Corrigendum: Biological Control Agents Against Fusarium Wilt of Banana
 Giovanni Bubici1*
Giovanni Bubici1* Manoj Kaushal2
Manoj Kaushal2 Maria Isabella Prigigallo1
Maria Isabella Prigigallo1 Carmen Gómez-Lama Cabanás3
Carmen Gómez-Lama Cabanás3 Jesús Mercado-Blanco3
Jesús Mercado-Blanco3- 1Consiglio Nazionale delle Ricerche (CNR), Istituto per la Protezione Sostenibile delle Piante (IPSP), Bari, Italy
- 2International Institute of Tropical Agriculture (IITA), Dar es Salaam, Tanzania
- 3Department of Crop Protection, Institute for Sustainable Agriculture (CSIC), Córdoba, Spain
In the last century, the banana crop and industry experienced dramatic losses due to an epidemic of Fusarium wilt of banana (FWB), caused by Fusarium oxysporum f.sp. cubense (Foc) race 1. An even more dramatic menace is now feared due to the spread of Foc tropical race 4. Plant genetic resistance is generally considered as the most plausible strategy for controlling effectively such a devastating disease, as occurred for the first round of FWB epidemic. Nevertheless, with at least 182 articles published since 1970, biological control represents a large body of knowledge on FWB. Remarkably, many studies deal with biological control agents (BCAs) that reached the field-testing stage and even refer to high effectiveness. Some selected BCAs have been repeatedly assayed in independent trials, suggesting their promising value. Overall under field conditions, FWB has been controlled up to 79% by using Pseudomonas spp. strains, and up to 70% by several endophytes and Trichoderma spp. strains. Lower biocontrol efficacy (42–55%) has been obtained with arbuscular mycorrhizal fungi, Bacillus spp., and non-pathogenic Fusarium strains. Studies on Streptomyces spp. have been mostly limited to in vitro conditions so far, with very few pot-experiments, and none conducted in the field. The BCAs have been applied with diverse procedures (e.g., spore suspension, organic amendments, bioformulations, etc.) and at different stages of plant development (i.e., in vitro, nursery, at transplanting, post-transplanting), but there has been no evidence for a protocol better than another. Nonetheless, new bioformulation technologies (e.g., nanotechnology, formulation of microbial consortia and/or their metabolites, etc.) and tailor-made consortia of microbial strains should be encouraged. In conclusion, the literature offers many examples of promising BCAs, suggesting that biocontrol can greatly contribute to limit the damage caused by FWB. More efforts should be done to further validate the currently available outcomes, to deepen the knowledge on the most valuable BCAs, and to improve their efficacy by setting up effective formulations, application protocols, and integrated strategies.
Introduction
Among plant pathologists, everybody knows the story of Fusarium wilt of banana (FWB), also known as Panama disease. Until the 1950s, the cultivar Gros Michel had dominated the panorama of cultivated banana worldwide. The global banana production was threatened by a destructive soil-borne fungus, namely Fusarium oxysporum f. sp. cubense (Foc) (Figure 1). Fortunately, a resistant cultivar was identified, the “Cavendish.” Hence, due to its resistance to the races 1 and 2 of Foc (Foc R1 and R2), “Cavendish” was used to replace universally the “Gros Michel,” which unique fruit flavor has become virtually a distant memory. The global banana industry was saved until the 1990s, when a Foc strain virulent on “Cavendish” emerged, the race 4 (R4). First identified in Taiwan, Foc race 4 (R4) rapidly spread to South East Asian countries (e.g., Indonesia, Malaysia, and the Philippines), China, northern Australia, India, Pakistan, Middle East countries (e.g., Jordan, Israel, and Lebanon) and Africa (Mozambique) (Vézina, 2018a). Foc R4 strains are further separated into tropical race 4 (TR4) and subtropical race 4 (STR4), based on the evidence that the latter group necessitates predisposing factors, such as low temperatures, to cause the disease. The race 4 affects not only “Cavendish,” but also R1- and R2-susceptible varieties, while R1 affects “Gros Michel,” “Silk” and “Pisang Awak,” and R2 affects “Bluggoe” (Ploetz and Pegg, 2000). The race 3 (R3), which infects Heliconia spp., is no longer included in the Foc species (Ploetz, 2005). Currently, Foc TR4 is present in 19 of the 135 countries producing bananas (Dusunceli, 2017; Zheng et al., 2018), and its alarming spread has gained a remarkable interest from world media and cultural community (Butler, 2013; Gittleson, 2018).
FIGURE 1

Figure 1. Typical symptoms of Fusarium wilt on a banana plant cv. Pequeña Enana (or “Dwarf Cavendish”; AAA genome) in Tenerife.
Nowadays, bananas (including plantains and other cooking bananas) are the most produced fruit on the Earth (148 million tons produced in 2016 in 135 countries) and provide a staple for some 400 million people worldwide (Dusunceli, 2017). Its production and cultivated lands have progressively increased over the years (FAO, 2018b). Under the climate change scenario, the temperature increase will make conditions more favorable for banana production in the subtropical and tropical highlands. Land area suitable for bananas is estimated to augment 50% by 2070 (FAO, 2016). Currently, Cavendish varieties cover ca. 40% of the global production (the export trade accounts for 15% of the global production), and may be well the only bananas present on supermarket shelves of non-producer countries, because they dominate the export trade for a 12 billion dollars affair (FruiTrop, 2018). Therefore, it is clear that the economic impact of a second Foc epidemic would be more dramatic than the first one. As demonstrated in the past for the Foc R1 plague, the host genetic resistance is the better way to tackle the virulent strain. So far, however, no commercial varieties displaying an effective resistance against Foc R4 are available. Several research projects (Kema, 2018) and international initiatives, such as Promusa (Van den Bergh et al., 2018), World Banana Forum (Liu and Prada, 2018), the African Consortium for TR4 (Viljoen et al., 2018), and the four banana regional research networks MUSALAC, Innovate Plantain, BARNESA, and BAPNET (Bioversity International, 2018) have been started with the aim to save this crop. Lastly, the research project “Microbial Uptakes for Sustainable Management of Major Banana Pests and Diseases (MUSA)” (Horizon 2020 framework), in which we are involved, aims at improving the sustainable protection of banana crops from three major biotic constraints: FWB, nematodes, and banana weevil. The MUSA project holistically encompasses integrated pest management methods based on microbial consortia and available banana and enset germplasms, including newly developed elite hybrids (Ciancio, 2017).
Biological control has gained great interest in the last years in many pathosystems, including Foc/banana. This has been mainly due to the large input of pesticides, which cause economic, environmental and safety concerns. Biological control must not be a strategy limited to organic farming (adopted in 1% of banana cultivated lands) (Liu and Prada, 2018), but included within integrated disease management frameworks implemented in agricultural systems. Historically, biological control has suffered from inconsistent results over the seasons and the environments, most likely due to interacting variables (e.g., environmental, genetic, physiological, etc.) present in any given agro-ecosystem, and that are not fully understood or difficult to control. Factors affecting the biocontrol efficacy of FWB have been summarized by Guo et al. (2013).
In the present article, we report the impact of FWB in different continents and the overall disease management strategies with special regard to biological control. Thus, we report a comprehensive literature review of the biological control agents (BCAs) used against Foc, we also analyze critically the most relevant results achieved, and identify gaps in our current knowledge of this control strategy in order to (i) show the actual potential of biological control of FWB, and (ii) foster new research lines based on currently-available powerful technologies that will aid in developing novel and more effective biocontrol tools.
Disease Impact on Banana Production …
… in Asia and Australia
Since 1967, the pathogen has caused severe damage in Taiwan, the major exporter of banana to Japan till that time, and the lands cultivated with banana have passed from 50,000 ha in the 1960s to about 6,000 ha in the 2000s. Banana plantations have been also decimated in Indonesia and Malaysia in the early 1990s. The spread of Foc TR4 has aggravated the condition in Asia (Molina et al., 2009). In the Philippines, Foc has been present since 1970, but TR4 has been detected in 2006. Then, hectares of banana plantations have been abandoned by farmers because of Foc TR4, resulting in an annual loss of 3 billion dollars, and approximately 66,000 families lost their livelihood (Molina et al., 2009). As a consequence, in 2014 the Federation of Cooperatives in Mindanao (FEDCO) advised banana growers to shift to oil palm in fields compromised by Foc, a crop suitable for the Philippines lands and with increasing market demand (Carillo, 2014). During a survey in 2006, about 6,700 ha of banana plantations were found severely affected by the pathogen in the Guangdong province, southern China (Yi et al., 2007), and in 2012 an extensive damage was also observed around the Guangxi's capital and the Hainan island (Farquhar, 2012). India is the largest producer of bananas in the world with 28.6 million tons produced annually (FAO, 2018b), 70% of which is of the Cavendish cultivar Grand Naine (AAA genome group). There, six vegetative compatibility groups (VCGs) of Foc have been found, and disease severity has been as high as 80–90% on susceptible cultivars such as “Silk” (AAB), “Ney Poovan” (AB), “Pisang Awak” (ABB), “Pome” (AAB), “Bluggoe” (ABB), “Monthan” (ABB), and “Mysore” (AAB) (the latter was resistant to Foc, but then found infected by VCG 0124/0125) (Mustaffa and Thangavelu, 2011). Foc TR4 has been first found in 2015 in the state of Bihar, in the northeastern part of the country, and then detected in the states of Uttar Pradesh, Madhya Pradesh, and Gujarat, on the west coast. It has been officially confirmed only in 2017 (Damodaran et al., 2018). It has been estimated that Foc TR4 can inflict losses for 500 billion Indian rupees (ca. 7 billion American dollars) to the country's banana industry (Kulkarni, 2018).
The most recent records of TR4 have come from Jordan, Lebanon, Pakistan, Laos, Vietnam and Myanmar (Molina et al., 2009; Zheng et al., 2018). In 2016, two TR4 outbreak areas were identified in Israel, where the affected farms were promptly fenced in, and diseased plants destroyed. In May 2018, the Israel's National Plant Protection Organization officially declared the TR4 eradicated from Israel (EPPO Reporting Service, 2018). Although the public declaration, scientists know that Foc cannot be eradicated from the soil.
FWB in Australia has been reviewed previously by Pegg et al. (1996). Australia was the continent where Foc was reported for the first time (Bancroft, 1876), whereas Foc TR4 has been reported in the Northern Territory since 1997 (Bentley et al., 2001; Conde and Pitkethley, 2001). Later, in Queensland, three TR4 spots were identified between 2015 and 2018. In Australia, the severe biosecurity regulations have been very effective, and have made the Foc TR4 spread extremely slower than elsewhere. In September 2018, Biosecurity Queensland in partnership with Biosecurity Solutions Australia has announced the development of a certification system for TR4-infested farms that meet the requirements for severe food safety and biosecurity standards (Northern Queensland Register, 2018). The lack of resistant cultivar has drastically reduced the banana production in Australia, with a commercial industry loss above 90% and an augmented price of bananas in domestic markets (Cook et al., 2015).
… in Latin America and the Caribbean
Latin America and the Caribbean (LAC) support 28% of the global banana production and provide more than 80% of the banana exports (FAO, 2018b). According to Dita et al. (2013), FWB in LAC impact on certain production systems more than others. Particularly affected systems are the monocultures such as “Prata” (AAB) in Brazil, “Isla” and “Palillo” (AAB, Pacific plantain) in Peru, and the banana plantations intercropped with coffee and cocoa. Due to the Foc R1 epidemic, not only “Gros Michel” (AAA) was replaced with “Cavendish” (AAA), but also “Prata Ana” (AAB) (in Brazil) and apple banana (AAB). The cooking banana “Bluggoe” (ABB) was replaced with “Pelipita” (ABB), which is also resistant to Moko (Ralstonia solanacearum race 2). With the “Cavendish” advent, FWB disappeared as a problem for the trades (Buddenhagen, 1990), while the black leaf streak (or black Sigatoka, caused by Mycosphaerella fijiensis) assumed the primary importance (Ploetz, 2015a). Then, the rapid spread of Foc TR4 has recovered the importance of FWB, though black leaf streak remains of high economic relevance, especially in tropical regions. At present, the banana industry in LAC is almost totally based on Cavendish varieties, and the limited production for national markets is based on apple banana, plantain and “Gros Michel.” In countries of LAC, Foc TR4 is not present, therefore the pathogen exclusion by means of strong quarantine procedures is compulsory. Farmers, technicians, and politicians of LAC became sensitized of the FWB menace from the past, and they continue to become aware thanks to dissemination and technical events, workshops and meetings (Clercx, 2013; FAO, 2018c). An awareness campaign has been launched by a consortium of institutes in order to emphasize the importance of quarantine measures in preventing the entrance of Foc TR4 into LAC (Pocasangre et al., 2011).
… in Africa
Banana has been present on the continent for over 1400 years (Blomme et al., 2013). It is an important staple food for African people, as in countries such as Uganda, Rwanda, and Cameroon per capita consumption exceeds well 200 kg of bananas, so providing up to 25–35% of the daily nutrient intake. About 70–80% of banana production is consumed locally (FAO, 2018a; Viljoen et al., 2018).
Foc race 1 has first been reported from West Africa in 1924, and then from Tanzania in 1951 (Blomme et al., 2013; Viljoen et al., 2018). Vegetative compatibility groups and phylogenies of Foc populations in Africa have been extensively studied (reviewed by Blomme et al., 2013). Until the year 2000, FWB was reported in two out of the six production areas of South Africa: Kiepersol and southern KwaZulu-Natal. In KwaZulu-Natal, the disease appeared in 1940 and spread to Kiepersol with infected plant material, where it resulted in a 30% loss of banana plantations during the 1990s. At that time, only Foc STR4 occurred in South Africa (Viljoen, 2002). In Uganda, the major producer of cooking bananas worldwide with 3.7 million tons in 2016 (FAO, 2018b), FWB is less relevant than weevil [Cosmopolites sordidus (Germar)] and Xanthomonas wilt (Xanthomonas campestris pv. musacearum) and is present at altitudes above 1,300 m (Vézina, 2018b). With the occurrence in one farm in northern Mozambique, Foc TR4 officially entered the African continent in 2013, although it was likely an earlier presence (Butler, 2013). There, Foc TR4 destroyed about one million plants at a rate of approximately 15,000 plants per week (Jansen, 2017). After the detection of Foc TR4 in the six farms of Metochéria (about 1,500 ha cultivated with banana), the access of people and the movement of farm personnel and international staff have been restricted and controlled in order to avoid further spread of the pathogen (Viljoen et al., 2018).
The “Cavendish” varieties are not widely grown in Africa (about 10% of the region's production), whereas plantains (AAB) predominate in West Africa (71% of the production) with about 100 varieties, and highland banana (AAA; East Africa Highland Banana or EAHB) and other cooking types (ABB) predominate in East Africa (71% of production) with more than 50 varieties (FruiTrop, 2018; Viljoen et al., 2018). “Cavendish” was used in countries like Kenya to replace the susceptible varieties “Kampala” (“Gros Michel”) and “Bokoboto” (“Bluggoe,” ABB), susceptible to Foc R1. Besides these exceptions and few others (Sebasigari and Stover, 1988; Rutherford, 2001), African germplasm is largely resistant to Foc R1 and R2, but its reaction to Foc TR4 is still unknown. This scenario would make the Foc TR4 diffusion subtler than in a “Cavendish” monoculture, since new TR4 spots could be undetected properly or assumed to be caused by already established races but not TR4 (Vézina, 2018c). In addition, the fact that sucker-derived plant material is still used more than tissue culture plants in Africa (Dubois et al., 2013; Niere et al., 2014) may contribute to exacerbating the problem.
… in the Canary Islands
Banana is the most important intensive agricultural crop in the Canary Islands, an archipelago located in the Atlantic Ocean between 27° 37′-29° 25′ N and 13° 20′-18° 10′ W. The orography and pedological conditions have conditioned banana cultivation in these islands, shaping a particular landscape of terraces built up over volcanic stones and debris. Banana farms are usually small and about 80% of them have <1 hectare (Instituto Canario de Calidad Agroalimentaria, 2018). To check historical and current figures on banana production and cultivated area per island and municipalities, interested readers can consult the Canary Islands government's website (Gobierno de Canarias, 2018). Cultivar “Pequeña Enana” (or “Dwarf Cavendish,” AAA) is overwhelmingly predominant in the islands, with a minor and more recent presence of “Gran Enana” (or “Grande Naine” or “Grand Nain”; AAA) as well as local selections from “Pequeña Enana” like “Gruesa,” “Brier,” and “Negrita” (Azkolain Olaondo, 2016). Banana production in the archipelago is highly technified with practices such as artificial soil preparation, use of in vitro propagated plants from selected cultivars, modern fertigation approaches and extended use of greenhouse cultivation systems (Azkolain Olaondo, 2016). In Tenerife, Fusarium wilt was detected for the first time in the 1920s (Blomme et al., 2013). It is currently present in any area of the archipelago where banana is cultivated, showing an incidence of affected plants ranging from 2 to 12%. In some cases, however, incidence has been reported to be much higher causing more than 30% of crop loss in specific spots (Rodríguez Serrano, 2012). Studies carried out to determine the racial structure and pathogenicity of Foc isolates obtained from infected plants and soils of “Dwarf Cavendish” and “Grande Naine” plantations pointed to the fact that there are no differences among pathogen populations (Regalado Guijarro and Hernández Hernández, 1998). The presence and virulence of Foc STR4 have been well determined in Tenerife, and this race has also been claimed to be responsible for Dwarf Cavendish infections occurring in Gran Canaria island (Ploetz et al., 1990; Domínguez-Hernández et al., 2008).
Overall Control Strategies Currently Adopted
Available and currently-implemented measures to control FWB have been recently compiled (Ploetz, 2015b; Dita et al., 2018; Siamak and Zheng, 2018), and comprehensive information are also available in web portals such as World Banana Forum (Clercx, 2013) and Promusa (Van den Bergh et al., 2018). Previously, other authors have also produced similar reviews (e.g., Figueroa, 1987; Jeger et al., 1996; Wui, 2000; Murray, 2001; Pocasangre et al., 2011; Pérez-Vicente, 2015).
Foc is particularly difficult to control for a number of reasons: (a) it is a soil-borne fungus with a long survival in the soil (more than 20 years), even in the absence of plant hosts (Stover, 1962; Buddenhagen, 2009), or within alternate hosts which do not necessarily show disease symptoms (Waite and Stover, 1960; Pittaway et al., 1999; Hennessy et al., 2005); (b) being a vascular pathogen, it escapes the contact with the control means (e.g., non-systemic fungicides, non-endophytic BCAs, etc.) once it penetrates into the plant; (c) it can be spread by banana vegetative propagation material, soil vectored by workers and machinery, irrigation water, etc.; and (d) the banana monoculture, especially Cavendish varieties in the case of Foc TR4, facilitates the pathogen spread.
Overall, fungicides have provided unsatisfactory control levels. In vitro, toxicity against Foc has been proved for phosphonate, ambuic acid, organotin mandelates, carbendazim, carboxin, propiconazole, benomyl, and difenoconazole (Davis et al., 1994; Davis and Grant, 1996; Li et al., 2001; Araujo et al., 2004; Somu et al., 2014). In planta, only a few research articles have reported a significant disease control by using fungicides (e.g., carbendazim) (Lakshmanan and Selvaraj, 1984; Eswaramurthy et al., 1988; Roy et al., 1998) or resistance inducers (e.g., indoleacetic acid and menadione sodium bisulphite) (Fernández-Falcón et al., 2003; Borges et al., 2004). Until now, however, these pot-experiments have not been validated under field conditions.
In the lack of highly effective control means, like available sources of host genetic resistance, plant diseases are usually managed by integrated frameworks, with an emphasis in preventive measures. This is particularly true for soil-borne diseases like FWB and Verticillium wilts, whose causal pathogens cannot be eradicated once contaminate the soil. With few context-specific differences, integrated disease management strategies of diverse vascular diseases have much in common (Bubici and Cirulli, 2008; Cirulli et al., 2010; López Escudero and Mercado-Blanco, 2011; Jiménez-Díaz et al., 2012).
In the case of FWB, Dita et al. (2018) stressed that the implementation and integration of some disease management practices may vary according to four main farm-level scenarios, which indeed correspond to four stages of the disease epidemic: (i) absence of Foc, (ii) first incursion of Foc, (iii) low FWB prevalence, and (iv) high FWB prevalence (Figure 2). In the absence of a devastating pathogen like Foc, it is imperative to adopt exclusion measures to prevent the pathogen entrance, both at farm level using proper practices adopted by the personnel, and at the regional or national level using legal initiatives such as quarantine, certification, etc. This is the case of Foc TR4 in LAC and other countries where this race is not yet present. At the first incursion of Foc, exclusion methods still have great importance, but containment measures must be rapidly initiated and scrupulously applied. Farms in countries where Foc TR4 has been detected recently would be in this second situation (e.g., Mozambique, Lebanon, Pakistan, Israel, Laos, Vietnam, and Myanmar). Once Foc is established, exclusion tactics make no longer sense, but containment measures must be implemented, and integrated disease management can be adopted under low disease pressure. With high disease prevalence, containment measures are obviously not effective, and integrated disease management may be questionable.
FIGURE 2
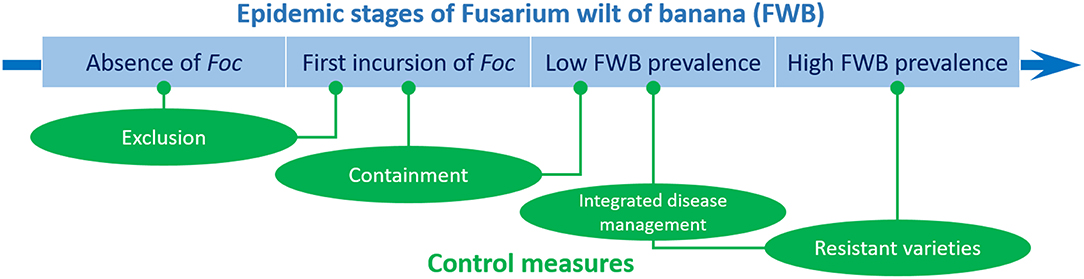
Figure 2. Control strategies of Fusarium wilt of banana at different epidemic stages. In the absence of Fusarium oxysporum f. sp. cubense (Foc), exclusion measures must be used to prevent the pathogen entrance. At the first incursion of Foc, containment measures must be rapidly initiated. Once Foc is established, containment measures must be continued, and integrated disease management can be adopted. With a high disease prevalence, the use of resistant varieties is the only way to successfully combat FWB.
Thus, the use of resistant varieties is the only way for successfully confronting FWB. As mentioned above, this was already experienced for the Foc R1 epidemic. On the other hand, Dita et al. (2018) have reported two examples where the integrated disease management against Foc R1 allows some farmers to grow the susceptible varieties “Prata”-type, in Brazil, and “Gros Michel,” in Colombia.
Unfortunately, no commercial varieties displaying high resistance to Foc TR4 coupled with good agronomic traits and fruit characteristics are yet available. However, the research on breeding for resistance has been particularly active and, besides conventional breeding and screening of genotypes, several mutant, and transgenic bananas have been developed (Figure 3). Resistance sources to Foc TR4 have been found in banana wild relatives, especially M. basjoo and M. itinerans, but also in M. yunnanensis, M. nagensium, M. ruiliensis, and M. velutina, whereas higher disease intensity was observed in M. balbisiana and M. acuminata subsp. burmannica (Li et al., 2015). Within M. acuminata, the genotype DH-Pahang, which genome was sequenced, has been found resistant to Foc TR4 (D'Hont et al., 2012; Zhang et al., 2018). Other promising varieties resistant to Foc TR4 are being tested under field conditions. Some of them were developed by the Fundación Hondureña de Investigación Agrícola (FHIA), and others by the Taiwan Banana Research Institute (TBRI). Other resistant genotypes have been identified within the world's largest collection of bananas, owned by the International Musa Germplasm Transit Centre, which is managed by Bioversity International and hosted by the Katholieke Universiteit Leuven in Belgium (Vézina, 2018a). The so-called “Giant Cavendish” tissue-culture variants (GCTCV) have been selected for the resistance against Foc TR4 by the Taiwan Banana Research Institute (TBRI). Four GCTCVs and three important Philippine local varieties were assayed over two cropping seasons in a heavily Foc-infested field in the southern Philippines. While the commercially-grown varieties “Grand Naine” and “Lakatan” showed disease incidence up to 92%, the GCTCV varieties were largely resistant, with a disease incidence of 0–8%. Moreover, “Saba” (ABB) was completely resistant in the two seasons (Molina et al., 2016a). The TR4-susceptibility of African banana germplasm, generally known to be resistant to Foc R1, has been started to be evaluated. A collection of 14 genetically diverse EAHB and plantain varieties were evaluated in fields of China and the Philippines and found all resistant with disease incidence as low as 0–5%, except EAHB “Ibwi” which showed a disease incidence of 32% (Molina et al., 2016b).
