
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Microbiol., 27 August 2018
Sec. Terrestrial Microbiology
Volume 9 - 2018 | https://doi.org/10.3389/fmicb.2018.01929
This article is part of the Research TopicElucidating Microbial Processes in Soils and Sediments: Microscale Measurements and ModelingView all 24 articles
 Philippe C. Baveye1*
Philippe C. Baveye1* Wilfred Otten2
Wilfred Otten2 Alexandra Kravchenko3
Alexandra Kravchenko3 María Balseiro-Romero1,4
María Balseiro-Romero1,4 Éléonore Beckers5
Éléonore Beckers5 Maha Chalhoub6Christophe Darnault7
Maha Chalhoub6Christophe Darnault7 Thilo Eickhorst8Patricia Garnier6Simona Hapca9Serkan Kiranyaz10Olivier Monga11
Thilo Eickhorst8Patricia Garnier6Simona Hapca9Serkan Kiranyaz10Olivier Monga11 Carsten W. Mueller12
Carsten W. Mueller12 Naoise Nunan13
Naoise Nunan13 Valérie Pot6
Valérie Pot6 Steffen Schlüter14
Steffen Schlüter14 Hannes Schmidt15
Hannes Schmidt15 Hans-Jörg Vogel14,16
Hans-Jörg Vogel14,16Over the last 60 years, soil microbiologists have accumulated a wealth of experimental data showing that the bulk, macroscopic parameters (e.g., granulometry, pH, soil organic matter, and biomass contents) commonly used to characterize soils provide insufficient information to describe quantitatively the activity of soil microorganisms and some of its outcomes, like the emission of greenhouse gasses. Clearly, new, more appropriate macroscopic parameters are needed, which reflect better the spatial heterogeneity of soils at the microscale (i.e., the pore scale) that is commensurate with the habitat of many microorganisms. For a long time, spectroscopic and microscopic tools were lacking to quantify processes at that scale, but major technological advances over the last 15 years have made suitable equipment available to researchers. In this context, the objective of the present article is to review progress achieved to date in the significant research program that has ensued. This program can be rationalized as a sequence of steps, namely the quantification and modeling of the physical-, (bio)chemical-, and microbiological properties of soils, the integration of these different perspectives into a unified theory, its upscaling to the macroscopic scale, and, eventually, the development of new approaches to measure macroscopic soil characteristics. At this stage, significant progress has been achieved on the physical front, and to a lesser extent on the (bio)chemical one as well, both in terms of experiments and modeling. With regard to the microbial aspects, although a lot of work has been devoted to the modeling of bacterial and fungal activity in soils at the pore scale, the appropriateness of model assumptions cannot be readily assessed because of the scarcity of relevant experimental data. For significant progress to be made, it is crucial to make sure that research on the microbial components of soil systems does not keep lagging behind the work on the physical and (bio)chemical characteristics. Concerning the subsequent steps in the program, very little integration of the various disciplinary perspectives has occurred so far, and, as a result, researchers have not yet been able to tackle the scaling up to the macroscopic level. Many challenges, some of them daunting, remain on the path ahead. Fortunately, a number of these challenges may be resolved by brand new measuring equipment that will become commercially available in the very near future.
Over the last decade, soils have become increasingly central to a number of crucial debates on issues of great societal concern. Because they contain a huge amount of carbon, soils could lead to a dramatic acceleration of global climate change, as mean temperatures increase and rainfall patterns are altered (Baveye, 2007; Baveye et al., 2011; Hamdi et al., 2013; Crowther et al., 2016). The idea, advocated by some (Paustian et al., 1997; Lal and Bruce, 1999), that on the contrary, with proper management, soils could store even more carbon than at present, and thereby help mitigate the production of greenhouse gasses resulting from the consumption of fossil fuels, has been adopted enthusiastically by politicians in a number of countries but has stirred intense discussions among scientists (Powlson et al., 2011; Cheng et al., 2012; Dungait et al., 2012; Kowalchuk, 2012; Verbruggen et al., 2012; Minasny et al., 2017, 2018; van Groenigen et al., 2017; Baveye et al., 2018; White et al., 2018). At the same time, humanity is faced with the prospect of having to significantly increase food production to feed the world population, which is expected to rise to 9 or 10 billion people by 2050 (Godfray et al., 2010). Since soil and water resources are already used at the maximum level of what some consider ecologically safe, a consensus seems to be emerging that as long as the focus is kept on land-based agricultural production, the best option to insure food security lies in exploiting plant-microbe partnerships to improve biomass production (Weyens et al., 2009; Glick, 2012, 2014; Blaser et al., 2016), or in stimulating so-called plant-soil feedback processes, whereby plants induce soil microbial communities to release nutrients and store water in the rhizosphere (Sposito, 2013; Baveye, 2015). In addition, even though the issue of soil contamination does not appear at the moment to be at the forefront of environmental concerns in many countries, the question remains of what to do with millions of severely polluted sites around the globe, especially given the fact that this number is ever increasing, as a result of practices like shale gas production (Baveye, 2013c; Meckenstock et al., 2015). Given the prospect of a progressive warming of soils in decades to come, renewed threats caused by soil contamination will undoubtedly need to be addressed at some point in the near future.
The intimately connected microbial and physico-chemical processes at the core of all these soil-related issues have posed daunting challenges to researchers. Until a decade ago, in spite of sustained research efforts, progress was very slow or even non-existent, and in several cases serious hurdles arose, which no one had anticipated. Kirschbaum (2006) admitted that in the 10 years prior to the publication of his review of the field, no real advance had been made in understanding and predicting quantitatively the effect of temperature on the decomposition of soil organic matter (OM). Available models also routinely underestimated the pulses of CO2 flux occurring when large rainfall events follow drafts (Blagodatsky and Smith, 2012; Evans et al., 2016). Recent work by Rabot et al. (2015) suggests that many of the previous measurements of the production by soil bacteria and fungi of nitrous oxide, a very potent greenhouse gas (Laughlin and Stevens, 2002; Crenshaw et al., 2008; Hu et al., 2015), probably missed very short emission bursts that occur at the onset of drying of soils, and therefore underestimated total N2O production by soils. Concomitantly, research on carbon sequestration in soils provided evidence of the problematic “priming” effect, identified early on (Macura et al., 1965; Arsjad and Giddens, 1966), but routinely overlooked until a decade ago (Fontaine et al., 2007; Kuzyakov, 2010; Tian et al., 2015) and still poorly understood (Nunan et al., 2015; van der Wal and de Boer, 2017). Through this effect, the addition of fresh OM to soils can lead to the mineralization of very old humic substances, previously thought to be utterly stable and recalcitrant to further degradation. In a similar fashion, in polluted soils, experiments showed that a slight change, for example brought about by the addition of a source of nutrients for microorganisms, could easily make supposedly “sequestered” contaminants once again bioavailable (Li et al., 2005). Some of these areas of ignorance remain “terra incognita” at this point, even with regard to the much ballyhooed biodiversity of soils (Baveye et al., 2016a,b). There is still no satisfactory explanation for the observation, made more than 60 years ago, that the mineralization of soil OM continues at the same rate even if 90% of soil microorganisms are wiped out by CHCl3 fumigation (Jenkinson, 1966; Powlson et al., 2017; Baveye, 2018). A final example of a situation where our understanding of soil systems is still insufficient is related to the links between the diversity of soil microbial communities and various soil parameters. Some authors have found a close correlation between this diversity and specific parameters, like soil pH (Fierer and Jackson, 2006), but more detailed statistical analyses sometimes present a different picture. In a recent study, Terrat et al. (2017) use some of the most sophisticated molecular techniques currently available to analyze the biodiversity of soil samples across France, and try to relate it to various parameters of soils and of their environment. The results are systematically underwhelming. They find that less than half (48.2%) of the observed variance of the biodiversity could be accounted for by using soil parameters that are routinely measured. Clearly, at least in this particular study, something fundamental about soils is being missed.
In virtually all these instances, a common observation is that soil samples that appear alike in most of their overall measured characteristics can behave very differently, making replicated observations and good correlations difficult to achieve. Obviously, it is not sufficient to describe soils solely on the basis of traditional macroscopic measurements, such as the volumetric water content, microbial density, or contaminant concentration. Quantitative information on the spatial heterogeneity manifested at the micron scale, at which microorganisms operate, is also absolutely required.
In some respects, this is not as novel a perspective as it may appear. In another era, in literature that unfortunately seems to have become largely ignored since, soil microbiologists already reached the same conclusion. Sixty years ago, Rovira and Greacen (1957) subjected moist soil samples to compression and shearing to simulate tillage, and concluded, after ruling out other possible explanations, that the enhanced oxygen consumption observed in the soils after disruption was due to exposure of organisms to OM that was previously inaccessible to them. These and a number of other early observations pointing in the same direction prompted Alexander (1964, p. 219) to conclude that “microorganisms apparently in the same habitat are, in fact, often exposed to entirely different environmental influences and population pressures. To understand the forces actually affecting the organisms, a microenvironmental concept rather than the gross macroscopic view of interactions must be adopted.” The review by Griffith (1965) of the extensive work carried out in the 40s and 50s on the opposite effect of microorganisms on their physical environment, and in particular on the development of soil architecture, also raises many questions that could be addressed only from a microscopic perspective. Experimental evidence obtained since the mid-sixties has provided steadily strengthening support for this perspective (Hattori, 1973; Cheshire, 1977; Elliott et al., 1980; Tiedje et al., 1984; Stotzky, 1986; Crozat et al., 1987; Darrah et al., 1987; Parkin et al., 1987; Postma and Altemuller, 1990; Postma and van Veen, 1990; Killham et al., 1993; Renault and Stengel, 1993; Strong et al., 1997; Wachinger et al., 2000; Chenu and Stotzky, 2001; Attard et al., 2011; Chapman et al., 2012; Johnson et al., 2013; Vos et al., 2013; Uroz et al., 2015; Xun et al., 2015; Barcenas-Moreno et al., 2016; Keiluweit et al., 2017, 2018).
In the 50s and 60s, very little could be done to come up with better measurements, unfortunately. Alexander (1964, p. 219), again, observed that “because of inherent technical difficulties in biochemical experimentation at the microscopic level, progress in understanding of the microenvironment has been painfully slow.” Even though more and more experiments over the years confirmed the significance of microenvironments, for a long time it was not feasible practically to characterize them in quantitative terms. The advent of transmission or scanning electron microscopes, and later of confocal laser microscopes as well, provided a wealth of qualitative information about microbial habitats in the form of micrographs of increasingly high quality (Foster, 1988; Vandevivere and Baveye, 1992a,b,c,d; DeLeo et al., 1997; Baveye et al., 1998), but the lack of related quantitative data prevented for several decades the development of satisfactory predictive models of soil microbial processes, accounting explicitly for the microheterogeneity of soils.
This situation has changed dramatically in the last decade and is continuing to evolve at a rapid pace. Significant technological advances have provided soil researchers, for example, with routine access to X-ray computed tomography (CT) systems, which provide increasingly reliable information about the geometry of pores and solids in soils at resolutions as small as 0.05 μm. Progress in near-edge X-ray spectromicroscopy (NEXAFS), scanning transmission X-ray microscopy (STXM), X-ray absorption spectroscopy, micro-fluorescence spectroscopy, and Nano-SIMS, applied to soil thin sections, has led to observations of sharp spatial heterogeneity in the chemical make-up of soils over minute distances, and in the accumulation of trace metals. Significant advances related to biological markers now allow specific bacteria to be identified in soils, and their spatial distribution at the micrometer scale to be determined in thin sections. This information can be translated into 3-dimensional distributions using recently developed statistical algorithms. In addition, very efficient modeling tools, like the Lattice-Boltzmann approach, allow the description of transport and physico-chemical processes occurring in soil pores at scales that are directly relevant to microorganisms, whereas individual-based or agent-based models, also developing rapidly, can describe the dynamics of microorganisms inhabiting the pore space (Gras et al., 2010, 2011; Muci et al., 2012; Hellweger et al., 2016; Kim and Or, 2016).
In the last few years, the application of each of these technologies and modeling methods to soils has been the object of a sizeable literature. Progress achieved in the use of each technology has already been expertly reviewed (O’Donnell et al., 2007; Taina et al., 2008; Young et al., 2008; Behrens et al., 2012; Rennert et al., 2012; Helliwell et al., 2013; Tuller et al., 2013; Wildenschild and Sheppard, 2013; Schlüter et al., 2014; Calistru and Jitareanu, 2015; Kuzyakov and Blagodatskaya, 2015; Prosser, 2015; Roose et al., 2016; Xiong et al., 2016; Totsche et al., 2017). For some technologies, since advances are extremely rapid, it would be useful, conceivably, to provide an updated coverage of recent work, and no doubt new reviews will fill the gaps in the near future. Yet, a different type of critical overview might be even more fruitful at this stage, one that keeps sight firmly on what started out as the ultimate goal of the research: A thorough understanding of what one needs to measure at the macroscale in order to adequately describe emergent microbial processes. Instead of surveying the increasingly widespread application of specific technologies to soils, it is worth taking a step back and analyzing how the use of these technologies and their continual improvements help us, or are expected to help us, move steadily on paths leading to the goal we seek. For each path, we can try to assess how far along we are at present and, to the best of our knowledge, to estimate how much distance remains to be covered. Also, since at the scale of bacterial and archaeal cells, it is virtually impossible to dissociate physical, (bio)chemical, and biological aspects of soils, another key point of interest is the extent to which the combined uses of different technologies, meant to access information on these complementary aspects, make us now, or at least promise to make us soon, converge consistently toward meaningful insights. In this reflection on what remains to be done, it makes sense to try to gauge as well how much assistance we could derive from measurement technologies that are barely emerging at the moment but will in all likelihood become routinely available to us in the next few years. It is to scrutiny along these different directions that the present review article is devoted.
First things first. As a famous microbiologist once wrote, “without the proper technological advances the road ahead is blocked. Without a proper vision, there is no road ahead” (Woese, 2004). So, it is vital to start from a clear perception of the goal that is being pursued, and then outline what paths lead to it. As pointed out above, it has been known for half a century at least that the type of macroscopic measurements that are carried out routinely on soils and sediments at the moment do not inform in a satisfactory way about the parameters that appear to be controlling the activity of microorganisms in these systems. Experience has shown clearly that knowledge of, e.g., the total microbial biomass and the total amount of OM present in a given volume of soil or sediment does not allow us to make reliable predictions about the activity of microorganisms or the fate of OM. Somehow, our usual measurements do not capture enough of the huge complexity that soils manifest at the microscopic scale to enable us to predict accurately various properties of soils, like the activity of microorganisms, at the macroscopic scale.
To describe the process by which microscale heterogeneity influences and generates macroscopic behaviors, researchers have used alternatively the terms of “emergence” (Holland, 1990; Addiscott, 2011) or “self-organization” (Smagin, 1989; Hallet, 1990; Phillips, 1995, 2000; Manson, 2001; Young and Crawford, 2004; Barot et al., 2007; Lavelle et al., 2007, 2016; Ebrahimi and Or, 2016; Tecon and Or, 2017a,b). For a number of reasons, explained in detail in Appendix 1 (Supplementary Information), “emergence,” implying a reality that is less than the sum of its parts and is therefore much simpler to describe, is far more appropriate than the term of “self-organization” to describe the type of soil-borne processes on which this review article focuses. In the following, we shall therefore refer consistently to “emergence.”
This point of terminology being resolved, the crux of the matter is that information of an entirely different nature than that currently available is needed to describe soil microbial processes adequately. We clearly need new macroscopic measurements. There are probably different ways to envisage the paths that will lead us eventually to this “Holy Grail.” Figure 1 proposes one of these perspectives, which has served as a general strategy map to a number of us in our research efforts. It starts on the left with information about basic soil features. What we understand at this point of emergent processes in soils indicates that this topic has (at least) three clear, resolutely interdependent facets, associated, respectively, with physical-, (bio)chemical-, and microbiological aspects of soils. For each of them, it is crucial to gather experimental information, either on static properties (dealt with in the boxes “physical characterization,” etc.), or on their dynamics. Alongside this evidence gathering, it is also important to develop theoretical and modeling frameworks that encapsulate experimental information and allow predictions to be made. In each case, experimental data should serve to refine theories and models, which in turn (e.g., through sensitivity analyses) can provide guidance in the procurement of additional data. The outcome of this type of iterative approach, hopefully, is a satisfactory description of each dynamic, which can then be integrated at first pairwise, and eventually all together, into a comprehensive model of soil processes at the microscale.
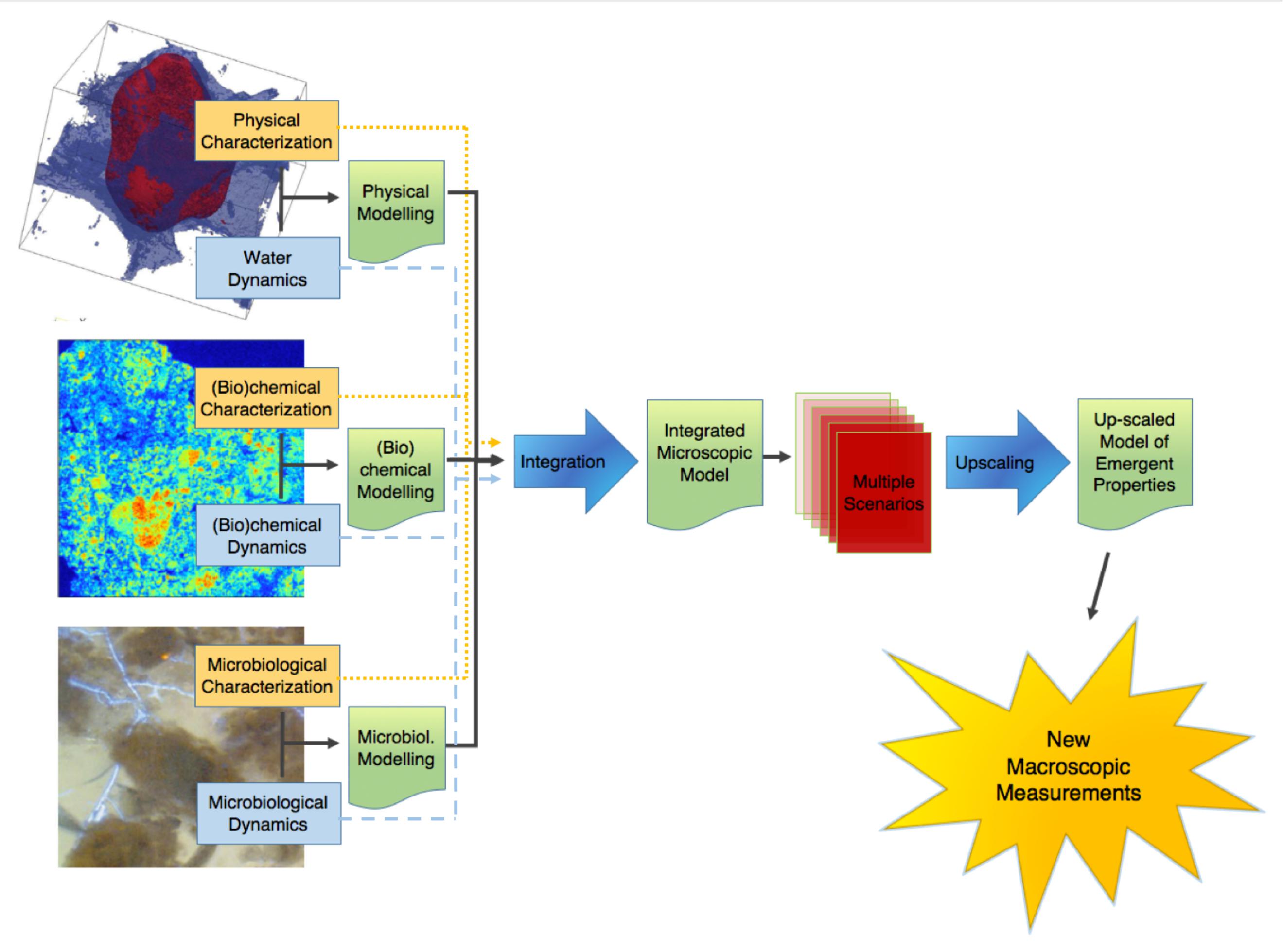
FIGURE 1. Schematic representation of the sequence of steps in the research on the emergent properties of soils, leading from a characterization of the various properties and dynamics at the microscale, onward to an upscaled macroscopic model, and finally to the ultimate goal of identifying macroscopic measurements that can be carried out routinely.
At that point, we are still somewhat far from the goal. Indeed, when this integrated model becomes available, running it on any given soil sample will require a tremendous amount of microscale information, which may take weeks or even months to gather, not to mention that the integrated model itself will likely take quite some time to run, which means that with this integrated microscale description of soils, only very few soil samples will ever be characterized and modeled. What we need instead is to come up with simple macroscopic measurements that can be carried out routinely.
One way to find out what these macroscopic measurements should be is suggested in Figure 1. It consists of expanding the available experimental database by simulating many different scenarios under different conditions of microscale heterogeneity of the soils, and of their properties. From these multiple scenarios, one can try to find out how one can simplify the description, in other words upscale the microscale model to the macroscale, while making sure that, in the process, the macroscopic parameters that appear in the resulting upscaled description are amenable to routine measurement in practice. This prospect of course rests on the assumption, at this stage very much open, that the simplification implied by the notion of “emergence” indeed occurs in soils. As discussed later, there is fortunately some circumstantial evidence that such simplification can be expected.
The research program, depicted in Figure 1, comprises a number of clear paths, which are discussed in the following. To the extent that some of the steps along these paths involve advanced technologies and elaborate methods of analysis or simulation, there is a definite risk of drift, i.e., to focus excessively on tools, perfect them, and progressively forget over time the reason for doing all this work in the first place, as one could argue has unfortunately happened occasionally in the past in other contexts in soil science (Baveye and Laba, 2015). One might argue that switching progressively from actual soils to very coarse sands or collections of clean 500 μm (or bigger) glass beads constitutes an example of such a drift. These systems admittedly pose far fewer technological challenges, which enable researchers to identify and isolate various microscale mechanisms, but, as experience acquired in the past (e.g., in the sixties, when glass beads were used to research several soil processes, like water retention hysteresis) has shown, the relevance of the information acquired in these idealized systems for the functioning of real soils is unclear, at best. To avoid such drift, as much as possible, the descriptions of the various paths of Figure 1 will focus exclusively on progress made to date with actual, living soils, in all their wonderful complexity and messiness.
One last comment that needs to be made before we embark in the description of the program of Figure 1 is that there is no reason to be so wedded to it as not to be open to alternatives that may surface. If tomorrow, an experimentalist comes up with a robust empirical relationship among novel macroscopic measurements, similar to what is envisaged as the ultimate outcome of the program of Figure 1, every researcher interested in the field should probably rejoice, change gear, and adopt an entirely different perspective, for example to try to understand why the solution works. This is reminiscent of the debate about top-down vs. bottom-up approaches in hydrology (Basu et al., 2011; Baveye and Laba, 2015). Regardless of how strongly held one’s philosophical beliefs are, what matters most is to find a satisfactory answer to a number of questions, not necessarily the manner in which the answers are obtained. This being said, no experimentalist has stepped forward yet with a ready answer, and the stepwise plan of Figure 1 appears to be our best bet at this point to ever obtain one in the limited time we have to do so.
Any overview of the quantitative research of the past 10 years on microscale processes in soils needs to start with their physical characterization. Indeed, soil physicists have undeniably led the charge. The pioneering work carried out in the early 1980s with medical and custom-made X-ray and gamma-ray computed tomography systems brought to the attention of the soil physics community the potential of this technology, then still in its infancy (Petrovic et al., 1982; Hainsworth and Aylmore, 1983; Crestana et al., 1985; Pires et al., 2010). The low (millimeter) resolution of scanners available at the time enabled researchers to characterize the geometry of macropores (e.g., earthworm burrows) in soils (Warner et al., 1989; Joschko et al., 1991; Heijs et al., 1995; Capowiez et al., 1998; Rogasik et al., 2003; Luo et al., 2010), but was much too coarse to provide information relevant to microorganisms. In the mid-1990s, various synchrotron facilities around the world began to devote beam time to soils, and researchers immediately took advantage of the significantly higher spatial resolution (down to a few μm) these facilities afforded, as well as the fact that the synchrotron X-ray beams are monochromatic (single-energy) (Anderson and Hopmans, 1994; Spanne et al., 1994; Garnier et al., 1998; Wildenschild et al., 2002; Feeney et al., 2006). However, access to synchrotron beam time was, and still is to a large extent, somewhat scarce and difficult to obtain, so that the extent of adoption of synchrotron-X-ray tomography has remained limited. The commercialization, around 2002, of the first tabletop, non-medical X-ray tomography systems, which were not excessively onerous and could therefore be entirely dedicated to soil science research, marked the beginning of a new era. The X-rays produced by these machines are polychromatic (i.e., are a mixture of X-rays of different energies), which in a number of ways is a disadvantage compared to the monochromatic X-ray produced by synchrotrons, but the resolution of these tabletop scanners has steadily improved since 2002 and several machines now allow resolutions that, in small soil samples of a few cm3, can be as low as 0.3 μm, i.e., commensurate with the resolution afforded by synchrotrons (Voltolini et al., 2017) and with the size of some of the “ultra-small” bacteria and archaea found in soils. The very high resolution of X-ray CT has for a time at least made other types of measuring instruments, like dual-energy gamma-ray scanners, neutron radiography, or nuclear magnetic resonance micro-imaging systems, fall off the radar screen, at least in applications to soils. Nevertheless, as we shall see later, these instruments afford advantages over X-ray CT, and are therefore likely to play a more significant role in the future.
The “3-dimensional” soil images that CT scanners provide are in fact stacks of 2-dimensional, grayscale images associated with virtual slices within the soil sample. Very early on in the use of these images, researchers came to the conclusion that these grayscale images would not be very useful to quantify the geometry of the soil pore space and that it was necessary to derive binary (black and white) images from the original grayscale ones, a process alternatively referred to as “thresholding” or “segmentation.” Significant progress has occurred over the years in how this thresholding is approached. Initially, it was carried out slice by slice, either manually by simple visual inspection (“eye-balling”) or with the assistance of one of a number of available 2D algorithms (e.g., Nunan et al., 2006). The first improvement consisted of thresholding the whole 3-d image at once, using an algorithm to calculate a unique, global threshold value. Then, various researchers showed that in the presence of textural heterogeneities (e.g., stones) within the samples, it was preferable to instead use local thresholds, which can vary from location to location within a sample (Iassonov et al., 2009; Schlüter et al., 2014). Up to that point, all thresholding algorithms required operator input, to adjust one or more parameters. This introduced unavoidable subjectivity in the process, which in principle would make it improbable for different individuals to threshold a given soil sample the same way, or even for a single individual to threshold different soil samples (e.g., associated with different agricultural practices or with successive times) in a consistent manner (Baveye et al., 2010).
The question of objectivity in the generation of X-ray CT images of soils is in fact much broader than just this issue regarding thresholding/segmentation. Indeed, as a number of authors have pointed out (Vaz et al., 2011; Houston et al., 2013b), the process of obtaining CT images of soils requires many decisions to be made by operators, concerning in particular the value of scanning parameters (e.g., energy level, choice of filter, scanning resolution), the selection of one among a number of alternative image reconstruction and artifact correction algorithms, the format (8- or 16 bit) used to store the images, and the use of a method to increase image sharpness or reduce the noise that is unavoidably present in the images after reconstruction. As with thresholding 10 years ago, different groups, and sometimes even different individuals within a group, adopt alternative perspectives with respect to the various decisions that need to be made by operators, which can lead to sometimes significant differences in some of the metrics that are associated eventually with CT images (see, e.g., Houston et al., 2013b). Nevertheless, at this point, there appears to be no effort underway to develop a set of materials that could be used as “scanning standards,” as suggested by Baveye et al. (2010), or simply to standardize analyzes. One way out of the difficulty would be to document exhaustively the parameter values used at each and every step of the image acquisition process, as well as, through detailed sensitivity analyses, the extent to which conclusions that are reached on the basis of CT images are affected by these parameter values.
