
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 22 January 2019
Sec. Virology
Volume 9 - 2018 | https://doi.org/10.3389/fmicb.2018.03327
 Mauricio Teixeira Lima1
Mauricio Teixeira Lima1 Graziele Pereira Oliveira1
Graziele Pereira Oliveira1 José Augusto Bastos Afonso2Rodolfo José Cavancanti Souto2Carla Lopes de Mendonça2Antonio Flavio Medeiros Dantas3
José Augusto Bastos Afonso2Rodolfo José Cavancanti Souto2Carla Lopes de Mendonça2Antonio Flavio Medeiros Dantas3 Jonatas Santos Abrahao1
Jonatas Santos Abrahao1 Erna Geessien Kroon1*
Erna Geessien Kroon1*Even nearly forty years after the eradication of smallpox, members of the Poxviridae family continue to be the focus of an increasing number of studies. Among these studies, prominently stands vaccinia virus, an orthopoxvirus that is associated with bovine vaccinia outbreaks. Although more frequently associated with infections in cattle and humans, the host range of vaccinia virus is not restricted only to these hosts. There are several instances of molecular and serological evidence of circulation of vaccinia virus among wildlife species. In addition, viral isolation has confirmed a broad spectrum of vaccinia virus hosts. In this report, we provide a brief update on the host range of Brazilian vaccinia virus, and present a case description of an outbreak in domestic buffalo calves from Northeastern Brazil that corroborates previous serological and molecular studies. Furthermore, in the present study, vaccinia virus has been isolated for the first time in buffaloes, and referred to as vaccinia virus Pernambuco (VACV-PE). Phylogenetic reconstruction was based on A56R clustered VACV-PE with vaccinia virus isolates belonging to group 1 Brazilian vaccinia virus. Furthermore, the vaccinia virus genome was detected in the milk of a lactating cow, which thereby revealed a pathway for future studies on the possible impact of vaccinia virus on buffalo milk and milk products. Taken together, these results provide the first description of clinical disease caused by vaccinia virus in buffaloes in South America. They also raise new questions about the chain of transmission of this virus.
Forty years after the eradication of smallpox, many species of genus Orthopoxvirus (OPV) are still relevant and have a considerable impact on human and veterinary health (Essbauer and Meyer, 2010). Currently, OPV has emerged and has been re-emerging around the world as zoonotic agents, including cowpox virus (CPXV) in Europe (Coras et al., 2005; Eder et al., 2017), monkeypox virus (MPXV) primarily in Africa (Reynolds et al., 2012; Wilson et al., 2014), and vaccinia virus (VACV) in Asia and South America (Singh et al., 2007; Kroon et al., 2011). In contrast to variola virus, which is restricted to humans, CPXV, MPXV, and VACV can infect a large range of hosts (Oliveira et al., 2017). These species are able to infect different groups of mammals, including exotic hosts, as occurred during outbreaks of CPXV in zoos of Europe (Marennikova et al., 1977; Pilaski and Rösen-Wolff, 1988; Kik and Luten, 2009). In addition, MPXV is able to infect prairie dogs from North America, indicating that these viruses can infect new hosts in new environments (Guarner et al., 2004; Kile et al., 2005). Thus, in a region with great biodiversity and wildlife, in addition to the presence of several domestic and synanthropic species, these viruses could have an impressive number of host species. The natural history of VACV in Brazil reflects this, and new hosts have been described over the years, as reviewed in Figure 1.
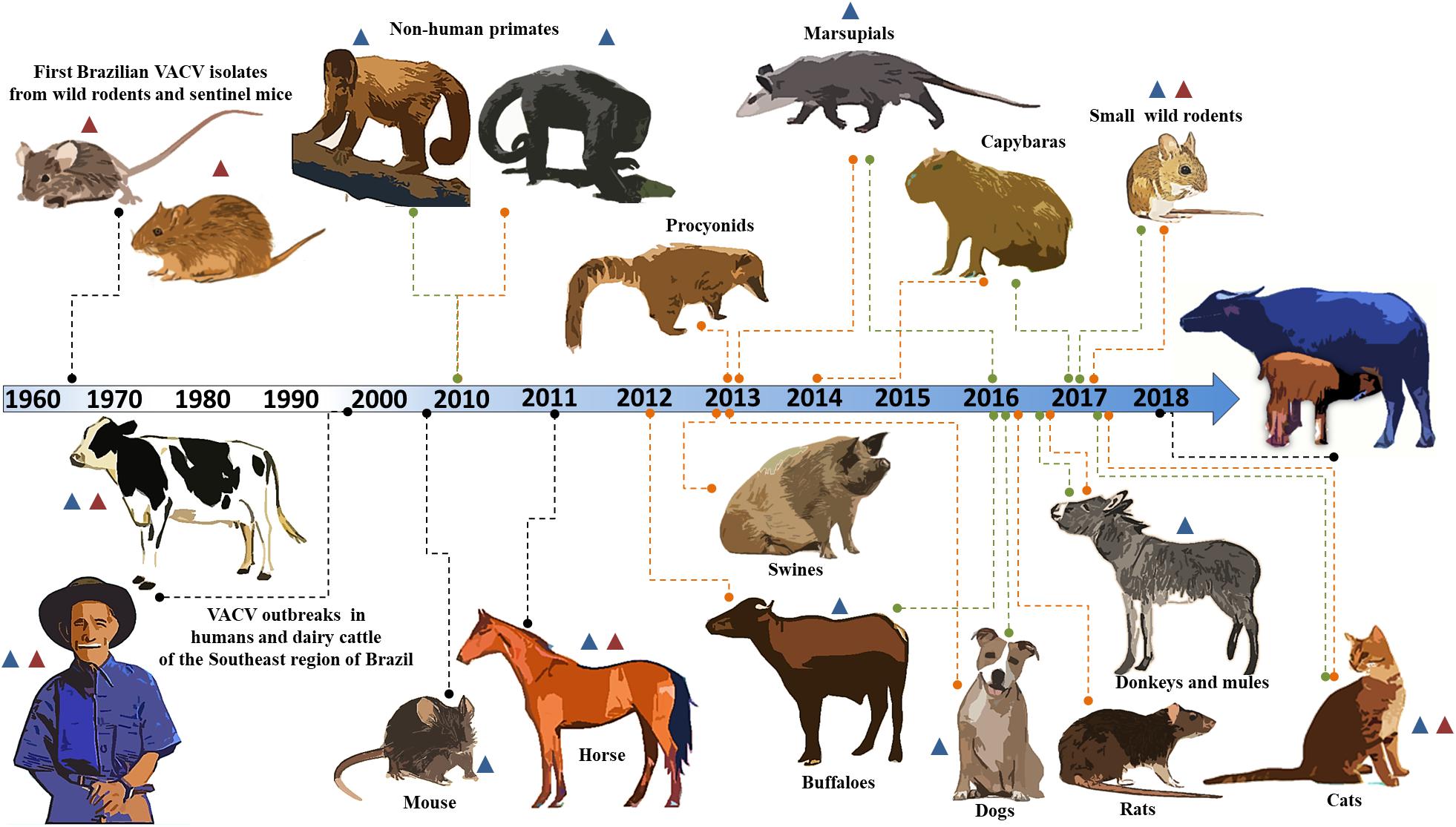
Figure 1. Timeline of identification of the host range of the Brazilian vaccinia virus. A timeline of Brazilian VACV circulation is shown. The orange and green lines indicate serological and molecular evidence, respectively. The black lines indicate viral isolation. The blue and red triangles represent circulation of the VACV GI (group I) and VACV GII (group II), respectively. The years for each study are indicated along the timeline.
Here, we present a brief update on the host range of Brazilian VACV, in addition to a case description of an outbreak in buffalo calves in Brazil. In the 1960s, the Rockefeller Institute for Research on Arboviruses in Brazil first isolated Brazilian VACV from wild rodents and sentinel mice in Pará and São Paulo (Fonseca et al., 1998; da Fonseca et al., 2002; Figure 1). More than three decades had passed before other isolates were described in association with humans and dairy cattle (Bos taurus) in the Southeast region of Brazil (Damaso et al., 2000; de Souza Trindade et al., 2003; Figure 1). Vesiculopustular exanthematous disease caused by VACV in these hosts was referred to as bovine vaccinia (BV).
Since then, many outbreaks have been described in Brazil, and related viruses have been characterized biologically and phylogenetically (Damaso et al., 2000; de Souza Trindade et al., 2003; Leite et al., 2005; Megid et al., 2008; Medaglia et al., 2009; Oliveira et al., 2013; Matos et al., 2018). Previous studies have demonstrated that circulating viruses belong to at least two distinct clusters, and these groups were referred to as group I (GI) and group II (GII) of the Brazilian VACV (Trindade et al., 2006; Drumond et al., 2008; Assis et al., 2012; de Souza Trindade et al., 2016). The host range of Brazilian VACV is not restricted to humans and bovines, and several species present molecular and serological evidence of the circulation of the virus.
