
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 18 December 2018
Sec. Infectious Agents and Disease
Volume 9 - 2018 | https://doi.org/10.3389/fmicb.2018.03011
 Isabella Dias da Silveira1Martha Thieme Petersen1Gabriel Sylvestre1Gabriela Azambuja Garcia1
Isabella Dias da Silveira1Martha Thieme Petersen1Gabriel Sylvestre1Gabriela Azambuja Garcia1 Mariana Rocha David1
Mariana Rocha David1 Márcio Galvão Pavan1,2
Márcio Galvão Pavan1,2 Rafael Maciel-de-Freitas1,2*
Rafael Maciel-de-Freitas1,2*A Zika virus (ZIKV) pandemic started soon after the first autochthonous cases in Latin America. Although Aedes aegypti is pointed as the primary vector in Latin America, little is known about the fitness cost due to ZIKV infection. We investigated the effects of ZIKV infection on the life-history traits of Ae. aegypti females collected in three districts of Rio de Janeiro, Brazil (Barra, Deodoro, and Porto), equidistant ~25 km each other. Aedes aegypti mosquitoes were classified into infected (a single oral challenge with ZIKV) and superinfected (two ZIKV-infected blood meals spaced by 7 days each other). ZIKV infection reduced Ae. aegypti survival in two of the three populations tested, and superinfection produced a sharper increase in mortality in one of those populations. We hypothesized higher mortality with the presence of more ZIKV copies in Ae. aegypti females from Porto. The number of eggs laid per clutch was statistically similar between vector populations and infected and uninfected mosquitoes. Infection by ZIKV not affected female oviposition success. ZIKV infection impacted Ae. aegypti vectorial capacity by reducing its lifespan, although female fecundity remained unaltered. The outcome of these findings to disease transmission intensity still needs further evaluation.
Aedes aegypti (Diptera: Culicidae) is known as the primary vector of dengue virus (DENV) and chikungunya virus (CHIKV) and was also found recently to be naturally infected with Zika virus (ZIKV), which reinforces the paramount role of this species in arthropod-borne virus transmission (Ferreira-de-Brito et al., 2016;Patterson et al., 2016). The emergence of an arbovirus followed by an outbreak depends upon several factors such as high density of primary vectors, efficient viral replication, and infectivity in vertebrate and invertebrate hosts and the presence of immunologically naïve hosts (Liu-Helmersson et al., 2014). For instance, recent studies concerning virus infectivity demonstrated that a single amino acid substitution in the NS1 gene improved the efficiency of ZIKV transmission from humans to Ae. aegypti and thus increasing its prevalence in mosquitoes (Liu et al., 2017; Rossi et al., 2018).
One of the most critical aspects to arbovirus epidemiology is related to the interaction between the genetic backgrounds of insect vectors and virus strains, which may influence vectorial capacity and transmission rates (Weaver and Reisen, 2010). For instance, Brazilian mosquitoes challenged with a ZIKV strain isolated from a patient in New Caledonia exhibited low vector competence (Chouin-Carneiro et al., 2016) but showed high susceptibility when infected with local ZIKV strains (Dutra et al., 2016; Fernandes et al., 2016). Therefore, it is of utmost relevance to investigate the interaction between mosquito vectors and locally isolated virus strains to provide a more realistic dataset.
The biology and behavior of infected mosquitoes have been little explored, although it is an important aspect of vector-borne diseases. Transmission models and the vectorial capacity formula often overlook that arbovirus infections may alter the behavior and/or pose a fitness cost in life-history traits of mosquitoes and thus affect transmission [cf. Kramer and Ciota (2015) for a comprehensive discussion about this topic]. There is a series of experimental data regarding Plasmodium and Anopheles mosquitoes that could be interpreted as host behavior manipulation by the parasite to enhance its transmission (Rossignol et al., 1986; Koella et al., 1998; Schwartz and Koella, 2001; Robinson et al., 2018). However, very limited information is available regarding the biology of Aedes mosquitoes infected with arboviruses. A few reports have demonstrated that DENV and ZIKV are capable to alter the expression of genes potentially involved in host-seeking behavior, especially odorant-binding protein transcripts (Sim et al., 2012; Etebari et al., 2017). The first two evidences of arbovirus impact on Ae. aegypti biting rate came from the 1990s, but produced conflicting results. First, there was no evidence of alteration in the biting behavior of Ae. aegypti after intrathoracic inoculation of DENV-2 (Putnam and Scott, 1995). Later, it was observed that dengue-infected mosquitoes required more time to feed on blood than uninfected ones (Platt et al., 1997). More recently, some papers reported a negative effect of DENV-2 on Ae. aegypti life-history traits, such as reduced lifespan and fecundity, and also an increase in the time to complete a blood meal (Maciel-de-Freitas et al., 2011, 2013; Sylvestre et al., 2013).
Even though negative effects were revealed for dengue virus, little is known about the potential cost of ZIKV infection in vector biology. The recent ZIKV emergence and its explosive outbreak across the Pacific and Latin America caused thousands of cases in cities such as Rio de Janeiro. Therefore, it sounds reasonable to speculate that, occasionally, susceptible mosquitoes are likely to blood feed more than once in ZIKV-infected hosts during the course of an outbreak. Beyond that, opportunities for arboviral exposure in mosquitoes already infected are considerable, since the viral infection persists throughout the insect life. This phenomenon is known as superinfection (Salas-Benito and De Nova-Ocampo, 2015). Considering adult Ae. aegypti females have a lifespan of 15–30 days measured by mark, release, recapture studies (Maciel-De-Freitas et al., 2007; Maciel-de-Freitas et al., 2008; David et al., 2009; Maciel-de-Freitas and Lourenço-de-Oliveira, 2009), the likelihood of ingesting blood from two infected hosts during its lifespan and its effects on mosquito-virus interactions has been receiving little attention. The present study aims to investigate the susceptibility of three field Ae. aegypti populations to ZIKV, and the infection effects on its life-history traits after mosquitoes were exposed to a single or two ZIKV-infected blood meals.
Eggs were collected through 80 ovitraps (Fay and Eliason, 1966) placed roughly every 25 m each other in each of three different regions in Rio de Janeiro—Barra (22°58′77″ S, 43°23′41″ W), Deodoro (22°51′01″ S, 43°23′52″ W), and Porto (22°53′43″ S, 43°11′03″ W), distant 15–25 km from each other. Ovitraps were installed over an extensive geographic area to assure we sampled the local Ae. aegypti genetic variability. Ae. aegypti females are considered a limited flyer with mean distance traveled inferior to 200 m (Maciel-De-Freitas et al., 2007), and thus, it is highly unlike that a single mosquito would cross all field sites analyzed here. A minimum of 500 eggs were collected per site and were hatched in the insectary. Adults were maintained at the insectary under a relative humidity of 80 ± 5% and a temperature of 25 ± 3°C, with ad libitum access to a 10% sucrose solution. Experiments were performed with F1 generation mosquitoes. A second collection of eggs was done 5–6 months later using the same protocol, and immature mosquitoes were reared to adults to perform a second experimental infection to evaluate the effect of ZIV on Ae. aegypti life-history traits.
