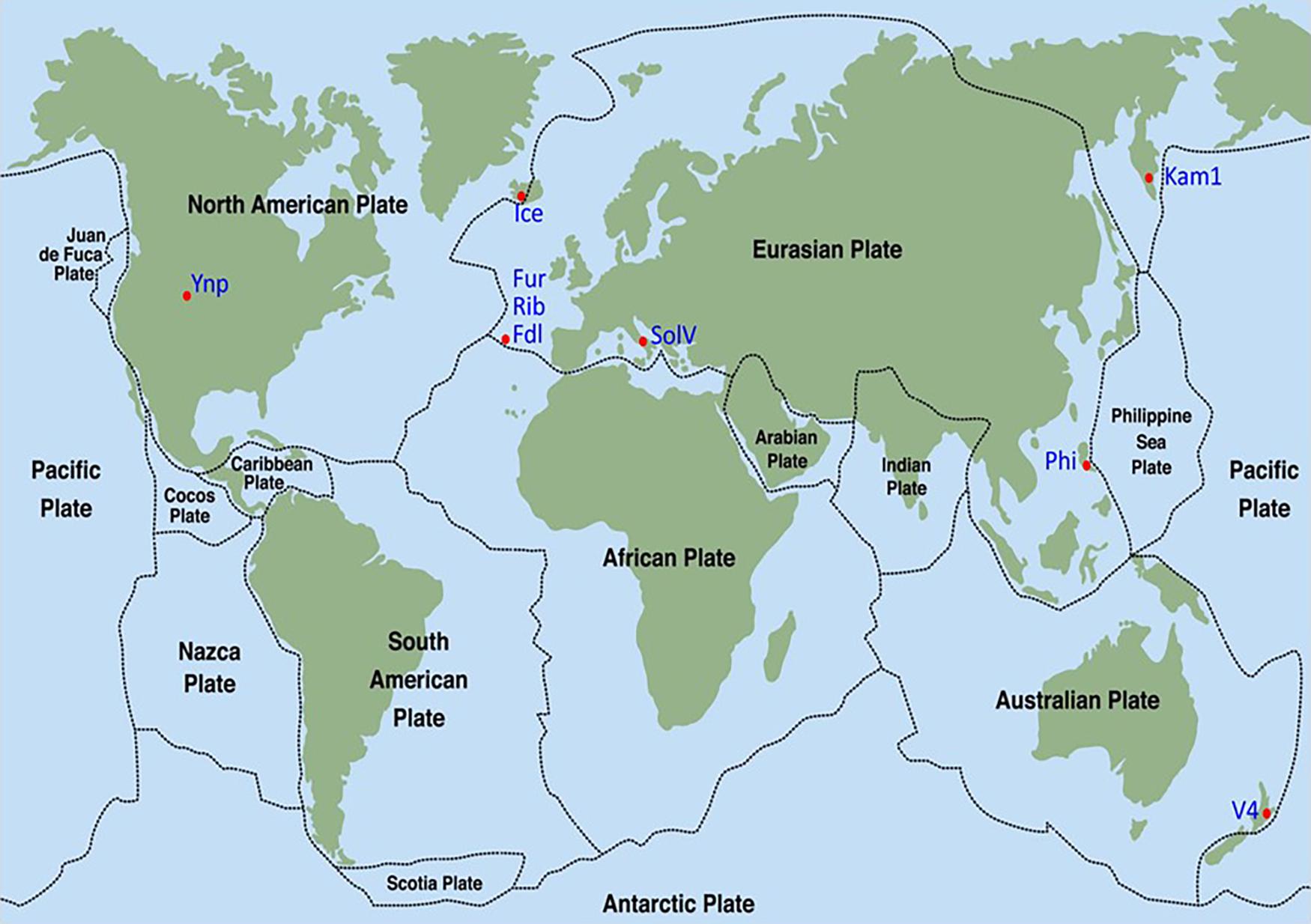
Figure 1. Sampling sites for isolation of thermoacidophilic methanotrophic Verrucomicrobia (indicated by red dots). Strain designations are indicated by blue color. Strippled lines indicate boundaries between tectonic plates.
Morphologically and physiologically, the six strains are very similar. Optimal growth was at 53–57°C and pH 2.5–3.5 (data not shown). Scanning electron microscopy images of the isolates confirmed that the cells are short rods, on average about 1–2 μm in length and 0.5 μm in diameter (Figure 2). Under transmission electron microscopy (TEM), multiple electron-dense inclusions, which are suspected stores of glycogen, are visible in all of the new isolates (Figure 2). These morphological and physiological data are consistent with previous reports. For example, in strain SolV similar inclusions were observed and were subsequently shown to contain glycogen (Khadem et al., 2012).
