
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 22 August 2018
Sec. Evolutionary and Genomic Microbiology
Volume 9 - 2018 | https://doi.org/10.3389/fmicb.2018.02007
 Imen Nouioui1†
Imen Nouioui1† Lorena Carro1†
Lorena Carro1† Marina García-López2†
Marina García-López2† Jan P. Meier-Kolthoff2
Jan P. Meier-Kolthoff2 Tanja Woyke3
Tanja Woyke3 Nikos C. Kyrpides3
Nikos C. Kyrpides3 Rüdiger Pukall2
Rüdiger Pukall2 Hans-Peter Klenk1
Hans-Peter Klenk1 Michael Goodfellow1
Michael Goodfellow1 Markus Göker2*
Markus Göker2*The application of phylogenetic taxonomic procedures led to improvements in the classification of bacteria assigned to the phylum Actinobacteria but even so there remains a need to further clarify relationships within a taxon that encompasses organisms of agricultural, biotechnological, clinical, and ecological importance. Classification of the morphologically diverse bacteria belonging to this large phylum based on a limited number of features has proved to be difficult, not least when taxonomic decisions rested heavily on interpretation of poorly resolved 16S rRNA gene trees. Here, draft genome sequences of a large collection of actinobacterial type strains were used to infer phylogenetic trees from genome-scale data using principles drawn from phylogenetic systematics. The majority of taxa were found to be monophyletic but several orders, families, and genera, as well as many species and a few subspecies were shown to be in need of revision leading to proposals for the recognition of 2 orders, 10 families, and 17 genera, as well as the transfer of over 100 species to other genera. In addition, emended descriptions are given for many species mainly involving the addition of data on genome size and DNA G+C content, the former can be considered to be a valuable taxonomic marker in actinobacterial systematics. Many of the incongruities detected when the results of the present study were compared with existing classifications had been recognized from 16S rRNA gene trees though whole-genome phylogenies proved to be much better resolved. The few significant incongruities found between 16S/23S rRNA and whole genome trees underline the pitfalls inherent in phylogenies based upon single gene sequences. Similarly good congruence was found between the discontinuous distribution of phenotypic properties and taxa delineated in the phylogenetic trees though diverse non-monophyletic taxa appeared to be based on the use of plesiomorphic character states as diagnostic features.
Prokaryotic systematics is the study of the kinds and diversity of Archaea and Bacteria and relationships within and between them. It encompasses the related disciplines of classification, nomenclature and identification, the concept of the taxonomic trinity (Cowan, 1955). Classification and identification are core scientific disciplines that are practiced by few but their applications are relevant to most, if not all, of the microbiological community. Nomenclature is also central to all aspects of microbiology as it deals with the correct use of names by following rules embodied in the International Code of Nomenclature of Prokaryotes, notably the nomenclatural type concept and the requirement to deposit type strains in two public culture collections in different countries (Stackebrandt et al., 2014; Parker et al., 2015). Classification and identification are markedly data-dependent and thereby in a perpetual state of development driven by new taxonomic concepts and practices.
Early classification of prokaryotes, including actinomycetes, which are now known as actinobacteria, was based on morphology and a few biochemical and physiological properties, as exemplified by Nocardia, the oldest name in current use for an aerobic actinomycete genus (Lechevalier, 1976). However, the subsequent application of chemotaxonomic, molecular systematic and numerical taxonomic procedures (Goodfellow et al., 1985) showed that classifications dependent on form and function tended to result in the delineation of heterogeneous taxa. Nocardia, for instance, as morphologically defined in the eighth edition of Bergey's Manual of Determinative Bacteriology (McClung, 1974) contained species that were later assigned taxonomically to diverse genera, including Actinomadura, Amycolatopsis, Oerskovia, Rhodococcus, and Rothia (Goodfellow and Minnikin, 1977; Lechevalier et al., 1986). Similarly, a series of more broadly based studies showed that coryneform bacteria, at the time an ill-assorted group in search of a taxonomic home, were actinomycetes (Bousfield and Callely, 1978). Conversely, Thermoactinomyces, which had, long been classified with the actinomycetes, was found to be related to aerobic, endospore-forming bacilli (Cross and Unsworth, 1981).
The new advances, especially in molecular systematics, led to the view that classifications at all levels in the taxonomic hierarchy should be based on the integrated use of genotypic and phenotypic data (Wayne et al., 1987; Stackebrandt, 1992). This approach, known as polyphasic taxonomy (Colwell, 1970), has become the lodestone of prokaryotic systematics (Vandamme et al., 1996; Gillis et al., 2005; Kämpfer and Glaeser, 2012). Methods selected for polyphasic studies tend to be somewhat subjective as they reflect the biology of taxa under study and available facilities. However, a prevailing theme in such studies is the comparative analysis of 16S rRNA gene sequences designed (Kim and Chun, 2014; Yarza and Munoz, 2014) to infer relationships between strains at generic and suprageneric ranks (Ludwig et al., 2012).
The widespread application of polyphasic taxonomy led to sweeping changes in the classification of prokaryotes (Brenner et al., 2005b,c; De Vos et al., 2009), including actinomycetes, as exemplified by the current classification of the phylum Actinobacteria, as shown in Bergey's Manual of Systematic Bacteriology (Goodfellow et al., 2012a). Despite the success of 16S rRNA gene sequencing in providing the phylogenetic backbone for the classification of prokaryotes (Brenner et al., 2005a; Ludwig and Klenk, 2005) problems remain in circumscribing actinobacterial taxa, including at family and genus levels. Nocardiaceae (Castellani and Chalmers, 1919), for instance, encompass the type genus Nocardia as well as Gordonia, Millisia, Rhodococcus, Skermania, and Smaragdicoccus (Goodfellow, 2012a) though a case was also made for the continued recognition of Gordoniaceae (Stackebrandt et al., 1997) to include not only the type genus, Gordonia, but also Millisia, Skermania, and Williamsia (Goodfellow, 2012a). Other families considered to be problematic include Cellulomonadaceae, Dermacoccaceae, Dermatophilaceae, Intrasporangiaceae, and Nocardioidaceae (Goodfellow, 2012a). The current assignment of genera to such families should be seen as staging posts to better classifications.
The situation at the genus level is not dissimilar to that outlined above as 16S rRNA gene trees often also lack the resolution to distinguish between closely related genera, as shown by Lechevalieria (Labeda et al., 2001) and Lentzea (Yassin et al., 1995), which are difficult to distinguish when assigning new species to either genus (Okoro et al., 2010; Idris et al., 2017). In turn, the observation that Kitasatospora (Omura et al., 1982) and Streptacidiphilus (Kim et al., 2003) fall within the Streptomyces 16S rRNA gene tree led some to question their taxonomic status (Kämpfer, 2012a). Along the same lines, the monospecific genera Jiangella (Song et al., 2005), Plantactinospora (Qin et al., 2009a), and Polymorphospora (Tamura et al., 2006) were interspersed within the Micromonospora 16S rRNA gene tree together with authentic representatives of Salinispora (Maldonado et al., 2005) and Verrucosispora (Rheims et al., 1998) in a recent study (Trujillo et al., 2014). It is also difficult to unravel the internal structure of complex actinobacterial genera using data acquired from polyphasic taxonomic studies, notable examples include Actinomyces (Schaal and Yassin, 2012a), Amycolatopsis (Tan and Goodfellow, 2012), Arthrobacter (Busse et al., 2012), Micromonospora (Carro et al., 2018), Nocardioides (Evtushenko et al., 2012), Rhodococcus (Jones and Goodfellow, 2012), and Streptomyces (Labeda et al., 2012). Polyphasic studies as currently conducted largely depend on the 16S rRNA gene (Montero-Calasanz et al., 2017) but despite its usefulness for resolving taxonomic questions in the past, the gene contains only a limited number of characters and thus, much like any other single gene, can yield trees with many statistically unsupported branches (Klenk and Göker, 2010; Breider et al., 2014).
There is clearly a compelling need to develop an improved framework for the classification of actinobacteria, a goal in its own right (Klenk and Göker, 2010) but also one driven by biotechnological, bioprospecting, and ecological imperatives (Bull et al., 2016; Katz and Baltz, 2016; Benito et al., 2017) and by fundamental research interests (Chandra and Chater, 2014; Barka et al., 2016). Given the rapid and ongoing progress in sequencing technologies (Mavromatis et al., 2012), classifications based on whole genome sequences and associated bioinformatic tools are addressing these needs as they are based on millions of unit characters and thereby provide a step change in reliability, as evidenced by significantly high average bootstrap support in phylogenomic trees (Breider et al., 2014; Meier-Kolthoff et al., 2014a). In contrast, trees based on a few thousand nucleotides (Tang et al., 2016) tend to have branches with low bootstrap values; the same limitation applies, albeit to a lesser extent, to multi-locus sequence analyses of conserved housekeeping genes (Glaeser and Kämpfer, 2015), which can hardly be called genome-scale (Klenk and Göker, 2010). Phylogenomic methods have already been applied to elucidate the classification of complex actinobacterial taxa, such as the genera Amycolatopsis, Micromonospora, Rhodococcus, and Salinispora (Sangal et al., 2016; Tang et al., 2016; Jensen, 2017; Carro et al., 2018), and in some cases has led to marked reclassification (Montero-Calasanz et al., 2017). However, a comprehensive analysis of the phylum Actinobacteria based on truly genome-scale methods has not been undertaken.
Experimental methods that indirectly determine DNA G+C composition (Mesbah et al., 1989; Moreira et al., 2011) can now be replaced by calculating it directly from accurate genome sequences. Claims in the literature that the variation in G+C content within bacterial species is at most 3 mol% (Mesbah et al., 1989) or even 5% (Rosselló-Mora and Amann, 2001) can be attributed to experimental error as within-species variation is at most 1% when G+C content is calculated from genome sequences (Meier-Kolthoff et al., 2014c). The G+C values cited in many species descriptions are often out of sync with those directly calculated from genome sequences, which also have a significantly better fit to the phylogeny than the former (Hahnke et al., 2016). Emendations to species descriptions are needed where estimates from experimental methods differ by more than 1% from in silico derived values (Meier-Kolthoff et al., 2014c). Similarly, description of higher ranked taxa should be emended if the range of G+C values are shown to be in conflict with corresponding values determined from genome sequences. In the same vein, digital DNA:DNA hybridization (DDH) values provide a more accurate way of establishing relationships between closely related species than corresponding data derived from experimental methods (Auch et al., 2010a,b; Meier-Kolthoff et al., 2013a), whereas genome size has as yet been less well elucidated as taxonomic marker.
Results from incorporating new technologies into systematics often raise the question whether discrepancies to earlier outcomes, if any, are caused by conflicts in the data themselves, such as between the phenotype, single genes or entire genomes, or by conflicting approaches of data interpretation such as those between the distinct schools of taxonomy (Klenk and Göker, 2010). In phylogenetic systematics only monophyletic taxa can be accepted in taxonomic classifications as they are designed to summarize the phylogeny of the classified organisms (Hennig, 1965; Wiley and Lieberman, 2011). While principles of phylogenetic systematics should also be used to guide microbial classification (Klenk and Göker, 2010; Hahnke et al., 2016), they have specific implications on how single characters, such as phenotypic ones, should be interpreted, even though this has not been widely considered in current taxonomic practice in microbiology (Montero-Calasanz et al., 2017).
The present study was designed to provide an improved framework for the classification of actinobacteria based on the principles of phylogenetic systematics, in line with our earlier study on Bacteroidetes (Hahnke et al., 2016). To this end, a comprehensive sampling of publicly available whole genome sequences of actinobacterial type strains was used to construct genome-scale phylogenetic trees and to address the following questions: (i) to what extent are phylogenies calculated from whole genome sequences in conflict with current actinobacterial classification and with 16S (or 23S) rRNA gene phylogenies? (ii) Which taxa need to be revised because they are evidently non-monophyletic? (iii) What is the historical cause of these discrepancies? (iv) Which taxon descriptions should be modified because of inaccurate or missing G+C values? and (v) How does genome size relate to phylogeny and genomic G+C content and how can it serve as a taxonomic marker?
A total number of 1,142 actinobacterial (ingroup) and 25 chloroflexi (outgroup) type-strain genomes were downloaded mainly from GenBank and to a smaller degree from the Integrated Microbial Genomes platform (Markowitz et al., 2009), that originated from the Genomic Encyclopedia of Archaea and Bacteria pilot phase or phase 1 of the One Thousand Microbial Genomes project (Kyrpides et al., 2014; Mukherjee et al., 2017). GenBank sequences comprising more than 500 contigs were discarded. The complete genome list is found in Supplementary Table 1. Further processing of these genome sequences closely followed an earlier study on genome-based classification (Hahnke et al., 2016). The high-throughput version (Meier-Kolthoff et al., 2014a) of the Genome BLAST Distance Phylogeny (GBDP) approach (Auch et al., 2006) was used to infer genome-scale phylogenies from whole proteomes in conjunction with BLAST+ (v2.2.30) (Camacho et al., 2009) in BLASTP mode with default parameters except for an e-value filter of 10−8 (Meier-Kolthoff et al., 2014a). The greedy-with-trimming GBDP algorithm was applied in conjunction with formula d5 and subjected to 100 pseudo-bootstrap replicates (Meier-Kolthoff et al., 2013a, 2014a). FastME (Lefort et al., 2015) was used to infer phylogenetic trees from the original and pseudo-bootstrapped intergenomic distance matrices, and trees and support values were visualized using Interactive Tree Of Life (Letunic and Bork, 2016) in conjunction with the script found at https://github.com/mgoeker/itol for facilitating tree annotation. Digital DNA:DNA hybridization (dDDH) was conducted with the recommended settings of the Genome-To-Genome Distance Calculator (GGDC) version 2.1 (Meier-Kolthoff et al., 2013a) to clarify species and subspecies affiliations.
Genomic G+C content values and genome sizes (approximate genome sizes in the case of incompletely sequenced genomes) were calculated as previously described (Hahnke et al., 2016); they were also recorded for single contigs to detect potential contamination of genome sequences. For the subsequent statistical tests, which were conducted without the outgroup, G+C content values were logit-transformed and genome sizes were log-transformed to account for the dependence of the variance on the mean in the case of proportion and count data (Crawley, 2007). Phylogenetic conservation was determined using the phylosignal function from the picante package (Kembel et al., 2010) for the R statistical environment (R Development Core Team, 2014) with 9,999 permutations. Phylogenetically independent contrasts (Felsenstein, 1985) were determined using the pic function of the ape package (Paradis et al., 2004). The relationship between G+C content and genome size was analyzed with Pearson and Kendall correlations. Loess-based bootstrap aggregating, that is bagging (Breiman, 1996), was applied in R with 100 replicates to increase stability and reduce overfitting.
Comprehensive, aligned, near full-length 16S rRNA gene sequence data for the Actinobacteria and Chloroflexi were initially taken from the All-Species Living Tree Project (Yarza et al., 2008) and considerably augmented by screening the more recent taxonomic literature (the complete DSMZ reference collection comprises c. 16,000 type-strain 16S rRNA gene sequences of Archaea and Bacteria). All of the rRNA genes were extracted from the genome sequences using RNAmmer version 1.2 (Lagesen et al., 2007) and the extracted 16S rRNA genes were compared using BLAST search (and in ambiguous cases also with phylogenetic trees) with the 16S rRNA gene reference data base (Chun et al., 2018). Non-matching genome sequences were discarded. Sequences were aligned using MAFFT version 7.271 with the “localpair” option (Katoh et al., 2005). Depending on the length and number of ambiguous bases, either the sequences extracted from the genome sequences or the previously published 16S sequences were used. Trees were inferred from the alignment with RAxML under the maximum-likelihood (ML) criterion using GTR+CAT as model approximation (Stamatakis, 2014) and with TNT under maximum-parsimony (MP) (Goloboff et al., 2008), as previously described (Hahnke et al., 2016). In addition to these unconstrained, comprehensive 16S rRNA gene trees (UCT), constrained comprehensive trees (CCT) were inferred with ML and MP using the bipartitions of the GBDP tree with ≥95% support as backbone constraint. Finally, unconstrained 16S rRNA gene trees reduced to genome-sequenced strains were inferred, as well as unconstrained 23S (i.e., large subunit) rRNA gene trees (ULT).
All of the trees were compared with the current classification used in LTP version s123, which was cleaned from inconsistencies such as mismatches between species and genus names and subsequently modified manually by incorporating newer literature sources. For instance, we followed the recent abandonment of subclass and suborder categories in Bergey's Manual of Systematic Bacteriology (Ludwig et al., 2012). Taxa were checked to determine whether they were monophyletic, paraphyletic or polyphyletic (Farris, 1974; Wood, 1994) as previously described (Hahnke et al., 2016).
Taxa non-monophyletic according to the GBDP tree were tested for evidence for their monophyly in the ULT, UCT, and the 16S rRNA gene trees, if any, in the original publication. In the case of a significant conflict (i.e., high support values for contradicting bipartitions) or low support in the GBDP tree, additional phylogenomic analyses of selected taxa were conducted. To this end, the reciprocal best hits from GBDP/BLAST were clustered with MCL under default settings and an e-value filter of 10−5 in analogy to OrthoMCL (Li et al., 2003a). The resulting sets of orthologous proteins were aligned with MAFFT and concatenated to form a supermatrix. The few clusters that still contained more than a single protein for at least one genome were discarded. Core-genome supermatrices were constructed for the orthologs that occurred in all of the genomes, whereas comprehensive supermatrices were compiled from all the orthologs that occurred in at least four genomes. Supermatrices were analyzed with TNT, and with RAxML under the PROTCATLGF model (Le and Gascuel, 2008), in conjunction with 100 partition bootstrap replicates, i.e., by sampling (with replacement) the orthologs instead of the single alignment positions (Siddall, 2010; Simon et al., 2017).
Unambiguously non-monophyletic taxa according to the genome-scale analyses were screened for published phenotypic evidence of their monophyly. Published evidence was judged as inconclusive when based on probably homoplastic characters or on probable plesiomorphic character states. Importantly, “diagnostic” features alone are insufficient in phylogenetic systematics, as plesiomorphies might well be diagnostic but just for paraphyletic groups (Hennig, 1965; Wiley and Lieberman, 2011). Finally, taxonomic consequences were proposed to fix all obviously non-monophyletic taxa by new taxon delineations sufficiently supported by the CCT, i.e., not hindered by the uncertain phylogenetic placement of taxa whose genome sequences were not available at the time of writing.
The phylogenetic tree inferred with GBDP is shown in Figures 1–8. The tree with all of the branches expanded is shown in Supplementary File 2 together with results from additional 16S rRNA gene (UCT, CCT, reduced tree), 23S rRNA gene (ULT), and supermatrix analyses, as well as a juxtaposition of GBDP and 16S rRNA gene support values. Supplementary Table 1 also contains a tabular overview of taxonomically relevant phenotypic features taken from the literature for taxa of interest. All of the classes and most of the orders, families, and genera appear to be monophyletic, mainly with high support but other taxa are in need of taxonomic revision according to the GBDP analysis. These discrepancies are considered below in decreasing order of taxonomic rank.
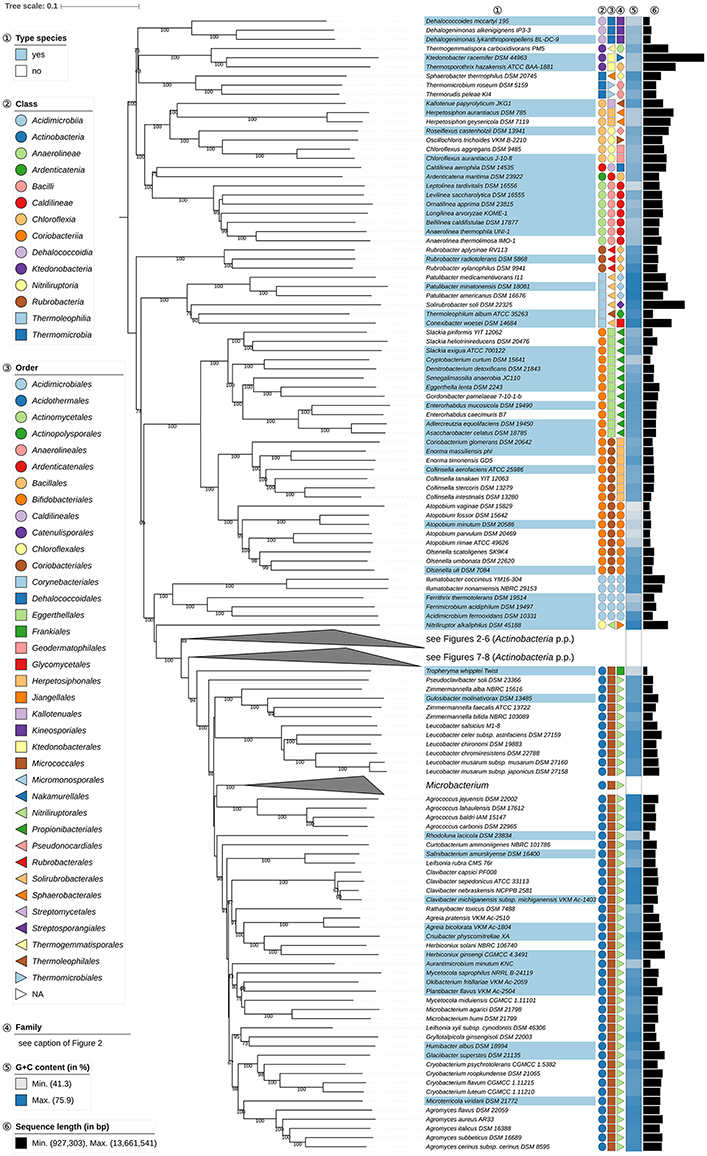
Figure 1. First part of the phylogenomic tree inferred with GBDP. Tree inferred with FastME from GBDP distances calculated from whole proteomes. The numbers above branches are GBDP pseudo-bootstrap support values from 100 replications. Tip colors indicate type species of genera, colors to the right of the tips indicate, from left to right, class, order and family (see the embedded legend for details, Figure 2 for the families). The blue gradient toward the far right indicates the exact G+C content as calculated from the genome sequences, followed by black bars indicating the (approximate) genome size in bp. The parts of the tree which have been collapsed here are shown in Figures 2–8.
Solirubrobacterales (Reddy and Garcia-Pichel, 2009) appear to be paraphyletic because of the position of Thermoleophilum album (Zarilla and Perry, 1984) of Thermoleophilales (Figure 1). However, the two branches responsible for this arrangement are poorly supported whereas the CCT, ULT, and an additional supermatrix analysis show that Solirubrobacterales are monophyletic (Supplementary File 2). It is evident that these taxa have phenotypic features in common, albeit unspecific ones (Supplementary Table 1). It can be concluded that taxonomic changes are not presently needed with respect to Solirubrobacterales and Thermoleophilales.
The position of Actinopolysporales (and Actinopolysporaceae) in the phylogenomic tree (Figure 2) causes Pseudonocardiales (and Pseudonocardiaceae) to be paraphyletic, as Actinopolyspora (Gochnauer et al., 1975), apart for A. iraqiensis (Ruan et al., 1994), appears to be a highly supported as sister group to Saccharopolyspora (Lacey and Goodfellow, 1975). Actinopolysporales arose from the elevation of Actinopolysporineae which in turn was based on signature nucleotides and the original 16S rRNA gene tree though the latter was defined with rather low backbone support; it was not clear whether these properties represent apomorphies (Zhi et al., 2009). In the CCT Bounagaea (Meklat et al., 2015) of Pseudonocardiales and Halopolyspora (Lai et al., 2014) of Actinopolysporaceae were also found to be part of the clade encompassing representative Actinopolysporaceae and Pseudonocardiaceae for which the UCT provided reasonable support. Mzabimyces algeriensis, for which Mzabimycetaceae fam. nov. had been suggested based on a largely unresolved 16S rRNA gene tree (Saker et al., 2014), was later regarded as a heterotypic synonym of Halopolyspora algeriensis (Lai et al., 2017). The phylogenetic results, even including those based only on the 16S rRNA gene, indicate that Actinopolysporaceae and Mzabimycetaceae should be classified in Pseudonocardiaceae, a taxonomically most conservative solution, and that Actinopolysporales be disbanded. It is interesting in this context that Actinopolyspora was initially classified in Pseudonocardiaceae (Embley et al., 1988), and that the phenotypic differences between Actinopolysporaceae and Pseudonocardiaceae are not pronounced (Supplementary Table 1).
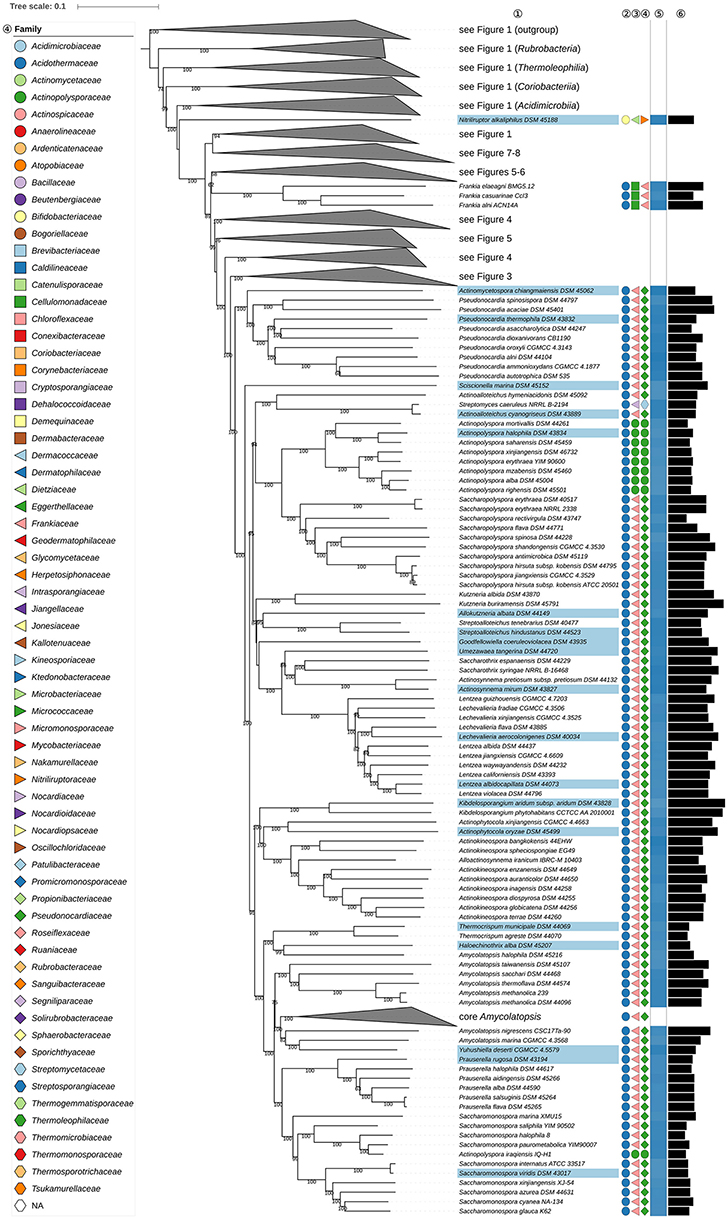
Figure 2. Second part of the phylogenomic tree inferred with GBDP. A detailed description is provided in the caption of Figure 1. The parts of the tree which have been collapsed here are shown in other figures as indicated.
Frankiales, as effectively published (Normand and Benson, 2012), encompasses in addition to Fodinicola (genus incertae sedis) nine genera, which are classified into Frankiaceae (Becking, 1970), Acidothermaceae (Stackebrandt et al., 1997), Cryptosporangiaceae (Zhi et al., 2009), Geodermatophilaceae (Normand, 2006), Motilibacteraceae (Lee, 2013b), Nakamurellaceae (Tao et al., 2004), and Sporichthyaceae (Stackebrandt et al., 1997). A phylogenomic study (Sen et al., 2014) assigned six of these families to four new orders, Frankiales, Acidothermales, Geodermatophilales, and Nakamurellales. The two remaining genera were Cryptosporangium (Tamura et al., 1998) and Sporichthya (Lechevalier et al., 1968). In the phylogenetic tree (Figures 4, 6), these genera were found to be only distantly related to the type genus of Frankiales, Frankia (Nouioui et al., 2016). Cryptosporangium forms a well-supported clade together with Glycomycetales (Labeda, 2012) and Micromonosporales (Genilloud, 2012) hence raising the prospect that the genus be assigned to a new order, Cryptosporangiales, a move in line with the isolated position of the genus in the CCT and ULT (Supplementary File 2). As detailed in Supplementary Table 1, there are no common phenotypic features between Cryptosporangium, Frankia, Glycomycetales, and Micromonosporales, apart from the unspecified peptidoglycan component DL-A2pm, i.e., meso-diaminopimelic acid (Genilloud, 2012; Labeda, 2012; Normand and Benson, 2012).
It is evident from Figure 5 that the position of Jatrophihabitans (Madhaiyan et al., 2013) is at variance with its assignment to Frankiales. Phenotypical synapomorphies of Frankia and Jatrophihabitans are not known. Jatrophihabitans forms short rods while frankiae form hyphae, multilocular sporangia and vesicles in which nitrogen is fixed (Supplementary Table 1). The position of Jatrophihabitans as the sister group to the clade comprising Antricoccus (Lee, 2015) and Geodermatophilaceae is only weakly supported (Figure 5). In the ULT (Supplementary File 2), Antricoccus and Jatrophihabitans were found to be sister groups albeit with weak support within Geodermatophilales. In light of these results, it is proposed that Jatrophihabitans be recognized as the type genus of Jatrophihabitandaceae fam. nov. without an assignment to an order. This proposal is not in conflict with the 16S rRNA gene tree shown in the initial description of the genus because the tree was poorly resolved.
Antricoccus (Lee, 2015), which is currently in search of a family, was found to be more closely related in the phylogenomic tree to Geodermatophilales than to Frankiales (Figure 5). Indeed, given its distinct position in the tree, it is best seen as a separate family within Geodermatophilales rather than placed within Geodermatophilaceae, from which it also differs in genomic G+C content (Figure 5). This proposition is not contradicted by the poorly resolved 16S rRNA gene tree presented in the original description of Antricoccus. Phenotypical synapomorphies of Antricoccus, which forms non-motile, asporogenous cocci, and Frankia are not known either (Supplementary Table 1).
It is evident from the GBDP tree (Figure 6) that Sporichthya (Lechevalier et al., 1968) of Sporichthyaceae (Stackebrandt et al., 1997) is separated from Frankiaceae. Sporichthya appears more closely related to Acidothermus cellulolyticus (Mohagheghi et al., 1986) of Acidothermaceae but without support. The sister-group relationship between Acidothermus and Sporichthya is not supported by either the CCT or ULT (Supplementary File 2) nor are synapomorphies of these taxa known. Sporichthya is motile and has LL-A2pm in the peptidoglycan whereas Acidothermus is non-motile, thermophilic with DL-A2pm in the peptidoglycan. Given these findings it is proposed that Sporichthya be recognized as the type genus of Sporichthyales ord. nov.
The relationship between Fodinicola (Carlsohn et al., 2008) and Cryptosporangiaceae was not resolved in the UCT (Supplementary File 2); the CCT showed it to have an isolated position, albeit with weak support. Likewise, the affiliation of Motilibacter (Lee, 2012, 2013b) to Frankiales is questionable; the UCT and CCT place this genus close to Streptomycetales, but with low support. Fodinicola and Motilibacter, originally assigned to Frankiales, might best be classified as genera incertae sedis until their position is clarified once more genome sequences become available.
Kineosporiales (Kämpfer, 2012b) and Micrococcales (Prévot, 1940) did not appear to be monophyletic though support to this effect is low in all the phylogenetic analyses (Figures 1–8, Supplementary File 2). The taxonomy of these orders needs to be revisited once more genome sequences become available.
Within Corynebacteriales (Goodfellow and Jones, 2012), Mycobacteriaceae (Chester, 1897; Stackebrandt et al., 1997; Zhi et al., 2009) contains the type genus Mycobacterium (Lehmann and Neumann, 1896) and Hoyosella (Jurado et al., 2009). Mycobacteriaceae appear to be paraphyletic in the phylogenomic tree (Figure 3) given the position of H. subflava (Hamada et al., 2016). Support for the monophyly of Mycobacteriaceae is strong in the 16S rRNA gene tree reduced to strains with genome sequences, but decreases when more sequences are considered; neither the UCT nor the ULT resolve these relationships (Supplementary File 2). Additional supermatrix analyses of Corynebacteriales type species confirm the non-monophyly of Mycobacteriaceae, as indicated in the GBDP tree. Tomitella (Katayama et al., 2010), currently in search of a family, was, like H. subflava, found within Nocardiaceae with high support in the phylogenomic tree (Figure 3). The proposed transfer of Hoyosella and Tomitella to Nocardiaceae is not contradicted by phenotypic features (Supplementary Table 1); for instance, mycolic acid carbon chain lengths of these genera are within the range known for the family.
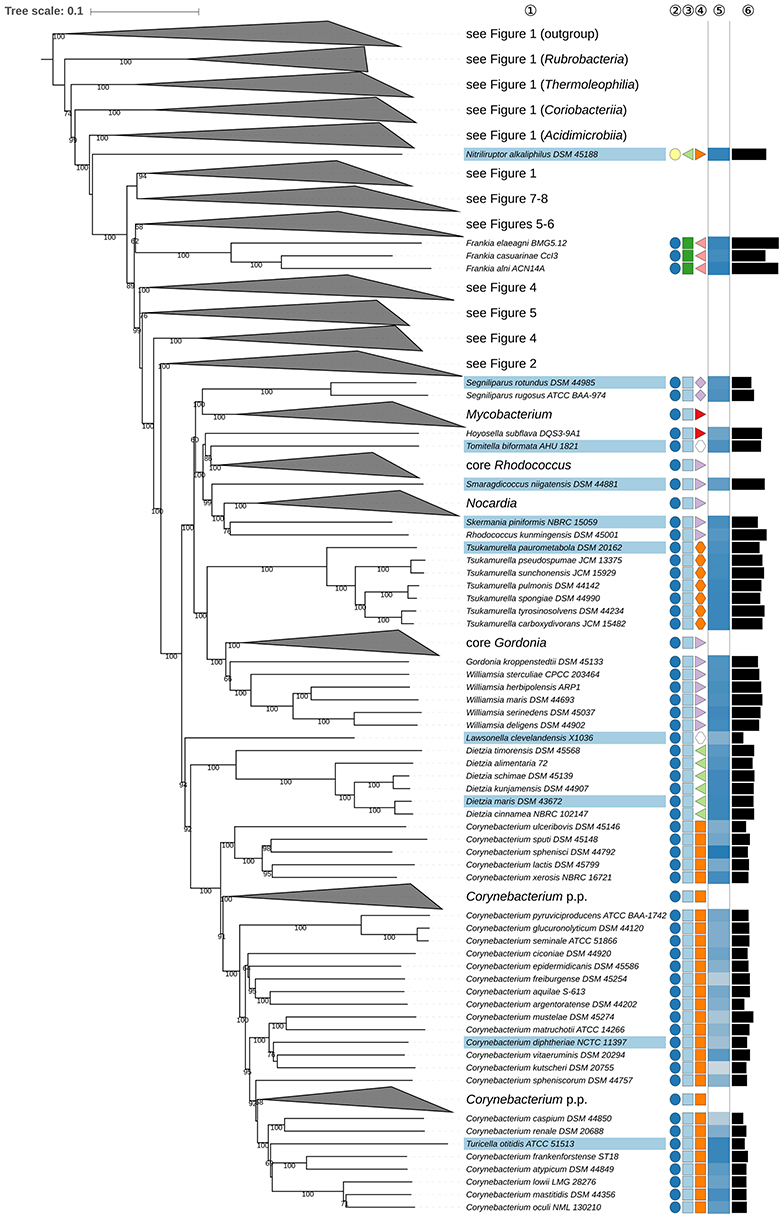
Figure 3. Third part of the phylogenomic tree inferred with GBDP. A detailed description is provided in the caption of Figure 1. The parts of the tree which have been collapsed here are shown in other figures as indicated.
The unresolved phylogenetic relationships of Gordonia (Tsukamura, 1971; Stackebrandt et al., 1988) and Williamsia (Kämpfer et al., 1999) relative to other Nocardiaceae as inferred from 16S rRNA gene sequences were noted previously (Goodfellow and Jones, 2012). In the present study, Gordonia and Williamsia form a well-supported clade together with Tsukamurella (Collins et al., 1988b) which was distinct from core Nocardiaceae (Figure 3). In the UCT and the ULT (Supplementary File 2) the backbone of the Corynebacteriales subtree was not resolved. The supermatrix analyses of the type species of Corynebacteriales confirmed the sister-group relationship of the Gordonia-Williamsia clade with Tsukamurella. A case can be made for assigning Gordonia and Williamsia to Tsukamurellaceae, but this prospect is less conservative than reassigning them to Gordoniaceae (Stackebrandt et al., 1997). Moreover, their inclusion in Nocardiaceae was mainly based on a 16S rRNA gene tree with low backbone support and on signature nucleotides but it was not clear whether these properties represented apomorphies (Zhi et al., 2009). Gordonia and Williamsia share phenotypic features with Tsukamurellaceae (Supplementary Table 1) though such properties are not necessarily apomorphic. Consequently, it is proposed that Gordonia and Williamsia be classified, as before, in Gordoniaceae (Stackebrandt et al., 1997).
The monospecific genus Lawsonella was proposed previously (Bell et al., 2016) for a clade that appeared most closely related in 16S rRNA gene trees to a clade that encompassed Corynebacterium, Dietzia, and Tsukamurella. It is evident from the GBDP tree (Figure 3) that L. clevelandensis (Bell et al., 2016) does not belong to any of the known Corynebacteriales families. It forms, with moderate support, the sister group of a clade that includes Corynebacteriaceae (Lehmann and Neumann, 1907; Stackebrandt et al., 1997; Zhi et al., 2009) and Dietziaceae (Stackebrandt et al., 1997), relationships underpinned by supermatrix analyses of Corynebacteriales type species (Supplementary File 2). Given these phylogenetic results it is proposed that Lawsonella be seen as the type genus of Lawsonellaceae fam. nov.
Propionibacteriales (Patrick and McDowell, 2012) contain Nocardioidaceae (Nesterenko et al., 1985; Stackebrandt et al., 1997; Zhi et al., 2009) and Propionibacteriaceae (Delwiche, 1957; Stackebrandt et al., 1997; Zhi et al., 2009). In addition to Nocardioides (Prauser, 1976), the type genus, Nocardioidaceae encompass genera such as Actinopolymorpha (Wang et al., 2001) Flindersiella (Kaewkla and Franco, 2011), Kribbella (Park et al., 1999; Sohn et al., 2003; Everest et al., 2013), Tenggerimyces (Sun et al., 2015), and Thermasporomyces (Yabe et al., 2011). Nocardioidaceae appear to be paraphyletic with respect to Propionibacteriaceae given the placing of Actinopolymorpha and Kribbella (Figure 5) though the branch support for the distant positions of these genera in the GBDP tree is not pronounced. However, there is a need to revise the classification of the Nocardioidaceae due to the genetic divergence it contains.
The positions of Actinopolymorpha and Kribbella are underpinned with relatively high support in the ULT (Supplementary File 2). The CCT and the UCT show Propionibacteriaceae and core Nocardioidaceae to be sister groups given the exclusion of Actinopolymorpha and Kribbella. In this context, it is interesting that Actinopolymorpha and Kribbella have much larger genomes than the clade composed of core Nocardioidaceae and Propionibacteriaceae indicating that reduced genome size is a synapomorphy in the latter (Figure 5). The heterogeneity of the Nocardioidaceae with respect to genomic and phenotypic properties has been noted (Evtushenko and Ariskina, 2012): Actinopolymorpha, Kribbella, Flindersiella, Tenggerimyces, and Thermasporomyces are non-motile and form branched hyphae whereas most of the other Nocardioidaceae form motile rods or cocci; Kribbella is the only one of these genera in which teichuronic and teichulosonic acids have been detected (Tul'skaya et al., 2011). Finally, a highly supported clade in the CCT (Supplementary File 2) includes Actinopolymorpha, Flindersiella, Tenggerimyces and Thermasporomyces, but not Kribbella. Given this wealth of genomic and phenotypic data, it is proposed that two additional families be recognized within Propionibacteriales.
The monospecific genus Thermobispora (Wang et al., 1996) appears as a distinct lineage within Streptosporangiales based on an analysis of 16S rRNA gene sequences, but without a clear association to any of the described families (Ludwig et al., 2012) hence its assignment to an order incertae sedis in the current edition of Bergey's Manual of Systematic Bacteriology (Goodfellow et al., 2012a). It is apparent from the phylogenomic tree (Figure 6) that T. bispora (Henssen, 1957; Wang et al., 1996) forms the sister group to a clade encompassing Microbispora (Nonomura and Ohara, 1957) and Sphaerimonospora (Katayama et al., 2010), together these taxa form the sister group to Microtetraspora (Thiemann et al., 1968). All three of these genera belong to Streptosporangiaceae (Goodfellow et al., 1990; Stackebrandt et al., 1997). Thermobispora can be distinguished from the current Streptosporangiaceae genera given its thermophilic nature, formation of pairs of spores on aerial hyphae and presence of MK-9(H0) as the predominant menaquinone (Wang et al., 1996). In contrast, Sphaerimonospora mesophila (Mingma et al., 2016), the type species of the genus, forms single spores on aerial hyphae and has MK-9(H4) as the predominant isoprenologue (Supplementary Table 1). Given these developments it is proposed that Thermobispora be classified in Streptosporangiaceae but kept in a genus of its own.
Sanguibacteraceae (Stackebrandt and Schumann, 2000; Zhi et al., 2009) appear paraphyletic because Jonesiaceae (Stackebrandt et al., 1997; Zhi et al., 2009), represented by Jonesia (Rocourt et al., 1987), were more closely related to Timonella (Mishra et al., 2013), and to some Sanguibacter species (Fernández-Garayzábal et al., 1995) than to other species in this genus with high support in the GBDP tree (Figure 7). The Jonesia-Timonella clade also shows lower genomic G+C content and genome size compared to the surrounding genera, a probable synapomorphy. Additional supermatrix analyses of selected species confirmed these relationships (Supplementary File 2). In the CCT the Jonesia and Timonella clade also includes Populibacterium (Li et al., 2016) from the same family and Rarobacter (Yamamoto et al., 1988; Li et al., 2016) of Rarobacteraceae (Stackebrandt and Schumann, 2000). Jonesiaceae and Sanguibacteraceae share key chemotaxonomic markers, as exemplified by the presence of L-Lysine as the cell wall diamino acid, MK-9 or MK-9(H4) as the predominant isoprenologue and anteiso-C15:0 and C16:0 as major fatty acids (Supplementary Table 1). In light of the genomic and phenotypic data, it is proposed that Sanguibacteraceae be reduced to a synonym of Jonesiaceae. It seems likely that further comparative taxonomic studies will show Rarobacteraceae, which like the other two families was based on signature nucleotides, for which it remained unclear whether they are apomorphies, and a largely unresolved 16S rRNA tree (Stackebrandt and Schumann, 2000), to belong to Jonesiaceae (Zhi et al., 2009). The presence of L-ornithine in the Rarobacter peptidoglycan does not preclude this, as its absence in the other genera might simply be the plesiomorphic state in the Jonesiaceae-Rarobacteraceae-Sanguibacteraceae clade.
Cellulomonadaceae (Stackebrandt and Prauser, 1991; Zhi et al., 2009) appear polyphyletic because Paraoerskovia (Khan et al., 2009) and Oerskovia (Prauser et al., 1970) are phylogenetically located within Promicromonosporaceae (Stackebrandt et al., 1997; Zhi et al., 2009), forming a highly supported clade together with Cellulosimicrobium (Schumann et al., 2001), whereas Tropheryma (La Scola et al., 2001) is placed in a completely different position as sister group to Microbacteriaceae (Park et al., 1993) albeit with not particularly high support (Figures 1, 7). The ULT does not resolve the position of Oerskovia and Paraoerskovia but shows the Microbacteriaceae-Tropheryma clade with maximum support (Supplementary File 2). Thus, none of the three genera appear phylogenetically closely related to the type genus Cellulomonas (Bergey et al., 1923). The transfer of Paraoerskovia (including Koreibacter, a known later heterotypic) and Oerskovia (excluding O. xanthineolytica, also a later heterotypic synonym) to Promicromonosporaceae is supported by several common phenotypic and genotypic features (Supplementary Table 1). Luteimicrobium (Hamada et al., 2010), currently in search of a family, is shown in Figure 7 as the sister group of an accordingly revised Promicromonosporaceae with strong support; hence we propose to also include it in this family. Except for a minor deviation regarding the isoprenologues their phenotypic features are in good agreement (Supplementary Table 1). The enigmatic, poorly characterized Tropheryma should, given its isolated phylogenetic position, best be transferred to it own family, currently without an assignment to an order.
Dermacoccaceae (Stackebrandt and Schumann, 2000; Zhi et al., 2009; Ruckmani et al., 2011) is not shown as monophyletic because of the position of Kytococcus (Stackebrandt et al., 1995) in the GBDP tree (Figure 8). The CCT (Supplementary File 2) shows the type genus, Dermacoccus, the genomes of which have not yet been sequenced, located, with high support, within the Branchiibius-Demetria-Luteipulveratus clade whereas Kytococcus does not form a well-supported sister group to this taxon. The ULT does not resolve the position of Kytococcus and hence does not support the monophyly of Dermacoccaceae. The probable misclassification of Kytococcus within Dermacoccaceae has been noted (Stackebrandt and Schumann, 2014) based on chemotaxonomic evidence (Supplementary Table 1) and on its position within the 16S rRNA gene tree. In light of all these results, it is proposed that Kytococcus be classified in Kytococcaceae fam. nov., within Micrococcales (Prévot, 1940).
Intrasporangiaceae (Stackebrandt et al., 1997; Zhi et al., 2009) also belong to Micrococcales, currently encompassing genera such as Intrasporangium (Kalakoutskii et al., 1967), the type genus, Arsenicicoccus (Collins et al., 2004b), Kribbia (Jung et al., 2006), Ornithinibacter (Xiao et al., 2011a), Ornithinicoccus (Groth et al., 1999a; Zhang et al., 2016b), Serinicoccus (Yi et al., 2004; Traiwan et al., 2011; Xiao et al., 2011b), and Ornithinimicrobium (Groth et al., 2001). It is evident from the GBDP tree (Figure 8) that the family is paraphyletic as Serinicoccus and Ornithinimicrobium appear to form the sister group to Kytococcus and are more distantly related to other Intrasporangiaceae; similar relationships are apparent in the ULT, albeit without strong support. The CCT (Supplementary File 2) shows that O. humiphilum (Groth et al., 2001), the type species of the genus, also belongs to the Ornithinimicrobium-Serinicoccus clade. These genera have ornithine as the diamino acid of the peptidoglycan, apart from S. chungangensis (Traiwan et al., 2011), which contains DL-A2pm whereas, in the main, Intrasporangiaceae lack ornithine (Supplementary Table 1). Consequently, it is proposed that Ornithinimicrobium and Serinicoccus be classified in Ornithinimicrobiaceae fam. nov.; it remains to be seen whether the other ornithine-containing genera, Ornithinibacter (Xiao et al., 2011a) and Ornithinicoccus (Groth et al., 1999a; Zhang et al., 2016b) belong to this family; the CCT does not settle this question.
In Figure 8 Arsenicicoccus (Collins et al., 2004b) appears to form the sister group to Dermatophilaceae (Austwick, 1958; Stackebrandt et al., 1997; Stackebrandt and Schumann, 2000; Zhi et al., 2009) (Figure 8). These taxa share a number of phenotypic properties (Supplementary Table 1), notably a facultatively anaerobic lifestyle (Jung et al., 2006). All of the other Intrasporangiacae are aerobic, apart from Kribbia which is a facultative anaerobe. The assignment of Arsenicicoccus to Intrasporangiaceae was based on chemotaxonomic data, notably the presence of LL-A2pm in the peptidoglycan, and on a poorly resolved 16S rRNA gene tree. In line with the recent evidence, it is proposed that Arsenicicoccus be classified in Dermatophilaceae. It is possible that Kribbia may be recovered in this family when genomic data are generated for this genus.
It can be seen from the phylogenomic tree (Figures 1–8) that a broad range of genera are non-monophyletic, as exemplified by Corynebacterium (Lehmann and Neumann, 1896; Bernard et al., 2010), Kocuria (Stackebrandt et al., 1995), Lechevalieria (Labeda et al., 2001), Kitasatospora (Omura et al., 1982; Wellington et al., 1992; Zhang et al., 1997), Streptomyces (Waksman and Henrici, 1943; Witt and Stackebrandt, 1990; Wellington et al., 1992), Actinomyces (Harz, 1877), Lawsonella (Bell et al., 2016), Rhodococcus (Zopf, 1891), Actinomadura (Lechevalier and Lechevalier, 1970; Zhao et al., 2015), Thermomonospora (Henssen, 1957; Zhang et al., 1998), Zimmermannella (Lin et al., 2004), and Amycolatopsis (Lechevalier et al., 1986; Lee, 2009; Tang et al., 2010a). In most of these cases, 16S rRNA gene sequence analyses (Supplementary File 2) had revealed the same taxonomic problems albeit with low support for delineated clades (Supplementary File 2).
In the GBDP tree the position of Gulosibacter (Manaia et al., 2004) makes Zimmermannella (Lin et al., 2004) appear paraphyletic (Figure 1). However, the name Zimmermannella has no standing in nomenclature as it is a later homotypic synonym of Pseudoclavibacter (Manaia et al., 2004) because its type species, Z. helvola (Lin et al., 2004) is a homotypic synonym of P. helvolus (Manaia et al., 2004) which means that Z. alba (Lin et al., 2004), Z. bifida (Lin et al., 2004), and Z. faecalis (Lin et al., 2004) and Z. helvola are also illegitimate. The UCT did not resolve the status of Zimmermannella species, but the CCT showed an unambiguously supported clade composed of G. molinativorax (Manaia et al., 2004), Z. bifida, and Z. faecalis though the placement of the other Zimmermannella species was uncertain (Supplementary File 2); the reported phenotypic differences between the species (Manaia et al., 2012) are not pronounced (Supplementary Table 1). Given these clear-cut phylogenetic relationships and the need to provide legitimate names for Zimmermannella species, it is proposed that Z. bifida and Z. faecalis be transferred to Gulosibacter. The taxonomic status of Z. alba may be resolved when genomic sequences become available for Pseudoclavibacter.
Atopobium (Collins and Wallbanks, 1992; Cools et al., 2014), the type genus of Atopobiaceae (Gupta et al., 2013) appears to be paraphyletic in the phylogenomic tree (Figure 1) as Olsenella (Dewhirst et al., 2001; Kraatz et al., 2011) forms the sister group to A. parvulum and A. rimae with high support; this relationship is also apparent in 16S rRNA gene phylogenies (Cools et al., 2014), in the latter case with insufficient resolution unless constrained (Supplementary File 2). The CCT shows a core Atopobium composed of A. minutum, the type species, A. deltae and A. fossor (the genome of A. deltae has yet to be sequenced); this clade was highly supported in the UCT. The ULT underpins the distinctness of the three Atopobium clades, but surprisingly shows Atopobium to be monophyletic and Olsenella paraphyletic. Supermatrix analyses undertaken to resolve this situation showed a different topology to that in the GBDP tree, but confirmed Atopobium as paraphyletic, whereas monophyly of Olsenella was unresolved due to the uncertain location of O. scatoligenes (Supplementary File 2). Consequently, it seems that the ULT rather than the GBDP topology is anomalous, a situation that offers two taxonomic ways of achieving monophyletic taxa, namely merging the genera Atopobium and Olsenella or splitting Atopobium. Differences in genomic G+C content and genome size (Figure 1) and oxygen requirements (Supplementary Table 1) argue against combining the two genera, as does the overall genomic divergence encompassed by the clade compared to its sister group. We thus will propose one new genus to accommodate A. vaginae and another new genus to accommodate A. parvulum and A. rimae. In contrast to the genus description, A. vaginae was described as facultatively anaerobic (Rodriguez Jovita et al., 1999). The culture condition used in that study point to microaerophilic preferences, however, while the type strain is successfully cultivated at DSMZ under anoxic conditions (DSMZ; unpublished data).
Eggerthellaceae (Gupta et al., 2013) encompass several genera, including the type genus Eggerthella (Wade et al., 1999), Adlercreutzia (Maruo et al., 2008), Asaccharobacter (Minamida et al., 2008), Enterorhabdus (Clavel et al., 2009), and Parvibacter (Clavel et al., 2013). The GBDP tree (Figure 1) shows that Adlercreutzia, Asaccharobacter, and Enterorhabdus form a clade which stands out as its genetic divergence is lower than that of adjacent clades, including closely related ones corresponding to individual genera, as exemplified by Collinsella and Slackia. In the 16S rRNA gene tree, a fourth genus, Parvibacter, was assigned to this clade with Enterorhabdus appearing to be paraphyletic, albeit with low support. All four genera have an anaerobic lifestyle and share chemotaxonomic features, notably with respect to major fatty acids and predominant menaquinones. Indeed, Adlercreutzia and Asaccharobacter, were undistinguishable at the species level with respect to their chemotaxonomic, genomic and physiological features (Table 1, Supplementary Table 1); these taxa also have similar G+C values (Figure 1). Given these similarities, it is proposed that Asaccharobacter, Enterorhabdus and Parvibacter be seen as synonyms of the earlier described Adlercreutzia.

Table 1. Outcome of applying GGDC to calculate intergenomic dDDH values.
Within Pseudonocardiaceae (Embley et al., 1988; Stackebrandt et al., 1997; Zhi et al., 2009; Labeda et al., 2011), Lentzea (Yassin et al., 1995) appears to be paraphyletic in the GBDP tree (Figure 2) due to the inclusion of Lechevalieria (Labeda et al., 2001). The CCT does not differentiate between these taxa and shows that they are closely associated with other genera, such as Actinorectispora and Actinosynnema (Supplementary File 2). It can also be seen from Figure 2 that the genetic diversity encompassed by the Lechevalieria and Lentzea clade is comparable to that shown by adjacent clades corresponding to single genera, as exemplified by Actinopolyspora and Actinosynnema. Lechevalieria and Lentzea have chemotaxonomic features in common, including DL-A2pm in the peptidoglycan, galactose, mannose, and ribose as whole cell sugars, MK-9(H4) as the predominant isoprenologue and PE (diagnostic component), DPG, PG, and PI as major polar lipids (Supplementary Table 1). In contrast, Lechevalieria is rich in saturated and mono-unsaturated iso- and anteiso- fatty acids and Lentzea in saturated iso- and anteiso-components and tuberculostearic acid though it is not clear whether these characters states are apomorphies. Lechevalieria and Lentzea have genomes of a similar size with G+C values within a narrow range (Figure 2); estimates based on experimental methods were much wider: 68.0–71.4% (Lechevalieria) and 68.6–79.6% (Lentzea). In light of these genomic and phenotypic data, it is proposed that Lechevalieria be seen as a subjective synonym of Lentzea.
Amycolatopsis (Pseudonocardiaceae) does not appear to be monophyletic (Figure 2) as A. halophila (Tang et al., 2010a) forms a clade together with Haloechinothrix and Thermocrispum (Korn-Wendisch et al., 1995) Indeed, genera such as Saccharomonospora (Nonomura and Ohara, 1971b) and Prauserella (Kim and Goodfellow, 1999) appear to be more closely related to core Amycolatopsis including A. orientalis (Lechevalier et al., 1986), the type species, than to A. halophila. In addition, Yuhushiella (Mao et al., 2011) forms the sister group to A. marina (Bian et al., 2009).
Given the genomic divergence encompassed by these genera it does not make sense to classify them into a single genus, nor would this be the most conservative solution taxonomically. In the CCT (Supplementary File 2), A. halophila (Tang et al., 2010a) and A. salitolerans (Guan et al., 2012) form a highly supported clade which forms the sister group to Haloechinothrix with high support; this clade forms the sister group of Thermocrispum (Figure 2), confirmed by the ULT. The genome sizes of A. halophila, Haloechinothrix and Thermocrispum show more similarity to one another than to core Amycolatopsis; increased genome size appears to be an apomorphy of the latter (Figure 2). Available phenotypic features of A. halophila and A. salitolerans are in good agreement with those of Haloechinothrix alba; all of these species are halophilic and filamentous with similar chemotaxonomic traits (Supplementary Table 1). Moreover, support for the inclusion of A. halophila and A. salitolerans within Amycolatopsis was uniformly low at the backbone of the respective 16S rRNA gene trees nor were these taxa compared with H. alba which was described around the same time. Given all the these results, it is proposed that A. halophila and A. salitolerans be transferred to the genus Haloechinothrix. The original description of A. halophila had made it necessary to emend the description of Amycolatopsis; these changes can now be reversed. Again, the original descriptions of A. halophila and A. salitolerans had not considered Haloechinothrix, and support was uniformly low at the backbone of the 16S rRNA gene trees.
In the original description of Yuhushiella (Mao et al., 2011) it was compared with a restricted number of Amycolatopsis species which did not include A. marina, while support for the backbone of the 16S rRNA gene tree was uniformly low. Yuhushiella has many features in common with Amycolatopsis, notably with respect to fatty-acid and polar-lipid profiles (Supplementary Table 1) and genomic G+C content (Figure 2). Consequently, it is proposed that Yuhushiella be regarded as a subjective synonym of Amycolatopsis. This solution is safer and more conservative taxonomically than further splitting Amycolatopsis, which is difficult because the genus is large and despite the strong resolution of the phylogenomic tree there were hardly any well-supported subgroups in the CCT (Supplementary File 2).
Actinokineospora (Hasegawa, 1988; Labeda et al., 2010; Tang et al., 2012) appears to be paraphyletic in the phylogenomic tree (Figure 2) given the position of Alloactinosynnema iranicum (Nikou et al., 2014). The ULT confirmed the paraphyly of Actinokineospora, albeit with moderate support (Supplementary File 2), while the CCT shows that the type species of Alloactinosynnema, A. album (Yuan et al., 2010), is the sister group to A. iranicum. The relationship between these species was not clear either in the UCT or in the original description of A. album. The chemotaxonomic characteristics of A. iranicum are more in line with those of Actinokineospora, as it contains arabinose, galactose and ribose as whole-organism sugars and PE (diagnostic component) in its polar-lipid profile whereas A. album is rich in PC and contains only arabinose as whole-organism sugar (Nikou et al., 2014). However, the value of these chemotaxonomic features in separating these genera is doubtful as the character states of A. album are most likely to be its autapomorphies; this means that the other states are plesiomorphic and hence cannot be used to separate a taxon. The CCT also shows that A. album falls within Actinokineospora. Consequently, it is proposed that Alloactinosynnema be seen as a subjective synonym of Actinokineospora.
Corynebacterium (Lehmann and Neumann, 1896; Bernard et al., 2010) appeared paraphyletic due to the position of Turicella otitidis (Funke et al., 1994a). It also appears from the CCT, UCT, and ULT (Supplementary File 2) that Corynebacterium is paraphyletic given the position of Turicella otitidis, a species found to be closely related to C. atypicum (Hall et al., 2003a) and C. frankenforstense (Wiertz et al., 2013). Both genera have DL-A2pm in the peptidoglycan and arabinose and galactose as whole-cell sugars (Supplementary Table 1), but can be distinguished based on menaquinone and fatty-acid composition. Corynebacterium has either MK-8(H2) or MK-9(H2) as predominant menaquinone whereas T. otitidis contains major amounts of MK-10 or MK-11 (Busse, 2012). Further, Corynebacterium mostly has short-chain mycolic acids and tuberculostearic acid whereas T. otitidis lacks these compounds, as do some species of Corynebacterium, as exemplified by C. atypicum, C. caspium (Collins et al., 2004a), and C. lactis (Wiertz et al., 2013). In turn, C. frankenforstense synthesizes mycolic acids but not tuberculostearic acid while the reverse is the case in C. ciconiae (Fernández-Garyzábal et al., 2004) and C. kroppenstedtii (Collins et al., 1998).
A recent study (Baek et al., 2018) revealed that genes that play an essential role in mycolic acid biosynthesis are absent in Turicella and other mycolate-less Corynebacterium species and proposed that T. otitidis be classified in the genus Corynebacterium. Moreover, menaquinone reductase was also found to be absent in Turicella. While these authors concluded from their results that chemotaxonomic features such as menaquinones and mycolic acids should be treated with caution when drawing taxonomic conclusions, they may not have taken into account the consequences of the fact that the presence of mycolic acids in Corynebacteriales (Goodfellow and Jones, 2012) is a synapomorphy of the taxa included in the order. It follows that this character state is plesiomorphic within the order and hence should not have been used to justify the separation of Corynebacterium and Turicella in the first place. The same holds for menaquinones, as the widespread distribution of MK-8(H2) and MK-9(H2) in Corynebacteriales (Supplementary Table 1) indicates that their presence is plesiomorphic within the order and in Corynebacterium, whereas their replacement by MK-10/11 is an autapomorphy of Turicella. If the separation of the two genera was incorrectly based on these characters, then one cannot conclude from the need to include Turicella in Corynebacterium that these characters are not trustworthy per se.
It appears from the GBDP tree (Figure 8) that Kocuria (Stackebrandt et al., 1995) is paraphyletic with respect to Rothia (Georg and Brown, 1967) as K. kristinae (Kloos et al., 1974) is more closely related to Rothia than to the remaining Kocuria species (Figure 8), an arrangement that carries maximum support in the ULT (Supplementary File 2) and is underpinned in the CCT. It is also apparent from the CCT that K. rosea (Flügge, 1886; Stackebrandt et al., 1995), the type species of the genus, belongs to a clade in the phylogenomic tree (Figure 8) that is composed of K. flava, K. polaris, and K. turfanensis, whereas K. halotolerans, K. koreensis, and K. kristinae form a clade that appears as the sister group to Rothia. There do not appear to be any pronounced phenotypic differences between the three Kocuria species and Rothia (Supplementary Table 1). Indeed, Kocuria also appears to be paraphyletic in a previously published 16S rRNA gene tree, albeit without much support (Collins et al., 2000b). In light of these observations, it is proposed that K. halotolerans, K. koreensis, and K. kristinae be classified in Rothia.
Streptomycetaceae (Waksman and Henrici, 1943; Stackebrandt et al., 1997; Kim et al., 2003; Zhi et al., 2009; Huang et al., 2017) contain four genera, namely Streptomyces (Waksman and Henrici, 1943; Witt and Stackebrandt, 1990; Wellington et al., 1992), the type genus, Allostreptomyces (Huang et al., 2017), Kitasatospora (Omura et al., 1982; Zhang et al., 1997), and Streptacidiphilus (Kim et al., 2003) though the status of Kitasatospora and Streptacidiphilus have been questioned (Kämpfer, 2012a). In the phylogenomic tree (Figure 6) the monophyly of Kitasatospora and Streptomyces is in conflict with the inclusion of S. aureofaciens (Duggar, 1948), S. avellaneus (Baldacci and Grein, 1966), and S. purpeofuscus (Yamaguchi and Saburi, 1955) within Kitasatospora and the insertion of Kitasatospora and Streptacidiphilus within Streptomyces, both with maximum support (Figure 6). In the 16S rRNA gene tree, the aforementioned Streptomyces species form a well-supported clade with Kitasatospora and S. aburaviensis (Nishimura et al., 1957), S. herbaricolor (Kawato and Shinobu, 1959), S. indigoferus (Shinobu and Kawato, 1960), S. xanthocidicus (Asahi et al., 1966), S. purpureus (Matsumae et al., 1968; Goodfellow et al., 1986c), S. chrysomallus (Lindenbein, 1952), S. psammoticus (Virgilio and Hengeller, 1960), and S. alboverticillatus (Witt and Stackebrandt, 1990) (Supplementary File 2); the ULT shows this group though the situation is not well-resolved. These results confirm those reported previously (Labeda et al., 2017) in a multi-locus sequence analysis of Streptomycetaceae, but additionally clarify the status of S. indigoferus and S. xanthocidicus. Consequently, it is proposed that these species be transferred to the genus Kitasatospora. However, the taxonomic status of S. alboverticillatus needs to be checked as the 16S rRNA gene sequence of this organism (AY999766), derived from strain JCM 5010T, was found within Kitasatospora in the CCT whereas S. alboverticillatus NRRL B 24281T was previously placed within Streptomyces (Labeda et al., 2017).
The assignment of Kitasatospora species, including K. cystarginea (Kusakabe and Isono, 1988), K. griseola (Takahashi et al., 1984), K. mediocidica (Labeda, 1988), K. phosalacinea (Takahashi et al., 1984), and the type species K. setae (Omura et al., 1982) to Streptomyces (Wellington et al., 1992) was based on the realization that these groups share high 16 rRNA gene sequence similarities and key chemotaxonomic and morphological properties (Supplementary Table 1). However, Kitasatospora and Streptacidiphilus were considered to differ from Streptomyces as their whole-organism hydrolysates contain DL- and LL-A2pm and galactose though these features were not available for most Streptomyces species, including those mentioned above. However, even if Kitasatospora and Streptomyces were merged, the resulting genus would still be paraphyletic as Streptacidiphilus forms the sister group to the Kitasatospora clade with high support (Figure 6). The results of the present analyses are in line with those reported previously (Labeda et al., 2017), as they support the distinctiveness of Kitasatospora and Streptacidiphilus as the considerable genetic divergence of the entire group also indicates the three genera should remain distinct; an earlier case was made for the separation of Kitasatospora from Streptomyces (Zhang et al., 1997).
It is evident from the GBDP tree (Figure 6) that Streptomyces scabrisporus (Ping et al., 2004) branches from core Streptomyces before Kitasatospora and Streptacidiphilus (Kim et al., 2003). S. scabrisporus forms the sister group to the core Streptomyces-Kitasatospora-Streptacidiphilus clade in the phylogenomic tree, and also in the CCT with high support and the ULT (Supplementary File 2). In addition, the organism can be distinguished from Kitasatospora, Streptacidiphilus, and core Streptomyces by a number of phenotypic features (Supplementary Table 1). Consequently, it is proposed that S. scabrisporus be classified as a genus in its own right.
S. thermoautotrophicus (Gadkari et al., 1990) branched even earlier than S. scabrisporus in the phylogenomic tree (Figure 6); its isolated position is underpinned in the CCT and ULT analyses with high support in the former (Supplementary File 2). This species shares chemotaxonomic and morphological features with core Streptomyces (Supplementary Table 1) but can be distinguished from the latter and from Kitasatospora and Streptacidiphilus given its obligate thermophilic nature, its inability grow below 40°C and its chemolithotrophic lifestyle; it is able to use CO2 or H2 plus CO2.. Indeed, this species is unique in its exclusive use of lithotrophic substrates. Given these observations, it is proposed that S. thermoautotrophicus merits generic status. An earlier multi-gene phylogeny (Mackellar et al., 2016) yielded the same outcome. However, the type strain of the species is currently available from a single culture collection only (DSM 100163), hence a new genus with S. thermoautotrophicus as type species cannot be proposed (Parker et al., 2015).
A highly supported clade composed of S. aomiensis (Nagai et al., 2011), S. catbensis (Sakiyama et al., 2014), and S. seranimatus (Wang et al., 2007c) was found to branch before S. scabrisporus but after S. thermoautotrophicus in the CCT. The three species share biochemical, cultural, and morphological features, notably the formation of smooth spores in straight to flexuous or looped spore chains, properties that distinguished them from their nearest phylogenetic neighbor, S. scabrisporus, which produces rugose ornamented spores in spiral chains (Ping et al., 2004; Wang et al., 2007c; Sakiyama et al., 2014). In turn, S. seranimatus can be distinguished from all other Streptomycetaceae given the presence of PC in its polar lipid profile (Supplementary Table 1). Given this information, it is proposed that S. aomiensis, S. catbensis, and S. seranimatus be assigned to a new genus. This is immediately feasible for S. aomiensis only because the type strains of the other two species are currently available from only a single culture collection (Parker et al., 2015).
The proposals for the recognition of four new genera leaves Streptomyces as a monophyletic taxon. Indeed, these developments provide a more taxonomically conservative way of addressing the paraphyly of Streptomyces than lumping Kitasatospora, Streptacidiphilus, and Streptomyces sensu lato into a single genomically diverse taxon. Moreover, the set of phenotypic character states common to the early branching Streptomyces and core Streptomyces, given their phylogenetic distribution, is probably plesiomorphic within Streptomycetaceae and hence do not justify retaining Streptomyces in its present form.
Actinomycetaceae (Buchanan, 1918; Stackebrandt et al., 1997; Zhi et al., 2009) encompass genera such as Actinomyces (Harz, 1877), the type genus, Actinobaculum (Lawson et al., 1997; Yassin et al., 2015), Actinotignum (Yassin et al., 2015), Arcanobacterium (Collins et al., 1982; Lehnen et al., 2006), Mobiluncus (Spiegel and Roberts, 1984), Trueperella (Yassin et al., 2011), and Varibaculum (Hall et al., 2003d). In the phylogenomic tree, the genomically diverse Actinomyces appears to be paraphyletic as it encompasses Mobiluncus and Varibaculum (Figure 7); the ULT shows a similar arrangement, but with lower support. The phylogenomic tree does not include Actinomyces bovis (Harz, 1877), the type species of the genus, as a genomic sequence of its type strain was not available at the time of analyzing the data. It is apparent from the CCT (Supplementary File 2) that A. bovis falls within the clade that encompasses most of the Actinomyces species; this clade in the phylogenomic tree ranges from A. slackii to A. massiliensis (Figure 7). This core Actinomyces clade has larger genomes and a higher genomic G+C content than is found in other Actinomyces species, apart for A. hordeovulneris and A. nasicola. The CCT also shows that the balance of Actinomyces species can be assigned to an additional seven distinct, maximally supported or monotypic clades (Supplementary File 2). A. neuii formed the sister group to Varibaculum cambriense, the type species of the genus, and V. anthropi with moderate support; the ULT showed strong support for the relationship between A. neui and V. cambriense; V. anthropi was not included in this analysis. The sister-group relationship between this latter group and Mobiluncus, including M. curtisii (Spiegel and Roberts, 1984), the type species of the genus, and its heterotypic synonym Falcivibrio vaginalis (Hammann et al., 1984) was unambiguous in the GBDP tree (Figure 7). The remaining six Actinomyces clades occupied isolated positions in the CCT (Supplementary File 2). The overall proteomic differences between several pairs of Actinomyces species, e.g., A. hordeovulneris and A. nasicola, are greater than those between Arcanobacterium and Trueperella and between Actinobaculum and Actinotignum. These results illustrate the inconsistencies of the current classification of Actinomycetaceae in quantitative terms.
In general, the results outlined above are in good agreement with those of previous studies which showed Actinomyces to be a heterogeneous taxon containing distinct subgroups (Schofield and Schaal, 1981; Schaal and Schofield, 1984; Schaal and Gatzer, 1985; Lawson et al., 1997; Ramos et al., 1997; Schaal et al., 2006) though two main groups were recognized based on 16S rRNA gene signature nucleotides (Schaal et al., 2006). However, in a recent phylogenomic analysis (Zhao et al., 2014) Actinomyces species were recovered in major groups A, B, and C, which correspond to three of the clades in the CCT (Supplementary File 2). Indeed, the taxonomic status of some of the clades defined in the CCT are supported by the discontinuous distribution of chemotaxonomic traits (Supplementary Table 1). Thus, the clade represented by A. meyeri to A. radingae in Figure 7 contains mannose and rhamnose as whole organism sugars, MK-9(H4) as the predominant menaquinone and DPG and PG as major polar lipids whereas core Actinomyces species are characterized by the presence of 6 deoxytalose, fucose, galactose, and glucose in whole organism hydrolysates, MK-10 as the predominant manaquinone and a polar lipid pattern that features PC. In turn, A. neuii subsp. anitratus (Funke et al., 1994b), A. neuii subsp. neuii (Funke et al., 1994b), and Varibaculum cambriense (Hall et al., 2003d), the type species of the genus, have an A5α (L-lysine-L-lysine-D-glutmic acid) peptidoglycan and a fatty-acid profile with major proportions of oleic, palmitic and stearic acids. In contrast, little chemotaxonomic data are currently available for several Actinomyces species, notably A. hongkongensis, A. marimammalium, and A. nasicola. The lack of chemotaxonomic features specific to the other clades might simply be due to their combination of features representing the plesiomorphic states. Given the phylogenomic and associated data outlined here, we propose to split Actinomyces according to the well-supported or monotypic, genomically well-differentiated clades mentioned above, yielding seven novel genera.
Leifsonia (Evtushenko et al., 2000a) appears to be polyphyletic in the GBDP tree (Figure 1) as L. rubra (Reddy et al., 2003) is placed distantly from L. xyli subsp. cynodontis (Evtushenko et al., 2000a), as is the case with the ULT and the CCT. It is also apparent, from the CCT that L. aquatica (Evtushenko et al., 2000a), the type species of Leifsonia, forms a clade with L. xyli subsp. cynodontis while a well supported second clade includes L. rubra and the type species of Rhodoglobus, R. vestalii (Sheridan et al., 2003). The 16S rRNA gene similarity between the L. rubra sequence AJ438585 and the R. vestalii sequence AJ459101 was 99.44% when calculated using the recommended settings (Meier-Kolthoff et al., 2013b), indicating that DNA:DNA relatedness must determine whether they should or should not remain as separate species. Because genome sequence data for R. vestalii were not available at the time of writing, we here refrain from considering the taxonomic consequences for L. rubra. Traditional DDH values between L. rubra DSM 21193 and R. vestalii DSM 21947 were determined as 73.8–75.0% (DSMZ, unpublished data) but while DSM 21193 is a replacement of the incorrect type-strain deposit DSM 15304 it is not included in the DSMZ online catalog because it was not clear whether it shows the properties indicated in the species description.
The mycolic-acid containing genera Nocardia (Trevisan, 1889), Skermania (Chun et al., 1997a), and Smaragdicoccus (Adachi et al., 2007) formed a maximally supported clade in the GBDP tree (Figure 3), which also included Rhodococcus kunmingensis (Wang et al., 2008). R. kunmingensis was also set apart from core Rhodococcus (Zopf, 1891) in the CCT and ULT (Supplementary File 2). However, in both the UCT and original publication R. kunmingensis was found to be closely related to R. equi (Goodfellow and Alderson, 1977), which is now included in R. hoagii (Kämpfer et al., 2014a); in the original 16S rRNA gene tree they formed a clade supported by a 92% bootstrap value using the neighbor-joining algorithm and the Kimura-2-parameter model whereas in the corresponding ML and MP analyses there was only low support for this association. Additional supermatrix analyses confirmed the distant position of R. kunmingensis relative to core Rhodococcus, particularly with the ML criterion (Supplementary File 2). The relatively isolated position of R. kunmingensis was noted in an extensive phylogenomic analysis of Rhodococcus (Sangal et al., 2016). The genome sequence of R. kunmingensis in the present study was incomplete but did not show any sign of contamination; the 5S, 16S, and 23S rRNA genes were located on the same contig (data not shown). Consequently, confidence can be placed in its position in the phylogenomic tree. There is only low support for the sister-group relationship between R. kunmingensis and Skermania piniformis in the GBDP tree (Figure 3) which also shows them separated by long branches. S. piniformis and R. kunmingensis contain DL-A2pm, arabinose, and galactose in the peptidoglycan and have DPG and PE as major polar lipids, but can be distinguished by marked differences in fatty acid, menaquinone, mycolic acid composition, and in genomic G+C content (Figure 3, Supplementary Table 1). In short, the various datasets show that R. kunmingensis merits recognition as a new genus within Nocardiaceae.
Gordonia appears to be paraphyletic in the GBDP tree (Figure 3) as G. kroppenstedtii (Kim et al., 2009) forms the sister group of Williamsia (Kämpfer et al., 1999) though support for this is low in both the GBDP tree and the CCT. An additional supermatrix analysis using Tsukamurella as outgroup shows, with strong support, that G. kroppenstedtii branches below the base of the core Gordonia and Williamsia clade thereby rendering Gordonia paraphyletic (Supplementary File 2). In the original description, G. kroppenstedtii appeared as the sister group of the other Gordonia species though bootstrap support was low and not all Williamsia species were represented while in a recent 16S rRNA gene analysis G. kroppenstedtii was recovered outside core Gordonia (Tsang et al., 2016). Gordonia and Williamsia have very similar phenotypic profiles (Supplementary Table 1) and there is no evidence that character states should be regarded as apomorphies of one or other of these taxa. The polar lipid profile, for instance, of Gordonia matches that of Tsukamurellaceae hence according to these features Gordonia may well be paraphyletic, as found in this study. Our proposal to assign G. kroppenstedtii to a new genus does not conflict with the phenotypic data and is taxonomically more conservative than merging Gordonia and Williamsia, a move that would run counter to their overall genetic divergence (Figure 3).
It has recently been suggested to split Mycobacterium into five distinct genera (Gupta et al., 2018). That study deserves credit for identifying molecular synapomorphies for each of these five clades, but since such synapomorphies were also identified for a monophyletic Mycobacterium as currently circumscribed, the decision to split the genus may be regarded as somehow arbitrary. In our tree (Figure 3, Supplementary File 2) Mycobacterium also appears to be monophyletic and not as the genomically most divergent genus within Corynebacteriales (Figure 3, Supplementary File 2). It remains to be seen which of the two concepts for Mycobacterium will be adopted by future taxonomic studies.
Micromonospora (Ørskov, 1923), the type genus of Micromonosporaceae (Krasil'nikov, 1938; Zhi et al., 2009), appears to be paraphyletic in the GBDP tree (Figure 4) as the clade composed of Verrucosispora and Xiangella appears to be the sister group to M. nigra (Weinstein et al., 1968; Kasai et al., 2000), while Verrucosispora (Rheims et al., 1998; Xi et al., 2012) appears to be paraphyletic given the position of Xiangella phaseoli (Wang et al., 2013a), the type species of the genus. Further, Salinispora (Maldonado et al., 2005) would seen to be more closely related to core Micromonospora than to M. pattaloongensis (Thawai et al., 2008). The ULT is in agreement with these relationships, albeit with lower support. Although the genome sequence of M. chalcea (Ørskov, 1923), the type species of the genus, was not available at the time of running the phylogenomic analysis, this organism was found in the CCT, with high support, in the M. aurantiaca-marina-tulbaghiae clade (Supplementary File 2). It is difficult to distinguish between the genera Micromonospora, Salinispora, Verrucosispora, and Xiangella as they have many phenotypic features in common (Supplementary Table 1), notably chemotaxonomic and morphological properties, as exemplified by their ability to form single non-motile spores on branched substrate mycelia and the typical absence of aerial hyphae (Supplementary Table 1). The original description of Verrucosispora was based on an ability to synthesize 10-methyl C17:0 and the absence of arabinose in whole-cell hydrolysates, but M. citrea (Kroppenstedt et al., 2005) and M. echinaurantiaca (Kroppenstedt et al., 2005) exhibit these features while M. endolithica (Hirsch et al., 2004), M. inositola (Kawamoto et al., 1974), and M. viridifaciens (Kroppenstedt et al., 2005) contain moderate proportions of 10-methyl C17:0 (Kroppenstedt et al., 2005) and M. cremea (Carro et al., 2012) and M. mirobrigensis (Trujillo et al., 2005) do not have arabinose in the cell wall. One of the key arguments for proposing the genus Xiangella was the presence of PC in the polar-lipid profile of X. phaseoli, however, the original polar lipid profile is not fully conclusive in this respect (Wang et al., 2013a) (Supplementary Table 1). In addition, single characters with two states are, even in the absence of homoplasies, insufficient to separate two taxa, as only one of the states can be apomorphic. It can be concluded from what is said above that there is insufficient evidence from both the phenotype and the single-gene phylogenies to separate Verrucosispora and Xiangella from Micromonospora, an outcome supported by 16S rRNA gene sequence evidence (Supplementary File 2). Consequently, it is proposed that Verrucosispora and Xiangella be included in Micromonospora.