Females were orally challenged with a ZIKV strain isolated from the urine of a patient in Rio de Janeiro. The ZIKV strain belongs to the Asian genotype Rio-U1 (GenBank accession number KU926309) and showed to have high infectivity to Rio Ae. aegypti mosquito populations (Fernandes et al., 2016). Viral titers in supernatants were previously determined by serial dilutions in Vero cells, expressed in plaque-forming unit per milliliters (PFU/ml). All the assays were performed with samples containing 3.55 × 106 PFU/ml. Viral stocks were maintained at −80°C until its use.
Two rounds of experimental infection assays were conducted, using the same protocol for mosquito superinfection. Thirty-six hours before infection, 6–7 days old inseminated Ae. aegypti females from each of the three populations (Barra, Deodoro, and Porto) were separated in 18 cylindrical plastic cages (70 mosquitoes/cage) for blood feeding. Sugar supply was removed 36-h before mosquitoes were challenged with the infective blood meal to increase female’s avidity. The oral infection procedures were performed through a membrane feeding system (Hemotek, Great Harwood, UK), adapted with a pig-gut covering, which gives access to the defibrinated rabbit blood. The infective blood meals consisted of 1 ml of supernatant of infected cell culture, 2 ml of washed rabbit erythrocytes, and 0.5 mM of ATP as phagostimulant. The same procedure and membrane feeding apparatus were used to feed control mosquitoes, but they received a noninfectious blood meal, with 1 ml of cell culture medium replacing the viral supernatant.
The explosive emergence of ZIKV across the Pacific and Latin America caused thousands of cases in cities such as Rio de Janeiro. Therefore, it is feasible that mosquitoes may feed more than once on ZIKV-infected hosts during their lifespan. Herein, we simulated Ae. aegypti superinfection with ZIKV by offering a second infective blood meal to a fraction of the infected mosquitoes. Seven days after the first oral infection (minimum extrinsic incubation period expected for ZIKV) (Roundy et al., 2017), mosquitoes from Deodoro and Porto populations were challenged with a new infective blood meal, using the same viral strain at the same titration (herein called “superinfection”). The other groups (control and infected) received a blood meal without the virus. Aedes aegypti individuals from Barra were not included in this procedure, since there were not sufficient egg stocks.
Those females that were visually completely engorged after oral infection assays were individualized in cylindrical plastic tubes (a height of 6.5 cm and a diameter of 3 cm) containing moistened cotton overlaid with filter paper as oviposition substrate on the bottom. Tubes were covered on the top with mosquito netting. Once a week, we offered uninfected anesthetized mice to mosquitoes to all groups as blood source. Filter papers were checked for eggs, and those were counted every third day after a blood meal, when a new filter paper was added. Mosquito survival was checked daily at 09:00 h. Each dead mosquito was removed from the plastic tube, and wing length was measured as the distance from the axillary incision to the apical margin, excluding the fringe (Harbach and Knight, 1980).
Infected mosquitoes were kept in an incubator under an artificial photoperiodic regime of 12 h of light and 12 h of dark, with 10% sucrose solution. Temperature was maintained at 28°C, with a relative humidity of 70–80% throughout the course of experiment.
In total, 3–10 mosquitoes from each population were sampled at 7 and 14 days postinfection (dpi) and on 7 days post superinfection (dpsi) to check for infection. For that, bodies and heads of 45 sampled individuals were separated before the RNA extraction procedure to assess ZIKV infection and dissemination, respectively. The 44 selected mosquitoes belonged to all three populations: Barra (n = 3), Porto (n = 22), and Deodoro (n = 20), from infected (n = 24) and superinfected (n = 20) groups.
Total RNA was extracted from mosquitoes with the QIAamp Viral RNA Mini Kit (Qiagen, Hilden, Germany). Viral RNA detection and quantification were performed individually through RT-qPCR with SuperScriptTM III PlatinumTM One-Step qRT-PCR Kit (Invitrogen, Carlsbad, CA, USA) in QuantStudio 6 Flex Real-Time PCR System (Applied Biosystems, Foster City, CA, USA). Each reaction was made with 600 nM forward primer (5′-CTTGGAGTGCTTGTGATT-3′, genome position 3451–3468), 600 nM reverse primer (5′-CTCCTCCAGTGTTCATTT-3′, genome position 3637–3620), and 800 nM probe (5′FAM-AGAAGAGAATGACCACAAAGATCA-3′TAMRA, genome position 3494–3517), previously published (Ferreira-de-Brito et al., 2016). Cycling conditions were as follows: 95°C for 2 minutes, followed by 40 amplification cycles of 95°C for 15 s, 58°C for 5 s, and 60°C for 30 s. Virus copy numbers were calculated by interpolation onto an internal standard curve made up of a seven-point dilution series (102–108 copies/ml) of in vitro transcribed ZIKV RNA (Bonaldo et al., 2016).
Mosquitoes from the two experimental infection assays were analyzed as belonging to three treatments: controls, which did not feed on ZIKV infectious blood; infected, which received one single infected blood meal on 6–7 days old; and superinfected, which received an additional infected blood meal on 13–14 days old.
The association among survival and ZIKV infection, population and wing length was analyzed using Cox proportional hazard regression models to obtain hazard ratios and 95% confidence intervals. First, we computed separate univariate Cox regression analyses with population (Barra, Deodoro, and Porto), treatment (control, infected, and superinfected), and wing length as covariates. Only statistically significant variables according to Wald statistic values were included in the multivariate Cox model. Kaplan-Meier (KM) survival curves were created to each mosquito population according to the different treatments (control, infected, or superinfected). We performed log-rank tests to address the global effects of treatment in each mosquito population. If significant, we confronted infected and superinfected against controls. Significance level was adjusted for multiple comparisons with the Bonferroni criteria. Those analyses were performed in the R environment (R Development Core Team, 2011).
Fecundity was analyzed by considering the first four clutches of eggs laid, as only a small number of females laid eggs when they were more than 4 weeks old, precluding adequate numbers for analysis. We considered two aspects of fecundity: 1) oviposition success: the likelihood of laying at least one egg (at a given clutch) with a logistic analysis that included treatment, population, wing length and clutch number (i.e., age); and 2) fecundity: the egg number of the successful mosquitoes with a repeated analysis. We included clutch number as the repeat and estimated the effects of treatment, wing length and population. These analyses were carried out with the statistical software JMP v. 13.0 (SAS Institute Inc., 2007).
The abundance of RNA ZIKV was not normally distributed (Shapiro-Wilk W = 0.4560, p < 0.001) and, therefore, the three treatments were compared through Wilcoxon-Mann-Whitney tests in the R environment (R Development Core Team, 2011). Significance level was adjusted for multiple comparisons with the false discovery rate (FDR) method (Benjamini and Yekutieli, 2001). The effect size of comparisons (r) was performed with Cohen’s d calculations (Cohen, 1988).
A total of 767 Ae. aegypti F1 females from three districts of Rio de Janeiro (Barra, Deodoro, and Porto), Brazil, were used in the experiments. From that, 44 individuals were used for vector competence tests, 399 females were orally infected with a local ZIKV strain (345 were infected once and 54 were superinfected, i.e., received a second infective blood meal at 7 days postinfection (dpi) with the same ZIKV strain). Of the 723 mosquitoes monitored daily during fitness cost assays, 90 belonged to Barra (60 infected and 30 controls), 308 to Deodoro (140 infected, 147 controls, and 21 superinfected), and 325 (145 infected, 147 controls, and 33 superinfected) to Porto field populations.