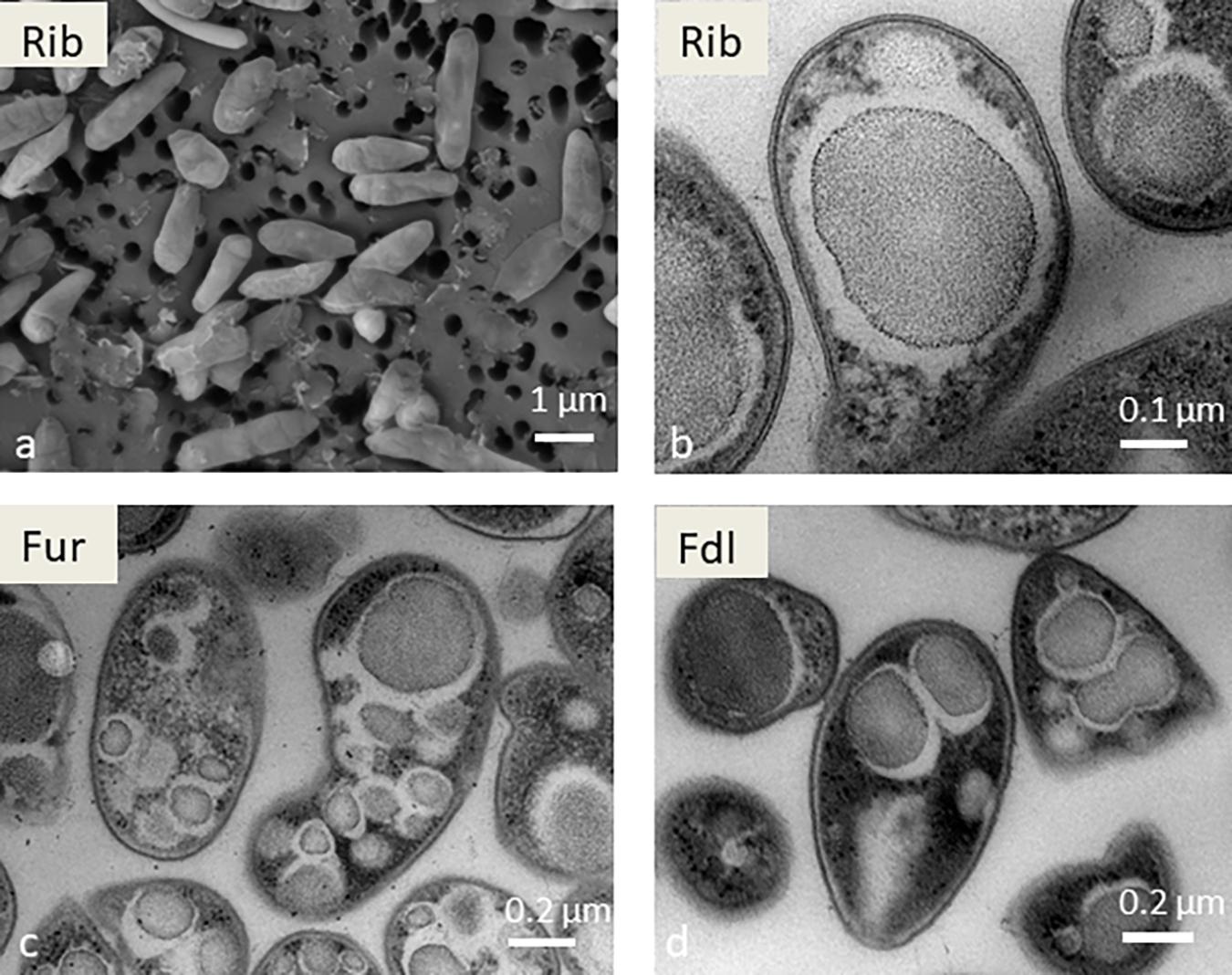
Figure 2. Scanning electron microscopy image of strain Rib (a) and transmission electron microscopy images of strains Rib (b), Fur (c), and Fdl (d). Scale bars are shown at the bottom right in all images.
Genome Analysis and Comparison
Like the three previously characterized strains, all of the six new isolates contain a single ribosomal RNA operon. All of the nine isolates are phylogenetically closely related, sharing ≥98.6% 16S rRNA sequence identity, which is clearly above the generally accepted species threshold value of 97% (Stackebrandt and Goebel, 1994) but close to the recently suggested species cutoff value of 98.7% (Chun et al., 2018). A phylogenetic tree based on concatenated sequences of seven housekeeping genes (gyrA, gyrB, ftsZ, rho, dnaG, rpoE, and groEL) illustrates that the European isolates group into a European clade with strain Kam1 and strain Yel as the closest relatives. Methylacidiphilum infernorum strain V4 (from New Zealand) and strain Phi appear to be phylogenetically distinct species (Figure 3), as also supported by the 16S rRNA tree (Supplementary Figure S1). Given the biogeographic boundaries of isolates from New Zealand (V4) and the Philippines and their relative proximity to one another when compared to the distance between these mid-to-south Pacific islands to Europe, the phylogenetic relationships are convincing. Moreover, in silico genome homology analyses (i.e., DNA-DNA hybridization or DDH) reveal that the strains Yel and Kam1 may also be regarded as separate species since DDH values were below the 70% DNA-DNA threshold line (see Table 2) considered to represent distinct species (Wayne et al., 1987). This is also supported by the pairwise average nucleotide identity (ANI) values (Supplementary Table S2). The European isolates (strains: Ice, Fur, Rib, Fdl, and Methylacidiphilum fumariolicum strain SolV) share more than 88% in silico gene homology while strains Phi and V4 only share less than 21.6% gene homology to the other strains.
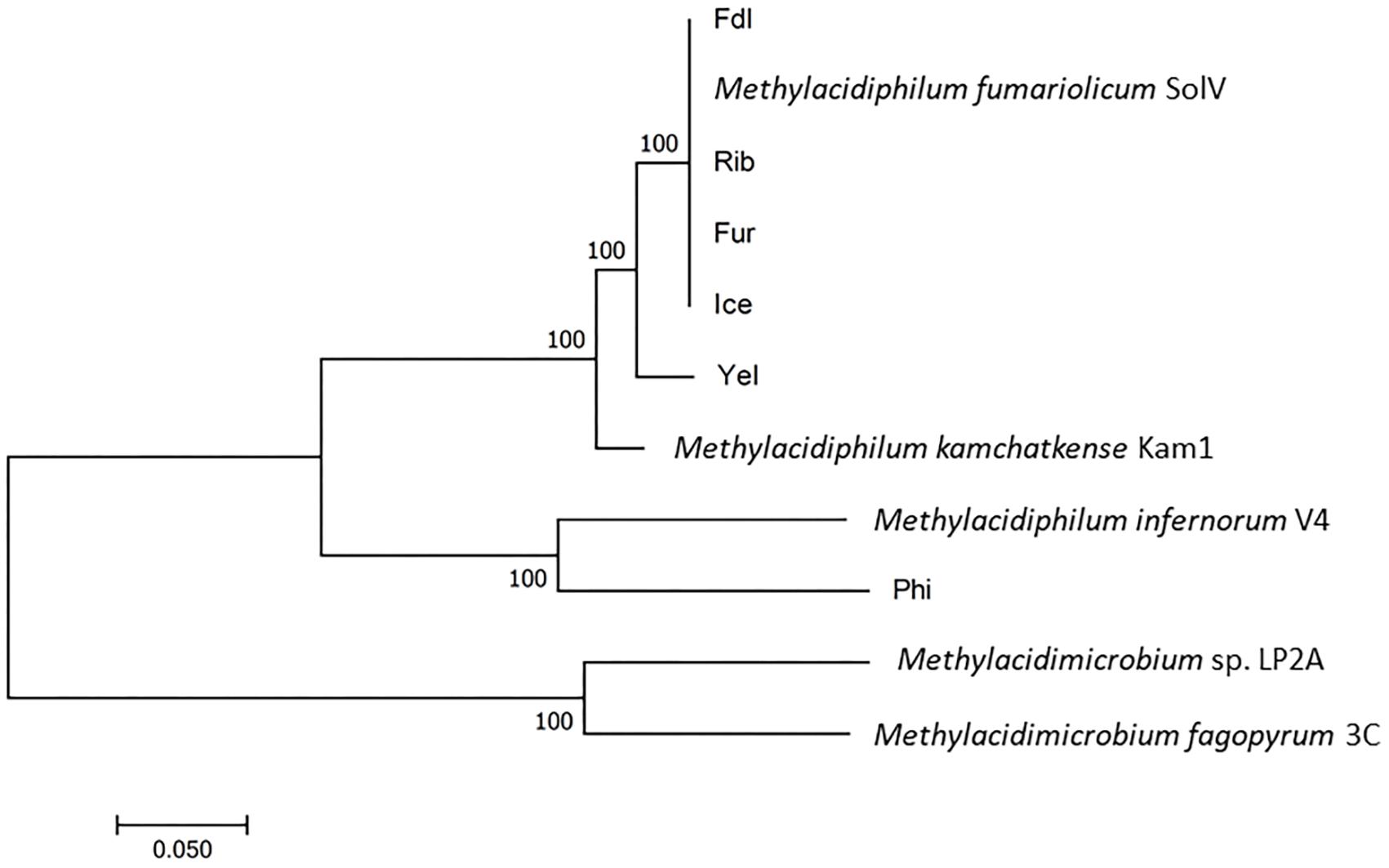
Figure 3. Phylogenetic analysis of Methylacidiphilum and Methylacidimicrobium strains by the Maximum Likelihood method using concatemers of seven housekeeping genes. The percentage of trees in which the associated taxa clustered together is shown next to the branches. The tree is drawn to scale, with branch lengths measured as the number of substitutions per site. Bootstrap values are indicated at nodes. All positions containing gaps and missing data were eliminated.
