 Yuxing Wu
Yuxing Wu Zhiyuan Yin
Zhiyuan Yin Liangsheng Xu
Liangsheng Xu Hao Feng
Hao Feng Lili Huang
Lili Huang- State Key Laboratory of Crop Stress Biology for Arid Areas, China–Australia Joint Research Centre for Abiotic and Biotic Stress Management, College of Plant Protection, Northwest A&F University, Yangling, China
The role of the transcription factor PacC has been characterised in several pathogenic fungi, and it affects virulence via several mechanisms. In this study, we examined the role of the PacC homolog VmPacC in Valsa mali, the causal agent of apple canker disease. We found that the expression of VmPacC was up-regulated in neutral and alkaline pH and during infection. At pH 6–10, the radial growth of a VmPacC deletion mutant decreased compared to wild-type. In addition, the sensitivity to oxidative stress of the VmPacC deletion mutant was impaired, as its growth was more severely inhibited by H2O2 than that of the wild-type. The lesion size caused by the VmPacC deletion mutant was smaller than that of the wild-type on apple leaves and twigs. Interestingly, expression of pectinase genes increased in deletion mutant during infection. To further confirm the negative regulation, we generated dominant activated C-27 allele mutants that constitutively express VmPacC. The pectinase activity of activated mutants was reduced at pH 4. We further observed that V. mali can acidify the pH during infection, and that the capacity for acidification was impaired after VmPacC deletion. Furthermore, VmPacC is involved in the generation of citric acid, which affects virulence. These results indicate that VmPacC is part of the fungal responses to neutral and alkaline pH and oxidative stress. More importantly, VmPacC is required for acidification of its environment and for full virulence in V. mali.
Introduction
To adapt to changing environmental conditions, fungi require an intracellular pH homeostasis system and regulatory mechanism that ensures that directly exposed molecules such as secreted enzymes can function at optimal pH (Penalva and Arst, 2002). An important system to regulate ambient pH in fungi is the Pal signalling pathway. This pathway includes seven components: PalA, PalB, PalC, PalF, PalH, PalI, and PacC. PacC is a transcription factor that regulates pH-dependent gene expression at the end of the pH signalling pathway (Peñalva et al., 2008). In Aspergillus nidulans, the PacC polypeptide shows a closed conformation with a processing protease domain inaccessible at acidic conditions. However, the full-length polypeptide is cleaved after a shift to neutral or alkaline conditions, and an intermediate 53 kDa fragment is generated (Díez et al., 2002; Peñalva et al., 2008). The 53-kDa fragment is cleaved again by the proteasome, and results in the final active form of 27-kDa PacC27 (Hervas-Aguilar et al., 2007). In addition, the binding motif of the PacC transcription factor is GCCARG. Transcription of alkaline-expressed genes is activated by PacC at alkaline pH, whereas transcription of acid-expressed genes is repressed at the same condition (Tilburn et al., 1995; Penalva and Espeso, 1996).
In several plant pathogenic fungi, the involvement of PacC in virulence has been demonstrated. Deletion of PacC homologs reduces fungal virulence in Colletotrichum gloeosporioides (Yakoby et al., 2000), Penicillium digitatum (Zhang et al., 2013), Magnaporthe oryzae (Landraud et al., 2013), Fusarium oxysporum (Caracuel et al., 2003), and Sclerotinia sclerotiorum (Rollins, 2003). PacC not only detects environment conditions, but also modifies these conditions via acid or ammonia secretion. Some plant pathogens are therefore grouped into two categories. The first group can alkalinise their host tissue and enhance virulence by producing ammonia. Examples include Colletotrichum spp. and Alternaria alternata (Eshel et al., 2002; Alkan et al., 2009). The other group of pathogens, including Penicillium spp., Botrytis cinerea, and S. sclerotiorum, can acidify the host tissue by producing acids (Wubben et al., 2000; Rollins and Dickman, 2001; Barad et al., 2012; Zhang et al., 2013). This modulation of host environment provides a suitable condition for the function of virulence factors like secreted enzymes. It may also serve as a signal to activate the production of these virulence factors. In addition to modify environment pH, PacC is also known to regulate some cell wall degrading enzymes such as polygalacturonase (Rollins, 2003), pectate lyase (Kramer-Haimovich et al., 2006), and endoglucanase to affect fungal virulence (Eshel et al., 2002).
Apple Valsa canker is a bark disease of apple trees (Malus sp.) that seriously imperils apple production in China (Wang et al., 2014), Korea (Lee et al., 2006), and Japan (Abe et al., 2007). This disease is caused by the parasite fungus Valsa mali. V. mali infects the bark of apple trees through various wounds such as pruning ends, sunscalds, frostbites, and some mechanical injuries (Ke et al., 2013). Following colonisation of the wounded tissue, the hyphae spread to all bark tissues, resulting in severe tissue maceration and necrosis. As a plant pathogenic fungus without specialised penetration structures, V. mali secretes hydrolytic enzymes, particularly pectinases, involved in disease development and pathogenesis (Ke et al., 2013, 2014; Feng et al., 2017b). Pectinases are considered virulence factors because the virulence of several V. mali pectinase gene deletion mutants is attenuated (Yin et al., 2015). However, these findings do not clarify the role of PacC in the regulation of virulence in this important woody plant fungal pathogens. Understanding the role of PacC during V. mali infection might allow the development of novel sustainable apple Valsa canker management strategies. In the present study, we have investigated the role of PacC in pH acidification, oxidative stress, and pectinase gene expression during V. mali infection.
Materials and Methods
Strains and Culture Conditions
The wild-type strain of V. mali named 03-8 (Laboratory of Integrated Management of Plant Diseases in College of Plant Protection, Northwest A&F University) was used (Yin et al., 2015). All strains including the wild-type and the transformants generated in this work were propagated on potato dextrose agar (PDA, 20% potato extract, 2% dextrose, 1.5% agar) when necessary. Potato dextrose broth (PDB, 20% potato extract, 2% dextrose) was used for measuring the dynamic pH change or for liquid culture.
Growth of wild-type and mutant strains on PDA media at different pH were assayed by measuring colony diameters 2 days post-inoculation (dpi) at 25°C. PDA medium was adjusted with HCl or NaOH to pH 3–10. The PDA media mended NaCl (0.1 M), H2O2 (3 mM), congo red (CR, 300 mg/L), or sodium dodecyl sulfonate (SDS 0.01%) were used for stress response assays. For selection of transformants during gene deletion, TB3 medium (0.3% yeast extract, 0.3% casamino acids, 20% sucrose, 1.5% agar) supplemented with 100 μg/mL hygromycin B (Calbiochem, La Jolla, CA, United States) or 100 μg/mL geneticin (Sigma, St. Louis, MO, United States) was used. To determine protein concentration and pectinase activity, synthetic medium (SM) (Srivastava et al., 2012) with 1% pectin as the sole carbon source was used. Buffered SM medium was obtained using 0.05 M Na2HPO4 and 0.05 M C4H2O7 to maintain pH 4 (38.5% volume ratio of Na2HPO4) or pH 7 (82.4% volume ratio of Na2HPO4).
Twig and Leave Infection Assays
Wild-type and mutant strains were cultured on PDA for 2 days. Five-mm agar plugs were taken from the edge of a colony and were inoculated in the armature wound on the fourth or fifth leave from the top of a branch at 25°C for 3 days. Lesion diameters were recorded. For twig assays, the agar plugs were inoculated in the scald wound on 1-year old twigs of Malus domestica borkh. ‘Fuji’ at 25°C for 9 days. The lesion lengths were recorded (Wu et al., 2017b).
Nucleic Acid Isolation and Manipulation
VmPacC genes were originally identified through homology searches in the V. mali genomic sequence (Accession No. JUIY00000000.1) using the PacC amino acid sequence of A. nidulans as query (Q002.2.1). The neighbour-joining method was used to construct phylogenetic trees of Pal pathway proteins (Tamura et al., 2007). Supplementary Table S1 shows the pectinase genes annotated in the V. mali genome.
Gene disruption constructs were generated by replacing the complete open reading frame (ORF) of VmPacC gene (Supplementary Figure S2A). The primer pairs VmPacC-1F/2R and VmPacC-3F/4R were used to amplify the upstream and downstream flanking sequences, respectively, of the target gene (primer sequences are shown in Supplementary Table S2). The hygromycin resistance gene (hph) fragment was amplified with the primer pair HYG/F and HYG/R. Deletion cassettes were constructed using double-joint PCR as described (Yu et al., 2004). The deletion cassettes were transformed into protoplasts of the 03-8 strain using the polyethylene glycol (PEG) method as described (Gao et al., 2011). Following screening in hygromycin-containing medium, the transformants were tested using PCR with primer pairs VmPacC-5F/6R, VmPacC-7F/H855R, or H856F/VmPacC-8R to confirm gene replacement events (Supplementary Table S2). Putative gene deletion mutants were further confirmed using Southern blotting with target (probe p) and hygromycin (probe h) gene probes and the DIG-High Prime DNA Labelling and Detection Starter Kit II (Roche, Penzberg, Germany).