The detection of anti-OPV antibodies and the VACV genome in wildlife, such as procyonids (Peres et al., 2013), non-human primates (Abrahão et al., 2010), marsupials (Peres et al., 2013, 2016; Miranda et al., 2017), and several species of wild rodents, including capybaras (Peres et al., 2013; Barbosa et al., 2014; Dutra et al., 2017; Miranda et al., 2017), has been described (Figure 1). In domestic and peridomestic environments, VACV has been isolated in a mouse (Abrahão et al., 2009a) and in horses (Brum et al., 2010; Campos et al., 2011); and serologically and/or molecularly detected in cats (Costa et al., 2017), dogs (Peres et al., 2016), swine (Peres et al., 2013), rats (Babolin et al., 2016), donkeys (Abrahão et al., 2017), and buffaloes (de Assis et al., 2012; Franco-Luiz et al., 2016a; Figure 1). The VACV GI has been associated with most of the hosts described to date; however, VACV GII has also been shown to be circulating in humans, cattle, horses, cats, and wild rodents (Fonseca et al., 1998; Trindade et al., 2006; Campos et al., 2011; de Souza Trindade et al., 2016; Costa et al., 2017; Miranda et al., 2017; Figure 1). In addition, previous studies have demonstrated co-circulation of VACV from these two groups during the same outbreak, and VACV co-infection of humans, cattle, and horses (Trindade et al., 2006; Campos et al., 2011; Oliveira et al., 2015; Lima et al., 2018).
During the last few years, VACV has been described in other South American countries besides Brazil. These include Argentina and Uruguay (Franco-Luiz et al., 2014, 2016b), where evidence of VACV circulation in dairy cattle has been described; and Colombia, where BV outbreaks have affected dairy workers (Usme-Ciro et al., 2017). Similar to the situation in South America, circulation of VACV has been described in Asian countries (Singh et al., 2012). Asiatic VACV isolates have been referred to as buffalopox virus (BPXV), and have caused similar BV outbreaks in rural areas of India and Pakistan (Singh et al., 2007; Zafar et al., 2007). The outbreaks caused by BPXV affect both humans and cattle, but primarily domestic water buffaloes (Bubalus bubalis) (Yadav et al., 2010; Singh et al., 2012). The water buffalo is a bovid that originated in Asia, and is currently found on all continents, except Antarctica (Cockrill, 1981). Studies show that buffaloes have greater resistance to common bovine diseases and show superior weight gain in comparison to cattle, making it also economically superior (Sheikh et al., 2006). Introduced in Brazil in 1895, the water buffalo presented feral and domestic populations that have been extensively used there for meat and dairy production (Long, 2003; Sheikh et al., 2006).
In the present study, we describe a VACV outbreak in domestic buffalo calves from Northeastern Brazil that corroborates previous serological and molecular studies. Furthermore, we describe the first isolation of Brazilian VACV from buffaloes.
In September 2017, three buffalo calves on a farm in Ribeirão County, Pernambuco State, Brazil (08°30′11″S and 35°22′26″W) (Figure 2A), presented with vesicular lesions on the lips, gums, and tongue. The lesions were restricted to the oral cavity and lasted for approximately 25 days (Figure 2B). The calves were all female, aged 45 to 90 days and were of the Mediterranean and Mestizo breeds. The region in which the farm is located is a rural area in the Mata Atlântica (Atlantic Forest) biome, a tropical rainforest on the Brazilian Atlantic coast. The region encompasses mainly sugar cane plantations and buffalo livestock areas. The calves sucked the same lactating cow, which had been previously affected by mastitis. Samples of the lesions and serum of each calf were collected, as well as the milk of the cow. All clinical specimens were derived from domestic buffaloes on private properties, and collected by a veterinarian, according to standard sanitary protocols in accordance with the requirements of the National Livestock Agency (Ministério da Agricultura, Pecuária e Abastecimento) and the requirements for animal research of Universidade Federal de Minas Gerais (UFMG), Minas Gerais, Brazil (protocol number 207/2010).
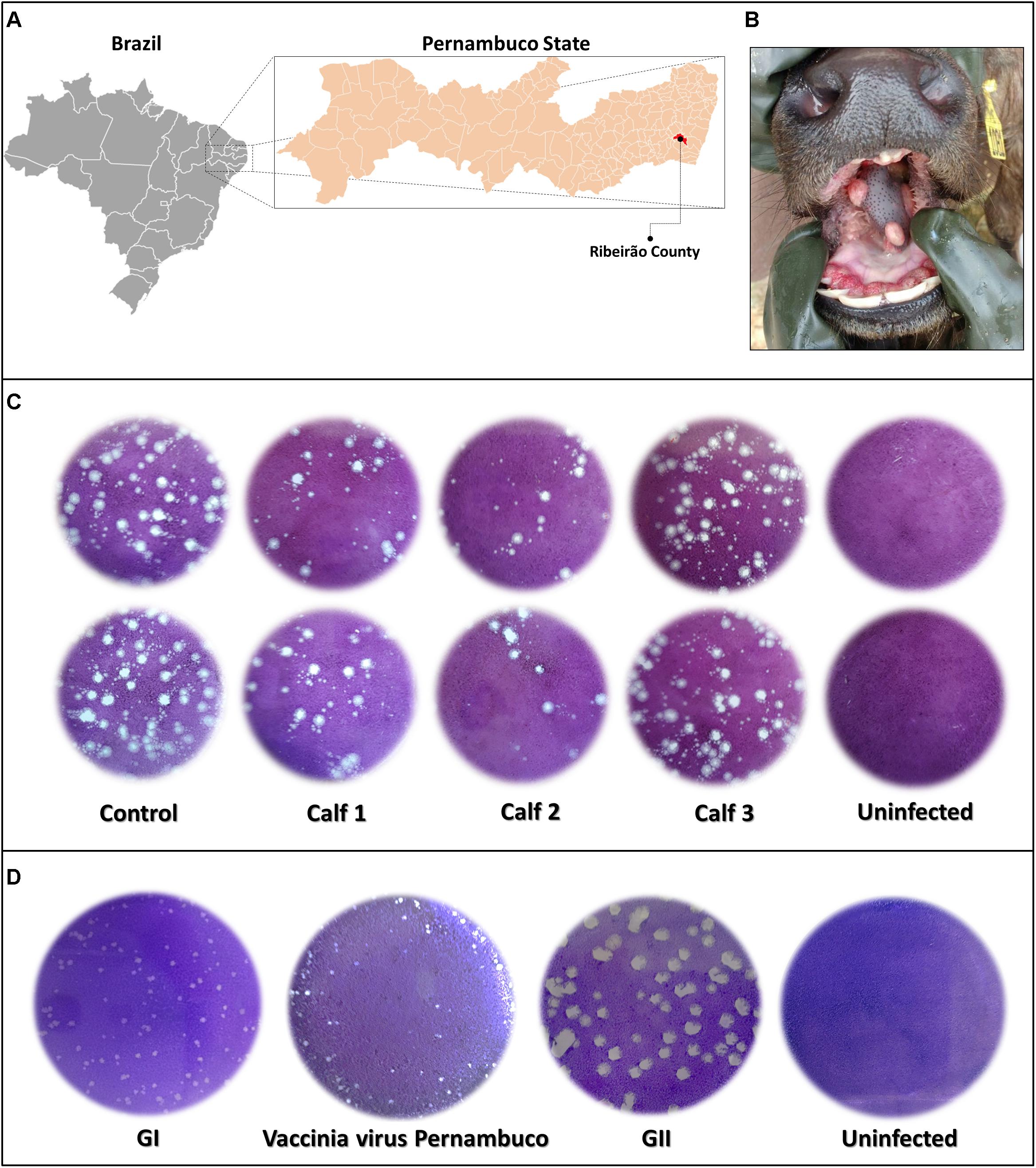
Figure 2. Outbreak localization, lesions, and viral lytic plaque. (A) Map displaying localization of the outbreak in Ribeirão County, Pernambuco State, Brazil (08°30′11″S and 35°22′26″W) and incidence of vesicular lesions on the tongue of buffalo calves (B). Plaque reduction neutralization test of calves’ serum samples diluted in a 1:40 ratio (C), and plaque phenotypic assay of vaccinia Pernambuco isolate and control Brazilian vaccinia viruses of GI and GII (groups I and II) (D).