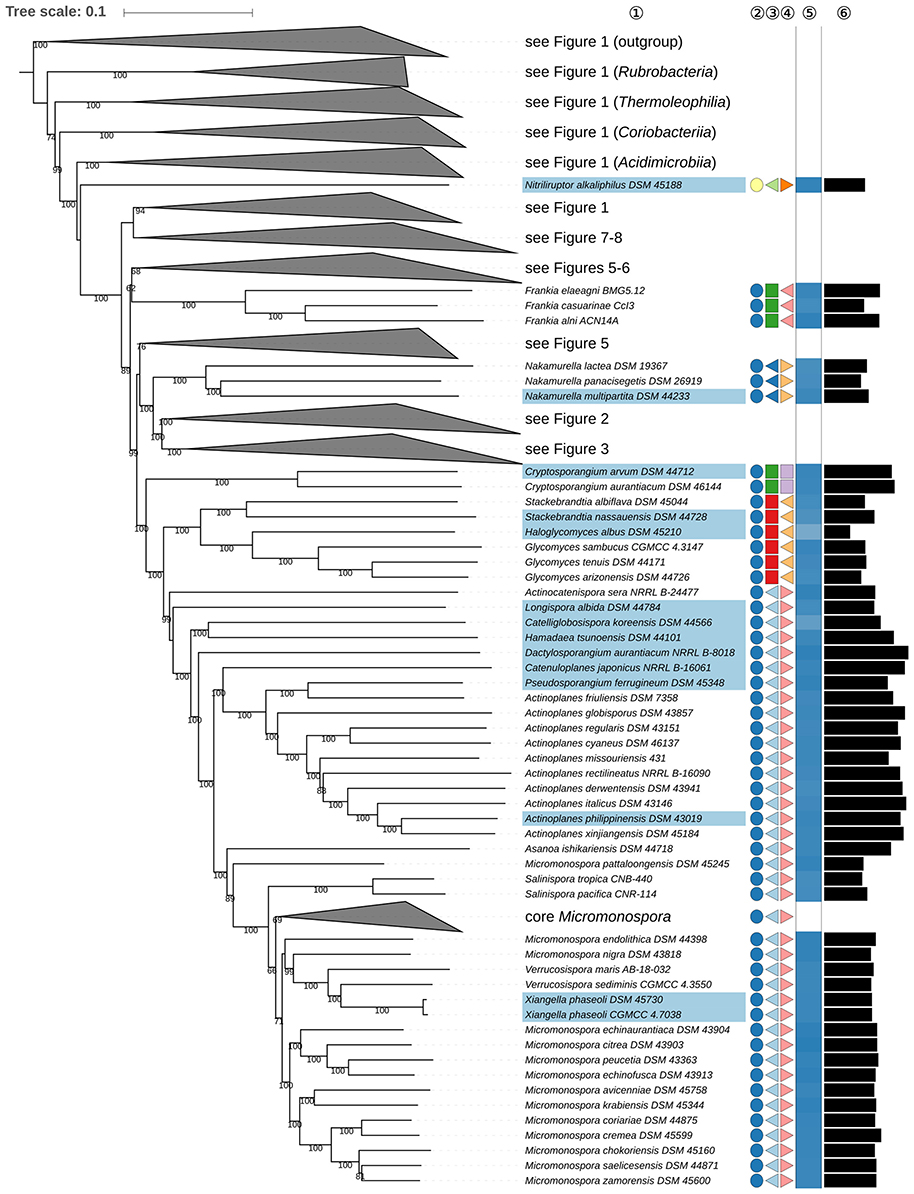
Figure 4. Fourth part of the phylogenomic tree inferred with GBDP. A detailed description is provided in the caption of Figure 1. The parts of the tree which have been collapsed here are shown in other figures as indicated.
Whereas an earlier study (Girard et al., 2013) suggested that Micromonospora, Salinispora, and Verrucosispora might represent different clades in the same genus based on an analysis of conserved protein sequences, Micromonospora and Salinispora were quite sharply separated, albeit closely related, in a genome-based phylogeny of Micromonosporaceae (Carro et al., 2018), although M. pattaloongensis was not considered in this study. The GBDP tree suggests that this species should better be removed from Micromonospora (Figure 4) though, the UCT and CCT were not clear on this point (Supplementary File 2) hence additional Micromonosporaceae genome sequences need to be examined to determine a more precise relationship between these taxa.
Actinoplanes (Couch, 1950) appears to be paraphyletic in the GBDP tree (Figure 4) as Pseudosporangium ferrugineum (Ara et al., 2008b) forms the sister group to A. friuliensis (Wink et al., 2014), a relationship underpinned in the UCT, ULT, and CCT (Supplementary File 2), but with low support. In general, P. ferrugineum has many phenotypic features in common with Actinoplanes (Supplementary Table 1); it can be distinguished from the latter based on menaquinone and whole-cell sugar composition but it remains unclear whether these character states yield apomorphies for both groups. Moreover, menaquinone composition can vary according to age and culture conditions (Saddler et al., 1986). Genome size and G+C content are consistent within the Actinoplanes-Pseudosporangium clade. Consequently, it is proposed that P. ferrugineum be transferred to Actinoplanes. The CCT indicated that Couchioplanes (Tamura et al., 1994) and Krasilnikovia (Ara and Kudo, 2007) form the sister group to Pseudosporangium but with low support only, which suggests that they may also be seen as synonyms of Actinoplanes though further comparative taxonomic studies are needed to address this point.
Microlunatus (Nakamura et al., 1995) and Friedmanniella (Schumann et al., 1997) belong to Propionibacteriaceae (Delwiche, 1957). In the original description of Friedmanniella, F. antarctica, the type species (Schumann et al., 1997), shared chemotaxonomic properties with its counterpart in the genus Microlunatus, M. phosphovorus (Nakamura et al., 1995), but was distinguished from the latter based on growth and phenotypic properties though it was not clear which character states were apomorphic. Moreover, several of the differential properties shown by F. antarctica were found to be variable when additional species were added to Friedmanniella (Iwai et al., 2010). It is evident from the GBDP tree (Figure 5) that Microlunatus is paraphyletic as it includes Friedmanniella; it is also clear that the genetic diversity shown within the clade is low. Although a genomic sequence is not available for F. antarctica it is evident from the CCT (Supplementary File 2) that it forms a well-supported clade with F. flava and F. sagamiharensis. Consequently, it is proposed that Friedmanniella be seen as a subjective synonym of Microlunatus.

Figure 5. Fifth part of the phylogenomic tree inferred with GBDP. A detailed description is provided in the caption of Figure 1. The parts of the tree which have been collapsed here are shown in other figures as indicated.
Propionibacterium (Orla-Jensen, 1909; Charfreitag et al., 1988) of Propionibacteriaceae (Delwiche, 1957; Zhi et al., 2009) was recently split by generating three novel genera, Acidipropionibacterium, Cutibacterium, and Pseudopropionibacterium, based on the analysis of genomic data (Scholz and Kilian, 2016). In the present study, Propionibacterium namnetense (Aubin et al., 2016), which was described at around the same time as the revision of Propionibacterium, was found to form the sister group to C. acnes in the phylogenomic tree (Figure 5), a position in line with phenotypic data (Supplementary Table 1). Consequently, it is proposed that P. namnetense be classified in Cutibacterium. P. acnes subsp. defendens (McDowell et al., 2016) also needs to be transferred to Cutibacterium.
Thermomonospora (Henssen, 1957; Zhang et al., 1998), the type genus of Thermomonosporaceae (Stackebrandt et al., 1997; Zhang et al., 2001; Zhi et al., 2009), appears to be paraphyletic in the GBDP tree (Figure 6) as T. curvata (Henssen, 1957), the type species, is phylogenetically located in the Actinomadura-Spirillospora clade. In turn, T. chromogena (Krasil'nikov and Agre, 1965; McCarthy and Cross, 1984) is phylogenetically placed within Streptosporangiaceae. While the problematic positioning of T. chromogena was recently fixed by taxonomically assigning the species to the genus Thermostaphylospora (Wu et al., 2018), the classification of Thermomonospora remained unsatisfactory.
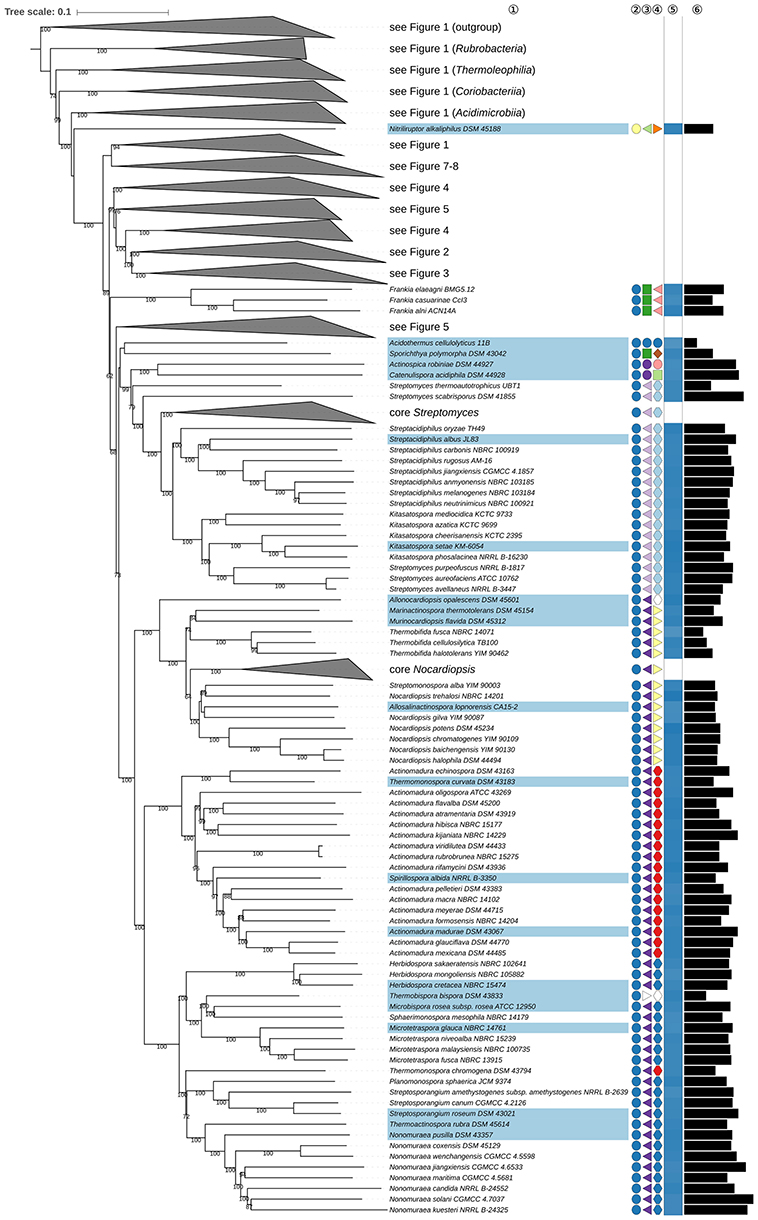
Figure 6. Sixth part of the phylogenomic tree inferred with GBDP. A detailed description is provided in the caption of Figure 1. The parts of the tree which have been collapsed here are shown in other figures as indicated.
Indeed, Actinomadura (Lechevalier and Lechevalier, 1970) also appears to be paraphyletic in the phylogenomic tree (Figure 6) as T. curvata formed a well-supported clade with A. echinospora (Nonomura and Ohara, 1971b; Kroppenstedt et al., 1990). A. amylolytica (Jiao et al., 2015), A. cellulosilytica (Jiao et al., 2015), and A. umbrina (Galatenko et al., 1981) were also placed in this clade in the CCT (Supplementary File 2). An earlier study based on 16S and 23S rRNA gene sequences showed that A. echinospora and T. curvata were assigned to a clade that was clearly separate from other Actinomadura species (Trujillo and Goodfellow, 2012). Much like T. curvata the four Actinomadura species are thermophilic and also have chemotaxonomic and other phenotypic features in common; their genomic G+C content is also similar (Supplementary Table 1). Taking all of these data into account it is proposed that the four species be classified in an amended genus Thermomonospora.
Despite the changes mentioned above Actinomadura still appears to be paraphyletic in the GBDP tree (Figure 6) given the strong support for the inclusion of Spirillospora albida, the type species of the genus (Couch, 1963), among the Actinomadura species, including the type species, A. madurae (Vicent, 1894; Kroppenstedt et al., 1990). This situation is compounded by the uncertain position of some Actinomadura species, notably A. alba (Wang et al., 2007b) and A. scrupuli (Lee and Lee, 2010a) in the CCT (Supplementary File 2). This situation is complicated even further as in the CCT Actinocorallia (Iinuma et al., 1994) is found within the Actinomadura clade. Comparative studies based on additional genome sequences are needed before the taxonomic issues touched upon here can be addressed though it should be noted that the name Spirillospora has priority over Actinomadura.
The type genus Nocardiopsis (Brocq-Rousseau, 1904; Meyer, 1976) appears paraphyletic because two other Nocardiopsaceae genera (Rainey et al., 1996), Allosalinactinospora (Guo et al., 2015), and Streptomonospora (Cui et al., 2001; Li et al., 2003d), are phylogenetically located within Nocardiopsis (Figure 6). This causes core Nocardiopsis, including the type species N. dassonvillei (Brocq-Rousseau, 1904; Meyer, 1976), to be located separately from a clade containing N. halophila, N. baichengensis, N. chromatogenes, and N. potens, a second clade consisting of N. gilva and a third one containing N. trehalosi. Except for core Nocardiopsis at the backbone of the Nocardiopsaceae subtree support was low in the CCT and UCT. This situation is complicated even further as additional genera appear to be intermixed with the deviating Nocardiopsis species, they include Haloactinospora (Tang et al., 2008), Lipingzhangella (Zhang et al., 2016a), and Salinactinospora (Chang et al., 2012). The complex taxonomic relationships outlined above need to be tackled but it is not practical to do so until additional Nocardiopsaceae genome sequences become available.
The type genus of Sanguibacteraceae (Stackebrandt and Schumann, 2000; Zhi et al., 2009) is Sanguibacter (Fernández-Garayzábal et al., 1995; Pikuta et al., 2017). This genus appears paraphyletic in the phylogenomic tree (Figure 7) as Jonesia (Rocourt et al., 1987) and Timonella (Mishra et al., 2013) were found, with maximal support, to be the sister group to a clade composed of the majority of Sanguibacter species while S. marinus (Huang et al., 2005; Pikuta et al., 2017) and S. soli (Kim et al., 2008; Pikuta et al., 2017) formed a distinct clade. A similar arrangement could be observed in the CCT (Supplementary File 2). In the UCT and the ULT maximum-likelihood tree the monophyly of Sanguibacter was unresolved, much like in the published 16S rRNA gene trees (Kim et al., 2008). In contrast, in the ULT maximum-parsimony analysis Sanguibacter was found to be monophyletic, but additional supermatrix analyses underlined the topology found in the GBDP tree (Supplementary File 2). The fact that Jonesia and Timonella have lower genomic G+C contents than Sanguibacter (Figure 7) argues against merging these taxa into a single genus, whereas S. marinus and S. soli differ in genome size from the other Sanguibacter species. There is also evidence that other genera, such as Rarobacter (Yamamoto et al., 1988), might be more closely related to some of the groups in the Sanguibacter-Jonesia-Timonella clade so the monophyly or otherwise of this assemblage cannot be established at present. Hence it is proposed that S. marinus and S. soli be assigned to a new genus.
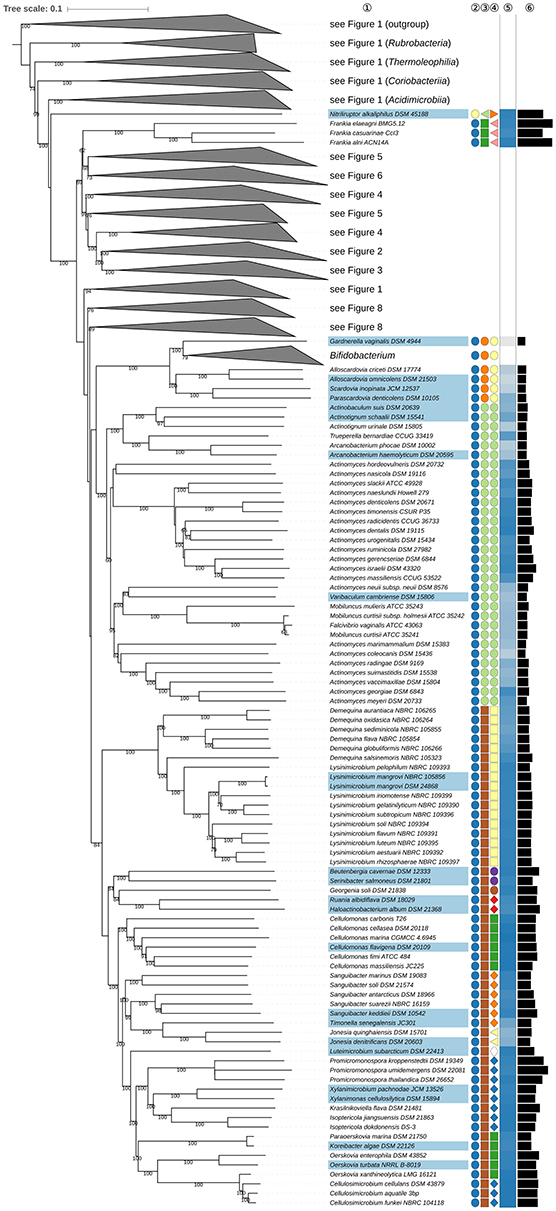
Figure 7. Seventh part of the phylogenomic tree inferred with GBDP. A detailed description is provided in the caption of Figure 1. The parts of the tree which have been collapsed here are shown in other figures as indicated.
The position of Demequina salsinemoris (Matsumoto et al., 2010) in the phylogenomic tree makes Demequina (Yi et al., 2007; Ue et al., 2011) paraphyletic due to the inclusion of Lysinimicrobium (Hamada et al., 2012, 2015); this relationship was underscored in the ULT albeit with low support (Supplementary File 2). Demequina and Lysinimicrobium have similar chemotaxonomic profiles, notably the presence of an unusual isoprenoid quinone, demethylmenaquinone [DMK(H4)], but differ in G+C content and oxygen preference (Supplementary Table 1). However, the genomic G+C content of D. salsinemoris is near to those of Lysinimicrobium species (Fig. 7) while D. lutea (Ue et al., 2011) is facultatively anaerobic like Lysinimicrobium; Demequina has L-lysine as the diamino acid in the peptidoglycan whereas Lysinimicrobium mostly has L-ornithine, though D. globuliformis contains both L-lysine and L-ornithine (Ue et al., 2011). Moreover, it has already been pointed out that single characters with two characters states do not provide sufficient evidence for separating two taxa according to the principles of phylogenetic systematics. Given all of this information it is proposed that Lysinimicrobium be considered as a subjective synonym of the earlier described taxon Demequina.
The Intrasporangiaceae genera Phycicoccus (Lee, 2006c; Zhang et al., 2011) and Tetrasphaera (Maszenan et al., 2000; Ishikawa and Yokota, 2006) are non-monophyletic in the GBDP tree (Figure 8) as P. jejuensis, the type species of Phycicoccus, is not only placed far from P. cremeus and P. dokdonensis but also forms a well-supported clade with T. duodecadis. In turn, T. japonica, the type species of Tetrasphaera (Maszenan et al., 2000) was separated from at least three of the other Tetrasphaera species. These relationships were also found in the CCT in which P. jejuensis formed a clade with P. endophyticus (Liu et al., 2016). The positioning of P. jejuensis differs significantly between the published 16S rRNA gene trees (Azman et al., 2016), which show a monophyletic Phycicoccus with strong support, and the GBDP tree. Additional supermatrix analyses clearly confirmed the sister-group relationship between P. jejuensis and T. duodecadis as well as between T. elongata and this clade. The ULT showed the same topology but with moderate support only. The genome sequence of P. jejuensis used was not complete but did not show any sign of contamination; moreover, the 5S, 16S, and 23S rRNA genes were located on the same contig (data not shown). It can be concluded, therefore, that Tetrasphaera is intermixed not only with Phycicoccus due to the position of T. duodecadis but also with Knoellia (Groth et al., 2002) due to the position of T. remsis. Although Knoellia, Phycicoccus, and Tetrasphaera display quite similar chemotaxonomic and morphological characteristics (Supplementary Table 1) it is too early to consider merging them into a single taxon, partly because the monophyly of the entire group remains unresolved in the GBDP tree but also because it is evident from the CCT that further Intrasporangiaceae genera, such as Lapillicoccus (Lee and Lee, 2007) and Oryzihumus (Kageyama et al., 2005), might also be intermixed. Despite these reservations it is timely to clarify some of the relationships found between and within Knoellia, Phycicoccus, and Tetrasphaera based on phylogenetic and associated phenotypic data in order to achieve better supported taxa. Toward this end, it is proposed that P. cremeus and P. dokdonensis as well as the other Phycicoccus species that unambiguously formed a clade in the CCT together with these two species (Supplementary File 2) be assigned to a new genus. It is also proposed that T. remsis be transferred to Knoellia and that T. duodecadis and T. elongata be reclassified in Phytococcus.

Figure 8. Eighth part of the phylogenomic tree inferred with GBDP. A detailed description is provided in the caption of Figure 1. The parts of the tree which have been collapsed here are shown in other figures as indicated.
It is evident from the phylogenetic analyses that the classification of Monashia flava (Azman et al., 2016), the type and only species of the genus, needs to be revised. In the phylogenomic tree (Figure 8), it was placed within a paraphyletic Intrasporangium (Figure 8), a relationship also found in the ULT, but with lower support (Supplementary File 2). In both the UCT and original 16S rRNA gene tree the position of M. flava was unresolved (Azman et al., 2016). Whereas, Monashia could well form the sister group of the three Intrasporangium species according to the 16S rRNA gene tree, a significant conflict with the other data sets is observed regarding the sister group of I. oryzae, which is either M. flava or I. calvum. Additional supermatrix analyses unambiguously confirmed M. flava to be most closely related to I. oryzae within the genus Intrasporangium (Supplementary File 2). The genome sequence of Monashia flava, while incomplete, showed no signs of contamination; additionally, the 5S, 16S, and 23S rRNA genes were located on the same contig (data not shown). Common chemotaxonomic features found between Monashia and Intrasporangium include a peptidoglycan containing LL-A2pm as the diagnostic diamino acid, an A3γ peptidoglycan type, major amounts of iso- and anteiso-fatty acids and MK-8(H4) as the predominant isoprenologue (Supplementary Table 1). In light of these phenotypic and phylogenetic data it is proposed that M. flava be assigned to Intrasporangium.
A recent study (Busse, 2016) led to an extensive revision of the classification of Arthrobacter (Conn and Dimmick, 1947; Koch et al., 1995) based primarily on chemotaxonomic traits though it remained unclear which ones where apomorphic character states. This strategy resulted in proposals for five new genera, namely Glutamicibacter, Paenarthrobacter, Paeniglutamicibacter, Pseudarthrobacter, and Pseudoglutamicibacter. However, it is apparent from the phylogenomic tree (Figure 8) that there are problems with this revised classification as the genera Arthrobacter and Pseudoarthrobacter do not form monophyletic groups. Further, the CCT does not yield strong support for the revised genera, including Paenarthrobacter, though it does not strongly suggest alternative groupings (Supplementary File 2). Additional genome sequences are needed to help chart a way forward.
Known and confirmed heterotypic synonyms and dDDH values found to be higher or lower than expected given current species and subspecies thresholds of 70% (Wayne et al., 1987) and 79%, respectively (Meier-Kolthoff et al., 2014b) are shown in Table 1 for pairs of closely related strains. Only Amycolatopsis keratiniphila subsp. keratiniphila and A. keratiniphila subsp. nogabecina (Wink et al., 2003) displayed a value above the 79% threshold, namely 90.3%, hence it is proposed that the latter be recognized as a heterotypic synonym of the former; an amended description of A. keratiniphila (Al-Mussallam et al., 2003) is given. Conversely, several subspecies were shown to merit species status, they included Bifidobacterium animalis subsp. lactis (Masco et al., 2004), B. longum subsp. infantis, (Mattareli et al., 2008), B. pseudolongum subsp. globosum (Yaeshima et al., 1992), and Nocardiopsis dassonvillei subsp. albirubida (Evtushenko et al., 2000b). It is proposed that in all of these cases the originally species epithets should be reintroduced, namely B. lactis (Meile et al., 1997), B. infantis (Reuter, 1963), B. globosum (Biavati et al., 1982), and N. alborubida (Grund and Kroppenstedt, 1990). Other subspecies that require species status include B. thermacidophilum subsp. porcinum (Zhu et al., 2003), Leucobacter musarum subsp. japonicus (Clark and Hodgkin, 2015), and all three subspecies of Clavibacter michiganensis (Davis et al., 1984) in accordance with recent results (Li et al., 2017).
It was not possibly to distinguish between some pairs of validly named species even at the rank of subspecies since they showed a dDDH value >79% (Meier-Kolthoff et al., 2014b). These included Cellulosimicrobium aquatile (Sultanpuram et al., 2015) vs. C. funkei (Brown et al., 2006); Dietzia cinnamea (Yassin et al., 2006) vs. D. maris (Rainey et al., 1995b); and several Bifidobacterium, Nocardiopsis, and Streptomyces species. In the case of Streptomyces the results included synonyms well-known from the literature (Lanoot et al., 2005; Labeda et al., 2017). Similarly, Saccharopolyspora jiangxiensis (Zhang et al., 2009a) was shown to belong to the same species as S. hirsuta subsp. kobensis (Iwasaki et al., 1979; Lacey, 1989). Although a genome sequence is not currently available for S. hirsuta subsp. hirsuta (Lacey and Goodfellow, 1975) the two subspecies share a 16S rRNA gene sequence similarity of 98.23% (sequences DQ381814 and EU267029, both from LTP) indicating that they merit species status (Meier-Kolthoff et al., 2013b). Moreover, the two subspecies were placed apart in the CCT (Supplementary File 2) where S. hirsuta subsp. hirsuta grouped together with S. hordei (Goodfellow et al., 1989). In light of these findings it is proposed that S. hirsuta subsp. kobensis be elevated to species level and S. jiangxiensis be seen as its heterotypic synonym.
The dDDH results also showed that several species contain strains that need to be assigned to subspecies because they displayed values between 70 and 79% (Wayne et al., 1987; Meier-Kolthoff et al., 2014b). Consequently, it is proposed that Bifidobacterium gallinarum (Watabe et al., 1983), B. kashiwanohense (Morita et al., 2011), and B. saeculare (Biavati et al., 1991) be classified as B. pullorum subsp. saeculare comb. nov., B. pullorum subsp. gallinarum comb. nov., and B. catenulatum subsp. kashiwanohense comb. nov. Actinopolyspora iraqiensis (Ruan et al., 1994) was considered to be a heterotypic synonym of Saccharomonospora halophila (Al-Zarban et al., 2002b) on the basis of experimental DDH results (Tang et al., 2011). However, using dDDH, it was found to be more closely related to Saccharomonospora paurometabolica (Li et al., 2003e), a relationship confirmed in the GBDP tree (Figure 2). Since A. iraqiensis has priority over S. paurometabolica, it is proposed that the former should be reclassified as S. iraqiensis comb. nov., with constituent strains recognized as S. iraqiensis subsp. iraqiensis comb. nov. and S. iraqiensis subsp. paurometabolica comb. nov. The results regarding Oerskovia xanthineolytica (Lechevalier, 1972) were unexpected because it is considered to be a heterotypic synonym of C. cellulans not of C. aquatile (Schumann et al., 2001) and because the GBDP tree indicated a distinct arrangement (Figure 7). In contrast to the other genomes investigated, the contigs of the genome sequence of O. xanthineolytica showed a somewhat heterogeneous G+C content distribution (data not shown) hence it cannot be ruled out that the sequence used was impure. Consequently, the relationships between O. xanthineolytica to related taxa needs to be revisited.
Finally, in the present study, all of the pairs of strains considered to represent distinct deposits of the same type strain were found to have d dDDH similarities of 99.0% or above with the exception of Amycolatopsis methanolica (De Boer et al., 1990) strain 239 vs. DSM 44096 (98.3%), Bifidobacterium longum subsp. longum (Mattareli et al., 2008) strain DSM 20219 vs. JCM 1217 (89.2%) and Pseudarthrobacter chlorophenolicus (Busse, 2016) strain A6 vs. DSM 12829 (97.3%), results which may account for the separation of each pair of these strains.
Genome size and G+C content appear to be phylogenetically conserved (p = 0.0001), a result that is in good agreement with those shown in Figures 1–8 where many of the clades encompassed strains that had similar genome sizes and G+C contents. In addition, genome size and G+C content were positively correlated (Pearson, 0.612, p < 10−15; Kendall, 0.339, p < 10−15) though the relationship was not linear (Figure 9A). In the case of the phylogenetically independent contrasts significant but weaker correlations (Pearson, 0.298, p < 10−15; Kendall, 0.043, p = 0.0409) were found (Figure 9B).
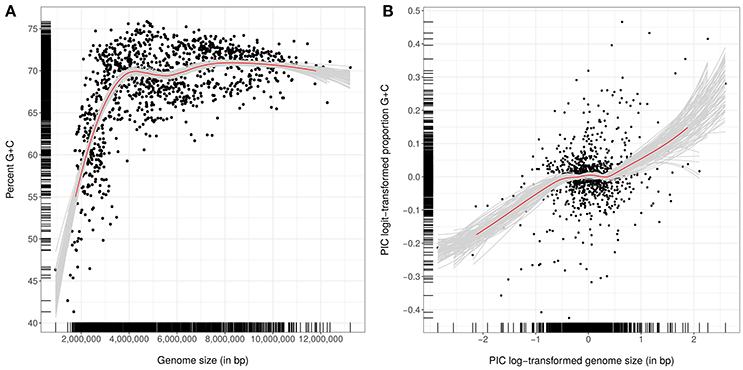
Figure 9. (A) Percent G+C content in dependence on genome size (in numbers of base pairs). (B) Phylogenetically independent contrasts (PICs) derived from logit-transformed proportions of G+C content in dependence on PICs derived from log-transformed genome sizes. PICs were calculated to account for the significant effect of the phylogeny on both G+C content and genome size. In both panels, Loess-based bootstrap aggregation (100 replicates) was applied; the resulting red curve denotes the average from 100 smoothers, each fitted to a subset of the original data set, to increase stability and reduce overfitting.
Experimentally derived G+C values cited in the descriptions of many species were found to be incorrect given corresponding data drawn from whole genome sequences while in other species descriptions G+C values were not available. These errors and omissions are addressed for many species using data drawn from genome sequences (see section on taxonomic revision).
Conflict with monophyly in a range of taxa was frequently seen in our genome-scale analyses though instances where a taxon was strongly supported in the GBDP tree but seriously out of kilter with corresponding 16S (or 23S) rRNA gene trees, or vice versa, were rarely observed (Supplementary File 2). As in previous studies, analyses of the genome-scale data resulted in more strongly resolved trees though it is known that such studies may increase incongruities between distinct analyses (Jeffroy et al., 2006; Klenk and Göker, 2010). Indeed, topological incongruities between analyses of single genes attributed to horizontal gene transfer have been used to argue against the concept of hierarchical classification (Bapteste and Boucher, 2009; Klenk and Göker, 2010). However, the addition of more genes, up to virtually all available ones, increases support in phylogenomic trees (Breider et al., 2014) thereby indicating a strong hierarchical signal. Phylogenetic inferences drawn from limited studies on few genes can hardly be called genome-scale, such studies rely on assumptions about the relative suitability of the selected genes (Lienau and DeSalle, 2009; Klenk and Göker, 2010). In contrast, whole-genome methods, such as GBDP, yield truly genome-based phylogenies though overestimating phylogenetic confidence from genome-scale data must be avoided (Taylor and Piel, 2004). Bootstrapping entire genes instead of single alignment positions can reduce incongruities between trees and thereby provide more realistic phylogenomic support values (Siddall, 2010; Simon et al., 2017) hence in this study partition bootstrapping was applied throughout in our supermatrix analyses. GBDP pseudo-bootstrapping in conjunction with the greedy-with-trimming algorithm (Meier-Kolthoff et al., 2014a) is akin to partition bootstrap.
It was particularly interesting that in the present study there were three cases where the 16S rRNA gene tree was somewhat misleading, two with respect to Intrasporangiaceae and one regarding the monophyly of Mycobacteriaceae. In all of these instances the 23S rRNA gene trees showed better congruence with the GBDP tree, albeit with low support, whereas the corresponding supermatrix analyses fully confirmed the results of the GBDP analyses. Potential causes for the probable incorrect 16S rRNA gene phylogenies include insufficient taxon sampling and misspecified models but it cannot be ruled out that in these cases the 16S rRNA gene truly conflicts with the organism tree. Atopobium and Olsenella also provided an interesting case because here the 23S rRNA gene analyses were strongly in conflict with the corresponding GBDP analyses. These differences might be due to horizontal gene transfer, particularly in light of the conflicting signal between protein-coding genes indicated by low partition bootstrap support in the supermatrix analyses (Supplementary File 2). However, irrespective of the conflicting topologies the ML and MP supermatrix trees indicate that Atopobium is not monophyletic.
It is encouraging that the taxonomic conclusions drawn from the GBDP tree were confirmed by corresponding supermatrix analyses in all of the cases investigated. Consequently, we believe that the approach used to assess whether conflict was evident between trees inferred from distinct data sets followed, where necessary, by additional analyses was a robust procedure. This also holds for the application of backbone constraint to augment comprehensible sampled single-gene data with information drawn from analyses of additional genes but fewer organisms (Liu et al., 2015; Hahnke et al., 2016). While the CCT allowed us to safely place type species lacking genome sequences and in one case to delineate an additional genus, it also highlighted other instances where taxonomic conclusions would have been premature.
The origin of the non-monophyletic taxa highlighted in this study cannot be understood without understanding the taxonomic practices used to generate these taxa. The current intense debate on how to describe new taxa focusses mainly on the question of which kind of characters should be investigated and in this context primarily on the contrast between phenotypic and genomic data (Vandamme and Peeters, 2014; Sutcliffe, 2015; Thompson et al., 2015). Based on the principles of phylogenetic systematics (Hennig, 1965; Wiley and Lieberman, 2011) and observations made in the course of this study we believe that the focus should be on character interpretation rather than on character sampling. In the current dominant practice of polyphasic taxonomy (Vandamme et al., 1996) 16S rRNA gene trees are usually inferred as the initial step following the identification of the taxa of interest; subsequently, new taxa are chosen from such trees and “diagnostic” features presented. Phenotypic characters still tend to be interpreted manually, they are rarely analyzed by clustering, let alone by using phylogenetic methods (Montero-Calasanz et al., 2017).
As for the initial step, almost all of the taxonomic problems observed in this study concerned taxa for which statistical support for their monophyly was not apparent in 16S rRNA gene trees, irrespective of whether they were inferred by us or depicted in the literature. In these single-gene trees the questionable taxa appeared to be monophyletic but with low bootstrap support or paraphyletic with weak (e.g., Amycolatopsis) to moderate (e.g., Corynebacterium) support against their monophyly. Thus, calculating branch support from single genes in an appropriate manner (or from genome-scale data as described above) is a necessary, but not a sufficient prerequisite, for safely generating monophyletic taxa; indeed taxa must also be chosen from trees so as to correspond to highly supported clades (Vences et al., 2013).
The use of diagnostic features in support for taxa is a practice that clashes with the principles of phylogenetic systematics which emphasize apomorphies as evidence for the monophyly of a group (Hennig, 1965; Wiley and Lieberman, 2011). Since we were unable to detect the use of the term “apomorphy” in any of the taxonomic publications evaluated in the course of this study, we would like to draw attention to this issue by reference to Figure 10. The pitfalls of relying on diagnostic features do not originate from the extent of homoplasy, which is zero in the case of the trees and characters in Figure 10, hence the issue at hand is not one of data quality. In the case of binary characters, such as the presence of mycolic acids in Corynebacterium against their absence in Turicella, even when homoplasies are lacking only one of the two characters states can be used to support the monophyly of a group as the other state is plesiomorphic. As shown in Figure 10A, the group recognized by the plesiomorphic state might accidentally be monophyletic, but it is more likely to be paraphyletic as depicted in Figure 10B. Reptiles were originally the best known example of a paraphyletic group, as they are perfectly defined by plesiomorphic features (Wiley and Lieberman, 2011). A single feature with two character states cannot be used to justify the separation of two distinct taxa because only the apomorphic character state can argue for the monophyly of one of the two taxa. The same holds for multiple binary characters provided one of the two taxa is plesiomorphic regarding all of them. Thus, contrary to a recent study (Baek et al., 2018), we conclude that discrepancies between traditional taxonomy and phylogenomic trees do not necessarily indicate that the features on which the traditional classification was based are in true conflict with the tree and that these features per se need to be treated with caution.
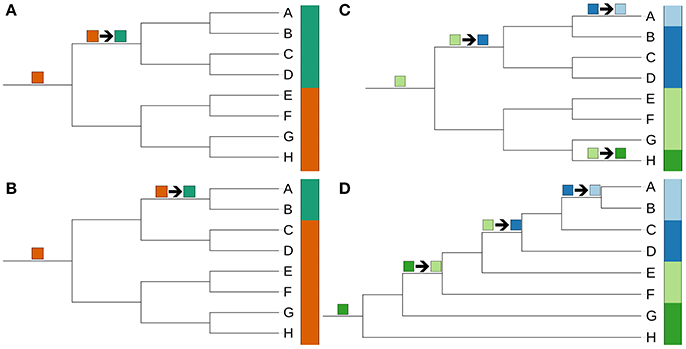
Figure 10. Hypothetical trees and character-state distributions illustrating the impossibility to detect “diagnostic” character states for monophyletic groups even in the case of a perfect fit of the character to the phylogeny. Most-parsimonious reconstructions of changes between character states are indicated by arrows. (A) Maximally symmetric tree with eight tips and a binary character without homoplasies. Each of the two character states is diagnostic for a clade. The orange-red state is actually plesiomorphic but accidentally happens to occur in a single clade only. (B) Same tree as before together with another binary character without homoplasies. Only one of the two character states is diagnostic for a monophyletic group. The orange-red state is again plesiomorphic, hence it comes as no surprise that it characterizes a paraphyletic group. (C) Same tree as before in conjunction with a multi-state character without homoplasies. Only two of the four states, light blue and dark green, are diagnostic, each for a distinct clade. (C) Maximally asymmetric tree with eight tips in conjunction with another multi-state character without homoplasies. Only one of the four states, light blue, is diagnostic for a clade.
In the case of multi-state characters, such as 16S rRNA gene signatures (Zhi et al., 2009) or peptidoglycan types [the latter have been particularly important in the classification of Gram-positive taxa, notably with respect to Actinobacteria (Schleifer and Kandler, 1972)], the difference between diagnostic features and apomorphies can be even more striking. Figure 10C illustrates a case of four characters states only two of which (light blue, dark green) define monophyletic groups whereas the other two states highlight paraphyletic groups. In the asymmetric tree (Figure 10D) only one of the four character states (light blue) defines a clade. The practice of only recognizing genera or higher taxa that are uniform with respect to single characters, such as peptidoglycan types and 16S rRNA gene signatures, should be discontinued as it is not sufficiently flexible to account for the various possible ways of character evolution even when homoplasies can be ruled out. The dark blue character state shown in Figure 10D, for instance, is actually a synapomorphy of strains A-D and hence plesiomorphic within that group; its change to light blue in the last common ancestor of A+B does not imply a homoplasy. Considering, the character to be diagnostic would either yield a paraphyletic taxon (C+D) or three taxa (A+B, C, and D) two of which cannot be distinguished using this character. The latter solution might amount to excessive splitting. Consequently, it would be better if taxa were allowed to have distinct character states even if the characters were previously considered to be diagnostic. An additional problem of 16S rRNA gene signatures is that they represent part of the information used to infer 16S rRNA gene trees and hence cannot provide independent evidence.
Attempts to classify large numbers of taxa using low numbers of characters (e.g., phenotypic ones) may frequently lead to the circumscription of “waste bin” taxa composed of groups that have no apparent apomorphy. This not only holds true for prokaryotic systematics but also for large non-monophyletic yeast genera, such as Candida (Daniel et al., 2014), Cryptococcus (Liu et al., 2015), and Rhodotorula (Wang et al., 2015), which in recent years have been split based on data derived from phylogenetic analyses. Indeed, in such instances the goal that taxonomic classification should be based on monophyletic taxa can be in conflict with the goal of providing apomorphies for all taxa, let alone diagnostic features, particularly for low numbers of previously selected (phenotypic or other) characters. As long as small numbers of characters are the limiting factor, taxonomic classification cannot fully benefit from the high resolution of phylogenetic trees based on available genome-scale data (Klenk and Göker, 2010). In cases such as Actinomyces we have favored the creation of taxa phylogenetically well-supported as monophyletic over insisting on recognizable apomorphies of selected characters. Researchers primarily interested in the differences in such characters could well emend taxon descriptions provided such differences were found (and, hopefully, synapomorphies would be among them). In the meantime those using taxonomic classifications can benefit from taxa that now more likely represent natural groups.
Since single characters do not yield bootstrapping or other statistical support it is not clear how to apply them in rigorous tests of the plausibility of certain phylogenetic outcomes. According to Dollo's law complex features arise only once in evolution but may be lost several times (Farris, 1977), hence a group of organisms displaying a complex feature should be monophyletic or paraphyletic in a tree, but not polyphyletic. The acid-fastness of most Corynebacteriales, indicating a multi-layered cell wall containing mycolic acids (Barry et al., 1998), might well-represent such a complex feature. If so, our GBDP tree (Figure 3) passed the test because Corynebacteriales appear to be monophyletic whereas the strains containing mycolic acids appear to be paraphyletic. Independent losses of mycolic acids within Corynebacteriales (Baek et al., 2018) indeed yield homoplasy but this is as expected under Dollo's law. The presence of mycolic acids, the apparent apomorphy of Corynebacteriales, was thereby shown to be plesiomorphic within the order and lost again in groups such as Turicella. A similar reasoning can be applied to rare peptidoglycan types which might characterize monophyletic or paraphyletic groups as in Figure 9D.
The results of the dDDH analyses revealed a number of new subspecies. In many cases this came as no surprise as these taxa had already regarded as later heterotypic synonyms while a dDDH subspecies boundary has only recently been introduced (Meier-Kolthoff et al., 2014b). In our view, applying a firm dDDH boundary for subspecies is a logical development analogous to cut-off values for the delineation of species. Moreover, dDDH similarities are more likely to yield monophyletic subspecies (Meier-Kolthoff et al., 2014c) than a selection of phenotypic differences based on a few characters. In other cases, heterotypic synonyms postulated in the literature or species regarded as distinct based on traditional DDH values, could not be confirmed. The most likely reason for this is that traditional DDH procedures are more error-prone than dDDH (Meier-Kolthoff et al., 2013a). Much like Average Nucleotide Identity (ANI), the choice of a similarity method and a species cut-off for dDDH was based on mimicking traditional DDH as closely as possible, albeit without its problematic reproducibility; however, according to this criterion dDDH yielded better results than ANI and can thereby be seen as the method of choice (Meier-Kolthoff et al., 2013a). For instance, the recent controversy regarding the subspecies of Mycobacterium abscessus (Adekambi et al., 2017; Tortoli et al., 2017) may have been avoided if dDDH had been preferred or pairwise comparison had been restricted to those involving type strains (Meier-Kolthoff et al., 2014c).
The significant correlation found between genome size and G+C content was not unexpected as trends to both larger genomes and higher G+C content in free-living organisms have been postulated previously (Mann and Chen, 2010). Potential mechanisms that increase genomic G+C content include positive selection (Hildebrand et al., 2010) and G+C-biased gene conversion (Lassalle et al., 2015). In contrast, symbiotic bacteria tend to have smaller genomes due to reductive evolution and to be richer in A+T content due to nutrient limitations as A+T synthesis is energetically less demanding (Rocha and Danchin, 2002; Mann and Chen, 2010). This can be easily confirmed for mutualistic actinobacteria such as Bifidobacterium (Supplementary File 2) and for parasitic ones like Tropheryma (Figure 1). In addition, we have shown that the relationship between genome size and G+C content is non-linear at least within the phylum Actinobacteria (Figure 9). The ability to conduct such analyses for an entire phylum underlines the advantage of comprehensive taxonomic treatments over ones restricted to specific taxa (Hahnke et al., 2016). A non-linear relationship has also been observed between genome size and G+C content of monocotyledonous plants (Šmarda et al., 2014) where it even appeared to be regularly quadratic.
As for the use of genome size as a taxonomic marker, it can be regarded as non-independent of G+C content. However, due to the non-linear relationship this dependency is much lower for higher genome sizes (Figure 9). Indeed, if one takes into account that, on average, larger genomes also display a greater variance as they represent count data (Crawley, 2007), genome sizes often appeared to be genus-specific (Figures 1–8). The correlation between G+C content and genome size was still significant after accounting for the impact of the phylogeny albeit considerably weaker (Figure 9). Both bacterial G+C content and bacterial genome size appeared to be strongly phylogenetically conserved, an observation that may have been underestimated in previous studies. It is for these reasons that genome sizes have been added to the description of the reclassified and emended species as shown below. As in the case of the Bacteroidetes (Hahnke et al., 2016), many species descriptions were found to be inaccurate or too imprecise now that it has been shown that within-species deviation in genomic G+C content is at most 1% (Meier-Kolthoff et al., 2014c). It is good practice to strengthen species descriptions in this way as such values not only assist in highlighting strains that do not belong to the same species but also show significant correlation to phylogenetic trees. In contrast, it is premature to redefine actinobacterial genera and higher taxa in this way; as additional type-strain genome sequences are needed before this issue can be addressed.
The results of this comparative phylogenomic study provide a much improved framework for the classification of the phylum Actinobacteria, one of the largest taxonomic lineages recognized in the domain Bacteria. The improved classification provides a sound basis for future studies on actinobacteria, not least on those of agricultural, biotechnological, clinical, and ecological interest. In particular, the clarification of relationships within taxa that, until now, have confounded the best efforts of actinobacterial systematists (Ludwig et al., 2012) will be of especial interest to those involved in developing biotechnological applications of strains belonging to taxa classified in emended families such as the Gordoniaceae, Micromonosporaceae, Nocardiaceae, and Streptomycetaceae. Similarly, improvements in the classification of taxa assigned to emended families, notably Actinomycetaceae and Corynebacteriaceae, will provide valuable leads to those working on organisms relevant to human and veterinary medicine. Indeed, it can be concluded that comparative genomic analyses not only provide “grist to the taxonomic mill” (Klenk and Göker, 2010; Sutcliffe et al., 2012), as exemplified in this study, but also provide invaluable insights into actinobacterial biology in its entirely thereby helping to revitalize prokaryotic systematics as a fundamental scientific discipline.
Cryp.to.spo.ran.gi.a'les (N.L. neut. n. Cryptosporangium, type genus of the order; -ales, ending to denote an order; N.L. fem. pl. n. Cryptosporangiales, the Cryptosporangium order).
The description is as given for Cryptosporangiaceae (Zhi et al., 2009; Sen et al., 2014). The type and only genus of the order is Cryptosporangium (Tamura et al., 1998).
Spo.rich.thy.a'les (N.L. fem. n. Sporichthya, type genus of the order; -ales, ending to denote an order; N.L. fem. pl. n. Sporichthyales, the Sporichthya order).
The description is as given for Sporichthyaceae (Stackebrandt et al., 1997; Zhi et al., 2009). The type and sole genus of the order is Sporichthya (Lechevalier et al., 1968).
Ac.ti.no.po.ly.mor.pha'ce.ae (N.L. fem. n. Actinopolymorpha, type genus of the family; -aceae, ending to denote a family; N.L. fem. pl. n. Actinopolymorphaceae, the Actinopolymorpha family).
Aerobic, Gram-stain-positive, non-acid-alcohol-fast, non-motile organisms that form irregular cells. Aerial mycelium is absent or scant. The diamino acid of the peptidoglycan is LL-A2pm. The predominant respiratory quinones are either MK-9 or MK-10 and the major polar lipids DPG, PG and PI together with phosphatidylglycolipids. The genomic G+C content is around 66–73%. The family comprises Actinopolymorpha (Wang et al., 2001), the type genus, Flindersiella, Tenggerimyces, and Thermasporomyces.
An.tri.coc.ca'ce.ae (N.L. masc. n. Antricoccus, type genus of the family; -aceae, ending to denote a family; N.L. fem. pl. n. Antricoccaceae, the Antricoccus family).
The description is as given for Antricoccus (Lee, 2015), the type and sole genus of the family.
Ja.tro.phi.ha.bi.tan.ta'ce.ae (N.L. masc. n. Jatrophihabitans, type genus of the family; -aceae, ending to denote a family; N.L. fem. pl. n. Jatrophihabitantaceae, the Jatrophihabitans family).
The description is as given for Jatrophihabitans (Madhaiyan et al., 2013), the type and sole genus of the family.
Krib.bel.la'ce.ae (N.L. fem. n. Kribbella, type genus of the family; -aceae, ending to denote a family; N.L. fem. pl. n. Kribbellaceae, the Kribbella family).
Aerobic, Gram-stain-positive organisms which form extensively branched hyphae that penetrate into agar media. Typically produce aerial hyphae; substrate and aerial hyphae usually show various degrees of fragmentation eventually resulting in the production of coccoid to irregular, non-motile, rod-shaped elements. Anaerobic growth has been detected. The wall peptidoglycan contains LL-A2pm, alanine, glutamic acid, and glycine; muramic acid residues of the peptidoglycan are N-acetylated; the main cell-wall polymer is either teichuronic or teichulosonic acid. The cellular fatty acids are mainly saturated, iso- and anteiso-branched acids, and the predominant isoprenoid quinone is MK-9(H4). An ubiquitous polar lipid is PC, additional components include DPG, PG, and PI. The genomic G+C content is around 65–77%. The family comprises only Kribbella (Park et al., 1999; Sohn et al., 2003; Everest et al., 2013), the type genus.
Ky.to.coc.ca'ce.ae (N.L. masc. n. Kytococcus, type genus of the family; -aceae, ending to denote a family; N.L. fem. pl. n. Kytococcaceae, the Kytococcus family).
Aerobic, Gram-stain-positive, asporogenous organisms which form coccoid cells. The peptidoglycan contains L-lysine as the diaminoacid and is of the A4α type. The predominant isoprenologues are MK-7, MK-8, MK-9, and MK-10 and the major polar lipids DPG, PG, and PI. The predominant cellular fatty acids are anteiso-C17:0 and iso-C17:1. Does not contain mycolic acids or teichoic acids. The genomic G+C content is around 68–73%. The family only contains Kytococcus (Stackebrandt et al., 1995), the type genus.
Law.so.nel.la'ce.ae (N.L. fem. n. Lawsonella, type genus of the family; -aceae, ending to denote a family; N.L. fem. pl. n. Lawsonellaceae, the Lawsonella family).
The description is as given for Lawsonella (Bell et al., 2016), the type and sole genus of the family.
Or.ni.thi.ni.mi.cro.bi.a'ce.ae (N.L. neut. n. Ornithinimicrobium, type genus of the family; -aceae, ending to denote a family; N.L. fem. pl. n. Ornithinimicrobiaceae, the Ornithinimicrobium family).
Aerobic to microaerobic, Gram-stain-positive, non-motile organisms which form short irregular rods and/or cocci. The peptidoglycan type is A4β, and muramic acid residues of the peptidoglycan are N-acetylated. The main cellular fatty acids are anteiso- and iso-branched acids, the predominant isoprenologue is MK-8(H4) and the major whole-cell sugars are arabinose, rhamnose and xylose. The polar lipid profile contains DPG, PG and PI. Does not have mycolic acids. Catalase positive and oxidase negative. The genomic G+C content is around 68–74%. The family includes Ornithinimicrobium (Groth et al., 2001), the type genus, and Serinicoccus.
Tro.phe.ry.ma.ta'ce.ae (N.L. neut. n. Tropheryma, type genus of the family; -aceae, ending to denote a family; N.L. fem. pl. n. Tropherymataceae, the Tropheryma family).
The description is as given for Tropheryma (La Scola et al., 2001) with the following additions. Grows in tissue culture media in co-culture with human fibroblasts axenically in such media enriched with amino acids. Does not grow on standard laboratory media. Forms straight regular small rods. Gram-stain-negative in culture but variably Gram-stain-positive in infected human tissues. The genomic G+C content is around 46%. The type and only genus is Tropheryma (La Scola et al., 2001).
Al.der.so'ni.a (N.L. fem. n. Aldersonia, named after Grace Alderson, a British microbiologist who made significant contributions to rhodococcal systematics).
Aerobic, Gram-stain-positive, acid-fast, non-motile organism which forms filaments or shows elementary branching in early growth phase and produces mainly cocci in stationary phase. Displays a rod -coccus growth cycle. Circular, convex, pink colonies are formed on yeast extract-malt extract agar after incubation for 7 days at 28°C. Whole-cell hydrolysates contain DL-A2pm, arabinose and galactose, the predominant isoprenologue is MK-8(H2) and DPG, PE, PI and PIM's are evident in polar lipid profiles. The major fatty acids are C16:0, C18:1 ω9c, and C16:1 ω7c. The genomic G+C content is about 66%. The type species is Aldersonia kunmingensis comb. nov.
Bou.da.bou'si.a (N.L. fem. n. Boudabousia, named after Abdellatif Boudabous Chihi, the first microbiologist in Tunisia, for his contributions to prokaryotic systematics).
Facultatively anaerobic, Gram-stain-positive, non-acid-fast, non-motile, catalase-negative, non-hemolytic organisms which form straight to slightly curved rods, some show branching. Colonies are gray, entire, convex, and pinpoint to 0.5 mm in diameter after 48 h. The major fatty acids are C16:0, C18:0, and C18:1 cis9. The genomic G+C content is about 54%. The type and only species is Boudabousia marimammalium comb. nov.
Bow.de'ni.a (N.L. fem. n. Bowdenia, named after George H. Bowden, a Canadian microbiologist, for his work on oral microbiology).
Facultatively anaerobic, Gram-stain-positive, catalase-negative organisms which form short diphtheroid shaped rods; branched elements and coccoid forms may be found. Pinpoint, white or gray, opaque, shiny, entire, and convex colonies produced after 48 h under anaerobic conditions. The major isoprenologue is MK-9. The fatty-acid profile contains major amounts of anteiso-C15:0, C16:0, C18:0, and C18:1 w9c. The genomic G+C content is around 65%. The type species is Bowdenia nasicola comb. nov.
Bu.cha.na.nel'la (L. fem. dim. ending -ella; N.L. fem. n. Buchananella, named after Robert E. Buchanan, an American microbiologist who made significant contributions to prokaryotic systematics).
Facultatively anaerobic, Gram-stain-positive, weakly catalase-positive organisms which form pleomorphic rods and filaments; the latter show branching on blood agar. Microcolonies are filamentous on brain-heart-infusion agar supplemented with bovine serum. Mature colonies on bovine blood agar are white, adhere to the agar and are molar-toothed. The peptidoglycan contains L-lysine, alanine, and glutamic acid; some strains also have L-ornithine. The muramic acid moiety of the peptidoglycan is N-glycosylated. Cell-wall sugars include galactose and glucose, but not 6-deoxytalose or rhamnose. The G+C content is around 67–68%. The type species is Buchananella hordeovulneris comb. nov.
Jong.sik.chu'ni.a (N.L. fem. n. Jongsikchunia, named after Jongsik Chun, a Korean microbiologist, for his extensive work on actinobacterial systematics).
Aerobic, Gram-stain-positive, non-motile organisms that form non-spore-forming rods and display a rod-coccus growth cycle. Colonies are white, opaque and polymorphic on complex media. Catalase positive and oxidase negative. Whole-cell hydrolysates contain DL-A2pm, arabinose and galactose. The muramic acid residues of the peptidoglycan are N-glycolated. The major fatty acids are C16:1 ω7c, C16:0, C18:1 ω9c, and 10-methyl C18:0; the predominant isoprenologue MK-9(H2); the polar lipid profile contains DPG, PE, PI, and PIMs. The principal mycolic acids have a chain length of 58-60 carbon atoms. The genomic G+C content is around 67%. The type species is Jongsikchunia kroppenstedtii comb. nov.
Fla.vi.mo'bi.lis (L. adj. flavus, yellow, pertaining to the yellow color of the colonies; L. adj. mobilis, mobile; N.L. masc. n. Flavimobilis, a yellow motile bacterium).
Aerobic or facultative anaerobic, Gram-stain-positive, motile, asporogenous organisms which form regular of irregular short rods. They grow from 15 to 42°C; arbutin and starch are degraded. The peptidoglycan contains meso-A2pm as the diamino acid and is of the A4α type. The cellular fatty-acid profile contains major amounts of anteiso-methyl branched and straight-chain saturated components; the predominant isoprenologue is MK-9(H4). Does not form mycolic acids. The genomic G+C content is around 72%. The type species is Flavimobilis marinus comb. nov.
Em.ble'ya (N.L. fem. n. Embleya, named after the British microbiologist T. Martin Embley for his contribution to actinobacterial systematics).
Well-developed, irregularly branched substrate and aerial mycelium formed. A gray aerial spore mass is produced on oatmeal agar. Mature spore chains are spiral with more than 20 cylindrical, rugose ornamented spores per chain. Whole-cell hydrolysates contain LL-A2pm and traces of arabinose but lack galactose. The muramic acid residues of the peptidoglycan are N-glycolated. The major isoprenologue is MK-9(H4) and the predominant phospholipids PE and an unidentified glucosamine component. The genomic G+C content is about 70%. The type and only species is Embleya scabrispora comb. nov.
Glei'mi.a (N.L. fem. n. Gleimia, named after Dorothea Gleim, a German microbiologist, for her work on the DSMZ Prokaryotic Nomenclature up-to-date Service).
Facultatively anaerobic, non-motile, Gram-stain-positive, slightly curved rods or short rods or coryneform cells are formed. The peptidoglycan type is A5α. The fatty-acid profile contains major amounts of C16:0, C18:1 ω9, C18:0, and C14:0. The type species is Gleimia europaea comb. nov.
Fan.ny.hes'se.a (N.L. fem. n. Fannyhessea, of Hesse, named to honor Angelina Fanny Hesse for her great contribution to microbiology, the use of agar to solidify media).
Microaerophilic or anaerobic, Gram-stain-positive cocci which occur singly, in pairs or short chains. Acid is not produced from either mannose or raffinose. Indole negative. Nitrate is not reduced. Gelatin and aesculin are not hydrolysed. Predominant fatty acids are C16:0, C18:0, and C18:1 ω9c. The genomic G+C content is around 43%. The type species is Fannyhessea vaginae.
Lan.ce.fiel.del'la (L. fem. dim. ending -ella; N.L. fem. n. Lancefieldella, named after Rebecca Lancefield, an American microbiologist known for her significant contributions to Streptococcus biology).
Obligately anaerobic, Gram-stain-positive, non-motile, asporogenous, small rods which occur singly, in pairs or short chains. Central swellings may occur particularly in cells grown on solid media without Tween. Colonies on brain-heart blood agar range from pinpoint to 2 mm in diameter and are raised or low convex, entire and translucent or transparent after anaerobic incubation for 2 to 5 days at 37°C. Growth is stimulated by 0.02% (v/v) Tween 80 with 10% (v/v) rabbit serum. Acid is produced from mannose and glucose. Indole, gelatin and catalase negative. Predominant fatty acids are C18:1 ω9c and C18:1 dimethylacetyl. The genomic G+C content is around 39–45%. The type species is Lancefieldella parvula comb. nov.
Paul.jen.se'ni.a (N.L. fem. n. Pauljensenia, named after Paul Jensen, an American microbiologist, for his work on Salinispora from ecological and taxonomic perspectives).
Aerobic, Gram-stain-positive, catalase-negative, non-hemolytic organisms that form straight short to medium rods. Pinpoint colonies are formed on sheep blood agar after anaerobic incubation for 24 h. The type and only species is Pauljensenia hongkongensis comb. nov.
Schaa'li.a (N.L. fem. n. Schaalia, named after Klaus P. Schaal, a German medical microbiologist, for his extensive work on Actinomyces systematics).
Aerobic, Gram-stain-positive organisms which form straight to slightly curved rods some of which may show branching; some species only produce coccoid or coccobacillary cells. Microcolonies are not filamentous. Peptidoglycan contains L-lysine as the diamino acid and is either of the A5α or A5β type; the muramic acid residue of the peptidoglycan is N-acetylated. The major cell-wall sugars are mannose and rhamnose; the predominant menaquinone is MK-9(H4), minor amounts of MK-10(H4) and MK-8(H4) also occur. The major polar lipids are DPG and PG. Fatty-acid profiles are rich in saturated, unsaturated and iso- and anteiso-branched-chain components The genomic G+C content is 56–70%. The type species is Schaalia odontolytica comb. nov.
Pe.do.coc'cus (Gr. n. pedon, the ground, earth; N.L. masc. n. coccus, (from Gr. n. kokkos) a grain or berry; N.L. masc. n. Pedococcus, coccus from soil).
Aerobic, Gram-stain-positive, asporogenous, catalase-positive organisms which form cocci or short rods. Circular, smooth, convex, cream, white or yellow colonies are formed. The peptidoglycan contains DL-A2pm as the diamino acid, demethylmenaquinone MK-8(H4) is the predominant isoprenologue and the main fatty acids are iso-C15:0 and iso-C16:0 acids. The genomic G+C content is about 69–74%. The type species is Pedococcus dokdonensis comb. nov.
Win'ki.a (N.L. fem. n. Winkia, named after Joachim Wink, a German microbiologist who has made a significant contributions to actinobacterial systematics).
Facultatively anaerobic, Gram-stain-positive, asporogenous, catalase-positive organisms that are predominantly diphtheroid though coccobacilli may occur, rods are mainly arranged in clusters or as V or Y forms. Colonies are circular, smooth, convex, more white than cream colored with entire edges and 0.5 to 1.5 mm in diameter following incubation for 48 h. Straight-chain fatty acids are mainly palmitic and stearic acids. The genomic G+C content is around 55–58%. The type species is Winkia neuii comb. nov.
Ying.hu.an'gi.a (N.L. fem. n. Yinghuangia, named after Ying Huang, a Chinese microbiologist, for her work on the reclassification of Streptomyces based on MLSA).
Aerobic, Gram-stain-positive organisms which form an extremely branched substrate mycelium that bears an abundant aerial spore mass on yeast extract/malt extract agar. Produces imperfect spiral chains (rectinaculiapetri) or flexuous to long chains of smooth-surfaced spores. The wall peptidoglycan contains LL-A2pm as the diamino acid; whole-organism hydrolysates contain galactose, mannose and ribose or arabinose, glucose, rhamnose, and ribose. The predominant isoprenologues are MK-9(H4, H6, H8) and the major polar lipids DPG, PE, and PI; PC has also been reported. The fatty-acid profiles contain major amount of saturated, iso-and anteiso-fatty acids. The genomic G+C content is around 70–75%. The type species is Yinghuangia aomiensis comb. nov.
A. fer.ru.gi'ne.us (L. masc. adj. ferrugineus, rusty brown, referring to the orange-dark brown to clove brown color of the substrate mycelium).
Basonym: Pseudosporangium ferrugineum Ara et al. 2008
The description is as given for Pseudosporangium ferrugineum (Ara et al., 2008b) with the following modification. The G+C content of the type-strain genome is 72.5%, its approximate size 8.65 Mbp, its IMG deposit 2728369278. The type strain is 3-44-a(19) = JCM 14710 = DSM 45348.
A. cae.ci'co.la (N.L. n. caecum, caecum; L. suff. -cola, inhabitant of; N.L. n. caecicola, caecum-dweller).
Basonym: Parvibacter caecicola Clavel et al. 2013
The description is as given for Parvibacter caecicola (Clavel et al., 2013). The type strain is NR06 = DSM 22242 = CCUG 57646.
A. cae.ci.mu'ris (N.L. n. caecum, caecum; L. n. mus/muris, mouse; N.L. gen. n. caecimuris, of the caecum of a mouse).
Basonym: Enterorhabdus caecimuris Clavel et al. 2010
The description is as given for Enterorhabdus caecimuris (Clavel et al., 2010). The type strain is B7 = CCUG 56815 = DSM 21839.
A. mu.co.si'co.la (N.L. n. mucosa, mucosa; L. suff. -cola, inhabitant of; N.L. n. mucosicola, inhabitant of the intestinal mucosa).
Basonym: Enterorhabdus mucosicola Clavel et al. 2009
The description is as given for Enterorhabdus mucosicola (Clavel et al., 2009). The type strain is Mt1B8 = CCUG 54980 = DSM 19490.
A. mu'ris (L. gen. n. muris, of a mouse).
Basonym: Enterorhabdus muris Lagkouvardos et al. 2016
The description is as given for Enterorhabdus muris (Lagkouvardos et al., 2016). The type strain is WCA-131-CoC-2 = DSM 29508 = KCTC 15543.
A. al'ba (N.L. fem. adj. alba, white).
Basonym: Alloactinosynnema album Yuan et al. 2010
The description is as given for Alloactinosynnema album (Yuan et al., 2010) with the following addition. The G+C content of the type-strain genome is 69.7%, its approximate size 7.29 Mbp, its IMG deposit 2775506708. The type strain is 03-9939 = DSM 45114 = KCTC 19294 = CCM 7461.
A. i.ra'ni.ca (N.L. fem. adj. iranica, of or belonging to Iran).
Basonym: Alloactinosynnema iranicum Nikou et al. 2014
The description is as given for Alloactinosynnema iranicum (Nikou et al., 2014). The type strain is Chem10 = IBRC-M 10403 = CECT 8209.
A. kun.min.gen'sis (N.L. fem. adj. kunmingensis, pertaining to Kunming, a city in Yunnan province in south-west China).
Basonym: Rhodococcus kunmingensis Wang et al. 2008
The description is as given for Rhodococcus kunmingensis (Wang et al., 2008) with the following modification. The G+C content of the type-strain genome is 66.2%, its approximate size 5.62 Mbp, its GenBank deposit SAMN04357316. The type strain is DSM 45001 = JCM 15626 = KCTC 19149.
A. a'ri.da (L. fem. adj. arida, referring to the isolation of the strain from arid soil).
Basonym: Yuhushiella deserti Mao et al. 2011 (The name Amycolatopsis deserti has already been published (Busarakam et al., 2016) hence a new epithet has been chosen to avoid homonyms).
The description is as given for Yuhushiella deserti (Mao et al., 2011) with the following modification and addition. The G+C content of the type-strain genome is 72.7%, its approximate size 5.96 Mbp, its GenBank deposit SAMN05421810. The type strain is RA45 = DSM 45648 = JCM 16584.
B. por.ci'num (L. neut. adj. porcinum, of a hog).
The description is as given for Bifidobacterium thermacidophilum subsp. porcinum (Zhu et al., 2003) with the following addition. The G+C content of the type-strain genome is 60.2%, its approximate size 2.08 Mbp, its GenBank deposit SAMN02673456. The type strain is P3-14 = CGMCC 1.3009 = JCM 16945 = LMG 21689.
B. ma.ri.mam.ma'li.um (L. neut. n. mare, the sea; N.L. fem. gen. pl. n. mammalium, of mammals; N.L. fem. gen. pl. n. marimammalium, of marine mammals).
Basonym: Actinomyces marimammalium Hoyles et al. 2001
The description is as given for Actinomyces marimammalium (Hoyles et al., 2001b) with the following modification. The G+C content of the type-strain genome is 54.2%, its approximate size 1.84 Mbp, its GenBank deposit SAMN06006700. The type strain is M1749/98/1 = CCUG 41710 = CIP 106509 = DSM 15383.
B. na.si'co.la (L. n. nasus, nose; L. suff. -cola, inhabitant of; N.L. n. nasicola, inhabitant of the nose, referring to the place of isolation of the type strain).
Basonym: Actinomyces nasicola Hall et al. 2003
The description is as given for Actinomyces nasicola (Hall et al., 2003c) with the following modification. The G+C content of the type-strain genome is 65.2%, its approximate size 2.64 Mbp, its GenBank deposit SAMN06092712. The type strain is R2014 = CCUG 46092 = CIP 107668.
B. hor.de.o.vul'ne.ris (L. n. hordeum, barley; N.L. n. Hordeum, a grass genus; L. n. vulnus/vulneris, wound, injury; N.L. gen. n. hordeovulneris, isolated from injuries of awns of Hordeum).
Basonym: Actinomyces hordeovulneris Buchanan et al. 1984
The description is as given for Actinomyces hordeovulneris (Buchanan et al., 1984). The type strain is ATCC 35275 = CCUG 32937 = CIP 103149 = DSM 20732 = UCD 81-332-9.
C. krop.pen.sted'ti.i (N.L. gen. n. kroppenstedtii, of Kroppenstedt, in honor of Reiner Michael Kroppenstedt, a German microbiologist, for his contributions to the taxonomy of Actinobacteria).
Basonym: Gordonia kroppenstedtii Kim et al. 2009
The description is as given for Gordonia kroppenstedtii (Kim et al., 2009) with the following modification. The G+C content of the type-strain genome is 67.1%, its approximate size 4.23 Mbp, its GenBank deposit SAMN02441055. The type strain is NP8-5 = DSM 45133 = JCM 16948 = KCTC 19360.
C. cap'si.ci (N.L. neut. gen. n. capsici, referring to Capsicum, the genus name of pepper).
The description is as given for Clavibacter michiganensis subsp. capsici (Oh et al., 2016). The type strain is PF008 = KACC 18448 = LMG 29047.
C. ne.bras.ken'sis (N.L. masc. adj. nebraskensis, of or belonging to Nebraska).
The description is as given for Clavibacter michiganensis subsp. nebraskensis (Davis et al., 1984). The type strain is ATCC 27794 = CCUG 38894 = CIP 105362 = DSM 7483 = ICMP 3298 = JCM 9666 = LMG 3700 = LMG 5627 = LMG 7223 = NCPPB 2581 = VKM Ac-1404.
C. se.pe.do'ni.cus (Gr. n. sepedon, rottenness, decay, putrefaction; L. neut. suff. -icum, suffix used with the sense of pertaining to; N.L. masc. adj. sepedonicus, pertaining to decay).
The description is as given for Clavibacter michiganensis subsp. sepedonicus (Davis et al., 1984) with the following modification.The G+C content of the type-strain genome is 72.4%, its approximate size 3.40 Mbp, its GenBank deposit SAMEA1705948. The type strain is ATCC 33113 = CCUG 23908 = CIP 104844 = CFBP 2049 = DSM 20744 = ICMP 2535 = JCM 9667 = LMG 2889 = NCPPB 2137 = VKM Ac-1405.
C. nam.ne.ten'se (N.L. neut. adj. namnetense, of Namnetum, the Latin name of Nantes).
Basonym: Propionibacterium namnetense Aubin et al. 2016
The description is as given for Propionibacterium namnetense (Aubin et al., 2016). The type strain is NTS 31307302 = DSM 29427 = CCUG 66358.
D. ge.la.ti.ni.ly'ti.ca (N.L. neut. n. gelatinum, gelatin; N.L. adj. lyticus, able to dissolve; N.L. fem. adj. gelatinilytica, gelatin-dissolving).
Basonym: Lysinimicrobium gelatinilyticum Hamada et al. 2015
The description is as given for Lysinimicrobium gelatinilyticum (Hamada et al., 2015). The type strain is HI12-44 = NBRC 109390 = DSM 28149.
D. i.ri.o.mo.ten'sis (N.L. fem. adj. iriomotensis, pertaining to Iriomote Island, Japan, where the type strain was isolated).
Basonym: Lysinimicrobium iriomotense Hamada et al. 2015
The description is as given for Lysinimicrobium iriomotense (Hamada et al., 2015). The type strain is HI12-143 = NBRC 109399 = DSM 28146.
D. man.gro'vi (N.L. gen. n. mangrovi, of a mangrove).
Basonym: Lysinimicrobium mangrovi Hamada et al. 2012
The description is as given for Lysinimicrobium mangrovi (Hamada et al., 2012). The type strain is HI08-69 = NBRC 105856 = DSM 24868.
D. ma'ris (L. gen. n. maris, of the sea).
Basonym: Lysinimicrobium aestuarii Hamada et al. 2015 (The name Demequina aestuarii has already been validly published (Yi et al., 2007) hence a new epithet has been chosen to avoid homonyms).
The description is as given for Lysinimicrobium aestuarii (Hamada et al., 2015). The type strain is HI12-104 = NBRC 109392 = DSM 28144.
D. pe.lo'phi.la (Gr. n. pelos, mud; Gr. adj. philos, loving; N.L. fem. adj. pelophila, mud-loving).
Basonym: Lysinimicrobium pelophilum Hamada et al. 2015
The description is as given for Lysinimicrobium pelophilum (Hamada et al., 2015). The type strain is HI12-111 = NBRC 109393 = DSM 28148.
D. phy.to'phi.la (N.L. n. phyto, plant; Gr. adj. phila, loving; N.L. fem. adj. phytophila, plant-loving).
Basonym: Lysinimicrobium flavum Hamada et al. 2015 (The name Demequina flava has already been validly published (Hamada et al., 2013) hence a new epithet has been chosen to avoid homonyms).
The description is as given for Lysinimicrobium flavum (Hamada et al., 2015). The type strain is HI12-45 = NBRC 109391 = DSM 28150.
D. rhi.zos.phae'rae (Gr. neut. n. rhiza, root; L. n. sphaera, ball, sphere; N.L. gen. n. rhizosphaerae, of the rhizosphere).
Basonym: Lysinimicrobium rhizosphaerae Hamada et al. 2015
The description is as given for Lysinimicrobium rhizosphaerae (Hamada et al., 2015). The type strain is HI12-135 = NBRC 109397 = DSM 28152.
D. sil.va'ti.ca (N.L. fem. adj. silvatica, of the forest).
Basonym: Lysinimicrobium luteum Hamada et al. 2015 (The name Demequina lutea has already been validly published (Finster et al., 2009) hence a new epithet has been chosen to avoid homonyms).
The description is as given for Lysinimicrobium luteum (Hamada et al., 2015). The type strain is HI12-123 = NBRC 109395 = DSM 28147.
D. so'li (L. gen. n. soli, of soil).
Basonym: Lysinimicrobium soli Hamada et al. 2015
The description is as given for Lysinimicrobium soli (Hamada et al., 2015). The type strain is HI12-122 = NBRC 109394 = DSM 28151.
D. sub.tro'pi.ca (L. pref. sub, under, below, slightly; L. neut. adj. tropicum, tropical; N.L. fem. adj. subtropica, subtropical, referring to the subtropical region).
Basonym: Lysinimicrobium subtropicum Hamada et al. 2015
The description is as given for Lysinimicrobium subtropicum (Hamada et al., 2015). The type strain is HI12-128 = NBRC 109396 = DSM 28145.
E. sca.bri'spo.ra (L. adj. scaber/-bra/-brum, scabby, rough; Gr. fem. n. spora, seed; N.L. fem. adj. scabrispora, referring to the rugose surface of the spores).
Basonym: Streptomyces scabrisporus Ping et al. 2004
The description is as given for Streptomyces scabrisporus (Ping et al., 2004). The type strain is KM-4927 = JCM 11712 = NBRC 100760 = NRRL B-24202 = DSM 41855.
F. ma.ri'nus (L. masc. adj. marinus, pertaining to the sea).
Basonym: Sanguibacter marinus Huang et al. 2005
The description is as given for Sanguibacter marinus (Huang et al., 2005) with the following modification. The G+C content of the type-strain genome is 72.0%, its approximate size 2.90 Mbp, its GenBank deposit SAMN04488035. The type strain is 1-19 = CGMCC 1.3457 = JCM 12547 = DSM 19083.
F. so'li (L. gen. n. soli, of soil).
Basonym: Sanguibacter soli Kim et al. 2008
The description is as given for Sanguibacter soli (Kim et al., 2008) with the following modification. The G+C content of the type-strain genome is 71.9%, its approximate size 2.89 Mbp, its GenBank deposit SAMN04490250. The type strain is DCY22 = KCTC 13155 = JCM 14841.
G. co.le.o.ca'nis (Gr. n. koleos, vagina; L. gen. n. canis, of the dog; N.L. gen. n. coleocanis, of the vagina of a dog).
Basonym: Actinomyces coleocanis Hoyles et al. 2002
The description is as given for Actinomyces coleocanis (Hoyles et al., 2002a) with the following addition. The G+C content of the type-strain genome is 49.6%, its approximate size 1.72 Mbp, its GenBank deposit SAMN00001853. The type strain is M343/98/2 = CCUG 41708 = CIP 106873 = DSM 15436.
G. eu.ro.pae'a (L. fem. adj. europaea, of or belonging to Europe, European, referring to the fact that six different European laboratories contributed to the description of the species).
Basonym: Actinomyces europaeus Funke et al. 1997
The description is as given for Actinomyces europaeus (Funke et al., 1997a). The type strain is ATCC 700353 = CCUG 32789 A = CIP 105308 = LMG 18454.
G. ho'mi.nis (L. n. homo/-inis, a human being, man, person; L. gen. n. hominis, of man, indicating that the type strain was isolated from a human).
Basonym: Actinomyces hominis Funke et al. 2010
The description is as given for Actinomyces hominis (Funke et al., 2010a). The type strain is 1094 = 7894GR = CCUG 57540 = DSM 22168.
G. bi'fi.dus (L. masc. adj. bifidus, divided into two parts).
Basonym: Zimmermannella bifida Lin et al. 2004
The description is as given for Zimmermannella bifida (Lin et al., 2004). The type strain is CCUG 50209 = JCM 12920 = NBRC 103089 = TISTR 1511 = DSM 17450.
G. fae.ca'lis (N.L. masc. adj. faecalis, faecal).
Basonym: Zimmermannella faecalis Lin et al. 2004
The description is as given for Zimmermannella faecalis (Lin et al., 2004) with the following modification. The G+C content of the type-strain genome is 68.5%, its approximate size 2.85 Mbp, its GenBank deposit SAMN02440490. The type strain is ATCC 13722 = CCUG 37873 = JCM 12866 = NBRC 15706 = TISTR 1514.
H. ha.lo'phi.la (Gr. n. halos, salt; Gr. adj. philos, loving; N.L. fem. adj. halophila, salt-loving, referring to the ability of the type strain to grow at high NaCl concentrations).
Basonym: Amycolatopsis halophila Tang et al. 2010
The description is as given for Amycolatopsis halophila (Tang et al., 2010a) with the following modification. The G+C content of the type-strain genome is 67.8%, its approximate size 5.55 Mbp, its GenBank deposit SAMN02597165. The type strain is DSM 45216 = JCM 18118 = KCTC 19403.
H. sa.li.to'le.rans (L. n. sal/salis, salt; L. part. adj. tolerans, tolerating, enduring; N.L. part. adj. salitolerans, salt tolerating).
Basonym: Amycolatopsis salitolerans Guan et al. 2012
The description is as given for Amycolatopsis salitolerans (Guan et al., 2012). The type strain is TRM F103 = CCTCC AB 208326 = JCM 15899 = DSM 45783.
H. va.gi'nae (L. n. vagina, sheath, vagina; L. gen. n. vaginae, of the vagina).
Basonym: Atopobium vaginae Rodriguez Jovita et al. 1999
The description is as given for Atopobium vaginae (Rodriguez Jovita et al., 1999) with the following modification. The G+C content of the type-strain genome is 42.7%, its approximate size 1.43 Mbp, its GenBank deposit SAMN00001474. The type strain is CCUG 38953 = ATCC BAA-55 = CIP 106431.
I. fla'vum (L. neut. adj. flavum, yellow).
Basonym: Monashia flava Azman et al. 2016
The description is as given for Monashia flava (Azman et al., 2016). The type strain is MUSC 78 = DSM 29621 = MCCC 1K00454 = NBRC 110749.
K. in.di.go'fe.ra (N.L. n. indigo, the dye indigo; L. suff. -fer/-fera/-ferum, bearing; N.L. fem. adj. indigofera, bearing indigo, referring to production of blue to green diffusible pigments on chemically defined media).
Basonym: Streptomyces indigoferus Shinobu and Kawato 1960
The description is as given for Streptomyces indigoferus (Shinobu and Kawato, 1960). The type strain is ATCC 23924 = CBS 908.68 = BCRC (formerly CCRC) 13773 = DSM 40124 = IFO (now NBRC) 12878 = IFO (now NBRC) 3868 = JCM 4646 = NCIMB 9718 = NRRL B-3301.
K. xan.tho.ci'di.ca (Gr. adj. xanthos, yellow; L. v. caedo, to kill; N.L. fem. adj. xanthocidica, pertaining to yellow and to cut, probably referring to the name given the antibiotic produced which, in turn, was probably derived from its activity against Xanthomonas oryzae).
Basonym: Streptomyces xanthocidicus Asahi et al. 1966 (Approved Lists 1980)
The description is as given for Streptomyces xanthocidicus (Asahi et al., 1966). The type strain is CGMCC 4.1424 = ATCC 27480 = CBS 770.72 = BCRC (formerly CCRC) 11874 = DSM 40575 = IFO (now NBRC) 13469 = JCM 4243 = JCM 4862 = NRRL B-12504 = NRRL-ISP 5575 = VKM Ac-872.
K. rem'sis (N.L. gen. n. remsis, of REMS, acronym of “Regenerative Enclosed Life Support Module Simulator”).
Basonym: Tetrasphaera remsis Osman et al. 2007
The description is as given for Tetrasphaera remsis (Osman et al., 2007) with the following modification. The G+C content of the type-strain genome is 70.7%, its approximate size 3.57 Mbp, its GenBank deposit SAMN05421826. The type strain is 3-M5-R-4 = ATCC BAA-1496 = CIP 109413 = JCM 15662.
L. par'vu.la (L. fem. dim. adj. parvula, very small).
Basonym: Atopobium parvulum (Weinberg et al. 1937) Collins and Wallbanks 1993
The description is as given for Atopobium parvulum (Collins and Wallbanks, 1992) with the following modification. The G+C content of the type-strain genome is 45.7%, its approximate size 1.54 Mbp, its GenBank deposit SAMN00001907. The type strain is JCM 10300 = ATCC 33793 = DSM 20469.
L. ri'mae (L. gen. n. rimae, of a fissure, pertaining to the gingival crevice).
Basonym: Atopobium rimae (Olsen et al. 1991) Collins and Wallbanks 1993
The description is as given for Atopobium rimae (Collins and Wallbanks, 1992) with the following modification. The G+C content of the type-strain genome is 49.3%, its approximate size 1.63 Mbp, its GenBank deposit SAMN00000717. The type strain is ATCC 49626 = CCUG 31168 = DSM 7090 = IFO (now NBRC) 15546 = JCM 10299 = LMG 11476.
L. ae.ro.co.lo.ni'ge.nes (Gr. n. aer, air; L. n. colonia, a colony; N.L. suff. -genes, producing; N.L. fem. adj. aerocolonigenes, producing aerial colonies).