Nevertheless, recognition a few years ago that the subjectivity in thresholding operations and in the manipulation of CT images could be substantial, prompted the development of a number of automated thresholding algorithms requiring no operator input (Schlüter et al., 2010; Hapca et al., 2013; Houston et al., 2013a), regardless of the level of “supervision” (learning from training data) adopted. These objective algorithms have been used in a number of investigations (e.g., Beckers et al., 2014a,b; Houston et al., 2017), and new algorithms are appearing that do not require any parameter tuning (e.g., West et al., 2018), but so far they have not stopped the development of operator-dependent approaches (Kulkarni et al., 2012; Hashemi et al., 2014; Ojeda-Magana et al., 2014; Martin-Sotoca et al., 2017). Therefore, further progress is needed in this area, especially in order to segment images containing multiple distinct populations of voxels.
Another approach that has recently been explored to obtain basically the same physical information as with X-ray CT consists of using broad- or focused ion beam scanning electron microscopy (BIB- or FIB-SEM). The ion beam can directly modify or “mill” a specimen surface, and this milling can be controlled with nanometer precision. By carefully controlling the energy and intensity of the ion beam, it is possible to perform very precise nano-machining to remove very thin layers of material, for example in a block of soil impregnated with resin. BIB milling produces cross-sections of a few mm2 to cm2, whereas FIB deals with surfaces that at most are a few hundred μm2. Once a new surface has been exposed, it can be imaged via SEM, at resolutions typically between 10 and 500 nm (Cantoni and Holzer, 2014). The sequence of images obtained in successive layers can be assembled into a 3D image, similar to those resulting from X-ray CT tomography, and subsequently segmented (Salzer et al., 2015; Liu et al., 2017). In the last few years, this approach has been used extensively to investigate the morphological characteristics of dolomite rocks, shales, and clays using BIB alone (Houben et al., 2013), a combination of BIB- and FIB-SEM (Hemes et al., 2015), or the joint use of micro-CT and FIB-SEM (Devarapalli et al., 2017). In soils, FIB-SEM presents a tremendous potential, but its use appears to have been limited so far to observations of microbially induced calcite precipitation in sandy soils (Li et al., 2017) and to obtain high-resolution images of the colonization of soil–root interfaces (Vidal et al., 2018).
Early in the use of CT scanners to characterize the physical properties of soils, it became apparent that this technology afforded a convenient response to the age-old question of how to best quantify soil “structure,” this term being understood either as “the arrangement or organization of the particles in the soil” (Hillel, 2004), or, following Dexter (1988), as “the spatial heterogeneity of the different components or properties of soils.”
For many decades, the vast majority of the research on the topic has viewed soil structure as intimately linked with the fact that it is possible to fragment soils into distinct aggregates upon the application of mechanical stress (Rabot et al., 2018). Undoubtedly this perspective has its roots in the soil surveyors’ traditional poking of exposed soil profiles with knives, leading to the detachment of chunks of soils, called “aggregates,” whose size and shape is used to diagnose the types of pedogenetic processes that might have taken place at that location, to classify soils, and to evaluate their agronomic potential. Since the 1940s, an extensive body of literature has been devoted to the assessment of the stability of soil aggregates under a variety of operational conditions, for example under dry or wet sieving. As Young et al. (2001) point out, “the ease and seeming reproducibility of the many standard stability tests are the main drivers behind the prevalence of this type of research.”
A common criticism of the concept of aggregate in soils is that it is little more than an artifact. The hierarchical organization of aggregates, identified and described in detail by Tisdall and Oades (1982), suggests that the distribution of sizes of aggregates one obtains might depend on the amount of energy that is applied to take soils apart. This operational issue, discussed by Amézketa (1999), is particularly well illustrated by the experimental results of Díaz-Zorita et al. (2002), who show that the size of fragments obtained by sieving soils is inversely related to the mechanical stress applied. Hallett et al. (2013) also point out that breakdown of soils by dynamic or static mechanical loading yields different fragmentations of soil aggregates. This dependence of the aggregate size distribution on the operational conditions under which it is measured raises the question of whether aggregates exist in soils in their natural state (Young et al., 2001), calling into question the extensive literature that tries to analyze the influence of aggregate size on various processes, e.g., in terms of the sequestration of OM, the distribution of bacteria, a wide range of geochemical processes, or the release of greenhouse gasses (Ranjard and Richaume, 2001; Jasinska et al., 2006; Nunan et al., 2006; Razafimbelo et al., 2008; Goebel et al., 2009; Pallud et al., 2010; Chivenge et al., 2011; Masue-Slowey et al., 2011, 2013; Blaud et al., 2014; Rabbi et al., 2014, 2016; Ebrahimi and Or, 2015; Jiang et al., 2015; San José Martínez et al., 2015; Sheehy et al., 2015; Hausladen and Fendorf, 2017; Rillig et al., 2017; Zhao et al., 2017; Bocking and Blyth, 2018; Li et al., 2018), and explaining perhaps why some authors have failed to observe anticipated correlations between OM content and aggregation (Razafimbelo et al., 2013). Nevertheless, one might argue that this dependence problem can be alleviated somewhat by standardizing methods, and that, in any event, it does not particularly affect attempts to understand at a very local scale in soils the interactions between pore geometry, chemical composition, and microbial activity. As long as aggregates are viewed as chunks of soil that are convenient to manipulate because they do not fall apart too easily, e.g., when they are rotated on the stage of a CT scanner, and to the extent that no particular significance is associated with their external surfaces, which might just have been failure planes in some larger aggregate, no harm is done in using aggregates to gain insight into microscale processes, as various authors have done successfully (Remusat et al., 2012; Ananyeva et al., 2013; Kravchenko et al., 2013, 2015; Voltolini et al., 2017; Yu et al., 2017).
One could also consider that there is no problem either with repacking aggregates extracted from a soil, and trying to find out experimentally or through simulation how this now entirely artificial system behaves (e.g., Daly and Roose, 2014; Ebrahimi and Or, 2016). We are often forced by journals to use repacked soil columns in order to have actual replicates, and be able to calculate statistics, which some reviewers view as sacred and indispensable. However, it is entirely unclear at this point to what extent the conclusions that one reaches from this kind of exercise relate to the behavior of real soils, including the very soil from which the aggregates that are used originated. The reason for this has to do fundamentally with the absence of any theoretical framework or set of procedures to, as it were, put the pieces of the puzzle back together, once a soil has been disaggregated and its aggregates have been characterized, e.g., relative to their size distribution and individual geometries. In the process of disaggregating a soil sample, as long as no information is obtained about the geometry and topology of the interstices that may have existed originally between what eventually becomes distinct aggregates, there is no way practically to “reconstruct” the original soil, even for computational purpose, and in particular to guarantee that the pores between aggregates in the repacked system be similar in shape to those that existed originally1. One could draw parallels here with architecture (Letey, 1991; Baveye, 2006) or even with card games: Indeed, one cannot say anything about the size and shape of a house of cards after it has been torn down, simply by looking at the pile of cards that is left.
Aware of these obstacles already many years ago, a number of authors argued for a different way to approach the structure of soils. Dexter (1988), in a thorough review of the then available methodology in this field, recommends that preference be given to methods involving direct observation of structural features by scanning electron microscopy and by optical scanning of resin-impregnated sections and fracture surfaces. A few years later, Letey (1991) vents his frustration in the face of many failed attempts to link soil structure, defined in terms of aggregates, to functionality within the soil system. He suggests that instead of focusing on the solid components of soil structure, as had been the tradition for decades, one should emphasize instead the arrangement of voids, and the properties that these voids confer to soils, just as to describe a building, it is not primarily the shape of the bricks or stones that matters, or the thickness of the walls, but the size of the rooms and openings (windows, door frames). Reiterating these same messages, Young et al. (2001) argue that “an investigation of discrete aggregates or distributions of aggregates does not offer any spatial information. Functional traits of soil structure, at all scales, rely on the connectivity, tortuosity, and heterogeneity of pore space in 3D.” The same message is echoed in the recent thorough review of the literature by Rabot et al. (2018), who conclude that “although appealing, the aggregate perspective does not seem to be the most appropriate to link soil structure with soil functions and processes.” Because of the historically close connection between “soil structure” and aggregates, Young et al. (2001) propose to drop the expression of “soil structure” in favor of that, less history-laden, of “soil architecture.” This terminology has been routinely adopted since (e.g., Baveye, 2006; Lin et al., 2010; de Jonge et al., 2012; Lin, 2012; Bouckaert et al., 2013a,b; Cazelles et al., 2013; Helliwell et al., 2013; Kravchenko and Guber, 2017; San José Martínez et al., 2017) and will be used consistently in the following.
In principle, it is feasible to analyze this architecture by taking 2D images of sequences of thin sections in resin-impregnated blocks of undisturbed soil, and then using dedicated software to reconstruct from these images a full 3D picture of the geometry of soil pores. This tedious, time-consuming approach has been adopted with success by Cousin et al. (1996, 1999), Vogel (1997), and Vogel and Roth (2001). However, access to X-ray beams at various synchrotron facilities, and especially the availability of table-top X-ray CT scanners, have allowed the work in this area to experience a quantum leap around the turn of the century. The new technology has made it possible to obtain 3D images of the pore space in intact soil cores much more rapidly, and at resolutions that have gradually improved over time (Mooney, 2002; Rozenbaum et al., 2012; Bouckaert et al., 2013a; Calistru and Jitareanu, 2015; Rabot et al., 2015).
The gradual conceptual shift from the aggregate-based “structure” to the “architecture” of soils has been accompanied by a refocus of the discourse on the voids within this architecture, following in that respect the suggestion of Letey (1991). Another conceptual shift as well is occurring in that respect. Conditioned to think in terms of a traditional analogy between the pore space of soils and a bundle or network of capillaries, soil physicists used to be concerned about the size of “pores” in soils. It is clear from CT images that there are no identifiable pores in soils, and that the delineation of individual pores is necessarily somewhat subjective. Some authors have tried to make the concept of pore size distribution less arbitrary by using automatic algorithms to determine locally the radii of maximum balls that are fully inside the pore space. Partly because of the historical weight of the capillary analogy and partly with the help of these “inscribed balls” algorithms, pore size distributions are still being computed (e.g., Kuka et al., 2007; Papadopoulos et al., 2009; Ostadi et al., 2010; Bouckaert et al., 2013a; Peng et al., 2014; Houston et al., 2017; Meira Cassaro et al., 2017). Yet, clearly, researchers have increasingly turned in recent years to other approaches to describe quantitatively the make-up of soils. Indeed, a whole panoply of mathematical tools is now available, and is steadily expanded, to characterize a number of aspects of the pore space. These tools include various algorithms to calculate the tortuosity and connectivity of the pore space on the basis of grayscale or binary 3D CT images (Gommes et al., 2009; Houston et al., 2017; Meira Cassaro et al., 2017). Another approach to describe the pore space quantitatively is provided by the fundamental set of Minkowski functional measures (Lehmann et al., 2006; Vogel et al., 2010; Falconer et al., 2012). These functional measures comprise the volume, surface area, integral mean curvature, and the Euler-Poincaré characteristic (or topological measure Chi). Karsanina et al. (2015) propose another set of descriptors, including two-point probability functions, linear functions, and two-point cluster functions, and they used the first two in simulated annealing optimization procedures to reconstruct soil architecture artificially, based on original images of soil thin sections. For a number of years, fractal geometry was thought to be an ideal tool to characterize the inner space of soils, since according to the way the theory was interpreted, a single parameter, the fractal dimension, could inform about the make-up of soils over a range of scales. This vision of fractal geometry has since been confronted with the reality that a single dimension, whose value turns out to be itself scale- and resolution-dependent, does not suffice. As explained by Mandelbrot from the start, at least one other parameter, either the lacunarity (e.g., Pendleton et al., 2005; San José Martínez et al., 2017) or the succolarity (de Melo and Conci, 2013) is required to obtain an accurate description. Since the lacunarity (Pendleton et al., 2005) and likely also the succolarity are affected by the resolution of images, it is not clear at this point whether fractal geometry still offers much interest. Another approach that might (but has not yet) provide a solution to the resolution-dependence derives from the application of the theory of multifractal measures to CT images of soils (Lafond et al., 2012; Wang et al., 2016; Torre et al., 2018). From a different perspective, various authors have also tried to extract networks from 3D images of soils, to which graph theory principles can be applied to characterize the connectivity and topology of the pore space (Vogel and Roth, 2001; Perez-Reche et al., 2012).
Since the ultimate objective of the research reviewed here is to eventually be able to predict the activity of microorganisms in soils, for whom the presence of water and readily biodegradable OM is crucial, it is important to be able to detect in what portion of the pore space they are located. In this respect, researchers have been confronted with the difficulty that, typically, if one places a wet soil sample in a table-top X-ray CT scanner, the outcome is a 3D grayscale image characterized by a histogram with a single, broad peak that is not suitable at all to tease apart the water from the solids without resorting to arbitrary assumptions (Tracy et al., 2015), nor to identify in the solid phase the portion that corresponds to OM. So far, to avoid this obstacle, researchers have either shifted their attention toward artificial media, or they have worked with actual soils but under special conditions that allow the identification of water and OM.
In terms of artificial porous media, researchers have used glass beads (Culligan et al., 2006; Schaap et al., 2007) and coarse sands (Brusseau et al., 2007) to quantify the 3-dimensional distribution of water in the pore space. If one scans these systems under partially saturated conditions, as these researchers did, evidence suggests that it is not very difficult to locate air-water interfaces. Another way to proceed, made possible by the low reactivity of glass beads or sands compared to soils, is to increase the contrast between the attenuation of X-rays in the solid phase and the liquid phase by using a contrast agent that increases X-ray attenuation in the liquid. Although the results obtained with glass beads and sands are definitely interesting and probably applicable to coarse aquifer materials, it is not clear at this stage how they help us identify water and OM in actual soils, which as a rule tend to be tremendously more heterogeneous, and have much smaller pores. There is no real answer at this point to the question of how one can transition from glass beads to actual soils. This is a perennial problem, also faced by researchers who for a time carried out extensive work in the 1960s on the hysteresis of water retention in glass beads systems (Topp and Miller, 1966; Topp, 1971).
Another approach that can conceivably work in some soils, consists of scanning a soil sample when it is dry, and then re-scan it when it has been brought to the desired moisture content (e.g., Tracy et al., 2015). Comparison between the non-air phases in the “before” and “after” images yields the distribution of water in the system. In principle, this approach could work very well if the soil does not swell at all when its moisture content is increased. This apparently was the case in the experiments carried out by Tracy et al. (2015), who report that “no significant evidence of shrinkage was observed.” Yet the problem is that most soils in the world do shrink/swell to some extent (Garnier et al., 1997), including soils like those described by Radulovich et al. (1992) whose kaolinitic mineralogy one does not traditionally associate with this phenomenon. The question remains at this point of what is significant enough evidence of shrinkage or swelling in a soil sample to prevent this “subtraction” method to be used to visualize the distribution of water.
Yet another strategy is to carry out CT measurements on real soils under conditions where water and OM are not intimately mixed with the solid constituents at very fine scales. This approach has been adopted by a number of researchers in the last few years who worked on plant residues within soils (De Gryze et al., 2006; Negassa et al., 2015; Kravchenko et al., 2017) or attempted to directly visualize soil moisture (Carminati et al., 2008; Tippkötter et al., 2009; Pot et al., 2015). Working with a real clay-loam soil material near water saturation, Carminati et al. (2008) focused on the water that occupies part of the volume in the larger pores. They were able under these conditions to clearly observe pendular rings of water in images at a resolution close to 6 μm. Tippkötter et al. (2009) adopted a similar focus, in undisturbed soil samples, and were able with a table-top X-ray CT scanner to visualize the presence of water films coating the inner surfaces of meso- and macropores. Similarly, Pot et al. (2015), working with synchrotron X-rays, were able to generate CT images of repacked aggregates in whose histograms there was a good separation of voxels associated with the air, liquid, and solid phases (Figure 2).
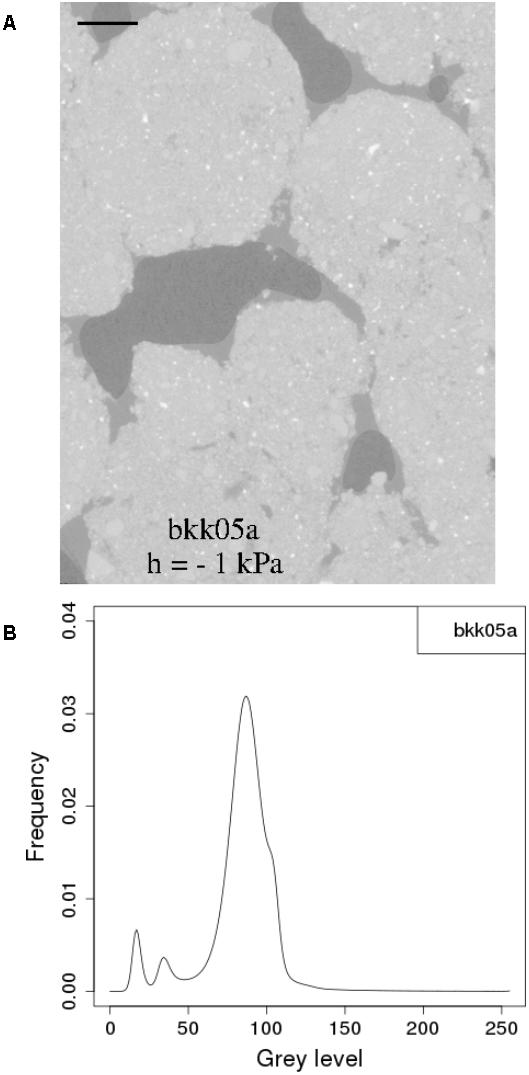
FIGURE 2. (A) two dimensional section of synchrotron X-ray computed tomography image of a soil cube equilibrated at –1 kPa and (B) histogram of the corresponding 3D SR-μCT image. In the tomographic sections, black is the air phase, dark gray is the water phase and light gray to white is the matrix phase. The scale bars represent 500 μm. (Reprinted with permission from Pot et al., 2015).
A last approach that could work in principle to see the moisture in soils consists of adding various contrast agents to the water, to modify its X-ray attenuation (Van Loo et al., 2014). However, in practice, contrast agents need to diffuse sufficiently for the method to work, which again, in many cases, in undisturbed soil samples, might be operationally workable only to image the largest pores near saturation.
It might thus be tempting to look elsewhere for a possible answer. Indeed, over the last decade, the resolution associated with 3D nuclear magnetic resonance (NMR) micro-imaging has become steadily better. Lee and Lee (2017), for example, managed to obtain images of 1.2 mm by 1.2 mm columns of glass beads and crushed silica gels particles, respectively, with a spatial resolution of 46.875 μm, which is still coarser than the resolution of CT scanners for this column width, but is not as far from it as it used to be. As encouraging as these results are, however, NMR micro-imaging as currently implemented still suffers from a major obstacle when it comes to real soils, and therefore is not a real solution in that context. Because of the very powerful magnets that are used to generate the signal, only soils that do not contain paramagnetic elements can be imaged. Since many if not most soils contain some iron, at least, this limits tremendously the conditions under which NMR is a viable alternative to X-ray CT to generate 3D images. An alternative to NMR would be to use neutron computed tomography (Tumlinson et al., 2008) to observe the distribution of water in soils, but at this stage the resolution of images that can be generated is still relatively low, comparable to that obtainable with medical or table-top X-ray CT equipment 15 or 20 years ago (Perfect et al., 2014).
The best option to “see” water at this point, even though it has not been implemented very much of late, appears to be the use of dual energy X-rays in CT scanners. With gamma rays of two different energies, typically produced by 241Am and 137Cs sources, it has been possible for a while to simultaneous assess the moisture content and bulk density of soils (Soane, 1967; Hopmans and Dane, 1986; Biassusi et al., 1999), but the measurements are extremely slow, and their spatial resolution is low. Garnier et al. (1998) applied dual-energy synchrotron X-rays for the first time to soils, to assess rapid vertical soil density and water content changes in swelling soils during infiltration. Shortly thereafter, Rogasik et al. (1999) used a medical scanner that allowed them to scan silt loam subsoil samples at two energy levels (80 and 120 kV) to evaluate the distributions of water, air, and solids, as well as the voxel dry bulk density. The spatial resolution during scanning was 0.25 mm in the horizontal and 1 mm in the vertical direction, which was (and still is) standard for scanners routinely used in hospital settings. Since this work almost 20 years ago, there has been to our knowledge no application of dual-energy X-ray tomography to soils. Several table-top X-ray scanners currently commercialized offer the possibility to carry out dual-energy scanning sequentially on soil samples. The fact that nobody so far has reported on the use of this feature with table-top scanners suggests that polychromatic X-rays are not suitable for dual-energy scanning to work in the case of soil samples. Further research is needed to determine if with monochromatic X-ray beams, at synchrotron facilities, dual-energy scanning produces promising results.
Part of the reason for the limited use of dual-energy scanning - and it would be true as well for attempts to scan soils rich in OM with dual-energy gamma-rays - is that until not too long ago, it would have been difficult to tease apart water from soil OM in CT images (Taina et al., 2008). The problem is not OM per se. Kettridge and Binley (2011)demonstrate that X-ray CT can image beautifully the structure of peat samples of various compositions. The difficulty has to do with the fact that at the high X-ray energies required to penetrate through soil materials, there is very little difference in X-ray attenuation between water and water-filled OM, whose peaks in grayscale image histograms are often not clearly distinguishable from a broad peak associated with mineral constituents. This problem was resolved, at least in part, in 2014, when Van Loo et al. (2014) tested 52 different chemical compounds. They perfused aqueous solutions saturated with the compounds through undisturbed soil samples under partial vacuum and found that 4 of these chemicals [phosphomolybdic acid (PMA), silver nitrate, lead nitrate and lead acetate] successfully enhance the X-ray attenuation contrast of OM relative to soil minerals and allow particulate organic matter (POM) to be easily detected. Peth et al. (2014) tried to take advantage of the fact that osmium has a marked absorption K-edge2 at a photon energy ∼74 keV. They exposed air-dry soil aggregates to a 25 w/w OsO4 solution for 48 h at room temperature in a closed vial under a fume hood (because of the very high toxicity of OsO4), and scanned these aggregates at a synchrotron facility below and above the absorption K-edge, respectively. Preliminary results, obtained by Peth et al. (2014) and Rawlins et al. (2016), suggest that this technique makes it possible to visualize the distribution of OM in soils, and to distinguish between POM and OM that is distributed more diffusely throughout the soil architecture (Figure 3). One promising approach to identify POM in CT images consists of building on both the attenuation, thus gray scale, values of the organic materials and on the spatial distribution patterns of POM grayscale values, which uniquely separate it from the rest of the soil solids (Kravchenko et al., 2014a). Indeed, even from a “naked eye” examination, POM often stands out on CT images due to much greater uniformity of its grayscale values. Kravchenko et al. (2014a) successfully used geostatistical parameters of POM fragments as indicators of the presence of POM in intact soil samples. This approach has advantages over POM identification via Os staining, since, unlike Os staining, CT scanning has minimal effect on soil microorganisms (Bouckaert et al., 2013b; Kravchenko et al., 2014b; Schmidt et al., 2015). Thus, the samples can be used for exploring the decomposition of the identified POM fragments in a sequence of initial CT scanning, incubation, and post-incubation CT scanning activities, as done by Kravchenko et al. (2015). However, as of now the process of POM identification using this approach is time consuming and requires a substantial user input.

FIGURE 3. Illustrative two-dimensional spatial distributions of osmium-stained OM mapped onto the reconstructed image cross-sections of aggregates. In (A), red colors are typically associated with particulate OM. Green colors reflect lower concentrations (Modified from Peth et al., 2014. Reprinted with permission). In (B), the red patches correspond to OM (Modified from Rawlins et al., 2016. Reprinted with permission).
Another issue that, at the moment, is still awaiting a definite solution, is related to the soil pores having dimensions smaller than the resolution of CT images. From the mid-1980s to the early 2000s, the resolution of X-ray CT scanners accessible to soil scientists improved by about 3 orders of magnitude, from a few hundred microns to a fraction of a micron in the best of cases. However, no matter how small this resolution is, a portion of the pore space unavoidably remains invisible to scanners. The practical significance of sub-resolution pores depends strongly on the resolution of CT images, of course, but also, critically, on the type of soil considered. For a coarse sandy soil, it is possible that at a 20 or 30 μm resolution, only a minute portion of the total porosity would not be visible in thresholded CT images. Contrastedly, in other types of soil, the portion of sub-resolution pores can be substantial. In microaggregated tropical soils like those investigated by Sollins and Radulovich (1988), pores with a diameter less than 5 μm make up approximately 70% of the pore space, and are key to understanding the unique physical and chemical properties of these systems (e.g., Radulovich et al., 1992). In the silty soils investigated by Piccoli et al. (2017), 82% of the pores a dimensions smaller than the resolution of 30 μm at which the CT scans were made. In clayey or clay-loam soils, like those whose pore size distribution was determined via mercury intrusion porosimetry by Churchman and Payne (1983), a resolution of 10 μm would be too large to identify any pore at all, and it would be of paramount importance to be able to somehow characterize the sub-resolution porosity in some fashion.
In some very special cases, grayscale CT images, before any thresholding or segmentation is carried out, may contain some information about sub-resolution features. In porous media like sandstone samples or columns filled with glass beads, which consist solely of a homogeneous mineral phase, grayscale values intermediate between those of the pores and solids in CT images result from partial-volume effect if the solid particles are not porous. If, on the contrary, the solid particles are themselves porous, intermediate grayscale values are associated both with partial-volume effects and with sub-resolution pores, and blind application of a ternary segmentation approach, like that used by Scheibe et al. (2015), may yield meaningless results if interpreted solely as sub-resolution porosity. However, in these same simple systems, as long as partial-volume effects are negligible, grayscale values contain unambiguous information about sub-resolution pores, information that is lost entirely when the images are thresholded (Gommes et al., 2009; Baveye et al., 2010). To refine the analysis, insight obtained from images of these systems at different resolutions can be combined, as was done recently by various authors (Sok et al., 2010; Pini and Madonna, 2016; Shah et al., 2016) and applied to soils by others (Vogel et al., 2010; Dal Ferro et al., 2014), to try to get a better handle on predicting the properties of sub-resolution voids. Unfortunately, in real soils, in most cases of practical interest, both these single- and multiple-resolution approaches run into as yet insurmountable hurdles, related to the intimate, fine-scale mixing of minerals, OM, and water. The contribution of these different phases and constituents to the grayscale value of voxels in reconstructed CT images cannot at this stage be differentiated, and as a result, it is not possible in general to correlate this grayscale level with the porosity of the volume of soil to which it corresponds. It might be necessary to wait until tunable X-ray and gamma-ray scanners become routinely available, to resolve this issue.
In order to get a dynamical picture of physical processes in soils, one needs to transition from 3D to 4D, the fourth dimension of course being time. In many disciplines outside soil science, this transition has captured the attention of researchers over the last few years, and very interesting results have been obtained, in particular for very coarse-textured porous media (Berg et al., 2013; Dobson et al., 2016). Yet, as far as soils are concerned, forays along these lines have been timid. At the mesoscale, very interesting work, starting already 25 years ago, describing how earthworm burrow systems evolve over time (Joschko et al., 1991; Capowiez et al., 1998), how the geometry of macropores in paddy soils evolves during soil shrinkage (Bottinelli et al., 2016), or how loamy soils are compacted during centrifugation (Schlüter et al., 2016). Also, various researchers have used MRI systems to monitor the infiltration of water in clay and coarse sandy loam columns (Amin et al., 1994, 1996; Preston et al., 2001; Votrubová et al., 2003), γ-ray CT equipment to quantify the swelling of vertisols over time (Biassusi et al., 1999), or neutron CT systems to investigate the dynamics of water flows in soil, especially in the rhizosphere (Badorreck et al., 2010; Perfect et al., 2014; Tötzke et al., 2017). However, virtually all of this work has been carried out at relatively low resolutions, at best of 15 μm but more often than not of several tens or even hundreds of microns.
