
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 24 May 2019
Sec. Food Microbiology
Volume 10 - 2019 | https://doi.org/10.3389/fmicb.2019.01046
This article is part of the Research Topic Insights of Fermented Foods and Beverages: Microbiology and Health-Promoting Benefits View all 19 articles
 S. González1,2
S. González1,2 T. Fernández-Navarro1,2
T. Fernández-Navarro1,2 S. Arboleya2,3
S. Arboleya2,3 C. G. de los Reyes-Gavilán2,3
C. G. de los Reyes-Gavilán2,3 N. Salazar2,3
N. Salazar2,3 M. Gueimonde2,3*
M. Gueimonde2,3*The intake of fermented foods is gaining increasing interest due to their health-promoting benefits. Among them, fermented dairy foods have been associated with obesity prevention, and reduction of the risk of metabolic disorders and immune-related pathologies. Fermented foods could lead to these health benefits by providing the consumer with both easily metabolizable nutrients and beneficial microorganisms. Our aim was to evaluate the relationship between the consumption of fermented dairy products and the intestinal microbiota, serum lipid profile, and the pro-oxidant/inflammatory status. 130 healthy adults were evaluated. Dietary fermented food intake was assessed by an annual food frequency questionnaire (FFQ), including 26 fermented dairy products. Levels of the major phylogenetic types of the intestinal microbiota were determined by qPCR, and concentration of fecal short chain fatty acids were assessed by gas chromatography. Serum glucose and lipid profile, as well as serum malondialdehyde (MDA), C-reactive protein (CRP), and leptin levels were determined by standardized protocols. Among fermented dairy foods, natural yogurt, sweetened yogurt and matured/semi-matured cheese were the most consumed. While natural yogurt consumers showed increased fecal levels of Akkermansia with respect to non-consumers, sweetened yogurt intake was associated to lower levels of Bacteroides. Serum levels of CRP were also significantly reduced in yogurt consumers. Our results underline the interest in exploring the potential effects of the different yogurt types and the role the microbiota may play in such effects.
Fermented foods have played an important role in human diet since the development of civilization and represent a special feature of some dietary patterns, such as the Mediterranean one. The initial goal of the fermentation process was to prolong the useful-life of some foods and beverages, and improving their safety, digestibility and organoleptic properties, however, nowadays fermented products have become more popular than ever before due to their health-promoting benefits (Şanlier et al., 2017). Fermented dairy foods have received special attention because of their association in epidemiological studies with obesity prevention, and with the reduction on the risk of different diseases, including metabolic disorders, cardiovascular and immune-related diseases or cognitive decline, among others (Guo et al., 2017; Salas-Salvadó et al., 2017; Kok and Hutkins, 2018; Sivamaruthi et al., 2018). Apart from their content of fatty acids, vitamins, and minerals, these products contain bioactive peptides and living microorganisms that could modulate the immune responses and impact on the intestinal microbiota (IM) composition and functionality (Chakrabarti et al., 2014; Severyn and Bhatt, 2018). The human IM is a complex and dynamic community, represented by trillions of microorganisms, that plays an important role in the maintenance of health. Indeed, recent studies have consistently identified disease-specific microbiota signatures in different health disorders (Duvallet et al., 2017). The microbiota of healthy adults is represented mainly by anaerobic bacteria from the Firmicutes and Bacteroidetes phyla (Eckburg et al., 2005). While the genera Clostridium, Enterococcus, Lactobacillus and Faecalibacterium are predominant within the Firmicutes phylum, others such as Bacteroides and Prevotella are the most representative of the Bacteroidetes phylum (Eckburg et al., 2005). All of them are present in different proportions depending on the specific microbial composition of each individual. The disruption and alteration of the microbiota may be related to different pathologies and, for this reason, the search for strategies capable of reversing the IM dysbiosis in order to improve the health status of the host has become a key area of interest for the scientific community. In this regard, long-term dietary habits, as well as specific food constituents, such as fiber or phenolics, have been identified as critical drivers of gut microbiota composition (Wu et al., 2011; Fernández-Navarro et al., 2018). Fermented products may also modulate the IM (Kato-Kataoka et al., 2016), however, the association between fermented foods as part of the regular diet and the IM composition has not been sufficiently studied yet (Alvaro et al., 2007; Uyeno et al., 2008). In this regard, a recent work examining the impact of consuming a fermented milk containing microorganisms from the genera Lactobacillus and Bifidobacterium on the IM has reported a gender-specific increase in the levels of these two bacteria in the feces of volunteers (Lisko et al., 2017). The administration of a probiotic fermented milk, containing Streptococcus thermophilus, Lactobacillus bulgaricus, Lactobacillus acidophilus LA5 and Bifidobacterium animalis subsp. lactis BB12, during the third trimester of pregnancy has been related with a reduced risk of maternal insulin resistance (Asemi et al., 2013). Yogurt consumption has been associated with immune effects, including a reduced concentration of inflammatory markers in pregnant woman (Asemi et al., 2011). It has also been reported that yogurt modulates both humoral (Meyer et al., 2007) and cellular (Chaves et al., 2011) immunity. Unfortunately, very often observational nutritional studies do not inform us as to whether the positive effect of fermented dairy foods is mediated by the microorganisms present, by some specific components of the product, or by the potential role of some of these products, i.e., yogurt, as a marker of a good overall diet (Kok and Hutkins, 2018). Nevertheless, it is worth underlining that some studies draw attention to the impact yogurt could have, independent of diet (Panahi et al., 2018).
Based on this evidence, it seems reasonable to hypothesize that some of the described beneficial effects of fermented dairy product on several pathologies, such as those affecting the cardiovascular and metabolic systems, might be partly explained by the potential changes induced in the gut microbiota (Marco et al., 2017; Kok and Hutkins, 2018). Thus, in this study we aimed at evaluating the relationship between the consumption of fermented dairy products within the regular diet and the intestinal microbiota. In addition, selected blood markers related with the metabolic profile of the subjects were also analyzed.
This cross-sectional study sample comprised of 130 subjects from the Principality of Asturias Region (Northern Spain). Inclusion criteria were: not being diagnosed with diseases related to intestinal function, not being currently treated with corticoids, nor having consumed pro- and prebiotic supplements or antibiotics during the previous month. Participants were mentally and physically able to participate in the study and gave written informed consent. Ethical approval was obtained from the Bioethics Committee of CSIC and from the Regional Ethics Committee for Clinical Research of the Principality of Asturias in compliance with the Declaration of Helsinki of 1964. All experiments were carried out in accordance with approved guidelines and regulations.