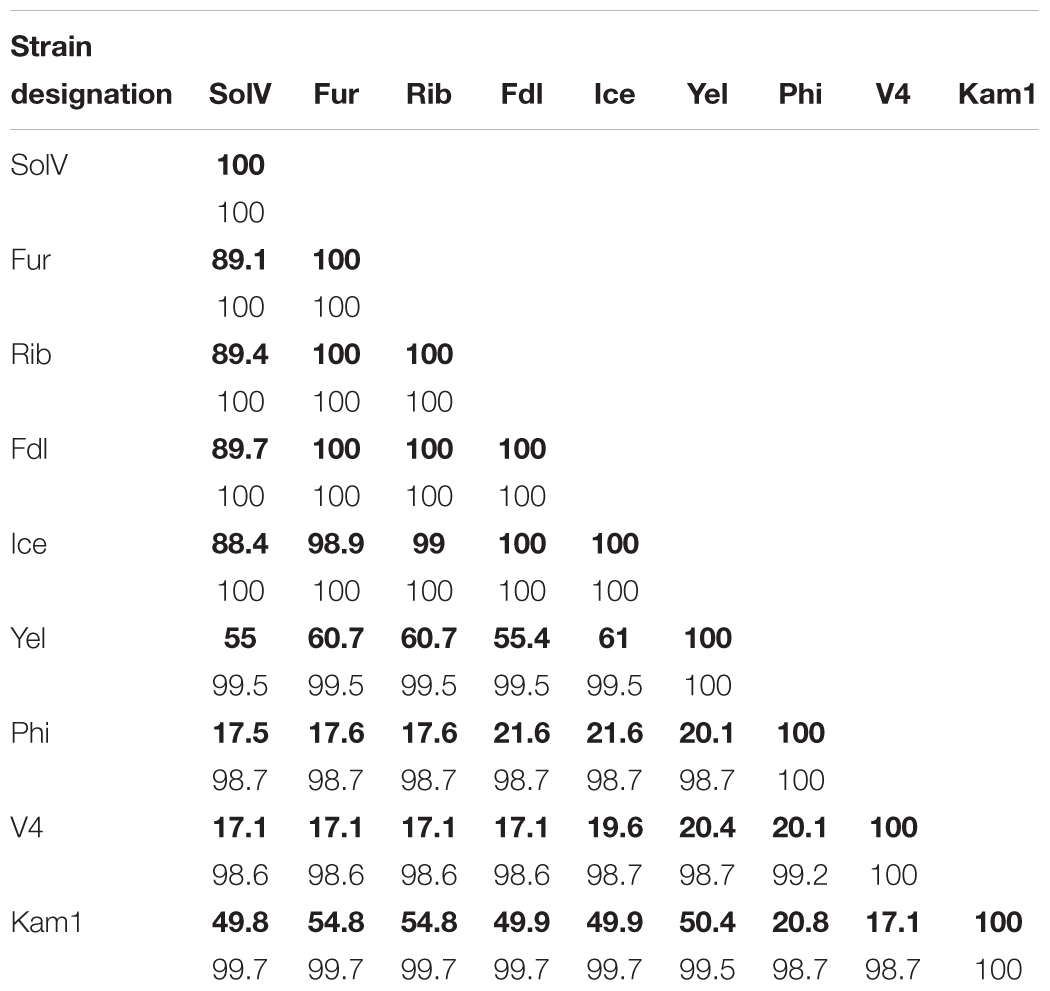
Table 2. Pairwise genomic DNA-DNA in silico hybridization values (in bold) and 16S rRNA sequence identities of all thermoacidophilic methanotrophic Verrucomicrobia isolates.
After reordering of genome contigs of strains Fdl, Yel, Phi, and Kam1 against the completed M. fumariolicum SolV genome (Anvar et al., 2014), all the new strains aligned reasonably well with the SolV genome, indicating a large degree of gene synteny among the genomes (Figure 4) despite a very low overall genome sequence similarity between some strains. Larger reorganizations and inversions are only seen for strain Phi. A few larger regions of unique non-homologous sequences can also be discerned in strains Kam1, Yel, and Phi. As compared to SolV, the four new genomes appear to be somewhat shorter, but a firm conclusion should await a complete PacBio sequence analysis.