At the micron scale sensu stricto, very little 4D work has been carried out so far. None of this research includes water movement, which is not very surprising, given the difficulties mentioned earlier concerning the detection of water at a sufficiently high resolution to be relevant to the microscale. Even under the various conditions where this detection is possible, water movement tends to be too fast to be monitored by X-ray CT, even at the fastest scanning times (of the order to 10–15 min, typically) available with table-top scanners. Ultrafast scanning techniques have been used recently with columns filled with gravel (Dobson et al., 2016), but similar research has yet to be conducted with soils. Because of these constraints, dynamic microscale measurements have been limited to situations that involve ice formation, or slow changes in the architecture of soils. Using a table-top X-ray CT scanner, Torrance et al. (2008) investigated the changes in structure and the redistribution of water to form ice lenses in saturated samples of an Aurora silt loam frost-susceptible soil that were thoroughly mixed to produce an initially homogeneous material, and of a Honeywood silt loam that was deliberately contaminated with motor oil. The soils were subjected to relatively rapid, downward freezing, with access to water at their base. The results indicate that CT can produce excellent images of the ice lens distribution within a frozen silt loam soil, the consolidation of soil between the ice lenses, and the effects of hydrocarbon contamination on ice formation. Also using X-ray CT in freezing soils, Starkloff et al. (2017) assessed the impact of a succession of freezing-thawing cycles on the pore network of a silty clay loam and a loamy sand topsoil. Also recently, Schlüter and Vogel (2016) quantified soil architecture turnover by labeling soil constituents in place with small garnet particles and tracking their fate in successive CT images. The particles adhere to pore boundaries at the beginning of the experiment but gradually change their position relative to the nearest pore as structure formation progresses and pores are destructed or newly formed.
Over the last 2 decades, a significant body of literature has been devoted to the mathematical modeling of water retention and transport within the complex geometry of soil pores, revealed with increasing resolution by X-ray CT scanners. The bulk of this literature has dealt with the development and application of the Lattice-Boltzmann method (Martys and Chen, 1996; Genty and Pot, 2013, 2014; Liu et al., 2016; Cruz et al., 2017; Zhou et al., 2018), but in recent years, other methods have also been adopted, based on finite element or finite difference schemes, or on geometric primitives.
Most of the work in this area has involved a number of variants of the Lattice-Boltzmann method, in which a fluid is viewed as a collection of fictitious particles that, alternatively, propagate from node to node on a regularly spaced grid (lattice mesh), then collide with the particles that end up on the same nodes. In the modeling of soils, the nodes correspond to the centers of voxels in 3D CT images. The method originates from a molecular description of a fluid and can directly incorporate physical terms stemming from a knowledge of the interaction between molecules. Hence, in principle, it keeps the cycle between the elaboration of a theory and the formulation of a corresponding numerical model short, which undoubtedly explains the enthusiasm it incited as soon as 3D CT images of soils became available. The key mathematical ingredient of the method is the probability of finding a particle at position in one of the microscopic directions envisaged within the lattice, at time t, where the subscript q is an index associated with a set of microscopic directions that are selected arbitrarily. Several discretizations of space can be used and are traditionally classified via the DnQm scheme, where “Dn” stands for “n dimensions” and “Qm” denotes “m speeds.” A common choice is D3Q19, in 3 dimensions and with 18 nearest neighbors considered around each node, described by the unit microscopic velocity vectors, . In this case, the subscript q takes on 19 different values (including rest particles).
Classical Lattice-Boltzmann models applied to soils require CT images to be thresholded and assume that voxels associated with pores in binary 3-D images are totally permeable to water molecules, whereas those associated with solids are completely impermeable. Recognition of the significance of the sub-resolution pore space has prompted a sizeable number of researchers in the last couple of years to investigate ways to take this pore space into account explicitly in Lattice-Boltzmann models of water movement in soils, following Gao and Sharma (1994) and Freed (1998). The resulting “Gray” or “Partial-Bounce-Back” (PBB) Lattice-Boltzmann models consider that each voxel in the original, grayscale CT images has a given probability of penetration by water or solutes, and therefore a complementary probability that water or solute particles that penetrate the voxel eventually bounce back to their previous positions (e.g., Sukop and Thorne, 2006; Chen and Zhu, 2008; Han et al., 2008; Walsh et al., 2009; Jones and Feng, 2011; El Ganaoui et al., 2012; Gottardi et al., 2013; Walsh and Saar, 2013; Zalzale et al., 2013; Chen et al., 2014; Li et al., 2014; Yoshida and Hayashi, 2014; Ginzburg et al., 2015; Xie et al., 2015; Yehya et al., 2015; Apourvari and Arns, 2016; Bultreys et al., 2016; McDonald and Turner, 2016; Pereira, 2016; Zhang et al., 2016). In all this work, considerable advances have been made recently and a number of technical issues have been clarified (Ginzburg, 2016), yet a major experimental hurdle related to the evaluation of the penetrability of sub-resolution pores, which at this point remains an arbitrary parameter in the models. As discussed in detail by Baveye et al. (2017), this penetrability cannot be deduced simply from grayscale values in CT images, and there is no practical alternative yet available.
The Lattice-Boltzmann method has indisputably become the de-facto standard in pore-scale studies of water retention and transport in soils. One of the drawbacks of the method, however, is the very long (sometimes weeks-long) computational time it typically requires on personal computers. Open Lattice-Boltzmann environments like Palabos3 or OpenLB4 offer options to run the code on massively parallel computers and arrays of graphics processing units (GPUs), or to decompose the flow domain into manageable subportions, and researchers are increasingly resorting to these speeding techniques in applications of the Lattice-Boltzmann method to soils. Nevertheless, the relative slowness of the original method, unless one has access to large computer clusters, has encouraged various authors to explore other avenues to model soils at the microscale.
One of these avenues encompasses a technique called “smoothed particle hydrodynamics” (SPH) (e.g., Tartakovsky et al., 2007), which works by dividing a fluid into a set of discrete elements, referred to as particles. To these particles is associated a spatial distance (known as the “smoothing length”), over which their properties are “smoothed” by a kernel function. This means that the physical quantity of any particle can be obtained by summing the relevant properties of all the particles that lie within the range of the kernel.
Finite element or finite difference schemes are also among the alternative techniques that have been selected to solve Stokes’ equation within the pore space of soils (e.g., Liu et al., 2016). In a recent article, for example, Gerke et al. (2018) introduce the free software Finite-Difference Method Stokes Solver (FDMSS) that solves Stokes’ equation using a finite-difference method (FDM) directly on voxelized 3D pore geometries (i.e., without meshing). Based on explicit convergence studies, validation on sphere packings with analytically known permeabilities, and comparison against lattice-Boltzmann and other published FDM studies, these authors conclude that FDMSS provides a computationally efficient and accurate basis for single-phase pore-scale flow simulations. By implementing an efficient parallelization and code optimization scheme, permeability inferences can now be made from 3D images of up to 109 voxels using modern desktop computers. Tracy et al. (2015) use another numerical technique, based on the SIMPLE (Semi-Implicit Method for Pressure-Linked Equations) algorithm (Patankar, 1980) to solve Stokes’ equation in the pore space of loamy sand and clayey loam soil samples. These authors did so with OpenFOAM, an open source Computational Fluid Dynamics toolbox.
A different path, beside the Lattice-Boltzmann method and the various numerical schemes just alluded to, consists of using a morphological model. Such a model involves the approximation of the soil pore space by a network of so-called volume primitives, i.e., simple geometric shapes that can be transformed at will and combined to represent more complex geometries (Monga et al., 2007; Ngom et al., 2012). One way to do so consists of using a geometrical algorithm based on Delaunay triangulation to determine the maximal balls of the pore space segmented from the 3D CT images. Maximal balls are defined as the balls included in the pore space but not included in any other ball included in the pore space. Then, a minimal set of maximal balls is extracted in order to obtain a compact representation of the pore space (Monga et al., 2009). The key advantage of the method is that it requires far fewer balls than voxels to cover the pore space, and one might hope in principle that this drastic simplification will carry over to the various processes (e.g., water retention, transport) that one wants to simulate. There is no guarantee in this respect, however, especially when models encompass not just physical processes but also (bio)chemical and microbiological ones. If the geometric primitives become too large, it may be necessary to divvy them up in smaller subcomponents in order to account adequately for the spatial heterogeneity exhibited by chemical and microbial processes in soils. The added computational time that would result from this division might very well negate the speeding up that theoretically results from the scheme.
Since there are different ways to simulate the retention and transport of water in soil pores, one might ask which of these methods performs best. The intercomparison of models (e.g., Yang et al., 2016) provides some general idea of the agreement, or lack thereof, among the models, but clearly, benchmarking model predictions against actual experimental data is by far the most desirable approach. At this juncture, since dynamic data about the movement of water (or other liquid phases, e.g., NAPLs) are not (yet) readily available, direct comparison with experimental data is feasible only for water retention in the pore space. Pot et al. (2015) carried out the only such comparison to date, on the basis of quantitative data of the distribution of water and air in soil samples constituted of repacked aggregates, equilibrated at three matric potentials (-0.5, -1, and -2 kPa). The phase distribution data were derived from synchrotron X-ray CT images at a resolution of 4.6 μm. Water distribution was simulated by a two-phase Lattice-Boltzmann model (LBM) and a morphological model (MOSAIC). Results indicate that the lattice-Boltzmann model is able to predict remarkably well the location of air–water interfaces (Figure 4). When one lifts the assumption, motivated by capillary theory, that a pore can drain only if a connecting pore is already full of air, MOSAIC gives an acceptable approximation of the observed air–water interfaces (Figure 4). However, discretization of pores as geometrical primitives causes interfaces predicted by MOSAIC to have non-physical bulbous shapes. Nevertheless, given the huge difference in computing time required to run these two models (minutes for MOSAIC versus tens of hours for Lattice-Boltzmann), Pot et al. (2015) recommend that further research be carried out on the development of both modeling approach, in parallel. One might argue that the same recommendation applies to other numerical schemes as well.
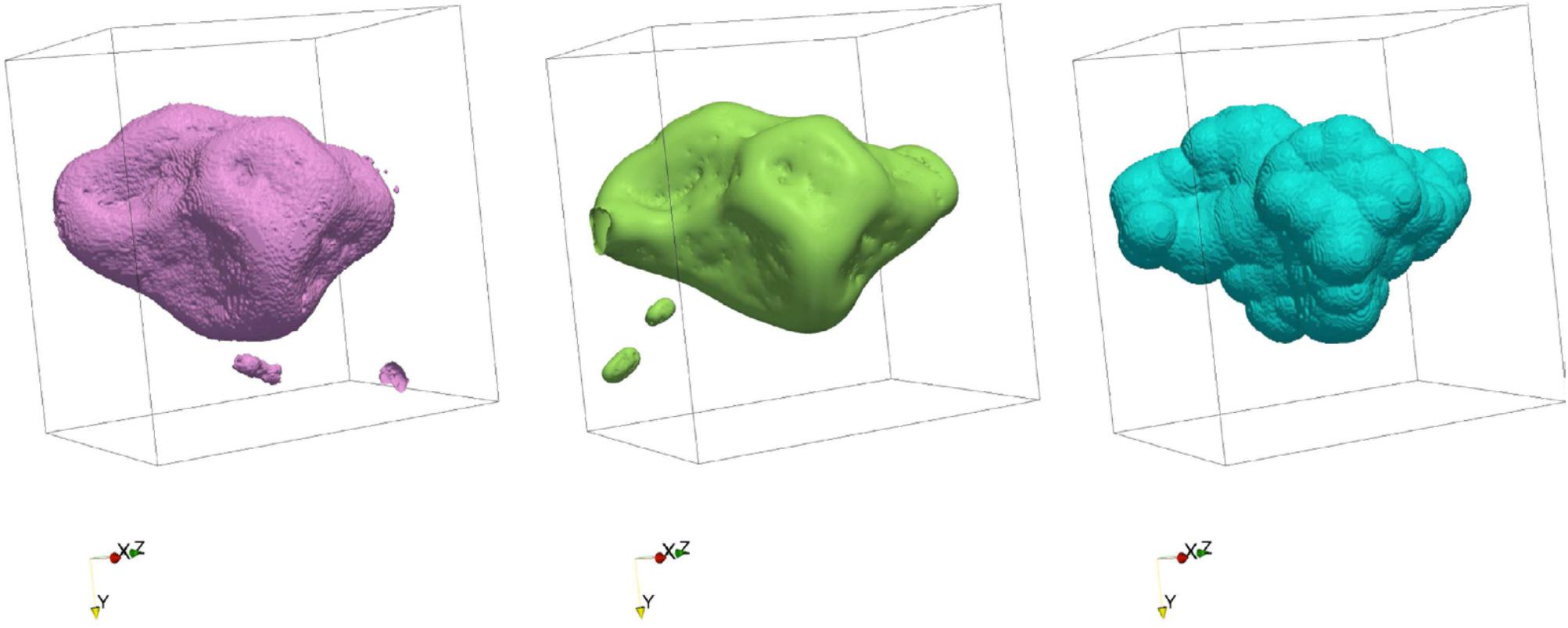
FIGURE 4. Air-water interface surfaces of a region of interest (region p1bkk04a) in a silt-loam soil, measured via synchrotron X-ray computed tomography (left), predicted using the Lattice-Boltzmann method (center), and predicted by the geometric primitive-based model MOSAIC (right) (Modified from Pot et al., 2015. Reprinted with permission).
Now that we have covered in some detail the progress achieved to date in the description and modeling of the physics of soils, it is probably a good idea, and a nice way to summarize things, to go back to the schematic diagram of Figure 1, and, with it, attempt to represent visually where we are at the moment. In Figure 5, this is done by shading in the diagram of Figure 1 the parts that correspond to work yet to be carried out. Admittedly, this is a subjective exercise, and different researchers, depending on how pessimistic or optimistic they are, may come up with contrasting evaluations. Yet, based on the detailed account provided above, the depiction in Figure 5 of the status of the physical front seems reasonable.
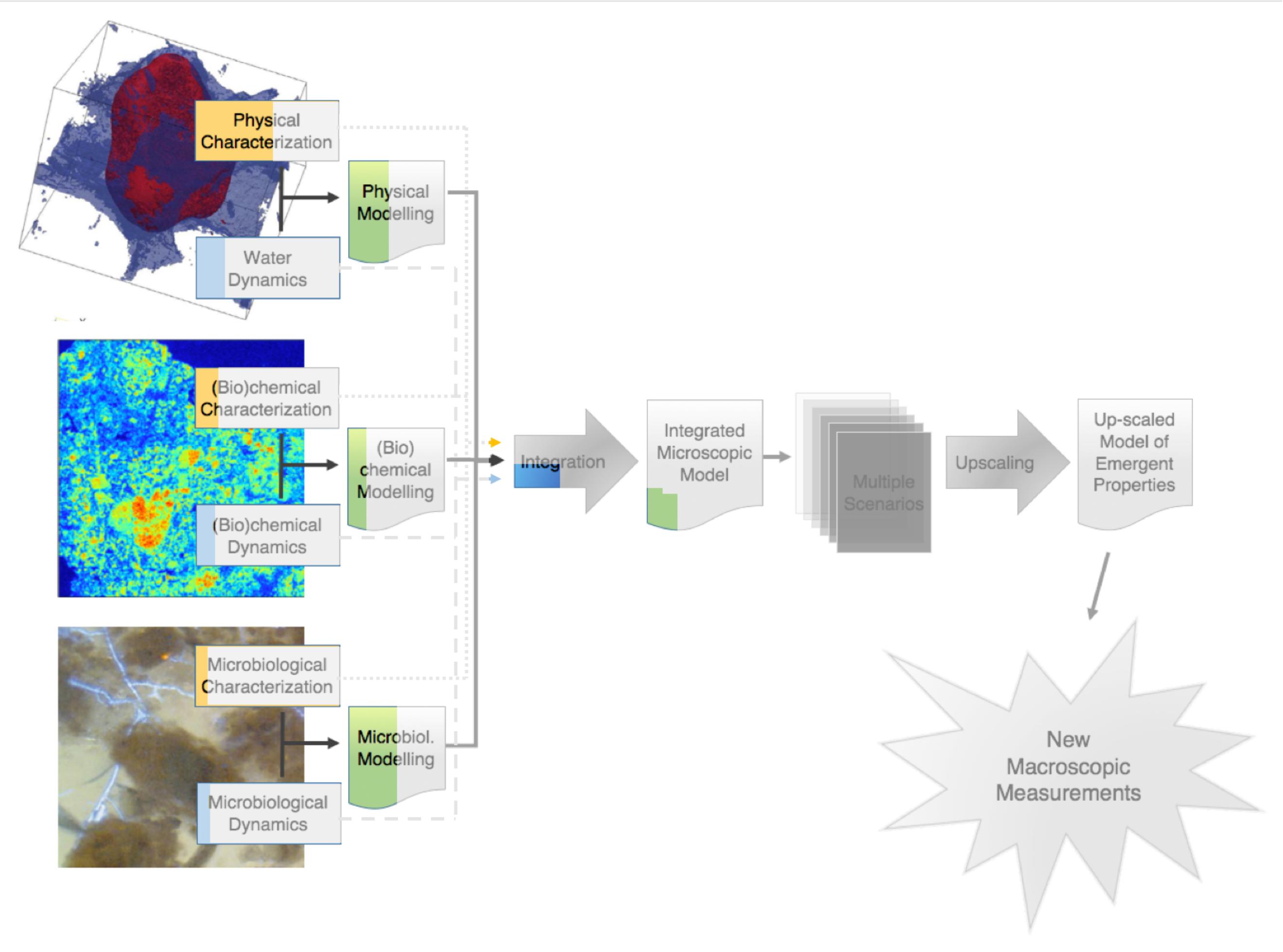
FIGURE 5. Visual assessment of the level of progress on the various steps in the research on the emergent properties of soils. The un-shaded parts (relative to the schematic diagram of Figure 1) correspond to the authors’ estimate of the progress achieved to date on each step. The shaded portion of the diagram still largely remain to be tackled.
In terms of the physics, Figure 5 summarizes visually the conclusion that work is relatively well advanced. Certainly, some areas require further research. Among others, the sub-resolution porosity of soils needs to be better apprehended. Yet, overall, significant progress has already been achieved on the experimental side. This is true to a lesser extent insofar as the dynamics of water (and other liquid phases) is concerned, due to difficulties in measuring changes in water content over sufficiently short times. In terms of modeling of the physics, some success has been achieved in the past decade, but there is room for improvement, in particular relative to the speed of computations.
Physical information is not sufficient to characterize microenvironments. Information about the chemistry, and in particular about the nature of reactive surfaces (e.g., Kotani-Tanoi et al., 2007) and of dissolved or adsorbed (bio)chemical species, is also important. Therefore, to complement the information available about the geometry and topology of the pore space in soils with similar information about the chemical properties, one should ideally be able, as a start, to measure in 3D the chemical composition of soils. A logical and relatively simple option in this respect, in principle (Egan et al., 2015), would be to take advantage systematically of the X-ray absorption K edge of all the elements we might be interested in, in a manner similar to the determination of the distribution of soil OM by adsorption of osmium (Peth et al., 2014). Unfortunately, the situation with osmium is somewhat unique. First, Os happens to have its K edge at 73.87 keV, in the middle of the range of energies that is typically required to deal with soil samples of a volume larger than a few cm3. In the literature so far, that range has extended from 30 keV for synchrotron X-rays (Pot et al., 2015) up to nominally 225 keV for polychromatic X-rays (Houston et al., 2013b). Practically, the low end of this energy range means that, in soil samples of a reasonably large size, it is not possible to detect elements with an atomic number lower than 51 in the periodic table, i.e., before antimony (Sb), which has an X-ray absorption K edge at 30.49 keV (Bearden and Burr, 1967). This constraint entails that only a few elements, like I, Cs, Ba, Hg, Tl, and Pb, can be mapped in 3D in soil samples, but even then, yet another condition has to be met, namely that these elements be present in soils in such high concentration that they affect appreciably the level of X-ray absorption in the voxels where they are located. In soils containing only trace-level concentrations of these elements, voxel grayscale values would be unlikely to differ much, if at all, immediately below and above the various K edges. This means in effect that, naturally or artificially, soils would have to be heavily laden, at least locally, with these elements for their spatial distribution to be detectable. For example, to be able to use the X-ray absorption K-edge of Cs to detect the distribution and movement of water in fine sand samples, Willson et al. (2012) had to use 10% (by mass) CsCl solutions. Similarly, to detect the transport of CaI2 solutions within small sand columns using the K-edge of iodine, Shokri (2014) had to use concentrated (5% by mass) CaI2 solutions. Altman et al. (2005) submerged soil aggregates in a 507 g L-1 CsCl solution in order to saturate the exchange complex with Cs, prior to scanning samples above and below the Cs K edge. Similarly, Keck et al. (2017) used a 0.3 mol L-1 BaCl2 solution to assess the distribution of cation adsorption sites in four undisturbed soils, three of which have a very high clay content (53.9–79.9% clay). In both Altman et al.’s (2005) and Keck et al.’s (2017) cases, the clay content of their soils makes one wonder whether the amount of CsCl and BaCl2, respectively, needed for complete saturation of the exchange complex with Cs+ and Ba2+, which appears necessary to detect significant differences in X-ray attenuation, may have also caused, respectively, a dispersion of clay particles or a shrinking of clayey aggregates, and therefore, changes in the scanned images that could have been misinterpreted.
Another option for the 3D determination of soil composition is X-ray fluorescence. It is far less constraining in terms of the elements it can map (elements starting with Na, atomic number 11, are possible candidates), but unfortunately it suffers from a similar limitation on the size of soil samples that can be analyzed. The principle of the method is simple. When an atom is irradiated with X- rays of sufficient energy, it ejects an inner orbital electron. An electron from higher orbitals then falls to fill the vacancy in the lower energy state, resulting in the release of a fluorescent X-ray. The energy of the fluorescent X-ray given off is characteristic of the energy difference between the two orbital energy levels, which is specific to each element, while the intensity of the emitted X-rays is related to the elemental abundance in the sample being analyzed. In X-ray fluorescence tomography systems (Bleuet et al., 2010), also called confocal XRF scanners (Patterson et al., 2010; Lühl et al., 2013) or spectrometers (Smolek et al., 2012), the presence of focusing optic both on the path of the incoming X-ray beam and between the sample and the detector allows the 3D elemental profiling of the sample, provided the travel path of fluorescence X-rays within the sample, once they are produced, is not too long, so that their absorption is minimized. This re-absorption of X-rays limits the 3D measurements to minute samples, extending to at most a few mm in any direction. In that context, McIntosh et al. (2015) used 3D micro X-ray fluorescence spectroscopy to determine non-destructively the elemental composition of minute aggregates of a plutonium-contaminated soil, within which they could identify distinct 30 μm-size Pu particles with a limit of detection <15 ng.
Until direct 3D mapping of the chemical properties of soils becomes technically feasible and more accessible, an alternative approach to obtain 3-dimensional chemical information about soils at the microscale is to carry out the same procedure used a couple of decades ago by Cousin et al. (1996, 1999), Vogel (1997), and Vogel and Roth (2001), to obtain insight into the physical properties of soils. The idea is to perform multiple cuts through soil samples, analyze in turn the (bio)chemical make-up of each exposed surface within the soil, then, using an interpolation technique, generate a 3-D picture from the data associated with the various surfaces. This procedure is routinely used for biological samples, such as human tissues, in which the serial removal of layers can be carried out easily, either by using a traditional microtome or a cryo-ultramicrotome, or via ion-beam ablation.
To these exposed surfaces, it is now possible to apply a panoply of different spectroscopic techniques, a luxury that not too long ago, researchers would not even have dreamt of. Indeed, until the mid-1990s, beside the standard bulk analytical methods, requiring a sizeable sample of soils and therefore precluding microscale analysis, the only method that was available to researchers to determine local (bio)chemical properties of soils was based on energy dispersive X-ray spectroscopy, either in the so-called “electron microprobe” or SEM-EDX equipment, or on electron energy loss spectroscopy (EELS) (Villemin et al., 1995). The first two of these instruments, in scanner mode, can in principle produce elemental maps like that of Figure 6A. Starting in the mid- to late 1990s, synchrotron facilities around the world began offering soil scientists the opportunity to run various types of analyses, including X-ray absorption near-edge structure (XANES) and near-edge X-ray absorption fine structure (NEXAFS) spectroscopies, which rapidly became popular because of the very useful information it is able to provide on the molecular environment of atoms, and therefore on element speciation (Prietzel et al., 2003; Schumacher et al., 2005; Solomon et al., 2005, 2012; Kinyangi et al., 2006; Christl and Kretzschmar, 2007; Wan et al., 2007; Strawn and Baker, 2009; Hesterberg et al., 2011; Milne et al., 2011; Jassogne et al., 2012; Kopittke et al., 2017). In most cases, the target of interest in this type of analysis is extremely minute in extent, a most a few μm2, but occasionally researchers have attempted to map properties over a slightly larger area, among other things to try to assess the heterogeneity of the composition of OM in soils (Figure 6B). Another synchrotron-based technique that has been used to some extent to obtain elemental maps involves X-ray micro-fluorescence (Hitchcock et al., 2004; Jacobson et al., 2007; Jassogne et al., 2012), which unfortunately does not provide information about speciation, but has the advantage that it can cover bigger surface areas (Figure 6C). Several other spectroscopic methods provide molecular- to micro-scale distributions of elements and isotopes and thus soil properties. Among them, the most commonly applied to soils in recent years is dynamic nanoscale secondary ion mass spectroscopy (NanoSIMS, Figure 6D). It uses a high-energy beam of ions (either Cs+ or O-) to eject secondary ions from a sample surface, which are then analyzed using a mass spectrometer, at a very high spatial resolution typically of the order of 100 nm for soil samples (Herrmann et al., 2007; Mueller et al., 2012, 2013, 2017). Slightly larger areas can be sampled with Static- or Time-of-flight Secondary Ion Mass Spectroscopy (Static SIMS or TofSIMS), which can target ions and small molecular fragments (Watrous and Dorrestein, 2011; Cerqueira et al., 2015; Worrich et al., 2017). Other spectroscopic methods, also working at spatial scales slightly larger than that of individual cells include Laser desorption/ionization (LDI), laser ablation inductively coupled plasma (LA-ICP), matrix-assisted laser desorption/ionization (MALDI) and desorption electrospray ionization (DESI) spectroscopies (Watrous and Dorrestein, 2011).