A total of 45 mosquitoes confirmed the susceptibility of the three populations to the locally isolated ZIKV strain Rio-U1. From that, 44 (97.7%) mosquitoes presented ZIKV at their bodies, and 41 (91.1%) had positive heads. As expected, the number of viral copies increased in bodies and heads over time for Porto and Deodoro. Regarding ZIKV quantification in bodies, although marginal and not significant, we observed a difference between Deodoro and Porto infected mosquitoes occurred at 14 dpi, with the former population presenting more ZIKV genome copies (W = 23, p = 0.032; Figure 1).
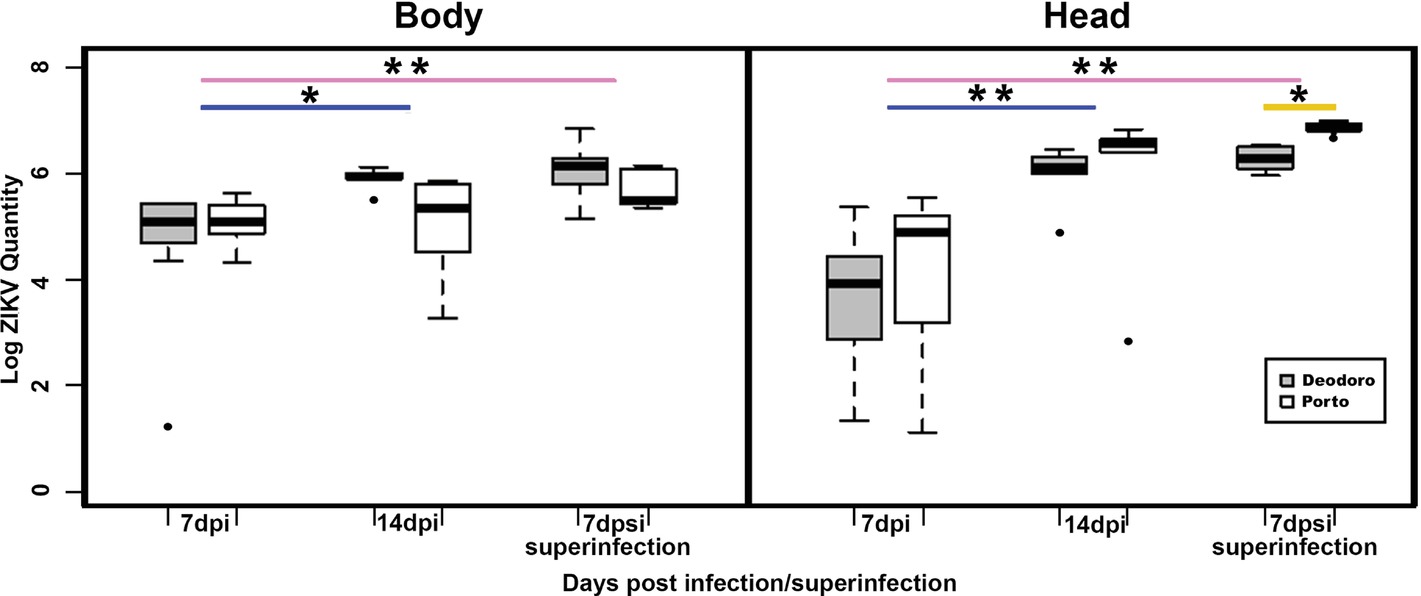
Figure 1. The viral load in the body and head of Aedes aegypti mosquitoes infected and superinfected with ZIKV from Deodoro and Porto field populations. Dpi: days postinfection; dpsi: days post superinfection. *p < 0.05; ** p < 0.001. Due to low-sample sizes, Barra population data were not included in the analysis.
However, no significant difference in the number of viral copies was detected when the heads of Porto- and Deodoro-infected mosquitoes were compared at 7 and 14 dpi (Figure 1). Superinfected mosquitoes from Deodoro and Porto had statistically similar viral copies in their bodies. Interestingly, superinfected Ae. aegypti from Porto showed significantly more ZIKV RNA loads at their heads at 7 days post superinfection (dpsi) than Deodoro population, reaching 107 copies (W = 0, p = 0.007; Figure 1).
Wing length had no detectable influence in mosquito survival (W = 0.15, df = 1, p = 0.7), while population and treatment exhibited significant effects (W = 28.3, df = 2, p < 0.01; W = 13.8, df = 2, p < 0.01, respectively) and thus were included in the multivariate analysis. The multivariate Cox model confirmed that populations exhibited different survival curves under laboratory conditions, despite the ZIKV infection (Table 1). In total, only eight individuals survived longer than 60 dpi, five from Deodoro and three from Porto. The median survival was 16 (95% confidence interval, CI: 15–19), 23 (95% CI: 22–25), and 20 (95% CI: 19–23) days to the Barra, Deodoro, and Porto populations, respectively. The Cox model also revealed that the treatment (i.e., exposure to ZIKV) produced a decrease in survival, regardless of population. These results suggest that ZIKV infection negatively affects the longevity of Ae. aegypti, with a stronger effect (i.e., a higher hazard ratio) in superinfected mosquitoes (Table 1).
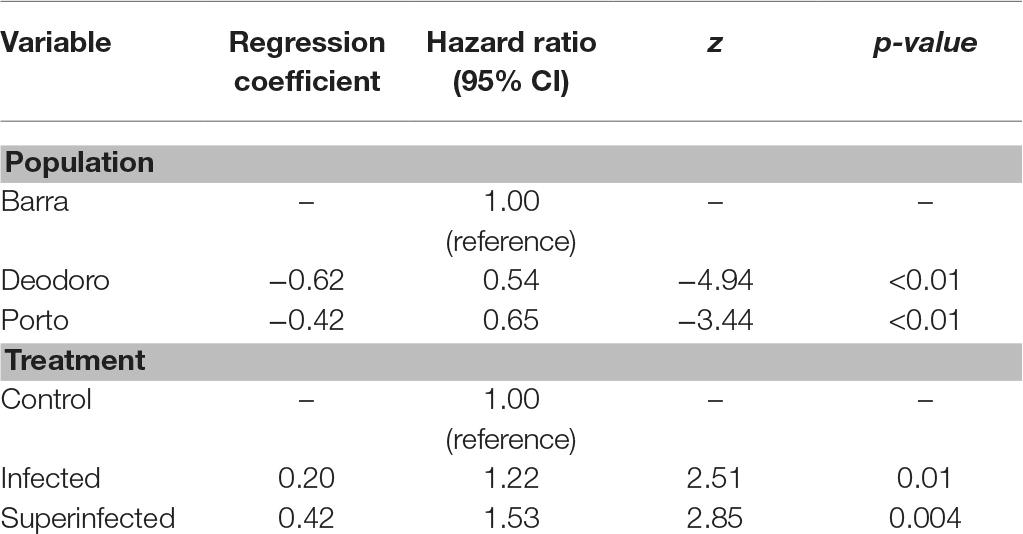
Table 1. Associations between mosquito survival and population (Barra, Deodoro, and Porto) and also survival and treatment (control, infected, and superinfected).