Blood samples were kept on ice and centrifuged (1000 ×g, 15 min) within 2–4 h after collection. Plasma and serum aliquots were kept at -20°C until analyses were performed. Plasma glucose, cholesterol, and triglycerides were determined by standard methods. Serum levels of C-reactive protein (CRP) were assessed using a CRP Human Instant ELISA kit (eBioscience, San Diego, CA, United States), and those of malondialdehyde (MDA) with a colorimetric assay of lipid peroxidation (Bioxytech LPO-586, Oxis International SA, Paris, France); the within-run coefficient of variation ranged from 1.2 to 3.4%, depending on the concentration of MDA (Gerard-Monnier et al., 1998). Serum leptin was measured by a sensitive ELISA test (Human Leptin ELISA Development Kit, PeproTech Inc., Rocky Hill, CT, United States); the detectable concentration range was 63–4000 pg/mL and the intra-assay and inter-assay coefficients of variation were 5.21 and 5.20%, respectively.
Fecal samples were immediately frozen at -20°C and transported to the laboratory. For analyses fecal samples were melted, weighed, diluted 1/10 in sterile PBS, and homogenized at full-speed in a LabBlender 400 Stomacher (Seward Medical, London, United Kingdom) for 4 min. The samples were then centrifuged and the supernatant was taken for SCFA analyses whereas the fecal pellet was used for DNA extraction using the QIAamp DNA stool mini kit (Qiagen, Hilden, Germany) as previously described (Arboleya et al., 2012).
Quantification of different bacterial populations was performed with a 7500 Fast Real-Time PCR System (Applied Biosystems, Foster City, CA, United States) using SYBR Green PCR Master Mix (Applied Biosystems), and covered the major bacterial groups present in the gut microbial ecosystem. One microliter of template fecal DNA (∼5 ng) and 0.2 μM of each primer were added to the 25 μL reaction mixture. PCR cycling consisted of an initial cycle of 95°C 10 min, followed by 40 cycles of 95°C 15 s, and 1 min at the appropriate primer-pair temperature. The number of cells was determined by comparing the Ct values obtained from a standard curve. Fecal DNA extracts were analyzed and the mean quantity per gram of fecal wet weight was calculated as indicated elsewhere (Arboleya et al., 2012).
The analysis of SCFA was performed by gas chromatography in system composed of a 6890N GC injection module (Agilent Technologies Inc., Palo Alto, CA, United States) connected to a flame injection detector (FID) and a mass spectrometry (MS) 5973N detector (Agilent), as described previously (Arboleya et al., 2016).
Dietary intake was assessed in a personal interview by means of an annual semi-quantitative food frequency questionnaire (FFQ) method validated in previous studies (Cuervo et al., 2014). The FFQ was organized by food groups and open-ended, allowing foods consumed by the subject and not present in the questionnaire to be recorded. Among the dairy products group, 26 items were listed, including the three major fermented food groups: yogurt, cheese, and fermented milk. Food intake was analyzed for energy, macronutrients, and total dietary fiber content by using the nutrient Food Composition Tables developed by CESNID (Centro de Enseñanza Superior de Nutrición Humana y Dietética [CESNID], 2008). Additionally, the following fiber components were ascertained using (Marlett and Cheung, 1997) food composition tables: soluble fiber, insoluble fiber based on the enzymatic-chemical method developed by Theander and Westerlund (1986).
Height and weight were recorded after an overnight fast, using the standardized procedures described previously (Fernández-Navarro et al., 2017) for BMI [weight (Kg)/height (m2)]. Body fat percentage was measured by bioelectrical impedance (BIA) with ± 1% variation (Tanita Corporation of America, Inc., Arlington Heights, IL, United States).
Statistical analysis was performed using the IBM SPSS program version 22.0 (IBM SPSS, Inc., Chicago, IL, United States). Goodness of fit to the normal distribution was analyzed by means of the Kolmogorov-Smirnov test. Categorical variables were summarized with percentages while continuous variables were summarized using mean and standard deviations. The chi-squared test and independent samples t-test were used for group comparisons where appropriate. Pearson bivariate correlation was used to investigate linear association between the intake of total fermented dairy products and each subgroup, with the major food groups consumed in the diet and to describe the relationship between the consumption of fermented dairy foods with fecal microbial levels and serum health-related biomarkers. Heatmap was generated under R version 3.5.1 package heatmap.2. The conventional probability value for significance (0.05) was used in the interpretation of results.
The general characteristics of the study sample defined a group of 130 healthy adults with a mean age of 58.2 ± 17.1 years, and a moderate overweight (Table 1). 12.3% of the sample were smokers, and 55% lived sedentary lifestyles. To avoid potential confounding factors, like age or gender, these variables were included as covariables in any further analysis.