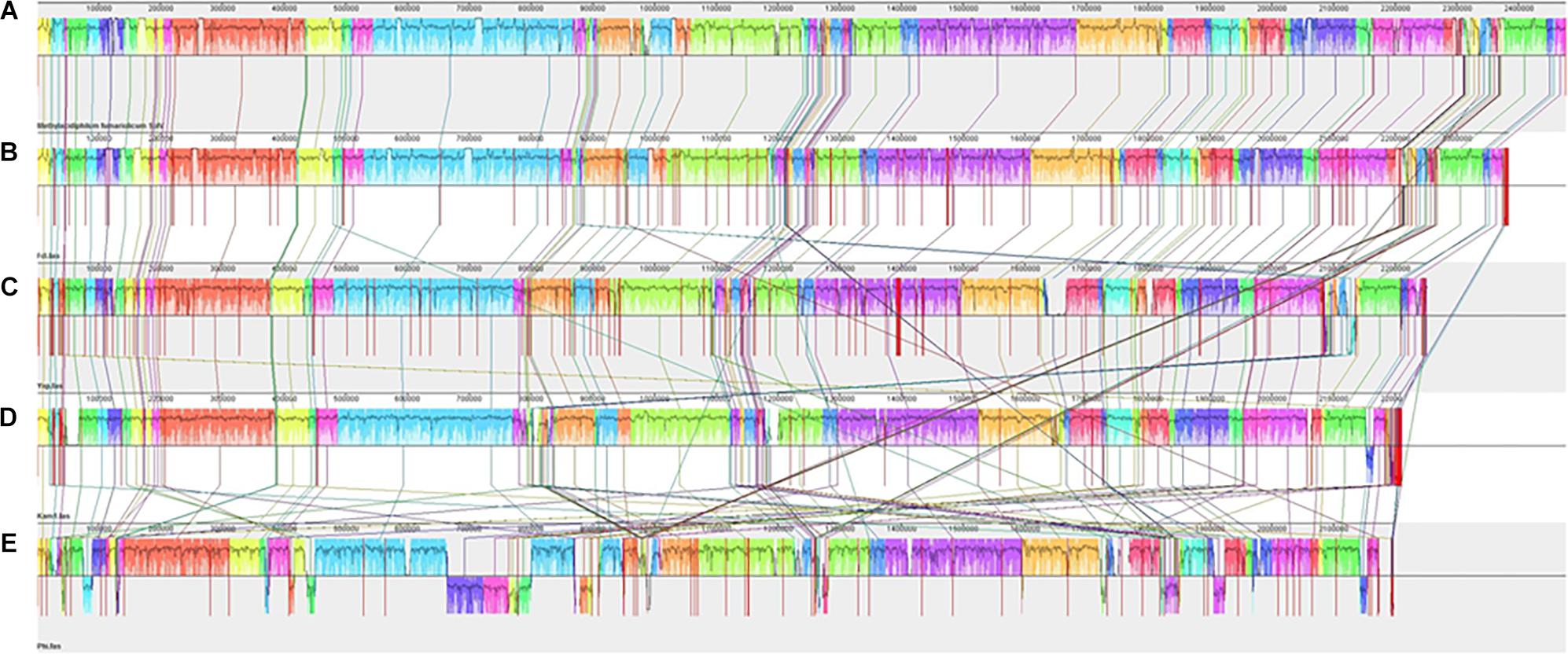
Figure 4. Alignment of the contigs of Methylacidiphilum spp. strains Fdl (B), Yel (C), Kam1 (D), and Phi (E) against the Methylacidiphilum fumariolicum SolV (A) chromosome using MAUVE 2.4.0. Vertical red lines indicate borders between contigs.
All of the six new isolates possess Rubisco-encoding genes, and thus appear to assimilate carbon from CO2 like has been shown for strain SolV (Khadem et al., 2011), as well as a complete set of nitrogen fixation genes. Strains Phi, Fur, and Fdl carry three copies of the pmoCAB genes like the three other described Methylacidiphilum species, while in strain Rib and Ice, only two complete pmoCAB operons could be identified from the draft genome sequences in addition to an apparently truncated pmoAB cluster. In strain Yel, only one complete pmoCAB cluster was identified, and two truncated pmoAB and pmoCA clusters. All strains contain one methanol dehydrogenase gene encoding the lanthanide-dependent XoxF type as described for SolV (Pol et al., 2014).
When internal transcribed spacer regions (ITS) and segments of the 23S rRNA sequence were included in the phylogenetic analysis, percent identity is somewhat reduced (≥91.75%) but remains above 90% (Figure 5). Comparisons made with a phylum-specific pmoA marker (i.e., pmoA2), sequence similarities are ≥79.9% (see Figure 5). Based on ongoing work in hyperthermophilic archaea and the hypothesis that there is correlation between genetic similarity and tectonic plate sympatry, the Mantel test was used to test for correlations between genetic divergence (among isolates) and geographic distance (i.e., distance between habitats from which isolates were derived). Alternatively, to