The ZIKV infection effect on mosquito survival was further investigated with survival curves (Figures 2A–C) and paired comparisons between infected and control groups. The treatment did not alter the survival of Barra mosquitoes ( χ2 = 3.5, df = 1, p = 0.062), but produced significant effects on Ae. aegypti populations from Deodoro ( χ2 = 8, df = 2, p = 0.01) and Porto ( χ2 = 20.3, df = 2, p < 0.01). In Deodoro, ZIKV infection (but not superinfection) altered mosquito mortality ( χ2 = 7.6, df = 1, p = 0.01). However, a different trend was observed for Porto mosquitoes—only the superinfection promoted a reduction in survivorship when compared to the control group ( χ2 = 22.1, df = 1, p < 0.001).
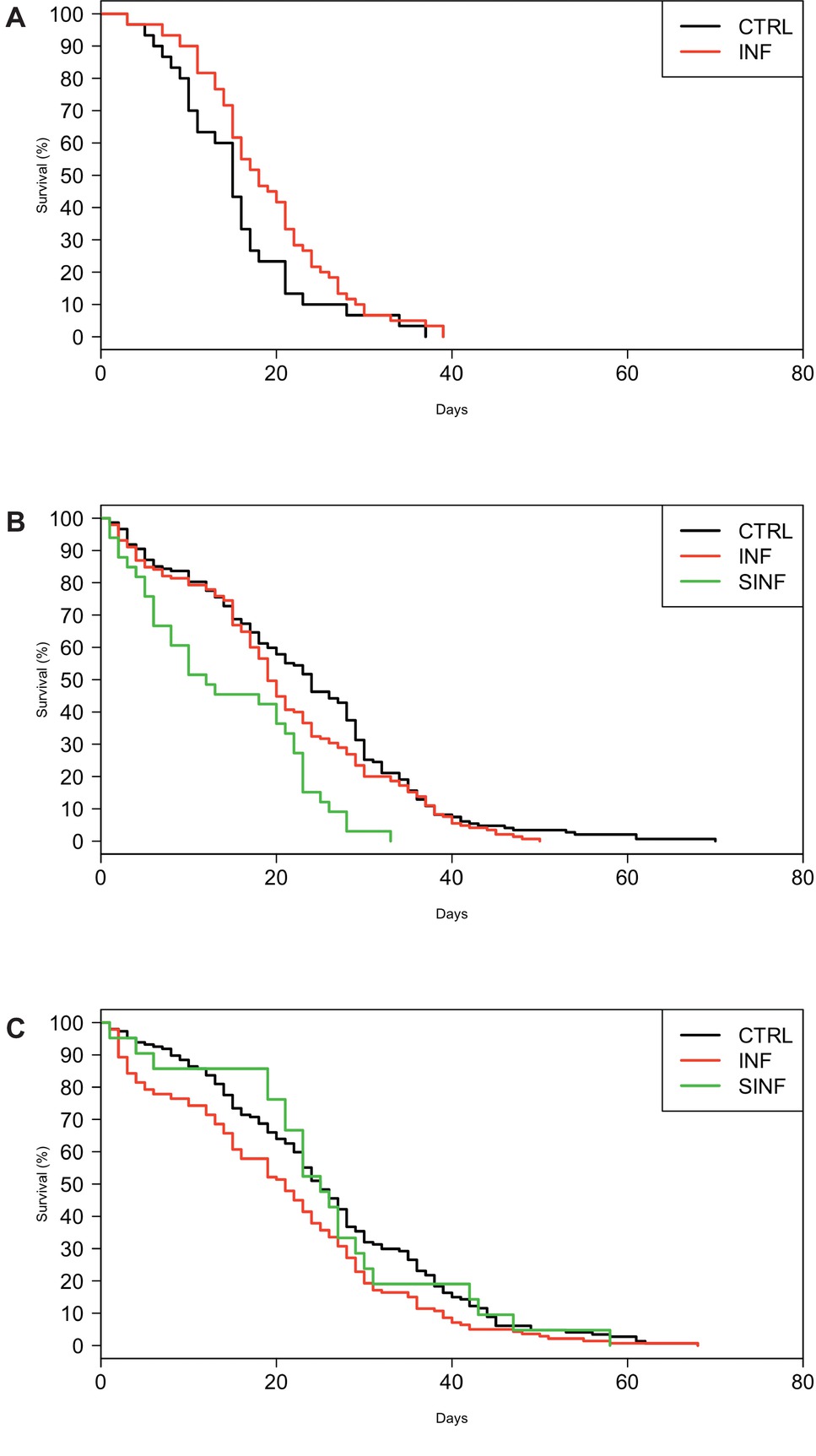
Figure 2. Aedes aegypti survival curves according to treatment (control, infected, and superinfected). Data for (A) Barra, (B) Porto, and (C) Deodoro populations. CTR: control (noninfected mosquitoes); INF: infected mosquitoes; SINF: superinfected mosquitoes.
The likelihood of Ae. aegypti females laying at least one egg per gonotrophic cycle was strongly affected by mosquito age, with older insects having lower oviposition success (Table 2). Deodoro females exhibited a significant higher oviposition success (an average of 50% considering the four first batches), while the mosquitoes from Barra and Porto exhibited lower averages (27% for both populations). On the other hand, the treatment (control, infection, or superinfection with ZIKV) not affected the oviposition success of Ae. aegypti (Table 2).
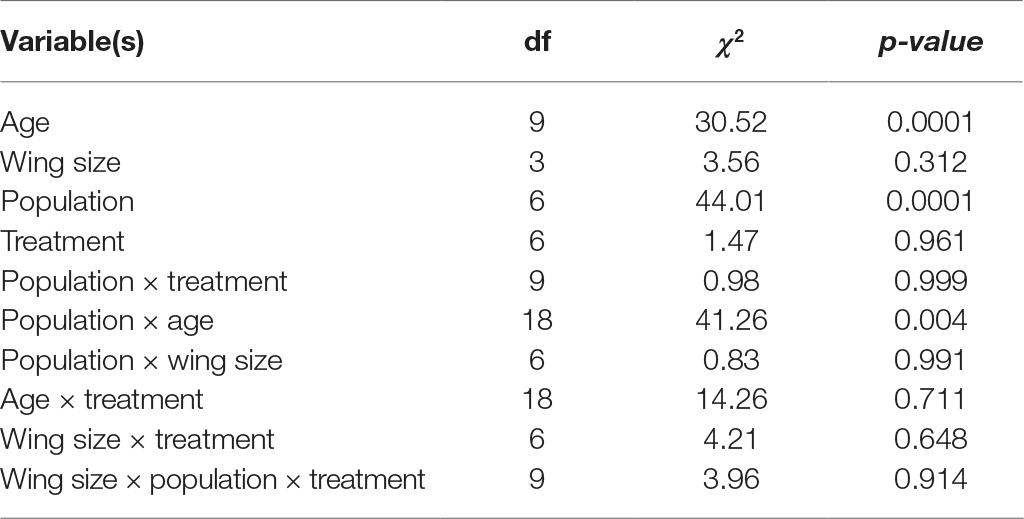
Table 2. Logistic regression analysis of the mosquito population, treatment, wing size, and age when they lay eggs on the success of oviposition of Aedes aegypti females.
Fecundity analysis considered the egg number of Ae. aegypti females that laid at least one egg. Interestingly, the number of eggs laid by females presented a slight increase over time (i.e., with age), from 49.2 in the first clutch to 55.1 in the fourth, in average (Table 3). Mosquito population, treatment and wing size had no effect on the number of eggs laid by Ae. aegypti females (Table 3).