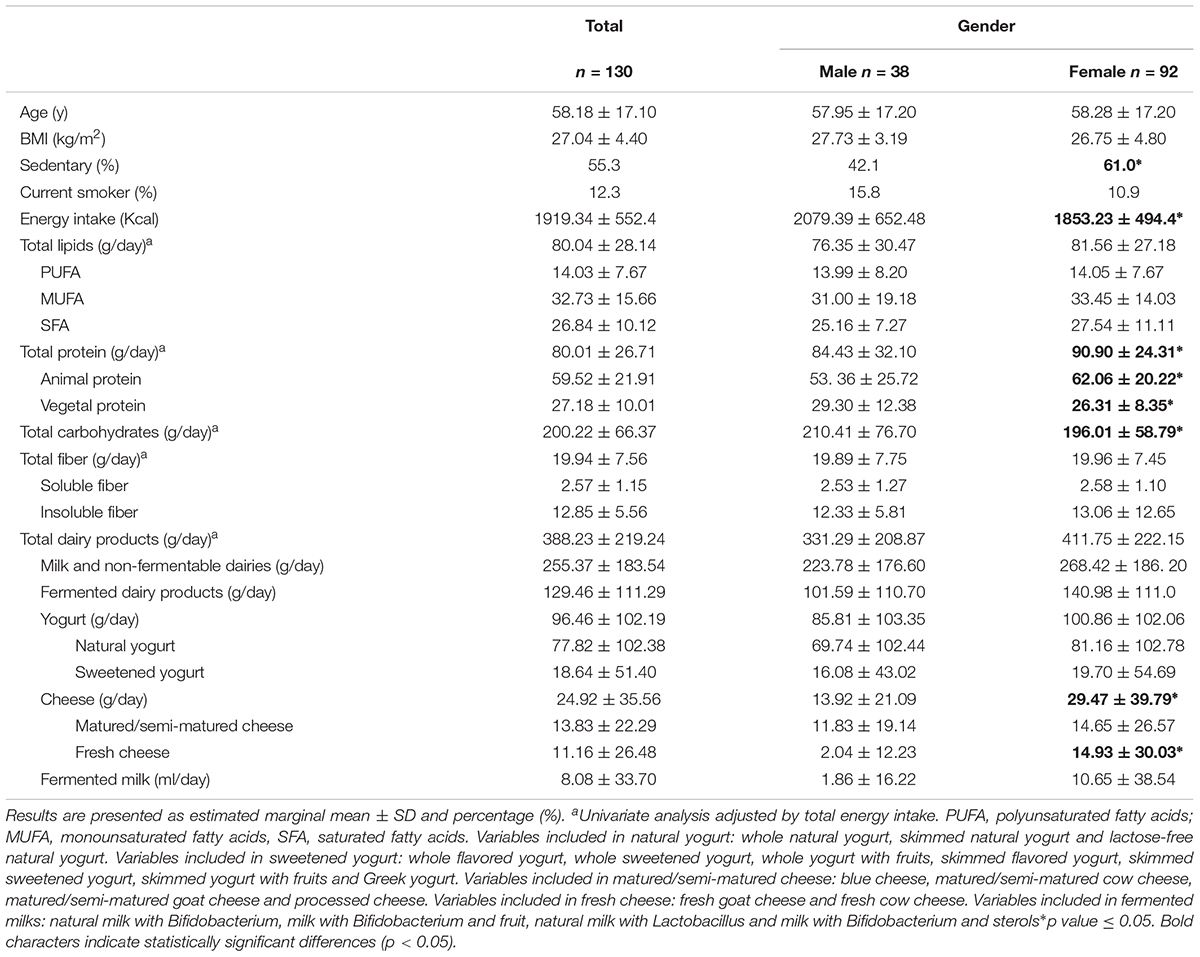
Table 1. General description of the study sample.
The total consumption of milk and dairy products (388.23 g/day) (Table 1) corresponded, in 33% of the sample, to the intake of fermented dairy foods, mainly yogurt and cheese (75 and 19%, respectively), as shown in Figure 1A. Among fermented dairy foods, natural yogurt (77.82 ± 102.38 g/day), sweetened yogurt (18.64 ± 51.40 g/day) and matured/semi-matured cheese (13.83 ± 22.29 g/day) were the most consumed (Table 1). Among them, natural yogurt was the main contributor (Figure 1B). The relationship between fermented dairy products and major food groups from the diet is shown in Figure 2. The consumption of fermented dairy foods presented a significant positive association with the intake of total dairy products, oils and fats, and dried fruits. In more detail, yogurt was negatively related to the intake of non-alcoholic beverages, and the consumption of cheese presented a direct relation with cereals, and fruits from the regular diet. Focusing on yogurt types, natural yogurt was directly related to the intake of dairy products and fruits, and negatively associated with sugars, sauces and non-alcoholic beverages; on the contrary, the intake of sweetened yogurt was positively related to these latter food groups (Figure 2). In the case of cheese, matured/semi-matured cheese consumption presented a positive relationship with the intake of cereals, while fresh cheese did it with fruits. Fermented milk has not been significantly associated with the intake of none of the other assessed food groups (Figure 2).
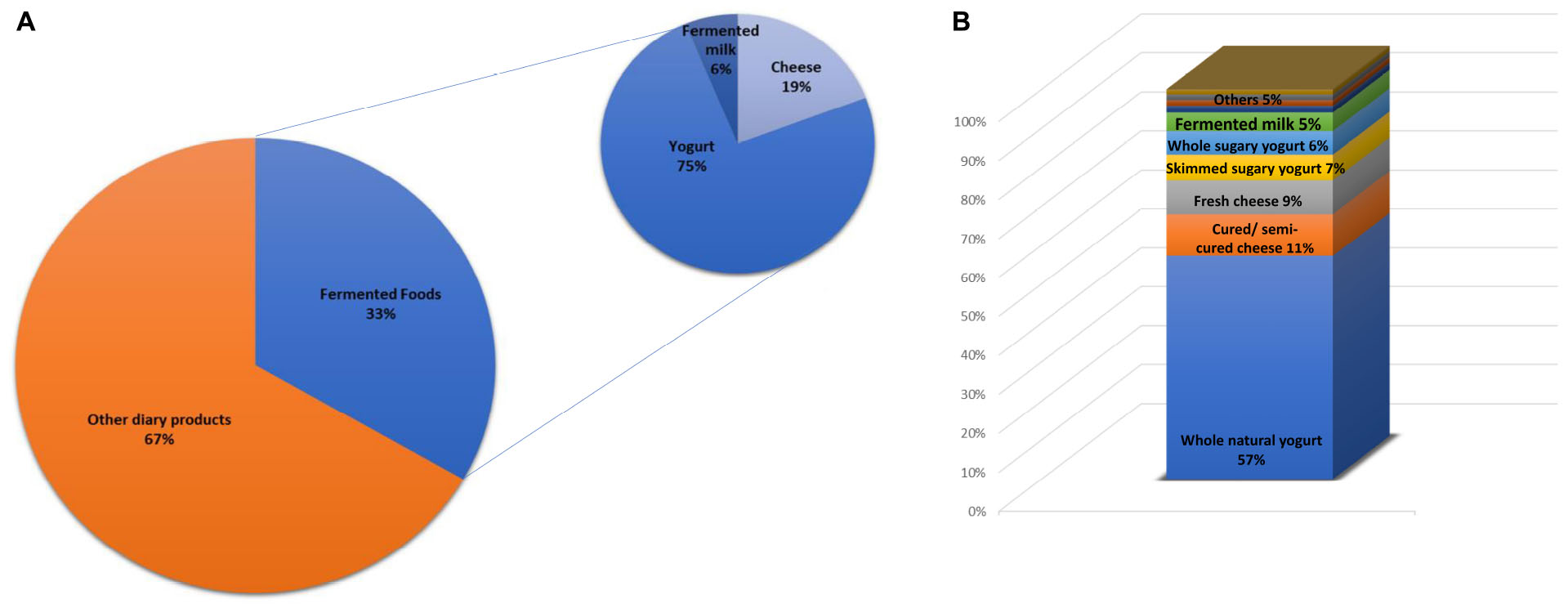
Figure 1. (A) Contribution (%) of the fermented dairy foods and main subgroups to the total intake of dairy products. (B) Intake proportion of detailed fermented dairy foods in the sample.