FIGURE 3
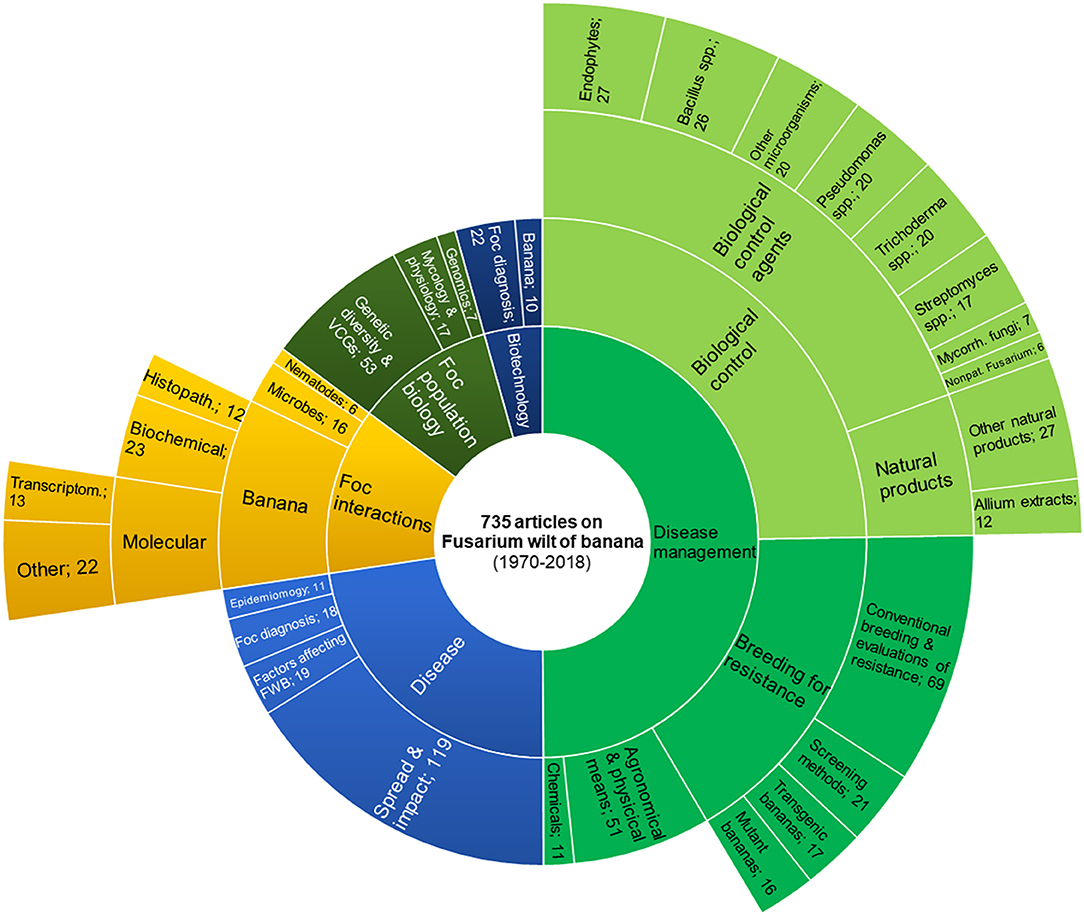
Figure 3. Main topics of scientific articles dealing with Fusarium wilt of banana. Articles were retrieved from the CAB Direct database (1970–2018) by searching the keywords “Fusarium cubense” or “Panama disease” in the title and abstract. Foc: Fusarium oxysporum f. sp. cubense; FWB: Fusarium wilt of banana; VCGs: vegetative compatibility groups.
When resistant varieties are not available, the management of FWB relies on an integrated approach. In the Canary Islands, for example, where FWB is caused by Foc STR4 and has a low prevalence, farmers try to contrast the pathogen by a number of palliative practices, sometimes based on empirical knowledge or deduced from basic research (Rodríguez Serrano, 2012; López-Cepero et al., 2014).
Soil amendment with calcium hydroxide or agricultural lime is practiced at the base of both diseased plants and symptomless neighboring ones with the aim to increase the pH and thus hinder Foc proliferation. The pH increase, however, may have deleterious effects on the physicochemical properties of soil, but probably the technicians and farmers choose the lesser evil. It has been observed that soil applications of CaCO3, Ca(OH)2, CaSO4, or Fe-EDDHA reduced Foc conidia germination and FWB severity, though the calcium amount was insufficient to change the soil pH. Nevertheless, this observation was not considered conclusive enough by the authors (Peng et al., 1999). A higher FWB incidence has been associated with low soil pH (Alvarez et al., 1981), but such observation has not been reproduced experimentally (da Silva Junior et al., 2000). Also, ammonia fumigation and biofertilization have been reported to reduce FWB incidence in a pot-experiment, with concurrent increases in soil pH, nutrient contents, and beneficial microbial community (Shen et al., 2019). Although no clear experimental evidence exists about the effectiveness against FWB of raising soil pH, it is known that Foc prefers low pH. For instance, a semi-selective agar medium widely used for isolation and enumeration of Foc from soil must be adjusted to pH 3.8–4.0 to favor Foc and hamper the growth of other fungi (Komada, 1975).
Adequate irrigation and fertilization regimes are also important to combat FWB. Waterlogging and acidification of nutritive solutions are generally avoided, and monthly treatments with zinc sulfate recommended. Stimulated by practical evidence that indoleacetic acid sprays combined with zinc supply reduced FWB, Hecht-Buchholz et al. (1998) observed ultrastructural changes of chloroplasts and mitochondria in banana leaves associated with the zinc-deficiency and, thus, with the FWB reduction. In a later work, the same research team confirmed the involvement of zinc nutrition in FWB development. Plants fertilized with a Zn-deficient solution were all diseased, whereas only 25% of plants treated with a normal zinc solution showed FWB symptoms (Fernández-Falcón et al., 2004). Overall, zinc seems to have a detrimental effect on several species of Fusarium, as observed in F. moniliforme (reduced fusarin C biosynthesis), in F. oxysporum f. sp. radicis-lycopersici (suppressed fusaric acid production) and F. verticillioides (reduced fumonisin production) (Jackson et al., 1989; Duffy and Défago, 1997; Savi et al., 2013). Similarly, silicon can decrease the intensity of several crop diseases (Datnoff and Rodrigues, 2015), including FWB (Fortunato et al., 2012b). This element may accumulate in the plant cell walls and act as a mechanical barrier, while it can also induce the phenylpropanoid pathway (e.g., lignin) and activate defense-related enzymes (e.g., chitinase, β-1,3-glucanase, phenylalanine ammonia-lyase, glucanase, peroxidase, polyphenoloxidase, etc.) (Smith et al., 2005; Fortunato et al., 2012a, 2014). Besides zinc and silicon, several other micro-nutrients like boron, iron, copper, and sodium, are known to affect FWB development (Qi et al., 2008; Sanjeev and Eswaran, 2008; Ji et al., 2012).
In the Canary Islands, it is also recommended (i) to increase the soil organic matter content above 3%, (ii) to avoid mulching for favoring soil aeration, (iii) to avoid high plant densities, and (iv) to apply plant resistance elicitors among other measures. Such practices arise from technical reports and, sometimes, from scientific articles (Alvarez et al., 1981; Aguilar et al., 2000; da Silva Junior et al., 2000; Rodríguez Serrano, 2012; López-Cepero et al., 2014). To the best of our knowledge, no detailed, long-term scientific studies have been conducted on the actual outcome of most of these practices, although they seem to help farmers to control FWB in a reasonable way.
Recently, Azkolain Olaondo (2016) investigated the use of biofumigation and soil solarization to manage FWB on a commercial farm in La Gomera island. However, results showed no differences in FWB incidence and severity among the treatments used (sheep manure or packing debris vs. control), although slight growth promotion was observed compared to the control. Moreover, solarization did not improve biofumigation treatments. An early trial from South Africa reported soil solarization as ineffective against FWB (Herbert and Marx, 1990). Nevertheless, in Indonesia, soil mulching with transparent polyethylene plastic for 10 months provided a 60% disease control until 14 months after planting. This treatment was superior to crop rotation with maize and bare soil treatment in terms of disease control and Foc TR4 suppression in the soil (Hermanto et al., 2012).
Crop rotation has provided attractive results in some cases. Rotation with rice reduced FWB to 8.1–17.6% after 1 year and to 0.8–6.3% after 2.5–3 years, compared to an initial level of 30–50%. Foc R4 population in the top layer of soil (20 cm) was undetectable after the treatment (Hwang, 1985). Furthermore, rotation with Chinese leek (Allium tuberosum) reduced FWB incidence and severity by 88–97% and 91–96%, respectively, and increased yield by 36–86% (Huang et al., 2012). In vitro assays showed that crude extracts of Chinese leek inhibit Foc R4 growth, suppress conidia proliferation and germination, and inhibit the activity of two cell wall degrading enzymes produced by Foc, polygalacturonase, and cellulase (Huang et al., 2012; Yang J. et al., 2015). Aqueous leachates and volatiles from Chinese leek exhibited strong inhibitory activity against Foc. Five volatiles including 2-methyl-2-pentenal and four organosulfur compounds (dimethyl trisulfide, dimethyl disulfide, dipropyl disulfide, and dipropyl trisulfide) were identified from the leaves and roots and found particularly active against the pathogen in vitro (Zhang et al., 2013). The pineapple-banana rotation has also given promising results. A 2-year crop rotation culminated with an 81% FWB reduction, and such control level was also attributed to changes in the bacterial and, more importantly, fungal communities in the soil (Wang B. et al., 2015). Lower disease control levels have been obtained by using rotations with maize, sugarcane, sunflower or eggplant (Hwang, 1985; Hermanto et al., 2012; Wang B. et al., 2015; Hong et al., 2017).
Biological (or reductive) soil disinfestation (BSD) has surfaced as another encouraging control means against FWB. Field experiments have shown that FWB can be reduced up to 82% in flooded soil incorporated with 0.5% rice straw (Huang et al., 2015c). The BSD simultaneously reduced the Foc inoculum in the soil and ameliorated the beneficial microbial communities, making the soil more suppressive to FWB (Huang et al., 2015a). Also, some microbes (e.g., Clostridium spp.) producing organic acids toxic for Foc (acetic, butyric, isovaleric and propionic acids) were found more abundant after BSD (Huang et al., 2015b).
Biological Control Agents and Their Modes of Action: Actual and Effective Tools to Confront Fusarium Wilt of Banana
From a historical perspective, biocontrol of FWB has been studied for more than 70 years (Thaysen and Butlin, 1945). Studies have reported that disease suppressive sites showed microbial communities displaying higher richness and diversity (Shen et al., 2015b; Köberl et al., 2017), and possibly a higher number of antagonistic members, as observed for streptomycetes (Kinkel et al., 2012; Jauri et al., 2017). Moreover, differences in the composition of these communities correlated with the disease suppressiveness (abundance of Acidobacteria) or conduciveness (abundance of Bacteroidetes) of soil (Shen et al., 2015b). The manipulation of banana rhizosphere microbiota by the introduction of well-characterized antagonists alone or in combination with organic amendments (bio-organic fertilizers) has already yielded very promising results against Foc TR4 in China (Shen Z. et al., 2013; Shen et al., 2015a; Xue et al., 2015). This strategy also leads to changes in the structure and composition of the microbial community that can be harnessed for more effective control of FWB (Shen et al., 2015a; Fu et al., 2016b).
The mechanisms underlying the BCAs' biocontrol activity are many and variegated (Narayanasamy, 2013; Singh, 2014). It is essential to know the BCAs' modes of action, including weakness and requirements, in order to exploit their potential for disease management in the most effective manner. Also, the combination of BCAs with different modes of action might result in a better biocontrol due to additive, or even synergistic, interactions between BCAs (Parnell et al., 2016; De Vrieze et al., 2018). Figure 4 schematizes how BCAs can act directly or indirectly against Foc. Direct antagonism may be due to antibiosis (e.g., antibiotics, lytic enzymes, volatile organic compounds, etc.), parasitism, or competition (for space and/or nutrients). Induction of plant local/systemic resistance, plant growth promotion, or changes of soil/plant microbiota in favor of more beneficial microbial taxa are typical mechanisms that indirectly act against the pathogen, or at least contribute to reducing the infections or the disease. Antibiosis is one of the primary mechanisms possessed by BCAs. Indeed, initial in vitro selection of new BCAs often relies on the evaluation of the sole anti-microbial activity against the pathogen (Figure 5A), while other mechanisms are studied later on, possibly once the BCA effectiveness is demonstrated at least under controlled conditions (Figure 5B).
FIGURE 4
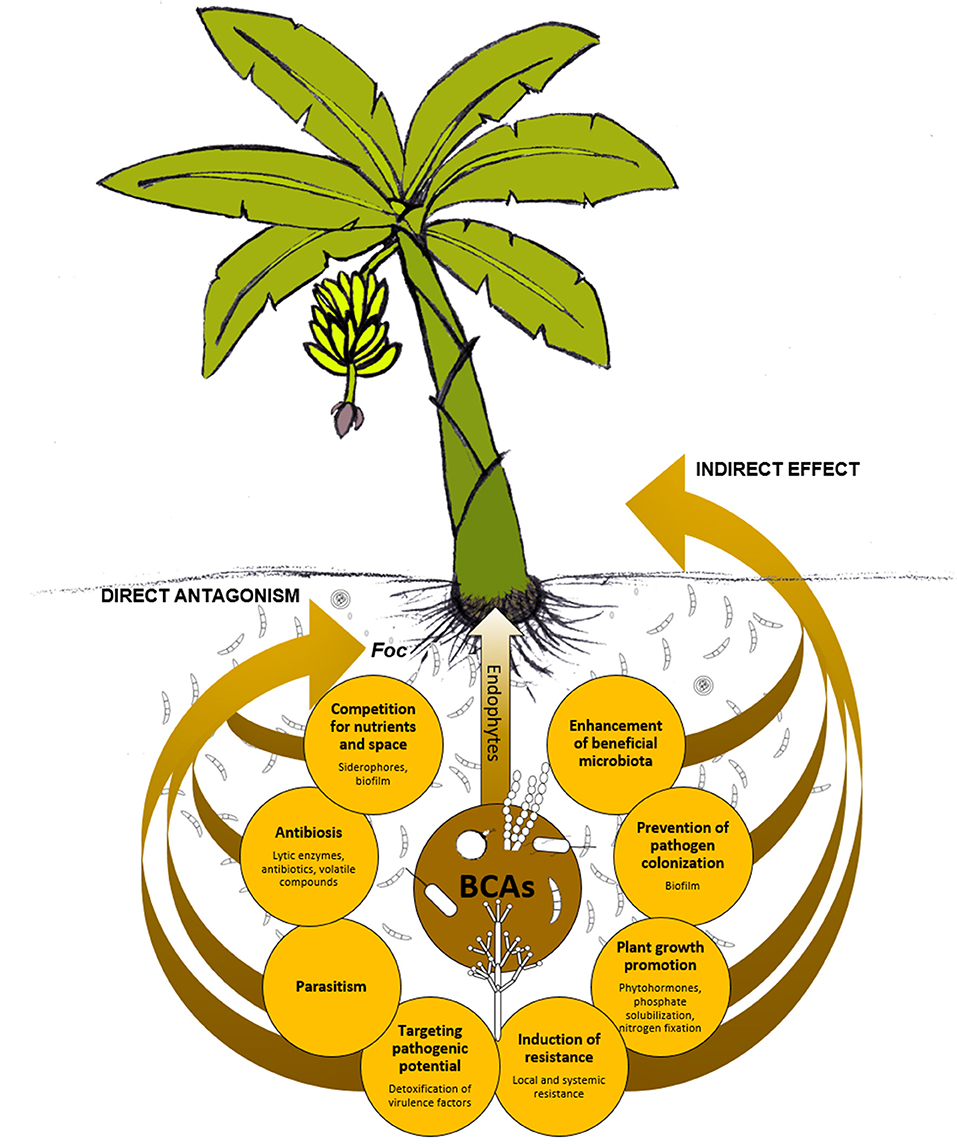
Figure 4. Possible modes of action of biological control agents (BCAs). Beneficial microorganisms can exhibit direct antagonism against Fusarium oxysporum f. sp. cubense (Foc) and can affect the plant physiology and/or the microbiota with a consequent, indirect effect against the pathogen.
FIGURE 5

Figure 5. Effectiveness of biological control agents against Fusarium oxysporum f. sp. cubense (Foc). The dual culture method (A) allows testing the in vitro antifungal activity of metabolites produced by biological control agents (BCAs). Pot experiments (B) provide first evidence for the in planta efficacy of BCAs.
Literature Overview Shows That Biocontrol of Fusarium Wilt of Banana Is a Widely Studied Research Topic
In order to understand the relevance of the research on biological control of FWB, we searched the keywords “Fusarium cubense” or “Panama disease” in the title and abstract of articles indexed by the CAB Direct database (CAB Direct, 2018). The search yielded a comprehensive overview of the literature from 1970 to date. Amongst 735 retrieved articles, 367 were focused on disease management and, with 182 articles (ca. 25% of the total), biological control represents the largest research sub-topic on FWB (Figure 3). Records of FWB presence, distribution or impact on the crop productivity rank second for their abundance (119 articles), suggesting how much the disease is feared worldwide (Figure 3). With 123 retrieved articles, breeding for resistance is the third relevant FWB-related research topic (17% of the total articles), which in our article categorization included conventional breeding, screening for resistance, development, and evaluation of mutant and transgenic bananas. Therefore, biological control and host genetic resistance have been considered the most important strategies for the management of FWB. It should be noted that conventional breeding in banana is particularly laborious because cultivated varieties are polyploid; hence, diploid parents must be used (or generated) for crosses, and then promising progeny must be polyploidized. In the late 70's, the research on breeding was much more intense than that on biological control, which increased only since 2000s (Supplemental Figure 1). At least 143 articles deal with BCAs. Endophytes and strains of Trichoderma spp., Pseudomonas spp., and Bacillus spp. have been the most studied BCAs, followed by arbuscular mycorrhizal fungi. Streptomyces spp. have been investigated mainly in vitro, with very few in planta experiments, and none conducted in the field (Figures 3 and Supplemental Figure 2). In 39 papers, the effects of natural products against Foc is showed and, interestingly, 12 articles published since 2011 report studies on the efficacy of A. tuberosum extracts (Supplemental Figure 1). The remaining articles are focused on the interactions of Foc with other microorganisms and its population biology, where the “-omics” approach has been used since the 2010s. Experiments conducted under field, pot and in vitro conditions are listed in Tables 1, 2 and Supplemental Table 1, respectively, and their by-taxon abundance is summarized in Supplemental Figure 2.
TABLE 1
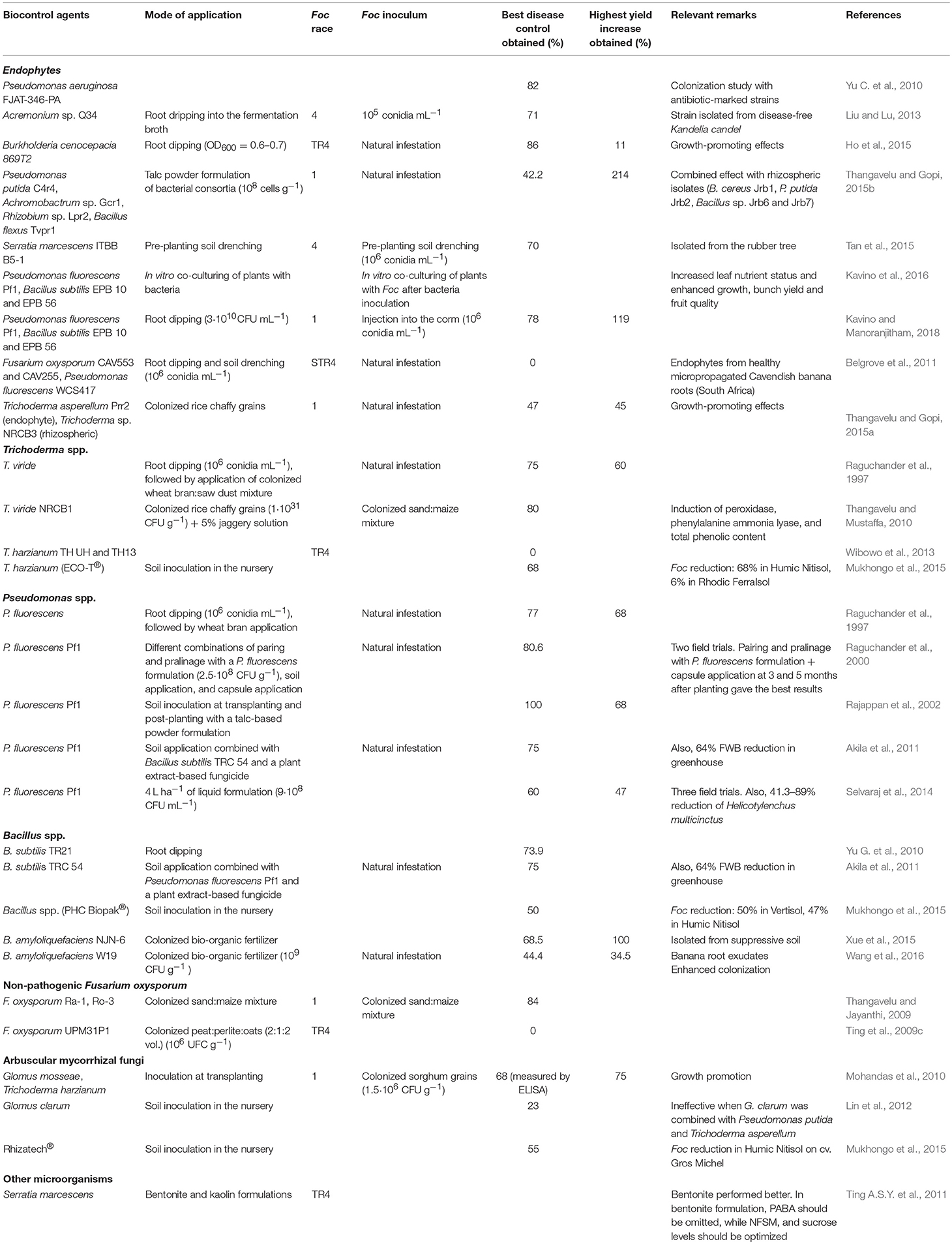
Table 1. List of biocontrol field trials against Fusarium wilt of banana.
TABLE 2
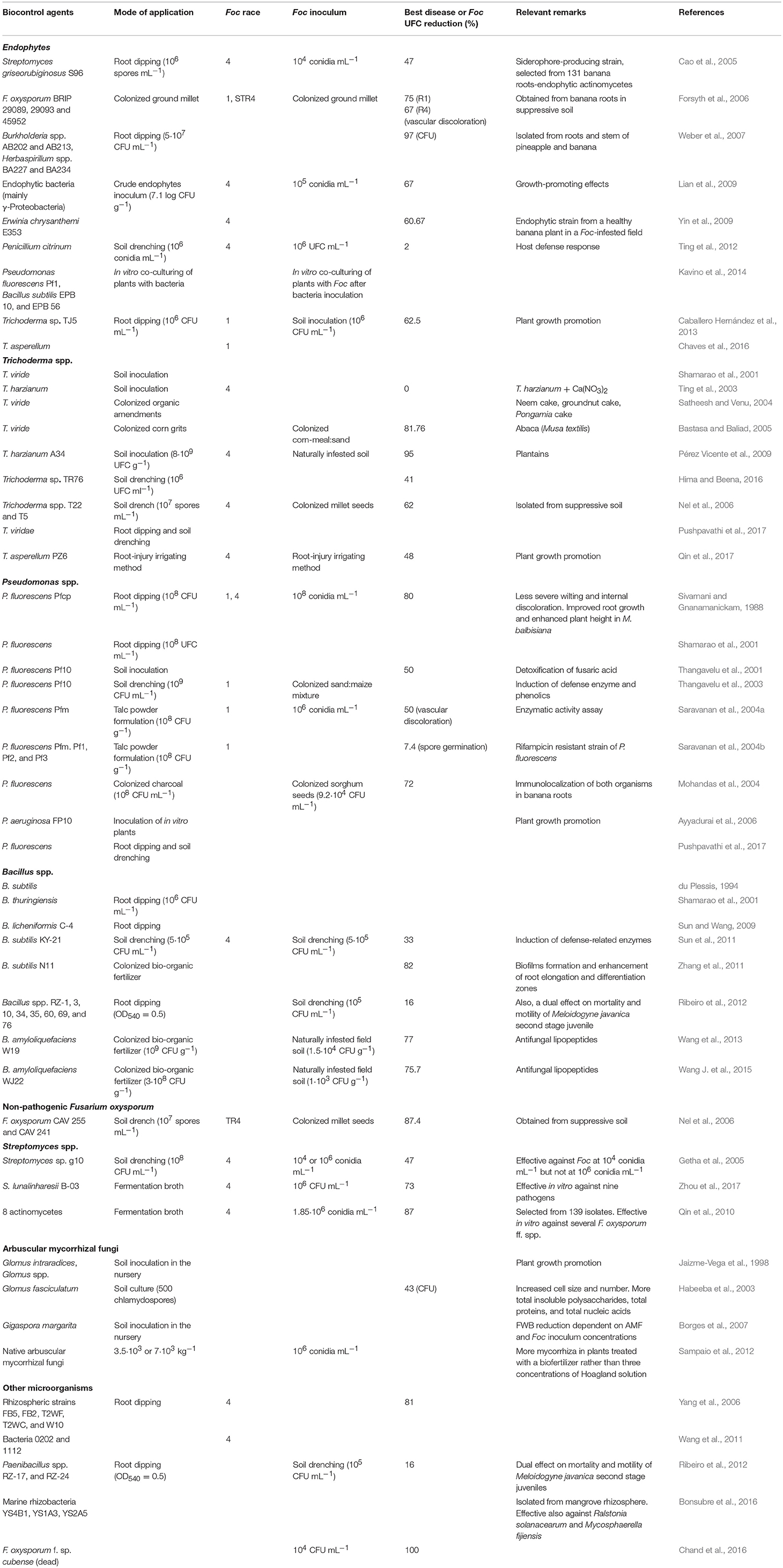
Table 2. List of biocontrol pot-experiments against Fusarium wilt of banana.
