 Mariana Domínguez1
Mariana Domínguez1 Claudio D. Miranda2,3*
Claudio D. Miranda2,3* Oliver Fuentes1,4Mery de la Fuente4,5
Oliver Fuentes1,4Mery de la Fuente4,5 Félix A. Godoy6
Félix A. Godoy6 Helia Bello-Toledo1
Helia Bello-Toledo1 Gerardo González-Rocha1
Gerardo González-Rocha1- 1Laboratorio de Investigación en Agentes Antibacterianos, Departamento de Microbiología, Universidad de Concepción, Concepción, Chile
- 2Laboratorio de Patobiología Acuática, Departamento de Acuicultura, Universidad Católica del Norte, Coquimbo, Chile
- 3Centro AquaPacífico, Coquimbo, Chile
- 4Facultad de Medicina Veterinaria, Universidad San Sebastián, Concepción, Chile
- 5Departamento de Ciencias Biológicas, Facultad de Ciencias de la Vida, Universidad Andres Bello, Talcahuano, Chile
- 6Centro i∼mar, Universidad de Los Lagos, Puerto Montt, Chile
Salmon farming industry in Chile currently uses a significant quantity of antimicrobials to control bacterial pathologies. The main aims of this study were to investigate the presence of transferable sulfonamide- and trimethoprim-resistance genes, sul and dfr, and their association with integrons among bacteria associated to Chilean salmon farming. For this purpose, 91 Gram-negative strains resistant to sulfisoxazole and/or trimethoprim recovered from various sources of seven Chilean salmonid farms and mainly identified as belonging to the Pseudomonas genus (81.0%) were studied. Patterns of antimicrobial resistance of strains showed a high incidence of resistance to florfenicol (98.9%), erythromycin (95.6%), furazolidone (90.1%) and amoxicillin (98.0%), whereas strains exhibited minimum inhibitory concentrations (MIC90) values of sulfisoxazole and trimethoprim of >4,096 and >2,048 μg mL−1, respectively. Strains were studied for their carriage of these genes by polymerase chain reaction, using specific primers, and 28 strains (30.8%) were found to carry at least one type of sul gene, mainly associated to a class 1 integron (17 strains), and identified by 16S rRNA gene sequencing as mainly belonging to the Pseudomonas genus (21 strains). Of these, 22 strains carried the sul1 gene, 3 strains carried the sul2 gene, and 3 strains carried both the sul1 and sul2 genes. Among these, 19 strains also carried the class 1 integron-integrase gene intI1, whereas the dfrA1, dfrA12 and dfrA14 genes were detected, mostly not inserted in the class 1 integron. Otherwise, the sul3 and intI2 genes were not found. In addition, the capability to transfer by conjugation these resistance determinants was evaluated in 22 selected strains, and sul and dfr genes were successfully transferred by 10 assayed strains, mainly mediated by a 10 kb plasmid, with a frequency of transfer of 1.4 × 10−5 to 8.4 × 10−3 transconjugant per recipient cell, and exhibiting a co-transference of resistance to florfenicol and oxytetracycline, currently the most used in Chilean salmon industry, suggesting an antibacterial co-selection phenomenon. This is the first report of the characterization and transferability of integrons as well as sul and dfr genes among bacteria associated to Chilean salmon farms, evidencing a relevant role of this environment as a reservoir of these genes.
Introduction
Intensive fish farming favors the development of infectious diseases and consequently the use of antimicrobial agents for the treatment of bacterial infections has increased. One of the consequences of this practice is the selection of resistant bacteria carrying a wide variety of antimicrobial resistance encoding genes (Miranda, 2012). Although Chile is the second largest salmon producer in the world, only a few studies concerning this problem have been developed, mostly studying the occurrence of genetic determinants encoding for resistance to oxytetracycline and florfenicol (Miranda et al., 2003; Fernández-Alarcón et al., 2010; Buschmann et al., 2012), the most used drugs in Chilean salmon farms, accounting for the 99.3% of the antimicrobials used in this industry during 2016 (SERNAPESCA, 2017).
The salmon farming industry in Chile used a high amount of antibiotics, and among these, potentiated sulfonamides were included. Evidence exists that these antimicrobial agents are accumulated in the environments during treatments, leading to a change in the native bacterial composition. Thus, the selection of antibiotic resistant bacteria carrying different resistance genes is expected. In the case of those inserted in plasmids and/or integrons, they can be disseminated to susceptible bacteria in different environments (Cabello et al., 2016; Watts et al., 2017).
Despite the fact that sulfonamides are not longer authorized for used in Chilean salmon farms they were extensively used prior to introduction of drug use regulations. According to the SERNAPESCA reports (SERNAPESCA, 2011), 2009 was the last year when potentiated sulfonamides were used in the Chilean salmon industry, mainly to treat the pathogens Flavobacterium psychrophilum and Streptococcus phocae, but using only very low quantities, corresponding to the 0.01% (2008 and 2009) and 0.02% (2007) of the total amount of drug used (SERNAPESCA, 2011; Miranda et al., 2018). Furthermore, despite sulfonamides were banned for their use in Chilean salmon industry many years ago, the presence of bacteria carrying genes encoding for their resistance could be still prevalent in Chilean salmon farm environments considering the feasibility of their co-selection when other antibacterial agents such as where florfenicol or oxytetracycline are administered in fish farms.
The most frequent mechanism of bacterial resistance to sulfonamides is the acquisition of the dihydropteroate synthase enzyme encoded by the genes sul1, sul2 and sul3 (Grape et al., 2003; Perreten and Boerlin, 2003; Bean et al., 2009; Vinué et al., 2010). While the sul1 gene is mainly associated with class 1 integrons, the sul2 gene has been detected on various plasmids but not associated with integrons (Bean et al., 2009; Vinué et al., 2010), whereas the sul3 gene has been linked to class 1 integrons lacking their 3′ CS region (Bean et al., 2009). Integrons are composed of two very conserved DNA regions, located at their ends, which are known as 5′ CS and 3′ CS (5′ and 3′ conserved segments), and between both zones one or more antibiotic resistance genes could be inserted (Ploy et al., 2000; Cambray et al., 2010; Deng et al., 2015). Acquisition and dissemination of these genes located within the integron structure results in an increase in antimicrobial resistance. Three classes of integron structure have been described (Cambray et al., 2010; Deng et al., 2015). Class 1 integrons are of principal importance in clinical isolates. The 5′-CS of class 1 integrons includes an intI1 gene, which encodes for a specific recombinase. This gene contains the att1 recombination site, required for specifically integrating gene cassettes. The 3′-CS region contains several open reading frames (ORFs). These include qacEΔ1, which confers resistance to quaternary ammonium compounds, often associated with antiseptics, along with a sul1 gene expressing resistance to sulfonamides (Paulsen et al., 1993; Partridge et al., 2009). Resistance to trimethoprim is mainly mediated by a dihydrofolate reductase enzyme encoded by the dfr genes, which are usually associated with class 1 and class 2 integrons or plasmids (Yu et al., 2004; Ho et al., 2009).
This association of sul and dfr genes with mobile genetic elements such as plasmids and integrons, is highly relevant for the increase in the emergence, evolution, and dissemination of sulfonamide resistance in aquatic environments. It has been extensively reported that a high number of strains resistant to florfenicol and oxytetracycline usually exhibit simultaneous resistance to sulfonamides (Miranda and Zemelman, 2002; Miranda and Rojas, 2007; Fernández-Alarcón et al., 2010), currently the most used drugs in the Chilean salmonid industry (SERNAPESCA, 2017).
In a recent study some sulfonamide-resistance genes were studied among several isolates recovered close to a Chilean salmon farm. A low number of isolates carrying a sul1 gene and positive for the intI1 integrase gene were detected, but their ability to be transferred was not detected (Shah et al., 2014). Considering the role of environmental bacteria as reservoirs of antimicrobial resistance genes and the importance of salmon farming in Chile, in this study, we aimed to investigate the occurrence of sul and dfr genes, their relationship with integrons, as well as assess their capacity to be transferred by conjugation among sulfonamide- and/or trimethoprim-resistant strains recovered from various sources of Chilean salmon farms.
Materials and Methods
Bacterials Strains
The study included 91 Gram-negative bacilli strains resistant to sulfisoxazole and/or trimethoprim recovered from a collection of bacteria obtained from seven salmon farms located in the South of Chile where the Chilean salmonid industry is concentrated (Figure 1). Fourteen strains were recovered from the land-based farms (facilities for salmonid growth until smoltification carried out in tanks located in land-based hatcheries), F1 and F2 (7 strains from each farm), and 77 strains were recovered from the lake-based farms (salmonids raised in fish cages located in freshwater lakes), C1 (6 strains), C2 (7 strains), C3 (32 strains), C4 (14 strains), and C5 (18 strains). Land-based salmonid farms F1 and F2 were located near Lake Llanquihue and Quellón Bay, respectively, whereas lake-based farms were located in the Lake Puyehue (C1), Lake Rupanco (C2 and C3) and Lake Llanquihue (C4 and C5) (Figure 1).
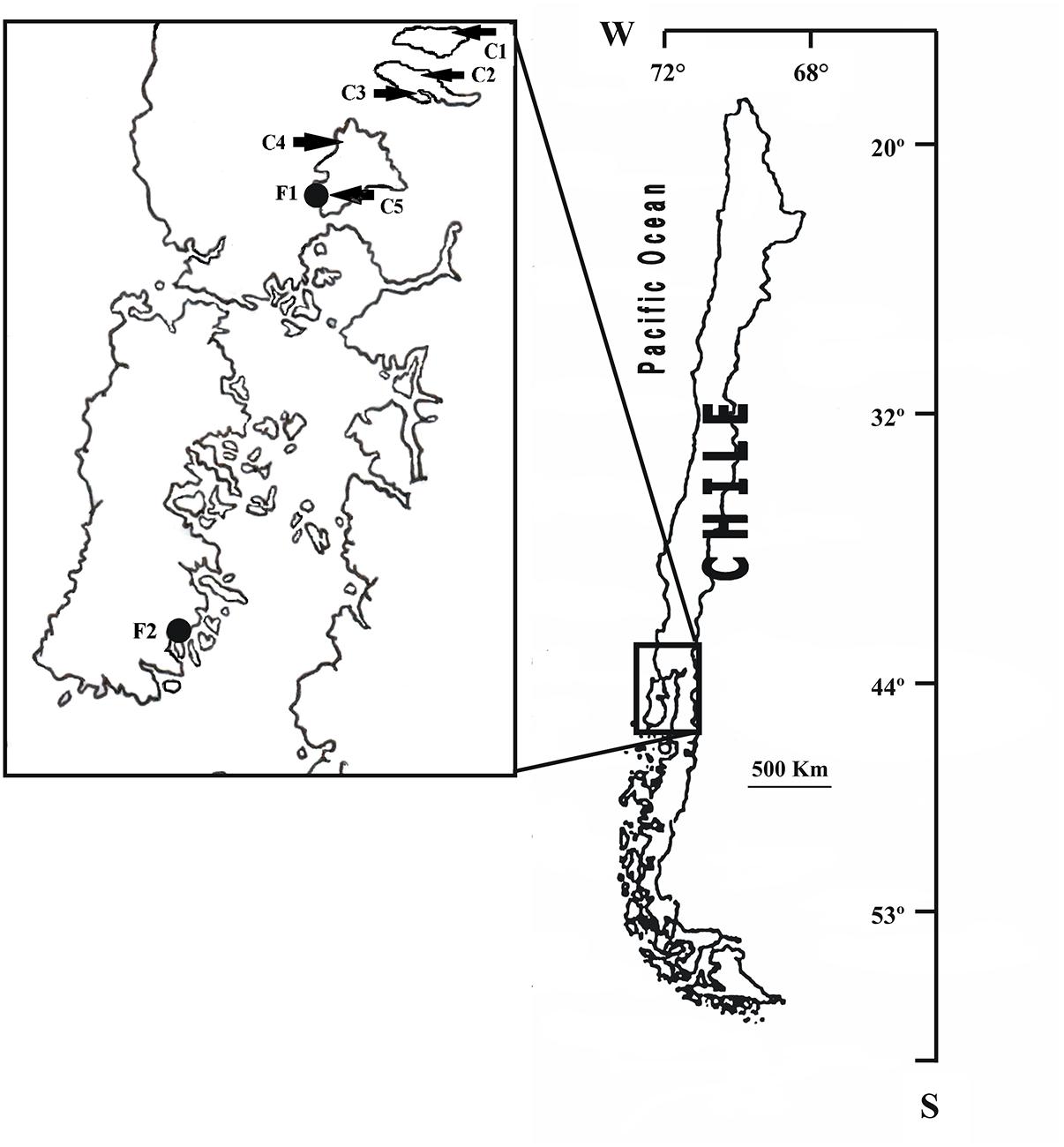
Figure 1. Geographic location of Chilean salmon farms, where sulfonamide- and/or trimethoprim-resistant strains were recovered, including land-based farms (F1–F2) and lake-based farms (C1–C5).