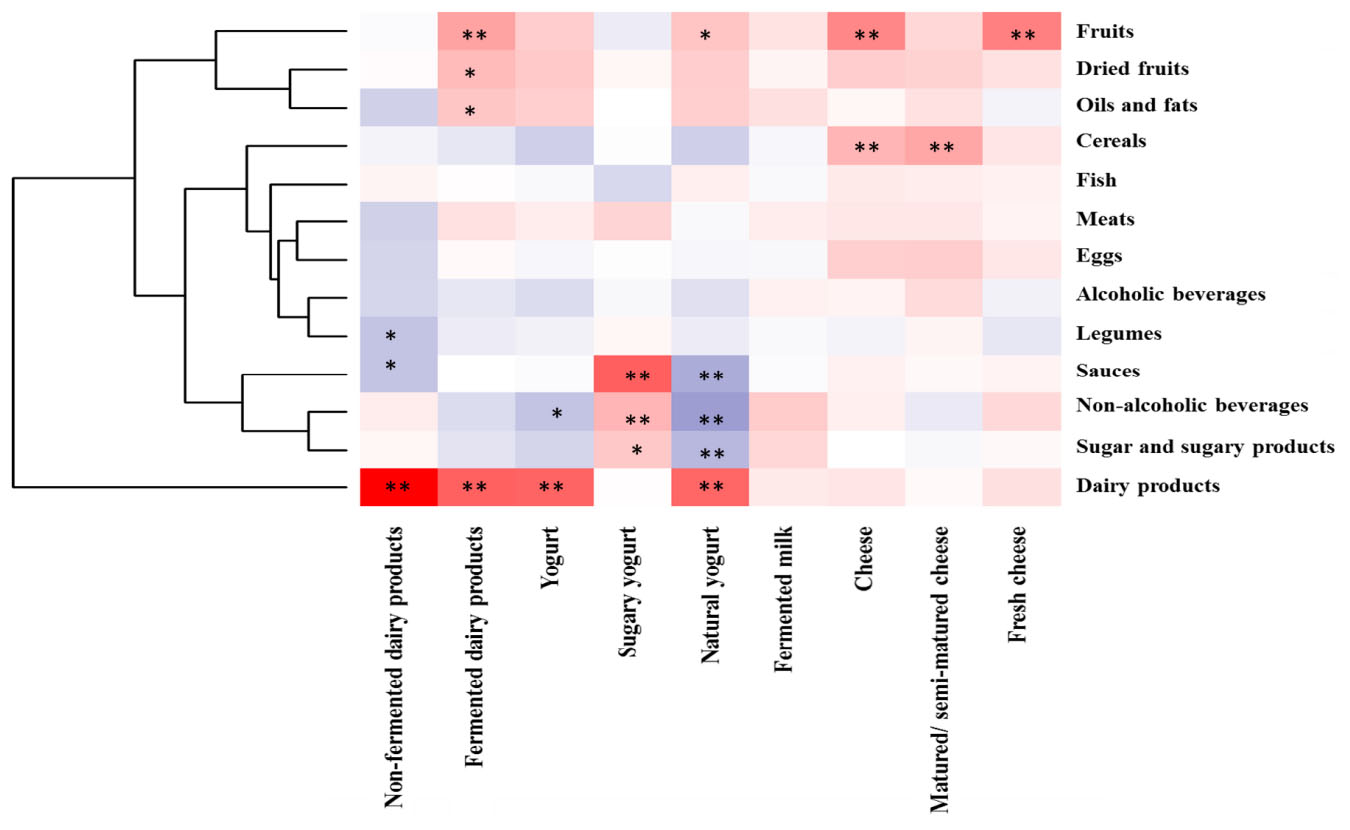
Figure 2. Pearson correlation between the intake of major food groups (g/day) with fermented dairy foods (g/day) in the sample. Columns correspond to main fermented dairy products whereas rows correspond to food groups. Blue and red colors denote negative and positive association, respectively. The intensity of the color represents the degree of association between the fermented dairies consumed in the sample and major food groups in the diet. Asterisks indicate significant associations: ∗p < 0.05; ∗∗p ≤ 0.01.
Regarding fecal microbial composition, natural yogurt consumers showed significantly higher fecal levels of Akkermansia, and sweetened yogurt consumers displayed significantly lower fecal levels of Bacteroides than non-consumers. Moreover, cheese consumers (considering all types jointly) presented significantly higher levels of the major fecal SCFA, acetate, propionate and butyrate, whereas the consumers of fresh cheese specifically presented higher levels of propionate and butyrate than non-consumers (Table 2).
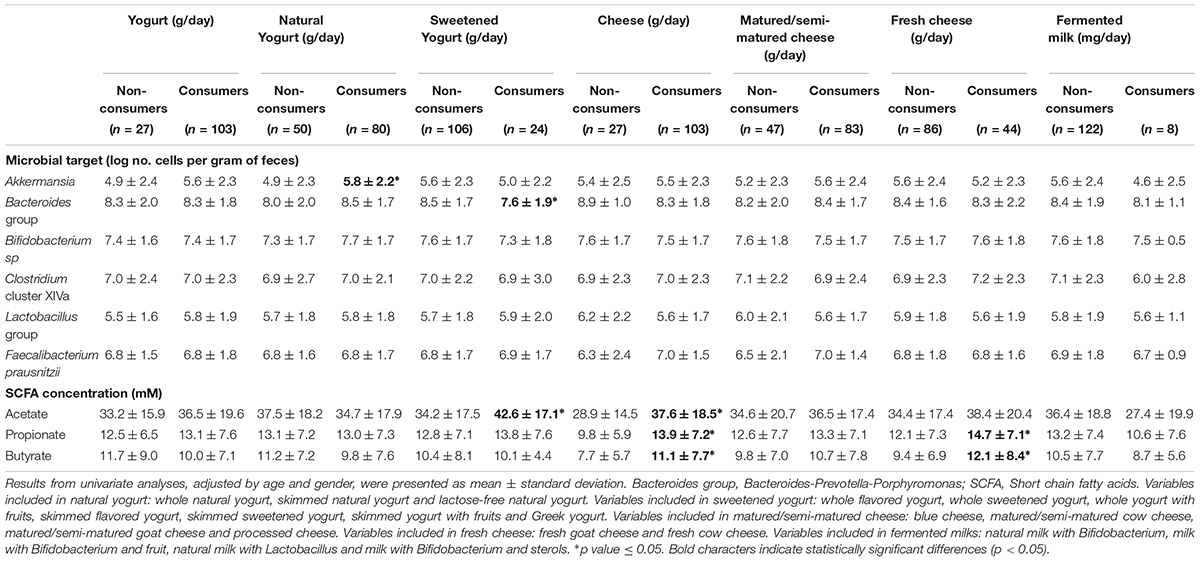
Table 2. Differences in the concentration of major microbial groups and short chain fatty acids according to the intake of the different types of fermented dairy foods consumed by the sample.