test potential correlation between genetic divergence and tectonic sympatry, the Spearman correlation method (i.e., rs statistic) was employed. A positive correlation between genetic “distance” versus geographic distance emerged from the Mantel test (rM = 0.885, p < 0.001) (Figure 6). In addition, the Spearman analysis was able to support a relationship between genetic similarity and tectonic plate sympatry (rs = 0.532, p < 0.001, N = 9).
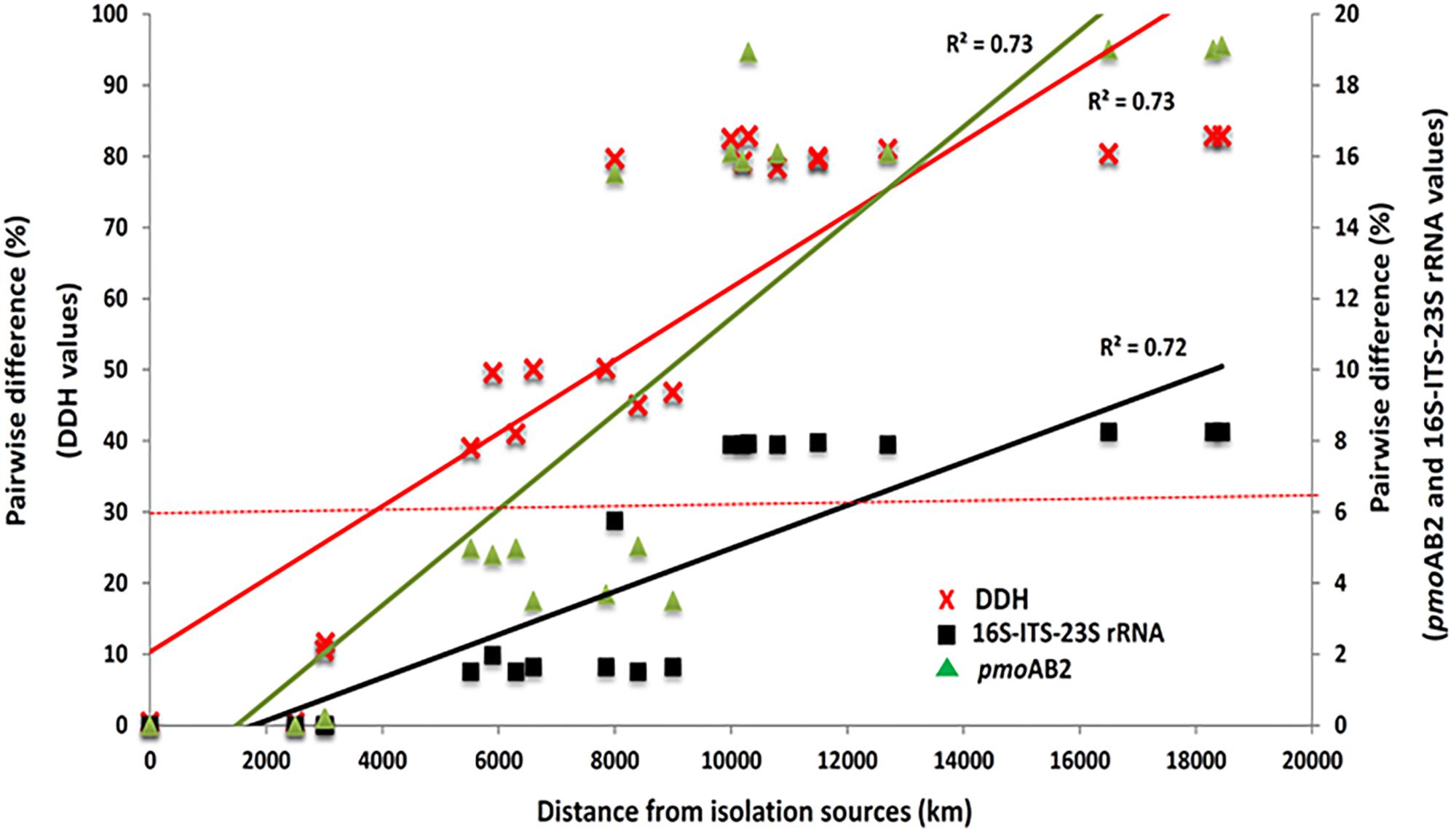
Figure 5. Scatter plot of pairwise nucleotide differences of the 16S-ITS-23S rRNA (black squares) and pmoAB2 (green triangles) gene sequences of the Methylacidiphilum spp. isolates and pairwise genome sequence differences (red crosses) based on in silico DNA-DNA hybridization (DDH) values. The left y-axis indicates genomic differences while the right y-axis indicates 16S-ITS-23S rRNA and pmoAB2 differences. The x-axis indicates the geographic distance between the isolation source for each pairwise strain comparison. A stippled demarcation line for DDH threshold for species separation (>30% difference or 70% > similarity) is included.
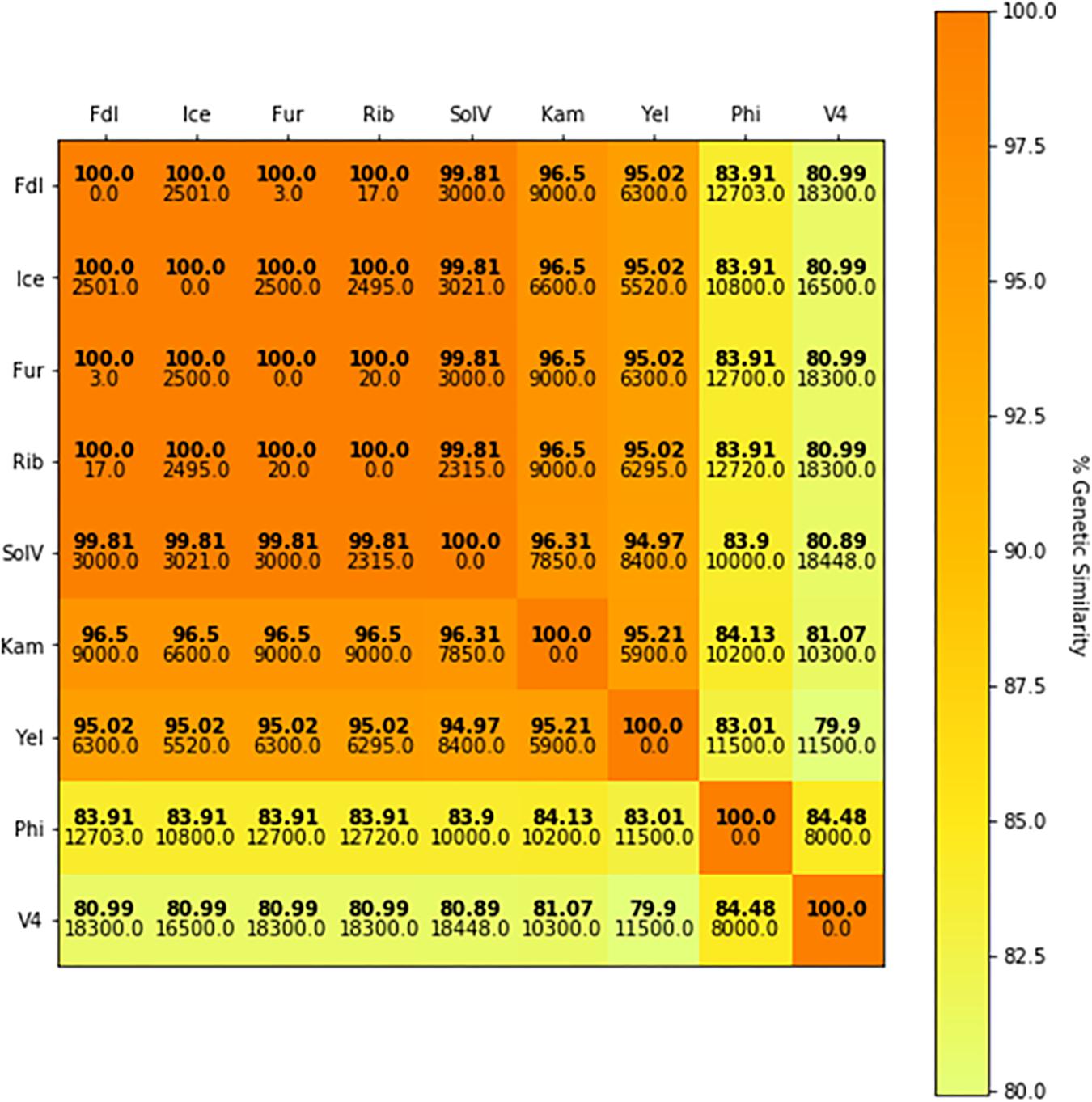
Figure 6. pmoAB2 sequence identity matrix. The color in the heat map represents percent sequence identity (%) for pmoAB2 across isolates (bold) with darker colors indicating higher sequence similarity between two isolates and lighter colors representing less sequence similarity. Geographic distances (km) between habitats from each pair of comparisons is shown below the genetic identity value. The Mantel statistic (rM) shows a significant correlation between genetic divergence and geographic distance (rM = 0.885, p < 0.001).