African green monkey kidney BSC-40 cells [American Type Culture Collection (ATCC) CRL-2761] and Vero cells (ATCC CCL-81) were maintained in a 5% CO2 atmosphere at 37°C, in Eagle’s minimum essential medium (MEM) (Gibco BRL, Invitrogen, Carlsbad, CA, United States), supplemented with 5% fetal bovine serum (Cultilab, Brazil); 2.5 μg/mL amphotericin B (Fungizone) (Cristalia, São Paulo, Brazil); 500 U/mL penicillin (Cristalia); and 50 μg/mL streptomycin (Schering-Plough, São Paulo, Brazil). Vero cells were used for viral isolation. The BSC-40 cells were used for the neutralization test. The vaccinia virus Western Reserve (VACV-WR) virus was kindly provided by Dr. C. Jungwirth (Universitat Wurzburg, Germany), and was used in the plaque reduction neutralization test (PRNT). The VACV-WR was purified on a sucrose gradient as described by Joklik (1962).
Fragments of the tongue lesions on buffalo calves were macerated in phosphate-buffered saline (PBS), containing amphotericin B (20 μg/mL), penicillin (1000 U/mL), and streptomycin (500 μg/mL) in a ratio of 0.1 g sample/0.9 mL PBS. The macerated fragments were then homogenized using a Mini-BeadBeater-24 (BioSpec, United States), and centrifuged at 3,000 × g for 5 min. The milk was diluted 10× in PBS and homogenized in a vortex apparatus. Vero cells were cultured in 25-cm2 flasks and infected with the specimen supernatants, to isolate the virus at 37°C, until a cytopathic effect was detected. The isolates were obtained after three additional rounds of plaque purification in Vero cells.
Tissue fragments excised from tongue lesions were collected and fixed in 10% buffered formalin. The fragments were routinely processed, sectioned at 5 μm, and then stained with hematoxylin and eosin (H&E).
For the plaque phenotypic assay, BSC40 cells seeded in six-well plates at 90–95% confluence were infected with specific plaque-purified viruses. After 1 h of adsorption (37°C, 5% CO2), monolayers were washed twice with PBS, and overlain with solid medium, prepared by mixing equal proportions of 1% agarose and 2× Eagle’s MEM (Gibco, São Paulo, Brazil), supplemented with 2% FBS (Gibco, São Paulo, Brazil). After 48 h of incubation (37°C, 5% CO2), cells were fixed with formaldehyde and stained with crystal violet for plaque size analysis. The method was developed using control viruses with large and small plaque phenotypes, as previously described (Lima et al., 2018).
For the PRNT, serum samples were heat-inactivated at 56°C for 30 min, initially diluted in a 1:20 ratio in MEM, and incubated at 37°C for 15 h (Newman et al., 2003) with the same volume of MEM containing 100 plaque forming units (pfu) of VACV-WR (1:40 ratio). At the same time, the viral suspension was also incubated with MEM to serve as a control. Bovine serum samples were used as positive and negative controls. Furthermore, 400 μL of this mixture was added to BSC-40 cells seeded in six-well plates, which were incubated for 1 h at 37°C, in a 5% CO2 atmosphere. Thereafter, 2 mL of MEM was added to each well, and they were further incubated under similar conditions for 48 h. The cells were then stained with a solution of crystal violet for 20 min, and the viral plaques were counted. The results were expressed as the highest serum dilution that was able to neutralize at least 70% of the viral plaques (PRNT70).
The DNA from the fragments of tongue lesions, sera, and milk samples was extracted using the phenol/chloroform/isoamyl alcohol method (Kroon et al., 2016). For molecular screening, DNA was subjected to quantitative PCR (qPCR), to amplify the highly conserved OPV vaccinia growth factor gene/C11R (F-5′CGCTACAACAGATATTCCAGCTATCAG3′-R-5′AGCGTGGATACAGTCACCGTGTAA3′) and viral hemagglutinin gene/ A56R (F-5′CATCATCTGGAATTGTCACTACTAAA3′- R-5′ACGGCCGACAATATAATTAATGC3′), as previously described by de Souza Trindade et al. (2008) and Kroon et al. (2016), respectively. The qPCR was performed using the SYBr Green Mix (Applied Biosystems, United States). The PCR amplification of viral gene chemokine binding protein gene (C23L) was achieved by two different reactions previously described by Oliveira et al. (2015): reaction 1 (F-5′GCGTGTCCCCAGGACAAGGT3′-R-5′ATGTCGCTGTCTTTCTCTTCTTCGC3′) amplifying a 124 bp DNA fragment, found in both Brazilian VACV groups; and reaction 2 (F-5′GCGTGTCCCCAGGACAAGGT3′-R-5′CTGGATGGGTCTTG3′), amplifying a 138 bp DNA fragment of GII viruses, but not from GI viruses. The PCR products were fractionated in 8% silver-stained polyacrylamide gel electrophoresis (Sambrook and Russell, 2001). The VACV-PE A56R sequence was amplified using the forward primer (5′TGGATCTACACATTCACCGGA3′) and the reverse primer was previously described by Ropp et al. (1995) (5′-CTAGACTTTGTTTTCTG-3′). The PCR conditions were as follows: 95°C for 10 min; 30 cycles at 95°C for 1 min; 55°C for 1 min; and 72°C for 1 min; followed by 72°C for 10 min. In addition, the EEV type-I membrane glycoprotein gene/B5R (F-5′TTTTAGTGCTGCACAGTG3′-R-5′AGTAAAAATGCTCTAACG3′) was amplified (Drumond et al., 2008). The PCR-amplified A56R and B5R fragments were directly sequenced in both orientations (GenBank B5R Accession Number MK210281 and A56R Accession Number MK210282), and in triplicate using an ABI3730 sequencer (Thermo Fisher Scientific, Waltham, MA, United States). The sequences were aligned with previously published OPV sequences from GenBank using the ClustalW method, and they were manually aligned using the MEGA software version X (Arizona State University, Phoenix, AZ, United States). The jModelTest 2.1.9 software was used to determine which model of evolution was most appropriate for our datasets (Darriba et al., 2012). Phylogenetic trees were constructed according to the maximum likelihood method, using the Hasegawa–Kishino–Yano model of nucleotide substitutions, gamma distribution, 1000 bootstrap replicates, and the MEGA software version X (Arizona State University).
Orthopoxvirus-specific PCR, targeting the C11R gene or A56R genes showed that fragments of the tongue lesions of calves and the milk of the buffalo cow were able to amplify DNA for both genes. The threshold cycle ranged between 26 and 30 for the C11R gene and between 27 and 29 for the A56R gene. However, the serum samples were negative. Serum samples were screened for neutralizing antibodies, using a PRNT70. Two of the calves were positive, showing neutralizing antibodies (Figure 2C).
A fragment of a tongue lesion from one calf was inoculated in Vero cells, and showed the formation of characteristic VACV viral plaques after 3 days of the second round passage. This isolate, which we named vaccinia virus Pernambuco (VACV-PE), was submitted to three additional rounds of plaque purification followed by viral replication in Vero cells and subjected to the plaque phenotypic assay. Only the small plaque phenotype was evident (Figure 2D).
Histopathology of the tongue lesions showed ulcerative superficial glossitis at multifocal areas, with superficial necrosis of the epithelium. This was associated with pustular formations characterized by the presence of eosinophilic debris, interspersed with cellular debris and degenerated neutrophils (Figure 3A). In the lamina propria and adjacent musculature, there was a moderate infiltrate of intact and degenerated neutrophils, sparse eosinophils, and lymphocytes. In addition, hyperplasia was detected, ballooning degeneration of the remaining epithelium, and the presence of circular eosinophilic structures of varying sizes with a light halo in the cytoplasm of keratinocytes interspersed with viral inclusion corpuscles (Figure 3B).