Delving into the impact of fermented dairy foods on health status, the association between them and serum health biomarkers was analyzed. While the intake of yogurt, especially natural yogurt, showed a direct association with LDL/HDL ratio values, serum CRP was significantly lower in yogurt consumers (5.5 ± 10.5 vs. 2.1 ± 4.6 mg/L). Moreover, natural yogurt was associated with the oxidant status, the consumers of this product showing also lower levels of serum MDA (2.80 ± 1.33 vs. 2.28 ± 0.59 μM) than non-consumers (Table 3). The intake of cheese and its different types or fermented milk did not show any association with any health-related biomarker (Table 3).
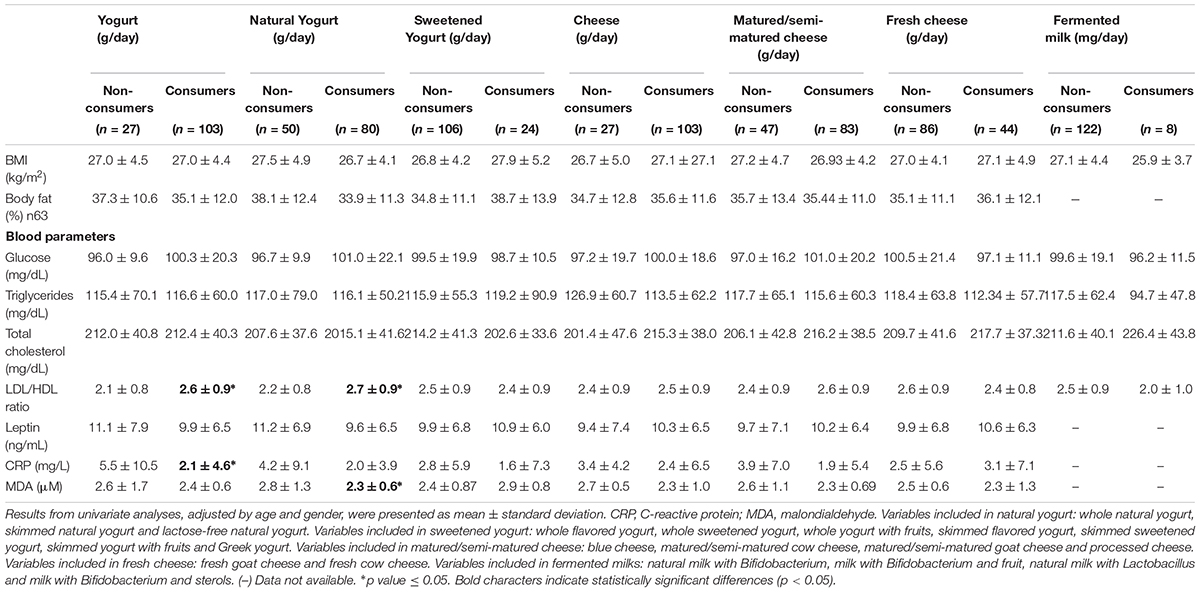
Table 3. Differences in anthropometric parameters and mean concentrations of serum health related biomarkers according to the intake of the different types of fermented dairy foods consumed by the sample.
The present study is a pioneer report analyzing the relationship between the intake of fermented dairy foods within the regular diet, the gut microbial profile and health related biomarkers, considering the subject’s global diet. Previous studies identified diets rich in fruits, vegetables or whole grains as critical modulators of the gut microorganisms, based on their content in fibers, phenolic compounds and prebiotics (Cuervo et al., 2014; Fernández-Navarro et al., 2018). However, the association between the different live microorganisms provided by the diet within the intestinal ecosystem offers a novel way to look into gut microbiota composition and its metabolic activity (Kok and Hutkins, 2018). In this regard, our results showed that, among the fermented dairy products assessed, yogurt was the product which showed higher ability to modulate the fecal microbiota. Interestingly, while the consumption of natural yogurt was directly associated with Akkermansia levels, the sweetened yogurt was inversely related with Bacteroides counts. The consumption of yogurt has been correlated with a good quality diet and some studies pointed out differences among yogurt types (Gómez-Gallego et al., 2018). A Danish cohort study suggested that consumption of whole-fat yogurt instead of low-fat products may be associated with a lower risk of type-2 diabetes (Ibsen et al., 2017). In the present sample, unfortunately, the low consumption of skimmed yogurt (consumed by only 6 out of the 80 volunteers consuming natural yogurt) precluded a skimmed vs. whole-fat comparison, however, it is worth mentioning that we have observed differences among the yogurt types assessed (natural vs. sweetened) with regards to the microbiota profile. These results underline the need for a full subcategorization of yogurt types in intervention and epidemiological studies, since different types may differ in their effects on health.
Given the descriptive nature of our study, we are not able to elucidate the mechanism of action explaining the observed associations. In spite of the lack of information about the modulation of intestinal Akkermansia in humans, recent research in mice treated with antibiotics has reported an increase in this bacterial group after the administration of a probiotic mix of Lactobacillus (Shi et al., 2018). Therefore, it may be plausible that the intake of such microorganisms, present in yogurt, might play a role in this association (Hill et al., 2014; Rezac et al., 2018). At this point, it should be mentioned that since labels of products do not provide information about the viable microorganisms present, we cannot know the exact amount and specific strains consumed by the study sample. According to the CODEX regulation (CODEX STAN 243-2003), yogurt must include a minimum bacterial counts of 107 cfu per gram from the symbiotic cultures of Streptococcus thermophilus and Lactobacillus bulgaricus. This, according to the intake data obtained, would correspond with intakes between 5 × 108 and 109 bacterial cells/day of each of these microorganisms. Nevertheless, although as shown in this study these levels can be easily reached within the context of a normal diet, it is also true that in interventional studies higher levels have been often used (Meyer et al., 2007; Asemi et al., 2011).