Data mining from the literature allowed us to infer on the potential and actual efficacy of several microbial genera in the control of FWB. In particular, from each retrieved article we mined the best FWB control value obtained, but not all the data. Although the experiments are not so numerous, plotting these data provides some evidence for microbial genera more prone to control FWB than others (Figure 6A). Interestingly, we realized that several biocontrol trials reached the field stage, and results even showed high effectiveness. In the field, FWB was controlled up to 77% (median of 5 articles) by Pseudomonas spp. strains, and up to 71% by several endophytic strains (8 articles) and other Trichoderma spp. strains (4 articles) (Figure 6A). Lower biocontrol was obtained with Bacillus spp. (69% as the median of 5 articles), arbuscular mycorrhizal fungi (55%, 3 articles), and non-pathogenic Fusarium strains (42%, 2 articles), whereas most studies on Streptomyces spp. have been limited to in vitro conditions. Biocontrol under field conditions has been often reported to be less effective than under (semi)controlled, pot experimental conditions. Surprisingly, a different scenario appears for FWB biocontrol. In fact, pot- (median = 65) and field-experiments (median = 70) resulted substantially similar in terms of disease control efficacy. Moreover, articles dealing with Pseudomonas spp. reported on average a higher efficacy in the field (median = 77%) than in pot-experiments (median = 50%), as also observed for endophytes, viz. 71% in the field and 65% in pots. Non-pathogenic Fusarium strains, however, resulted more effective in pot-experiments (median = 87%) than in the field (median = 42%). Finally, in planta effectiveness of BCAs was not dependent on the target Foc race (R1 and R4), meaning that Foc R1 and R4 showed overall comparable sensitivity to BCAs (median disease control efficacy of 62%; Figure 6B).
FIGURE 6
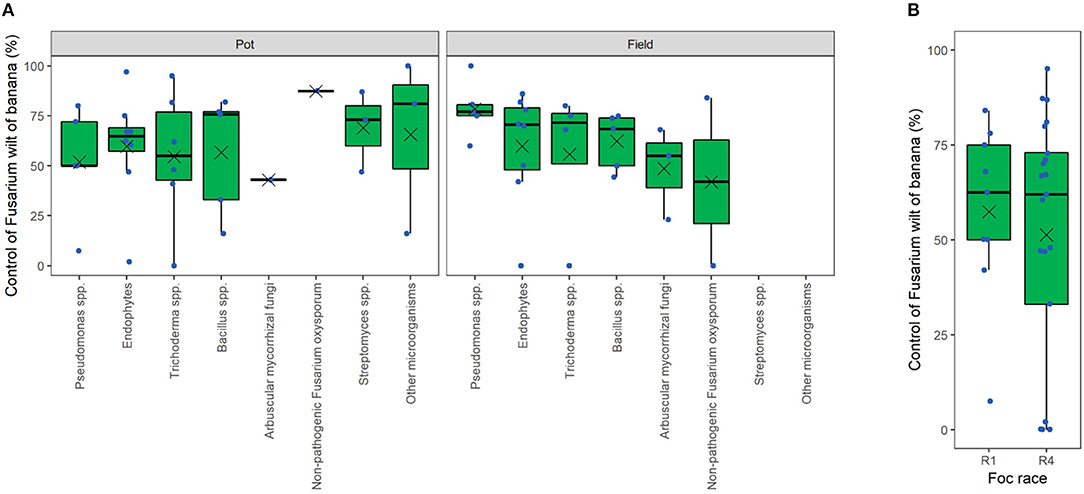
Figure 6. Overview of the efficacy of biological control agents against Fusarium wilt of banana. Data of pot and field trials separated by microbial groups (A) or by Fusarium oxysporum f. sp. cubense (Foc) races (B). Dots correspond to the best control levels (viz. reduction of disease incidence or severity) obtained in each scientific article in the literature. In the boxes, the mean is reported as a cross and the median as a horizontal line.
Endophytes: The Help From Inside
All plants harbor a huge diversity of beneficial or neutral microorganisms living inside their tissues without causing any deleterious effect in the host (reviewed by Hardoim et al., 2015). Since beneficial endophytes can promote plant fitness and growth through a range of different mechanisms (i.e., phytohormones synthesis, nitrogen fixation, phosphate solubilization, induction of defense responses, alleviation of abiotic stress by reducing ethylene level, etc.) (Compant et al., 2016), they have a considerable agro-biotechnological potential yet to be fully exploited (Mercado-Blanco and Lugtenberg, 2014, and references therein). Endophytes have found evolutionary solutions to live within the plant interior (i.e., nutrient availability, evading/modulating host defense responses, etc.), where they can also deploy biocontrol activity against pathogens. Thus, an increasing number of studies are available on the isolation, characterization, and assessment as BCAs of specific culturable members of indigenous endophytic communities. Endophytic bacteria and fungi from banana plants have received early attention as BCAs against Foc and other biotic constraints (Ortiz and Pocasangre, 2012; Niere et al., 2014). Although many of them belong to genera that will be described in more detail in the following sections, we would like to summarize some reports focused on endophytes to highlight the importance of this special group of the plant-associated microbes.
It is interesting that many experiments with endophytes against FWB have been conducted in the field during the last years. One of the early research showed how P. aeruginosa FJAT-346-PA reduced FWB by 82–84% in semi-field and field conditions. The strain was proved to colonize the roots and stems of banana and to promote the plant growth (Yu C. et al., 2010). Cao et al. (2004) reported on the actinomycete communities found in the interior of leaves and roots of both healthy and diseased banana plants. Most of the isolates were Streptomyces griseorubiginosus-like strains. Isolates displaying antagonistic activity against Foc originated from roots of healthy plants, whereas no difference in this phenotype was reported between antagonists isolated from healthy and wilted leaves. In vitro, the antagonism of S. griseorubiginosus S96 was lost when FeCl3 was introduced in the Petri plates, suggesting that it relies on siderophore production. In vivo, FWB severity (R4) was reduced by 47%, and plant fresh weight increased in plantlets treated with strain S96 compared to the control (Cao et al., 2005). Similarly, three endophytic Bacillus spp. isolated from different Musa cultivars in Brazil showed antagonistic activity against Foc and Colletotrichum guaranicola (Souza et al., 2014). More recently, endophytic bacteria from the classes Actinobacteria (genera Arthrobacter, Brevibacterium, Corynebacterium, Curtobacterium, Kocuria, Kytococcus, Micrococcus, Naumanella, Rothia, and Tessaracoccus), α- and γ-Proteobacteria (Brevundimonas, Enterobacter, Klebsiella, Pseudomonas, Serratia, and Sphingomonas) and Firmicutes (Bacillus and Staphylococcus) were isolated from the shoot-tips of banana cv. Grand Naine (Sekhar and Pious, 2015). From the collection generated in this study, strains of Pseudomonas aeruginosa, Klebsiella variicola, and Enterobacter cloacae showed variable antagonistic activity against Foc, but their potential as effective BCAs remained to be proved. Other studies reported on fungal and bacterial endophytes originating from plants other than banana such as weeds and medicinal plants (Ting et al., 2009b) or Capsicum frutescens (He H. et al., 2002), making them interesting reservoirs of Foc antagonists worth to be explored. Finally, some studies go a step forward, exploring the potential mechanism involved in the antagonistic effect. For instance, Ting et al. (2010) investigated the role of volatiles produced by several fungal endophytes in the growth inhibition of Foc R4. Even if with inconsistent disease control, the same authors also suggested the induction of host resistance as a mechanism involved in FWB control mediated by Penicillium citrinum BTF08 isolated from banana internal stem tissues (Ting et al., 2012).
Interestingly, some of the reports were focused on the inoculation of BCAs at the in vitro propagation stage of banana plants. Banana tissue culturing is widely used as a source of pathogen-free planting material. Since banana plantlets produced by these micropropagation schemes are more susceptible to diseases due to the absence of the natural-associated microbiota, the reintroduction of endophytic and rhizospheric microorganisms to protect the plant against subsequent challenges by Foc is an approach that yielded promising results (Kavino et al., 2014, 2016; Kavino and Manoranjitham, 2018). The in vitro co-culturing of banana plants with Pseudomonas fluorescens Pf1, Bacillus subtilis EPB 10 and EPB 56 provided a successful control of FWB in the field, combined with increased leaf nutrient status, vegetative growth, bunch yield and fruit quality (Kavino et al., 2016). In two other field trials, the endophyte and rhizobacteria strain led to a FWB control of 78% and a significant higher bunch weight (Kavino and Manoranjitham, 2018). Furthermore, this strategy was also proved to be successful when inoculating banana tissue culture plantlets with a mixture of naturally-occurring uncultivated endophytes from healthy banana plants of a commercial plantation (Lian et al., 2009). In this latter study, the re-introduction of endophytes to the banana tissue culture led to a significant reduction of wilt disease (67%) caused by Foc R4 (artificial inoculation) and growth promotion under greenhouse conditions. Ten non-pathogenic F. oxysporum isolates obtained from healthy micro-propagated “Cavendish” banana roots were able to significantly reduce FWB under greenhouse conditions, but none of them nor P. fluorescens WCS417 gave protection from the disease (STR4) in the field (Belgrove et al., 2011). Some researchers tried to have more chances to obtain effective BCAs by isolating from FWB-suppressive soil, e.g., non-pathogenic F. oxysporum strains (Forsyth et al., 2006), or even from healthy banana plants located in Foc-infested soil, e.g., Erwinia chrysanthemi E353 (Yin et al., 2009).
Furthermore, endophytes effective against FWB did not originate only from banana plants. For instance, Ho et al. (2015) isolated Burkholderia cenocepacia 869T2 from surface-sterilized vetiver grass (Chrysopogon zizanioides) roots. Banana tissue culture plantlets inoculated with 869T2 showed a lower disease incidence caused by Foc TR4 (86% incidence reduction) as well as significant plant growth promotion under field conditions. The endophytic strain Serratia marcescens ITBB B5-1 was isolated from the rubber tree (Hevea brasiliensis) (Tan et al., 2015). A sharp reduction in disease severity caused by Foc R4 was scored under both greenhouse (79%) and field conditions (70%) upon inoculation of banana plants with this strain. Moreover, chitinase and glucanase activities were suggested to be involved in its antifungal activity (Tan et al., 2015). Endophytic diazotrophic bacteria (strains of Burkholderia sp. and Herbaspirillum sp.) isolated from roots and stems of pineapple (Weber et al., 2007), and an Acremonium sp. strain isolated from Kandelia candel (Liu and Lu, 2013) are additional examples of promising biocontrol and biofertilizer candidates for the banana crop that originate from a different host. Finally, effective control of FWB and production increase (number of banana hands and bunch weight) was reported under field conditions when different combinations of endophytic (P. putida C4r4, Achromobactrum sp. Gcr1, Rhizobium sp. Lpr2, and B. flexus Tvpr1) and rhizospheric bacteria (B. cereus Jrb1, P. putida Jrb2, Bacillus sp. Jrb6, and Jrb7), in this case isolated from different banana accessions, were applied to a naturally infested soil (Thangavelu and Gopi, 2015b). In another field trial, combined applications of the endophytic T. asperellum prr2 together with the rhizospheric Trichoderma sp. NRCB3 resulted in a 47% reduction of FWB incidence and a 45% increase of the bunch weight (Thangavelu and Gopi, 2015a).
Bacillus spp.: The Endospore-Forming Bacteria
The use and number of Bacillus spp. strains displaying suppressive effect against plant diseases caused by soil-borne phytopathogens has been increased rapidly, and a large body of knowledge on the biocontrol mechanisms involved as well as on their application and effectiveness under diverse conditions is available for the interested reader (Fira et al., 2018; Aloo et al., 2019). The spore-forming ability of Bacillus species confers them an important advantage over other beneficial microorganisms in the field of biological control. On the one hand, this capability enables these bacteria to endure adverse environmental conditions. On the other hand, and from the agro-biotechnological point of view, it favors the development and manufacturing of commercial formulations more stable over time. In addition, many Bacillus species show rapid growth rates and the ability to synthesize a large number of secondary metabolites which play a key role in the antibiosis against many deleterious microorganisms (Radhakrishnan et al., 2017; Fira et al., 2018). Some species, such as B. subtilis, are also able to produce volatile organic compounds (VOCs), which are important sometimes for plant growth promotion and the activation of plant defense mechanisms by triggering induced systemic resistance (Raaijmakers et al., 2010; Cawoy et al., 2014). Bacillus-mediated plant growth promotion can also be due to the capacity to promote phytohormone (i.e., gibberellic acid and indole-3-acetic acid) biosynthesis, thereby enhancing nutrient uptake ability in the host and stimulating plant defense responses against biotic and abiotic stresses (Chen et al., 2007; Harman, 2011). Besides the production of antibiotics and the elicitation of systemic resistance in plants against pathogens, Bacillus species are also able to produce lytic enzymes like chitinase and β-1,3-glucanase, involved in the degradation of the fungal cell wall (Kumar et al., 2012). Considering their versatility, the combination of different Bacillus spp. strains (or with other BCAs) displaying different biocontrol mechanisms appears as an interesting approach to improve biocontrol effectiveness under different cropping scenarios and environmental conditions.
Bacillus spp. are commonly found in the banana rhizosphere (Xue et al., 2015), and many members of this genus have already been investigated as BCAs of diverse Fusarium-induced plant diseases (Khan et al., 2017). Among them, representatives of B. subtilis, B. amyloliquefaciens, B. pumilus and B. thuringiensis are found in the literature. Bacillus subtilis is well known for its antagonistic activity against several fungal and bacterial plant diseases. Its biocontrol activity is mainly attributed to antibiotic production (Cawoy et al., 2015), and its enzymatic products are highly active against many fungal pathogens. The biocontrol effect of the plant endophytic B. subtilis strain TR21 against FWB was investigated in Brazilian fields, and promising results (74% effectiveness) were reported (Yu G. et al., 2010). Likewise, B. subtilis strain N11 isolated from the rhizosphere of a healthy banana plant showed biocontrol activity in pot experiments under greenhouse conditions (Zhang et al., 2011). Addition of 10% (v/v) of culture filtrate of the endophytic B. subtilis strain EBT1 to the plant growth medium increased bud multiplication, plantlet weight, pseudo-stem height, and conferred resistance to plantlets against Foc conidia and toxin (Yang et al., 2010). Bacillus subtilis strain B25, isolated from banana rhizosphere soil in Hainan, is another example of an effective antagonist, not only against Foc, but also against other plant pathogenic fungi including Corynespora cassiicola, Alternaria solani, Botrytis cinerea, and Colletotrichum gloeosporioides (Tan et al., 2013; Yu et al., 2016). Results on its capability to control FWB under greenhouse and field conditions can be found in the literature, although they are not easily accessible (Liu, 2011). The antifungal protein of B25, identified as a disease-resistance protein, produced mycelium and spore tumescence and abnormal growth of the pathogen (Tan et al., 2013). The chitinolytic and heat tolerant strain B. subtilis TSA3 showed in vitro inhibition of Foc growth although effectiveness in planta and under field conditions has not been demonstrated yet (Nawangsih and Purba, 2013). Similarly to previous examples, B. subtilis strain S-1 not only inhibited in vitro Foc growth but also antagonized fungal pathogens such as the formae speciales lycopersici, vasinfectum and niveum of F. oxysporum, as well as Curvularia lunata, C. gloeosporioides, Verticillium dahliae, and Gibberella zeae (Sun et al., 2008).
The B. amyloliquefaciens strain NJN-6 was isolated from the rhizosphere of a healthy banana plant in a FWB-suppressive soil. Field plots with plants pre-treated (in nursery pots) with a bio-organic fertilizer colonized by NJN-6 showed a decreased disease incidence by 68.5%, resulting in doubled yield (Xue et al., 2015). The mode of action of this strain relies on several metabolites. The lipopeptide iturin A, a powerful antifungal surfactant, is produced by several Bacillus strains including NJN-6 (Yuan et al., 2011). Two homologs of bacillomycin D and three homologs of members of the macrolactin family were identified in NJN-6 using HPLC/electrospray ionization mass spectrometry. Bacillomycin D and macrolactin exhibited significant antagonistic effects against Foc and R. solanacearum, respectively (Yuan et al., 2012a). Finally, among 36 VOCs detected in NJN-6, 11 compounds completely inhibited the Foc growth (Yuan et al., 2012b). Wang et al. (2013) isolated 57 bacterial strains from the rhizosphere of healthy banana plants grown in a field severely affected by the disease, all showing antagonism against Foc. Six strains (W2, W10, W14, W15, W17, and W19) displaying the best survival abilities in the rhizosphere soil were tested in greenhouse experiments, B. amyloliquefaciens W19 being the most effective against FWB. Moreover, and even more suggestive, biocontrol effectiveness of a bio-organic fertilizer colonized by W19 was then proved in a naturally infested field, where it reduced FWB by 44% and increased yield by 35% (Wang et al., 2016). Similarly to B. amyloliquefaciens NJN-6, the strain W19 produces several antifungal metabolites, including lipopeptides (e.g., iturin, bacillomycin D, and surfactin), 18 VOCs (Wang et al., 2013), and indole-3-acetic acid (Wang et al., 2016). Interestingly, banana root exudates seemed to enhance the ability of this strain to colonize roots by augmenting bacteria biofilm formation, due to surfactin production (Wang et al., 2016).
Munimbazi and Bullerman (1998) reported on extracellular antifungal metabolites produced by B. pumilus which inhibited the mycelial growth of different strains of Aspergillus sp., Paenibacillus sp., and Fusarium sp. The chitinolytic B. pumilus strain CH4 caused inhibition of Foc mycelial growth under in vitro conditions, but its effectiveness in planta has not been tested yet (Nawangsih and Purba, 2013).
The interest of B. thuringiensis as a BCA has been mainly focused on the Cry protein and its effect as bio-insecticide, a topic outside the scope of this review (Bravo et al., 2013, 2017). Nevertheless, it has also potential in biocontrol of phytopathogenic fungi due to the chitinase production. In fact, two chitinolytic strains of B. thuringiensis (50E and 48F) caused complete growth inhibition of Foc R4 in vitro. Indeed, uneven thickening and swelling of hyphae tips were observed at the sites where interaction with the bacterial crude chitinase took place (Usharani and Gowda, 2011).
Pseudomonas spp.: The Metabolically-Versatile Biological Control Agents
Pseudomonas is a genus that comprises more than a hundred species (Mulet et al., 2010; Loper et al., 2012; Hesse et al., 2018). Many Pseudomonas spp. strains are indigenous inhabitants of the plant endosphere, rhizosphere, and/or phyllosphere, mostly established in these niches as commensals. Some of them are able to suppress the deleterious effects caused by different phytopathogens thereby promoting plant growth and health, and have thus been successfully used as plant inoculants (Arshad and Frankenberger, 1997; Haas and Défago, 2005; Mercado-Blanco and Bakker, 2007; Lugtenberg and Kamilova, 2009; Pliego et al., 2011; Schreiter et al., 2018). These bacteria displayed characteristics such as: (i) high colonization competence for plant surface, internal plant tissues (endophytism), and/or phytopathogen structures; (ii) versatility in the production of antibiotics suppressing diverse phytopathogens; (iii) ability to use specific nutrients in the target niche like plant exudates, that enable them to outcompete many components of the plant-associated microbiota; and (iv) capability to trigger (systemic) defense responses in the host plants (Mercado-Blanco, 2015, and references therein).
A relatively large number of Pseudomonas spp. strains have been studied as antagonists of Foc, the majority of which has been focused on species of the fluorescens group. A screening for the antagonistic activity of fluorescent pseudomonads isolated from the rhizoplane of several crops led to the identification of P. fluorescens Pf1 (Vidhyasekaran and Muthamilan, 1995). Since then, this strain has been studied, and proved effective, on several plant diseases, such as Fusarium wilt of chickpea (F. oxysporum f. sp. ciceris) (Vidhyasekaran and Muthamilan, 1995), rice blast (Pyricularia oryzae) (Vidhyasekaran et al., 1997), and rice sheath blight (Rhizoctonia solani) (Vidhyasekaran and Muthamilan, 1999; Nandakumar et al., 2001). Pseudomonas fluorescens Pf1 produces siderophores, hydrogen cyanide, the antibiotics 2,4-diacetylphloroglucinol (DAPG) and pyoluteorin, and induces resistance-associated enzymes (e.g., PO and PPO) in banana roots (Akila et al., 2011; Selvaraj et al., 2014). Repeated field trials have also demonstrated the effectiveness of P. fluorescens Pf1 against FWB, using diverse application protocols and formulations. Carriers such as wheat bran-saw dust (Raguchander et al., 1997), paring and pralinage, capsule (Raguchander et al., 2000; Rajappan et al., 2002), and talc (Rajappan et al., 2002; Saravanan et al., 2004b) have been explored. However, it is known that liquid formulations of P. fluorescens offer numerous advantages over solid formulations, e.g., high cell count, zero contamination, longer shelf life, greater protection against environmental stresses, and increased field efficacy (Hegde, 2002). Different chemicals, such as trehalose, polyvinylpyrrolidone, and glycerol, were tested for the development of a liquid formulation, and glycerol supported the highest Pf1 survival until 6 months of storage (Manikandan et al., 2010). A liquid formulation of Pf1 provided good results against FWB in multiple trials and locations (Selvaraj et al., 2014).
Saravanan et al. (2004a) reported a significant in vitro inhibitory effect on Foc R1 when testing five different P. fluorescens strains (Pf1, Pf2, Pf3, Pf4, and Pfm) isolated from banana rhizosphere, with the strain Pfm showing the highest antagonist effect against the pathogen growth. When greenhouse experiments were conducted using a talc-based formulation of strain Pfm, a significant reduction in vascular discoloration of the banana rhizome was observed (Saravanan et al., 2004a). Additionally, P. fluorescens Pfm systemically induced the accumulation of three key defense enzymes (PAL, PO, and polyphenol oxidase or PPO) in roots that contributed to induce resistance against Foc (Saravanan et al., 2004b).
Sivamani and Gnanamanickam (1988) investigated the possibility of suppressing FWB by bacterization with different P. fluorescens strains originating from roots of rice (strain Pfrl3), peanut (Pfgn), banana (Pfb), black gram leaves (Pfbg), citrus (Pfcp), and cotton (Pfco). These strains were tested for their in vitro antagonism ability against Foc R1 and R4. Results showed that strain Pfcp showed maximum inhibition of Foc mycelial growth, and was chosen to bacterize seedlings of Musa balbisiana. Seedlings treated with Pfcp showed less severe wilting symptoms and internal discoloration under greenhouse conditions. In addition, they also showed enhanced root growth and overall plant height.
In another study, 11 strains of P. fluorescens isolated from the banana rhizosphere were tested for their in vitro antagonistic effect against Foc. Among the tested isolates, strain Pf10 was the most effective in inhibiting the pathogen mycelial growth (Thangavelu et al., 2001). Further studies were focused on the effect of strain Pf10 treatment and Foc inoculation on the induction of banana plant enzymes and compounds known to be related with defense responses (e.g., PAL, POX, chitinase, β-1,3 glucanase and phenolics) (Thangavelu et al., 2003).
In vitro growth inhibition of Foc was also observed with P. fluorescens strain IIHRPf12. In greenhouse experiments using banana cv. Neeypovan, this strain reduced Foc colonization and FWB severity symptoms. Interestingly, structural modifications in the cortical cells at the site of fungal entry were observed, indicating that bacterized root cells were somehow “alerted” to mobilize a number of defense structures aiming to hinder the pathogen progress (Mohandas et al., 2004).
Goswami et al. (2015) reported on the P. aeruginosa strain BG, isolated from marine water from the Gulf of Khambhat in Gujarat, and its ability to inhibit Foc growth under in vitro conditions. Pseudomonas aeruginosa BG displayed plant growth promotion, biocontrol abilities, secretion of enzymes such as catalase, urease, and phosphatase, as well as the synthesis of metabolites such indole-3-acetic acid, siderophores, ammonia, and hydrogen cyanide. Other P. aeruginosa were also tested against Foc in vitro (Ayyadurai et al., 2006; Sekhar and Pious, 2015) and in planta (Yu C. et al., 2010).