Strains from land-based culture centers were isolated from various sources, including un-medicated fish food pellet, salmonid fingerling and water samples from fish farm effluent and fish rearing tanks. Strains recovered from lake-based salmonid cultures were isolated from samples of mucus and intestinal content of reared fish, surface water samples from salmon cages and un-impacted control sites, as well as samples of sediments beneath salmonid cages and control sites. Samples were collected and processed as previously described (Miranda and Zemelman, 2002; Miranda and Rojas, 2007). Purified strains were stored at −85°C in CryoBank (Mast Diagnostic) vials until use.
Bacterial Identification
Phenotypical characteristics, Gram stain, oxidase production and oxidative/fermentative utilization of glucose were determined for the 91 studied strains. The Gram reaction of strains was determined using the Gram stain (Barrow and Feltham, 1993), oxidase production was determined by the method described by Barrow and Feltham (1993), and the oxidative/fermentative (O/F) test was carried out according to Hugh and Leifson (1953). Strains were further characterized by using the GN Microplate system (Biolog, Inc., Hayward, CA, United States) according to Miranda and Rojas (2007) and a number of strains (20%) were reexamined to check reproducibility of the assay. Bacterial identification was performed by using the Microlog System 4.2 identification software (Biolog, Inc.).
The identity of thirty sulfisoxazole- and/or trimethoprim-resistant strains positive for any of the studied genes was confirmed by bacterial 16S rRNA gene sequence analysis. For DNA extraction, the bacterial strains were cultured in Tryptic Soy Agar (TSA, Difco labs) for 2 days, and total genomic DNA was extracted from each strain using the PureLink genomic DNA kit (Invitrogen). The extracted DNA was amplified by polymerase chain reaction (PCR) using the eubacterial 16S rDNA primers 27F (5′-AGAGTTTGATCCTGGCTCAG-3′) and 1492R (5′-GGTTACCTTGTTACGACTT-3′) described by Lane (1991) and PCR products were purified using the Wizard SV Gel kit and PCR Clean-up System (Promega). Sequencing of amplicons was performed by Macrogen using the ABI PRISM 373 DNA Sequencer (Applied Biosystems) with ABI PRISM cycle sequencing kit (Macrogen, South Korea). Primers 27F (5′-AGAGTTTGATCMTGGCTCAG-3′), 907R (5′-CCGTCAATTCCTTTRAGTTT-3′) and 1492R (5′-TACGGYTACCTTGTTACGACTT-3′) were used for sequencing. Sequence data was aligned and compared with available standard sequences of bacterial lineage in the GenBank database (NCBI) using Basic Local Alignment Search Tool. The partial sequences of the 16S rDNA gene belonging to each strain were deposited in the GenBank database under accession numbers KX279647 to KX279675, and MH424518.
Minimum Inhibitory Concentrations (MICs)
Minimum inhibitory concentrations (MICs) of sulfisoxazole and trimethoprim against all strains were determined by an agar dilution method, as recommended by the Clinical and Laboratory Standards Institute (CLSI) guideline M07-A9 (CLSI, 2012a). A serial twofold dilution pattern of the antibiotic was added into Mueller–Hinton agar (Difco) so as to obtain final concentrations ranging from 0.5 to 2,048 μg mL−1. Bacterial suspensions were prepared in sterile 0.85% saline and triplicate plates were inoculated using a Steers replicator apparatus (Steers et al., 1959), delivering approximately 104 colony-forming units per spot, and incubated for 48 h at 22°C. The first and last agar plates did not contain any antibiotic in order to detect possible contamination of the strains or antibiotic carryover. MIC was defined as the lowest concentration of the antibacterial agent producing absence of growth at least in two of the three plates after 48 h. Reference strain Escherichia coli ATCC 25922, recommended by CLSI (CLSI, 2012a), was used as quality control organism for verification of MIC ranges on Mueller–Hinton agar plates.
Antimicrobial Resistance Patterns
Sulfisoxazole- and/or trimethoprim-resistant strains were tested for their susceptibility to 15 antimicrobials by an agar disk diffusion method as described in the CLSI guideline VET03-A (CLSI, 2006), using Müeller–Hinton agar (Difco). The antibacterial susceptibility patterns of resistant strains were performed using disks containing the antibacterial agents: amoxicillin (AML, 25 μg), cefotaxime (CTX, 30 μg), streptomycin (S, 10 μg), kanamycin (K, 30 μg), gentamicin (CN, 10 μg), erythromycin (E, 15 μg), chloramphenicol (CM, 30 μg), florfenicol (FFC, 30 μg), oxytetracycline (OT, 30 μg), oxolinic acid (OA, 2 μg), flumequine (UB, 30 μg), enrofloxacin (ENR, 5 μg), furazolidone (FR, 100 μg), sulfisoxazole (SFX, 300 μg), and trimethoprim (TMP, 5 μg). All disks were obtained from Oxoid, Ltd. (Basingstoke, Hampshire, England). Bacterial strains were suspended in sterile 0.85% saline to a turbidity to match a McFarland No. 2 standard (bioMerieux S.A.), diluted 1:20, and streaked on the used media. Plates were incubated for 24–48 h at 22°C and E. coli ATCC 25922 was used as the control strain, as recommended by the CLSI (CLSI, 2012b). Characterization of strains as resistant was stated according to standards suggested by CLSI (CLSI, 2008) or by Miranda and Rojas (2007). A number of strains (20%) were re-examined to check reproducibility of the assay.
Detection of sul and dfr Genes
The detection of genes encoding for resistance to sulfonamide (sul1, sul2, and sul3) and trimethoprim (dfrA1, dfrA5, dfrA6, dfrA7, dfrA8, dfrA9, dfrA10, dfrA12, dfrA13, dfrA14, dfrA17, dfrA20, dfrB1, dfrB2, dfrB3) was made using PCR using specific primers previously described (Grape et al., 2003; Perreten and Boerlin, 2003) or designed in this study (Tables 1, 2). Colonies from an overnight culture on Mueller Hinton agar at 22°C were resuspended in 200 μL of 5% Chelex (BioRad) and 25 μL of 20 mg mL−1 proteinase K (Invitrogen) were added. The suspension was incubated for 45 min at 56°C followed by 8 min at 100°C, then centrifuged at 14,000 rpm and the supernatant was used as template. The PCR mixtures (25 μL of total volume) consisted of 2.5 μL of Taq reaction buffer 10× (Invitrogen), 0.75 μL of MgCl2 (50 mM, Invitrogen), 0.25 μL dNTPs (25 mM each, Invitrogen), 0.125 μL of each primer (25 pmoL μL−1, Invitrogen), 0.2 μL Taq polymerase (5 U μL−1, Invitrogen), and 2 μL of template DNA.

Table 1. Primers used to detect sul genes.
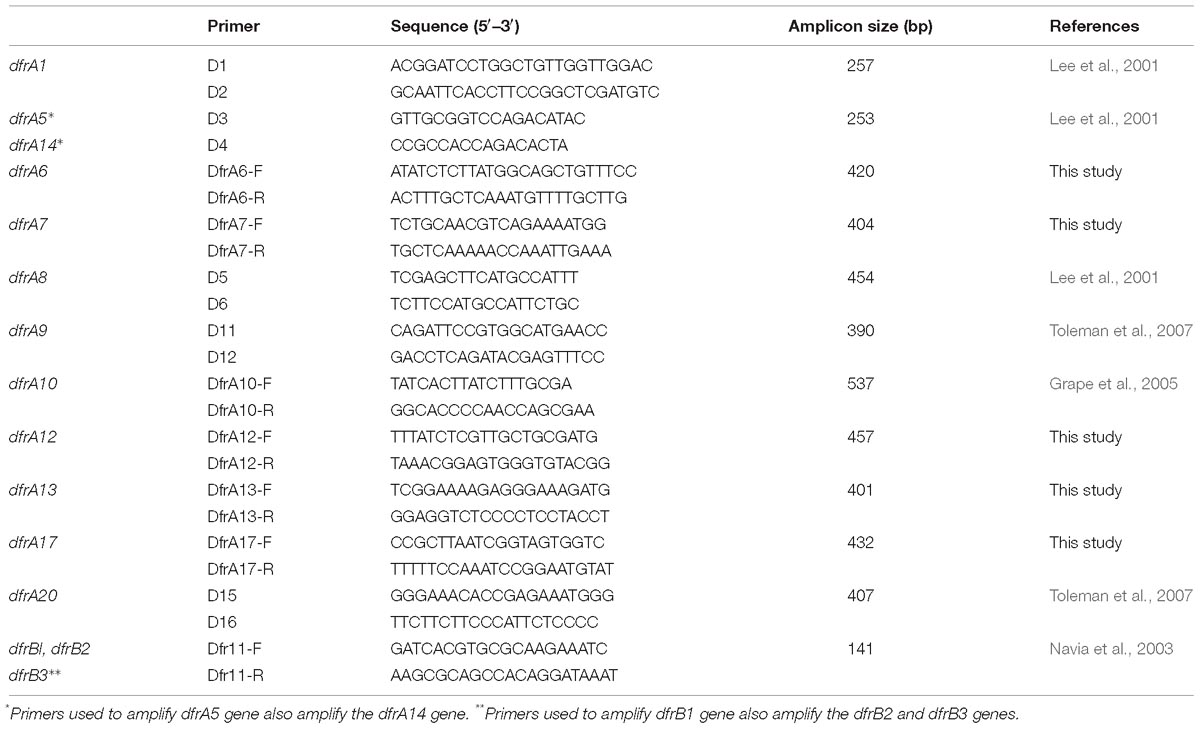
Table 2. Primers used to detect dfr genes.
Polymerase chain reaction products were detected by electrophoresis in 1.5% agarose gels and visualized by ultraviolet illumination after staining with ethidium bromide (0.5 mg L−1).
Detection and Characterization of Class 1 and Class 2 Integrons
The occurrence of class 1 and class 2 integrons was detected using PCR, amplifying intragenic fragments using specific primers for each gene (Table 3), according to previously described methodologies (Rosser and Young, 1999; Mazel et al., 2000). PCR products for intI1 (class 1 integron) gene were confirmed using restriction enzymes, considering that SphI enzyme produces two fragments (393 and 499 bp), whereas no strains positive for intI2 were found. Additionally, in strains carrying class 1 integrons their whole integron gene cassettes were amplified using a combination of pair of primers which align in the zone of the integrase gene and the possible gene inserted as cassette in reverse. Similarly, combinations of primers which amplify the possible gene inserted as cassette and primers which align in the 3′ CS end of the integron were used. The obtained amplification products were purified by using Wizard PCR Preps (Promega) and sequenced using the Macrogen, United States sequencing service.