Chemotaxonomy and Serology
The PLFA profiles of the six new isolates and strain Kam1 displayed a dominance of saturated phospholipids (Supplementary Table S3). PLFAs i14:0, a15:0, and 18:0 account for more than 61% of the total PLFA content. Only trace amounts of unsaturated PLFAs: a13:1 12–13; 18:1 ω9c; and, 18:1 ω7c – were detected. Overall, the major PLFAs were present across most strains with some variability in abundances. One notable exception was the Icelandic strain (Ice), which exhibited a different PLFA profile: 15:0; i16:0; 16:0; 16:0 3OH; and, 17:0.
Antiserum raised against strain Kam1 showed strong strain-specificity in an ELISA experiment using whole cells of the isolates as antigens at high antiserum dilutions (≥51,200-fold) (Supplementary Figure S2). This demonstrates the presence of highly specific cell surface antigens in Kam1 as compared to the other isolates. The antigenic determinant is not known, but it might be LPS. At lower antiserum dilutions considerable cross-reactions can be observed, which are most likely caused by unspecific binding reactions.
Discussion
Methane from natural sources comprises an estimated ∼36% of total global CH4 emissions, while anthropogenic activities account for ∼64% (Bousquet et al., 2006). Fossil fuel use and farming activity have resulted in the doubling of methane emissions over the past 150 years (IPCC, 2007). Microbiological mitigation of natural methane release is accomplished, in part, by several taxonomic groups of methanotrophs (see review by Op den Camp et al., 2009). Although emissions from geothermal features worldwide may only account for up to 1% of the total methane release (Etiope and Klusman, 2002), methanotrophy by mesophilic and/or neutrophilic Alphaproteobacteria and Gammaproteobacteria of three well-established families: Methylocystaceae (α-proteobacteria), Beijerinckiaceae (α-proteobacteria), and Methylococcaceae (γ-proteobacteria) – is limited at extremes of low pH and high temperature. Reports of a few methanotrophs (Dunfield et al., 2007; Pol et al., 2007; Islam et al., 2008), which are capable of thriving in acidic (pH 0–4) and high temperature (∼65°C) environments, have led to the proposed family Methylacidiphilaceae (phylum: Verrucomicrobia). The global distribution and genetic relatedness of species within this family are still not well-understood. Given the distance, water bodies, and other geographic boundaries that exist between geothermal habitats worldwide, it is not known how conserved this taxon may be. From this study, all six previously uncharacterized isolates (i.e., Fdl, Rib, Fur, Yel, Ice, Phi) are shown to be closely related to one another as determined by 16S rRNA gene sequence comparisons as well as to the three previously described isolates of the family, specifically: SolV (Pol et al., 2007), V4 (Dunfield et al., 2007), and Kam1 (Islam et al., 2008). In addition to sharing high 16S rRNA gene sequence similarity (≥98.6%), morphological (i.e., size and shape) and physiological (e.g., growth rates and metabolism) properties of these newly reported geographically-distinct isolates suggest close relatedness.
However, DNA-DNA hybridization (DDH) studies indicate that not all of these new isolates are likely to be of the same species (and, perhaps, not even of the same genus). Indeed, employing the standard DDH cut-off (≥70%) for “same species” determination (Wayne et al., 1987; Goris et al., 2007), it appears isolates Fur, Fdl, Rib, SolV, and Ice are all geographic variants or geovars of the same species (see Table 2). Despite geographic barriers (i.e., waterways) between the Fur/Fdl/Rib group and SolV and Ice, all five geovars are within the same tectonic plate (i.e., the Eurasian Plate). Notably, isolate Ice originates from a geothermal region that borders the Eurasian Plate and the North American Plate (see Figure 1). By the proposed 16S rRNA gene identity threshold of >98.6% for “same species” determination (Chun et al., 2018), Yel may be the same species as Ice and the rest of the Eurasian group. However, the DDH value between isolate Yel and Ice is well below the standard 70% cut-off for DDH-based same species determination. Still, Yel shows a higher DDH value to Ice (61%) and other Eurasian Plate isolates (55–60.7%) than to isolates from other tectonic plate areas – V4 (20.4%), Phi (20.1%), and Kam1 (50.4%). Although the sampling site from which Phi was derived also sits at the edge of the Eurasian Plate (adjacent to the Philippine Sea Plate), DDH shows more distant relatedness between Phi and members of the Eurasian Plate group (i.e., Fur, Fdl, Rib, Ice, and SolV). Moreover, it appears that isolates Phi, V4, Kam1, and Yel, which each are derived from geothermal features in distinct tectonic plate regions are also more distantly-related in pairwise comparisons to one another than the within-group pairwise comparisons of isolates in Eurasian Plate clade. In short, this system exhibits biogeographic structure and tectonic boundaries may play a role in allopatric evolution and speciation. The proposed relationships between geography and genetic divergence are further supported by examination of phylogenetics of taxon-specific genes (i.e., pmoAB2) and concatenated contigs (i.e., 16S-ITS-23S) (see Figures 3–5).
Based on these results, we suggest that isolates Fur, Rib, and Fdl are the same species and that SolV and Ice are also geovars of that same species. However, Yel, Phi, V4, and Kam1 are all distinct species, perhaps within the same genus. We further suggest that species divergence from a common ancestor is the result of allopatric evolution within geographically-distinct geothermal habitats, which are not only separated by distance and water bodies but also by tectonic plate boundaries. As further isolates are collected from other regions in the world, we anticipate that novel species of the genus Methylacidiphilium will be characterized and that descriptions of additional isolates from other tectonic regions may resolve multiple genera within the family Methylacidiphilaceae.
Author Contributions
H-AE, N-KB, and RC conceived and designed the experiments and did the sampling. H-AE and NS performed the experiments under supervision of N-KB. All authors interpreted the results and contributed in writing the manuscript.
Funding
This work was supported by grants from the Research Council of Norway to the University of Bergen (Award Nos. 261923 and 204797; Principal Investigator: N-KB) and by a U.S. National Science Foundation RCN grant to the University of Arkansas (Award No. 1624171; Principal Investigator: RC).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Kelsey Bevenour (Ph.D. student in the Ceballos Lab) for assistance with statistical analyses. We are grateful to Gyri Teien Haugland (UiB) for assistance with the serological analyses.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2019.01129/full#supplementary-material
Footnotes
- ^ https://www.google.com/maps/
- ^ http://www.gatc-biotech.com
- ^ http://rast.nmpdr.org/
- ^ http://www.ncbi.nlm.nih.gov/genome/annotation_prok/
- ^ http://ggdc.dsmz.de/
- ^ http://darlinglab.org/mauve/mauve.html
- ^ https://www.dsmz.de/home.html
- ^ http:/www.biogenes.de
References
Anvar, S. Y., Frank, J., Pol, A., Schmitz, A., Kraaijeveld, K., den Dunnen, J. T., et al. (2014). The genomic landscape of the verrucomicrobial methanotroph Methylacidiphilum fumariolicum SolV. BMC Genomics 15:914. doi: 10.1186/1471-2164-15-914
Baas-Becking, L. G. M. (1934). Geobiologie of Inleiding tot de Milieukunde. The Hague: Van Stockum and Zoon.