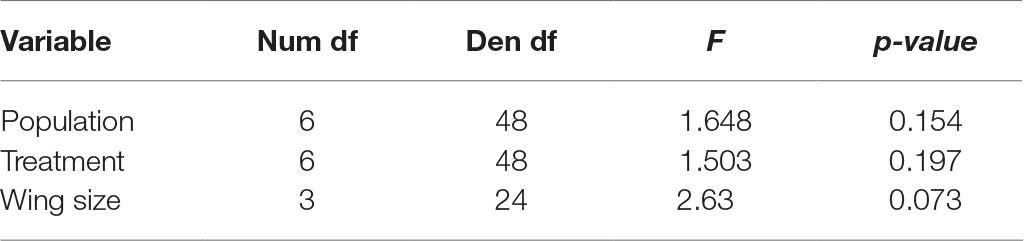
Table 3. Repeated measure analysis (with clutch taken as the repeat) of the number of eggs laid by Aedes aegypti females.
The vectorial capacity (VC) is defined as the entomological component of the basic reproduction rate (R0) of vector-borne diseases such as malaria, dengue, and Zika viruses (Service, 1993). Therefore, appropriate estimates of VC parameters may provide valuable insights into disease epidemiology and also yield the establishment of more efficient vector control activities to mitigate transmission (Brady et al., 2016). Previous studies have shown that a given pathogen can modify life-history traits of vectors and, therefore, directly influence the VC and ultimately the R0 (Scott and Lorenz, 1998; Moncayo et al., 2000; Martin et al., 2010; Vezilier et al., 2012). This article describes the effects of a locally isolated ZIKV strain (Asian genotype Rio-U1) on the biology of three Ae. aegypti field populations from Rio de Janeiro. Our results showed that a single (infection) or two ZIKV infected blood meals (superinfection) posed significant effects on the longevity, but not on the fecundity of female mosquitoes. Moreover, mosquito populations differed in their response to virus infection regarding survival rates and viral loads in the body and head.
Shortened lifespan due to pathogen infection was observed in different insect models, such as Culiseta melanura orally challenged with Eastern equine encephalomyelitis virus, Aedes albopictus infected with chikungunya, and Anopheles stephensi mosquitoes infected with Plasmodium berghei (Scott and Lorenz, 1998; Dawes et al., 2009; Martin et al., 2010). Negative effects on Ae. aegypti longevity were also detected after challenging females with a DENV-2 strain that has never circulated in the region where mosquitoes were collected (Maciel-de-Freitas et al., 2011; Sylvestre et al., 2013). Interestingly, our study pointed that one of the three tested populations (Barra) not had any reduction in lifespan due to ZIKV infection, while the other two populations exhibited an increase in mortality when infected (Deodoro) or superinfected (Porto). However, considering that Barra had only a few mosquitoes, conclusions about this population must be taken carefully. Although we did not accessed all the variables that affect the VC, mortality rate is one of the most important entomological parameters for its estimation (Luz et al., 2003) and thus is reasonable to assume that changes on mosquito lifespan may affect disease transmission under natural settings.
Despite distant each other less than 25 km, the response of mosquito populations to the laboratory environment and to ZIKV infection differed substantially. Deodoro mosquitoes had a higher significant lifespan than Porto and Barra populations, while Porto had a greater survival than Barra, despite the infection status. The treatment also produced different outcomes in Deodoro and Porto populations. The mortality of infected Deodoro mosquitoes decreases in comparison with the control group, while only the superinfected females from Porto had a significant decline in survival. We hypothesized that the higher mortality in superinfected Ae. aegypti from Porto is related to a greater viral load in the heads when contrasted to Deodoro mosquitoes. Virulence is tightly coupled to parasite load in An. stephensi and to higher RNA copy numbers in Drosophilidae, which means that the extent of the harm to a host might be partially explained by pathogen accumulation (Dawes et al., 2009; Longdon et al., 2015).
Vector competence seems to be influenced by specific interactions between mosquitoes and arbovirus genotypes. By challenging three isofemale families of field-derived Ae. aegypti from Thailand with three contemporaneous low-passage DENV-1 isolates, Lambrechts et al. (2009) evinced vector competence is likely governed to a large extent by virus and mosquito genetic interactions in natural populations. Moreover, the susceptibility of Ae. aegypti to pathogens is a highly dynamic feature, dependent on both genetic and environmental factors. Thus, it is expected that both vector competence and fitness cost of an infection also vary among geographically close field mosquito populations (Gubler et al., 1979; Failloux et al., 2002; Tabachnick, 2013; Severson and Behura, 2016). Nuclear markers of Aedes aegypti populations collected in Rio de Janeiro, including areas overlapping Deodoro, Barra, and Porto, revealed multiple introductions and extensive gene flow among populations. However, a strong spatial structuring was found considering mitochondrial markers and 25 genes related to mosquito immune response and insecticide resistance (Rašić et al., 2015). Therefore, these data may explain why the populations studied here exhibited different outcomes in longevity and superinfected with ZIKV.
Regardless of the extensive studies about the immune pathways activated in mosquitoes during a single flavivirus exposure, it remains unclear whether the immune system activation through a primary virus infection has any effect on a secondary infection (Xi et al., 2008; Souza-Neto et al., 2009; Sim and Dimopoulos, 2010). In the case of the infection with phylogenetic closely related viruses (at least from the same genus) in a single specimen, a superinfection exclusion is expected (i.e., a primary virus infection inhibits a secondary infection), since common host immune factors might be formerly activated by the first infection (Bolling et al., 2012). Mosquito cells previously infected with DENV and then exposed to the four DENV serotypes displayed a reduction in the viral titer (Igarashi, 1979; Kuno, 1982). A similar outcome was observed in Culex quinquefasciatus mosquitoes with a persistent infection by a Culex flavivirus and subsequently exposed to West Nile virus (WNV) (Bolling et al., 2012). However, Kuwata et al. (2015) found no evidence of superinfection exclusion of Japanese Encephalitis virus or DENV in cells previously infected with a Culex flavivirus. In the present study, no evidence of superinfection exclusion was identified, since viral titers did not decrease at 7 dpsi. Indeed, mosquitoes from Porto had a sharp decline on their survival when superinfected.
One limitation of this study is the impossibility of analyzing the effect of arbovirus genotypes in the susceptibility and life-history traits of the mosquitoes, since we infected them with a single ZIKV local strain. Genomic analyses of ZIKV epidemic strains of 2016 revealed the coexistence of at least seven phylogenetically diversified virus clusters circulating in Brazil (Shi et al., 2016; Wang et al., 2017). Although there is no information available about the diversity of ZIKV in small geographic scales such as those studied here, high levels of genetic variability of the flavivirus WNV among local strains were observed in near counties of the New York State infecting Culex mosquitoes (Ehrbar et al., 2017). In line with the idea of vector competence being population-specific, the possible genetic variability of ZIKV strains could contribute to the prevalence and transmission on local levels.