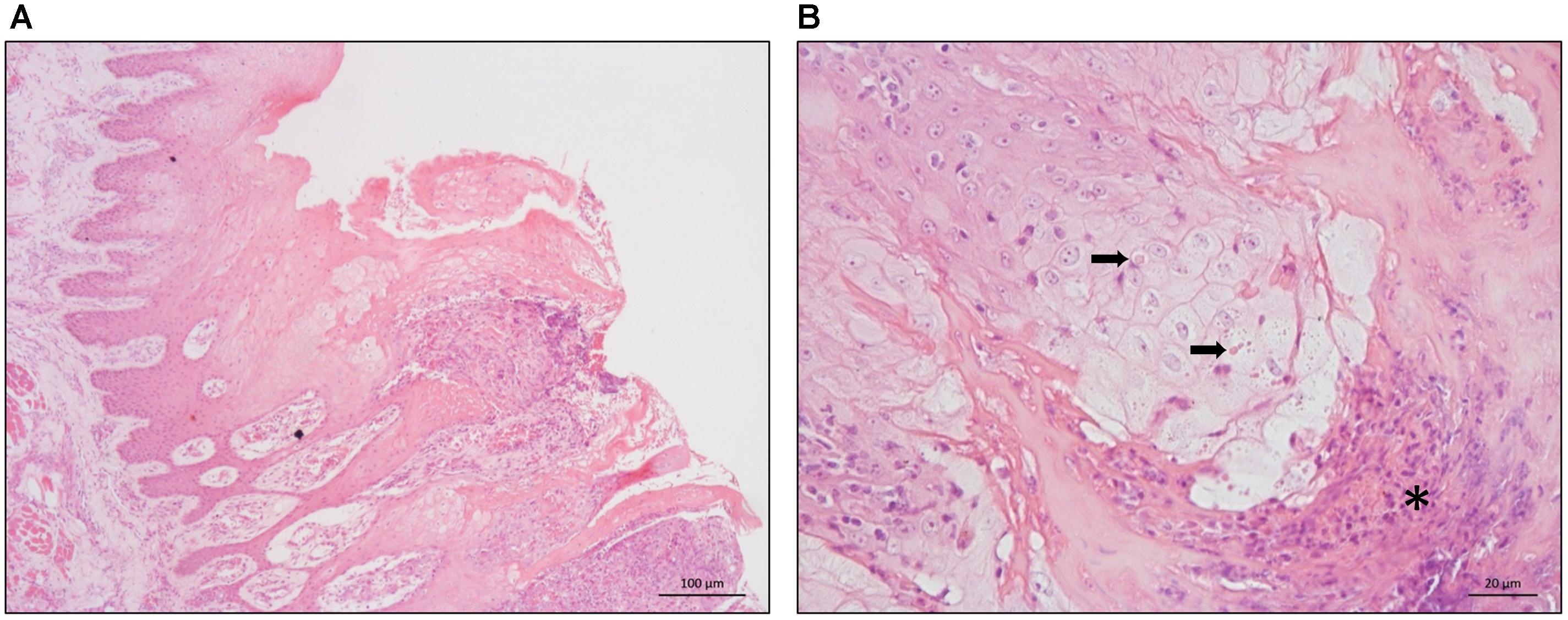
Figure 3. Histopathology of buffalo tongue lesions. (A) Hyperplasia of the epithelium associated with hydropic degeneration of the keratinocytes and pustular formations – 100 μm scale bar. (B) Hydropic degeneration of the epithelium associated with eosinophilic inclusions in the cytoplasm of keratinocytes (arrows) and pustular formations (asterisks) – 20 μm scale bar.
Amplified DNA from the two different reactions for the C23L gene showed a profile similar to that of Brazilian VACV GI, which corroborated the small plaque phenotype (Figure 2D). The DNA amplified in the A56R and B5R PCR was directly sequenced in both orientations. The A56R and B5R genes were analyzed by alignment with sequences from other OPV isolates deposited in GenBank (115 and 36 sequences, respectively). Partial A56R gene nucleotide alignment showed that the VACV-PE sequence contained the signature deletion (18 nt) that was also present in other Brazilian sequences of VACV isolates in GI, but not in GII. Interestingly, this signature was also absent in the sequences of the buffalopox virus of India (Figure 4A). Phylogenetic reconstruction, based on the A56R (Figure 4B) and B5R nucleotide sequences (Figure 4C), also clustered VACV-PE with VACV isolates of GI Brazilian VACV.
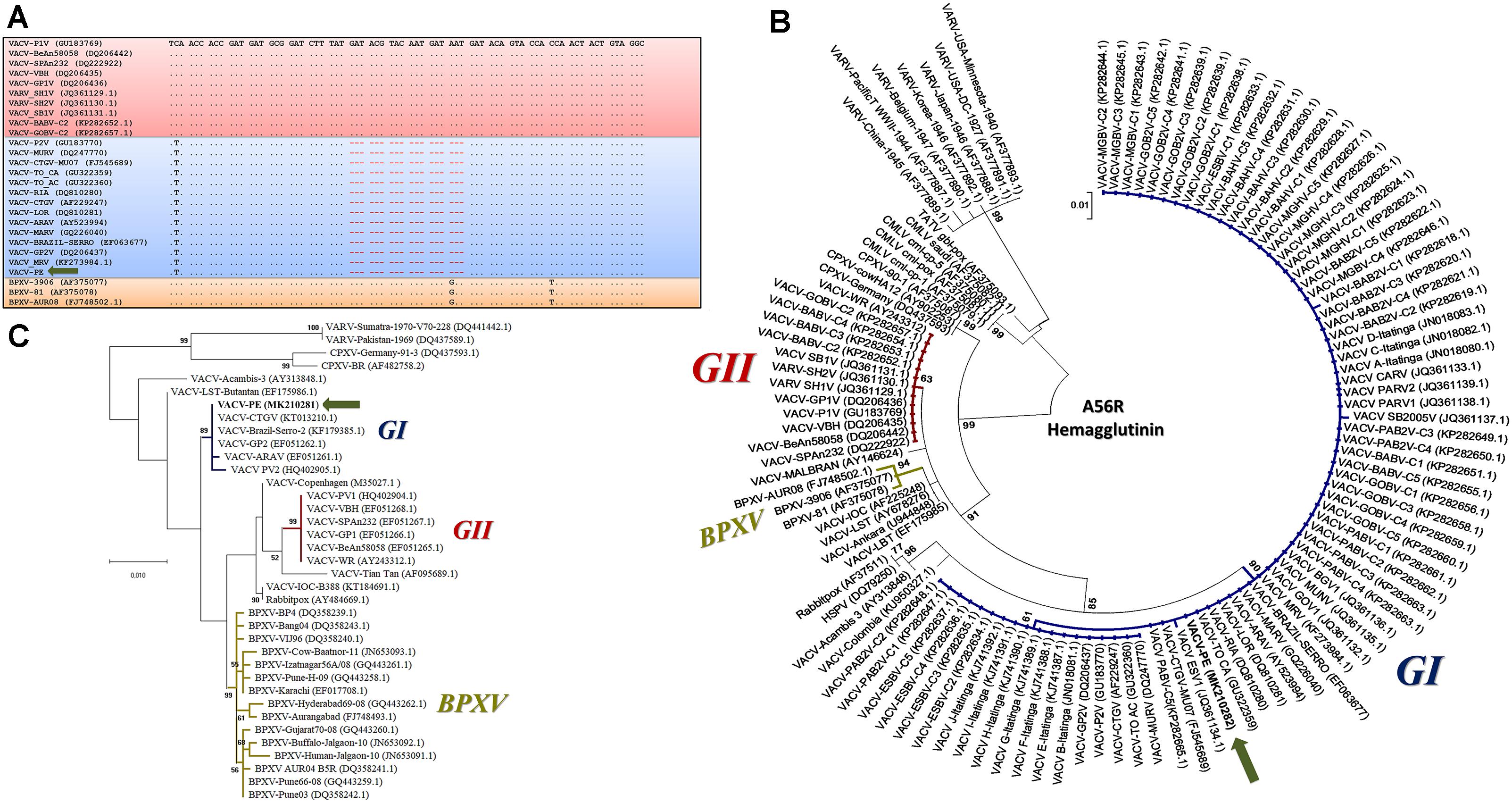
Figure 4. Phylogenetic analyses based on the A56R and B5R gene sequences from vaccinia virus Pernambuco (VACV-PE) and other Orthopoxviruses (OPVs). (A) Alignment of nucleotide sequences from a fragment of the VACV-PE A56R gene with other OPV sequences. The sequences were obtained from GenBank and aligned using the ClustalW method. Nucleotide positions are shown according to the VACV-PV1 (GU183769). The (.) indicates identity and (-) indicates nucleotide deletions. The blue and red boxes highlight GI and GII VACV-BR, respectively, and the orange box highlights the Asiatic buffalopox virus sequences. The VACV-PE isolate is indicated by a green arrow. (B) A56R Phylogenetic tree constructed by the maximum likelihood method, using the Hasegawa–Kishino–Yano model of nucleotide substitutions, gamma distribution, 1000 bootstrap replicates, and the MEGA software version X. The VACV-PE highlights and accession numbers are indicated. The blue, red, and gold subtrees highlight GI and GII VACV-BR, and the Asiatic buffalopox virus sequences, respectively. The VACV-PE isolate is indicated by a green arrow. (C) B5R phylogenetic tree constructed by the maximum likelihood method, using the Hasegawa–Kishino–Yano model of nucleotide substitutions, 1000 bootstrap replicates, and the MEGA software version X. The VACV-PE highlights and accession numbers are indicated. The blue, red, and gold subtrees highlight the GI and GII VACV-BR, and the Asiatic buffalopox virus sequences, respectively. The VACV-PE isolate is identified by a green arrow. The accession numbers are presented between parentheses in respective sequence.