Table 3. Primers used for PCR detection of integrons and associated gene cassette regions.
Isolation of Plasmid DNA
Bacterial strains (30 strains) harboring a studied gene (sul1, sul2, dfrA1, dfrA12, dfrA14, and intI1) were screened for their plasmid content. The plasmid isolation was made using a geneJET Plasmid miniprep kit (Thermo Scientific) according to the manufacturer’s instructions. The plasmid DNA obtained was run on 1.5% agarose gel electrophoresis for plasmids less than 20 kb and 0.8% agarose gel for plasmid greater than 20 kb. Gels were stained with GelRedTM (Biotium) and viewed by UV transillumination. The size was estimated by comparing with known plasmid weight standards and standard molecular weight markers.
Mating Experiments
Twenty-two strains were selected according their resistance pattern, MIC levels and carriage of sul and/or dfr genes and integrons, to evaluate their ability to transfer these determinants by conjugation in liquid medium according to the methodology described by Adams et al. (1998). A mutant of E. coli K-12 resistant to nalidixic acid, rifampicin and sodium azide was used as the recipient. Transconjugants were selected on MacConkey agar plates containing sodium azide (300 μg mL−1) and sulfisoxazole (512 μg mL−1) and tested for the presence of the antimicrobial resistance genes by PCR as previously described.
Results
Bacterial Identification
Out of the 91 Gram-negative bacilli strains studied, 85 (93.4%) were glucose non-fermenting bacteria, and only 6 (18.6%) were glucose fermenters. Most of the strains belonged to the Pseudomonas genus (81 strains), with a high frequency of P. fluorescens and P. putida (21 and 18 strains, respectively), whereas strains belonging to other non-fermenting genre included Acinetobacter johnsonii, Brevundimonas vesicularis and Sphingobacterium multivorum and were isolated at low frequency (Table 4). The glucose-fermenting strains were all recovered from lake-based farms and were identified as Citrobacter gillenii, C. freundii, Kluyvera intermedia, Comamonas sp., Hafnia sp., and Raoultella terrigena (Table 4). When the 30 sulfonamide- and/or trimethoprim-resistant strains positive for any of the assayed genes were identified by amplifying their 16S rRNA genes, a high predominance of representatives of the Pseudomonas genus (22 strains), mainly belonging to the P. fluorescens (6), P. putida (6), and P. baetica (3) species was observed, whereas most of the enteric strains included in the study, identified as C. gillenii, C. freundii, Hafnia sp., Comamonas sp. and K. intermedia, harbored at least one of the detected genes (Table 4).
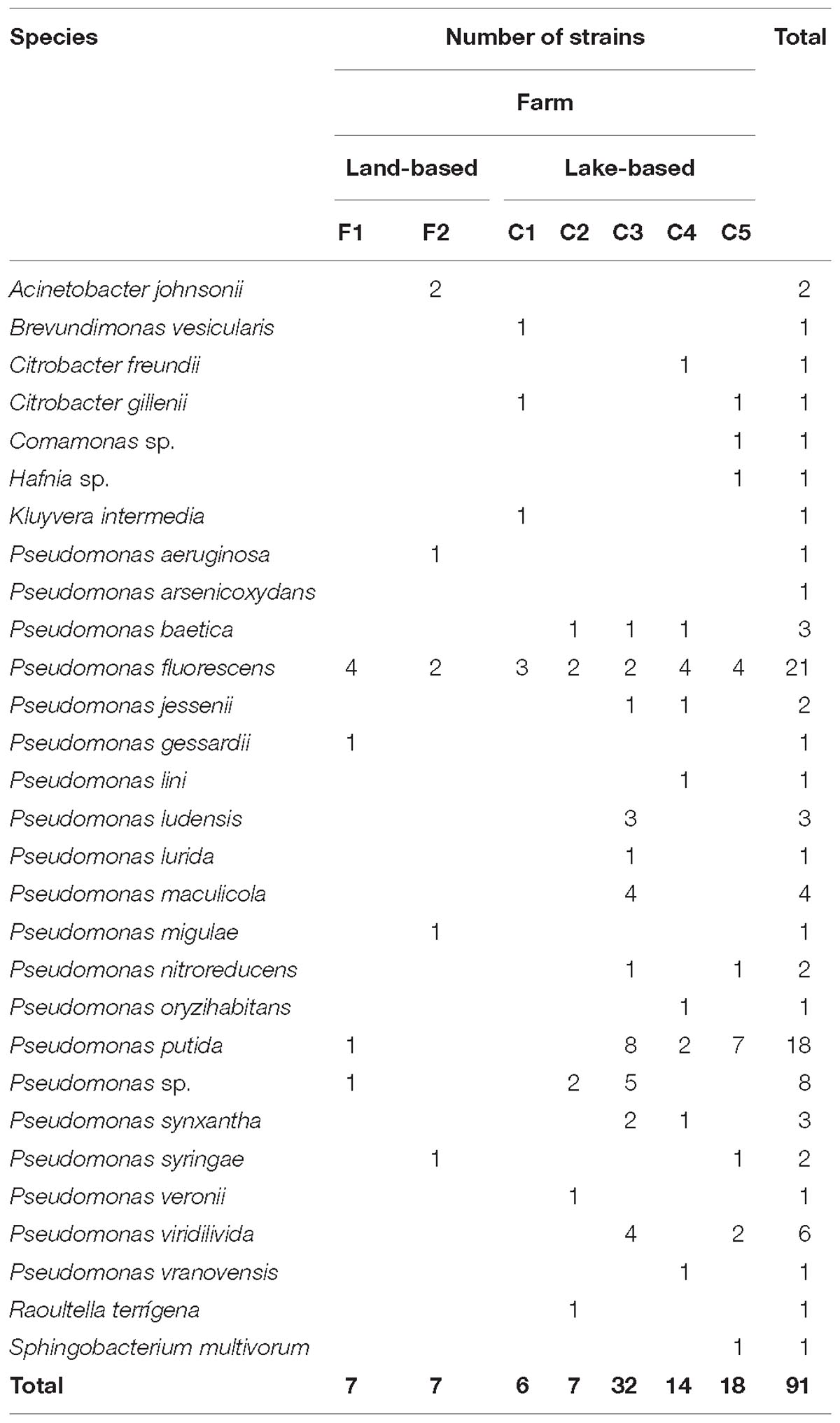
Table 4. Identification of resistant strains recovered from Chilean salmonid farms.
Minimum Inhibitory Concentrations (MICs)
Resistant strains exhibited high MIC values of sulfisoxazole and trimethoprim, not observing remarkable differences among strains from different farms and sources (Table 5). Sulfisoxazole MIC values of strains varied between 64 and ≥4,016 μg mL−1, with MIC50 and MIC90 values of 2,046 and ≥ 4,096 μg mL−1, respectively, whereas trimethoprim MIC values ranged from 4 to ≥2,048 μg mL−1, with MIC50 and MIC90 values of 2,046 and ≥2,046 μg mL−1, respectively (Table 5). Thus, strains showed high levels of resistance, considering that breakpoint values for categorizing resistance for these antibacterials are 16 μg mL−1 for trimethoprim and 512 μg mL−1 for sulfisoxazole (CLSI, 2015). Reference strain E. coli ATCC 25922, used for quality control exhibited MIC values of sulfisoxazole and trimethoprim of 8 and 0.5 μg mL−1, respectively, which agrees with the values recommended by CLSI (2015).
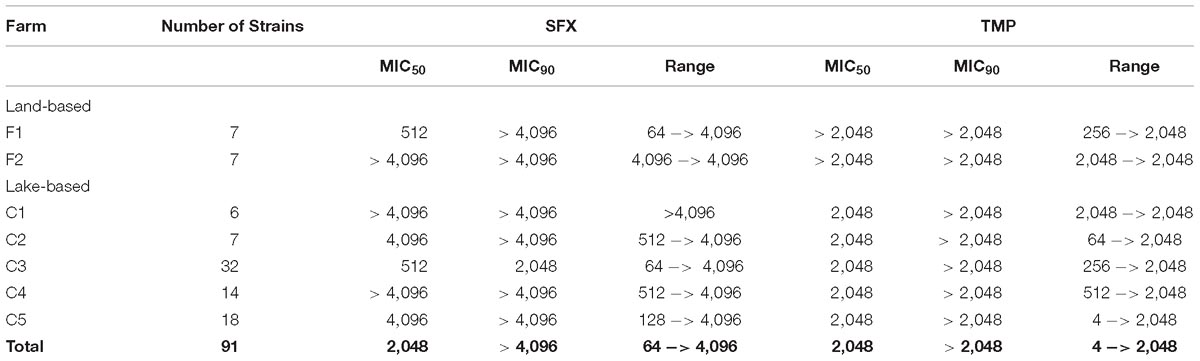
Table 5. Sulfixosazole (SFX) and trimethoprim (TMP) minimum inhibitory concentrations (MIC, in μg mL−1) of resistant strains recovered from Chilean salmonid farms.
Antimicrobial Resistance Patterns
Strains used in the study were resistant to sulfisoxazole (85.7%) and/or trimethoprim (98.9%). A high percentage of strains exhibited resistance to chloramphenicol and florfenicol (98.9%), erythromycin (95.6%) and furazolidone (90.1%). In addition, a high incidence of resistance to amoxicillin (78.0%), as well as an intermediate resistance to oxolinic acid (45.1%), oxytetracycline (34.1%), gentamicin (34.1%), and cefotaxime (26.4%) was observed in the strains (Figure 2). Otherwise, a low incidence of resistance to enrofloxacin (3.3%), kanamycin (12.1%), and flumequine (19.8%), was detected among the studied strains (Figure 2). None of the strains was susceptible to less than four antimicrobial agents. Moreover, 51.9% and 26.7% of the strains showed simultaneous resistance to 7 and 8 antibacterial agents, respectively. Among the 13 strains exhibiting susceptibility to sulfisoxazole, were those recovered from farms F1 (2), C3 (9), and C5 (2), whereas the strain susceptible to trimethoprim was recovered from farm C5.