Several papers reported the impact of pathogen infection on host fecundity, which associated with a reduction in survival rate, would negatively impact VC by reducing the offspring size (Kramer and Ciota, 2015). For instance, An. stephensi and Anopheles gambiae seem to produce smaller egg batches after infected with Plasmodium yoelii nigeriensis (Hogg and Hurd, 1995; Jahan and Hurd, 1998). WNV-infected Culex tarsalis presented smaller egg rafts, mainly in the first oviposition cycle (Styer et al., 2007). Aedes aegypti females exposed to DENV-2 exhibited an impairment in fecundity, which varied over mosquito lifespan. Overall, DENV-2 infection seems to interfere with mosquitoes’ fecundity by both reducing egg-laying success and batch sizes (Maciel-de-Freitas et al., 2011; Sylvestre et al., 2013). Bearing in mind that ZIKV reaches mosquito ovaries on the second day postinfection (Li et al., 2017), we sought that the virus would exert a resembling impact over fecundity traits on the first clutch onward. Surprisingly, ZIKV infection did not influence directly on the oviposition success and fecundity, although increased mortality early after infection may result in lower reproduction rates.
Aedes aegypti vector competence seems to be genetically determined, which means that coadaptation between mosquitoes and viruses in a local setting may provide a more efficient transmission by sympatric vector genotypes with lower fitness cost (Lambrechts et al., 2009). Overall, our findings suggest that ZIKV infection causes a reduction in Ae. aegypti survival but did not alter fecundity, different from what has been observed for DENV and other arboviruses. It is worth mentioning, however, that mosquito populations varied in their response to ZIKV infection considering survival and viral loads, highlighting that fitness outcomes may be governed by the interaction between host and parasite genotypes. For example, superinfected mosquitoes from one of the populations exhibited more ZIKV copies in the head at 7 dpsi and increased mortality. These different outcomes toward field-derived Ae. aegypti might provide additional information regarding local epidemiological settings. In conclusion, ZIKV infection yields a reduction in Ae. aegypti survival but produced no effects on mosquito fecundity and oviposition success. Therefore, the presence of ZIKV negatively affected the Ae. aegypti vectorial capacity by reducing mosquito lifespan.
ZIKV-infected and uninfected mosquitoes were blood fed once a week on anesthetized mice. This study was carried out in accordance with the recommendations of Fiocruz Ethical Committee for Animal Use. The protocol was approved by the Fiocruz Ethical Committee for Animal Use (CEUA LW-32/14).
IS, GG, MGP, and RM were responsible for the conception and design of study; IS, MTP, GS, and MTP were responsible for the acquisition of data; MD, MGP, and RM performed the data analysis; IS, MD, MGP, and RM wrote the manuscript. All authors participated in interpretation of the findings, and all authors read and approved the final version of the manuscript. All authors confirm that the content has not been published elsewhere and does not overlap with or duplicate their published work.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
This work was supported by the Brazilian Research Councils MCTIC/FNDCT-CNPq/MEC-CAPES/MS-Decit E14/2016 (440929/2016-4), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - Brasil (CAPES, 23038.007199/2012-17 and Finance Code 001) and FAPERJ E18/2015. Funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
We would like to thank Dr. Rafaela Vieira Bruno for her valuable comments.
Benjamini, Y., and Yekutieli, D. (2001). The control of the false discovery rate in multiple testing under dependency. Ann. Stat. 29, 1165–1188. doi: 10.1214/aos/1013699998
Bolling, B. G., Olea-Popelka, F. J., Eisen, L., Moore, C. G., and Blair, C. D. (2012). Transmission dynamics of an insect-specific flavivirus in a naturally infected Culex pipiens laboratory colony and effects of co-infection on vector competence for West Nile virus. Virology 427, 90–97. doi: 10.1016/j.virol.2012.02.016
Bonaldo, M. C., Ribeiro, I. P., Lima, N. S., dos Santos, A. A. C., Menezes, L. S. R., da Cruz, S. O. D., et al. (2016). Isolation of infective Zika virus from urine and saliva of patients in Brazil. PLoS Negl. Trop. Dis. 10:e0004816. doi: 10.1371/journal.pntd.0004816
Brady, O, J., Godfray, H. C., Tatem, A. J., Gething, P. W., Cohen, J. M., McKenzie, F. E., et al. (2016). Vectorial capacity and vector control: reconsidering sensitivity to parameters for malaria elimination. Trans. R. Soc. Trop. Med. Hyg. 110, 107–117. doi: 10.1093/trstmh/trv113
Chouin-Carneiro, T., Vega-Rua, A., Vazeille, M., Yebakima, A., Girod, R., Goindin, D., et al. (2016). Differential susceptibilities of Aedes aegypti and Aedes albopictus from the Americas to Zika Virus. PLoS Negl. Trop. Dis. 10:e0004543. doi: 10.1371/journal.pntd.0004543
Cohen, J. (1988). Statistical Power Analysis for the Behavioral Sciences . 2nd Edn. New York: Lawrence Erlbaum Associates.
David, M. R., Lourenço-de-Oliveira, R., and De Freitas, R. M. (2009). Container productivity, daily survival rates and dispersal of Aedes aegypti mosquitoes in a high income dengue epidemic neighbourhood of Rio de Janeiro: presumed influence of differential urban structure on mosquito biology. Mem. Inst. Oswaldo Cruz 104, 927–932. Available at: http://www.ncbi.nlm.nih.gov/pubmed/19876569, doi: doi: 10.1590/S0074-02762009000600019
Dawes, E. J., Churcher, T. S., Zhuang, S., Sinden, R. E., and Basáñez, M.-G. (2009). Anopheles mortality is both age- and Plasmodium-density dependent: implications for malaria transmission. Malar. J. 8:228. doi: 10.1186/1475-2875-8-228
Dutra, H. L. C., Rocha, M. N., Dias, F. B. S., Mansur, S. B., Caragata, E. P., and Moreira, L. A. (2016). Wolbachia blocks currently circulating Zika virus isolates in Brazilian Aedes aegypti mosquitoes. Cell Host Microbe 19, 771–774. doi: 10.1016/j.chom.2016.04.021
Ehrbar, D. J., Ngo, K. A., Campbell, S. R., Kramer, L. D., and Ciota, A. T. (2017). High levels of local inter- and intra-host genetic variation of West Nile virus and evidence of fine-scale evolutionary pressures. Infect. Genet. Evol. 51, 219–226. doi: 10.1016/j.meegid.2017.04.010