The present study reports the first description of clinical manifestation of VACV in buffaloes in the Western Hemisphere, and the first isolation of Brazilian VACV in a bubaline host. Here, we detected the presence of neutralizing antibodies in buffalo calves (Figure 2C). Only two out of three calves showed neutralizing antibodies in sera. This fact was possibly due to differences in the stage of infection at which each calf was sampled. In infected cows, the VACV antibody response detected by the PRNT was observed 15–21 days post-infection (Gerber et al., 2012). Thus, positive calves could have been sampled at a later stage of infection, whereas the negative calf could have been sampled at a comparatively earlier stage. Previous serologic studies have demonstrated OPV seropositivity in buffalo herds in Southeastern and Northern regions of Brazil (de Assis et al., 2012; Franco-Luiz et al., 2016a). In addition, molecular data have demonstrated that circulating OPV in buffaloes from Northern Brazil is the VACV (Franco-Luiz et al., 2016a). Complementing these data, we demonstrated the presence of VACV in domestic buffaloes in the state of Pernambuco in Northeastern Brazil, further supporting other studies conducted in Brazil. A few VACV studies have been conducted in this region, related only to infections in humans and cattle (Oliveira et al., 2013, 2015; Assis et al., 2015; da Silva et al., 2018).
Analysis of the VACV-PE A56R gene showed that the 18 nt deletion can be used as a molecular signature of Brazilian VACV GI (Figure 4A). The A56R sequences of buffaloes from Ilha do Marajó also showed this deletion and were grouped in GI (Franco-Luiz et al., 2016a). A broad range of hosts has been associated with GI VACV, including humans, cows, cats, dogs, horses, donkeys, marsupials, small rodents, non-human primates, and buffaloes (Damaso et al., 2000; de Souza Trindade et al., 2003; Abrahão et al., 2010, 2017; Campos et al., 2011; Peres et al., 2016; Costa et al., 2017; Miranda et al., 2017; Figure 1). However, phylogenetic analyses of A56R and B5R genes and others, such as C18L, E3L, K3L, and C7L, have demonstrated that BPXV isolates from India are grouped separately in a single cluster apart from the Brazilian isolates and vaccine isolates (Bera et al., 2012; Singh et al., 2012). Our phylogenetic data corroborate these studies, as they demonstrated that the origin of VACV-PE is more closely related to other Brazilian VACV isolates than Asian isolates from buffaloes (Figures 4B,C).
Another noteworthy issue was the detection of viral DNA in the milk of the cow that suckled the calves, as previous studies point to VACV as a possible foodborne pathogen (Abrahão et al., 2009b; Matos et al., 2018). The VACV genome and infectious particles have been previously found in milk samples collected from cows during BV outbreaks (Abrahão et al., 2009b; de Oliveira et al., 2015), and in cows experimentally infected with VACV through healed teat lesions. In those cases, viral DNA was detected in the milk up until 67 days post-infection (de Oliveira et al., 2015). In addition, a study on the ingestion of contaminated milk by mice demonstrated systemic infection in the absence of clinical signs (Rehfeld et al., 2015). Reinforcing this possible route of infection, viable VACV has been detected in artisanal cheese samples produced with the milk of experimentally infected dairy cows (de Oliveira et al., 2018). We suggest that similar circumstances might be evident with dairy products derived from water buffaloes. However, further studies are needed to better characterize and clarify the real impact of VACV in buffalo milk and milk products.
Generally, during BV outbreaks in lactating cows, calves comprise the most severely affected group among the herd showing clinical signs of the disease (Lobato et al., 2005; Matos et al., 2018). Similarly, buffalopox outbreaks occur frequently in female buffaloes and their calves (Singh et al., 2007). In addition, the main route of VACV transmission among cows is through the handling of cow teats by milkers, and a similar trend can be identified in buffalopox cases (Lobato et al., 2005; Singh et al., 2007; Kroon et al., 2011). These similarities between species may indicate that cattle and buffaloes play a similar role in the VACV transmission chain.
In support of these observations, we propose an update of the hypothetical model previously described (Abrahão et al., 2009a; de Oliveira et al., 2017), that highlights the dynamics of VACV circulation and the inclusion of water buffaloes and cattle within the same niche (Figure 5). In this model, rural VACV outbreaks affect buffaloes, humans, and dairy cattle. In addition, other domestic animals, such as horses, cats, and dogs could be implicated in the VACV transmission chain. Peridomestic rodents possibly act as a link for VACV spread between wild and rural environments, thereby promoting transmission among buffaloes, humans, and other farm animals. Furthermore, peridomestic rodents and other wild species, such as capybaras and coatis, can spread VACV and connect urban, wild, and rural areas. Another route of transmission for the VACV of bovine and bubaline animals to humans is through dairy products. This route might be associated with the movement of VACV from the countryside to urban areas (Figure 5). This way, we inserted buffaloes into the hypothetical model of the VACV transmission chain.
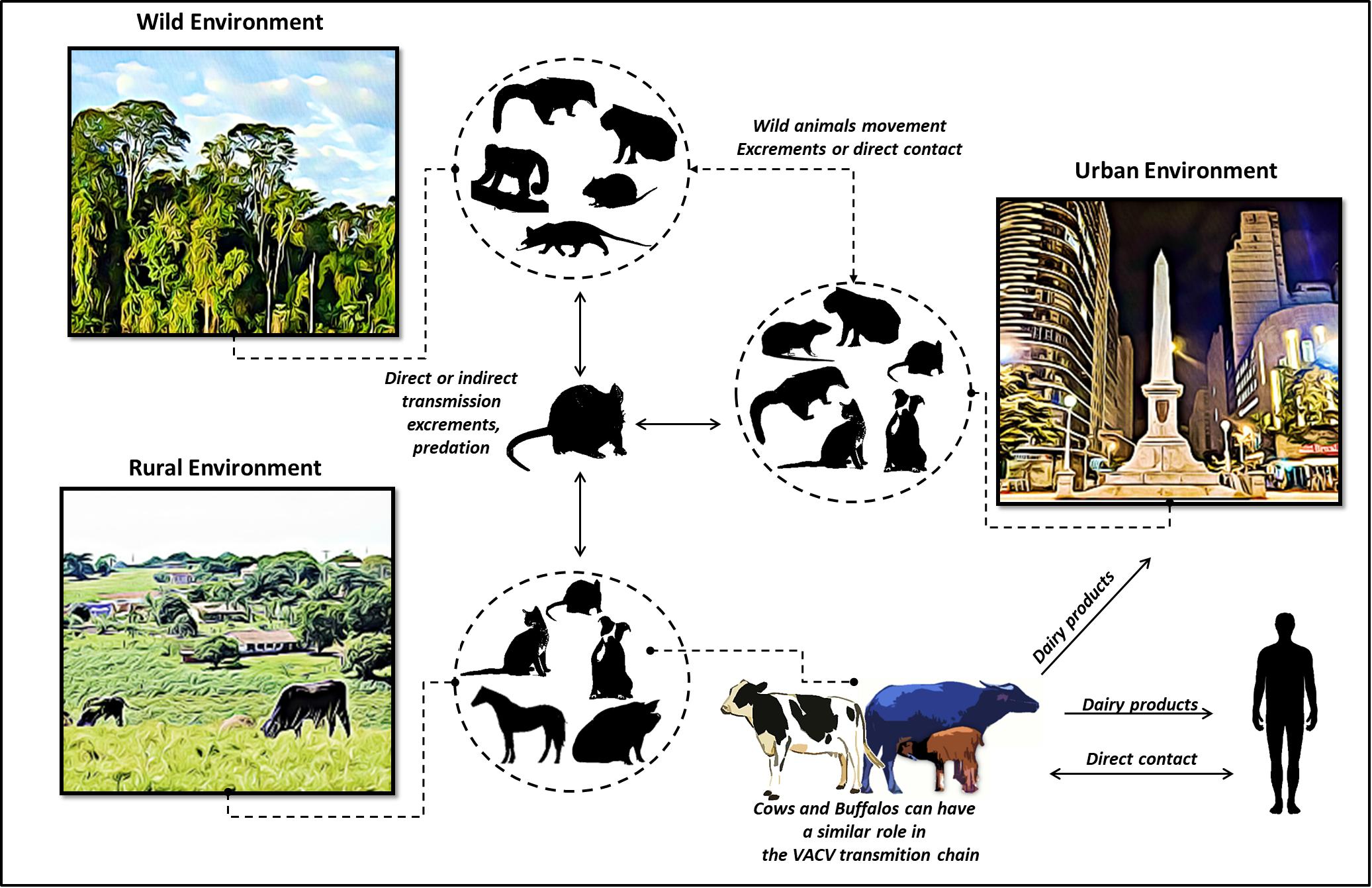
Figure 5. Update of the dynamics of vaccinia virus circulation, represented by a hypothetical model. Wild, rural, and urban environments are shown, and viral association among the different hosts is highlighted. In this updated model, rural VACV outbreaks affecting buffaloes, humans, and dairy cattle are shown. In the VACV transmission cycle, peridomestic rodents can transmit VACV directly or indirectly among buffaloes, humans, and other farm animals, thereby forming links between these environments. Peridomestic rodents and other wild species (capybaras and coatis) could also spread VACV, thus facilitating the connections among urban, wild, and rural areas. In addition, other domestic animals represented in the figure could be implicated in the VACV transmission chain. Another suggested route is that of VACV spread from bovine and bubaline animals to humans through dairy products.