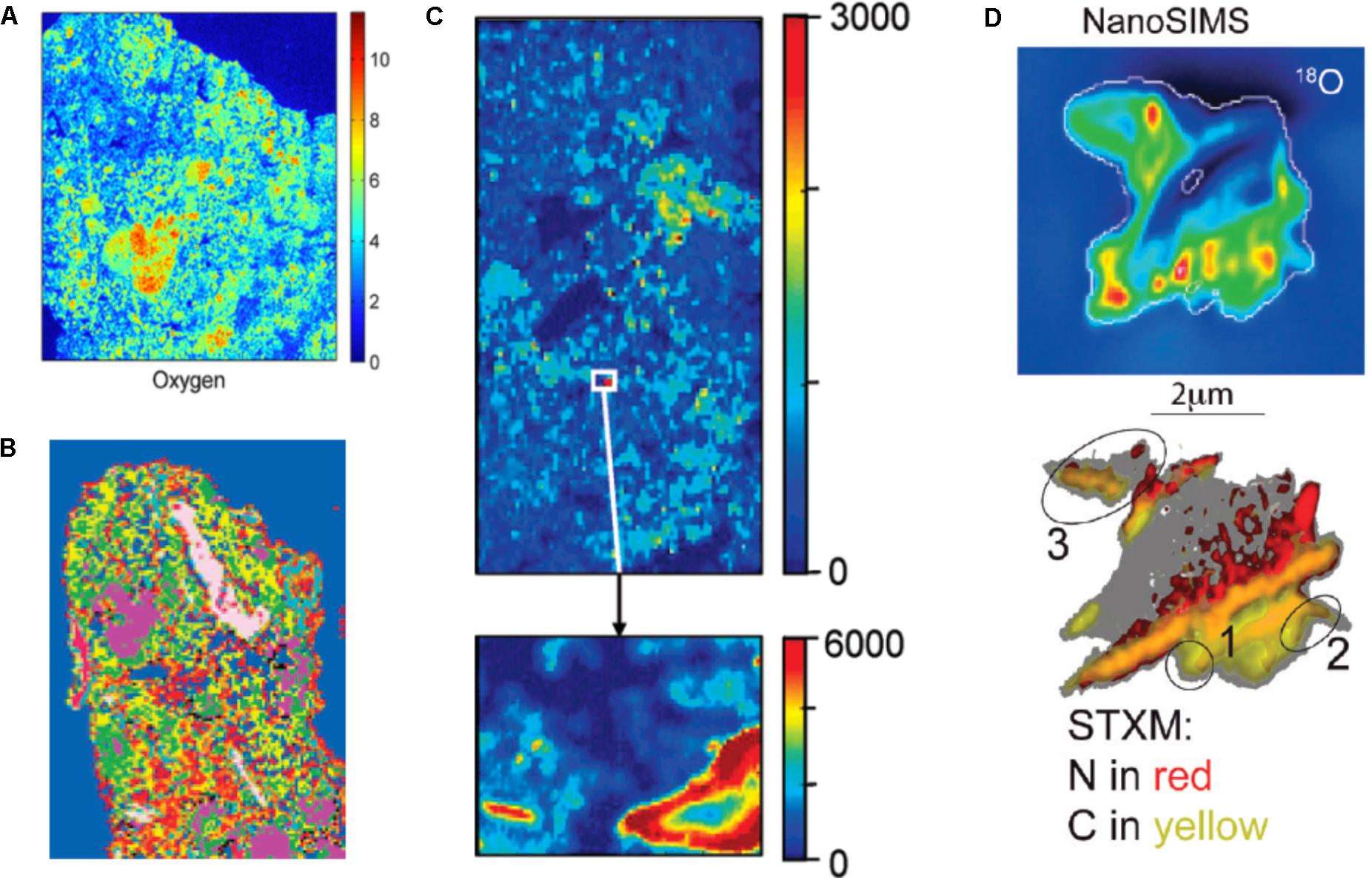
FIGURE 6. Illustrative examples of various chemical measurements that are now routinely carried out on 2D cross-sections through soil samples: (A) SEM-EDX mapping of the distribution of oxygen in a section through a calcareous soil from Scotland. The intensity of the color indicates the concentration of oxygen (Adapted from Hapca et al., 2015). (B) Cluster map showing the large heterogeneity of carbon forms within a soil micro-assemblage from Nandi Forest (Kenya) determined by NEXAFS in combination with scanning transmission X-ray microscopy (STXM). The field of view is 5.4 μmm by 7.5 μm (adapted from Lehmann et al., 2008. Reprinted with permission). (C) Synchrotron-based μXRF maps of Cu in a calcareous soil vineyard soil from Burgundy (France). The large map on top (2 by 4 cm) was obtained with a 0.3-mm spot size, the small map at the bottom (1.5 by 1 mm) with a 20-μm spot size. The color, from blue to red, is correlated with the Cu concentration (Adapted from Jacobson et al., 2007. Reprinted with permission). (D) NanoSIMS map of 18O in a soil aggregate (top) and C and N map of the same aggregate obtained by STXM (bottom). NEXAFS spectra were obtained in the three circled regions, whereas in the gray zones the sample was too thick to get NEXAFS spectra or was free of OM (Reprinted with permission from Remusat et al. (2012). Copyright [2012] American Chemical Society).
The fact that several of these techniques involve synchrotron X-ray beams, which are generally in extremely high demand, may explain why until now, their use has resulted in extremely few actual property maps of soil surfaces larger than microaggregates. Normally, experimentalists get “beam time” of 48–72 h at the most, during which it may be difficult to do a full scan of a thin section, for example. Due to the growing number of available instruments (soon to reach 50 worldwide), the situation for the use of NanoSIMS is improving, but access to all of them is highly coveted by researchers in many fields and only a hand full of laboratories routinely analyze soils. Fortunately, things might be getting better relatively soon in terms of NEXAFS since various groups of researchers (Peth et al., 2008; Müller et al., 2014; Kühl et al., 2016) have recently developed laser-based, benchtop-scale NEXAFS instruments, one of which is now commercially available. The results obtained to date, including on soil clays and OM, are very promising (Gleber et al., 2011; Sedlmair et al., 2012).
As exciting as the use of these various spectroscopic types of equipment might be, they afford measurements only of the concentration of various elements or their speciation, but not at all of the physico-chemical conditions in which specific (bio)chemical species are located. In particular, one would absolutely need information about the pH or redox potential locally in a porous medium, among other “thermodynamic” variables, to have a full picture of what is going on. Unfortunately, once a block of soil has been impregnated with resin and has cured, none of these variables is accessible any more. If somehow, one could cut through a soil, and obtain a relatively flat surface in the process, without having to solidify the soil and denature it in any way, it would be possible to obtain information on pH and redox potential through the application of microelectrodes, microsensor probes or planar optodes (Pedersen et al., 2015; Rubol et al., 2016; Keiluweit et al., 2018; Wanzek et al., 2018). Other gel-based approaches such as diffusive gradients in thin films (DGT, Santner et al., 2015) could be used to obtain 2D maps of the distribution of labile chemical species. Zymography is a methodology similar to “optodes” in that a planar membrane is brought to contact with exposed soil to measure the activity of various enzymes (Spohn et al., 2013; Razavi et al., 2016). The development of new sensors is a very active field, offering perhaps interesting opportunities for microscale soil characterization in a few years.
Since many 2D measurements can be carried out on cuts through soil samples, it is feasible under certain circumstances to produce from them a 3D image of the soil (bio)chemical characteristics. As with anything one does, it is useful to inquire whether this step is absolutely required. If methods were readily available to provide us with 3D images of the (bio)chemical composition of soils, we would not necessarily ask the question of why we need 3D images in the first place, but since the best we can get experimentally is 2D images of cuts through soils, it is worthwhile asking ourselves whether we really need to go through the added effort of the transition to 3D. Some researchers may be interested mostly in the relative distribution of chemical elements with respect to the pore system, to evaluate local gradients and accessibility of substrate for soil biota. They may consider that information in this respect can be obtained by single 2D slices of chemical maps that are projected on the 3D pore structure. From this standpoint, serial sectioning and interpolation are not necessary. A different perspective on the question, held for example by Hapca et al. (2011, 2015) is that, just like the degree of connectivity or tortuosity of the pore space in 2-dimensional cuts through a soil are generally different than in 3-dimensions, the spatial characteristics of the chemical make-up of soils, the distribution and local concentration gradients of targeted (bio)chemical compounds, also need to be estimated in 3D if one is to understand their influence on microbial processes. Experience will show in the future which one of these two perspectives is most conducive to progress.
Nevertheless, to obtain 3D information on (bio)chemical properties, the process of interpolation between 2D maps is complicated by the fact that cutting through soils is not as straightforward as it may seem. In mineral soils, the frequent presence of dense constituents reduces the range of techniques that can be used to cut or scrape away successive layers with minimal disturbance. Particularly when operating microtomes, the presence of constituents with markedly different densities often causes blades to deviate from their set course, so that eventually the exposed surfaces are not perfectly flat. Because of that, the correspondence of 2-D chemical or microbiological maps with the physical information obtained via computed tomography is likely to be poor, unless artifacts generated during soil cutting are accounted for.
Therefore, the first step in any attempt to simultaneously evaluate in 3D the physical, chemical, and biological characteristics of soil samples is to find a way to correct for any distortion that may occur when cutting or grinding down soil samples to successively expose surfaces on which 2-D chemical mapping is carried out, and to geo-reference these 2-D maps within the geometry of the soil solid phase, determined via X-ray computed tomography. A practical, automated procedure to accomplish these tasks has been developed by Hapca et al. (2011). This procedure, depicted in Figure 7, involves three successive steps, namely the reconstitution of the physical structure of a given soil layer surface, the alignment of the chemical maps with the reconstituted soil surface image, and finally the 3D alignment of the 2D chemical maps with the internal structure of the soil cube. Once this alignment is carried out satisfactorily, one can proceed to a statistical interpolation between successive geo-referenced 2D planes. Hapca et al. (2015) suggested that, for this interpolation, the 3D information produced via X-ray CT could be used as a guide. They proposed a method based on a regression tree method and ordinary kriging applied to residuals, and used it to predict the 3D spatial distribution of carbon, silicon, iron, and oxygen at the microscale. The spatial correlation between the X-ray grayscale intensities and the chemical maps made it possible to use a regression-tree model as an initial step to predict the 3D chemical composition. For chemical elements, e.g., iron, that have high attenuation and are sparsely distributed in a soil sample, the regression-tree model provides a good prediction, explaining as much as 90% of the variability in some of the data. However, for chemical elements with lower attenuation coefficients that are more homogenously distributed, such as carbon, silicon, or oxygen, the additional kriging of the regression tree residuals improved significantly the prediction with an increase in the R2 value from 0.221 to 0.324 for carbon, 0.312 to 0.423 for silicon, and 0.218 to 0.374 for oxygen, respectively. In principle, this method could be used for any (bio)chemical parameter that can be mapped on 2D cuts.
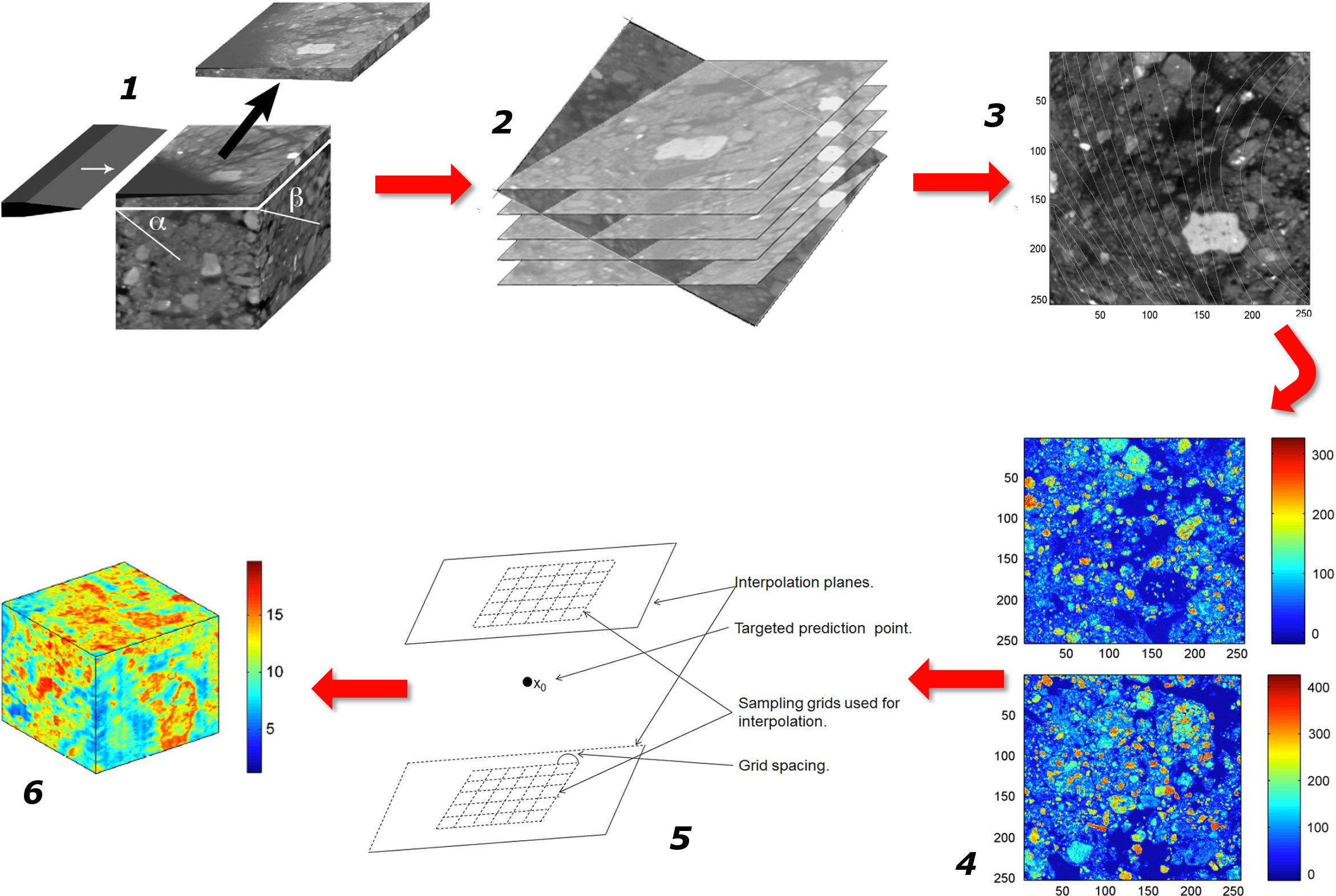
FIGURE 7. Schematic diagram of the successive steps in the 2D–3D interpolation method proposed by Hapca et al. (2011, 2015). (1) Illustration of a typical method to isolate a layer from a soil cube, with a microtome blade. The cut may be at angles α and β, respectively, with the x–y plane in the x and y directions, respectively, resulting in layer surfaces that are not strictly parallel to each other, (2) rotation of the chemical analysis plane within the 3D CT image, (3) Reconstituted CT image of the soil surface. The dotted lines correspond to the limits of the different masks applied to the successive layers during the zonation process, (4) Spatial distribution, measured with SEM–EDX, of silicon in the top and bottom soil surfaces of an individual slice through the soil sample, (5) schematic representation of the interpolation layers and the corresponding sampling grids for the selection of the interpolation points, (6) 3D prediction of the silicon distribution in soil sample.
Given the need to be able to work with 2D cuts through soils, e.g., by impregnating soils with resin and cutting through the resulting blocks in one way or another, to carry out measurements of (bio)chemical characteristics, it should come as no surprise that dynamical measurements at the microscale have been so far, and may remain for some time, impossible to achieve (Blaser et al., 2016). Of course, dynamical measurements of properties as they emerge at the macroscopic scale can be achieved relatively easily, for example breakthrough curves in column experiments serving as evidence of chemical transport. To some extent and for specific purposes, such macroscopic properties may suffice, but experience has shown time and again that in an of themselves, these macroscopic observations are not adequate to assess the soundness of microscale models of (bio)chemical dynamics.
Much of the energy, nutrient and information (signal molecules) flows in soil occur in aqueous phase. A broad range of organic and inorganic molecular forms are released into solution, taken up and metabolized, transformed by enzymes or immobilized on surfaces continuously. These constitute the most dynamic portion of the microbial environment. Although much research has been carried out on the soluble OM of soil (e.g., Rousk and Jones, 2010; Lerch et al., 2011), the scale at which the measurements have been made is inappropriate for understanding how the flows of energy, nutrients and information vary throughout the soil pore network; only an average value is obtained. Furthermore, extraction methods tend to introduce biases related to the choice of extractant and the disruption of the physical architecture of soil may release molecules not previously in solution (Inselsbacher et al., 2014). The use of miniaturized sampling devices (millimeter scale) such as microdialysis probes or micro-suction-cups offer the possibility of detecting gradients in the soil solution at scales that are getting closer to those relevant to microbial communities (Inselsbacher et al., 2014; Oburger and Schmidt, 2016), particularly in the rhizosphere where gradients are stronger. With microdialysis the soil solution is sampled by passive diffusion and is therefore likely to better reflect the soluble environment perceived by microbial communities. Recent research has shown that there are significant differences in the size and composition of soluble organic and inorganic N pools measured by microdialysis and those measured by the traditional extraction methods (Inselsbacher et al., 2014). The small size of the probes makes it possible to locate them precisely in soil samples using micro-CT imaging and to map diffusive fluxes in real time (Brackin et al., 2017).
Over the last decade, a very significant amount of work has been devoted by geochemists and environmental engineers to the development of computer models able to describe the fate of a number of chemical species of interest in porous and fractured media (Tartakovsky et al., 2007; Valocchi, 2012; Steefel et al., 2013; Yoon et al., 2015). These models typically combine a transport component with a chemical speciation algorithm. Chemical transport is described using a variety of approaches, including the Lattice-Boltzmann method, smooth particle hydrodynamics (Tartakovsky et al., 2007), hybrid Lattice-Boltzmann-direct numerical simulation (DNS) (Yoon et al., 2012), and pore network models (Li et al., 2006). In terms of the chemical reactions that the speciation algorithms describe, as Iliev et al. (2017) accurately point out, many models focus on chemical reactions occurring in solution, with only a few models dealing with reactions controlled by the reactivity of the surfaces, like the dissolution of mineral phases. This bias makes sense for the type of systems researchers have been trying to describe, namely aquifer materials, calcareous formations, sandstones, or simply laboratory set-ups filled with glass beads. In that general context, one of the key predictions of these models has been that under a wide range of situations, macroscopic-scale descriptions with “effective” (i.e., volume-averaged) parameters do not account adequately for model predictions when non-linear reactive transport processes are associated with highly localized chemical reactions and incomplete mixing within the porous medium (Li et al., 2007a,b,c; Battiato et al., 2011; Steefel et al., 2013).
The different features of current microscale geochemical models of reactive transport probably explain to a large extent why none of them has been used so far to describe soil processes at the microscale. Given the extremely high specific surface area, and the significant surface reactivity of many soils, in addition to the fact that the reactions that take place in soils are complicated by the presence of very heterogeneous OM, the speciation portion of typical microscale geochemical models would have to be entirely overhauled before it could be applied to soils. Nobody, as far as we are aware, seems to have launched into this work yet. In addition, even if someone had done that work, model predictions could not at the moment be compared with actual microscale measurements at this stage, as discussed in detail in the previous two subsections.
If we try to summarize graphically the state-of-the-art of the (bio)chemical characterization and modeling of soils at the microscale, it is clear that work in this area is far less advanced than on the physical front (Figure 5). Measurements of static features or of the dynamics of (bio)chemical species are still very limited. Some of this scarcity of data can, however, be addressed. There is indeed a great potential to generate many static, 2D data, using a wide array of experimental methods, and to extrapolate them to three dimensions. Modeling frameworks are available to describe the transport of reactive chemical species in porous or fractured media, but they would need to be modified substantially before they could be applied to soils, and that significant effort has not taken place yet.
To complement the 3D data related to the geometry and topology of soil pores, as well as 3D data about the (bio)chemical properties of soils, generated by interpolation among 2D pictures, it would be ideal if detailed 3D information could be obtained about the distribution of microorganisms in soils. Such information can be readily obtained in the case of wood, at least for fungi (Van den Bulcke et al., 2009). But unfortunately, for exactly the same reasons that hinder the direct 3D determination of the distribution of OM in soils, it has proven impossible so far to quantify the spatial heterogeneity of the distribution of microorganisms in actual soils.
When direct 3D measurements of biomass distribution in porous media have been obtained, it has so far always been under conditions that bear little similarity to real soils. Lilje et al. (2013) describe the development of a culture system and staining protocol they have used to obtain 3D quantitative data of filamentous and zoosporic soil fungi in an artificial matrix that was “developed to simulate the particulate nature of soil.” This artificial matrix consists of 500–900 μm diameter X-ray translucent polystyrene beads, which might be morphologically similar, to some extent, to coarse sand particles, but would likely have very different surface and hydration properties than typically highly heterogeneous soils. In many ways, the same comment pertains to the use of nuclear resonance imaging to detect “biofilms” in systems composed of polystyrene beads (Vogt et al., 2013). Sanderlin et al. (2013) pioneered the use of a very promising low-field magnetic resonance system to visualize the 3D distribution of biofilms in glass beads and sand particles, whereas a number of other authors used X-ray tomography to assess the distribution of biofilms in systems of glass beads (Davit et al., 2011; Iltis et al., 2011; Peszynska et al., 2016) or 2.5 mm-diameter Nafion pellets (Carrel et al., 2017). In all these cases, the properties and geometry of the systems investigated are drastically different from those of actual soils, which are generally characterized by a spatially dispersed- rather than concentrated biomass, and it is not clear at all at this stage how the transition from artificial media to actual soils will be made.
Since direct methods are lacking to quantify the 3D microscale distribution of bacteria in whole soil samples in one go, a number of authors have developed sampling techniques to obtain 3D information in other ways. Dechesne et al. (2003) developed such a technique and tested it in repacked soil columns. Their approach consists of a number of steps. The soil is first micro-sampled within several small subunit volumes of roughly the same volume (minimum sample side: length of 50 μm) within the columns, and these microsamples are subsequently tested for the presence or absence of targeted microorganisms, which in the original study were two bacterial strains but could equally easily have been archaea or fungi. A subsequent statistical analysis involves a comparison of experimental sampling data with data expected from limited sampling of numerous theoretical spatial distributions. Since the exact spatial location of the microsamples was not determined by Dechesne et al. (2003), they could identify only which statistical distributions of patches occupied by bacteria were possible within their sample. However, now that with CT, it might be possible to geolocalize small subsamples within soil columns, a similar approach could now be used to determine spatial distributions of various microorganisms as well, albeit at a relatively low resolution. An implementation of this approach is reported by Kravchenko et al. (2014b). In that work, ≈5 mm-sized soil fragments (referred to as macro-aggregates) were subjected to CT scanning, which provided information on pore architecture. Scanning was followed by cutting the macro-aggregates into geolocalized subsections, that is, the position of each subsection on the CT images was determined. Then, microbial community analyses of each geolocalized subsection via 16S rRNA pyrosequencing was conducted, enabling exploration of associations between presence of certain groups of microorganisms and abundances of soil pores of different sizes.
Given the technical difficulties associated with 3D measurements, it is natural that researchers attempted to find out what information could be obtained from 2D cuts through soils. Alexander and Jackson (1954, 1955) were apparently the first to suggest that thin sections of resin impregnated soil samples could be useful to observe algae, fungal hyphae, and bacteria, using either light or phase-contrast microscopy. They indicated that staining the soil before impregnation enhances the detection of hyaline mycelia and bacteria. Nevertheless, as soon as transmission and scanning electron microscope became available, microbiologists turned to the machines to obtain information about soil microorganisms. Consistently, they confirmed Clark’s (1951) observations. Bacteria, often coated by clay platelets, were generally present, not as “biofilms,” but as small colonies of a few cells, with many bacterial cells being dispersed in the rest of the soil as individual cells (Foster, 1988). As enlightening as these and other similar observations have been and still are5, a drawback with TEM and SEM is the fact that at least until recently they could provide only qualitative information. To obtain quantitative data about the distribution of microorganisms, researchers found it necessary to return to staining cells in thin sections that could be georeferenced easily and viewed in their entirety (e.g., Jones and Griffiths, 1964; White et al., 1994; Nunan et al., 2001, 2002, 2003; Li et al., 2003, 2004). Aside from non-specific stains like calcofluor white M2R applied before impregnation (Postma and Altemuller, 1990), or basic fuchsin and methylene blue applied after impregnation (Tippkötter et al., 1986), researchers also have been interested in selective staining techniques of specific cells, e.g., using fluorescence-conjugated antibody techniques (Postma and Altemuller, 1990), to observe the distribution of bacteria and fungal hyphae in soils. In some cases, problem arose because of the crystallization of the stains when in contact with soils (Harris et al., 2002, 2003). Nevertheless, after these slight technical issues got resolved, images of soil thin sections obtained with these various staining techniques showed clearly that for fungal hyphae, given their size, it is relatively straightforward to identify them (Figure 8A). But for bacteria and archaea, as clearly indicated in Figure 8B, a tremendous amount of skill (or faith, or both) is required to be able to identify a cell conclusively. Experience shows that part of the problem is related to the difficulty, with traditional light microscopes to focus precisely on a specific depth. It is possible, but tricky, to focus on the top surface of a thin section, hoping that one would then have a sharp image of the first 1 or 2 μm at the surface (Nunan et al., 2001). An easier solution consists of using a confocal laser microscope (e.g., Caldwell et al., 1992; DeLeo et al., 1997; Li et al., 2004), which can produce sharp 2D images at selected shallow depths within a soil thin section. With special software, 3D images can be reconstructed from a set of z-dependent 2D images. In principle, an extension of Hapca et al.’s (2015) statistical interpolation technique, described earlier, should make it possible to assemble these very thin 3D images into a full 3D picture of microbial microscale distribution in soil columns.
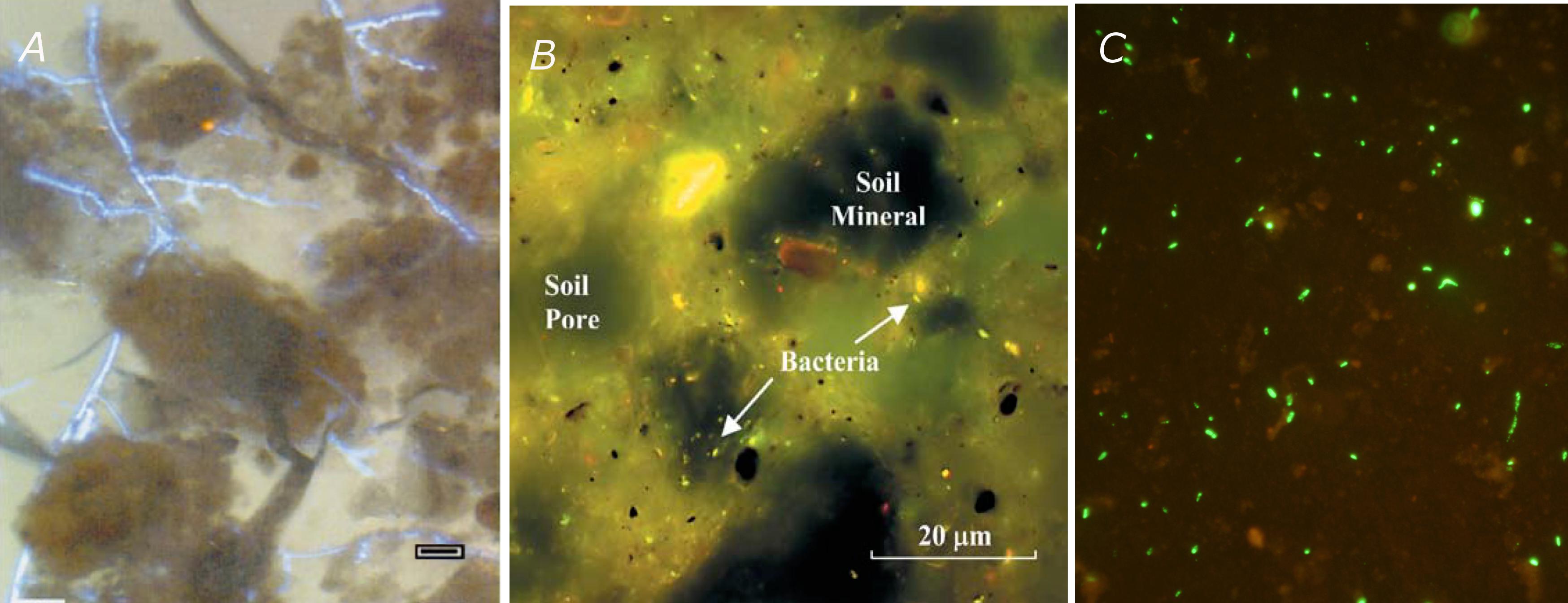
FIGURE 8. Examples of experimentally determined microbial distribution in soils: (A) microscopic image of hyphae of the fungus Rhizoctonia solani growing in the pore space of a sandy loam. Scale bar 20 μm (Harris et al., 2002. Reproduced with permission of the British Mycological Society). (B) Micrograph of ethidium bromide-stained thin sections of a silt loam soil after inoculation by Escherichia coli. Image obtained using an epifluorescence microscope with blue excitation (Modified from Li et al., 2004. Reprinted with permission). (C) CARD-FISH stained Bacillus subtilis cells in soil filter sections under double excitation filter 643 (465–505 and 564–892 nm) (Modified from Juyal et al., 2018).
A more difficult problem to resolve with traditional stains is related to their non-specific binding to OM or other soil constituents. As a result, many microbial cells may be undetectable against a very bright background of fluorescing soil constituents (Figure 8B). Luckily, that stumbling block too has found a solution in recent years, with the development of very reliable fluorescence in situ hybridization (FISH) techniques, which use fluorescent-labeled oligonucleotide probes (Pickup, 1995; Mcnaughton et al., 1996; Li et al., 2004; Eickhorst and Tippkötter, 2008a). Eickhorst and Tippkötter (2008b) have shown that when FISH is combined with tyramide signal amplification, in what is referred to as Catalyzed reporter deposition (CARD)-FISH, one typically obtains higher signal intensity and reduced interference of the background fluorescence of the soil. This methodology, or variants of it, have been used by a few researchers in the past decade to assess the distribution of bacteria in soils (Schmidt et al., 2012a,b, 2015; Schmidt and Eickhorst, 2014). The combination of CARD-FISH, or one of its variants, with confocal laser microscopy, affords a very powerful tool to visualize and quantify the distribution of microorganisms in soils, with a satisfactory depth-resolution.