Etebari, K., Hegde, S., Saldaña, M. A., Widen, S. G., Wood, T. G., Asgari, S., et al. (2017). Global transcriptome analysis of Aedes aegypti mosquitoes in response to Zika virus infection. mSphere 2, pii:e00456–17. doi: 10.1128/mSphere.00456-17
Failloux, A.-B., Vazeille, M., and Rodhain, F. (2002). Geographic genetic variation in populations of the dengue virus vector Aedes aegypti. J. Mol. Evol. 55, 653–663. doi: 10.1007/s00239-002-2360-y
Fay, R. W., and Eliason, D. A. (1966). A preferred oviposition site as a surveillance method for Aedes aegypti. Mosq. News 26, 531–535
Fernandes, R. S., Campos, S. S., Ferreira-de-Brito, A., de Miranda, R. M., Barbosa da Silva, K. A., de Castro, M. G., et al. (2016). Culex quinquefasciatus from Rio de Janeiro is not competent to transmit the local Zika virus. PLoS Negl. Trop. Dis. 10, 1–13. doi: 10.1371/journal.pntd.0004993
Ferreira-de-Brito, A., Ribeiro, I. P., de Miranda, R. M., Fernandes, R. S., Campos, S. S., da Silva, K. A. B., et al. (2016). First detection of natural infection of Aedes aegypti with Zika virus in Brazil and throughout South America. Mem. Inst. Oswaldo Cruz 111, 655–658. doi: 10.1590/0074-02760160332
Gubler, D. J., Nalim, S., Tan, R., Saipan, H., and Sulianti Saroso, J. (1979). Variation in susceptibility to oral infection with dengue viruses among geographic strains of Aedes aegypti. Am. J. Trop. Med. Hyg. 28, 1045–1052. Available at: http://www.ncbi.nlm.nih.gov/pubmed/507282, doi: 10.4269/ajtmh.1979.28.1045
Harbach, R., and Knight, K. (1980). Taxonomists’ Glossary of Mosquito Anatomy . Marlton, New Jersey: Plexus Publishing
Hogg, J. C., and Hurd, H. (1995). Malaria-induced reduction of fecundity during the first gonotrophic cycle of Anopheles stephensi mosquitoes. Med. Vet. Entomol. 9, 176–180. Available at: http://www.ncbi.nlm.nih.gov/pubmed/7787226, doi: 10.1111/j.1365-2915.1995.tb00175.x
Igarashi, A. (1979). Characteristics of Aedes albopictus cells persistently infected with dengue viruses. Nature 280, 690–691. Available at: http://www.ncbi.nlm.nih.gov/pubmed/471048, doi: 10.1038/280690a0
Jahan, N., and Hurd, H. (1998). Effect of Plasmodium yoelii nigeriensis (Haemosporidia: Plasmodiidae) on Anopheles stephensi (Diptera: Culicidae) vitellogenesis. J. Med. Entomol. 35, 956–961. Available at: http://www.ncbi.nlm.nih.gov/pubmed/9835686, doi: 10.1093/jmedent/35.6.956
Koella, J. C., Sorensen, F. L., and Anderson, R. A. (1998). The malaria parasite, Plasmodium falciparum, increases the frequency of multiple feeding of its mosquito vector, Anopheles gambiae. Proc. R. Soc. B Biol. Sci. 265, 763–768. doi: 10.1098/rspb.1998.0358
Kramer, L. D., and Ciota, A. T. (2015). Dissecting vectorial capacity for mosquito-borne viruses. Curr. Opin. Virol. 15, 112–118. doi: 10.1016/j.coviro.2015.10.003
Kuno, G. (1982). Persistent infection of a nonvector mosquito cell line (TRA-171) with dengue viruses. Intervirology 18, 45–55. doi: 10.1159/000149303
Kuwata, R., Isawa, H., Hoshino, K., Sasaki, T., Kobayashi, M., Maeda, K., et al. (2015). Analysis of mosquito-borne flavivirus superinfection in Culex tritaeniorhynchus (Diptera: Culicidae) cells persistently infected with culex flavivirus (Flaviviridae). J. Med. Entomol. 52, 222–229. doi: 10.1093/jme/tju059
Lambrechts, L., Chevillon, C., Albright, R. G., Thaisomboonsuk, B., Richardson, J. H., Jarman, R. G., et al. (2009). Genetic specificity and potential for local adaptation between dengue viruses and mosquito vectors. BMC Evol. Biol. 9:160. doi: 10.1186/1471-2148-9-160
Li, C.-X., Guo, X.-X., Deng, Y.-Q., Xing, D., Sun, A.-J., Liu, Q.-M., et al. (2017). Vector competence and transovarial transmission of two Aedes aegypti strains to Zika virus. Emerg. Microbes Infect. 6:e23. doi: 10.1038/emi.2017.8
Liu-Helmersson, J., Stenlund, H., Wilder-Smith, A., and Rocklöv, J. (2014). Vectorial capacity of Aedes aegypti: effects of temperature and implications for global dengue epidemic potential. PLoS One 9:e89783. doi: 10.1371/journal.pone.0089783
Liu, Y., Liu, J., Du, S., Shan, C., Nie, K., Zhang, R., et al. (2017). Evolutionary enhancement of Zika virus infectivity in Aedes aegypti mosquitoes. Nature 545, 482–486. doi: 10.1038/nature22365
Longdon, B., Hadfield, J. D., Day, J. P., Smith, S. C. L., McGonigle, J. E., Cogni, R., et al. (2015). The causes and consequences of changes in virulence following pathogen host shifts. PLoS Pathog. 11:e1004728. doi: 10.1371/journal.ppat.1004728
Luz, P. M., Codeço, C. T., Massad, E., and Struchiner, C. J. (2003). Uncertainties regarding dengue modeling in Rio de Janeiro, Brazil. Mem. Inst. Oswaldo Cruz 98, 871–878. Available at: http://www.ncbi.nlm.nih.gov/pubmed/21519709, doi: 10.1590/S0074-02762003000700002
Maciel-De-Freitas, R., Codeço, C. T., and Lourenço-De-Oliveira, R. (2007). Daily survival rates and dispersal of Aedes aegypti females in Rio de Janeiro, Brazil. Am. J. Trop. Med. Hyg. 76, 659–665. doi: 76/4/659 [pii]
Maciel-de-Freitas, R., Eiras, A. E., and Lourenço-de-Oliveira, R. (2008). Calculating the survival rate and estimated population density of gravid Aedes aegypti (Diptera, Culicidae) in Rio de Janeiro, Brazil, Cad. Saude Publica 24, 2747–2754. Available at: http://www.ncbi.nlm.nih.gov/pubmed/19082265, doi: 10.1590/S0102-311X2008001200003
Maciel-de-Freitas, R., Koella, J. C., and Lourenço-de-Oliveira, R. (2011). Lower survival rate, longevity and fecundity of Aedes aegypti (Diptera: Culicidae) females orally challenged with dengue virus serotype 2. Trans. R. Soc. Trop. Med. Hyg. 105, 452–458. doi: 10.1016/j.trstmh.2011.05.006
Maciel-de-Freitas, R., and Lourenço-de-Oliveira, R. (2009). Presumed unconstrained dispersal of Aedes aegypti in the city of Rio de Janeiro, Brazil, Rev. Saude Publica 43, 8–12. Available at: http://www.ncbi.nlm.nih.gov/pubmed/19169571, doi: 10.1590/S0034-89102009000100002
Maciel-de-Freitas, R., Sylvestre, G., Gandini, M., and Koella, J. C. (2013). The Influence of Dengue Virus Serotype-2 Infection on Aedes aegypti (Diptera: Culicidae) Motivation and Avidity to Blood Feed. PLoS One 8:e65252. doi: 10.1371/journal.pone.0065252
Martin, E., Moutailler, S., Madec, Y., and Failloux, A.-B. (2010). Differential responses of the mosquito Aedes albopictus from the Indian Ocean region to two chikungunya isolates. BMC Ecol. 10:8. doi: 10.1186/1472-6785-10-8