Trichoderma spp.: Antagonists and Plant Growth Promoters
Trichoderma is a genus of asexually-reproducing fungi widely distributed in nearly all temperate and tropical soils. The sexual teleomorph (genus Hypocrea) can be found frequently, but many strains, including most biocontrol strains, have no known sexual stage. Trichoderma spp. show a wide genetic diversity, and are producers of several extracellular proteins, enzymes such as cellulase and chitinase, and more than 100 different metabolites with antibiotic activities. This genus can also parasitize a range of other fungi (e.g., R. solani). Besides antibiosis and mycoparasitism, Trichoderma-mediated biocontrol also relies on the induction of plant resistance (Harman et al., 2004). Therefore, due to their metabolite arsenal, rhizosphere-competence, and ability to stimulate plant growth, Trichoderma species have long been recognized as BCAs (Harman et al., 2004; Vinale et al., 2008; Woo et al., 2014), and they are widely studied against FWB (Supplemental Figure 2). These fungi are efficient colonizers of plant roots, where they establish intense interactions with plants (Vinale et al., 2008). They colonize root surfaces and invade the root epidermis, usually not beyond the first or second layer of cells (Yedidia et al., 2000), but some authors have claimed the endophytic nature of the strains tested in their research, even in banana (Caballero Hernández et al., 2013; Thangavelu and Gopi, 2015a; Chaves et al., 2016).
A rhizospheric strain, namely T. viride NRCB1, was identified in vitro among 37 isolates and tested in the pot- and field-experiments, where it reduced FWB by a maximum of 75–80% in terms of external symptoms and vascular browning (Thangavelu and Mustaffa, 2010). The authors proved that a bioformulation based on the rice chaffy grains conferred a higher efficacy over the talc cum powder formulation. Finally, the strain was able to induce the peroxidase (PO), phenylalanine ammonia lyase (PAL), and total phenolic content in treated plants (Thangavelu and Mustaffa, 2010). Later, the same authors identified a new rhizospheric strain (Trichoderma sp. NRCB3) which was combined with the endophyte Trichoderma asperellum Prr2, and successfully tested against FWB in the field (Thangavelu and Gopi, 2015a). Significant disease protection was obtained in the field using a T. viride strain applied by root dipping at transplanting and by a colonized wheat bran:saw dust mixture 3 months later. The treatment reduced FWB incidence by 75% and increased the yield by 60% (Raguchander et al., 1997). Interesting results were also obtained using a commercial product namely ECO-T® (containing T. harzianum; Plant Health Products, South Africa), though they were context-specific. In fact, the product reduced by 68% the Foc inoculum in Humic Nitisol, but only by 6% in Rhodic Ferralsol (Mukhongo et al., 2015). With the assumption that potential microbial antagonists are more abundant in soil with a history of low disease incidence, Nel et al. (2006) selected two Trichoderma spp. isolates, T22 and T5, from FWB-suppressive soil in South Africa, and obtained with them a 62% FWB control in a glasshouse (Foc R4). These strains were among the most effective strains tested in the experiment (33 strains) and were superior to a Trichoderma-based commercial product. Interesting results (efficacy higher than 80%) were also obtained on abaca (Musa textilis) and plantain (Bastasa and Baliad, 2005; Pérez Vicente et al., 2009). The literature, however, also reports examples of failure in the control of Foc (R4) using Trichoderma spp. strains, albeit resulting effective in vitro or able to delay the disease onset (Ting et al., 2003; Wibowo et al., 2013).
Arbuscular Mycorrhizal Fungi: Not Only Nutrient Providers for the Plants
The root cortical tissues enable the host plant to live in a symbiotic association (intercellularly or intracellularly) with fungi of the phylum Glomeromycota, which develop morphological structures called arbuscules and transform the infected root into the so-called “mycorrhiza.” These arbuscular mycorrhizal fungi (AMF) gain some nutrients from the plant and return others to it, thus enhancing the plant growth. Additionally, AMF protect plants against phytopathogens and abiotic stresses (Parniske, 2008; Bonfante and Genre, 2010; Lenoir et al., 2016). The AMF's beneficial effects have been also demonstrated in banana, albeit sometimes with inconstant results. While AMF are generally considered as plant growth promoters rather than pathogen antagonists, it is fascinating to see how AMF inoculated in the banana nursery could provide, in some cases, protection from FWB in the field during several subsequent weeks. Indeed, an early study reported that either Glomus intraradices or Glomus spp. stimulated the growth of banana plants cv. Grande Naine and reduced both rhizome necrosis and external FWB symptoms (Jaizme-Vega et al., 1998). Inoculation with Gigaspora margarita was found to reduce FWB in pot-grown plants cv. Maçã, especially under low concentration of Foc inoculum (Borges et al., 2007). Also, the pathogen population in roots of banana plants at 7 months after treatment with a combination of G. mosseae and T. harzianum was significantly reduced, as measured by ELISA (Mohandas et al., 2010). In the field, banana plants pre-treated in the nursery with G. clarum had higher biomass than untreated plants, and showed lower FWB incidence (67%), compared to the untreated control (88%). But plants pre-treated with G. clarum and then inoculated at transplanting with a commercial product based on P. putida and T. asperellum did not differ from the untreated control in FWB incidence and severity (Lin et al., 2012). The lack of knowledge about the interactions among those BCAs might have led to unexpected results.
It is known that plant colonization by AMF is stimulated by the soil organic matter and hindered by mineral fertilization. In banana cv. Maçã, the application of a bio-fertilizer promoted abundant mycorrhizal colonization and was associated to lower expression of FWB symptoms, compared to applications of the Hoagland solution at three concentrations (non-fertilized control was not established) (Sampaio et al., 2012). However, inconsistent results obtained with AMF have been a common experience. In a factorial experiment, the effectiveness of a commercial AMF-based product and two other commercial BCAs was context-specific. Three products were inoculated in the nursery and their efficacy against FWB evaluated on “Gros Michel” banana grown in fields with three different soil types. Rhizatech® (Dudutech, Kenya) reduced Foc inoculum by 55% in Humic Nitisol. In this soil type, PHC Biopak® (Bacillus spp.; Plant Health Care Inc., USA) reduced Foc by 47%, a level similarly reached in Vertisol (50%). ECO-T® (T. harzianum; Plant Health Products, South Africa), also showed the best efficacy in Humic Nitisol (68% disease control), while it was almost ineffective in Rhodic Ferralsol (6%) (Mukhongo et al., 2015).
Non-pathogenic Fusarium oxysporum: Beneficial Relatives
The species F. oxysporum includes pathogenic strains as well as plant beneficial endophytes and saprophytes living in soil and on organic debris (Di Pietro et al., 2003). Non-pathogenic F. oxysporum (npFo) strains are primarily recognized upon their inability to infect plants. Since about 120 formae speciales are known for F. oxysporum, npFo should be validated on as many species by pathogenicity tests. Nevertheless, it is generally accepted that a limited number of plant species is enough to declare an F. oxysporum as npFo (Fravel et al., 2003). Determining the vegetative compatibility group (VCG) cannot be used as a universal tool to identify npFo isolates. Nel et al. (2006) developed a PCR-based restriction fragment length polymorphism (RFLP) analysis of the rRNA intergenic spacer (IGS) region for discriminating npFo strains from Foc among 100 isolates obtained from banana rhizosphere in South Africa. The mechanisms underlying the biocontrol exerted by npFo strains are based on the competition for infection sites or nutrients, as well as the induction of systemic resistance (He C. Y. et al., 2002; Fravel et al., 2003; Olivain et al., 2006). The fact that npFo share the same niche with the pathogen is advantageous from the biocontrol perspective (Larkin and Fravel, 2002).
Two npFo namely Ro-3 and Ra-1 reduced FWB (Foc R1) by 80% on cv. “Rasthali” in a field trial. They were selected in vitro, proved to be effective in reducing the disease (up to 89% reduced severity) and promoting the plant growth when applied three times on both tissue-cultured and sucker-derived plants under greenhouse conditions (Thangavelu and Jayanthi, 2009). Another npFo isolate, UPM31P1, alone or in combination with S. marcescens isolate UPM39B3, resulted effective in vitro against Foc TR4, and reduced FWB under greenhouse condition. In the field, this strain only delayed the onset of FWB (Foc TR4), as it reduced the percentage of diseased plants by 75% at 15 weeks post-transplanting, but no significant difference between treated and untreated plants was observed at 28 weeks post-transplanting (Ting et al., 2009c). Plants treated with two npFo isolates obtained from disease suppressive soils in South Africa, CAV 255 and CAV 241, showed a FWB incidence reduced by 87.4 and 75.0%, respectively (Belgrove et al., 2011). In the same trial, the widely studied strain Fo47 did not suppress significantly the disease. Forsyth et al. (2006) pointed out that npFo isolates may unexpectedly be synergistic with Foc, thus they are not necessarily antagonists. In fact, although more evidence would be needed, the strain BRIP 45952 increased Fusarium wilt disease severity on “Cavendish.” Nevertheless, another isolate, BRIP 29089, reduced disease severity in artificially inoculated “Lady Finger” (Foc R1) and “Cavendish” (Foc STR4) plants. At least three more research works have dealt with npFo isolated from FWB suppressive soil and/or banana plants, though their assays remained at the laboratory stage (Nita and Harsh, 2015) or were not related to biocontrol (Nel et al., 2006; Deltour et al., 2018).
Streptomyces spp. and Other Actinomycetes: Natural Antibiotics Factories
Streptomycetes are the most important antibiotic-producing microbes. They also produce a broad range of additional secondary metabolites and lytic enzymes. For this reason, they receive attention for biotechnological, pharmaceutical and agricultural purposes. Streptomycetes are widely distributed in the soil, where they are strong competitors and antagonists. The use of streptomycetes as BCAs is largely documented in the literature and, recently, their application against plant diseases incited by Fusarium species has been reviewed by Bubici (2018). In the literature, we found only four pot trials where streptomycetes were tested against FWB (Table 1). All four experiments were conducted against Foc R4 and showed that FWB could be reduced between 46 and 87%. Interestingly, the highest disease reductions were obtained using the streptomycete fermentation broth (Qin et al., 2010; Zhou et al., 2017), compared to the experiments where spore suspensions were applied by drenching or root dipping (Cao et al., 2005; Getha et al., 2005). Qin et al. (2010) selected 8 out of 139 isolates using in vitro assays against several F. oxysporum formae speciales and demonstrated that the application of their fermentation broth provided a FWB control ranging from 78 to 87% in pot experiments. In particular, using 1.85·106 conidia mL−1 of Foc R4, plants treated with the best streptomycete strain, ZJ-E1-2, showed FWB incidence of 12%, while it was 76% on untreated trees. With a similar disease pressure, viz. 78% incidence on the control plants upon inoculation with 1·106 CFU mL−1 of Foc R4, Zhou et al. (2017) observed a FWB reduction of 73% after treatment with S. lunalinharesii B-03. The application of fermentation broth introduces into the soil both the microbial cells and their metabolites and, hence, it has a stronger impact on soil Foc inoculum than the sole microbial cells. In fact, when introduced alone, the cells must first proliferate to produce antifungal metabolites enough for effective control of Foc. Trees treated before planting with 106 CFU mL−1 of Streptomyces sp. strain S96, and later inoculated with 104 conidia mL−1 of Foc R4, showed significant reductions in FWB incidence, severity, and vascular browning. Streptomyces sp. strain S96 was selected from 131 endophytic actinomycetes isolated from surface-sterilized banana roots (Cao et al., 2005). The soil application of a spore suspension (108 CFU mL−1) of Streptomyces sp. strain g10 reduced FWB severity index by 47% and rhizome discoloration by 53% when banana plantlets were inoculated with 104 conidia mL−1 of Foc R4. The same treatment was ineffective under higher pathogen pressure, i.e., 106 conidia mL−1 (Getha et al., 2005). Streptomyces sp. strain g10 was effective in vitro against several phytopathogenic fungi, including different physiological races of Foc (Getha and Vikineswary, 2002). Nevertheless, Foc and R. solani were more resistant than other fungi (i.e., P. oryzae and Phytophthora palmivora) to the antagonistic streptomycete (Getha et al., 2004). Crude fractions containing antifungal metabolites excreted in liquid media by g10 produced swelling, distortion and excessive branching of Foc R4 hyphae, as well as inhibition of spore germination. Antibiosis-mediated Foc antagonism was also demonstrated in sterile soils for the strain g10 by using an indirect method, i.e., the paper disc method (Gunji et al., 1983).
Several articles reported on in vitro experiments with Streptomyces spp. strains, but assays under in vivo conditions to fully demonstrate their biocontrol effectiveness have not made yet (Table 2). In these studies new antifungal metabolites were discovered, such as (6S,8aS,9S,11S,12aR)-6-hydroxy-9,10-dimethyldecahydrobenzo[d]azecine-2,4,12(3H)-trione (termed as 210-A) (Wu et al., 2009), and fungichromin (Wei et al., 2011). Other three compounds were isolated from S. albospinus 15-4-2: 2-methyl-2,5,6-bornantriol, 4,4′-(3-hydroxypropane-1,1-diyl)diphenol, and 7-(4-methoxybenzyl)-4,5,6,7-tetrahydro-1,3-oxazepine-5,6-diol. These compounds did not show an inhibitory effect against Foc R4, though the streptomycete was effective against the same pathogen (Yu et al., 2011). Two other studies demonstrated the efficacy of crude culture filtrate or methanol extracts of streptomycetes, but the effective metabolites were not identified (Shih et al., 2013; Wang L. et al., 2015). Soil inoculation with S. griseus St 4 viable cells was more effective in suppressing Foc TR4 (6 log10 CFU g−1 soil of Foc) than cell-free crude extracts (7 log10 CFU g−1 soil) at 20 days after inoculation (Zacky and Ting, 2013). The formulation of S. griseus St 4 with kaolin clay, sodium alginate, or a kaolin-alginate combination increased the effectiveness of the streptomycete, compared to non-formulated cells. The kaolin clay formulation reduced Foc TR4 soil inoculum from 6 to 5.4 log10 CFU g−1 soil (Zacky and Ting, 2015).
Other Genera or Unidentified Species of Biological Control Agents
A Serratia marcescens strain, isolated from roots of wild bananas, has shown plant growth promoting effect both in glasshouse and field, and suppressed FWB, though only in the glasshouse. The loss of control efficacy stimulated the evaluation of diverse formulations in an attempt to improve its viability and efficacy in field applications. Results showed that bentonite performed better and that further advantage could come from optimization of non-fat skimmed milk and sucrose levels, whereas para-aminobenzoic acid should be omitted from bentonite formulations (Ting et al., 2009a; Ting A.S.Y. et al., 2011). Other studies in greenhouse evaluating several rhizospheric bacterial strains (FB5, FB2, T2WF, T2WC, and W10) of unidentified species culminated in the successful control of FWB (81% reduction) (Yang et al., 2006). Moderate control (16%) was obtained using Paenibacillus spp. strains RZ-17 and RZ-24 which also had additional effects on mortality and motility of Meloidogyne javanica second stage juveniles (Ribeiro et al., 2012). Multiple beneficial effects have been also reported for strains of marine rhizobacteria isolated from mangrove (YS4B1, YS1A3, and YS2A5), which were also effective against R. solanacearum and Mycosphaerella fijiensis (Bonsubre et al., 2016). Finally, a number of studies identified Foc antagonists among strains of Talaromyces spp., Eutypella sp., Paenibacillus polymyxa, Herbaspirillum spp., Tsukamurella paurometabola, Brevibacillus brevis, and Streptoverticillium lavenduligriseum (Sun et al., 2010; Manoch and Dethoup, 2011; Ting S. et al., 2011; Marín et al., 2013; Shen L. et al., 2013; Sun and Hsieh, 2015; Qi et al., 2017). A spectacular control of FWB was claimed by Chand et al. (2016), who applied a dead Foc to plant roots before inoculation with live Foc. Although no data were showed, the authors stated that inoculated plants, grown in a sick plot, did not show disease symptoms even 2 months after inoculation, while they occurred on control plants 1 week after inoculation. This approach appears as interesting as it is unusual, and no other researchers have replicated the technique against Foc until now.
Fusarium Wilt of Banana and the Banana-Associated Microbiomes
The development of next-generation sequencing (NGS) technologies, along with advanced bioinformatics tools, is rapidly increasing our knowledge on many biological processes. Diverse “-omics” techniques such as (meta)genomics, (meta)transcriptomics, proteomics, metabolomics, microbiomics, etc., are currently available to better understand plant-microbe(s) interactions from a holistic perspective (Massart et al., 2015). However, the implementation of “-omics” in the study of BCAs effective against FWB as well as their interaction with the banana-Foc pathosystem is still very scant.
The NGS-based approaches are very useful for the in-depth study of the structure, composition, and diversity of plant-associated microbiomes. Yet, microbiomics is waiting to be used in a more frequent way in the research field of banana, FWB, and biocontrol. Recent studies (16S rRNA and ITS amplicon sequencing profiling) have been focused on endophytic bacterial communities present in different parts and micro-environments of banana plants (Köberl et al., 2015, 2017; Zhai et al., 2016; Suhaimi et al., 2017), as well as in the microbiota of banana rhizosphere (Fu et al., 2016a) and soil (Xue et al., 2015; Rames et al., 2018).
Healthy plants and healthy soils have higher microbial diversity and more abundant beneficial microbes, which can improve nutrients uptake, promote plant growth, and control soil-borne diseases (Bulluck and Ristaino, 2002; Bailey and Lazarovits, 2003; Raaijmakers et al., 2009; Luan et al., 2015). Köberl et al. (2015) studied the impact of biogeography and agroforestry on the banana-associated microbiome, mostly γ-proteobacteria. Banana plants grown under agroforestry systems showed a higher abundance of potentially beneficial plant-associated bacteria and lower presence of phytopathogenic bacteria. Thus, γ-proteobacteria diversity and community members were identified as potential health indicators. Healthy plants revealed an increase in potentially beneficial microbes like Pseudomonas and Stenotrophomonas, while diseased plants showed a preferential occurrence of Enterobacteriaceae (Köberl et al., 2017). Another study correlated FWB positively with the abundance of Proteobacteria, Ascomycota, Fusarium, Cylindrocarpon, Gymnascella, Monographella, Pochonia, and Sakaguchia, but negatively with Acidobacteria, Firmicutes, Leptosphaeria, and Phaeosphaeriopsis (Shen et al., 2015a). In such pot-experiment, 2 years of biofertilizer application manipulated the composition of the rhizosphere microbial community and induced the FWB suppression. The relationship among suppression of Foc under field conditions, the use of ground cover management, and changes in the soil microbiome was also investigated in “Ducasse” banana (synonym “Pisang Awak,” ABB) (Pattison et al., 2014; Rames et al., 2018). Results showed that suppression of FWB tended to increase over time when the banana was cultivated with ground covers compared to bare soil conditions. Statistically significant changes over time in the structure of soil microbial communities in the vegetated treatment were observed, and potential biomarkers related to disease suppression were identified. In addition, fungal amplicon sequencing demonstrated that reduction of Foc in the vegetated treatment was associated with disease suppression.
Similarly, analyzing the banana/R. solanacearum pathosystem (bacterial wilt of banana or Moko), five major microbial genera were found in both symptomatic and non-symptomatic plant samples: Sphingomonas, Methylobacterium, Flavobacterium, Pseudomonas, and Ralstonia, the latter being more abundant in symptomatic (59% out of the entire genera) than in non-symptomatic plants (only 36%). In addition, several genera were only assigned to non-symptomatic plants (Suhaimi et al., 2017). Another experiment showed that the soil cultivated with tobacco and infested by R. solanacearum had lower microbial diversity than the soil free from the pathogen, which harbored more abundant beneficial microbes such as Bacillus, Agromyces, Micromonospora, Pseudonocardia, Acremonium, Lysobacter, Mesorhizobium, Microvirga, Bradyrhizobium, Acremonium, and Chaetomium. Also, the activities of catalase, invertase, and urease, as well as soil pH, available phosphorus and potassium content were lower in the infested soils (Wang et al., 2017).
Soil microbial community varies because of many factors. The main drivers of the rhizosphere microbiome are soil type and plant genotype (Berg and Smalla, 2009), but fertilizers (Ikeda et al., 2011), crop rotation (Hilton et al., 2013), and pesticides (Jacobsen and Hjelmso, 2014) may also play a significant role. Soil microbiota also fluctuates with the plant growth stages, mostly due to changes in the root exudates (Yang and Crowley, 2000; Okubo et al., 2016; Wang et al., 2017). Finally, the soil microbial assembly may be influenced by BCAs artificially introduced in the system (Xue et al., 2015). However, several studies have revealed that edaphic and anthropic factors had a deeper and more durable effect on the rhizosphere microbiota than a BCA application. For example, compared to untreated plants, R. solani had a much higher impact on lettuce rhizosphere bacterial communities than the applications of diverse BCAs such as Trichoderma sp. (Grosch et al., 2006), P. jessenii RU47 (Adesina et al., 2009), or B. amyloliquefaciens FZB42 (Chowdhury et al., 2013; Erlacher et al., 2014). On the other hand, co-inoculation of different BCAs may cause a more pronounced impact on the microbial community structure compared to the single strain application, as demonstrated in the lettuce rhizosphere (Grosch et al., 2012). Nonetheless, some systems may be more reluctant to changes, sometimes even contrasting the applied BCAs (Garbelotto et al., 2019).
The “-omics” technologies do not only provide a global overview of the banana-associated microbiota but may also yield useful information to develop more effective biological control strategies. For instance, dominant bacterial groups can be identified in FWB suppressive soils, thereby leading to the development of strains or consortia of strains serving as new biocontrol tools. For example, the above mentioned B. amyloliquefaciens strain NJN-6 was isolated from a FWB-suppressive soil after the NGS analysis had evidenced Bacillus as the dominant taxon (Xue et al., 2015). A metagenomic study has been targeted to microbes that harbor the non-ribosomal peptide-synthetase (NRPS) gene, which encodes for one of the largest groups of natural microbial secondary metabolites, such as the antibiotics vancomycin and gramicidin, as well as the lipopeptides surfactin, iturin A, and bacillomycin. The research evidenced that these microbes were more abundant in FWB-suppressive soil than in FWB-conducive soil. The main microbial taxa harboring the NRPS gene and related to FWB suppression were Pseudomonas spp. and Streptomyces spp. (Zhao et al., 2018). As potential probiotic candidates, plant vertically transmitted actinobacteria are beneficial to the growth and health of host plants (Du et al., 2018). The majority of bacteria from healthy banana shoot tips were affiliated with actinobacteria, being Mycobacterium and Nocardia the dominant taxa. The streptomycetes were isolated from shoot tips and proved to enhance the growth and resistance to Foc of pot-grown banana plants. The research elegantly presented how microbiomics can foster the selection for probiotic agents (Du et al., 2018). Another study has shown that the endophytic root microbiome of healthy banana plants was dominated by Nocardioidaceae (56.37%), Pseudonocardiaceae (14.36%) and Nocardiaceae (9.77%) (Zhai et al., 2016).
But metagenomics and microbiomics are not the sole “-omics” technologies that can help the research on biocontrol of FWB. Nevertheless, the implementation of “-omics” techniques other than microbiomics for studying biocontrol mechanisms of Foc is still absent or very limited. The study of a tripartite interaction among banana, Foc, and T. asperellum strain Prr2 is one of the few examples available, however with a non-NGS approach, viz. the suppression subtractive hybridization or SSH (Thangavelu et al., 2016).
Gaps in the Knowledge and Concluding Remarks
Banana production systems lack phytosanitary certification schemes. In Australia, where Foc TR4 is considered a quarantine pest, the organization that governs standardization and certification of agricultural practices, GlobalG.A.P., launched in 2017 an “add-on” in its standards which encourages farmers to take preventive biosecurity measures following a strict protocol (GLOBALG.A.P, 2019). However, it is only a preventive awareness measure rather than a true certification. Phytosanitary certification would be necessary either for tissue culture- or sucker-derived plants, because Foc-contamination may occur at different stages of plant production. In fact, undesired infected plants can derive not only from infected asymptomatic suckers but also from pathogen-free tissue culture plantlets, which can be later contaminated by the pathogen during the acclimation period (e.g., by contaminated irrigation water or substrate).
It is worth noting that, despite its dangerousness, Foc is not yet a quarantine pathogen in several countries, and where it is considered as such, the regulation is limited to TR4 only. For example, in countries joining to the European and Mediterranean Plant Protection Organization (EPPO), Foc is not yet reported in the A1 or A2 lists, which include pests and pathogens absent or present, respectively, in the EPPO region and recommended for quarantine regulations (EPPO, 2019). Among the Fusarium species, only F. circinatum, F. euwallaceae, F. foetens and F. oxysporum f. sp. albedinis are included in the A2 list, while F. oxysporum f. sp. lactucae is provisionally placed in an alert list because it is pending for a risk assessment that will designate or not it as a quarantine pathogen. Therefore, in the EPPO region, no evaluation for the inclusion of Foc among the quarantine pathogens seems to have initiated yet. Nevertheless, awareness campaigns, pest risk assessments, and research works have largely emphasized the importance of quarantine measures against Foc TR4 (Baker et al., 2008; Pocasangre et al., 2011; Blomme et al., 2013; Sánchez, 2013; Anses, 2018). Both quarantine measures and phytosanitary certification schemes require huge efforts by the governments in terms of legal framework, personnel training, and protocols for inspection, sampling, diagnosis, etc.