The (few) measurements that have been carried out in thin sections have shed some light on the spatial pattern of microbial distribution in soils, but many questions remain. Based on the analysis of 744 images of observed bacterial distributions in soil thin sections taken at different depths, Raynaud and Nunan (2014) found that the distance between neighboring bacterial cells was, on average 12.46 μm and that these inter-cell distances were shorter near the soil surface (10.38 μm) than at depth (>18 μm), due to changes in cell densities. These authors’ analyses suggest that despite the very high number of cells and species in soil, bacteria only interact with a few other individuals. For example, at bacterial densities commonly found in bulk soil (108 cells per gram of soil), the number of neighbors a single bacterium has within an interaction distance of ca. 20 μm is relatively limited (120 cells on average). This analysis is based on calculations of Euclidean distances, which as Raynaud and Nunan (2014) acknowledge do not take into account the presence of solids, nor the tortuosity of the pore space.
A slightly different perspective on the distribution of bacteria is obtained when one proceeds to a simple back-of the-envelope calculation focused on the surfaces of soil pores, which may indicate somewhat better than Euclidean distances the degree of separation among bacterial cells. The specific surface area of soils varies between a low of 0.1 m2 per gram, for coarse sand, to a high of 800 m2 for a smectite clay, with most soils falling in between these extremes (Pennell, 2016). In a soil with a relatively low specific surface area of 10 m2 per gram, a population of 108 bacteria, each having on average a 1 μm2 cross section, would occupy a mere 0.0001 m2 per gram of soil, i.e., about 1/100,000th of the specific surface area. Even if one assumes bacterial cells to be much bigger, with a longitudinal cross-sectional area of 4 μm2, they would still cover only 1/25,000th of the specific surface. In other words, in either case, it is as if on the surface of soil solids, each cell would sit in the middle of an exclusion zone with an average radius of 178 μm. These numbers, in line with earlier estimates (Postma and van Veen, 1990; Grundmann, 2004; Young and Crawford, 2004; O’Donnell et al., 2007; Vos et al., 2013; Kuzyakov and Blagodatskaya, 2015) suggest a very lonely existence indeed, but of course, they are very crude estimates at best, ignoring any tendency cells may have to aggregate. Nevertheless, as more micrographs like that of Figure 8C become available in the coming years, and are combined with detailed information about the geometry of the pore space, it will become possible to refine our understanding of the patterns of spatial distribution not only of bacterial cells, but also of archaea, fungi, and bacteriophages.
The relatively low number of articles dealing with the distribution of microorganisms in soils may surprise, especially given the tools that have been at our disposal for at least a decade (CARD-FISH) or even two (confocal laser microscopes). To understand why that has been the case, and especially to try to change this unfortunate state of affairs, it is useful to describe briefly in what context this work has been carried out. This background is of course familiar to soil microbiologists, but researchers in other disciplines may not necessarily be aware of it.
In the mid-1960s, contrary to the views that had prevailed earlier, a number of researchers, seeking to make their life easier, decided to start viewing the soil microbial biomass as a blackbox, which meant ignoring entirely both the diversity of microorganisms present in soils and their relation to their immediate physico-chemical environment (Baveye, 2018). This approach became dominant for a few years, but in the 1980s and 1990s, a slew of molecular methods were developed to characterize DNA or RNA extracted from soils (Maron et al., 2011; Mendes et al., 2015). Microbiologists in growing numbers jumped on these methods enthusiastically, with the hope that they would give them the opportunity to get information about the diversity of soil microorganisms, i.e., would allow them access inside the blackbox of soil biomass, but still with the convenience of not having to worry about where exactly microorganisms are located. Indeed, in what became known as “metagenomic” and, more recently, “high throughput sequencing” methods, all that was needed to carry out the analysis of a given soil was to extract its microbial DNA or RNA. Indeed, virtually all researchers adopting this approach have entirely ignored the geometry of the pore space in soils or the characteristics of microenvironments (e.g., Nannipieri et al., 2003). Even the so-called “high-resolution” metagenomics (Kalyuzhnaya et al., 2008) ignores the physico-chemical environment of microorganisms. The claim was also made initially, at least by some, that metagenomic methods would enable researchers to avoid having to culture microorganisms in the laboratory, a process that for an estimated 98% of soil microbes, had proven impossible until then (Vogel et al., 2009).
In terms of actual benefits of work carried out along those lines, one should mention the fact that knowledge of the diversity of nucleic acids present in soils paved the way for the design of oligonucleotide probes used in FISH. In itself, this is an important outcome, but in most other ways, experience over the years has demonstrated that many if not all of the initial claims made by proponents of metagenomics were unrealistically optimistic. Scores of researchers have shown that the extraction of DNA or RNA from soils in many cases manages to get at only a fraction of the total amount present (Terrat et al., 2012; Knauth et al., 2013; Dlott et al., 2015; Wagner et al., 2015), that some of this DNA or RNA material is associated with dead or dormant cells, or is extracellular (Carini et al., 2016), and finally, that the information yielded by DNA or RNA analysis provides a picture of the genetic potential of microorganisms in a soil, not at all of what microorganisms actually do (Prosser et al., 2007; Blazewicz et al., 2013). Furthermore, it has become obvious that, far from alleviating the need to culture microorganisms, the metagenomic approach has increased the urgency of finding ways to identify and characterize vastly more organisms than is the case at the moment (Oremland et al., 2005; Baveye, 2009a,b; Pham and Kim, 2012; Puspita et al., 2012; Prakash et al., 2013). In spite of all these false hopes, limitations, and biases (Lombard et al., 2011; Prosser, 2015), it is fair to say that, at the moment, bulk “meta”-something-“omics” approaches (metagenomics, metatranscriptomics, metaproteomics, metabolomics) capture virtually all the funding dedicated to soil microbial diversity, and their use seems to be crucial to insure microbiologists’ career advancement. Year after year, articles praising the merits of metagenomics to uncover the secrets of soils (e.g., van Elsas et al., 2008; Vogel et al., 2009; Delmont et al., 2011; Fierer, 2017; Popescu and Cao, 2018) still make headlines6 and attract record numbers of citations.
But things may be changing. Roughly 12 years ago, partly in response to the inability of metagenomics to link functions to species and also because information needed to make sense of metagenomics data is lacking for a multitude of still uncultured microorganisms (Su et al., 2012; Shi et al., 2015), researchers started investigating ways to isolate and sequence the DNA and RNAs of individual cells. A number of articles (e.g., Wang and Bodovitz, 2010; Lasken, 2012, 2013; Pamp et al., 2012; Stepanauskas, 2012, 2015; Yilmaz and Singh, 2012; Blainey, 2013; Woyke et al., 2017) have recently retraced some of the key breakthroughs that have enabled what could probably be viewed as a fundamental revolution, in particular in the application of molecular biology techniques to environmental systems (Ishoey et al., 2008). The onset of that revolution is generally considered to be Raghunathan et al.’s (2005) proof-of-principle demonstration that it is possible to use the multiple displacement amplification (MDA) reaction to amplify genomic DNA from a single bacterium several billion fold, with a recovery of about 30% of the genome in the process. Marcy et al. (2007), Podar et al. (2007), and Kvist et al. (2007) applied MDA to environmental cells and established the feasibility of single-cell genome sequencing from uncultivated targets. Woyke et al. (2010) showed that it is possible to produce a completely closed genome from an individual cell. Progress has been very rapid since (Supplementary Figure S1), including in RNA sequencing (e.g., Pan et al., 2013; Svensson et al., 2017), single-cell transcriptomics (e.g., Kang et al., 2011), and single-cell metabolomics (e.g., Heinemann and Zenobi, 2011).
All these single-cell techniques offer tremendous potential for the study of soil microorganisms as various researchers have already pointed out (e.g., Ishii et al., 2010; Pedersen et al., 2015), provided two key challenges can be overcome. The first is related to the fact that FISH, the technique that seems most promising at the moment to locate bacteria and archaea in 2D cuts through soils, has been documented to interfere with single-cell genome recovery (Woyke et al., 2017). This problem may be partly avoided by complementing FISH with other methods to detect and characterize microorganisms, like Raman spectroscopy (see below). The second challenge is related to isolating individual cells from their microenvironments in soils. In the past, various researchers have used micromanipulators of different types over the years, to extract hyphae fragments from soils (Söderström and Erland, 1986) or to sample bacteria on soil surfaces (Dennis et al., 2008). Ashida et al. (2010) and Nishizawa et al. (2012) adopted a micromanipulator originally developed by Fröhlich and König (1999, 2000) and Ishøy et al. (2006), and consisting of a microcapillary (with an outside tip diameter of 60–100 μm), to extract individual, artificially elongated bacteria from a rice paddy soil sample, and subsequently proceed to 16S rRNA gene analysis. More recently, Ringeisen et al. (2015) have used a laser printing technique, called BioLP, to isolate viable microorganisms from a thin layer of soil spread over a titanium-coated quartz plate. For both the micromanipulator and laser printing technologies, the technological challenge at this point is to design a sampling method that would have a far smaller footprint than is currently achievable, to make it possible to zero in on a single cell or a very small group of cells in a soil microhabitat. Since intracellular capillary microsensors with tip diameters less than 1 μm have been used by microbiologists for at least 60 years (Draper and Weidmann, 1951) and the technology of “optical tweezers” has evolved tremendously in the last 2 decades (e.g., Fröhlich and König, 2006; Whitley et al., 2017), it may not be foolish to imagine that we could come up with a way to extract single bacterial cells from soils in a very efficient manner in the not too distant future.
An argument that could be put forth to downplay the interest of this type of single-cell analysis is that such a detailed description of microbial communities is an unnecessary luxury for understanding a large number of microbial functions in soil. It is widely accepted that microbial communities are characterized by a functional redundancy with respect to a range of functions, such as organic C mineralization (e.g., Wertz et al., 2006, 2007; Allison and Martiny, 2008), meaning that the loss of a large number of species does not have a significant effect on functions of interest. Where functional redundancy is apparent, it is possible that viewing microbial communities in soil as a distribution of active sites rather than a distribution of species might suffice. Further research is needed to determine under what conditions information about individual microbial cells is crucial and when it is superfluous.
Given the need to impregnate soils with resin and to cut through the resulting block in one way or another to obtain information about the distribution of microorganisms and the (bio)chemical features of the microenvironments where they reside, it is clear that it is not possible at this stage to monitor in real time, at the microscale, the dynamics of microbial processes in actual soils. And, to be realistic in our expectations, for bacteria, archaea, and definitely viruses, it may be that we shall never be able to monitor their activity directly in soils.
We might be able, however, to observe the dynamics of these organisms or viruses in 2-dimensional, manufactured soil-like structures, generally referred to as micromodels or “microfluidic” devices. The development of these micromodels has been the object of significant research over the last 20 years (Karadimitriou and Hassanizadeh, 2012; Stanley et al., 2014, 2016; Stanley and van der Heijden, 2017; Aleklett et al., 2018). Early generations of micromodels, still in use to some extent (e.g., Dupin and McCarty, 1999; Stewart and Fogler, 2001; Lanning and Ford, 2002; Coyte et al., 2017; Borer et al., 2018) had idealized geometric properties, being basically two-dimensional networks of straight cylindrical segments, etched in glass or plexiglass. But as technology matured, second- and third-generation structures have become progressively closer to what one would find in a typical fine- to medium sandy soil. Of course, the material these micromodels are made of, often polydimethylsiloxane (PDMS), does not have the same surface properties as sand or silt particles. For some microbial processes this may be an issue, and it makes it impossible to reproduce surface chemical properties of soils, but at least the geometry of the pore space is realistic. With such a soil-like micromodel, Deng et al. (2015) have been able to observe the effect of extracellular polymeric substances (EPS), released by bacteria, on the drying kinetics of the pore space, whereas Rubinstein et al. (2015) have used it to demonstrate the effect a protist, the ciliate Colpoda sp., can have on the transport of nanoparticles through soils.
The use of micromodels opens up a number of very interesting avenues for further research, which may provide useful insight. For example, micromodels would seem to be ideal systems to instrument with optodes (Pedersen et al., 2015; Rubol et al., 2016), in order to access some of the physico-chemical parameters (pH, redox potential) that at the moment we cannot measure in soils at the microscale. The use of micromodels might also allow us to better understand how the moisture content of soils influences the activity of bacteria, archaea, and fungi. There is macroscopic evidence that these organisms react very differently to high or low moisture contents (e.g., Otten et al., 1999; Otten and Gilligan, 2006; Kaisermann et al., 2015; Baveye et al., 2016b), as do their predators, which we should not forget (Stefana et al., 2014), and it would be very useful to obtain direct evidence of this at the pore scale.
However, it is likely that for micromodels to give us valuable insight about actual soils, at least two significant challenges will have to be addressed and resolved. The first concerns the connectivity of the pore space. In nature, microorganisms evolve in a 3-dimensional space, which is significantly more connected than is achievable in 2D (e.g., discussion in Hapca et al., 2011). It will therefore be crucial to find a way to relate 2D observation made in micromodels with the more complex situation found in soils. The second challenge is related to what was referred to as “sub-resolution” pores in CT images. The issue, still very much an object of debate, is whether these pores are important to understand microbial activities in soils, and therefore whether they should be present in micromodels. A body of literature, published over the last few decades, argues that pores in the 30 to 150 μm size range are particularly crucial to understand microbial activity (e.g., Kravchenko and Guber, 2017). Specifically, pores of this size group were found to harbor greater abundance of a number of bacteria groups, such as copiotrophic actinobacteria, firmicutes, and proteobacteria (Kravchenko et al., 2014b) and presence of such pores was associated with greater microbial activity and greater OM decomposition (Killham et al., 1993; Chenu et al., 2001; Strong et al., 2004; Ruamps et al., 2011, 2013), in spite of a higher predation pressure in larger pores, compared to small ones (e.g., Wright et al., 1995). Recently, it was also found that dissolved OM contained within such pores is more labile, having less lignin and tannin-like compounds, than that in small (<6 μm) pores (Bailey et al., 2017; Smith et al., 2017). One perspective on these data is that pores of this size range offer better micro-environmental conditions, e.g., O2 and water supply, while providing enough space not only for individual organisms but for formation of microbial colonies, which then generate these sizeable experimentally detectable activities and changes in soil characteristics.
Based on this evidence, one would be temped to conclude that when constructing micromodels, one could safely ignore small pores, which would undoubtedly make everyone’s life simpler. However, the much lower connectivity of the pore space that would result from that may prevent us from describing correctly some of the processes occurring in soils, both in terms of microbial movement and metabolism. To resist predation, it may be vital for bacteria and archaea to be able to find refuge in smaller pores in which amoebae and particularly ciliates are not able to penetrate. One expects motile bacterial and archaeal cells, sometimes as small as 0.3 μm in diameter or width in soils, to be able to move relatively easily in and out of 2–3 μm-wide pores filled with water. But, as the experiments of Männik et al. (2009) with micro-fabricated channels show, some bacterial cells (of Escherichia coli, but not of Bacillus subtilis), can penetrate pores smaller than themselves. Although organisms constricted in narrow channels had no mobility and were squeezed, they could still penetrate the channel by growth and division (Hallett et al., 2013). Perhaps more important still is the fact that, given their even smaller size, exoenzymes that bacteria and archaea, as well as fungal hyphae, release into the soil solution can move in and out of tiny pores. Likewise, solutes present in the soil solution can diffuse in and out of the smaller pores, including the very narrow 1.8 nm-wide spaces between clay particles (Dumestre et al., 2000, 2006). In particular, dissolved components of the OM that is located, and possibly to some extent is physically protected, in small pores can also diffuse out into wider pores, where they can be taken up by microorganisms or be transported with the percolating water. Results obtained by Michelson et al. (2017) using a microfluidic device also suggest that members of the Geobacteraceae family produce nanowires that are able to penetrate in pore spaces too small for cell passage and, there, up to 15 μm away from cell bodies, reduce Mn(IV) and Fe(III) oxides via long-range extracellular electron transport. Finally, the (so far virtually ignored) bacteriophages swarming in the pore space of soils in huge numbers (Ashelford et al., 2003), with sizes sometimes as small as a few tens of nanometers, are likely to diffuse through tiny pores as well, and it may turn out that to understand the dynamics of phages in soils, a topic of increasing interest at the moment, it will be necessary to deal with sub-resolution pores in one way or another.
For all these reasons, inclusion of sub-micron pores in micromodels is a technological challenge that may need to be met if we want to use micromodels to gain knowledge about a range of soil processes, but in the meantime, a number of interesting processes, which are not or are only marginally influenced by sub-resolution pores, can still be studied with existing soil-like micromodels, such as the proliferation of fungal hyphae (as long as the release of exoenzymes is not the key mechanism by which fungi metabolize food sources), or the effect of bacterial activity on water or particle retention and movement in larger pores (e.g., Deng et al., 2015; Rubinstein et al., 2015).
Contrary to what has happened on the (bio)chemical scene, the least one can say is that the lack of experimental data about the spatial distribution and activity of microorganisms in soils has not discouraged at all a number of researchers from developing increasingly more sophisticated biokinetic models. On the contrary, work in this area, overwhelmingly carried out by soil physicists, mathematicians, and computer scientists, has been extensive over the last decade (e.g., Thullner and Baveye, 2008; Heße et al., 2009; Gras et al., 2010, 2011; Wang and Or, 2010; Gharasoo et al., 2012; Ebrahimi and Or, 2014, 2015, 2016, 2017; Vogel et al., 2015; Tecon and Or, 2017a,b; Wilmoth et al., 2018; Vogel H.J. et al., 2018; Vogel L.E. et al., 2018). Some of this research has consisted at first of a relatively straightforward extension to the microscale of macroscopic modeling approaches originally developed for saturated porous media, with the biomass consisting exclusively of bacteria, attached to surfaces and growing in response to the influx of a substrate, according to Monod’s equation in its simplest formulation (e.g., Widdowson et al., 1988; Baveye and Valocchi, 1989; Loehle and Johnson, 1994; Vandevivere et al., 1995). Over the years, in addition to being extended to the microscale in variably saturated porous media, the original biokinetic model has also been greatly improved (e.g., Hron et al., 2015). Description of bacterial growth has included an explicit account of endogenous metabolism. Bacteria have been allowed to move via chemotaxis, in response to substrate concentration gradients (Olson et al., 2004; Ebrahimi and Or, 2014; Son et al., 2015), and to become dormant (Gras et al., 2011; Resat et al., 2012; Joergensen and Wichern, 2018), under a range of conditions. Instead of simply relying on population-level kinetic equations like Monod’s, researchers have progressively turned to individual- or agent-based models, recognizing that locally in soils, the number of bacterial cells tends to be very small, and therefore the large-number assumption embodied in Monod’s equation is no longer met (Hellweger et al., 2016). In all these respects, progress in the development of the models over the last decade has been very significant, although from a strictly bacteriological perspective, the models currently available still fail to include a number of processes that might be very significant in soils, like conjugation, quorum sensing, siderophore production, exopolymer and exoenzyme production, filamentous growth of some bacterial strains, or the release of antibiotics to compete with other bacteria or archea (Wolf et al., 2013; Abrudan et al., 2015; DeAngelis, 2016).
In parallel with this modeling effort related to bacteria, various researchers have endeavored to develop computer models to describe the 3-dimensional proliferation of fungi in various types of environments (Otten et al., 2001; Falconer et al., 2005, 2007, 2012, 2015; Boswell and Hopkins, 2008; Jeger et al., 2008; Pajor et al., 2010; Kravchenko A. et al., 2011; Kravchenko A.N. et al., 2011; Hopkins and Boswell, 2012; Cazelles et al., 2013; de Ulzurrun et al., 2017). These models include a number of processes, which for soils might be very relevant, like biomass recycling and the release of exo-enzymes. In applications of some of these models to soils, thresholded CT images can be used to establish the boundaries of the geometric domain in which fungal growth occurs.
Since most soils simultaneously harbor bacteria, archaea, and fungi (among many other organisms), one would expect that the two families of models developed so far to describe specifically the activity of these organisms would have been combined at some stage. This would seem to make a lot of sense, especially as far as bacteria are concerned. One might argue that, under a number of circumstances (e.g., discrete POM serving as exclusive carbon source to fungi), the presence or not of bacteria in the pore space is in general pretty much irrelevant for the proliferation of fungal hyphae. Exceptions occur when bacteria have fungicidal activity (Stanley et al., 2014), and influence the propagation of hyphae. But it is more common for fungi to exert an influence on the behavior and spread of bacteria. Over the last few years, evidence has accumulated that bacteria, “Hitchhikers on the fungal highway” as Warmink et al. (2011) put it, can hop on, or at least be passively carried by, fungal hyphae as they propagate through the pore space (Kohlmeier et al., 2005; Warmink et al., 2011; Ellegaard-Jensen et al., 2014; Stanley et al., 2014), with the consequence that mycelia may be having a very significant role in gene transfer in soils (Berthold et al., 2016; Nazir et al., 2017). Recent ToF- and NanoSIMS measurements carried out by Worrich et al. (2017) also demonstrate that fungal or fungal-like (oomycete) mycelia can reduce water and nutrient stresses experienced by bacteria in otherwise dry and nutrient-poor microhabitats. All these recent observations seem to run counter to previous research suggesting that the high biodiversity of bacterial populations in soils, as well as their community structure, could be accounted for by the low connectivity of the water-filled pore space (e.g., Tiedje et al., 2001; Fierer et al., 2003; Treves et al., 2003; Carson et al., 2010; Ebrahimi and Or, 2015).
Therefore, it would seem important for models describing the activity of microorganisms in soils to simultaneously involve bacteria, archaea, and fungi, and in particular describe the transport of bacterial or archaeal cells by fungal hyphae or the transfer of water and nutrients by mycelia in the pore space. Unfortunately, this is far easier said than done, because of the difference in scale at which these various groups of organisms operate. In order to describe the activity of fungi realistically, one needs to model a volume of soil in which typically very large numbers of bacterial or archaeal cells would be located, making the prediction of microbial activity extremely CPU intensive, especially when using individual-based models. Within the context of this type of model, there is a possibility to deal with groups of cells, or “super-individuals,” as if they were single individuals, to save computing time (e.g., Scheffer et al., 1995) but the approach does not appear to have been used yet to describe bacterial or archeal populations in soils, and it remains to be seen whether it really makes sense.
This hurdle we need to resolve, somehow, about including both bacteria and fungi in the same simulations, raises a broader question of how many other similar hurdles we need to face. How much biodiversity needs to be included in models, in order to have meaningful insights into what is occurring in soils, and in order for the label of “microbial” used abusively in the title of many articles dealing only with bacteria (e.g., Or, 2002; Ebrahimi and Or, 2015), is really justified? In principle, there is no problem in developing models that involve only one type of microorganism, as long as the conclusions reached are restricted to the organism(s) involved, under the conditions assumed in the modeling, and are not considered generally applicable to soils, which contain a multitude of other organisms beside the targeted one(s) (this point is discussed in detail in Baveye et al., 2016b). Clearly, however, such limited models, from which crucial components are missing, are not likely at all to be very useful in the long run in the context of the program defined in Figure 1. To make real progress, we need a model that includes as many as possible of the organisms that are relevant to the goal that is being pursued. In general, bacteria, archaea, fungi probably all need to be included, but so do their predators (e.g., DeLeo and Baveye, 1997; Ronn et al., 2012), as well as bacterial and archaeal phages that are present in the soil in large numbers and are more and more suspected to have a very significant, yet still largely misunderstood, influence on microbial dynamics (Williamson et al., 2017; Pratama and van Elsas, 2018). Likewise, the too often ignored aspects of mesofaunal and macrofaunal activity in soils (Briones, 2014), which directly relate to the growth and metabolism of microorganisms, probably also ought to be accounted for, somehow. Depending on the specific questions we try to address, it may be that, in addition to microorganisms, phages and the mesofauna all need to be taken into account in our description of soils, in which case individual-based techniques might not be workable, or only some organisms need to be involved explicitly. Further research is needed to enlighten us in this respect.
As we navigate among all these additional components that may, or may not, need to be added to current microbial models to make them encompass more of the known biodiversity of soils, it soon becomes apparent that progress vitally requires being able to compare model predictions with actual measurements, which at this point, as was discussed in previous sections, are sadly lacking… Over the years, various soil scientists have reacted strongly, sometimes eloquently (Thomas, 1992), sometimes caustically (Philip, 1991), against modeling efforts that are not systematically backed by sound experimental support. A well-known philosopher and writer, David Henry Thoreau, offered a long time ago a more positive take on a similar situation (in a different context), when he wrote: “If you have built castles in the air, your work need not be lost; that is where they should be. Now put the foundations under them.” (Thoreau, 1854). Clearly, in the case of models of microbial activity in the hugely complicated environment that soils constitute for microorganisms, it seems essential to heed this advice, and to obtain relatively quickly the type of experimental data that would enable us to establish our modeling efforts on a much stronger foundation.
In terms of measurements, the situation on the microbiological front is very similar to that found on the (bio)chemical one (Figure 5). Quantitative measurements of microbial distribution or dynamics are extremely limited and related only to a very small portion of the biodiversity found in soils. Unlike on the (bio)chemical front, however, efforts to model the activity of microorganisms in soils have been extensive, especially regarding bacteria, and have produced some interesting predictions. Nevertheless, this effort has so far been entirely focused on selected bacteria and just a few species of non-sporulating fungi, and in the absence of actual measurements, it is not clear at all how close to reality model predictions are.
The next step in the program of Figure 1 consists of integrating disciplinary insights into a coherent integrated picture of microbial processes in soils. This integration should take place at both the static and dynamic experimental levels, and in terms of modeling, with the understanding that what is needed eventually, going into the next step, is a comprehensive, thoroughly tested microscale model of microbial activity. Right from the onset, one should expect this integration to pose significant challenges. Besides the usual institutional impediments to any kind of interdisciplinary research (see, e.g., Baveye, 2013b, 2014; Baveye et al., 2014), this integration is complicated by the fact that separate measurements that need to be made on the same soil samples often require heavy pieces of equipment that are not commonly found in a single location, causing logistic issues.
At the experimental level, one would expect that since data are still scanty on the (bio)chemical and microbiological scenes, very little integration would have taken place. And yet, encouragingly, some countries have set up a framework for efforts along these lines (e.g., Kögel-Knabner et al., 2008) and there have already been several attempts at integrating various types of experimental approaches. A case in point is the very interesting article by Rawlins et al. (2016). These authors attempt to determine how soil heterotrophic respiration (SHR) is related to the accessibility of OM to microbes in aggregates of a soil from the United Kingdom. They use a combination of synchrotron X-ray CT, osmium staining, and total organic carbon (TOC) content measurements to quantify the 3D distribution of OM, pore space, and mineral phases, and eventually find a weak correlation (r = 0,12) between SHR and a measure of accessibility of OM, which they define as the probability that a given voxel, “filled” with OM be adjacent to a pore voxel. More recently, Yu et al. (2017) combine synchrotron-based 3D X-ray micro-computed tomography with scanning electron microscopy of 2D slices of two different soils, coupled with an energy –dispersive X-ray spectrometer (SEM-EDX) to establish the relation between pore architecture and cementing substances (iron oxide, carbon) in soil aggregates. In an even more recent article, Vidal et al. (2018) combine Nano-SIMS to FIB-SEM to gain information about the distribution of minerals and biomass in the vicinity of roots. They show in particular that bacteria near roots are surrounded by iron oxides, and that some microaggregates are intimately associated with the surface of fungal hyphae.
In terms of the integration of models, some limited work has been carried out as well. Falconer et al. (2012) focus on combining predictions of water retention in a soil, using a LB approach, with a model of the growth of fungal hyphae. Simulation results, based on X-ray CT images of three different soils, show that the water distribution in the soils is affected more by the pore size distribution than by the total porosity of the soils. The presence of water decreases the colonization efficiency of the fungi, as evinced by a decline in the magnitude of all fungal biomass functional measures, in all three samples. The architecture of the soils and water distribution have an effect on the general morphology of the hyphal network, with a “looped” configuration in one soil, due to growing around water droplets. These morphologic differences are satisfactorily discriminated by Minkowski functionals, applied to the fungal biomass.