Beeder, J., Nilsen, R. K., Rosnes, J. T., Torsvik, T., and Lien, T. (1994). Archaeoglobus fulgidus isolated from hot north sea oil field waters. Appl. Environ. Microbiol. 60, 1227–1231.
Birkeland, N. K., Schönheit, P., Poghosyan, L., Fiebig, A., and Klenk, H. P. (2017). Complete genome sequence analysis of Archaeoglobus fulgidus strain 7324 (DSM 8774), a hyperthermophilic archaeal sulfate reducer from a North Sea oil field. Stand. Genomic Sci. 12:79. doi: 10.1186/s40793-017-0296-5
Bousquet, P., Ciais, P., Miller, J. B., Dlugokencky, E. J., Hauglustaine, D. A., Prigent, C., et al. (2006). Contribution of anthropogenic and natural sources to atmospheric methane variability. Nature 443, 439–443.
Brewer, T. E., Handley, K. M., Carini, P., Gilbert, J. A., and Fierer, N. (2017). Genome reduction in an abundant and ubiquitous soil bacterium ‘Candidatus Udaeobacter copiosus’. Nat. Microbiol. 2, 2–7. doi: 10.1038/nmicrobiol.2016.198
Ceballos, R. M., Marceau, C. D., Marceau, J. O., Morris, S., Clore, A. J., and Stedman, K. M. (2012). Differential virus host-ranges of the Fuselloviridae of hyperthermophilic Archaea: implications for evolution in extreme environments. Front. Microbiol. 3:295. doi: 10.3389/fmicb.2012.00295
Christensen, B., Torsvik, T., and Lien, T. (1992). Immunomagnetically captured thermophilic sulfate-reducing bacteria from North Sea oil field waters. Appl. Environ. Microbiol. 58, 1244–1248.
Chun, J., Oren, A., Ventosa, A., Christensen, H., Arahal, D. R., Da Costa, M. S., et al. (2018). Proposed minimal standards for the use of genome data for the taxonomy of prokaryotes. Int. J. Syst. Evol. Microbiol. 68, 461–466. doi: 10.1099/ijsem.0.002516
Dahle, H., Garshol, F., Madsen, M., and Birkeland, N. K. (2008). Microbial community structure analysis of produced water from a high-temperature North Sea oil-field. Antonie Van Leeuwenhoek 93, 37–49.
Darling, A. C., Mau, B., Blattner, F. R., and Perna, N. T. (2004). Mauve: multiple alignment of conserved genomic sequence with rearrangements. Genome Res. 14, 1394–1403.
Dodge, Y. (ed.). (2008). “Spearman rank correlation coefficient,” in The Concise Encyclopedia of Statistics, (New York, NY: Springer), 502–505.
Dunfield, P. F., Yuryev, A., Senin, P., Smirnova, A. V., Stott, M. B., Hou, S. B., et al. (2007). Methane oxidation by an extremely acidophilic bacterium of the phylum Verrucomicrobia. Nature 450, 879–882.
Erikstad, H. A., and Birkeland, N. K. (2015). Draft genome sequence of “Candidatus Methylacidiphilum kamchatkense” strain Kam1, a thermoacidophilic methanotrophic verrucomicrobium. Genome Announc. 3:e00065-15. doi: 10.1128/genomeA.00065-15
Erikstad, H. A., Jensen, S., Keen, T. J., and Birkeland, N. K. (2012). Differential expression of particulate methane monooxygenase genes in the verrucomicrobial methanotroph ‘Methylacidiphilum kamchatkense’ Kam1. Extremophiles 16, 405–409. doi: 10.1007/s00792-012-0439-y
Etiope, G., and Klusman, R. W. (2002). Geologic emissions of methane to the atmosphere. Chemosphere 49, 777–789. doi: 10.1016/S0045-6535(02)00380-6
Glöckner, F. O., Zaichikov, E., Belkova, N., Denissova, L., Pernthaler, J., Pernthaler, A., et al. (2000). Comparative 16S rRNA analysis of lake bacterioplankton reveals globally distributed phylogenetic clusters including an abundant group of Actinobacteria. Appl. Environ. Microbiol. 66, 5053–5065.
Goris, J., Konstantinidis, K. T., Klappenbach, J. A., Coenye, T., Vandamme, P., and Tiedje, J. M. (2007). DNA-DNA hybridization values and their relationship to whole-genome sequence similarities. Int. J. Syst. Evol. Microbiol. 57, 81–91.
He, S. T., Zhi, X. Y., Jiang, H., Yang, L. L., Wu, J. Y., Zhang, Y. G., et al. (2015). Biogeography of Nocardiopsis strains from hypersaline environments of Yunnan and Xinjiang Provinces, western China. Sci. Rep. 5:13323. doi: 10.1038/srep13323
Held, N. L., and Whitaker, R. J. (2009). Viral biogeography revealed by signatures in Sulfolobus islandicus genomes. Environ. Microbiol. 11, 457–466. doi: 10.1111/j.1462-2920.2008.01784.x
Hou, S., Makarova, K. S., Saw, J. H., Senin, P., Ly, B. V., Zhou, Z., et al. (2008). Complete genome sequence of the extremely acidophilic methanotroph isolate V4, Methylacidiphilum infernorum, a representative of the bacterial phylum Verrucomicrobia. Biol. Direct 3:26. doi: 10.1186/1745-6150-3-26
Huber, R., Woese, C. R., Langworthy, T. A., Fricke, H., and Stetter, K. O. (1989). Thermosipho africanus gen-nov, represents a new genus of thermophilic eubacteria within the Thermotogales. Syst. Appl. Microbiol. 12, 32–37.