Following the pathogen exclusion and quarantine measures, which are critical to hinder the current Foc TR4 expansion, the use of pathogen-free certified plating material is certainly one of the first key preventive steps toward successful management of FWB. It has been implementing well in countries of Latin America, where almost 100% of plants derived from tissue culture, but only partially in Africa, where suckers are still largely used to establish new (especially small) plantations, very likely for economic and cultural reasons. On the other hand, the introduction of beneficial and well-characterized microorganisms during banana propagation protocols can be an excellent strategy to “prepare” (or “pre-condition,” or “prime”) plantlets to cope with Foc inoculum in the field.
A large number of studies have contributed to select BCAs with variable effectiveness against Foc, and many of them reached the field-testing stage (Table 1 and Supplemental Figure 2). Interestingly, the disease control degree obtained experimentally has been surprisingly high, even considering that Foc is a vascular pathogen enduring in the soil for a long time and, thus, it is generally considered difficult to control using BCAs. Bio-formulation, which is one of the key factors affecting the BCA efficacy, has been tested in several cases, but new aspects, such as nanotechnology and formulation of microbial consortia and/or their metabolites (Keswani et al., 2016), merit to be studied. Moreover, the effectiveness of these formulations could be improved by the amendment with specific nutrients/factors aiming to enhance the biosynthesis of these metabolites as well as the survival of the BCAs in the banana rhizosphere. Microbial metabolites have proved their value in nutrition, agriculture, and healthcare, but poorly evaluated so far against FWB. In contrast, plant extracts like A. tuberosum (Figure 3) have been repeatedly tested, especially during the last years (Singh et al., 2017). Bioformulations combining different strains, nutrients, metabolites and/or other natural products must take into account the compatibility (i.e., the absence of antagonism) of the different components (Sarma et al., 2015; Gómez-Lama Cabanás et al., 2018). Blends of microorganisms beyond their mere mixture, but based on tailor-made combinations of strains with complementary and/or synergistic modes of action are encouraged (Lutz et al., 2004). Solidly supported by the powerful currently-available “-omics” methodologies, biocontrol strategies can now aim to provide novel tools based on ad hoc tailored consortia of BCAs originating from the indigenous microbiota associated with the target plant (Gopal et al., 2013; Berg et al., 2017; Mercado-Blanco et al., 2018), thereby overcoming problems and inconsistencies frequently observed when biopesticides are based on formulations of a single microorganism or combinations of few of them. Furthermore, successful colonization and endurance in the target niche must be guaranteed as the first requisite for successful biocontrol. Therefore, a comprehensive understanding of the mode of action, ecology, and trophic interactions established upon the application of BCAs is instrumental for their success (Saraf et al., 2014; Eljounaidi et al., 2016; Shafi et al., 2017). Monitoring the microbial strains in the field after their application should be addressed in order to understand their fate in the environment and optimize their application protocol. For these aspects, “-omics”-based approaches are of great help.
The availability of the M. acuminata draft genome marked a milestone in the genetic research of banana (D'Hont et al., 2012), and the genomic sequence of several Foc races is also available. Furthermore, transcriptomics has been implemented in studies on Musa spp. (e.g., Backiyarani et al., 2015) enhancing our knowledge on physiological processes such as the fruit ripening (Asif et al., 2014) and the responses to abiotic stresses such as low-temperature (Yang Q. S. et al., 2015). This “-omics” has been largely used to study the interaction between BCAs, plant and/or microbiome, but no studies are available on the banana/Foc pathosystem. Moreover, integrating genomics, metagenomics and (meta)transcriptomics would allow understanding the microbiota structure and the roles and functions of its members, as well as the intricate interactions between BCAs, microbiota, plant and the environment. This approach has not been applied to plant microbiomes yet, but it is providing new insight in other fields like the human microbiome (Massart et al., 2015). Proteomics and metabolomics analyses have been also conducted in banana, but they have been focused mainly on the plant-pathogen interaction while BCAs have not been involved (Li et al., 2013, 2017; Lu et al., 2013; Sun et al., 2014; Ramu et al., 2016; Gopalakrishnan, 2017; Yuan et al., 2017). Therefore, at least to the best of our knowledge, there is still an important gap in our knowledge of biocontrol against Foc, including insight on the mechanisms involved in the antagonism, plant colonization, plant growth promotion, etc.
Biocontrol should not be considered as an independent tool, but adequately implemented in an integrated management framework. Actually, besides the combination with organic fertilizers, very little has been investigated on the integration of BCAs with other control means. For instance, the effect of an organic amendment against FWB was enhanced by the combination with a BCA, B. amyloliquefaciens strain NJN-6, namely biofertilizer (Shen et al., 2015a). Furthermore, the biocontrol efficacy of such biofertilizer resulted even higher when it was applied after ammonia fumigation (Shen et al., 2019). Other studies have shown that combinations of biocontrol organisms with silicon and mulching, or with neem cake can be advantageous compared to the individual applications, and therefore can provide a better control option for banana growers who have to deal with FWB in their plantations (Saravanan et al., 2003; Kidane and Laing, 2010). Diverse combinations of treatments with silicon, T. harzianum, compost, various sources of nitrogen, phosphorus and potassium, and the cover crop Crotalaria juncea were applied in the field to banana varieties differing in the susceptibility level to Foc. The results highlighted the advantages of integrated disease management, especially the combination of different control means with the host genetic resistance. In fact, the treatment including all the control means was more effective than those including only some of them. Also, while in the highly susceptible cultivar Silk (AAB) the treatment was not effective in reducing FWB during the first crop cycle, in the moderately susceptible variety Prata Anã (AAB) it reduced the disease by 58%, with a yield increment of 157.3% (Haddad et al., 2018).
We showed that the literature offers numerous examples of encouraging results, suggesting that biocontrol can greatly contribute to limit the damage caused by FWB. More efforts should be done to further validate the currently available outcomes, to deepen the knowledge on the most valuable BCAs, and to improve their efficacy by setting up effective formulations, application protocols, and integrated strategies.
Author Contributions
All the authors wrote sections of the manuscript, contributing equally to its first draft. GB coordinated and merged the individual contributions from the authors. All the authors read, revised, and approved the submitted version.
Funding
This review was supported by the Horizon 2020 Project entitled Microbial Uptakes for Sustainable Management of Major Banana Pests and Diseases (MUSA; grant number 727624).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2019.00616/full#supplementary-material
Supplemental Figure 1. Publication timeline of scientific articles dealing with Fusarium wilt of banana. Articles were retrieved from the CAB Direct database (1970–2018) by searching the keywords “Fusarium cubense” or “Panama disease” in the title and abstract (735 articles). Foc: Fusarium oxysporum f. sp. cubense; FWB: Fusarium wilt of banana.
Supplemental Figure 2. Biological control agents studied for the control of Fusarium oxysporum f. sp. cubense. The numbers indicate the scientific articles retrieved from CAB Direct database (1970–2018) by searching the keywords “Fusarium cubense” or “Panama disease” in the title and abstract.
Supplemental Table 1. List of in vitro experiments conducted against Fusarium oxysporum f. sp. cubense (Foc) using beneficial microorganisms.
References
Adesina, M. F., Grosch, R., Lembke, A., Vatchev, T. D., and Smalla, K. (2009). In vitro antagonists of Rhizoctonia solani tested on lettuce: rhizosphere competence, biocontrol efficiency and rhizosphere microbial community response. FEMS Microbiol. Ecol. 69, 62–74. doi: 10.1111/j.1574-6941.2009.00685.x
Aguilar, E. A., Turner, D. W., and Sivasithamparam, K. (2000). Fusarium oxysporum f. sp. cubense inoculation and hypoxia alter peroxidase and phenylalanine ammonia lyase activities in nodal roots of banana cultivars (Musa sp.) differing in their susceptibility to Fusarium wilt. Aust. J. Bot. 48, 589–596. doi: 10.1071/BT99009
Akila, R., Rajendran, L., Harish, S., Saveetha, K., Raguchander, T., and Samiyappan, R. (2011). Combined application of botanical formulations and biocontrol agents for the management of Fusarium oxysporum f. sp. cubense (Foc) causing Fusarium wilt in banana. Biol. Control 57, 175–183. doi: 10.1016/j.biocontrol.2011.02.010
Aloo, B. N., Makumba, B. A., and Mbega, E. R. (2019). The potential of Bacilli rhizobacteria for sustainable crop production and environmental sustainability. Microbiol. Res. 219, 26–39. doi: 10.1016/j.micres.2018.10.011
Alvarez, C. E., Garcia, V., Robles, J., and Diaz, A. (1981). Influence of soil characteristics on the incidence of Panama disease. Fruits 36, 71–81.
Anses (2018). “Risque Phytosanitaire Portant sur Fusarium oxysporum f. sp. cubense Pour les Départements D'outre-mer”, Report. Available online at: https://www.anses.fr/fr/system/files/SANTVEG2017SA0050Ra.pdf (accessed February 20, 2019).
Araujo, E. T. D., Melo, W. D. C., Guerreiro, M. C., Barbiéri, R. S., and Abreu, C. M. P. D. (2004). Biocidal effect of organotin mandelates on Fusarium oxysporum f. sp. cubense. Ciênc. Agrotecnol, 28, 34–41. doi: 10.1590/S1413-70542004000100004
Arshad, M., and Frankenberger, W. T. (1997). Plant growth-regulating substances in the rhizosphere: microbial production and functions. Adv. Agron. 62, 45–151. doi: 10.1016/S0065-2113(08)60567-2
Asif, M. H., Lakhwani, D., Pathak, S., Gupta, P., Bag, S. K., Nath, P., et al. (2014). Transcriptome analysis of ripe and unripe fruit tissue of banana identifies major metabolic networks involved in fruit ripening process. BMC Plant Biol. 14:316. doi: 10.1186/s12870-014-0316-1
Ayyadurai, N., Naik, P. R., Rao, M. S., Kumar, R. S., Samrat, S. K., Manohar, M., et al. (2006). Isolation and characterization of a novel banana rhizosphere bacterium as fungal antagonist and microbial adjuvant in micropropagation of banana. J. Appl. Microbiol. 100, 926–937. doi: 10.1111/j.1365-2672.2006.02863.x
Azkolain Olaondo, J. A. (2016). Biofumigación y Biosolarización Para el Manejo del mal de Panamá en la Platanera de Canarias. [Master's thesis]. Pamplona: ETSIA, Universidad Pública de Navarra.
Backiyarani, S., Uma, S., Saraswathi, M. S., Saravanakumar, A. S., and Chandrasekar, A. (2015). Transcriptome analysis of banana (Musa balbisiana) based on next-generation sequencing technology. Turk. J. Agric. For. 39, 705–717. doi: 10.3906/tar-1406-171
Bailey, K. L., and Lazarovits, G. (2003). Suppressing soil-borne diseases with residue management and organic amendments. Soil Tillage Res. 72, 169–180. doi: 10.1016/S0167-1987(03)00086-2
Baker, R., Caffier, D., Choiseul, J. W., De Clercq, P., Dormannsné-Simon, E., Gerowitt, B., et al. (2008). Scientific opinion of the panel on plant health on a request from the European Commission on pest risk assessment made by France on Fusarium oxysporum f. sp. cubense considered by France as harmful in French overseas departments of French Guiana, Guadeloupe, Martinique and Réunion. EFSA J. 668, 1–22. doi: 10.2903/j.efsa.2008.668
Bancroft, J. (1876). Report of the board appointed to enquire into the cause of disease affecting livestock and plants. Votes Proc. 3, 1011–1038.
Bastasa, G. N., and Baliad, A. A. (2005). Biological control of Fusarium wilt of abaca (Fusarium oxysporum) with Trichoderma and yeast. Philipp. J. Crop Sci. 30, 29–37.
Belgrove, A., Steinberg, C., and Viljoen, A. (2011). Evaluation of nonpathogenic Fusarium oxysporum and Pseudomonas fluorescens for Panama disease control. Plant Dis. 95, 951–959. doi: 10.1094/PDIS-06-10-0409
Bentley, S., Moore, N. Y., Pegg, K. G., Gerlach, K. S., and Smith, L. J. (2001). “Genetic characterization and detection of Fusarium wilt,” in Proceedings of the International Workshop ‘Banana Fusarium Wilt Management: Towards Sustainable Cultivation’, eds A. B. Molina, N. H. Nik Masdesk, and K. W. Liew (Los Baños: INIBAP-ASPNET), 143–151.
Berg, G., Köberl, M., Rybakova, D., Müller, H., Grosch, R., and Smalla, K. (2017). Plant microbial diversity is suggested as the key to future biocontrol and health trends. FEMS Microbiol. Ecol. 93:fix50 doi: 10.1093/femsec/fix050
Berg, G., and Smalla, K. (2009). Plant species and soil type cooperatively shape the structure and function of microbial communities in the rhizosphere. FEMS Microbiol. Ecol. 68, 1–13. doi: 10.1111/j.1574-6941.2009.00654.x
Bioversity International (2018). Banana Regional Research Networks (Montpellier, France: Bioversity International). Available online at: http://banana-networks.org/ (accessed December 3, 2018).
Blomme, G., Ploetz, R., Jones, D., De Langhe, E., Price, N., Gold, C., et al. (2013). A historical overview of the appearance and spread of Musa pests and pathogens on the African continent: highlighting the importance of clean Musa planting materials and quarantine measures. Ann. Appl. Biol. 162, 4–26. doi: 10.1111/aab.12002
Bonfante, P., and Genre, A. (2010). Mechanisms underlying beneficial plant-fungus interactions in mycorrhizal symbiosis. Nature Commun. 1:48. doi: 10.1038/ncomms1046
Bonsubre, J. A., Montaos, E., Cadiz, D. A. J., Beldad, F. D., Alburo, G., Papa, I., et al. (2016). “Preventing moko, Panama, and black Sigatoka diseases in bananas using rhizobacteria from mangrove rhizospheres as biological control agent,” in Proceedings of the ‘4th Asian PGPR Conference - Recent Trends in PGPR Research for Sustainable Crop Productivity’ (Hanoi: Asian PGPR Society), 229–237.
Borges, A. A., Borges-Pérez, A., and Fernández-Falcón, M. (2004). Induced resistance to Fusarial wilt of banana by menadione sodium bisulphite treatments. Crop Prot. 23, 1245–1247. doi: 10.1016/j.cropro.2004.05.010
Borges, A. J. D. S., Trindade, A. V., Matos, A. P. D., and Peixoto, M. D. F. D. S. (2007). Reduction of Fusarium wilt of “banana-Maçã” by inoculation of arbuscular mycorrhizal fungi. Pesq. Agropec. Bras. 42, 35–41. doi: 10.1590/S0100-204X2007000100005
Bravo, A., Gómez, I., Porta, H., García-Gómez, B. I., Rodriguez-Almazan, C., Pardo, L., et al. (2013). Evolution of Bacillus thuringiensis Cry toxins insecticidal activity. Microb. Biotechnol. 6, 17–26. doi: 10.1111/j.1751-7915.2012.00342.x
Bravo, A., Pacheco, S., Gómez, I., Garcia-Gómez, B., Onofre, J., and Soberón, M. (2017). “Insecticidal proteins from Bacillus thuringiensis and their mechanism of action,” in Bacillus thuringiensis and Lysinibacillus sphaericus: Characterization and Use in the Field of Biocontrol, eds L. M. Fiuza, R. A. Polanczyk, and N. Crickmore (Cham: Springer International Publishing), 53–66. doi: 10.1007/978-3-319-56678-8_4
Bubici, G. (2018). Streptomyces spp. as biological control agents against Fusarium species. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 13:50. doi: 10.1079/PAVSNNR201813050
Bubici, G., and Cirulli, M. (2008). “Integrated management of Verticillium wilt of tomato,” in Integrated Management of Diseases Caused by Fungi, Phytoplasma and Bacteria, eds A. Ciancio, and K. G. Mukerji (Dordrecht: Springer), 225–242. doi: 10.1007/978-1-4020-8571-0_12
Buddenhagen, I. (2009). Understanding strain diversity in Fusarium oxysporum f. sp. cubense and history of introduction of “Tropical Race 4” to better manage banana production. Acta Hortic. 193–204. doi: 10.17660/ActaHortic.2009.828.19
Buddenhagen, I. W. (1990). “Banana breeding and Fusarium wilt,” in Fusarium Wilt of Banana, ed R.C. Ploetz (St. Paul, MN: The American Phytopathological Society), 107–113.
Bulluck, L. R., and Ristaino, J. B. (2002). Effect of synthetic and organic soil fertility amendments on southern blight, soil microbial communities, and yield of processing tomatoes. Phytopathology 92, 181–189. doi: 10.1094/PHYTO.2002.92.2.181
CAB Direct (2018). CAB Direct Database (Wallingford: CAB International). Available online at: https://www.cabdirect.org/ (accessed October 4, 2018).
Caballero Hernández, Á. J., Pocasangre Enamorado, L. E., Casanoves, F., Avelino, J., Tapia Fernández, A. C., and Ortiz, J. L. (2013). Use of endophytic insulation of Trichoderma spp., for biocontrol of Panama disease (Fusarium oxysporum f. sp. cubense) race 1, in vitro plants of banana, Gros Michel variety (AAA) under greenhouse. La Calera 13, 16–23. doi: 10.5377/calera.v13i20.1620
Cao, L., Qiu, Z., Dai, X., Tan, H., Lin, Y., and Zhou, S. (2004). Isolation of endophytic actinomycetes from roots and leaves of banana (Musa acuminata) plants and their activities against Fusarium oxysporum f. sp. cubense. World J. Microbiol. Biotechnol. 20, 501–504. doi: 10.1023/B:WIBI.0000040406.30495.48
Cao, L., Qiu, Z., You, J., Tan, H., and Zhou, S. (2005). Isolation and characterization of endophytic streptomycete antagonists of Fusarium wilt pathogen from surface-sterilized banana roots. FEMS Microbiol. Lett. 247, 147–152. doi: 10.1016/j.femsle.2005.05.006
Carillo, C. A. (2014). “Davao eyes oil palm as alternative export crop,” in Business World online (Manila, The Philippines: BusinessWorld Publishing). Available online at: http://www.bworldonline.com/content.php?section=Economy&title=davao-eyes-oil-palm-as-alternative-export-crop&id=85863 (accessed December 22, 2018).
Cawoy, H., Debois, D., Franzil, L., De Pauw, E., Thonart, P., and Ongena, M. (2015). Lipopeptides as main ingredients for inhibition of fungal phytopathogens by Bacillus subtilis/amyloliquefaciens. Microb. Biotechnol. 8, 281–295. doi: 10.1111/1751-7915.12238
Cawoy, H., Mariutto, M., Henry, G., Fisher, C., Vasilyeva, N., Thonart, P., et al. (2014). Plant defense stimulation by natural isolates of bacillus depends on efficient surfactin production. Mol. Plant Microbe Interact. 27, 87–100. doi: 10.1094/MPMI-09-13-0262-R
Chand, G., Kumar, A., Kumar, S., Gupta, R. N., Jaiswal, U. S., Maru, A. K., et al. (2016). Induction of resistance against Fusarium wilt of banana by application of live RKN, live and dead pathogenic strain of Fusarium oxysporum f. sp. cubense. J. Pure Appl. Microbiol. 10, 2307–2314.
Chaves, N. P., Staver, C., and Dita, M. A. (2016). Potential of Trichoderma asperellum for biocontrol of Fusarium wilt in banana. Acta Hortic. 1114, 261–265. doi: 10.17660/ActaHortic.2016.1114.35
Chen, X. H., Koumoutsi, A., Scholz, R., Eisenreich, A., Schneider, K., Heinemeyer, I., et al. (2007). Comparative analysis of the complete genome sequence of the plant growth-promoting bacterium Bacillus amyloliquefaciens FZB42. Nat. Biotechnol. 25, 1007–1014. doi: 10.1038/nbt1325
Chowdhury, S. P., Dietel, K., Randler, M., Schmid, M., Junge, H., Borriss, R., et al. (2013). Effects of Bacillus amyloliquefaciens FZB42 on lettuce growth and health under pathogen pressure and its impact on the rhizosphere bacterial community. PLoS ONE 8:e68818. doi: 10.1371/journal.pone.0068818
Ciancio, A. (2017). Microbial Uptakes for Sustainable Management of Major Banana Pests and Diseases (MUSA). Available online at: http://www.projectmusa.eu/ (accessed December 3, 2018).
Cirulli, M., Bubici, G., Amenduni, M., Armengol, J., Berbegal, M., Jiménez-Gasco, M. D., et al. (2010). Verticillium wilt: a threat to artichoke production. Plant Dis. 94, 1176–1187. doi: 10.1094/PDIS-12-09-0852
Clercx, L. (2013). “Task force on Fusarium wilt tropical race 4 (TR4),” in World Banana Forum (Rome: Food and Agriculture Organization of the United Nations, FAO). Available online at: http://www.fao.org/world-banana-forum/projects/fusarium-tr4/en/ (accessed December 3, 2018).
Compant, S., Saikkonen, K., Mitter, B., Campisano, A., and Mercado-Blanco, J. (2016). Editorial special issue: soil, plants and endophytes. Plant Soil 405, 1–11. doi: 10.1007/s11104-016-2927-9
Conde, B. D., and Pitkethley, R. N. (2001). “The discovery, identification and management of banana Fusarium wilt outbreaks in the Northern Territory of Australia,” in Proceedings of the International Workshop ‘Banana Fusarium Wilt Management: Towards Sustainable Cultivation’, eds A. B. Molina, N. H. Nik Masdesk, and K. W. Liew (Los Baños: INIBAP-ASPNET), 260–265.
Cook, D. C., Taylor, A. S., Meldrum, R. A., and Drenth, A. (2015). Potential economic impact of Panama Disease (tropical race 4) on the Australian banana industry. J. Plant Dis. Prot. 122, 229–237. doi: 10.1007/BF03356557
da Silva Junior, J. F., Cordeiro, Z. J. M., and Oliveira, A. M. G. (2000). Soil chemical parameters in relation to the incidence and intensity of Panama disease. Infomusa 9, 13–16.
Damodaran, T., Rajan, S., Mishra, V. K., Jha, S. K., Ahmad, I., and Gopal, R. (2018). First report of Fusarium wilt in banana caused by Fusarium oxysporum f. sp. cubense tropical race 4 in India. Plant Dis. doi: 10.1094/PDIS-07-18-1263-PDN
Datnoff, L. E., and Rodrigues, F. A. (2015). Silicon and Plant Diseases, eds. F. A. Rodrigues and L. E. Datnoff Cham: Springer International Publishing. doi: 10.1007/978-3-319-22930-0_1
Davis, A. J., and Grant, B. R. (1996). The effect of phosphonate on the sporulation of Fusarium oxysporum f. sp. cubense. Australas. Plant Pathol. 25, 31–35. doi: 10.1071/AP96007
Davis, A. J., Say, M., Snow, A. J., and Grant, B. R. (1994). Sensitivity of Fusarium oxysporum f. sp. cubense to phosphonate. Plant Pathol. 43, 200–205. doi: 10.1111/j.1365-3059.1994.tb00571.x
De Vrieze, M., Germanier, F., Vuille, N., and Weisskopf, L. (2018). Combining different potato-associated Pseudomonas strains for improved biocontrol of Phytophthora infestans. Front. Microbiol. 9:2573. doi: 10.3389/fmicb.2018.02573
Deltour, P., França, S. C., Heyman, L., Pereira, O. L., and Höfte, M. (2018). Comparative analysis of pathogenic and nonpathogenic Fusarium oxysporum populations associated with banana on a farm in Minas Gerais, Brazil. Plant Pathol. 67, 707–718. doi: 10.1111/ppa.12757
D'Hont, A., Denoeud, F., Aury, J.-M., Baurens, F.-C., Carreel, F., Garsmeur, O., et al. (2012). The banana (Musa acuminata) genome and the evolution of monocotyledonous plants. Nature 488, 213–217. doi: 10.1038/nature11241
Di Pietro, A., Madrid, M. P., Caracuel, Z., Delgado-Jarana, J., and Roncero, M. I. (2003). Fusarium oxysporum: exploring the molecular arsenal of a vascular wilt fungus. Mol. Plant Pathol. 4, 315–325. doi: 10.1046/j.1364-3703.2003.00180.x
Dita, M., Barquero, M., Heck, D., Mizubuti, E. S. G., and Staver, C. P. (2018). Fusarium wilt of banana: current knowledge on epidemiology and research needs toward sustainable disease management. Front. Plant Sci. 9:1468. doi: 10.3389/fpls.2018.01468
Dita, M. A., Garming, H., Bergh, I. V. D., Staver, C., and Lescot, T. (2013). Banana in Latin America and the Caribbean: current state, challenges and perspectives. Acta Hortic. 986, 365–380. doi: 10.17660/ActaHortic.2013.986.39
Domínguez-Hernández, J., Negrín, M. A., and Rodríguez, C. M. (2008). Soil potassium indices and clay-sized particles affecting banana-wilt expression caused by soil fungus in banana plantation development on transported volcanic soils. Commun. Soil Sci. Plant Anal. 39, 397–412. doi: 10.1080/00103620701826522
du Plessis, M. C. (1994). Biological control of Fusarium oxysporum f. sp. cubense. Inligtingsbulletin - Instituut vir Tropiese en Subtropiese Gewasse. 262, 12–13.