Two other articles, also combining different models, demonstrate the large benefits that can be derived from the availability of models. Once an X-ray CT image of a soil have been obtained, one can artificially create all kinds of “what-if” scenarios, in which one can imagine that the OM or the microorganisms are distributed in the soil in a multitude of different manners, and one can determine the effect that these relative distributions have on some macroscopic outcome, like the amount of CO2 evolved from a given soil sample. Of course, such “what-if” scenarios do not alleviate the need to secure actual measurements, of microbial and OM distribution, as well as of any macroscopic outcome one is interested in, but the scenarios can definitely complement and expand the experimental data set in very advantageous ways, if only for the purposes of testing statistically various types of novel metrics of microscale heterogeneity (discussed later on). For example, Falconer et al. (2015) obtain strikingly different predictions of evolved CO2 and fungal biomass production in soils, depending on how an identical amount of POM is distributed spatially in the pore space (Figure 9). When POM is present in relatively large chunks (>200 μm in diameter, in scenarios 1 and 4), results in terms of both CO2 evolution and biomass C produced show a large variability and in some cases a high level of production. On the contrary, when an identical amount of POM is more finely dispersed in the soil sample (scenarios 2 and 3), CO2 evolution and biomass C production both vanish. This result may surprise, but as discussed by Falconer et al. (2015), it is entirely consistent with the foraging pattern of fungi. In another article also published just a few years ago, Vogel et al. (2015) analyze numerically the role of meso- and macropore topology on the biodegradation of a soluble carbon substrate in variably saturated and pure diffusion conditions. The simulations involve the coupling of a LB model to describe the retention of water in the pore space, and a simplified compartmental biodegradation model that does not allow bacterial motility. Not unexpectedly, Vogel et al. (2015) show that under these conditions, the biodegradation of the solute is strongly dependent on the separation distance between bacteria and solute, and is influenced by the moisture content of the soil.
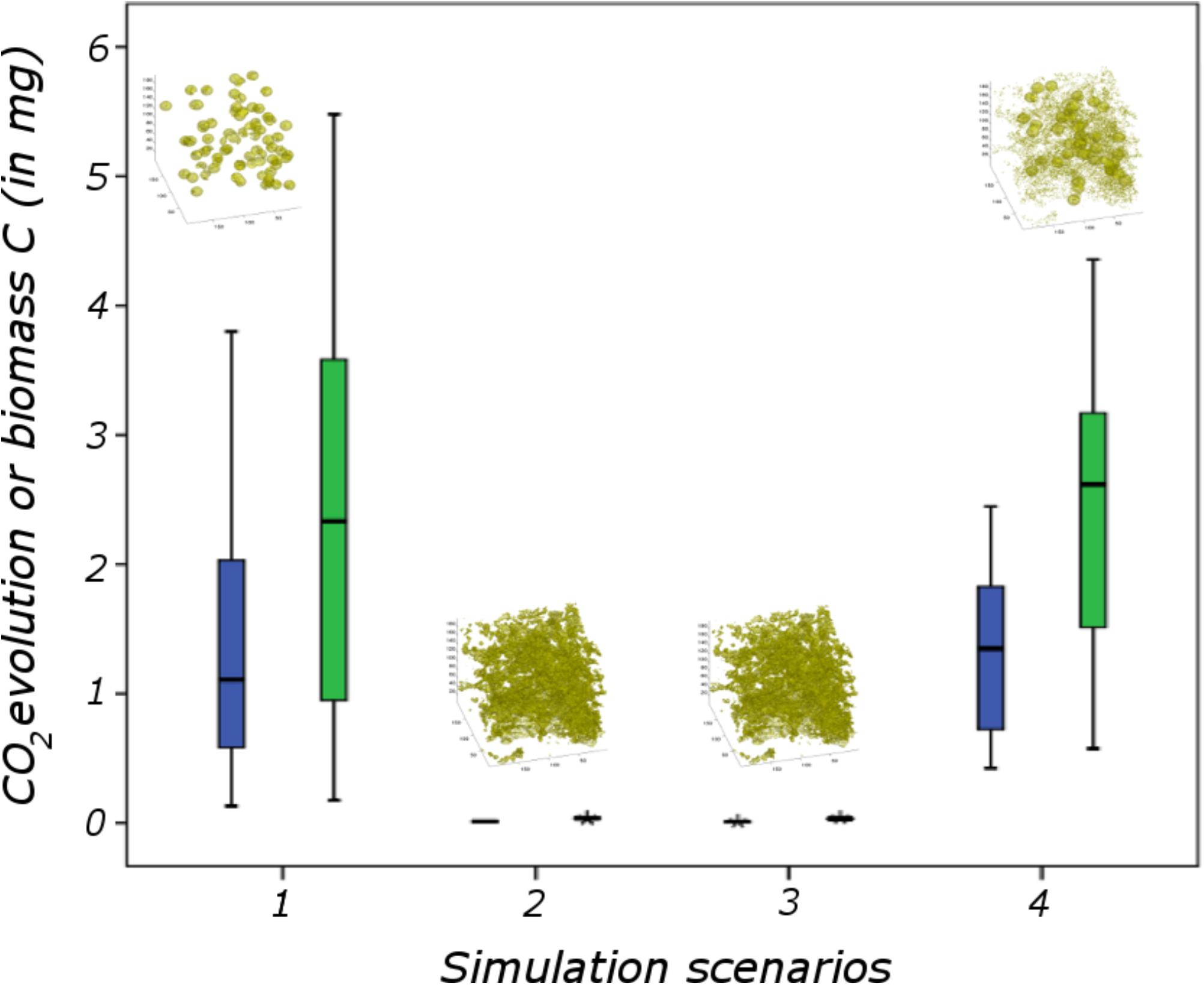
FIGURE 9. Boxplot diagram of simulated CO2 production by fungi in soil samples with an identical Particulate Organic Matter (POM) content of 3%. The abscissa represents different scenarios with, respectively, (from left to right) 49.95, 51.06, 56.73, and 60.12% of POM accessible at pore-solid interfaces. Individual samples differ in the way POM is distributed in the pore space (Modified from Falconer et al., 2015).
To summarize this inevitably quick overview of the work done so far on the integration of disciplinary perspectives on soils at the microscale (Figure 5), it seems fair to say that very little progress has been achieved, in large part because very few research projects have so far focused on integration. Such an integration is already largely feasible, for example by measuring in the same soil samples, both the characteristics of the pore space and the spatial heterogeneity of physico-chemical parameters, or either one of these parameters and the spatial distribution of microorganisms. And yet, at the time of the writing of this review article, very little integration at all has occurred at the experimental level. The tiny bit of work that has been carried out is encouraging, but a whole lot of research remains to be done. This is even more so in the case of upscaling, which remains a virtual terra incognita.
To achieve a high resolution when scanning soil samples, it is necessary to restrict the size of the samples. Typically, with table-top scanners, samples cannot have a volume larger than 100 cm3 to obtain micrometer resolution, and they have to be significantly less than that if one wants to reach the highest resolution (of about 0.3 μm) that is advertised by manufacturers. Meanwhile, the modeling of soil samples with lattice Boltzmann models is often limited at the moment to handling images of at most 500 × 500 × 500 voxels, which at a resolution of say, 20 μm, corresponds to a physical volume of only 1 cm3.
This small size of soil samples has occasionally raised questions in terms of the “representativeness” of measurements or simulations carried out with these samples (e.g., Al-Raoush and Willson, 2005; Costanza-Robinson et al., 2011; Gao et al., 2014; Rab et al., 2014). These questions stem from the concept of Representative Elementary Volume (REV), which has served over the last few decades as a conceptual foundation for much of the description of transport processes in soils (Fowler, 1997; Vogel and Ippisch, 2008; Hemes et al., 2015; Tracy et al., 2015; Daly et al., 2016; Liu et al., 2016; Cooper et al., 2017; Gonzalez et al., 2018). From that standpoint, soil samples scanned via X-ray CT or simulated via LB, which are of a much smaller size than the REV, would not be sufficiently representative, and one should therefore try to work with larger samples, be it physically, or virtually by aggregating together, mosaic-style, images obtained on a number of small juxtaposed samples. The difficulty with this approach is that in general one has no idea how big an REV is in any particular situation or whether an REV indeed exists (Baveye and Sposito, 1984, 1985; Vogel et al., 2002; Koestel, 2017). One can try computationally to consider sequentially larger volumes of soil in CT images to ascertain that a given property, e.g., porosity or bulk density, tends to become constant as the volume grows, as was done by various authors (Baveye et al., 2002; Vogel et al., 2002). However, there is no guarantee that the REV associated with a particular soil property applies to any other parameter of interest, so that the volume-growing procedure has to be repeated in principle for every single parameter needed to fully describe soil dynamics. Therefore, what at first appears to be a sound physically based constraint on the size of soil samples turns out often not to be operationally meaningful, and the best one can do, as suggested by Baveye and Sposito (1984), is to carefully reference any observation that is made on a soil sample to the volume and shape of this sample.
Nevertheless, regardless of how one feels concerning the need to invoke the notion of REV, it is clear that observations made on cm3-sized soil samples are not directly relevant to answering the questions raised by soil management, all of which relate to significantly larger spatial scales. Even the root zone of individual crop plants at maturation often encompasses several m3 (Baveye and Laba, 2015) and many societal concerns at the moment relate to the kilometric cells of typical climate and general circulation models used to predict global environmental change, or even to the much larger scales of watersheds and continents. Therefore, there is a definite need to upscale the observations made on small soil samples to the much larger scales at which answers are needed. In the words of Wachinger et al. (2000), “a path for translating small-scale understanding into large-scale phenomenology is required.” At the moment, no solution is available for this upscaling, which turns out to be an extremely challenging step, but different options have been suggested, some of which can be eliminated right off the bat.
One of the approaches that could be considered as an upscaling option consists of the process of mosaicking images obtained on small-sized soil samples, so as to obtain a virtual sample of much larger size. If this procedure results in soil samples of decimetric dimensions, one could be led to assume that the microscopic information and description relevant to the soil samples has been somehow “upscaled” to the macroscopic scale. However, even though the final sample considered may indeed be macroscopic in extent, the information one gets about it essentially remains microscopic in nature and does not necessarily provide the type of simplified description of reality that is sought in Figure 1 and that corresponds to the notion of emergence.
Another option, which has been used by numerous authors over the last two decades, consists of averaging microscopic descriptions of porous media, over either a Representative Elementary Volume or an arbitrary volume (e.g., associated with a measuring instrument), in order to obtain macroscopic variables (Ayub and Bentsen, 1999; Lichtner and Kang, 2007a,b; Golfier et al., 2009; Valdes-Parada et al., 2009; Wood, 2009, 2010; Davit et al., 2010; Baveye, 2013a; Lugo-Mendez et al., 2015). Davit et al. (2013) have shown that in terms of outcome, this approach is equivalent to another popular upscaling method involving homogenization through multiscale asymptotics (e.g., Roose et al., 2016). In the classical literature on scales in hydrology, both methods are closely associated with what is often referred to as “coarse-graining,” in analogy to a common practice in image analysis (Kitanidis, 2015). When applied to the type of soil processes we are interested in, the upshot of volume averaging, however, one looks at it, is a massive loss of information (Baveye, 2010), which takes us several steps backward in our understanding of emerging microbial processes. Indeed, if in a given soil, we perform a simple volume averaging of the concentration of a carbon source and, separately, of the biomass density, and if in so doing we ignore all the microscopic-scale information about the relative distributions of both, we are back to a situation we used to be in, with macroscopic parameters that have no causal relationship any more, and do not allow us to describe emerging processes accurately. Even if, as envisaged, e.g., by Wood (2010) and Porta et al. (2016), one goes beyond simple volume averaging, and somehow takes into account spatial fluctuations or variance within the volume in which averages are computed, leading to non-local integrodifferential equations, the results still miss some of the key ingredients that we recognize intuitively that an upscaled description of emerging microbial processes should have, in particular a quantification of the disconnect between microorganisms and their carbon/energy sources.
Occasionally, in discussions, the suggestion is made that the very popular “machine” or “deep” learning techniques (LeCun et al., 2015; Willcock et al., 2018) could perhaps provide a way to upscale microscale modeling of soils to the macroscopic scale (Veres et al., 2015). Machine learning explores the study and construction of algorithms that can learn from data and make data-driven predictions. Machine learning algorithms have started to be employed in soil science, in particular for pattern analysis and image classification to predict material classes in single channel X-ray CT images (Chauhan et al., 2016) and multi-channel nanoSIMS images (Steffens et al., 2017; Schweizer et al., 2018). Deep learning is a class of machine learning algorithms that use a cascade of multiple layers of non-linear processing units for feature extraction and transformation; learn in supervised and/or unsupervised (e.g., pattern analysis) manners; learn multiple levels of representations that correspond to different levels of abstraction; and use some form of gradient descent for training via back-propagation.
The application of machine or deep learning techniques to soils might consist of feeding a computer with detailed information about a multitude of scenarios, like those depicted in Figure 1, as well as results of simulations carried out for each scenario with the integrated model. With this supply of “big data,” deep learning algorithms would in principle search for patterns through all the simulations. Based on these patterns, it would then become feasible to predict the macroscopic behavior of a soil sample on the basis of microscopic data, without having to go through the likely time-consuming effort of re-running the integrated model. This type of outcome might conceivably be useful under specific circumstances, but it clearly does not correspond to what is expected of an upscaled model in Figure 1, namely the ability to predict the macroscopic behavior of soil samples based on macroscopic data. In other words, deep learning in itself does not automatically result in true upscaling. Nevertheless, deep learning algorithms might still be useful if somehow the patterns they identify in the data could (1) be revealed explicitly, (2) be related to specific macroscopic features of the soils, and (3) help in the development of appropriate macroscopic measurement techniques. At this point, further research is needed to determine whether any one of these different conditions can be met.
The research carried out to date, and in particular some of the scenario modeling alluded to earlier, point to a “disconnect” between microorganisms and their carbon/energy sources as being one of the keys to a proper understanding of emergent microbial processes in soils. In principle, this disconnect could be quantified in a number of ways. The Euclidean distance between microorganisms and OM might be a logical candidate, but it does not suffice, since closeness does not guarantee that OM be accessible either directly to a microorganism or indirectly to its extracellular enzymes (see illustration in Supplementary Figure S2). To convey the degree of direct or indirect accessibility of OM to microorganisms, a possibility is to consider the length of the most direct path through the pore space that connects a given bacterial cell or segment of fungal hyphae to a blob of OM, if pore connectivity allows such a path to exist at all. This shortest path, generally referred to as the “geodesic” distance, can be computed easily for individual pairs of points, using a number of algorithms developed in graph theory. In principle, a statistical mean of all relevant geodesic distances can then be generated within a specific soil sample. Within a range of CT image resolutions (which influence the apparent connectivity of soils, and therefore the calculation of shortest paths), the mean geodesic distance may prove to have merit, in particular if its use to characterize soil samples in investigations on the effect of temperature and precipitation on carbon mineralization manages to reduce the unexplained experimental variability observed so far.
Unfortunately, the geodesic distance in itself does not provide a complete answer. One issue with it is the fact that it does not take into account the geometry of the pore space along the shortest path, with which it is associated. In practice, this geometry matters tremendously. If a given geodesic path such as the one in Supplementary Figure S2 goes through a tiny constriction (which used to be referred to as a “pore neck”) between two adjacent voids in a soil, not only might bacterial cells or fungal hyphae have great difficulties passing through it, but chemical species (dissolved OM, exoenzymes, byproducts of enzymatic reactions) diffusing randomly through the pore space might also have a reduced likelihood of crossing over. From this perspective, instead of computing the geodesic distance, it might make more sense to quantify the average length of the path taken by molecules diffusing through the pore space. Even though conceptually, the geodesic and diffusion distances are very different, computationally they are not as distinct. Indeed, in order to compute the geodesic distance, algorithms typically track the diffusion paths of large number of random walkers, from which they eventually retain the shortest path. Therefore, the computation of an average diffusion distance between two points does not take a lot more time than the estimation of the geodesic distance.
Computer simulations, using the models under development at the moment and with a wide range of scenarios, could help determine under what conditions metrics like the mean geodesic distance, the mean diffusion distance, or some refinement of them, could be useful. The challenge at that point, then, will be to find a way to relate the metric that eventually turns out to be most suitable, to actual macroscopic measurements that make sense operationally. This is clearly a formidable challenge, whose practical importance cannot be downplayed. The end result of the program of Figure 1 absolutely cannot require extensive microbiological, chemical, and physical measurements at the microscopic scale. To be useful, the research needs to come up with simple measurement techniques, which can be used routinely, in a fully automated mode. It is far too early to have even a vague idea of what these routine measurements might be, but they need to remain front and center on our radar screen.
One way to perceive the overall message conveyed by the visual assessment of Figure 5 is that we are not very far along the way, and that a tremendous amount of work remains to be done. One could easily argue that this “half-empty glass” perspective is more than warranted. There is indeed a lot of work left, and a long way to go. From a more optimistic, “half-full glass” viewpoint, one could contend that, given the incredible complexity of soils and the fact that suitable technologies to deal with the various components of this complexity have been available for only a little over a decade, the progress achieved to date is remarkable.
Regardless of how one feels about the current state of affairs, it seems clear what the next steps in the research should be. The first step needs to address the clear imbalance that exists among the three core disciplines in the level of effort made to secure measurements in soils at the microscale. The current uneven level of knowledge, with some aspects of the research program that are far more advanced than others, if it is not alleviated in some way, is likely to dramatically hinder the credibility of any effort to make the basic disciplinary outlooks converge into a fully integrated microscopic model. At the moment, some integration of models has taken place, but one cannot actually assess how reliable the integrated descriptions are in practice, because in most situations, relevant microbiological observations are utterly lacking. Therefore, it seems fair to say that one of the key priorities of the research in this field will be to come up with the kind of microscale observations of the distribution and activity of microorganisms that are needed, whether that work be carried out by soil microbiologists or, as it has often happened in the last few decades, by non-microbiologists who have managed to gain the required expertise. When more precise information about the location and activity of microorganisms in soils becomes available, it will be useful to try to characterize as accurately as possible the physical and (bio)chemical nature of their microenvironments, and to determine how these microenvironments co-evolve with microorganisms over time.
A second step, which should be initiated now already, without waiting for the first step to be completed, consists of running multiple analyses on the same soil samples, in order to obtain an integrated view of the different parameters that control their functioning at the microscale. Some timid efforts have been made in this respect, but we have to shift to higher speed. In most cases, given the fact that the heavy equipment (e.g., scanners, NanoSIMS) used for some of these analyses are not located in the same institutions, this integration will require soil samples and possibly also researchers to travel from one institution to another. For some time, it has become well accepted that to run synchrotron-based analysis of soils (e.g., μXRF, XANES, or NEXAFS), one had to take soil samples to one of the handful of synchrotrons around the world. But now, this same attitude will have to be generalized to a much wider range of investigations, including microbiological analyses.
The next activity we should delve into at this point, much more forcefully than has been the case so far, is to use the existing microscale models of soils to run multiple “what-if” scenarios, and thereby try to understand how, for example, a spatial disconnect between microorganisms and the POM in soils affects the mineralization of this POM. Little by little, as more and more scenarios are run, it is likely that we will progressively get a sharper idea of the features that control the emergent properties of microbial activity in heterogeneous soil microenvironments, and eventually guide us in terms of the still somewhat fuzzy (but crucial) upscaling to the macroscopic scale.
These steps should keep us busy over the next 5 years. Besides funding, several factors will determine how fast we can make progress. In particular, much could depend on how quickly we can take advantage of a number of tremendous technological advances that should become readily available to researchers in the next few years.
As the preceding sections have documented in some detail, research on microscale aspects of emergent soil properties has been greatly stimulated by a number of major technological breakthroughs achieved at the turn of the century, especially in terms of X-ray CT but also with respect to other measurement techniques (e.g., CARD-FISH). The literature published in the last few years suggests that research is currently paving the way for another wave of phenomenal technological advances, which in several ways can be expected to be even more revolutionary than the previous one. Several of the new technologies are still at the development stage, such as zero-field nuclear magnetic resonance (Ledbetter and Budker, 2013) or quantum microscopes using molecular-scale MRI sensors built from diamonds (Reardon, 2017), but others have already become commercially available and could conceivably cause a huge leap in our ability to visualize and quantify processes in soils.
Years of efforts have been devoted to the development of near-synchrotron quality X-ray sources in facilities that are much smaller than the football stadium size of synchrotrons, and cost significantly less than the billions of euros a typical synchrotron does. Some of these efforts have resulted in 2015 in the installation in Munich (Germany) of the first commercially available mini particle accelerator, or “compact light source” (Eggl et al., 2016). With a very small 5 by 3 m footprint, it produces X-rays through Compton scattering, resulting from the interaction of low energy electrons and a high-powered laser pulse. The X-rays have high-brightness, intermediate between that of X-ray tube sources, used in table-top CT scanners, and large-scale synchrotrons. Another machine based on a similar principle, the ThomX compact light source is currently under completion at Orsay (France). It has an 18 m long storage ring, and will produce photons with energies up to 90 keV with a maximum flux of 1013 photons per second, i.e., with a brightness similar to that of synchrotrons. Undoubtedly, this type of machine will become widely available in years to come, and will eventually afford soil scientists far more access to nearly monochromatic, tunable X-ray beams than is currently the case.
X-ray beams produced using a very different approach may prove to be of even greater interest to soil scientists, because of the very small footprint and, potentially, cost, of the technology. The principle of laser-wakefield accelerators (LWFAs) was proposed more than three decades ago, and technological advances are progressively bringing them closer and closer to practical applications. In a LWFA, not much larger than a shoebox, where an intense laser pulse focused onto a plasma forms an electromagnetic wave in its wake, electrons can be trapped and are now routinely accelerated to GeV energies. Betatron motion, Compton scattering, and undulators produce tunable x-rays or gamma-rays by oscillating relativistic electrons in the wakefield behind the laser pulse, a counter-propagating laser field, or a magnetic undulator (Malka et al., 2008; Ben-Ismail et al., 2011; Mourou et al., 2013; Albert et al., 2014; Cole et al., 2015a,b; Albert and Thomas, 2016). LWFAs still need to be improved, and in particular their brightness needs to increase significantly to the level of synchrotron sources. Nevertheless, progress has been very rapid in recent years, so much so that a number of researchers have been able to use the technology for microtomography of bones, insects, and small mammals (Cole et al., 2015a,b; Wenz et al., 2015; Döpp et al., 2018). It may not be very long before LWFAs are commercialized and become viable options to image soils.
Perhaps as a result of the appearance of novel sources of X-rays, there has also been an upsurge of interest in developing a variety of novel X-ray detectors (Gruner, 2012), such as the photon-counting silicon-strip detector allowing energy-resolved CT (Persson et al., 2014). Various research groups have also been keen to look beyond simply taking advantage of X-ray attenuation to produce 3D images of materials, including soils. Phase-contrast is a very good candidate in this respect (Bhreasail et al., 2012). Techniques that have received attention recently are edge-illumination phase-contrast tomography (Zamir et al., 2017), dark-field scatter tomography (Bech et al., 2010) and ptychographic X-ray computed tomography. In the latter, phase-contrast information can be used to generate high-contrast 3D electron density maps without having to invoke the usual assumptions of a weak phase object or negligible adsorption (Chapman, 2010; Dierolf et al., 2010). To our knowledge, even though they have been mentioned in the literature on natural porous media, these techniques have not yet been applied to soil samples, but this is bound to happen in the not too distant future, at which point it will be possible to determine exactly how much promise they hold.
A possible danger with these tremendous technological advances that are now on the not-very-distant horizon is that they will allow major progress to be made chiefly in the quantification of the physical aspects of soils, thereby widening the already large gap that exists relative to the (bio)chemical and microbiological aspects. But that does not need to be the case. High-brightness, monochromatic, tunable X-ray and gamma-ray beams, possible with LWFAs, could prove extremely useful to visualize water and OM in soils. Also, it may turn out that some of the alternative X-ray techniques, like phase-contrast or dark-field imaging, may offer great advantages to visualize OM or even fungal hyphae in soils. Techniques like 3D micro-XRF or micro-XANES (Silversmit et al., 2010), whose application to characterize the (bio)chemical make-up of soils is currently handicapped by the limited access to synchrotron facilities, may also benefit greatly from the widespread availability of much cheaper, versatile X-ray sources.
Progress is being achieved not only in terms of X-ray or gamma-ray. One area where progress has been tremendous, and that, clearly, holds a lot of promise to assess the distribution of microorganisms in soil thin sections is related to fluorescence microscopy. Light microscopy, including fluorescence microscopy, has experienced phenomenal advances in the last decade. Until about 40 years ago (Cremer and Cremer, 1978), the resolution achievable with light microscopy was strictly constrained by the diffraction of light. Over time, increasing numbers of “super-resolution” microscopes have been developed, relying either on deterministic super-resolution techniques, like the stimulated emission depletion (STED) and saturated structured illumination microscopy (SSIM), or on stochastic functional techniques, like the super-resolution optical fluctuation imaging (SOFI) and the omnipresent localization microscopy (OLM) (Min et al., 2011; Cremer and Masters, 2013; Duwé and Dedecker, 2017; Ji, 2017; Power and Huisken, 2017; Yang and Yuste, 2017). Again, most of these techniques have yet to be applied to soil samples. With more and more of this super-resolution equipment becoming commercially available, there is little doubt that this application to soils will occur in the near future. When super-resolution images become available, we might be surprised (or not) to find out that the individual spots of light in images like those of Figure 8, which at the moment are identified as single bacterial or archaeal cells, are in fact small groups of cells.
In terms of the identification of microorganisms, significant progress has also been achieved recently, which could be very helpful in soils. In parallel with single-cell omics methods, a number of other techniques have been developed in the last few years, which allow the less-detailed, but much more rapid, characterization of single microbial cells. For example, Single Cell Raman Spectroscopy (SCRS) allows the direct measurement of intrinsic information about single cells in a non-invasive, label-free, and in vivo manner (Li et al., 2012; Smith et al., 2016). SCRS measures vibrations of biomolecules resulting from the inelastic scattering of incident laser light, producing a Raman spectrum, which is associated with a small physical volume (<1 μm3), of about the size of a bacterium. A typical single-cell Raman spectrum contains more than 1000 bands that can be assigned to different cellular compounds such as nucleic acids, protein, carbohydrates and lipids. With this information, SCRS enables the characterization of different cell types and can show physiological and phenotypic changes in living single cells. At its inception, the SCRS technique was afflicted by weak Raman signal and significant difficulty in the interpretation of the spectral data, however, recent work on multi-laser beams techniques like the Coherent anti-Stokes Raman Spectroscopy (CARS) (Min et al., 2011) and stimulated Raman spectroscopy (SRS) (Freudiger et al., 2011) has allowed a three order of magnitude increase in the strength of the signal, and significant improvements in signal interpretation. Any of these techniques could be routinely used to complement single-cell omics analysis. Prior to carrying out such an analysis on a particular microorganism in a soil thin section, one could ascertain whether its Raman spectrum is similar to one obtained for another organism already analyzed. In the affirmative, there may not be a need to perform a full single-cell omics protocol, resulting in considerable time saving.
The key take-home message of this article is visualized in Figure 5. It presents our assessment of progress achieved to date toward what we view as the ultimate objective of the research about emerging soil microbial processes, namely the development of macroscopic measurement techniques that would provide us with the information needed to make reasonably accurate predictions. This Figure 5 contains good news and bad news. The good news is that we have made significant progress. For forty years after prominent microbiologists argued in the mid 1960s that the quantitative microscale description of soil microbial processes was essential, the lack of suitable measurement techniques prevented the research from advancing at all. As a result, in spite of the publication of numerous articles on soil OM and on microbial processes in soils, very little progress has been achieved since the 1960s on several key questions in these areas. In the past 15 years, major technological breakthroughs have changed all that, with the result that our understanding of soils at the microscale has improved significantly on a number of fronts, experimentally as well as in terms of computer modeling. The bad news is that progress is very uneven. At the extremes of the spectrum, whereas research on the physical characteristics of soils at the microscale is moving full speed ahead, the (arguably more complicated) experimental observation of microbial processes is lagging far behind, casting doubt on the soundness on some of the extensive modeling that has been carried out in this field over the last decade, and hindering the needed integration of physical, (bio)chemical, and microbiological perspectives. Clearly, there is still a long way before reaching the holy grail, with many daunting challenges on the different paths leading to it.