Basonym: Lechevalieria aerocolonigenes (Labeda, 1986) Labeda et al. 2001
The description is as given for Lechevalieria aerocolonigenes (Labeda, 1986; Labeda et al., 2001) with the following modification. The G+C content of the type-strain genome is 68.9%, its approximate size 10.80 Mbp, its GenBank deposit SAMN05661031. The type strain is Shinobu 701 = ATCC 23870 = BCRC (formerly CCRC) 13661 = CIP 107109 = DSM 40034 = IFO (now NBRC) 13195 = ISP 5034 = JCM 4150 = JCM 4614 = NRRL B-3298 = NRRL ISP-5034 = VKM Ac-1081.
L. a.ta.ca.men'sis (N.L. fem. adj. atacamensis, of or pertaining to the Atacama Desert of northern Chile, the source of the soil from which the type strain was isolated).
Basonym: Lechevalieria atacamensis Okoro et al. 2010
The description is as given for Lechevalieria atacamensis (Okoro et al., 2010) with the following addition. The G+C content of the type-strain genome is 68.9%, its approximate size 9.31 Mbp, its IMG deposit 2756170238. The type strain is C61 = CGMCC 4.5536 = JCM 17492 = NRRL B-24706 = DSM 45479.
L. de.ser'ti (L. gen. n. deserti, of a desert).
Basonym: Lechevalieria deserti Okoro et al. 2010
The description is as given for Lechevalieria deserti (Okoro et al., 2010) with the following addition. The G+C content of the type-strain genome is 68.8%, its approximate size 9.53 Mbp, its IMG deposit 2756170278. The type strain is C68 = CGMCC 4.5535 = JCM 17493 = NRRL B-24707 = DSM 45480.
L. fla'va (L. fem. adj. flava, yellow, referring to the color of the substrate mycelium).
Basonym: Lechevalieria flava (Gauze et al. 1974) Labeda et al. 2001
The description is as given for Lechevalieria flava (Labeda et al., 2001) with the following addition. The G+C content of the type-strain genome is 69.0%, its approximate size 9.75 Mbp, its GenBank deposit SAMN05660974. The type strain is ATCC 29533 = BCRC (formerly CCRC) 13328 = CIP 107110 = DSM 43885 = IFO (now NBRC) 14521 = JCM 3296 = NRRL B-16131 = VKM Ac-906.
L. fra'di.ae (N.L. gen. n. fradiae, of Fradia, a patronymic).
Basonym: Lechevalieria fradiae Zhang et al. 2007
The description is as given for Lechevalieria fradiae (Zhang et al., 2007b) with the following modification. The G+C content of the type-strain genome is 70.5%, its approximate size 8.51 Mbp, its GenBank deposit SAMN05216553. The type strain is Z6 = CGMCC 4.3506 = JCM 14205 = DSM 45099.
L. ni.ge.ri'a.ca (N.L. fem. adj. nigeriaca, of or belonging to Nigeria).
Basonym: Lechevalieria nigeriaca Camas et al. 2013
The description is as given for Lechevalieria nigeriaca (Camas et al., 2013). The type strain is NJ2035 = DSM 45680 = KCTC 29057 = NRRL B-24881 = JCM 31207.
L. ro.se.ly'ni.ae (N.L. fem. gen. n. roselyniae, of Roselyn, named after Roselyn Brown for her many practical contributions to actinomycete systematics).
Basonym: Lechevalieria roselyniae Okoro et al. 2010
The description is as given for Lechevalieria roselyniae (Okoro et al., 2010). The type strain is C81 = CGMCC 4.5537 = JCM 17494 = NRRL B-24708 = DSM 45481.
L. xin.ji.an.gen'sis (N.L. fem. adj. xinjiangensis, referring to Xinjiang, north-western China, the source of the isolate).
Basonym: Lechevalieria xinjiangensis Wang et al. 2007
The description is as given for Lechevalieria xinjiangensis (Wang et al., 2007a) with the following modification. The G+C content of the type-strain genome is 70.7%, its approximate size 8.68 Mbp, its GenBank deposit SAMN05216188. The type strain is R24 = CGMCC 4.3525 = DSM 45081 = JCM 15473.
L. ja.po'ni.cus (N.L. masc. adj. japonicus, Japanese).
The description is as given for Leucobacter musarum subsp. japonicus (Clark and Hodgkin, 2015) with the following addition. The G+C content of the type-strain genome is 66.8%, its approximate size 3.59 Mbp, its GenBank deposit SAMN02673066. The type strain is CBX130 = DSM 27158 = CIP 110719 = JCM 31936.
M. ae.ro.la'tus (Gr. n. aer, air; L. part. adj. latus, carried; N.L. part. adj. aerolatus, airborne).
Basonym: Friedmanniella aerolata Kim et al. 2016
The description is as given for Friedmanniella aerolata (Kim et al., 2016). The type strain is 7515T-26 = KACC 17306 = DSM 27139.
M. an.tarc'ti.cus (M.L. masc. adj. antarcticus, isolated from Antarctica).
Basonym: Friedmanniella antarctica Schumann et al. 1997
The description is as given for Friedmanniella antarctica (Schumann et al., 1997). The type strain is AA-1042 = NBRC 16127 = DSM 11053 = KACC 14501.
M. cap.su.la'tus (L. masc. adj. capsulatus, capsulated).
Basonym: Friedmanniella capsulata Maszenan et al. 1999
The description is as given for Friedmanniella capsulata (Maszenan et al., 1999a). The type strain is Ben 108 = ACM 5120 = JCM 13522 = KACC 14261.
M. fla'vus (L. masc. adj. flavus, yellow, pertaining to the yellow color of the colonies).
Basonym: Friedmanniella flava Zhang et al. 2013
The description is as given for Friedmanniella flava (Zhang et al., 2013b) with the following modification. The G+C content of the type-strain genome is 73.5%, its approximate size 4.79 Mbp, its GenBank deposit SAMN05421756. The type strain is W6 = CGMCC 4.6856 = JCM 17701 = KACC 17380.
M. la.cus'tris (L. masc. adj. lacustris, belonging to the lake).
Basonym: Friedmanniella lacustris Lawson et al. 2000
The description is as given for Friedmanniella lacustris (Lawson et al., 2000). The type strain is EL-17A = DSM 11465 = ATCC BAA-165 = KACC 15021.
M. lu'ci.dus (L. masc. adj. lucidus, brilliant, shining, referring to the shiny colony surface).
Basonym: Friedmanniella lucida Iwai et al. 2010
The description is as given for Friedmanniella lucida (Iwai et al., 2010). The type strain is FA2 = DSM 21742 = NBRC 104964 = KACC 15053.
M. lu.te'o.lus (L. masc. adj. luteolus, referring to the yellow color of the colonies).
Basonym: Friedmanniella luteola Iwai et al. 2010
The description is as given for Friedmanniella luteola (Iwai et al., 2010) with the following modification. The G+C content of the type-strain genome is 73.4%, its approximate size 4.41 Mbp, its GenBank deposit SAMN04488544. The type strain is FA1 = DSM 21741 = NBRC 104963 = KACC 15054.
M. o.ki.na.wen'sis (N.L. masc. adj. okinawensis, pertaining to Okinawa, a prefecture in Japan, where the organism was first isolated).
Basonym: Friedmanniella okinawensis Iwai et al. 2010
The description is as given for Friedmanniella okinawensis (Iwai et al., 2010). The type strain is FB1 = DSM 21744 = NBRC 104966 = KACC 15055.
M. sa.ga.mi.ha.ren'sis (N.L. masc. adj. sagamiharensis, pertaining to Sagamihara, a city in Japan, where the organism was first isolated).
Basonym: Friedmanniella sagamiharensis Iwai et al. 2010
The description is as given for Friedmanniella sagamiharensis (Iwai et al., 2010) with the following modification. The G+C content of the type-strain genome is 73.4%, its approximate size 4.41 Mbp, its GenBank deposit SAMN04488544. The type strain is FB2 = DSM 21743 = NBRC 104965 = KACC 15056.
M. spu.mi'co.lus (L. fem. n. spuma, foam; L. suff. -colus, inhabitant of; N.L. n. spumicolus, inhabitant of foam).
Basonym: Friedmanniella spumicola Maszenan et al. 1999
The description is as given for Friedmanniella spumicola (Maszenan et al., 1999a). The type strain is Ben 107 = ACM 5121 = JCM 16540 = KACC 14247.
M. an.da.ma.nen'sis (N.L. fem. adj. andamanensis, referring to the Andaman Sea, Thailand, the source of the type strain).
Basonym: Verrucosispora andamanensis Supong et al. 2013
The description is as given for Verrucosispora andamanensis (Supong et al., 2013). The type strain is SP03-05 = BCC 45620 = NBRC 109075 = DSM 46721.
M. fie'dle.ri (N.L. fem. gen. n. fiedleri, after Hans-Peter Fiedler in recognition of his contributions to the search and discovery of new antibiotics from Actinobacteria).
Basonym: Verrucosispora fiedleri Goodfellow et al. 2013
The description is as given for Verrucosispora fiedleri (Goodfellow et al., 2013). The type strain is MG-37 = NCIMB 14794 = NRRL-B-24892 = DSM 46741 = KACC 18210.
M. gif.hor.nen'sis (N.L. fem. n. gifhornensis, referring to the city of Gifhorn, adjacent to the peat bog from which the organisms was isolated).
Basonym: Verrucosispora gifhornensis Rheims et al. 1998
The description is as given for Verrucosispora gifhornensis (Rheims et al., 1998). The type strain is HR1-2 = DSM 44337 = NBRC 16317 = JCM 10457 = KACC 20946.
M. lu'te.a (L. fem. adj. lutea, orange colored).
Basonym: Verrucosispora lutea Liao et al. 2009
The description is as given for Verrucosispora lutea (Liao et al., 2009). The type strain is CCTCC AA207012 = JCM 16959 = KCTC 19195 = DSM 45424.
M. ma'ris (L. gen. n. maris, of the sea).
Basonym: Verrucosispora maris Goodfellow et al. 2012
The description is as given for Verrucosispora maris (Goodfellow et al., 2012b). The type strain is AB-18-032 = DSM 45365 = NRRL B-24793 = JCM 31040.
M. pha.se.o'li (N.L. masc. n. Phaseolus, a genus of plants; N.L. gen. n. phaseoli, of Phaseolus, referring to the isolation of the first strains from Phaseolus vulgaris).
Basonym: Xiangella phaseoli Wang et al. 2013
The description is as given for Xiangella phaseoli (Wang et al., 2013a) with the following modification. Major polar lipids are PE, diPE, PI and PG. The type strain is NEAU-J5 = CGMCC 4.7038 = DSM 45730 = NBRC 110907.
M. qi.u.i'ae (N.L. fem. gen. n. qiuiae, of Qiu, in honor of Danheng Qiu, for her devotion to the investigation of Micromonospora-like Actinobacteria).
Basonym: Verrucosispora qiuiae Xi et al. 2012
The description is as given for Verrucosispora qiuiae (Xi et al., 2012). The type strain is RtIII47 = CGMCC 4.5826 = NBRC 106684 = DSM 45781 = JCM 19682.
M. se.di.mi.ni.ma'ris (L. n. sedimen/-inis, sediment; L. n. mare/-is, the sea; N.L. gen. n. sediminimaris, from the marine sediment).
Basonym: Verrucosispora sediminis Dai et al. 2010 (The name Micromonospora sediminis has already been validly published (Muangham et al., 2016), hence a new epithet has been chosen to avoid homonyms).
The description is as given for Verrucosispora sediminis (Dai et al., 2010) with the following modification. The G+C content of the type-strain genome is 71.0%, its approximate size 6.40 Mbp, its GenBank deposit SAMN05216284. The type strain is MS426 = CGMCC 4.3550 = JCM 15670 = DSM 45558.
M. tru.jil.lo'ni.a.e (N.L. gen. n. trujilloniae, after Martha E. Trujillo in recognition of her contributions to the discovery of new Micromonospora strains).
Basonym: Verrucosispora wenchangensis Xie et al. 2012 (The name Micromonospora wenchangensis has already been validly published (Ren et al., 2013) hence a new epithet has been chosen to avoid homonyms).
The description is as given for Verrucosispora wenchangensis (Xie et al., 2012). The type strain is 234402 = CCTCC AA 2011018 = DSM 45674 = JCM 31041.
M. hol.me'si.i (N.L. gen. n. holmesii, named after K. K. Holmes, a researcher of sexually transmitted diseases).
The description is as given for Mobiluncus curtisii subsp. holmesii (Spiegel and Roberts, 1984) with the following addition. The G+C content of the type-strain genome is 55.6%, its approximate size 2.08 Mbp, its GenBank deposit SAMN00253295. The type strain is BV376-6 = ATCC 35242 = CCUG 17762 = LMG 7786 = NCTC 11657.
P. hong.kon.gen'sis (N.L. fem. adj. hongkongensis, of or belonging to Hong Kong, where the type strain was isolated).
Basonym: Actinomyces hongkongensis Woo et al. 2004
The description is as given for Actinomyces hongkongensis (Woo et al., 2003). The type strain is HKU8 = CCUG 48484 = DSM 15629 = CIP 107949 = LMG 21939.
P. du.o.de'ca.dis (L. n. duodecas/-adis, the number twelve; L. gen. n. duodecadis, of twelve, referring to the requirement of the organism for vitamin B12).
Basonym: Tetrasphaera duodecadis (Lochhead 1958) Ishikawa and Yokota 2006
The description is as given for Tetrasphaera duodecadis (Ishikawa and Yokota, 2006). The type strain is ATCC 13347 = NCIMB 9222 = NBRC 12959.
P. e.lon.ga'tus (L. masc. adj. elongatus, elongated, pertaining to the formation of elongated clumps).
Basonym: Tetrasphaera elongata Hanada et al. 2002
The description is as given for Tetrasphaera elongata (Hanada et al., 2002a). The type strain is Lp2 = DSM 14184 = JCM 11141.
R. ha.lo.to'le.rans (Gr. n. halos, salt; L. part. adj. tolerans, tolerating, enduring; N.L. part. adj. halotolerans, salt-tolerating).
Basonym: Kocuria halotolerans Zucchi et al. 2009
The description is as given for Kocuria halotolerans (Tang et al., 2009). The type strain is CCTCC AB 206069 = DSM 18442 = KCTC 19172 = JCM 31975.
R. ko.re.en'sis (N.L. fem. adj. koreensis, pertaining to Korea).
Basonym: Kocuria koreensis Park et al. 2010
The description is as given for Kocuria koreensis (Park et al., 2010). The type strain is P31 = JCM 15915 = KCTC 19595 = DSM 23367.
R. kris.ti'nae (M.L. gen. n. kristinae, of Kristin).
Basonym: Kocuria kristinae (Kloos et al., 1974) Stackebrandt et al. 1995
The description is as given for Kocuria kristinae (Kloos et al., 1974; Stackebrandt et al., 1995) with the following modification. The G+C content of the type-strain genome is 71.9%, its approximate size 2.36 Mbp, its GenBank deposit SAMD00046484. The type strain is ATCC 27570 = CCM 2690 = CCUG 33026 = CIP 81.69 = DSM 20032 = IEGM 390 = IFO (now NBRC) 15354 = JCM 7237 = LMG 14215 = NCTC 11038 = NRRL B-14835 = VKM B-1811.
S. i.ra.qi.en'sis (N.L. fem. adj. iraqiensis, of or belonging to Iraq, the source of the soil sample from which the organism was isolated).
Basonym: Actinopolyspora iraqiensis Ruan et al. 1994
The description is as given for Actinopolyspora iraqiensis (Ruan et al., 1994) with the following addition. The G+C content of the type-strain genome is 71.5%, its approximate size 3.84 Mbp, its GenBank deposit SAMN02441411. The type strain is IQ-H1 = DSM 44640 = JCM 9891 = NBRC 103187.
S. ko.ben'sis (N.L. fem. adj. kobensis, pertaining to Kobe, a city in Japan where the organism was isolated).
The description is as given for Saccharopolyspora hirsuta subsp. kobensis (Lacey, 1989) with the following addition. The G+C content of the type-strain genome is 70.9%, its approximate size 7.74 Mbp, its GenBank deposit SAMN02982929. The type strain is ATCC 20501 = DSM 44795 = FERM-P 3912 = NBRC 15151 = JCM 9109.
S. ca'nis (L. gen. n. canis, of the dog).
Basonym: Actinomyces canis Hoyles et al. 2000
The description is as given for Actinomyces canis (Hoyles et al., 2000). The type strain is M2289/98/2 = CCUG 41706 = DSM 15536.
S. car.diff.en'sis (L. fem. adj. cardiffensis, pertaining to Cardiff, a city in Wales).
Basonym: Actinomyces cardiffensis Hall et al. 2003
The description is as given for Actinomyces cardiffensis (Hall et al., 2002). The type strain is R10394 = CCUG 44997 = CIP 107323.
S. fun'ke.i (N.L. gen. n. funkei, of Funke, to honor the microbiologist Guido Funke).
Basonym: Actinomyces funkei Lawson et al. 2001
The description is as given for Actinomyces funkei (Lawson et al., 2001). The type strain is CCUG 42773 = CIP 106713.
S. ge.or'gi.ae (N.L. gen. n. georgiae, of Georg, named in honor of Lucile Georg, a pioneer in Actinomyces taxonomy).
Basonym: Actinomyces georgiae Johnson et al. 1990
The description is as given for Actinomyces georgiae (Johnson et al., 1990) with the following restriction. The G+C content of the type-strain genome is 69.9%, its approximate size 2.50 Mbp, its GenBank deposit SAMN02441198. The type strain is ATCC 49285 = CCUG 32935 = CIP 104749 = DSM 6843.
S. hy.o.va.gi.na'lis (Gr. n. hys, pig; L. n. vagina, sheath, vagina; N.L. fem. adj. hyovaginalis, pertaining to the vagina of a pig).
Basonym: Actinomyces hyovaginalis Collins et al. 1993
The description is as given for Actinomyces hyovaginalis (Collins et al., 1993). The type strain is BM 1192/5 = ATCC 51367 = CCUG 35604 = CCUG 35715 = CIP 103923 = DSM 10695 = NCIMB 702983.
S. me'ye.ri (N.L. gen. n. meyeri, of Meyer, to honor Kurt F. Meyer, a German bacteriologist, who described the organism as a “new anaerobic Streptothrix species” in 1911).
Basonym: Actinomyces meyeri (ex Prévot 1938) Cato et al. 1984
The description is as given for Actinomyces meyeri (Cato et al., 1984) with the following modification. The G+C content of the type-strain genome is 65.6%, its approximate size 2.03 Mbp, its GenBank deposit SAMN04489715. The type strain is ATCC 35568 = CCUG 21024 = CIP 103148 = DSM 20733 = LMG 16161.
S. na.tu'rae (L. gen. n. naturae, of nature, intended to mean from the environment).
Basonym: Actinomyces naturae Rao et al. 2012
The description is as given for Actinomyces naturae (Rao et al., 2012). The type strain is BL-79 = CCUG 56698 = NRRL B-24670 = DSM 26713.
S. o.don.to.ly'ti.ca (Gr. n. odous/-ontos, tooth; N.L. fem. adj. lytica, able to dissolve; N.L. fem. adj. odontolytica, tooth-dissolving).
Basonym: Actinomyces odontolyticus Batty 1958 (Approved Lists 1980)
The description is as given for Actinomyces odontolyticus (Batty, 1958). The type strain is ATCC 17929 = CCUG 20536 = CIP 101124 = DSM 43760 = JCM 14871 = LMG 18080 = NCTC 9935.
S. ra.din'gae (M.L. masc. n. Radinga, the Latin name for Reading, UK; M.L. gen. n. radingae, of Reading).
Basonym: Actinomyces radingae Wüst et al. 1995
The description is as given for Actinomyces radingae (Wüst et al., 1995; Vandamme et al., 1998) with the following modification. The G+C content of the type-strain genome is 58.9%, its approximate size 2.43 Mbp, its GenBank deposit SAMN04489714. The type strain is APL1 = ATCC 51856 = CCUG 32394 = CCUG 34270 = CIP 105358 = DSM 9169 = LMG 15960.
S. su.i.mas.ti'ti.dis (N.L. n. sus, pig; M.L. n. mastitis, infection of the milk glands; N.L. gen. n. suimastitidis, of porcine mastitis).
Basonym: Actinomyces suimastitidis Hoyles et al. 2001
The description is as given for Actinomyces suimastitidis (Hoyles et al., 2001a) with the following addition. The G+C content of the type-strain genome is 56.4%, its approximate size 2.29 Mbp, its GenBank deposit SAMN02440611. The type strain is CCUG 39276 = CIP 106779.
S. tu.ri.cen'sis (L. fem. adj. turicensis, of Turicum, the Latin name for Zürich, referring to the place where the type strain had been isolated).
Basonym: Actinomyces turicensis Wüst et al. 1995
The description is as given for Actinomyces turicensis (Wüst et al., 1995; Vandamme et al., 1998). The type strain is APL10 = ATCC 51857 = CCUG 32401 = CCUG 34269 = CIP 105357 = DSM 9168 = LMG 15961.
S. vac.ci.ma.xil'lae (L. n. vacca, cow; L. n. maxilla, jaw; N.L. gen. n. vaccimaxillae, of the cow's jaw).
Basonym: Actinomyces vaccimaxillae Hall et al. 2003
The description is as given for Actinomyces vaccimaxillae (Hall et al., 2003b) with the following addition. The G+C content of the type-strain genome is 57.6%, its approximate size 2.34 Mbp, its GenBank deposit SAMN02441246. The type strain is R10176 = CCUG 46091 = CIP 107423.
S. ae.ro'phi.lus (Gr. n. aer, air; Gr. adj. philos, loving; N.L. masc. adj. aerophilus, air-loving).
Basonym: Phycicoccus aerophilus Weon et al. 2008
The description is as given for Phycicoccus aerophilus (Weon et al., 2008). The type strain is 5516T-20 = KACC 20658 = DSM 18548 = JCM 16378.
S. ba.di.i.sco.ri'ae (L. adj. badius, brown; L. gen. n. scoriae, of scoria, i.e., volvanic ash; N.L. gen. n. badiiscoriae, of brown-colored scoria).
Basonym: Phycicoccus badiiscoriae Lee 2013
The description is as given for Phycicoccus badiiscoriae (Lee, 2013a). The type strain is Sco-B23 = KCTC 19807 = KACC 15111 = DSM 23987 = NBRC 107918.
S. bi.geu.men'sis (N.L. masc. adj. bigeumensis, pertaining to Bigeum Island, Korea, from where the type strain was isolated).
Basonym: Phycicoccus bigeumensis Dastager et al. 2008
The description is as given for Phycicoccus bigeumensis (Dastager et al., 2008). The type strain is MSL-03 = DSM 19264 = KCTC 19266 = JCM 16023.
S. cre.me'us (N.L. masc. adj. cremeus, cream-white).
Basonym: Phycicoccus cremeus Zhang et al. 2011
The description is as given for Phycicoccus cremeus (Zhang et al., 2011). The type strain is V2M29 = CGMCC 1.6963 = NBRC 104261 = DSM 28108 = JCM 17739.
S. dok.do.nen'sis (N.L. masc. adj. dokdonensis, of Dokdo, Korea, from where the type strain was isolated).
Basonym: Phycicoccus dokdonensis Yoon et al. 2008
The description is as given for Phycicoccus dokdonensis (Yoon et al., 2008). The type strain is DS-8 = KCTC 19248 = CCUG 54521 = DSM 22329 = JCM 19120.
S. gin.se.no.si.di.mu'tans (N.L. n. ginsenosidum, ginsenoside; L. part. adj. mutans, transforming, converting; N.L. part. adj. ginsenosidimutans, ginsenoside-converting).
Basonym: Phycicoccus ginsenosidimutans Wang et al. 2011
The description is as given for Phycicoccus ginsenosidimutans (Wang et al., 2011). The type strain is BXN5-13 = DSM 21006 = KCTC 19419 = JCM 18961.
S. so'li (L. gen. n. soli, of soil).
Basonym: Phycicoccus soli Singh et al. 2015
The description is as given for Phycicoccus soli (Singh et al., 2015). The type strain is THG-a14 = KACC 17892 = JCM 19837 = DSM 103360.
T. a.my.lo.ly'ti.ca (Gr. n. amylon, starch; Gr. adj. lytikos, dissolving; N.L. fem. adj. amylolytica, starch dissolving).
Basonym: Actinomadura amylolytica Jiao et al. 2015
The description is as given for Actinomadura amylolytica (Jiao et al., 2015). The type strain is CCTCC AA 2012024 = DSM 45822 = JCM 30324.
T. cel.lu.lo.si.ly'ti.ca (N.L. n. cellulosum, cellulose; N.L. adj. lyticus, able to dissolve; N.L. fem. adj. cellulosilytica, cellulose-dissolving).
Basonym: Actinomadura cellulosilytica Jiao et al. 2015
The description is as given for Actinomadura cellulosilytica (Jiao et al., 2015). The type strain is CCTCC AA 2012023 = DSM 45823 = JCM 30326.
T. e.chi.no'spo.ra (L. n. echinus, sea urchin; Gr. fem. n. spora, seed; N.L. fem. n. echinospora, spiny spore).
Basonym: Actinomadura echinospora (Nonomura and Ohara 1971) Kroppenstedt et al. 1991
The description is as given for Actinomadura echinospora (Nonomura and Ohara, 1971a; Kroppenstedt et al., 1990) with the following addition. The G+C content of the type-strain genome is 71.8%, its approximate size 8.64 Mbp, its GenBank deposit SAMN04489712. The type strain is ATCC 27300 = CCRC 12547 = DSM 43163 = HUT 6548 = NBRC 14042 = JCM 3148 = KCTC 9313.
T. um.bri'na (N.L. fem. adj. umbrina, wood brown).
Basonym: Actinomadura umbrina Galatenko et al. 1987
The description is as given for Actinomadura umbrina (Galatenko et al., 1981) with the following addition. The G+C content of the type-strain genome is 71.9%, its approximate size 7.40 Mbp, its IMG deposit 2778260945. The type strain is ATCC 49502 = CIP 105485 = DSM 43927 = IFO (now NBRC) 14346 = INA 2309 = JCM 6837 = NRRL B-16244 = VKM Ac-1086.
W. neu'i.i (N.L. gen. n. neuii, of Neu, named to honor Harols Neu, an authority in antimicrobial chemotherapy and infectious diseases).
Basonym: Actinomyces neuii Funke et al. 1994
The description is as given for Actinomyces neuii (Funke et al., 1994b) with the following restriction. The G+C content of the type-strain genome is 56.2%, its approximate size 2.27 Mbp, its GenBank deposit SAMN02441245. The type strain is 97/90 = ATCC 51847 = CCUG 32252 = CIP 104015 = DSM 8576.
Y. a.o.mi.en'sis (N.L. fem. adj. aomiensis, of or pertaining to Aomi, the place from where the strain was isolated).
Basonym: Streptomyces aomiensis Nagai et al. 2011
The description is as given for Streptomyces aomiensis (Nagai et al., 2011). The type strain is M24DS4 = M24DS04 = JCM 17986 = KACC 14925 = NBRC 106164.
A. e.quo.li.fa'ci.ens subsp. ce.la'tus (L. masc. adj. celatus, conceal, hide, keep secret).
The description is as given for Asaccharobacter celatus (Minamida et al., 2008). The type strain is do03 = AHU 1763 = DSM 18785 = JCM 14811.
B. ca.te.nu.la'tum subsp. ka.shi.wa.no.hen'se (N.L. neut. adj. kashiwanohense, of or pertaining to Kashiwanoha in Japan).
The description is as given for Bifidobacterium kashiwanohense (Morita et al., 2011) with the following restriction. The G+C content of the type-strain genome is 56.3%, its approximate size 2.34 Mbp, its GenBank deposit SAMD00061044. The type strain is HM2-2 = DSM 21854 = JCM 15439.
B. pul.lo'rum subsp. gal.li.na'rum (L. n. gallina, a hen; L. gen. pl. n. gallinarum, of hens).
The description is as given for Bifidobacterium gallinarum (Watabe et al., 1983) with the following restriction. The G+C content of the type-strain genome is 64.2%, its approximate size 2.16 Mbp, its GenBank deposit SAMN02673435. The type strain is Ch206-5 = AS 1.2283 = ATCC 33777 = BCRC (formerly CCRC) 14679 = CCUG 34990 = DSM 20670 = JCM 6291 = LMG 11586.
B. pul.lo'rum subsp. sae.cu.la're (L. neut. adj. saeculare, of or belonging to a saeculum, commemorating the ninth centenary of the foundation of Bologna University).
The description is as given for Bifidobacterium saeculare (Biavati et al., 1991). The type strain is RA161 = AS 1.2278 = ATCC 49392 = CCUG 34977 = DSM 6531 = JCM 8223 = LMG 14934.
C. ac'nes subsp. de.fen'dens (L. part. adj. defendens, defending, guarding, protecting, referring to the CRISPR/Cas system of the strains).
Basonym: Propionibacterium acnes subsp. defendens McDowell et al. 2016
The description is as given for Propionibacterium acnes subsp. defendens (McDowell et al., 2016). The type strain is ATCC 11828 = CCUG 6369 = JCM 6473.
D. kun.ja.men'sis subsp. schi'mae (N.L. gen. n. schimae, of the plant genus Schima).
The description is as given for Dietzia schimae (Li et al., 2008a) with the following modification. The G+C content of the type-strain genome is 70.5%, its approximate size 3.65 Mbp, its IMG deposit 2724679793. The type strain is CCTCC AA 207015 = DSM 45139 = JCM 16003.
L. al.bi.do.ca.pil.la'ta subsp. vi.o.la'ce.a (L. fem. adj. violacea, violet-colored).
The description is as given for Saccharothrix violacea (Lee et al., 2000). The type strain is LM 036 = DSM 44796 = JCM 10975.
M. in.tra.cel.lu.la're subsp. chi.mae'ra (L. n. chimaera, the chimaera, the mythological being made up of parts of three different animals).
The description is as given for Mycobacterium chimaera (Tortoli et al., 2004) with the following addition. The G+C content of the type-strain genome is 67.7%, its approximate size 6.09 Mbp, its GenBank deposit SAMN04216918. The type strain is FI-01069 = CCUG 50989 = CIP 107892 = DSM 44623 = JCM 14737.
M. in.tra.cel.lu.la're subsp. yon.go.nen'se (N.L. neut. adj. yongonense, of or belonging to Yongon-Dong, the location of the department performing the taxonomic research on the species).
The description is as given for Mycobacterium yongonense (Kim et al., 2013) with the following addition. The G+C content of the type-strain genome is 67.9%, its approximate size 5.66 Mbp, its GenBank deposit SAMN02603187. The type strain is 05-1390 = DSM 45126 = KCTC 19555.
N. sal.mo.ni'ci.da subsp. cum.mi.de'lens (L. n. cummis, gum, rubber; L. part. adj. delens, destroying; N.L. part. adj. cummidelens, rubber destroying).
The description is as given for Nocardia cummidelens (Maldonado et al., 2000) with the following modification. Colony color can be brown to pale orange or pale pink. The G+C content of the type-strain genome is 67.1%, its approximate size 7.50 Mbp, its GenBank deposit SAMD00040649. The type strain is R89 = CCUG 46121 = CCUG 46295 = CIP 107225 = DSM 44490 = JCM 11439 = NBRC 100378 = NCIMB 13758.
S. i.ra.qi.en'sis subsp. pau.ro.me.ta.bo'li.ca (Gr. adj. pauros, little; Gr. adj. metabolikos, changeable; N.L. fem. adj. paurometabolica, little changeable, referring to the poor utilization of carbon sources).
The description is as given for Saccharomonospora paurometabolica (Li et al., 2003e). The type strain is BCRC (formerly CCRC) 16315 = CCTCC AA 001018 = DSM 44619 = JCM 13241.
W. neu'i.i subsp. a.ni.tra'ta (Gr. pref. a, not; N.L. n. nitratus, nitrate; N.L. fem. adj. anitrata, not reducing nitrate).
Basonym: Actinomyces neuii subsp. anitratus Funke et al. 1994
The description is as given for Actinomyces neuii subsp. anitratus (Funke et al., 1994b). The type strain is 50/90 = ATCC 51849 = CCUG 32253 = CIP 104016 = DSM 8577 = LMG 14788.
The original proposal of the class “Actinobacteria” (Stackebrandt et al., 1997) did not designate a type order hence this name is not validly published under Rule 27 of the International Code of Nomenclature of Prokaryotes (Parker et al., 2015). However, since this name is in common currency it is used here. The type order is likely to be Actinomycetales Buchanan 1917 162AL emend. Zhi et al. 2009, 594, a move that would require the approval of the Judicial Commission. Actinobacteria show amazingly diverse morphologies that range from cocci (e.g., Lapillicoccus and Micrococcus), short rods (e.g., Mycobacterium and Tropheryma), irregular rods (e.g., Arcanobacterium and Cellulomonas), rods and cocci (e.g., Arthrobacter and Brevibacterium) to mycelia that fragment into coccoid and rod-like forms (e.g., Nocardia and Promicromonospora). Others show more pronounced morphological differentiation which ranges from extensively branched substrate hyphae that carry spores (e.g., Catellatospora and Micromonospora) or spores vesicles (e.g., Actinoplanes and Dactylosporangium) to strains that produce a stable branched substrate mycelium which bears aerial hyphae which differentiate into long chains of spores (e.g., Nonomuraea and Streptomyces) or into spore vesicles (e.g., Planotetraspora and Streptosporangium). In general, spores are non-motile though ones released from spore vesicles tend to be motile. Actinobacteria comprise the orders Actinomycetales, Bifidobacteriales, Catenulisporales, Corynebacteriales, Cryptosporangiales, Frankiales, Glycomycetales, Jiangellales, Kineosporiales, Micrococcales, Micromonosporales, Propionibacteriales, Pseudonocardiales, Sporichthyales, Streptomycetales and Streptosporangiales.
The description is as given before (Goodfellow and Jones, 2012) with the following additions. Corynebacteriales are found in diverse habitats, notably in the soil ecosystem. Some strains are serious pathogens of humans (e.g., Mycobacterium tuberculosis), animals (e.g., Rhodococcus equi = R. hoagii) and plants (e.g., Rhodococcus fascians). The order comprises the families Corynebacteriaceae, Dietziaceae, Gordoniaceae, Lawsonellaceae, Mycobacteriaceae, Nocardiaceae, Segniliparaceae and Tsukamurellaceae. The type genus is Corynebacterium.
The description is as given before (Prévot, 1940) with the following additions. The isoprenoid quinone systems are composed of menaquinones with 6–14 isoprenoid units in the side chain; one or two subunits may be saturated. The order consists of the families Micrococcaceae, Beutenbergiaceae, Bogoriellaceae, Brevibacteriaceae, Cellulomonadaceae, Demequinaceae, Dermabacteraceae, Dermacoccaceae, Dermatophilaceae, Intrasporangiaceae, Jonesiaceae, Kytococcaceae, Microbacteriaceae, Micrococcaceae, Ornithinimicrobiaceae, Promicromonosporaceae, Rarobacteraceae and Ruaniaceae. The new families assigned to this order are Kytococcaceae, Ornithinimicrobiaceae and Tropherymataceae: in contrast Sanguibacteraceae has been assigned to the family Jonesiaceae. The type genus is Micrococcus.
The description is as given before (Patrick and McDowell, 2012) with the following additions. Cellular fatty acids are composed of different combinations of straight-chain saturated and iso-and anteiso-methyl branched acids; mycolic acids are absent. The order consists of the established families Nocardioidaceae and Propionibacteriaceae as well as Actinopolymorphaceae fam. nov. and Kribbellaceae fam. nov. The type genus is Propionibacterium.
The description is as given before (Goodfellow, 2012b) with the following additions. Some strains are thermophilic. The order contains the families Nocardiopsaceae, Streptosporangiaceae and Thermomonosporaceae. The type genus is Streptosporangium.
The description is as given before (Zhi et al., 2009) together with extensive additions made earlier (Schaal and Yassin, 2012b) and as a result of the present study. In addition to the revised type genus Actinomyces, the family contains the established genera Actinobaculum, Arcanobacterium, Mobiluncus and Varibaculum and seven novel genera, namely Boudabousia, Bowdenia, Buchananella, Gleimia, Pauljensenia, Schaalia and Winkia.
The description is as given before (Gupta et al., 2013) with the following additions. Obligately or facultative anaerobe, Gram-stain-positive, non-motile, catalase-negative organisms that form short rods, which often show central swellings or small cocci which may be elliptical. The main fermentation products from glucose are lactic acid with acetic and fumaric acids. Growth is enhanced by Tween 80. The G+C content is around 35–46%. The family includes the type genus Atopobium, Olsenella and the novel genera Fannyhessea and Lancefieldella.
The description is as given before (Zhi et al., 2009) with a number of additions. Aerobic, Gram-stain-positive, asporogenous organisms which form slender, irregular rods; some rods are arranged at an angle to one another giving V-shaped formations. Strains typically show anaerobic growth in static cultures. Chemoorganotrophic metabolism is primarily respiration but may be fermentative. Most strains form acid from glucose both aerobically and anaerobically. Opaque, usually convex, white, yellowish or yellow colonies formed on peptone-yeast extract agar at neutral pH. Most strains are cellulolytic. The peptidoglycan contains L-lysine as the L-diamino acid; the predominant isoprenologue is MK-9(H4). The G+C content is around 70%. The type and only genus is Cellulomonas.
The description is as given before (Zhi et al., 2009) with the following additions. Gram-stain-positive, non-acid fast, non-motile, asporogenous, catalase-positive organisms which form straight to slightly curved rods with tapered ends. Club-shaped elements may be seen. Most species are facultatively anaerobic, others are aerobic. Whole-cell hydrolysates contain DL-A2pm, arabinose and galactose. The muramic acid residues of the peptidoglycan are N-acetylated. The peptidoglycan type is A1α with directly cross-linked DL-A2pm. The predominant isoprenologues are MK-8(H2) and MK-9(H2) or a mixture of these components. The major fatty acids are C16:0 and C18:1 ω9c; tuberculostearic acid may be present. Mycolic acids have 22–38 carbon atoms though some species lack these compounds. The G+C content is around 45–75%. The type and only genus is Corynebacterium, now that Turicella has been merged with Corynebacterium.
The emended description is drawn from several sources (Ue et al., 2011). Aerobic or facultatively anaerobic, Gram-stain-positive, non-acid fast, asporogenous, non-motile, catalase-positive organisms that form rods though ovoid to coccoid cells may be present in older cultures. Whole-cell hydrolysates contain arabinose, galactose and mannose. The peptidoglycan has either L-lysine or L-ornithine as the diamino acid though some species have both components; the peptidoglycan type is either A4α or A4β. The predominant isoprenologue is demethylmenaquinone DMK-9(H2), and fatty-acid profiles consist mainly of iso- and anteiso-branched and straight-chain components with anteiso-C15:0 as the main acid. The principal polar lipids are DPG, PG and PI. The G+C content is around 60–75%. The type and only genus is Demequina, now that Lysinimicrobium has been merged with Demequina.
The description of this family needs to be radically changed since the previous one (Ruckmani et al., 2011) given the addition of eight genera and the reclassification of the genus Kytococcus in Kytococcaceae. The revised family encompasses aerobic to microaerophilic, Gram-stain-positive, non-acid-fast, asporogenous, non-motile, predominately catalase-positive organisms that occur either as cocci or short rods. The peptidoglycan is of the A4α type with a L-lysine-glycine-L-serine-D-aspartic acid interpeptide bridge. The peptidoglycan is N-acetylated. Whole-cell hydrolysates contain L-lysine, arabinose, galactose, glucose, mannose, rhamnose, ribose may occur. The major fatty acids are anteiso-, methyl- and iso-methyl branched components though straight-chain saturated and monounsaturated acids may occur. The common polar lipids are DPG, PG, and PI; some genera also contain PE, PS and PIMs. Diverse menaquinones are found though MK-8(H4), MK-8(H4) and MK-8(H6) are the most frequently found isoprenologues. The G+C content is around 65-80%. The family includes Dermacoccus, the type genus, Barrientosiimonas, Branchiibius, Calidifontibacter, Demetria, Dermacoccus, Flexivirga, Luteipulveratus, Rudaeicoccus, Tamlicoccus, and Yimella.
The description of this family needs to be revised since the last emendation (Zhi et al., 2009) given the addition of five genera. It includes aerobic to facultatively anaerobic, Gram-stain-positive, acid-fast-negative, catalase-positive organisms which form cocci or branched mycelia; the mature mycelia of dermatophili undergo transverse and longitudinal divisions leading to the production of zoospores which germinate into filaments. Cocci may be motile or non-motile. The peptidoglycan contains DL-A2pm as the diamino acid. Whole-cell hydrolysates contain mannose and ribose, the predominant isoprenologue is either MK-8(H2) or MK-8(H4); fatty acids are mainly unsaturated and straight-chain components. The major polar lipids are DPG, PG, and PI; PE and lyso-PE may occur. The G+C content is around 60–75%. The family includes Dermatophilus, the type genus, Arsenicicoccus, Austwickia, Kineosphaera, Mobilicoccus, Piscicoccus and Tonsilliphilus.
The description is as given before (Gupta et al., 2013) with changes reflecting the merging of some genera. The family includes obligatory anaerobic, Gram-stain-positive, asporogenous, asaccharolytic organisms which form cocci and/or short rods. Cells may be motile or non-motile. The peptidoglycan contains DL-A2pm as the diamino acid, the predominant isoprenologue is MK-6; fatty acids are mainly saturated and unsaturated and common polar lipids are DPG and PG. The G+C content is around 55–65%. The family includes Eggerthella, the type genus, Adlercreutzia, Cryptobacterium, Denitrobacterium, Gordonibacter, Paraeggerthella and Slackia.
The description is as given before (Stackebrandt et al., 1997) with a number of additional criteria. Aerobic, Gram-stain-positive, asporogenous, non-motile, catalase-positive organisms which form short rods and cocci; elementary branched hyphae that fragment into rod and coccoid-like elements may occur. Whole-organism hydrolysates contain DL-A2pm, arabinose and galactose. The peptidoglycan type is A1α. The muramic acid residues of the peptidoglycan are N-glycolated. The predominant isoprenologue is either MK-9(H2) or MK-9 and the common major polar lipids are DPG, PE and PI; PG and PIMs may occur. The fatty-acid profiles contain major amounts of straight-chain saturated, unsaturated and 10-methyloctadecanoic (tuberculeostearic acids). Mycolic acids have 46–70 carbon atoms. The G+C content is around 60–80%. This family includes Gordonia, the type genus, Jongsikchunia gen. nov. and Williamsia.
The description is as given before (Zhi et al., 2009) with the following additions due to the inclusion of recently described genera and the transfer of Arsenicicoccus to Dermatophilaceae and that of Ornithinimicrobium and Serinicoccus to Ornithinimicrobiaceae; Monashia has been merged with Intrasporangium. The revised family encompasses aerobic and facultatively anaerobic, Gram-stain-positive, asporogenous organisms that typically form short rods and cocci though some strains form branched mycelia, others cocci and some a rod-coccus cycle. Generally non-motile (some strains of Terrabacter are motile). The peptidoglycan contains either LL- or DL-A2pm as diamino acid. The predominant isoprenologue is typically MK-8(H4) but major amounts of MK-8 may also be found; Aquipuribacter has MK-10(H4) as the major component. Common polar lipids are DPG and PI though PE and PG may occur. Fatty-acid profiles consist of major amounts of iso- and anteiso-branched-chain acids together with saturated and unsaturated straight-chain components. Mycolic acids are absent. The G+C content is around 65–75%. The family includes Intrasporangium, the type genus, Aquipuribacter, Fodinibacter, Humibacillus, Humihabitans, Intrasporangium, Janibacter, Knoellia, Kribbia, Lapillicoccus, Marihabitans, Ornithinibacter, Ornithinicoccus, Oryzihumus, Oryzobacter, Phycicoccus, Terrabacter, Terracoccus, Tetrasphaera, and Pedococcus gen. nov.
The description is as given before (Zhi et al., 2009) together with additions that reflect the inclusion of additional genera. The family includes aerobic and facultative anaerobic, Gram-stain-positive, asporogenous, motile organisms which form irregular shaped short rods. The peptidoglycan type is A4α with L-lysine as the diamino acid. The major fatty acids are anteiso-C15:0 and C16:0; the predominant isoprenologue is typically MK-9 and the common major polar lipids are DPG, PG, and PI. The G+C content is around 55–75%. The family includes Jonesia, the type genus, Populibacterium, Sanguibacter and Flavimobilis gen. nov.
The description is as given before (Zhi et al., 2009) together with changes that reflect the merging of some genera. The family encompasses aerobic, Gram-stain-positive, non-acid-fast, mesophilic organisms that form a stable branched and septate mycelium that rarely carries scant aerial hyphae though Actinocatenispora forms aerial hyphae that differentiate into chains of around 10 spores. Single spores, short spore chains or spore vesicles are formed on substrate hyphae; spores may be motile with tuffs of polar flagellae and are either smooth or ornamented. The peptidoglycan typically contains DL-A2pm (some taxa have L-lysine or 3-OH A2pm as the major diamino acid). The muramic acid residues of the peptidoglycan are N-glycolated. Predominant isoprenologues may include all types in the MK-9 and MK-10 series. PE is an ubiquitous polar lipid though DPG, PG, PI and PIMs may occur. Saturated iso- and anteiso-branched fatty acids predominate in nearly all of the genera. Mycolic acids are absent. The G+C content is around 65–75%. The family includes Micromonospora, the type genus, Actinaurispora, Actinocatenispora, Actinoplanes, Allocatelliglobosispora, Amorphosporangium, Ampullariella, Asanoa, Catellatospora, Catelliglobosispora, Catenuloplanes, Couchioplanes, Dactylosporangium, Hamadaea, Krasilnikovia, Longispora, Luedemannella, Mangrovihabitans, Phytohabitans, Phytomonospora, Pilimelia, Planopolyspora, Planosporangium, Plantactinospora, Polymorphospora, Rhizocola, Rugosimonospora, Salinispora, Spirilliplanes, and Virgisporangium.
The description is as given before (Zhi et al., 2009) with a number of additions. The family contains aerobic to mesophilic, Gram-stain-positive, acid-fast, non-motile, asporogenous organisms which form slightly curved or straight rods though some strains produce branched filaments. The peptidoglycan is of the A1α type. The muramic acid residues of the peptidoglycan are N-glycolated. Whole-cell hydrolysates contain DL-A2pm, arabinose and galactose. The predominant isoprenologue is MK-9(H2), the major polar lipids are DPG, PE, PI, and PIMs and the main fatty acids straight-chain saturated, unsaturated and 10-methyloctadecanoic (tuberculostearic) acids. Mycolic acids have 60–90 carbon atoms; fatty acid esters released on pyrolysis mass spectrometry (MS) of mycolic acid esters have 22–26 carbon atoms. The G+C content is around 55–75%. The type and only genus is Mycobacterium.
The description is as given before (Zhi et al., 2009) with changes following the inclusion of Aldersonia, Hoyosella and Tomitella and the exclusion of Gordonia (Stackebrandt et al., 1988) and Williamsia (Kämpfer et al., 1999). The family encompasses strictly aerobic and facultatively aerobic, Gram-stain-positive to Gram-stain variable, non-motile, catalase-positive organisms some of which form a extensively branched substrate mycelium that fragments into coccoid to rod-shaped elements, others are coccoid or rod-shaped or show a rod-coccus, rod-coccus-mycelium growth cycle. Strains are typically acid-alcohol or partially acid-alcohol fast at some stage of the growth cycle. Chemo-organotrophic with an oxidative metabolism. The pepetidoglycan is of the A1α type. The muramic acid residues of the peptidoglycan are N-glycolated. Predominant isoprenologues are MK-8(H2) or MK-9(H2) or MK-8(H4); major polar lipids are DPG PE, PI and PIMs, and fatty acids are mainly straight-chain saturated, unsaturated and 10-methyloctadecanoic (tuberculostearic) acids. Mycolic acids have between 34 and 64 carbon atoms; fatty-acid esters released on pyrolysis MS of mycolic acid esters have 12–18 carbon atoms. The G+C content is about 60–75%. The family includes Nocardia, the type genus, Millisia, Rhodococcus, Skermania, Tomitella and Aldersonia gen. nov.
The description of the family requires substantial modifications from that given previously (Zhi et al., 2009), notably the addition of Mumia and the reclassification of Actinopolymorpha in Actinopolymorphaceae and that of Kribbella, Flindersiella, Tenggerimyces, and Thermasporomyces in Kribbellaceae. Nocardioidaceae include aerobic, Gram-stain-positive, non-acid-fast organisms which form branched substrate hyphae or irregular rods or cocci; substrate hyphae and aerial hyphae, when formed, fragment into rod-like and coccoid cells. Rod-shaped cells may be motile. The peptidoglycan contains LL-A2pm and glycine as amino acids and is of the A3α type; muramic acid residues of the peptidoglycan are N-acetylated; cell walls contain teichoic acids. The predominant isoprenologue is MK-8(H4), the common polar lipids are DPG and PG though PC, PE, PI and PIMs may occur; the fatty acids consist of saturated and monounsaturated straight-chain and iso-, anteiso- and 10-methyl-branched components. The G+C content is around 65-75%. The family includes Nocardioides, the type genus, Marmoricola and Mumia.
The description is as given before (Zhi et al., 2009) with the following additions. The family includes aerobic to facultatively anaerobic, Gram-stain positive, acid-fast-negative organisms which form a substrate mycelium that fragments into irregular elements or presents as cocci or short rods. The peptidoglycan is of the A4α type. Whole-cell hydrolysates contain L-lysine as the diamino acid and combinations of different sugars. The predominant menaquinone is MK-9(H2) or MK-9(H4) sometimes with MK-8(H2) or MK-8(H4), and major polar lipids are DPG and PG; PI and PIMs may occur. Branching fatty acids are of the iso- and anteiso-type, notably iso-C15:0 and anteiso-C15:0. Mycolic acids are absent. The G+C content is around 70-80%. The family includes Promicromonospora, the type genus, Cellulosimicrobium, Isoptericola, Luteimicrobium, Krasilnikoviella, Myceligenerans, Oerskovia, Paraoerskovia, Xylanibacterium, Xylanimicrobium, and Xylanimonas.
The description is as given before (Labeda et al., 2011) with the following additions. Cell-wall hydrolysates contain DL-A2pm as diaminoacid and galactose or arabinose as whole-cell sugars. PE or DPG, PC or PG and lyso-PG are evident in polar-lipid patterns. Fatty acids are composed of straight-chain saturated and monounsaturated and iso-and anteiso-branched compounds. The family includes Pseudonocardia, the type genus, Actinoalloteichus, Actinobispora, Actinokineospora, Actinomycetospora, Actinophytocola, Actinosynnema, Alloactinosynnema Allokutzneria Amycolata, Amycolatopsis, Crossiella, Faenia, Goodfellowiella, Haloechinothrix, Kibdelosporangium, Kutzneria, Labedaea, Lentzea, Longimycelium, Prauserella, Pseudoamycolata, Pseudonocardia, Saccharomonospora, Saccharopolyspora, Saccharothrix, Sciscionella, Streptoalloteichus, Thermobispora, Thermocrispum, and Umezawaea. Lechevalieria and Yuhushiella have been merged with Lentzea and Amycolatopsis, respectively.
The description is as given before (Huang et al., 2017) with changes following the addition of several new genera. The family encompasses aerobic, Gram-stain-positive, non-acid-alcohol-fast, non-motile organisms that form a stable, extensively branched substrate mycelium and aerial hyphae that differentiate into two to many spores. Short chains of spores may be observed on substrate hyphae. Most strains produce a wide range of pigments which are responsible for the color of substrate and aerial mycelia. Strains are generally chemoautotrophic and mesophilic but some are autotrophic and thermophilic. The peptidoglycan of the substrate mycelium contains either LL-or DL-A2pm as the predominant diamino acid, aerial hyphae and submerged spores have LL-A2pm; the peptidoglycan type is A3α. The predominant isoprenologues are typically MK-9(H6, H8), common polar lipids are DPG and PIMs though PE and PI also occur; fatty acids are rich in saturated and iso- and anteiso-compounds. The G+C content is around 65–75%. The family contains Streptomyces, the type genus, Allostreptomyces, Kitasatospora and Streptacidiphilus and the novel genera Embleya and Yinghuangia.
The description is as given before (Zhi et al., 2009) with changes reflecting inclusion of additional genera, notably Pauljensenia and Thermobispora. The family includes aerobic, Gram-stain-positive, non-acid-alcohol-fast, chemoorganotrophic organisms which form a stable, branched susbstrate mycelium. When produced, aerial hyphae differentiate into either two or more spores or into spore vesicles that enclose one to many spores. Spores may be motile or non-motile. Most strains are mesophilic but some are thermophilic. The muramic acid residues of the peptidoglycan are N-acetylated. The peptidoglycan contains DL-A2pm and is of the A2α type. Predominant isoprenologues include all types in the MK-9 series, PE occurs as polar lipid; cells contain major amounts of iso-, anteiso-saturated, unsaturated and 10-methyl-branched components. Mycolic acids are absent. The G+C content is around 60–80%. The family contains Streptosporangium, the type genus, Acrocarpospora, Herbidospora, Microbispora, Microtetraspora, Nonomuraea, Planobispora, Planomonospora, Planotetraspora, Pauljensenia, Sphaerisporangium, Thermoactinospora, Thermobispora, Thermocatellispora, and Thermopolyspora.
The description is as given before (Tang et al., 2012) with additions following the inclusion of Alloactinosynnema. The ubiquitous whole-cell sugar is galactose but strains can also contain arabinose, galactose, mannose or ribose. The polar-lipid profile includes DPG, PE, PG, PI, or PC. Cells can be filamentous or rod shaped. The G+C content is about 65–75%. The type species is Actinokineospora riparia.
The description is as given before (Stackebrandt and Kroppenstedt, 1987) with additions following the inclusion of Pseudosporangium. Whole-cell hydrolysates are rich in arabinose, galactose, glucose, mannose, ribose and xylose or arabinose, galactose and xylose; fatty-acid profiles contain iso-C15:0, C18:1 ω9c and C16:0 or iso-C15:0 and iso-C16:0; the predominant menaquinone is either MK-9(H4) or MK-9(H6). The DNA G+C content is around 69–74%. The type species is Actinoplanes philippinensis.
The description of the genus needs to be revised since the initial description (Harz, 1877) following the assignment of certain Actinomyces species to novel genera. Aerobic to facultatively anaerobic, Gram-stain-positive, asporogenous, non-motile organisms which tend to form short straight to slightly curved rods that occur singly or in pairs or as Y-, V-, or T-forms; filaments and branched rods may be evident; some strains present as cocci or coccoid rods. The peptidoglycan contains either L-lysine or L-ornithine as the diamino acid. Whole-cell hydrolysates contain 6-deoxytalase, fucose, galactose and glucose. MK-10 is the predominant isoprenologue and PC the predominant polar lipid. The G+C content is 57–71%. The type species is Actinomyces bovis.
The description is as given before (Maruo et al., 2008) with changes reflecting the addition of Asaccharobacter, Enterorhabdus and Parvibacter. Whole-cell hydrolysates contain DL-or LL-A2pm as the diamino acid and galactose with either ribose or glucose. The predominant isoprenologues are methylmenaquinone-6 and demethylmenaquinone-6. Major fatty acids are either C16:0, iso-C15:0 or C16:0, iso-C17:0 and C18:1 ω9c. The G+C content is about 60–70%. The type species is Adlercreutzia equolifaciens.
The description is as given before (Lee, 2009) with additions that reflect developments in the composition of the genus, notably the removal of A. halophila (Tang et al., 2010a) and A. salitolerans (Guan et al., 2012) and the inclusion of Yuhushiella (Mao et al., 2011). The revised genus contains aerobic to facultatively anaerobic organisms that form branching substrate hyphae that fragment into squarish and rod-shaped elements. When formed, aerial hyphae may be sterile or differentiated into chains of smooth surfaced, squarish to ellipsoidal spore-like structures. The peptidoglycan is of the A1α type, and muramic acid moieties are N-acetylated. Whole-cell hydrolysates are rich in DL-A2pm, arabinose and galactose and the major polar lipids DPG, PE, PG, PI and PME; the fatty acids consist of complex mixtures of saturated and branched-chain components. The G+C content is about 65-75%. The type species is Amycolatopsis orientalis.
The description is as given before (Bernard et al., 2010) with additions that reflect developments in the composition of the genus, notably the inclusion of the genus Turicella (Funke et al., 1994a). The emended description is the same as that given earlier for the family Corynebacteriaceae. The type species is Corynebacterium diphtheriae.
The description is as given before (Park et al., 2016) with additions following the inclusion of Lysinimicrobium. The emended description is as given earlier for the family Demequinaceae. The type species is Demequina aestuarii.
The description is as given before (Park et al., 2012) with additions following the inclusion of Zimmermannella bifida and Z. faecalis. The revised genus encompasses irregular, non-motile, non-acid-fast, irregular rod-shaped cells that have a tendency to form short filaments and branched elements. Cell-wall peptidoglycan contains DAB or D-ornithine as a diamino acid. Whole-cell hydrolysates contain mainly glucose and ribose or rhamnose; predominant menaquinones are MK-8, MK-9, and MK-10, and fatty acid profiles are rich in C16:0, iso-C16:0, anteiso-C15:0, iso-C16:0, anteiso-C17:0 and anteiso-C15:0. The G+C content is around 60–70%. The type species is Gulosibacter molinativorax.
The description is as given before (Tang et al., 2010b) with additions following the inclusion of Amycolatopsis halophila and A. salitolerans. Substrate mycelium fragments into squarish rod-like elements; aerial mycelium may differentiate into long chains of spore-like elements at maturity. Whole-cell hydrolysates are rich in galactose and glucose (ribose is variable) or in glucose, glucosamine and mannose. Common phospholipids are DPG, PE, OH-PE, PI, PIM, and PL or DPG, PME, PE and glucosamine-containing PL or DPG, PG, PE, and PIM. Major fatty acids are iso-C16:0 or C16:0; mycolic acids are absent. The G+C content is around 65–70%. The type species is Haloechinothrix alba.
The description is as given before (Yang et al., 2012) with additions following the inclusion of Monashia flava. Produces irregular coccoid to short rods or a branching substrate mycelium that fragments into pleomorphic elements. Does not form aerial mycelium. Whole-cell hydrolysates are rich in mannose with lower amounts of galactose and rhamnose. The cellular fatty-acid profile is dominated by iso-C14:0, iso-C15:0, anteiso-C15:0 and iso-C16:0 or iso-C15:0, anteiso-C15:0, and iso-C16:0 components. The G+C content is around 65–75%. The type species is Intrasporangium calvum.
The description is as given before (Zhang et al., 1997) with the following additions. Whole-cell hydrolysates are rich in galactose or ribose, glucose and mannose; the predominant isoprenologues are MK-9(H4, H6, H8) or MK-9(H6, H8); the major polar lipids DPG, PE, PI and PIMs; fatty acids are rich in saturated, iso- and anteiso-components. The G+C content is around 65–80%. The type species is Kitasatospora setae.
The description is as given before (Groth et al., 2002) with additions following the inclusion of Tetrasphaera remsis. Cells are irregular rods and cocci that occur, singly, in pairs, tetrades, clusters and as short-rods. Fatty-acids are mainly iso- and anteiso-branched components. Mycolic acids are absent. The G+C content is around 65–75%. The type species is Knoellia sinensis.
The description is as given before (Fang et al., 2017) with additions following the inclusion of Lechevalieria. Aerial mycelium is produced but may be scant. The G+C content is around 65-80%. The type species is Lentzea albidocapillata.
The description is as given before (Nakamura et al., 1995) with additions following the inclusion of Friedmanniella. Coccoid or spherical cells occur singly, in pairs and sometimes in clusters. Major fatty acids are anteiso-C15:0 and iso-C15:0; iso-C16:0 may occur. The G+C content is around 60–75%. The type species is Microlunatus phosphovorus.
The initial description (Ørskov, 1923) has to be changed dramatically (Gao et al., 2014) given the inclusion of many additional Micromonospora species as well as Verrucosispora and Xiangella. The genus contains aerobic to microaerophilic, Gram-stain-positive, non-acid-fast organisms that form a well developed, branched, substrate mycelium. Non-motile spores are borne singly and are either sessile or carried on short or long sporophores. Aerial hyphae are usually absent. The peptidoglycan contains DL-A2pm and/or hydroxy-A2pm and muramic acid residues of the peptidoglycan are N-glycolated. Whole-cell hydrolysates are rich in xylose with either mannose or galactose, glucose and mannose; predominant isoprenologues are MK-9(H4, H6), MK-10(H4, H6) or MK-12(H2) and common major polar lipids PE and PIMs; DPG, PI and PS may occur. The fatty-acid profile contains iso-C16:0 and iso-C15:0 or iso-C15:0 and C16:0. Mycolic acids are absent. The G+C content is around 65–75%. The type species is Micromonospora chalcea.
The description is as given before (Zhang et al., 2011) with the following changes. The major cell-wall diamino acid is DL-DAP acid or 3-hydroxy DL-DAP. Galactose and glucose are the main whole-cell sugars. The G+C content is around 70-75%. The type species is Phycicoccus jejuensis.
The initial description of the genus (Georg and Brown, 1967) has been changed given the inclusion of additional Rothia species as well as Kocuria halotolerans, K. koreensis and K. kristinae. The revised genus contains Gram-stain-positive, acid-fast-negative, asporogenous organisms that may be coccoid, diphtheroid or filamentous or a mixture of these forms. The peptidoglycan contains L-lysine, as the diamino acid and is of the A3α type. Menaquinone patterns vary though most strains have MK-6(H2), MK-7, MK-8, MK-9(H2) components either singly or in combination, though others have MK-7(H2) and in some cases MK-8(H2) as the predominant isoprenologue. The major fatty acids are anteiso- and iso-methyl-branched though straight-chain saturated and mono-unsaturated components may occur; mycolic acids are absent. The G+C content is around 50-75%. The type species is Rothia dentocariosa.
The last description of the genus (Zhang et al., 1998) needs to be revised following the exclusion of Thermomonospora chromogena and the inclusion of Actinomadura amylolytica, A. cellulosilytica, A. echinospora and A. umbrina. The revised genus encompasses aerobic, Gram-stain-positive, non-acid-fast, chemoorganotrophic organisms which form a branched substrate mycelium that carries aerial hyphae. Single or short chains of spores formed on the aerial mycelium have either a spiny or rugose ornamentation; spores may be borne on sporophores. The temperature range for growth is 25–55°C. The diamino acid of the peptidoglycan is DL-A2pm. Whole-cell hydrolysates may contain glucose, madurose, mannose and rhamnose; ribose, xylose, madurose, galactose and glucose; or just ribose. The predominant isoprenologues are MK-9(H2, H4, H6, H8) and the major polar lipids are DPG, PI, PG, and PIM. Fatty-acid profiles are rich in straight-chain, iso-and anteiso-components. Mycolic acids are absent. The G+C contant is around 65–75%. The type species is Thermomonospora curvata.
The description is as before (Pukall et al., 2006) with the following modification. The G+C content of the type-strain genome is 62.3%, its approximate size 2.42 Mbp, its GenBank deposit SAMN02441381.
The description is as before (Clark and Norris, 1996) with the following restriction. The G+C content of the type-strain genome is 68.3%, its approximate size 2.16 Mbp, its GenBank deposit SAMN02598466.
The description is as before (Scholz and Kilian, 2016) with the following addition. The G+C content of the type-strain genome is 68.8%, its approximate size 3.59 Mbp, its GenBank deposit SAMN02441222.
The description is as before (Scholz and Kilian, 2016) with the following addition. The G+C content of the type-strain genome is 68.7%, its approximate size 3.03 Mbp, its GenBank deposit SAMN02441538.
The description is as before (Scholz and Kilian, 2016) with the following addition. The G+C content of the type-strain genome is 68.0%, its approximate size 2.94 Mbp, its GenBank deposit SAMN02440839.
The description is as before (Mohagheghi et al., 1986) with the following restriction. The G+C content of the type-strain genome is 66.9%, its approximate size 2.44 Mbp, its GenBank deposit SAMN02598358.
The description is as before (Zhang et al., 2006) with the following addition. The G+C content of the type-strain genome is 68.1%, its approximate size 6.31 Mbp, its GenBank deposit SAMN03295676.
The description is as before (Yassin et al., 2015) with the following modification. The G+C content of the type-strain genome is 57.8%, its approximate size 2.20 Mbp, its GenBank deposit SAMN05421878.
The description is as before (Tamura et al., 1995) with the following modification. The G+C content of the type-strain genome is 70.3%, its approximate size 8.11 Mbp, its GenBank deposit SAMN03080616.
The description is as before (Tamura et al., 1995) with the following modification. The G+C content of the type-strain genome is 70.8%, its approximate size 7.64 Mbp, its GenBank deposit SAMN03080607.
The description is as before (Tamura et al., 1995) with the following modification. The G+C content of the type-strain genome is 70.2%, its approximate size 7.28 Mbp, its GenBank deposit SAMN02441427.
The description is as before (Kämpfer et al., 2015) with the following addition. The G+C content of the type-strain genome is 72.8%, its approximate size 7.53 Mbp, its GenBank deposit SAMN02382011.
The description is as before (Tamura et al., 1995) with the following modification. The G+C content of the type-strain genome is 71.0%, its approximate size 7.58 Mbp, its GenBank deposit SAMN04487818.
The description is as before (Miyadoh et al., 1987) with the following modification. The G+C content of the type-strain genome is 73.7%, its approximate size 6.71 Mbp, its GenBank deposit SAMN02441315.
The description is as before (Qin et al., 2009b) with the following modification. The G+C content of the type-strain genome is 73.3%, its approximate size 6.17 Mbp, its GenBank deposit SAMN02441028.
The description is as before (Tomita et al., 1990) with the following addition. The G+C content of the type-strain genome is 72.4%, its approximate size 9.02 Mbp, its GenBank deposit SAMD00046515.
The description is as before (Horan and Brodsky, 1982) with the following addition. The G+C content of the type-strain genome is 73.2%, its approximate size 10.26 Mbp, its GenBank deposit SAMD00046512.
The description is as before (Huang, 1980) with the following addition. The G+C content of the type-strain genome is 70.7%, its approximate size 9.05 Mbp, its GenBank deposit SAMD00046507.
The description is as before (Lechevalier and Lechevalier, 1970) with the following addition. The G+C content of the type-strain genome is 72.1%, its approximate size 10.25 Mbp, its GenBank deposit SAMN04489713.
The description is as before (Quintana et al., 2003) with the following addition. The G+C content of the type-strain genome is 72.3%, its approximate size 8.79 Mbp, its IMG deposit 2724679706.
The description is as before (Quintana et al., 2003) with the following addition. The G+C content of the type-strain genome is 72.9%, its approximate size 8.57 Mbp, its GenBank deposit SAMN05443665.
The description is as before (Mertz and Yao, 1986) with the following addition. The G+C content of the type-strain genome is 72.0%, its approximate size 9.35 Mbp, its GenBank deposit SAMN02584999.
The description is as before (Lechevalier and Lechevalier, 1970) with the following addition. The G+C content of the type-strain genome is 70.4%, its approximate size 7.53 Mbp, its IMG deposit 2728369498.
The description is as before (Kroppenstedt et al., 1990) with the following addition. The G+C content of the type-strain genome is 72.8%, its approximate size 6.72 Mbp, its GenBank deposit SAMD00046516.
The description is as before (Zhang et al., 2001) with the following addition. The G+C content of the type-strain genome is 72.8%, its approximate size 6.72 Mbp, its IMG deposit 2728369515.
The description is as before (Hall et al., 2005) with the following modification. The G+C content of the type-strain genome is 73.1%, its approximate size 3.53 Mbp, its GenBank deposit SAMN02440612.
The description is as before (Dent and Williams, 1984) with the following restriction. The G+C content of the type-strain genome is 71.2%, its approximate size 2.90 Mbp, its GenBank deposit SAMD00059584.
The description is as before (Lachner-Sandoval, 1898) with the following addition. The G+C content of the type-strain genome is 71.4%, its approximate size 4.03 Mbp, its GenBank deposit SAMN02745698.
The description is as before (Renvoise et al., 2009) with the following addition. The G+C content of the type-strain genome is 67.8%, its approximate size 3.35 Mbp, its GenBank deposit SAMN02469892.
The description is as before (Henssge et al., 2009) with the following modification. The G+C content of the type-strain genome is 67.9%, its approximate size 3.11 Mbp, its GenBank deposit SAMN00828754.
The description is as before (Collins et al., 2000a) with the following addition. The G+C content of the type-strain genome is 72.6%, its approximate size 3.05 Mbp, its GenBank deposit SAMN04435863.
The description is as before (An et al., 2006) with the following modification. The G+C content of the type-strain genome is 69.9%, its approximate size 3.09 Mbp, its GenBank deposit SAMN05216355.
The description is as before (Dent and Williams, 1986) with the following restriction. The G+C content of the type-strain genome is 70.1%, its approximate size 3.17 Mbp, its GenBank deposit SAMN02441219.
The description is as before (Renvoise et al., 2010) with the following addition. The G+C content of the type-strain genome is 71.2%, its approximate size 2.91 Mbp, its GenBank deposit SAMN02472053.
The description is as before (Nikolaitchouk et al., 2000) with the following modification. The G+C content of the type-strain genome is 68.7%, its approximate size 2.61 Mbp, its GenBank deposit SAMN00002545.
The description is as before (Jiang et al., 2008) with the following modification. The G+C content of the type-strain genome is 74.0%, its approximate size 5.86 Mbp, its GenBank deposit SAMN02256421.
The description is as before (Terekhova et al., 1977) with the following addition. The G+C content of the type-strain genome is 71.0%, its approximate size 10.42 Mbp, its IMG deposit 2724679919.
The description is as before (Goodfellow et al., 1990) with the following addition. The G+C content of the type-strain genome is 69.4%, its approximate size 10.65 Mbp, its GenBank deposit SAMN04489716.
The description is as before (Wink et al., 2014) with the following addition. The G+C content of the type-strain genome is 70.4%, its approximate size 9.38 Mbp, its GenBank deposit SAMN02603031.
The description is as before (Stackebrandt and Kroppenstedt, 1987) with the following addition. The G+C content of the type-strain genome is 70.9%, its approximate size 10.99 Mbp, its GenBank deposit SAMN02256449.
The description is as before (Beretta, 1973) with the following addition. The G+C content of the type-strain genome is 70.4%, its approximate size 11.15 Mbp, its IMG deposit 2728369518.
The description is as before (Couch, 1963) with the following addition. The G+C content of the type-strain genome is 70.8%, its approximate size 8.77 Mbp, its GenBank deposit SAMD00060941.
The description is as before (Couch, 1950) with the following addition. The G+C content of the type-strain genome is 71.4%, its approximate size 10.38 Mbp, its GenBank deposit SAMN05421541.
The description is as before (Lechevalier and Lechevalier, 1975) with the following addition. The G+C content of the type-strain genome is 70.6%, its approximate size 10.32 Mbp, its GenBank deposit SAMN03327476.
The description is as before (Stackebrandt and Kroppenstedt, 1987) with the following addition. The G+C content of the type-strain genome is 70.3%, its approximate size 10.03 Mbp, its IMG deposit 2724679777.
The description is as before (Cao et al., 2009) with the following modification. The G+C content of the type-strain genome is 67.9%, its approximate size 7.98 Mbp, its GenBank deposit SAMN02440435.
The description is as before (Wang et al., 2001) with the following modification. The G+C content of the type-strain genome is 70.6%, its approximate size 6.40 Mbp, its GenBank deposit SAMN04489717.
The description is as before (Tang et al., 2011) with the following modification. The G+C content of the type-strain genome is 68.8%, its approximate size 5.34 Mbp, its GenBank deposit SAMN02911260.
The description is as before (Gochnauer et al., 1975) with the following modification. The G+C content of the type-strain genome is 68.0%, its approximate size 5.35 Mbp, its GenBank deposit SAMN02261305.
The description is as before (Meklat et al., 2013a) with the following addition. The G+C content of the type-strain genome is 67.7%, its approximate size 5.00 Mbp, its GenBank deposit SAMN04487820.
The description is as before (Meklat et al., 2013b) with the following addition. The G+C content of the type-strain genome is 67.5%, its approximate size 4.92 Mbp, its GenBank deposit SAMN04487904.
The description is as before (Meklat et al., 2013c) with the following addition. The G+C content of the type-strain genome is 69.5%, its approximate size 4.68 Mbp, its GenBank deposit SAMN04489718.
The description is as before (Hasegawa et al., 1978) with the following addition. The G+C content of the type-strain genome is 73.7%, its approximate size 8.25 Mbp, its GenBank deposit SAMN00001904.
The description is as before (Hasegawa et al., 1983) with the following restriction. The G+C content of the type-strain genome is 73.4%, its approximate size 9.35 Mbp, its GenBank deposit SAMN03080609.
The description is as before (Miller et al., 1991) with the following restriction. The G+C content of the type-strain genome is 72.1%, its approximate size 3.63 Mbp, its GenBank deposit SAMN03701813.
The description is as before (Evtushenko et al., 2001) with the following modification. The G+C content of the type-strain genome is 65.2%, its approximate size 3.92 Mbp, its GenBank deposit SAMN03333328.
The description is as before (Zlamala et al., 2002) with the following restriction. The G+C content of the type-strain genome is 71.2%, its approximate size 2.90 Mbp, its GenBank deposit SAMN04487783.
The description is as before (Mayilraj et al., 2006b) with the following modification. The G+C content of the type-strain genome is 72.5%, its approximate size 2.68 Mbp, its GenBank deposit SAMN02440814.
The description is as before (Zgurskaya et al., 1992) with the following modification. The G+C content of the type-strain genome is 70.0%, its approximate size 4.19 Mbp, its GenBank deposit SAMN05443544.
The description is as before (Jurado et al., 2005) with the following modification. The G+C content of the type-strain genome is 69.1%, its approximate size 4.30 Mbp, its GenBank deposit SAMN02441229.
The description is as before (Labeda and Kroppenstedt, 2008) with the following modification. The G+C content of the type-strain genome is 70.3%, its approximate size 8.57 Mbp, its GenBank deposit SAMN04489726.
The description is as before (Du et al., 2013) with the following modification. The G+C content of the type-strain genome is 73.9%, its approximate size 6.97 Mbp, its IMG deposit 2728369266.
The description is as before (Killer et al., 2013) with the following modification. The G+C content of the type-strain genome is 50.1%, its approximate size 1.88 Mbp, its GenBank deposit SAMN02441737.
The description is as before (Huys et al., 2007) with the following modification. The G+C content of the type-strain genome is 46.6%, its approximate size 1.85 Mbp, its GenBank deposit SAMN02441248.
The description is as before (Mertz and Yao, 1993) with the following addition. The G+C content of the type-strain genome is 68.7%, its approximate size 9.81 Mbp, its GenBank deposit SAMN02261260.
The description is as before (Tan et al., 2006) with the following addition. The G+C content of the type-strain genome is 71.9%, its approximate size 9.31 Mbp, its GenBank deposit SAMN04489730.
The description is as before (Henssen et al., 1987) with the following modification. The G+C content of the type-strain genome is 68.9%, its approximate size 9.22 Mbp, its GenBank deposit SAMN01939083.
The description is as before (Wink et al., 2003) with the following addition. The G+C content of the type-strain genome is 70.8%, its approximate size 10.86 Mbp, its GenBank deposit SAMN02256403.
The description is as before (Majumdar et al., 2006) with the following addition. The G+C content of the type-strain genome is 70.0%, its approximate size 8.70 Mbp, its GenBank deposit SAMN02261299.
The description is as before (Labeda, 1995) with the following modification. The G+C content of the type-strain genome is 68.4%, its approximate size 9.05 Mbp, its GenBank deposit SAMN04384194.
The description is as before (Wink et al., 2004) with the following addition. The G+C content of the type-strain genome is 68.6%, its approximate size 8.53 Mbp, its GenBank deposit SAMN02470008.
The description is as before (Lee, 2006a) with the following modification. The G+C content of the type-strain genome is 69.1%, its approximate size 10.10 Mbp, its GenBank deposit SAMN02645327.
The description is as before (Al-Mussallam et al., 2003) with the following addition. Produces light-gray aerial mycelium where present. The G+C content of the type-strain genome is 69.0%, its approximate size 9.09 Mbp, its GenBank deposit SAMN04384022.
The description is as before (Stackebrandt et al., 2004) with the following modification. The G+C content of the type-strain genome is 68.7%, its approximate size 9.05 Mbp, its GenBank deposit SAMN04489729.
The description is as before (Bian et al., 2009) with the following modification. The G+C content of the type-strain genome is 68.2%, its approximate size 7.02 Mbp, its GenBank deposit SAMN05216266.
The description is as before (Lechevalier et al., 1986) with the following restriction. The G+C content of the type-strain genome is 71.3%, its approximate size 10.24 Mbp, its GenBank deposit SAMN02604164.
The description is as before (Groth et al., 2007) with the following addition. The G+C content of the type-strain genome is 70.0%, its approximate size 9.11 Mbp, its GenBank deposit SAMN02261308.
The description is as before (Ding et al., 2007) with the following modification. The G+C content of the type-strain genome is 69.5%, its approximate size 9.31 Mbp, its GenBank deposit SAMN04489734.
The description is as before (Lechevalier et al., 1986) with the following modification. The G+C content of the type-strain genome is 69.0%, its approximate size 9.06 Mbp, its GenBank deposit SAMN02142155.
The description is as before (Labeda et al., 2003) with the following addition. The G+C content of the type-strain genome is 71.2%, its approximate size 10.30 Mbp, its GenBank deposit SAMN05421837.
The description is as before (Tan et al., 2007) with the following addition. The G+C content of the type-strain genome is 68.5%, its approximate size 8.29 Mbp, its GenBank deposit SAMN04489731.
The description is as before (Bala et al., 2004) with the following addition. The G+C content of the type-strain genome is 71.8%, its approximate size 9.20 Mbp, its GenBank deposit SAMN02745639.
The description is as before (Huang et al., 2001) with the following modification. The G+C content of the type-strain genome is 69.8%, its approximate size 9.87 Mbp, its GenBank deposit SAMN05421854.
The description is as before (Carlsohn et al., 2007) with the following addition. The G+C content of the type-strain genome is 71.0%, its approximate size 9.86 Mbp, its GenBank deposit SAMN04489732.
The description is as before (Goodfellow et al., 2001) with the following addition. The G+C content of the type-strain genome is 71.3%, its approximate size 7.59 Mbp, its GenBank deposit SAMN05421835.
The description is as before (Lechevalier et al., 1986) with the following modification. The G+C content of the type-strain genome is 69.4%, its approximate size 6.86 Mbp, its GenBank deposit SAMN04489728.
The description is as before (Chun et al., 1999) with the following modification. The G+C content of the type-strain genome is 71.6%, its approximate size 8.69 Mbp, its GenBank deposit SAMN02441454.
The description is as before (Wink et al., 2003) with the following addition. The G+C content of the type-strain genome is 71.7%, its approximate size 10.36 Mbp, its GenBank deposit SAMN04489727.
The description is as before (Wink et al., 2003) with the following addition. The G+C content of the type-strain genome is 72.0%, its approximate size 9.84 Mbp, its GenBank deposit SAMN02645326.
The description is as before (Chen et al., 2010) with the following modification. The G+C content of the type-strain genome is 68.9%, its approximate size 9.41 Mbp, its GenBank deposit SAMN05421504.
The description is as before (Yamada et al., 2006) with the following modification. The G+C content of the type-strain genome is 54.6%, its approximate size 4.10 Mbp, its GenBank deposit SAMD00034885.
The description is as before (Lee, 2015) with the following modification. The G+C content of the type-strain genome is 64.1%, its approximate size 4.68 Mbp, its IMG deposit 2728369516.
The description is as before (Collins et al., 1982) with the following restriction. The G+C content of the type-strain genome is 53.1%, its approximate size 1.99 Mbp, its GenBank deposit SAMN02598519.
The description is as given before (Ramos et al., 1997) with the following addition. The G+C content of the type-strain genome is 50.0%, its approximate size 2.00 Mbp, its GenBank deposit SAMN04489737.
The description is as before (Kawaichi et al., 2013) with the following modification. The G+C content of the type-strain genome is 59.6%, its approximate size 3.56 Mbp, its GenBank deposit SAMN03274860.
The description is as before (Zhang et al., 2010a) with the following modification. The G+C content of the type-strain genome is 60.6%, its approximate size 4.62 Mbp, its GenBank deposit SAMN04489740.
The description is as before (Heyrman et al., 2005) with the following modification. The G+C content of the type-strain genome is 63.6%, its approximate size 4.58 Mbp, its GenBank deposit SAMN02441477.
The description is as before (Ensign and Rittenberg, 1963) with the following addition. The G+C content of the type-strain genome is 64.4%, its approximate size 5.20 Mbp, its GenBank deposit SAMN04489742.
The description is as before (Zhang et al., 2012) with the following modification. The G+C content of the type-strain genome is 67.0%, its approximate size 4.06 Mbp, its GenBank deposit SAMN05216555.
The description is as before (Dastager et al., 2014b) with the following modification. The G+C content of the type-strain genome is 67.1%, its approximate size 4.22 Mbp, its GenBank deposit SAMN04263892.
The description is as before (Conn and Dimmick, 1947) with the following addition. The G+C content of the type-strain genome is 66.2%, its approximate size 4.95 Mbp, its GenBank deposit SAMD00041813.
The description is as before (Kageyama et al., 1999) with the following modification. The G+C content of the type-strain genome is 45.4%, its approximate size 1.66 Mbp, its GenBank deposit SAMN02440747.