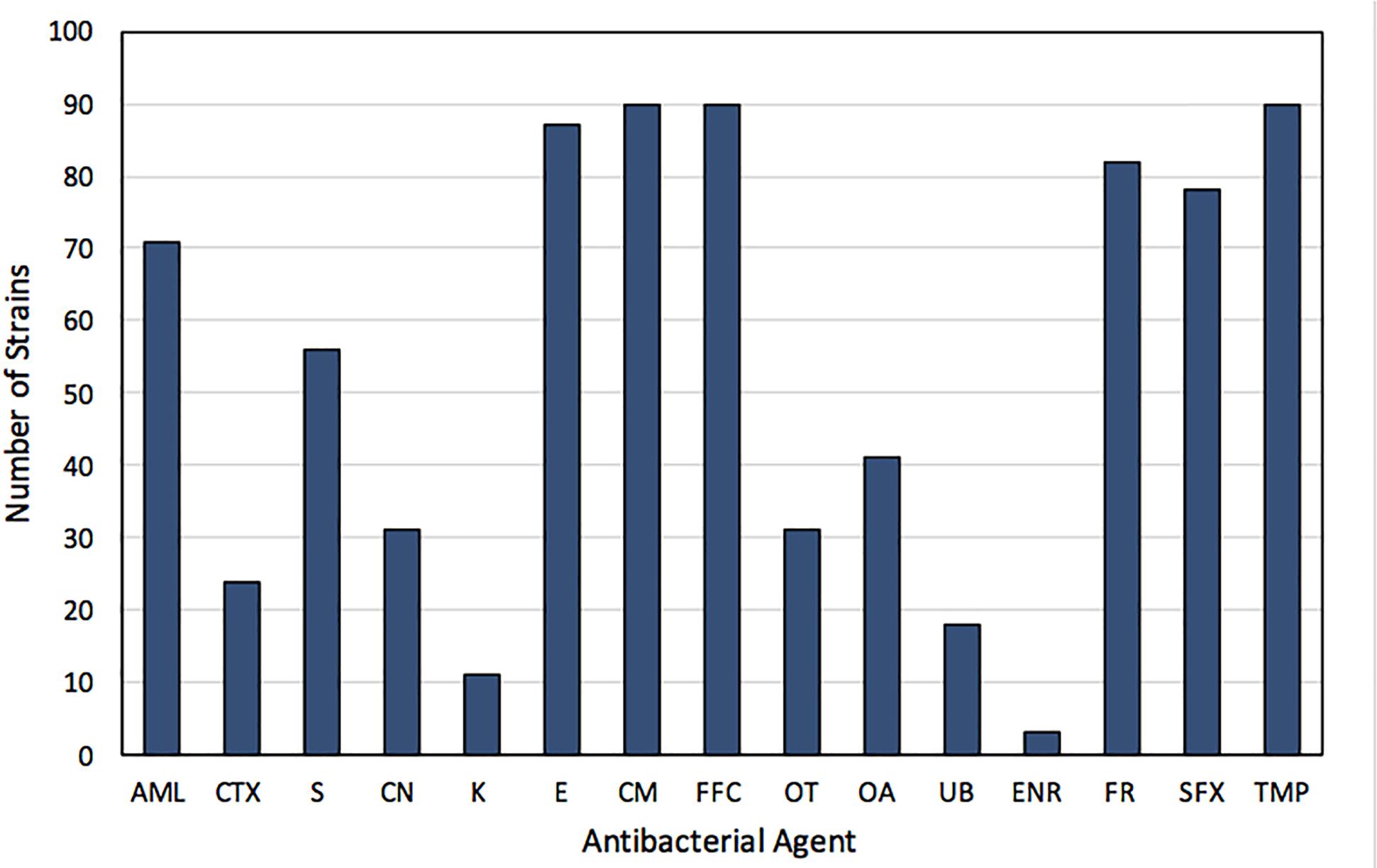
Figure 2. Frequency of resistance to antibacterials of 91 sulfonamide- and/or trimethoprim-resistant strains isolated from Chilean salmon farms. AML, amoxicillin; CTX, cefotaxime; S, streptomycin; CN, gentamicin; K, kanamycin; E, erythromycin; CM, chloramphenicol; FFC, florfenicol; OT, oxytetracycline; OA, oxolinic acid; UB, flumequine; ENR, enrofloxacin; FR, furazolidone; SFX, sulfisoxazole; TMP, trimethoprim.
Detection of sul and dfr Genes
An important number of the sulfonamide-resistant strains (28 out of 78 strains; 35.9%) carried a sul gene, and among them 18 strains carried a class 1 integron (Figure 3). Among these, 22 harbored the sul1 gene, 3 carried the sul2 gene and 3 strains carried both genes (Figure 3). The sul1 gene was detected in strains recovered from all 7 salmon farms, distributed in 8 out of 12 (66.7%) and 17 out of 66 (25.8%) from land-based and lake-based farms, respectively, whereas sul2 was found only in strains from four farms (Figure 3). On the other hand, none of the studied strains carried the sul3 gene. The majority of sul-positive strains were found among the strains exhibiting the highest MICs of sulfisoxazole; considering that 20/22 strains carrying sul1 as well as 4 strains harboring sul2 had MIC values of ≥4,098 μg mL−1 (Supplementary Table S1). It must be noted that 18 strains recovered from surface water (3 strains) and sediments (15 strains) from non-aquaculture unpolluted sites but exhibiting resistance to sulfisoxazole and/or trimethoprim were included in the study, but none of them were positive for any of the assayed genes.
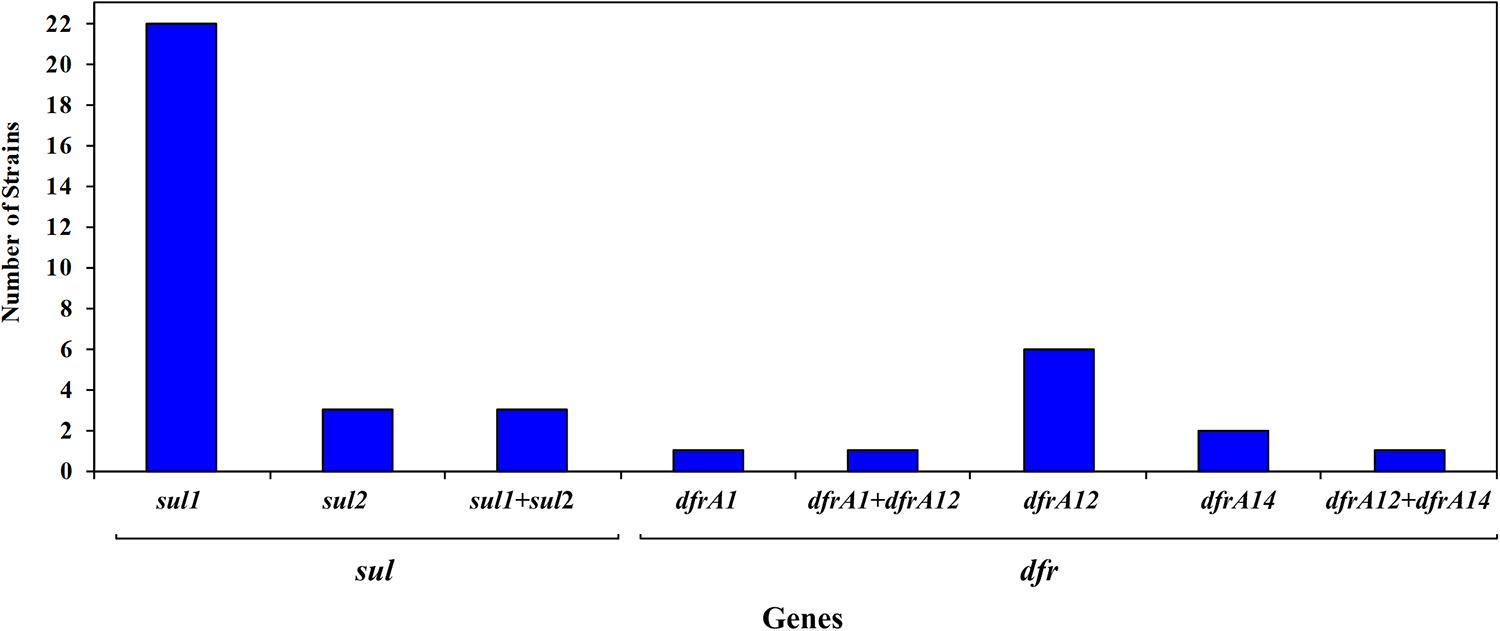
Figure 3. Frequency of sul and dfr genes among 91 sulfonamide- and/or trimethoprim-resistant strains isolated from Chilean salmon farms.
When the presence of dfrA and dfrB genes was investigated, only the dfrA1, dfrA12, and dfrA14 genes were detected, distributed in 12 of the 90 trimethoprim-resistant strains, with a predominance of dfrA12 gene (8/12 strains). The distribution of dfr genes was similar in strains isolated from land-based (13.6%) and lake-based (11.7%) farms. In addition, it must be noted that two strains belonging to the Citrobacter genus, simultaneously carried two different genes encoding for resistance to trimethoprim (C. gillenii FP75 carried dfrA1 and dfrA12 genes and strain C. freundii FB98 carried dfrA12 and dfrA14 genes) (Figure 3).
Detection and Characterization of Integrons
In this study, 21 out of 91 strains (23.1%) harbored a class 1 integron, whereas intl2 integrase was not detected among the studied strains. This genetic structure was present in bacterial strains from land-based (6/14; 42.9%) and from lake-based (19/77; 24.7%) farms. Among strains carrying a class 1 integron, 17 strains showed a 3′ CS end with the qacEΔ1 gene adjacent to the sul1 gene, but 4 of them had a modified gene qacEΔ1, whereas in another four strains positive for the integrase intI1 it was not possible to amplify the 3′ CS end of their integrons. As observed with strains carrying the sul and dfr genes, the integrons were concentrated in strains with high levels of resistance to trimethoprim and/or sulfisoxazole (Table 6). An important number of strains (17 strains) carried a sul gene inserted in integrons (Table 6).
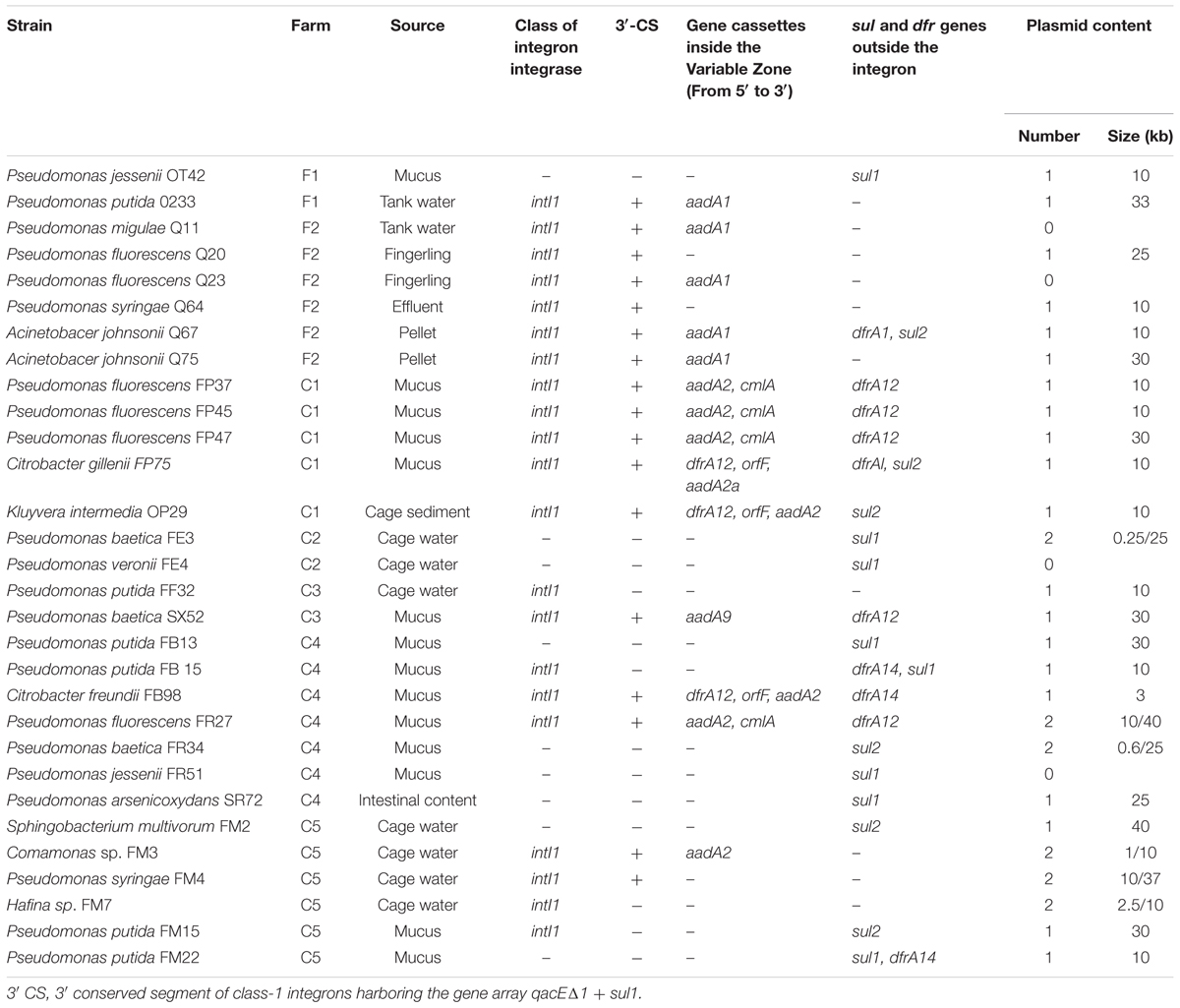
Table 6. Identification, integron structure, location of sul and dfr genes and plasmid content among sulfonamide- and/or trimethoprim-resistant strains.