There are reasons to be optimistic, however, and not to be intimidated by these challenges. For one thing, technological breakthroughs did not stop a decade ago. New measuring devices and new technologies in other respects as well (e.g., single-cell “omics”) are being developed and, for some of them, even getting commercialized, which should lead to many quantum leaps in our ability to carry out microscale measurements in soils. In addition, one can always hope that as we run more and more experiments to try to understand the emergent microbial properties of soils, someone will come up with an empirical equation that will provide a simple answer to all the questions we have at the moment, a little bit like what Henry Darcy did in his day for water movement in sand filters. Such an empirical description, if and when it becomes available, would completely change the game plan. But in the meantime, we need to keep in mind that we do not really have a choice but to move forward, no matter how challenging that might be. As was mentioned at the beginning of this article, the unresolved questions the research addresses are the object of extreme societal concern and it is not overblown to consider that they need to be answered urgently if we ultimately want humanity to survive. This message is not yet understood by decision makers in most countries, but as time goes by and it becomes more and more urgent to get answers, we should hope that even politicians will realize it will be in everyone’s best interest to devote to this research more than the shoe-string budgets that have been allocated to it so far.
No new data were generated during the writing of this article.
PCB initiated the writing of this article, based on in-depth conversations he had over an extended period of time with all co-authors, and he revised several successive drafts of the text, after receiving substantial input from everyone (and very helpful suggestions from the associate editor and reviewers). All authors have approved publication of the final version.
The research described in this article was made possible in part through NPRP grant #9-390-1-088 from the Qatar National Research Fund (Project Simupor), as well as through a grant from the Agence Nationale de la Recherche (ANR, France) to project Soilμ3D. WO acknowledges funding from the National Environment and Research Council (NE/P014208/1). MB-R’s contribution was made possible by a postdoctoral fellowship (Programa de axudas á etapa posdoutoral; ED481B 2017/073) awarded by the Consellería de Cultura, Educación e Ordenación Universitaria (Xunta de Galicia, Spain).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer PN and handling Editor declared their shared affiliation.
TE acknowledges gratefully the financial support received from the German Research Foundation (DFG; ZK/14/M8-Plus) and German Academic Research Service (DAAD; K/15/464342). Finally, PCB, SS, TE, and CM acknowledge the financial support by the German Soil Science Society (DBG) for the workshop “Combined analysis of biochemistry and structural complexity of soil via imaging”, held at the UFZ in July 2016 and during which many aspects of the topic reviewed in this article were discussed.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.01929/full#supplementary-material
Abrudan, M. I., Smakman, F., Grimbergen, A. J., Westhoff, S., Miller, E. L., van Wezel, G. P., et al. (2015). Socially mediated induction and suppression of antibiosis during bacterial coexistence. Proc. Natl. Acad. Sci. U.S.A. 112, 11054–11059. doi: 10.1073/pnas.1504076112
Addiscott, T. M. (2011). Emergence or self-organization? Look to the soil population. Commun. Integr. Biol. 4, 469–470. doi: 10.4161/cib.15547
Albert, F., and Thomas, A. G. R. (2016). Applications of laser wakefield accelerator-based light sources. Plasma Phys. Control. Fusion 58. doi: 10.1088/0741-3335/58/10/103001
Albert, F., Thomas, A. G. R., Mangles, S. P. D., Banerjee, S., Corde, S., Flacco, A., et al. (2014). Laser wakefield accelerator based light sources: potential applications and requirements. Plasma Phys. Control. Fusion 56:084015. doi: 10.1088/0741-3335/56/8/084015
Aleklett, K., Kiers, E. T., Ohlsson, P., Shimizu, T. S., Caldas, V. E. A., and Hammer, E. C. (2018). Build your own soil: exploring microfluidics to create microbial habitat structures. ISME J. 12, 312–319. doi: 10.1038/ismej.2017.184
Alexander, F. E. S., and Jackson, R. M. (1954). Examination of soil micro-organisms in their natural environment. Nature 174, 750–751. doi: 10.1038/174750b0
Alexander, F. E. S., and Jackson, R. M. (1955). “Preparation of sections for study of soil microorganisms,” in Proceedings of the University of Nottingham Second Easter School in Agriculture Science: Soil Zoology, ed. D. K. Kevan (London: Butterworths), 433–441.
Alexander, M. (1964). Biochemical ecology of soil microorganisms. Annu. Rev. Microbiol. 18, 217–250. doi: 10.1146/annurev.mi.18.100164.001245
Allison, S. D., and Martiny, J. B. H. (2008). Resistance, resilience, and redundancy in microbial communities. Proc. Natl. Acad. Sci. U.S.A. 105, 11512–11519. doi: 10.1073/pnas.0801925105
Al-Raoush, R. I., and Willson, C. S. (2005). A pore-scale investigation of a multiphase porous media system. J. Contam. Hydrol. 77, 67–89. doi: 10.1016/j.jconhyd.2004.12.001
Altman, S. J., Rivers, M. L., Reno, M. D., Cygan, R. T., and McLain, A. A. (2005). Characterization of adsorption sites on aggregate soil samples using synchrotron X-ray computerized microtomography. Environ. Sci. Technol. 39, 2679–2685. doi: 10.1021/es049103y
Amézketa, E. (1999). Soil aggregate stability: a review. J. Sustain. Agri. 14, 83–151. doi: 10.1300/J064v14n02_08
Amin, M. H., Hall, L. D., Chorley, R. J., Carpenter, T. A., Richards, K. S., and Bache, B. W. (1994). Magnetic resonance imaging of soil-water phenomena. Magn. Reson. Imaging 12, 319–321. doi: 10.1016/0730-725X(94)91546-6
Amin, M. H., Richards, K. S., Chorley, R. J., Gibbs, S. J., Carpenter, T. A., and Hall, L. D. (1996). Studies of soil-water transport by MRI. Magn. Reson. Imaging 14, 879–882. doi: 10.1016/S0730-725X(96)00171-3
Ananyeva, K., Wang, W., Smucker, A. J. M., Rivers, M. L., and Kravchenko, A. N. (2013). Can intra-aggregate pore structures affect the aggregate’s effectiveness in protecting carbon? Soil Biol. Biochem. 57, 868–875. doi: 10.1016/j.soilbio.2012.10.019
Anderson, S. E., and Hopmans, J. W. eds (1994). Tomography of Soil Water Root Processes, Vol. 36. Madison, WI: Soil Science Society of America.
Apourvari, S. N., and Arns, C. H. (2016). Image-based relative permeability upscaling from the pore scale. Adv. Water Resour. 95, 161–175. doi: 10.1016/j.advwatres.2015.11.005
Arsjad, S., and Giddens, J. (1966). Effect of added plant tissue on decomposition of soil organic matter under different wetting and drying cycles. Soil Sci. Soc. Am. Proc. 30, 457. doi: 10.2136/sssaj1966.03615995003000040018x
Ashelford, K. E., Day, M. J., and Fry, J. C. (2003). Elevated abundance of bacteriophage infecting bacteria in soil. Appl. Environ. Microbiol. 69, 285–289. doi: 10.1128/AEM.69.1.285-289.2003
Ashida, N., Ishii, S., Hayano, S., Tago, K., Tsuji, T., Yoshimura, Y., et al. (2010). Isolation of functional single cells from environments using a micromanipulator: application to study denitrifying bacteria. Appl. Microbiol. Biotechnol. 85, 1211–1217. doi: 10.1007/s00253-009-2330-z
Attard, E., Recous, S., Chabbi, A., De Berranger, C., Guillaumaud, N., Labreuche, J., et al. (2011). Soil environmental conditions rather than denitrifier abundance and diversity drive potential denitrification after changes in land uses. Global Change Biol. 17, 1975–1989. doi: 10.1111/j.1365-2486.2010.02340.x
Ayub, M., and Bentsen, R. G. (1999). Interfacial viscous coupling: a myth or reality? J. Petrol. Sci. Eng. 23, 13–26. doi: 10.1016/S0920-4105(99)00003-0
Badorreck, A., Gerke, H. H., and Vontobel, P. (2010). Noninvasive observations of flow patterns in locally heterogeneous mine soils using neutron radiation. Vadose Zone J. 9, 362–372. doi: 10.2136/vzj2009.0100
Bailey, V. L., Smith, A. P., Tfaily, M., Fansler, S. J., and Bond-Lamberty, B. (2017). Differences in soluble organic carbon chemistry in pore waters sampled from different pore size domains. Soil Biol. Biochem. 107, 133–143. doi: 10.1016/j.soilbio.2016.11.025
Barcenas-Moreno, G., Baath, E., and Rousk, J. (2016). Functional implications of the pH-trait distribution of the microbial community in a re-inoculation experiment across a pH gradient. Soil Biol. Biochem. 93, 69–78. doi: 10.1016/j.soilbio.2015.10.024
Barot, S., Rossi, J. P., and Lavelle, P. (2007). Self-organization in a simple consumer-resource system, the example of earthworms. Soil Biol. Biochem. 39, 2230–2240. doi: 10.1016/j.soilbio.2007.03.021
Basu, N. B., Thompson Sally, E., and Rao, P. S. C. (2011). Hydrologic and biogeochemical functioning of intensively managed catchments: a synthesis of top-down analyses. Water Resour. Res. 47:W00J15. doi: 10.1029/2011WR010800
Battiato, I., Tartakovsky, D. M., Tartakovsky, A. M., and Scheibe, T. D. (2011). Hybrid models of reactive transport in porous and fractured media. Adv. Water Resour. 37, 1140–1150. doi: 10.1016/j.advwatres.2011.01.012
Baveye, P. (2006). Comment on “Soil structure and management: a review” by C.J. Bronick R. Lal. Geoderma 134, 231–232. doi: 10.1016/j.geoderma.2005.10.003
Baveye, P. (2010). Comment on “the role of scaling laws in upscaling” by BD Wood. Adv. Water Resour. 33, 123–124. doi: 10.1016/j.advwatres.2009.11.003
Baveye, P., Rogasik, H., Wendroth, O., Onasch, I., and Crawford, J. W. (2002). Effect of sampling volume on the measurement of soil physical properties: simulation with x-ray tomography data. Measure. Sci. Technol. 13, 775–784. doi: 10.1088/0957-0233/13/5/316
Baveye, P., and Sposito, G. (1984). The operational significance of the continuum-hypothesis in the theory of water-movement through soils and aquifers. Water Resour. Res. 20, 521–530. doi: 10.1029/WR020i005p00521
Baveye, P., and Sposito, G. (1985). Macroscopic balance-equations in soils and aquifers – The case of space-dependent and time-dependent instrumental response. Water Resour. Res. 21, 1116–1120. doi: 10.1029/WR021i008p01116
Baveye, P., and Valocchi, A. (1989). An evaluation of mathematical models of the transport of biologically reacting solutes in saturated soils and aquifers. Water Resour. Res. 25, 1413–1421. doi: 10.1029/WR025i006p01413
Baveye, P., Vandevivere, P., Hoyle, B. L., DeLeo, P. C., and de Lozada, D. S. (1998). Environmental impact and mechanisms of the biological clogging of saturated soils and aquifer materials. Crit. Rev. Environ. Sci. Technol. 28, 123–191. doi: 10.1080/10643389891254197
Baveye, P. C. (2007). Soils and runaway global warming: terra incognita. J. Soil Water Conserv. 62, 139A–143A.
Baveye, P. C. (2009a). Comment on “Conservation of protists: Is it needed at all?” by Cotterill et al. Biodiv. Conserv. 18, 503–505. doi: 10.1007/s10531-008-9525-y
Baveye, P. C. (2009b). To sequence or not to sequence the whole-soil metagenome? Nat. Rev. Microbiol. 7:756. doi: 10.1038/nrmicro2119-c2
Baveye, P. C. (2013a). Comment on “Averaging theory for description of environmental problems: what have we learned?” by William G. Gray, Cass T. Miller, and Bernhard A. Schrefler. Adv. Water Resour. 52, 328–330. doi: 10.1016/j.advwatres.2012.08.012
Baveye, P. C. (2013b). Addressing key challenges to interdisciplinary research on water-related issues: biologists’ engagement and funding structure. Biologia 68, 1087–1088. doi: 10.2478/s11756-013-0280-5
Baveye, P. C. (2013c). Hydrology and the looming water crisis: It is time to think, and act, outside the box. J. Hydrol. Hydromech. 61, 89–96. doi: 10.2478/johh-2013-0012
Baveye, P. C. (2014). Learned publishing: who still has time to read? Learn. Publish. 27, 48–51. doi: 10.1087/20140107
Baveye, P. C. (2015). Grand challenges in the research on soil processes. Front. Environ. Sci. 3:10. doi: 10.3389/fenvs.2015.00010
Baveye, P. C. (2018). Book review: shifting paradigms on soil microbial biomass. Front. Environ. Sci. 6:10. doi: 10.3389/fenvs.2018.00010
Baveye, P. C., Baveye, J., and Gowdy, J. (2016a). Soil “ecosystem” services and natural capital: critical appraisal of research on uncertain ground. Front. Environ. Sci. 4:41. doi: 10.3389/fenvs.2016.00041
Baveye, P. C., Berthelin, J., and Munch, J.-C. (2016b). Too much or not enough: reflection on two contrasting perspectives on soil biodiversity. Soil Biol. Biochem. 103, 320–326. doi: 10.1016/j.soilbio.2016.09.008
Baveye, P. C., Berthelin, J., Tessier, D., and Lemaire, G. (2018). The “4 per 1000” initiative: A credibility issue for the soil science community? Geoderma 309, 118–123. doi: 10.1016/j.geoderma.2017.05.005
Baveye, P. C., and Laba, M. (2015). Moving away from the geostatistical lamppost: why, where, and how does the spatial heterogeneity of soils matter? Ecol. Modell. 298, 24–38. doi: 10.1016/j.ecolmodel.2014.03.018
Baveye, P. C., Laba, M., Otten, W., Bouckaert, L., Dello Sterpaio, P., Goswami, R. R., et al. (2010). Observer-dependent variability of the thresholding step in the quantitative analysis of soil images and X-ray microtomography data. Geoderma 157, 51–63. doi: 10.1016/j.geoderma.2010.03.015
Baveye, P. C., Palfreyman, J., and Otten, W. (2014). Research efforts involving several disciplines: adherence to a clear nomenclature Is needed. Water Air Soil Pollut. 225:1997. doi: 10.1007/s11270-014-1997-7
Baveye, P. C., Pot, V., and Garnier, P. (2017). Accounting for sub-resolution pores in models of water and solute transport in soils based on computed tomography images: are we there yet? J. Hydrol. 555, 253–256. doi: 10.1016/j.jhydrol.2017.10.021
Baveye, P. C., Rangel, D., Jacobson, A. R., Laba, M., Darnault, C., Otten, W., et al. (2011). From dust bowl to dust bowl: soils are still very much a frontier of science. Soil Sci. Soc. Am. J. 75, 2037–2048. doi: 10.2136/sssaj2011.0145
Bearden, J. A., and Burr, A. F. (1967). Reevaluation of X-ray atomic energy levels. Rev. Mod. Phys. 39, 125–142. doi: 10.1103/RevModPhys.39.125
Bech, M., Bunk, O., Donath, T., Feidenhans’l, R., David, C., and Pfeiffer, F. (2010). Quantitative x-ray dark-field computed tomography. Phys. Med. Biol. 55, 5529–5539. doi: 10.1088/0031-9155/55/18/017
Beckers, E., Plougonven, E., Gigot, N., Leonard, A., Roisin, C., Brostaux, Y., et al. (2014a). Coupling X-ray microtomography and macroscopic soil measurements: a method to enhance near-saturation functions? Hydrol. Earth Syst. Sci. 18, 1805–1817. doi: 10.5194/hess-18-1805-2014
Beckers, E., Plougonven, E., Roisin, C., Hapca, S., Leonard, A., and Degre, A. (2014b). X-ray microtomography: a porosity-based thresholding method to improve soil pore network characterization? Geoderma 219, 145–154. doi: 10.1016/j.geoderma.2014.01.004
Behrens, S., Kappler, A., and Obst, M. (2012). Linking environmental processes to the in situ functioning of microorganisms by high-resolution secondary ion mass spectrometry (NanoSIMS) and scanning transmission X-ray microscopy (STXM). Environ. Microbiol. 14, 2851–2869. doi: 10.1111/j.1462-2920.2012.02724.x
Ben-Ismail, A., Lundh, O., Rechatin, C., Lim, J. K., Faure, J., Corde, S., et al. (2011). Compact and high-quality gamma-ray source applied to 10 mu m-range resolution radiography. Appl. Phys. Lett. 98:264101. doi: 10.1063/1.3604013
Berg, S., Ott, H., Klapp, S. A., Schwing, A., Neiteler, R., Brussee, N., et al. (2013). Real-time 3D imaging of haines jumps in porous media flow. Proc. Natl. Acad. Sci. U.S.A. 110, 3755–3759. doi: 10.1073/pnas.1221373110
Berthold, T., Centler, F., Huebschmann, T., Remer, R., Thullner, M., Harms, H., et al. (2016). Mycelia as a focal point for horizontal gene transfer among soil bacteria. Sci. Rep. 6:36390. doi: 10.1038/srep36390
Bhreasail, A. N., Lee, P. D., O’Sullivan, C., Fenton, C. H., Hamilton, R., Rockett, P., et al. (2012). In-Situ observation of cracks in frozen soil using synchrotron tomography. Permafrost Periglacial Process. 23, 170–176. doi: 10.1002/ppp.1737
Biassusi, M., Pauletto, E. A., and Crestana, S. (1999). Estudo da deformação de um vertissolo por meio da tomografia computadorizada de dupla energia simultânea. Rev. Brasi. Ciência Solo 23, 1–7. doi: 10.1590/S0100-06831999000100001
Blagodatsky, S., and Smith, P. (2012). Soil physics meets soil biology: towards better mechanistic prediction of greenhouse gas emissions from soil. Soil Biol. Biochem. 47, 78–92. doi: 10.1016/j.soilbio.2011.12.015
Blainey, P. C. (2013). The future is now: single-cell genomics of bacteria and archaea. FEMS Microbiol. Rev. 37, 407–427. doi: 10.1111/1574-6976.12015
Blaser, M. J., Cardon, Z. G., Cho, M. K., Dangl, J. L., Donohue, T. J., Green, J. L., et al. (2016). Toward a predictive understanding of earth’s microbiomes to address 21st century challenges. MBio 7:e00714-16. doi: 10.1128/mBio.00714-16
Blaud, A., Chevallier, T., Virto, I., Pablo, A.-L., Chenu, C., and Brauman, A. (2014). Bacterial community structure in soil microaggregates and on particulate organic matter fractions located outside or inside soil macroaggregates. Pedobiologia 57, 191–194. doi: 10.1016/j.pedobi.2014.03.005
Blazewicz, S. J., Barnard, R. L., Daly, R. A., and Firestone, M. K. (2013). Evaluating rRNA as an indicator of microbial activity in environmental communities: limitations and uses. ISME J. 7, 2061–2068. doi: 10.1038/ismej.2013.102
Bleuet, P., Gergaud, P., Lemelle, L., Tucoulou, R., Cloetens, P., Susini, J., et al. (2010). 3D chemical imaging based on a third-generation synchrotron source. TrAC Trends Anal. Chem. 29, 518–527. doi: 10.1016/j.trac.2010.02.011
Bocking, C. R., and Blyth, M. G. (2018). Oxygen uptake and denitrification in soil aggregates. Acta Mech. 229, 595–612. doi: 10.1007/s00707-017-2042-x
Borer, B., Tecon, R., and Or, D. (2018). Spatial organization of bacterial populations in response to oxygen and carbon counter-gradients in pore networks. Nat. Commun. 9:769. doi: 10.1038/s41467-018-03187-y
Boswell, G. P., and Hopkins, S. (2008). Linking hyphal growth to colony dynamics: spatially explicit models of mycelia. Fung. Ecol. 1, 143–154. doi: 10.1016/j.funeco.2008.10.003
Bottinelli, N., Zhou, H., Boivin, P., Zhang, Z. B., Jouquet, P., Hartmann, C., et al. (2016). Macropores generated during shrinkage in two paddy soils using X-ray micro-computed tomography. Geoderma 265, 78–86. doi: 10.1016/j.geoderma.2015.11.011
Bouckaert, L., Sleutel, S., Van Loo, D., Brabant, L., Cnudde, V., Van Hoorebeke, L., et al. (2013a). Carbon mineralisation and pore size classes in undisturbed soil cores. Soil Res. 51, 14–22. doi: 10.1071/sr12116
Bouckaert, L., Van Loo, D., Ameloot, N., Buchan, D., Van Hoorebeke, L., and Sleutel, S. (2013b). Compatibility of X-ray micro-Computed tomography with soil biological experiments. Soil Biol. Biochem. 56, 10–12. doi: 10.1016/j.soilbio.2012.02.002
Brackin, R., Atkinson, B. S., Sturrock, C. J., and Rasmussen, A. (2017). Roots-eye view: using microdialysis and microCT to non-destructively map root nutrient depletion and accumulation zones. Plant Cell Environ. 40, 3135–3142. doi: 10.1111/pce.13072
Briones, M. J. I. (2014). Soil fauna and soil functions: a jigsaw puzzle. Front. Environ. Sci. 2:7. doi: 10.3389/fenvs.2014.00007
Brusseau, M. L., Peng, S., Snaar, G., and Murao, A. (2007). Measuring air-water interfacial areas with X-ray microtomography and interfacial partitioning tracer tests. Environ. Sci. Technol. 41, 1956–1961. doi: 10.1021/es061474m
Bultreys, T., De Boever, W., and Cnudde, V. (2016). Imaging and image-based fluid transport modeling at the pore scale in geological materials: a practical introduction to the current state-of-the-art. Earth-Sci. Rev. 155, 93–128. doi: 10.1016/j.earscirev.2016.02.001
Caldwell, D. E., Korber, D. R., and Lawrence, J. R. (1992). Imaging of bacterial cells by fluorescence exclusion using scanning confocal laser microscopy. J. Microb. Methods 15, 249–261. doi: 10.1016/0167-7012(92)90045-6
Calistru, A.-E., and Jitareanu, G. (2015). Applications of X-ray computed tomography for examining soil structure: a review. Bull. UASVM Agric. 72, 31–36. doi: 10.15835/buasvmcn-agr:11145
Cantoni, M., and Holzer, L. (2014). Advances in 3D focused ion beam tomography. MRS Bull. 39, 354–360. doi: 10.1557/mrs.2014.54
Capowiez, Y., Pierret, A., Daniel, O., Monestiez, P., and Kretschmar, A. (1998). 3D skeleton reconstructions of natural earthworm burrow systems using CAT scan images of soil cores. Biol. Fertil. Soils 27, 51–59. doi: 10.1007/s003740050399
Carini, P., Marsden, P. J., Leff, J. W., Morgan, E. E., Strickland, M. S., and Fierer, N. (2016). Relic DNA is abundant in soil and obscures estimates of soil microbial diversity. Nat. Microbiol. 2:16242. doi: 10.1038/nmicrobiol.2016.242
Carminati, A., Kaestner, A., Lehmann, P., and Flühler, H. (2008). Unsaturated water flow across soil aggregate contacts. Adv. Water Resour. 31, 1221–1232. doi: 10.1016/j.advwatres.2008.01.008
Carrel, M., Beltran, M. A., Morales, V. L., Derlon, N., Morgenroth, E., and Kaufmann, R. (2017). Biofilm imaging in porous media by laboratory X-Ray tomography: combining a non-destructive contrast agent with propagation-based phase-contrast imaging tools. PLoS One 12:e0180374. doi: 10.1371/journal.pone.0180374
Carson, J. K., Gonzalez-Quinones, V., Murphy, D. V., Hinz, C., Shaw, J. A., and Gleeson, D. B. (2010). Low pore connectivity increases bacterial diversity in soil. Appl. Environ. Microbiol. 76, 3936–3942. doi: 10.1128/aem.03085-09
Cazelles, K., Otten, W., Baveye, P. C., and Falconer, R. E. (2013). Soil fungal dynamics: parameterisation and sensitivity analysis of modelled physiological processes, soil architecture and carbon distribution. Ecol. Modell. 248, 165–173. doi: 10.1016/j.ecolmodel.2012.08.008
Cerqueira, B., Arenas-Lago, D., Andrade, M. L., and Vega, F. A. (2015). Using time of flight secondary ion mass spectrometry and field emission scanning electron microscopy with energy dispersive X-ray spectroscopy to determine the role of soil components in competitive copper and cadmium migration and fixation in soils. Geoderma 25, 65–77. doi: 10.1016/j.geoderma.2015.03.026
Chapman, N., Miller, A. J., Lindsey, K., and Whalley, W. R. (2012). Roots, water, and nutrient acquisition: let’s get physical. Trends Plant Sci. 17, 701–710. doi: 10.1016/j.tplants.2012.08.001
Charles, D. (2018). Scientists Peek Inside the ‘Black Box’ of Soil Microbes to Learn Their Secrets. Available at: https://www.npr.org/sections/thesalt/2018/01/18/578924748/scientists-peek-inside-the-black-box-of-soil-microbes-to-learn-their-secrets
Chauhan, S., Ruehaak, W., Anbergen, H., Kabdenov, A., Freise, M., Wille, T., et al. (2016). Phase segmentation of X-ray computer tomography rock images using machine learning techniques: an accuracy and performance study. Solid Earth 7, 1125–1139. doi: 10.5194/se-7-1125-2016
Chen, C., Geer, J., and Sammakia, B. (2014). Sub-continuum thermal transport modeling using diffusion in the lattice boltzmann transport equation. Int. J. Heat Mass Trans. 79, 666–675. doi: 10.1016/j.ijheatmasstransfer.2014.08.052
Chen, Y., and Zhu, K. (2008). A study of the upper limit of solid scatters density for gray lattice boltzmann method. Acta Mech. Sin. 24, 515–522. doi: 10.1007/s10409-008-0167-9
Cheng, L., Booker, F. L., Tu, C., Burkey, K. O., Zhou, L., Shew, H. D., et al. (2012). Arbuscular mycorrhizal fungiincrease organic carbon decomposition under elevated CO2. Science 337, 1084–1087. doi: 10.1126/science.1224304
Chenu, C., Hassink, J., and Bloem, J. (2001). Short-term changes in the spatial distribution of microorganisms in soil aggregates as affected by glucose addition. Soil Biol. Biochem. 34, 349–356.
Chenu, C., and Stotzky, G. (2001). “Interactions between microorganisms and soil particles: an overview,” in Interactions between Soil Particles and Microorganisms: Impact on the Terrestrial Ecosystem, eds P. M. Huang, J. M. Bollag, and N. Senesi (London: Wiley), 3–40.
Cheshire, M. V. (1977). Origins and stability of soil polysaccharide. J. Soil Sci. 28, 1–10. doi: 10.1111/j.1365-2389.1977.tb02290.x
Chivenge, P., Vanlauwe, B., Gentile, R., and Six, J. (2011). Comparison of organic versus mineral resource effects on short-term aggregate carbon and nitrogen dynamics in a sandy soil versus a fine textured soil. Agric. Ecosyst. Environ. 140, 361–371. doi: 10.1016/j.agee.2010.12.004
Christl, I., and Kretzschmar, R. (2007). C-1s NEXAFS spectroscopy reveals chemical fractionation of humic acid by cation-induced coagulation. Environ. Sci. Technol. 41, 1915–1920. doi: 10.1021/es062141s
Churchman, G. J., and Payne, D. (1983). Mercury intrusion porosimetry of some New Zealand soils in relation to clay mineralogy and texture. J. Soil Sci. 34, 437–451. doi: 10.1111/j.1365-2389.1983.tb01047.x
Cole, J. M., Wood, J. C., Lopes, N. C., Poder, K., Abel, R. L., Alatabi, S., et al. (2015a). Laser-wakefield accelerators as hard x-ray sources for 3D medical imaging of human bone. Sci. Rep. 5:13244. doi: 10.1038/srep13244
Cole, J. M., Wood, J. C., Lopes, N. C., Poder, K., Abel, R. L., Alatabi, S., et al. (2015b). Tomography of human trabecular bone with a laser-wakefield driven x-ray source. Plasma Phys. Control. Fusion 58:14008. doi: 10.1088/0741-3335/58/1/014008
Cooper, L. J., Daly, K. R., Hallett, P. D., Naveed, M., Koebernick, N., Bengough, A. G., et al. (2017). Fluid flow in porous media using image-based modelling to parametrize Richards’ equation. Proc. R. Soc. Lond. Ser. A Mathemat. Phys. Eng. Sci. 473:20170178. doi: 10.1098/rspa.2017.0178
Costanza-Robinson, M. S., Estabrook, B. D., and Fouhey, D. F. (2011). Representative elementary volume estimation for porosity, moisture saturation, and air-water interfacial areas in unsaturated porous media: data quality implications. Water Resour. Res. 47, 1–12. doi: 10.1029/2010WR009655
Cousin, I., Levitz, P., and Bruand, A. (1996). Three-dimensional analysis of a loamy-clay soil using pore and solid chord distributions. Eur. J. Soil Sci. 47, 439–452. doi: 10.1111/j.1365-2389.1996.tb01844.x
Cousin, I., Porion, P., Renault, P., and Levitz, P. (1999). Gas diffusion in a silty-clay soil: experimental study on an undisturbed soil core and simulation in its three-dimensional reconstruction. Eur. J. Soil Sci. 50, 249–259. doi: 10.1046/j.1365-2389.1999.00215.x
Coyte, K. Z., Tabuteau, H., Gaffney, E. A., Foster, K. R., and Durham, W. M. (2017). Microbial competition in porous environments can select against rapid biofilm growth. Proc. Natl. Acad. Sci. U.S.A. 114, E161–E170. doi: 10.1073/pnas.1525228113
Cremer, C., and Cremer, T. (1978). Considerations on a laser-scanning-microscope with high resolution and depth of field. Micros. Acta 81, 31–44.