The description is as before (Collins and Wallbanks, 1992) with the following modification. The G+C content of the type-strain genome is 48.7%, its approximate size 1.78 Mbp, its GenBank deposit SAMN04489746.
The description is as before (Yamada et al., 2007) with the following modification. The G+C content of the type-strain genome is 52.8%, its approximate size 3.66 Mbp, its GenBank deposit SAMN03842211.
The description is as before (Groth et al., 1999b) with the following modification. The G+C content of the type-strain genome is 73.1%, its approximate size 4.67 Mbp, its GenBank deposit SAMN02598436.
The description is as before (Killer et al., 2011) with the following modification. The G+C content of the type-strain genome is 62.7%, its approximate size 1.83 Mbp, its GenBank deposit SAMN02442035.
The description is as before (Reuter, 1963) with the following addition. The G+C content of the type-strain genome is 59.2%, its approximate size 2.09 Mbp, its GenBank deposit SAMD00061080.
The description is as before (Scardovi and Trovatelli, 1969) with the following addition. The G+C content of the type-strain genome is 60.0%, its approximate size 2.14 Mbp, its GenBank deposit SAMN02442012.
The description is as given before (Orla-Jensen, 1924) with the following addition. The G+C content of the type-strain genome is 62.7%, its approximate size 2.21 Mbp, its GenBank deposit SAMD00061040.
The description is as before (Killer et al., 2011) with the following modification. The G+C content of the type-strain genome is 57.5%, its approximate size 2.05 Mbp, its GenBank deposit SAMN02673427.
The description is as before (Killer et al., 2009) with the following modification. The G+C content of the type-strain genome is 56.1%, its approximate size 1.90 Mbp, its GenBank deposit SAMN02952076.
The description is as before (Reuter, 1963) with the following addition. The G+C content of the type-strain genome is 58.9%, its approximate size 2.27 Mbp, its GenBank deposit SAMD00061041.
The description is as before (Scardovi and Crociani, 1974) with the following modification. The pH range sustaining growth is 6–8. The G+C content of the type-strain genome is 56.2%, its approximate size 2.08 Mbp, its GenBank deposit SAMD00061042.
The description is as before (Biavati et al., 1982) with the following addition. The G+C content of the type-strain genome is 60.5%, its approximate size 1.76 Mbp, its GenBank deposit SAMN02666223.
The description is as before (Delcenserie et al., 2007) with the following modification. The G+C content of the type-strain genome is 57.7%, its approximate size 2.36 Mbp, its GenBank deposit SAMN02699097.
The description is as before (Scardovi et al., 1979) with the following modification. The G+C content of the type-strain genome is 64.9%, its approximate size 2.53 Mbp, its GenBank deposit SAMN02673433.
The description is as before (Scardovi and Crociani, 1974) with the following modification. The G+C content of the type-strain genome is 58.6%, its approximate size 2.67 Mbp, its GenBank deposit SAMN05192536.
The description is as before (Lauer, 1990) with the following modification. The G+C content of the type-strain genome is 57.5%, its approximate size 2.02 Mbp, its GenBank deposit SAMN02299421.
The description is as before (Biavati et al., 1982) with the following restriction. The G+C content of the type-strain genome is 63.4%, its approximate size 1.91 Mbp, its GenBank deposit SAMN02743881.
The description is as before (Scardovi and Trovatelli, 1969) with the following addition. The G+C content of the type-strain genome is 60.5%, its approximate size 1.73 Mbp, its GenBank deposit SAMN05771123.
The description is as before (Modesto et al., 2015) with the following modification. The G+C content of the type-strain genome is 62.6%, its approximate size 2.91 Mbp, its GenBank deposit SAMD00065376.
The description is as before (Mattareli et al., 2008) with the following modification. The G+C content of the type-strain genome is 60.0%, its approximate size 2.39 Mbp, its GenBank deposit SAMN04489749.
The description is as before (Scardovi and Zani, 1974) with the following restriction. The G+C content of the type-strain genome is 58.7%, its approximate size 1.83 Mbp, its GenBank deposit SAMN02441586.
The description is as before (Biavati and Mattarelli, 1991) with the following modification. The G+C content of the type-strain genome is 60.3%, its approximate size 2.28 Mbp, its GenBank deposit SAMN02673440.
The description is as before (Biavati et al., 1982) with the following addition. The G+C content of the type-strain genome is 62.7%, its approximate size 1.87 Mbp, its GenBank deposit SAMN02440587.
The description is as before (Watanabe et al., 2009) with the following modification. The G+C content of the type-strain genome is 62.8%, its approximate size 2.17 Mbp, its GenBank deposit SAMN02673442.
The description is as before (Scardovi et al., 1979) with the following modification. The G+C content of the type-strain genome is 56.4%, its approximate size 2.31 Mbp, its GenBank deposit SAMD00061045.
The description is as before (Mitsuoka, 1969) with the following addition. The G+C content of the type-strain genome is 63.1%, its approximate size 1.90 Mbp, its GenBank deposit SAMN02673445.
The description is as before (Trovatelli et al., 1974) with the following restriction. pH range of growth from 4.5 to 7.3. The G+C content of the type-strain genome is 64.2%, its approximate size 2.15 Mbp, its GenBank deposit SAMN02673447.
The description is as before (Biavati and Mattarelli, 1991) with the following modification. The G+C content of the type-strain genome is 59.2%, its approximate size 2.22 Mbp, its GenBank deposit SAMN02743880.
The description is as before (Hoyles et al., 2002b) with the following modification. The G+C content of the type-strain genome is 64.6%, its approximate size 3.16 Mbp, its GenBank deposit SAMD00061046.
The description is as before (Kim et al., 2010) with the following modification. The G+C content of the type-strain genome is 59.4%, its approximate size 2.30 Mbp, its GenBank deposit SAMN02673454.
The description is as before (Biavati et al., 1982) with the following addition. The G+C content of the type-strain genome is 60.9%, its approximate size 2.77 Mbp, its GenBank deposit SAMN02440843.
The description is as before (Zhu et al., 2003) with the following restriction. The G+C content of the type-strain genome is 60.4%, its approximate size 2.22 Mbp, its GenBank deposit SAMN02441522.
The description is as before (Mitsuoka, 1969) with the following addition. The G+C content of the type-strain genome is 59.9%, its approximate size 2.10 Mbp, its GenBank deposit SAMN02673459.
The description is as before (Collins et al., 1988a) with the following modification. The G+C content of the type-strain genome is 72.0%, its approximate size 3.61 Mbp, its GenBank deposit SAMN00002685.
The description is as before (Park et al., 2011) with the following modification. The G+C content of the type-strain genome is 72.8%, its approximate size 3.19 Mbp, its GenBank deposit SAMN02470219.
The description is as before (Collins et al., 1983) with the following modification. The G+C content of the type-strain genome is 64.3%, its approximate size 3.70 Mbp, its GenBank deposit SAMD00046483.
The description is as before (Roux and Raoult, 2009) with the following addition. The G+C content of the type-strain genome is 62.3%, its approximate size 2.35 Mbp, its GenBank deposit SAMEA3138417.
The description is as before (McBride et al., 1993) with the following modification. The G+C content of the type-strain genome is 58.0%, its approximate size 2.56 Mbp, its GenBank deposit SAMN00139181.
The description is as before (Kämpfer et al., 2010b) with the following addition. The G+C content of the type-strain genome is 63.3%, its approximate size 4.44 Mbp, its GenBank deposit SAMN04489751.
The description is as before (Sekiguchi et al., 2003) with the following addition. The G+C content of the type-strain genome is 58.8%, its approximate size 5.14 Mbp, its GenBank deposit SAMN00707155.
The description is as before (Ara et al., 2008a) with the following modification. The G+C content of the type-strain genome is 64.4%, its approximate size 7.69 Mbp, its GenBank deposit SAMN02256447.
The description is as before (Busti et al., 2006) with the following modification. The G+C content of the type-strain genome is 69.8%, its approximate size 10.47 Mbp, its GenBank deposit SAMN02598445.
The description is as before (Shi et al., 2012) with the following modification. The G+C content of the type-strain genome is 73.0%, its approximate size 3.98 Mbp, its GenBank deposit SAMN03113093.
The description is as before (Bergey et al., 1923) with the following addition. The G+C content of the type-strain genome is 74.6%, its approximate size 3.91 Mbp, its GenBank deposit SAMN03112937.
The description is as before (Bergey et al., 1923) with the following addition. The G+C content of the type-strain genome is 74.7%, its approximate size 4.27 Mbp, its GenBank deposit SAMN00713615.
The description is as before (Bergey et al., 1923) with the following addition. The G+C content of the type-strain genome is 74.3%, its approximate size 4.12 Mbp, its GenBank deposit SAMN02598424.
The description is as before (Zhang et al., 2013a) with the following modification. The G+C content of the type-strain genome is 75.6%, its approximate size 3.96 Mbp, its GenBank deposit SAMN05421867.
The description is as before (Lagier et al., 2012) with the following modification. The G+C content of the type-strain genome is 74.8%, its GenBank deposit SAMEA2272357.
The description is as before (Pierson and Castenholz, 1974) with the following restriction. The G+C content of the type-strain genome is 56.7%, its approximate size 5.26 Mbp, its GenBank deposit SAMN02598539.
The description is as before (Davis et al., 1984) with the following addition. The G+C content of the type-strain genome is 72.7%, its approximate size 3.30 Mbp, its GenBank deposit SAMN06265879.
The description is as before (Zhou et al., 2016) with the following modification. The G+C content of the type-strain genome is 70.8%, its approximate size 4.35 Mbp, its GenBank deposit SAMN06622498.
The description is as before (Kageyama and Benno, 2000) with the following modification. The G+C content of the type-strain genome is 62.5%, its approximate size 1.80 Mbp, its GenBank deposit SAMN00008813.
The description is as before (Kageyama and Benno, 2000) with the following modification. The G+C content of the type-strain genome is 63.2%, its approximate size 2.40 Mbp, its GenBank deposit SAMN00008814.
The description is as before (Kim et al., 2012b) with the following modification. The G+C content of the type-strain genome is 69.3%, its approximate size 3.88 Mbp, its IMG deposit 2731639183.
The description is as before (Monciardini et al., 2003) with the following modification. The G+C content of the type-strain genome is 72.7%, its approximate size 6.36 Mbp, its GenBank deposit SAMN00002582.
The description is as before (Neubauer et al., 1991) with the following modification. The G+C content of the type-strain genome is 59.7%, its approximate size 2.41 Mbp, its GenBank deposit SAMN00002226.
The description is as before (Riegel et al., 1993) with the following modification. The G+C content of the type-strain genome is 64.9%, its approximate size 2.33 Mbp, its GenBank deposit SAMN05421802.
The description is as before (Collins, 1987) with the following restriction. The G+C content of the type-strain genome is 55.6%, its approximate size 2.76 Mbp, its GenBank deposit SAMN00189098.
The description is as before (Yassin et al., 2002a) with the following modification. The G+C content of the type-strain genome is 64.3%, its approximate size 2.25 Mbp, its GenBank deposit SAMN05444817.
The description is as given before (Fernández-Garayzábal et al., 2003) with the following addition. The G+C content of the type-strain genome is 60.9%, its approximate size 2.93 Mbp, its GenBank deposit SAMN02996496.
The description is as before (Riegel et al., 1995a) with the following modification. The G+C content of the type-strain genome is 58.9%, its approximate size 2.03 Mbp, its GenBank deposit SAMN02603032.
The description is as before (Hall et al., 2003a) with the following addition. The G+C content of the type-strain genome is 65.4%, its approximate size 2.36 Mbp, its GenBank deposit SAMN02911287.
The description is as before (Collins et al., 1999) with the following modification. The G+C content of the type-strain genome is 58.5%, its approximate size 2.57 Mbp, its GenBank deposit SAMN03106126.
The description is as before (Yamada and Komagata, 1972) with the following restriction. The G+C content of the type-strain genome is 52.4%, its approximate size 2.89 Mbp, its GenBank deposit SAMN02441249.
The description is as before (Fernández-Garayzábal et al., 1998) with the following addition. The G+C content of the type-strain genome is 59.4%, its approximate size 2.45 Mbp, its GenBank deposit SAMN03365263.
The description is as before (Collins et al., 2001a) with the following addition. The G+C content of the type-strain genome is 64.5%, its approximate size 1.96 Mbp, its GenBank deposit SAMN02256495.
The description is as before (Brennan et al., 2001a) with the following modification. The G+C content of the type-strain genome is 55.7%, its approximate size 3.13 Mbp, its GenBank deposit SAMN03081454.
The description is as before (Collins et al., 2004a) with the following addition. The G+C content of the type-strain genome is 49.7%, its approximate size 1.84 Mbp, its GenBank deposit SAMN02256419.
The description is as before (Fernández-Garyzábal et al., 2004) with the following addition. The G+C content of the type-strain genome is 62.1%, its approximate size 2.55 Mbp, its GenBank deposit SAMN02256507.
The description is as before (Yanagawa and Honda, 1978) with the following modification. The G+C content of the type-strain genome is 57.0%, its approximate size 2.94 Mbp, its GenBank deposit SAMN05661109.
The description is as before (Zhou et al., 2012b) with the following modification. The G+C content of the type-strain genome is 55.3%, its approximate size 3.03 Mbp, its GenBank deposit SAMN02950576.
The description is as before (Lehmann and Neumann, 1896) with the following addition. The G+C content of the type-strain genome is 53.5%, its approximate size 2.46 Mbp, its GenBank deposit SAMEA2517360.
The description is as before (Lee et al., 2009) with the following modification. The G+C content of the type-strain genome is 66.9%, its approximate size 2.65 Mbp, its GenBank deposit SAMN02256506.
The description is as before (Fudou et al., 2002) with the following restriction. The G+C content of the type-strain genome is 63.0%, its approximate size 3.22 Mbp, its GenBank deposit SAMD00061103.
The description is as before (Frischmann et al., 2012) with the following addition. The G+C content of the type-strain genome is 58.1%, its approximate size 2.69 Mbp, its GenBank deposit SAMN03462986.
The description is as before (Sjödén et al., 1998) with the following addition. The G+C content of the type-strain genome is 63.2%, its approximate size 2.72 Mbp, its GenBank deposit SAMN02641485.
The description is as before (Barksdale et al., 1979) with the following modification. The G+C content of the type-strain genome is 59.9%, its approximate size 2.76 Mbp, its GenBank deposit SAMN02996497.
The description is as before (Wiertz et al., 2013) with the following addition. The G+C content of the type-strain genome is 71.5%, its approximate size 2.60 Mbp, its GenBank deposit SAMN02991553.
The description is as before (Funke et al., 2009) with the following addition. The G+C content of the type-strain genome is 49.8%, its approximate size 2.91 Mbp, its GenBank deposit SAMN02441205.
The description is as before (Funke et al., 1995) with the following modification. The G+C content of the type-strain genome is 59.1%, its approximate size 2.82 Mbp, its GenBank deposit SAMN02850907.
The description is as before (Abe et al., 1967) with the following addition. The G+C content of the type-strain genome is 53.8%, its approximate size 3.28 Mbp, its GenBank deposit SAMEA3138338.
The description is as before (Chen et al., 2004) with the following modification. The G+C content of the type-strain genome is 68.3%, its approximate size 3.22 Mbp, its GenBank deposit SAMN02603027.
The description is as before (Wu et al., 2011) with the following modification. The G+C content of the type-strain genome is 68.6%, its approximate size 2.68 Mbp, its GenBank deposit SAMN03283197.
The description is as before (Funke et al., 1997b) with the following modification. The G+C content of the type-strain genome is 64.3%, its approximate size 2.57 Mbp, its GenBank deposit SAMN02950575.
The description is as before (Jackman et al., 1987) with the following modification. The G+C content of the type-strain genome is 61.6%, its approximate size 2.43 Mbp, its GenBank deposit SAMN00001506.
The description is as before (Collins et al., 1998) with the following modification. The G+C content of the type-strain genome is 57.5%, its approximate size 2.45 Mbp, its GenBank deposit SAMN02603033.
The description is as before (Bergey et al., 1925) with the following addition. The G+C content of the type-strain genome is 46.5%, its approximate size 2.35 Mbp, its GenBank deposit SAMN03365283.
The description is as before (Wiertz et al., 2013) with the following addition. The G+C content of the type-strain genome is 60.5%, its approximate size 2.77 Mbp, its GenBank deposit SAMN04012704.
The description is as before (Kämpfer et al., 2009a) with the following addition. The G+C content of the type-strain genome is 58.6%, its approximate size 2.95 Mbp, its GenBank deposit SAMN02256424.
The description is as before (Du et al., 2010) with the following modification. The G+C content of the type-strain genome is 67.8%, its approximate size 2.73 Mbp, its GenBank deposit SAMN02800399.
The description is as before (Merhej et al., 2009) with the following addition. The G+C content of the type-strain genome is 65.0%, its approximate size 2.18 Mbp, its GenBank deposit SAMN02441587.
The description is as before (Bernard et al., 2016) with the following restriction. The G+C content of the type-strain genome is 69.0%, its approximate size 2.37 Mbp, its GenBank deposit SAMN02441393.
The description is as before (Collins, 1982b) with the following restriction. The G+C content of the type-strain genome is 57.1%, its approximate size 2.86 Mbp, its GenBank deposit SAMN00001942.
The description is as before (Yassin et al., 2002b) with the following modification. The G+C content of the type-strain genome is 60.0%, its approximate size 2.66 Mbp, its GenBank deposit SAMN03140311.
The description is as before (Brennan et al., 2001a) with the following modification. The G+C content of the type-strain genome is 67.1%, its approximate size 3.43 Mbp, its GenBank deposit SAMN02603088.
The description is as before (Funke et al., 2010b) with the following addition. The G+C content of the type-strain genome is 52.6%, its approximate size 3.47 Mbp, its GenBank deposit SAMN03568800.
The description is as before (Collins, 1982a) with the following modification. The G+C content of the type-strain genome is 66.6%, its approximate size 2.27 Mbp, its GenBank deposit SAMN04488535.
The description is as before (Shukla et al., 2003) with the following addition. The G+C content of the type-strain genome is 60.6%, its approximate size 2.82 Mbp, its GenBank deposit SAMN02603064.
The description is as before (Shin et al., 2011) with the following modification. The G+C content of the type-strain genome is 69.5%, its approximate size 3.11 Mbp, its GenBank deposit SAMN02470217.
The description is as before (Lehmann and Neumann, 1896) with the following addition. The G+C content of the type-strain genome is 55.3%, its approximate size 2.26 Mbp, its GenBank deposit SAMN02743909.
The description is as before (Eberson, 1918) with the following addition. The G+C content of the type-strain genome is 52.2%, its approximate size 2.34 Mbp, its GenBank deposit SAMN06701041.
The description is as before (Ernst, 1906) with the following addition. The G+C content of the type-strain genome is 59.1%, its approximate size 2.35 Mbp, its GenBank deposit SAMN04488536.
The description is as before (Otsuka et al., 2005) with the following modification. The G+C content of the type-strain genome is 57.1%, its approximate size 2.60 Mbp, its GenBank deposit SAMN02603065.
The description is as before (Riegel et al., 1995b) with the following modification. The G+C content of the type-strain genome is 59.0%, its approximate size 2.85 Mbp, its GenBank deposit SAMN00001463.
The description is as before (Riegel et al., 1997) with the following modification. The G+C content of the type-strain genome is 60.1%, its approximate size 2.83 Mbp, its GenBank deposit SAMN03177398.
The description is as before (Goyache et al., 2003a) with the following addition. The G+C content of the type-strain genome is 74.7%, its approximate size 2.59 Mbp, its GenBank deposit SAMN02996499.
The description is as before (Goyache et al., 2003b) with the following addition. The G+C content of the type-strain genome is 57.5%, its approximate size 2.45 Mbp, its GenBank deposit SAMN05660282.
The description is as before (Yassin and Siering, 2008) with the following addition. The G+C content of the type-strain genome is 61.5%, its approximate size 2.92 Mbp, its GenBank deposit SAMN02441223.
The description is as before (Eberson, 1918) with the following addition. The G+C content of the type-strain genome is 59.4%, its approximate size 2.72 Mbp, its GenBank deposit SAMN00001507.
The description is as before (Collins et al., 2001b) with the following addition. The G+C content of the type-strain genome is 63.1%, its approximate size 2.72 Mbp, its GenBank deposit SAMN03480629.
The description is as before (Merhej et al., 2009) with the following addition. The G+C content of the type-strain genome is 66.6%, its approximate size 2.63 Mbp, its GenBank deposit SAMN04488539.
The description is as before (Yassin, 2009) with the following addition. The G+C content of the type-strain genome is 59.2%, its approximate size 2.30 Mbp, its GenBank deposit SAMN02256494.
The description is as before (Pitcher et al., 1992) with the following modification. The G+C content of the type-strain genome is 64.2%, its approximate size 2.37 Mbp, its GenBank deposit SAMEA3138282.
The description is as before (Yassin, 2007) with the following addition. The G+C content of the type-strain genome is 65.0%, its approximate size 2.33 Mbp, its GenBank deposit SAMN02953970.
The description is as before (Hoyles et al., 2013) with the following addition. The G+C content of the type-strain genome is 65.5%, its approximate size 2.42 Mbp, its GenBank deposit SAMN03480647.
The description is as before (Lehmann and Neumann, 1899) with the following addition. The G+C content of the type-strain genome is 69.7%, its approximate size 2.69 Mbp, its GenBank deposit SAMD00046521.
The description is as before (Liu et al., 2012) with the following modification. The G+C content of the type-strain genome is 65.1%, its approximate size 3.83 Mbp, its GenBank deposit SAMN05216281.
The description is as before (Zhang et al., 2007a) with the following modification. The G+C content of the type-strain genome is 68.3%, its approximate size 3.25 Mbp, its GenBank deposit SAMN05216282.
The description is as before (Tamura et al., 1998) with the following modification. The G+C content of the type-strain genome is 71.7%, its approximate size 9.20 Mbp, its GenBank deposit SAMN02849401.
The description is as before (Tamura and Hatano, 2001) with the following addition. The G+C content of the type-strain genome is 71.2%, its approximate size 9.58 Mbp, its GenBank deposit SAMN05443668.
The description is as before (Scholz and Kilian, 2016) with the following addition. The G+C content of the type-strain genome is 60.1%, its approximate size 2.48 Mbp, its GenBank deposit SAMN02440711.
The description is as before (Scholz and Kilian, 2016) with the following addition. The G+C content of the type-strain genome is 63.4%, its approximate size 2.53 Mbp, its GenBank deposit SAMN02299449.
The description is as before (Scholz and Kilian, 2016) with the following addition. The G+C content of the type-strain genome is 64.2%, its approximate size 3.52 Mbp, its GenBank deposit SAMN02798018.
The description is as before (Thiemann et al., 1967) with the following addition. The G+C content of the type-strain genome is 73.2%, its approximate size 11.45 Mbp, its GenBank deposit SAMN02645310.
The description is as before (Löffler et al., 2013) with the following restriction. The G+C content of the type-strain genome is 48.9%, its GenBank deposit SAMN02603990.
The description is as before (Moe et al., 2009) with the following modification. The G+C content of the type-strain genome is 55.0%, its approximate size 1.69 Mbp, its GenBank deposit SAMN02598529.
The description is as before (Ue et al., 2011) with the following modification. The G+C content of the type-strain genome is 64.1%, its approximate size 2.62 Mbp, its GenBank deposit SAMD00022567.
The description is as before (Matsumoto et al., 2010) with the following restriction. The G+C content of the type-strain genome is 70.2%, its approximate size 3.21 Mbp, its GenBank deposit SAMD00022568.
The description is as before (Groth et al., 1997) with the following modification. The G+C content of the type-strain genome is 64.7%, its approximate size 3.57 Mbp, its GenBank deposit SAMN02440416.
The description is as before (Jones and Collins, 1988) with the following modification. The G+C content of the type-strain genome is 63.2%, its approximate size 2.16 Mbp, its GenBank deposit SAMD00046478.
The description is as before (Gordon, 1964) with the following addition. The G+C content of the type-strain genome is 59.4%, its approximate size 2.62 Mbp, its GenBank deposit SAMN02440871.
The description is as before (Jang et al., 2012) with the following modification. The G+C content of the type-strain genome is 70.7%, its approximate size 3.49 Mbp, its IMG deposit 2731639227.
The description is as before (Kim et al., 2011) with the following modification. The G+C content of the type-strain genome is 67.3%, its approximate size 3.35 Mbp, its GenBank deposit SAMN02470218.
The description is as before (Yassin et al., 2006) with the following modification. The G+C content of the type-strain genome is 70.8%, its approximate size 3.60 Mbp, its GenBank deposit SAMD00046472.
The description is as before (Mayilraj et al., 2006a) with the following modification and addition. Tolerates up to 15% NaCl, grows at temperatures between 10 and 45°C and from pH 6–10. Main fatty acids are C18:1 ω9c, C16:0 and 10-methyl-C18:0 (tuberculostearic acid). The G+C content of the type-strain genome is 70.6%, its approximate size 3.54 Mbp, its IMG deposit 2728369259.
The description is as before (Rainey et al., 1995b) with the following modification. The G+C content of the type-strain genome is 70.9%, its approximate size 3.51 Mbp, its GenBank deposit SAMN04543680.
The description is as before (Maruo et al., 2008) with the following modification. The G+C content of the type-strain genome is 64.2%, its approximate size 3.63 Mbp, its GenBank deposit SAMN00002594.
The description is as before (Hammann et al., 1984) with the following modification. The G+C content of the type-strain genome is 55.4%, its approximate size 2.15 Mbp, its GenBank deposit SAMN00001505.
The description is as before (Johnson et al., 2009) with the following restriction. The G+C content of the type-strain genome is 51.1%, its approximate size 2.49 Mbp, its GenBank deposit SAMN02745225.
The description is as before (Greenwood and Pickett, 1980) with the following restriction. The G+C content of the type-strain genome is 41.3%, its approximate size 1.68 Mbp, its GenBank deposit SAMN04488545.
The description is as before (Kämpfer et al., 2010a) with the following addition. The G+C content of the type-strain genome is 72.7%, its approximate size 4.27 Mbp, its GenBank deposit SAMN04488547.
The description is as before (Katayama et al., 2009) with the following modification. The G+C content of the type-strain genome is 64.4%, its approximate size 4.80 Mbp, its GenBank deposit SAMN02440580.
The description is as before (Busse, 2016) with the following addition. The G+C content of the type-strain genome is 59.3%, its approximate size 3.92 Mbp, its GenBank deposit SAMEA2272213.
The description is as before (Busse, 2016) with the following addition. The G+C content of the type-strain genome is 62.0%, its approximate size 3.46 Mbp, its GenBank deposit SAMN04489744.
The description is as before (Labeda and Kroppenstedt, 2004) with the following addition. The G+C content of the type-strain genome is 70.1%, its approximate size 5.05 Mbp, its GenBank deposit SAMN02441107.
The description is as before (Gu et al., 2007) with the following modification. The G+C content of the type-strain genome is 71.8%, its approximate size 5.60 Mbp, its GenBank deposit SAMN05216298.
The description is as before (Evtushenko et al., 1991) with the following modification. The G+C content of the type-strain genome is 70.4%, its approximate size 5.73 Mbp, its GenBank deposit SAMN02441561.
The description is as before (Labeda et al., 2008) with the following modification. The G+C content of the type-strain genome is 72.4%, its approximate size 8.77 Mbp, its GenBank deposit SAMN05660962.
The description is as before (Klatte et al., 1994) with the following restriction. The G+C content of the type-strain genome is 67.4%, its approximate size 5.31 Mbp, its GenBank deposit SAMD00041822.
The description is as before (Kim et al., 2000) with the following modification. The G+C content of the type-strain genome is 67.4%, its approximate size 4.92 Mbp, its GenBank deposit SAMD00041764.
The description is as before (Kageyama et al., 2006) with the following addition. The G+C content of the type-strain genome is 68.0%, its approximate size 3.91 Mbp, its GenBank deposit SAMD00041783.
The description is as before (Stackebrandt et al., 1988) with the following addition. The G+C content of the type-strain genome is 67.1%, its approximate size 5.21 Mbp, its GenBank deposit SAMN00002600.
The description is as before (Kim et al., 1999) with the following modification. The G+C content of the type-strain genome is 68.1%, its approximate size 5.43 Mbp, its GenBank deposit SAMD00018679.
The description is as before (Kageyama et al., 2006) with the following addition. The G+C content of the type-strain genome is 62.5%, its approximate size 4.70 Mbp, its GenBank deposit SAMD00041782.
The description is as before (Bendinger et al., 1995) with the following modification. The G+C content of the type-strain genome is 67.5%, its approximate size 4.58 Mbp, its GenBank deposit SAMD00047212.
The description is as before (le Roes et al., 2008) with the following addition. The G+C content of the type-strain genome is 68.1%, its approximate size 5.76 Mbp, its GenBank deposit SAMN06928587.
The description is as before (Yassin et al., 2007a) with the following addition. The G+C content of the type-strain genome is 66.2%, its approximate size 4.71 Mbp, its GenBank deposit SAMN04488550.
The description is as before (Liu et al., 2011) with the following addition. The G+C content of the type-strain genome is 68.2%, its approximate size 4.26 Mbp, its GenBank deposit SAMN02470983.
The description is as before (Linos et al., 1999) with the following addition. The G+C content of the type-strain genome is 66.9%, its approximate size 6.29 Mbp, its GenBank deposit SAMD00041829.
The description is as before (Stackebrandt et al., 1988) with the following restriction. The G+C content of the type-strain genome is 67.4%, its approximate size 5.20 Mbp, its GenBank deposit SAMD00041793.
The description is as before (Luo et al., 2007) with the following addition. The G+C content of the type-strain genome is 69.3%, its approximate size 3.33 Mbp, its GenBank deposit SAMN02440656.
The description is as before (Stackebrandt et al., 1988) with the following addition. The G+C content of the type-strain genome is 67.8%, its approximate size 5.67 Mbp, its GenBank deposit SAMD00041763.
The description is as before (Linos et al., 2002) with the following addition. The G+C content of the type-strain genome is 66.9%, its approximate size 6.41 Mbp, its GenBank deposit SAMN04488548.
The description is as before (Guan et al., 2009) with the following addition. The G+C content of the type-strain genome is 60.4%, its approximate size 3.54 Mbp, its GenBank deposit SAMN02597195.
The description is as before (Behrendt et al., 2011) with the following modification. The G+C content of the type-strain genome is 68.4%, its approximate size 4.85 Mbp, its GenBank deposit SAMN05216554.
The description is as before (Behrendt et al., 2011) with the following addition. The G+C content of the type-strain genome is 70.2%, its approximate size 3.85 Mbp, its GenBank deposit SAMD00046479.
The description is as before (Kudo et al., 1993) with the following restriction. The G+C content of the type-strain genome is 70.7%, its approximate size 8.28 Mbp, its GenBank deposit SAMD00029379.
The description is as before (Boondaeng et al., 2011) with the following modification. The G+C content of the type-strain genome is 71.0%, its approximate size 8.58 Mbp, its GenBank deposit SAMD00029380.
The description is as before (Holt and Lewin, 1968) with the following restriction. The G+C content of the type-strain genome is 50.9%, its approximate size 6.79 Mbp, its GenBank deposit SAMN00623048.
The description is as before (Lewin, 1970) with the following modification. The G+C content of the type-strain genome is 50.7%, its approximate size 6.14 Mbp, its GenBank deposit SAMN03274862.
The description is as before (Matsumoto et al., 2013) with the following modification. The G+C content of the type-strain genome is 67.0%, its approximate size 4.28 Mbp, its GenBank deposit SAMD00036665.
The description is as before (Yang et al., 2012) with the following addition. The G+C content of the type-strain genome is 70.7%, its approximate size 4.02 Mbp, its GenBank deposit SAMN00713569.
The description is as before (Yang et al., 2012) with the following modification. The G+C content of the type-strain genome is 71.4%, its approximate size 4.64 Mbp, its GenBank deposit SAMN02952955.
The description is as before (Wu et al., 2010) with the following modification. The G+C content of the type-strain genome is 73.8%, its approximate size 4.04 Mbp, its GenBank deposit SAMN04488560.
The description is as before (Shivaji et al., 2009) with the following modification. The G+C content of the type-strain genome is 71.3%, its approximate size 3.14 Mbp, its GenBank deposit SAMN02470608.
The description is as before (Zhang et al., 2014) with the following modification. The G+C content of the type-strain genome is 71.2%, its approximate size 3.42 Mbp, its GenBank deposit SAMN06296429.
The description is as before (Lee et al., 2018) with the following modification. The G+C content of the type-strain genome is 72.9%, its approximate size 4.48 Mbp, its GenBank deposit SAMN05443575.
The description is as before (Kämpfer et al., 2011) with the following addition. The G+C content of the type-strain genome is 72.3%, its approximate size 7.26 Mbp, its GenBank deposit SAMN03801508.
The description is as before (Rocourt et al., 1987) with the following addition. The G+C content of the type-strain genome is 58.4%, its approximate size 2.75 Mbp, its GenBank deposit SAMN02598439.
The description is as before (Cole et al., 2013) with the following modification. The G+C content of the type-strain genome is 65.8%, its approximate size 4.48 Mbp, its GenBank deposit SAMN02584935.
The description is as before (Xing et al., 2012) with the following modification. The G+C content of the type-strain genome is 68.4%, its approximate size 11.76 Mbp, its GenBank deposit SAMN03840758.
The description is as before (Phillips et al., 2002) with the following addition. The G+C content of the type-strain genome is 74.2%, its approximate size 4.96 Mbp, its GenBank deposit SAMN02598259.
The description is as before (Liu et al., 2009a) with the following modification. The G+C content of the type-strain genome is 74.6%, its approximate size 4.62 Mbp, its IMG deposit 2731639219.
The description is as before (Matsumae et al., 1968) with the following addition. The G+C content of the type-strain genome is 71.5%, its approximate size 7.46 Mbp, its GenBank deposit SAMN02261248.
The description is as before (Zhang et al., 1997) with the following modification. The G+C content of the type-strain genome is 71.6%, its approximate size 8.27 Mbp, its GenBank deposit SAMN02745392.
The description is as before (Chung et al., 1999) with the following modification. The G+C content of the type-strain genome is 73.5%, its approximate size 8.04 Mbp, its GenBank deposit SAMN02796068.
The description is as before (Labeda, 1988) with the following modification. The G+C content of the type-strain genome is 71.9%, its approximate size 8.68 Mbp, its GenBank deposit SAMN02745411.
The description is as before (Takahashi et al., 1984) with the following modification. The G+C content of the type-strain genome is 74.1%, its approximate size 7.62 Mbp, its GenBank deposit SAMN02645204.
The description is as before (Omura et al., 1982) with the following addition. The G+C content of the type-strain genome is 74.2%, its approximate size 8.78 Mbp, its GenBank deposit SAMD00060928.
The description is as before (Weon et al., 2007) with the following modification. The G+C content of the type-strain genome is 71.3%, its approximate size 4.09 Mbp, its GenBank deposit SAMN03144715.
The description is as before (Zhou et al., 2008) with the following modification. The G+C content of the type-strain genome is 73.9%, its approximate size 3.64 Mbp, its GenBank deposit SAMN04270876.
The description is as before (Dastager et al., 2014c) with the following modification. The G+C content of the type-strain genome is 68.9%, its approximate size 2.88 Mbp, its GenBank deposit SAMN06296028.
The description is as before (Camacho et al., 2017) with the following modification. The G+C content of the type-strain genome is 72.8%, its approximate size 4.16 Mbp, its GenBank deposit SAMN04455734.
The description is as before (Lee and Lee, 2010b) with the following modification. The G+C content of the type-strain genome is 70.7%, its approximate size 2.99 Mbp, its GenBank deposit SAMN04489860.
The description is as before (Urz et al., 2008) with the following addition. The G+C content of the type-strain genome is 67.5%, its approximate size 9.63 Mbp, its GenBank deposit SAMN02256503.
The description is as before (Suriyachadkun et al., 2013) with the following modification. The G+C content of the type-strain genome is 70.4%, its approximate size 11.97 Mbp, its GenBank deposit SAMN05421845.
The description is as before (Kämpfer et al., 2009b) with the following addition. The G+C content of the type-strain genome is 72.3%, its approximate size 2.40 Mbp, its IMG deposit 2734482797.
The description is as before (Stackebrandt et al., 1995) with the following addition. The G+C content of the type-strain genome is 71.6%, its approximate size 2.79 Mbp, its GenBank deposit SAMN02598443.
The description is as before (Lee, 2007) with the following modification. The G+C content of the type-strain genome is 69.2%, its approximate size 3.84 Mbp, its IMG deposit 2734482166.
The description is as before (Reddy et al., 2003) with the following modification. The G+C content of the type-strain genome is 59.2%, its approximate size 2.78 Mbp, its GenBank deposit SAMN02469437.
The description is as before (Labeda et al., 2001) with the following addition. The G+C content of the type-strain genome is 70.2%, its approximate size 9.44 Mbp, its GenBank deposit SAMN04488000.
The description is as before (Labeda et al., 2001) with the following addition. The G+C content of the type-strain genome is 69.3%, its approximate size 9.00 Mbp, its GenBank deposit SAMN05660743.
The description is as before (Labeda et al., 2007) with the following addition. The G+C content of the type-strain genome is 68.8%, its approximate size 10.21 Mbp, its GenBank deposit SAMN06294978.
The description is as before (Labeda et al., 2001) with the following modification. The G+C content of the type-strain genome is 68.9%, its approximate size 10.15 Mbp, its GenBank deposit SAMN04488564.
The description is as before (Yamada et al., 2006) with the following modification. The G+C content of the type-strain genome is 46.8%, its approximate size 3.68 Mbp, its GenBank deposit SAMN03856817.
The description is as before (Halpern et al., 2009) with the following addition. The G+C content of the type-strain genome is 69.9%, its approximate size 2.96 Mbp, its GenBank deposit SAMN02441232.
The description is as before (Sturm et al., 2011) with the following addition. The G+C content of the type-strain genome is 70.3%, its approximate size 3.21 Mbp, its GenBank deposit SAMN04488565.
The description is as before (Yun et al., 2011) with the following modification. The G+C content of the type-strain genome is 64.5%, its approximate size 3.19 Mbp, its GenBank deposit SAMN02470216.
The description is as before (Juboi et al., 2015) with the following modification. The G+C content of the type-strain genome is 70.1%, its approximate size 4.46 Mbp, its GenBank deposit SAMN03421050.
The description is as before (Lee and Lee, 2010c) with the following modification. The G+C content of the type-strain genome is 73.5%, its approximate size 4.06 Mbp, its GenBank deposit SAMN04488570.
The description is as before (Young et al., 2010) with the following addition. The G+C content of the type-strain genome is 64.5%, its approximate size 3.23 Mbp, its GenBank deposit SAMN04488572.
The description is as before (Mawlankar et al., 2015) with the following modification. The G+C content of the type-strain genome is 70.4%, its approximate size 3.66 Mbp, its GenBank deposit SAMN04263881.
The description is as before (Behrendt et al., 2001) with the following modification. The G+C content of the type-strain genome is 68.7%, its approximate size 3.56 Mbp, its GenBank deposit SAMN03256325.
The description is as before (Brennan et al., 2001b) with the following restriction. The G+C content of the type-strain genome is 68.1%, its approximate size 3.02 Mbp, its GenBank deposit SAMN02440649.
The description is as before (Young et al., 2010) with the following addition. The G+C content of the type-strain genome is 67.3%, its approximate size 3.36 Mbp, its GenBank deposit SAMN04489806.
The description is as before (Schippers et al., 2005) with the following addition. The G+C content of the type-strain genome is 68.6%, its approximate size 3.59 Mbp, its GenBank deposit SAMN04489807.
The description is as before (Shivaji et al., 2007) with the following modification. The G+C content of the type-strain genome is 71.4%, its approximate size 2.81 Mbp, its GenBank deposit SAMN02440686.
The description is as before (Vaz-Moreira et al., 2008) with the following modification. The G+C content of the type-strain genome is 70.7%, its approximate size 3.11 Mbp, its GenBank deposit SAMN02440801.
The description is as before (Lee et al., 2014a) with the following restriction. The G+C content of the type-strain genome is 70.0%, its approximate size 4.42 Mbp, its GenBank deposit SAMN03070121.
The description is as before (Schippers et al., 2005) with the following addition. The G+C content of the type-strain genome is 69.0%, its approximate size 2.98 Mbp, its GenBank deposit SAMD00046492.
The description is as before (Behrendt et al., 2001) with the following addition. The G+C content of the type-strain genome is 71.2%, its approximate size 3.98 Mbp, its GenBank deposit SAMD00046494.
The description is as before (Takeuchi and Hatano, 1998) with the following modification. The G+C content of the type-strain genome is 70.2%, its approximate size 4.52 Mbp, its GenBank deposit SAMN03266141.
The description is as before (Nonomura and Ohara, 1957) with the following addition. The G+C content of the type-strain genome is 71.2%, its approximate size 8.86 Mbp, its GenBank deposit SAMN05421833.
The description is as before (Wieser et al., 2002) with the following addition. The G+C content of the type-strain genome is 73.0%, its approximate size 2.50 Mbp, its GenBank deposit SAMN00000281.
The description is as before (Wieser et al., 2002) with the following modification. The G+C content of the type-strain genome is 71.3%, its approximate size 2.69 Mbp, its GenBank deposit SAMD00046485.
The description is as before (Zhang et al., 2010b) with the following modification. The G+C content of the type-strain genome is 68.9%, its approximate size 3.09 Mbp, its GenBank deposit SAMN04487966.
The description is as before (Kämpfer et al., 2010d) with the following addition. The G+C content of the type-strain genome is 67.4%, its approximate size 6.73 Mbp, its GenBank deposit SAMN04489812.
The description is as before (Li et al., 2013) with the following modification. The G+C content of the type-strain genome is 71.5%, its approximate size 6.82 Mbp, its GenBank deposit SAMN05444858.
The description is as before (Horan and Brodsky, 1986) with the following modification. The G+C content of the type-strain genome is 73.4%, its approximate size 7.38 Mbp, its GenBank deposit SAMN04440589.
The description is as before (Ren et al., 2013) with the following modification. The G+C content of the type-strain genome is 73.0%, its approximate size 7.51 Mbp, its GenBank deposit SAMN06554453.
The description is as before (Thiemann et al., 1968) with the following addition. The G+C content of the type-strain genome is 70.9%, its approximate size 9.02 Mbp, its GenBank deposit SAMD00035248.
The description is as before (Thiemann et al., 1968) with the following addition. The G+C content of the type-strain genome is 70.2%, its approximate size 9.27 Mbp, its GenBank deposit SAMD00035249.
The description is as before (Nakajima et al., 2003) with the following modification. The G+C content of the type-strain genome is 70.7%, its approximate size 8.82 Mbp, its GenBank deposit SAMD00035251.
The description is as before (Nonomura and Ohara, 1971a) with the following addition. The G+C content of the type-strain genome is 71.7%, its approximate size 8.47 Mbp, its GenBank deposit SAMD00035250.
The description is as before (Hoyles et al., 2004) with the following restriction. The G+C content of the type-strain genome is 55.7%, its approximate size 2.14 Mbp, its GenBank deposit SAMN00117214.
The description is as before (Hoyles et al., 2004) with the following restriction. The G+C content of the type-strain genome is 55.1%, its approximate size 2.40 Mbp, its GenBank deposit SAMN00139192.
The description is as before (Lee, 2012) with the following modification. The G+C content of the type-strain genome is 73.6%, its approximate size 4.19 Mbp, its IMG deposit 2731639228.
The description is as before (Kämpfer et al., 2010c) with the following addition. The G+C content of the type-strain genome is 72.4%, its approximate size 7.38 Mbp, its IMG deposit 2728369519.
The description is as before (Tsukamoto et al., 2001) with the following modification. The G+C content of the type-strain genome is 66.1%, its approximate size 3.45 Mbp, its GenBank deposit SAMN02645226.
The description is as before (Tortoli et al., 2016a) with the following restriction. The G+C content of the type-strain genome is 64.1%, its approximate size 5.05 Mbp, its GenBank deposit SAMN02470911.
The description is as before (Tortoli et al., 2016a) with the following restriction. The G+C content of the type-strain genome is 64.1%, its approximate size 4.98 Mbp, its GenBank deposit AP014547.
The description is as before (Sahraoui et al., 2011) with the following addition. The G+C content of the type-strain genome is 68.3%, its approximate size 4.62 Mbp, its GenBank deposit SAMN06064225.
The description is as before (Tortoli et al., 2016b) with the following addition. The G+C content of the type-strain genome is 69.3%, its approximate size 5.69 Mbp, its GenBank deposit SAMN06064226.
The description is as before (Hennessee et al., 2009) with the following addition. The G+C content of the type-strain genome is 66.4%, its approximate size 6.30 Mbp, its GenBank deposit SAMN02585141.
The description is as before (Bang et al., 2008) with the following addition. The G+C content of the type-strain genome is 66.8%, its approximate size 5.98 Mbp, its GenBank deposit SAMN06064229.
The description is as before (Cloud et al., 2006) with the following addition. The G+C content of the type-strain genome is 67.4%, its approximate size 4.44 Mbp, its GenBank deposit SAMN06064230.
The description is as before (Weiszfeiler et al., 1971) with the following addition. The G+C content of the type-strain genome is 66.3%, its approximate size 5.91 Mbp, its GenBank deposit SAMEA3138999.
The description is as before (Tsukamura, 1966) with the following addition. The G+C content of the type-strain genome is 67.5%, its approximate size 6.02 Mbp, its GenBank deposit SAMEA3216266.
The description is as before (Tsukamura et al., 1983) with the following addition. The G+C content of the type-strain genome is 67.7%, its approximate size 6.68 Mbp, its GenBank deposit SAMEA3138973.
The description is as before (Schinsky et al., 2004) with the following modification. The G+C content of the type-strain genome is 66.8%, its approximate size 6.51 Mbp, its GenBank deposit SAMEA92028418.
The description is as before (Reischl et al., 1998) with the following modification. The G+C content of the type-strain genome is 68.8%, its approximate size 5.08 Mbp, its GenBank deposit SAMEA3305052.
The description is as before (Ben Salah et al., 2009) with the following addition. The G+C content of the type-strain genome is 68.6%, its approximate size 5.90 Mbp, its GenBank deposit SAMN06064234.
The description is as before (Koukila-Kähkölä et al., 1995) with the following addition. The G+C content of the type-strain genome is 66.5%, its approximate size 5.90 Mbp, its GenBank deposit SAMN06064235.
The description is as before (Butler et al., 1993) with the following addition. The G+C content of the type-strain genome is 67.0%, its approximate size 4.72 Mbp, its GenBank deposit SAMN04216916.
The description is as before (Shahraki et al., 2015) with the following addition. The G+C content of the type-strain genome is 66.9%, its approximate size 4.95 Mbp, its GenBank deposit SAMN06064236.
The description is as before (Kim et al., 2017) with the following addition. The G+C content of the type-strain genome is 63.9%, its approximate size 5.03 Mbp, its GenBank deposit SAMN02630629.
The description is as before (Hägglblom et al., 1994) with the following restriction. The G+C content of the type-strain genome is 68.4%, its approximate size 7.33 Mbp, its GenBank deposit SAMD00046518.
The description is as before (Tsukamura, 1981) with the following addition. The G+C content of the type-strain genome is 69.2%, its approximate size 5.94 Mbp, its GenBank deposit SAMN03375743.
The description is as before (Murcia et al., 2006) with the following addition. The G+C content of the type-strain genome is 68.1%, its approximate size 5.58 Mbp, its GenBank deposit SAMN00622208.
The description is as before (Adékambi et al., 2006) with the following restriction. The G+C content of the type-strain genome is 66.3%, its approximate size 6.38 Mbp, its GenBank deposit SAMEA3305051.
The description is as before (Kirschner et al., 1992) with the following addition. The G+C content of the type-strain genome is 67.5%, its approximate size 5.84 Mbp, its GenBank deposit SAMN04216920.
The description is as before (Springer et al., 1996) with the following addition. The G+C content of the type-strain genome is 67.4%, its approximate size 6.20 Mbp, its GenBank deposit SAMN04216921.
The description is as before (Cooksey et al., 2004) with the following addition. The G+C content of the type-strain genome is 68.2%, its approximate size 6.46 Mbp, its GenBank deposit SAMEA3138996.
The description is as before (Tortoli et al., 2001) with the following modification. The G+C content of the type-strain genome is 67.6%, its approximate size 3.95 Mbp, its GenBank deposit SAMN04216922.
The description is as before (Tortoli et al., 2013) with the following addition. The G+C content of the type-strain genome is 68.5%, its approximate size 4.52 Mbp, its GenBank deposit SAMN04216923.
The description is as before (Tortoli et al., 2011) with the following addition. The G+C content of the type-strain genome is 68.5%, its approximate size 5.63 Mbp, its GenBank deposit SAMN04216924.
The description is as before (Lévy-Frébault et al., 1983) with the following addition. The G+C content of the type-strain genome is 70.3%, its approximate size 4.23 Mbp, its GenBank deposit SAMN04216925.
The description is as before (Tortoli et al., 2005) with the following addition. The G+C content of the type-strain genome is 66.4%, its approximate size 6.18 Mbp, its GenBank deposit SAMN04216926.
The description is as before (da Costa Cruz, 1938) with the following addition. The G+C content of the type-strain genome is 66.4%, its approximate size 5.90 Mbp, its GenBank deposit SAMN04295031.
The description is as before (Ramos et al., 2013) with the following addition. The G+C content of the type-strain genome is 66.0%, its approximate size 4.73 Mbp, its GenBank deposit SAMN04216927.
The description is as before (Nogueira et al., 2015) with the following addition. The G+C content of the type-strain genome is 64.1%, its approximate size 5.43 Mbp, its GenBank deposit SAMN05186797.
The description is as before (Böttger et al., 1993) with the following addition. The G+C content of the type-strain genome is 66.9%, its approximate size 4.94 Mbp, its GenBank deposit SAMN02585006.
The description is as before (Bojalil et al., 1962) with the following addition. The G+C content of the type-strain genome is 66.6%, its approximate size 7.60 Mbp, its GenBank deposit SAMN04216929.
The description is as before (Sompolinsky et al., 1978) with the following modification. The G+C content of the type-strain genome is 63.9%, its approximate size 4.24 Mbp, its GenBank deposit SAMN01918491.
The description is as before (Schröder et al., 1997) with the following modification. The G+C content of the type-strain genome is 69.4%, its approximate size 5.08 Mbp, its GenBank deposit SAMN02256416.
The description is as before (Haas et al., 1997) with the following addition. The G+C content of the type-strain genome is 68.0%, its approximate size 5.00 Mbp, its GenBank deposit SAMN06064240.
The description is as before (Kazda et al., 1993) with the following addition. The G+C content of the type-strain genome is 68.5%, its approximate size 4.34 Mbp, its GenBank deposit SAMN04216930.
The description is as before (Schinsky et al., 2004) with the following modification. The G+C content of the type-strain genome is 67.0%, its approximate size 6.38 Mbp, its GenBank deposit SAMEA3913388.
The description is as before (Wilson et al., 2001) with the following addition. The G+C content of the type-strain genome is 64.3%, its approximate size 5.57 Mbp, its GenBank deposit SAMN03733495.
The description is as before (Springer et al., 1993) with the following addition. The G+C content of the type-strain genome is 67.9%, its approximate size 5.85 Mbp, its GenBank deposit SAMEA3913390.
The description is as before (Meier et al., 1993) with the following addition. The G+C content of the type-strain genome is 65.8%, its approximate size 6.82 Mbp, its GenBank deposit SAMN06064242.
The description is as before (Shojaei et al., 2013) with the following addition. The G+C content of the type-strain genome is 66.1%, its approximate size 6.34 Mbp, its GenBank deposit SAMN04216933.
The description is as before (Hauduroy, 1955) with the following addition. The G+C content of the type-strain genome is 66.2%, its approximate size 6.58 Mbp, its GenBank deposit SAMN02603596.
The description is as before (Kim et al., 2012a) with the following addition. The G+C content of the type-strain genome is 69.4%, its approximate size 4.08 Mbp, its GenBank deposit SAMN06651658.
The description is as before (Floyd et al., 2000) with the following addition. The G+C content of the type-strain genome is 66.0%, its approximate size 5.83 Mbp, its GenBank deposit SAMN04216934.
The description is as before (Masaki et al., 2006) with the following addition. The G+C content of the type-strain genome is 68.1%, its approximate size 4.82 Mbp, its GenBank deposit SAMN06064243.
The description is as before (Okazaki et al., 2009) with the following addition. The G+C content of the type-strain genome is 66.9%, its approximate size 5.30 Mbp, its GenBank deposit SAMD00019087.
The description is as before (Turenne et al., 2002) with the following addition. The G+C content of the type-strain genome is 66.9%, its approximate size 4.91 Mbp, its GenBank deposit SAMN04216936.
The description is as before (Tortoli et al., 2013) with the following addition. The G+C content of the type-strain genome is 67.9%, its approximate size 4.81 Mbp, its GenBank deposit SAMN04216937.
The description is as before (Domenech et al., 1997) with the following addition. The G+C content of the type-strain genome is 66.9%, its approximate size 7.97 Mbp, its GenBank deposit SAMEA3138978.
The description is as before (van Ingen et al., 2009d) with the following addition. The G+C content of the type-strain genome is 66.9%, its approximate size 6.12 Mbp, its GenBank deposit SAMN06064245.
The description is as before (Ben Salah et al., 2009) with the following addition. The G+C content of the type-strain genome is 67.7%, its approximate size 5.46 Mbp, its GenBank deposit SAMN06064246.
The description is as before (Hannigan et al., 2013) with the following addition. The G+C content of the type-strain genome is 67.1%, its approximate size 4.19 Mbp, its GenBank deposit SAMN06064248.
The description is as before (Reischl et al., 2006) with the following addition. The G+C content of the type-strain genome is 68.4%, its approximate size 6.00 Mbp, its GenBank deposit SAMN06064249.
The description is as before (Tsukamura et al., 1986) with the following modification. The G+C content of the type-strain genome is 66.0%, its approximate size 6.22 Mbp, its GenBank deposit SAMN06064251.
The description is as before (Mohamed et al., 2004) with the following addition. The G+C content of the type-strain genome is 66.6%, its approximate size 6.73 Mbp, its GenBank deposit SAMN04216938.
The description is as before (Schinsky et al., 2004) with the following modification. The G+C content of the type-strain genome is 66.8%, its approximate size 6.29 Mbp, its GenBank deposit SAMEA3481574.
The description is as before (van Ingen et al., 2009b) with the following addition. The G+C content of the type-strain genome is 65.8%, its approximate size 4.74 Mbp, its GenBank deposit SAMN06064252.
The description is as before (Tsukamura et al., 1981) with the following addition. The G+C content of the type-strain genome is 68.0%, its approximate size 5.71 Mbp, its GenBank deposit SAMN02952053.
The description is as before (Torkko et al., 2002) with the following addition. The G+C content of the type-strain genome is 68.5%, its approximate size 6.04 Mbp, its GenBank deposit SAMN04216940.
The description is as before (Fusco da Costa et al., 2015) with the following addition. The G+C content of the type-strain genome is 69.3%, its approximate size 5.62 Mbp, its GenBank deposit SAMN04216942.
The description is as before (Lee et al., 2016) with the following addition. The G+C content of the type-strain genome is 68.1%, its approximate size 5.50 Mbp, its GenBank deposit SAMN02603185.
The description is as before (Turenne et al., 2004a) with the following addition. The G+C content of the type-strain genome is 68.4%, its approximate size 6.30 Mbp, its GenBank deposit SAMN00189881.
The description is as before (Lee et al., 2010) with the following addition. The G+C content of the type-strain genome is 67.9%, its approximate size 6.08 Mbp, its GenBank deposit SAMN06064254.
The description is as before (Fanti et al., 2004) with the following addition. The G+C content of the type-strain genome is 68.4%, its approximate size 5.89 Mbp, its GenBank deposit SAMN04216945.
The description is as before (Kusunoki and Ezaki, 1992) with the following addition. The G+C content of the type-strain genome is 66.3%, its approximate size 7.05 Mbp, its GenBank deposit SAMN04216946.
The description is as before (Lehmann and Neumann, 1899) with the following addition. The G+C content of the type-strain genome is 69.4%, its approximate size 5.32 Mbp, its GenBank deposit SAMN04537322.
The description is as before (Rhodes et al., 2005) with the following addition. The G+C content of the type-strain genome is 65.7%, its approximate size 5.94 Mbp, its GenBank deposit SAMD00042842.
The description is as before (Tsukamura, 1981) with the following addition. The G+C content of the type-strain genome is 66.6%, its approximate size 6.01 Mbp, its GenBank deposit SAMN06064257.
The description is as before (van Ingen et al., 2009a) with the following addition. The G+C content of the type-strain genome is 65.3%, its approximate size 6.27 Mbp, its GenBank deposit SAMN04216947.
The description is as before (Hennessee et al., 2009) with the following addition. The G+C content of the type-strain genome is 69.2%, its approximate size 6.18 Mbp, its GenBank deposit SAMN02777056.
The description is as before (Hennessee et al., 2009) with the following addition. The G+C content of the type-strain genome is 68.4%, its approximate size 5.99 Mbp, its GenBank deposit SAMN04489835.
The description is as before (Whipps et al., 2007) with the following addition. The G+C content of the type-strain genome is 64.3%, its approximate size 4.77 Mbp, its GenBank deposit SAMN05186798.
The description is as before (Turenne et al., 2004b) with the following addition. The G+C content of the type-strain genome is 68.3%, its approximate size 5.93 Mbp, its GenBank deposit SAMN04216948.
The description is as before (Prissick and Masson, 1956) with the following addition. The G+C content of the type-strain genome is 68.4%, its approximate size 6.17 Mbp, its GenBank deposit SAMN06064259.
The description is as before (Mun et al., 2008) with the following addition. The G+C content of the type-strain genome is 68.7%, its approximate size 4.53 Mbp, its GenBank deposit SAMN04216949.
The description is as before (Schinsky et al., 2000) with the following modification. The G+C content of the type-strain genome is 66.7%, its approximate size 6.87 Mbp, its GenBank deposit SAMEA2272699.
The description is as before (Lamy et al., 2008) with the following addition. The G+C content of the type-strain genome is 66.4%, its approximate size 6.27 Mbp, its GenBank deposit SAMN01174788.
The description is as before (van Ingen et al., 2011) with the following addition. The G+C content of the type-strain genome is 67.0%, its approximate size 5.56 Mbp, its GenBank deposit SAMN04216950.
The description is as before (Tsukamura, 1982a) with the following addition. The G+C content of the type-strain genome is 65.8%, its approximate size 4.71 Mbp, its GenBank deposit SAMN04216951.
The description is as before (Saito et al., 2011) with the following addition. The G+C content of the type-strain genome is 67.8%, its approximate size 4.41 Mbp, its GenBank deposit SAMN06064260.
The description is as before (Karassova et al., 1965) with the following addition. The G+C content of the type-strain genome is 66.2%, its approximate size 5.69 Mbp, its GenBank deposit SAMEA2272207.
The description is as before (Lehmann and Neumann, 1899) with the following addition. The G+C content of the type-strain genome is 67.3%, its approximate size 6.98 Mbp, its GenBank deposit SAMEA2517361.
The description is as before (Marks and Jenkins, 1972) with the following addition. The G+C content of the type-strain genome is 65.8%, its approximate size 6.67 Mbp, its GenBank deposit SAMN04216953.
The description is as before (Tsukamura, 1966) with the following addition. The G+C content of the type-strain genome is 69.0%, its approximate size 4.87 Mbp, its GenBank deposit SAMN02471087.
The description is as before (Floyd et al., 1996) with the following addition. The G+C content of the type-strain genome is 66.6%, its approximate size 6.38 Mbp, its GenBank deposit SAMEA3139001.
The description is as before (Bönicke and Juhasz, 1964) with the following addition. The G+C content of the type-strain genome is 68.6%, its approximate size 6.24 Mbp, its GenBank deposit SAMN03702434.
The description is as before (Khan et al., 2002) with the following modification. The G+C content of the type-strain genome is 67.8%, its approximate size 6.49 Mbp, its GenBank deposit SAMN02598347.
The description is as before (van Ingen et al., 2009c) with the following addition. The G+C content of the type-strain genome is 66.8%, its approximate size 6.27 Mbp, its GenBank deposit SAMN06651660.
The description is as before (Brown et al., 1999) with the following addition. The G+C content of the type-strain genome is 66.4%, its approximate size 7.52 Mbp, its GenBank deposit SAMN04216957.
The description is as before (Schwabacher, 1959) with the following addition. The G+C content of the type-strain genome is 65.9%, its approximate size 4.93 Mbp, its GenBank deposit SAMN04216958.
The description is as before (Tao et al., 2004) with the following modification. The G+C content of the type-strain genome is 70.9%, its approximate size 6.06 Mbp, its GenBank deposit SAMN02598467.
The description is as before (Luo et al., 2009) with the following modification. The G+C content of the type-strain genome is 63.7%, its approximate size 2.59 Mbp, its GenBank deposit SAMN02440588.
The description is as before (Sorokin et al., 2009) with the following modification. The G+C content of the type-strain genome is 72.1%, its approximate size 5.56 Mbp, its GenBank deposit SAMN02745916.
The description is as before (Hamid et al., 2001) with the following addition. The G+C content of the type-strain genome is 67.9%, its approximate size 7.81 Mbp, its GenBank deposit SAMD00018765.
The description is as before (Ezeoke et al., 2013) with the following restriction. The G+C content of the type-strain genome is 68.5%, its approximate size 7.66 Mbp, its GenBank deposit SAMN04244555.
The description is as before (Kageyama et al., 2004a) with the following restriction. The G+C content of the type-strain genome is 68.4%, its approximate size 8.46 Mbp, its GenBank deposit SAMD00041765.
The description is as before (Blanchard, 1896) with the following addition. The G+C content of the type-strain genome is 69.9%, its approximate size 6.95 Mbp, its GenBank deposit SAMN05444423.
The description is as before (Pinoy, 1913) with the following addition. The G+C content of the type-strain genome is 68.2%, its approximate size 8.90 Mbp, its GenBank deposit SAMD00016800.
The description is as before (Goodfellow and Pirouz, 1982) with the following addition. The G+C content of the type-strain genome is 67.0%, its approximate size 7.01 Mbp, its GenBank deposit SAMD00041812.
The description is as before (Castellani and Chalmers, 1913) with the following addition. The G+C content of the type-strain genome is 67.1%, its approximate size 7.49 Mbp, its GenBank deposit SAMD00041818.
The description is as before (Friedman et al., 1998) with the following modification. The G+C content of the type-strain genome is 67.7%, its approximate size 8.29 Mbp, its GenBank deposit SAMD00040648.
The description is as before (Trevisan, 1889) with the following addition. The G+C content of the type-strain genome is 70.7%, its approximate size 6.41 Mbp, its GenBank deposit SAMN05421776.
The description is as before (Maldonado et al., 2000) with the following addition. The G+C content of the type-strain genome is 67.5%, its approximate size 8.06 Mbp, its GenBank deposit SAMN04489843.
The description is as before (Kageyama et al., 2004c) with the following modification. The G+C content of the type-strain genome is 67.8%, its approximate size 8.12 Mbp, its GenBank deposit SAMD00040655.
The description is as before (Lee, 2006b) with the following modification. The G+C content of the type-strain genome is 67.6%, its approximate size 8.65 Mbp, its GenBank deposit SAMD00018769.
The description is as before (Cui et al., 2005) with the following addition. The G+C content of the type-strain genome is 66.8%, its approximate size 10.45 Mbp, its GenBank deposit SAMD00041792.
The description is as before (Conville et al., 2004) with the following addition. The G+C content of the type-strain genome is 68.0%, its approximate size 7.32 Mbp, its GenBank deposit SAMD00040657.
The description is as before (Kageyama et al., 2004c) with the following modification. The G+C content of the type-strain genome is 68.2%, its approximate size 8.22 Mbp, its GenBank deposit SAMD00041766.
The description is as before (Tsukamura, 1982b) with the following addition. The G+C content of the type-strain genome is 67.9%, its approximate size 7.85 Mbp, its GenBank deposit SAMD00040661.
The description is as before (Snijders, 1924) with the following addition. The G+C content of the type-strain genome is 69.0%, its approximate size 7.48 Mbp, its GenBank deposit SAMD00041819.
The description is as before (Kim et al., 2002) with the following addition. The G+C content of the type-strain genome is 65.7%, its approximate size 9.94 Mbp, its GenBank deposit SAMD00018770.
The description is as before (Everest et al., 2011) with the following addition. The G+C content of the type-strain genome is 68.5%, its approximate size 7.67 Mbp, its GenBank deposit SAMN02645234.
The description is as before (Kageyama et al., 2004b) with the following modification. The G+C content of the type-strain genome is 69.1%, its approximate size 6.33 Mbp, its GenBank deposit SAMD00040665.
The description is as before (Maldonado et al., 2000) with the following addition. The G+C content of the type-strain genome is 67.0%, its approximate size 7.55 Mbp, its GenBank deposit SAMD00018782.
The description is as before (Pijper and Pullinger, 1927) with the following addition. The G+C content of the type-strain genome is 69.2%, its approximate size 8.38 Mbp, its GenBank deposit SAMD00041825.
The description is as before (Isik et al., 1999) with the following addition. The G+C content of the type-strain genome is 66.0%, its approximate size 8.77 Mbp, its GenBank deposit SAMD00040667.
The description is as before (Demaree and Smith, 1952) with the following addition. The G+C content of the type-strain genome is 66.7%, its approximate size 9.22 Mbp, its GenBank deposit SAMD00040668.
The description is as before (Gürtler et al., 2001) with the following addition. The G+C content of the type-strain genome is 68.2%, its approximate size 6.79 Mbp, its GenBank deposit SAMD00041771.
The description is as before (Kageyama et al., 2004c) with the following modification. The G+C content of the type-strain genome is 68.1%, its approximate size 9.10 Mbp, its GenBank deposit SAMD00018773.
The description is as before (Park et al., 2008) with the following modification. The G+C content of the type-strain genome is 72.3%, its approximate size 4.38 Mbp, its GenBank deposit SAMN04606834.
The description is as before (Li et al., 2007) with the following modification. The G+C content of the type-strain genome is 71.8%, its approximate size 4.60 Mbp, its GenBank deposit SAMN04489844.
The description is as before (Dastager et al., 2009) with the following modification. The G+C content of the type-strain genome is 72.0%, its approximate size 4.85 Mbp, its GenBank deposit SAMN02440651.
The description is as before (Liu et al., 2013) with the following modification. The G+C content of the type-strain genome is 70.6%, its approximate size 4.52 Mbp, its GenBank deposit SAMN05216561.
The description is as before (O'Donnell et al., 1982) with the following modification. The G+C content of the type-strain genome is 72.9%, its approximate size 5.63 Mbp, its GenBank deposit SAMN05421671.
The description is as before (Liu et al., 2013) with the following modification. The G+C content of the type-strain genome is 71.0%, its approximate size 4.22 Mbp, its GenBank deposit SAMN05192576.
The description is as before (Grund and Kroppenstedt, 1990) with the following addition. The G+C content of the type-strain genome is 69.8%, its approximate size 5.82 Mbp, its GenBank deposit SAMN02472146.
The description is as before (Hozzein et al., 2004) with the following modification. The G+C content of the type-strain genome is 67.5%, its approximate size 5.21 Mbp, its GenBank deposit SAMN02472156.
The description is as before (Grund and Kroppenstedt, 1990) with the following addition. The G+C content of the type-strain genome is 72.0%, its approximate size 7.18 Mbp, its GenBank deposit SAMD00046506.
The description is as before (Al-Tai and Ruan, 1994) with the following additions. Urea hydrolysis is variable, The predominant menaquinone is MK-10; the major phospholipids are PC and DPG, although PME, PI and PG may occur. The G+C content of the type-strain genome is 73.7%, its approximate size 6.32 Mbp, its GenBank deposit SAMN02472157.
The description is as before (Li et al., 2006) with the following modification. The G+C content of the type-strain genome is 73.8%, its approximate size 6.91 Mbp, its GenBank deposit SAMN02472161.
The description is as before (Meyer, 1976) with the following addition. The G+C content of the type-strain genome is 72.7%, its approximate size 6.54 Mbp, its GenBank deposit SAMN02598425.
The description is as before (Fang et al., 2011) with the following modification. The G+C content of the type-strain genome is 74.1%, its approximate size 7.17 Mbp, its GenBank deposit SAMN05421803.
The description is as before (Li et al., 2006) with the following modification. The G+C content of the type-strain genome is 69.7%, its approximate size 6.04 Mbp, its GenBank deposit SAMN02472152.
The description is as before (Al-Zarban et al., 2002a) with the following modification. The G+C content of the type-strain genome is 71.4%, its approximate size 6.26 Mbp, its GenBank deposit SAMN02472154.
The description is as before (Chun et al., 2000) with the following modification. The G+C content of the type-strain genome is 69.6%, its approximate size 5.31 Mbp, its GenBank deposit SAMN02472160.
The description is as before (Grund and Kroppenstedt, 1990) with the following addition. The G+C content of the type-strain genome is 68.9%, its approximate size 5.69 Mbp, its GenBank deposit SAMD00046482.
The description is as before (Yassin et al., 2009) with the following addition. The G+C content of the type-strain genome is 74.8%, its approximate size 6.91 Mbp, its GenBank deposit SAMN02472155.
The description is as before (Yassin et al., 1997) with the following addition. The G+C content of the type-strain genome is 70.8%, its approximate size 6.00 Mbp, its GenBank deposit SAMN02472147.
The description is as before (Li et al., 2006) with the following modification. The G+C content of the type-strain genome is 70.4%, its approximate size 5.81 Mbp, its GenBank deposit SAMN02472151.
The description is as before (Yassin et al., 1997) with the following modification. The G+C content of the type-strain genome is 72.3%, its approximate size 7.36 Mbp, its GenBank deposit SAMN02472153.
The description is as before (Evtushenko et al., 2000b) with the following addition. The G+C content of the type-strain genome is 75.2%, its approximate size 6.38 Mbp, its GenBank deposit SAMD00046510.
The description is as before (Li et al., 2003b) with the following modification. The G+C content of the type-strain genome is 69.7%, its approximate size 5.33 Mbp, its GenBank deposit SAMN02472150.
The description is as before (le Roes and Meyers, 2008) with the following addition. The G+C content of the type-strain genome is 72.1%, its approximate size 9.63 Mbp, its GenBank deposit SAMN02645361.
The description is as before (Li et al., 2012) with the following modification. The G+C content of the type-strain genome is 70.6%, its approximate size 11.81 Mbp, its GenBank deposit SAMN05421869.
The description is as before (Kämpfer et al., 2005) with the following addition. The G+C content of the type-strain genome is 70.4%, its approximate size 12.10 Mbp, its GenBank deposit SAMN02645367.
The description is as before (Zhang et al., 1998) with the following addition. The G+C content of the type-strain genome is 72.4%, its approximate size 9.20 Mbp, its GenBank deposit SAMN05660976.
The description is as before (Wang et al., 2013b) with the following modification. The G+C content of the type-strain genome is 70.4%, its approximate size 13.24 Mbp, its GenBank deposit SAMN05444920.
The description is as before (Stackebrandt et al., 2002) with the following restriction. The G+C content of the type-strain genome is 72.2%, its approximate size 4.03 Mbp, its GenBank deposit SAMN02645273.
The description is as before (Lechevalier, 1972) with the following addition. The G+C content of the type-strain genome is 74.3%, its approximate size 4.28 Mbp, its GenBank deposit SAMEA2272518.
The description is as before (Kraatz et al., 2011) with the following addition. The G+C content of the type-strain genome is 64.9%, its approximate size 2.35 Mbp, its GenBank deposit SAMN04489857.
The description is as before (Liu et al., 2008) with the following modification. The G+C content of the type-strain genome is 72.9%, its approximate size 3.85 Mbp, its GenBank deposit SAMN02441240.
The description is as before (Busse, 2016) with the following modification. The G+C content of the type-strain genome is 63.0%, its approximate size 4.32 Mbp, its GenBank deposit SAMN02469439.
The description is as before (Jian and Dong, 2002) with the following restriction. The G+C content of the type-strain genome is 58.3%, its approximate size 1.89 Mbp, its GenBank deposit SAMN02299441.
The description is as before Pichel (Reddy and Garcia-Pichel, 2009) with the following modification. The G+C content of the type-strain genome is 74.4%, its approximate size 4.47 Mbp, its GenBank deposit SAMN02440564.
The description is as before (Takahashi et al., 2006) with the following modification. The G+C content of the type-strain genome is 74.1%, its approximate size 5.51 Mbp, its GenBank deposit SAMN02584907.
The description is as before (Mertz, 1994) with the following addition. The G+C content of the type-strain genome is 72.7%, its approximate size 8.13 Mbp, its GenBank deposit SAMD00048613.
The description is as before (Li et al., 2003c) with the following modification. The G+C content of the type-strain genome is 70.1%, its approximate size 5.62 Mbp, its GenBank deposit SAMN05660435.
The description is as before (Li et al., 2003c) with the following modification. The G+C content of the type-strain genome is 69.8%, its approximate size 5.05 Mbp, its GenBank deposit SAMN05660332.
The description is as before (Kim and Goodfellow, 1999) with the following restriction. The G+C content of the type-strain genome is 70.0%, its approximate size 5.28 Mbp, its GenBank deposit SAMN02946344.
The description is as before (Thawai and Kudo, 2012) with the following modification. The G+C content of the type-strain genome is 73.1%, its approximate size 5.38 Mbp, its GenBank deposit SAMN04244578.
The description is as before (Martin et al., 2010) with the following addition. The G+C content of the type-strain genome is 70.8%, its approximate size 6.60 Mbp, its GenBank deposit SAMN04324259.
The description is as before (van Niel, 1928) with the following addition. The G+C content of the type-strain genome is 67.3%, its approximate size 2.65 Mbp, its GenBank deposit SAMN03263638.
The description is as before (Johnson and Cummins, 1972) with the following addition. The G+C content of the type-strain genome is 56.1%, its approximate size 2.04 Mbp, its GenBank deposit SAMN02441171.
The description is as before (Warwick et al., 1994) with the following restriction. The G+C content of the type-strain genome is 74.2%, its approximate size 5.99 Mbp, its GenBank deposit SAMN04490211.
The description is as before (Liu et al., 2006) with the following modification. The G+C content of the type-strain genome is 73.5%, its approximate size 7.36 Mbp, its GenBank deposit SAMN05216207.
The description is as before (Reichert et al., 1998) with the following addition. The G+C content of the type-strain genome is 71.8%, its approximate size 5.05 Mbp, its GenBank deposit SAMN02441500.
The description is as before (Warwick et al., 1994) with the following modification. The G+C content of the type-strain genome is 73.0%, its approximate size 7.35 Mbp, its GenBank deposit SAMN05722966.
The description is as before (Gu et al., 2006) with the following modification. The G+C content of the type-strain genome is 73.0%, its approximate size 6.11 Mbp, its GenBank deposit SAMN05216377.
The description is as before (Henssen, 1957) with the following addition. The G+C content of the type-strain genome is 72.9%, its approximate size 6.10 Mbp, its GenBank deposit SAMN05443637.
The description is as before (Killer et al., 2014) with the following modification. The G+C content of the type-strain genome is 60.5%, its approximate size 2.25 Mbp, its IMG deposit 2731639185.
The description is as before (Maszenan et al., 2005) with the following modification. The G+C content of the type-strain genome is 72.6%, its approximate size 4.88 Mbp, its IMG deposit 2724679791.
The description is as before (Sasaki et al., 1998) with the following modification. The G+C content of the type-strain genome is 61.5%, its approximate size 2.29 Mbp, its GenBank deposit SAMN02440627.
The description is as before (Sanders and Fryer, 1980) with the following modification. The G+C content of the type-strain genome is 56.3%, its approximate size 3.16 Mbp, its GenBank deposit SAMN02603684.
The description is as before (Rowbotham and Cross, 1977) with the following restriction. The G+C content of the type-strain genome is 66.9%, its approximate size 4.55 Mbp, its GenBank deposit SAMD00046763.
The description is as before (Yassin and Schaal, 2005) with the following addition. The G+C content of the type-strain genome is 70.2%, its approximate size 3.90 Mbp, its GenBank deposit SAMN04357305.
The description is as before (Kämpfer et al., 2014a) with the following addition. The G+C content of the type-strain genome is 68.7%, its approximate size 5.13 Mbp, its GenBank deposit SAMN02910065.
The description is as before (Dastager et al., 2014a) with the following modification. The G+C content of the type-strain genome is 62.3%, its approximate size 7.46 Mbp, its GenBank deposit SAMN04263893.
The description is as before (Goodfellow and Alderson, 1977) with the following modification. The G+C content of the type-strain genome is 68.8%, its approximate size 5.20 Mbp, its GenBank deposit SAMN02471952.
The description is as before (Goodfellow and Alderson, 1977) with the following modification. The G+C content of the type-strain genome is 62.4%, its approximate size 6.59 Mbp, its GenBank deposit SAMD00046519.
The description is as before (Goodfellow, 1984) with the following restriction. The G+C content of the type-strain genome is 64.4%, its approximate size 5.77 Mbp, its GenBank deposit SAMN02570003.
The description is as before (Goodfellow et al., 1982) with the following restriction. The G+C content of the type-strain genome is 61.7%, its approximate size 6.74 Mbp, its GenBank deposit SAMD00047211.
The description is as before (Jones et al., 2004) with the following addition. The G+C content of the type-strain genome is 67.9%, its approximate size 4.82 Mbp, its GenBank deposit SAMN04357310.
The description is as before (Kämpfer et al., 2014a) with the following addition. The G+C content of the type-strain genome is 68.8%, its approximate size 4.97 Mbp, its GenBank deposit SAMN04357304.
The description is as before (Ghosh et al., 2006) with the following modification. The G+C content of the type-strain genome is 67.2%, its approximate size 8.23 Mbp, its GenBank deposit SAMN02470011.
The description is as before (Yoon et al., 2000a) with the following modification. The G+C content of the type-strain genome is 67.4%, its approximate size 10.31 Mbp, its GenBank deposit SAMN04490239.
The description is as before (Zhang et al., 2002) with the following modification. The G+C content of the type-strain genome is 69.2%, its approximate size 5.67 Mbp, its GenBank deposit SAMN05444583.
The description is as before (Rehfuss and Urban, 2005) with the following modification. The G+C content of the type-strain genome is 68.4%, its approximate size 6.28 Mbp, its GenBank deposit SAMN04357313.
The description is as before (Yoon et al., 2000b) with the following modification. The G+C content of the type-strain genome is 67.8%, its approximate size 5.26 Mbp, its GenBank deposit SAMN04490240.
The description is as before (Xu et al., 2007) with the following modification. Vogues-Proskauer and reduction of nitrate tests are variable. The G+C content of the type-strain genome is 62.4%, its approximate size 7.26 Mbp, its GenBank deposit SAMN04357315.
The description is as before (Goodfellow and Alderson, 1977) with the following modification. The G+C content of the type-strain genome is 69.7%, its approximate size 4.46 Mbp, its GenBank deposit SAMD00046764.
The description is as before (Rainey et al., 1995a) with the following restriction. The G+C content of the type-strain genome is 68.2%, its approximate size 5.20 Mbp, its GenBank deposit SAMD00034188.
The description is as before (Goodfellow and Alderson, 1977) with the following addition. The G+C content of the type-strain genome is 70.7%, its approximate size 5.30 Mbp, its GenBank deposit SAMN04357318.
The description is as before (Yassin, 2005) with the following addition. The G+C content of the type-strain genome is 68.7%, its approximate size 4.73 Mbp, its GenBank deposit SAMN05444695.
The description is as before (Matsuyama et al., 2003) with the following modification. The G+C content of the type-strain genome is 69.8%, its approximate size 5.49 Mbp, its GenBank deposit SAMN05444580.
The description is as before (Goodfellow et al., 2002) with the following restriction. The G+C content of the type-strain genome is 66.8%, its approximate size 10.40 Mbp, its GenBank deposit SAMD00041784.
The description is as before (Stoecker et al., 1994) with the following modification. The G+C content of the type-strain genome is 68.2%, its approximate size 6.30 Mbp, its GenBank deposit SAMD00046765.
The description is as before (Hanada et al., 2002b) with the following modification. The G+C content of the type-strain genome is 60.7%, its approximate size 5.72 Mbp, its GenBank deposit SAMN02598306.
The description is as before (Daneshvar et al., 2004) with the following addition. The G+C content of the type-strain genome is 53.7%, its approximate size 2.51 Mbp, its GenBank deposit SAMN00120582.
The description is as before (Collins et al., 2000b) with the following restriction. The G+C content of the type-strain genome is 59.5%, its approximate size 2.26 Mbp, its GenBank deposit SAMN00001919.
The description is as before (Kämpfer et al., 2014b) with the following addition. The G+C content of the type-strain genome is 65.1%, its approximate size 3.19 Mbp, its GenBank deposit SAMN03729479.
The description is as before (Suzuki et al., 1988) with the following modification. The G+C content of the type-strain genome is 66.5%, its approximate size 3.40 Mbp, its GenBank deposit SAMN00767673.
The description is as before (Carreto et al., 1996) with the following modification. The G+C content of the type-strain genome is 70.5%, its approximate size 3.23 Mbp, its GenBank deposit SAMN02598258.
The description is as before (Runmao, 1987) with the following addition. The G+C content of the type-strain genome is 70.1%, its approximate size 4.75 Mbp, its GenBank deposit SAMN02261398.
The description is as before (Runmao et al., 1988) with the following addition. The G+C content of the type-strain genome is 69.7%, its approximate size 5.41 Mbp, its GenBank deposit SAMN02256534.