Complementation mutants and activated mutants were generated using the gap repair approach by co-transformation of VmPacC gene or activated fragments (Supplementary Figure S3A) amplified with primer pairs VmPacC-C (27)-F/R and ClaI-digested plasmid pFL2 into yeast strain XK1-25 (Zhou et al., 2012; Wu et al., 2017a). Trp+ yeast transformants were screened for the desired fusion constructs, which were confirmed by sequencing and transformed into the VmPacC deletion mutant. Geneticin-resistant transformants of complementation with the desired constructs were identified using PCR with primer pairs VmPacC-C-F/R and Southern blotting. The activated geneticin-resistant transformants were detected using PCR with primer pairs VmPacC27-F/R (Supplementary Figure S3B). The activated mutants were further identified by measuring the growth at pH 9 in unbuffered PDA for 3 days.
Transcript levels of VmPacC and pectinase-related genes were determined using real-time PCR. For expression of VmPacC in different pH, strains were cultured on PDA for 2 days. Five-mm agar plugs were taken from the edge of a colony and were cultured in a non-buffered PDB medium (pH 5.5) for 48 h. Mycelia were then transferred to fresh media buffered to pH 4, pH 7, or pH 9 for 6 h. The colonised apple tree bark was sampled as described (Ke et al., 2014). RNA was isolated with the TRIzol reagent (Invitrogen, Carlsbad, CA, United States) as described (Yin et al., 2013). For qRT-PCR assays, we used the Fermentas (Hanover, MD, United States) 1st strand cDNA synthesis kit following instructions of the manufacturer. The glyceraldehyde-6-phosphate dehydrogenase (G6PDH) gene of V. mali was used as internal control (Yin et al., 2013). Relative transcript levels of each gene were calculated by the 2-ΔΔCT method (Livak and Schmittgen, 2001). Data from three replicates were used to calculate means and standard deviations. Statistical analysis was done using the Student’s t-test implemented in the SAS software package (SAS Institute), P < 0.05. Primers used for gene expression are listed in Supplementary Table S2.
Pectinase Activity and Protein Concentration Assays
Five-mm agar plugs were taken from the edge of a colony and were cultured in PDB medium for 48 h. Mycelia were transferred to SM buffered to pH 4 or pH 7 and supplemented with 1% pectin as the sole carbon source for 12 h. Pectinase activity was quantified using the 3,5-dinitrosalicylic acid (DNS) method as described (Wu et al., 2017b).
Total extracellular protein content in culture supernatant was measured using the Bradford Protein Assay Kit (TIANGEN, China) with absorbance at 595 nm, and using bovine serum albumin as the standard.
pH and Citric Acid Measurements
The pH value of liquid PDB was measured with a pH electrode (Mettler Toledo, Shanghai, China) at different times after incubation. The pH of inoculation sites on the apple tree bark was measured using a flathead pH electrode as described (Schmidt et al., 2001). Three replicates were tested for each treatment.
High-performance liquid chromatography (HPLC) was used to determine citric acid content (Chinnici et al., 2005). Five-mm agar plugs were taken from the edge of a colony and were cultured in PDB medium for 48 h. Mycelia were transferred to SM supplemented with 0.5% saccharose as the sole carbon source for 48 h. The supernatant was filtered through a 0.22 μm cellulose acetate membrane before injection and measured using ion-exclusion chromatography. A Waters 600E HPLC apparatus with an attached refractive index detector (2414 RI) was used. Samples were separated in a Sepax Carbomix H-NP column under the following conditions: 2.5 mM H2SO4 mobile phase, 55°C column temperature, 0.6 mL/min flow-rate, and 30 min or less of analysis time. Peak data were collected with Empower LC solution. Citric acid standard (Solarbio, China) at 0.01–1 mg/mL was used for the standard curve.
Results
Gene Expression Analysis of VmPacC in V. mali
The V. mali genome contains single copy genes for pH-signalling pathway proteins designated VmPalA, VmPalB, VmPalC, VmPalF, VmPalI, VmPalH, and VmPacC. Analysis of the amino acid sequences revealed significant similarities to four fungi pH signalling pathway protein sequences (Supplementary Figure S1A). VmPacC codes for 560 amino acid residues interrupted by two introns, being smaller than the PacC/Rim101 orthologs of A. nidulans (678 aa) and yeast (635 aa). In addition, VmPacC contains classical zinc-finger domain and Zinc-finger double domains at its N-terminal region (residues 116–138 and 102–127) (Supplementary Figure S1B).
Expression of VmPacC in different pH and infection conditions was determined. After transferring to different pH for 6 h, qRT-PCR analysis showed that VmPacC gene expression was upregulated 9.9- and 17-fold in pH 7 and pH 9, respectively, compared to that of pH 4 (Figure 1A). These results show the important regulatory role of this gene in alkaline conditions. Upregulated VmPacC expression was also observed during infection. After inoculating V. mali in 1 year-old twigs, the VmPacC gene expression increased and reached a peak 12 h post-inoculation, and then decreased and remained stable at twofold (Figure 1B). These results suggest that VmPacC might be involved in V. mali virulence.
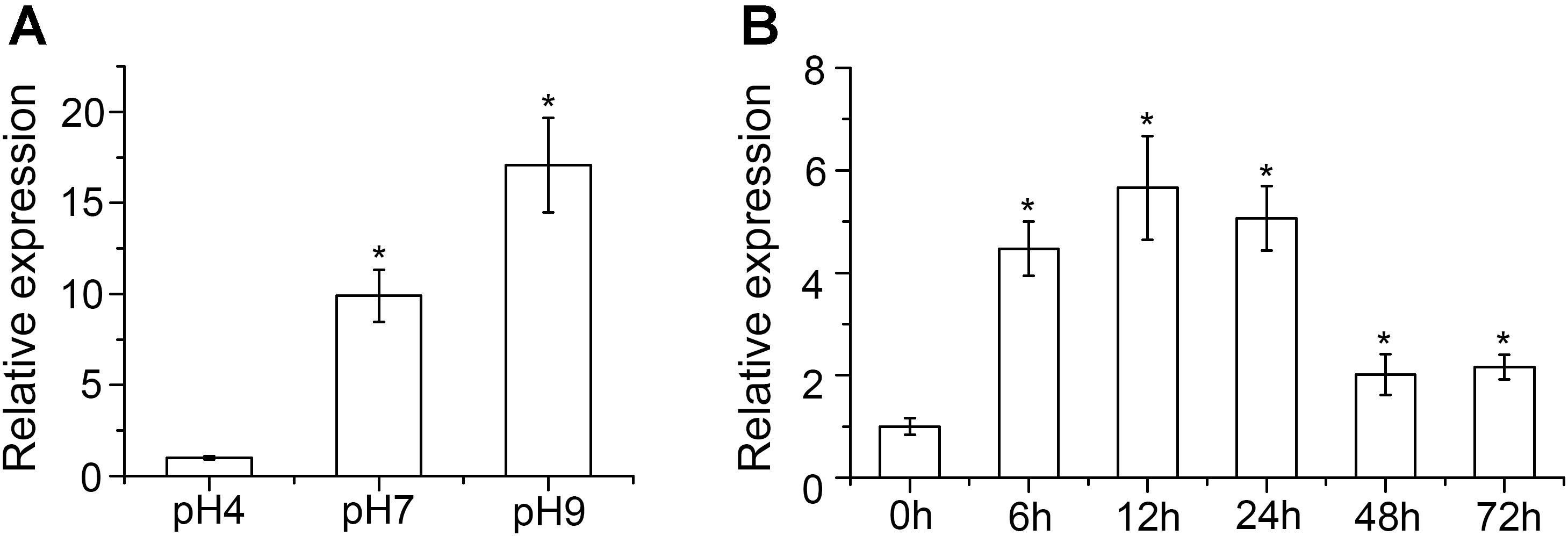
FIGURE 1. Expression profiles of VmPacC in Valsa mali. (A) Quantitative real-time PCR analysis of VmPacC expression in fungus grown at different pH conditions. RNA samples were isolated from mycelia of the wild-type strain on potato dextrose broth (PDB) medium at pH 4, 7, and 9 after pre-growth on unbuffered PDB for 48 h. (B) Quantitative real-time PCR analysis of VmPacC expression during infection. RNA samples were isolated from apple tree barks around the inoculation site at 0, 6, 12, 24, 48, and 72 h post-inoculation. Transcript abundances (mean and standard deviation) were calculated using data from three independent biological replicates. The asterisk represents a statistically significant difference (P < 0.05).