Results from intervention studies, both in animals and humans, have shown that the increase in Akkermansia muciniphila is associated with lower adiposity and a better metabolic status, suggesting this microorganism could be a potential candidate for obesity control (Everard et al., 2013; Dao et al., 2016; Rodríguez-Carrio et al., 2017). In the current study, we found that natural yogurt consumers presented not only higher intestinal Akkermansia levels with respect to non-consumers, but also a “healthier metabolic profile” based on lower inflammation and serum lipid peroxidation, measured through serum CRP and MDA. These immune variables have been reported to be moderately reduced in intervention studies with probiotic yogurt by other authors (Mohamadshahi et al., 2014; Burton et al., 2017). These findings are coherent with recent data from the Kuopio Ischaemic Heart Disease Risk Factor Study showing a cardiovascular protective effect in men consuming fermented dairy products (Koskinen et al., 2018), and with several epidemiological studies supporting a protective role of fermented dairy products against the chronic “low-grade” inflammation associated with the metabolic syndrome and related diseases (Baothman et al., 2016; Kim et al., 2017; Salas-Salvadó et al., 2017). Despite the values of LDL/HDL ratio in our sample were higher for yogurt consumers than for non-consumers, these are far from the established levels of atherogenic risk (>4.5). It is also important to underline that, in contrast to some of the previous studies (Asemi et al., 2013), age and gender have been introduced as covariates in the analyses performed in our study, and global diet has been determined.
Fermented dairy foods may present nutritional properties independent of the presence of microorganisms, as seems to occur with the sweetened yogurts. Although the lower levels of Bacteroides observed in the consumers of sweetened yogurt in our sample could be a priori surprisingly; this result is in consonance with previous reports indicating a reduction in the intestinal level of Bacteroides associated with the consumption of certain sweeteners such as sucralose (Uebanso et al., 2017). Therefore, it could be interesting to examine if the addition of additives (flavors, sweeteners, etc.) to traditionally considered healthy products, such as yogurt, could influence on the gut microbiota and, therefore, on the health status of the host.
No statistical differences were found in the levels of intestinal microbial groups as related to cheese consumption. However, cheese consumers showed higher fecal concentrations of the major SCFA. These compounds have been widely related with different metabolic effects, directly modulating host health through a range of tissue-specific mechanisms (den Besten et al., 2013; Rios-Covian et al., 2016; Uebanso et al., 2017). From a nutritional point of view, differences in the relationship with health may be expected depending on the types of cheese considered. Notwithstanding, we have not observed differences in our sample in health-related parameters according to cheese intake.
It is also important to be aware that this study contains some limitations. As mentioned before, although the FFQ has been carried out with a high grade of detail, it has not been possible to collect information on the specific microbial strains contained in the products. On the other hand, even though the multivariate models were adjusted by age and gender, we cannot rule out possible residual confounders often present in this sort of study. In spite of this, the present work has the strength of being conducted within the context of the habitual and global dietary pattern of the volunteers, and points out natural yogurt as a healthy product that, as previously suggested (Gómez-Gallego et al., 2018), should have a more visible role in dietary recommendations and guidelines. Our data suggests that fermented dairy products in general, and yogurt in particular, could be a key element affecting the relationship between diet and health by means of the modulation of gut microbial composition and functionality.
The datasets generated for this study are available on request to the corresponding author.
This study was carried out in accordance with the recommendations of the Bioethics Committee from CSIC and the Regional Bioethics Committee from the Principality of Asturias (Spain) with written informed consent from all subjects. All subjects gave written informed consent in accordance with the Declaration of Helsinki. The protocol was approved by the Bioethics Committee from CSIC and the Regional Bioethics Committee from the Principality of Asturias (Spain).
MG and SG had the primary responsibility in the study design and protocol development, and confirm that they had full access to the data in the study and final responsibility for the decision to submit for publication and drafted the manuscript. SG and TF-N were involved in data collection and contributed to the dietary and nutritional data analysis and interpretation. CR-G, SA, and NS conducted the microbial analysis and data processing and supervised the execution of the study and data analysis. All authors critically reviewed the manuscript and approved the final version submitted for publication.
This work was funded through the Grant GRUPIN14-043 “Microbiota Humana, Alimentación y Salud” funded by “Plan Regional de Investigación del Principado de Asturias,” Asturias, Spain and by the Alimerka Foundation. NS is the recipient of a postdoctoral contract awarded by the Fundación para la Investigación Biosanitaria de Asturias (FINBA). Public National and Regional grants received co-funding from European Union FEDER funds. SA is the recipient of a postdoctoral Maria Curie contract funded by the EU.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We show our greatest gratitude to all the volunteers participating in the study. This work was presented as an abstract at the X Workshop of the Spanish Society for Probiotics and Prebiotics.