Du, X., Zhai, Y., Deng, Q., Tan, H., and Cao, L. (2018). Illumina-based sequencing analysis directed selection for Actinobacterial probiotic candidates for banana plants. Probiotics Antimicrob. Proteins 10, 284–292. doi: 10.1007/s12602-017-9293-7
Dubois, T., Dusabe, Y., Lule, M., Van Asten, P., Coyne, D., Hobayo, J. C., et al. (2013). Tissue culture banana (Musa spp.) for smallholder farmers: lessons learnt from East Africa. Acta Hortic. 986, 51–59. doi: 10.17660/ActaHortic.2013.986.3
Duffy, B. K., and Défago, G. (1997). Zinc improves biocontrol of Fusarium crown and root rot of tomato by Pseudomonas fluorescens and represses the production of pathogen metabolites inhibitory to bacterial antibiotic biosynthesis. Phytopathology 87, 1250–1257. doi: 10.1094/PHYTO.1997.87.12.1250
Dusunceli, F. (2017). Global Programme on Banana Fusarium Wilt Disease: Protecting Banana Production From the Disease With Focus on Tropical Race 4 (TR4). Rome: FAO. Available online at: http://www.fao.org/3/a-i7921e.pdf (accessed December 3, 2018).
Eljounaidi, K., Lee, S. K., and Bae, H. (2016). Bacterial endophytes as potential biocontrol agents of vascular wilt diseases - Review and future prospects. Biol. Control 103, 62–68. doi: 10.1016/j.biocontrol.2016.07.013
EPPO (2019). EPPO A1 List of Pests Recommended for Regulation as Quarantine Pests. Available online at: https://www.eppo.int/ACTIVITIES/plant_quarantine/A1_list (accessed February 20, 2019).
EPPO Reporting Service (2018). Incursion and Eradication of Fusarium oxysporum f. sp. cubense Tropical Race 4 From Israel. Available online at: https://gd.eppo.int/reporting/article-6300 (accessed December 5, 2018).
Erlacher, A., Cardinale, M., Grosch, R., Grube, M., and Berg, G. (2014). The impact of the pathogen Rhizoctonia solani and its beneficial counterpart Bacillus amyloliquefaciens on the indigenous lettuce microbiome. Front. Microbiol. 5:175. doi: 10.3389/fmicb.2014.00175
Eswaramurthy, S., Muthusamy, M., Muthusamy, S., Jayasekar, R. R., and Natarajan, S. (1988). Effect of Bavistin, aureofungin application on Panama wilt and Sigatoka leaf spot of banana. Hindustan Antibiot. Bull. 30, 25–26.
FAO (2016). All About Bananas: Things You Should Know About the Tropical Fruit. Rome: Food and Agriculture Organization of the United Nations, FAO. Available online at: http://www.fao.org/zhc/detail-events/en/c/446573/ (accessed December 3, 2018).
FAO (2018a). Banana Facts and Figures. Rome, Italy: Food and Agriculture Organization of the United Nations, FAO. Available online at: http://www.fao.org/economic/est/est-commodities/bananas/bananafacts/en/#.XAUCkGhKhPY (accessed December 3, 2018).
FAO (2018b). FAOSTAT. Rome, Italy: Food and Agriculture Organization of the United Nations, FAO. Available online at: http://www.fao.org/faostat/en/#home (accessed November 14, 2018).
FAO (2018c). Fusarium Wilt TR4 - The Caribbean Carries Out Awareness-Raising and Training Activities. Rome: Food and Agriculture Organization of the United Nations, FAO. Available online at: http://www.fao.org/agriculture/crops/news-events-bulletins/detail/en/item/235419/ (accessed November 30, 2018).
Farquhar, I. (2012). Bananas in China. Banana Link. Available online at: http://www.bananalink.org.uk/sites/default/files/Bananas%20in%20China%202012.pdf (accessed December 5, 2018).
Fernández-Falcón, M., Borges, A. A., and Borges-Pérez, A. (2003). Induced resistance to Fusarium wilt of banana by exogenous applications of indoleacetic acid. Phytoprotection 84, 149–153. doi: 10.7202/008492ar
Fernández-Falcón, M., Borges, A. A., and Borges-Pérez, A. (2004). Response of Dwarf Cavendish banana plantlets to inoculation with races 1 and 4 of Fusarium oxysporum f. sp. cubense at different levels of Zn nutrition. Fruits 59, 319–323. doi: 10.1051/fruits:2004030
Figueroa, L. (1987). Control of diseases and nematodes of banana and plantain. Rev. Mex. Fitopatol. 5, 181–184.
Fira, D., Dimkic, I., Beric, T., Lozo, J., and Stankovic, S. (2018). Biological control of plant pathogens by Bacillus species. J. Biotechnol. 285, 44–55. doi: 10.1016/j.jbiotec.2018.07.044
Forsyth, L. M., Smith, L. J., and Aitken, E. A. B. (2006). Identification and characterization of non-pathogenic Fusarium oxysporum capable of increasing and decreasing Fusarium wilt severity. Mycol. Res. 110, 929–935. doi: 10.1016/j.mycres.2006.03.008
Fortunato, A. A., Rodrigues, F. Á., Baroni, J. C. P., Soares, G. C. B., Rodriguez, M. A. D., and Pereira, O. L. (2012b). Silicon suppresses Fusarium wilt development in banana plants. J. Phytopathol. 160, 674–679. doi: 10.1111/jph.12005
Fortunato, A. A., Rodrigues, F. Á., and Nascimento, K. J. T. D. (2012a). Physiological and biochemical aspects of the resistance of banana plants to Fusarium wilt potentiated by silicon. Phytopathology 102, 957–966. doi: 10.1094/PHYTO-02-12-0037-R
Fortunato, A. A., Silva, W. L. D., and Rodrigues, F. Á. (2014). Phenylpropanoid pathway is potentiated by silicon in the roots of banana plants during the infection process of Fusarium oxysporum f. sp. cubense. Phytopathology 104, 597–603. doi: 10.1094/PHYTO-07-13-0203-R
Fravel, D., Olivain, C., and Alabouvette, C. (2003). Fusarium oxysporum and its biocontrol. New Phytol. 157, 493–502. doi: 10.1046/j.1469-8137.2003.00700.x
FruiTrop (2018). FruiTrop Online, Fruit & Vegetables Market Expertise. Available online at: http://www.fruitrop.com/en/Produits/Banana (accessed November 30, 2018).
Fu, L., Penton, C., Ruan, Y., Shen, Z., Xue, C., Rong, L., et al. (2016a). Inducing the rhizosphere microbiome by biofertilizer application to suppress banana Fusarium wilt disease. Soil Biol. Biochem. 104, 39–48. doi: 10.1016/j.soilbio.2016.10.008
Fu, L., Ruan, Y., Tao, C., Li, R., and Shen, Q. (2016b). Continous application of bioorganic fertilizer induced resilient culturable bacteria community associated with banana Fusarium wilt suppression. Sci. Rep. 6, 27731–27731. doi: 10.1038/srep27731
Garbelotto, M., Lowell, N., Chen, I. Y., and Osmundson, T. W. (2019). Evidence for inhibition of a fungal biocontrol agent by a plant microbiome. J. Plant Pathol. 102. doi: 10.1007/s42161-019-00247-0
Getha, K., and Vikineswary, S. (2002). Antagonistic effects of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f. sp. cubense race 4: indirect evidence for the role of antibiosis in the antagonistic process. J. Ind. Microbiol. Biotechnol. 28, 303–310. doi: 10.1038/sj.jim.7000247
Getha, K., Vikineswary, S., Wong, W. H., Seki, T., Ward, A., and Goodfellow, M. (2004). Characterization of selected isolates of indigenous Streptomyces species and evaluation of their antifungal activity against selected plant pathogenic fungi. Malay. J. Sci. 23, 37–47.
Getha, K., Vikineswary, S., Wong, W. H., Seki, T., Ward, A., and Goodfellow, M. (2005). Evaluation of Streptomyces sp. strain g10 for suppression of Fusarium wilt and rhizosphere colonization in pot-grown banana plantlets. J. Ind. Microbiol. Biotechnol. 32, 24–32. doi: 10.1007/s10295-004-0199-5
Gittleson, K. (2018). Battling to Save the World's Bananas. Available online at: https://www.bbc.com/news/business-42777803 (accessed December 3, 2018).
GLOBALG.A.P (2019). TR4 Biosecurity Add-on For Bananas. Available online at: https://www.globalgap.org/ (accessed February 20, 2019).
Gobierno de Canarias (2018). Producciones Agrícolas del Plátano. Available online at: http://www.gobiernodecanarias.org/agricultura/sgt/temas/estadistica/agricultura/index.html (accessed November 8, 2018).
Gómez-Lama Cabanás, C., Ruano-Rosa, D., Legarda, G., Pizarro-Tobías, P., Valverde-Corredor, A., Triviño, J., et al. (2018). Bacillales members from the olive rhizosphere are effective biological control agents against the defoliating pathotype of Verticillium dahliae. Agriculture 8:90. doi: 10.3390/agriculture8070090
Gopal, M., Gupta, A., and Thomas, G. (2013). Bespoke microbiome therapy to manage plant diseases. Front. Microbiol. 4:355. doi: 10.3389/fmicb.2013.00355
Gopalakrishnan, V. (2017). Optimization of protein isolation and preliminary comparative proteomics of pathogenic Fusarium oxysporum f. sp. cubense (p-Foc) and non-pathogenic Fusarium oxysporum (np-Fo). J. Plant Pathol. 99, 361–369. doi: 10.4454/jpp.v99i2.3883
Goswami, D., Patel, K., Parmar, S., Vaghela, H., Muley, N., Dhandhukia, P., et al. (2015). Elucidating multifaceted urease producing marine Pseudomonas aeruginosa BG as a cogent PGPR and bio-control agent. Plant Growth Regul. 75, 253–263. doi: 10.1007/s10725-014-9949-1
Grosch, R., Dealtry, S., Schreiter, S., Berg, G., Mendonca-Hagler, L., and Smalla, K. (2012). Biocontrol of Rhizoctonia solani: complex interaction of biocontrol strains, pathogen and indigenous microbial community in the rhizosphere of lettuce shown by molecular methods. Plant Soil 361, 343–357. doi: 10.1007/s11104-012-1239-y
Grosch, R., Scherwinski, K., Lottmann, J., and Berg, G. (2006). Fungal antagonists of the plant pathogen Rhizoctonia solani: selection, control efficacy and influence on the indigenous microbial community. Mycol. Res. 110, 1464–1474. doi: 10.1016/j.mycres.2006.09.014
Gunji, S., Arima, K., and Beppu, T. (1983). Screening of antifungal antibiotics according to activities inducing morphological abnormalities. Agric. Biol. Chem. 47, 2061–2069.
Guo, G., Wang, B., Ma, W., Li, X., Yang, X., Zhu, C., et al. (2013). Biocontrol of Fusarium wilt of banana: key influence factors and strategies. Afr. J. Microbiol. Res. 7, 4835–4843. doi: 10.5897/AJMR2012.2392
Haas, D., and Défago, G. (2005). Biological control of soil-borne pathogens by fluorescent pseudomonads. Nat. Rev. Microbiol. 3, 307–319. doi: 10.1038/nrmicro1129
Habeeba, T., Ravichandra, N. G., Krishnappa, K., and Reddy, B. M. R. (2003). “Interaction of Glomus fasciculatum in the management of burrowing nematode, Radopholus similis and wilt fungus, Fusarium oxysporum f. sp. cubense on banana,” in Proceedings of the ‘National Symposium on Biodiversity and Management of Nematodes in Cropping Systems for Sustainable Agriculture’ (New Delhi: Division of Nematology, Indian Agricultural Research Institute), 244–250.
Haddad, F., Rocha, L. S., Soares, A. C. F., Martins, I. P. S., Teixeira, L. A. J., Staver, C., et al. (2018). Management of Fusarium wilt of bananas in Minas Gerais, Brazil. Acta Hortic. 1196, 137–145. doi: 10.17660/ActaHortic.2018.1196.16
Hardoim, P. R., van Overbeek, L. S., Berg, G., Pirttilä, A. M., Compant, S., Campisano, A., et al. (2015). The hidden world within plants: ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol. Mol. Biol. Rev. 79, 293–320. doi: 10.1128/MMBR.00050-14
Harman, G. E. (2011). Multifunctional fungal plant symbionts: new tools to enhance plant growth and productivity. New Phytol. 189, 647–649. doi: 10.1111/j.1469-8137.2010.03614.x
Harman, G. E., Howell, C. R., Viterbo, A., Chet, I., and Lorito, M. (2004). Trichoderma species - opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2, 43–56. doi: 10.1038/nrmicro797
He, C. Y., Hsiang, T., and Wolyn, D. J. (2002). Induction of systemic disease resistance and pathogen defence responses in Asparagus officinalis inoculated with nonpathogenic strains of Fusarium oxysporum. Plant Pathol. 51, 225–230. doi: 10.1046/j.1365-3059.2002.00682.x
He, H., Cai, X., Hong, Y., Guan, X., and Hu, F. (2002). Selection of endophytic antifungal bacteria from Capsicum. Chin. J. Biol. Control 18, 171–175.
Hecht-Buchholz, C., Borges-Pérez, A., Fernández Falcón, M., and Borges, A. A. (1998). Influence of zinc nutrition on Fusarium wilt of banana - an electron microscopic investigation. Acta Hortic. 490, 277–283. doi: 10.17660/ActaHortic.1998.490.27
Hegde, S. V. (2002). Liquid biofertilizers in Indian agricultural. Biofertilizers News Lett. 12, 17–22. doi: 10.13140/RG.2.1.1876.9367
Hennessy, C., Walduck, G., Daly, A., and Padovan, A. (2005). Weed hosts of Fusarium oxysporum f. sp. cubense tropical race 4 in northern Australia. Australas. Plant Pathol. 34, 115–117. doi: 10.1071/AP04091
Herbert, J. A., and Marx, D. (1990). Short-term control of Panama disease of bananas in South Africa. Phytophylactica 22, 339–340.
Hermanto, C., Eliza Djatnika, I., Emilda, D., and Mujiman, Subhana (2012). Pre-planting treatments for management of banana fusarium wilt. J. Agric. Biol. Sci. 7, 260–265.
Hesse, C., Schulz, F., Bull, C. T., Shaffer, B. T., Yan, Q., Shapiro, N., et al. (2018). Genome-based evolutionary history of Pseudomonas spp. Environ. Microbiol. 20, 2142–2159. doi: 10.1111/1462-2920.14130
Hilton, S., Bennett, A. J., Keane, G., Bending, G. D., Chandler, D., Stobart, R., et al. (2013). Impact of shortened crop rotation of oilseed rape on soil and rhizosphere microbial diversity in relation to yield decline. PLoS ONE 8:e59859. doi: 10.1371/journal.pone.0059859
Hima, V. M., and Beena, S. (2016). Bioefficacy of Trichoderma spp. for the management of soil borne fungal pathogens. Indian Phytopathol. 69, 316–318.
Ho, Y., Chiang, H., Chao, C., Su, C., Hsu, H., Guo, C., et al. (2015). In planta biocontrol of soilborne Fusarium wilt of banana through a plant endophytic bacterium, Burkholderia cenocepacia 869T2. Plant Soil 387, 295–306. doi: 10.1007/s11104-014-2297-0
Hong, S., Ju, H., Ruan, Y., Lu, M., Wang, B., Zhao, Y., et al. (2017). Effect of eggplant-banana rotation with bioorganic fertilizer treatment on soil microflora in banana continuous cropping orchard. J. Eco Agric. 25, 78–85. doi: 10.13930/j.cnki.cjea.160609
Huang, X., Liu, L., Wen, T., Zhu, R., Zhang, J., and Cai, Z. (2015a). Illumina MiSeq investigations on the changes of microbial community in the Fusarium oxysporum f. sp. cubense infected soil during and after reductive soil disinfestation. Microbiol. Res. 181, 33–42. doi: 10.1016/j.micres.2015.08.004
Huang, X., Wen, T., Zhang, J., Meng, L., Zhu, T., and Cai, Z. (2015b). Toxic organic acids produced in biological soil disinfestation mainly caused the suppression of Fusarium oxysporum f. sp. cubense. Bio Control 60, 113–124. doi: 10.1007/s10526-014-9623-6
Huang, X., Wen, T., Zhang, J., Meng, L., Zhu, T., Liu, L., et al. (2015c). Control of soil-borne pathogen Fusarium oxysporum by biological soil disinfestation with incorporation of various organic matters. Eur. J. Plant Pathol. 143, 223–235. doi: 10.1007/s10658-015-0676-x
Huang, Y. H., Wang, R. C., Li, C. H., Zuo, C. W., Wei, Y. R., Zhang, L., et al. (2012). Control of Fusarium wilt in banana with Chinese leek. Eur. J. Plant Pathol. 134, 87–95. doi: 10.1007/s10658-012-0024-3
Hwang, S. H. (1985). Ecology and control of fusarial wilt of banana. Plant Prot. Bull. Taiwan 27, 233–245.
Ikeda, S., Anda, M., Inaba, S., Eda, S., Sato, S., Sasaki, K., et al. (2011). Autoregulation of nodulation interferes with impacts of nitrogen fertilization levels on the leaf-associated bacterial community in soybeans. Appl. Environ. Microbiol. 77, 1973–1980. doi: 10.1128/AEM.02567-10
Instituto Canario de Calidad Agroalimentaria, (2018). Platano de Canarias IGP. Available online at: http://www.gobiernodecanarias.org/agricultura/icca/temas_calidad/platano (accessed November 8, 2018).
Jackson, M. A., Slininger, P. J., and Bothast, R. J. (1989). Effects of zinc, iron, cobalt, and manganese on Fusarium moniliforme NRRL 13616 growth and fusarin C biosynthesis in submerged cultures. Appl. Environ. Microbiol. 55, 649–655.
Jacobsen, C. S., and Hjelmso, M. H. (2014). Agricultural soils, pesticides and microbial diversity. Curr. Opin. Biotechnol. 27, 15–20. doi: 10.1016/j.copbio.2013.09.003
Jaizme-Vega, M. C., Sosa Hernández, B., and Hernández Hernández, J. M. (1998). Interaction of arbuscular mycorrhizal fungi and the soil pathogen Fusarium oxysporum f. sp. cubense on the first stages of micropropagated Grande Naine banana. Acta Hortic. 490, 285–295. doi: 10.17660/ActaHortic.1998.490.28
Jansen, C. (2017). Fears around containment of Fusarium TR4 in Africa and Asia. Available online at: https://www.freshplaza.com/article/2184078/fears-around-containment-of-fusarium-tr-in-africa-and-asia/ (accessed October 31, 2017).
Jauri, P. V., Altier, N., Pérez, C. A., and Kinkel, L. (2017). Cropping history effects on pathogen suppressive and signaling dynamics in Streptomyces communities. Phytobiomes J. 2, 14–23. doi: 10.1094/PBIOMES-05-17-0024-R
Jeger, M. J., Waller, J. M., Johanson, A., and Gowen, S. R. (1996). Monitoring in banana pest management. Crop Prot. 15, 391–397. doi: 10.1016/0261-2194(96)00011-7
Ji, H., Zheng, Q., Dong, X., Zhou, J., Shen, Q., and Guo, S. (2012). Effects and mechanism of copper and zinc elements on controlling Fusarium-wilt disease of banana. Acta Hortic. Sinica 39, 1064–1072.
Jiménez-Díaz, R. M., Cirulli, M., Bubici, G., Jiménez-Gasco, M. D., Antoniou, P. P., and Tjamos, E. C. (2012). Verticillium wilt, a major threat to olive production: current status and future prospects for its management. Plant Dis. 96, 304–329. doi: 10.1094/PDIS-06-11-0496
Kavino, M., and Manoranjitham, S. K. (2018). In vitro bacterization of banana (Musa spp.) with native endophytic and rhizospheric bacterial isolates: novel ways to combat Fusarium wilt. Eur. J. Plant Pathol. 151, 371–387. doi: 10.1007/s10658-017-1379-2
Kavino, M., Manoranjitham, S. K., Balamohan, T. N., Kumar, N., Karthiba, L., and Samiyappan, R. (2014). Enhancement of growth and Panama wilt resistance in banana by in vitro co-culturing of banana plantlets with PGPR and endophytes. Acta Hortic. 1024, 277–282. doi: 10.17660/ActaHortic.2014.1024.37
Kavino, M., Manoranjitham, S. K., Kumarand, N., and Vijayakumar, R. M. (2016). “Plant growth stimulation and biocontrol of Fusarium wilt (Fusarium oxysporum f. sp. cubense) by co-inoculation of banana (Musa spp.) plantlets with PGPR and endophytes,” in Proceedings of the ‘4th Asian PGPR - Recent Trends in PGPR Research for Sustainable Crop Productivity’ (Hanoi: Asian PGPR Society), 77–83.
Kema, G. H. J. (2018). Panama Disease. Wageningen: Wageningen University and Research. Available online at: https://fusariumwilt.org/ (accessed December 3, 2018).
Keswani, C., Bisen, K., Singh, V., Sarma, B. K., and Singh, H. B. (2016). Bioformulations: For Sustainable Agriculture. New Delhi: Springer, 299.
Khan, N., Maymon, M., and Hirsch, A. M. (2017). Combating Fusarium infection using Bacillus-based antimicrobials. Microorganisms 5:E75. doi: 10.3390/microorganisms5040075
Kidane, E. G., and Laing, M. D. (2010). Integrated control of Fusarium wilt of banana (Musa spp.). Acta Hortic. 879, 315–321. doi: 10.17660/ActaHortic.2010.879.32
Kinkel, L. L., Schlatter, D. C., Bakker, M. G., and Arenz, B. E. (2012). Streptomyces competition and co-evolution in relation to plant disease suppression. Res. Microbiol. 163, 490–499. doi: 10.1016/j.resmic.2012.07.005
Köberl, M., Dita, M., Martinuz, A., Staver, C., and Berg, G. (2015). Agroforestry leads to shifts within the gammaproteobacterial microbiome of banana plants cultivated in Central America. Front. Microbiol. 6:91. doi: 10.3389/fmicb.2015.00091
Köberl, M., Dita, M., Martinuz, A., Staver, C., and Berg, G. (2017). Members of Gammaproteobacteria as indicator species of healthy banana plants on Fusarium wilt-infested fields in Central America. Sci. Rep. 7:45318. doi: 10.1038/srep45318
Komada, H. (1975). Development of a selective medium for quantitative isolation of Fusarium oxysporum from natural soil. Rev. Plant Prot. Res. 8, 114–124.
Kulkarni, V. (2018). “India in a race against wilt in Cavendish banana,” in Agri Business (India: The Hindu). Available online at: https://www.thehindubusinessline.com/economy/agri-business/india-in-a-race-against-wilt-in-cavendish-banana/article23650060.ece (accessed February 20, 2019).
Kumar, D. P., D, A. P., Singh, R. K., Thenmozhi, R., Nagasathya, A., Thajuddin, N., et al. (2012). Evaluation of extracellular lytic enzymes from indigenous Bacillus isolates. J. Microbiol. Biotechnol. Res. 2, 129–137.
Lakshmanan, P., and Selvaraj, P. (1984). Effect of fungicides on Panama disease of Rasthali banana. South Indian Hortic. 32, 249–251.