VmPacC Is Required for Mycelial Growth at Alkaline pH
To investigate the roles of VmPacC genes in V. mali, we generated a deletion mutant in which the entire ORF of VmPacC was deleted. We constructed a gene replacement cassette by placing upstream and downstream flanking sequences next to hph resistance gene sequences (Supplementary Figure S2A). The cassette was introduced in protoplasts of the wild-type strain. Three VmPacC deletion mutants (ΔVmPacC) were obtained from 215 hygromycin-resistant transformants and identified using PCR (Supplementary Figure S2B). When hybridised with the VmPacC ORF probe (probe p), a 5.6-kb ClaI fragment was detected in the wild-type strain but not in the ΔVmPacC mutant. In addition, ΔVmPacC showed a single locus homologous recombination event after hybridisation with the hygromycin probe (probe h) (Supplementary Figure S2C). Complementation mutant strains (ΔVmPacC-C) were also generated by re-introducing a wild-type allele of VmPacC in deletion mutants at an ectopic locus.
Wild-type, ΔVmPacC, and ΔVmPacC-C strains were grown on unbuffered PDA at pH 3 to pH 10. The wild-type strain of V. mali showed a more wide-range adaptation to pH 3 to pH 10. The colony diameters of the ΔVmPacC strain on PDA were similar to those of the wild-type at pH 3 to pH 5, but were reduced at pH 6 to pH 10. The deletion mutant hardly grew at pH 8 or higher (Figure 2). Normal growth was restored in the ΔVmPacC-C mutant, indicating that VmPacC regulates growth in alkaline conditions.
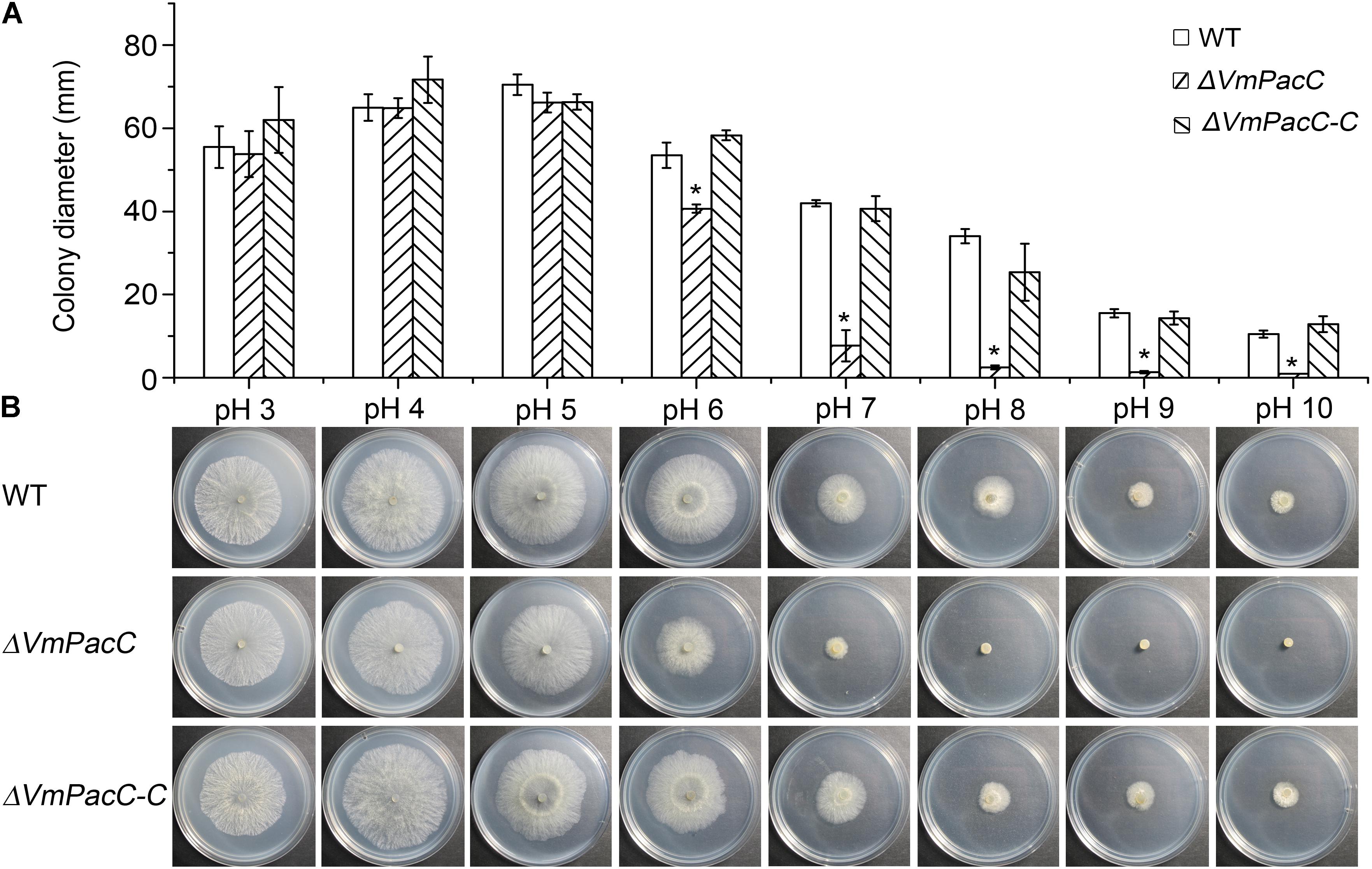
FIGURE 2. VmPacC is required for adaptation to ambient pH in V. mali. (A) Quantification of growth rate incubated for 2 days at 25°C. Results are the average of three independent experiments and standard deviations are shown. The asterisk represents a statistically significant difference compared to wild-type (P < 0.05). (B) Growth of wild-type, VmPacC deletion, and complemented mutant strains in unbuffered potato dextrose agar (PDA) adjusted to pH 3 to pH 10 with HCl or NaOH.
VmPacC Is Involved in the Oxidative Stress Response
To test whether VmPacC is involved in the response to abiotic stress, we measured the colony diameters of wild-type, ΔVmPacC, and ΔVmPacC-C strains on PDA supplemented with NaCl, H2O2, CR, or SDS. Results showed that the growth inhibition of NaCl on the ΔVmPacC mutant was higher than that on the wild-type or complemented mutant strains. However, the sensitivity of the VmPacC deletion mutant was reduced on PDA supplemented with H2O2. In addition, no significant differences were observed between wild-type, ΔVmPacC, and ΔVmPacC-C on PDA supplemented with CR or SDS (Figure 3). These results suggest that VmPacC is involved in the response to oxidative stress.
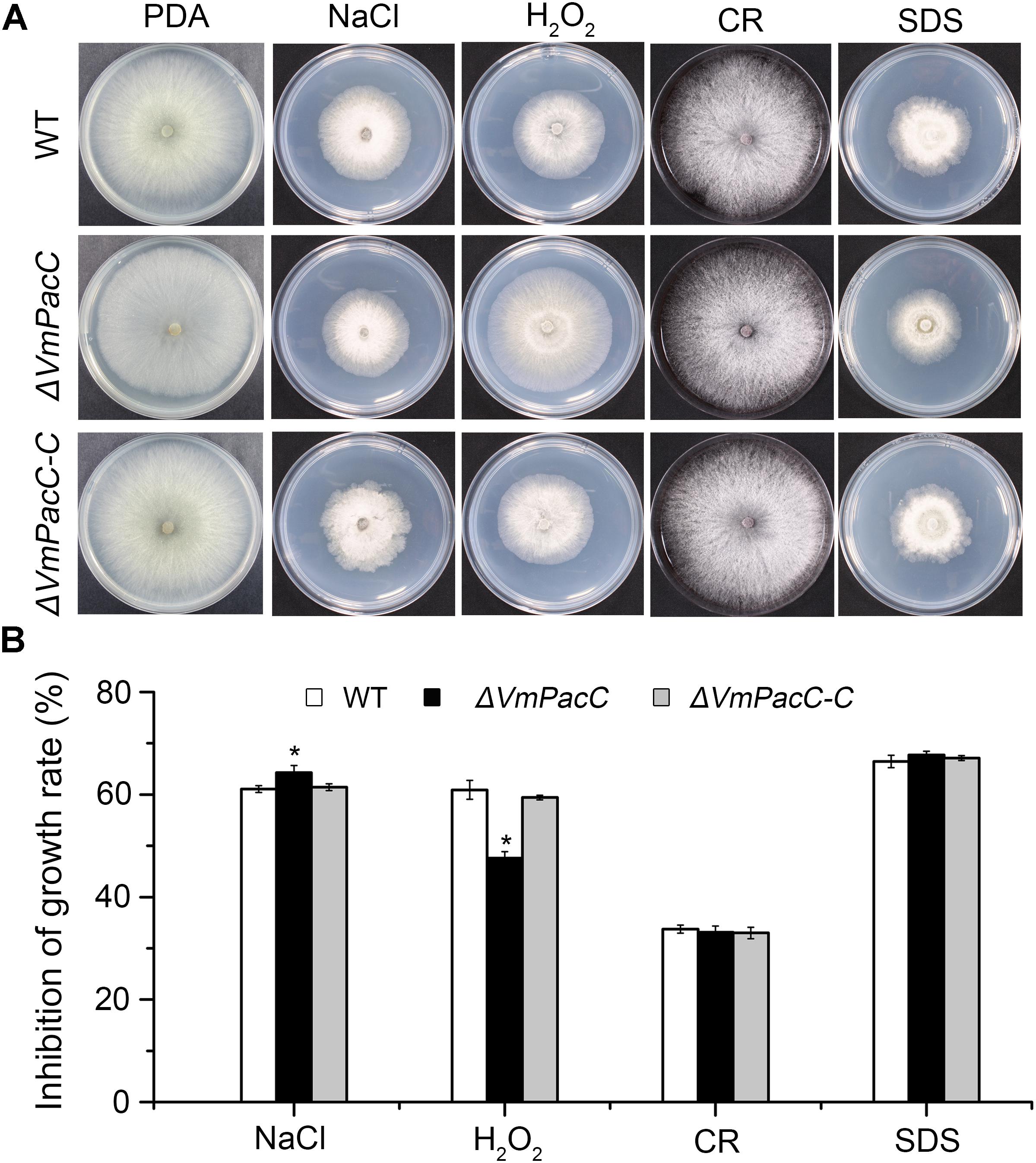
FIGURE 3. Effects of inactivation of VmPacC gene on the response of V. mali to stress. (A) Cultures of the wild-type, VmPacC gene null mutant, and complemented strains grown on PDA supplemented with 1 M sorbitol, 0.1 M NaCl, 3 mM H2O2, 300 mg/L Congo red (CR), or 0.01% SDS. Images were taken after 2.5 days on PDA and 3.5 days in inhibitor medium. (B) Inhibition of growth (colony diameter) on PDA supplemented with inhibitor compared with PDA without stress. The asterisk represents a statistically significant difference compared to wild-type (P < 0.05). Bars indicate standard deviations of the mean of three replicates.