Alvaro, E., Andrieux, C., Rochet, V., Rigottier-Gois, L., Lepercq, P., Sutren, M., et al. (2007). Composition and metabolism of the intestinal microbiota in consumers and non-consumers of yogurt. Br. J. Nutr. 97, 126–133. doi: 10.1017/S0007114507243065
Arboleya, S., Binetti, A., Salazar, N., Fernandez, N., Solis, G., Hernandez-Barranco, A., et al. (2012). Establishment and development of intestinal microbiota in preterm neonates. FEMS Microbiol. Ecol. 79, 63–72. doi: 10.1111/j.1574-6941.2011.01261.x
Arboleya, S., Sánchez, B., Solís, G., Fernández, N., Suárez, M., Hernández-Barranco, A. M., et al. (2016). Impact of prematiruty and perinatal antibiotics on the developing intestinal microbiota: a functional inference study. Int. J. Mol. Sci. 17:649. doi: 10.3390/ijms17050649
Asemi, Z., Jazayeri, S., Najafi, M., Samimi, M., Mofid, V., Shidfar, F., et al. (2011). Effects of daily consumption of probiotic yoghurt on inflammatory factors in pregnant women: a randomized controlled trial. Pakistan J. Biol. Sci. 14, 476–482. doi: 10.3923/pjbs.2011.476.482
Asemi, Z., Samimi, M., Tabassi, Z., Naghibi Rad, M., Rahimi Foroushani, A., Khorammian, H., et al. (2013). Effect of daily consumption of probiotic yoghurt on insulin resistance in pregnant women: a randomized controlled trial. Eur. J. Clin. Nutr. 67, 71–74. doi: 10.1038/ejcn.2012.189
Baothman, O. A., Zamzami, M. A., Taher, I., Abubaker, J., and Abu-Farha, M. (2016). The role of gut microbiota in the development of obesity and diabetes. Lipids Health Dis. 15:108. doi: 10.1186/s12944-016-0278-4
Burton, K. J., Rosikiewicz, M., Pimentel, G., Bütikofer, U., Von Ah, U., Voirol, M. J., et al. (2017). Probiotic yogurt and acidified milk similarly reduce postprandial inflammation and both alter the gut microbiota of healthy, young men. Br. J. Nutr. 117, 1312–1322. doi: 10.1017/S0007114517000885
Centro de Enseñanza Superior de Nutrición Humana y Dietética [CESNID] (2008). Tablas de Composición de Alimentos por Medidas Caseras de Consumo Habitual en España. Barcelona: Publicaciones y ediciones de la Universidad de Barcelona.
Chakrabarti, S., Jahandideh, F., and Wu, J. (2014). Food-derived bioactive peptides on inflammation and oxidative stress. Biomed. Res. Int. 2014:608979. doi: 10.1155/2014/608979
Chaves, S., Perdigon, G., De Moreno, A., and De Leblanc, A. (2011). Yoghurt consumption regulates the immune cells implicated in acute intestinal inflammation and prevents the recurrence of the inflammatory process in a mouse model. J. Food Prot. 74, 801–811. doi: 10.4315/0362-028X.JFP-10-375
Cuervo, A., Valdés, L., Salazar, N., De Los Reyes-Gavilán, C. G., Ruas-Madiedo, P., Gueimonde, M., et al. (2014). Pilot study of diet and microbiota: interactive associations of fibers and polyphenols with human intestinal bacteria. J. Agric. Food Chem. 62, 5330–5336. doi: 10.1021/jf501546a
Dao, M. C., Everard, A., Aron-Wisnewsky, J., Sokolovska, N., Prifti, E., Verger, E. O., et al. (2016). Akkermansia muciniphila and improved metabolic health during a dietary intervention in obesity: relationship with gut microbiome richness and ecology. Gut 65, 426–436. doi: 10.1136/gutjnl-2014-308778
den Besten, G., van Eunen, K., Groen, A. K., Venema, K., Reijngoud, D. J., and Bakker, B. M. (2013). The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 54, 2325–2340. doi: 10.1194/jlr.R036012
Duvallet, C., Gibbons, S. M., Gurry, T., Irizarry, R. A., and Alm, E. J. (2017). Meta-analysis of gut microbiome studies identifies disease-specific and shared responses. Nat. Commun. 8:1784. doi: 10.1038/s41467-017-01973-8
Eckburg, P. B., Lauber, C. L., Costello, E. K., Berg-Lyons, D., Gonzalez, A., Stombaugh, J., et al. (2005). Diversity of the human intestinal microbial flora. Science 308, 1635–1638. doi: 10.1126/science.1110591
Everard, A., Belzer, C., Geurts, L., Ouwerkerk, J. P., Druart, C., Bindels, L. B., et al. (2013). Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc. Natl. Acad. Sci. U.S.A. 110, 9066–9071. doi: 10.1073/pnas.1219451110
Fernández-Navarro, T., Salazar, N., Gutiérrez-Díaz, I., de los Reyes-Gavilán, C. G., Gueimonde, M., and González, S. (2017). Different intestinal microbial profile in over-weight and obese subjects consuming a diet with low content of fiber and antioxidants. Nutrients 9:E551. doi: 10.3390/nu9060551
Fernández-Navarro, T., Salazar, N., Gutiérrez-Díaz, I., Sánchez, B., Rúas-Madiedo, P., de los Reyes-Gavilán, C. G., et al. (2018). Bioactive compounds from regular diet and faecal microbial metabolites. Eur. J. Nutr. 57, 487–497. doi: 10.1007/s00394-016-1332-8
Gerard-Monnier, D., Erdelmeier, I., Regnard, K., Moze-Henry, N., Yadan, J. C., and Chaudiere, J. (1998). Reactions of 1-methyl-2-phenylindole with malondialdehyde and 4-hydroxyalkenals. Analytical applications to a colorimetric assay of lipid peroxidation. Chem. Res. Toxicol. 11, 1176–1183. doi: 10.1021/tx9701790
Gómez-Gallego, C., Gueimonde, M., and Salminen, S. (2018). The role of yogurt in food-based dietary guidelines. Nutr. Rev. 76, 29–39. doi: 10.1093/nutrit/nuy059
Guo, J., Astrup, A., Lovegrove, J. A., Gijsbers, L., Givens, D. I., and Soedamah-Muthu, S. S. (2017). Milk and dairy consumption and risk of cardiovascular diseases and all-cause mortality: dose–response meta-analysis of prospective cohort studies. Eur. J. Epidemiol. 32, 269–287. doi: 10.1007/s10654-017-0243-1
Hill, C., Guarner, F., Reid, G., Gibson, G. R., Merenstein, D. J., Pot, B., et al. (2014). Expert consensus document: the international scientific association for probiotics and prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 11, 506–514. doi: 10.1038/nrgastro.2014.66
Ibsen, D. B., Laursen, A. S. D., Lauritzen, L., Tjønneland, A., Overvad, K., and Jakobsen, M. U. (2017). Substitutions between dairy product subgroups and risk of type 2 diabetes: the Danish diet, cancer and health cohort. Br. J. Nutr. 118, 989–997. doi: 10.1017/S0007114517002896
Kato-Kataoka, A., Nishida, K., Takada, M., Kawai, M., Kikuchi-Hayakawa, H., Suda, K., et al. (2016). Fermented milk containing Lactobacillus casei strain Shirota preserves the diversity of the gut microbiota and relieves abdominal dysfunction in healthy medical students exposed to academic stress. Appl. Environ. Microbiol. 82, 3649–3658. doi: 10.1128/AEM.04134-15
Kim, D. H., Kim, H., Jeong, D., Kang, I. B., Chon, J. W., Kim, H. S., et al. (2017). Kefir alleviates obesity and hepatic steatosis in high-fat diet-fed mice by modulation of gut microbiota and mycobiota: targeted and untargeted community analysis with correlation of biomarkers. J. Nutr. Biochem. 44, 35–43. doi: 10.1016/j.jnutbio.2017.02.014
Kok, C. R., and Hutkins, R. (2018). Yogurt and other fermented foods as sources of health-promoting bacteria. Nutr. Rev. 76, 4–15. doi: 10.1093/nutrit/nuy056
Koskinen, T. T., Virtanen, H. E. K., Voutilainen, S., Tuomainen, T. P., Mursu, J., and Virtanen, J. K. (2018). Intake of fermented and non-fermented dairy products and risk of incident CHD: the kuopio ischaemic heart disease risk factor study. Br. J. Nutr. 120, 1288–1297. doi: 10.1017/S0007114518002830
Lisko, D., Johnston, G., and Johnston, C. (2017). Effects of dietary yogurt on the healthy human gastrointestinal (GI) microbiome. Microorganisms 5:E6. doi: 10.3390/microorganisms5010006
Marco, M. L., Heeney, D., Binda, S., Cifelli, C. J., Cotter, P. D., Foligné, B., et al. (2017). Health benefits of fermented foods: microbiota and beyond. Curr. Opin. Biotechnol. 44, 94–102. doi: 10.1016/j.copbio.2016.11.010
Marlett, J. A., and Cheung, T. F. (1997). Database and quick methods of assessing typical dietary fiber intakes using data for 228 commonly consumed foods. J. Am. Diet. Assoc. 97, 1139–1148.