Moncayo, A. C., Edman, J. D., and Turell, M. J. (2000). Effect of eastern equine encephalomyelitis virus on the survival of Aedes albopictus, Anopheles quadrimaculatus, and Coquillettidia perturbans (Diptera: Culicidae). J. Med. Entomol. 37, 701–706. Available at: http://www.ncbi.nlm.nih.gov/pubmed/11004781, doi: 10.1603/0022-2585-37.5.701
Patterson, J., Sammon, M., and Garg, M. (2016). Dengue, Zika and Chikungunya: emerging arboviruses in the new world. West J. Emerg. Med. 17, 671–679. doi: 10.5811/westjem.2016.9.30904
Platt, K. B., Linthicum, K. J., Myint, K. S., Innis, B. L., Lerdthusnee, K., and Vaughn, D. W. (1997). Impact of dengue virus infection on feeding behavior of Aedes aegypti. Am. J. Trop. Med. Hyg. 57, 119–125. Available at: http://www.ncbi.nlm.nih.gov/pubmed/9288801, doi: 10.4269/ajtmh.1997.57.119
Putnam, J. L., and Scott, T. W. (1995). Blood-feeding behavior of dengue-2 virus-infected Aedes aegypti. Am. J. Trop. Med. Hyg. 52, 225–227. Available at: http://www.ncbi.nlm.nih.gov/pubmed/7694963, doi: 10.4269/ajtmh.1995.52.225
R Development Core Team, R. (2011). R: a language and environment for statistical computing. R Found. Stat. Comput. 1:409. doi: 10.1007/978-3-540-74686-7
Rašić, G., Schama, R., Powell, R., Maciel-de Freitas, R., Endersby-Harshman, N. M., Filipović, I., et al. (2015). Contrasting genetic structure between mitochondrial and nuclear markers in the dengue fever mosquito from Rio de Janeiro: implications for vector control. Evol. Appl. 8, 901–915. doi: 10.1111/eva.12301
Robinson, A., Busula, A. O., Voets, M. A., Beshir, K. B., Caulfield, J. C., Powers, S. J., et al. (2018). Plasmodium -associated changes in human odor attract mosquitoes. Proc. Natl. Acad. Sci. E4209–E4218. doi: 10.1073/pnas.1721610115
Rossi, S. L., Ebel, G. D., Shan, C., Shi, P.-Y., and Vasilakis, N. (2018). Did Zika Virus Mutate to Cause Severe Outbreaks?Trends Microbiol. 26, 877–885. doi: 10.1016/j.tim.2018.05.007
Rossignol, P. A., Ribeiro, J. M., and Spielman, A. (1986). Increased biting rate and reduced fertility in sporozoite-infected mosquitoes. Am. J. Trop. Med. Hyg. 35, 277–279. Available at: http://www.ncbi.nlm.nih.gov/pubmed/3953943, doi: 10.4269/ajtmh.1986.35.277
Roundy, C. M., Azar, S. R., Rossi, S. L., Huang, J. H., Leal, G., Yun, R., et al. (2017). Variation in Aedes aegypti mosquito competence for Zika virus transmission. Emerg. Infect. Dis. 23, 625–632. doi: 10.3201/eid2304.161484
Salas-Benito, J. S., and De Nova-Ocampo, M. (2015). Viral interference and persistence in mosquito-borne flaviviruses. J. Immunol. Res. 2015, 1–14. doi: 10.1155/2015/873404
Schwartz, A., and Koella, J. C. (2001). Trade-offs, conflicts of interest and manipulation in Plasmodium-mosquito interactions. Trends Parasitol. 17, 189–194. Available at: http://www.ncbi.nlm.nih.gov/pubmed/11282509, doi: 10.1016/S1471-4922(00)01945-0
Scott, T. W., and Lorenz, L. H. (1998). Reduction of Culiseta melanura fitness by eastern equine encephalomyelitis virus. Am. J. Trop. Med. Hyg. 59, 341–346. Available at: http://www.ncbi.nlm.nih.gov/pubmed/9715958, doi: 10.4269/ajtmh.1998.59.341
Service, M. W. (1993). Mosquito Ecology . Dordrecht: Springer Netherlands doi: 10.1007/978-94-015-8113-4
Severson, D. W., and Behura, S. K. (2016). Genome investigations of vector competence in Aedes aegypti to inform novel arbovirus disease control approaches. Insects 7. doi: 10.3390/insects7040058
Shi, W., Zhang, Z., Ling, C., Carr, M. J., Tong, Y., and Gao, G. F. (2016). Increasing genetic diversity of Zika virus in the Latin American outbreak. Emerg. Microbes Infect. 5:e68. doi: 10.1038/emi.2016.68
Sim, S., and Dimopoulos, G. (2010). Dengue virus inhibits immune responses in Aedes aegypti Cells. PLoS One 5:e10678. doi: 10.1371/journal.pone.0010678
Sim, S., Ramirez, J. L., and Dimopoulos, G. (2012). Dengue virus infection of the Aedes aegypti salivary gland and chemosensory apparatus induces genes that modulate infection and blood-feeding behavior. PLoS Pathog. 8:e1002631. doi: 10.1371/journal.ppat.1002631
Souza-Neto, J. A., Sim, S., and Dimopoulos, G. (2009). An evolutionary conserved function of the JAK-STAT pathway in anti-dengue defense. Proc. Natl. Acad. Sci. 106, 17841–17846. doi: 10.1073/pnas.0905006106
Styer, L. M., Meola, M. A., and Kramer, L. D. (2007). West Nile virus infection decreases fecundity of Culex tarsalis females. J. Med. Entomol. 44, 1074–1085. Available at: http://www.ncbi.nlm.nih.gov/pubmed/18047209. doi: 10.1093/jmedent/44.6.1074
Sylvestre, G., Gandini, M., and Maciel-de-Freitas, R. (2013). Age-dependent effects of oral infection with dengue virus on Aedes aegypti (Diptera: Culicidae) feeding behavior, survival, oviposition success and fecundity. PLoS One 8:e59933. doi: 10.1371/journal.pone.0059933
Tabachnick, W. (2013). Nature, nurture and evolution of intra-species variation in mosquito arbovirus transmission competence. Int. J. Environ. Res. Public Health 10, 249–277. doi: 10.3390/ijerph10010249
Vezilier, J., Nicot, A., Gandon, S., and Rivero, A. (2012). Plasmodium infection decreases fecundity and increases survival of mosquitoes. Proc. R. Soc. B Biol. Sci. 279, 4033–4041. doi: 10.1098/rspb.2012.1394
Wang, A., Thurmond, S., Islas, L., Hui, K., and Hai, R. (2017). Zika virus genome biology and molecular pathogenesis. Emerg. Microbes Infect. 6:e13. doi: 10.1038/emi.2016.141
Weaver, S. C., and Reisen, W. K. (2010). Present and future arboviral threats. Antivir. Res. 85, 328–345. doi: 10.1016/j.antiviral.2009.10.008
Keywords: Aedes aegypti, Zika, vectorial capacity, survival, fecundity, disease transmission
Citation: da Silveira ID, Petersen MT, Sylvestre G, Garcia GA, David MR, Pavan MG and Maciel-de-Freitas R (2018) Zika Virus Infection Produces a Reduction on Aedes aegypti Lifespan but No Effects on Mosquito Fecundity and Oviposition Success. Front. Microbiol. 9:3011. doi: 10.3389/fmicb.2018.03011
Edited by:
Lisa Sedger, University of Technology Sydney, AustraliaReviewed by:
Matthew H. Collins, Emory University, United StatesCopyright © 2018 da Silveira, Petersen, Sylvestre, Garcia, David, Pavan and Maciel-de-Freitas. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rafael Maciel-de-Freitas, ZnJlaXRhc0Bpb2MuZmlvY3J1ei5icg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.