Buffaloes exhibited clinical signs of viral infection that could be confused with those of BV. The possible etiological agents that produce similar clinical signs include foot-and-mouth disease virus, bluetongue virus, vesicular stomatitis viruses, bovine viral diarrhea virus types 1 and 2, bovine papular stomatitis virus, pseudocowpox virus, ovine herpesvirus-2, caprine herpesvirus-2, and bovine herpesviruses 1 and 2 (Klein et al., 2008; Amoroso et al., 2013; Stahel et al., 2013; Laguardia-Nascimento et al., 2016). Based on the economic relevance of strategies to prevent the emergence of BV outbreaks in water buffaloes, efforts to differentiate this viral disease from other similar vesicular diseases are necessary. Thus, proper differential diagnosis is important for all vesicular diseases, mainly to differentiate from foot-and-mouth disease, for which economic and sanitary barriers are necessary (Laguardia-Nascimento et al., 2016).
ML, JA, RS, CdM, AD, JSA, and EK conceived and designed the experiments. ML, GO, JA, and AD performed the experiments. ML, GO, JA, JSA, and EK analyzed the data. ML and EK wrote the manuscript.
This study was supported by the Conselho Nacional de Desenvolvimento Científico e Tecnológico, Pro-Reitoria de Pesquisa da UFMG (PRPq-UFMG), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG), and Ministério da Agricultura, Pecuária e Abastecimento (MAPA). AD, JSA, and EK are CNPq researchers.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank the veterinarian Carlos Urbano Freire and all colleagues from Laboratório de Vírus – UFMG for excellent technical support.
ATCC, American Type Culture Collection; BPXV, buffalopox; BV, bovine vaccinia; CPXV, cowpox virus; GI, Group I; GII, Group II; MEM, minimum essential medium; MPXV, monkeypox virus; OPV, orthopoxvirus; PRNT, plaque reduction neutralization test; VACV, vaccinia virus.
Abrahão, J. S., de Souza Trindade, G., Pereira-Oliveira, G., de Oliveira Figueiredo, P., Costa, G., Moreira Franco-Luiz, A. P., et al. (2017). Detection of Vaccinia virus during an outbreak of exanthemous oral lesions in Brazilian equids. Equine Vet. J. 49, 221–224. doi: 10.1111/evj.12571
Abrahão, J. S., Guedes, M. I. M., Trindade, G. S., Fonseca, F. G., Campos, R. K., Mota, B. F., et al. (2009a). One more piece in the VACV ecological puzzle: could peridomestic rodents be the link between wildlife and bovine vaccinia outbreaks in Brazil? PLoS One 4:e7428. doi: 10.1371/journal.pone.0007428
Abrahão, J. S., Oliveira, T. M. L., Campos, R. K., Madureira, M. C., Kroon, E. G., and Lobato, Z. I. P. (2009b). Bovine vaccinia outbreaks: detection and isolation of Vaccinia virus in milk samples. Foodborne Pathog. Dis. 6, 1141–1146. doi: 10.1089/fpd.2009.0324
Abrahão, J. S., Silva-Fernandes, A. T., Lima, L. S., Campos, R. K., Guedes, M. I. M. C., Cota, M. M. G., et al. (2010). Vaccinia virus infection in monkeys, Brazilian Amazon. Emerg. Infect. Dis. 16, 976–979. doi: 10.3201/eid1606.091187
Amoroso, M. G., Corrado, F., De Carlo, E., Lucibelli, M. G., Martucciello, A., and Galiero, G. (2013). Bubaline herpesvirus 1 associated with abortion in a mediterranean water buffalo. Res. Vet. Sci. 94, 813–816. doi: 10.1016/j.rvsc.2012.12.009
Assis, F. L., Borges, I. A., Peregrino Ferreira, P. C., Bonjardim, C. A., de Souza Trindade, G., Portela Lobato, Z. I., et al. (2012). Group 2 vaccinia virus, Brazil. Emerg. Infect. Dis. 18, 2035–2038. doi: 10.3201/eid1812.120145
Assis, F. L., Franco-Luiz, A. P. M., Paim, L. M., Oliveira, G. P., Pereira, A. F., de Almeida, G. M. F., et al. (2015). Horizontal study of Vaccinia virus infections in an endemic area: epidemiologic, phylogenetic and economic aspects. Arch. Virol. 160, 2703–2708. doi: 10.1007/s00705-015-2549-1
Babolin, L. D S., Almeida-Silva, M. J. F., de Potenza, M. R., Fava, C. Del, Castro, V., Harakava, R., et al. (2016). Zoonosis associated to Rattus rattus and the impacts of the public actions to control the species. Arq. Inst. Biol. 83, 1–7. doi: 10.1590/1808-1657000832014
Barbosa, A. V., Medaglia, M. L. G., Soares, H. S., Quixabeira-Santos, J. C., Gennari, S. M., and Damaso, C. R. (2014). Presence of neutralizing antibodies to orthopoxvirus in capybaras (Hydrochoerus hydrochaeris) in Brazil. J. Infect. Dev. Ctries. 8, 1646–1649. doi: 10.3855/jidc.5216
Bera, B. C., Shanmugasundaram, K., Barua, S., Anand, T., Riyesh, T., Vaid, R. K., et al. (2012). Sequence and phylogenetic analysis of host-range (E3L, K3L, and C7L) and structural protein (B5R) genes of buffalopox virus isolates from buffalo, cattle, and human in India. Virus Genes 45, 488–498. doi: 10.1007/s11262-012-0788-8
Brum, M. C. S., Anjos, B. L., Dos Nogueira, C. E. W., Amaral, L. A, Weiblen, R., and Flores, E. F. (2010). An outbreak of orthopoxvirus-associated disease in horses in southern Brazil. J. Vet. Diagn. Invest. 22, 143–147. doi: 10.1177/104063871002200132
Campos, R. K., Brum, M. C. S., Nogueira, C. E. W., Drumond, B. P., Alves, P. A., Siqueira-Lima, L., et al. (2011). Assessing the variability of Brazilian Vaccinia virus isolates from a horse exanthematic lesion: coinfection with distinct viruses. Arch. Virol. 156, 275–283. doi: 10.1007/s00705-010-0857-z
Cockrill, W. R. (1981). The water buffalo: a review. Br. Vet. J. 137, 8–16. doi: 10.1016/S0007-1935(17)31782-7
Coras, B., Eßbauer, S., Pfeffer, M., Meyer, H., Schröder, J., Stolz, W., et al. (2005). Cowpox and a cat. Lancet 365:446. doi: 10.1016/S0140-6736(05)17836-2
Costa, G. B., Miranda, J. B., Almeida, G. G., de Oliveira, J. S., Pinheiro, M. S., Gonçalves, S. A., et al. (2017). Detection of Vaccinia virus in urban domestic cats, Brazil. Emerg. Infect. Dis. 23, 360–362. doi: 10.3201/eid2302.161341