The description is as before (Greiner-Mai et al., 1988) with the following addition. The G+C content of the type-strain genome is 69.1%, its approximate size 4.56 Mbp, its GenBank deposit SAMN02441787.
The description is as before (Al-Zarban et al., 2002b) with the following addition. The G+C content of the type-strain genome is 70.9%, its approximate size 3.63 Mbp, its GenBank deposit SAMN02256397.
The description is as before (Greiner-Mai et al., 1987) with the following addition. The G+C content of the type-strain genome is 67.4%, its approximate size 4.31 Mbp, its GenBank deposit SAMN02982918.
The description is as before (Nonomura and Ohara, 1971a) with the following addition. The G+C content of the type-strain genome is 67.3%, its approximate size 4.31 Mbp, its GenBank deposit SAMN02598440.
The description is as before (Jin et al., 1998) with the following addition. The G+C content of the type-strain genome is 68.9%, its approximate size 4.77 Mbp, its GenBank deposit SAMN02261402.
The description is as before (Yuan et al., 2008) with the following modification. The G+C content of the type-strain genome is 70.4%, its approximate size 8.27 Mbp, its GenBank deposit SAMN04490245.
The description is as before (Labeda, 1987) with the following modification. The G+C content of the type-strain genome is 71.1%, its approximate size 8.21 Mbp, its GenBank deposit SAMEA2272514.
The description is as before (Lu et al., 2001) with the following modification. The G+C content of the type-strain genome is 70.8%, its approximate size 6.29 Mbp, its GenBank deposit SAMN05660874.
The description is as before (Korn-Wendisch et al., 1989) with the following modification. The G+C content of the type-strain genome is 68.9%, its approximate size 3.98 Mbp, its GenBank deposit SAMN02472042.
The description is as before (Mertz and Yao, 1990) with the following modification. The G+C content of the type-strain genome is 67.9%, its approximate size 9.02 Mbp, its GenBank deposit SAMN05216473.
The description is as before (Grund and Kroppenstedt, 1990) with the following addition. The G+C content of the type-strain genome is 73.5%, its approximate size 10.88 Mbp, its GenBank deposit SAMN02645315.
The description is as before (Ahmed et al., 2013) with the following restriction. The G+C content of the type-strain genome is 69.7%, its approximate size 5.87 Mbp, its GenBank deposit SAMN02597245.
The description is as before (Maldonado et al., 2005) with the following addition. The G+C content of the type-strain genome is 69.5%, its approximate size 5.18 Mbp, its GenBank deposit SAMN02598367.
The description is as before (Pikuta et al., 2017) with the following modification. The G+C content of the type-strain genome is 71.9%, its approximate size 4.25 Mbp, its GenBank deposit SAMN02598426.
The description is as before (Downes et al., 2011) with the following modification. The G+C content of the type-strain genome is 48.6%, its approximate size 1.80 Mbp, its GenBank deposit SAMD00061049.
The description is as before (Tian et al., 2009) with the following modification. The G+C content of the type-strain genome is 67.8%, its approximate size 8.54 Mbp, its GenBank deposit SAMN02256422.
The description is as before (Butler et al., 2005) with the following modification. The G+C content of the type-strain genome is 66.8%, its approximate size 3.16 Mbp, its GenBank deposit SAMN02598516.
The description is as before (Butler et al., 2005) with the following modification. The G+C content of the type-strain genome is 68.2%, its approximate size 3.59 Mbp, its GenBank deposit SAMN02463810.
The description is as before (Lagier et al., 2013) with the following modification. The G+C content of the type-strain genome is 62.1%, its GenBank deposit SAMEA2272806.
The description is as before (Zhou et al., 2009) with the following modification. The G+C content of the type-strain genome is 71.4%, its approximate size 4.49 Mbp, its GenBank deposit SAMN04508113.
The description is as before (Prabhu et al., 2015) with the following modification. The G+C content of the type-strain genome is 71.1%, its approximate size 4.01 Mbp, its GenBank deposit SAMN06212359.
The description is as before (Chun et al., 1997a) with the following modification. The G+C content of the type-strain genome is 68.6%, its approximate size 4.21 Mbp, its GenBank deposit SAMD00046514.
The description is as before (Wade et al., 1999) with the following restriction. The G+C content of the type-strain genome is 62.2%, its approximate size 2.09 Mbp, its GenBank deposit SAMN00008861.
The description is as before (Wade et al., 1999) with the following modification. The G+C content of the type-strain genome is 60.2%, its approximate size 3.17 Mbp, its GenBank deposit SAMN02598438.
The description is as before (Mingma et al., 2016) with the following modification. The G+C content of the type-strain genome is 70.0%, its approximate size 7.34 Mbp, its GenBank deposit SAMD00046509.
The description is as before (Houghton et al., 2015) with the following modification. The G+C content of the type-strain genome is 68.1%, its approximate size 3.99 Mbp, its GenBank deposit SAMN02598446.
The description is as before (Couch, 1963) with the following addition. The G+C content of the type-strain genome is 72.6%, its approximate size 6.03 Mbp, its GenBank deposit SAMN02645261.
The description is as before (Lechevalier et al., 1968) with the following addition. The G+C content of the type-strain genome is 71.4%, its approximate size 5.50 Mbp, its GenBank deposit SAMN02261321.
The description is as before (Labeda and Kroppenstedt, 2005) with the following modification. The G+C content of the type-strain genome is 68.1%, its approximate size 6.84 Mbp, its GenBank deposit SAMN02598427.
The description is as before (Kim et al., 2003) with the following addition. The G+C content of the type-strain genome is 71.8%, its approximate size 9.92 Mbp, its GenBank deposit SAMN02745413.
The description is as before (Cho et al., 2008) with the following addition. The G+C content of the type-strain genome is 72.2%, its approximate size 9.32 Mbp, its GenBank deposit SAMD00022877.
The description is as before (Kim et al., 2003) with the following addition. The G+C content of the type-strain genome is 71.5%, its approximate size 8.41 Mbp, its GenBank deposit SAMD00022873.
The description is as before (Cho et al., 2008) with the following addition. The G+C content of the type-strain genome is 72.0%, its approximate size 8.72 Mbp, its GenBank deposit SAMD00022876.
The description is as before (Kim et al., 2003) with the following addition. The G+C content of the type-strain genome is 72.2%, its approximate size 8.33 Mbp, its GenBank deposit SAMD00022875.
The description is as before (Wang et al., 2006) with the following addition. The G+C content of the type-strain genome is 73.4%, its approximate size 7.81 Mbp, its GenBank deposit SAMN02745412.
The description is as before (Cho et al., 2008) with the following addition. The G+C content of the type-strain genome is 71.8%, its approximate size 9.00 Mbp, its GenBank deposit SAMN02745414.
The description is as before (Tomita et al., 1987) with the following addition. The G+C content of the type-strain genome is 72.6%, its approximate size 7.28 Mbp, its GenBank deposit SAMN05444320.
The description is as before (Li et al., 2003d) with the following modification. The G+C content of the type-strain genome is 71.9%, its approximate size 5.91 Mbp, its GenBank deposit SAMN03074668.
The description is as before (Umezawa et al., 1953) with the following addition. The G+C content of the type-strain genome is 72.5%, its approximate size 8.11 Mbp, its GenBank deposit SAMN02645217.
The description is as before (Xia et al., 2013) with the following modification. The G+C content of the type-strain genome is 73.2%, its approximate size 6.47 Mbp, its GenBank deposit SAMN05421773.
The description is as before (Rong et al., 2009) with the following restriction. The G+C content of the type-strain genome is 73.4%, its approximate size 7.08 Mbp, its GenBank deposit SAMN02645503.
The description is as before (Waksman and Henrici, 1948) with the following addition. The G+C content of the type-strain genome is 72.2%, its approximate size 9.72 Mbp, its GenBank deposit SAMN02645294.
The description is as before (Labeda et al., 2014) with the following addition. The G+C content of the type-strain genome is 72.7%, its approximate size 7.59 Mbp, its GenBank deposit SAMD00023485.
The description is as before (Liu et al., 2009b) with the following modification. The G+C content of the type-strain genome is 72.1%, its approximate size 8.27 Mbp, its GenBank deposit SAMN05216251.
The description is as before (Pinnert-Sindico, 1954) with the following addition. The G+C content of the type-strain genome is 72.2%, its approximate size 8.39 Mbp, its GenBank deposit SAMN03921868.
The description is as before (Sharma et al., 2014) with the following restriction. The G+C content of the type-strain genome is 72.6%, its approximate size 7.82 Mbp, its GenBank deposit SAMN04386232.
The description is as before (Waksman and Henrici, 1948) with the following addition. The G+C content of the type-strain genome is 70.8%, its approximate size 10.12 Mbp, its GenBank deposit SAMN04193346.
The description is as before (Pridham et al., 1958) with the following addition. The G+C content of the type-strain genome is 70.7%, its approximate size 8.22 Mbp, its GenBank deposit SAMN02645281.
The description is as before (Pridham, 1970) with the following addition. The G+C content of the type-strain genome is 71.2%, its approximate size 9.32 Mbp, its GenBank deposit SAMN02645370.
The description is as before (Kelly et al., 1959) with the following addition. The G+C content of the type-strain genome is 70.9%, its approximate size 8.79 Mbp, its GenBank deposit SAMD00031377.
The description is as before (Witt and Stackebrandt, 1990) with the following addition. The G+C content of the type-strain genome is 72.4%, its approximate size 7.21 Mbp, its GenBank deposit SAMN02645263.
The description is as before (Johstone and Waksman, 1947) with the following addition. The G+C content of the type-strain genome is 73.1%, its approximate size 7.39 Mbp, its GenBank deposit SAMN02645190.
The description is as before (Waksman, 1961) with the following addition. The G+C content of the type-strain genome is 71.2%, its approximate size 8.91 Mbp, its GenBank deposit SAMN02471210.
The description is as before (Eguchi et al., 1993) with the following modification. The G+C content of the type-strain genome is 72.7%, its approximate size 8.33 Mbp, its GenBank deposit SAMN04193354.
The description is as before (Lanoot et al., 2002) with the following modification. The G+C content of the type-strain genome is 72.5%, its approximate size 6.03 Mbp, its GenBank deposit SAMN02645219.
The description is as before (Waksman and Henrici, 1948) with the following addition. The G+C content of the type-strain genome is 72.7%, its approximate size 7.79 Mbp, its GenBank deposit SAMN02645354.
The description is as before (Heinemann et al., 1953) with the following addition. The G+C content of the type-strain genome is 70.2%, its approximate size 11.50 Mbp, its GenBank deposit SAMN04193350.
The description is as before (Waksman, 1961) with the following addition. The G+C content of the type-strain genome is 73.0%, its approximate size 7.05 Mbp, its GenBank deposit SAMN02645222.
The description is as before (Hamada, 1958) with the following addition. The G+C content of the type-strain genome is 71.0%, its approximate size 9.68 Mbp, its GenBank deposit SAMN04193341.
The description is as before (Nishimura et al., 1953) with the following addition. The G+C content of the type-strain genome is 71.7%, its approximate size 8.68 Mbp, its GenBank deposit SAMN02645237.
The description is as before (Pridham et al., 1958) with the following addition. The G+C content of the type-strain genome is 70.7%, its approximate size 9.35 Mbp, its GenBank deposit SAMN02645291.
The description is as before (Gause et al., 1983) with the following addition. The G+C content of the type-strain genome is 70.3%, its approximate size 9.79 Mbp, its GenBank deposit SAMN04193344.
The description is as before (Pridham, 1970) with the following addition. The G+C content of the type-strain genome is 71.3%, its approximate size 8.40 Mbp, its GenBank deposit SAMN02645293.
The description is as before (Higgens and Kastner, 1971) with the following addition. The G+C content of the type-strain genome is 72.5%, its approximate size 8.53 Mbp, its GenBank deposit SAMN02471788.
The description is as before (Lanoot et al., 2005) with the following addition. The G+C content of the type-strain genome is 72.0%, its approximate size 10.27 Mbp, its GenBank deposit SAMN04193345.
The description is as before (Trejo and Bennett, 1963) with the following addition. The G+C content of the type-strain genome is 71.0%, its approximate size 8.39 Mbp, its GenBank deposit SAMN04193339.
The description is as before (Pridham et al., 1958) with the following addition. The G+C content of the type-strain genome is 71.6%, its approximate size 7.90 Mbp, its GenBank deposit SAMN02645238.
The description is as before (Gordon and Lapa, 1966) with the following addition. The G+C content of the type-strain genome is 71.7%, its approximate size 9.08 Mbp, its GenBank deposit SAMN02645292.
The description is as before (Sun et al., 2007) with the following modification. The G+C content of the type-strain genome is 72.9%, its approximate size 7.12 Mbp, its GenBank deposit SAMN05216505.
The description is as before (Waksman and Henrici, 1948) with the following addition. The G+C content of the type-strain genome is 72.0%, its approximate size 7.53 Mbp, its GenBank deposit SAMN02645344.
The description is as before (Gause et al., 1983) with the following addition. The G+C content of the type-strain genome is 71.2%, its approximate size 8.76 Mbp, its GenBank deposit SAMN02645208.
The description is as before (Bartz et al., 1951) with the following addition. The G+C content of the type-strain genome is 72.7%, its approximate size 7.90 Mbp, its GenBank deposit SAMN02645357.
The description is as before (Pridham, 1970) with the following addition. The G+C content of the type-strain genome is 71.3%, its approximate size 8.40 Mbp, its GenBank deposit SAMN02645251.
The description is as before (Wallhausser et al., 1965) with the following addition. The G+C content of the type-strain genome is 72.2%, its approximate size 8.22 Mbp, its GenBank deposit SAMN02595233.
The description is as before (Sakamoto et al., 1962) with the following addition. The G+C content of the type-strain genome is 71.7%, its approximate size 8.95 Mbp, its GenBank deposit SAMN02645272.
The description is as before (Waksman and Henrici, 1948) with the following addition. The G+C content of the type-strain genome is 72.0%, its approximate size 7.54 Mbp, its GenBank deposit SAMN02645254.
The description is as before (Yamaguchi and Saburi, 1955) with the following addition. The G+C content of the type-strain genome is 71.7%, its approximate size 10.11 Mbp, its GenBank deposit SAMN04193353.
The description is as before (Pridham et al., 1958) with the following addition. The G+C content of the type-strain genome is 70.9%, its approximate size 9.83 Mbp, its GenBank deposit SAMN04193351.
The description is as before (Liu et al., 2005) with the following modification. The G+C content of the type-strain genome is 72.2%, its approximate size 8.63 Mbp, its GenBank deposit SAMN04490359.
The description is as before (Waksman and Henrici, 1948) with the following addition. The G+C content of the type-strain genome is 71.9%, its approximate size 7.74 Mbp, its GenBank deposit SAMN02645380.
The description is as before (Luo et al., 2011) with the following restriction. The G+C content of the type-strain genome is 71.4%, its approximate size 8.23 Mbp, its GenBank deposit SAMN05421806.
The description is as before (Sarmin et al., 2013) with the following modification. The G+C content of the type-strain genome is 71.6%, its approximate size 8.22 Mbp, its GenBank deposit SAMN03254380.
The description is as before (Trejo et al., 1977) with the following addition. The G+C content of the type-strain genome is 72.3%, its approximate size 8.03 Mbp, its GenBank deposit SAMD00050872.
The description is as before (Waksman and Henrici, 1948) with the following addition. The G+C content of the type-strain genome is 72.7%, its approximate size 8.47 Mbp, its GenBank deposit SAMN02645248.
The description is as before (Mason et al., 1963) with the following addition. The G+C content of the type-strain genome is 71.0%, its approximate size 10.32 Mbp, its GenBank deposit SAMN05335414.
The description is as before (Prosser and Palleroni, 1976) with the following addition. The G+C content of the type-strain genome is 73.1%, its approximate size 8.34 Mbp, its GenBank deposit SAMN04193348.
The description is as before (Luo et al., 2017) with the following modification. The G+C content of the type-strain genome is 71.6%, its approximate size 7.83 Mbp, its GenBank deposit SAMN02803977.
The description is as before (De Boer et al., 1956) with the following addition. The G+C content of the type-strain genome is 72.1%, its approximate size 8.18 Mbp, its GenBank deposit SAMN02645328.
The description is as before (Gause et al., 1983) with the following addition. The G+C content of the type-strain genome is 72.3%, its approximate size 5.85 Mbp, its GenBank deposit SAMN02645209.
The description is as before (Arcamone et al., 1959) with the following addition. The G+C content of the type-strain genome is 71.0%, its approximate size 10.77 Mbp, its GenBank deposit SAMN04490356.
The description is as before (Cercos et al., 1962) with the following addition. The G+C content of the type-strain genome is 72.4%, its approximate size 8.47 Mbp, its GenBank deposit SAMN04490357.
The description is as before (Witt and Stackebrandt, 1990) with the following addition. The G+C content of the type-strain genome is 73.3%, its approximate size 7.48 Mbp, its GenBank deposit SAMN02470112.
The description is as before (Gause et al., 1983) with the following addition. The G+C content of the type-strain genome is 72.1%, its approximate size 7.80 Mbp, its GenBank deposit SAMN02645312.
The description is as before (Goodfellow et al., 1986b) with the following addition. The G+C content of the type-strain genome is 71.7%, its approximate size 8.49 Mbp, its GenBank deposit SAMN02645268.
The description is as before (Waksman, 1961) with the following addition. The G+C content of the type-strain genome is 70.9%, its approximate size 7.71 Mbp, its GenBank deposit SAMN03020625.
The description is as before (Brown et al., 1953) with the following addition. The G+C content of the type-strain genome is 71.5%, its approximate size 9.82 Mbp, its GenBank deposit SAMN03287657.
The description is as before (Pridham, 1970) with the following addition. The G+C content of the type-strain genome is 71.4%, its approximate size 8.41 Mbp, its GenBank deposit SAMN02645378.
The description is as before (Waksman and Henrici, 1948) with the following addition. The G+C content of the type-strain genome is 72.4%, its approximate size 8.58 Mbp, its GenBank deposit SAMN02645259.
The description is as before (Waksman and Henrici, 1948) with the following addition. The G+C content of the type-strain genome is 70.2%, its approximate size 11.61 Mbp, its GenBank deposit SAMN04193347.
The description is as before (Xu et al., 2006) with the following modification. The G+C content of the type-strain genome is 72.3%, its approximate size 8.16 Mbp, its GenBank deposit SAMN05216499.
The description is as before (Lanoot et al., 2004) with the following modification. The G+C content of the type-strain genome is 71.0%, its approximate size 9.54 Mbp, its GenBank deposit SAMN04193340.
The description is as before (Madhaiyan et al., 2016) with the following modification. The G+C content of the type-strain genome is 73.0%, its approximate size 6.34 Mbp, its GenBank deposit SAMN05192584.
The description is as before (Tresner and Backus, 1956) with the following addition. The G+C content of the type-strain genome is 71.2%, its approximate size 8.40 Mbp, its GenBank deposit SAMN05722965.
The description is as before (Lee et al., 2014b) with the following restriction. The G+C content of the type-strain genome is 70.0%, its approximate size 7.48 Mbp, its GenBank deposit SAMN03070117.
The description is as before (Pridham et al., 1958) with the following addition. The G+C content of the type-strain genome is 69.7%, its approximate size 11.76 Mbp, its GenBank deposit SAMD00041815.
The description is as before (Pridham, 1970) with the following addition. The G+C content of the type-strain genome is 71.0%, its approximate size 9.58 Mbp, its GenBank deposit SAMN04193342.
The description is as before (Hu et al., 2012) with the following modification. The G+C content of the type-strain genome is 72.7%, its approximate size 7.17 Mbp, its GenBank deposit SAMN05421870.
The description is as before (Lindenbein, 1952) with the following addition. The G+C content of the type-strain genome is 71.0%, its approximate size 9.15 Mbp, its GenBank deposit SAMN04193356.
The description is as before (Waksman and Lechevalier, 1953) with the following addition. The G+C content of the type-strain genome is 71.9%, its approximate size 9.50 Mbp, its GenBank deposit SAMN02471950.
The description is as before (Xu et al., 2006) with the following modification. The G+C content of the type-strain genome is 72.9%, its approximate size 9.01 Mbp, its GenBank deposit SAMN05216267.
The description is as before (Pridham, 1970) with the following addition. The G+C content of the type-strain genome is 71.9%, its approximate size 8.23 Mbp, its GenBank deposit SAMN02645383.
The description is as before (Chun et al., 1997b) with the following modification. The G+C content of the type-strain genome is 71.9%, its approximate size 5.87 Mbp, its GenBank deposit SAMN02645290.
The description is as before (Waksman and Henrici, 1948) with the following addition. The G+C content of the type-strain genome is 74.1%, its approximate size 5.18 Mbp, its GenBank deposit SAMN02471954.
The description is as before (Kämpfer et al., 2008) with the following addition. The G+C content of the type-strain genome is 72.7%, its approximate size 5.86 Mbp, its GenBank deposit SAMEA3696889.
The description is as before (Wallick et al., 1956) with the following addition. The G+C content of the type-strain genome is 70.7%, its approximate size 8.57 Mbp, its GenBank deposit SAMN02469390.
The description is as before (Gause et al., 1983) with the following addition. The G+C content of the type-strain genome is 72.0%, its approximate size 10.15 Mbp, its GenBank deposit SAMD00040622.
The description is as before (Waksman and Lechevalier, 1953) with the following addition. The G+C content of the type-strain genome is 71.7%, its approximate size 7.10 Mbp, its GenBank deposit SAMN02440478.
The description is as before (Kurylowicz and Wóznicka, 1967) with the following addition. The G+C content of the type-strain genome is 72.4%, its approximate size 8.59 Mbp, its GenBank deposit SAMN02645386.
The description is as before (Ehrlich et al., 1948) with the following addition. The G+C content of the type-strain genome is 72.5%, its approximate size 8.22 Mbp, its GenBank deposit SAMEA3138410.
The description is as before (Zhu et al., 2007) with the following modification. The G+C content of the type-strain genome is 72.0%, its approximate size 9.15 Mbp, its GenBank deposit SAMN02730132.
The description is as before (Gause et al., 1983) with the following addition. The G+C content of the type-strain genome is 72.2%, its approximate size 7.65 Mbp, its GenBank deposit SAMN02645210.
The description is as before (Grundy et al., 1952) with the following addition. The G+C content of the type-strain genome is 72.4%, its approximate size 8.32 Mbp, its GenBank deposit SAMN02645365.
The description is as before (Goodfellow et al., 1986a) with the following addition. The G+C content of the type-strain genome is 72.0%, its approximate size 6.43 Mbp, its GenBank deposit SAMN02256407.
The description is as before (Gause et al., 1983) with the following addition. The G+C content of the type-strain genome is 72.1%, its approximate size 9.38 Mbp, its GenBank deposit SAMN02645189.
The description is as before (Lindenbein, 1952) with the following addition. The G+C content of the type-strain genome is 72.1%, its approximate size 6.29 Mbp, its GenBank deposit SAMN02645271.
The description is as before (Xu et al., 2006) with the following modification. The G+C content of the type-strain genome is 72.6%, its approximate size 9.59 Mbp, its GenBank deposit SAMN05216223.
The description is as before (Kim et al., 2004) with the following addition. The G+C content of the type-strain genome is 73.6%, its approximate size 7.82 Mbp, its GenBank deposit SAMN02745173.
The description is as before (Nakamura, 1961) with the following addition. The G+C content of the type-strain genome is 71.2%, its approximate size 10.01 Mbp, its GenBank deposit SAMN04193343.
The description is as before (Nonomura and Ohara, 1960) with the following addition. The G+C content of the type-strain genome is 69.4%, its approximate size 9.44 Mbp, its GenBank deposit SAMN02645242.
The description is as before (Zhang et al., 2009b) with the following modification. The G+C content of the type-strain genome is 70.7%, its approximate size 9.26 Mbp, its GenBank deposit SAMN05216275.
The description is as before (Couch, 1955) with the following addition. The G+C content of the type-strain genome is 70.9%, its approximate size 10.37 Mbp, its GenBank deposit SAMN00002585.
The description is as before (Maszenan et al., 1999b) with the following modification. The G+C content of the type-strain genome is 66.7%, its approximate size 4.13 Mbp, its GenBank deposit SAMN02745244.
The description is as before (Kumari et al., 2016) with the following modification. The G+C content of the type-strain genome is 68.0%, its approximate size 3.19 Mbp, its GenBank deposit SAMN05428934.
The description is as before (Cai et al., 2011) with the following restriction. The G+C content of the type-strain genome is 69.7%, its approximate size 3.16 Mbp, its GenBank deposit SAMN04488242.
The description is as before (McKenzie et al., 2006) with the following addition. The G+C content of the type-strain genome is 68.2%, its approximate size 3.79 Mbp, its GenBank deposit SAMEA3146295.
The description is as before (Zhou et al., 2012a) with the following modification. The G+C content of the type-strain genome is 71.7%, its approximate size 8.22 Mbp, its GenBank deposit SAMN06041775.
The description is as before (Kukolya et al., 2002) with the following modification. The G+C content of the type-strain genome is 72.1%, its approximate size 4.32 Mbp, its GenBank deposit SAMN03840822.
The description is as before (Zhang et al., 1998) with the following addition. The G+C content of the type-strain genome is 67.5%, its approximate size 3.64 Mbp, its GenBank deposit SAMD00046774.
The description is as before (Yang et al., 2008) with the following modification. The G+C content of the type-strain genome is 71.2%, its approximate size 5.43 Mbp, its GenBank deposit SAMN04009169.
The description is as before (Wang et al., 1996) with the following addition. The G+C content of the type-strain genome is 72.4%, its approximate size 4.19 Mbp, its GenBank deposit SAMN00002587.
The description is as before (Korn-Wendisch et al., 1995) with the following addition. The G+C content of the type-strain genome is 69.9%, its approximate size 4.20 Mbp, its GenBank deposit SAMN02441562.
The description is as before (Korn-Wendisch et al., 1995) with the following addition. The G+C content of the type-strain genome is 68.9%, its approximate size 4.53 Mbp, its GenBank deposit SAMN02440599.
The description is as before (Dodsworth et al., 2014) with the following modification. The G+C content of the type-strain genome is 67.3%, its approximate size 3.22 Mbp, its GenBank deposit SAMN02746019.
The description is as before (King and King, 2014) with the following modification. The G+C content of the type-strain genome is 60.9%, its approximate size 5.61 Mbp, its GenBank deposit SAMN02841144.
The description is as before (Zarilla and Perry, 1984) with the following modification. The G+C content of the type-strain genome is 69.1%, its approximate size 2.21 Mbp, its GenBank deposit SAMN02745716.
The description is as before (Henssen, 1957) with the following addition. The G+C content of the type-strain genome is 71.6%, its approximate size 5.64 Mbp, its GenBank deposit SAMN02598435.
The description is as before (Wu et al., 2018) with the following modification. The G+C content of the type-strain genome is 71.0%, its approximate size 5.99 Mbp, its GenBank deposit SAMN04489764.
The description is as before (Katayama et al., 2010) with the following modification. The G+C content of the type-strain genome is 68.1%, its approximate size 4.71 Mbp, its GenBank deposit SAMD00036785.
The description is as before (La Scola et al., 2001) with the following modification. The G+C content of the type-strain genome is 46.3%, its approximate size 0.93 Mbp, its GenBank deposit SAMN02603248.
The description is as before (Yassin et al., 2011) with the following restriction. The G+C content of the type-strain genome is 65.4%, its approximate size 2.03 Mbp, its GenBank deposit SAMN04196844.
The description is as before (Park et al., 2009) with the following modification. The G+C content of the type-strain genome is 71.0%, its approximate size 4.95 Mbp, its GenBank deposit SAMN04457857.
The description is as before (Collins et al., 1988b) with the following modification. The G+C content of the type-strain genome is 68.4%, its approximate size 4.48 Mbp, its GenBank deposit SAMN00002597.
The description is as before (Olson et al., 2007) with the following modification. The G+C content of the type-strain genome is 71.0%, its approximate size 4.59 Mbp, its GenBank deposit SAMN04457855.
The description is as before (Seong et al., 2003) with the following modification. The G+C content of the type-strain genome is 70.1%, its approximate size 5.21 Mbp, its GenBank deposit SAMN04457858.
The description is as before (Labeda and Kroppenstedt, 2007) with the following modification. The G+C content of the type-strain genome is 71.6%, its approximate size 10.68 Mbp, its IMG deposit 2728369483.
The description is as before (Hall et al., 2003d) with the following modification. The G+C content of the type-strain genome is 53.4%, its approximate size 2.02 Mbp, its GenBank deposit SAMN02440916.
The description is as before (Yassin and Hupfer, 2006) with the following addition. The G+C content of the type-strain genome is 70.2%, its approximate size 4.41 Mbp, its GenBank deposit SAMN05661029.
The description is as before (Stach et al., 2004) with the following addition. The G+C content of the type-strain genome is 68.2%, its approximate size 4.93 Mbp, its GenBank deposit SAMN05660695.
The description is as before (Yassin et al., 2007b) with the following addition. The G+C content of the type-strain genome is 70.4%, its approximate size 4.64 Mbp, its GenBank deposit SAMN05660736.
The description is as before (Fang et al., 2013) with the following modification. The G+C content of the type-strain genome is 67.2%, its approximate size 4.44 Mbp, its GenBank deposit SAMN05445060.
The description is as before (Stackebrandt and Schumann, 2004) with the following modification. The G+C content of the type-strain genome is 71.9%, its approximate size 4.00 Mbp, its GenBank deposit SAMD00047215.
The description is as before (Li et al., 2008b) with the following modification. The G+C content of the type-strain genome is 55.6%, its approximate size 2.78 Mbp, its GenBank deposit SAMN02440577.
The description is as before (Lin et al., 2004) with the following modification. The G+C content of the type-strain genome is 64.7%, its approximate size 2.14 Mbp, its GenBank deposit SAMD00046486.
TW, NK, H-PK, and MaG collected the genome sequences. JM-K and MaG phylogenetically and statistically analyzed the data. MG-L and MaG collected and interpreted the G+C content and genome-size information. MG-L, IN, LC, and MiG collected the phenotypic information. IN, LC, MiG, MaG and RP interpreted the phenotypic information. MiG, MaG, IN, LC, MG-L, RP and JM-K wrote the manuscript. All authors read and approved the final manuscript.
The work conducted by the Joint Genome Institute, a United States Department of Energy Office of Science User Facility, is supported under Contract No. DE-AC02-05CH11231.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We are grateful to Peter Schumann (DSMZ), Brian J. Tindall (DSMZ) and Maria del Carmen Montero Calasanz (Newcastle University) for helpful comments, as well as to Evelyne Brambilla, Iris Brandes, Meike Döppner, Katja Steenblock, and Beatrice Trümper (all DSMZ) for technical assistance.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.02007/full#supplementary-material
Abe, S., Takayama, K.-I., and Kinoshita, S. (1967). Taxonomical studies on glutamic acid-producing bacteria. J. Gen. Appl. Microbiol. 13, 279–301.
Adachi, K., Katsuta, A., Matsuda, S., Peng, X., Misawa, N., Shizuri, Y., et al. (2007). Smaragdicoccus niigatensis gen. nov., sp. nov., a novel member of the suborder Corynebacterineae. Int. J. Syst. Evol. Microbiol. 57, 297–301. doi: 10.1099/ijs.0.64254-0
Adekambi, T., Sassi, M., van Ingen, J., and Drancourt, M. (2017). Reinstating Mycobacterium massiliense and Mycobacterium bolletii as species of the Mycobacterium abscessus complex. Int. J. Syst. Evol. Microbiol. 67, 2726–2730. doi: 10.1099/ijsem.0.002011
Adékambi, T., Stein, A., Carvajal, J., Raoult, D., and Drancourt, M. (2006). Description of Mycobacterium conceptionense sp. nov., a Mycobacterium fortuitum group organism isolated from a posttraumatic osteitis inflammation. J. Clin. Microbiol. 44, 1268–1273. doi: 10.1128/JCM.44.4.1268-1273.2006
Ahmed, L., Jensen, P. R., Freel, K. C., Broen, R., Jones, A. L., Kim, B.-Y., et al. (2013). Salinispora pacifica sp. nov., an actinomycete from marine sediments. Antonie Van Leeuwenhoek 103, 1069–1078. doi: 10.1007/s10482-013-9886-4
Al-Mussallam, A. A., Al-Zarban, S. S., Fasasi, Y. A., Kroppenstedt, R. M., and Stackebrandt, E. (2003). Amycolatopsis keratiniphila sp. nov., a novel keratinolytic soil actinomycete from Kuwait. Int. J. Syst. Evol. Microbiol. 53, 871–874. doi: 10.1099/ijs.0.02515-0
Al-Tai, A. M., and Ruan, J.-S. (1994). Nocardiopsis halophila sp. nov., a new halophilic actinomycete isolated from soil. Int. J. Syst. Bacteriol. 44, 474–478. doi: 10.1099/00207713-44-3-474
Al-Zarban, S. S., Abbas, I., Al-Musallam, A. A., Steiner, U., Stackebrandt, E., and Kroppenstedt, R. M. (2002a). Nocardiopsis halotolerans sp. nov., isolated from salt marsh soil in Kuwait. Int. J. Syst. Evol. Microbiol. 52, 525–529. doi: 10.1099/ijs.0.01658-0
Al-Zarban, S. S., Al-Musallam, A. A., Abbas, I., Stackebrandt, E., and Kroppenstedt, R. M. (2002b). Saccharomonospora halophila sp. nov., a novel halophilic actinomycete isolated from marsh soil in Kuwait. Int. J. Syst. Evol. Microbiol. 52, 555–558. doi: 10.1099/ijs.0.01894-0
An, D., Cai, S., and Dong, X. (2006). Actinomyces ruminicola sp. nov., isolated from cattle rumen. Int. J. Syst. Evol. Microbiol. 56, 2043–2048. doi: 10.1099/ijs.0.64059-0
Ara, I., Bakir, M. A., and Kudo, T. (2008a). Transfer of Catellatospora koreensis Lee et al. 2000 as Catelliglobosispora koreensis gen. nov., comb. nov. and Catellatospora tsunoense Asano et. al. 1989 as Hamadaea tsunoensis gen. nov., comb. nov., and emended description of the genus Catellatospora Asano and Kawamoto 1986 emend. Lee and Hah 2002. Int. J. Syst. Evol. Microbiol. 58, 1950–1960. doi: 10.1099/ijs.0.65548-0
Ara, I., and Kudo, T. (2007). Krasilnikovia gen. nov., a new member of the family Micromonosporaceae and description of Krasilnikovia cinnamonea sp. nov. Actinomycetologica 21, 1–10. doi: 10.3209/saj.SAJ210101
Ara, I., Matsumoto, A., Bakir, M. A., Kudo, T., Omura, S., and Takahashi, Y. (2008b). Pseudosporangium ferrugineum gen. nov., sp. nov., a new member of the family Micromonosporaceae. Int. J. Syst. Evol. Microbiol. 58, 1644–1652. doi: 10.1099/ijs.0.65680-0
Arcamone, F. M., Bertazzoli, C., Ghione, M., and Scotti, T. (1959). Melanosporin and elaiophylin, new antibiotics from Streptomyces melanosporus (sirve Melanosporofaciens) n. sp. G. di Microbiol. 7, 207–216.
Asahi, K., Nagatsu, J., and Suzuki, S. (1966). Xanthocidin, a new antibiotic. J. Antibiot. Ser. A 19, 195–199.
Aubin, G. G., Bémer, P., Kambarev, S., Patel, N. B., Lemenand, O., Caillon, J., et al. (2016). Propionibacterium namnetense sp. nov., isolated from a human bone infection. Int. J. Syst. Evol. Microbiol. 66, 3393–3399. doi: 10.1099/ijsem.0.001204
Auch, A. F., Henz, S. R., Holland, B. R., and Göker, M. (2006). Genome BLAST distance phylogenies inferred from whole plastid and whole mitochondrion genome sequences. BMC Bioinformatics 7:350. doi: 10.1186/1471-2105-7-350
Auch, A. F., Klenk, H.-P., and Göker, M. (2010a). Standard operating procedure for calculating genome-to-genome distances based on high-scoring segment pairs. Stand. Genomic Sci. 2, 142–148. doi: 10.4056/sigs.541628
Auch, A. F., von Jan, M., Klenk, H.-P., and Göker, M. (2010b). Digital DNA-DNA hybridization for microbial species delineation by means of genome-to-genome sequence comparison. Stand. Genomic Sci. 2, 117–134. doi: 10.4056/sigs.531120
Austwick, P. K. C. (1958). Cutaneous streptothricosis, mycotic dermatitis and strawberry foot rot and the genus Dermatophilus Van Saceghem. Vet. Rev. Annot. 4, 33–48.
Azman, A.-S., Zainal, N., Ab Mutalib, N.-S., Yin, W.-F., Chan, K.-G., and Lee, L.-H. (2016). Monashia flava gen. nov., sp nov., an actinobacterium of the family Intrasporangiaceae. Int. J. Syst. Evol. Microbiol. 66, 554–561. doi: 10.1099/ijsem.0.000753
Baek, I., Kim, M., Lee, I., Na, S.-I., Goodfellow, M., and Chun, J. (2018). Phylogeny trumps chemotaxonomy: a case study involving Turicella otitidis. Front. Microbiol. 9:834. doi: 10.3389/fmicb.2018.00834
Bala, S., Khanna, R., Dadhwal, M., Prabagaran, S. R., Shivaji, S., Cullum, J., et al. (2004). Reclassification of Amycolatopsis mediterranei DSM 46095 as Amycolatopsis rifamycinica sp. nov. Int. J. Syst. Evol. Microbiol. 54, 1145–1149. doi: 10.1099/ijs.0.02901-0
Baldacci, E., and Grein, A. (1966). Streptomyces avellaneus and Streptomyces libani: two new species characterized by a hazel-nut brown (avellaneus) aerial mycelium. G. di Microbiol. 14, 185–198.
Bang, D., Herlin, T., Stegger, M., Andersen, A. B., Torkko, P., Tortoli, E., et al. (2008). Mycobacterium arosiense sp. nov., a slowly growing, scotochromogenic species causing osteomyelitis in an immunocompromised child. Int. J. Syst. Evol. Microbiol. 58, 2398–2402. doi: 10.1099/ijs.0.65503-0
Bapteste, E., and Boucher, Y. (2009). Epistemological impacts of horizontal gene transfer on classification in microbiology, in Horizontal Gene Transfer. Methods in Molecular Biology, eds M. B. Gogarten, J. P. Gogarten, and L. C. Olendzenski (New York, NY: Humana Press), 55–72.
Barka, E. A., Vatsa, P., Sanchez, L., Gaveau-Vaillant, N., Jacquard, C., Klenk, H.-P., et al. (2016). Taxonomy, physiology, and natural products of Actinobacteria. Microbiol. Mol. Biol. Rev. 80, 1–43. doi: 10.1128/MMBR.00019-15.Address
Barksdale, L., Lanéelle, M.-A., Pollice, M, C., Asselineau, J., Welby, M., and Norgard, M. V. (1979). Biological and chemical basis for the reclassification of Microbacterium flavum Orla-Jensen as Corynebacterium flavescens nom. nov. Int. J. Syst. Bacteriol. 29, 222–233.
Barry, C. E., Lee, R. E., Mdluli, K., Sampson, A. E., Schroeder, B. G., Slayden, R. A., et al. (1998). Mycolic acids: structure, biosynthesis and physiological functions. Prog. Lipid Res. 37, 143–179. doi: 10.1016/S0163-7827(98)00008-3
Bartz, Q. R., Ehrlich, J., Mold, J. D., Penner, M. A., and Smith, R. M. (1951). Viomycin, a new tuberculostatic antibiotic. Am. Rev. Tuberc. 63, 4–6.
Batty, I. (1958). Actinomyces odontolyticus, a new species of actinomycete regularly isolated from deep carious dentine. J. Pathol. Bacteriol. 75, 455–459.
Becking, J. H. (1970). Frankiaceae fam. nov. (Actinomycetales) with one new combination and six new species of the genus Frankia Brunchorst 1886,174. Int. J. Syst. Bacteriol. 20, 201–220.
Behrendt, U., Schumann, P., Hamada, M., Suzuki, K., Spröer, C., and Ulrich, A. (2011). Reclassification of Leifsonia ginsengi (Qiu et al. 2007) as Herbiconiux ginsengi gen. nov., comb. nov. and description of Herbiconiux solani sp. nov., an actinobacterium associated with the phyllosphere of Solanum tuberosum L. Int. J. Syst. Evol. Microbiol. 61, 1039–1047. doi: 10.1099/ijs.0.021352-0
Behrendt, U., Ulrich, A., and Schumann, P. (2001). Description of Microbacterium foliorum sp. nov. and Microbacterium phyllosphaerae sp. nov., isolated from the phyllosphere of grasses and the surface litter after mulching the sward, and reclassification of Aureobacterium resistens (Funke et al. 1998) as Microbacterium resistens comb. nov. Int. J. Syst. Evol. Microbiol. 51, 1267–1276.
Bell, M. E., Bernard, K. A., Harrington, S. M., Patel, N. B., Tucker, T.-A., Metcalfe, M. G., et al. (2016). Lawsonella clevelandensis gen. nov., sp. nov., a new member of the suborder Corynebacterineae isolated from human abscesses. Int. J. Syst. Evol. Microbiol. 66, 2929–2935. doi: 10.1099/ijsem.0.001122
Ben Salah, I., Cayrou, C., Raoult, D., and Drancourt, M. (2009). Mycobacterium marseillense sp. nov., Mycobacterium timonense sp. nov. and Mycobacterium bouchedurhonense sp. nov., members of the Mycobacterium avium complex. Int. J. Syst. Evol. Microbiol. 59, 2803–2808. doi: 10.1099/ijs.0.010637-0
Bendinger, B., Rainey, F. A., Kroppenstedt, R. M., Moormann, M., and Klatte, S. (1995). Gordona hydrophobica sp. nov., isolated from biofilters for waste gas treatment. Int. J. Syst. Bacteriol. 45, 544–548.
Benito, P., Alonso-Vega, P., Aguado, C., Luján, R., Anzai, Y., Hirsch, A. M., et al. (2017). Monitoring the colonization and infection of legume nodules by Micromonospora in co-inoculation experiments with rhizobia. Sci. Rep. 7, 11051. doi: 10.1038/s41598-017-11428-1
Beretta, G. (1973). Actinoplanes italicus, a new red-pigmented species. Int. J. Syst. Bacteriol. 23, 37–42.
Bergey, D. H., Harrison, F. C., Breed, R. S., Hammer, B. W., and Huntoon, F. M. (eds.). (1923). Bergey's Manual of Determinative Bacteriology, 1st Edn. Baltimore, MD: The Williams & Wilkins Co.
Bergey, D. H., Harrison, F. C., Breed, R. S., Hammer, B. W., and Huntoon, F. M. (eds.). (1925). Bergey's Manual of Determinative Bacteriology, 2nd Edn. Baltimore, MD: The Williams & Wilkins Co.
Bernard, K. A., Pacheco, A. L., Loomer, C., Burdz, T., Wiebe, D., Huynh, C., et al. (2016). Corynebacterium lowii sp. nov. and Corynebacterium oculi sp. nov., derived from human clinical disease and an emended description of Corynebacterium mastitidis. Int. J. Syst. Evol. Microbiol. 66, 2803–2812. doi: 10.1099/ijsem.0.001059
Bernard, K. A., Wiebe, D., Burdz, T., Reimer, A., Ng, B., Singh, C., et al. (2010). Assignment of Brevibacterium stationis (ZoBell and Upham 1944) Breed 1953 to the genus Corynebacterium, as Corynebacterium stationis comb. nov., and emended description of the genus Corynebacterium to include isolates that can alkalinize citrate. Int. J. Syst. Evol. Microbiol. 60, 874–879.
Bian, J., Li, Y., Wang, J., Song, F.-H., Liu, M., Dai, H.-Q., et al. (2009). Amycolatopsis marina sp. nov., an actinomycete isolated from an ocean sediment. Int. J. Syst. Evol. Microbiol. 59, 477–481. doi: 10.1099/ijs.0.000026-0
Biavati, B., and Mattarelli, P. (1991). Bifidobacterium ruminantium sp. nov. and Bifidobacterium merycium sp. nov. from the rumens of cattle. Int. J. Syst. Bacteriol. 41, 163–168.
Biavati, B., Mattarelli, P., and Crociani, F. (1991). Bifidobacterium saeculare: a new species isolated from feces of rabbit. Syst. Appl. Microbiol. 14, 389–392.
Biavati, B., Scardovi, V., and Moore, W. E. C. (1982). Electrophoretic pattern of proteins in the genus Bifidobacterium and proposal of four new species. Int. J. Syst. Bacteriol. 32, 358–373.
Blanchard, R. (1896). Parasites végétaux à l'exclusion des bactéries, in Traité de Pathologie Générale, Vol. 2, ed C. Bouchard (Paris: G. Masson), 1–932.
Bojalil, L. F., Cerbón, J., and Trujillo, A. (1962). Adansonian classification of mycobacteria. J. Gen. Microbiol. 28, 333–346. doi: 10.1099/00221287-28-2-333
Bönicke, R., and Juhasz, S. E. (1964). Beschreibung der neuen Species Mycobacterium vaccae n. sp. Zbl. Bakt. Hyg. I Abt. Orig. 192, 133–135.
Boondaeng, A., Suriyachadkun, C., Ishida, Y., Tamura, T., Tokuyama, S., and Kitpreechavanich, V. (2011). Herbidospora sakaeratensis sp. nov., isolated from soil, and reclassification of Streptosporangium claviforme as a later synonym of Herbidospora cretacea. Int. J. Syst. Evol. Microbiol. 61, 777–780. doi: 10.1099/ijs.0.024315-0
Böttger, E. C., Hirschel, B., and Coyle, M. B. (1993). Mycobacterium genavense sp. nov. Int. J. Syst. Bacteriol. 43, 841–843. doi: 10.1099/00207713-43-4-841
Breider, S., Scheuner, C., Schumann, P., Fiebig, A., Petersen, J., Pradella, S., et al. (2014). Genome-scale data suggest reclassifications in the Leisingera-Phaeobacter cluster including proposals for Sedimentitalea gen. nov. and Pseudophaeobacter gen. nov. Front. Microbiol. 5:416. doi: 10.3389/fmicb.2014.00416
Brennan, N. M., Brown, R., Goodfellow, M., Ward, A. C., Beresford, T. P., Simpson, P. J., et al. (2001a). Corynebacterium mooreparkense sp. nov. and Corynebacterium casei sp. nov., isolated from the surface of a smear-ripened cheese. Int. J. Syst. Evol. Microbiol. 51, 843–852. doi: 10.1099/00207713-51-3-843
Brennan, N. M., Brown, R., Goodfellow, M., Ward, A. C., Beresford, T. P., Vancanneyt, M., et al. (2001b). Microbacterium gubbeenense sp. nov., from the surface of a smear-ripened cheese. Int. J. Syst. Evol. Microbiol. 51, 1969–1976.
Brenner, D. J., Krieg, N. R., Staley, J. T., and Garrity, G. M. (eds.). (2005a). Bergey's Manual of Systematic Bacteriology, Vol. 2, The Proteobacteria (Part A), 2nd Edn. New York, NY: Springer.
Brenner, D. J., Krieg, N. R., Staley, J. T., and Garrity, G. M. (eds.). (2005b). Bergey's Manual of Systematic Bacteriology, Vol. 2, The Proteobacteria (Part B). 2nd Edn. New York, NY: Springer.
Brenner, D. J., Krieg, N. R., Staley, J. T., and Garrity, G. M. (eds.). (2005c). Bergey's Manual of Systematic Bacteriology, Vol. 2, The Proteobacteria (Part C), 2nd Edn. New York, NY: Springer.
Brocq-Rousseau, D. (1904). Sur un Streptothrix cause de l'altération des avoines moisies. Rev. Générale Bot. 16, 219–230.
Brown, B. A., Springer, B., Steingrube, V. A., Wilson, R. W., Pfyffer, G. E., Garcia, M. J., et al. (1999). Mycobacterium wolinskyi sp. nov. and Mycobacterium goodii sp. nov., two new rapidly growing species related to Mycobacterium smegmatis and associated with human wound infections: a cooperative study from the international working group on Mycobacterial taxonomy. Int. J. Syst. Bacteriol. 49, 1493–1511. doi: 10.1099/00207713-49-4-1493
Brown, J. M., Steigerwalt, A. G., Morey, R. E., Daneshvar, M. I., Romero, L.-J., and McNeil, M. M. (2006). Characterization of clinical isolates previously identified as Oerskovia turbata: proposal of Cellulosimicrobium funkei sp. nov. and emended description of the genus Cellulosimicrobium. Int. J. Syst. Evol. Microbiol. 56, 801–804. doi: 10.1099/ijs.0.63882-0
Brown, R., Hazen, E. L., and Mason, A. (1953). Effect of fungicidin (Nystatin) in mice injected with lethal mixtures of aureomycin and Candida albicans. Science 117, 609–610.
Buchanan, A. M., Scott, J. L., Gerencser, M. A., Beaman, B. L., Jang, S., and Biberstein, E. L. (1984). Actinomyces hordeovulneris sp. nov. an agent of canine Actinomycosis. Int. J. Syst. Bacteriol. 34, 439–443. doi: 10.1099/00207713-34-4-439
Buchanan, R. E. (1918). Studies in the classification and nomenclature of the bacteria. VIII. The subgroups and genera of the Actinomycetales. J. Bacteriol. 3, 403–406.
Bull, A. T., Asenjo, J. A., Goodfellow, M., and Gómez-Silva, B. (2016). The Atacama Desert: technical resources and the growing importance of novel microbial diversity. Annu. Rev. Microbiol. 70, 215–234. doi: 10.1146/annurev-micro-102215-095236
Busarakam, K., Brown, R., Bull, A. T., Tan, G. Y. A., Zucchi, T. D., da Silva, L. J., et al. (2016). Classification of thermophilic actinobacteria isolated from arid desert soils, including the description of Amycolatopsis deserti sp. nov. Antonie Van Leeuwenhoek 109, 319–334. doi: 10.1007/s10482-015-0635-8
Busse, H.-J. (2012). Order X. Micrococcales Prévot 1940, 223AL, in Bergey's Manual of Systematic Bacteriology, Vol. 5, The Actinobacteria, eds M. Goodfellow, P. Kämpfer, H.-J. Busse, M. E. Trujillo, K. Suzuki, W. Ludwig, et al. (New York, NY: Springer), 569–570.
Busse, H.-J. (2016). Review of the taxonomy of the genus Arthrobacter, emendation of the genus Arthrobacter sensu lato, proposal to reclassify selected species of the genus Arthrobacter in the novel genera Glutamicibacter gen. nov., Paeniglutamicibacter gen. nov., Pseudoglutamicibacter gen. nov., Paenarthrobacter gen. nov. and Pseudarthrobacter gen. nov., and emended description of Arthrobacter roseus. Int. J. Syst. Evol. Microbiol. 66, 9–37. doi: 10.1099/ijsem.0.000702
Busse, H.-J., Wieser, M., and Buczolits, S. (2012). Genus III. Arthrobacter Conn and Dimmick 1947, 301 AL emend. Koch, Schumann and Stackbrandt 1995, 838, in Bergey's Manual of Systematic Bacteriology, Vol. 5, The Actinobacteria, eds M. Goodfellow, P. Kämpfer, H.-J. Busse, M. E. Trujillo, K. Suzuki, W. Ludwig, et al. (New York, NY: Springer), 578–624.
Busti, E., Cavaletti, L., Monciardini, P., Schumann, P., Rohde, M., Sosio, M., et al. (2006). Catenulispora acidiphila gen. nov., sp. nov., a novel, mycelium-forming actinomycete, and proposal of Catenulisporaceae fam. nov. Int. J. Syst. Evol. Microbiol. 56, 1741–1746. doi: 10.1099/ijs.0.63858-0
Butler, W. R., Floyd, M. M., Brown, J. M., Toney, S. R., Daneshvar, M. I., Cooksey, R. C., et al. (2005). Novel mycolic acid-containing bacteria in the family Segniliparaceae fam. nov., including the genus Segniliparus gen. nov., with descriptions of Segniliparus rotundus sp. nov. and Segniliparus rugosus sp. nov. Int. J. Syst. Evol. Microbiol. 55, 1615–1624. doi: 10.1099/ijs.0.63465-0
Butler, W. R., O'Connor, S. P., Yakrus, M. A., Smithwick, R. W., Plikaytis, B. B., Moss, C. W., et al. (1993). Mycobacterium celatum sp. nov. Int. J. Syst. Bacteriol. 43, 539–548. doi: 10.1099/00207713-43-3-539
Cai, M., Wang, L., Cai, H., Li, Y., Wang, Y.-N., Tang, Y.-Q., et al. (2011). Salinarimonas ramus sp. nov. and Tessaracoccus oleiagri sp. nov., isolated from a crude oil-contaminated saline soil. Int. J. Syst. Evol. Microbiol. 61, 1767–1775. doi: 10.1099/ijs.0.025932-0
Camacho, C., Coulouris, G., Avagyan, V., Ma, N., Papadopoulos, J., Bealer, K., et al. (2009). BLAST+: architecture and applications. BMC Bioinformatics 10:421. doi: 10.1186/1471-2105-10-421
Camacho, M., Redondo-Gomez, S., Rodríguez-Llorente, I., Rohde, M., Spröer, C., Schumann, P., et al. (2017). Kocuria salina sp. nov., an actinobacterium isolated from the rhizosphere of the halophyte Arthrocnemum macrostachyum and emended description of Kocuria turfanensis. Int. J. Syst. Evol. Microbiol. 67, 5006–5012. doi: 10.1099/ijsem.0.002401
Camas, M., Veyisoglu, A., Tatar, D., Saygin, H., Cetin, D., Sazak, A., et al. (2013). Lechevalieria nigeriaca sp. nov., isolated from arid soil. Int. J. Syst. Evol. Microbiol. 63, 3750–3754. doi: 10.1099/ijs.0.052266-0
Cao, Y.-R., Jiang, Y., Wu, J.-Y., Xu, L.-H., and Jiang, C.-L. (2009). Actinopolymorpha alba sp. nov., isolated from a rhizosphere soil. Int. J. Syst. Evol. Microbiol. 59, 2200–2203. doi: 10.1099/ijs.0.010280-0
Carlsohn, M. R., Groth, I., Saluz, H.-P., Schumann, P., and Stackebrandt, E. (2008). Fodinicola feengrottensis gen. nov., sp. nov., an actinomycete isolated from a medieval mine. Int. J. Syst. Evol. Microbiol. 58, 1529–1536. doi: 10.1099/ijs.0.65512-0
Carlsohn, M. R., Groth, I., Tan, G. Y. A., Schütze, B., Saluz, H.-P., Munder, T., et al. (2007). Amycolatopsis saalfeldensis sp. nov., a novel actinomycete isolated from a medieval alum slate mine. Int. J. Syst. Evol. Microbiol. 57, 1640–1646. doi: 10.1099/ijs.0.64903-0
Carreto, L., Moore, E., Nobre, M. F., Wait, R., Riley, P. W., Sharp, R. J., et al. (1996). Rubrobacter xylanophilus sp. nov., a new thermophilic species isolated from a thermally polluted efluent. Int. J. Syst. Bacteriol. 46, 460–465.
Carro, L., Nouioui, I., Sangal, V., Meier-Kolthoff, J. P., Trujillo, M. E., Montero-Calasanz, M., del, C., et al. (2018). Genome-based classification of micromonosporae with a focus on their biotechnological and ecological potential. Sci. Rep. 8, 525. doi: 10.1038/s41598-017-17392-0
Carro, L., Pukall, R., Spröer, C., Kroppenstedt, R. M., and Trujillo, M. E. (2012). Micromonospora cremea sp. nov. and Micromonospora zamorensis sp. nov., isolated from the rhizosphere of Pisum sativum. Int. J. Syst. Evol. Microbiol. 62, 2971–2977. doi: 10.1099/ijs.0.038695-0
Castellani, A., and Chalmers, A. J. (1913). Manual of Tropical Medicine, 2nd Edn. London: Bailliere, Tindall and Cox.
Castellani, A., and Chalmers, A. J. (1919). Manual of Tropical Medicine, 3rd Edn. New York, NY: William Wood and Co.
Cato, E. P., Moore, W. E. C., Nygaard, G., and Holdeman, L. V. (1984). Actinomyces meyeri sp. nov., specific epithet rev. Int. J. Syst. Bacteriol. 34, 487–489. doi: 10.1099/00207713-34-4-487
Cercos, A. P., Eilberg, B. L., Goyena, J. G., Souto, J., Vautier, E. E., and Widuczynski, I. (1962). Misionina: antibiótico poliénico producido por Streptomyces misionensis n. sp. Rev. Investig. Agric. 16, 5–27.
Chandra, G., and Chater, K. F. (2014). Developmental biology of Streptomyces from the perspective of 100 actinobacterial genome sequences. FEMS Microbiol. Rev. 38, 345–379. doi: 10.1111/1574-6976.12047
Chang, X., Liu, W., and Zhang, X.-H. (2012). Salinactinospora qingdaonensis gen. nov., sp. nov., a halophilic actinomycete isolated from a salt pond. Int. J. Syst. Evol. Microbiol. 62, 954–959. doi: 10.1099/ijs.0.031088-0
Charfreitag, O., Collins, M. D., and Stackebrandt, E. (1988). Reclassification of Arachnia propionica as Propionibacterium propionicus comb. nov. Int. J. Syst. Bacteriol. 38, 354–357.
Chen, H.-H., Li, W.-J., Tang, S.-K., Kroppenstedt, R. M., Stackebrandt, E., Xu, L.-H., et al. (2004). Corynebacterium halotolerans sp. nov., isolated from saline soil in the west of China. Int. J. Syst. Evol. Microbiol. 54, 779–782. doi: 10.1099/ijs.0.02919-0
Chen, J., Su, J.-J., Wei, Y.-Z., Li, Q.-P., Yu, L.-Y., Liu, H.-Y., et al. (2010). Amycolatopsis xylanica sp. nov., isolated from soil. Int. J. Syst. Evol. Microbiol. 60, 2124–2128. doi: 10.1099/ijs.0.016865-0
Chester, F. D. (1897). Report of mycologist: bacteriological work. Delaware Agric. Exp. Stn. Bull. 9, 38–145.
Cho, S.-H., Han, J.-H., Ko, H.-Y., and Kim, S. B. (2008). Streptacidiphilus anmyonensis sp. nov., Streptacidiphilus rugosus sp. nov. and Streptacidiphilus melanogenes sp. nov., acidophilic actinobacteria isolated from Pinus soils. Int. J. Syst. Evol. Microbiol. 58, 1566–1570. doi: 10.1099/ijs.0.65480-0
Chun, J., Bae, K. S., Moon, E. Y., Jung, S.-O., Lee, H. K., and Kim, S.-J. (2000). Nocardiopsis kunsanensis sp. nov., a moderately halophilic actinomycete isolated from a saltern. Int. J. Syst. Evol. Microbiol. 50, 1909–1913. doi: 10.1099/00207713-50-5-1909
Chun, J., Blackall, L. L., Kang, S.-O., Hah, Y. C., and Goodfellow, M. (1997a). A proposal to reclassify Nocardia pinensis Blackall et al. as Skermania piniformis gen. nov., comb. nov. Int. J. Syst. Bacteriol. 47, 127–131.
Chun, J., Kim, S. B., Oh, K. Y., Seong, C.-N., Lee, D.-H., Bae, K. S., et al. (1999). Amycolatopsis thermoflava sp. nov., a novel soil actinomycete from Hainan Island, China. Int. J. Syst. Bacteriol. 49, 1369–1373.
Chun, J., Oren, A., Ventosa, A., Christensen, H., Arahal, D. R., da Costa, M. S., et al. (2018). Proposed minimal standards for the use of genome data for the taxonomy of prokaryotes. Int. J. Syst. Evol. Microbiol. 68, 461–466. doi: 10.1099/ijsem.0.002516
Chun, J., Youn, H.-D., Yim, Y.-I., Lee, H., Kim, M. Y., Hah, Y. C., et al. (1997b). Streptomyces seoulensis sp. nov. Int. J. Syst. Bacteriol. 47, 492–498.
Chung, Y. R., Sung, K. C., Mo, H. K., Son, D. Y., Nam, J. S., Chun, J., et al. (1999). Kitasatospora cheerisanensis sp. nov., a new species of the genus Kitasatospora that produces an antifungal agent. Int. J. Syst. Bacteriol. 49, 753–758.
Clark, D. A., and Norris, P. R. (1996). Acidimicrobium ferrooxidans gen. nov., sp. nov.: mixed-culture ferrous iron oxidation with Sulfobacillus species. Microbiology 142, 785–790.
Clark, L. C., and Hodgkin, J. (2015). Leucobacter musarum subsp. musarum sp. nov., subsp. nov., Leucobacter musarum subsp. japonicus subsp. nov., and Leucobacter celer subsp. astrifaciens subsp. nov., three nematopathogenic bacteria isolated from Caenorhabditis, with an emended description of Leucobacter celer. Int. J. Syst. Evol. Microbiol. 65, 3977–3984. doi: 10.1099/ijsem.0.000523
Clavel, T., Charrier, C., Braune, A., Wenning, M., Blaut, M., and Haller, D. (2009). Isolation of bacteria from the ileal mucosa of TNFdeltaARE mice and description of Enterorhabdus mucosicola gen. nov., sp. nov. Int. J. Syst. Evol. Microbiol. 59, 1805–1812. doi: 10.1099/ijs.0.003087-0
Clavel, T., Charrier, C., Wenning, M., and Haller, D. (2013). Parvibacter caecicola gen. nov., sp. nov., a bacterium of the family Coriobacteriaceae isolated from the caecum of a mouse. Int. J. Syst. Evol. Microbiol. 63, 2642–2648. doi: 10.1099/ijs.0.045344-0
Clavel, T., Duck, W., Charrier, C., Wenning, M., Elson, C., and Haller, D. (2010). Enterorhabdus caecimuris sp. nov., a member of the family Coriobacteriaceae isolated from a mouse model of spontaneous colitis, and emended description of the genus Enterorhabdus Clavel et al. 2009. Int. J. Syst. Evol. Microbiol. 60, 1527–1531. doi: 10.1099/ijs.0.015016-0
Cloud, J. L., Meyer, J. J., Pounder, J. I., Jost, K. C., Sweeney, A., Carroll, K. C., et al. (2006). Mycobacterium arupense sp. nov., a non-chromogenic bacterium isolated from clinical specimens. Int. J. Syst. Evol. Microbiol. 56, 1413–1418. doi: 10.1099/ijs.0.64194-0
Cole, J. K., Gieler, B. A., Heisler, D. L., Palisoc, M. M., Williams, A. J., Dohnalkova, A. C., et al. (2013). Kallotenue papyrolyticum gen. nov., sp. nov., a cellulolytic and filamentous thermophile that represents a novel lineage (Kallotenuales ord. nov., Kallotenuaceae fam. nov.) within the class Chloroflexia. Int. J. Syst. Evol. Microbiol. 63, 4675–4682. doi: 10.1099/ijs.0.053348-0
Collins, M. D. (1982a). Corynebacterium mycetoides sp. nov., nom. rev. Zbl. Bakt. Hyg. I Abt. Orig. C3, 399–400.
Collins, M. D. (1982b). Reclassification of Bacterionema matruchotii (Mendel) in the genus Corynebacterium, as Corynebacterium matruchotii comb. nov. Zbl. Bakt. Hyg. I Abt. Orig. C3, 364–367. doi: 10.1016/S0721-9571(82)80016-1
Collins, M. D. (1987). Transfer of Brevibacterium ammoniagenes (Cooke and Keith) to the genus Corynebacterium as Corynebacterium ammoniagenes comb. nov. Int. J. Syst. Bacteriol. 37, 442–443.
Collins, M. D., Brown, J., and Jones, D. (1988a). Brachybacterium faecium gen. nov., sp. nov., a coryneform bacterium from poultry deep litter. Int. J. Syst. Bacteriol. 38, 45–48.
Collins, M. D., Falsen, E., Åkervall, E., Sjöden, B., and Alvarez, A. (1998). Corynebacterium kroppenstedtii sp. nov., a novel corynebacterium that does not contain mycolic acids. Int. J. Syst. Bacteriol. 48, 1449–1454.
Collins, M. D., Farrow, J. A. E., Goodfellow, M., and Minnikin, D. E. (1983). Brevibacterium casei sp. nov. and Brevibacterium epidermidis sp. nov. Syst. Appl. Microbiol. 4, 388–395. doi: 10.1016/S0723-2020(83)80023-X
Collins, M. D., Hoyles, L., Foster, G., and Falsen, E. (2004a). Corynebacterium caspium sp. nov., from a Caspian seal (Phoca caspica). Int. J. Syst. Evol. Microbiol. 54, 925–928. doi: 10.1099/ijs.0.02950-0
Collins, M. D., Hoyles, L., Foster, G., Sjödén, B., and Falsen, E. (2001a). Corynebacterium capitovis sp. nov., from a sheep. Int. J. Syst. Evol. Microbiol. 51, 857–860. doi: 10.1099/00207713-51-3-857
Collins, M. D., Hoyles, L., Hutson, R. A., Foster, G., and Falsen, E. (2001b). Corynebacterium testudinoris sp. nov., from a tortoise, and Corynebacterium felinum sp. nov., from a Scottish wild cat. Int. J. Syst. Evol. Microbiol. 51, 1349–1352. doi: 10.1099/00207713-51-4-1349
Collins, M. D., Hoyles, L., Kalfas, S., Sundquist, G., Monsen, T., Nikolaitchouk, N., et al. (2000a). Characterization of Actinomyces isolates from infected root canals of teeth: description of Actinomyces radicidentis sp. nov. J. Clin. Microbiol. 38, 3399–3403.
Collins, M. D., Hoyles, L., Lawson, P. A., Falsen, E., Robson, R. L., and Foster, G. (1999). Phenotypic and phylogenetic characterization of a new Corynebacterium species from dogs: description of Corynebacterium auriscanis sp. nov. J. Clin. Microbiol. 37, 3443–3447.
Collins, M. D., Hutson, R. A., Båverud, V., and Falsen, E. (2000b). Characterization of a Rothia-like organism from a mouse: description of Rothia nasimurium sp. nov. and reclassification of Stomatococcus mucilaginosus as Rothia mucilaginosa comb. nov. Int. J. Syst. Evol. Microbiol. 50, 1247–1251. doi: 10.1099/00207713-50-3-1247
Collins, M. D., Jones, D., and Schofield, G. M. (1982). Reclassification of ‘Corynebacterium haernolyticum’' (MacLean, Liebow & Rosenberg) in the genus Arcanobacterium gen. nov. as Arcanobacterium haemolyticum nom. rev., comb. nov.' J. Gen. Microbiol. 128, 1279–1281.
Collins, M. D., Routh, J., Saraswathy, A., Lawson, P. A., Schumann, P., Welinder-Olsson, C., et al. (2004b). Arsenicicoccus bolidensis gen. nov., sp. nov., a novel actinomycete isolated from contaminated lake sediment. Int. J. Syst. Evol. Microbiol. 54, 605–608. doi: 10.1099/ijs.0.02918-0
Collins, M. D., Smida, J., Dorsch, M., and Stackebrandt, E. (1988b). Tsukamurella gen. nov. harboring Corynebacterium paurometabolum and Rhodococcus aurantiacus. Int. J. Syst. Bacteriol. 38, 385–391.
Collins, M. D., Stubbs, S., Hommez, J., and Devriese, L. A. (1993). Molecular taxonomic studies of Actinomyces-like bacteria isolated from purulent lesions in pigs and description of Actinomyces hyovaginalis sp. nov. Int. J. Syst. Bacteriol. 43, 471–473.
Collins, M. D., and Wallbanks, S. (1992). Comparative sequence analyses of the 16S rRNA genes of Lactobacillus minutus, Lactobacillus rimae and Streptococcus parvulus: proposal for the creation of a new genus Atopobium. FEMS Microbiol. Lett. 95, 235–240. doi: 10.1016/0378-1097(92)90435-Q
Colwell, R. R. (1970). Polyphasic taxonomy of the genus vibrio: numerical taxonomy of Vibrio cholerae, Vibrio parahaemolyticus, and related Vibrio species. J. Bacteriol. 104, 410–433.
Conn, H. J., and Dimmick, I. (1947). Soil bacteria similar in morphology to Mycobacterium and Corynebacterium. J. Bacteriol. 54, 291–303.
Conville, P. S., Brown, J. M., Steigerwalt, A. G., Lee, J. W., Anderson, V. L., Fishbain, J. T., et al. (2004). Nocardia kruczakiae sp. nov., a pathogen in immunocompromised patients and a member of the “N. nova complex.” J. Clin. Microbiol. 42, 5139–5145. doi: 10.1128/JCM.42.11.5139
Cooksey, R. C., de Waard, J. H., Yakrus, M. A., Rivera, I., Chopite, M., Toney, S. R., et al. (2004). Mycobacterium cosmeticum sp. nov., a novel rapidly growing species isolated from a cosmetic infection and from a nail salon. Int. J. Syst. Evol. Microbiol. 54, 2385–2391. doi: 10.1099/ijs.0.63238-0
Cools, P., Oyaert, M., Vaneechoutte, M., De Laere, E., and Vervaeke, S. (2014). Atopobium deltae sp. nov., isolated from the blood of a patient with Fournier's gangrene. Int. J. Syst. Evol. Microbiol. 64, 3140–3145. doi: 10.1099/ijs.0.065243-0
Couch, J. N. (1950). Actinoplanes a new genus of the Actinomycetales. J. Elisha Mitchell Sci. Soc. 66, 87–92.
Couch, J. N. (1955). A new genus and family of the Actinomycetales, with a revision of the genus Actinoplanes. J. Elisha Mitchell Sci. Soc. 71, 148–155.
Couch, J. N. (1963). Some new genera and species of the Actinoplanaceae. J. Elisha Mitchell Sci. Soc. 79, 53–70.
Cowan, S. T. (1955). The principles of microbial classification. Introduction: the philosophy of classification. J. Gen. Microbiol. 12, 314–321.
Cross, T., and Unsworth, B. A. (1981). The taxonomy of the endospore-forming actinomycetes, in The Aerobic, Endospore-forming Bacteria, Classification and Identification, eds R. C. W. Berkeley and M. Goodfellow (London: Academic Press), 17–32.
Cui, Q., Wang, L., Huang, Y., Liu, Z., and Goodfellow, M. (2005). Nocardia jiangxiensis sp. nov. and Nocardia miyunensis sp. nov., isolated from acidic soils. Int. J. Syst. Evol. Microbiol. 55, 1921–1925. doi: 10.1099/ijs.0.63644-0
Cui, X.-L., Mao, P.-H., Zeng, M., Li, W.-J., Zhang, L.-P., Xu, L.-H., et al. (2001). Streptimonospora salina gen. nov., sp. nov., a new member of the family Nocardiopsaceae. Int. J. Syst. Evol. Microbiol. 51, 357–363. doi: 10.1099/00207713-51-2-357
da Costa Cruz, J. C. (1938). Mycobacterium fortuitum um novo bacillo acido-resistente patogenico para o homen. Acta Medica (Rio Janeiro) 1, 297–301.
Dai, H.-Q., Wang, J., Xin, Y.-H., Pei, G., Tang, S.-K., Ren, B., et al. (2010). Verrucosispora sediminis sp. nov., a cyclodipeptide-producing actinomycete from deep-sea sediment. Int. J. Syst. Evol. Microbiol. 60, 1807–1812. doi: 10.1099/ijs.0.017053-0
Daneshvar, M. I., Hollis, D. G., Weyant, R. S., Jordan, J. G., MacGregor, J. P., Morey, R. E., et al. (2004). Identification of some charcoal-black-pigmented CDC fermentative coryneform group 4 isolates as Rothia dentocariosa and some as Corynebacterium aurimucosum: proposal of Rothia dentocariosa emend. Georg and Brown 1967, Corynebacterium aurimucosum emend. Yassin et al. 2002, and Corynebacterium nigricans Shukla et al. 2003 pro synon. Corynebacterium aurimucosum. J. Clin. Microbiol. 42, 4189–4198. doi: 10.1128/JCM.42.9.4189
Daniel, H.-M., Lachance, M.-A., and Kurtzman, C. P. (2014). On the reclassification of species assigned to Candida and other anamorphic ascomycetous yeast genera based on phylogenetic circumscription. Antonie Van Leeuwenhoek 106, 67–84. doi: 10.1007/s10482-014-0170-z
Dastager, S. G., Lee, J.-C., Ju, Y.-J., Park, D.-J., and Kim, C.-J. (2008). Phycicoccus bigeumensis sp. nov., a mesophilic actinobacterium isolated from Bigeum Island, Korea. Int. J. Syst. Evol. Microbiol. 58, 2425–2428. doi: 10.1099/ijs.0.65563-0
Dastager, S. G., Lee, J.-C., Ju, Y.-J., Park, D.-J., and Kim, C.-J. (2009). Nocardioides sediminis sp. nov., isolated from a sediment sample. Int. J. Syst. Evol. Microbiol. 59, 280–284. doi: 10.1099/ijs.0.002162-0
Dastager, S. G., Mawlankar, R., Tang, S.-K., Krishnamurthi, S., Venkata Ramana, V., Joseph, N., et al. (2014a). Rhodococcus enclensis sp. nov., a novel member of the genus Rhodococcus. Int. J. Syst. Evol. Microbiol. 64, 2693–2699. doi: 10.1099/ijs.0.061390-0
Dastager, S. G., Qin, L., Tang, S.-K., Krishnamurthi, S., Lee, J.-C., and Li, W.-J. (2014b). Arthrobacter enclensis sp. nov., isolated from sediment sample. Arch. Microbiol. 196, 775–782. doi: 10.1007/s00203-014-1016-9
Dastager, S. G., Tang, S.-K., Srinivasan, K., Lee, J.-C., and Li, W.-J. (2014c). Kocuria indica sp. nov., isolated from a sediment sample. Int. J. Syst. Evol. Microbiol. 64, 869–874. doi: 10.1099/ijs.0.052548-0
Davis, M. J., Gillaspie, A. G., Vidaver, A. K., and Harris, R. W. (1984). Clavibacter: a new genus containing some phytopathogenic coryneform bacteria, including Clavibacter xyli subsp. xyli sp. nov., subsp. nov. and Clavibacter xyli subsp. cynodontis subsp. nov., pathogens that cause ratoon stunting disease of sugarcane and bermudagrass stunting disease. Int. J. Syst. Bacteriol. 34, 107–117. doi: 10.1099/00207713-34-2-107
De Boer, C., Dietz, A., Silver, W. S., and Savage, G. M. (1956). Streptolydigin, a new antimicrobial antibiotic. 1. Biologic studies of streptolydigin. Antibiot. Annu. 1955–1956, 886–892.
De Boer, L., Dijkhuizen, L., Grobben, G., Goodfellow, M., Stackebrandt, E., Parlett, J. H., et al. (1990). Amycolatopsis methanolica sp. nov., a facultatively methylotrophic actinomycete. Int. J. Syst. Bacteriol. 40, 194–204. doi: 10.1099/00207713-40-2-194
De Vos, P., Garrity, G. M., Jones, D., Krieg, N. R., Ludwig, W., Rainey, F. A., et al. (eds.). (2009). Bergey's Manual of Systematic Bacteriology, Vol. 3, The Firmicutes, 2nd Edn. New York, NY: Springer.
Delcenserie, V., Gavini, F., Beerens, H., Tresse, O., Franssen, C., and Daube, G. (2007). Description of a new species, Bifidobacterium crudilactis sp. nov., isolated from raw milk and raw milk cheeses. Syst. Appl. Microbiol. 30, 381–389. doi: 10.1016/j.syapm.2007.01.004
Delwiche, E. A. (1957). Family XI. Propionibacteriaceae Delwiche, fam. nov., in Bergey's Manual of Determinative Bacteriology, eds R. S. Breed, E. G. D. Murray, and N. R. Smith (Baltimore, MD: The Williams & Wilkins Co.), 569.
Demaree, J. B., and Smith, N. R. (1952). Nocardia vaccinii n. sp. causing galls on blueberry plants. Phytopathology 42, 249–252.
Dent, V. E., and Williams, R. A. (1984). Actinomyces denticolens Dent & Williams sp. nov.: a new species from the dental plaque of cattle. J. Appl. Bacteriol. 56, 183–192.
Dent, V. E., and Williams, R. A. D. (1986). Actinomyces slackii sp. nov. from dental plaque of dairy cattle. Int. J. Syst. Bacteriol. 36, 392–395.
Dewhirst, F. E., Paster, B. J., Tzellas, N., Coleman, B., Downes, J., Spartt, D. A., et al. (2001). Characterization of novel human oral isolates and cloned 16S rDNA sequences that fall in the family Coriobacteriaceae: description of Olsenella gen. nov., reclassification of Lactobacillus uli as Olsenella uli comb. nov. and description of Olsenella profusa sp. nov. Int. J. Syst. Evol. Microbiol. 51, 1797–1804. doi: 10.1099/00207713-51-5-1797
Ding, L., Hirose, T., and Yokota, A. (2007). Amycolatopsis echigonensis sp. nov. and Amycolatopsis niigatensis sp. nov., novel actinomycetes isolated from a filtration substrate. Int. J. Syst. Evol. Microbiol. 57, 1747–1751. doi: 10.1099/ijs.0.64791-0
Dodsworth, J. A., Gevorkian, J., Despujos, F., Cole, J. K., Murugapiran, S. K., Ming, H., et al. (2014). Thermoflexus hugenholtzii gen. nov., sp. nov., a thermophilic, microaerophilic, filamentous bacterium representing a novel class in the Chloroflexi, Thermoflexia classis nov., and description of Thermoflexaceae fam. nov. and Thermoflexales ord. nov. Int. J. Syst. Evol. Microbiol. 64, 2119–2127. doi: 10.1099/ijs.0.055855-0
Domenech, P., Jimenez, M. S., Menendez, M. C., Bull, T. J., Samper, S., Manrique, A., et al. (1997). Mycobacterium mageritense sp. nov. Int. J. Syst. Bacteriol. 47, 535–540.
Downes, J., Mantzourani, M., Beighton, D., Hooper, S., Wilson, M. J., Nicholson, A., et al. (2011). Scardovia wiggsiae sp. nov., isolated from the human oral cavity and clinical material, and emended descriptions of the genus Scardovia and Scardovia inopinata. Int. J. Syst. Evol. Microbiol. 61, 25–29. doi: 10.1099/ijs.0.019752-0
Du, H.-J., Zhang, Y.-Q., Liu, H.-Y., Su, J., Wei, Y.-Z., Ma, B.-P., et al. (2013). Allonocardiopsis opalescens gen. nov., sp. nov., a new member of the suborder Streptosporangineae, from the surface-sterilized fruit of a medicinal plant. Int. J. Syst. Evol. Microbiol. 63, 900–904. doi: 10.1099/ijs.0.041491-0
Du, Z.-J., Jordan, E. M., Rooney, A. P., Chen, G.-J., and Austin, B. (2010). Corynebacterium marinum sp. nov. isolated from coastal sediment. Int. J. Syst. Evol. Microbiol. 60, 1944–1947. doi: 10.1099/ijs.0.018523-0
Duggar, B. M. (1948). Aureomycin: a product of the continuing search for new antibiotics. Ann. N. Y. Acad. Sci. 51, 177–181.
Eberson, F. (1918). A bacteriologic study of the diphtheroid organisms with special reference to Hodgkin's disease. J. Infect. Dis. 23, 1–43.
Eguchi, T., Takada, N., Nakamura, S., Tanaka, T., Makino, T., and Oshima, Y. (1993). Streptomyces bungoensis sp. nov. Int. J. Syst. Bacteriol. 43, 794–798.
Ehrlich, J., Gottlieb, D., Burkholder, P. R., Anderson, L. E., and Pridham, T. G. (1948). Streptomyces venezuelae n. sp. the source of chloromycetin. J. Bacteriol. 56, 467–477.
Embley, M. T., Smida, J., and Stackebrandt, E. (1988). The phylogeny of mycolate-less wall chemotype IV Actinomycetes and description of Pseudonocardiaceae fam. nov. Syst. Appl. Microbiol. 11, 44–52. doi: 10.1016/S0723-2020(88)80047-X
Ensign, J. C., and Rittenberg, S. C. (1963). A crystalline pigment produced from 2-hydroxypyridine by Arthrobacter crystallopoietes n. sp. Arch. Mikrobiol. 47, 137–153.
Ernst, W. (1906). Über Pyelonephritis diphtherica bovis und die Pyelonephritisbazillen. Zbl. Bakt. Hyg. I Abt. Orig. 40, 79–91.
Everest, G. J., Cook, A. E., le Roes-Hill, M., and Meyers, P. R. (2011). Nocardia rhamnosiphila sp. nov., isolated from soil. Syst. Appl. Microbiol. 34, 508–512. doi: 10.1016/j.syapm.2011.03.006
Everest, G. J., Curtis, S. M., De Leo, F., Urzì, C., and Meyers, P. R. (2013). Kribbella albertanoniae sp. nov., isolated from a Roman catacomb, and emended description of the genus Kribbella. Int. J. Syst. Evol. Microbiol. 63, 3591–3596. doi: 10.1099/ijs.0.050237-0
Evtushenko, L. I., and Ariskina, E. V. (2012). Family II: Nocardioidaceae Nesterenko, Kvasnikov and Nogina 1990, 320VP (Effective publication: Nesterenko, Kvasnikov and Nogina 1985a, 9) emend. Rainey, Ward-Rainey and Stackebrandt 1997, 484 emend. Zhi, Li and Stackebrandt 2009, 599, in Bergey's Manual of Systematic Bacteriology, Vol. 5, The Actinobacteria, eds M. Goodfellow, P. Kämpfer, H.-J. Busse, M. E. Trujillo, K. Suzuki, W. Ludwig, et al. (New York, NY: Springer), 1189–1197.