IPCC (2007). Climate Change 2007. Synthesis Report. Contribution of Working Groups I, II & III to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Geneva: IPCC. doi: 10.1017/CBO9780511546013
Islam, T., Jensen, S., Reigstad, L. J., Larsen, O., and Birkeland, N. K. (2008). Methane oxidation at 55o C and pH 2 by a thermoacidophilic bacterium belonging to the Verrucomicrobia phylum. Proc. Natl. Acad. Sci. U.S.A. 105, 300–304. doi: 10.1073/pnas.0704162105
Khadem, A. F., Pol, A., Wieczorek, A., Mohammadi, S. S., Francoijs, K.-J., Stunnenberg, H. G., et al. (2011). Autotrophic methanotrophy in verrucomicrobia: Methylacidiphilum fumariolicum SolV uses the Calvin-Benson-Bassham cycle for carbon dioxide fixation. J. Bacteriol. 193, 4438–4446. doi: 10.1128/JB.00407-11
Khadem, A. F., van Teeseling, M. C. F., van Niftrik, L., Jetten, M. S. M., Op den Camp, H. J. M., and Pol, A. (2012). Genomic and physiological analysis of carbon storage in the verrucomicrobial methanotroph “Ca. Methylacidiphilum fumariolicum” SolV. Front. Microbiol. 3:345. doi: 10.3389/fmicb.2012.00345
Kumar, S., Stecher, G., and Tamura, K. (2016). MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 33, 1870–1874. doi: 10.1093/molbev/msw054
Kuykendall, L. D., Roy, M. A., Oneill, J. J., and Devine, T. E. (1988). Fatty-acids, antibiotic-resistance, and deoxyribonucleic-acid homology groups of Bradyrhizobium japonicum. Int. J. Syst. Bacteriol. 38, 358–361.
Mantel, N. (1967). The detection of disease clustering and a generalized regression approach. Cancer Res. 27, 209–220.
Massana, R., Delong, E. F., and Pedros-Alio, C. (2000). A few cosmopolitan phylotypes dominate planktonic archaeal assemblages in widely different oceanic provinces. Appl. Environ. Microbiol. 66, 1777–1787.
Miller, L. T. (1982). Single derivatization method for routine analysis of bacterial whole-cell fatty acid methyl esters, including hydroxy acids. J. Clin. Microbiol. 16, 584–586.
Musser, J. M., Kroll, J. S., Granoff, D. M., Moxon, E. R., Brodeur, B. R., Campos, J., et al. (1990). Global genetic structure and molecular epidemiology of encapsulated Haemophilus influenzae. Rev. Infect. Dis. 12, 75–111.
Op den Camp, H. J. M., Islam, T., Stott, M. B., Harhangi, H. R., Hynes, A., Schouten, S., et al. (2009). Environmental, genomic and taxonomic perspectives on methanotrophic Verrucomicrobia. Environ. Microbiol. Rep. 1, 293–306. doi: 10.1111/j.1758-2229.2009.00022.x
Pol, A., Barends, T. R. M., Dietl, A., Khadem, A. F., Eygensteyn, J., Jetten, M. S. M., et al. (2014). Rare earth metals are essential for methanotrophic life in volcanic mudpots. Environ. Microbiol. 16, 255–264. doi: 10.1111/1462-2920.12249
Pol, A., Heijmans, K., Harhangi, H. R., Tedesco, D., Jetten, M. S. M., and Op Den Camp, H. J. M. (2007). Methanotrophy below pH1 by a new Verrucomicrobia species. Nature 450, 874–878.
Reno, M. L., Held, N. L., Fields, C. J., Burke, P. V., and Whitaker, R. J. (2009). Biogeography of the Sulfolobus islandicus pan-genome. Proc. Natl. Acad. Sci. U.S.A. 106, 8605–8610. doi: 10.1073/pnas.0808945106
Roberts, M. S., and Cohan, F. M. (1995). Recombination and migration rates in natural populations of Bacillus subtilis and Bacillus mojavensis. Evolution 49, 1081–1094. doi: 10.1111/j.1558-5646.1995.tb04435.x
Smith, J. M., Dowson, C. G., and Spratt, B. G. (1991). Localized sex in bacteria. Nature 349, 29–31.
Snyder, J. C., Wiedenheft, B., Lavin, M., Roberto, F. F., Spuhler, J., Ortmann, A. C., et al. (2007). Virus movement maintains local virus population diversity. Proc. Natl. Acad. Sci. U.S.A. 104, 19102–19107.
Stackebrandt, E., and Goebel, B. M. (1994). Taxonomic note: a place for DNA-DNA reassociation and 16S rRNA sequence analysis in the present species definition in bacteriology. Int. J. Syst. Bacteriol. 44, 846–849. doi: 10.1099/00207713-44-4-846
Stetter, K. O. (1988). Archaeoglobus fulgidus gen-nov, sp-nov - a new taxon of extremely thermophilic Archaebacteria. Syst. Appl. Microbiol. 10, 172–173.
Tamura, K., and Nei, M. (1993). Estimation of the number of nucleotide substitutions in the control region of mitochondrial-DNA in humans and chimpanzees. Mol. Biol. Evol. 10, 512–526.
Van Teeseling, M. C. F., Pol, A., Harhangi, H. R., Van Der Zwart, S., Jetten, M. S. M., and Op Den Camp, H. J. M. (2014). Expanding the verrucomicrobial methanotrophic world: description of three novel species of Methylacidimicrobium gen. nov. Appl. Environ. Microbiol. 80, 6782–6791. doi: 10.1128/AEM.01838-14
Wayne, L. G., Brenner, D. J., Colwell, R. R., Grimont, P. A. D., Kandler, O., Krichevsky, M. I., et al. (1987). Report of the Ad-Hoc-committee on reconciliation of approaches to bacterial systematics. Int. J. Syst. Bacteriol. 37, 463–464.
Whitaker, R. J., Grogan, D. W., and Taylor, J. W. (2003). Geographic barriers isolate endemic populations of hyperthermophilic Archaea. Science 301, 976–978.
Keywords: Methylacidiphilum, biogeography, Verrucomicrobia, methanotrophs, acidophiles, PVC superphylum, allopatric evolution
Citation: Erikstad H-A, Ceballos RM, Smestad NB and Birkeland N-K (2019) Global Biogeographic Distribution Patterns of Thermoacidophilic Verrucomicrobia Methanotrophs Suggest Allopatric Evolution. Front. Microbiol. 10:1129. doi: 10.3389/fmicb.2019.01129
Received: 10 January 2019; Accepted: 03 May 2019;
Published: 29 May 2019.
Edited by:
Peter Dunfield, University of Calgary, CanadaReviewed by:
Sayaka Mino, Hokkaido University, JapanArjan Pol, Radboud University Nijmegen, Netherlands
Copyright © 2019 Erikstad, Ceballos, Smestad and Birkeland. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nils-Kåre Birkeland, bmlscy5iaXJrZWxhbmRAdWliLm5v