VmPacC Is Required for Virulence
To determine whether VmPacC plays an important role in disease development, virulence assays were performed on leaves and twigs using the wild-type, ΔVmPacC, and ΔVmPacC-C strains. The diameter of the lesions in leaves inoculated with the wild-type was 34 mm at 3 dpi, whereas that of ΔVmPacC-inoculated leaves was 17 mm. When twigs were used as host, the length of lesions was also smaller in samples infected with the ΔVmPacC strain. In addition, infection with the complementation strains restored the wild-type phenotype (Figure 4). These results clearly indicate that VmPacC plays a significant role in controlling virulence in V. mali.
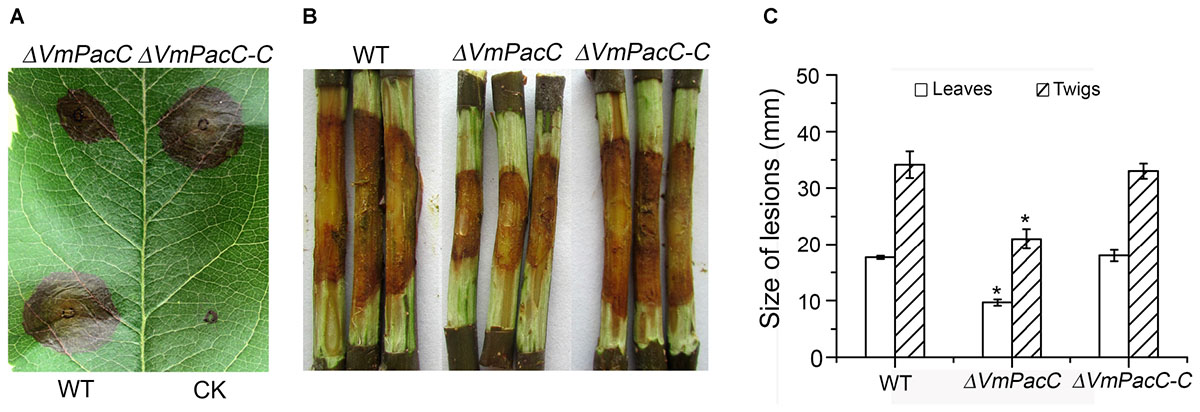
FIGURE 4. Phenotype of leaves and twigs inoculated with VmPacC deleted mutants of V. mali. (A) Apple leaves were inoculated with mycelium agar plugs from the wild-type, deletion mutant, and complemented strains. Lesion sizes are shown at 3 days post-inoculation (dpi). (B) Apple twigs were scald inoculated with mycelium agar plugs from the wild-type, deletion mutant, and complemented mutant strains. Lesion sizes are shown at 9 dpi. (C) Quantitation of the lesion size on apple leaves is lesion diameter at 3 dpi. Quantitation of the lesion size on twigs is lesion length at 9 dpi. The asterisks represent a statistically significant difference compared to wild-type (P < 0.05). Bars indicate standard deviations of the mean of three replicates.
VmPacC Suppresses the Production of Pectinase
To further evaluate the role of VmPacC on the virulence factor pectinase, we determined the transcript levels of a dozen pectinase genes in the deletion mutant and compared to those of the wild-type strain in colonised apple tree bark. Unexpectedly, transcript levels of most pectinase genes increased to different degrees (Figure 5). These results suggest that VmPacC negatively regulates pectinase expression.
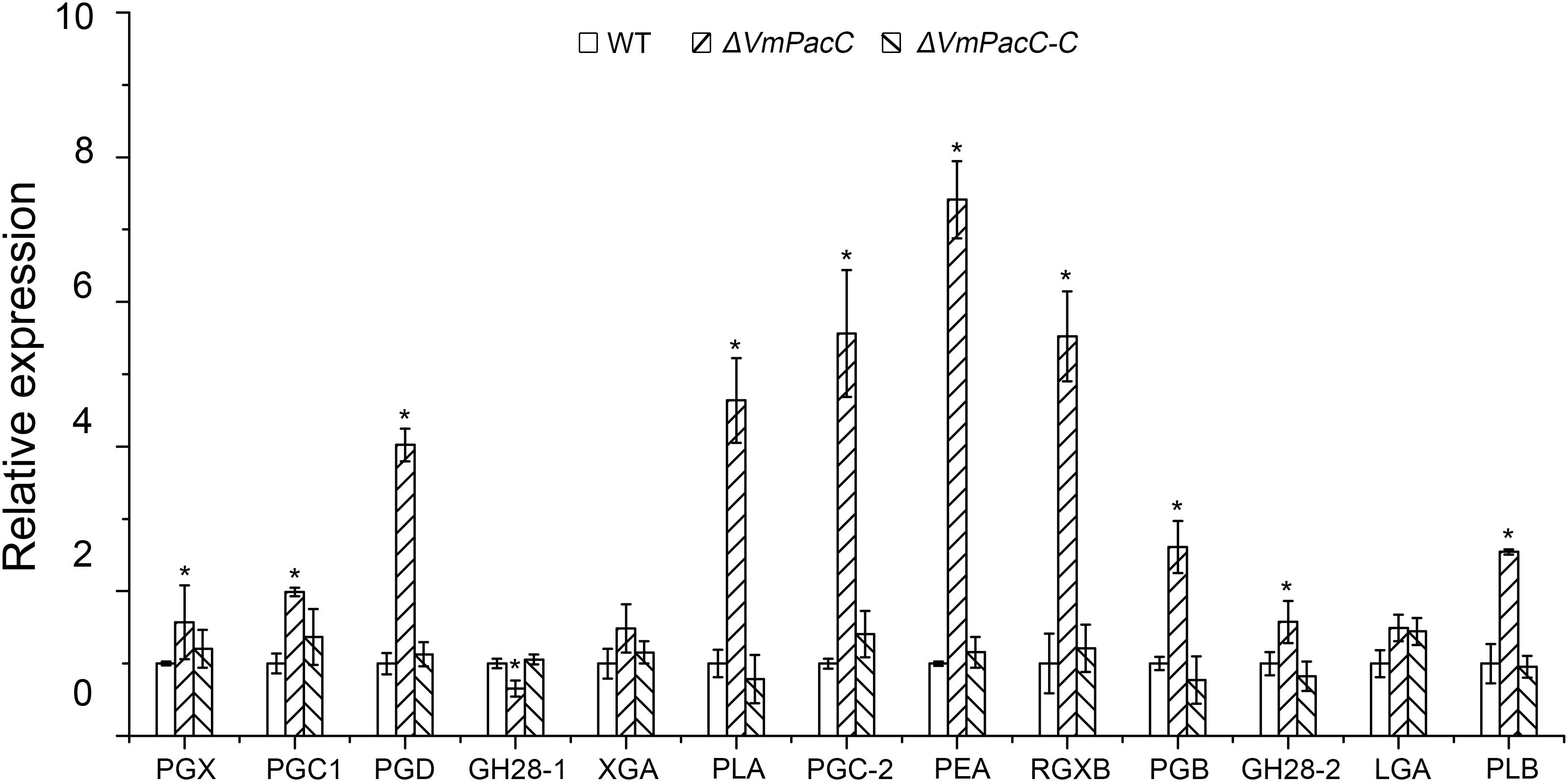
FIGURE 5. Expression levels of pectinase genes in wild-type, deletion mutant, and complemented mutant strains determined by qRT-PCR. RNA samples were isolated from lesion borders of apple tree bark at 3 days post-inoculation. Wild-type expression levels were arbitrarily set to 1. The mean was calculated using data from three independent biological replicates. The asterisk represents a statistically significant difference compared to wild-type.