Fourteen variable zones were found among the detected class 1 integrons, carrying 1 to 3 gene cassettes. Five genes encoding for resistance to trimethoprim (dfrA12), aminoglycosides (aadA1, aadA2, and aadA9), and chloramphenicol (cmlA) were identified as part of the integron structures, being found either alone or in combination with other gene cassettes (Table 6). The variable regions with one cassette harbored the genes aadA1 (five strains), aadA2 (one isolate) and aadA9 (one isolate). The strains with two cassettes inserted in its variable region carried aadA2-cmlA (four strains). Finally, three strains were found to carry three gene cassettes in their variable zones containing the genes dfrA12-orf-aadA2 (two strains) and dfrA12-orf-aadA2a (one strain). Otherwise, in three strains the variable zone of their integrons could not be determined, despite using various primer combinations (Table 6).
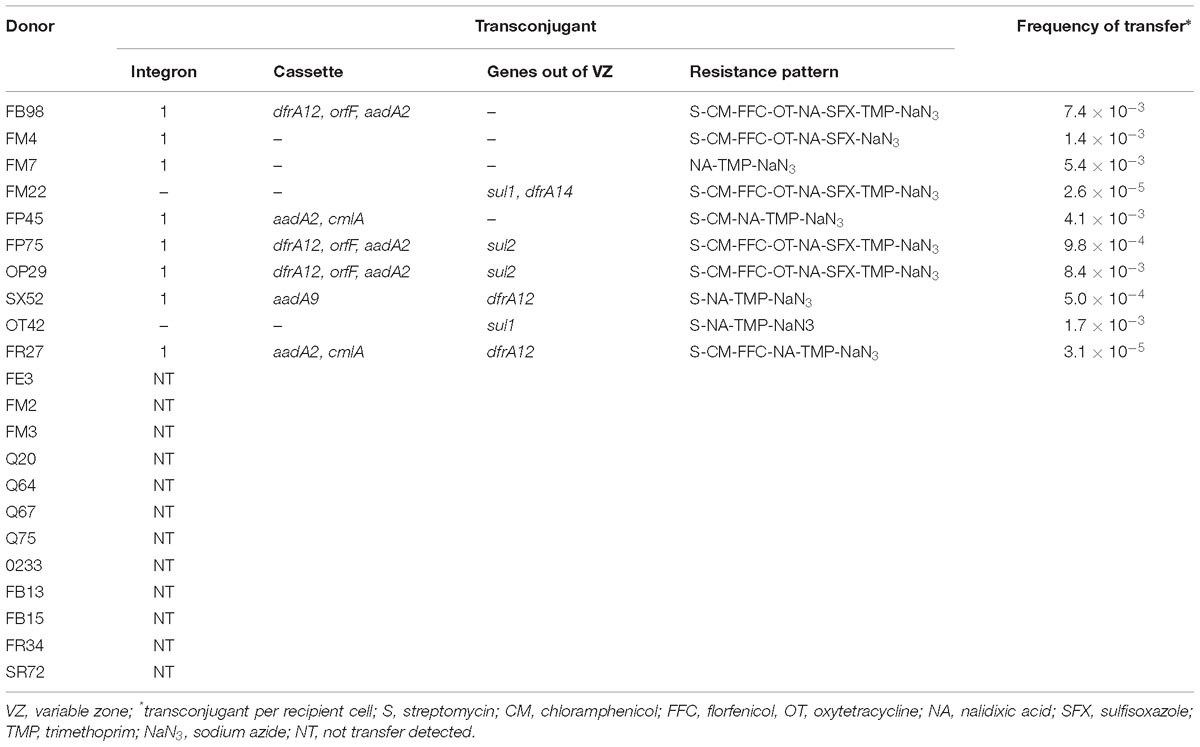
Table 7. Transfer of sul and dfr genes and integrons from resistant strains to Escherichia coli K-12.
It must be noted that various strains harboring a class 1 integron carried genes encoding for resistance to sulfonamides and trimethoprim not inserted in the integron structure. For the dfr genes two situations were seen, the presence of dfr genes in strains with integrons, but not associated with these structures, and the presence of more than one dfr gene per strain (associated and not associated with integron). In the case of sul genes, these were found mainly associated with class 1 integrons, with sul1 as part of the integron, whereas sul2 was not present as a gene cassette, but rather associated with other structures (Table 6).
The dfrA12 gene encoding for the enzyme dehidrofolate reductase XII, carried by the FP75, FB98 and OP29 strains, was analyzed using the 5′ CS/dfrA12 R, dfrA12 F/3′ CS and dfrA12F/sul1R primers, and the cassette was found to be located adjacent to the integrase gene. In addition, when amplicons were sequenced, two cassettes were detected, corresponding to the orf and aadA2a genes (FP75 strain), and orf and aadA2 genes (FB98 and OP29 strains). These genes were adjacent to the qacEΔ1 gene in the 3′ CS end, presenting a 100% similarity to the dfrA12, orfF, and aadA2a genes included in the GenBank database under the Accession No. DQ322593.
Plasmid Content
Plasmid DNA was detected in 25 out of 30 resistant strains examined, and from these, six strains contained two plasmid bands. The number and size of plasmid bands found are shown in Table 6. The most commonly found plasmid band had a molecular weight of 10 kb (13 strains) followed by plasmids with a range from 25 to 40 kb (12 strains), with a predominance of plasmids of 30 kb (four strains). Among the four strains carrying two plasmids, all of them carried a 10 kb plasmid and were identified as Hafnia sp., Comamonas sp., P. syringae, and P. fluorescens (Table 6).
Transfer of Resistance Genes
Among the 22 strains assayed for conjugation experiments, in 10 strains it was demonstrated that sul and dfr genes, as well as the integrons were transferable. A high frequency of transfer of resistance to sulfonamides was observed in the positive strains, ranging from 10−3 to 10−5 (transconjugant cells per recipient cells). The majority of these strains (6 out of 10) showed a frequency of transfer of 10−3, whereas only two strains showed a frequency of 10−5 (Table 7). In addition, strains FP75, OP29, OT42, FM7, FM22, and FB98 were able to co-transfer resistance to oxytetracycline, chloramphenicol and florfenicol, whereas strain FR27 co-transferred resistance to chloramphenicol and florfenicol. It was observed that all transconjugant strains acquired similar levels of resistance to antimicrobials to those exhibited by the donor strains.
Six strains used as donors and carrying a class 1 integron were able to transfer this structure as well as their associated gene cassettes (Table 7). Furthermore, sul and dfrA genes not associated with integrons were transferred, corresponding to the sul1 gene carried by OT42 and FM22 strains, sul2 gene carried by the FP75 and OP29 strains, and dfrA12 carried by the SX52 and FR27 strains (Table 7). On the contrary, other dfr genes not associated with integrons such as the dfrA12, dfrA14, and dfrA1 genes carried by the FP45, FB98 and FP75 strains, respectively were not transferred by conjugation (Table 7). The simultaneous transfer of sul1 and sul2 genes from strains C. gillenii FP75 and K. intermedia OP29, which carried both genes suggests the location of these genes on the same plasmid, considering that both strains carried only one plasmid. Otherwise, an important number of the assayed strains, both carrying and not carrying a class 1 integron (7 and 5 strains, respectively) were not able to transfer by conjugation their resistance to sulfonamides and/or trimethoprim, as well as the associated resistance encoding genes (Table 7).
Discussion
In previous studies investigating resistant strains from Chilean salmon farms with different histories of antimicrobial usage the occurrence of resistance to trimethoprim/sulfametoxazole, accompanied with resistance to florfenicol and oxytetracycline, as well as the carriage of floR and tet genes respectively has been reported (Miranda and Zemelman, 2002; Miranda et al., 2003; Miranda and Rojas, 2007; Fernández-Alarcón et al., 2010). This was regardless of whether sulfonamides had been used or not, thus suggesting that sulfonamide-resistance may be selected and maintained by the use of other antimicrobials, such as florfenicol or oxytetracycline, the most used antimicrobials in Chilean salmon farms (SERNAPESCA, 2017; Miranda et al., 2018). Furthermore, Sköld (2001) concluded that susceptibility to sulfonamides and trimethoprim will not return after suspending their use, mainly due to the reported high variety of mechanisms of bacterial resistance to these antibacterials, helping to explain the finding of the persistence of these resistances even without selective pressure.
The high incidence of multiresistance exhibited by the studied strains is according with previous studies of antibiotic resistant bacteria recovered from Chilean salmon farms and sites near salmon farms (Miranda and Zemelman, 2002; Miranda and Rojas, 2007; Shah et al., 2014), in which most of strains showed simultaneous resistance to 4–10 antibacterial agents. Otherwise, MIC values of sulfisoxazole and trimethoprim of strains were remarkable high, but no previous MIC values of strains recovered from Chilean salmon farms are available. The high MIC values of these antibacterials suggest the concurrence of sul and dfr genes.
Among the sul-carrying strains, 19 of these strains carried the sul1 gene associated with the 3′ conserved end, proper of a class 1 integron, as was reported by Rosser and Young (1999), whereas the occurrence of nine strains carrying the sul1 gene not associated with an integron structure, is in accordance with that reported by Grape et al. (2005) and Infante et al. (2005) in clinical strains. On the contrary the detected sul2 genes were not integron inserted, in agreement with Agersø and Petersen (2007), who studied Acinetobacter strains resistant to sulfametoxazole recovered from fish farms in Thailand. Thus, the positive transfer of sul2 genes is consistent with a location on another mobile element, such as small size plasmids as was previously reported (Rådström and Swedberg, 1988; Enne et al., 2004; Bean et al., 2005; Agersø and Petersen, 2007). As sul genes were not detected in 64 sulfonamide-resistant strains, other resistance mechanisms must be involved, most probably multidrug efflux pumps, considering the multiresistant phenotypes of studied strains as well as the high predominance of non-fermenters of glucose, which usually exhibit this mechanism of resistance (Maseda et al., 2000).
In this study, the dfrA1, dfrA12, and dfrA14 genes were found, with dfrA12 the most frequent. Lee et al. (2001) reported these genes encoding for enzymes conferring the highest levels of resistance to trimethoprim, in accordance with this study, because strains carrying dfr genes exhibited the highest levels of resistance to trimethoprim. Otherwise, the feasibility of carrying other dfr genes cannot be discarded, especially in those strains negative for any of the assayed dfr genes and exhibiting high levels of resistance to trimethoprim. It was observed that the majority of assayed strains from F1 and F2 land-based farms (5/6) was not capable of transfer by conjugation their plasmids and sul or dfr genes, whereas an important number of strains recovered from C1 and C5 lake-based farms (6/8) was able to transfer their integrons as well as their sul and dfr genes, being included or not in a class 1 integron. Furthermore, considering that transfer of sulfonamide resistance was usually accompanied with a transfer of resistance to the antimicrobials most intensively used in Chilean salmon farms, such as florfenicol and oxytetracycline, these antibacterials might promote the selection and spread of antibiotic resistance genes encoding for sulfonamide and trimethoprim resistance via co-selection.