da Fonseca, F. G., Trindade, G. S., Silva, R. L., Bonjardim, C. A, Ferreira, P. C., and Kroon, E. G. (2002). Characterization of a vaccinia-like virus isolated in a Brazilian forest. J. Gen. Virol. 83(Pt 1), 223–228 doi: 10.1099/0022-1317-83-1-223
da Silva, T. G., Lima, M. S., de Castro, A. M. M. G., Martins, M. S. N., Castiglioni, V. C., Del Fava, C., et al. (2018). Bovine vaccinia in dairy cattle and suspicion of vesicular disease on milkers in Brazil. Cienc. Rural. 48:e20170723 doi: 10.1590/0103-8478cr20170723
Damaso, C. R. A., Esposito, J. J., Condit, R. C., and Moussatché, N. (2000). An emergent poxvirus from humans and cattle in Rio de Janeiro state: Cantagalo virus may derive from brazilian smallpox vaccine. Virology 277, 439–449. doi: 10.1006/viro.2000.0603
Darriba, D., Taboada, G. L., Doallo, R., and Posada, D. (2012). JModelTest 2: more models, new heuristics and parallel computing. Nat. Methods 9:772. doi: 10.1038/nmeth.2109
de Assis, F. L., Pereira, G., Oliveira, C., Rodrigues, G. O. L., Cotta, M. M. G., Silva-Fernandes, A. T., et al. (2012). Serologic evidence of orthopoxvirus infection in buffaloes, Brazil. Emerg. Infect. Dis. 18, 698–700. doi: 10.3201/eid1804.111800
de Oliveira, J. S., Figueiredo, P. O., Costa, G. B., De Assis, F. L., Drumond, B. P., Da Fonseca, F. G., et al. (2017). Vaccinia virus natural infections in Brazil: the good, the bad, and the ugly. Viruses 9:E340. doi: 10.3390/v9110340
de Oliveira, T. M., Guedes, M. I., Rehfeld, I. S., Matos, A. C., Rivetti, A. V., Alves, P. A., et al. (2015). Detection of Vaccinia Virus in milk: evidence of a systemic and persistent infection in experimentally infected cows. Foodborne Pathog. Dis. 12, 898–903. doi: 10.1089/fpd.2015.1974
de Oliveira, T. M., Guedes, M. I. M., Rehfeld, I. S., Matos, A. C., Rivetti Júnior, A. V., da Cunha, A. F., et al. (2018). Vaccinia virus detection in dairy products made with milk from experimentally infected cows. Transbound. Emerg. Dis. 65, e40–e47. doi: 10.1111/tbed.12666
de Souza Trindade, G., da Fonseca, F. G., Marques, J. T., Nogueira, M. L., Mendes, L. C., Borges, A S., et al. (2003). Aracatuba virus: a vaccinialike virus associated with infection in humans and cattle. Emerg. Infect. Dis. 9, 155–160. doi: 10.3201/eid0902.020244
de Souza Trindade, G., Emerson, G. L., Sammons, S., Frace, M., Govil, D., Mota, B. E. F., et al. (2016). Serro 2 virus highlights the fundamental genomic and biological features of a natural Vaccinia virus infecting humans. Viruses 8:E328. doi: 10.3390/v8120328
de Souza Trindade, G., Li, Y., Olson, V. A., Emerson, G., Regnery, R. L., da Fonseca, F. G., et al. (2008). Real-time PCR assay to identify variants of Vaccinia virus: implications for the diagnosis of bovine vaccinia in Brazil. J. Virol. Methods 152, 63–71. doi: 10.1016/j.jviromet.2008.05.028
Drumond, B. P., Leite, J. A., da Fonseca, F. G., Bonjardim, C. A., Ferreira, P. C. P., and Kroon, E. G. (2008). Brazilian Vaccinia virus strains are genetically divergent and differ from the Lister vaccine strain. Microbes Infect. 10, 185–197. doi: 10.1016/j.micinf.2007.11.005
Dutra, L. A., de Freitas Almeida, G. M., Oliveira, G. P., Abrahão, J. S., Kroon, E. G., and Trindade, G. S. (2017). Molecular evidence of orthopoxvirus DNA in capybara (Hydrochoerus hydrochaeris) stool samples. Arch. Virol. 162, 439–448. doi: 10.1007/s00705-016-3121-3
Eder, I., Vollmar, P., Pfeffer, M., Naether, P., Rodloff, A. C., and Meyer, H. (2017). Two distinct clinical courses of human cowpox, Germany, 2015. Viruses 9:E375. doi: 10.3390/v9120375
Essbauer, S., and Meyer, H. (2010). Zoonotic poxviruses. Vet. Microbiol. 140, 229–236. doi: 10.1016/J.VETMIC.2009.08.026
Fonseca, F. G., Lanna, M. C. S., Campos, M. A. S., Kitajima, E. W., Peres, J. N., Golgher, R. R., et al. (1998). Morphological and molecular characterization of the poxvirus BeAn 58058. Arch. Virol. 143, 1171–1186. doi: 10.1007/s007050050365
Franco-Luiz, A. P. M., Fagundes Pereira, A., de Oliveira, C. H. S., Barbosa, J. D., Oliveira, D. B., Bonjardim, C. A., et al. (2016a). The detection of Vaccinia virus confirms the high circulation of Orthopoxvirus in buffaloes living in geographical isolation, Marajó Island, Brazilian Amazon. Comp. Immunol. Microbiol. Infect. Dis. 46, 16–19. doi: 10.1016/j.cimid.2016.03.003
Franco-Luiz, A. P. M., Oliveira, D. B., Fagundes Pereira, A., Gasparini, M. C. S., Bonjardim, C. A., Ferreira, P. C. P., et al. (2016b). Detection of vaccinia virus in dairy cattle serum samples from 2009, Uruguay. Emerg. Infect. Dis. 22, 2174–2177. doi: 10.3201/eid2212.160447
Franco-Luiz, A. P. M., Fagundes-Pereira, A., Costa, G. B., Alves, P. A., Oliveira, D. B., Bonjardim, C. A., et al. (2014). Spread of Vaccinia virus to cattle herds, Argentina, 2011. Emerg. Infect. Dis. 20, 1576–1578. doi: 10.3201/eid2009.140154
Gerber, P. F., Matos, A. C. D., Guedes, M. I., Madureira, M. C., Silva, M. X., and Lobato, Z. I. (2012). Validation of an immunoperoxidase monolayer assay for total anti-Vaccinia virus antibody titration. J. Vet. Diagn. Investig. 24, 355–358. doi: 10.1177/1040638711435231
Guarner, J., Johnson, B. J., Paddock, C. D., Shieh, W. J., Goldsmith, C. S., Reynolds, M. G., et al. (2004). Monkeypox transmission and pathogenesis in prairie dogs. Emerg. Infect. Dis. 10, 426–431. doi: 10.3201/eid1003.030878
Joklik, W. K. (1962). The purification of four strains of poxvirus. Virology 18, 9–18. doi: 10.1016/0042-6822(62)90172-1
Kik, M., and Luten, H. (2009). “Cowpoxvirus infection in Asian elephants (Elephas maximus) in a zoo,” In Proceedings of the International Conference Disease Zoology Wild Animal, (Berlin: Leibniz Institute for Zoo and Wildlife Research).