To further confirm the repressive effect of VmPacC on pectinase expression, we generated dominant activated allele mutants, strains C-27, which constitutively express VmPacC in an acid environment. C-27 strains showed normal growth compared to wild-type on alkaline PDA indicating the VmPacC was expressive (Supplementary Figure S3C). However, C-27 strains showed significantly reduced virulence (Figure 6A). When inoculated on solid medium with pectin as the sole carbon source, the growth rate of C-27 was reduced compared with the wild-type and ΔVmPacC strains (Figure 6B). To determine the requirement of VmPacC for pH-dependent expression of pectinase genes, pectinase transcript levels and activity in the wild-type, ΔVmPacC, and C-27 strains were assayed (Figures 6C,D). The results show that pectinase activity in the supernatant of C-27 strains was reduced compared to that of wild-type and ΔVmPacC strains at pH 4 (Figure 6E). At pH 7, the pectinase genes of ΔVmPacC were up regulated 3- to 7-fold, and protein content was also increased (Figure 6F). However, pectinase activity was similar in all strains possible because the optimum pH value of pectinases is 3.5 (Figure 6E) (Feng et al., 2017a). These results indicate that pectinases are suppressed by VmPacC.
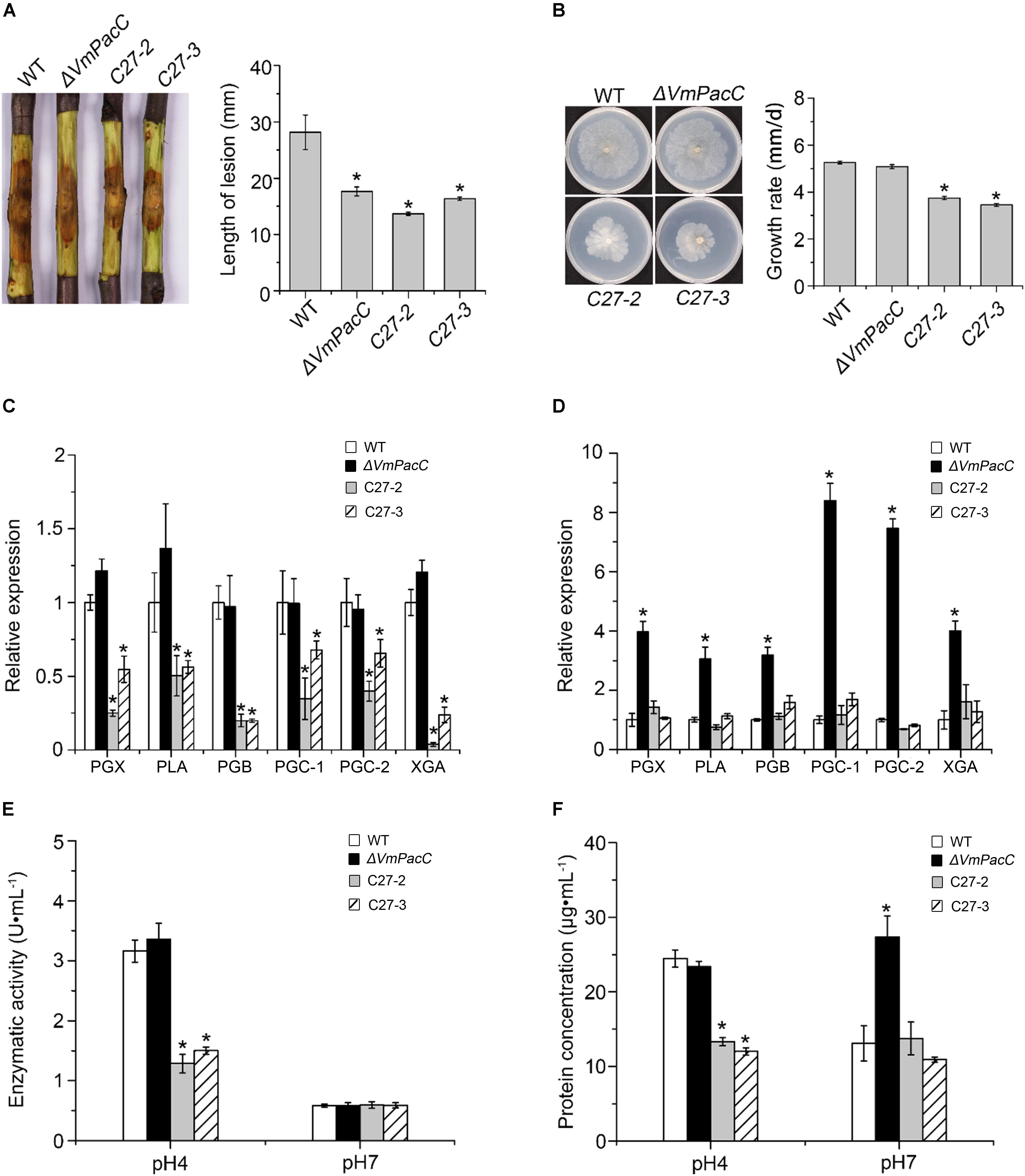
FIGURE 6. Negative regulation of pectinase production by VmPacC dominant activated mutants C27-2 and C27-3. (A) Phenotypes of twigs inoculated with wild-type, VmPacC deletion mutant, and activated mutants C27-2 and C27-3; length of lesions was measured at 9 days post-inoculation. (B) Mycelial growth on SM supplemented with pectin for 6 days. Expression levels of pectinase genes in wild-type, deletion mutant, and activated mutant strains cultured in SM medium supplemented with pectin at pH 4 (C) and pH 7 (D) determined using qRT-PCR. (E) Pectinase activity of different strains cultured in MS supplemented pectin for 12 h. (F) Assays for protein concentration of wild-type, deletion mutant, and activated mutant strains in 1% pectin inducing medium after culture for 12 h. Bars marked by asterisks in each group differ significantly from wild-type (LSD, P < 0.05). Error bars represent standard deviations from three replicates.
VmPacC Is Required for Acidification of the V. mali Environment
To investigate the involvement of VmPacC in the modification of the environment, the dynamic change in pH of the wild-type, ΔVmPacC, and ΔVmPacC-C strains was measured in an unbuffered liquid medium. The pH value of the wild-type strain decreased from the initial pH 5.8 to pH 3.4 in 120 h. Even though the same decrease in pH units was observed after 96 h in the VmPacC deletion mutant, the rate of descent was slower than in the wild-type (Figure 7A).