Evtushenko, L. I., Dorofeeva, L. V., Dobrovolskaya, T. G., Streshinskaya, G. M., Subbotin, S. A., and Tiedje, J. M. (2001). Agreia bicolorata gen. nov., sp. nov., to accommodate actinobacteria isolated from narrow reed grass infected by the nematode Heteroanguina graminophila. Int. J. Syst. Evol. Microbiol. 51, 2073–2079. doi: 10.1099/00207713-51-6-2073
Evtushenko, L. I., Dorofeeva, L. V., Subbotin, S. A., Cole, J. R., and Tiedje, J. M. (2000a). Leifsonia poae gen. nov., sp. nov., isolated from nematode galls on Poa annua, and reclassification of “Corynebacterium aquaticum” Leifson 1962 as Leifsonia aquatica (ex Leifson 1962) gen. nov., nom. rev., comb. nov. and Clavibacter xyli Davis et al. 1984. Int. J. Syst. Evol. Microbiol. 50, 371–380. doi: 10.1099/00207713-50-1-371
Evtushenko, L. I., Krausova, V. I., and Yoon, J.-H. (2012). Genus I. Nocardioides Prauser 1976, 61 AL, in Bergey's Manual of Systematic Bacteriology, Vol. 5, The Actinobacteria, eds M. Goodfellow, P. Kämpfer, H.-J. Busse, M. E. Trujillo, K. Suzuki, W. Ludwig, et al. (New York, NY: Springer), 1197–1251.
Evtushenko, L. I., Taptykova, S. D., Akimov, V. N., Semyonova, S. A., and Kalakoutskii, L. V. (1991). Glycomyces tenuis sp. nov. Int. J. Syst. Bacteriol. 41, 154–157.
Evtushenko, L. I., Taran, V. V., Akimov, V. N., Kroppenstedt, R. M., Tiedje, J. M., and Stackebrandt, E. (2000b). Nocardiopsis tropica sp. nov., Nocardiopsis trehalosi sp. nov., nom. rev. and Nocardiopsis dassonvillei subsp. albirubida subsp. nov., comb. nov. Int. J. Syst. Evol. Microbiol. 50, 73–81. doi: 10.1099/00207713-50-1-73
Ezeoke, I., Klenk, H.-P., Pötter, G., Schumann, P., Moser, B. D., Lasker, B. A., et al. (2013). Nocardia amikacinitolerans sp. nov., an amikacin-resistant human pathogen. Int. J. Syst. Evol. Microbiol. 63, 1056–1061. doi: 10.1099/ijs.0.039990-0
Fang, B.-Z., Han, M.-X., Liu, L., Zhang, Z.-T., Liu, W.-L., Shen, J.-T., et al. (2017). Lentzea cavernae sp. nov., an actinobacterium isolated from a karst cave sample, and emended description of the genus Lentzea. Int. J. Syst. Evol. Microbiol. 67, 2357–2362. doi: 10.1099/ijsem.0.001958
Fang, C., Zhang, J., Pang, H., Li, Y., Xin, Y., and Zhang, Y. (2011). Nocardiopsis flavescens sp. nov., an actinomycete isolated from marine sediment. Int. J. Syst. Evol. Microbiol. 61, 2640–2645. doi: 10.1099/ijs.0.027987-0
Fang, X.-M., Su, J., Wang, H., Wei, Y.-Z., Zhang, T., Zhao, L.-L., et al. (2013). Williamsia sterculiae sp. nov., isolated from a Chinese medicinal plant. Int. J. Syst. Evol. Microbiol. 63, 4158–4162. doi: 10.1099/ijs.0.052688-0
Fanti, F., Tortoli, E., Hall, L., Roberts, G. D., Kroppenstedt, R. M., Dodi, I., et al. (2004). Mycobacterium parmense sp. nov. Int. J. Syst. Evol. Microbiol. 54, 1123–1127. doi: 10.1099/ijs.0.02760-0
Farris, J. S. (1974). Formal definitions of paraphyly and polyphyly. Syst. Zool. 23, 548–554. doi: 10.2307/2412474
Fernández-Garayzábal, J. F., Collins, M. D., Hutson, R. A., Gonzalez, I., Fernández, E., and Domínguez, L. (1998). Corynebacterium camporealensis sp. nov., associated with subclinical mastitis in sheep. Int. J. Syst. Bacteriol. 48, 463–468.
Fernández-Garayzábal, J. F., Dominguez, L., Pascual, C., Jones, D., and Collins, M. D. (1995). Phenotypic and phylogenetic characterization of some unknown coryneform bacteria isolated from bovine blood and milk: description of Sanguibacter gen. nov. Lett. Appl. Microbiol. 20, 69–75.
Fernández-Garayzábal, J. F., Egido, R., Vela, A. I., Briones, V., Collins, M. D., Mateos, A., et al. (2003). Isolation of Corynebacterium falsenii and description of Corynebacterium aquilae sp. nov., from eagles. Int. J. Syst. Evol. Microbiol. 53, 1135–1138. doi: 10.1099/ijs.0.02533-0
Fernández-Garyzábal, J. F., Vela, A. I., Egido, R., Hutson, R. A., Lanzarot, M. P., Fernández-García, M., et al. (2004). Corynebacterium ciconiae sp. nov., isolated from the trachea of black storks (Ciconia nigra). Int. J. Syst. Evol. Microbiol. 54, 2191–2195. doi: 10.1099/ijs.0.63165-0
Finster, K. W., Herbert, R. A., Kjeldsen, K. U., Schumann, P., and Lomstein, B. A. (2009). Demequina lutea sp. nov., isolated from a high Arctic permafrost soil. Int. J. Syst. Evol. Microbiol. 59, 649–653. doi: 10.1099/ijs.0.004929-0
Floyd, M. M., Gross, W. M., Bonato, D. A., Silcox, V. A., Smithwick, R. W., Metchock, B., et al. (2000). Mycobacterium kubicae sp. nov., a slowly growing, scotochromogenic Mycobacterium. Int. J. Syst. Evol. Microbiol. 50, 1811–1816. doi: 10.1099/00207713-50-5-1811
Floyd, M. M., Guthertz, L. S., Silcox, V. A., Duffey, P. S., Jang, Y., Desmond, E. P., et al. (1996). Characterization of an SAV organism and proposal of Mycobacterium triplex sp. nov. J. Clin. Microbiol. 34, 2963–2967.
Friedman, C. S., Beaman, B. L., Chun, J., Goodfellow, M., Gee, A., and Hedrickl, R. P. (1998). Nocardia crassostreae sp. nov., the causal agent of nocardiosis in Pacific oysters. Int. J. Syst. Bacteriol. 48, 237–246.
Frischmann, A., Knoll, A., Hilbert, F., Zasada, A. A., Kämpfer, P., and Busse, H.-J. (2012). Corynebacterium epidermidicanis sp. nov., isolated from skin of a dog. Int. J. Syst. Evol. Microbiol. 62, 2194–2200. doi: 10.1099/ijs.0.036061-0
Fudou, R., Jojima, Y., Seto, A., Yamada, K., Kimura, E., Nakamatsu, T., et al. (2002). Corynebacterium efficiens sp. nov., a glutamic acid-producing species from soil and vegetables. Int. J. Syst. Evol. Microbiol. 52, 1127–1131. doi: 10.1099/ijs.0.02086-0
Funke, G., Alvarez, N., Pascual, C., Falsen, E., Akervall, E., Sabbe, L., et al. (1997a). Actinomyces europaeus sp. nov., isolated from human clinical specimens. Int. J. Syst. Bacteriol. 47, 687–692.
Funke, G., Bernard, K. A., Bucher, C., Pfyffer, G. E., and D, C. M. (1995). Corynebacterium glucuronolyticum sp. nov., isolated from male patients with genitourinary infections. Med. Microbiol. Lett. 4, 204–215.
Funke, G., Efstratiou, A., Kuklinska, D., Hutson, R. A., De Zoysa, A., Engler, K. H., et al. (1997b). Corynebacterium imitans sp. nov. isolated from patients with suspected diphtheria. J. Clin. Microbiol. 35, 1978–1983.
Funke, G., Englert, R., Frodl, R., Bernard, K. A., and Stenger, S. (2010a). Actinomyces hominis sp. nov., isolated from a wound swab. Int. J. Syst. Evol. Microbiol. 60, 1678–1681. doi: 10.1099/ijs.0.015818-0
Funke, G., Frodl, R., and Bernard, K. A. (2010b). Corynebacterium mustelae sp. nov., isolated from a ferret with lethal sepsis. Int. J. Syst. Evol. Microbiol. 60, 871–873. doi: 10.1099/ijs.0.010942-0
Funke, G., Frodl, R., Bernard, K. A., and Englert, R. (2009). Corynebacterium freiburgense sp. nov., isolated from a wound obtained from a dog bite. Int. J. Syst. Evol. Microbiol. 59, 2054–2057. doi: 10.1099/ijs.0.008672-0
Funke, G., Stubbs, S., Altwegg, M., Carlotti, A., and Collins, M. D. (1994a). Turicella otitidis gen. nov., sp. nov., a coryneform bacterium isolated from patients with otitis media. Int. J. Syst. Bacteriol. 44, 270–273.
Funke, G., Stubbs, S., von Graevenitz, A., and Collins, M. D. (1994b). Assignment of human-derived CDC group 1 coryneform bacteria and CDC group 1-like coryneform bacteria to the genus Actinomyces as Actinomyces neuii subsp. neuii sp. nov., subsp. nov., and Actinomyces neuii subsp. anitratus subsp. nov. Int. J. Syst. Bacteriol. 44, 167–171.
Fusco da Costa, A. R., Fedrizzi, T., Lopes, M. L., Pecorari, M., da Costa, W. L. O., Giacobazzi, E., et al. (2015). Characterization of 17 strains belonging to the Mycobacterium simiae complex and description of Mycobacterium paraense sp. nov. Int. J. Syst. Evol. Microbiol. 65, 656–662. doi: 10.1099/ijs.0.068395-0
Gadkari, D., Schricker, K., Acker, G., Kroppenstedt, R. M., and Meyer, O. (1990). Streptomyces thermoautotrophicus sp. nov., a thermophilic CO- and H2-oxidizing obligate chemolithoautotroph. Appl. Environ. Microbiol. 56, 3727–3734.
Galatenko, O. A., Terekhova, L. P., and Preobrazhenskaya, T. P. (1981). New Actinomadura species isolated from turkmen soil samples and their antagonostic properties. Antibiotiki 26, 803–807.
Gao, R., Liu, C., Zhao, J., Jia, F., Yu, C., Yang, L., et al. (2014). Micromonospora jinlongensis sp. nov., isolated from muddy soil in China and emended description of the genus Micromonospora. Antonie Van Leeuwenhoek 105, 307–315. doi: 10.1007/s10482-013-0074-3
Gause, G. F., Preobrazhenskaya, T. P., Sveshnikova, M. A., Terekhova, L. P., and Maximova, T. S. (1983). A Guide for the Determination of Actinomycetes. Genera Streptomyces, Streptoverticillium, and Chainia. Moscow: Nauka.
Genilloud, O. (2012). Order XI. Micromonosporales ord. nov., in Bergey's Manual of Systematic Bacteriology, Vol. 5, The Actinobacteria, eds M. Goodfellow, P. Kämpfer, H.-J. Busse, M. E. Trujillo, K. Suzuki, W. Ludwig, et al. (New York, NY: Springer), 1035.
Georg, L. K., and Brown, J. M. (1967). Rothia, gen. nov., an aerobic genus of the family Actinomycetaceae. Int. J. Syst. Bacteriol. 17, 79–88.
Ghosh, A., Paul, D., Prakash, D., Mayilraj, S., and Jain, R. K. (2006). Rhodococcus imtechensis sp. nov., a nitrophenol-degrading actinomycete. Int. J. Syst. Evol. Microbiol. 56, 1965–1969. doi: 10.1099/ijs.0.63939-0
Gillis, P., Vandamme, P., De Vos, P., Swings, J., and Kersters, K. (2005). Polyphasic taxonomy, in Bergey's Manual of Systematic Bacteriology, Vol. 2, The Proteobacteria (Part A), eds D. J. Brenner, N. R. Krieg, J. T. Staley, and G. M. Garrity (New York, NY: Springer Verlag), 43–48.
Girard, G., Traag, B. A., Sangal, V., Mascini, N., Hoskisson, P. A., Goodfellow, M., et al. (2013). A novel taxonomic marker that discriminates between morphologically complex actinomycetes. Open Biol. 3:130073. doi: 10.1098/rsob.130073
Glaeser, S. P., and Kämpfer, P. (2015). Multilocus sequence analysis (MLSA) in prokaryotic taxonomy. Syst. Appl. Microbiol. 38, 237–245. doi: 10.1016/j.syapm.2015.03.007
Gochnauer, M. B., Leppard, G. G., Komaratat, P., Kates, M., Novitsky, T., and Kushner, D. J. (1975). Isolation and characterization of Actinopolyspora halophila, gen. et sp. nov., an extremely halophilic actinomycete. Can. J. Microbiol. 21, 1500–1511.
Goloboff, P. A., Farris, J. S., and Nixon, K. C. (2008). TNT, a free program for phylogenetic analysis. Cladistics 24, 774–786. doi: 10.1111/j.1096-0031.2008.00217.x
Goodfellow, M. (1984). Reclassification of Corynebacterium fascians (Tilford) Dowson in the genus Rhodococcus, as Rhodococcus fascians comb. nov. Syst. Appl. Microbiol. 5, 225–229. doi: 10.1016/S0723-2020(84)80023-5
Goodfellow, M. (2012a). Family IV. Nocardiaceae, in Bergey's Manual of Systematic Bacteriology, Vol. 5, The Actinobacteria, eds M. Goodfellow, P. Kämpfer, H.-J. Busse, M. E. Trujillo, K. Suzuki, W. Ludwig, et al. (New York, NY: Springer), 376.
Goodfellow, M. (2012b). Order XV. Streptosporangiales ord. nov., in Bergey's Manual of Systematic Bacteriology, Vol. 5, The Actinobacteria, eds M. Goodfellow, P. Kämpfer, H.-J. Busse, M. E. Trujillo, K. Suzuki, W. Ludwig, et al. (New York, NY: Springer), 1805.
Goodfellow, M., and Alderson, G. (1977). The Actinomycete-genus Rhodococcus: a home for the “rhodochrous” complex. J. Gen. Microbiol. 100, 99–122.
Goodfellow, M., Brown, R., Ahmed, L., Pathom-aree, W., Bull, A. T., Jones, A. L., et al. (2013). Verrucosispora fiedleri sp. nov., an actinomycete isolated from a fjord sediment which synthesizes proximicins. Antonie Van Leeuwenhoek 103, 493–502. doi: 10.1007/s10482-012-9831-y
Goodfellow, M., Chun, J., Stackebrandt, E., and Kroppenstedt, R. M. (2002). Transfer of Tsukamurella wratislaviensis Goodfellow et al. 1995 to the genus Rhodococcus as Rhodococcus wratislaviensis comb. nov. Int. J. Syst. Evol. Microbiol. 52, 749–755. doi: 10.1099/ijs.0.01969-0
Goodfellow, M., Embley, T. M., and Austin, B. (1985). Numerical taxonomy and emended description of Renibacterium salmoninarum. J. Gen. Microbiol. 131, 2739–2752.
Goodfellow, M., and Jones, A. L. (2012). Order V. Corynebacteriales ord. nov., in Bergey's Manual of Systematic Bacteriology, Vol. 5, The Actinobacteria, eds M. Goodfellow, P. Kämpfer, H.-J. Busse, M. E. Trujillo, K. Suzuki, W. Ludwig, et al. (New York, NY: Springer), 234.
Goodfellow, M., Kämpfer, P., Busse, H.-J., Trujillo, M. E., Suzuki, K., Ludwig, W., et al. (eds.). (2012a). Bergey's Manual of Systematic Bacteriology, Vol. 5, The Actinobacteria, 2nd Edn. New York, NY: Springer.
Goodfellow, M., Kim, S. B., Minnikin, D. E., Whitehead, D., Zhou, Z.-H., and Mattinson-Rose, A. D. (2001). Amycolatopsis sacchari sp. nov., a moderately thermophilic actinomycete isolated from vegetable matter. Int. J. Syst. Evol. Microbiol. 51, 187–193. doi: 10.1099/00207713-51-1-187
Goodfellow, M., Lacey, J., Athalye, M., Embley, T. M., and Bowen, T. (1989). Saccharopolyspora gregorri and Saccharopolyspora hordei: two new Actinomycete species from fodder. J. Gen. Microbiol. 135, 2125–2139.
Goodfellow, M., and Minnikin, D. E. (1977). Nocardioform bacteria. Ann. Rev. Microbiol. 31, 159–180. doi: 10.1146/annurev.mi.31.100177.001111
Goodfellow, M., and Pirouz, T. (1982). Numerical classification of Sporoactinomycetes containing meso-diaminopimelic acid in the cell wall. J. Gen. Microbiol. 128, 503–527.
Goodfellow, M., Stach, J. E., Brown, R., Venkata Bonda, A. N., Jones, A. L., Mexson, J., et al. (2012b). Verrucosispora maris sp. nov., a novel deep-sea actinomycete isolated from a marine sediment which produces abyssomicins. Antonie Van Leeuwenhoek 101, 185–193. doi: 10.1007/s10482-011-9651-5
Goodfellow, M., Stanton, L. J., Simpson, K. E., and Minnikin, D. E. (1990). Numerical and chemical classification of Actinoplanes and some related actinomycetes. J. Gen. Microbiol. 136, 19–36.
Goodfellow, M., Weaver, C. R., and Minnikin, D. E. (1982). Numerical classification of some Rhodococci, Corynebacteria and related organisms. J. Gen. Microbiol. 128, 731–745.
Goodfellow, M., Williams, S. T., and Alderson, G. (1986a). Transfer of Actinosporangium violaceum Krasil'nikov and Yuan, Actinosporangium vitaminophilum Shomura et al. and Actinopycnidium caeruleum Krasil'nikov to the genus Streptomyces, with amended descriptions of the species. Syst. Appl. Microbiol. 8, 61–64.
Goodfellow, M., Williams, S. T., and Alderson, G. (1986b). Transfer of Chainia species to the genus Streptomyces with emended description of species. Syst. Appl. Microbiol. 8, 55–60. doi: 10.1016/S0723-2020(86)80148-5
Goodfellow, M., Williams, S. T., and Alderson, G. (1986c). Transfer of Kitasatoa purpurea Matsumae and Hata to the genus Streptomyces as Streptomyces purpureus comb. nov. Syst. Appl. Microbiol. 8, 65–66.
Gordon, M. A., and Lapa, E. W. (1966). Durhamycin, a pentaene antifungal antibiotic from Streptomyces durhamensis sp. n. Appl. Microbiol. 14, 754–760.
Goyache, J., Ballesteros, C., Vela, A. I., Collins, M. D., Briones, V., Hutson, R. A., et al. (2003a). Corynebacterium sphenisci sp. nov., isolated from wild penguins. Int. J. Syst. Evol. Microbiol. 53, 1009–1012. doi: 10.1099/ijs.0.02502-0
Goyache, J., Vela, A. I., Collins, M. D., Ballesteros, C., Briones, V., Moreno, J., et al. (2003b). Corynebacterium spheniscorum sp. nov., isolated from the cloacae of wild penguins. Int. J. Syst. Evol. Microbiol. 53, 43–46. doi: 10.1099/ijs.0.02343-0
Greenwood, J. R., and Pickett, M. J. (1980). Transfer of Haemophilus vaginalis Gardner and Dukes to a new genus, Gardnerella: G. vaginalis (Gardner and Dukes) comb. nov. Int. J. Syst. Bacteriol. 30, 170–178.
Greiner-Mai, E., Korn-Wendisch, F., and Kutzner, H. J. (1988). Taxonomic revision of the genus Saccharomonospora and description of Saccharomonospora glauca sp. nov. Int. J. Syst. Bacteriol. 38, 398–405.
Greiner-Mai, E., Kroppenstedt, R., Korn-Wendisch, F., and Kutzner, H. J. (1987). Morphological and biochemical characterization and emended descriptions of thermophilic Actinomycetes species. Syst. Appl. Microbiol. 9, 97–109. doi: 10.1016/S0723-2020(87)80062-0
Groth, I., Schumann, P., Martin, K., Schuetze, B., Augsten, K., Kramer, I., et al. (1999a). Ornithinicoccus hortensis gen. nov., sp. nov., a soil actinomycete which contains L-ornithine. Int. J. Syst. Bacteriol. 49, 1717–1724.
Groth, I., Schumann, P., Rainey, F. A., Martin, K., Schuetze, B., and Augsten, K. (1997). Demetria terragena gen. nov., sp. nov., a new genus of Actinomycetes isolated from compost soil. Int. J. Syst. Bacteriol. 47, 1129–1133.
Groth, I., Schumann, P., Schuetze, B., Augsten, K., Kramer, I., and Stackebrandt, E. (1999b). Beutenbergia cavernae gen. nov., sp. nov., an L-lysine-containing actinomycete isolated from a cave. Int. J. Syst. Bacteriol. 49, 1733–1740.
Groth, I., Schumann, P., Schütze, B., Augsten, K., and Stackebrandt, E. (2002). Knoellia sinensis gen. nov., sp. nov. and Knoellia subterranea sp. nov., two novel actinobacteria isolated from a cave. Int. J. Syst. Evol. Microbiol. 52, 77–84.
Groth, I., Schumann, P., Weiss, N., Schuetze, B., Augsten, K., and Stackebrandt, E. (2001). Ornithinimicrobium humiphilum gen. nov., sp. nov., a novel soil actinomycete with L-ornithine in the peptidoglycan. Int. J. Syst. Evol. Microbiol. 51, 81–87. doi: 10.1099/00207713-51-1-81
Groth, I., Tan, G. Y. A., González, J. M., Laiz, L., Carlsohn, M. R., Schütze, B., et al. (2007). Amycolatopsis nigrescens sp. nov., an actinomycete isolated from a Roman catacomb. Int. J. Syst. Evol. Microbiol. 57, 513–519. doi: 10.1099/ijs.0.64602-0
Grund, E., and Kroppenstedt, R. M. (1990). Chemotaxonomy and numerical taxonomy of the genus Nocardiopsis Meyer 1976. Int. J. Syst. Bacteriol. 40, 5–11. doi: 10.1099/00207713-40-1-5
Grundy, W. E., Whitman, A. L., Rdzok, E. G., Rdzok, E. J., Hanes, M. E., and Sylvester, J. C. (1952). Actithiazic acid. I. Microbiological studies. Antibiot. Chemother. 2, 399–408.
Gu, Q., Luo, H., Zheng, W., Liu, Z., and Huang, Y. (2006). Pseudonocardia oroxyli sp. nov., a novel actinomycete isolated from surface-sterilized Oroxylum indicum root. Int. J. Syst. Evol. Microbiol. 56, 2193–2197. doi: 10.1099/ijs.0.64385-0
Gu, Q., Zheng, W., and Huang, Y. (2007). Glycomyces sambucus sp. nov., an endophytic actinomycete isolated from the stem of Sambucus adnata wall. Int. J. Syst. Evol. Microbiol. 57, 1995–1998. doi: 10.1099/ijs.0.65064-0
Guan, T.-W., Tang, S.-K., Wu, J.-Y., Zhi, X.-Y., Xu, L.-H., Zhang, L.-L., et al. (2009). Haloglycomyces albus gen. nov., sp. nov., a halophilic, filamentous actinomycete of the family Glycomycetaceae. Int. J. Syst. Evol. Microbiol. 59, 1297–1301. doi: 10.1099/ijs.0.006270-0
Guan, T.-W., Xia, Z.-F., Tang, S.-K., Wu, N., Chen, Z.-J., Huang, Y., et al. (2012). Amycolatopsis salitolerans sp. nov., a filamentous actinomycete isolated from a hypersaline habitat. Int. J. Syst. Evol. Microbiol. 62, 23–27. doi: 10.1099/ijs.0.030031-0
Guo, L., Tuo, L., Habden, X., Zhang, Y., Liu, J., Jiang, Z., et al. (2015). Allosalinactinospora lopnorensis gen. nov., sp. nov., a new member of the family Nocardiopsaceae isolated from soil. Int. J. Syst. Evol. Microbiol. 65, 206–213. doi: 10.1099/ijs.0.055160-0
Gupta, R. S., Chen, W. J., Adeolu, M., and Chai, Y. (2013). Molecular signatures for the class Coriobacteriia and its different clades; proposal for division of the class Coriobacteriia into the emended order Coriobacteriales, containing the emended family Coriobacteriaceae and Atopobiaceae fam. nov., and Eggerthellales ord. nov., containing the family Eggerthellaceae fam. nov. Int. J. Syst. Evol. Microbiol. 63, 3379–3397. doi: 10.1099/ijs.0.048371-0
Gupta, R. S., Lo, B., and Son, J. (2018). Phylogenomics and comparative genomic studies robustly support division of the genus Mycobacterium into an emended genus Mycobacterium and four novel genera. Front. Microbiol. 9:67. doi: 10.3389/fmicb.2018.00067
Gürtler, V., Smith, R., Mayall, B. C., Pötter-Reinemann, G., Stackbrandt, E., and Kroppenstedt, R. M. (2001). Nocardia veterana sp. nov., isolated from human bronchial lavage. Int. J. Syst. Evol. Microbiol. 51, 933–936.
Haas, W. H., Butler, W. R., Kirschner, P., Plikaytis, B. B., Coyle, M. B., Amthor, B., et al. (1997). A new agent of mycobacterial lymphadenitis in children: Mycobacterium heidelbergense sp. nov. J. Clin. Microbiol. 35, 3203–3209.
Hägglblom, M. M., Nohynek, L. J., Palleroini, N. J., Kronqvist, K., Nurmiaho-Lassila, E.-L., Salkinoja-Salonen, M. S., et al. (1994). Transfer of polychlorophenol-degrading Rhodococcus chlorophenolicus (Apajalahti et al. 1986) to the genus Mycobacterium as Mycobacterium chlorophenolicum comb. nov. Int. J. Syst. Bacteriol. 44, 485–493.
Hahnke, R. L., Meier-Kolthoff, J. P., García-López, M., Mukherjee, S., Huntemann, M., Ivanova, N. N., et al. (2016). Genome-based taxonomic classification of Bacteroidetes. Front. Microbiol. 7:2003. doi: 10.3389/fmicb.2016.02003
Hall, V., Collins, M. D., Hutson, R., Falsen, E., and Duerden, B. I. (2002). Actinomyces cardiffensis sp. nov. from human clinical sources. J. Clin. Microbiol. 40, 3427–3431. doi: 10.1128/JCM.40.9.3427-3431.2002
Hall, V., Collins, M. D., Hutson, R., Inganäs, E., Falsen, E., and Duerden, B. I. (2003b). Actinomyces vaccimaxillae sp. nov., from the jaw of a cow. Int. J. Syst. Evol. Microbiol. 53, 603–606. doi: 10.1099/ijs.0.02439-0
Hall, V., Collins, M. D., Hutson, R. A., Lawson, P. A., Falsen, E., and Duerden, B. I. (2003a). Corynebacterium atypicum sp. nov., from a human clinical source, does not contain corynomycolic acids. Int. J. Syst. Evol. Microbiol. 53, 1065–1068. doi: 10.1099/ijs.0.02442-0
Hall, V., Collins, M. D., Lawson, P. A., Falsen, E., and Duerden, B. I. (2003c). Actinomyces nasicola sp. nov., isolated from a human nose. Int. J. Syst. Evol. Microbiol. 53, 1445–1448. doi: 10.1099/ijs.0.02582-0
Hall, V., Collins, M. D., Lawson, P. A., Falsen, E., and Duerden, B. I. (2005). Actinomyces dentalis sp. nov., from a human dental abscess. Int. J. Syst. Evol. Microbiol. 55, 427–431. doi: 10.1099/ijs.0.63376-0
Hall, V., Collins, M. D., Lawson, P. A., Hutson, R. A., Falsen, E., Inganas, E., et al. (2003d). Characterization of some Actinomyces-like isolates from human clinical sources: description of Varibaculum cambriensis gen. nov., sp. nov. J. Clin. Microbiol. 41, 640–644. doi: 10.1128/JCM.41.2.640
Halpern, M., Shakéd, T., Pukall, R., and Schumann, P. (2009). Leucobacter chironomi sp. nov., a chromate-resistant bacterium isolated from a chironomid egg mass. Int. J. Syst. Evol. Microbiol. 59, 665–670. doi: 10.1099/ijs.0.004663-0
Hamada, M., Otoguro, M., Yamamura, H., Tamura, T., Suzuki, K., and Hayakawa, M. (2010). Luteimicrobium subarcticum gen. nov., sp. nov., a new member of the suborder Micrococcineae. Int. J. Syst. Evol. Microbiol. 60, 796–800. doi: 10.1099/ijs.0.014597-0
Hamada, M., Shibata, C., Saitou, S., Tamura, T., Komaki, H., Ichikawa, N., et al. (2015). Proposal of nine novel species of the genus Lysinimicrobium and emended description of the genus Lysinimicrobium. Int. J. Syst. Evol. Microbiol. 65, 4394–4402. doi: 10.1099/ijsem.0.000587
Hamada, M., Shibata, C., Sakurai, K., Hosoyama, A., Oji, S., Teramoto, K., et al. (2016). Reclassification of Amycolicicoccus subflavus as Hoyosella subflava comb. nov. and emended descriptions of the genus Hoyosella and of Hoyosella altamirensis. Int. J. Syst. Evol. Microbiol. 66, 4711–4715. doi: 10.1099/ijs.0.001415
Hamada, M., Tamura, T., Yamamura, H., Suzuki, K., and Hayakawa, M. (2012). Lysinimicrobium mangrovi gen. nov., sp. nov., an actinobacterium isolated from the rhizosphere of a mangrove. Int. J. Syst. Evol. Microbiol. 62, 1731–1735. doi: 10.1099/ijs.0.035493-0
Hamada, M., Tamura, T., Yamamura, H., Suzuki, K., and Hayakawa, M. (2013). Demequina flava sp. nov. and Demequina sediminicola sp. nov., isolated from sea sediment. Int. J. Syst. Evol. Microbiol. 63, 249–253. doi: 10.1099/ijs.0.039297-0
Hamada, S. (1958). A study of a new antitumor substance, cellostatin. Tohoku J. Exp. Med. 67, 173–179.
Hamid, M. E., Maldonado, L., Eldin, G. S. S., Mohamed, M. F., Saeed, N. S., and Goodfellow, M. (2001). Nocardia africana sp. nov., a new pathogen isolated from patients with pulmonary infections. J. Clin. Microbiol. 39, 625–630. doi: 10.1128/JCM.39.2.625
Hammann, R., Kronibus, A., Viebahn, A., and Brandis, H. (1984). Falcivibrio grandis gen. nov. sp. nov., and Falcivibrio vaginalis gen. nov. sp. nov., a new genus and species to accommodate anaerobic motile curved rods formerly described as “Vibrio mulieris” (Prévot 1940) Breed et at. 1948. Syst. Appl. Microbiol. 5, 81–96. doi: 10.1016/S0723-2020(84)80053-3
Hanada, S., Liu, W.-T., Shintani, T., Kamagata, Y., and Nakamura, K. (2002a). Tetrasphaera elongata sp. nov., a polyphosphate-accumulating bacterium isolated from activated sludge. Int. J. Syst. Evol. Microbiol. 52, 883–887. doi: 10.1099/ijs.0.01990-0
Hanada, S., Takaichi, S., Matsuura, K., and Nakamura, K. (2002b). Roseiflexus castenholzii gen. nov., sp. nov., a thermophilic, filamentous, photosynthetic bacterium that lacks chlorosomes. Int. J. Syst. Evol. Microbiol. 52, 187–193.
Hannigan, G. D., Krivogorsky, B., Fordice, D., Welch, J. B., and Dahl, J. L. (2013). Mycobacterium minnesotense sp. nov., a photochromogenic bacterium isolated from sphagnum peat bogs. Int. J. Syst. Evol. Microbiol. 63, 124–128. doi: 10.1099/ijs.0.037291-0
Harz, C. O. (1877). Actinomyces bovis ein Neuer Schimmel in den Geweben des Rindes. Dtsch. Zeitschrift für Thiermedizin. 5, 125–140.
Hasegawa, T. (1988). Actinokineospora: a new genus of the Actinomycetales. Actinomycetologica 2, 31–45.
Hasegawa, T., Lechevalier, M. P., and Lechevalier, H. A. (1978). New genus of the Actinomycetales: Actinosynnema gen. nov. Int. J. Syst. Bacteriol. 28, 304–310.
Hasegawa, T., Tanida, S., Hatano, K., Higashide, E., and Yoneda, M. (1983). Motile Actinomycetes: Actinosynnema pretiosum subsp. pretiosum sp. nov., subsp. nov., and Actinosynnema subsp. auranticum subsp. nov. Int. J. Syst. Bacteriol. 33, 314–320.
Heinemann, B., Kaplan, M. A., Muir, R. D., and Hooper, I. R. (1953). Amphomycin, a new antibiotic. Antibiot. Chemother. 3, 1239–1242.
Hennessee, C. T., Seo, J.-S., Alvarez, A. M., and Li, Q. X. (2009). Polycyclic aromatic hydrocarbon-degrading species isolated from Hawaiian soils: Mycobacterium crocinum sp. nov., Mycobacterium pallens sp. nov., Mycobacterium rutilum sp. nov., Mycobacterium rufum sp. nov. and Mycobacterium aromaticivorans sp. nov. Int. J. Syst. Evol. Microbiol. 59, 378–387. doi: 10.1099/ijs.0.65827-0
Henssen, A. (1957). Beiträge zur Morphologie und Systematik der thermophilen Actinomyceten. Arch. Mikrobiol. 26, 373–414. doi: 10.1007/BF00407588
Henssen, A., Kothe, H. W., and Kroppenstedt, R. M. (1987). Transfer of Pseudonocardia azurea and “Pseudonocardia fastidiosa” to the genus Amycolatopsis, with emended species description. Int. J. Syst. Bacteriol. 37, 292–295.
Henssge, U., Do, T., Radford, D. R., Gilbert, S. C., Clark, D., and Beighton, D. (2009). Emended description of Actinomyces naeslundii and descriptions of Actinomyces oris sp. nov. and Actinomyces johnsonii sp. nov., previously identified as Actinomyces naeslundii genospecies 1, 2 WVA 963. Int. J. Syst. Evol. Microbiol. 59, 509–516. doi: 10.1099/ijs.0.000950-0
Heyrman, J., Verbeeren, J., Schumann, P., Swings, J., and De Vos, P. (2005). Six novel Arthrobacter species isolated from deteriorated mural paintings. Int. J. Syst. Evol. Microbiol. 55, 1457–1464. doi: 10.1099/ijs.0.63358-0
Higgens, C. E., and Kastner, R. E. (1971). Streptomyces clavuligerus sp. nov., a ß-lactam antibiotic producer. Int. J. Syst. Bacteriol. 21, 326–331.
Hildebrand, F., Meyer, A., and Eyre-Walker, A. (2010). Evidence of selection upon genomic GC-content in bacteria. PLoS Genet. 6:e1001107. doi: 10.1371/journal.pgen.1001107
Hirsch, P., Mevs, U., Kroppenstedt, R. M., Schumann, P., and Stackebrandt, E. (2004). Cryptoendolitihic Actinomycetes from Antarctic sandstone rock samples: Micromonospora endolithica sp. nov. and two isolates related to Micromonospora coerulea Jensen 1932. Syst. Appl. Microbiol. 27, 166–174.
Holt, J. G., and Lewin, R. A. (1968). Herpetosiphon aurantiacus gen. et sp. n., a new filamentous gliding organism. J. Bacteriol. 95, 2407–2408.
Horan, A. C., and Brodsky, B. C. (1982). A novel antibiotic-producing Actinomadura, Actinomadura kijaniata sp. nov. Int. J. Syst. Bacteriol. 32, 195–200.
Horan, A. C., and Brodsky, B. C. (1986). Micromonospora rosaria sp. nov., nom. rev., the rosaramicin producer. Int. J. Syst. Bacteriol. 36, 478–480.
Houghton, K. M., Morgan, X. C., Lagutin, K., MacKenzie, D., Vyssotskii, M., Mitchell, K. A., et al. (2015). Thermorudis pharmacophila sp. nov., a novel member of the class Thermomicrobia isolated from geothermal soil, and emended descriptions of Thermomicrobium roseum, Thermomicrobium carboxidum, Thermorudis peleae and Sphaerobacter thermophilus. Int. J. Syst. Evol. Microbiol. 65, 4479–4487. doi: 10.1099/ijsem.0.000598
Hoyles, L., Collins, M. D., Falsen, E., Nikolaitchouk, N., and McCartney, A. L. (2004). Transfer of members of the genus Falcivibrio to the genus Mobiluncus, and emended description of the genus Mobiluncus. Syst. Appl. Microbiol. 27, 72–83.
Hoyles, L., Falsen, E., Foster, G., and Collins, M. D. (2002a). Actinomyces coleocanis sp. nov., from the vagina of a dog. Int. J. Syst. Evol. Microbiol. 52, 1201–1203. doi: 10.1099/ijs.0.01934-0
Hoyles, L., Falsen, E., Foster, G., Pascual, C., Greko, C., and Collins, M. D. (2000). Actinomyces canis sp. nov., isolated from dogs. Int. J. Syst. Evol. Microbiol. 50, 1547–1551. doi: 10.1099/00207713-50-4-1547
Hoyles, L., Falsen, E., Holmström, G., Persson, A., Sjödén, B., and Collins, M. D. (2001a). Actinomyces suimastitidis sp. nov., isolated from pig mastitis. Int. J. Syst. Evol. Microbiol. 51, 1323–1326. doi: 10.1099/00207713-51-4-1323
Hoyles, L., Inganäs, E., Falsen, E., Drancourt, M., Weiss, N., McCartney, A. L., et al. (2002b). Bifidobacterium scardovii sp. nov., from human sources. Int. J. Syst. Evol. Microbiol. 52, 995–999. doi: 10.1099/ijs.0.02065-0
Hoyles, L., Ortman, K., Cardew, S., Foster, G., Rogerson, F., and Falsen, E. (2013). Corynebacterium uterequi sp. nov., a non-lipophilic bacterium isolated from urogenital samples from horses. Vet. Microbiol. 165, 469–474. doi: 10.1016/j.vetmic.2013.03.025
Hoyles, L., Pascual, C., Falsen, E., Foster, G., Grainger, J. M., and Collins, M. D. (2001b). Actinomyces marimammalium sp. nov., from marine mammals. Int. J. Syst. Evol. Microbiol. 51, 151–156. doi: 10.1099/00207713-51-1-151
Hozzein, W. N., Li, W.-J., Ali, M. I. A., Hammouda, O., Mousa, A. S., Xu, L.-H., et al. (2004). Nocardiopsis alkaliphila sp. nov., a novel alkaliphilic actinomycete isolated from desert soil in Egypt. Int. J. Syst. Evol. Microbiol. 54, 247–252. doi: 10.1099/ijs.0.02832-0
Hu, H., Lin, H.-P., Xie, Q., Li, L., Xie, X.-Q., and Hong, K. (2012). Streptomyces qinglanensis sp. nov., isolated from mangrove sediment. Int. J. Syst. Evol. Microbiol. 62, 596–600. doi: 10.1099/ijs.0.032201-0
Huang, L. H. (1980). Actinomadura macra sp. nov., the producer of antibiotics CP-47,433 and CP-47,434. Int. J. Syst. Bacteriol. 30, 565–568.
Huang, M.-J., Narsing Rao, P. M., Salam, N., Xiao, M., Huang, H.-Q., and Li, W.-J. (2017). Allostreptomyces psammosilenae gen. nov. sp. nov. an endophytic actinobacterium isolated from the roots of Psammosilene tunicoides and emended description of the family Streptomycetaceae [Waksman and Henrici (1943) AL] emend. Rainey et al. 1997, em. Int. J. Syst. Evol. Microbiol. 67, 288–293. doi: 10.1099/ijsem.0.001617
Huang, Y., Dai, X., He, L., Wang, Y.-N., Wang, B.-J., Liu, Z., et al. (2005). Sanguibacter marinus sp. nov., isolated from coastal sediment. Int. J. Syst. Evol. Microbiol. 55, 1755–1758. doi: 10.1099/ijs.0.63471-0
Huang, Y., Qi, W., Lu, Z., Liu, Z., and Goodfellow, M. (2001). Amycolatopsis rubida sp. nov., a new Amycolatopsis species from soil. Int. J. Syst. Evol. Microbiol. 51, 1093–1097. doi: 10.1099/00207713-51-3-1093
Huys, G., Vancanneyt, M., D'Haene, K., Falsen, E., Wauters, G., and Vandamme, P. (2007). Alloscardovia omnicolens gen. nov., sp. nov., from human clinical samples. Int. J. Syst. Evol. Microbiol. 57, 1442–1446. doi: 10.1099/ijs.0.64812-0
Idris, H., Nouioui, I., Asenjo, J. A., Bull, A. T., and Goodfellow, M. (2017). Lentzea chajnantorensis sp. nov., an actinobacterium from a very high altitude Cerro Chajnantor gravel soil in northern Chile. Antonie Van Leeuwenhoek 110, 795–802. doi: 10.1007/s10482-017-0851-5
Iinuma, S., Yokota, A., Hasegawa, T., and Kanamaru, T. (1994). Actinocorallia gen. nov., a new genus of the order Actinomycetales. Int. J. Syst. Bacteriol. 44, 230–234. doi: 10.1099/00207713-44-2-230
Ishikawa, T., and Yokota, A. (2006). Reclassification of Arthrobacter duodecadis Lochhead 1958 as Tetrasphaera duodecadis comb. nov. and emended description of the genus Tetrasphraera. Int. J. Syst. Evol. Microbiol. 56, 1369–1373. doi: 10.1099/ijs.0.02984-0
Isik, K., Chun, J., Hah, Y. C., and Goodfellow, M. (1999). Nocardia uniformis nom. rev. Int. J. Syst. Bacteriol. 49, 1227–1230.
Iwai, K., Aisaka, K., and Suzuki, M. (2010). Friedmanniella luteola sp. nov., Friedmanniella lucida sp. nov., Friedmanniella okinawensis sp. nov. and Friedmaniella sagamiharensis sp. nov., isolated from spiders. Int. J. Syst. Evol. Microbiol. 60, 113–120. doi: 10.1099/ijs.0.007815-0
Iwasaki, A., Itoh, H., and Mori, T. (1979). A new broad-spectrum aminoglycoside antibiotic complex, sporaricin. II. Taxonomic studies on the sporaricin producing strain Saccharopolyspora hirsuta subsp. kobensis nov. subsp. J. Antibiot. (Tokyo). 32, 180–186.
Jackman, P. J. H., Pitcher, D. G., Pelczynska, S., and Borman, P. (1987). Classification of corynebacteria associated with endocarditis (Group JK) as Corynebacterium jeikeium sp. nov. Syst. Appl. Microbiol. 9, 83–90. doi: 10.1016/S0723-2020(87)80060-7
Jang, Y.-H., Kim, S.-J., Hamada, M., Tamura, T., Ahn, J.-H., Weon, H.-Y., et al. (2012). Diaminobutyricimonas aerilata gen. nov., sp. nov., a novel nember of the family Microbacteriaceae isolated from an air sample in Korea. J. Microbiol. 50, 1047–1052. doi: 10.1007/s12275-012-2118-1
Jeffroy, O., Brinkmann, H., Delsuc, F., and Philippe, H. (2006). Phylogenomics: the beginning of incongruence? Trends Genet. 22, 225–231. doi: 10.1016/j.tig.2006.02.003
Jian, W., and Dong, X. (2002). Transfer of Bifidobacterium inopinatum and Bifidobacterium denticolens to Scardovia inopinata gen. nov., comb. nov., and Parascardovia denticolens gen. nov., comb. nov., respectively. Int. J. Syst. Evol. Microbiol. 52, 809–812. doi: 10.1099/ijs.0.02054-0
Jiang, Y., Wiese, J., Tang, S.-K., Xu, L.-H., Imhoff, J. F., and Jiang, C.-L. (2008). Actinomycetospora chiangmaiensis gen. nov., a new member of the family Pseudonocardiaceae. Int. J. Syst. Evol. Microbiol. 58, 408–413. doi: 10.1099/ijs.0.64976-0
Jiao, J.-Y., Liu, L., Zhou, E.-M., Wei, D.-Q., Ming, H., Xian, W.-D., et al. (2015). Actinomadura amylolytica sp. nov. and Actinomadura cellulosilytica sp. nov., isolated from geothermally heated soil. Antonie Van Leeuwenhoek 108, 75–83. doi: 10.1007/s10482-015-0465-8
Jin, X., Xu, L.-H., Mao, P.-H., Hseu, T.-H., and Jiang, C.-L. (1998). Description of Saccharomonospora xinjiangensis sp. nov. based on chemical and molecular classification. Int. J. Syst. Bacteriol. 48, 1095–1099.
Johnson, D. B., Bacelar-Nicolau, P., Okibe, N., Thomas, A., and Hallberg, K. B. (2009). Ferrithrix thermotolerans gen. nov., sp. nov.: heterotrophic, iron-oxidizing, extremely acidophilic actinobacteria. Int. J. Syst. Evol. Microbiol. 59, 1082–1089. doi: 10.1099/ijs.0.65409-0
Johnson, J. L., and Cummins, C. S. (1972). Cell wall composition and deoxyribonucleic acid similarities among the anaerobic coryneforms, classical propionibacteria, and strains of Arachnia propionica. J. Bacteriol. 109, 1047–1066.
Johnson, J. L., Moore, L. V. H., Kaneko, B., and Moore, W. E. C. (1990). Actinomyces georgiae sp. nov., Actinomyces gerencseriae sp. nov., designation of two genospecies of Actinomyces naeslundii, and inclusion of A. naeslundii serotypes II and III and Actinomyces viscosus serotype II in A. naeslundii genospecies 2. Int. J. Syst. Bacteriol. 40, 273–286.
Johstone, D. B., and Waksman, S. A. (1947). Streptomycin II, an antibiotic substance produced by a new species of Streptomyces. Proc. Soc. Exp. Biol. Med. 65, 294–295.
Jones, A. L., Brown, J. M., Mishra, V., Perry, J. D., Steigerwalt, A. G., and Goodfellow, M. (2004). Rhodococcus gordoniae sp. nov., an actinomycete isolated from clinical material and phenol-contaminated soil. Int. J. Syst. Evol. Microbiol. 54, 407–411. doi: 10.1099/ijs.0.02756-0
Jones, A. L., and Goodfellow, M. (2012). Genus IV. Rhodococcus (Zopf 1891) emend. Goodfellow, Alderson and Chun 1998a, in Bergey's Manual of Systematic Bacteriology, Vol. 5, The Actinobacteria, eds M. Goodfellow, P. Kämpfer, H.-J. Busse, M. E. Trujillo, K. Suzuki, W. Ludwig, et al. (New York, NY: Springer), 437–464.
Jones, D., and Collins, M. D. (1988). Taxonomic studies on some human cutaneous coryneform bacteria: description of Dermabacter hominis gen. nov., sp. nov. FEMS Microbiol. Lett. 51, 51–56.
Juboi, H., Basik, A. A., Gill Shamsul, S. S., Arnold, P., Schmitt, E. K., Sanglier, J.-J., et al. (2015). Luteipulveratus halotolerans sp. nov., an actinobacterium (Dermacoccaceae) from forest soil. Int. J. Syst. Evol. Microbiol. 65, 4113–4120. doi: 10.1099/ijsem.0.000548
Jung, S.-Y., Kim, H.-S., Song, J. J., Lee, S.-G., Oh, T.-K., and Yoon, J.-H. (2006). Kribbia dieselivorans gen. nov., sp. nov., a novel member of the family Intrasporangiaceae. Int. J. Syst. Evol. Microbiol. 56, 2427–2432. doi: 10.1099/ijs.0.64459-0
Jurado, V., Groth, I., Gonzalez, J. M., Laiz, L., and Saiz-Jimenez, C. (2005). Agromyces subbeticus sp. nov., isolated from a cave in southern Spain. Int. J. Syst. Evol. Microbiol. 55, 1897–1901. doi: 10.1099/ijs.0.63637-0
Jurado, V., Kroppenstedt, R. M., Saiz-Jimenez, C., Klenk, H.-P., Mouniée, D., Laiz, L., et al. (2009). Hoyosella altamirensis gen. nov., sp. nov., a new member of the order Actinomycetales isolated from a cave biofilm. Int. J. Syst. Evol. Microbiol. 59, 3105–3110. doi: 10.1099/ijs.0.008664-0
Kaewkla, O., and Franco, C. M. M. (2011). Flindersiella endophytica gen. nov., sp. nov., an endophytic actinobacterium isolated from the root of Grey Box, an endemic eucalyptus tree. Int. J. Syst. Evol. Microbiol. 61, 2135–2140. doi: 10.1099/ijs.0.026757-0
Kageyama, A., and Benno, Y. (2000). Emendation of genus Collinsella and proposal of Collinsella stercoris sp. nov. and Collinsella intestinalis sp. nov. Int. J. Syst. Evol. Microbiol. 50, 1767–1774.
Kageyama, A., Benno, Y., and Nakase, T. (1999). Phylogenic and phenotypic evidence for the transfer of Eubacterium fossor to the genus Atopobium as Atopobium fossor comb. nov. Microbiol. Immunol. 43, 389–395.
Kageyama, A., Iida, S., Yazawa, K., Kudo, T., Suzuki, S., Koga, T., et al. (2006). Gordonia araii sp. nov., and Gordonia effusa sp. nov., isolated from patients in Japan. Int. J. Syst. Evol. Microbiol. 56, 1817–1821. doi: 10.1099/ijs.0.64067-0
Kageyama, A., Poonwan, N., Yazawa, K., Mikami, Y., and Nishimura, K. (2004a). Nocardia asiatica sp. nov., isolated from patients with nocardiosis in Japan and clinical specimens from Thailand. Int. J. Syst. Evol. Microbiol. 54, 125–130. doi: 10.1099/ijs.0.02676-0
Kageyama, A., Takahashi, Y., Seki, T., Tomoda, H., and Omura, S. (2005). Oryzihumus leptocrescens gen. nov., sp. nov. Int. J. Syst. Evol. Microbiol. 55, 2555–2559. doi: 10.1099/ijs.0.63799-0
Kageyama, A., Yazawa, K., Mukai, A., Kinoshita, M., Takata, N., Nishimura, K., et al. (2004b). Nocardia shimofusensis sp. nov., isolated from soil, and Nocardia higoensis sp. nov., isolated from a patient with lung nocardiosis in Japan. Int. J. Syst. Evol. Microbiol. 54, 1927–1931. doi: 10.1099/ijs.0.63061-0
Kageyama, A., Yazawa, K., Nishimura, K., and Mikami, Y. (2004c). Nocardia inohanensis sp. nov., Nocardia yamanashiensis sp. nov. and Nocardia niigatensis sp. nov., isolated from clinical specimens. Int. J. Syst. Evol. Microbiol. 54, 563–569. doi: 10.1099/ijs.0.02794-0
Kalakoutskii, L. V., Kirillova, I. P., and Krassilnikov, N. A. (1967). A new genus of the Actinomycetales-Intrasporangium gen. nov. J. Gen. Microbiol. 48, 79–85.
Kämpfer, P. (2012a). Genus I. Streptomyces Waksman and Henrici 1943, 339 AL emend. Witt and Stackebrandt 1990, 370 emend. Wellington, Stackebrandt, Sanders, Wolstrup and Jorgensen 1992, 159, in Bergey's Manual of Systematic Bacteriology, Vol. 5, The Actinobacteria, eds M. Goodfellow, P. Kämpfer, H.-J. Busse, M. E. Trujillo, K. Suzuki, W. Ludwig, et al. (New York, NY: Springer), 1455–1767.
Kämpfer, P. (2012b). Order IX. Kineosporiales ord. nov., in Bergey's Manual of Systematic Bacteriology, Vol. 5, The Actinobacteria, eds M. Goodfellow, P. Kämpfer, H.-J. Busse, M. E. Trujillo, K. Suzuki, W. Ludwig, et al. (New York, NY: Springer), 561.
Kämpfer, P., Andersson, M. A., Rainey, F. A., Kroppenstedt, R. M., and Salkinoja-Salonen, M. (1999). Williamsia muralis gen. nov., sp. nov., isolated from the indoor environment of a children's day care centre. Int. J. Syst. Bacteriol. 49, 681–687.
Kämpfer, P., Arun, A. B., Busse, H.-J., Langer, S., Young, C.-C., Chen, W.-M., et al. (2010a). Georgenia soli sp. nov., isolated from iron-ore-contaminated soil in India. Int. J. Syst. Evol. Microbiol. 60, 1027–1030. doi: 10.1099/ijs.0.014902-0
Kämpfer, P., Dott, W., Martin, K., and Glaeser, S. P. (2014a). Rhodococcus defluvii sp. nov., isolated from wastewater of a bioreactor and formal proposal to reclassify [Corynebacterium hoagii] and Rhodococcus equi as Rhodococcus hoagii comb. nov. Int. J. Syst. Evol. Microbiol. 64, 755–761. doi: 10.1099/ijs.0.053322-0
Kämpfer, P., and Glaeser, S. P. (2012). Prokaryotic taxonomy in the sequencing era – the polyphasic approach revisited. Environ. Microbiol. 14, 291–317. doi: 10.1111/j.1462-2920.2011.02615.x
Kämpfer, P., Glaeser, S. P., Busse, H.-J., Abdelmohsen, U. R., and Hentschel, U. (2014b). Rubrobacter aplysinae sp. nov., isolated from the marine sponge Aplysina aerophoba. Int. J. Syst. Evol. Microbiol. 64, 705–709. doi: 10.1099/ijs.0.055152-0
Kämpfer, P., Glaeser, S. P., Busse, H.-J., Abdelmohsen, U. R., Safwat, A., and Hentschel, U. (2015). Actinokineospora spheciospongiae sp. nov., isolated from the marine sponge Spheciospongia vagabunda. Int. J. Syst. Evol. Microbiol. 65, 879–884. doi: 10.1099/ijs.0.000031
Kämpfer, P., Huber, B., Buczolits, S., Thummes, K., Grün-Wollny, I., and Busse, H.-J. (2008). Streptomyces specialis sp. nov. Int. J. Syst. Evol. Microbiol. 58, 2602–2606. doi: 10.1099/ijs.0.2008/001008-0
Kämpfer, P., Kroppenstedt, R. M., and Grün-Wollny, I. (2005). Nonomuraea kuesteri sp. nov. Int. J. Syst. Evol. Microbiol. 55, 847–851. doi: 10.1099/ijs.0.63380-0
Kämpfer, P., Lodders, N., Warfolomeow, I., Falsen, E., and Busse, H.-J. (2009a). Corynebacterium lubricantis sp. nov., isolated from a coolant lubricant. Int. J. Syst. Evol. Microbiol. 59, 1112–1115. doi: 10.1099/ijs.0.006379-0
Kämpfer, P., Martin, K., Schäfer, J., and Schumann, P. (2009b). Kytococcus aerolatus sp. nov., isolated from indoor air in a room colonized with moulds. Syst. Appl. Microbiol. 32, 301–305. doi: 10.1016/j.syapm.2009.05.004
Kämpfer, P., Schäfer, J., Lodders, N., and Busse, H.-J. (2010b). Brevibacterium sandarakinum sp. nov., isolated from a wall of an indoor environment. Int. J. Syst. Evol. Microbiol. 60, 909–913. doi: 10.1099/ijs.0.014100-0
Kämpfer, P., Schäfer, J., Lodders, N., and Martin, K. (2010c). Murinocardiopsis flavida gen. nov., sp. nov., an actinomycete isolated from indoor walls. Int. J. Syst. Evol. Microbiol. 60, 1729–1734. doi: 10.1099/ijs.0.015990-0
Kämpfer, P., Schäfer, J., Lodders, N., and Martin, K. (2011). Jiangella muralis sp. nov., from an indoor environment. Int. J. Syst. Evol. Microbiol. 61, 128–131. doi: 10.1099/ijs.0.022277-0
Kämpfer, P., Young, C.-C., Busse, H.-J., Chu, J.-N., Schumann, P., Arun, A. B., et al. (2010d). Microlunatus soli sp. nov., isolated from soil. Int. J. Syst. Evol. Microbiol. 60, 824–827. doi: 10.1099/ijs.0.013540-0
Karassova, V., Weissfeiler, J., and Krasznay, E. (1965). Occurrence of atypical mycobacteria in Macacus rhesus. Acta Microbiol. Acad. Sci. Hung. 12, 275–282.
Kasai, H., Tamura, T., and Harayama, S. (2000). Intrageneric relationships among Micromonospora species deduced from gyrB-based phylogeny and DNA relatedness. Int. J. Syst. Evol. Microbiol. 50, 127–134.
Katayama, T., Kato, T., Tanaka, M., Douglas, T. A., Brouchkov, A., Abe, A., et al. (2010). Tomitella biformata gen. nov., sp. nov., a new member of the suborder Corynebacterineae isolated from a permafrost ice wedge. Int. J. Syst. Evol. Microbiol. 60, 2803–2807. doi: 10.1099/ijs.0.017962-0
Katayama, T., Kato, T., Tanaka, M., Douglas, T. A., Brouchkov, A., Fukuda, M., et al. (2009). Glaciibacter superstes gen. nov., sp. nov., a novel member of the family Microbacteriaceae isolated from a permafrost ice wedge. Int. J. Syst. Evol. Microbiol. 59, 482–486. doi: 10.1099/ijs.0.001354-0
Katoh, K., Kuma, K., Miyata, T., and Toh, H. (2005). Improvement in the accuracy of multiple sequence alignment program MAFFT. Genome Inf. 16, 22–33.
Katz, L., and Baltz, R. H. (2016). Natural product discovery: past, present, and future. J. Ind. Microbiol. Biotechnol. 43, 155–176. doi: 10.1007/s10295-015-1723-5
Kawaichi, S., Ito, N., Kamikawa, R., Sugawara, T., Yoshida, T., and Sako, Y. (2013). Ardenticatena maritima gen. nov., sp. nov., a ferric iron- and nitrate-reducing bacterium of the phylum ‘Chloroflexi’ isolated from an iron-rich coastal hydrothermal field, and description of Ardenticatenia classis nov. Int. J. Syst. Evol. Microbiol. 63, 2992–3002. doi: 10.1099/ijs.0.046532-0
Kawamoto, I., Okachi, R., Kato, H., Yamamoto, S., Takahashi, I., Takasawa, S., et al. (1974). The antibiotic XK-41 complex. I. production, isolation and characterization. J. Antibiot. (Tokyo). 27, 493–501.
Kawato, N., and Shinobu, R. (1959). On Streptomyces herbaricolor, nov. sp. Supplement: a simple technique for the microscopic observation. Nat. Sci. 8, 114–119.
Kazda, J., Cooney, R., Monaghan, M., Quinn, P. J., Stackebrandt, E., Dorsch, M., et al. (1993). Mycobacterium hiberniae sp. nov. Int. J. Syst. Bacteriol. 43, 352–357.
Kelly, J., Kutscher, A. H., and Tuoti, F. (1959). Thiostrepton, a new antibiotic: tube dilution sensitivity studies. Oral Surg. Oral Med. Oral Pathol. 12, 1334–1339.
Kembel, S. W., Cowan, P. D., Helmus, M. R., Cornwell, W. K., Morlon, H., Ackerly, D. D., et al. (2010). Picante: R tools for integrating phylogenies and ecology. Bioinformatics 26, 1463–1464. doi: 10.1093/bioinformatics/btq166
Khan, A. A., Kim, S.-J., Paine, D. D., and Cerniglia, C. E. (2002). Classification of a polycyclic aromatic hydrocarbon-metabolizing bacterium, Mycobacterium sp. strain PYR-1, as Mycobacterium vanbaalenii sp. nov. Int. J. Syst. Evol. Microbiol. 52, 1997–2002. doi: 10.1099/ijs.0.02163-0
Khan, S. T., Harayama, S., Tamura, T., Ando, K., Takagi, M., and Kazuo, S. (2009). Paraoerskovia marina gen. nov., sp. nov., an actinobacterium isolated from marine sediment. Int. J. Syst. Evol. Microbiol. 59, 2094–2098. doi: 10.1099/ijs.0.007666-0
Killer, J., Havlik, J., Bunešová, V., Vlková, E., and Benada, O. (2014). Pseudoscardovia radai sp. nov., a representative of the family Bifidobacteriaceae isolated from the digestive tract of a wild pig (Sus scrofa scrofa). Int. J. Syst. Evol. Microbiol. 64, 2932–2938. doi: 10.1099/ijs.0.063230-0
Killer, J., Kopecný, J., Mrázek, J., Koppová, I., Havlík, J., Benada, O., et al. (2011). Bifidobacterium actinocoloniiforme sp. nov. and Bifidobacterium bohemicum sp. nov., from the bumblebee digestive tract. Int. J. Syst. Evol. Microbiol. 61, 1315–1321. doi: 10.1099/ijs.0.022525-0
Killer, J., Kopecný, J., Mrázek, J., Rada, V., Benada, O., Koppová, I., et al. (2009). Bifidobacterium bombi sp. nov., from the bumblebee digestive tract. Int. J. Syst. Evol. Microbiol. 59, 2020–2024. doi: 10.1099/ijs.0.002915-0
Killer, J., Rocková, Š., Vlková, E., Rada, V., Havlik, J., Kopečný, J., et al. (2013). Alloscardovia macacae sp. nov., isolated from the milk of a macaque (Macaca mulatta), emended description of the genus Alloscardovia and proposal of Alloscardovia criceti comb. nov. Int. J. Syst. Evol. Microbiol. 63, 4439–4446. doi: 10.1099/ijs.0.051326-0
Kim, B.-J., Jeong, J., Lee, S. H., Kim, S.-R., Yu, H.-K., Park, Y.-G., et al. (2012a). Mycobacterium koreense sp. nov., a slowly growing non-chromogenic species closely related to Mycobacterium triviale. Int. J. Syst. Evol. Microbiol. 62, 1289–1295. doi: 10.1099/ijs.0.033274-0
Kim, B.-J., Kim, G.-N., Kim, B.-R., Jeon, C. O., Jeong, J., Lee, S. H., et al. (2017). Description of Mycobacterium chelonae subsp. bovis subsp. nov., isolated from cattle (Bos taurus coreanae), emended description of Mycobacterium chelonae and creation of Mycobacterium chelonae subsp. chelonae subsp. nov. Int. J. Syst. Evol. Microbiol. 67, 3882–3887. doi: 10.1099/ijsem.0.002217
Kim, B.-J., Math, R. K., Jeon, C. O., Yu, H.-K., Park, Y.-G., Kook, Y.-H., et al. (2013). Mycobacterium yongonense sp. nov., a slow-growing non-chromogenic species closely related to Mycobacterium intracellulare. Int. J. Syst. Evol. Microbiol. 63, 192–199. doi: 10.1099/ijs.0.037465-0
Kim, J., Roh, S. W., Choi, J.-H., Jung, M.-J., Nam, Y.-D., Kim, M.-S., et al. (2011). Dietzia alimentaria sp. nov., isolated from a traditional Korean food. Int. J. Syst. Evol. Microbiol. 61, 2254–2258. doi: 10.1099/ijs.0.021501-0
Kim, K. K., Lee, K. C., Klenk, H.-P., Oh, H.-M., and Lee, J.-S. (2009). Gordonia kroppenstedtii sp. nov., a phenol-degrading actinomycete isolated from a polluted stream. Int. J. Syst. Evol. Microbiol. 59, 1992–1996. doi: 10.1099/ijs.0.005322-0
Kim, K. K., Roth, A., Andrees, S., Lee, S. T., and Kroppenstedt, R. M. (2002). Nocardia pseudovaccinii sp. nov. Int. J. Syst. Evol. Microbiol. 52, 1825–1829. doi: 10.1099/00207713-52-5-1825
Kim, M., and Chun, J. (2014). 16S rRNA gene based identification of Bacteria and Archaea using the EzTaxon server. Meth. Microbiol. 41, 61–74. doi: 10.1016/bs.mim.2014.08.001
Kim, M.-S., Roh, S. W., and Bae, J.-W. (2010). Bifidobacterium stercoris sp. nov., isolated from human faeces. Int. J. Syst. Evol. Microbiol. 60, 2823–2827. doi: 10.1099/ijs.0.019943-0
Kim, M. K., Pulla, R. K., Kim, S.-Y., Yi, T.-H., Soung, N.-K., and Yang, D.-C. (2008). Sanguibacter soli sp. nov., isolated from soil of a ginseng field. Int. J. Syst. Evol. Microbiol. 58, 538–541. doi: 10.1099/ijs.0.65399-0
Kim, S., Lonsdale, J., Seong, C.-N., and Goodfellow, M. (2003). Streptacidiphilus gen. nov., acidophilic actinomycetes with a wall chemotype I and emendation of the family Streptomycetaceae (Waksman & Henrici 1943 emend Rainey et al. 1997). Antonie Van Leeuwenhoek 83, 107–116.
Kim, S.-J., Hamada, M., Ahn, J.-H., Weon, H.-Y., Suzuki, K.-I., and Kwon, S.-W. (2016). Friedmanniella aerolata sp. nov., isolated from air. Int. J. Syst. Evol. Microbiol. 66, 1970–1975. doi: 10.1099/ijsem.0.000973
Kim, S.-J., Tamura, T., Hamada, M., Ahn, J.-H., Weon, H.-Y., Park, I.-C., et al. (2012b). Compostimonas suwonensis gen. nov., sp. nov., isolated from spent mushroom compost. Int. J. Syst. Evol. Microbiol. 62, 2410–2416. doi: 10.1099/ijs.0.036343-0
Kim, S. B., Brown, R., Oldfield, C., Gilbert, S. C., and Goodfellow, M. (1999). Gordonia desulfuricans sp. nov., a benzothiophene-desulphurizing actinomycete. Int. J. Syst. Bacteriol. 49, 1845–1851.
Kim, S. B., Brown, R., Oldfield, C., Gilbert, S. C., Iliarionov, S., and Goodfellow, M. (2000). Gordonia amicalis sp. nov., a novel dibenzothiophene-desulphurizing actinomycete. Int. J. Syst. Evol. Microbiol. 50, 2031–2036. doi: 10.1099/00207713-50-6-2031
Kim, S. B., and Goodfellow, M. (1999). Reclassification of Amycolatopsis rugosa Lechevalier et al. 1986 as Prauserella rugosa gen. nov., comb. nov. Int. J. Syst. Bacteriol. 49, 507–512.
Kim, S. B., Seong, C. N., Jeon, S. J., Bae, K. S., and Goodfellow, M. (2004). Taxonomic study of neutrotolerant acidophilic actinomycetes isolated from soil and description of Streptomyces yeochonensis sp. nov. Int. J. Syst. Evol. Microbiol. 54, 211–214. doi: 10.1099/ijs.0.02519-0
King, C. E., and King, G. M. (2014). Description of Thermogemmatispora carboxidivorans sp. nov., a carbon-monoxide-oxidizing member of the class Ktedonobacteria isolated from a geothermally heated biofilm, and analysis of carbon monoxide oxidation by members of the class Ktedonobacteria. Int. J. Syst. Evol. Microbiol. 64, 1244–1251. doi: 10.1099/ijs.0.059675-0
Kirschner, P., Teske, A., Schröder, K.-H., Kroppenstedt, R. M., Wolters, J., and Böttger, E. C. (1992). Mycobacterium confluentis sp. nov. Int. J. Syst. Bacteriol. 42, 257–262.
Klatte, S., Rainey, F. A., and Kroppenstedt, R. M. (1994). Transfer of Rhodococcus aichiensis Tsukamura 1982 and Nocardia amarae Lechevalier and Lechevalier 1974 to the genus Gordona as Gordona aichiensis comb. nov. and Gordona amarae comb. nov. Int. J. Syst. Bacteriol. 44, 769–773.
Klenk, H.-P., and Göker, M. (2010). En route to a genome-based classification of Archaea and Bacteria? Syst. Appl. Microbiol. 33, 175–182. doi: 10.1016/j.syapm.2010.03.003
Kloos, W. E., Tornabene, T. G., and Schleifer, K. H. (1974). Isolation and characterization of Micrococci from human skin, including two new species: Micrococcus lylae and Micrococcus kristinae. Int. J. Syst. Bacteriol. 24, 79–101. doi: 10.1099/00207713-24-1-79
Koch, C., Schumann, P., and Stackebrandt, E. (1995). Reclassification of Micrococcus agilis (Alii-Cohen 1889) to the genus Arthrobacter as Arthrobacter agilis comb. nov. and emendation of the genus Arthrobacter. Int. J. Syst. Bacteriol. 45, 837–839.
Korn-Wendisch, F., Kempf, A., Grund, E., Kroppenstedt, R. M., and Kutzner, H. J. (1989). Transfer of Faenia rectivirgula Kurup and Agre 1983 to the genus Saccharopolyspora Lacey and Goodfellow 1975, elevation of Saccharopolyspora hirsuta subsp. taberi Labeda 1987 to species level, and emended description of the genus Saccharopolyspora. Int. J. Syst. Bacteriol. 39, 430–441.
Korn-Wendisch, F., Rainey, F., Kroppenstedt, R. M., Kempf, A., Majazza, A., Kutzner, H. J., et al. (1995). Thermocrispum gen. nov., a new genus of the order Actinomycetales, and description of Thermocrispum municipale sp. nov. and Thermocrispum agreste sp. nov. Int. J. Syst. Bacteriol. 45, 67–77.
Koukila-Kähkölä, P., Springer, B., Böttger, E. C., Paulin, L., Jantzen, E., and Katila, M.-L. (1995). Mycobacterium branderi sp. nov., a new potential human pathogen. Int. J. Syst. Bacteriol. 45, 549–553.
Kraatz, M., Wallace, R. J., and Svensson, L. (2011). Olsenella umbonata sp. nov., a microaerotolerant anaerobic lactic acid bacterium from the sheep rumen and pig jejunum, and emended descriptions of Olsenella, Olsenella uli and Olsenella profusa. Int. J. Syst. Evol. Microbiol. 61, 795–803. doi: 10.1099/ijs.0.022954-0
Krasil'nikov, N. A. (1938). Ray Fungi and Related Organisms-Actinomycetales. Moscow: Akademii Nauk SSSR.
Krasil'nikov, N. A., and Agre, N. S. (1965). The brown group of Actinobifida chromogena n. sp. Mikrobiologiya 34, 284–291.
Kroppenstedt, R. M., Mayilraj, S., Wink, J. M., Kallow, W., Schumann, P., Secondini, C., et al. (2005). Eight new species of the genus Micromonospora, Micromonospora citrea sp. nov., Micromonospora echinaurantiaca sp. nov., Micromonospora echinofusca sp. nov. Micromonospora fulviviridis sp. nov., Micromonospora inyonensis sp. nov., Micromonospora peucetia sp. nov., Micromonospora sagamiensis sp. nov., and Micromonospora viridifaciens sp. nov. Syst. Appl. Microbiol. 28, 328–339. doi: 10.1016/j.syapm.2004.12.011
Kroppenstedt, R. M., Stackebrandt, E., and Goodfellow, M. (1990). Taxonomic revision of the actinomycete genera Actinomadura and Microtetraspora. Syst. Appl. Microbiol. 13, 148–160. doi: 10.1016/S0723-2020(11)80162-1
Kudo, T., Itoh, T., Miyadoh, S., Shomura, T., and Seino, A. (1993). Herbidospora gen. nov., a new genus of the family Streptosporangiaceae Goodfellow et al. 1990. Int. J. Syst. Bacteriol. 43, 319–328.
Kukolya, J., Nagy, I., Láday, M., Tóth, E., Oravecz, O., Márialigeti, K., et al. (2002). Thermobifida cellulolytica sp. nov., a novel lignocellulose-decomposing actinomycete. Int. J. Syst. Evol. Microbiol. 52, 1193–1199.
Kumari, R., Singh, P., Schumann, P., and Lal, R. (2016). Tessaracoccus flavus sp. nov., isolated from the drainage system of a lindane-producing factory. Int. J. Syst. Evol. Microbiol. 66, 1862–1868. doi: 10.1099/ijsem.0.000958
Kurylowicz, W., and Wóznicka, W. (1967). Actinomyces (Streptomyces) varsoviensis. I. Taxonomic studies. Med. Dosw. Mikrobiol. 19, 1–9.
Kusakabe, H., and Isono, K. (1988). Taxonomic studies on Kitasatosporia cystarginea sp. nov., which produces a new antifungal antibiotic cystargin. J. Antibiot. (Tokyo). 41, 1758–1762.
Kusunoki, S., and Ezaki, T. (1992). Proposal of Mycobacterium peregrinum sp. nov., nom. rev., and elevation of Mycobacterium chelonae subsp. abscessus (Kubica et al.) to species status: Mycobacterium abscessus comb. nov. Int. J. Syst. Bacteriol. 42, 240–245.
Kyrpides, N. C., Hugenholtz, P., Eisen, J. A., Woyke, T., Göker, M., Parker, C. T., et al. (2014). Genomic encyclopedia of Bacteria and Archaea: sequencing a myriad of type strains. PLoS Biol. 12:e1001920. doi: 10.1371/journal.pbio.1001920
La Scola, B., Fenollar, F., Fournier, P., Altwegg, M., and Mallet, M. N. (2001). Description of Tropheryma whipplei gen. nov., sp. nov., the Whipple's disease bacillus. Int. J. Syst. Evol. Microbiol. 51, 1471–1479. doi: 10.1099/00207713-51-4-1471
Labeda, D. P. (1986). Transfer of “Nocardia aerocolonigenes” (Shinobu and Kawato 1960) Pridham 1970 into the genus Saccharothrix Labeda, Testa, Lechevalier, and Lechevalier 1984 as Saccharothrix aerocolonigenes sp. nov. Int. J. Syst. Bacteriol. 36, 109–110.
Labeda, D. P. (1987). Transfer of the type strain of Streptomyces erythraeus (Waksman 1923) Waksman and Henrici 1948 to the genus Saccharopolyspora Lacey and Goodfellow 1975 as Saccharopolyspora erythraea sp. nov., and designation of a neotype strain for Streptomyces erythraeus. Int. J. Syst. Bacteriol. 37, 19–22.
Labeda, D. P. (1995). Amycolatopsis coloradensis sp. nov., the Avoparcin (LL-AV290)-producing strain. Int. J. Syst. Bacteriol. 45, 124–127.
Labeda, D. P. (2012). Order VII. Glycomycetales ord. nov., in Bergey's Manual of Systematic Bacteriology, Vol. 5, The Actinobacteria, eds M. Goodfellow, P. Kämpfer, H.-J. Busse, M. E. Trujillo, K. Suzuki, W. Ludwig, et al. (New York, NY: Springer), 546.
Labeda, D. P., Donahue, J. M., Sells, S. F., and Kroppenstedt, R. M. (2007). Lentzea kentuckyensis sp. nov., of equine origin. Int. J. Syst. Evol. Microbiol. 57, 1780–1783. doi: 10.1099/ijs.0.64245-0
Labeda, D. P., Donahue, J. M., Williams, N. M., Sells, S. F., and Henton, M. M. (2003). Amycolatopsis kentuckyensis sp. nov., Amycolatopsis lexingtonensis sp. nov. and Amycolatopsis pretoriensis sp. nov., isolated from equine placentas. Int. J. Syst. Evol. Microbiol. 53, 1601–1605. doi: 10.1099/ijs.0.02691-0
Labeda, D. P., Doroghazi, J. R., Ju, K.-S., and Metcalf, W. W. (2014). Taxonomic evaluation of Streptomyces albus and related species using multilocus sequence analysis and proposals to emend the description of Streptomyces albus and describe Streptomyces pathocidini sp. nov. Int. J. Syst. Evol. Microbiol. 64, 894–900. doi: 10.1099/ijs.0.058107-0
Labeda, D. P., Dunlap, C. A., Rong, X., Huang, Y., Doroghazi, J. R., Ju, K.-S., et al. (2017). Phylogenetic relationships in the family Streptomycetaceae using multi-locus sequence analysis. Antonie Van Leeuwenhoek 110, 563–583. doi: 10.1007/s10482-016-0824-0
Labeda, D. P., Goodfellow, M., Brown, R., Ward, A. C., Lanoot, B., Vanncanneyt, M., et al. (2012). Phylogenetic study of the species within the family Streptomycetaceae. Antonie Van Leeuwenhoek 101, 73–104. doi: 10.1007/s10482-011-9656-0
Labeda, D. P., Goodfellow, M., Chun, J., Zhi, X.-Y., and Li, W.-J. (2011). Reassessment of the systematics of the suborder Pseudonocardineae: transfer of the genera within the family Actinosynnemataceae Labeda and Kroppenstedt 2000 emend. Zhi et al. 2009 into an emended family Pseudonocardiaceae Embley et al. 1989 emend. Zhi et al. 2009. Int. J. Syst. Evol. Microbiol. 61, 1259–1264. doi: 10.1099/ijs.0.024984-0
Labeda, D. P., Hatano, K., Kroppenstedt, R. M., and Tamura, T. (2001). Revival of the genus Lentzea and proposal for Lechevalieria gen. nov. Int. J. Syst. Evol. Microbiol. 51, 1045–1050. doi: 10.1099/00207713-51-3-1045
Labeda, D. P., and Kroppenstedt, R. M. (2004). Emended description of the genus Glycomyces and description of Glycomyces algeriensis sp. nov., Glycomyces arizonensis sp. nov. and Glycomyces lechevalierae sp. nov. Int. J. Syst. Evol. Microbiol. 54, 2343–2346. doi: 10.1099/ijs.0.63089-0
Labeda, D. P., and Kroppenstedt, R. M. (2005). Stackebrandtia nassauensis gen. nov., sp. nov. and emended description of the family Glycomycetaceae. Int. J. Syst. Evol. Microbiol. 55, 1687–1691. doi: 10.1099/ijs.0.63496-0
Labeda, D. P., and Kroppenstedt, R. M. (2007). Proposal of Umezawaea gen. nov., a new genus of the Actinosynnemataceae related to Saccharothrix, and transfer of Saccharothrix tangerinus Kinoshita et al. 2000 as Umezawaea tangerina gen. nov., comb. nov. Int. J. Syst. Evol. Microbiol. 57, 2758–2761. doi: 10.1099/ijs.0.64985-0
Labeda, D. P., and Kroppenstedt, R. M. (2008). Proposal for the new genus Allokutzneria gen. nov. within the suborder Pseudonocardineae and transfer of Kibdelosporangium albatum Tomita et al. 1993 as Allokutzneria albata comb. nov. Int. J. Syst. Evol. Microbiol. 58, 1472–1475. doi: 10.1099/ijs.0.65474-0
Labeda, D. P., Kroppenstedt, R. M., Euzéby, J. P., and Tindall, B. J. (2008). Proposal of Goodfellowiella gen. nov. to replace the illegitimate genus name Goodfellowia Labeda and Kroppenstedt 2006. Int. J. Syst. Evol. Microbiol. 58, 1047–1048. doi: 10.1099/ijs.0.2008/000299-0
Labeda, D. P., Price, N. P., Tan, G. Y. A., Goodfellow, M., and Klenk, H.-P. (2010). Emended description of the genus Actinokineospora Hasegawa 1988 and transfer of Amycolatopsis fastidiosa Henssen et al. 1987 as Actinokineospora fastidiosa comb. nov. Int. J. Syst. Evol. Microbiol. 60, 1444–1449. doi: 10.1099/ijs.0.016568-0
Lacey, J. (1989). Genus Saccharopolyspora Lacey and Goodfellow 1975, 77AL, in Bergey's Manual of Systematic Bacteriology, Vol. 4, eds S. T. Williams, M. E. Sharpe, and J. G. Holt (Baltimore, MD: The Williams & Wilkins Co.), 2382–2386.
Lacey, J., and Goodfellow, M. (1975). A novel actinomycete from sugar-cane bagasse: Saccharopolyspora hirsuta gen. et. sp. nov. J. Gen. Microbiol. 88, 75–85. doi: 10.1099/00221287-88-1-75
Lachner-Sandoval, V. (1898). Über Strahlenpilze. Inaugural Dissertation Strassburg. Universitäts Buchdruckerei Von Carl Georgi, Bonn.
Lagesen, K., Hallin, P., Rødland, E. A., Stærfeldt, H.-H., Rognes, T., and Ussery, D. W. (2007). RNAmmer: consistent and rapid annotation of ribosomal RNA genes. Nucleic Acids Res. 35, 3100–3108. doi: 10.1093/nar/gkm160
Lagier, J.-C., Elkarkouri, K., Rivet, R., Couderc, C., Raoult, D., and Fournier, P.-E. (2013). Non contiguous-finished genome sequence and description of Senegalemassilia anaerobia gen. nov., sp. nov. Stand. Genomic Sci. 7, 343–356. doi: 10.4056/sigs.3246665
Lagier, J.-C., Ramasamy, D., Rivet, R., Raoult, D., and Fournier, P.-E. (2012). Non contiguous-finished genome sequence and description of Cellulomonas massiliensis sp. nov. Stand. Genomic Sci. 7, 258–270. doi: 10.4056/sigs.3316719
Lagkouvardos, I., Pukall, R., Abt, B., Foesel, B. U., Meier-Kolthoff, J. P., Kumar, N., et al. (2016). The Mouse Intestinal Bacterial Collection (miBC) provides host-specific insight into cultured diversity and functional potential of the gut microbiota. Nat. Microbiol. 1, 1–15. doi: 10.1038/nmicrobiol.2016.131
Lai, H., Wei, X., Jiang, Y., Chen, X., Li, Q., Jiang, Y., et al. (2014). Halopolyspora alba gen. nov., sp. nov., isolated from sediment. Int. J. Syst. Evol. Microbiol. 64, 2775–2780. doi: 10.1099/ijs.0.057638-0
Lai, X., Jiang, Y., Saker, R., Chen, X., Bouras, N., Klenk, H.-P., et al. (2017). Reclassification of Mzabimyces algeriensis Saker et al. 2015 as Halopolyspora algeriensis comb. nov. Int. J. Syst. Evol. Microbiol. 67, 2787–2790.