Meyer, A. L., Elmadfa, I., Herbacek, I., and Micksche, M. (2007). Probiotic, as well as conventional yogurt, can enhance the stimulated production of proinflammatory cytokines. J. Hum. Nutr. Diet. 20, 590–598. doi: 10.1111/j.1365-277X.2007.00807.x
Mohamadshahi, M., Veissi, M., Haidari, F., Shahbazian, H., Kaydani, G. A., and Mohammadi, F. (2014). Effects of probiotic yogurt consumption on inflammatory biomarkers in patients with type 2 diabetes. BioImpacts 4, 83–88. doi: 10.5681/bi.2014.007
Panahi, S., Doyon, C. Y., Després, J. P., Pérusse, L., Vohl, M. C., Drapeau, V., et al. (2018). Yogurt consumption, body composition, and metabolic health in the Québec family study. Eur. J. Nutr. 57, 1591–1603. doi: 10.1007/s00394-017-1444-9
Rezac, S., Kok, C. R., Heermann, M., and Hutkins, R. (2018). Fermented foods as a dietary source of live organisms. Front. Microbiol. 9:1785. doi: 10.3389/fmicb.2018.01785
Rios-Covian, D., Ruas-Madiedo, P., Margolles, A., Gueimonde, M., de los Reyes-Gavilan, C. G., and Salazar, N. (2016). Intestinal short chain fatty acids and their link with diet and human health. Front. Microbiol. 7:185. doi: 10.3389/fmicb.2016.00185
Rodríguez-Carrio, J., Salazar, N., Margolles, A., González, S., Gueimonde, M., de los Reyes-Gavilán, C. G., et al. (2017). Free fatty acids profiles are related to gut microbiota signatures and short-chain fatty acids. Front. Immunol. 8:823. doi: 10.3389/fimmu.2017.00823
Salas-Salvadó, J., Guasch-Ferré, M., Díaz-López, A., and Babio, N. (2017). Yogurt and diabetes: overview of recent observational studies. J. Nutr. 147, 1452S–1461S. doi: 10.3945/jn.117.248229
Şanlier, N., Gökcen, B. B., and Sezgin, A. C. (2017). Health benefits of fermented foods. Crit. Rev. Food Sci. Nutr. 25, 1–22. doi: 10.1080/10408398.2017.1383355
Severyn, C. J., and Bhatt, A. S. (2018). With probiotics, resistance is not always futile. Cell Host Microbe 24, 334–336. doi: 10.1016/j.chom.2018.08.014
Shi, Y., Zhao, X., Zhao, J., Zhang, H., Zhai, Q., Narbad, A., et al. (2018). A mixture of Lactobacillus species isolated from traditional fermented foods promote recovery from antibiotic-induced intestinal disruption in mice. J. Appl. Microbiol. 124, 842–854. doi: 10.1111/jam.13687
Sivamaruthi, B. S., Kesika, P., and Chaiyasut, C. (2018). Impact of fermented foods on human cognitive function—A review of outcome of clinical trials. Sci. Pharm. 86:E22. doi: 10.3390/scipharm86020022
Theander, O., and Westerlund, E. A. (1986). Studies on dietary fiber. 3. improved procedures for analysis of dietary fiber. J. Agric. Food Chem. 34, 330–336. doi: 10.1021/jf00068a045
Uebanso, T., Ohnishi, A., Kitayama, R., Yoshimoto, A., Nakahashi, M., Shimohata, T., et al. (2017). Effects of low-dose non-caloric sweetener consumption on gut microbiota in mice. Nutrients 9:E560. doi: 10.3390/nu9060560
Uyeno, Y., Sekiguchi, Y., and Kamagata, Y. (2008). Impact of consumption of probiotic lactobacilli-containing yogurt on microbial composition in human feces. Int. J. Food Microbiol. 122, 16–22. doi: 10.1016/j.ijfoodmicro.2007.11.042
Keywords: fermented foods, yogurt, inflammation, microbiota, oxidative stress
Citation: González S, Fernández-Navarro T, Arboleya S, de los Reyes-Gavilán CG, Salazar N and Gueimonde M (2019) Fermented Dairy Foods: Impact on Intestinal Microbiota and Health-Linked Biomarkers. Front. Microbiol. 10:1046. doi: 10.3389/fmicb.2019.01046
Received: 25 February 2019; Accepted: 25 April 2019;
Published: 24 May 2019.
Edited by:
Jyoti Prakash Tamang, Sikkim University, IndiaReviewed by:
Patricia Burns, CONICET Instituto de Lactología Industrial (INLAIN), ArgentinaCopyright © 2019 González, Fernández-Navarro, Arboleya, de los Reyes-Gavilán, Salazar and Gueimonde. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: M. Gueimonde, bWd1ZWltb25kZUBpcGxhLmNzaWMuZXM=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.