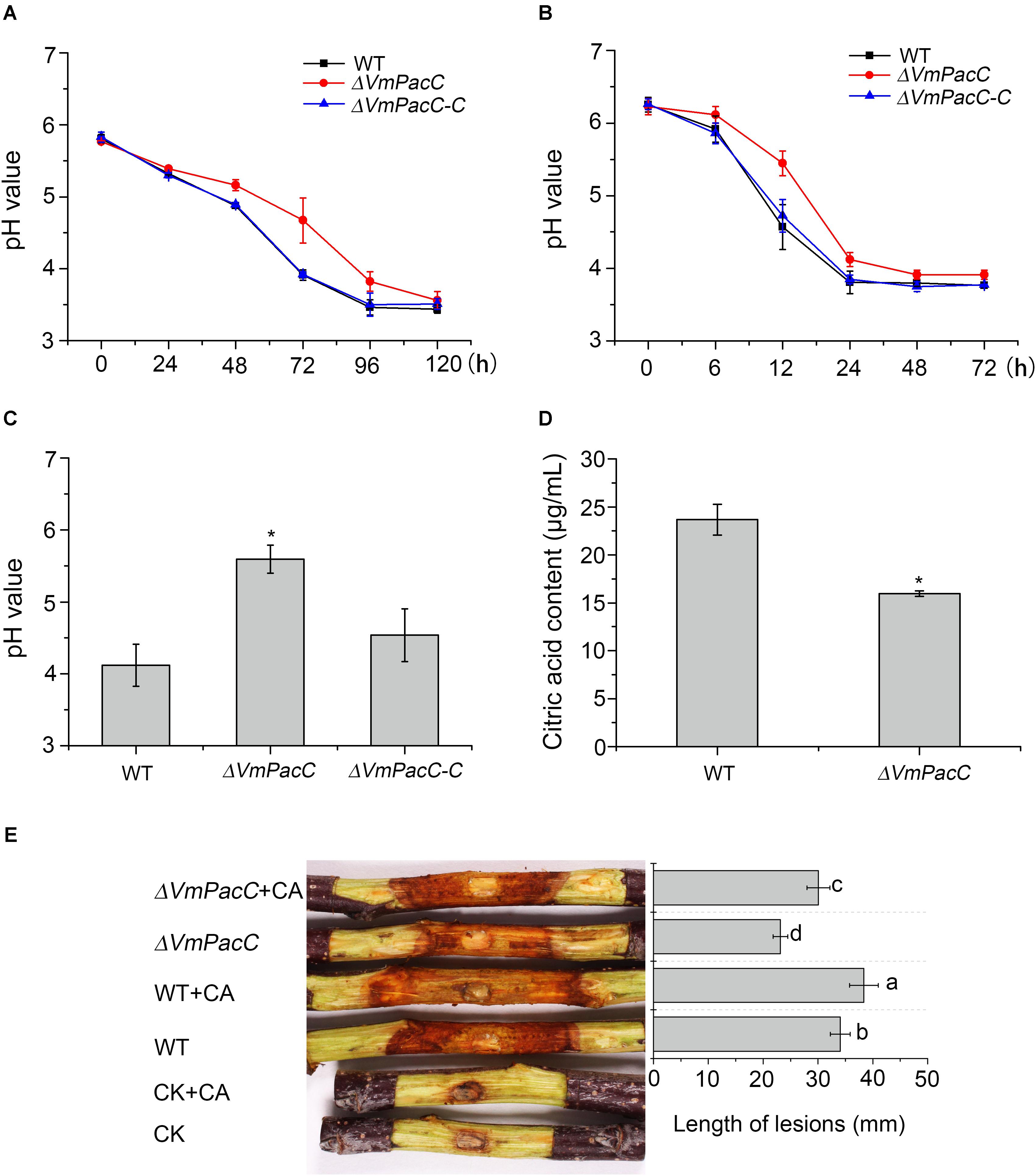
FIGURE 7. Valsa mali modulates its environmental pH. (A) Decrease in pH value during growth of different strains for 120 h. (B) pH values at the inoculated sites in the apple tree bark. (C) pH values at the lesion borders of the apple tree bark. (D) Citric acid production in wild-type and VmPacC deletion mutant strains grown in PDB determined by high-performance liquid chromatography (HPLC). The experiments were repeated three times. The asterisk represents a statistically significant difference compared to wild-type. (E) Virulence assays performed on twigs with wild-type and deletion mutant strains using citric acid (CA). The length of lesions was measured at 9 days post-inoculation. The experiments were repeated three times. Different letters represent a significant difference (P < 0.05).
When the wild-type strain of V. mali was inoculated in detached apple twigs, the initial pH at the inoculation sites was 6.2. The pH decreased to 3.8 when canker symptoms appeared 24 h after inoculation. Thereafter, the pH remained relatively stable. However, the same pH value (pH 3.9) was determined 48 h after inoculation in the VmPacC deletion mutant. The pH value of the deletion mutant decreased more slowly than in the wild-type strain. Furthermore, the pH value at the border of the lesion in the deletion mutant was higher than in the wild-type strain. The acidification capacity was impaired when VmPacC was deleted, and this effect could be reversed by complementation of the deletions (Figures 7B,C).
High-performance liquid chromatography analysis of organic acids secreted by V. mali indicated that the accumulation of citric acid was reduced in the VmPacC deletion mutant compared to wild-type (Figure 7D). To further test whether acidification of the environment contributes to virulence, we have inoculated the deletion mutant and externally treated with citric acid at pH 3.5. As expected, the virulence of the VmPacC deletion mutant and the wild-type strain increased when externally treated with citric acid (Figure 7E).
Our results clearly show that V. mali requires acidification of its environment for growth. In addition, VmPacC is involved in virulence partly via regulating the generation of citric acid.
Discussion
The Pal ambient pH response signalling pathways are essential for fungi to adapt to environmental and host conditions. This signalling cascade system ultimately activates the PacC transcription factor, which controls the environmental pH sensing and response (Peñalva et al., 2008). In this study, we found that the optimum pH of the apple bark pathogenic fungus, V. mali, is acid. Neutral or alkaline conditions did not favour mycelial growth, as indicated by the impaired vegetative growth of deletion mutant VmPacC at neutral and alkaline conditions (Figures 2A,B). These results indicate that VmPacC is required for V. mali adaptation to alkaline pH. This is consistent with what has been described in other fungi such as P. digitatum (Zhang et al., 2013), S. sclerotiorum (Rollins, 2003), F. graminearum (Merhej et al., 2011), and A. nidulans (Tilburn et al., 1995). In contrast, in Ustilago maydis, the null rim101 mutants show normal growth rate in vitro(Arechiga-Carvajal and Ruiz-Herrera, 2005). Moreover, in Metarhizium robertsii and M. oryzae, PacC affects growth under both acidic and alkaline conditions (Landraud et al., 2013; Huang et al., 2015). These results suggest that, in addition to adaptation to alkaline pH, alternative PacC signalling pathways may have evolved in some fungi.
We also found that VmPacC affected the sensitivity to salt and oxidative stress in V. mali (Figure 3). Similarly, deletion of PacC/Rim101 reduces ion tolerance because of repression of ion pump genes (Caracuel et al., 2003). In the insect pathogenic fungi M. robertsii, the reduced ion tolerance in ΔMrpacC results from smaller vacuoles (Huang et al., 2015). In pathogen–host interactions, the reactive oxygen species (ROS) play an important role. Plants use the oxidative burst as an early defence reaction to pathogen attack. Here, we show that VmPacC is involved in the sensitivity to oxidative stress. However, this issue is indeterminacy in necrotrophs. For example, in B. cinerea, deletion of the AP-1 transcription factor gene does not lead to impaired virulence (Temme and Tudzynski, 2009). Thus, for V. mali, the relation between abiotic stress responses and virulence needs further research.
The present study showed that deletion of the VmPacC gene significantly impaired the virulence of V. mali (Figure 4). Similar to what has been reported for PacC mutant strains of P. digitatum (Zhang et al., 2013), S. sclerotiorum (Rollins, 2003), C. gloeosporioides (Miyara et al., 2008), and M. oryzae (Landraud et al., 2013), PacC regulates the virulence in a positive manner in V. mali (Figure 3). However, PacC is a negative regulator of virulence in F. oxysporum and F. graminearum (Caracuel et al., 2003; Merhej et al., 2011). In addition, the pathogenicity of null rim101 mutants of U. maydis is not affected (Arechiga-Carvajal and Ruiz-Herrera, 2005). In phytopathogenic fungi, cell wall-degrading enzymes are important virulence factors during plant infection (Kubicek et al., 2014). The PacC is also a key regulator of secretion of these hydrolytic enzymes in several filamentous fungi (Miyara et al., 2010). In C. gloeosporioides, the reduced expression of a pectate lyase gene decreases virulence in a CgpacC-disrupted mutant (Miyara et al., 2008). In P. digitatum, the expression of cell wall degradation enzyme genes such as Pdpg2 and Pdpnl1 is regulated by PdPacC. In addition, this PdPacC regulation is associated with pathogenesis (Zhang et al., 2013). In S. sclerotiorum, the induced virulence of a PacC deletion mutant results from impaired expression of endopolygalacturonase (pg1) in higher ambient pH (Rollins, 2003). Pectinase is an important virulence factor in V. mali (Ke et al., 2013; Yin et al., 2015). Therefore, we determined the transcript levels of six pathogenesis-related pectinase genes and six up-regulated genes during infection (Yin et al., 2015). However, unexpectedly, the expression of most pectinase genes tested was up-regulated to different degrees in VmPacC deletion mutants (Figure 5). These results show that VmPacC is involved in repressing pectinase genes. This regulation model of virulence and pectinase expression is distinct from the above-mentioned mechanism. Therefore, VmPacC should not impact virulence via regulating the expression of pectinase genes.
To further investigate the repressive effect of VmPacC on pectinase gene expression, we generated dominant activated allele mutants. The mutants constitutively express PacC independent of pH. In agreement with our hypothesis, we found that dominant activated allele mutants showed significantly reduced growth on solid media supplemented with pectin as the sole carbon source. In addition, both the expression and activity of pectinase genes decreased and led to a reduction in the size of lesions on twigs. These results are consistent with the expression of two polygalacturonase genes, pg1 and pg5, being negatively regulated by PacC in F. oxysporum, although PacC functions as a negative regulator of virulence in this organism (Caracuel et al., 2003). In several fungi, PacC is responsible for up-regulation of alkaline-expressed genes and suppression of acid-expressed genes in alkaline pH (Alkan et al., 2013). Proteins need to be selectively expressed at the optimal external host pH environment to function correctly. We have demonstrated that the optimum pH value of pectinase activity is 3.5 (Feng et al., 2017a). The pectinase gene should be inhibited by PacC in neutral and alkaline conditions. Although transcript and protein levels in ΔVmPacC significantly increase at pH 7, the pectinase activity was the same as in the wild-type. These results indicate that the secreted pectinases do not contribute to pectinase activity in neutral and alkaline conditions.