Lamy, B., Marchandin, H., Hamitouche, K., and Laurente, F. (2008). Mycobacterium setense sp. nov., a Mycobacterium fortuitum-group organism isolated from a patient with soft tissue infection and osteitis. Int. J. Syst. Evol. Microbiol. 58, 486–490. doi: 10.1099/ijs.0.65222-0
Lanoot, B., Vancanneyt, M., Cleenwerck, I., Wang, L., Li, W., Liu, Z., et al. (2002). The search for synonyms among streptomycetes by using SDS-PAGE of whole-cell proteins. Emendation of the species Streptomyces aurantiacus, Streptomyces cacaoi subsp. cacaoi, Streptomyces caeruleus and Streptomyces violaceus. Int. J. Syst. Evol. Microbiol. 52, 823–829. doi: 10.1099/ijs.0.02008-0
Lanoot, B., Vancanneyt, M., Dawyndt, P., Cnockaert, M., Zhang, J., Huang, Y., et al. (2004). BOX-PCR fingerprinting as a powerful tool to reveal synonymous names in the genus Streptomyces. Emended descriptions are proposed for the species Streptomyces cinereorectus, S. fradiae, S. tricolor, S. colombiensis, S. filamentosus, S. vinaceus and S. phaeopurpureus. Syst. Appl. Microbiol. 27, 84–92.
Lanoot, B., Vancanneyt, M., Schoor, A., Van Liu, Z., and Swings, J. (2005). Reclassification of Streptomyces nigrifaciens as a later synonym of Streptomyces flavovirens; Streptomyces citreofluorescens, Streptomyces chrysomallus subsp. chrysomallus and Streptomyces fluorescens as later synonyms of Streptomyces anulatus; Streptomyces chibaensis as a later synonym of Streptomyces corchorusii; Streptomyces flaviscleroticus as a later synonym of Streptomyces minutiscleroticus; and Streptomyces lipmanii, Streptomyces griseus subsp. alpha, Streptomyces griseus subsp. cretosus and Streptomyces willmorei as later synonyms of Streptomyces microflavus. Int. J. Syst. Evol. Microbiol. 55, 729–731. doi: 10.1099/ijs.0.63391-0
Lassalle, F., Périan, S., Bataillon, T., Nesme, X., Duret, L., and Daubin, V. (2015). GC-Content evolution in bacterial genomes: the biased gene conversion hypothesis expands. PLoS Genet. 11:e1004941. doi: 10.1371/journal.pgen.1004941
Lauer, E. (1990). Bifidobacterium gallicum sp. nov. isolated from human feces. Int. J. Syst. Bacteriol. 40, 100–102.
Lawson, P. A., Collins, M. D., Schumann, P., Tindall, B. J., Hirsch, P., and Labrenz, M. (2000). New LL-diaminopimelic acid-containing actinomycetes from hypersaline, heliothermal and meromictic Antarctic Ekho Lake: Nocardioides aquaticus sp. nov. and Friedmannielly lacustris sp. nov. Syst. Appl. Microbiol. 23, 219–229. doi: 10.1016/S0723-2020(00)80008-9
Lawson, P. A., Falsen, E., Åkervall, E., Vandamme, P., and Collins, M. D. (1997). Characterization of some Actinomyces-like isolates from human clinical specimens: reclassification of Actinomyces suis (Soltys and Spratling) as Actinobaculum suis comb. nov. and description of Actinobaculum schaalii sp. nov. Int. J. Syst. Bacteriol. 47, 899–903.
Lawson, P. A., Nikolaitchouk, N., Falsen, E., Westling, K., and Collins, M. D. (2001). Actinomyces funkei sp. nov., isolated from human clinical specimens. Int. J. Syst. Evol. Microbiol. 51, 853–855.
le Roes, M., Goodwin, C. M., and Meyers, P. R. (2008). Gordonia lacunae sp. nov., isolated from an estuary. Syst. Appl. Microbiol. 31, 17–23. doi: 10.1016/j.syapm.2007.10.001
le Roes, M. L., and Meyers, P. R. (2008). Nonomuraea candida sp. nov., a new species from south African soil. Antonie Van Leeuwenhoek 93, 133–139. doi: 10.1007/s10482-007-9187-x
Le, S. Q., and Gascuel, O. (2008). An improved general amino acid replacement matrix. Mol. Biol. Evol. 25, 1307–1320. doi: 10.1093/molbev/msn067
Lechevalier, H. A., and Lechevalier, M. P. (1970). A critical evaluation of the genera of aerobic actinomycetes, in The Actinomycetales, ed Prauser (Jena: Gustav Fischer Verlag), 393–405.
Lechevalier, M. P. (1972). Description of a new species, Oerskovia xanthineolytica, and emendation of Oerskovia Prauser et al. Int. J. Syst. Bacteriol. 22, 260–264.
Lechevalier, M. P. (1976). The taxonomy of the genus Nocardia: Some light at the end of the tunnel?, in The Biology of the Nocardiae, eds M. Goodfellow, G. H. Brownell, and J. A. Serrano (London: Academic press), 1–38.
Lechevalier, M. P., Lechevalier, H., and Holbert, P. E. (1968). Sporichthya, un nouveau genre de streptomycetaceae. Ann. Inst. Pasteur 114, 277–285.
Lechevalier, M. P., and Lechevalier, H. A. (1975). Actinoplanete with cylindrical sporangia, Actinoplanes rectilineatus sp. nov. Int. J. Syst. Bacteriol. 25, 371–376.
Lechevalier, M. P., Prauser, H., Labeda, D. P., and Ruan, J.-S. (1986). Two new genera of nocardioform actinomycetes: Amycolata gen. nov. and Amycolatopsis gen. nov. Int. J. Syst. Bacteriol. 36, 29–37. doi: 10.1099/00207713-36-1-29
Lee, D. W., and Lee, S. D. (2010a). Actinomadura scrupuli sp. nov., isolated from rock. Int. J. Syst. Evol. Microbiol. 60, 2647–2651. doi: 10.1099/ijs.0.017608-0
Lee, D. W., and Lee, S. D. (2010b). Koreibacter algae gen. nov., sp. nov., isolated from seaweed. Int. J. Syst. Evol. Microbiol. 60, 1510–1515. doi: 10.1099/ijs.0.014647-0
Lee, D. W., and Lee, S. D. (2010c). Marmoricola scoriae sp. nov., isolated from volcanic ash. Int. J. Syst. Evol. Microbiol. 60, 2135–2139. doi: 10.1099/ijs.0.018242-0
Lee, H., Lee, S.-A., Lee, I.-K., Yu, H.-K., Park, Y.-G., Jeong, J., et al. (2010). Mycobacterium paraterrae sp. nov. recovered from a clinical specimen: novel chromogenic slow growing mycobacteria related to Mycobacterium terrae complex. Microbiol. Immunol. 54, 46–53. doi: 10.1111/j.1348-0421.2009.00184.x
Lee, H. J., Cho, S.-L., Jung, M. Y., Nguyen, T. H., Van Jung, Y.-C., Park, H. K., et al. (2009). Corynebacterium doosanense sp. nov., isolated from activated sludge. Int. J. Syst. Evol. Microbiol. 59, 2734–2737. doi: 10.1099/ijs.0.010108-0
Lee, K. C., Suh, M. K., Eom, M. K., Kim, K. K., Kim, J.-S., Kim, D.-S., et al. (2018). Jatrophihabitans telluris sp. nov., isolated from sediment soil of lava forest wetlands and the emended description of the genus Jatrophihabitans. Int. J. Syst. Evol. Microbiol. 68, 1107–1111. doi: 10.1099/ijsem.0.002639
Lee, L.-H., Azman, A.-S., Zainal, N., Eng, S.-K., Mutalib, N.-S. A., Yin, W.-F., et al. (2014a). Microbacterium mangrovi sp. nov., an amylolytic actinobacterium isolated from mangrove forest soil. Int. J. Syst. Evol. Microbiol. 64, 3513–3519. doi: 10.1099/ijs.0.062414-0
Lee, L.-H., Zainal, N., Azman, A.-S., Eng, S.-K., Mutalib, N.-S. A., Yin, W.-F., et al. (2014b). Streptomyces pluripotens sp. nov., a bacteriocin-producing streptomycete that inhibits meticillin-resistant Staphylococcus aureus. Int. J. Syst. Evol. Microbiol. 64, 3297–3306. doi: 10.1099/ijs.0.065045-0
Lee, S.-Y., Kim, B.-J., Kim, H., Won, Y.-S., Jeon, C. O., Jeong, J., et al. (2016). Mycobacterium paraintracellulare sp. nov., for the genotype INT-1 of Mycobacterium intracellulare. Int. J. Syst. Evol. Microbiol. 66, 3132–3141. doi: 10.1099/ijsem.0.001158
Lee, S. D. (2006a). Amycolatopsis jejuensis sp. nov. and Amycolatopsis halotolerans sp. nov., novel actinomycetes isolated from a natural cave. Int. J. Syst. Evol. Microbiol. 56, 549–553. doi: 10.1099/ijs.0.63881-0
Lee, S. D. (2006b). Nocardia jejuensis sp. nov., a novel actinomycete isolated from a natural cave on Jeju Island, Republic of Korea. Int. J. Syst. Evol. Microbiol. 56, 559–562. doi: 10.1099/ijs.0.63866-0
Lee, S. D. (2006c). Phycicoccus jejuensis gen. nov., sp., an actinomycete isolated from seaweed. Int. J. Syst. Evol. Microbiol. 56, 2369–2373. doi: 10.1099/ijs.0.64271-0
Lee, S. D. (2007). Labedella gwakjiensis gen. nov., sp. nov., a novel actinomycete of the family Microbacteriaceae. Int. J. Syst. Evol. Microbiol. 57, 2498–2502. doi: 10.1099/ijs.0.64591-0
Lee, S. D. (2009). Amycolatopsis ultiminotia sp. nov., isolated from rhizosphere soil, and emended description of the genus Amycolatopsis. Int. J. Syst. Evol. Microbiol. 59, 1401–1404. doi: 10.1099/ijs.0.006577-0
Lee, S. D. (2012). Motilibacter peucedani gen. nov., sp. nov., isolated from rhizosphere soil. Int. J. Syst. Evol. Microbiol. 62, 315–321. doi: 10.1099/ijs.0.030007-0
Lee, S. D. (2013a). Phycicoccus badiiscoriae sp. nov., a novel actinomycete isolated from scoria. Int. J. Syst. Evol. Microbiol. 63, 989–994. doi: 10.1099/ijs.0.041699-0
Lee, S. D. (2013b). Proposal of Motilibacteraceae fam. nov., with the description of Motilibacter rhizosphaerae sp. nov. Int. J. Syst. Evol. Microbiol. 63, 3818–3822. doi: 10.1099/ijs.0.052357-0
Lee, S. D. (2015). Antricoccus suffuscus gen. nov., sp. nov., isolated from a natural cave. Int. J. Syst. Evol. Microbiol. 65, 4410–4416. doi: 10.1099/ijsem.0.000588
Lee, S. D., Kim, E. S., Roe, J.-H., Kim, J., Kang, S.-O., and Hah, Y. C. (2000). Saccharothrix violacea sp. nov., isolated from a gold mine cave, and Saccharothrix albidocapillata comb. nov. Int. J. Syst. Evol. Microbiol. 50, 1315–1323. doi: 10.1099/00207713-50-3-1315
Lee, S. D., and Lee, D. W. (2007). Lapillicoccus jejuensis gen. nov., sp. nov., a novel actinobacterium of the family Intrasporangiaceae, isolated from stone. Int. J. Syst. Evol. Microbiol. 57, 2794–2798. doi: 10.1099/ijs.0.64911-0
Lefort, V., Desper, R., and Gascuel, O. (2015). FastME 2.0: a comprehensive, accurate, and fast distance-based phylogeny inference program. Mol. Biol. Evol. 32, 2798–2800. doi: 10.1093/molbev/msv150
Lehmann, K. B., and Neumann, R. O. (1896). Atlas und Grundriss der Bakteriologie und Lehrbuch der Speziellen Bakteriologischen Diagnostik, 1st Edn. München: J.F. Lehmann.
Lehmann, K. B., and Neumann, R. O. (1899). Lehmann's Medizin, Handatlanter X. Atlas und Grundriss der Bakteriologie und Lehrbuch der Speziellen Bakteriologischen Diagnostik, 2nd Edn. München: J. F. Lehmann.
Lehmann, K. B., and Neumann, R. O. (1907). Lehmann's Medizin, Handatlanten. X. Atlas und Grundriss der Bakteriologie und Lehrbuch der Speziellen Bakteriologischen Diagnostik, 4th Edn. München: J.F. Lehmann.
Lehnen, A., Busse, H.-J., Frölich, K., Krasinka, M., Kämpfer, P., and Speck, S. (2006). Arcanobacterium bialowiezense sp. nov. and Arcanobacterium bonasi sp. nov., isolated from the prepuce of European bison bulls (Bison bonasus) suffering from balanoposthitis, and emended description of the genus Arcanobacterium Collins et al. 1983. Int. J. Syst. Evol. Microbiol. 56, 861–866. doi: 10.1099/ijs.0.63923-0
Letunic, I., and Bork, P. (2016). Interactive tree of life (iTOL) v3: an online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 44, W242–W245. doi: 10.1093/nar/gkw290
Lévy-Frébault, V., Rafidinarivo, É., Promé, J.-C., Grandy, J., Boisvert, H., and David, H. L. (1983). Mycobacterium fallax sp. nov. Int. J. Syst. Bacteriol. 33, 336–343.
Lewin, R. A. (1970). New Herpetosiphon species (Flexibacterales). Can. J. Microbiol. 16, 517–520. doi: 10.1139/m70-087
Li, B., Xie, C.-H., and Yokota, A. (2007). Nocardioides exalbidus sp. nov., a novel actinomycete isolated from lichen in Izu-Oshima Island, Japan. Actinomycetologica 21, 22–26. doi: 10.3209/saj.SAJ210103
Li, J., Zhao, G.-Z., Zhang, Y.-Q., Klenk, H.-P., Pukall, R., Qin, S., et al. (2008a). Dietzia schimae sp. nov. and Dietzia cercidiphylli sp. nov., from surface-sterilized plant tissues. Int. J. Syst. Evol. Microbiol. 58, 2549–2554. doi: 10.1099/ijs.0.2008/000919-0
Li, L., Mao, Y.-J., Xie, Q.-Y., Deng, Z., and Kui, H. (2013). Micromonospora avicenniae sp. nov., isolated from a root of Avicennia marina. Antonie Van Leeuwenhoek 103, 1089–1096. doi: 10.1007/s10482-013-9888-2
Li, L., Stoeckert, C. J., and Roos, D. S. (2003a). OrthoMCL: identification of ortholog groups for eukaryotic genomes. Genome Res. 13, 2178–2189. doi: 10.1101/gr.1224503
Li, M.-G., Li, W.-J., Xu, P., Cui, X.-L., Xu, L.-H., and Jiang, C.-L. (2003b). Nocardiopsis xinjiangensis sp. nov., a halophilic actinomycete isolated from a saline soil sample in China. Int. J. Syst. Evol. Microbiol. 53, 317–321. doi: 10.1099/ijs.0.02055-0
Li, W.-J., Xu, P., Tang, S.-K., Xu, L.-H., Kroppenstedt, R. M., Stackebrandt, E., et al. (2003c). Prauserella halophila sp. nov. and Prauserella alba sp. nov., moderately halophilic actinomycetes from saline soil. Int. J. Syst. Evol. Microbiol. 53, 1545–1549. doi: 10.1099/ijs.0.02611-0
Li, W.-J., Xu, P., Zhang, L.-P., Tang, S.-K., Cui, X.-L., Mao, P.-H., et al. (2003d). Streptomonospora alba sp. nov., a novel halophilic actinomycete, and emended description of the genus Streptomonospora Cui et al. 2001. Int. J. Syst. Evol. Microbiol. 53, 1421–1425. doi: 10.1099/ijs.0.02543-0
Li, W.-J., Zhi, X.-Y., and Euzéby, J. P. (2008b). Proposal of Yaniellaceae fam. nov., Yaniella gen. nov. and Sinobaca gen. nov. as replacements for the illegitimate prokaryotic names Yaniaceae Li et al. 2005, Yania Li et al. 2004, emend Li et al. 2005, and Sinococcus Li et al. 2006, respectively. Int. J. Syst. Evol. Microbiol. 58, 525–527. doi: 10.1099/ijs.0.65792-0
Li, W. J., Kroppenstedt, R., Wang, D., Tang, S. K., Lee, J. C., Park, D. J., et al. (2006). Five novel species of the genus Nocardiopsis isolated from hypersaline soils and emended description of Nocardiopsis salina Li et al. 2004. Int. J. Syst. Evol. Microbiol. 56, 1089–1096. doi: 10.1099/ijs.0.64033-0
Li, W. J., Tang, S. K., Stackebrandt, E., Kroppenstedt, R. M., Schumann, P., Xu, L. H., et al. (2003e). Saccharomonospora paurometabolica sp. nov., a moderately halophilic actinomycete isolated from soil in China. Int. J. Syst. Evol. Microbiol. 53, 1591–1594. doi: 10.1099/ijs.0.02633-0
Li, X., Tambong, J., Yuan, K. X., Chen, W., Xu, H., Lévesque, C. A., et al. (2017). Re-classification of Clavibacter michiganensis subspecies on the basis of whole-genome and multi-locus sequence analyses. Int. J. Syst. Evol. Microbiol. 68, 234–240. doi: 10.1099/ijsem.0.002492
Li, X., Zhang, L., Ding, Y., Gao, Y., Ruan, J., and Ying, H. (2012). Nonomuraea jiangxiensis sp. nov., isolated from acidic soil. Int. J. Syst. Evol. Microbiol. 62, 1409–1413. doi: 10.1099/ijs.0.034520-0
Li, Y., Guo, M., Wang, L., Wang, X., and Lin, C. (2016). Populibacterium corticicola gen. nov., sp. nov., a member of the family Jonesiaceae, isolated from symptomatic bark of Populus × euramericana canker. Int. J. Syst. Evol. Microbiol. 66, 3743–3748. doi: 10.1099/ijsem.0.001261
Liao, Z.-L., Tang, S.-K., Guo, L., Zhang, Y.-Q., Tian, X.-P., Jiang, C.-L., et al. (2009). Verrucosispora lutea sp. nov., isolated from a mangrove sediment sample. Int. J. Syst. Evol. Microbiol. 59, 2269–2273. doi: 10.1099/ijs.0.008813-0
Lienau, K. E., and DeSalle, R. (2009). Evidence, content and corroboration and the tree of life. Acta Biotheor. 57, 187–199. doi: 10.1007/s10441-008-9066-5
Lin, Y.-C., Uemori, K., de Briel, D. A., Arunpairojana, V., and Yokota, A. (2004). Zimmermannella helvola gen. nov., sp. nov., Zimmermannella alba sp. nov., Zimmermannella bifida sp. nov., Zimmermannella faecalis sp. nov. and Leucobacter albus sp. nov., novel members of the family Microbacteriaceae. Int. J. Syst. Evol. Microbiol. 54, 1669–1676. doi: 10.1099/ijs.0.02741-0
Lindenbein, W. (1952). Über einige chemisch interessante Aktinomycetenstämme und ihre Klassifizierung und ihre Klassifizierung. Arch. Microbiol. 17, 361–383.
Linos, A., Berekaa, M. M., Steinbüchel, A., Kim, K. K., Spröer, C., and Kroppenstedt, R. M. (2002). Gordonia westfalica sp. nov., a novel rubber-degrading actinomycete. Int. J. Syst. Evol. Microbiol. 52, 1133–1139. doi: 10.1099/00207713-52-4-1133
Linos, A., Steinbüchel, A., Spröer, C., and Kroppenstedt, R. M. (1999). Gordonia polyisoprenivorans sp. nov., a rubber-degrading actinomycete isolated from an automobile tyre. Int. J. Syst. Bacteriol. 49, 1785–1791.
Liu, M., Peng, F., Wang, Y., Zhang, K., Chen, G., and Frang, C. (2009a). Kineococcus xinjiangensis sp. nov., isolated from desert sand. Int. J. Syst. Evol. Microbiol. 59, 1090–1093. doi: 10.1099/ijs.0.004168-0
Liu, N., Wang, H., Liu, M., Gu, Q., Zheng, W., and Huang, Y. (2009b). Streptomyces alni sp. nov., a daidzein-producing endophyte isolated from a root of Alnus nepalensis. Int. J. Syst. Evol. Microbiol. 59, 254–258. doi: 10.1099/ijs.0.65769-0
Liu, Q., Liu, H., Wen, Y., Zhou, Y., and Xin, Y. (2012). Cryobacterium flavum sp. nov., and Cryobacterium luteum isolated from glacier ice. Int. J. Syst. Evol. Microbiol. 62, 1296–1299. doi: 10.1099/ijs.0.033738-0
Liu, Q., Xin, Y., Liu, H., Zhou, Y., and Wen, Y. (2013). Nocardioides szechwanensis sp. nov. and Nocardioides psychrotolerans sp. nov. isolated from a glacier. Int. J. Syst. Evol. Microbiol. 63, 129–133. doi: 10.1099/ijs.0.038091-0
Liu, S.-W., Xu, M., Tuo, L., Li, X.-J., Hu, L., Chen, L., et al. (2016). Phycicoccus endophyticus sp. nov., an endophytic actinobacterium isolated from Bruguiera gymnorhiza. Int. J. Syst. Evol. Microbiol. 66, 1105–1111. doi: 10.1099/ijsem.0.000842
Liu, X.-Y., Wang, B.-J., Jiang, C.-Y., and Liu, S.-J. (2008). Ornithinimicrobium pekingense sp. nov., isolated from activated sludge. Int. J. Syst. Evol. Microbiol. 58, 116–119. doi: 10.1099/ijs.0.65229-0
Liu, X.-Z., Wang, Q.-M., Göker, M., Groenewald, M., Kachalkin, A. V., Lumbsch, H. T., et al. (2015). Towards an integrated phylogenetic classification of the Tremellomycetes. Stud. Mycol. 81, 85–147. doi: 10.1016/j.simyco.2015.12.001
Liu, Y., Ge, F., Chen, G., Li, W., Ma, P., Zhang, G., et al. (2011). Gordonia neofelifaecis sp. nov., a cholesterol side-chain-cleaving actinomycete isolated from the faeces of Neofelis nebulosa. Int. J. Syst. Evol. Microbiol. 61, 165–169. doi: 10.1099/ijs.0.020321-0
Liu, Z., Shi, Y., Zhang, Y., Zhou, Z., Lu, Z., Li, W., et al. (2005). Classification of Streptomyces griseus (Krainsky 1914) Waksman and Henrici 1948 and related species and the transfer of ‘Microstreptospora cinerea’ to the genus Streptomyces as Streptomyces yanii sp. nov. Int. J. Syst. Evol. Microbiol. 55, 1605–1610. doi: 10.1099/ijs.0.63654-0
Liu, Z.-P., Wu, J.-F., Liu, Z.-H., and Liu, S.-J. (2006). Pseudonocardia ammonioxydans sp. nov., isolated from coastal sediment. Int. J. Syst. Evol. Microbiol. 56, 555–558. doi: 10.1099/ijs.0.63878-0
Löffler, F. E., Yan, J., Ritalahti, K. M., Adrian, L., Edwards, E. A., Konstantinidis, K. T., et al. (2013). Dehalococcoides mccartyi gen. nov., sp. nov., obligately organohalide-respiring anaerobic bacteria relevant to halogen cycling and bioremediation, belong to a novel bacterial class, Dehalococcoidia classis nov., order Dehalococcoidales ord. nov. and. Int. J. Syst. Evol. Microbiol. 63, 625–635. doi: 10.1099/ijs.0.034926-0
Lu, Z., Liu, Z., Wang, L., Zhang, Y., Qi, W., and Goodfellow, M. (2001). Saccharopolyspora flava sp. nov. and Saccharopolyspora thermophila sp. nov., novel actinomycetes from soil. Int. J. Syst. Evol. Microbiol. 51, 319–325. doi: 10.1099/00207713-51-2-319
Ludwig, W., Euzéby, J., Schumann, P., Busse, H. J., Trujillo, M. E., Kämpfer, P., et al. (2012). Road map of the phylum Actinobacteria, in Bergey's Manual of Systematic Bacteriology, Vol. 5, The Actinobacteria, eds M. Goodfellow, P. Kämpfer, H.-J. Busse, M. E. Trujillo, K. Suzuki, W. Ludwig, et al. (New York, NY: Springer), 1–28.
Ludwig, W., and Klenk, H.-P. (2005). Overview: a phylogenetic backbone and taxonomic framework for procaryotic systematics, in Bergey's Manual of Systematic Bacteriology, Vol. 2, The Proteobacteria (Part A), eds. D. J. Brenner, N. R. Krieg, J. T. Staley, and G. M. Garrity (New York, NY: Springer), 49–66. doi: 10.1007/0-387-28021-9_8
Luo, H., Gu, Q., Xie, J., Hu, C., Liu, Z., and Huang, Y. (2007). Gordonia shandongensis sp. nov., isolated from soil in China. Int. J. Syst. Evol. Microbiol. 57, 605–608. doi: 10.1099/ijs.0.64536-0
Luo, H.-Y., Wang, Y.-R., Miao, L.-H., Yang, P.-L., Shi, P.-J., Fang, C.-X., et al. (2009). Nesterenkonia alba sp. nov., an alkaliphilic actinobacterium isolated from the black liquor treatment system of a cotton pulp mill. Int. J. Syst. Evol. Microbiol. 59, 863–868. doi: 10.1099/ijs.0.003376-0
Luo, X., Kai, L., Wang, Y., Wan, C., and Zhang, L.-L. (2017). Streptomyces luteus sp. nov., an actinomycete isolated from soil. Int. J. Syst. Evol. Microbiol. 67, 543–547. doi: 10.1099/ijsem.0.001611
Luo, Y., Xiao, J., Wang, Y., Xu, J., Xie, S., and Xu, J. (2011). Streptomyces indicus sp. nov. an actinomycete isolated from deep-sea sediment. Int. J. Syst. Evol. Microbiol. 61, 2712–2716. doi: 10.1099/ijs.0.029389-0
Mackellar, D., Lieber, L., Norman, J. S., Bolger, A., Tobin, C., Murray, J. W., et al. (2016). Streptomyces thermoautotrophicus does not fix nitrogen. Sci. Rep. 6, 20086. doi: 10.1038/srep20086
Madhaiyan, M., Hu, C. J., Kim, S.-J., Weon, H.-Y., Kwon, S.-W., and Ji, L. (2013). Jatrophihabitans endophyticus gen. nov., sp. nov., an endophytic actinobacterium isolated from a surface-sterilized stem of Jatropha curcas L. Int. J. Syst. Evol. Microbiol. 63, 1241–1248. doi: 10.1099/ijs.0.039685-0
Madhaiyan, M., Poonguzhali, S., Saravanan, V. S., Duraipandiyan, V., Al-Dhabi, N. A., Pragatheswari, D., et al. (2016). Streptomyces pini sp. nov., an actinomycete isolated from phylloplane of pine (Pinus sylvestris L.) needle-like leaves. Int. J. Syst. Evol. Microbiol. 66, 4204–4210. doi: 10.1099/ijsem.0.001336
Majumdar, S., Prabhagaran, S. R., Shivaji, S., and Lal, R. (2006). Reclassification of Amycolatopsis orientalis DSM 43387 as Amycolatopsis benzoatilytica sp. nov. Int. J. Syst. Evol. Microbiol. 56, 199–204. doi: 10.1099/ijs.0.63766-0
Maldonado, L., Hookey, J. V., Ward, A. C., and Goodfellow, M. (2000). The Nocardia salmonicida clade, including descriptions of Nocardia cummidelens sp. nov., Nocardia fluminea sp. nov. and Nocardia soli sp. nov. Ant. Van Leeuwenhoek 78, 367–377. doi: 10.1023/A:1010230632040
Maldonado, L. A., Fenical, W., Jensen, P. R., Kauffman, C. A., Mincer, T. J., Ward, A. C., et al. (2005). Salinispora arenicola gen. nov., sp. nov. and Salinispora tropica sp. nov., obligate marine actinomycetes belonging to the family Micromonosporaceae. Int. J. Syst. Evol. Microbiol. 55, 1759–1766. doi: 10.1099/ijs.0.63625-0
Manaia, C. L., Nogales, B., and Nunes, O. C. (2012). Genus XXI. Pseudoclavibacter Manaia, Nogales, Weiss and Nunes 2004, 787VP, in Bergey's Manual of Systematic Bacteriology, Vol. 5, The Actinobacteria, eds M. Goodfellow, P. Kämpfer, H.-J. Busse, M. E. Trujillo, K. Suzuki, W. Ludwig, et al. (New York, NY: Springer), 948.
Manaia, C. M., Nogales, B., Weiss, N., and Nunes, O. C. (2004). Gulosibacter molinativorax gen. nov., a molinate-degrading bacterium, and classification of “Brevibacterium helvolum” DSM 20419 as Pseudoclavibacter helvolus gen. nov., sp. nov. Int. J. Syst. Evol. Microbiol. 54, 783–789. doi: 10.1099/ijs.0.02851-0
Mann, S., and Chen, Y.-P. P. (2010). Bacterial genomic G + C composition-eliciting environmental adaptation. Genomics 95, 7–15. doi: 10.1016/j.ygeno.2009.09.002
Mao, J., Wang, J., Dai, H.-Q., Zhang, Z.-D., Tang, Q.-Y., Ren, B., et al. (2011). Yuhushiella deserti gen. nov., sp. nov., a new member of the suborder Pseudonocardineae. Int. J. Syst. Evol. Microbiol. 61, 621–630. doi: 10.1099/ijs.0.019588-0
Markowitz, V. M., Mavromatis, K., Ivanova, N. N., Chen, I.-M. A., Chu, K., and Kyrpides, N. C. (2009). IMG ER: a system for microbial genome annotation expert review and curation. Bioinformatics 25, 2271–2278. doi: 10.1093/bioinformatics/btp393
Martin, K., Schäfer, J., and Kämpfer, P. (2010). Promicromonospora umidemergens sp. nov., isolated from moisture from indoor wall material. Int. J. Syst. Evol. Microbiol. 60, 537–541. doi: 10.1099/ijs.0.014142-0
Maruo, T., Sakamoto, M., Ito, C., Toda, T., and Benno, Y. (2008). Adlercreutzia equolifaciens gen. nov., sp. nov., an equol-producing bacterium isolated from human faeces, and emended description of the genus Eggerthella. Int. J. Syst. Evol. Microbiol. 58, 1221–1227. doi: 10.1099/ijs.0.65404-0
Masaki, T., Ohkusu, K., Hata, H., Fujiwara, N., Iihara, H., Yamada-Noda, M., et al. (2006). Mycobacterium kumamotonense sp. nov. recovered from clinical specimen and the first isolation report of Mycobacterium arupense in Japan: novel slowly growing, nonchromogenic clinical isolates related to Mycobacterium terrae complex. Microbiol. Immunol. 50, 889–897. doi: 10.1111/j.1348-0421.2006.tb03865.x
Masco, L., Ventura, M., Zink, R., Huys, G., and Swings, J. (2004). Polyphasic taxonomic analysis of Bifidobacterium animalis and Bifidobacterium lactis reveals relatedness at the subspecies level: reclassification of Bifidobacterium animalis as Bifidobacterium animalis subsp. animalis subsp. nov and Bifidobacterium lactis as Bifidobacterium animalis subsp. lactis subsp. nov. Int. J. Syst. Evol. Microbiol. 54, 1137–1143. doi: 10.1099/ijs.0.03011-0
Mason, D. J., Dietz, A., and De Boer, C. (1963). Lincomycin, a new antibiotic. I. Discovery and biological properties. Antimicrob. Agents Chemother. 1962, 554–559.
Maszenan, A. M., Seviour, R. J., Patel, B. K. C., Schumann, P., Burghardt, J., Tokiwa, Y., et al. (2000). Three isolates of novel polyphosphate-accumulating Gram-positive cocci, obtained from activated sludge, belong to a new genus, Tetrasphaera gen. nov., and description of two new species, Tetrasphaera japonica sp. nov. and Tetrasphaera australiensis sp. nov. Int. J. Syst. Evol. Microbiol. 50, 593–603. doi: 10.1099/00207713-50-2-593
Maszenan, A. M., Seviour, R. J., Patel, B. K. C., Schumann, P., Burghardt, J., Webb, R. I., et al. (1999a). Friedmanniella spumicola sp. nov. and Friedmanniella capsulata sp. nov. from activated sludge foam: Gram-positive cocci that grow in aggregates of repeating groups of cocci. Int. J. Syst. Bacteriol. 49, 1667–1680.
Maszenan, A. M., Seviour, R. J., Patel, B. K. C., Schumann, P., and Rees, G. N. (1999b). Tessaracoccus bendigoensis gen. nov., sp. nov., a Gram-positive coccus occurring in regular packages or tetrads, isolated from activated sludge biomass. Int. J. Syst. Bacteriol. 49, 459–468.
Maszenan, A. M., Tay, J.-H., Schumann, P., Jiang, H.-L., and Tay, S. T.-L. (2005). Quadrisphaera granulorum gen. nov., sp. nov., a Gram-positive polyphospate-accumulating coccus in tetrads or aggregates isolated from aerobic granules. Int. J. Syst. Evol. Microbiol. 55, 1771–1777. doi: 10.1099/ijs.0.63583-0
Matsumae, A., Ohtani, M., Takeshima, H., and Hata, T. (1968). A new genus of the Actinomycetales: Kitasatoa gen. nov. J. Antibiot. 21, 616–625.
Matsumoto, A., Kasai, H., Matsuo, Y., Shizuri, Y., Ichikawa, N., Fujita, N., et al. (2013). Ilumatobacter nonamiense sp. nov. and Ilumatobacter coccineum sp. nov., isolated from seashore sand. Int. J. Syst. Evol. Microbiol. 63, 3404–3408. doi: 10.1099/ijs.0.047316-0
Matsumoto, A., Nakai, K., Morisaki, K., Omura, S., and Takahashi, Y. (2010). Demequina salsinemoris sp. nov., isolated on agar media supplemented with ascorbic acid or rutin. Int. J. Syst. Evol. Microbiol. 60, 1206–1209. doi: 10.1099/ijs.0.012617-0
Matsuyama, H., Yumoto, I., Kudo, T., and Shida, O. (2003). Rhodococcus tukisamuensis sp. nov., isolated from soil. Int. J. Syst. Evol. Microbiol. 53, 1333–1337. doi: 10.1099/ijs.0.02523-0
Mattareli, P., Bonaparte, C., Pot, B., and Biavati, B. (2008). Proposal to reclassify the three biotypes of Bifidobacterium longum as three subspecies: Bifidobacterium longum subsp. longum subsp. nov., Bifidobacterium longum subsp. infantis comb. nov. and Bifidobacterium longum subsp. suis comb. nov. Int. J. Syst. Evol. Microbiol. 58, 767–772. doi: 10.1099/ijs.0.65319-0
Mavromatis, K., Land, M. L., Brettin, T. S., Quest, D. J., Copeland, A., Clum, A., et al. (2012). The fast changing landscape of sequencing technologies and their impact on microbial genome assemblies and annotation. PLoS ONE 7:e48837. doi: 10.1371/journal.pone.0048837
Mawlankar, R. R., Mual, P., Sonalkar, V. V., Thorat, M. N., Verma, A., Srinivasan, K., et al. (2015). Microbacterium enclense sp. nov., isolated from sediment sample. Int. J. Syst. Evol. Microbiol. 65, 2064–2070. doi: 10.1099/ijs.0.000221
Mayilraj, S., Suresh, K., Kroppenstedt, R. M., and Saini, H. S. (2006a). Dietzia kunjamensis sp. nov., isolated from the Indian Himalayas. Int. J. Syst. Evol. Microbiol. 56, 1667–1671. doi: 10.1099/ijs.0.64212-0
Mayilraj, S., Suresh, K., Schumann, P., Kroppenstedt, R. M., and Saini, H. S. (2006b). Agrococcus lahaulensis sp. nov., isolated from a cold desert of the Indian Himalayas. Int. J. Syst. Evol. Microbiol. 56, 1807–1810. doi: 10.1099/ijs.0.64247-0
McBride, M. E., Ellner, K. M., Black, H. S., Clarridge, J. E., and Wolf, J. E. (1993). A new Brevibacterium sp. isolated from infected genital hair of patients with white piedra. J. Med. Microbiol. 39, 255–261.
McCarthy, A. J., and Cross, T. (1984). A taxonomic study of Thermomonospora and other monosporic actinomycetes. J. Gen. Microbiol. 130, 5–25.
McClung, N. M. (1974). Family VI. Nocardiaceae Cartellani and Chalmers 1919, 1040, in Bergey's Manual of Determinative Bacteriology, eds R. E. Buchanan and N. E. Gibbons (Baltimore, MD: The Williams & Wilkins Co.), 726–746.
McDowell, A., Barnard, E., Liu, J., Li, H., and Patrick, S. (2016). Proposal to reclassify Propionibacterium acnes type i as Propionibacterium acnes subsp. acnes subsp. nov. and Propionibacterium acnes type ii as Propionibacterium acnes subsp. defendens subsp. nov. Int. J. Syst. Evol. Microbiol. 66, 5358–5365. doi: 10.1099/ijsem.0.001521
McKenzie, C. M., Seviour, E. M., Schumann, P., Maszenan, A. M., Liu, J.-R., Webb, R. I., et al. (2006). Isolates of ‘Candidatus Nostocoida limicola’ Blackall et al. 2000 should be described as three novel species of the genus Tetrasphaera, as Tetrasphaera jenkinsii sp. nov., Tetrasphaera vanveenii sp. nov. and Tetrasphaera veronensis sp. Int. J. Syst. Evol. Microbiol. 56, 2279–2290. doi: 10.1099/ijs.0.63978-0
Meier, A., Kirschner, P., Schröder, K.-H., Wolters, J., Kroppenstedt, R. M., and Böttger, E. C. (1993). Mycobacterium intermedium sp. nov. Int. J. Syst. Bacteriol. 43, 204–209.
Meier-Kolthoff, J. P., Auch, A. F., Klenk, H.-P., and Göker, M. (2013a). Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinformatics 14:60. doi: 10.1186/1471-2105-14-60
Meier-Kolthoff, J. P., Auch, A. F., Klenk, H.-P., and Göker, M. (2014a). Highly parallelized inference of large genome-based phylogenies. Concurr. Comput. Pract. Exp. 26, 1715–1729. doi: 10.1002/cpe.3112
Meier-Kolthoff, J. P., Göker, M., Spröer, C., and Klenk, H.-P. (2013b). When should a DDH experiment be mandatory in microbial taxonomy? Arch. Microbiol. 195, 413–418. doi: 10.1007/s00203-013-0888-4
Meier-Kolthoff, J. P., Hahnke, R. L., Petersen, J., Scheuner, C., Michael, V., Fiebig, A., et al. (2014b). Complete genome sequence of DSM 30083(T), the type strain (U5/41(T)) of Escherichia coli, and a proposal for delineating subspecies in microbial taxonomy. Stand. Genomic Sci. 9:2. doi: 10.1186/1944-3277-9-2
Meier-Kolthoff, J. P., Klenk, H.-P., and Göker, M. (2014c). Taxonomic use of DNA G+C content and DNA-DNA hybridization in the genomic age. Int. J. Syst. Evol. Microbiol. 64, 352–356. doi: 10.1099/ijs.0.056994-0
Meile, L., Ludwig, W., Rueger, U., Gut, C., Kaufmann, P., Dasen, G., et al. (1997). Bifidobacterium lactis sp. nov., a moderately oxygen tolerant species isolated from fermented milk. Syst. Appl. Microbiol. 20, 57–64.
Meklat, A., Bouras, N., Mokrane, S., Zitouni, A., Schumann, P., Spröer, C., et al. (2015). Bounagaea algeriensis gen. nov., sp. nov., an extremely halophilic actinobacterium isolated from a Saharan soil of Algeria. Antonie Van Leeuwenhoek 108, 473–482. doi: 10.1007/s10482-015-0500-9
Meklat, A., Bouras, N., Zitouni, A., Mathieu, F., Lebrihi, A., Schumann, P., et al. (2013a). Actinopolyspora mzabensis sp. nov., a halophilic actinomycete isolated from an Algerian Saharan soil. Int. J. Syst. Evol. Microbiol. 63, 3787–3792. doi: 10.1099/ijs.0.046649-0
Meklat, A., Bouras, N., Zitouni, A., Mathieu, F., Lebrihi, A., Schumann, P., et al. (2013b). Actinopolyspora righensis sp. nov., a novel halophilic actinomycete isolated from Saharan soil in Algeria. Antonie Van Leeuwenhoek 104, 301–307. doi: 10.1007/s10482-013-9948-7
Meklat, A., Bouras, N., Zitouni, A., Mathieu, F., Lebrihi, A., Schumann, P., et al. (2013c). Actinopolyspora saharensis sp. nov., a novel halophilic actinomycete isolated from a Saharan soil of Algeria. Antonie Van Leeuwenhoek 103, 771–776. doi: 10.1007/s10482-012-9859-z
Merhej, V., Falsen, E., Raoult, D., and Roux, V. (2009). Corynebacterium timonense sp. nov. and Corynebacterium massiliense sp. nov., isolated from human blood and human articular hip fluid. Int. J. Syst. Evol. Microbiol. 59, 1953–1959. doi: 10.1099/ijs.0.005827-0
Mertz, F. P. (1994). Planomonospora alba sp. nov., two new species isolated from soil by baiting techniques. Int. J. Syst. Bacteriol. 44, 274–281.
Mertz, F. P., and Yao, R. C. (1986). Actinomadura oligospora sp. nov. the producer of a new polyether antibiotic. Int. J. Syst. Bacteriol. 36, 179–182.
Mertz, F. P., and Yao, R. C. (1990). Saccharopolyspora spinosa sp. nov. isolated from soil collected in a sugar mill rum still. Int. J. Syst. Bacteriol. 40, 34–39.
Mertz, F. P., and Yao, R. C. (1993). Amycolatopsis alba sp. nov., Isolated from soil. Int. J. Syst. Bacteriol. 43, 715–720.
Mesbah, M., Premachandran, U., and Whitman, W. B. (1989). Precise measurement of the G+C content of deoxyribonucleic acid by high-performance liquid chromatography. Int. J. Syst. Bacteriol. 39, 159–167. doi: 10.1099/00207713-39-2-159
Meyer, J. (1976). Nocardiopsis, a new genus of the order Actinomycetales. Int. J. Syst. Bacteriol. 26, 487–493.
Miller, E. S., Woese, C. R., and Brenner, S. (1991). Description of the erythromycin-producing bacterium Arthrobacter sp. strain NRRL B-3381 as Aeromicrobium erythreum gen. nov., sp. nov. Int. J. Syst. Bacteriol. 41, 363–368.
Minamida, K., Ota, K., Nishimukai, M., Tanaka, M., Abe, A., Sone, T., et al. (2008). Asaccharobacter celatus gen. nov., sp. nov., isolated from rat caecum. Int. J. Syst. Evol. Microbiol. 58, 1238–1240. doi: 10.1099/ijs.0.64894-0
Mingma, R., Duangmal, K., Také, A., Inahashi, Y., Omura, S., Takahashi, Y., et al. (2016). Proposal of Sphaerimonospora cavernae gen. nov., sp. nov. and transfer of Microbispora mesophila (Zhang et al., 1998) to Sphaerimonospora mesophila comb. nov. and Microbispora thailandensi (Duangmal et al., 2012) to Sphaerimonospora thailandensis comb. nov. Int. J. Syst. Evol. Microbiol. 66, 1735–1744. doi: 10.1099/ijsem.0.000935
Mishra, A. K., Lagier, J.-C., Robert, C., Raoult, D., and Fournier, P.-E. (2013). Genome sequence and description of Timonella senegalensis gen. nov., sp nov., a new member of the suborder Micrococcinae. Stand. Genomic Sci. 8, 318–335. doi: 10.4056/sigs.3476977
Mitsuoka, T. (1969). Vergleichende Untersuchungen über die Bifidobakterien aus dem Verdauungstakt von Menschen und Tieren. Zbl. Bakt. Hyg. I Abt. Orig. 210, 52–64.
Miyadoh, S., Amano, S., Tohyama, H., and Shomura, T. (1987). Actinomadura atramentaria, a new species of the Actinomycetales. Int. J. Syst. Bacteriol. 37, 342–346. doi: 10.1099/00207713-37-4-342
Modesto, M., Michelini, S., Stefanini, I., Sandri, C., Spiezio, C., Pisi, A., et al. (2015). Bifidobacterium lemurum sp. nov., from faeces of the ring-tailed lemur (Lemur catta). Int. J. Syst. Evol. Microbiol. 65, 1726–1734. doi: 10.1099/ijs.0.000162
Moe, W. M., Yan, J., Nobre, M. F., da Costa, M. S., and Rainey, F. A. (2009). Dehalogenimonas lykanthroporepellens gen. nov., sp. nov., a reductively dehalogenating bacterium isolated from chlorinated solvent-contaminated groundwater. Int. J. Syst. Evol. Microbiol. 59, 2692–2697. doi: 10.1099/ijs.0.011502-0
Mohagheghi, A., Grohmann, K., Himmel, M., Leighton, L. M., and Updegraff, D. M. (1986). Isolation and characterization of Acidothermus cellulolyticus gen. nov., sp. nov., a new genus of thermophilic, acidophilic, cellulolytic bacteria. Int. J. Syst. Bacteriol. 36, 435–443. doi: 10.1099/00207713-36-3-435
Mohamed, A. M., Iwen, P. C., Tarantolo, S., and Hinrichs, S. H. (2004). Mycobacterium nebraskense sp. nov., a novel slowly growing scotochromogenic species. Int. J. Syst. Evol. Microbiol. 54, 2057–2060. doi: 10.1099/ijs.0.63126-0
Monciardini, P., Cavaletti, L., Schumann, P., Rohde, M., and Donadio, S. (2003). Conexibacter woesei gen. nov., sp. nov., a novel representative of a deep evolutionary line of descent within the class Actinobacteria. Int. J. Syst. Evol. Microbiol. 53, 569–576. doi: 10.1099/ijs.0.02400-0
Montero-Calasanz, M. D. C., Meier-Kolthoff, J. P., Zhang, D.-F., Yaramis, A., Rohde, M., Woyke, T., et al. (2017). Genome-scale data call for a taxonomic rearrangement of Geodermatophilaceae. Front. Microbiol. 8:2501. doi: 10.3389/fmicb.2017.02501
Moreira, A. P. B., Pereira, N., Thompson, F. L., Pereira, N. Jr., and Thompson, F. L. (2011). Usefulness of a real-time PCR platform for G+C content and DNA-DNA hybridization estimations in vibrios. Int. J. Syst. Evol. Microbiol. 61, 2379–2383. doi: 10.1099/ijs.0.023606-0
Morita, H., Nakano, A., Onoda, H., Toh, H., Oshima, K., Takami, H., et al. (2011). Bifidobacterium kashiwanohense sp. nov., isolated from healthy infant faeces. Int. J. Syst. Evol. Microbiol. 61, 2610–2615. doi: 10.1099/ijs.0.024521-0
Muangham, S., Suksaard, P., Mingma, R., Matsumoto, A., Takahashi, Y., Duangmal, K., et al. (2016). Nocardiopsis sediminis sp. nov., isolated from mangrove sediment. Int. J. Syst. Evol. Microbiol. 66, 3835–3840. doi: 10.1099/ijsem.0.001273
Mukherjee, S., Seshadri, R., Varghese, N. J., Eloe-Fadrosh, E. A., Meier-Kolthoff, J. P., Göker, M., et al. (2017). 1,003 reference genomes of bacterial and archaeal isolates expand coverage of the tree of life. Nat. Biotechnol. 35, 676–685. doi: 10.1038/nbt.3886
Mun, H.-S., Park, J.-H., Kim, H., Yu, H.-K., Park, Y.-G., Cha, C.-Y., et al. (2008). Mycobacterium senuense sp. nov., a slowly growing, non-chromogenic species closely related to the Mycobacterium terrae complex. Int. J. Syst. Evol. Microbiol. 58, 641–646. doi: 10.1099/ijs.0.65374-0
Murcia, M. I., Tortoli, E., Menendez, M. C., Palenque, E., and Garcia, M. J. (2006). Mycobacterium colombiense sp. nov., a novel member of the Mycobacterium avium complex and description of MAC-X as a new ITS genetic variant. Int. J. Syst. Evol. Microbiol. 56, 2049–2054. doi: 10.1099/ijs.0.64190-0
Nagai, A., Khan, S. T., Tamura, T., Takagi, M., and Shin-Ya, K. (2011). Streptomyces aomiensis sp. nov., isolated from a soil sample using the membrane-filter method. Int. J. Syst. Evol. Microbiol. 61, 947–950. doi: 10.1099/ijs.0.020719-0
Nakajima, Y., Ho, C. C., and Kudo, T. (2003). Microtetraspora malaysiensis sp. nov., isolated from Malaysian primary dipterocarp forest soil. J. Gen. Appl. Microbiol. 49, 181–189. doi: 10.2323/jgam.49.181
Nakamura, G. (1961). Studies on antibiotic actinomycetes. III. On Streptomyces producing A-ß-D-ribofuranosylpurine. J. Antibiot. Ser. A 14, 94–97.
Nakamura, K., Hiraishi, A., Yoshimi, Y., Kawaharasaki, M., Masuda, K., and Kamagata, Y. (1995). Microlunatus phosphovorus gen. nov., sp. nov., a new gram-positive polyphosphate-accumulating bacterium isolated from activated sludge. Int. J. Syst. Bacteriol. 45, 17–22.
Nesterenko, O. A., Kvasnikov, E. I., and Nogina, T. M. (1985). Nocardioidaceae fam. nov., a new family of the order Actinomycetales Buchanan 1917. Mikrobiol. Zhurnal 47, 3–12.
Neubauer, M., Šourek, J., Rýc, M., Boháček, J., Mára, M., and Mnuková, J. (1991). Corynebacterium accolens sp. nov., a Gram-positive rod exhibiting satellitism, from clinical material. Syst. Appl. Microbiol. 14, 46–51. doi: 10.1016/S0723-2020(11)80360-7
Nikolaitchouk, N., Hoyles, L., Falsen, E., Grainger, J. M., and Collins, M. D. (2000). Characterization of Actinomyces isolates from samples from the human urogenital tract: description of Actinomyces urogenitalis sp. nov. Int. J. Syst. Evol. Microbiol. 50, 1649–1654. doi: 10.1099/00207713-50-4-1649
Nikou, M. M., Ramezani, M., Amoozegar, M. A., Fazeli, S. A. S., Schumann, P., Spröer, C., et al. (2014). Alloactinosynnema iranicum sp. nov., a rare actinomycete isolated from a hypersaline wetland, and emended description of the genus Alloactinosynnema. Int. J. Syst. Evol. Microbiol. 64, 1173–1179. doi: 10.1099/ijs.0.049189-0
Nishimura, H., Kimura, T., and Kuroya, M. (1953). On a yellow crystalline antibiotic, identical with aureothricin isolated from a new species of Streptomyces, 39a, and its taxonomic study. J. Antibiot. Ser. A 6, 57–65.
Nishimura, H., Kimura, T., Tawara, K., Sasaki, K., Nakajima, K., Shimaoka, N., et al. (1957). Aburamycin, a new antibiotic. J. Antibiot. Ser. A 10, 205–212.
Nogueira, C. L., Whipps, C. M., Matsumoto Kayoko, C., Chimara, E., Droz, S., Tortoli, E., et al. (2015). Mycobacterium saopaulense sp. nov., a rapidly growing mycobacterium closely related to members of the Mycobacterium chelonae – Mycobacterium abscessus group. Int. J. Syst. Evol. Microbiol. 65, 4403–4409. doi: 10.1099/ijsem.0.000590
Nonomura, H., and Ohara, Y. (1957). Distribution of Actinomycetes in the soil. II. Microbispora, a new genus of the Streptomycetaceae. J. Ferment. Technol. 35, 307–3011.
Nonomura, H., and Ohara, Y. (1960). Distribution of the Actinomycetes in soil. IV. The isolation and classification of the genus Streptosporangium. J. Ferment. Technol. 38, 405–409.
Nonomura, H., and Ohara, Y. (1971a). Distribution of Actinomycetes in soil. IX. New species of the genera Microbispora and Microtetaspora, and their isolation method. J. Ferment. Technol. 49, 887–894.
Nonomura, H., and Ohara, Y. (1971b). Distribution of Actinomycetes in soil. X. New genus and species of monosporic actinomycetes in soil. J. Ferment. Technol. 49, 895–903.
Normand, P. (2006). Geodermatophilaceae fam. nov., a formal description. Int. J. Syst. Evol. Microbiol. 56, 2277–2278. doi: 10.1099/ijs.0.64298-0
Normand, P., and Benson, D. R. (2012). Order VI. Frankiales ord. nov., in Bergey's Manual of Systematic Bacteriology, Vol. 5, The Actinobacteria, eds M. Goodfellow, P. Kämpfer, H.-J. Busse, M. E. Trujillo, K. Suzuki, W. Ludwig, et al. (New York, NY: Springer), 508.
Nouioui, I., Ghodhbane-Gtari, F., Montero-Calasanz, M. D., Göker, M., Meier-Kolthoff, J. P., Schumann, P., et al. (2016). Proposal of a type strain for Frankia alni (Woronin 1866) Von Tubeuf 1895, emended description of Frankia alni, and recognition of Frankia casuarinae sp. nov. and Frankia elaeagni sp. nov. Int. J. Syst. Evol. Microbiol. 66, 5201–5210. doi: 10.1099/ijsem.0.001496
O'Donnell, A. G., Goodfellow, M., and Minnikin, D. E. (1982). Lipids in the classification of Nocardioides: reclassification of Arthrobacter simples (Jensen) Lochhead in the genus Nocardioides (Prauser) emend. O'Donnell et al. as Nocardioides simplex comb. nov. Arch. Microbiol. 133, 323–329.
Oh, E.-J., Bae, C., Lee, H.-B., Hwang, I. S., Lee, H.-I., Yea, M. C., et al. (2016). Clavibacter michiganensis subsp. capsici subsp. nov., causing bacterial canker disease in pepper. Int. J. Syst. Evol. Microbiol. 66, 4065–4070. doi: 10.1099/ijsem.0.001311
Okazaki, M., Ohkusu, K., Hata, H., Ohnishi, H., Sugahara, K., Kawamura, C., et al. (2009). Mycobacterium kyorinense sp. nov., a novel, slow-growing species, related to Mycobacterium celatum, isolated from human clinical specimens. Int. J. Syst. Evol. Microbiol. 59, 1336–1341. doi: 10.1099/ijs.0.000760-0
Okoro, C. K., Bull, A. T., Mutreja, A., Rong, X., Huang, Y., and Goodfellow, M. (2010). Lechevalieria atacamensis sp. nov., Lechevalieria deserti sp. nov. and Lechevalieria roselyniae sp. nov., isolated from hyperarid soils. Int. J. Syst. Evol. Microbiol. 60, 296–300. doi: 10.1099/ijs.0.009985-0
Olson, J. B., Harmody, D. K., Bej, A. K., and Mccarthy, P. J. (2007). Tsukamurella spongiae sp. nov., a novel actinomycete isolated from a deep-water marine sponge. Int. J. Syst. Evol. Microbiol. 57, 1478–1481. doi: 10.1099/ijs.0.64837-0
Omura, S., Takahashi, Y., Iwai, Y., and Tanaka, H. (1982). Kitasatosporia, a new genus of the order Actinomycetales. J. Antibiot. 35, 1013–1019.
Orla-Jensen, S. (1909). Die Hauptlinien des natürlichen Bakteriensystems. Zbl. Bakt. Hyg. II Abt. 22, 305–346.
Ørskov, J. (1923). Investigations into the Morphology of the Ray Fungi. Copenhagen: Levin and Munksgaard.
Osman, S., Moissl, C., Hosoya, N., Briegel, A., Mayilraj, S., Satomi, M., et al. (2007). Tetrasphaera remsis sp. nov., isolated from the regenerative enclosed life support module simulator (REMS) air system. Int. J. Syst. Evol. Microbiol. 57, 2749–2753. doi: 10.1099/ijs.0.65137-0
Otsuka, Y., Kawamura, Y., Koyama, T., Iihara, H., Ohkusu, K., and Ezaki, T. (2005). Corynebacterium resistens sp. nov., a new multidrug-resistant coryneform bacterium isolated from human infections. J. Clin. Microbiol. 43, 3713–3717. doi: 10.1128/JCM.43.8.3713
Paradis, E., Claude, J., and Strimmer, K. (2004). APE: analyses of phylogenetics and evolution in R language. Bioinformatics 20, 289–290. doi: 10.1093/bioinformatics/btg412
Park, E.-J., Roh, S. W., Kim, M.-S., Jung, M.-J., Shin, K.-S., and Bae, J.-W. (2010). Kocuria koreensis sp. nov., isolated from fermented seafood. Int. J. Syst. Evol. Microbiol. 60, 140–143. doi: 10.1099/ijs.0.012310-0
Park, M.-H., Traiwan, J., Jung, M. Y., and Kim, W. (2012). Gulosibacter chungangensis sp. nov., an actinomycete isolated from a marine sediment, and emended description of the genus Gulosibacter. Int. J. Syst. Evol. Microbiol. 62, 1055–1060. doi: 10.1099/ijs.0.032268-0
Park, S., Jung, Y.-T., Won, S.-M., and Yoon, J.-H. (2016). Demequina litorisediminis sp. nov., isolated from a tidal flat, and emended description of the genus Demequina. Int. J. Syst. Evol. Microbiol. 66, 4197–4203. doi: 10.1099/ijsem.0.001335
Park, S.-K., Kim, M.-S., Jung, M.-J., Nam, Y.-D., Park, E.-J., Roh, S. W., et al. (2011). Brachybacterium squillarum sp. nov., isolated from salt-fermented seafood. Int. J. Syst. Evol. Microbiol. 61, 1118–1122. doi: 10.1099/ijs.0.022517-0
Park, S. C., Baik, K. S., Kim, M. S., Chun, J., and Seong, C. N. (2008). Nocardioides dokdonensis sp. nov., an actinomycete isolated from sand sediment. Int. J. Syst. Evol. Microbiol. 58, 2619–2623. doi: 10.1099/ijs.0.65835-0
Park, S. W., Kim, S. M., Park, S. T., and Kim, Y. M. (2009). Tsukamurella carboxydivorans sp. nov., a carbon monoxide-oxidizing actinomycete. Int. J. Syst. Evol. Microbiol. 59, 1541–1544. doi: 10.1099/ijs.0.005959-0
Park, Y.-H., Suzuki, K.-I., Yim, D.-G., Lee, K.-C., Kim, E., Yoon, J.-S., et al. (1993). Suprageneric classification of peptidoglycan group B actinomycetes by nucleotide sequencing of 5S ribosomal RNA. Antonie Van Leeuwenhoek 64, 307–313.
Park, Y.-H., Yoon, J.-H., Shin, Y. K., Suzuki, K., Kudo, T., Seino, A., et al. (1999). Classification of “Nocardioides fulvus” IFO 14399 and Nocardioides sp. ATCC 39419 in Kribbella gen. nov., as Kribbella flavida sp. nov. and Kribbella sandrarnycini sp. nov. Int. J. Syst. Bacteriol. 49, 743–752.
Parker, C. T., Tindall, B. J., and Garrity, G. M. (2015). International code of nomenclature of prokaryotes. Int. J. Syst. Evol. Microbiol. doi: 10.1099/ijsem.0.000778. [Epub ahead of print].
Patrick, S., and McDowell, A. (2012). Order XII. Propionibacteriales ord. nov., in Bergey's Manual of Systematic Bacteriology, Vol. 5, The Actinobacteria, eds M. Goodfellow, P. Kämpfer, H.-J. Busse, M. E. Trujillo, K. Suzuki, W. Ludwig, et al. (New York, NY: Springer), 1137.
Phillips, R. W., Wiegel, J., Berry, C. J., Fliermans, C., Peacock, A. D., White, D. C., et al. (2002). Kineococcus radiotolerans sp. nov., a radiation-resistant, Gram-positive bacterium. Int. J. Syst. Evol. Microbiol. 52, 933–938. doi: 10.1099/ijs.0.02029-0
Pierson, B. K., and Castenholz, R. W. (1974). A phototrophic gliding filamentous bacterium of hot springs, Chloroflexus aurantiacus, gen. and sp. nov. Arch. Microbiol. 100, 5–24.
Pikuta, E. V., Lyu, Z., Williams, M. D., Patel, N. B., Liu, Y., Hoover, R. B., et al. (2017). Sanguibacter gelidistatuariae sp. nov., a novel psychrotolerant anaerobe from an ice sculpture in Antarctica, and emendation of descriptions of the family Sanguibacteraceae, the genus Sanguibacter and species S. antarcticus, S. inulinus, S. kedie. Int. J. Syst. Evol. Microbiol. 67, 1442–1450. doi: 10.1099/ijsem.0.001838
Ping, X., Takahashi, Y., Seino, A., Iwai, Y., and Omura, S. (2004). Streptomyces scabrisporus sp. nov. Int. J. Syst. Evol. Microbiol. 54, 577–581. doi: 10.1099/ijs.0.02692-0
Pinnert-Sindico, M. S. (1954). Une nouvelle epèce de Streptomyces productrice d'antibiotiques: Streptomyces ambofaciens n. sp. caractères culturaux. Ann. Inst. Pasteur 87, 702–707.
Pitcher, D., Soto, A., Soriano, F., and Valero-Guillén, P. (1992). Classification of coryneform bacteria associated with human urinary tract infection (Group D2) as Corynebacterium urealyticum sp. nov. Int. J. Syst. Bacteriol. 42, 178–181.
Prabhu, D. M., Quadri, S. R., Cheng, J., Liu, L., Chen, W., Yang, Y., et al. (2015). Sinomonas mesophila sp. nov., isolated from ancient fort soil. J. Antibiot. 68, 318–321. doi: 10.1038/ja.2014.161
Prauser, H. (1976). Nocardioides, a new genus of the order Actinomycetales. Int. J. Syst. Bacteriol. 26, 58–65.
Prauser, H., Lechevalier, M. P., and Lechevalier, H. (1970). Description of Oerskovia gen. n. to harbor Ørskov's motile nocardia. Appl. Microbiol. 19:534.
Prévot, A. R. (1940). Manuel de Classification et de Détermination des Bactéries Anaérobies, 1st Edn. Paris: Masson et Cie.
Pridham, T. G. (1970). New names and new combinations in the order Actinomycetales Buchanan 1917. Bull. United States Dep. Agric. 1424, 1–55.
Pridham, T. G., Hesseltine, C. W., and Benedict, R. G. (1958). A guide for the classification of Streptomycetes according to selected groups: placement of strains in morphological sections. Appl. Microbiol. 6, 52–79.
Prissick, F. H., and Masson, A. M. (1956). Cervical lymphadentis in children caused by chromogenic mycobacteria. Can. Med. Assoc. J. 75, 798–803.
Prosser, B. L. T., and Palleroni, N. (1976). Streptomyces longwoodensis sp. nov. Int. J. Syst. Bacteriol. 26, 319–322.
Pukall, R., Schumann, P., Schütte, C., Gols, R., and Marcel, D. (2006). Acaricomes phytoseiuli gen. nov., sp. nov., isolated from the predatory mite Phytoseiulus persimilis. Int. J. Syst. Evol. Microbiol. 56, 465–469. doi: 10.1099/ijs.0.63930-0
Qin, S., Li, J., Zhang, Y., Zhu, W., Zhao, G., Xu, L., et al. (2009a). Plantactinospora mayteni gen. nov., sp. nov., a member of the family Micromonosporaceae. Int. J. Syst. Evol. Microbiol. 59, 2527–2533. doi: 10.1099/ijs.0.010793-0
Qin, S., Zhao, G. Z., Li, J., Zhu, W. Y., Xu, L. H., and Li, W. J. (2009b). Actinomadura flavalba sp. nov., an endophytic actinomycete isolated from leaves of Maytenus austroyunnanensis. Int. J. Syst. Evol. Microbiol. 59, 2453–2457. doi: 10.1099/ijs.0.010652-0
Quintana, E. T., Trujillo, M. E., and Goodfellow, M. (2003). Actinomadura mexicana and Actinomadura meyerii sp. nov., two novel soil Sporoactinomycetes. Syst. Appl. Microbiol. 26, 511–517. doi: 10.1078/072320203770865800
R Development Core Team (2014). R: A Language and Environment for Statistical Computing. Vienna: R Development Core Team.
Rainey, F. A., Burghardt, J., Kroppenstedt, R., Klatte, S., and Stackebrandt, E. (1995a). Polyphasic evidence for the transfer of Rhodococcus roseus to Rhodococcus rhodochrous. Int. J. Syst. Bacteriol. 45, 101–103.
Rainey, F. A., Klatte, S., Kroppenstedt, R. M., and Stackebrandt, E. (1995b). Dietzia, a new genus including Dietzia maris comb. nov., formerly Rhodococcus maris. Int. J. Syst. Bacteriol. 45, 32–36.
Rainey, F. A., Ward-Rainey, N., Kroppenstedt, R. M., and Stackebrandt, E. (1996). The genus Nocardiopsis represents a phylogenetically coherent taxon and a distinct actinomycete lineage: proposal of Nocardiopsaceae fam. nov. Int. J. Syst. Bacteriol. 46, 1088–1092. doi: 10.1099/00207713-46-4-1088
Ramos, C. P., Foster, G., and Collins, M. D. (1997). Phylogenetic analysis of the genus Actinomyces based on 16S rRNA gene sequences: description of Arcanobacterium phocae sp. nov., Arcanobacterium bernardiae comb. nov. and, Arcanobacterium pyogenes comb. nov. Int. J. Syst. Bacteriol. 47, 46–53.
Ramos, J. P., Dias Campos, C. E., de Souza Caldas, P. C., Ferreira, N. V., Boas da Silva, M. V., Redner, P., et al. (2013). Mycobacterium fragae sp. nov., a non-chromogenic species isolated from human respiratory specimens. Int. J. Syst. Evol. Microbiol. 63, 2583–2587. doi: 10.1099/ijs.0.046862-0
Rao, J. U., Rash, B. A., Nobre, M. F., da Costa, M. S., Rainey, F. A., and Moe, W. M. (2012). Actinomyces naturae sp. nov., the first Actinomyces sp. isolated from a non-human oranimal source. Antonie Van Leeuwenhoek 101, 155–168. doi: 10.1007/s10482-011-9644-4
Reddy, G. S., and Garcia-Pichel, F. (2009). Description of Patulibacter americanus sp. nov., isolated from biological soil crusts, emended description of the genus Patulibacter Takahashi et al. 2006 and proposal of Solirubrobacterales ord. nov. and Thermoleophilales ord. nov. Int. J. Syst. Evol. Microbiol. 59, 87–94. doi: 10.1099/ijs.0.64185-0
Reddy, G. S., Prakash, J. S. S., Srinivas, R., Matsumoto, G. I., and Shivaji, S. (2003). Leifsonia rubra sp. nov. and Leifsonia aurea sp. nov., psychrophiles from a pond in Antarctica. Int. J. Syst. Evol. Microbiol. 53, 977–984. doi: 10.1099/ijs.0.02396-0
Rehfuss, M., and Urban, J. (2005). Rhodococcus phenolicus sp. nov., a novel bioprocessor isolated actinomycete with the ability to degrade chlorobenzene, dichlorobenzene and phenol as sole carbon sources. Syst. Appl. Microbiol. 28, 695–701. doi: 10.1016/j.syapm.2005.05.011
Reichert, K., Lipski, A., Pradella, S., Stackebrandt, E., and Altendorf, K. (1998). Pseudonocardia asaccharolytica sp. nov. and Pseudonocardia sulfidoxydans sp. nov. new dimethyl disulfide-degrading actinomycetes and emended description of the genus Pseudonocardia. Int. J. Syst. Bacteriol. 48, 441–449.
Reischl, U., Emler, S., Horak, Z., Kaustova, J., Kroppenstedt, R. M., Lehn, N., et al. (1998). Mycobacterium bohernicum sp. nov., a new slow-growing scotochromogenic mycobacterium. Int. J. Syst. Bacteriol. 48, 1349–1355.
Reischl, U., Melzl, H., Kroppenstedt, R. M., Miethke, T., Naumann, L., Mariottini, A., et al. (2006). Mycobacterium monacense sp. nov. Int. J. Syst. Evol. Microbiol. 56, 2575–2578. doi: 10.1099/ijs.0.64527-0
Ren, J., Li, L., Wei, B., Tang, Y. L., Deng, Z. X., Sun, M., et al. (2013). Micromonospora wenchangensis sp. nov., isolated from mangrove soil. Int. J. Syst. Evol. Microbiol. 63, 2389–2395. doi: 10.1099/ijs.0.045476-0
Renvoise, A., Didier, R., and Véronique, R. (2009). Actinomyces massiliensis sp. nov., isolated from a patient blood culture. Int. J. Syst. Evol. Microbiol. 59, 540–544. doi: 10.1099/ijs.0.001503-0
Renvoise, A., Raoult, D., and Véronique, R. (2010). Actinomyces timonensis sp. nov., isolated from a human clinical osteo-articular sample. Int. J. Syst. Evol. Microbiol. 60, 1516–1521. doi: 10.1099/ijs.0.012914-0
Reuter, G. (1963). Vergleichende Untersuchungen über die Bifidus-Flora im Säuglings und Erwachsenenstuhl. Zbl. Bakt. Hyg. I Abt. Orig. 191, 486–507.
Rheims, H., Schumann, P., Rohde, M., and Stackebrandt, E. (1998). Verrucosispora gifhornensis gen. nov., sp. nov., a new member of the actinobacterial family Micromonosporaceae. Int. J. Syst. Bacteriol. 48, 1119–1127.
Rhodes, M. W., Kator, H., McNabb, A., Deshayes, C., Reyrat, J. M., Brown-Elliott, B. A., et al. (2005). Mycobacterium pseudoshottsii sp. nov., a slowly growing chromogenic species isolated from Chesapeake Bay striped bass (Morone saxatilis). Int. J. Syst. Evol. Microbiol. 55, 1139–1147. doi: 10.1099/ijs.0.63343-0
Riegel, P., Briel, D., De Prévost, G., Jehl, F., Monteil, H., and Minck, R. (1993). Taxonomic study of Corynebacterium group ANF-1 strains: proposal of Corynebacterium afementans sp. nov, containing the subspecies C. afermentans subsp. afermentans subsp. nov. and C. afermentans subsp. lipophilum subsp. nov. Int. J. Syst. Bacteriol. 43, 287–292.
Riegel, P., Ruimy, R., Briel, D., De Prevost, G., Jehl, F., Bimet, F., et al. (1995a). Corynebacterium argentoratense sp. nov., from the human throat. Int. J. Syst. Bacteriol. 45, 533–537.
Riegel, P., Ruimy, R., de Briel, D., Prévost, G., Jehl, F., Bimet, F., et al. (1995b). Corynebacterium seminale sp. nov., a new species associated with genital infections in male patients. J. Clin. Microbiol. 33, 2244–2249.
Riegel, P., Ruimy, R., Renaud, F. N. R., Freney, J., Prevost, G., Jehl, F., et al. (1997). Corynebacterium singulare sp. nov., a new species for urease-positive strains related to Corynebacterium minutissimum. Int. J. Syst. Bacteriol. 47, 1092–1096.
Rocha, E. P., and Danchin, A. (2002). Base composition bias might result from competition for metabolic resources. Trends Genet. 18, 291–294. doi: 10.1016/S0168-9525(02)02690-2
Rocourt, J., Wehmeyer, U., and Stackebrandt, E. (1987). Transfer of Listeria dentrificans to a new genus, Jonesia gen. nov., as Jonesia denitrificans comb. nov. Int. J. Syst. Bacteriol. 37, 266–270. doi: 10.1099/00207713-37-3-266
Rodriguez Jovita, M., Collins, M. D., Sjödén, B., and Falsen, E. (1999). Characterization of a novel Atopobium isolate from the human vagina: description of Atopobium vaginae sp. nov. Int. J. Syst. Bacteriol. 49, 1573–1576. doi: 10.1099/00207713-49-4-1573
Rong, X., Guo, Y., and Huang, Y. (2009). Proposal to reclassify the Streptomyces albidoflavus clade on the basis of multilocus sequence analysis and DNA–DNA hybridization, and taxonomic elucidation of Streptomyces griseus subsp. solvifaciens. Syst. Appl. Microbiol. 32, 314–322. doi: 10.1016/j.syapm.2009.05.003
Rosselló-Mora, R., and Amann, R. (2001). The species concept for prokaryotes. FEMS Microbiol. Rev. 25, 39–67. doi: 10.1016/S0168-6445(00)00040-1
Roux, V., and Raoult, D. (2009). Brevibacterium massiliense sp. nov., isolated from a human ankle discharge. Int. J. Syst. Evol. Microbiol. 59, 1960–1964. doi: 10.1099/ijs.0.007864-0
Rowbotham, T. J., and Cross, T. (1977). Rhodococcus coprophilus sp. nov.: an aerobic nocardioform Actinomycete belonging to the “rodochrous” complex. J. Gen. Microbiol. 100, 123–138.
Ruan, J.-S., Al-Tai, A. M., Zhou, Z.-H., and Qu, L.-H. (1994). Actinopolyspora iraqiensis sp. nov., a new halophilic actinomycete isolated from soil. Int. J. Syst. Bacteriol. 44, 759–763.
Ruckmani, A., Kaur, I., Schumann, P., Klenk, H.-P., and Mayilraj, S. (2011). Calidifontibacter indicus gen. nov., sp. nov., a member of the family Dermacoccaceae isolated from a hot spring, and emended description of the family Dermacoccaceae. Int. J. Syst. Evol. Microbiol. 61, 2419–2424. doi: 10.1099/ijs.0.025593-0
Runmao, H. U. (1987). Saccharomonospora azurea sp. nov. a new species from soil. Int. J. Syst. Bacteriol. 37, 60–61.
Runmao, H. U., Lin, C., and Guizhen, W. (1988). Saccharomonospora cyanea sp. nov. Int. J. Syst. Bacteriol. 38, 444–446.
Saddler, G. S., Goodfellow, M., Minnikin, D. E., and O'Donnell, A. G. (1986). Influence of the growth cycle on the fatty acid and menaquinone composition of Streptomyces cyaneus NCIB 9616. J. Appl. Bacteriol. 60, 51–56.
Sahraoui, N., Ballif, M., Zelleg, S., Yousfi, N., Ritter, C., Friedel, U., et al. (2011). Mycobacterium algericum sp. nov., a novel rapidly growing species related to the Mycobacterium terrae complex and associated with goat lung lesions. Int. J. Syst. Evol. Microbiol. 61, 1870–1874. doi: 10.1099/ijs.0.024851-0
Saito, H., Iwamoto, T., Ohkusu, K., Otsuka, Y., Akiyama, Y., Sato, S., et al. (2011). Mycobacterium shinjukuense sp. nov., a slowly growing, non-chromogenic species isolated from human clinical specimens. Int. J. Syst. Evol. Microbiol. 61, 1927–1932. doi: 10.1099/ijs.0.025478-0
Sakamoto, J. M. J., Kondo, S., Yumoto, H., and Arishina, M. (1962). Bundlins A and B, two antibiotics produced by Streptomyces griseofuscus nov. sp. J. Antibiot. Ser. A 15, 98–102.
Saker, R., Bouras, N., Zitouni, A., Ghoul, M., Rohde, M., Schumann, P., et al. (2014). Mzabimyces algeriensis gen. nov., sp. nov., a halophilic filamentous actinobacterium isolated from a Saharan soil, and proposal of Mzabimycetaceae fam. nov. Antonie Van Leeuwenhoek 106, 1021–1030. doi: 10.1007/s10482-014-0271-8
Sakiyama, Y., Giang, N. M., Miyadoh, S., Luong, D. T., Van Hop, D., and Ando, K. (2014). Streptomyces catbensis sp. nov., isolated from soil. Int. J. Syst. Evol. Microbiol. 64, 2146–2151. doi: 10.1099/ijs.0.048264-0
Sanders, J. E., and Fryer, J. L. (1980). Renibacterium salmoninarum gen. nov., sp. nov., the causative agent of bacterial kidney disease in salmonid fishes. Int. J. Syst. Bacteriol. 30, 496–502.
Sangal, V., Goodfellow, M., Jones, A. L., Schwalbe, E. C., Blom, J., Hoskisson, P. A., et al. (2016). Next-generation systematics: an innovative approach to resolve the structure of complex prokaryotic taxa. Sci. Rep. 6:38392. doi: 10.1038/srep38392
Sarmin, N. I., Tan, G. Y. A., Franco, C. M. M., Edrada-Ebel, R., Latip, J., and Zin, N. M. (2013). Streptomyces kebangsaanensis sp. nov., an endophytic actinomycete isolated from an ethnomedicinal plant, which produces phenazine-1-carboxylic acid. Int. J. Syst. Evol. Microbiol. 63, 3733–3738. doi: 10.1099/ijs.0.047878-0
Sasaki, J., Chijimatsu, M., and Suzuki, K. (1998). Taxonomic significance of 2,4-diaminobutyric acid isomers in the cell wall peptidoglycan of actinomycetes and reclassification of Clavibacter toxicus as Rathayibacter toxicus comb. nov. Int. J. Syst. Bacteriol. 48, 403–410.
Scardovi, V., and Crociani, F. (1974). Bifidobacterium catenulatum, Bifidobacterium dentium, and Bifidobacterium angulatum: three new species and their deoxyribonucleic acid homology relationships. Int. J. Syst. Bacteriol. 24, 6–20. doi: 10.1099/00207713-24-1-6
Scardovi, V., and Trovatelli, L. D. (1969). New species of bifidobacteria from Apis mellifica L. and Apis indica F. A contribution to the taxonomy and biochemistry of the genus Bifidobacterium. Zbl. Bakt. Hyg. II Abt. 123, 64–88.
Scardovi, V., Trovatelli, L. D., Biavati, B., and Zani, G. (1979). Bifidobacterium cuniculi, Bifidobacterium choerinum, Bifidobacterium boum, and Bifidobacterium pseudocatenulatum: four new species and their deoxyribonucleic acid homology relationships. Int. J. Syst. Bacteriol. 29, 291–311.
Scardovi, V., and Zani, G. (1974). Bifidobacterium magnum sp. nov., a large, acidophilic Bifidobacterium isolated from rabbit feces. Int. J. Syst. Bacteriol. 24, 29–34.
Schaal, K. P., and Gatzer, R. (1985). Serological and numerical phenetic classification of clinically significant fermentative actinomycetes, in Filamentous Microorganisms. Biomedical Aspects, ed K. Arai (Tokyo: Japan Scientific Societies Press), 85–109.
Schaal, K. P., and Schofield, G. (1984). Classification and identification of clinically significant Actinomycetaceae, in Biological, Biochemical, and Biomedical Aspects of Actinomycetes, eds B. Ortiz-Ortiz and Yakoleff (Orlando, FL; New York, NY; London: Academic Press), 505–520.
Schaal, K. P., and Yassin, A. A. (2012a). Genus I. Actinomyces Harz 1877, 133 AL emend. Georg, Pine and Gerencser 1969, 292 VP, in Bergey's Manual of Systematic Bacteriology, Vol. 5, The Actinobacteria, eds M. Goodfellow, P. Kämpfer, H.-J. Busse, M. E. Trujillo, K. Suzuki, W. Ludwig, et al. (New York, NY: Springer), 42–114.
Schaal, K. P., and Yassin, A. F. (2012b). Family I. Actinomycetaceae Buchanan 1918, 403 (emend. Stackebrandt, Rainey and Ward-Rainey 1997, 484), emend. Zhi, Li and Stackebrandt 2009, 594 VP, in Bergey's Manual of Systematic Bacteriology, Vol. 5, The Actinobacteria, eds. M. Goodfellow, P. Kämpfer, H.-J. Busse, M. E. Trujillo, K. Suzuki, W. Ludwig, et al. (New York, NY: Springer), 36.