Larkin, R. P., and Fravel, D. R. (2002). Effects of varying environmental conditions on biological control of Fusarium wilt of tomato by nonpathogenic Fusarium spp. Phytopathology 92, 1160–1166. doi: 10.1094/PHYTO.2002.92.11.1160
Lenoir, I., Fontaine, J., and Lounès-Hadj Sahraoui, A. (2016). Arbuscular mycorrhizal fungal responses to abiotic stresses: a review. Phytochemistry 123, 4–15. doi: 10.1016/j.phytochem.2016.01.002
Li, C. Q., Shao, J. F., Wang, Y. J., Li, W. B., Guo, D. J., Yan, B., et al. (2013). Analysis of banana transcriptome and global gene expression profiles in banana roots in response to infection by race 1 and tropical race 4 of Fusarium oxysporum f. sp cubense. BMC Genomics 14:851. doi: 10.1186/1471-2164-14-851
Li, J. Y., Harper, J. K., Grant, D. M., Tombe, B. O., Bashyal, B., Hess, W. M., et al. (2001). Ambuic acid, a highly functionalized cyclohexenone with antifungal activity from Pestalotiopsis spp. and Monochaetia sp. Phytochemistry 56, 463–468. doi: 10.1016/S0031-9422(00)00408-8
Li, W. B., Li, C. Q., Sun, J. B., and Peng, M. (2017). Metabolomic, biochemical, and gene expression analyses reveal the underlying responses of resistant and susceptible banana species during early infection with Fusarium oxysporum f. sp cubense. Plant Dis. 101, 534–543. doi: 10.1094/PDIS-09-16-1245-RE
Li, W. M., Dita, M., Wu, W., Hu, G. B., Xie, J. H., and Ge, X. J. (2015). Resistance sources to Fusarium oxysporum f. sp. cubense tropical race 4 in banana wild relatives. Plant Pathol. 64, 1061–1067. doi: 10.1111/ppa.12340
Lian, J., Wang, Z., Cao, L., Tan, H., Patrik, I., Jiang, Z., et al. (2009). Artificial inoculation of banana tissue culture plantlets with indigenous endophytes originally derived from native banana plants. Biol. Control 51, 427–434. doi: 10.1016/j.biocontrol.2009.08.002
Lin, S., Wang, C., and Su, C. (2012). Using arbuscular mycorrhizal fungus and other microorganisms for control of Fusarium wilt of banana. J. Taiwan Agric. Res. 61, 241–249.
Liu, H. (2011). Identification of Antagonistic Bacteria Against Banana Wilt and Its Preliminary Application. [Master's thesis]. Hainan: Hainan University.
Liu, P., and Prada, V. (2018). World Banana Forum. Rome: Food and Agriculture Organization of the United Nations, FAO. Available online at: http://www.fao.org/economic/worldbananaforum/en/ (accessed December 3, 2018).
Liu, Y., and Lu, Q. (2013). Inhibitory effects of Acremonium sp. on Fusarium wilt in bananas. Afr. J. Agric. Res. 8, 6241–6249. doi: 10.5897/AJAR2013.7082
Loper, J. E., Hassan, K. A., Mavrodi, D. V., Davis, E. W. II., Lim, C. K., Shaffer, B. T., et al. (2012). Comparative genomics of plant-associated Pseudomonas spp.: insights into diversity and inheritance of traits involved in multitrophic interactions. PLoS Genet. 8:e1002784. doi: 10.1371/journal.pgen.1002784
López Escudero, F. J., and Mercado-Blanco, J. (2011). Verticillium wilt of olive: a case study to implement an integrated strategy to control a soil-borne pathogen. Plant Soil 344, 1–50. doi: 10.1007/s11104-010-0629-2
López-Cepero, J., Puerta, M., and Piedra Buena, A. (2014). Guía para la Gestión Integrada de Plagas en Platanera, Vol. 2. Santa Cruz de Tenerife: COPLACA, 44.
Lu, Y., Liao, D. J., Pu, J. J., Qi, Y. X., and Xie, Y. X. (2013). Proteome analysis of resistant and susceptible Cavendish banana roots following inoculation with Fusarium oxysporum f. sp cubense. Physiol. Mol. Plant Pathol. 84, 163–171. doi: 10.1016/j.pmpp.2013.09.002
Luan, F. G., Zhang, L. L., Lou, Y. Y., Wang, L., Liu, Y. N., and Zhang, H. Y. (2015). Analysis of microbial diversity and niche in rhizosphere soil of healthy and diseased cotton at the flowering stage in southern Xinjiang. Genet. Mol. Res. 14, 1602–1611. doi: 10.4238/2015.March.6.7
Lugtenberg, B., and Kamilova, F. (2009). Plant-growth-promoting rhizobacteria. Annu. Rev. Microbiol. 63, 541–556. doi: 10.1146/annurev.micro.62.081307.162918
Lutz, M. P., Wenger, S., Maurhofer, M., Defago, G., and Duffy, B. (2004). Signaling between bacterial and fungal biocontrol agents in a strain mixture. FEMS Microbiol. Ecol. 48, 447–455. doi: 10.1016/j.femsec.2004.03.002
Manikandan, R., Saravanakumar, D., Rajendran, L., Raguchander, T., and Samiyappan, R. (2010). Standardization of liquid formulation of Pseudomonas fluorescens Pf1 for its efficacy against Fusarium wilt of tomato. Biol. Control 54, 83–89. doi: 10.1016/j.biocontrol.2010.04.004
Manoch, L., and Dethoup, T. (2011). A potential use of Talaromyces species as biological agents against plant pathogenic fungi. Thai J. Agric. Sci. 44, 81–91.
Marín, M., Wong, I., García, G., Morán, R., Basulto, R., Pimentel, E., et al. (2013). In vitro antagonistic activity of Tsukamurella paurometabola C-924 against phytopathogens. Rev. Protec. Vegetal 28, 132–137.
Massart, S., Martinez-Medina, M., and Jijakli, M. H. (2015). Biological control in the microbiome era: challenges and opportunities. Biol. Control 89, 98–108. doi: 10.1016/j.biocontrol.2015.06.003
Mercado-Blanco, J. (2015). “Pseudomonas strains that exert biocontrol of plant pathogens,” in Pseudomonas, New Aspects of Pseudomonas Biology, Vol. 7, eds J. L. Ramos, J. B. Goldberg, and A. Filloux (Dordrecht: Springer Netherlands), 121–172. doi: 10.1007/978-94-017-9555-5_6
Mercado-Blanco, J., Abrantes, I., Barra Caracciolo, A., Bevivino, A., Ciancio, A., Grenni, P., et al. (2018). Belowground microbiota and the health of tree crops. Front. Microbiol. 9:1006. doi: 10.3389/fmicb.2018.01006
Mercado-Blanco, J., and Bakker, P. A. H. M. (2007). Interactions between plants and beneficial Pseudomonas spp.: exploiting bacterial traits for crop protection. Antonie Van Leeuwenhoek 92, 367–389. doi: 10.1007/s10482-007-9167-1
Mercado-Blanco, J., and Lugtenberg, B. J. J. (2014). Biotechnological applications of bacterial endophytes. Curr. Biotechnol. 3, 60–75. doi: 10.2174/22115501113026660038
Mohandas, S., Manamohan, M., Rawal, R. D., Saikat, C., Sreekantappa, H., Manjula, R., et al. (2004). Interaction of Fusarium oxysporum f. sp. cubense with Pseudomonas fluorescens precolonized to banana roots. World J. Microbiol. Biotechnol. 20, 651–655. doi: 10.1023/B:WIBI.0000043197.54871.1b
Mohandas, S., Manjula, R., Rawal, R. D., Lakshmikantha, H. C., Saikat, C., and Ramachandra, Y. L. (2010). Evaluation of arbuscular mycorrhiza and other biocontrol agents in managing Fusarium oxysporum f. sp. cubense infection in banana cv. Neypoovan. Biocontrol Sci. Technol. 20, 165–181. doi: 10.1080/09583150903438439
Molina, A. B., Fabregar, E., Sinohin, V. G., Yi, G., and Viljoen, A. (2009). Recent occurrence of Fusarium oxysporum f. sp. cubense tropical race 4 in Asia. Acta Hortic. 828, 109–116. doi: 10.17660/ActaHortic.2009.828.10
Molina, A. B., Sinohin, V. O., Fabregar, E. G., Ramillete, E. B., Loayan, M. M., and Chao, C. P. (2016a). Field resistance of Cavendish somaclonal variants and local banana cultivars to tropical race 4 of Fusarium wilt in the Philippines. Acta Hortic. 1114, 227–230. doi: 10.17660/ActaHortic.2016.1114.31
Molina, A. B., Sinohin, V. O., Fabregar, E. G., Ramillete, E. B., Yi, G., Sheng, O., et al. (2016b). Resistance to Fusarium oxysporum f. sp. cubense tropical race 4 in African bananas. Acta Hortic. 1114, 107–110. doi: 10.17660/ActaHortic.2016.1114.15
Mukhongo, R. W., Kavoo-Mwangi, M. A., Kahangi, M. E., Ateka, E. M., Were, A. B., Okalebo, J. R., et al. (2015). Occurrence of arbuscular mycorrhizal fungi and Fusarium in TC banana rhizosphere inoculated with microbiological products in different soils in Kenya. Int. J. Soil Sci. 10, 45–62. doi: 10.3923/ijss.2015.45.62
Mulet, M., Lalucat, J., and García-Valdés, E. (2010). DNA sequence-based analysis of the Pseudomonas species. Environ. Microbiol. 12, 1513–1530. doi: 10.1111/j.1462-2920.2010.02181.x
Munimbazi, C., and Bullerman, L. B. (1998). Isolation and partial characterization of antifungal metabolites of Bacillus pumilus. J. Appl. Microbiol. 84, 959–968. doi: 10.1046/j.1365-2672.1998.00431.x
Mustaffa, M. M., and Thangavelu, R. (2011). Status of Fusarium wilt in India. Acta Hortic. 897, 323–329. doi: 10.17660/ActaHortic.2011.897.44
Nandakumar, R., Babu, S., Viswanathan, R., Sheela, J., Raguchander, T., and Samiyappan, R. (2001). A new bio-formulation containing plant growth promoting rhizobacterial mixture for the management of sheath blight and enhanced grain yield in rice. BioControl 46, 493–510. doi: 10.1023/A:1014131131808
Narayanasamy, P. (2013). Biological Management of Diseases of Crops: Vol. 1: Characteristics of Biological Control Agents, ed P. Narayanasamy (Dordrecht: Springer).
Nawangsih, A. A., and Purba, F. (2013). Isolation of fluorescent pseudomonads, heat tolerant and chitinolytic bacteria in banana rhizosphere with antagonistic activities against Fusarium oxysporum f. sp. cubense in vitro and molecular identification of selected isolates. J. Int. Soc. Southeast Asian Agr. Sci. 19, 30–40.
Nel, B., Steinberg, C., Labuschagne, N., and Viljoen, A. (2006). Isolation and characterization of nonpathogenic Fusarium oxysporum isolates from the rhizosphere of healthy banana plants. Plant Pathol. 55, 207–216. doi: 10.1111/j.1365-3059.2006.01343.x
Niere, B., Gold, C. S., Coyne, D., Dubois, T., and Sikora, R. (2014). Performance of tissue-cultured versus sucker-derived East African highland banana (Musa AAA-EA) under high and low input systems in Uganda. Field Crops Res. 156, 313–321. doi: 10.1016/j.fcr.2013.11.014
Nita, K., and Harsh, K. (2015). Association of nonpathogenic Fusarium oxysporum species with cultured shoot apices of banana (Musa acuminata) cultivars. Bioscan 10, 629–633.
Northern Queensland Register (2018). Bananas: TR4 Affected Farms Benefit from New Agreement. Available online at: https://www.northqueenslandregister.com.au/story/5643572/tr4-affected-farms-benefit-from-new-banana-agreement/?cs=4735 (Accessed: February 20, 2019)
Okubo, A., Matsusaka, M., and Sugiyama, S. (2016). Impacts of root symbiotic associations on interspecific variation in sugar exudation rates and rhizosphere microbial communities: a comparison among four plant families. Plant Soil 399, 345–356. doi: 10.1007/s11104-015-2703-2
Olivain, C., Humbert, C., Nahalkova, J., Fatehi, J., L'Haridon, F., and Alabouvette, C. (2006). Colonization of tomato root by pathogenic and nonpathogenic Fusarium oxysporum strains inoculated together and separately into the soil. Appl. Environ. Microbiol. 72, 1523–1531. doi: 10.1128/AEM.72.2.1523-1531.2006
Ortiz, R., and Pocasangre, L. E. (2012). Biological control of Panama disease (Fusarium oxysporum f. sp. cubense) using endophytic fungi. Tierra Trop. 8, 221–228.
Parnell, J. J., Berka, R., Young, H. A., Sturino, J. M., Kang, Y. W., Barnhart, D. M., et al. (2016). From the lab to the farm: an industrial perspective of plant beneficial microorganisms. Front. Plant Sci. 7:1110. doi: 10.3389/fpls.2016.01110
Parniske, M. (2008). Arbuscular mycorrhiza: the mother of plant root endosymbioses. Nat. Rev. Microbiol. 6:763. doi: 10.1038/nrmicro1987
Pattison, A. B., Wright, C. L., Kukulies, T. L., and Molina, A. B. (2014). Ground cover management alters development of Fusarium wilt symptoms in Ducasse bananas. Australas. Plant Pathol. 43, 465–476. doi: 10.1007/s13313-014-0296-5
Pegg, K., Moore, N., and Bentley, S. (1996). Fusarium wilt of banana in Australia: a review. Aust. J. Agric. Res. 47, 637–650. doi: 10.1071/AR9960637
Peng, H. X., Sivasithamparam, K., and Turner, D. W. (1999). Chlamydospore germination and Fusarium wilt of banana plantlets in suppressive and conducive soils are affected by physical and chemical factors. Soil Biol. Biochem. 31, 1363–1374. doi: 10.1016/S0038-0717(99)00045-0
Pérez Vicente, L., Batlle Viera, A., Chacón Benazet, J., and Montenegro Moracén, V. (2009). Efficacy of Trichoderma harzianum A34 in the biocontrol of Fusarium oxysporum f. sp. cubense, the causal agent of Fusarium wilt or Panama disease of bananas in Cuba. Fitosanidad 13, 259–263.
Pérez-Vicente, L. (2015). Banana farm best practices for prevention of Fusarium wilt TR4 and other exotic banana diseases. Fitosanidad 19, 243–250.
Pittaway, P. A., Pittaway, P. A., Nasir, N., and Pegg, K. G. (1999). Soil receptivity and host-pathogen dynamics in soils naturally infested with Fusarium oxysporum f. sp. cubense, the cause of Panama disease in bananas. Aust. J. Agric. Res. 50, 623–628. doi: 10.1071/A98152
Pliego, C., Kamilova, F., and Lugtenberg, B. (2011). “Plant growth-promoting bacteria: fundamentals and exploitation,” in Bacteria in Agrobiology: Crop Ecosystems, ed D. K. Maheshwari (Berlin; Heidelberg: Springer), 295–343. doi: 10.1007/978-3-642-18357-7_11
Ploetz, R. C. (2005). Panama Disease: An Old Nemesis Rears Its Ugly Head: Part 1. The Beginnings of the Banana Export Trades. Plant Health Progress. St. Paul, MN: Plant Management Network. Available online at: http://www.apsnet.org/publications/apsnetfeatures/Pages/PanamaDiseasePart1.aspx (accessed February 20, 2019).
Ploetz, R. C. (2015a). Fusarium wilt of banana. Phytopathology 105, 1512–1521. doi: 10.1094/PHYTO-04-15-0101-RVW
Ploetz, R. C. (2015b). Management of Fusarium wilt of banana: a review with special reference to tropical race 4. Crop Prot. 73, 7–15. doi: 10.1016/j.cropro.2015.01.007
Ploetz, R. C., Herbert, J., Sebasigari, K., Hernandez, J. H., Pegg, K. G., Ventura, J. A., et al. (1990). “Importance of Fusarium wilt in different banana growing regions,” in Proceedings ‘Fusarium Wilt of Banana’, ed R. C. Ploetz (St. Paul, MN: APS Press), 9–26.
Ploetz, R. C., and Pegg, K. G. (2000). “Fusarium wilt,” in Diseases of Banana, Abaca and Enset, ed D. R. Jones (Wallingford: CABI Publishing), 143–159.
Pocasangre, L. E., Ploetz, R. C., Molina, A. B., and Vicente, L. P. (2011). Raising awareness of the threat of Fusarium wilt tropical race 4 in Latin America and the Caribbean. Acta Hortic. 897, 331–337. doi: 10.17660/ActaHortic.2011.897.45
Pushpavathi, Y., Dash, S. N., Reddy, Y. A., and Triveni, V. (2017). Evaluation of fungicides and biocontrol agents for potential application in Fusarium wilt management of banana cv Bantal. Int. J. Farm Sci. 7, 115–118.
Qi, D., Zou, L., Zhou, D., Feng, R., Gao, Z., and Zhang, X. (2017). Isolation, identification of strain GA1-2 and its antifungal activity against Fusarium oxysporum f. sp. cubense. J. Plant Prot. 44, 809–816.
Qi, Y., Zhang, X., Pu, J., Zhang, H., and Xie, Y. (2008). Inactivation effect of 10 compounds on Fusarium oxysporum f. sp. cubense and its toxin. J. Fruit Sci. 25, 78–82.
Qin, H., Yang, L., Li, S., Xie, Y., and Huang, J. (2010). Isolation of antagonistic actinomyces against banana Fusarium wilt disease and primary evaluation on their inhibition effects. Chin. J. Biol. Control 26, 174–180.
Qin, L., Guo, C., Huang, S., Li, C., Wei, L., Wei, S., et al. (2017). Growth-promoting effects of Trichoderma asperellum strain PZ6 on banana and its indoor control effect against banana Fusarium wilt. J. South. Agric. 48, 277–283.
Raaijmakers, J. M., De Bruijn, I., Nybroe, O., and Ongena, M. (2010). Natural functions of lipopeptides from Bacillus and Pseudomonas: more than surfactants and antibiotics. FEMS Microbiol. Rev. 34, 1037–1062. doi: 10.1111/j.1574-6976.2010.00221.x
Raaijmakers, J. M., Paulitz, T. C., Steinberg, C., Alabouvette, C., and Moënne-Loccoz, Y. (2009). The rhizosphere: a playground and battlefield for soilborne pathogens and beneficial microorganisms. Plant Soil 321, 341–361. doi: 10.1007/s11104-008-9568-6
Radhakrishnan, R., Hashem, A., and Abd Allah, E. F. (2017). Bacillus: a biological tool for crop improvement through bio-molecular changes in adverse environments. Front. Physiol. 8:667. doi: 10.3389/fphys.2017.00667
Raguchander, T., Jayashree, K., and Samiyappan, R. (1997). Management of Fusarium wilt of banana using antagonistic microorganisms. J. Biol. Control 11, 101–105.
Raguchander, T., Shanmugam, V., and Samiyappan, R. (2000). Biological control of Panama wilt disease of banana. Madras Agric. J. 87, 320–321.
Rajappan, K., Vidhyasekaran, P., Sethuraman, K., and Baskaran, T. L. (2002). Development of powder and capsule formulations of Pseudomonas fluorescens strain Pf-1 for control of banana wilt. Z. Pflanzenk. Pflanzen. 109, 80–87.
Rames, E. K., Pattison, A. B., Czislowski, E., and Smith, M. K. (2018). Soil microbial community changes associated with ground cover management in cultivation of Ducasse banana (Musa sp. ABB, Pisang Awak subgroup) and suppression of Fusarium oxysporum. Australas. Plant Pathol. 47, 449–462. doi: 10.1007/s13313-018-0578-4
Ramu, V., Venkatarangaiah, K., Krishnappa, P., Rajanna, S. K. S., Deeplanaik, N., Pal, A. C., et al. (2016). Identification of biomarkers for resistance to Fusarium oxysporum f. sp. cubense infection and in silico studies in Musa paradisiaca cultivar Puttabale through proteomic approach. Proteomes 4:9. doi: 10.3390/proteomes4010009
Regalado Guijarro, V. M., and Hernández Hernández, J. M. (1998). Racial structure of Fusarium oxysporum f. sp. cubense in the Canary Islands. Acta Hortic. 490, 315–321. doi: 10.17660/ActaHortic.1998.490.31
Ribeiro, R. C. F., Campos, V. P., Xavier, A. A., Rocha, L. S., Ribeiro, H. B., Aguiar, F. M., et al. (2012). Control of Meloidogyne javanica and Panama disease with rhizobacteria. Nematropica 42, 218–226.
Rodríguez Serrano, M. (2012). Mal de Panamá. Medidas de Control y Prevención. Technical Report. Agrocabildo, Servicio Técnico de Agricultura y Desarrollo Rural. Santa Cruz de Tenerife (accessed November 8, 2018).
Roy, S., Ojha, P. K., Ojha, K. L., Pathak, K. N., and Upadhyay, J. P. (1998). Effects of pesticides on disease intensity of banana wilt complexes. J. Appl. Biol. 8, 36–38.
Rutherford, M. A. (2001). “Fusarium wilt of banana in East Africa,” in Proceedings of the International Workshop ‘Banana Fusarium Wilt Management: Towards Sustainable Cultivation’, eds A. B. Molina, N. H. Nik Masdesk, and K. W. Liew (Los Baños: INIBAP-ASPNET), 86–94.
Sampaio, D. B., Mendes Filho, P. F., Mascena, A. M., Gomes, V. F. F., and Guimarães, F. V. A. (2012). Colonisation of arbuscular mycorrhiza and tolerance to Panama disease in seedlings of the maçã banana. Rev. Ciênc. Agron. 43, 462–469. doi: 10.1590/S1806-66902012000300007
Sánchez, G. M. (2013). Quarantine importance of diseases of bananas and plantains. Rev. Mex. Fitopatol. 31, S4–S5.
Sanjeev, K. K., and Eswaran, A. (2008). Efficacy of micro nutrients on banana Fusarium wilt (Fusarium oxysporum f. sp. cubense) and its synergistic action with Trichoderma viride. Notulae Botanicae Horti Agrobotanici Cluj Napoca 36, 52–54. doi: 10.15835/nbha36191
Saraf, M., Pandya, U., and Thakkar, A. (2014). Role of allelochemicals in plant growth promoting rhizobacteria for biocontrol of phytopathogens. Microbiol. Res. 169, 18–29. doi: 10.1016/j.micres.2013.08.009
Saravanan, T., Bhaskaran, R., and Muthusamy, M. (2004a). Pseudomonas fluorescens induced enzymological changes in banana roots (cv. Rasthali) against Fusarium wilt disease. Plant Pathol. J. 3, 72–80. doi: 10.3923/ppj.2004.72.80
Saravanan, T., Muthusamy, M., and Marimuthu, T. (2003). Development of integrated approach to manage the fusarial wilt of banana. Crop Prot. 22, 1117–1123. doi: 10.1016/S0261-2194(03)00146-7
Saravanan, T., Muthusamy, M., and Marimuthu, T. (2004b). Effect of Pseudomonas fluorescens on Fusarium wilt pathogen in banana rhizosphere. J. Biol. Sci. 4, 192–198. doi: 10.3923/jbs.2004.192.198
Sarma, B. K., Yadav, S. K., Singh, S., and Singh, H. B. (2015). Microbial consortium-mediated plant defense against phytopathogens: readdressing for enhancing efficacy. Soil Biol. Biochem. 87, 25–33. doi: 10.1016/j.soilbio.2015.04.001
Satheesh, S., and Venu, D. V. (2004). Effect of various amendments and Trichoderma viride on the fusarial wilt of banana. Mysore J. Agr. Sci. 38, 546–550.
Savi, G., Vitorino, V. J, Bortoluzzi, A., and Scussel, V. (2013). Effect of zinc compounds on Fusarium verticillioides growth hyphae alterations, conidia and fumonisin production. J. Sci. Food Agric. 93, 3395–3402. doi: 10.1002/jsfa.6271
Schreiter, S., Babin, D., Smalla, K., and Grosch, R. (2018). Rhizosphere competence and biocontrol effect of Pseudomonas sp. RU47 independent from plant species and soil type at the field scale. Front. Microbiol. 9:97. doi: 10.3389/fmicb.2018.00097
Sebasigari, K., and Stover, R. H. (1988). Banana diseases and pests in East Africa: Report of a survey made in november 1987 (Montpellier: INIBAP), 15.
Sekhar, A. C., and Pious, T. (2015). Isolation and identification of shoot-tip associated endophytic bacteria from banana cv. Grand Naine and testing for antagonistic activity against Fusarium oxysporum f. sp. cubense. Am. J. Plant Sci. 6, 943–954. doi: 10.4236/ajps.2015.67101
Selvaraj, S., Ganeshamoorthi, P., Anand, T., Raguchander, T., Seenivasan, N., and Samiyappan, R. (2014). Evaluation of a liquid formulation of Pseudomonas fluorescens against Fusarium oxysporum f. sp. cubense and Helicotylenchus multicinctus in banana plantation. BioControl 59, 345–355. doi: 10.1007/s10526-014-9569-8
Shafi, J., Tian, H., and Ji, M. (2017). Bacillus species as versatile weapons for plant pathogens: a review. Biotechnol. Biotechnol. Equip. 31, 446–459. doi: 10.1080/13102818.2017.1286950
Shamarao, J., Siddaramaiah, A. L., and Ramaswamy, G. R. (2001). Influence of biocontrol agents and MPG-3 on Fusarium oxysporum f. sp. cubense, incitant of Panama disease of banana. Plant Dis. Res. 16, 68–72.
Shen, L., Xiong, G., Dong, L., Kong, R., Guo, T., and Zhang, S. (2013). Identification of a strain HN-1 against banana wilt disease and determination of its antagonism. Plant Dis. Pests 4, 12–16.