In plant pathogens, the capacity to modify the ambient pH seems to be associated with infection of the host plants. Several phytopathogenic fungi acidify or alkalinise their surrounding pH during infection. Some plant pathogenic fungi such as S. sclerotiorum and P. digitatum are known to acidify their environment during infection via the secretion of oxalic acid or other organic acids (Rollins, 2003; Zhang et al., 2013). Other fungi, such as M. oryzae and Colletotrichum species, tend to secrete ammonia to alkalinise the invaded plant tissues (Landraud et al., 2013; Ment et al., 2015). In the present study, V. mali showed similar behaviour to S. sclerotiorum and P. digitatum. V. mali can acidify the surrounding or host pH, and the acidification capacity was impaired after deletion of VmPacC. We further demonstrated that VmPacC is required for the accumulation of citric acid in V. mali, and that the virulence of the wild-type strain and deletion mutant can be increased by externally adding citric acid to twigs. These results show that VmPacC is involved in virulence partly owing to the regulation of citric acid generation. However, whether citric acid contributes to virulence via another pathway remains to be elucidated.
Conclusion
We determined that the optimum pH of V. mali is acid, and that this organism can acidify its surrounding pH. In addition, the capacity to acidify the environmental pH is regulated by VmPacC, which is also involved in the response to oxidative stress and modulates expression of pectinase genes. More importantly, VmPacC is involved in virulence partly owing to the regulation of citric acid generation. In the future, comparison of the transcriptomes of the wild-type strain and the VmPacC deletion mutant of V. mali at different pH or during infection of apple bark will contribute to a better understanding of pathways involved in PacC-related pathogenesis.
Author Contributions
YW and LH conceived and designed the experiments and wrote the paper. YW and LX performed the experiments. YW, ZY, and HF analyzed the experiment data. ZY and LH contributed reagents, materials, and analysis tools. All authors have read and approved the final manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (Grant Nos. 31471732, 31671982, and 31772115).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.01981/full#supplementary-material
FIGURE S1 | Structure and sequence analysis of pH-signalling pathway proteins in Valsa mali. (A) Phylogenetic analysis of pH-signalling pathway proteins of V. mali and their homologs from Magnaporthe oryzae, Aspergillus nidulans, and Neurospora crassa. The amino acid sequences of Pal pathway proteins were analysed using MEGA version 6 and neighbour-joining analysis with 1,000 bootstrap replicates. Numbers on the branches represent the percentage of replicates supporting each branch. Subclades containing pH-signalling pathway proteins of V. mali and orthologs from other species are shaded. The bar represents 20% sequence divergence. (B) The open reading frame (ORF) of VmPacC consists of 1,825 bp, interrupted by two introns, and encodes a predicted protein of 560 amino acids with classical zinc-finger domain and zinc-finger double domain at its N-terminal region.
FIGURE S2 | Generation of VmPacC gene deletion mutants. (A) Generation of gene replacement constructs. The small arrows mark the position and direction of primers used for PCR. (B) For PCR detection of deletion transformants, four primer pairs (VmPacC-5F/6R, H850F/H852R, VmPacC-7F/H855F, and H856F/VmPacC-8R) were used to detect VmPacC gene deletion mutants. (C) Southern blots of ClaI-digested genomic DNA of wild-type, deletion mutant (ΔVmPacC), and complemented mutant (ΔVmPacC-C) strains hybridised with probe p (VmPacC ORF or probe h (hph ORF)).
FIGURE S3 | Construction of dominant activated mutant of VmPacC27. (A) Prediction of activated sequence with A. nidulans. (B) PCR detection of positive transformants of activated mutants with primer pairs VmPacC27-F/R. (C) Wild-type (WT) deletion and activated mutants grow at pH 9 in unbuffered potato dextrose agar for 3 days. The normal growth was restored in the activated mutants, indicating the VmPacC was expressive.
TABLE S1 | Pectinase genes for expression.
TABLE S2 | Primers used for deletion, complementation, and gene expression.
References
Abe, K., Kotoda, N., Kato, H., and Soejima, J. (2007). Resistance sources to Valsa canker (Valsa ceratosperma) in a germplasm collection of diverse Malus species. Plant Breed. 126, 449–453. doi: 10.1111/j.1439-0523.2007.01379.x
Alkan, N., Davydov, O., Sagi, M., Fluhr, R., and Prusky, D. (2009). Ammonium secretion by Colletotrichum coccodes activates host NADPH oxidase activity enhancing host cell death and fungal virulence in tomato fruits. Mol. Plant Microbe Interact. 22, 1484–1491. doi: 10.1094/MPMI-22-12-1484
Alkan, N., Meng, X., Friedlander, G., Reuveni, E., Sukno, S., Sherman, A., et al. (2013). Global aspects of pacC regulation of pathogenicity genes in Colletotrichum gloeosporioides as revealed by transcriptome analysis. Mol. Plant Microbe Interact. 26, 1345–1358. doi: 10.1094/MPMI-03-13-0080-R
Arechiga-Carvajal, E. T., and Ruiz-Herrera, J. (2005). The RIM101/pacC homologue from the basidiomycete Ustilago maydis is functional in multiple pH-sensitive phenomena. Eukaryot. Cell 4, 999–1008. doi: 10.1128/EC.4.6.999-1008.2005
Barad, S., Horowitz, S. B., Moscovitz, O., Lichter, A., Sherman, A., and Prusky, D. (2012). A Penicillium expansum glucose oxidase–encoding gene, GOX 2, is essential for gluconic acid production and acidification during colonization of deciduous fruit. Mol. Plant Microbe Interact. 25, 779–788. doi: 10.1094/MPMI-01-12-0002
Caracuel, Z., Casanova, C., Roncero, M. I. G., Di Pietro, A., and Ramos, J. (2003). pH Response transcription factor PacC controls salt stress tolerance and expression of the P-Type Na+-ATPase Ena1 in Fusarium oxysporum. Eukaryot. Cell 2, 1246–1252. doi: 10.1128/ec.2.6.1246-1252.2003
Chinnici, F., Spinabelli, U., Riponi, C., and Amati, A. (2005). Optimization of the determination of organic acids and sugars in fruit juices by ion-exclusion liquid chromatography. J. Food Compos. Anal. 18, 121–130. doi: 10.1016/j.jfca.2004.01.005
Díez, E., Álvaro, J., Espeso, E. A., Rainbow, L., Suárez, T., Tilburn, J., et al. (2002). Activation of the Aspergillus PacC zinc finger transcription factor requires two proteolytic steps. Embo. J. 21, 1350–1359. doi: 10.1093/emboj/21.6.1350
Eshel, D., Miyara, I., Ailing, T., Dinoor, A., and Prusky, D. (2002). pH regulates endoglucanase expression and virulence of Alternaria alternata in persimmon fruit. Mol. Plant Microbe Interact. 15, 774–779. doi: 10.1094/MPMI.2002.15.8.774
Feng, H., He, Y., Zhen, W., Gao, X., Wang, H., and Huang, L. (2017a). Isolation, purification and characterization of extracellular pectinase produced by Valsa mali. Microbiol. China 44, 639–647. doi: 10.13344/j.microbiol.china.160236
Feng, H., Xu, M., Liu, Y., Dong, R., Gao, X., and Huang, L. (2017b). Dicer-Like genes are required for H2O2 and KCl stress responses, pathogenicity and small rna generation in Valsa mali. Front. Microbiol. 8:1166. doi: 10.3389/fmicb.2017.01166
Gao, J., Li, Y., Ke, X., Kang, Z., and Huang, L. (2011). Development of genetic transformation system of Valsa mali of apple mediated by PEG. Act. Microbiol. Sin. 51, 1194–1199. doi: 10.13343/j.cnki.wsxb.2011.09.007
Hervas-Aguilar, A., Rodriguez, J. M., Tilburn, J., Arst, H. N. Jr., and Penalva, M. A. (2007). Evidence for the direct involvement of the proteasome in the proteolytic processing of the Aspergillus nidulans zinc finger transcription factor PacC. J. Biol. Chem. 282, 34735–34747. doi: 10.1074/jbc.M706723200