Kile, J. C., Fleischauer, A. T., Beard, B., Kuehnert, M. J., Kanwal, R. S., Pontones, P., et al. (2005). Transmission of monkeypox among persons exposed to infected prairie dogs in Indiana in 2003. Arch. Pediatr. Adolesc. Med. 159, 1022–1025. doi: 10.1001/archpedi.159.11.1022
Klein, J., Hussain, M., Ahmad, M., Afzal, M., and Alexandersen, S. (2008). Epidemiology of foot-and-mouth disease in Landhi Dairy Colony, Pakistan, the world largest Buffalo colony. Virol. J. 5:53. doi: 10.1186/1743-422X-5-53
Kroon, E. G., Abrahão, J. S., Trindade, G. S., Oliveira, G. P., Luiz, A. P. M. F., Costa, G. B., et al. (2016). Natural Vaccinia virus infection: diagnosis, isolation, and characterization. Curr. Protoc. Microbiol. 42, 14A.5.1–14A.5.43. doi: 10.1002/cpmc.13
Kroon, E. G., Mota, B. E., Abrahão, J. S., da Fonseca, F. G., and de Souza Trindade, G. (2011). Zoonotic Brazilian Vaccinia virus: from field to therapy. Antivir. Res. 92, 150–163. doi: 10.1016/j.antiviral.2011.08.018
Laguardia-Nascimento, M., Sales, E. B., Gasparini, M. R., de Souza, N. M., da Silva, J. A. G., Souza, G. G., et al. (2016). Detection of multiple viral infections in cattle and buffalo with suspected vesicular disease in Brazil. J. Vet. Diagn. Invest. 28, 377–381. doi: 10.1177/1040638716645836
Leite, J. A., Drumond, B. P., Trindade, G. S., Lobato, Z. I. P., Da Fonseca, F. G., Dos Santos, J. R., et al. (2005). Passatempo virus, a Vaccinia virus strain, Brazil. Emerg. Infect. Dis. 11, 1935–1938. doi: 10.3201/eid1112.050773
Lima, M. T., Oliveira, G. P., Assis, F. L., de Oliveira, D. B., Vaz, S. M., de Souza Trindade, G., et al. (2018). Ocular vaccinia infection in dairy worker, Brazil. Emerg. Infect. Dis. 24, 161–162. doi: 10.3201/eid2401.170430
Lobato, Z. I. P., Trindade, G. S., Frois, M. C. M., Ribeiro, E. B. T., Dias, G. R. C., Teixeira, B. M., et al. (2005). Outbreak of exantemal disease caused by Vaccinia virus in human and cattle in Zona da Mata region, Minas Gerais. Arq. Bras. Med. Vet. Zootec. 57, 423–429
Long, J. L. (2003). Introduced Mammals of the world. Their History, Distribution and Influence. Wallingford: CABI Publication. doi: 10.1086/428221
Marennikova, S. S., Maltseva, N. N., Korneeva, V. I., and Garanina, N. M. (1977). Outbreak of pox disease among carnivora (Felidae) and edentata. J. Infect. Dis. 135, 358–366. doi: 10.1093/infdis/135.3.358
Matos, A. C. D., Rehfeld, I. S., Guedes, M. I. M. C., and Lobato, Z. I. P. (2018). Bovine vaccinia: insights into the disease in cattle. Viruses 10:E120. doi: 10.3390/v10030120
Medaglia, M. L. G., Pessoa, L. C. G. D., Sales, E. R. C., Freitas, T. R. P., and Damaso, C. R. (2009). Spread of Cantagalo virus to northern Brazil. Emerg. Infect. Dis. 15, 1142–1143. doi: 10.3201/eid1507.081702
Megid, J., Appolinário, C. M., Langoni, H., Pituco, E. M., and Okuda, L. H. (2008). Short report: Vaccinia virus in humans and cattle in Southwest region of São Paulo State, Brazil. Am. J. Trop. Med. Hyg. 79, 647–651. doi: 10.4269/ajtmh.2008.79.647
Miranda, J. B., Borges, I. A., Campos, S. P. S., Vieira, F. N., De Ázara, T. M. F., Marques, F. A., et al. (2017). Serologic and molecular evidence of Vaccinia virus circulation among small mammals from different biomes, Brazil. Emerg. Infect. Dis. 23, 931–938. doi: 10.3201/eid2306.161643
Newman, F. K., Frey, S. E., Blevins, T. P., Mandava, M., Bonifacio, A., Yan, L., et al. (2003). Improved assay to detect neutralizing antibody following vaccination with diluted or undiluted vaccinia (Dryvax) vaccine. J. Clin. Microbiol. 41, 3154–3157. doi: 10.1128/JCM.41.7.3154-3157.2003
Oliveira, D. B., Assis, F. L., Ferreira, P. C. P., Bonjardim, C. A., de Souza Trindade, G., Kroon, E. G., et al. (2013). Group 1 Vaccinia virus zoonotic outbreak in maranhao state, Brazil. Am. J. Trop. Med. Hyg. 89, 1142–1145. doi: 10.4269/ajtmh.13-0369
Oliveira, G., Assis, F., Almeida, G., Albarnaz, J., Lima, M., Andrade, A. C., et al. (2015). From lesions to viral clones: biological and molecular diversity amongst autochthonous brazilian Vaccinia virus. Viruses 7, 1218–1237. doi: 10.3390/v7031218
Oliveira, G. P., Rodrigues, R. A. L., Lima, M. T., Drumond, B. P., and Abrahão, J. S. (2017). Poxvirus host range genes and virus–host spectrum: a critical review. Viruses 9:E331. doi: 10.3390/v9110331
Peres, M. G., Bacchiega, T. S., Appolinário, C. M., Vicente, A. F., Allendorf, S. D., Antunes, J. M. A. P., et al. (2013). Serological study of Vaccinia virus reservoirs in areas with and without official reports of outbreaks in cattle and humans in São Paulo, Brazil. Arch. Virol. 153, 2433–2441. doi: 10.1007/s00705-013-1740-5
Peres, M. G., Barros, C. B., Appolinário, C. M., Antunes, J. M. A. P., Mioni, M. S. R., Bacchiega, T. S., et al. (2016). Dogs and opossums positive for Vaccinia virus during outbreak affecting cattle and humans, São Paulo State, Brazil. Emerg. Infect. Dis. 22, 271-273. doi: 10.3201/eid2202.140747
Pilaski, J., and Rösen-Wolff, A. (1988). “Poxvirus Infection in Zoo-kept mammals”, in Virus Diseases in Laboratory and Captive Animals, ed. Darai. G, (Berlin: Springer Science+Business Media). doi: 10.1007/978-1-4613-2091-3_5
Rehfeld, I. S., Guedes, M. I. M. C., Fraiha, A. L. S., Costa, A. G., Matos, A. C. D., Fiúza, A. T. L., et al. (2015). Vaccinia virus transmission through experimentally contaminated milk using a murine model. PLoS One 10:e0127350. doi: 10.1371/journal.pone.0127350
Reynolds, M. G., Carroll, D. S., and Karem, K. L. (2012). Factors affecting the likelihood of monkeypox’s emergence and spread in the post-smallpox era. Curr. Opin. Virol. 2, 335–343. doi: 10.1016/j.coviro.2012.02.004
Ropp, S. L., Jin, Q., Knight, J. C., Massung, R. F., and Esposito, J. J. (1995). PCR strategy for identification and differentiation of small pox and other orthopoxviruses. J. Clin. Microbiol. 33, 2069–2076
Sambrook, J., and Russell, D. W. (2001). Molecular Cloning: A Laboratory Manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press. doi: 10.1016/0092-8674(90)90210-6
Sheikh, P. A., Merry, F. D., and McGrath, D. G. (2006). Water buffalo and cattle ranching in the Lower Amazon Basin: comparisons and conflicts. Agric. Syst. 87, 313–330. doi: 10.1016/j.agsy.2005.02.003
Singh, R. K., Balamurugan, V., Bhanuprakash, V., Venkatesan, G., and Hosamani, M. (2012). Emergence and reemergence of vaccinia-like viruses: global scenario and perspectives. Indian J. Virol. 23, 1–11. doi: 10.1007/s13337-012-0068-1
Singh, R. K., Hosamani, M., Balamurugan, V., Bhanuprakash, V., Rasool, T. J., and Yadav, M. P. (2007). Buffalopox: an emerging and re-emerging zoonosis. Anim. Health Res. Rev. 8, 105–114. doi: 10.1017/S1466252307001259
Stahel, A. B. J., Baggenstos, R., Engels, M., Friess, M., and Ackermann, M. (2013). Two different Macaviruses, ovine herpesvirus-2 and caprine herpesvirus-2, behave differently in water buffaloes than in cattle or in their respective reservoir species. PLoS One 8:e83695. doi: 10.1371/journal.pone.0083695
Trindade, G. S., Lobato, Z. I., Drumond, B. P., Leite, J. A., Trigueiro, R. C., Guedes, M. I., et al. (2006). Short report: isolation of two Vaccinia virus strains from a single bovine vaccinia outbreak in rural area from Brazil: implications on the emergence of zoonotic orthopoxviruses. Am. J. Trop. Med. Hyg. 75, 486–490. doi: 10.4269/ajtmh.2006.75.486
Usme-Ciro, J. A., Paredes, A., Walteros, D. M., Tolosa-Pérez, E. N., Laiton-Donato, K., del Carmen Pinzón, M., et al. (2017). Detection and molecular characterization of zoonotic poxviruses circulating in the amazon region of Colombia, 2014. Emerg. Infect. Dis. 23, 649–653. doi: 10.3201/eid2304.161041
Wilson, M. E., Hughes, J. M., McCollum, A. M., and Damon, I. K. (2014). Human monkeypox. Clin. Infect. Dis. 58, 260–267. doi: 10.1093/cid/cit703
Yadav, S., Hosamani, M., Balamurugan, V., Bhanuprakash, V., and Singh, R. K. (2010). Partial genetic characterization of viruses isolated from pox-like infection in cattle and buffaloes: evidence of buffalo pox virus circulation in Indian cows. Arch. Virol. 155, 255–261. doi: 10.1007/s00705-009-0562-y
Keywords: vaccinia virus, buffalopox, bovine vaccinia, bubaline, buffalo diseases
Citation: Lima MT, Oliveira GP, Afonso JAB, Souto RJC, de Mendonça CL, Dantas AFM, Abrahao JS and Kroon EG (2019) An Update on the Known Host Range of the Brazilian Vaccinia Virus: An Outbreak in Buffalo Calves. Front. Microbiol. 9:3327. doi: 10.3389/fmicb.2018.03327
Received: 28 September 2018; Accepted: 21 December 2018;
Published: 22 January 2019.
Edited by:
Juliano Bordignon, Fundação Oswaldo Cruz (Fiocruz), BrazilReviewed by:
Brian J. Ferguson, University of Cambridge, United KingdomCopyright © 2019 Lima, Oliveira, Afonso, Souto, de Mendonça, Dantas, Abrahao and Kroon. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Erna Geessien Kroon, ZXJuYWdrcm9vbkBnbWFpbC5jb20=; a3Jvb25lQGljYi51Zm1nLmJy
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.