In a previous study, Shah et al. (2014) studied a total of 124 strains recovered from aquaculture impacted sediments located near Chilean salmon farms, founding 62 and 58 strains exhibiting resistance to sulfamethizole and trimethoprim, respectively, with 12 strains carrying the sul1 gene and the integrase-encoding int1 gene. Otherwise, dfrA1, dfrA5, and dfrA12 genes were detected in 17, 14, and 22 strains, respectively. These authors studied a group of quinolone resistant strains recovered from site near a Chilean salmon farm, and detected three strains identified as Dietzia sp., Rhodococcus sp., and Arcobacter sp., to be carrying class 1 integrons with the dfrA12-aadA2-qacEΔ1-sul1 array (Tomova et al., 2018), similar to those shown by the FP75, FB98 and OP29 strains, but lacking the orfF gene. Thus, this study reports for the first time the carriage of the gene dfrA14, encoding for resistance to trimethoprim as well as the genes aadA1 and aadA9, encoding for resistance to aminoglycosides, among bacteria associated to Chilean salmonid farms, whereas all the observed gene arrays within class 1 integron are described for the first time among bacteria associated to Chilean salmon farming. According to the sequencing analysis, all cassette arrays found in the study have been previously described, mainly associated to enteric bacteria (Chang et al., 2007; Domingues et al., 2015), whereas the gene cassettes identified in the study were not novel.
Most of previous studies dealing with the occurrence of integrons in Pseudomonas considered clinical strains belonging to the Pseudomonas aeruginosa species. It has been reported that class 1 integrons have been found to be widespread in P. aeruginosa isolated from environmental and clinical settings from various countries (Fonseca et al., 2005; Gu et al., 2007; Chen et al., 2009; Ruiz-Martínez et al., 2011; Martínez et al., 2012; Kiddee et al., 2013; Kor et al., 2013; Hsiao et al., 2014; Adesoji et al., 2015; Khosravi et al., 2017), in according with the results observed in this study, in which the majority of the strains carrying a class 1 integron belonged to the Pseudomonas genus.
After amplifying and DNA sequencing of whole class 1 and integron gene cassettes, a total of six different types of gene cassette arrays were identified, with a predominance of aadA1 (5 strains) and aadA2-cmlA (4 strains) (Table 6). It is interesting to note the high incidence of aadA genes (14 strains), encoding for the enzyme aminoglycoside adenyltransferase that confers resistance to aminoglycosides, inserted in the variable zone of a class 1 integron, considering that no aminoglycosides have been used in Chilean salmon farming, thus suggesting that these integrons have a human clinic origin. In other studies, most of the class 1 integrons identified in E. coli strains carried the dfr12-orfF-aadA2 cassette array (Kang et al., 2005; Chang et al., 2007), as was observed in this study for the enteric bacteria.
Otherwise, while the occurrence of int2 gene in Pseudomonas aeruginosa has been previously reported (Xu et al., 2009; Goudarzi et al., 2015), none of the Pseudomonas strains included in this study were found to carry the int2 gene, which was in concordant to other reports of P. aeruginosa from clinical settings (Rossolini and Mantengoli, 2005; Sun et al., 2014; Khosravi et al., 2017). It must be noted that class 2 integrons are not usually detected in Pseudomonas strains from environmental sources, being most commonly associated with members of the family Enterobacteriaceae, such as E. coli and Salmonella enterica (Ahmed et al., 2005; Antunes et al., 2006; Cocchi et al., 2007; Essen-Zandbergen et al., 2007; Laroche et al., 2009; Ozgumus et al., 2009).
Of the class 1 integron-containing strains, it was demonstrated the occurrence of an empty integron in seven strains, and possible causes could be the lack of the 3′ conserved segment or insufficient homology to the 3′ conserved segment primer to produce a product such as the observed in strains Hafnia sp. FM7, Pseudomonas putida FM15, P. putida FB15 and P. putida FF32, but strains P. fluorescens Q20, P. syringae Q64 and P. syringae FM4 possessed a 3′ conserved segment (Table 6). Thus, most probably, the lack of integrated genes cassettes may be a consequence of the excision of previously integrated cassettes from the integron when antibiotic selective pressure is diluted in the environment, as was concluded by Ruiz-Martínez et al. (2011). Furthermore, these empty structures could confer to these strains the capacity to adapt rapidly to the aquaculture environment, where antibiotics are intensively used, conferring to them a selective advantage by means of the acquisition of new antibiotic resistance genes. Otherwise, it was suggested that these empty structures could be used as indicators of the absence of a sustained antimicrobial pressure (Ruiz-Martínez et al., 2011).
Otherwise, the absence of the conserved end 3′-CS in four strains carrying class 1 integrons could be a consequence of the activity of insertion sequences which could produce non-functional genes (Laroche et al., 2009; Dawes et al., 2010). Furthermore, it must be noted that integrons of environmental bacteria commonly exhibit deletions in their structures, contrary to the observed in the integrons harbored by clinical strains. In this trend, complex integrons associated with the insertion sequence ISCR1 have been reported (Quiroga et al., 2007; Song et al., 2010), which frequently have non-cassette type resistance genes and a truncated version of 3′-CS region followed by an ISCR1 element (Toleman et al., 2006a,b). Other studies demonstrated that rearrangements produced by the insertion sequence IS26 in the integrons may change class 1 integron structure causing the loss of parts of the 3′-CS (Labbate et al., 2008; Doublet et al., 2009; Juan et al., 2010).
The association of integrons inserted in conjugative plasmids and antimicrobial resistance was confirmed by transfer experiments for 10 out of 22 assayed strains. Therefore, integrons appear to have an essential role in facilitating the dissemination of the resistance genes and contributing to the creation of multidrug resistant phenotypes. Furthermore, previous studies demonstrated that copy numbers between the sul1 and intI1 genes were significantly correlated suggesting that class 1 integrons may play a role in the prevalence and propagation of sul1 in various aquatic environments, including farm sediments and marine coastal areas (Muziasari et al., 2014; Na et al., 2014; Lin et al., 2015). It is well-known that intI1 genes are the most abundant gene capture and transmission system in clinical and environmental strains (Partridge et al., 2009; Shah et al., 2012; Domingues et al., 2015), and their abundance and structures are greatly influenced by anthropogenic contamination (Wright et al., 2008). Considering that class 1 integrons are common among multi-drug resistant bacteria inhabiting natural environments associated with human activities, where a strong selection pressure is imposed by the use of antimicrobials, integrons have been proposed as markers for the identification of multi-drug resistant strains, as well as for anthropogenic pollution (Gillings et al., 2008; Gillings et al., 2015).
Conclusion
The present results demonstrated that Chilean salmon farms play an important role as reservoirs of sulfonamide- and trimethoprim-resistant bacteria. The prevalence of conjugative plasmids and integrons among sul-carrying bacteria suggests these bacteria, mainly belonging to the Pseudomonas genus may contribute to high spread of bacterial resistance to sulfonamides and other antibacterials in environments associated with Chilean salmon farms. This is the first study reporting the occurrence of transferable sul and dfr genes and integrons among the antimicrobial resistant bacteria associated with Chilean salmonid farms and the results suggest that the increasing prevalence of antibiotic resistant bacteria in Chilean salmon farming resistance is partly attributable to the acquisition, dissemination and stable maintenance of a class 1 integron. Thus, a continuous surveillance of these resistance elements even in absence of antibacterial therapy is urgently required to evaluate the potential role of fish farming environments as reservoirs and source of elements composing the mobilome, such as conjugative plasmids and integrons, increasing the risk of their dissemination to human pathogens through horizontal transfer.
Author Contributions
MD planned the experiments, analyzed the data, and participated in manuscript drafting. CM isolated and provided the studied strains, studied antimicrobial susceptibility patterns of resistant bacteria, analyzed the data, wrote the manuscript and is the corresponding author and primary contact during the manuscript submission, review, and publication process. OF performed the MIC assays, detected the genes encoding for sulfonamide and trimethoprim resistance, as well as integrase and variable region of detected integrons, and also carried out their DNA sequence analysis. MF performed conjugation assays and participated in manuscript drafting. FG identified the studied strains by sequencing their 16S rRNA genes, carried out DNA sequence analysis and determined the plasmid content of strains. HB-T and GG-R participated in manuscript drafting, revisions and interpretation of data. All authors have made intellectual contribution to the work, and approved it for publication.
Funding
This study was supported by the Science and Technology National Council (CONICYT) of Chile by the Project (Grant No. 1040924).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Dr. Matthew Lee, Centro i∼mar, Universidad De Los Lagos, Puerto Montt, Chile, for editing the manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2019.00748/full#supplementary-material
References
Adams, C., Austin, B., Meaden, P., and McIntosh, D. (1998). Molecular characterization of plasmid-mediated oxytetracycline resistance in Aeromonas salmonicida. Appl. Environ. Microbiol. 64, 4194–4201.
Adesoji, A. T., Ogunjobi, A. A., and Olatoye, I. O. (2015). Molecular characterization of selected multidrug resistant Pseudomonas from water distribution systems in southwestern Nigeria. Ann.Clin. Microbiol. Antimicrob. 14:39. doi: 10.1186/s12941-015-0102-4
Agersø, Y., and Petersen, A. (2007). The tetracycline resistance determinant Tet 39 and the sulphonamide resistance gene sulII are common among resistant Acinetobacter spp. isolated from integrated fish farms in Thailand. J. Antimicrob. Chemother. 59, 23–27. doi: 10.1093/jac/dkl419
Ahmed, A. M., Nakano, H., and Shimamoto, T. (2005). Molecular characterization of integrons in non-typhoid Salmonella serovars isolated in Japan: description of an unusual class 2 integron. J. Antimicrob. Chemother. 55, 371–374. doi: 10.1093/jac/dkh534
Antunes, P., Machado, J., and Peixe, L. (2006). Characterization of antimicrobial resistance and class 1 and 2 integrons in Salmonella enterica isolates from different sources in Portugal. J. Antimicrob. Chemother. 58, 297–304. doi: 10.1093/jac/dkl242
Barrow, G. I., and Feltham, R. K. A. (1993). Cowan and Steel’s Manual for the Identification of Medical Bacteria, 3rd Edn. Cambridge: Cambridge University Press, 331. doi: 10.1017/CBO9780511527104
Bean, D. C., Livermore, D. M., and Hall, L. M. C. (2009). Plasmid imparting sulfonamide resistance in Escherichia coli: implications for persistence. Antimicrob. Agents Chemother. 53, 1088–1093. doi: 10.1128/AAC.00800-08
Bean, D. C., Livermore, D. M., Papa, I., and Hall, L. M. C. (2005). Resistance among Escherichia coli to sulphonamides and other antimicrobial now little used in man. J. Antimicrob. Chemother. 56, 962–964. doi: 10.1093/jac/dki332
Buschmann, A. H., Tomova, A., López, A., Maldonado, M. A., Henríquez, L. A., Ivanova, L., et al. (2012). Salmon aquaculture and antimicrobial resistance in the marine environment. PloS One 7:e42724. doi: 10.1371/journal.pone.0042724
Cabello, F. C., Godfrey, H. P., Buschmann, A. H., and Dölz, H. J. (2016). Aquaculture as yet another environmental gateway to the development and globalisation of antimicrobial resistance. Lancet Infect. Dis. 16, 127–133. doi: 10.1016/S1473-3099(16)00100-6
Cambray, G., Guerout, A.-M., and Mazel, D. (2010). Integrons. Annu. Rev. Genet. 44, 141–166. doi: 10.1146/annurev-genet-102209-163504
Chang, L.-L., Chang, T.-M., and Chang, C.-Y. (2007). Variable gene cassette patterns of class 1 integron-associated drug resistant Escherichia coli in Taiwan. Kaohsiung. J. Med. Sci. 23, 273–280. doi: 10.1016/S1607-551X(09)70409-7
Chen, J., Su, Z., Liu, Y., Wang, S., Dai, X., Li, Y., et al. (2009). Identification and characterization of class 1 integrons among Pseudomonas aeruginosa isolates from patients in Zhenjiang, China. Int. J. Infect. Dis. 13, 717–721. doi: 10.1016/j.ijid.2008.11.014
CLSI (2006). Methods for Antimicrobial Disk Susceptibility Testing of Bacteria Isolated from Aquatic Animals; Approved Guideline. CLSI document M42-A. Wayne, PA: Clinical and Laboratory Standards Institute.