Shen, Z., Ruan, Y., Chao, X., Zhang, J., Li, R., and Shen, Q. (2015a). Rhizosphere microbial community manipulated by 2 years of consecutive biofertilizer application associated with banana Fusarium wilt disease suppression. Biol. Fertil. Soils 51, 553–562. doi: 10.1007/s00374-015-1002-7
Shen, Z., Ruan, Y., Xue, C., Zhong, S., Li, R., and Shen, Q. (2015b). Soils naturally suppressive to banana Fusarium wilt disease harbor unique bacterial communities. Plant Soil 393, 21–33. doi: 10.1007/s11104-015-2474-9
Shen, Z., Xue, C., Penton, C. R., Thomashow, L. S., Zhang, N., Wang, B., et al. (2019). Suppression of banana Panama disease induced by soil microbiome reconstruction through an integrated agricultural strategy. Soil Biol. Biochem. 128, 164–174. doi: 10.1016/j.soilbio.2018.10.016
Shen, Z., Zhong, S., Wang, Y., Wang, B., Mei, X., Li, R., et al. (2013). Induced soil microbial suppression of banana Fusarium wilt disease using compost and biofertilizers to improve yield and quality. Eur. J. Soil Biol. 57, 1–8. doi: 10.1016/j.ejsobi.2013.03.006
Shih, H. D., Chung, W. C., Huang, H. C., Tseng, M., and Huang, J. W. (2013). Identification for Streptomyces padanus strain PMS-702 as a biopesticide agent. Plant Pathol. Bull. 22, 145–158.
Siamak, S. B., and Zheng, S. (2018). Banana Fusarium wilt (Fusarium oxysporum f. sp. cubense) control and resistance, in the context of developing wilt-resistant bananas within sustainable production systems. Hortic. Plant J. 4, 208–218. doi: 10.1016/j.hpj.2018.08.001
Singh, H. (2014). Management of plant pathogens with microorganisms. Proc. Indian Natn. Sci. Acad. 80, 443–454. doi: 10.16943/ptinsa/2014/v80i2/55120
Singh, R., Kumar, M., Mittal, A., and Mehta, P. K. (2017). Microbial metabolites in nutrition, healthcare and agriculture. 3 Biotech 7, 15–15. doi: 10.1007/s13205-016-0586-4
Sivamani, E., and Gnanamanickam, S. S. (1988). Biological control of Fusarium oxysporum f. sp. cubense in banana by inoculation with Pseudomonas fluorescens. Plant Soil 107, 3–9. doi: 10.1007/BF02371537
Smith, L., O'Neill, W., Kochman, J., Lehane, J., and Salmond, G. (2005). Silicon shows promise for Fusarium wilt suppression. Austr. Cottongr. 26, 50–52.
Somu, R., Thammaiah, N., Swamy, G. S. K., Kulkarni, M. S., and Devappa, V. (2014). In vitro evaluation of fungicides against Fusarium oxysporum f. sp. cubense. Int. J. Plant Prot. 7, 221–224.
Souza, A., Cruz, J. C., Sousa, N. R., Procópio, A. R. L., and Silva, G. F. (2014). Endophytic bacteria from banana cultivars and their antifungal activity. Genet. Mol. Res. 13, 8661–8670. doi: 10.4238/2014.October.27.6
Stover, R. H. (1962). Studies on Fusarium wilt of bananas: VIII. Differentiation of clones by cultural interaction and volatile substances. Can. J. Bot. 40, 1467–1471. doi: 10.1139/b62-142
Suhaimi, N. S. M., Goh, S. Y., Ajam, N., Othman, R. Y., Chan, K. G., and Thong, K. L. (2017). Diversity of microbiota associated with symptomatic and non-symptomatic bacterial wilt-diseased banana plants determined using 16S rRNA metagenome sequencing. World J. Microbiol. Biotechnol. 33:168. doi: 10.1007/s11274-017-2336-0
Sun, J., Wang, Y., Zhao, P., and Peng, M. (2010). Isolation, identification and antagonistic acitivity of biocontrol bacterium against Fusarium oxysporum f. sp. cubense. Chin. J. Biol. Control 26, 347–351.
Sun, J. B., Peng, M., Wang, Y. G., Zhao, P. J., and Xia, Q. Y. (2011). Isolation and characterization of antagonistic bacteria against fusarium wilt and induction of defense related enzymes in banana. Afr. J. Microbiol. Res. 5, 509–515. doi: 10.5897/AJMR10.607
Sun, T., and Hsieh, F. (2015). First record of Eutypella sp. as a mycoparasite on Fusarium oxysporum f. sp. cubense. Plant Prot. Bull. Taiwan 57, 25–30.
Sun, Y., Yi, X. P., Peng, M., Zeng, H. C., Wang, D., Li, B., et al. (2014). Proteomics of Fusarium oxysporum race 1 and race 4 reveals enzymes involved in carbohydrate metabolism and ion transport that might play important roles in banana Fusarium wilt. PLoS ONE 9:e113818. doi: 10.1371/journal.pone.0113818
Sun, Z., Ji, C., Li, Y., and Wang, Z. (2008). Antagonistic rhizobacteria strain Bacillus subtilis S-1 against banana Fusarium wilt. Chin. J. Biol. Control 24, 143–147.
Sun, Z., and Wang, Z. (2009). Characterization, identification and control effect against banana Fusarium wilt of antagonistic strain C-4. Acta Phytophylacica Sinica 36, 392–396.
Tan, D., Fu, L., Han, B., Sun, X., Zheng, P., and Zhang, J. (2015). Identification of an endophytic antifungal bacterial strain isolated from the rubber tree and its application in the biological control of banana Fusarium wilt. PLoS ONE 10:e0131974. doi: 10.1371/journal.pone.0131974
Tan, Z., Lin, B., and Zhang, R. (2013). A novel antifungal protein of Bacillus subtilis B25. SpringerPlus 2:543. doi: 10.1186/2193-1801-2-543
Thangavelu, R., and Gopi, M. (2015a). Combined application of native Trichoderma isolates possessing multiple functions for the control of Fusarium wilt disease in banana cv. Grand Naine. Biocontrol Sci. Technol. 25, 1147–1164. doi: 10.1080/09583157.2015.1036727
Thangavelu, R., and Gopi, M. (2015b). Field suppression of Fusarium wilt disease in banana by the combined application of native endophytic and rhizospheric bacterial isolates possessing multiple functions. Phytopathol. Mediterr. 54, 241–252. doi: 10.14601/Phytopathol_Mediterr-15160
Thangavelu, R., and Jayanthi, A. (2009). RFLP analysis of rDNA-ITS regions of native non-pathogenic Fusarium oxysporum isolates and their field evaluation for the suppression of Fusarium wilt disease of banana. Australas. Plant Pathol. 38, 13–21. doi: 10.1071/AP08071
Thangavelu, R., and Mustaffa, M. (2010). A potential isolate of Trichoderma viride NRCB1 and its mass production for the effective management of Fusarium wilt disease in banana. Tree For. Sci. Biotechnol. 4, 76–84.
Thangavelu, R., Palaniswami, A., Doraiswamy, S., and Velazhahan, R. (2003). The effect of Pseudomonas fluorescens and Fusarium oxysporum f. sp. cubense on induction of defense enzymes and phenolics in banana. Biol. Plant. 46, 107–112. doi: 10.1023/A:1022374520121
Thangavelu, R., Palaniswami, A., Ramakrishnan, G., Sabitha, D., Muthukrishnan, S., and Velazhahan, R. (2001). Involvement of fusaric acid detoxification by Pseudomonas fluorescens strain Pf10 in the biological control of Fusarium wilt of banana caused by Fusarium oxysporum f. sp. cubense. Z. Pflanzenk. Pflanzen. 108, 433–445.
Thangavelu, R., Varun, G., and Ganga Devi, P. (2016). Identification of differentially expressed genes from Fusarium oxysporum f. sp. cubense and Trichoderma asperellum (prr2) interaction in the susceptible banana cultivar Grand Naine. Turkish J. Bot. 40, 480–487. doi: 10.3906/bot-1511-19
Thaysen, A. C., and Butlin, K. R. (1945). Inhibition of the development of Fusarium oxysporum cubense by a growth substance produced by Meredith's actinomycetes. Nature 156, 781–782. doi: 10.1038/156781b0
Ting, A. S. Y., Fang, M. T., and Tee, C. S. (2009a). Assessment on the effect of formulative materials on the viability and efficacy of Serratia marcescens- a biocontrol agent against Fusarium oxysporum f. sp. cubense race 4. Am. J. Agr. Biol. Sci. 4, 283–288. doi: 10.3844/ajabssp.2009.283.288
Ting, A. S. Y., Fang, M. T., and Tee, C. S. (2011). Efficacy of clay-based formulated Serratia in reducing inoculum of Fusarium oxysporum f. sp. cubense tropical race 4. Acta Hortic., 421–426. doi: 10.17660/ActaHortic.2011.897.58
Ting, A. S. Y., Mah, S. W., and Tee, C. S. (2009b). Prevalence of endophytes antagonistic towards Fusarium oxysporum f. sp. cubense race 4 in various plants. Am.-Eurasian J. Sustain. Agr. 3, 399–406.
Ting, A. S. Y., Mah, S. W., and Tee, C. S. (2010). Identification of volatile metabolites from fungal endophytes with biocontrol potential towards Fusarium oxysporum f. sp. cubense race 4. Am. J. Agr. Biol. Sci. 5, 177–182. doi: 10.3844/ajabssp.2010.177.182
Ting, A. S. Y., Mah, S. W., and Tee, C. S. (2012). Evaluating the feasibility of induced host resistance by endophytic isolate Penicillium citrinum BTF08 as a control mechanism for Fusarium wilt in banana plantlets. Biol. Control 61, 155–159. doi: 10.1016/j.biocontrol.2012.01.010
Ting, A. S. Y., Meon, S., Jugah, K., and Anuar, A. R. (2003). Effect of artificially induced suppressive soil on Fusarium wilt. Infomusa 12, 33–34.
Ting, A. S. Y., Sariah, M., Kadir, J., and Gurmit, S. (2009c). Field evaluation of non-pathogenic Fusarium oxysporum isolates UPM31P1 and UPM39B3 for the control of Fusarium wilt in ‘Pisang Berangan’ (Musa, AAA). Acta Hortic. 828, 139–144. doi: 10.17660/ActaHortic.2009.828.13
Ting, S., Mah, S., and Tee, C. (2011). Detection of potential volatile inhibitory compounds produced by endobacteria with biocontrol properties towards Fusarium oxysporum f. sp. cubense race 4. World J. Microbiol. Biotechnol. 27, 229–235. doi: 10.1007/s11274-010-0447-y
Usharani, T. R., and Gowda, T. K. S. (2011). Cloning of chitinase gene from Bacillus thuringiensis. Indian J. Biotechnol. 10, 264–269. doi: 10.2323/jgam.43.341
Van den Bergh, I., Lehrer, K., and Vézina, A. (2018). Promusa. Montpellier: Bioversity International. Available online at: http://www.promusa.org/ (accessed December 3, 2018).
Vézina, A. (2018a). “Tropical race 4,” in Musapedia, the Banana Knowledge Compedium (Montpellier: Bioversity International). Available online at: http://www.promusa.org/Tropical+race+4+-+TR4 (accessed November 14, 2018).
Vézina, A. (2018b). “Uganda,” in Musapedia, the Banana Knowledge Compedium (Montpellier, France: Bioversity International). Available online at: http://www.promusa.org/Uganda (accessed December 23, 2018).
Vézina, A. (2018c). “The year of TR4,” in Musapedia, the Banana Knowledge Compedium (Montpellier: Bioversity International). Available online at: http://www.promusa.org/Tropical+race+4+-+TR4 (accessed: November 30, 2018).
Vidhyasekaran, P., and Muthamilan, M. (1995). Development of formulations of Pseudomonas fluorescens for control of chickpea wilt. Plant Dis. 79, 782–786. doi: 10.1094/PD-79-0782
Vidhyasekaran, P., and Muthamilan, M. (1999). Evaluation of a powder formulation of Pseudomonas fluorescens Pf1 for control of rice sheath blight. Biocontrol Sci. Technol. 9, 67–74. doi: 10.1080/09583159929910
Vidhyasekaran, P., Rabindran, R., Muthamilan, M., Nayar, K., Rajappan, K., Subramanian, N., et al. (1997). Development of a power formulation of Pseudomonas fluorescens for control of rice blast. Plant Pathol. 46, 291–297. doi: 10.1046/j.1365-3059.1997.d01-27.x
Viljoen, A. (2002). The status of Fusarium wilt (Panama disease) of banana in South Africa. South Afr. J. Sci. 98, 341–344.
Viljoen, A., Karamura, E., and Swennen, R. (2018). Banana Fusarium wilt in Africa. Stellenbosch: Stellenbosch University. Available online at: http://www.sun.ac.za/english/faculty/agri/plant-pathology/ac4tr4/Pages/default.aspx (accessed December 3, 2018).
Vinale, F., Sivasithamparam, K., Ghisalberti, E. L., Marra, R., Woo, S. L., and Lorito, M. (2008). Trichoderma-plant pathogen interactions. Soil Biol. Biochem. 40, 1–10. doi: 10.1016/j.soilbio.2007.07.002
Waite, B. H., and Stover, R. H. (1960). Studies on Fusarium wilt of bananas: VI. Variability and the cultivar concept in Fusarium oxysporum f. sp. cubense. Can. J. Bot. 38, 985–994. doi: 10.1139/b60-087
Wang, B., Li, R., Ruan, Y., Ou, Y., Zhao, Y., and Shen, Q. (2015). Pineapple-banana rotation reduced the amount of Fusarium oxysporum more than maize-banana rotation mainly through modulating fungal communities. Soil Biol. Biochem. 86, 77–86. doi: 10.1016/j.soilbio.2015.02.021
Wang, B., Shen, Z., Zhang, F., Raza, W., Yuan, J., Huang, R., et al. (2016). Bacillus amyloliquefaciens strain W19 can promote growth and yield and suppress Fusarium wilt in banana under greenhouse and field conditions. Pedosphere 26, 733–744. doi: 10.1016/S1002-0160(15)60083-2
Wang, B., Yuan, J., Zhang, J., Shen, Z., Zhang, M., Li, R., et al. (2013). Effects of novel bioorganic fertilizer produced by Bacillus amyloliquefaciens W19 on antagonism of Fusarium wilt of banana. Biol. Fertil. Soils 49, 435–446. doi: 10.1007/s00374-012-0739-5
Wang, J., Zhao, Y., and Ruan, Y. (2015). Effects of bio-organic fertilizers produced by four Bacillus amyloliquefaciens strains on banana fusarium wilt disease. Compost Sci. Util. 23, 185–198. doi: 10.1080/1065657X.2015.1020398
Wang, L., Xing, M., Di, R., and Luo, Y. (2015). Isolation, identification and antifungal activities of Streptomyces aureoverticillatus HN6. J. Plant Pathol. Microbiol. 6, 281. doi: 10.4172/2157-7471.1000281
Wang, R., Zhang, H., Sun, L., Qi, G., Chen, S., and Zhao, X. (2017). Microbial community composition is related to soil biological and chemical properties and bacterial wilt outbreak. Sci. Rep. 7:343. doi: 10.1038/s41598-017-00472-6
Wang, T., Duan, Y., Liu, B., Guo, G., Zhou, D., Tan, X., et al. (2011). The colonization of two strains of antagonistic bacteria of Fusarium oxysporum in banana. Genomics Appl. Biol. 30, 342–350.
Weber, O. B., Muniz, C. R., Vitor, A. O., Freire, F. C. O., and Oliveira, V. M. (2007). Interaction of endophytic diazotrophic bacteria and Fusarium oxysporum f. sp. cubense on plantlets of banana ‘Maçã’. Plant Soil 298, 47–56. doi: 10.1007/s11104-007-9335-0
Wei, S., Zhang, Z., Tu, X., He, J., and Tu, G. (2011). Studies on the isolation, identification and activity of anti-Fusarium oxysporum secondary metabolites produced by Streptomyces sp. 702. Acta Agric. Univ. Jiangxiensis 33, 982–986.
Wibowo, A., Santosa, A. T., Subandiyah, S., Hermanto, C., and Taylor, M. F. P. (2013). Control of Fusarium wilt of banana by using Trichoderma harzianum and resistant banana cultivars. Acta Hortic. 975, 173–177. doi: 10.17660/ActaHortic.2013.975.18
Woo, S. L., Ruocco, M., Vinale, F., Nigro, M., Marra, R., Lombardi, N., et al. (2014). Trichoderma-based products and their widespread use in agriculture. Open Mycol. J. 8, 71–126. doi: 10.2174/1874437001408010071
Wu, X., Huang, H., Chen, G., Sun, Q., Peng, J., Zhu, J., et al. (2009). A novel antibiotic produced by Streptomyces noursei Da07210. Antonie Van Leeuwenhoek 96, 109–112. doi: 10.1007/s10482-009-9333-8
Wui, L. (2000). Sustainable disease control approaches in some Malaysian crops. Mar. Highland Biosci. Center Rep. 11, 75–80.
Xue, C., Ryan Penton, C., Shen, Z., Zhang, R., Huang, Q., Li, R., et al. (2015). Manipulating the banana rhizosphere microbiome for biological control of Panama disease. Sci. Rep. 5:11124. doi: 10.1038/srep11124
Yang, C. H., and Crowley, D. E. (2000). Rhizosphere microbial community structure in relation to root location and plant iron nutritional status. Appl. Environ. Microbiol. 66, 345–351. doi: 10.1128/AEM.66.1.345-351.2000
Yang, J., Huang, J., Wang, C., and Feng, Y. (2015). Effects of Allium tuberosum juice on the activities of two cell wall degrading enzymes produced by Fusarium oxysporum f. sp. cubense. J. Zhongkai Univ. Agric. Eng. 28, 64–66.
Yang, Q. S., Gao, J., He, W. D., Dou, T. X., Ding, L. J., Wu, J. H., et al. (2015). Comparative transcriptomics analysis reveals difference of key gene expression between banana and plantain in response to cold stress. BMC Genomics 16:446. doi: 10.1186/s12864-015-1551-z
Yang, X., Chen, F., Gan, L., Du, Y., and Ruan, H. (2010). Effect of the endophytic Bacillus subtilis EBT1 isolated from banana on the growth and resistance to Fusarium wilt disease in banana. Acta Phytophylacica Sinica 37, 300–306.
Yang, X., Chen, F., and Ruan, H. (2006). Screening of antagonistic strains against Fusarium oxysporum f. sp. cubense on banana. Fujian J. Agric. Sci. 21, 105–108.
Yedidia, I., Benhamou, N., Kapulnik, Y., and Chet, I. (2000). Induction and accumulation of PR proteins activity during early stages of root colonization by the mycoparasite Trichoderma harzianum strain T-203. Plant Physiol. Biochem. 38, 863–873. doi: 10.1016/S0981-9428(00)01198-0
Yi, G. J., Huang, B. Z., Xu, L. B., Chen, H. B., Hu, G. B., Xu, C. X., et al. (2007). “Fusarium wilt threatens livelihoods of banana farmers in Southern China,” in RISBAP Bulletin, Vol. 11, eds A. B. Molina, J. T. Oliver, V. N. Roa, and V. G. O. Sinohin (Los Baños: Bioversity International Regional Office for Asia and the Pacific), 1–2.
Yin, X., Chen, D., Wu, H., and Zheng, F. (2009). An endophytic Erwinia chrysanthemi strain antagonistic against banana fusarium wilt disease. Chin. J. Biol. Control 25, 60–65.
Yu, C., Xiao, R., Liu, B., Lin, N., and Chen, L. (2010). Endophytic colonization of biocontrol bacterium FJAT-346-PA and its efficiency against banana Fusarium wilt. Acta Phytophylacica Sinica 37, 493–498.
Yu, G., Cheng, P., Wang, Y., Chen, Y., Chen, Y., and Li, Y. (2010). Control effect of Bacillus subtilis strain TR21 on Panama disease of banana Brazil (Musa spp.) in fields. Chin. J. Biol. Control 26, 497–500.
Yu, J., Zhang, R., Tan, Z., and Lin, B. (2016). Studies on antifungal activity and purification of antifungal substance from Bacillus subtilis B25 strain. Genomics Appl. Biol. 35, 629–634.
Yu, L., Dai, H., Zhao, Y., Zeng, Y., Jiang, W., Mei, W., et al. (2011). Three new compounds from soil actinomycete Streptomyces albospinus 15-4-2. J. Asian Nat. Prod. Res. 13, 901–906. doi: 10.1080/10286020.2011.599322
Yuan, J., Li, B., Zhang, N., Waseem, R., Shen, Q., and Huang, Q. (2012a). Production of bacillomycin- and macrolactin-type antibiotics by Bacillus amyloliquefaciens NJN-6 for suppressing soilborne plant pathogens. J. Agric. Food Chem. 60, 2976–2981. doi: 10.1021/jf204868z
Yuan, J., Raza, W., Huang, Q., and Shen, Q. (2011). Quantification of the antifungal lipopeptide iturin A by high performance liquid chromatography coupled with aqueous two-phase extraction. J. Chromatogr. B 879, 2746–2750. doi: 10.1016/j.jchromb.2011.07.041
Yuan, J., Raza, W., Shen, Q., and Huang, Q. (2012b). Antifungal activity of Bacillus amyloliquefaciens NJN-6 volatile compounds against Fusarium oxysporum f. sp. cubense. Appl. Environ. Microbiol. 78, 5942–5944. doi: 10.1128/AEM.01357-12
Yuan, Y. F., Zhao, Y. P., Yang, J. L., Jiang, Y. M., Lu, F., Jia, Y. X., et al. (2017). Metabolomic analyses of banana during postharvest senescence by H-1-high resolution-NMR. Food Chem. 218, 406–412. doi: 10.1016/j.foodchem.2016.09.080
Zacky, F. A., and Ting, A. S. Y. (2013). Investigating the bioactivity of cells and cell-free extracts of Streptomyces griseus towards Fusarium oxysporum f. sp. cubense race 4. Biol. Control 66, 204–208. doi: 10.1016/j.biocontrol.2013.06.001
Zacky, F. A., and Ting, S. (2015). Biocontrol of Fusarium oxysporum f. sp. cubense tropical race 4 by formulated cells and cell-free extracts of Streptomyces griseus in sterile soil environment. Biocontrol Sci. Technol. 25, 685–696. doi: 10.1080/09583157.2015.1007921
Zhai, Y., Wang, W., Tan, H., and Cao, L. (2016). A new approach to analyzing endophytic actinobacterial population in the roots of banana plants (Musa sp., AAA). J. Biochem. Mol. Biol. Res. 2, 180–184. doi: 10.17554/j.issn.2313-7177.2016.02.30
Zhang, H., Mallik, A., and Zeng, R. (2013). Control of Panama disease of banana by rotating and intercropping with Chinese chive (Allium tuberosum Rottler): role of plant volatiles. J. Chem. Ecol. 39, 243–252. doi: 10.1007/s10886-013-0243-x
Zhang, L., Yuan, T., Wang, Y., Zhang, D., Bai, T., Xu, S., et al. (2018). Identification and evaluation of resistance to Fusarium oxysporum f. sp. cubense tropical race 4 in Musa acuminata Pahang. Euphytica 214:106. doi: 10.1007/s10681-018-2185-4
Zhang, N., Wu, K., He, X., Li, S.-Q., Zhang, Z.-H., Shen, B., et al. (2011). A new bioorganic fertilizer can effectively control banana wilt by strong colonization with Bacillus subtilis N11. Plant Soil 344, 87–97. doi: 10.1007/s11104-011-0729-7
Zhao, M., Yuan, J., Zhang, R., Dong, M., Deng, X., Zhu, C., et al. (2018). Microflora that harbor the NRPS gene are responsible for Fusarium wilt disease-suppressive soil. Appl. Soil Ecol. 132, 83–90. doi: 10.1016/j.apsoil.2018.08.022
Zheng, S.-J., García-Bastidas, F. A., Li, X., Zeng, L., Bai, T., Xu, S., et al. (2018). New geographical insights of the latest expansion of Fusarium oxysporum f. sp. cubense tropical race 4 into the greater Mekong subregion. Front. Plant Sci. 9:457. doi: 10.3389/fpls.2018.00457
Keywords: Musa acuminata, Fusarium oxysporum f. sp. cubense, Panama disease, soil microbiota, beneficial microorganisms, biocontrol
Citation: Bubici G, Kaushal M, Prigigallo MI, Gómez-Lama Cabanás C and Mercado-Blanco J (2019) Biological Control Agents Against Fusarium Wilt of Banana. Front. Microbiol. 10:616. doi: 10.3389/fmicb.2019.00616
Received: 29 December 2018; Accepted: 11 March 2019;
Published: 05 April 2019.
Edited by:
Miguel Dita, Brazilian Agricultural Research Corporation (EMBRAPA), BrazilReviewed by:
Rong Li, Nanjing Agricultural University, ChinaSijun Zheng, Yunnan Academy of Agricultural Sciences, China
Copyright © 2019 Bubici, Kaushal, Prigigallo, Gómez-Lama Cabanás and Mercado-Blanco. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Giovanni Bubici, Z2lvdmFubmluaWNvbGEuYnViaWNpQGNuci5pdA==