Huang, W., Shang, Y., Chen, P., Gao, Q., and Wang, C. (2015). MrpacC regulates sporulation, insect cuticle penetration and immune evasion in Metarhizium robertsii. Environ. Microbiol. 17, 994–1008. doi: 10.1111/1462-2920.12451
Ke, X., Huang, L., Han, Q., Gao, X., and Kang, Z. (2013). Histological and cytological investigations of the infection and colonization of apple bark by Valsa mali var. mali. Australas. Plant. Pat. 42, 85–93. doi: 10.1007/s13313-012-0158-y
Ke, X., Yin, Z., Song, N., Dai, Q., Voegele, R. T., Liu, Y., et al. (2014). Transcriptome profiling to identify genes involved in pathogenicity of Valsa mali on apple tree. Fungal Genet. Biol. 68, 31–38. doi: 10.1016/j.fgb.2014.04.004
Kramer-Haimovich, H., Servi, E., Katan, T., Rollins, J., Okon, Y., and Prusky, D. (2006). Effect of ammonia production by Colletotrichum gloeosporioides on pelB activation, pectate lyase secretion, and fruit pathogenicity. Appl. Environ. Microbiol. 72, 1034–1039. doi: 10.1128/AEM.72.2.1034-1039.2006
Kubicek, C. P., Starr, T. L., and Glass, N. L. (2014). Plant cell wall-degrading enzymes and their secretion in plant-pathogenic fungi. Annu. Rev. Phytopathol. 52, 427–451. doi: 10.1146/annurev-phyto-102313-045831
Landraud, P., Chuzeville, S., Billon-Grande, G., Poussereau, N., and Bruel, C. (2013). Adaptation to pH and role of PacC in the rice blast fungus Magnaporthe oryzae. PLoS One 8:e69236. doi: 10.1371/journal.pone.0069236
Lee, D. H., Lee, S. W., Choi, K. H., Kim, D. A., and Uhm, J. Y. (2006). Survey on the occurrence of apple diseases in Korea from 1992 to 2000. Plant Pathol. J. 22, 375. doi: 10.5423/PPJ.2006.22.4.375
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT Method. Methods 25, 402–408. doi: 10.1006/meth.2001.1262
Ment, D., Alkan, N., Luria, N., Bi, F.-C., Reuveni, E., Fluhr, R., et al. (2015). A role of AREB in the regulation of PACC-dependent acid-expressed-genes and pathogenicity of Colletotrichum gloeosporioides. Mol. Plant Microbe Interact. 28, 154–166. doi: 10.1094/MPMI-09-14-0252-R
Merhej, J., Richard-Forget, F., and Barreau, C. (2011). The pH regulatory factor Pac1 regulates Tri gene expression and trichothecene production in Fusarium graminearum. Fungal Genet. Biol. 48, 275–284. doi: 10.1016/j.fgb.2010.11.008
Miyara, I., Shafran, H., Davidzon, M., Sherman, A., and Prusky, D. (2010). pH regulation of ammonia secretion by Colletotrichum gloeosporioides and its effect on appressorium formation and pathogenicity. Mol. Plant Microbe Interact 23, 304–316. doi: 10.1094/MPMI-23-3-0304
Miyara, I., Shafran, H., Kramer Haimovich, H., Rollins, J., Sherman, A., and Prusky, D. (2008). Multi-factor regulation of pectate lyase secretion by Colletotrichum gloeosporioides pathogenic on avocado fruits. Mol. Plant Pathol. 9, 281–291. doi: 10.1111/j.1364-3703.2007.00462.x
Penalva, M. A., and Arst, H. N. (2002). Regulation of gene expression by ambient pH in Filamentous Fungi and Yeasts. Microbiol. Mol. Biol. 66, 426–446. doi: 10.1128/mmbr.66.3.426-446.2002
Penalva, M. A., and Espeso, E. A. (1996). Three binding sites for the Aspergillus nidulans PacC Zinc-finger transcription factor are necessary and sufficient for regulation by ambient pH of the isopenicillin N synthase gene promoter. J. Biol. Chem. 271, 28825–28830. doi: 10.1074/jbc.271.46.28825
Peñalva, M. A., Tilburn, J., Bignell, E., and Arst, H. N. (2008). Ambient pH gene regulation in fungi: making connections. Trends Microbiol. 16, 291–300. doi: 10.1016/j.tim.2008.03.006
Rollins, J. A. (2003). The Sclerotinia sclerotiorum pac1 gene is required for sclerotial development and virulence. Mol. Plant Microbe Interact. 16, 785–795. doi: 10.1094/MPMI.2003.16.9.785
Rollins, J. A., and Dickman, M. B. (2001). pH signaling in Sclerotinia sclerotiorum: identification of a pacC/RIM1 homolog. Appl. Environ. Microbiol. 67, 75–81. doi: 10.1128/AEM.67.1.75-81.2001
Schmidt, J., Kricke, R., and Feige, G. B. (2001). Measurements of bark pH with a modified flathead electrode. Lichenologist 33, 456–460. doi: 10.1006/lich.2001.0341
Srivastava, A., Ohm, R. A., Oxiles, L., Brooks, F., Lawrence, C. B., Grigoriev, I. V., et al. (2012). A zinc-finger-family transcription factor, AbVf 19, is required for the induction of a gene subset important for virulence in Alternaria brassicicola. Mol. Plant Microbe Interact. 25, 443–452. doi: 10.1094/MPMI-10-11-0275
Tamura, K., Dudley, J., Nei, M., and Kumar, S. (2007). MEGA4: molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol. Biol. Evol. 24, 1596–1599. doi: 10.1093/molbev/msm092
Temme, N., and Tudzynski, P. (2009). Does Botrytis cinerea ignore H2O2-induced oxidative stress during infection? Characterization of Botrytis activator protein 1. Mol. Plant Microbe Interact. 22, 987–998. doi: 10.1094/MPMI-22-8-0987
Tilburn, J., Sarkar, S., Widdick, D., Espeso, E., Orejas, M., Mungroo, J., et al. (1995). The Aspergillus PacC zinc finger transcription factor mediates regulation of both acid-and alkaline-expressed genes by ambient pH. Embo. J. 14:779. doi: 10.1002/j.1460-2075.1995.tb07056.x
Wang, X., Zang, R., Yin, Z., Kang, Z., and Huang, L. (2014). Delimiting cryptic pathogen species causing apple Valsa canker with multilocus data. Ecol. Evol. 4, 1369–1380. doi: 10.1002/ece3.1030
Wu, Y., Xu, L., Liu, J., Yin, Z., Gao, X., Feng, H., et al. (2017a). A mitogen-activated protein kinase gene (VmPmk1) regulates virulence and cell wall degrading enzyme expression in Valsa mali. Microb. Pathog. 111, 298–306. doi: 10.1016/j.micpath.2017.09.003
Wu, Y., Xu, L., Yin, Z., Dai, Q., Gao, X., Feng, H., et al. (2017b). Two members of the velvet family, VmVeA and VmVelB, affect conidiation, virulence and pectinase expression in Valsa mali. Mol. Plant Pathol 19, 1639–1651. doi: 10.1111/mpp.12645
Wubben, J. P., ten Have, A., van Kan, J. A., and Visser, J. (2000). Regulation of endopolygalacturonase gene expression in Botrytis cinerea by galacturonic acid, ambient pH and carbon catabolite repression. Curr. Genet 37, 152–157. doi: 10.1007/s002940050022
Yakoby, N., Kobiler, I., Dinoor, A., and Prusky, D. (2000). pH Regulation of Pectate Lyase Secretion Modulates the Attack of Colletotrichum gloeosporioides on Avocado Fruits. Appl. Environ. Microbiol. 66, 1026–1030. doi: 10.1128/aem.66.3.1026-1030.2000
Yin, Z., Ke, X., Huang, D., Gao, X., Voegele, R. T., Kang, Z., et al. (2013). Validation of reference genes for gene expression analysis in Valsa mali var. mali using real-time quantitative PCR. World J. Microbiol. Biotechnol. 29, 1563–1571. doi: 10.1007/s11274-013-1320-6
Yin, Z., Liu, H., Li, Z., Ke, X., Dou, D., Gao, X., et al. (2015). Genome sequence of Valsa canker pathogens uncovers a potential adaptation of colonization of woody bark. New Phytol. 208, 1202–1216. doi: 10.1111/nph.13544
Yu, J. H., Hamari, Z., Han, K. H., Seo, J. A., Reyes-Dominguez, Y., and Scazzocchio, C. (2004). Double-joint PCR: a PCR-based molecular tool for gene manipulations in filamentous fungi. Fungal Genet. Biol. 41, 973–981. doi: 10.1016/j.fgb.2004.08.001
Zhang, T., Sun, X., Xu, Q., Candelas, L. G., and Li, H. (2013). The pH signaling transcription factor PacC is required for full virulence in Penicillium digitatum. Appl. Microbiol. Biotechnol. 97, 9087–9098. doi: 10.1007/s00253-013-5129-x
Keywords: Malus domestica, deletion mutant, pH regulation, pectinase, virulence
Citation: Wu Y, Yin Z, Xu L, Feng H and Huang L (2018) VmPacC Is Required for Acidification and Virulence in Valsa mali. Front. Microbiol. 9:1981. doi: 10.3389/fmicb.2018.01981
Received: 15 April 2018; Accepted: 06 August 2018;
Published: 23 August 2018.
Edited by:
Brigitte Mauch-Mani, University of Neuchâtel, SwitzerlandCopyright © 2018 Wu, Yin, Xu, Feng and Huang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lili Huang, aHVhbmdsaWxpQG53c3VhZi5lZHUuY24=