CLSI (2008). Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals. CLSI Supplement VET08, 4th Edn. Wayne, Pennsylvania: Clinical and Laboratory Standards Institute.
CLSI (2012a). Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically; Approved Standard – Ninth Edition.Guideline. CLSI document M07-A9. Wayne, PA: Clinical and Laboratory Standards Institute.
CLSI (2012b). Performance Standards for Antimicrobial Disk Susceptibility Tests; Approved Standard – Eleventh Edition. Guideline. CLSI document M02-A11. Wayne, PA: Clinical and Laboratory Standards Institute.
CLSI (2015). Performance Standards for Antimicrobial Susceptibility Testing; Twenty Fifth Informational Supplement M100-S25, Vol. 35. Wayne, PA: Clinical and Laboratory Standards Institute.
Cocchi, S., Grasselli, E., Gutacker, M., Benagli, M., Convert, M., and Piffaretti, J. C. (2007). Distribution and characterization of integrons in Escherichia coli strains of animal and human origin. FEMS Immunol. Med. Microbiol. 50, 126–132. doi: 10.1111/j.1574-695X.2007.00242.x
Dawes, F. E., Kuzevski, A., Bettelheim, K. A., Hornitzky, M. A., Djordjevic, S. P., and Walker, M. J. (2010). Distribution of class 1 integrons with IS26-mediated deletions in their 3’-conserved segments in Escherichia coli of human and animal origin. PLoS One 5:e12754. doi: 10.1371/journal.pone.0012754
Deng, Y., Bao, X., Ji, L., Chen, L., Liu, J., Miao, J., et al. (2015). Resistance integrons: class 1, 2 and 3 integrons. Ann. Clin. Microbiol. Antimicrob. 14:45. doi: 10.1186/s12941-015-0100-6
Domingues, S., da Silva, G. J., and Nielsen, K. M. (2015). Global dissemination patterns of common gene cassette arrays in class 1 integrons. Microbiology 161, 1313–1337. doi: 10.1099/mic.0.000099
Doublet, B., Praud, K., Weill, F.-X., and Cloeckaert, A. (2009). Association of IS26-composite transposons and complex In4-type integrons generates novel multidrug resistance loci in Salmonella genomic island 1. J. Antimicrob. Chemother. 63, 282–289. doi: 10.1093/jac/dkn500
Enne, V. I., Bennett, P. M., Livermore, D. M., and Hall, L. M. C. (2004). Enhancement of host fitness by the sul2-coding plasmid p9123 in the absence of selective pressure. J. Antimicrob. Chemother. 53, 958–963. doi: 10.1093/jac/dkh217
Essen-Zandbergen, A., Smith, H., Veldman, K., and Mevius, D. (2007). Occurrence and characteristics of class 1, 2 and 3 integrons in Escherichia coli, Salmonella and Campylobacter spp. in the Netherlands. J. Antimicrob. Chemother. 59, 746–750. doi: 10.1093/jac/dkl549
Fernández-Alarcón, C., Miranda, C. D., Singer, R. S., Lopez, Y., Rojas, R., Bello, H., et al. (2010). Detection of the floR gene in a diversity of florfenicol resistant gram-negative bacilli from freshwater salmon farms in Chile. Zoon. Publ. Health 57, 181–188. doi: 10.1111/j.1863-2378.2009.01243.x
Fonseca, E. L., Vieira, V. V., Cipriano, R., and Vicente, A. C. (2005). Class 1 integrons in Pseudomonas aeruginosa isolates from clinical settings in Amazon region, Brazil. FEMS Immunol. Med. Microbiol. 44, 303–309. doi: 10.1016/j.femsim.2005.01.004
Gillings, M., Boucher, Y., Labbate, M., Holmes, A., Krishnan, S., Holley, M., et al. (2008). The evolution of class 1 integrons and the rise of antibiotic resistance. J. Bacteriol. 190, 5095–5100. doi: 10.1128/JB.00152-08
Gillings, M. R., Gaze, W. H., Pruden, A., Smalla, K., Tiedje, J. M., and Zhu, Y. G. (2015). Using the class 1 integron-integrase gene as a proxy for anthropogenic pollution. ISME J. 9, 1269–1279. doi: 10.1038/ismej.2014.226
Goudarzi, M., Fazeli, M., Azad, M., Seyedjavadi, S., Mousavi, R., Rashidan, M., et al. (2015). Carriage of class 1 and class 2 integron in multidrug resistant Pseudomonas aeruginosa isolated from burn patients in Tehran hospitals, Iran. West Indian Med. J. 65, 32–39. doi: 10.7727/wimj.2014.315
Grape, M., Farra, A., Kronvall, G., and Sundström, L. (2005). Integrons and gene cassettes in clinical isolates of co-trimoxazole-resistant Gram-negative bacteria. Clin. Microbiol. Infect. 11, 185–192. doi: 10.1111/j.1469-0691.2004.01059.x
Grape, M., Sundstrüm, L., and Kronvall, G. (2003). Sulphonamide resistance gene sul3 found in Escherichia coli isolates from human sources. J. Antimicrob. Chemother. 52, 1022–1024. doi: 10.1093/jac/dkg473
Gu, B., Tong, M., Zhao, W., Liu, G., Ning, M., Pan, S., et al. (2007). Prevalence and characterization of class I integrons among Pseudomonas aeruginosa and Acinetobacter baumannii isolates from patients in Nanjing, China. J. Clin. Microbiol. 45, 241–243. doi: 10.1128/JCM.01318-06
Ho, P. L., Wong, R. C., Chow, K. H., and Que, T. L. (2009). Distribution of integron-associated trimethoprim–sulfamethoxazole resistance determinants among Escherichia coli from humans and food-producing animals. Lett. Appl. Microbiol. 49, 627–634. doi: 10.1111/j.1472-765X.2009.02717.x
Hsiao, K.-Y., Lee, M.-F., and Peng, C.-F. (2014). Detection and characterization of class 1 integron-associated gene cassettes from Pseudomonas aeruginosa isolates in southern Taiwan. Biomark. Genom. Med. 6, 74–78. doi: 10.1016/j.bgm.2014.02.004
Hugh, R., and Leifson, E. (1953). The taxonomic significance of fermentative versus oxidative metabolism of carbohydrates by various gram negative bacteria. J. Bacteriol. 66, 24–26.
Infante, B., Grape, M., Larsson, M., Kristiansson, C., Pallecchi, L., Rossolini, G., et al. (2005). Acquired sulphonamide resistance genes in faecal Escherichia coli from healthy children in Bolivia and Peru. J. Antimicrob. Agents 25, 308–312. doi: 10.1016/j.ijantimicag.2004.12.004
Juan, C., Zamorano, L., Mena, A., Albertí, S., Pérez, J. L., and Oliver, A. (2010). Metallo-b-lactamase-producing Pseudomonas putida as a reservoir of multidrug resistance elements that can be transferred to successful Pseudomonas aeruginosa clones. J. Antimicrob. Chemother. 65, 474–478. doi: 10.1093/jac/dkp491
Kang, H. Y., Jeong, Y. S., Oh, J. Y., Tae, S. H., Choi, C. H., Moon, D. C., et al. (2005). Characterization of antimicrobial resistance and class 1 integrons found in Escherichia coli isolates from humans and animals in Korea. J. Antimicrob. Chemother. 55, 639–644. doi: 10.1093/jac/dki076
Khosravi, A. D., Motahar, M., and Montazeri, E. A. (2017). The frequency of class1 and 2 integrons in Pseudomonas aeruginosa strains isolated from burn patients in a burn center of Ahvaz, Iran. PLoS One 12:e0183061. doi: 10.1371/journal.pone.0183061
Kiddee, A., Henghiranyawong, K., Yimsabai, J., Tiloklurs, M., and Niumsup, P. (2013). Nosocomial spread of class 1 integron-carrying extensively drug-resistant Pseudomonas aeruginosa isolates in a Thai hospital. Int. J. Antimicrob. Agents 42, 301–306. doi: 10.1016/j.ijantimicag.2013.05.009
Kor, S. B., Choo, Q. C., and Chew, C. H. (2013). New integron gene arrays from multiresistant clinical isolates of Enterobacteriaceae and Pseudomonas aeruginosa from hospitals in Malaysia. J. Med. Microbiol. 62, 412–420. doi: 10.1099/jmm.0.053645-0
Labbate, M., Roy Chowdhury, P., and Stokes, H. W. (2008). A class 1 integron present in a human commensal has a hybrid transposition module compared to Tn402: evidence of interaction with mobile DNA from natural environments. J. Bacteriol. 190, 5318–5327. doi: 10.1128/JB.00199-08
Lane, D. J. (1991). “16S/23S rRNA sequencing. pp. 115–176,” in Nucleic Acid Techniques in Bacterial Systematics, eds E. Stackebrandt and M. Goodfellow (New York, NY: John Wiley).