Schaal, K. P., Yassin, A. F., and Stackebrandt, E. (2006). The family Actinomycetaceae: the genera Actinomyces, Actinobaculum, Arcanobacterium, Varibaculum, and Mobiluncus, in The Prokaryotes. eds M. Dworkin, S. Falkow, E. Rosenberg, K. H. Schleifer, and E. Stackebrandt (New York, NY: Springer), 430–537. doi: 10.1007/0-387-30743-5
Schinsky, M. F., McNeil, M. M., Whitney, A. M., Steigerwalt, A. G., Lasker, B. A., Floyd, M. M., et al. (2000). Mycobacterium septicum sp. nov., a new rapidly growing species associated with catheter-related bacteraemia. Int. J. Syst. Evol. Microbiol. 50, 575–581. doi: 10.1099/00207713-50-2-575
Schinsky, M. F., Morey, R. E., Steigerwalt, A. G., Douglas, M. P., Wilson, R. W., Floyd, M. M., et al. (2004). Taxonomic variation in the Mycobacterium fortuitum third biovariant complex: description of Myobacterium boenickei sp. nov., Mycobacterium houstonense sp. nov., Mycobacterium neworleansense sp. nov. and Mycobacterium brisbanense sp. nov. and recognition of Mycobacterium porcinum from human clinical isolates. Int. J. Syst. Evol. Microbiol. 54, 1653–1667. doi: 10.1099/ijs.0.02743-0
Schippers, A., Bosecker, K., Spröer, C., and Schumann, P. (2005). Microbacterium oleivorans sp. nov. and Microbacterium hydrocarbonoxydans sp. nov., novel crude-oil-degrading Gram-positive bacteria. Int. J. Syst. Evol. Microbiol. 55, 655–660. doi: 10.1099/ijs.0.63305-0
Schleifer, K. H., and Kandler, O. (1972). Peptidoglycan types of bacterial cell walls and their taxonomic implications. Bacteriol. Rev. 36, 407–477.
Schofield, G. M., and Schaal, K. P. (1981). A numerical taxonomic study of members of the Actinomycetaceae and related taxa. J. Gen. Microbiol. 127, 237–259. doi: 10.1099/00221287-127-2-237
Scholz, C. F., and Kilian, M. (2016). The natural history of cutaneous propionibacteria, and reclassification of selected species within the genus Propionibacterium to the proposed novel genera Acidipropionibacterium gen. nov., Cutibacterium gen. nov. and Pseudopropionibacterium gen. nov. Int. J. Syst. Evol. Microbiol. 66, 4422–4432. doi: 10.1099/ijsem.0.001367
Schröder, K.-H., Naumann, L., Kroppenstedt, R. M., and Reischl, U. (1997). Mycobacterium hassiacum sp. nov., a new rapidly growing thermophilic Mycobacterium. Int. J. Syst. Bacteriol. 47, 86–91.
Schumann, P., Prauser, H., Rainey, F. A., Stackebrandt, E., and Hirsch, P. (1997). Friedmanniella antarctica gen. nov., sp. nov., an LL-Diaminopimelic acid-containing Actinomycete from Antarctic Sandstone. Int. J. Syst. Bacteriol. 47, 278–283.
Schumann, P., Weiss, N., and Stackebrandt, E. (2001). Reclassification of Cellulomonas cellulans (Stackebrandt and Keddie 1986) as Cellulosimicrobium cellulans gen. nov., comb. nov. Int. J. Syst. Evol. Microbiol. 51, 1007–1010. doi: 10.1099/00207713-51-3-1007
Schwabacher, H. (1959). A strain of Mycobacterium isolated from skin lesions of a cold-blooded animal, Xenopus laevis, and its relaton to atypical acid-fast bacilli occurring in man. J. Hyg. (Lond). 57, 57–67.
Sekiguchi, Y., Yamada, T., Hanada, S., Ohashi, A., Harada, H., and Kamagata, Y. (2003). Anaerolinea thermophila gen. nov., sp. nov. and Caldilinea aerophila gen. nov., sp. nov., novel filamentous thermophiles that represent a previously uncultured lineage of the domain Bacteria at the subphylum level. Int. J. Syst. Evol. Microbiol. 53, 1843–1851. doi: 10.1099/ijs.0.02699-0
Sen, A., Daubin, V., Abrouk, D., Gifford, I., Berry, A. M., and Normand, P. (2014). Phylogeny of the class Actinobacteria revisited in the light of complete genomes. The orders “Frankiales” and Micrococcales should be split into coherent entities: proposal of Frankiales ord. nov., Geodermatophilales ord. nov., Acidothermales ord. nov. an. Int. J. Syst. Evol. Microbiol. 64, 3821–3832. doi: 10.1099/ijs.0.063966-0
Seong, C. N., Kim, Y. S., Baik, K. S., Choi, S. K., Kim, M. B., Kim, S. B., et al. (2003). Tsukamurella sunchonensis sp. nov., a bacterium associated with foam in activated sludge. J. Microbiol. 41, 83–88.
Shahraki, A. H., Çavuşoglu, C., Borroni, E., Heidarieh, P., Koksalan, O. K., Cabibbe, A. M., et al. (2015). Mycobacterium celeriflavum sp. nov., a rapidly growing scotochromogenic bacterium isolated from clinical specimens. Int. J. Syst. Evol. Microbiol. 65, 510–515. doi: 10.1099/ijs.0.064832-0
Sharma, D., Mayilraj, S., and Manhas, R. K. (2014). Streptomyces amritsarensis sp. nov., exhibiting broad-spectrum antimicrobial activity. Antonie Van Leeuwenhoek 105, 943–949. doi: 10.1007/s10482-014-0151-2
Sheridan, P. P., Loveland-Curtze, J., Miteva, V. I., and Brenchley, J. E. (2003). Rhodoglobus vestalii gen. nov., sp. nov., a novel psychrophilic organism isolated from an Antarctic Dry Valley Lake. Int. J. Syst. Evol. Microbiol. 53, 985–994. doi: 10.1099/ijs.0.02415-0
Shi, Z., Luo, G., and Wang, G. (2012). Cellulomonas carbonis sp. nov., isolated from coal mine soil. Int. J. Syst. Evol. Microbiol. 62, 2004–2010. doi: 10.1099/ijs.0.034934-0
Shin, N.-R., Jung, M.-J., Kim, M.-S., Roh, S. W., Nam, Y.-D., and Bae, J.-W. (2011). Corynebacterium nuruki sp. nov., isolated from an alcohol fermentation starter. Int. J. Syst. Evol. Microbiol. 61, 2430–2434. doi: 10.1099/ijs.0.027763-0
Shinobu, R., and Kawato, M. (1960). On Streptomyces indigoferus nov. sp., producing blue to green soluble pigment on some synthetic media. Nat. Sci. 9, 49–53.
Shivaji, S., Bhadra, B., Rao, S. R., Chaturvedi, P., Pindi, P. K., and Raghukumar, C. (2007). Microbacterium indicum sp. nov., isolated from a deep-sea sediment sample from the Chagos Trench, Indian Ocean. Int. J. Syst. Evol. Microbiol. 57, 1819–1822. doi: 10.1099/ijs.0.64782-0
Shivaji, S., Chaturvedi, P., Begum, Z., Pindi, P. K., Manorama, R., Padmanaban, D. A., et al. (2009). Janibacter hoylei sp. nov., Bacillus isronensis sp. nov. and Bacillus aryabhattai sp. nov., isolated from cryotubes used for collecting air from the upper atmosphere. Int. J. Syst. Evol. Microbiol. 59, 2977–2986. doi: 10.1099/ijs.0.002527-0
Shojaei, H., Daley, C., Gitti, Z., Hashemi, A., Heidarieh, P., Moore, E. R. B., et al. (2013). Mycobacterium iranicum sp. nov., a rapidly growing scotochromogenic species isolated from clinical specimens on three different continents. Int. J. Syst. Evol. Microbiol. 63, 1383–1389. doi: 10.1099/ijs.0.043562-0
Shukla, S. K., Bernard, K. A., Harney, M., Frank, D. N., and Reed, K. D. (2003). Corynebacterium nigricans sp. nov.: proposed name for a black-pigmented Corynebacterium species recovered from the human female urogenital tract. J. Clin. Microbiol. 41, 4353–4358. doi: 10.1128/JCM.41.9.4353
Siddall, M. E. (2010). Unringing a bell: metazoan phylogenomics and the partition bootstrap. Cladistics 26, 444–452. doi: 10.1111/j.1096-0031.2009.00295.x
Simon, M., Scheuner, C., Meier-Kolthoff, J. P., Brinkhoff, T., Wagner-Döbler, I., Ulbrich, M., et al. (2017). Phylogenomics of Rhodobacteraceae reveals evolutionary adaptation to marine and non-marine habitats. ISME J. 11, 1483–1499. doi: 10.1038/ismej.2016.198
Singh, H., Won, K., Ngo, H. T. T., Du, J., Kook, M., and Yi, T.-H. (2015). Phycicoccus soli sp. nov., isolated from soil. Int. J. Syst. Evol. Microbiol. 65, 2351–2356. doi: 10.1099/ijs.0.000265
Sjödén, B., Guido, F., Izquierdo, A., Akervall, E., and Collins, M. D. (1998). Description of some coryneform bacteria isolated from human clinical specimens as Corynebacterium falsenii sp. nov. Int. J. Syst. Bacteriol. 48, 69–74.
Šmarda, P., Bureš, P., Horová, L., Leitch, I. J., Mucina, L., Pacini, E., et al. (2014). Ecological and evolutionary significance of genomic GC content diversity in monocots. Proc. Natl. Acad. Sci. U S A. 111, E4096–E4102. doi: 10.1073/pnas.1321152111
Snijders, E. P. (1924). Cavia-scheefkopperij, een nocardiose. Geneeskd. Tijdschr. voor Ned. 64, 85–87.
Sohn, K., Hong, S. G., Bae, K. S., and Chun, J. (2003). Transfer of Hongia koreensis Lee et al. 2000 to the genus Kribbella Park et al. 1999 as Kribbella koreensis comb. nov. Int. J. Syst. Evol. Microbiol. 53, 1005–1007. doi: 10.1099/ijs.0.02449-0
Sompolinsky, D., Lagziel, A., Naveh, D., and Yankilevitz, T. (1978). Mycobacterium haemophilum sp. nov., a new pathogen of humans. Int. J. Syst. Bacteriol. 28, 67–75.
Song, L., Li, W.-J., Wang, Q.-L., Chen, G.-Z., Zhang, Y.-S., and Xu, L.-H. (2005). Jiangella gansuensis gen. nov., sp. nov., a novel actinomycete from a desert soil in north-west China. Int. J. Syst. Evol. Microbiol. 55, 881–884. doi: 10.1099/ijs.0.63353-0
Sorokin, D. Y., van Pelt, S., Tourova, T. P., and Evtushenko, L. I. (2009). Nitriliruptor alkaliphilus gen. nov., sp. nov., a deep-lineage haloalkaliphilic actinobacterium from soda lakes capable of growth on aliphatic nitriles, and proposal of Nitriliruptoraceae fam. nov. and Nitriliruptorales ord. nov. Int. J. Syst. Evol. Microbiol. 59, 248–253. doi: 10.1099/ijs.0.002204-0
Spiegel, C. A., and Roberts, M. (1984). Mobiluncus gen. nov., Mobiluncus curtisii subsp. curtisii sp. nov., Mobiluncus curtisii subsp. holmesii subsp. nov., and Mobiluncus mulieris sp. nov., curved rods from the human vagina. Int. J. Syst. Bacteriol. 34, 177–184. doi: 10.1099/00207713-34-2-177
Springer, B., Kirschner, P., Rost-meyer, G., Schröder, K.-H., Kroppenstedt, R. M., and Böttger, E. C. (1993). Mycobacterium interjectum, a new species isolated from a patient with chronic lymphadenitis. J. Clin. Microbiol. 31, 3083–3089.
Springer, B., Wu, W.-K., Bodmer, T., Haase, G., Pfyffer, G. E., Kroppenstedt, R. M., et al. (1996). Isolation and characterization of a unique group of slowly growing Mycobacteria: description of Mycobacterium lentiflavum sp. nov. J. Clin. Microbiol. 34, 1100–1107.
Stach, J. E., Maldonado, L. A., Ward, A. C., Bull, A. T., and Goodfellow, M. (2004). Williamsia maris sp. nov., a novel actinomycete isolated from the sea of Japan. Int. J. Syst. Evol. Microbiol. 54, 191–194. doi: 10.1099/ijs.0.02767-0
Stackebrandt, E. (1992). Unifying phylogeny and phenotypic diversity, in The Prokaryotes, eds A. Balows, H. G. Trueper, M. Dworkin, W. Harder, and K. H. Schleifer (New York, NY: Springer), 19–47.
Stackebrandt, E., Breymann, S., Steiner, U., Prauser, H., Weiss, N., and Schumann, P. (2002). Re-evaluation of the status of the genus Oerskovia, reclassification of Promicromonospora enterophila (Jáger et al. 1983) as Oerskovia enterophila comb. nov. and Oerskovia paurometabola sp. nov. Int. J. Syst. Evol. Microbiol. 52, 1105–1111. doi: 10.1099/ijs.0.02084-0
Stackebrandt, E., Koch, C., Gvozdiak, O., and Schumann, P. (1995). Taxonomic dissection of the genus Micrococcus: Kocuria gen. nov., Nesterenkonia gen. nov., Kytococcus gen. nov., Dermacoccus gen. nov., and Micrococcus Cohn 1872 gen. emend. Int. J. Syst. Bacteriol. 45, 682–692.
Stackebrandt, E., and Kroppenstedt, R. M. (1987). Union of the genera Actinoplanes Couch, Ampullariella Couch, and Amorphosporangium Couch in a redefined genus Actinoplanes. Syst. Appl. Microbiol. 9, 110–114. doi: 10.1016/S0723-2020(87)80063-2
Stackebrandt, E., Kroppenstedt, R. M., Wink, J., and Schumann, P. (2004). Reclassification of Amycolatopsis orientalis subsp. lurida Lechevalier et al. 1986 as Amycolatopsis lurida sp. nov., comb. nov. Int. J. Syst. Evol. Microbiol. 54, 267–268. doi: 10.1099/ijs.0.02937-0
Stackebrandt, E., and Prauser, H. (1991). Assignment of the genera Cellulomonas, Oerskovia, Promicromonospora and Jonesia to Cellulomonadaceae fam. nov. Syst. Appl. Microbiol. 14, 261–265. doi: 10.1016/S0723-2020(11)80378-4
Stackebrandt, E., Rainey, F. A., and Ward-Rainey, N. L. (1997). Proposal for a new hierarchic classification system, Actinobacteria classis nov. Int. J. Syst. Bacteriol. 47, 479–491.
Stackebrandt, E., and Schumann, P. (2000). Description of Bogoriellaceae fam. nov., Dermacoccaceae fam. nov., Rarobacteraceae fam. nov. and Sanguibacteraceae fam. nov., and emendation of some families of the suborder Micrococcineae. Int. J. Syst. Evol. Microbiol. 50, 1279–1285. doi: 10.1099/00207713-50-3-1279
Stackebrandt, E., and Schumann, P. (2004). Reclassification of Promicromonospora pachnodae Cazemier et al. 2004 as Xylanimicrobium pachnodae gen. nov., comb. nov. Int. J. Syst. Evol. Microbiol. 54, 1383–1386. doi: 10.1099/ijs.0.63064-0
Stackebrandt, E., and Schumann, P. (2014). The family Dermacoccaceae, in The Prokaryotes, eds E. Rosenberg, E. F. DeLong, S. Lory, E. Stackebrandt, and F. Thompson (Berlin; Heidelberg: Springer), 301–315. doi: 10.1007/978-3-642-30138-4
Stackebrandt, E., Smida, J., and Collins, M. D. (1988). Evidence of phylogenetic heterogeneity within the genus Rhodococcus: revival of the genus Gordona (Tsukamura). J. Gen. Appl. Microbiol. 34, 341–348.
Stackebrandt, E., Smith, D., Casaregola, S., Varese, G. C., Verkleij, G., Lima, N., et al. (2014). Deposit of microbial strains in public service collections as part of the publication process to underpin good practice in science. Springerplus 3:208. doi: 10.1186/2193-1801-3-208
Stamatakis, A. (2014). RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 30, 1312–1313. doi: 10.1093/bioinformatics/btu033
Stoecker, M. A., Herwig, R. P., and Staley, J. T. (1994). Rhodococcus zopfii sp. nov., a toxicant-degrading bacterium. Int. J. Syst. Bacteriol. 44, 106–110.
Sturm, G., Jacobs, J., Spröer, C., Schumann, P., and Gescher, J. (2011). Leucobacter chromiiresistens sp. nov., a chromate-resistant strain. Int. J. Syst. Evol. Microbiol. 61, 956–960. doi: 10.1099/ijs.0.022780-0
Sultanpuram, V. R., Mothe, T., Chintalapati, S., and Chintalapati, V. R. (2015). Cellulosimicrobium aquatile sp. nov., isolated from Panagal reservoir, Nalgonda, India. Antonie Van Leeuwenhoek 108, 1357–1364. doi: 10.1007/s10482-015-0588-y
Sun, H.-M., Zhang, T., Wei, Y.-Z., Liu, H.-Y., Yu, L.-Y., and Zhang, Y.-Q. (2015). Tenggerimyces mesophilus gen. nov., sp. nov., a member of the family Nocardioidaceae. Int. J. Syst. Evol. Microbiol. 65, 3359–3364. doi: 10.1099/ijsem.0.000421
Sun, W., Huang, Y., Zhang, Y.-Q., and Liu, Z.-H. (2007). Streptomyces emeiensis sp. nov., a novel streptomycete from soil in China. Int. J. Syst. Evol. Microbiol. 57, 1635–1639. doi: 10.1099/ijs.0.64934-0
Supong, K., Suriyachadkun, C., Suwanborirux, K., Pittayakhajonwut, P., and Thawai, C. (2013). Verrucosispora andamanensis sp. nov., isolated from a marine sponge. Int. J. Syst. Evol. Microbiol. 63, 3970–3974. doi: 10.1099/ijs.0.050906-0
Suriyachadkun, C., Ngaemthao, W., Chunhametha, S., Tamura, T., and Sanglier, J.-J. (2013). Kutzneria buriramensis sp. nov., isolated from soil, and emended description of the genus Kutzneria. Int. J. Syst. Evol. Microbiol. 63, 47–52. doi: 10.1099/ijs.0.036533-0
Sutcliffe, I. C. (2015). Challenging the anthropocentric emphasis on phenotypic testing in prokaryotic species descriptions: rip it up and start again. Front. Genet. 6:218. doi: 10.3389/fgene.2015.00218
Sutcliffe, I. C., Trujillo, M. E., and Goodfellow, M. (2012). A call to arms for systematists: revitalising the purpose and practises underpinning the description of novel microbial taxa. Antonie Van Leeuwenhoek 101, 13–20. doi: 10.1007/s10482-011-9664-0
Suzuki, K., Collins, M. D., Iijima, E., and Komagata, K. (1988). Chemotaxonomic characterization of a radiotolerant bacterium, Arthrobacter radiotolerans: description of Rubrobacter radiotolerans gen. nov., comb nov. FEMS Microbiol. Lett. 52, 33–40.
Takahashi, Y., Iwai, Y., and Omura, S. (1984). Two new species of the genus Kitasatosporia, Kitasatosporia phosalacinea sp. nov. and Kitasatosporia griseola sp. nov. J. Gen. Appl. Microbiol. 30, 377–387.
Takahashi, Y., Matsumoto, A., Morisaki, K., and Omura, S. (2006). Patulibacter minatonensis gen. nov., sp. nov., a novel actinobacterium isolated using an agar medium supplemented with superoxide dismutase, and proposal of Patulibacteraceae fam. nov. Int. J. Syst. Evol. Microbiol. 56, 401–406. doi: 10.1099/ijs.0.63796-0
Takeuchi, M., and Hatano, K. (1998). Union of the genera Microbacterium Orla-Jensen and Aureobacterium Collins et al. in a redefined genus Microbacterium. Int. J. Syst. Bacteriol. 48, 739–747.
Tamura, T., and Hatano, K. (2001). Phylogenetic analysis of the genus Actinoplanes and transfer of Actinoplanes minutisporangius Ruan et al. 1986 and ‘Actinoplanes aurantiacus’ to Cryptosporangium minutisporangium comb. nov. and Cryptosporangium aurantiacum sp. nov. Int. J. Syst. Evol. Microbiol. 51, 2119–2125. doi: 10.1099/00207713-51-6-2119
Tamura, T., Hatano, K., and Suzuki, K. (2006). A new genus of the family Micromonosporaceae, Polymorphospora gen. nov., with description of Polymorphospora rubra sp. nov. Int. J. Syst. Evol. Microbiol. 56, 1959–1964. doi: 10.1099/ijs.0.64046-0
Tamura, T., Hayakawa, M., and Hatano, K. (1998). A new genus of the order Actinomycetales, Cryptosporangium gen. nov., with descriptions of Cryptosporangium arvum sp. nov. and Cryptosporangium japonicum sp. nov. Int. J. Syst. Bacteriol. 48, 995–1005.
Tamura, T., Hayakawa, M., Nonomura, H., Yokota, A., and Hatano, K. (1995). Four new species of the genus Actinokineospora: Actinokineospora inagensis sp. nov., Actinokineospora globicatena sp. nov., Actinokineospora terrae sp. nov., and Actinokineospora diospyrosa sp. nov. Int. J. Syst. Bacteriol. 45, 371–378.
Tamura, T., Nakagaito, Y., Nishii, T., Hasegawa, T., Stackebrandt, E., and Yokota, A. (1994). A new genus of the order Actinomycetales, Couchioplanes gen. nov., with descriptions of Couchioplanes caeruleus (Horan and Brodsky 1986) comb. nov. and Couchioplanes caeruleus subsp. azureus subsp. nov. Int. J. Syst. Bacteriol. 44, 193–203. doi: 10.1099/00207713-44-2-193
Tan, G. Y., Robinson, S., Lacey, E., Brown, R., Kim, W., and Goodfellow, M. (2007). Amycolatopsis regifaucium sp. nov., a novel actinomycete that produces kigamicins. Int. J. Syst. Evol. Microbiol. 57, 2562–2567. doi: 10.1099/ijs.0.64974-0
Tan, G. Y. A., and Goodfellow, M. (2012). Genus V. Amycolatopsis Lechevalier, Prauser, Labeda and Ruan 1986, 34 VP emend. Lee 2009, 1403, in Bergey's Manual of Systematic Bacteriology, Vol. 5, The Actinobacteria, eds M. Goodfellow, P. Kämpfer, H.-J. Busse, M. E. Trujillo, K. Suzuki, W. Ludwig, et al. (New York, NY: Springer), 1334–1358.
Tan, G. Y. A., Robinson, S., Lacey, E., and Goodfellow, M. (2006). Amycolatopsis australiensis sp. nov., an actinomycete isolated from arid soils. Int. J. Syst. Evol. Microbiol. 56, 2297–2301. doi: 10.1099/ijs.0.64260-0
Tang, H., Shi, X., Wang, X., Hao, H., Zhang, X.-M., and Zhang, L.-P. (2016). Environmental controls over Actinobacteria communities in ecological sensitive Yanshan mountains zone. Front. Microbiol. 7:343. doi: 10.3389/fmicb.2016.00343
Tang, S.-K., Tian, X.-P., Zhi, X.-Y., Cai, M., Wu, J.-Y., Yang, L.-L., et al. (2008). Haloactinospora alba gen. nov., sp. nov., a halophilic filamentous actinomycete of the family Nocardiopsaceae. Int. J. Syst. Evol. Microbiol. 58, 2075–2080. doi: 10.1099/ijs.0.65531-0
Tang, S.-K., Wang, Y., Guan, T.-W., Lee, J.-C., Kim, C.-J., and Li, W.-J. (2010a). Amycolatopsis halophila sp. nov., a halophilic actinomycete isolated from a salt lake. Int. J. Syst. Evol. Microbiol. 60, 1073–1078. doi: 10.1099/ijs.0.012427-0
Tang, S.-K., Wang, Y., Klenk, H.-P., Shi, R., Lou, K., Zhang, Y.-J., et al. (2011). Actinopolyspora alba sp. nov. and Actinopolyspora erythraea sp. nov., isolated from a salt field, and reclassification of Actinopolyspora iraqiensis Ruan et al. 1994 as a heterotypic synonym of Saccharomonospora halophila. Int. J. Syst. Evol. Microbiol. 61, 1693–1698. doi: 10.1099/ijs.0.022319-0
Tang, S.-K., Wang, Y., Lou, K., Mao, P.-H., Xu, L.-H., Jiang, C.-L., et al. (2009). Kocuria halotolerans sp. nov., an actinobacterium isolated from a saline soil in China. Int. J. Syst. Evol. Microbiol. 59, 1316–1320. doi: 10.1099/ijs.0.006627-0
Tang, S.-K., Wang, Y., Zhang, H., Lee, J.-C., Lou, K., Kim, C.-J., et al. (2010b). Haloechinothrix alba gen. nov., sp. nov., a halophilic, filamentous actinomycete of the suborder Pseudonocardineae. Int. J. Syst. Evol. Microbiol. 60, 2154–2158. doi: 10.1099/ijs.0.018531-0
Tang, X., Zhou, Y., Zhang, J., Ming, H., Nie, G.-X., Yang, L.-L., et al. (2012). Actinokineospora soli sp. nov., a thermotolerant actinomycete isolated from soil, and emended description of the genus Actinokineospora. Int. J. Syst. Evol. Microbiol. 62, 1845–1849. doi: 10.1099/ijs.0.035832-0
Tao, T., Yue, Y., Chen, W., and Chen, W. (2004). Proposal of Nakamurella gen. nov. as a substitute for the bacterial genus Microsphaera Yoshimi et al. 1996 and Nakamurellaceae fam. nov. as a substitute for the illegitimate bacterial family Microsphaeraceae Rainey et al. 1997. Int. J. Syst. Evol. Microbiol. 54, 999–1000. doi: 10.1099/ijs.0.02933-0
Taylor, D. J., and Piel, W. H. (2004). An assessment of accuracy, error, and conflict with support values from genome-scale phylogenetic data. Mol. Biol. Evol. 21, 1534–1537. doi: 10.1093/molbev/msh156
Terekhova, L. P., Sadikova, O. A., and Preobrazhenskaya, T. P. (1977). Actinoplanes cyaneus sp. nov. and its antagonistic properties. Antibiotiki 22, 1059–1063.
Thawai, C., and Kudo, T. (2012). Promicromonospora thailandica sp. nov., isolated from marine sediment. Int. J. Syst. Evol. Microbiol. 62, 2140–2144. doi: 10.1099/ijs.0.035972-0
Thawai, C., Tanasupawat, S., and Kudo, T. (2008). Micromonospora pattaloongensis sp. nov., isolated from a Thai mangrove forest. Int. J. Syst. Evol. Microbiol. 58, 1516–1521. doi: 10.1099/ijs.0.65410-0
Thiemann, J. E., Pagani, H., and Beretta, G. (1967). A new genus of the Actinoplanaceae: Dactylosporangium, gen. nov. Arch. Mikrobiol. 58, 42–52.
Thiemann, J. E., Pagani, H., and Beretta, G. (1968). A new genus of the Actinomycetales: Microtetraspora gen. nov. J. Gen. Microbiol. 50, 295–303.
Thompson, C. C., Amaral, G. R., Campeão, M., Edwards, R. A., Polz, M. F., Dutilh, B. E., et al. (2015). Microbial taxonomy in the post-genomic era: rebuilding from scratch? Arch. Microbiol. 197, 359–370. doi: 10.1007/s00203-014-1071-2
Tian, X.-P., Zhi, X.-Y., Qiu, Y.-Q., Zhang, Y.-Q., Tang, S.-K., Xu, L.-H., et al. (2009). Sciscionella marina gen. nov., sp. nov., a marine actinomycete isolated from a sediment in the northern south China sea. Int. J. Syst. Evol. Microbiol. 59, 222–228. doi: 10.1099/ijs.0.001982-0
Tomita, K., Nakakita, Y., Hoshino, Y., Numata, K., and Kawaguchi, H. (1987). New genus of the Actinomycetales: Streptoalloteichus hindustanus gen. nov., nom. rev.; sp. nov., nom. rev. Int. J. Syst. Bacteriol. 37, 211–213.
Tomita, K., Nishio, M., Saitoh, K., Yamamoto, H., Hoshino, Y., Ohkuma, H., et al. (1990). Pradimicins A, B, and C: new antifungal antibiotics. I. Taxonomy. Production, isolation and physico-chemical properties. J. Antibiot. 43, 755–762.
Torkko, P., Suomalainen, S., Iivanainen, E., Tortoli, E., Suutari, M., Seppa, J., et al. (2002). Mycobacterium palustre sp. nov., a potentially pathogenic, slowly growing mycobacterium isolated from clinical and veterinary specimens and from finnish stream waters. Int. J. Syst. Evol. Microbiol. 52, 1519–1525. doi: 10.1099/ijs.0.02089-0
Tortoli, E., Böttger, E. C., Fabio, A., Falsen, E., Gitti, Z., Grottola, A., et al. (2011). Mycobacterium europaeum sp. nov., a scotochromogenic species related to the Mycobacterium simiae complex. Int. J. Syst. Evol. Microbiol. 61, 1606–1611. doi: 10.1099/ijs.0.025601-0
Tortoli, E., Gitti, Z., Klenk, H.-P., Lauria, S., Mannino, R., Mantegani, P., et al. (2013). Survey of 150 strains belonging to the Mycobacterium terrae complex and description of Mycobacterium engbaekii sp. nov., Mycobacterium heraklionense sp. nov. and Mycobacterium longobardum sp. nov. Int. J. Syst. Evol. Microbiol. 63, 401–411. doi: 10.1099/ijs.0.038737-0
Tortoli, E., Kohl, T. A., Brown-Elliott, B. A., Trovato, A., Cardoso Leão, S., García, M. J., et al. (2016a). Emended description of Mycobacterium abscessus, Mycobacterium abscessus subs. abscessus, Mycobacterium abscessus subsp. bolletii and designation of Mycobacterium abscessus subsp. massiliense comb. nov. Int. J. Syst. Evol. Microbiol. 66, 4471–4479. doi: 10.1099/ijsem.0.001376
Tortoli, E., Kohl, T. A., Brown-Elliott, B. A., Trovato, A., Cardoso-Leão, S., Garcia, M. J., et al. (2017). Mycobacterium abscessus, a taxonomic puzzle. Int. J. Syst. Evol. Microbiol. 68, 467–469. doi: 10.1099/ijsem.0.002457
Tortoli, E., Piersimoni, C., Kroppenstedt, R. M., Montoya-Burgos, J. I., Reischl, U., Giacometti, A., et al. (2001). Mycobacterium doricum sp. nov. Int. J. Syst. Evol. Microbiol. 51, 2007–2012. doi: 10.1099/00207713-51-6-2007
Tortoli, E., Richter, E., Borroni, E., Cabibbe, A. M., Capitolo, E., Cittaro, D., et al. (2016b). Mycobacterium alsense sp. nov. a scotochromogenic slow grower isolated from clinical respiratory specimens. Int. J. Syst. Evol. Microbiol. 66, 450–455. doi: 10.1099/ijsem.0.000744
Tortoli, E., Rindi, L., García, M. J., Chiaradonna, P., Dei, R., Garzelli, C., et al. (2004). Proposal to elevate the genetic variant MAC-A included in the Mycobacterium avium complex, to species rank as Mycobacterium chimaera sp. nov. Int. J. Syst. Evol. Microbiol. 54, 1277–1285. doi: 10.1099/ijs.0.02777-0
Tortoli, E., Rindi, L., Goh, K. S., Katila, M. L., Mariottini, A., Mattei, R., et al. (2005). Mycobacterium florentinum sp. nov., isolated from humans. Int. J. Syst. Evol. Microbiol. 55, 1101–1106. doi: 10.1099/ijs.0.63485-0
Traiwan, J., Park, M. H., and Kim, W. (2011). Serinicoccus chungangensis sp. nov., isolated from tidal flat sediment, and emended description of the genus Serinicoccus. Int. J. Syst. Evol. Microbiol. 61, 1299–1303. doi: 10.1099/ijs.0.025635-0
Trejo, W. H., and Bennett, R. E. (1963). Streptomyces species comprising the blue-spore series. J. Bacteriol. 85, 676–690.
Trejo, W. H., Dean, L. D., Pluscec, J., Meyers, E., and Brown, W. E. (1977). Streptomyces laurentii, a new species producing thiostrepton. J. Antibiot. (Tokyo). 30, 639–643.
Tresner, H. D., and Backus, E. J. (1956). A broadened concept of the characteristics of Streptomyces hygroscopicus. Appl. Microbiol. 4, 243–250.
Trovatelli, L. D., Crociani, F., Pedinotti, M., and Scardovi, V. (1974). Bifidobacterium pullorum sp. nov.: a new species isolated from chicken feces and a related group of bifidobacteria isolated from rabbit feces. Arch. Microbiol. 98, 187–198. doi: 10.1007/BF00425281
Trujillo, M. E., Bacigalupe, R., Pujic, P., Igarashi, Y., Benito, P., Riesco, R., et al. (2014). Genome features of the endophytic actinobacterium Micromonospora lupini strain Lupac 08: on the process of adaptation to an endophytic life style? PLoS ONE 9:e108522. doi: 10.1371/journal.pone.0108522
Trujillo, M. E., Ferna, C., Kroppenstedt, R. M., Schumann, P., and Mateos, P. F. (2005). Micromonospora mirobrigensis sp. nov. Int. J. Syst. Evol. Microbiol. 55, 877–880. doi: 10.1099/ijs.0.63361-0
Trujillo, M. E., and Goodfellow, M. (2012). Genus III. Actinomadura, in Bergey's Manual of Systematic Bacteriology, Vol. 5, The Actinobacteria, eds. M. Goodfellow, P. Kämpfer, H.-J. Busse, M. E. Trujillo, K. Suzuki, W. Ludwig, et al. (New York, NY: Springer), 1940–1959.
Tsang, C.-C., Xiong, L., Poon, R. W. S., Chen, J. H. K., Leung, K.-W., Lam, J. Y. W., et al. (2016). Gordonia hongkongensis sp. nov., isolated from blood culture and peritoneal dialysis effluent of patients in Hong Kong. Int. J. Syst. Evol. Microbiol. 66, 3942–3950. doi: 10.1099/ijsem.0.001292
Tsukamoto, T., Takeuchi, M., Shida, O., Murata, H., and Shirata, A. (2001). Proposal of Mycetocola gen. nov. in the family Microbacteriaceae and three new species, Mycetocola saprophilus sp. nov., Mycetocola tolaasinivorans sp. nov. and Mycetocola lacteus sp. nov., isolated from cultivated mushroom, Pleurotus ostreatus. Int. J. Syst. Evol. Microbiol. 51, 937–944. doi: 10.1099/00207713-51-3-937
Tsukamura, M. (1971). Proposal of a new genus, Gordona, for slightly acid-fast organisms occurring in sputa of patients with pulmonary disease and in soil. J. Gen. Microbiol. 68, 15–26.
Tsukamura, M. (1981). Numerical analysis of rapidly growing, nonphotochromogenic Mycobacteria, including Mycobacteriurn agri (Tsukamura 1972) Tsukamura sp. nov., nom. rev. Int. J. Syst. Bacteriol. 31, 247–258.
Tsukamura, M. (1982a). Mycobacterium shimoidei sp. nov., nom. rev., a lung pathogen. Int. J. Syst. Bacteriol. 32, 67–69.
Tsukamura, M. (1982b). Numerical analysis of the taxonomy of Nocardiae and Rhodococci. Division of Nocardia asteroides sensu stricto into two species and descriptions of Nocardia paratuberculosis sp. nov. Tsukamura (formerly the Kyoto-I group of Tsukamura), Nocardia nova sp. nov. Tsukamura, Rhodococcus aichiensis sp. nov. Tsukamura, Rhodococcus chuhuensis sp. nov. Tsukamura, and Rhodococcus obuensis sp. nov. Tsukamura. Microbiol. Immunol. 26, 1101–1119.
Tsukamura, M., Mizuno, S., and Tsukamura, S. (1981). Numerical analysis of rapidly growing, scotochromogenic Mycobacteria, including Mycobacterium obuense sp. nov., chubuense sp. nov., nom. rev., and Mycobacterium tokaiense sp. nov., nom. rev. Int. J. Syst. Bacteriol. 31, 263–275.
Tsukamura, M., Van Der Meulen, H. J., and Grabow, W. O. K. (1983). Numerical taxonomy of rapidly growing, scotochromogenic mycobacteria of the Mycobacterium parafortuitum complex: Mycobacterium austroafricanum sp. nov. and Mycobacterium diernhoferi sp. nov., nom. rev. Int. J. Syst. Bacteriol. 33, 460–469. doi: 10.1099/00207713-33-3-460
Tsukamura, M., Yano, I., and Imaeda, T. (1986). Mycobacterium moriokaense sp. nov., a rapidly growing, nonphotochromogenic Mycobacterium. Int. J. Syst. Bacteriol. 36, 333–338.
Tul'skaya, E. M., Shashkov, A. S., Streshinskaya, G. M., Senchenkova, S. N., Potekhina, N. V., Kozlova, Y. I., et al. (2011). Teichuronic and teichulosonic acids of actinomycetes. Biochemistry 76, 736–744. doi: 10.1134/S0006297911070030
Turenne, C., Chedore, P., Wolfe, J., Jamieson, F., Broukhanski, G., May, K., et al. (2002). Mycobacterium lacus sp. nov., a novel slowly growing, non-chromogenic clinical isolate. Int. J. Syst. Evol. Microbiol. 52, 2135–2140. doi: 10.1099/ijs.0.02170-0
Turenne, C. Y., Cook, V. J., Burdz, T. V., Pauls, R. J., Thibert, L., Wolfe, J. N., et al. (2004a). Mycobacterium parascrofulaceum sp. nov., novel slowly growing, scotochromogenic clinical isolates related to Mycobacterium simiae. Int. J. Syst. Evol. Microbiol. 54, 1543–1551. doi: 10.1099/ijs.0.02940-0
Turenne, C. Y., Thibert, L., Williams, K., Burdz, T. V., Cook, V. J., Wolfe, J. N., et al. (2004b). Mycobacterium saskatchewanense sp. nov., a novel slowly growing scotochromogenic species from human clinical isolates related to Mycobacterium interjectum and accuprobe-positive for Mycobacterium avium complex. Int. J. Syst. Evol. Microbiol. 54, 659–667. doi: 10.1099/ijs.0.02739-0
Ue, H., Matsuo, Y., Kasai, H., and Yokota, A. (2011). Demequina globuliformis sp. nov., Demequina oxidasica sp. nov. and Demequina aurantiaca sp. nov., actinobacteria isolated from marine environments, and proposal of Demequinaceae fam. nov. Int. J. Syst. Evol. Microbiol. 61, 1322–1329. doi: 10.1099/ijs.0.024299-0
Umezawa, H., Takeuchi, T., Okami, Y., and Tazaki, T. (1953). On screening of antiviral substances produced by Streptomyces and on an antiviral substance achromoviromycin. Jpn. J. Med. Sci. Biol. 6, 261–268.
Urzì, C., De Leo, F., and Schumann, P. (2008). Kribbella catacumbae sp. nov. and Kribbella sancticallisti sp. nov., isolated from whitish-grey patinas in the catacombs of St Callistus in Rome, Italy. Int. J. Syst. Evol. Microbiol. 58, 2090–2097. doi: 10.1099/ijs.0.65613-0
van Ingen, J., Al-Hajoj, S. A. M., Boeree, M., Al-Rabiah, F., Enaimi, M., de Zwaan, R., et al. (2009a). Mycobacterium riyadhense sp. nov., a non-tuberculous species identified as Mycobacterium tuberculosis complex by a commercial line-probe assay. Int. J. Syst. Evol. Microbiol. 59, 1049–1053. doi: 10.1099/ijs.0.005629-0
van Ingen, J., Boeree, M. J., de Lange, W. C. M., de Haas, P. E. W., van der Zanden, A. G. M., Mijs, W., et al. (2009b). Mycobacterium noviomagense sp. nov.; clinical relevance evaluated in 17 patients. Int. J. Syst. Evol. Microbiol. 59, 845–849. doi: 10.1099/ijs.0.001511-0
van Ingen, J., Boeree, M. J., Kösters, K., Wieland, A., Tortoli, E., Dekhuijzen, P. N. R., et al. (2009c). Proposal to elevate Mycobacterium avium complex ITS sequevar MAC-Q to Mycobacterium vulneris sp. nov. Int. J. Syst. Evol. Microbiol. 59, 2277–2282. doi: 10.1099/ijs.0.008854-0
van Ingen, J., Lindeboom, J. A., Hartwig, N. G., de Zwaan, R., Tortoli, E., Dekhuijzen, P. N. R., et al. (2009d). Mycobacterium mantenii sp. nov., a pathogenic, slowly growing, Scotochromogenic species. Int. J. Syst. Evol. Microbiol. 59, 2782–2787. doi: 10.1099/ijs.0.010405-0
van Ingen, J., Tortoli, E., Selvarangan, R., Coyle, M. B., Crump, J. A., Morrissey, A. B., et al. (2011). Mycobacterium sherrisii sp. nov., a slow-growing non-chromogenic species. Int. J. Syst. Evol. Microbiol. 61, 1293–1298. doi: 10.1099/ijs.0.024752-0
Vandamme, P., Falsen, E., Vancanneyt, M., Esbroeck, M., Van, Van de Merwe, D., Bergmans, A., et al. (1998). Characterization of Actinomyces turicensis and Actinomyces radingae strains from human clinical samples. Int. J. Syst. Bacteriol. 48, 503–510.
Vandamme, P., and Peeters, C. (2014). Time to revisit polyphasic taxonomy. Antonie Van Leeuwenhoek 106, 57–65. doi: 10.1007/s10482-014-0148-x
Vandamme, P., Pot, B., Gillis, M., De Vos, P., Kersters, K., and Swings, J. (1996). Polyphasic taxonomy, a consensus approach to bacterial systematics. Microbiol. Rev. 60, 407–438.
Vaz-Moreira, I., Lopes, A. R., Falsen, E., Schumann, P., Nunes, O. C., and Manaia, C. M. (2008). Microbacterium luticocti sp. nov., isolated from sewage sludge compost. Int. J. Syst. Evol. Microbiol. 58, 1700–1704. doi: 10.1099/ijs.0.65494-0
Vences, M., Guayasamin, J. M., Miralles, A., and De La Riva, I. (2013). To name or not to name: criteria to promote economy of change in Linnaean classification schemes. Zootaxa 3636, 201–244. doi: 10.11646/zootaxa.3636.2.1
Virgilio, A., and Hengeller, C. (1960). Produzione di tetraciclina con Streptomyces psammoticus. Farm. Ed. Sci. 15, 164–174.
Wade, W. G., Downes, J., Dymock, D., Hiom, S. J., Weightman, A. J., Dewhirst, F. E., et al. (1999). The family Coriobacteriaceae: reclassification of Eubacterium exiguum (Poco et al. 1996) and Peptostreptococcus heliotrinreducens (Lanigan 1976) as Slackia exigua gen. nov., comb. nov. and Slackia heliotrinireducens gen. nov., comb. nov., and Eubacterium lentum (Prevot 1938) as Eggerthella Ienta gen. nov., comb. nov. Int. J. Syst. Bacteriol. 49, 595–600.
Waksman, S. A. (ed.). (1961). The Actinomycetes, Vol. 2, Classification, Identification and Descriptions of Genera and Species. Baltimore, MA: The Williams & Wilkins Co.
Waksman, S. A., and Henrici, A. (1943). The nomenclature and classification of the Actinomycetes. Actinomycetes 46, 337–341. doi: 10.1001/jama.1895.02430120030007
Waksman, S. A., and Henrici, A. T. (1948). Family III. Streptomycetaceae Waksman and Henrici, in Bergey's Manual of Determinative Bacteriology, eds R. S. Breed, E. G. D. Murray, and A. P. Hitchens (Baltimore, MA: The Williams & Wilkins Co.), 929–980.
Waksman, S. A., and Lechevalier, H. A. (eds.). (1953). A Guide to the Classification of the Actinomycetes and their Antibiotics. Baltimore, MA: The Williams & Wilkins Co.
Wallhausser, K. H., Nesemann, P., Prave, P., and Steigler, A. (1965). Moenomycin, a new antibiotic. I. Fermentation and isolation. Antimicrob. Agents Chemother. 5, 734–736.
Wallick, H., Harris, D. A., Reagan, M. A., Ruger, M., and Woodruff, H. B. (1956). Discovery and antimicrobial properties of cathomycin, a new antibiotic produced by Streptomyces spheroides, n. sp. Antibiot. Annu. 1955–56, 909–917.
Wang, L., An, D. S., Jin, F. X., Lee, S. T., Im, W. T., and Bae, H. M. (2011). Phycicoccus ginsenosidimutans sp. nov., isolated from soil of a ginseng field. Int. J. Syst. Evol. Microbiol. 61, 524–528. doi: 10.1099/ijs.0.022137-0
Wang, L., Huang, Y., Liu, Z., Goodfellow, M., and Rodríguez, C. (2006). Streptacidiphilus oryzae sp. nov., an actinomycete isolated form rice-field soil in Thailand. Int. J. Syst. Evol. Microbiol. 56, 1257–1261. doi: 10.1099/ijs.0.64165-0
Wang, Q.-M., Yurkov, A. M., Göker, M., Lumbsch, H. T., Leavitt, S. D., Groenewald, M., et al. (2015). Phylogenetic classification of yeasts and related taxa within Pucciniomycotina. Stud. Mycol. 81, 149–189. doi: 10.1016/j.simyco.2015.12.002
Wang, W., Zhang, Z., Tang, Q., Mao, J., Wei, D., Huang, Y., et al. (2007a). Lechevalieria xinjiangensis sp. nov., a novel actinomycete isolated from radiation-polluted soil in China. Int. J. Syst. Evol. Microbiol. 57, 2819–2822. doi: 10.1099/ijs.0.65134-0
Wang, X., Jia, F., Liu, C., Zhao, J., Wang, L., Shen, Y., et al. (2013a). Xiangella phaseoli gen. nov., sp. nov., a member of the family Micromonosporaceae. Int. J. Syst. Evol. Microbiol. 63, 2138–2145. doi: 10.1099/ijs.0.045732-0
Wang, X., Zhao, J., Liu, C., Wang, J., Shen, Y., Jia, F., et al. (2013b). Nonomuraea solani sp. nov., an actinomycete isolated from eggplant root (Solanum melongena L.). Int. J. Syst. Evol. Microbiol. 63, 2418–2423. doi: 10.1099/ijs.0.045617-0
Wang, Y., Zhang, Z., and Ruan, J. (1996). A proposal to transfer Microbispora bispora (Lechevalier 1965) to a new genus, Thermobispora gen. nov., as Thermobispora bispora comb. nov. Int. J. Syst. Bacteriol. 46, 933–938. doi: 10.1099/00207713-46-4-933
Wang, Y.-X., Wang, H.-B., Zhang, Y.-Q., Xu, L.-H., Jiang, C.-L., and Li, W.-J. (2008). Rhodococcus kunmingensis sp. nov., an actinobacterium isolated from a rhizosphere soil. Int. J. Syst. Evol. Microbiol. 58, 1467–1471. doi: 10.1099/ijs.0.65673-0
Wang, Y.-X., Zhi, X.-Y., Chen, H.-H., Zhang, Y.-Q., Tang, S.-K., Jiang, C.-L., et al. (2007b). Actinomadura alba sp. nov., isolated from soil in Yunnan, China. Int. J. Syst. Evol. Microbiol. 57, 1735–1739. doi: 10.1099/ijs.0.64864-0
Wang, Y.-X., Zhi, X.-Y., Chen, H.-H., Zhang, Y.-Q., Tang, S. K., Jiang, C.-L., et al. (2007c). Streptomyces serianimatus sp. nov., isolated from a rhizophere soil. Antonie Van Leeuwenhoek 92, 201–206. doi: 10.1007/s10482-007-9147-5
Wang, Y. M., Zhang, Z. S., Xu, X. L., Ruan, J. S., and Wang, Y. (2001). Actinopolymorpha singaporensis gen. nov., sp. nov., a novel actinomycete from the tropical rainforest of Singapore. Int. J. Syst. Evol. Microbiol. 51, 467–473. doi: 10.1099/00207713-51-2-467
Warwick, S., Bowen, T., McVeigh, H., and Embley, T. M. (1994). A phylogenetic analysis of the family Pseudonocardiaceae and the genera Actinokineospora and Saccharothrix with 16S rRNA sequences and a proposal to combine the genera Amycolata and Pseudonocardia in an emended genus Pseudonocardia. Int. J. Syst. Bacteriol. 44, 293–299.
Watabe, J., Benno, Y., and Mitsuoka, T. (1983). Bifidobacterium gallinarum sp. nov.: a new species isolated from the ceca of chickens. Int. J. Syst. Bacteriol. 33, 127–132. doi: 10.1099/00207713-33-2-127
Watanabe, K., Makino, H., Sasamoto, M., Kudo, Y., Fujimoto, J., and Demberel, S. (2009). Bifidobacterium mongoliense sp. nov., from airag, a traditional fermented mare's milk product from Mongolia. Int. J. Syst. Evol. Microbiol. 59, 1535–1540. doi: 10.1099/ijs.0.006247-0
Wayne, L. G., Brenner, D. J., Colwell, R. R., Grimont, P. A. D., Kandler, O., Krichevsky, M. I., et al. (1987). Report of the ad hoc committee on reconciliation of approaches to bacterial systematics. Int. J. Syst. Bacteriol. 37, 463–464.
Weinstein, M. J., Luedemann, G. M., Oden, E. M., and Wagman, G. H. (1968). Halomicin, a new Micromonospora-produced antibiotic. Antimicrob. Agents Chemother. 1967, 435–441.
Weiszfeiler, G., Karasseva, V., and Karczag, E. (1971). A new Mycobacterium species: Mycobacterium asiaticum n. sp. Acta Microbiol. Acad. Sci. Hung. 18, 247–252.
Wellington, E. M., Stackebrandt, E., Sanders, D., Wolstrup, J., and Jorgensen, N. O. (1992). Taxonomic status of Kitasatosporia, and proposed unification with Streptomyces on the basis of phenotypic and 16S rRNA analysis and emendation of Streptomyces Waksman and Henrici 1943, 339AL. Int. J. Syst. Bacteriol. 42, 156–160. doi: 10.1099/00207713-42-1-156
Weon, H.-Y., Kim, B.-Y., Schumann, P., Kroppenstedt, R. M., Noh, H.-J., Park, C.-W., et al. (2007). Knoellia aerolata sp. nov., isolated from an air sample in Korea. Int. J. Syst. Evol. Microbiol. 57, 2861–2864. doi: 10.1099/ijs.0.65293-0
Weon, H.-Y., Yoo, S.-H., Kim, B.-Y., Schumann, P., Kroppenstedt, R. M., Hong, S.-K., et al. (2008). Phycicoccus aerophilus sp. nov., isolated from air. Int. J. Syst. Evol. Microbiol. 58, 2389–2392. doi: 10.1099/ijs.0.65202-0
Whipps, C. M., Butler, W. R., Pourahmad, F., Watral, V. G., and Kent, M. L. (2007). Molecular systematics support the revival of Mycobacterium salmoniphilum (ex Ross 1960) sp. nov., nom. rev., a species closely related to Mycobacterium chelonae. Int. J. Syst. Evol. Microbiol. 57, 2525–2531. doi: 10.1099/ijs.0.64841-0
Wiertz, R., Schulz, S. C., Müller, U., Kämpfer, P., and Lipski, A. (2013). Corynebacterium frankenforstense sp. nov. and Corynebacterium lactis sp. nov., isolated from raw cow milk. Int. J. Syst. Evol. Microbiol. 63, 4495–4501. doi: 10.1099/ijs.0.050757-0
Wieser, M., Denner, E. B. M., Kämpfer, P., Schumann, P., Tindall, B., Steiner, U., et al. (2002). Emended descriptions of the genus Micrococcus, Micrococcus luteus (Cohn 1872) and Micrococcus lylae (Kloos et al. 1974). Int. J. Syst. Evol. Microbiol. 52, 629–637. doi: 10.1099/00207713-52-2-629
Wiley, E. O., and Lieberman, B. S. (2011). Phylogenetics: Theory and Practice of Phylogenetic Systematics, 2nd Edn. Hoboken, NJ: John Wiley & Sons, Inc.
Wilson, R. W., Steingrube, V. A., Böttger, E. C., Springer, B., Brown-Elliott, B. A., Vicent, V., et al. (2001). Mycobacterium immunogenum sp. nov., a novel species related to Mycobacterium abscessus and associated with clinical disease, pseudo-outbreaks and contaminated metalworking fluids: an international cooperative study on mycobacterial taxonomy. Int. J. Syst. Evol. Microbiol. 51, 1751–1764. doi: 10.1099/00207713-51-5-1751
Wink, J., Gandhi, J., Kroppenstedt, R. M., Seibert, G., Sträubler, B., Schumann, P., et al. (2004). Amycolatopsis decaplanina sp. nov., a novel member of the genus with unusual morphology. Int. J. Syst. Evol. Microbiol. 54, 235–239. doi: 10.1099/ijs.0.02586-0
Wink, J., Schumann, P., Spöer, C., Eisenbarth, K., Glaeser, S. P., Martin, K., et al. (2014). Emended description of Actinoplanes friuliensis and description of Actinoplanes nipponensis sp. nov., antibiotic-producing species of the genus Actinoplanes. Int. J. Syst. Evol. Microbiol. 64, 599–606. doi: 10.1099/ijs.0.057836-0
Wink, J. M., Kroppenstedt, R. M., Ganguli, B. N., Nadkarni, S. R., Schumann, P., Seibert, G., et al. (2003). Three new antibiotic producing species of the genus Amycolatopsis, Amycolatopsis balhimycina sp. nov., A. tolypomycina sp. nov., A. vancoresmycina sp. nov., and description of Amycolatopsis keratiniphila subsp. keratiniphila subsp. nov. and A. keratiniphila subsp. nogabecina subsp. nov. Syst. Appl. Microbiol. 26, 38–46. doi: 10.1078/072320203322337290
Witt, D., and Stackebrandt, E. (1990). Unification of the genera Streptoverticillum and Streptomyces, and amendation of Streptomyces Waksman and Henrici 1943, 339AL. Syst. Appl. Microbiol. 13, 361–371. doi: 10.1016/S0723-2020(11)80234-1
Woo, P. C. Y., Fung, A. M. Y., Lau, S. K. P., Teng, J. L. L., Wong, B. H. L., Wong, M. K. M., et al. (2003). Actinomyces hongkongensis sp. nov. a novel Actinomyces species Isolated from a patient with pelvic actinomycosis. Syst. Appl. Microbiol. 26, 518–522. doi: 10.1078/072320203770865819
Wood, S. (1994). Monophyly and comparison between trees. Cladistics 10, 339–346. doi: 10.1006/clad.1994.1024
Wu, C.-Y., Zhuang, L., Zhou, S.-G., Li, F.-B., and He, J. (2011). Corynebacterium humireducens sp. nov., an alkaliphilic, humic acid-reducing bacterium isolated from a microbial fuel cell. Int. J. Syst. Evol. Microbiol. 61, 882–887. doi: 10.1099/ijs.0.020909-0
Wu, H., Liu, B., Shao, Y., Ou, X., and Huang, F. (2018). Thermostaphylospora grisealba gen. nov., sp. nov., isolated from mushroom compost and transfer of Thermomonospora chromogena Zhang et al. 1998 to Thermostaphylospora chromogena comb. nov. Int. J. Syst. Evol. Microbiol. 68, 602–608. doi: 10.1099/ijsem.0.002551
Wu, Y., Li, W.-J., Tian, W., Zhang, L.-P., Xu, L., Shen, Q.-R., et al. (2010). Isoptericola jiangsuensis sp. nov., a chitin-degrading bacterium. Int. J. Syst. Evol. Microbiol. 60, 904–908. doi: 10.1099/ijs.0.012864-0
Wüst, J., Stubbs, S., Weiss, N., Funke, G., and Collins, M. D. (1995). Assignment of Actinomyces pyogenes-like (CDC coryneform group E) bacteria to the genus Actinomyces as Actinomyces radingae sp. nov. and Actinomyces turicensis sp. nov. Lett. Appl. Microbiol. 20, 76–81.
Xi, L., Zhang, L., Ruan, J., and Huang, Y. (2012). Description of Verrucosispora qiuiae sp. nov., isolated from mangrove swamp sediment, and emended description of the genus Verrucosispora. Int. J. Syst. Evol. Microbiol. 62, 1564–1569. doi: 10.1099/ijs.0.033787-0
Xia, Z.-F., Ruan, J.-S., Huang, Y., and Zhang, L.-L. (2013). Streptomyces aidingensis sp. nov., an actinomycete isolated from lake sediment. Int. J. Syst. Evol. Microbiol. 63, 3204–3208. doi: 10.1099/ijs.0.049205-0
Xiao, C., Huang, H., Ye, J., Wu, X., Zhu, J., Zhan, B., et al. (2011a). Ornithinibacter aureus gen. nov., sp. nov., a novel member of the family Intrasporangiaceae. Int. J. Syst. Evol. Microbiol. 61, 659–660. doi: 10.1099/ijs.0.022806-0
Xiao, J., Luo, Y., Xie, S., and Xu, J. (2011b). Serinicoccus profundi sp. nov., an actinomycete isolated from deep-sea sediment, and emended description of the genus Serinicoccus. Int. J. Syst. Evol. Microbiol. 61, 16–19. doi: 10.1099/ijs.0.019976-0
Xie, Q. Y., Lin, H., Li, L., Roselyn, B., Goodfellow, M., Deng, Z., et al. (2012). Verrucosispora wenchangensis sp. nov., isolated from mangrove soil. Antonie Van Leeuwenhoek 102, 1–7. doi: 10.1007/s10482-012-9707-1
Xing, K., Bian, G.-K., Qin, S., Klenk, H.-P., Yuan, B., Zhang, Y.-J., et al. (2012). Kibdelosporangium phytohabitans sp. nov., a novel endophytic actinomycete isolated from oil-seed plant Jatropha curcas L. containing 1-aminocyclopropane-1-carboxylic acid deaminase. Antonie Van Leeuwenhoek 101, 433–441. doi: 10.1007/s10482-011-9652-4
Xu, C., Wang, L., Cui, Q., Huang, Y., Liu, Z., Zheng, G., et al. (2006). Neutrotolerant acidophilic Streptomyces species isolated from acidic soils in China: Streptomyces guanduensis sp. nov., Streptomyces paucisporeus sp. nov., Streptomyces rubidus sp. nov. and Streptomyces yanglinensis sp. nov. Int. J. Syst. Evol. Microbiol. 56, 1109–1115. doi: 10.1099/ijs.0.63959-0
Xu, J. L., He, J., Wang, Z. C., Wang, K., Li, W. J., Tang, S. K., et al. (2007). Rhodococcus qingshengii sp. nov., a carbendazim-degrading bacterium. Int. J. Syst. Evol. Microbiol. 57, 2754–2757. doi: 10.1099/ijs.0.65095-0
Yabe, S., Aiba, Y., Sakai, Y., Hazaka, M., and Yokota, A. (2011). Thermasporomyces composti gen. nov., sp. nov., a thermophilic actinomycete isolated from compost. Int. J. Syst. Evol. Microbiol. 61, 86–90. doi: 10.1099/ijs.0.021741-0
Yaeshima, T., Fujisawa, T., and Mitsuoka, T. (1992). Bifidobacterium globosum, subjective synonym of Bifidobacterium pseudolongum, and description of Bifidobacterium pseudolongum subsp. pseudolongum comb. nov. and Bifidobacterium pseudolongum subsp. globosum comb. nov. Syst. Appl. Microbiol. 15, 380–385.
Yamada, K., and Komagata, K. (1972). Taxonomic studies on coryneform bacteria. V. Classification of coryneform bacteria. J. Gen. Appl. Microbiol. 18, 417–431.
Yamada, T., Imachi, H., Ohashi, A., Harada, H., Hanada, S., Kamagata, Y., et al. (2007). Bellilinea caldifistulae gen. nov., sp. nov. and Longilinea arvoryzae gen. nov., sp. nov., strictly anaerobic, filamentous bacteria of the phylum Chloroflexi isolated from methanogenic propionate-degrading consortia. Int. J. Syst. Evol. Microbiol. 57, 2299–2306. doi: 10.1099/ijs.0.65098-0
Yamada, T., Sekiguchi, Y., Hanada, S., Imachi, H., Ohashi, A., Harada, H., et al. (2006). Anaerolinea thermolimosa sp. nov., Levilinea saccharolytica gen. nov., sp. nov. and Leptolinea tardivitalis gen. nov., sp. nov., novel filamentous anaerobes, and description of the new classes Anaerolineae classis nov. and Caldilineae classis nov. in the bacterial phylum Chloroflexi. Int. J. Syst. Evol. Microbiol. 56, 1331–1340. doi: 10.1099/ijs.0.64169-0
Yamaguchi, T., and Saburi, Y. (1955). Studies on the anti-trichomonal actinomycetes and their classification. J. Gen. Appl. Microbiol. 1, 201–234.
Yamamoto, N., Sato, S.-I., Saito, K., Hasu, T., Tadenuma, M., Suzuki, K.-I., et al. (1988). Rarobacter faecitabidus gen. nov. sp. nov. a yeast-lysing coryneform bacterium. Int. J. Syst. Bacteriol. 38, 7–11.
Yanagawa, R., and Honda, E. (1978). Corynebacterium pilosum and Corynebacterium cystitidis, two new species from cows. Int. J. Syst. Bacteriol. 28, 209–216.
Yang, L.-L., Ao, T., Wang, X.-H., He, J., Klenk, H.-P., Tang, S.-K., et al. (2012). Proposal of Intrasporangium mesophilum sp. nov., and reclassification of Humihabitans oryzae Kageyama et al. 2007 as Intrasporangium oryzae comb. nov. Int. J. Syst. Evol. Microbiol. 62, 1037–1041. doi: 10.1099/ijs.0.029934-0
Yang, L.-L., Tang, S.-K., Zhang, Y.-Q., Zhi, X.-Y., Wang, D., Xu, L.-H., et al. (2008). Thermobifida halotolerans sp. nov., isolated from a salt mine sample, and emended description of the genus Thermobifida. Int. J. Syst. Evol. Microbiol. 58, 1821–1825. doi: 10.1099/ijs.0.65732-0
Yarza, P., and Munoz, R. (2014). The all-species living tree project. Meth. Microbiol. 41, 45–59. doi: 10.1016/bs.mim.2014.07.006
Yarza, P., Richter, M., Peplies, J., Euzeby, J., Amann, R., Schleifer, K.-H., et al. (2008). The All-Species Living Tree project: a 16S rRNA-based phylogenetic tree of all sequenced type strains. Syst. Appl. Microbiol. 31, 241–250. doi: 10.1016/j.syapm.2008.07.001
Yassin, A. F. (2005). Rhodococcus triatomae sp. nov., isolated from a blood-sucking bug. Int. J. Syst. Evol. Microbiol. 55, 1575–1579. doi: 10.1099/ijs.0.63571-0
Yassin, A. F. (2007). Corynebacterium ureicelerivorans sp. nov., a lipophilic bacterium isolated from blood culture. Int. J. Syst. Evol. Microbiol. 57, 1200–1203. doi: 10.1099/ijs.0.64832-0
Yassin, A. F. (2009). Corynebacterium ulceribovis sp. nov., isolated from the skin of the udder of a cow with a profound ulceration. Int. J. Syst. Evol. Microbiol. 59, 34–37. doi: 10.1099/ijs.0.65832-0
Yassin, A. F., and Hupfer, H. (2006). Williamsia deligens sp. nov., isolated from human blood. Int. J. Syst. Evol. Microbiol. 56, 193–197. doi: 10.1099/ijs.0.63856-0
Yassin, A. F., Hupfer, H., and Schaal, K. P. (2006). Dietzia cinnamea sp. nov., a novel species isolated from a perianal swab of a patient with a bone marrow transplant. Int. J. Syst. Evol. Microbiol. 56, 641–645. doi: 10.1099/ijs.0.63863-0
Yassin, A. F., Hupfer, H., Siering, C., and Schumann, P. (2011). Comparative chemotaxonomic and phylogenetic studies on the genus Arcanobacterium Collins et al. 1982 emend. Lehnen et al. 2006: proposal for Trueperella gen. nov. and emended description of the genus Arcanobacterium. Int. J. Syst. Evol. Microbiol. 61, 1265–1274. doi: 10.1099/ijs.0.020032-0
Yassin, A. F., Rainey, F. A., Brzezinka, H., Jahnke, K. D., Weissbrodt, H., Budzikiewicz, H., et al. (1995). Lentzea gen. nov., a new genus of the order Actinomycetales. Int. J. Syst. Bacteriol. 45, 357–363. doi: 10.1099/00207713-45-2-357
Yassin, A. F., Rainey, F. A., Burghardt, J., Gierth, D., Ungerechts, J., Lux, I., et al. (1997). Description of Nocardiopsis synnemataformans sp. nov., elevation of Nocardiopsis alba subsp. prasina to Nocardiopsis prasina comb. nov., and designation of Nocardiopsis antarctica and Nocardiopsis alborubida as later subjective synonyms of Nocardiopsis dassonvillei. Int. J. Syst. Bacteriol. 47, 983–988.
Yassin, A. F., and Schaal, K. P. (2005). Reclassification of Nocardia corynebacterioides Serrano et al. 1972 (Approved Lists 1980) as Rhodococcus corynebacterioides comb. nov. Int. J. Syst. Evol. Microbiol. 55, 1345–1348. doi: 10.1099/ijs.0.63529-0
Yassin, A. F., Shen, F.-T., Hupfer, H., Arun, A. B., Lai, W.-A., Rekha, P. D., et al. (2007a). Gordonia malaquae sp. nov., isolated from sludge of a wastewater treatment plant. Int. J. Syst. Evol. Microbiol. 57, 1065–1068. doi: 10.1099/ijs.0.64893-0
Yassin, A. F., and Siering, C. (2008). Corynebacterium sputi sp. nov., isolated from the sputum of a patient with pneumonia. Int. J. Syst. Evol. Microbiol. 58, 2876–2879. doi: 10.1099/ijs.0.2008/000414-0
Yassin, A. F., Spröer, C., Hupfer, H., Siering, C., and Klenk, H. P. (2009). Nocardiopsis potens sp. nov., isolated from household waste. Int. J. Syst. Evol. Microbiol. 59, 2729–2733. doi: 10.1099/ijs.0.011288-0
Yassin, A. F., Spröer, C., Pukall, R., Sylvester, M., Siering, C., and Schumann, P. (2015). Dissection of the genus Actinobaculum: reclassification of Actinobaculum schaalii Lawson et al. 1997 and Actinobaculum urinale Hall et al. 2003 as Actinotignum schaalii gen. nov., comb. nov. and Actinotignum urinale comb. nov., description of Actinotignum sanguinis sp. nov. and emended descriptions of the genus Actinobaculum and Actinobaculum suis; and re-examination of the culture deposited as Actinobaculum massiliense CCUG 47753 T (5DSM 19118 T), revealing that it does not represent a strain of this species. Int. J. Syst. Evol. Microbiol. 65, 615–624. doi: 10.1099/ijs.0.069294-0
Yassin, A. F., Steiner, U., and Ludwig, W. (2002a). Corynebacterium appendicis sp. nov. Int. J. Syst. Evol. Microbiol. 52, 1165–1169. doi: 10.1099/ijs.0.02141-0
Yassin, A. F., Steiner, U., and Ludwig, W. (2002b). Corynebacterium aurimucosum sp. nov. and emended description of Corynebacterium minutissimum Collins and Jones (1983). Int. J. Syst. Evol. Microbiol. 52, 1001–1005. doi: 10.1099/ijs.0.02045-0
Yassin, A. F., Young, C. C., Lai, W.-A., Hupfer, H., Arun, A. B., Shen, F.-T., et al. (2007b). Williamsia serinedens sp. nov., isolated from an oil-contaminated soil. Int. J. Syst. Evol. Microbiol. 57, 558–561. doi: 10.1099/ijs.0.64691-0
Yi, H., Schumann, P., and Chun, J. (2007). Demequina aestuarii gen. nov., sp. nov., a novel actinomycete of the suborder Micrococcineae, and reclassification of Cellulomonas fermentans Bagnara et al. 1985 as Actinotalea fermentans gen. nov., comb. nov. Int. J. Syst. Evol. Microbiol. 57, 151–156. doi: 10.1099/ijs.0.64525-0
Yi, H., Schumann, P., Sohn, K., and Chun, J. (2004). Serinicoccus marinus gen. nov., sp. nov., a novel actinomycete with L-ornithine and L-serine in the peptidoglycan. Int. J. Syst. Evol. Microbiol. 54, 1585–1589. doi: 10.1099/ijs.0.03036-0
Yoon, J.-H., Cho, Y.-G., Kang, S.-S., Kim, S. B., Lee, S. T., and Park, Y.-H. (2000a). Rhodococcus koreensis sp. nov., a 2, 4-dinitrophenol-degrading bacterium. Int. J. Syst. Evol. Microbiol. 50, 1193–1201. doi: 10.1099/00207713-50-3-1193
Yoon, J.-H., Kang, S.-S., Cho, Y.-G., Lee, S. T., Kho, Y. H., Kim, C.-J., et al. (2000b). Rhodococcus pyridinivorans sp. nov., a pyridine-degrading bacterium. Int. J. Syst. Evol. Microbiol. 50, 2173–2180. doi: 10.1099/00207713-50-6-2173
Yoon, J.-H., Lee, S.-Y., Kang, S.-J., and Oh, T.-K. (2008). Phycicoccus dokdonensis sp. nov., isolated from soil. Int. J. Syst. Evol. Microbiol. 58, 597–600. doi: 10.1099/ijs.0.65284-0
Young, C.-C., Busse, H.-J., Langer, S., Chu, J.-N., Schumann, P., Arun, A. B., et al. (2010). Microbacterium agarici sp. nov., Microbacterium humi sp. nov. and Microbacterium pseudoresistens sp. nov., isolated from the base of the mushroom Agaricus blazei. Int. J. Syst. Evol. Microbiol. 60, 854–860. doi: 10.1099/ijs.0.014092-0
Yuan, L.-J., Zhang, Y.-Q., Guan, Y., Wei, Y.-Z., Li, Q.-P., Yu, L.-Y., et al. (2008). Saccharopolyspora antimicrobica sp. nov., an actinomycete from soil. Int. J. Syst. Evol. Microbiol. 58, 1180–1185. doi: 10.1099/ijs.0.65532-0
Yuan, L.-J., Zhang, Y.-Q., Yu, L.-Y., Liu, H.-Y., Guan, Y., Lee, J.-C., et al. (2010). Alloactinosynnema album gen. nov., sp. nov., a member of the family Actinosynnemataceae isolated from soil. Int. J. Syst. Evol. Microbiol. 60, 39–43. doi: 10.1099/ijs.0.010744-0
Yun, J.-H., Roh, S. W., Kim, M.-S., Jung, M.-J., Park, E.-J., Shin, K.-S., et al. (2011). Leucobacter salsicius sp. nov., from a salt-fermented food. Int. J. Syst. Evol. Microbiol. 61, 502–506. doi: 10.1099/ijs.0.021360-0
Zarilla, K. A., and Perry, J. J. (1984). Thermoleophilum album gen. nov. and sp. nov., a bacterium obligate for thermophily and n-alkane substrates. Arch. Microbiol. 137, 286–290.
Zgurskaya, H. I., Evtushenko, L. I., Akimov, V. N., Voyevoda, H. V., Dobrovolskaya, T. G., Lysak, L. V., et al. (1992). Emended description of the genus Agromyces and description of Agromyces cerinus subsp. cerinus sp. nov., subp. nov., Agromyes cerinus subsp nitratus sp. nov., subsp. nov., Agromyces fucosus subsp. fucosus sp. nov., subsp. nov., and Agromyces fucosus subsp. hippuratus sp. nov., subsp. nov. Int. J. Syst. Bacteriol. 42, 635–641.
Zhang, D.-C., Schumann, P., Liu, H.-C., Xin, Y.-H., Zhou, Y.-G., Schinner, F., et al. (2010a). Arthrobacter alpinus sp. nov., a psychrophilic bacterium isolated from alpine soil. Int. J. Syst. Evol. Microbiol. 60, 2149–2153. doi: 10.1099/ijs.0.017178-0
Zhang, D.-C., Wang, H.-X., Cui, H.-L., Yang, Y., Liu, H.-C., Dong, X.-Z., et al. (2007a). Cryobacterium psychrotolerans sp. nov., a novel psychrotolerant bacterium isolated from the China No. 1 glacier. Int. J. Syst. Evol. Microbiol. 57, 866–869. doi: 10.1099/ijs.0.64750-0
Zhang, G., Ren, H., Wang, S., Chen, X., Yang, Y., Zhang, Y., et al. (2014). Janibacter indicus sp. nov., isolated from hydrothermal sediment of the Indian Ocean. Int. J. Syst. Evol. Microbiol. 64, 2353–2357. doi: 10.1099/ijs.0.059527-0
Zhang, H., Zheng, W., Huang, J., Luo, H., Jin, Y., Zhang, W., et al. (2006). Actinoalloteichus hymeniacidonis sp. nov., an actinomycete isolated from the marine sponge Hymeniacidon perleve. Int. J. Syst. Evol. Microbiol. 56, 2309–2312. doi: 10.1099/ijs.0.64217-0
Zhang, J., Ma, Y., and Yu, H. (2012). Arthrobacter cupressi sp. nov., an actinomycete isolated from the rhizosphere soil of Cupressus sempervirens. Int. J. Syst. Evol. Microbiol. 62, 2731–2736. doi: 10.1099/ijs.0.036889-0
Zhang, J., Wu, D., and Liu, Z. (2009a). Saccharopolyspora jiangxiensis sp. nov., isolated from grass-field soil. Int. J. Syst. Evol. Microbiol. 59, 1076–1081. doi: 10.1099/ijs.0.65863-0
Zhang, J., Xie, Q., Liu, Z., and Goodfellow, M. (2007b). Lechevalieria fradiae sp. nov., a novel actinomycete isolated from soil in China. Int. J. Syst. Evol. Microbiol. 57, 832–836. doi: 10.1099/ijs.0.64777-0
Zhang, J., Zhang, Y., Xiao, C., Liu, Z., and Goodfellow, M. (2002). Rhodococcus maanshanensis sp. nov., a novel actinomycete from soil. Int. J. Syst. Evol. Microbiol. 52, 2121–2126. doi: 10.1099/ijs.0.02253-0.Abbreviation
Zhang, J.-Y., Liu, X.-Y., and Liu, S.-J. (2010b). Agrococcus terreus sp. nov., isolated from forest soil. Int. J. Syst. Evol. Microbiol. 60, 1897–1903. doi: 10.1099/ijs.0.013235-0
Zhang, J.-Y., Liu, X.-Y., and Liu, S.-J. (2011). Phycicoccus cremeus sp. nov., isolated from forest soil, and emended description of the genus Phycicoccus. Int. J. Syst. Evol. Microbiol. 61, 71–75. doi: 10.1099/ijs.0.020842-0
Zhang, L., Xi, L., Qiu, D., Song, L., Dai, X., Ruan, J., et al. (2013a). Cellulomonas marina sp. nov., isolated from deep-sea water. Int. J. Syst. Evol. Microbiol. 63, 3014–3018. doi: 10.1099/ijs.0.048876-0
Zhang, L.-P., Zhang, L.-M., and Zhang, X.-M. (2009b). Streptosporangium canum sp. nov., isolated from soil. Int. J. Syst. Evol. Microbiol. 59, 1715–1719. doi: 10.1099/ijs.0.007401-0
Zhang, X., Zhang, J., Zhang, Y., Xin, Y., and He, H. (2013b). Friedmanniella flava sp. nov., a soil actinomycete. Int. J. Syst. Evol. Microbiol. 63, 1771–1775. doi: 10.1099/ijs.0.043984-0
Zhang, Y.-G., Lu, X.-H., Ding, Y.-B., Wang, S.-J., Zhou, X.-K., Wang, H.-F., et al. (2016a). Lipingzhangella halophila gen. nov., sp. nov., a new member of the family Nocardiopsaceae. Int. J. Syst. Evol. Microbiol. 66, 4071–4076. doi: 10.1099/ijsem.0.001312
Zhang, Y.-G., Wang, H.-F., Yang, L.-L., Guo, J.-W., Xiao, M., Huang, M.-J., et al. (2016b). Ornithinicoccus halotolerans sp. nov., and emended description of the genus Ornithinicoccus. Int. J. Syst. Evol. Microbiol. 66, 1894–1899. doi: 10.1099/ijsem.0.000964
Zhang, Z., Kudo, T., Nakajima, Y., and Wang, Y. (2001). Clarification of the relationship between the members of the family Thermomonosporaceae on the basis of 16S rDNA, 16S-23S rDNA internal transcribed spacer and 23S rDNA sequences and chemotaxonomic analyses. Int. J. Syst. Evol. Microbiol. 51, 373–383. doi: 10.1099/00207713-51-2-373
Zhang, Z., Wang, Y., and Ruan, J. (1997). A proposal to revive the genus Kitasatospora (Omura, Takahashi, Iwai, and Tanaka 1982). Int. J. Syst. Bacteriol. 47, 1048–1054. doi: 10.1099/00207713-47-4-1048
Zhang, Z., Wang, Y., and Ruan, J. (1998). Reclassification of Thermomonospora and Microtetraspora. Int. J. Syst. Bacteriol. 48, 411–422. doi: 10.1099/00207713-48-2-411
Zhao, J., Guo, L., Sun, P., Han, C., Bai, L., Liu, C., et al. (2015). Actinomadura jiaoheensis sp. nov. and Actinomadura sporangiiformans sp. nov., two novel actinomycetes isolated from muddy soil and emended description of the genus Actinomadura. Antonie Van Leeuwenhoek 108, 1331–1339. doi: 10.1007/s10482-015-0585-1
Zhao, K., Li, W., Kang, C., Du, L., Huang, T., Zhang, X., et al. (2014). Phylogenomics and evolutionary dynamics of the family Actinomycetaceae. Genome Biol. Evol. 6, 2625–2633. doi: 10.1093/gbe/evu211
Zhi, X. Y., Li, W. J., and Stackebrandt, E. (2009). An update of the structure and 16S rRNA gene sequence-based definition of higher ranks of the class Actinobacteria, with the proposal of two new suborders and four new families and emended descriptions of the existing higher taxa. Int. J. Syst. Evol. Microbiol. 59, 589–608. doi: 10.1099/ijs.0.65780-0
Zhou, E.-M., Tang, S.-K., Sjøholm, C., Song, Z.-Q., Yu, T.-T., Yang, L.-L., et al. (2012a). Thermoactinospora rubra gen. nov., sp. nov., a thermophilic actinomycete isolated from Tengchong, Yunnan province, south–west China. Antonie Van Leeuwenhoek 102, 177–185. doi: 10.1007/s10482-012-9725-z
Zhou, G., Luo, X., Tang, Y., Zhang, L., Yang, Q., Qiu, Y., et al. (2008). Kocuria flava sp. nov. and Kocuria turfanensis sp. nov., airborne actinobacteria isolated from Xinjiang, China. Int. J. Syst. Evol. Microbiol. 58, 1304–1307. doi: 10.1099/ijs.0.65323-0
Zhou, X., Guo, G. N., Wang, L. Q., Bai, S. L., and Li, Y. H. (2016). Cnuibacter physcomitrellae gen. nov., sp. nov., a novel member of the family Microbacteriaceae isolated from the moss of Physcomitrella patens. Int. J. Syst. Evol. Microbiol. 66, 680–688. doi: 10.1099/ijsem.0.000775
Zhou, Y., Wei, W., Wang, X., and Lai, R. (2009). Proposal of Sinomonas flava gen. nov., sp. nov., and description of Sinomonas atrocyanea comb. nov. to accommodate Arthrobacter atrocyaneus. Int. J. Syst. Evol. Microbiol. 59, 259–263. doi: 10.1099/ijs.0.000695-0
Zhou, Z., Yuan, M., Tang, R., Chen, M., Lin, M., and Zhang, W. (2012b). Corynebacterium deserti sp. nov., isolated from desert sand. Int. J. Syst. Evol. Microbiol. 62, 791–794. doi: 10.1099/ijs.0.030429-0
Zhu, H., Guo, J., Yao, Q., Yang, S., Deng, M., Phuong, L. T. B., et al. (2007). Streptomyces vietnamensis sp. nov., a streptomycete with violet–blue diffusible pigment isolated from soil in Vietnam. Int. J. Syst. Evol. Microbiol. 57, 1770–1774. doi: 10.1099/ijs.0.64774-0
Zhu, L., Li, W., and Dong, X. (2003). Species identification of genus Bifidobacterium based on partial HSP60 gene sequences and proposal of Bifidobacterium thermacidophilum subsp. porcinum subsp. nov. Int. J. Syst. Evol. Microbiol. 53, 1619–1623. doi: 10.1099/ijs.0.02617-0
Keywords: G+C content, genome size, Genome BLAST Distance Phylogeny, chemotaxonomy, morphology, phylogenetic systematics, phylogenomics
Citation: Nouioui I, Carro L, García-López M, Meier-Kolthoff JP, Woyke T, Kyrpides NC, Pukall R, Klenk H-P, Goodfellow M and Göker M (2018) Genome-Based Taxonomic Classification of the Phylum Actinobacteria. Front. Microbiol. 9:2007. doi: 10.3389/fmicb.2018.02007
Received: 31 January 2018; Accepted: 09 August 2018;
Published: 22 August 2018.
Edited by:
Martin G. Klotz, Washington State University Tri-Cities, United StatesReviewed by:
Nicola Segata, University of Trento, ItalyCopyright © 2018 Nouioui, Carro, García-López, Meier-Kolthoff, Woyke, Kyrpides, Pukall, Klenk, Goodfellow and Göker. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Markus Göker, bWFya3VzLmdvZWtlckBkc216LmRl
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.