Laroche, E., Pawlak, B., Berthe, T., Skurni, D., and Petit, F. (2009). Occurrence of antibiotic resistance and class 1, 2 and 3 integrons in Escherichia coli isolated from a densely populated estuary (Seine, France). FEMS Microbiol. Ecol. 68, 118–130. doi: 10.1111/j.1574-6941.2009.00655.x
Lee, J., Young, J., We, J., Chun, J., Min, J., Yong, S., et al. (2001). The prevalence of trimethoprim-resistance-conferring dihydrofolate reductase genes in urinary isolates of Escherichia coli in Korea. J. Antimicrob. Chemother. 47, 599–604. doi: 10.1093/jac/47.5.599
Lévesque, C., Piché, L., Larose, C., and Roy, P. H. (1995). PCR mapping of integrons reveals several novel combinations of resistance genes. Antimicrob. Agents Chemother. 39, 185–191. doi: 10.1128/AAC.39.1.185
Lin, L., Yuan, K., Liang, X., Chen, X., Zhao, Z., Yang, Y., et al. (2015). Occurrences and distribution of sulfonamide and tetracycline resistance genes in the Yangtze river estuary and nearby coastal area. Mar. Pollut. Bull. 100, 304–310. doi: 10.1016/j.marpolbul.2015.08.036
Martínez, T., Vazquez, G. J., Aquino, E. E., Goering, R. V., and Robledo, I. E. (2012). Two novel class I integron arrays containing IMP-18 metallo-b-lactamase gene in Pseudomonas aeruginosa clinical isolates from Puerto Rico. Antimicrob. Agents Chemother. 56, 2119–2121. doi: 10.1128/AAC.05758-11
Maseda, H., Yoneyama, H., and Nakae, T. (2000). Assignment of the substrate-selective subunits of the MexEF-OprN multidrug efflux pump of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 44, 658–664. doi: 10.1128/AAC.44.3.658-664.2000
Mazel, D., Dychinco, B., Webb, V. A., and Davies, J. (2000). Antibiotic resistance in the ECOR collection: integrons an identification of a novel aad gene. Antimicrob. Agents Chemother. 44, 1568–1574. doi: 10.1128/AAC.44.6.1568-1574.2000
Miranda, C. D. (2012). “Antimicrobial resistance associated with salmonid farming,” in Antimicrobial Resistance in the Environment, eds P. L. Keen and M. H. H. M. Montforts (Hoboken, NJ: Wiley-Blackwell), 423–451. doi: 10.1002/9781118156247.ch22
Miranda, C. D., Godoy, F. A., and Lee, M. R. (2018). Current status of the use of antibiotics and their antimicrobial resistance in the chilean salmon farms. Front. Microbiol. 9:1284. doi: 10.3389/fmicb.2018.01284
Miranda, C. D., Kehrenberg, C., Ulep, C., Schwarz, S., and Roberts, M. C. (2003). Diversity of tetracycline resistance genes in bacteria from Chilean salmon farms. Antimicrob. Agents Chemother. 47, 883–888. doi: 10.1128/AAC.47.3.883-888.2003
Miranda, C. D., and Rojas, R. (2007). Occurrence of florfenicol resistance in bacteria associated with two Chilean salmon farms with different history of antibacterial usage. Aquaculture 266, 39–46. doi: 10.1016/j.aquaculture.2007.02.007
Miranda, C. D., and Zemelman, R. (2002). Antimicrobial multiresistance in bacteria isolated from freshwater Chilean salmon farms. Sci. Total Environ. 293, 207–218. doi: 10.1016/S0048-9697(02)00022-0
Muziasari, W. I., Managaki, S., Pärnänen, K., Karkman, A., Lyra, C., Tamminen, M., et al. (2014). Sulfonamide and trimethoprim resistance genes persist in sediments at Baltic Sea aquaculture farms but are not detected in the surrounding environment. PloS One 9:e92702. doi: 10.1371/journal.pone.0092702
Na, G., Zhang, W., Zhou, S., Gao, H., Lu, Z., Wu, X. L., et al. (2014). Sulfonamide antibiotics in the Northern Yellow Sea are related to resistant bacteria: implications for antibiotic resistance genes. Mar. Pollut. Bull. 84, 70–75. doi: 10.1016/j.marpolbul.2014.05.039
Navia, M., Ruiz, J., Sanchez-Cespedes, J., and Vila, J. (2003). Detection of dihydrofolate reductase genes by PCR and RFLP. Diagn. Microbiol. Infect. Dis. 46, 295–298. doi: 10.1016/S0732-8893(03)00062-2
Ozgumus, O. B., Sandalli, C., Sevim, A., Celik-Sevim, E., and Sivri, N. (2009). Class 1 and class 2 integrons and plasmid-mediated antibiotic resistance in coliforms isolated from ten rivers in northern Turkey. J. Microbiol. 47, 19–27. doi: 10.1007/s12275-008-0206-z
Partridge, S. R., Tsafnat, G., Coiera, E., and Iredell, J. R. (2009). Gene cassettes and cassette arrays in mobile resistance integrons. FEMS Microbiol. Rev. 33, 757–784. doi: 10.1111/j.1574-6976.2009.00175.x
Paulsen, I. T., Littlejohn, T. G., Radstrom, P., Sundstrom, L., Sköld, O., Swedberg, G., et al. (1993). The 3′ conserved segment of integrons contains a gene associated with multidrug resistance to antiseptics and disinfectants. Antimicrob. Agents Chemother. 37, 761–768. doi: 10.1128/AAC.37.4.761
Perreten, V., and Boerlin, P. (2003). A new sulfonamide resistance gene (sul3) in Escherichia coli is widespread in the pig population of Switzerland. Antimicrob. Agents Chemother. 47, 1169–1172. doi: 10.1128/AAC.47.3.1169-1172.2003
Ploy, M.-C., Lambert, T., Couty, J.-P., and Denis, F. (2000). Integrons: an antibiotic resistance gene capture and expression system. Clin. Chem. Lab. Med. 38, 483–487. doi: 10.1515/CCLM.2000.070
Quiroga, M. P., Andres, P., Petroni, A., Soler, Bistué AJ, Guerriero, L., Vargas, L. J., et al. (2007). Complex class 1 integrons with diverse variable regions, including aac(6′)-lb-cr and a novel allele, qnrB10, associated with ISCR1 in clinical enterobacterial isolates from Argentina. Antimicrob. Agents Chemother. 51, 4466–4470. doi: 10.1128/AAC.00726-07
Rådström, P., and Swedberg, G. (1988). RSF1010 and conjugative plasmid contain sul1, one of two known genes for plasmid-borne sulfonamide resistance dihydropteroate syntetase. Antimicrob. Agents Chemother. 32, 1684–1692. doi: 10.1128/AAC.32.11.1684
Rosser, S., and Young, H.-K. (1999). Identification and characterization of class 1 integrons in bacteria from an aquatic environment. J. Antimicrob. Chemother. 44, 11–18. doi: 10.1093/jac/44.1.11
Rossolini, G., and Mantengoli, E. (2005). Treatment and control of severe infections caused by multiresistant Pseudomonas aeruginosa. Clin. Microbiol. Infect. Suppl. 4, 17–32. doi: 10.1111/j.1469-0691.2005.01161.x
Ruiz-Martínez, L., López-Jiménez, L., Fusté, E., Vinuésa, T., Martínez, J. P., and Viñas, M. (2011). Class 1 integrons in environmental and clinical isolates of Pseudomonas aeruginosa. Int. J. Antimicrob. Agents 38, 398–402. doi: 10.1016/j.ijantimicag.2011.06.016
SERNAPESCA (2011). Informe Sobre Uso de Antimicrobianos en la Salmonicultura Nacional 2005-2009. Available at: http://www.sernapesca.cl (accessed April 12, 2018).
SERNAPESCA (2017). Informe Sobre Uso de Antimicrobianos en la Salmonicultura Nacional 2016. Available at: http://www.sernapesca.cl (accessed April 12, 2018).
Shah, S. Q., Cabello, F. C., L’Abée-Lund, T. M., Tomova, A., Godfrey, H. P., Buschmann, A. H., et al. (2014). Antimicrobial resistance and antimicrobial resistance genes in marine bacteria from salmon aquaculture and non-aquaculture sites. Environ. Microbiol. 16, 1310–1320. doi: 10.1111/1462-2920.12421
Shah, S. Q. A., Colquhoun, D. J., Nikuli, H. L., and Sørum, H. (2012). Prevalence of antibiotic resistance genes in the bacterial flora of integrated fish farming environments of Pakistan and Tanzania. Environ. Sci. Technol. 46, 8672–8679. doi: 10.1021/es3018607
Sköld, O. (2001). Resistance to trimethoprim and sulfonamides. Vet. Res. 32, 261–273. doi: 10.1051/vetres:2001123
Song, J. S., Jang, S. J., Bae, I. K., Lee, H. J., Jeong, B. C., and Lee, S. H. (2010). New complex class 1 integron carrying an ISCR1 element in Escherichia coli clinical isolates harbouring the blaCMY -11 gene. J. Med. Microbiol. 59, 132–134. doi: 10.1099/jmm.0.012559-0
Steers, E., Foltz, E. L., Graves, B. S., and Riden, J. (1959). An inocula replicating apparatus for routine testing of bacterial susceptibility to antibiotics. Antibiot. Chemother. 9, 307–311.
Sun, G., Yi, M., Shao, C., Ma, J., Zhang, Q., and Shao, S. (2014). Novel class 1 integrons in multi-drug resistant isolates from Eastern China. Indian J. Microbiol. 54, 227–231. doi: 10.1007/s12088-013-0441-9
Toleman, M. A., Bennett, P. M., Bennett, M. C., Jones, R. N., and Walsh, T. R. (2007). Global emergence of trimethoprim/sulfamethoxazole resistance in Stenotrophomonas maltophilia mediated by acquisition of sul genes. Emerg. Infect. Dis. 13, 559–565. doi: 10.3201/eid1304.061378
Toleman, M. A., Bennett, P. M., and Walsh, T. R. (2006b). ISCR elements: novel gene-capturing systems of the 21st century? Microbiol. Mol. Biol. Rev. 70, 296–316. doi: 10.1128/MMBR.00048-05
Toleman, M. A., Bennett, P. M., and Walsh, T. R. (2006a). Common regions e.g. orf513 and antibiotic resistance: IS91-like elements evolving complex class 1 integrons. J. Antimicrob. Chemother. 58, 1–6. doi: 10.1093/jac/dkl204
Tomova, A., Ivanova, L., Buschmann, A. H., Godfrey, H. P., and Cabello, F. C. (2018). Plasmid-mediated quinolone resistance (PMQR) genes and class 1 integrons in quinolone-resistant marine bacteria and clinical isolates of Escherichia coli from an aquacultural area. Microb. Ecol. 75, 104–112. doi: 10.1007/s00248-017-1016-9
Vinué, L., Sáenz, Y., Rojo-Bezares, B., Olarte, I., Undabeitia, E., Somalo, S., et al. (2010). Genetic environment of sul genes and characterisation of integrons in Escherichia coli isolates of blood origin in a Spanish hospital. Int. J. Antimicrob. Agents 35, 492–496. doi: 10.1016/j.ijantimicag.2010.01.012
Watts, J. E. M., Schreier, H. J., Lanska, L., and Hale, M. S. (2017). The rising tide of antimicrobial resistance in aquaculture: sources, sinks and solutions. Mar. Drugs 15:E158. doi: 10.3390/md15060158
Wright, M. S., Baker-Austin, C., Lindell, A. H., Stepanauskas, R., Stokes, H. W., and McArthur, J. V. (2008). Influence of industrial contamination on mobile genetic elements: class 1 integron abundance and gene cassette structure in aquatic bacterial communities. ISME J. 2, 417–428. doi: 10.1038/ismej.2008.8
Xu, Z., Li, L., Shirtliff, M. E., Alam, M., Yamasaki, S., and Shi, L. (2009). Occurrence and characteristics of class 1 and 2 integrons in Pseudomonas aeruginosa isolates from patients in Southern China. J. Clin. Microbiol. 47, 230–234. doi: 10.1128/JCM.02027-08
Keywords: bacteria, sulfonamide resistance, sul, integron, salmon farming, Chile
Citation: Domínguez M, Miranda CD, Fuentes O, de la Fuente M, Godoy FA, Bello-Toledo H and González-Rocha G (2019) Occurrence of Transferable Integrons and sul and dfr Genes Among Sulfonamide-and/or Trimethoprim-Resistant Bacteria Isolated From Chilean Salmonid Farms. Front. Microbiol. 10:748. doi: 10.3389/fmicb.2019.00748
Received: 18 June 2018; Accepted: 25 March 2019;
Published: 12 April 2019.
Edited by:
Martin Stephen Llewellyn, University of Glasgow, United KingdomReviewed by:
Chunlei Shi, Shanghai Jiao Tong University, ChinaCemal Sandalli, Recep Tayyip Erdoǧan University, Turkey
Copyright © 2019 Domínguez, Miranda, Fuentes, de la Fuente, Godoy, Bello-Toledo and González-Rocha. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Claudio D. Miranda, Y2RtaXJhbmRAdWNuLmNs