Cremer, C., and Masters, B. R. (2013). Resolution enhancement techniques in microscopy. Eur. Phys. J. H 38, 281–344. doi: 10.1140/epjh/e2012-20060-1
Crenshaw, C. L., Lauber, C., Sinsabaugh, R. L., and Stavely, L. K. (2008). Fungal control of nitrous oxide production in semiarid grassland. Biogeochemistry 87, 17–27. doi: 10.1007/s10533-007-9165-4
Crestana, S., Mascarenhas, S., and Pozzimucelli, R. S. (1985). Static and dynamic 3-dimensional studies of water in soil using computed tomography scanning. Soil Sci. 140, 326–332. doi: 10.1097/00010694-198511000-00002
Crowther, T. W., Todd-Brown, K. E. O., Rowe, C. W., Wieder, W. R., Carey, J. C., Machmuller, M. B., et al. (2016). Quantifying global soil carbon losses in response to warming. Nature 540, 104–108. doi: 10.1038/nature20150
Crozat, Y., Cleyet-Marel, J. C., and Corman, A. (1987). Use of the fluorescentantibody technique to characterize equilibrium survival concentrations of Bradyrhizobium japonicum strains in soil. Biol. Fertil. Soils 4, 85–90.
Cruz, B. C., Furrer, J. M., Guo, Y. S., Dougherty, D., Hinestroza, H. F., Hernandez, J. S., et al. (2017). Pore-scale water dynamics during drying and the impacts of structure and surface wettability. Water Resour. Res. 53, 5585–5600. doi: 10.1002/2016WR019862
Culligan, K. A., Wildenschild, D., Christensen, B. S. B., Gary, W. G., and Rivers, M. L. (2006). Pore- scale characteristics of multiphase flow in porous media: a comparison of air-water and oil-water experiments. Adv. Water Resour. 29, 227–38. doi: 10.1016/j.advwatres.2005.03.021
Dal Ferro, N., Sartori, L., Simonetti, G., Berti, A., and Morari, F. (2014). Soil macro- and microstructure as affected by different tillage systems and their effects on maize root growth. Soil Tillage Res. 140, 55–65. doi: 10.1016/j.still.2014.02.003
Daly, K. R., Keyes, S. D., Masum, S., and Roose, T. (2016). Image-based modelling of nutrient movement in and around the rhizosphere. J. Exp. Bot. 67, 1059–1070. doi: 10.1093/jxb/erv544
Daly, K. R., and Roose, T. (2014). Multiscale modelling of hydraulic conductivity in vuggy porous media. Proc. R. Soc. Lond. Ser. A Mathemat. Phys. Eng. Sci. 470:20130383. doi: 10.1098/rspa.2013.0383
Darrah, P. R., White, R. E., and Nye, P. H. A. (1987). Theoretical consideration of the implications of cell clustering for the prediction of nitrification in soil. Plant Soil 99, 387–400. doi: 10.1007/BF02370884
Davit, Y., Bell, C. G., Byrne, H. M., Chapman, L. A. C., Kimpton, L. S., Lang, G. E., et al. (2013). Homogenization via formal multiscale asymptotics and volume averaging: how do the two techniques compare? Adv. Water Resour. 62, 178–206. doi: 10.1016/j.advwatres.2013.09.006
Davit, Y., Debenest, G., Wood, B. D., and Quintard, M. (2010). Modeling non-equilibrium mass transport in biologically reactive porous media. Adv. Water Resour. 33, 1075–1093. doi: 10.1016/j.advwatres.2010.06.013
Davit, Y., Iltis, G., Debenest, G., Veran-Tissoires, S., Wildenschild, D., Gerino, M., et al. (2011). Imaging biofilm in porous media using X-ray computed microtomography. J. Microsc. 242, 15–25. doi: 10.1111/j.1365-2818.2010.03432.x
De Gryze, S., Jassogne, L., Six, J., Bossuyt, H., Wevers, M., and Merckx, R. (2006). Pore structure changes during decomposition of fresh residue: X-ray tomography analyses. Geoderma 134, 82–96. doi: 10.1016/j.geoderma.2005.09.002
de Jonge, L. W., Moldrup, P., Vendelboe, A. L., Tuller, M., and Wildenschild, D. (2012). Soil rchitecture and physicochemical functions: an introduction. Vadose Zone J. 11:vzj2011.0185. doi: 10.2136/vzj2011.0185
de Melo, R. H. C., and Conci, A. (2013). How succolarity could be used as another fractal measure in image analysis. Telecommun. Syst. 52, 1643–1655. doi: 10.1007/s11235-011-9657-3
de Ulzurrun, G. V.-D., Baetens, J. M., Van den Bulcke, J., and De Baets, B. (2017). Modelling three-dimensional fungal growth in response to environmental stimuli. J. Theor. Biol. 414, 35–49. doi: 10.1016/j.jtbi.2016.11.020
DeAngelis, K. M. (2016). Chemical communication connects soil food webs. Soil Biol. Biochem. 102, 48–51. doi: 10.1016/j.soilbio.2016.06.024
Dechesne, A., Pallud, C., Debouzie, D., Flandrois, J. P., Vogel, T. M., Gaudet, J. P., et al. (2003). A novel method for characterizing the microscale 3D spatial distribution of bacteria in soil. Soil Biol. Biochem. 35, 1537–1546. doi: 10.1016/s0038-0717(03)00243-8
DeLeo, P. C., and Baveye, P. (1997). Factors affecting protozoan predation of bacteria clogging laboratory aquifer microcosms. Geomicrobiol. J. 14, 127–149. doi: 10.1080/01490459709378039
DeLeo, P. C., Baveye, P., and Ghiorse, W. C. (1997). Use of confocal laser scanning microscopy on soil thin-sections for improved characterization of microbial growth in unconsolidated soils and aquifer materials. J. Microbiol. Methods 30, 193–203. doi: 10.1016/s0167-7012(97)00065-1
Delmont, T. O., Robe, P., Cecillon, S., Clark, I. M., Constancias, F., Simonet, P., et al. (2011). Accessing the soil metagenome for studies of microbial diversity. Appl. Environ. Microbiol. 77, 1315–1324. doi: 10.1128/aem.01526-10
Deng, J., Orner, E. P., Chau, F., Anderson, E. M., Kadilak, A. L., Rubinstein, R. L., et al. (2015). Synergistic effects of soil microstructure and bacterial EPS on drying rate in emulated soil micromodels. Soil Biol. Biochem. 83, 116–124. doi: 10.1016/j.soilbio.2014.12.006
Dennis, P. G., Miller, A. J., Clark, I. M., Taylor, R. G., Valsami-Jones, E., and Hirsch, P. R. (2008). A novel method for sampling bacteria on plant root and soil surfaces at the microhabitat scale. J. Microbiol. Methods 75, 12–18. doi: 10.1016/j.mimet.2008.04.013
Devarapalli, R. S., Islam, A., Faisal, T. F., Sassi, M., and Jouiad, M. (2017). Micro-CT and FIB-SEM imaging and pore structure characterization of dolomite rock at multiple scales. Arabian J. Geosci. 10:361. doi: 10.1007/s12517-017-3120-z
Dexter, A. R. (1988). Advances in characterization of soil structure. Soil Tillage Res. 11, 199–238. doi: 10.1016/0167-1987(88)90002-5
Díaz-Zorita, M., Perfect, E., and Grove, J. H. (2002). Disruptive methods for assessing soil structure. Soil Tillage Res. 64, 3–22. doi: 10.1016/S0167-1987(01)00254-9
Dierolf, M., Menzel, A., Thibault, P., Schneider, P., Kewish, C. M., Wepf, R., et al. (2010). Ptychographic X-ray computed tomography at the nanoscale. Nature 467, 436–482. doi: 10.1038/nature09419
Dlott, G., Maul, J. E., Buyer, J., and Yarwood, S. (2015). Microbial rRNA:rDNA gene ratios may be unexpectedly low due to extracellular DNA preservation in soils. J. Microbiol. Methods 115, 112–120. doi: 10.1016/j.mimet.2015.05.027
Dobson, K. J., Coban, S. B., McDonald, S. A., Walsh, J. N., Atwood, R. C., and Withers, P. J. (2016). 4-D imaging of sub-second dynamics in pore-scale processes using real-time synchrotron X-ray tomography. Solid Earth 7, 1059–1073. doi: 10.5194/se-7-1059-2016
Döpp, A., Hehn, L., Götzfried, J., Wenz, J., Gilljohann, M., Ding, H., et al. (2018). Quick x-ray microtomography using a laser-driven betatron source. Optica 5, 199–203. doi: 10.1364/OPTICA.5.000199
Draper, M. H., and Weidmann, S. (1951). Cardiac resting and action potentials recorded with an intracellular electrode. J. Physiol. Lon. 115, 74–94. doi: 10.1113/jphysiol.1951.sp004653
Dumestre, A., McBride, M., and Baveye, P. (2000). Use of EPR To monitor the distribution and availability of organic xenobiotics in model soil systems. Environ. Sci. Technol. 34, 1259–1264. doi: 10.1021/es990824k
Dumestre, A., Spagnuolo, M., Bladon, R., Berthelin, J., and Baveye, P. (2006). EPR monitoring of the bioavailability of an organic xenobiotic (4-hydroxy-TEMPO) in model clay suspensions and pastes. Environ. Pollut. 143, 73–80. doi: 10.1016/j.envpol.2005.11.015
Dungait, J. A. J., Hopkins, D. W., Gregory, A. S., and Whitmore, A. P. (2012). Soil organic matter turnover is governed by accessibility not recalcitrance. Global Change Biol. 18, 1781–1796. doi: 10.1111/j.1365-2486.2012.02665.x
Dupin, H. J., and McCarty, P. L. (1999). Mesoscale and microscale observations of biological growth in a silicon pore imaging element. Environ. Sci. Technol. 33, 1230–1236. doi: 10.1021/es981146p
Duwé, S., and Dedecker, P. (2017). Super-resolution imaging goes fast and deep. Nat. Methods 14, 1041–1044. doi: 10.1038/nmeth.4484
Ebrahimi, A., and Or, D. (2015). Hydration and diffusion processes shape microbial community organization and function in model soil aggregates. Water Resour. Res. 51, 9804–9827. doi: 10.1002/2015wr017565
Ebrahimi, A., and Or, D. (2016). Microbial community dynamics in soil aggregates shape biogeochemical gas fluxes from soil profiles – Upscaling an aggregate biophysical model. Global Change Biol. 22, 3141–3156. doi: 10.1111/gcb.13345
Ebrahimi, A., and Or, D. (2017). Mechanistic modeling of microbial interactions at pore to profile scale resolve methane emission dynamics from permafrost soil. J. Geophys. Res. Biogeosci. 122, 1216–1238. doi: 10.1002/2016jg003674
Ebrahimi, A. N., and Or, D. (2014). Microbial dispersal in unsaturated porous media: characteristics of motile bacterial cell motions in unsaturated angular pore networks. Water Resour. Res. 50, 7406–7429. doi: 10.1002/2014wr015897
Egan, C. K., Jacques, S. D. M., Wilson, M. D., Veale, M. C., Seller, P., Beale, A. M., et al. (2015). 3D chemical imaging in the laboratory by hyperspectral X-ray computed tomography. Sci. Rep. 5:15979. doi: 10.1038/srep15979
Eggl, E., Dierolf, M., Achterhold, K., Jud, C., Günther, B., Braig, E., et al. (2016). The munich compact light source: initial performance measures. J. Synchrotron. Radiat. 23, 1137–1142. doi: 10.1107/S160057751600967X
Eickhorst, T., and Tippkötter, R. (2008a). Improved detection of soil microorganisms using fluorescence in situ hybridization (FISH) and catalyzed reporter deposition (CARD-FISH). Soil Biol. Biochem. 40, 1883–1891. doi: 10.1016/j.soilbio.2008.03.024
Eickhorst, T., and Tippkötter, R. (2008b). Detection of microorganisms in undisturbed soil by combining fluorescence in situ hybridization (FISH) and micropedological methods. Soil Biol. Biochem. 40, 1284–1293. doi: 10.1016/j.soilbio.2007.06.019
El Ganaoui, M., Addakiri, S., and Semma, E. (2012). On the aptitude of the lattice Boltzmann approach for the treatment of the transient heat transfer with crack resistance. Comptes Rendus Mecanique 340, 518–525. doi: 10.1016/j.crme.2012.03.010
Ellegaard-Jensen, L., Knudsen, B. E., Johansen, A., Albers, C. N., Aamand, J., and Rosendahl, S. (2014). Fungal-bacterial consortia increase diuron degradation in water-unsaturated systems. Sci. Total Environ. 466, 699–705. doi: 10.1016/j.scitotenv.2013.07.095
Elliott, E. T., Anderson, R. V., Coleman, D. C., and Cole, C. V. (1980). Habitable pore space and microbial trophic interactions. Oikos 35, 327–335. doi: 10.2307/3544648
Evans, S., Dieckmann, U., Franklin, O., and Kaiser, C. (2016). Synergistic effects of diffusion and microbial physiology reproduce the Birch effect in a micro-scale model. Soil Biol. Biochem. 93, 28–37. doi: 10.1016/j.soilbio.2015.10.020
Falconer, R. E., Battaia, G., Schmidt, S., Baveye, P., Chenu, C., and Otten, W. (2015). Microscale heterogeneity explains experimental variability and non-linearity in soil organic matter mineralisation. PLoS One 10:e0123774. doi: 10.1371/journal.pone.0123774
Falconer, R. E., Bown, J. L., White, N. A., and Crawford, J. W. (2005). Biomass recycling and the origin of phenotype in fungal mycelia. Proc. R. Soc. B-Biol. Sci. 272, 1727–1734. doi: 10.1098/rspb.2005.3150
Falconer, R. E., Bown, J. L., White, N. A., and Crawford, J. W. (2007). Biomass recycling: a key to efficient foraging by fungal colonies. Oikos 116, 1558–1568. doi: 10.1111/j.2007.0030-1299.15885.x
Falconer, R. E., Houston, A. N., Otten, W., and Baveye, P. C. (2012). Emergent behavior of soil fungal dynamics: influence of soil architecture and water distribution. Soil Sci. 177, 111–119. doi: 10.1097/SS.0b013e318241133a
Feeney, D. S., Crawford, J. W., Daniell, T., Hallett, P. D., Nunan, N., Ritz, K., et al. (2006). Three-dimensional microorganization of the soil–root–microbe system. Microb. Ecol. 52, 151–158. doi: 10.1007/s00248-006-9062-8
Fierer, N. (2017). Embracing the unknown: disentangling the complexities of the soil microbiome. Nat. Rev. Microbiol. 15, 579–590. doi: 10.1038/nrmicro.2017.87
Fierer, N., and Jackson, R. B. (2006). The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. U.S.A. 103, 626–631. doi: 10.1073/pnas.0507535103
Fierer, N., Schimel, J. P., and Holden, P. A. (2003). Influence of drying–rewetting frequency on soil bacterial community structure. Microb. Ecol. 45, 63–71. doi: 10.1007/s00248-002-1007-2
Fontaine, S., Barot, S., Barre, P., Bdioui, N., Mary, B., and Rumpel, C. (2007). Stability of organic carbon in deep soil layers controlled by fresh carbon supply. Nature 450, 277–281. doi: 10.1038/nature06275
Foster, R. C. (1988). Microenvironments of soil microorganisms. Biol. Fertil. Soils 6, 189–203. doi: 10.1007/BF00260816
Fowler, A. C. (1997). Mathematical Models in the Applied Sciences. Cambridge: Cambridge University Press.
Freed, D. M. (1998). Lattice-boltzmann method for macroscopic porous media modeling. Int. J. Modern Phys. C 9, 1491–1503. doi: 10.1142/s0129183198001357
Freudiger, C. W., Min, W., Holtom, G. R., Xu, B., Dantus, M., and Xie, X. S. (2011). Highly specific label-free molecular imaging with spectrally tailored excitation-stimulated Raman scattering (STE-SRS) microscopy. Nat. Photon. 5, 103–109. doi: 10.1038/nphoton.2010.294
Fröhlich, J., and König, H. (1999). Rapid isolation of single microbial cell from mixed natural and laboratory populations with the aid of a micromanipulator. Syst. Appl. Microbiol. 22, 249–257. doi: 10.1016/S0723-2020(99)80072-1
Fröhlich, J., and König, H. (2000). New techniques for isolation of single prokaryotic cells. FEMS Microbiol. Rev. 24, 567–572. doi: 10.1111/j.1574-6976.2000.tb00558.x
Fröhlich, J., and König, H. (2006). “Micromanipulation techniques for the isolation of single microorganisms,” in Intestinal Microorganisms of Termites and Other Invertebrates, Vol. 6, eds H. König and A. Varma (Berlin: Springer), 425–437. doi: 10.1007/3-540-28185-1_18
Gao, J. F., Xing, H. L., Tian, Z. W., and Muhlhaus, H. (2014). Lattice Boltzmann modeling and evaluation of fluid flow in heterogeneous porous media involving multiple matrix constituents. Comput. Geosci. 62, 198–207. doi: 10.1016/j.cageo.2013.07.019
Gao, Y., and Sharma, M. M. (1994). A lga model for fluid-flow in heterogeneous porous-media. Transp. Porous Media 17, 1–17. doi: 10.1007/BF00624047
Garnier, P., Angulo-Jaramillo, R., DiCarlo, D. A., Bauters, T. W. J., Darnault, C. J. G., Steenhuis, T. S., et al. (1998). Dual-energy synchrotron X ray measurements of rapid soil density and water content changes in swelling soils during infiltration. Water Resour. Res. 34, 2837–2842. doi: 10.1029/98wr02367
Garnier, P., Perrier, E., Jaramillo, R. A., and Baveye, P. (1997). Numerical model of 3-dimensional anisotropic deformation and 1-dimensional water flow in swelling soils. Soil Sci. 162, 410–420. doi: 10.1097/00010694-199706000-00003
Genty, A., and Pot, V. (2013). Numerical Simulation of 3D liquid-gas distribution in porous media by a two-phase TRT lattice boltzmann method. Transp. Porous Media 96, 271–294. doi: 10.1007/s11242-012-0087-9
Genty, A., and Pot, V. (2014). Numerical calculation of effective diffusion in unsaturated porous media by the TRT lattice boltzmann method. Transp. Porous Media 105, 391–410. doi: 10.1007/s11242-014-0374-8
Gerke, K. M., Vasilyev, R. V., Khirevich, S., Collins, D., Karsanina, M. V., Sizonenko, T. O., et al. (2018). Finite-difference method stokes solver (FDMSS) for 3D pore geometries: Software development, validation and case studies. Comput. Geosci. 114, 41–58. doi: 10.1016/j.cageo.2018.01.005
Gharasoo, M., Centler, F., Regnier, P., Harms, H., and Thullner, M. (2012). A reactive transport modeling approach to simulate biogeochemical processes in pore structures with pore-scale heterogeneities. Environ. Modell. Softw. 30, 102–114. doi: 10.1016/j.envsoft.2011.10.010
Ginzburg, I. (2016). Comment on “An improved gray Lattice Boltzmann model for simulating fluid flow in multi-scale porous media”: intrinsic links between LBE Brinkman schemes. Adv. Water Resour. 88, 241–249. doi: 10.1016/j.advwatres.2014.05.007
Ginzburg, I., Silva, G., and Talon, L. (2015). Analysis and improvement of Brinkman lattice Boltzmann schemes: bulk, boundary, interface. Similarity and distinctness with finite elements in heterogeneous porous media. Phys. Rev. E91:0233072. doi: 10.1103/PhysRevE.91.023307
Gleber, S. C., Vogt, S., Niemeyer, J., Finney, L., McNulty, I., and Thieme, J. (2011). “Hard X-ray fluorescence microscopy to determine the element distribution of soil colloids in aqueous environment,” in Proceedings of the 10th International Conference on X-Ray Microscopy, Vol. 1365, eds I. McNulty, C. Eyberger, and B. Lai (College Park, MD: American Institute of Physics), 365–368. doi: 10.1063/1.3625379
Glick, B. R. (2012). Plant growth-promoting bacteria: mechanisms and applications. Scientifica 2012, 1–15. doi: 10.6064/2012/963401
Glick, B. R. (2014). Bacteria with ACC deaminase can promote plant growth and help to feed the world. Spec. Issue Plant Growth Promot. 169, 30–39. doi: 10.1016/j.micres.2013.09.009
Godfray, H. C. J., Beddington, J. R., Crute, I. R., Haddad, L., Lawrence, D., Muir, J. F., et al. (2010). Food security: the challenge of feeding 9 billion people. Science 327, 812–818. doi: 10.1126/science.1185383
Goebel, M.-O., Woche, S. K., and Bachmann, J. (2009). Do soil aggregates really protect encapsulated organic matter against microbial decomposition? Biologia 64, 443–448. doi: 10.2478/s11756-009-0065-z
Golfier, F., Wood, B. D., Orgogozo, L., Quintard, M., and Bues, M. (2009). Biofilms in porous media: development of macroscopic transport equations via volume averaging with closure for local mass equilibrium conditions. Adv. Water Resour. 32, 463–485. doi: 10.1016/j.advwatres.2008.11.012
Gommes, C. J., Bons, A.-J., Blacher, S., Dunsmuir, J. H., and Tsou, A. H. (2009). Practical methods for measuring the tortuosity of porous materials from binary or gray-tone tomographic reconstructions. AICHE J. 55, 2000–2012. doi: 10.1002/aic.11812
Gonzalez, J. L., de Faria, E. L., Albuquerque, M. P., Albuquerque, M. P., Bom, C. R., Freitas, J. C. C., et al. (2018). Representative elementary volume for simulations based on X-ray microtomography of sedimentary rock. J. Petrol. Sci. Eng. 166, 906–912. doi: 10.1016/j.petrol.2018.03.104
Gottardi, R., Kao, P.-H., Saar, M. O., and Teyssier, C. (2013). Effects of permeability fields on fluid, heat, and oxygen isotope transport in extensional detachment systems. Geochem. Geophys. Geosyst. 14, 1493–1522. doi: 10.1002/ggge.20100
Gras, A., Ginovart, M., Portell, X., and Baveye, P. C. (2010). Individual-based modeling of carbon and nitrogen dynamics in soils: parameterization and sensitivity analysis of abiotic components. Soil Sci. 175, 363–374. doi: 10.1097/SS.0b013e3181eda507
Gras, A., Ginovart, M., Valls, J., and Baveye, P. C. (2011). Individual-based modelling of carbon and nitrogen dynamics in soils: parameterization and sensitivity analysis of microbial components. Ecol. Modell. 222, 1998–2010. doi: 10.1016/j.ecolmodel.2011.03.009
Griffith, E. (1965). Micro-organisms and soil structure. Biol. Rev. Camb. Philos. Soc. 40, 129–142. doi: 10.1111/j.1469-185X.1965.tb00799.x
Grundmann, G. L. (2004). Spatial scales of soil bacterial diversity – The size of a clone. FEMS Microbiol. Ecol. 48, 119–127. doi: 10.1016/j.femsec.2004.01.010
Hainsworth, J. M., and Aylmore, L. A. G. (1983). The use of computer-assisted tomography to determine spatial distribution of soil water content. Austr. J. Soil Res. 21, 435–443. doi: 10.1071/sr9830435
Hallet, B. (1990). Self-organization in freezing soils: from microscopic ice lenses to patterned ground. Can. J. Phys. 68, 842–852. doi: 10.1139/p90-122
Hallett, P. D., Karim, K. H., Bengough, A. G., and Otten, W. (2013). Biophysics of the vadose zone: from reality to model systems and back again. Vadose Zone J. 12. doi: 10.2136/vzj2013.05.0090
Hamdi, S., Moyano, F., Sall, S., Bernoux, M., and Chevallier, T. (2013). Synthesis analysis of the temperature sensitivity of soil respiration from laboratory studies in relation to incubation methods and soil conditions. Soil Biol. Biochem. 58, 115–126. doi: 10.1016/j.soilbio.2012.11.012
Han, K., Feng, Y. T., and Owen, D. R. J. (2008). Modelling of thermal contact resistance within the framework of the thermal lattice Boltzmann method. Int. J. Ther. Sci. 47, 1276–1283. doi: 10.1016/j.ijthermalsci.2007.11.007
Hapca, S., Baveye, P. C., Wilson, C., Lark, R. M., and Otten, W. (2015). Three-dimensional mapping of soil chemical characteristics at micrometric scale by combining 2D SEM-EDX Data and 3D X-Ray CT Images. PLoS One 10:e0137025. doi: 10.1371/journal.pone.0137205
Hapca, S. M., Houston, A. N., Otten, W., and Baveye, P. C. (2013). New local thresholding method for soil images by minimizing grayscale intra-class variance. Vadose Zone J. 12:vzj2012.0172. doi: 10.2136/vzj2012.0172
Hapca, S. M., Wang, Z. X., Otten, W., Wilson, C., and Baveye, P. C. (2011). Automated statistical method to align 2D chemical maps with 3D X-ray computed micro-tomographic images of soils. Geoderma 164, 146–154. doi: 10.1016/j.geoderma.2011.05.018
Harris, K., Crabb, D., Young, I. M., Weaver, H., Gilligan, C. A., Otten, W., et al. (2002). In situ visualisation of fungi in soil thin sections: problems with crystallisation of the fluorochrome FB 28 (CalcofluorM2R) and improved staining by SCRI Renaissance 2200. Mycol. Res. 106, 293–297. doi: 10.1017/S0953756202005749
Harris, K., Young, I. M., Gilligan, C. A., Otten, W., and Ritz, K. (2003). Effect of bulk density on the spatial organisation of the fungus Rhizoctonia solani in soil. FEMS Microbiol. Ecol. 44, 45–56. doi: 10.1111/j.1574-6941.2003.tb01089.x
Hashemi, M. A., Khaddour, G., Francois, B., Massart, T. J., and Salager, S. (2014). A tomographic imagery segmentation methodology for three-phase geomaterials based on simultaneous region growing. Acta Geotech. 9, 831–846. doi: 10.1007/s11440-013-0289-5
Hausladen, D. M., and Fendorf, S. (2017). Hexavalent chromium generation within naturally structured soils and sediments. Environ. Sci. Technol. 51, 2058–2067. doi: 10.1021/acs.est.6b04039
Heijs, A. W. J., de Lange, J., Schoute, J. F. T., and Bouma, J. (1995). Computed tomography as a tool for non-destructive analysis of flow patterns in macroporous clay soils. Geoderma 64, 183–196. doi: 10.1016/0016-7061(94)00020-B
Heinemann, M., and Zenobi, R. (2011). Single cell metabolomics. Curr. Opin. Biotechnol. 22, 26–31. doi: 10.1016/j.copbio.2010.09.008
Helliwell, J. R., Sturrock, C. J., Grayling, K. M., Tracy, S. R., Flavel, R. J., Young, I. M., et al. (2013). Applications of X-ray computed tomography for examining biophysical interactions and structural development in soil systems: a review. Eur. J. Soil Sci. 64, 279–297. doi: 10.1111/ejss.12028
Hellweger, F. L., Clegg, R. J., Clark, J. R., Plugge, C. M., and Kreft, J.-U. (2016). Advancing microbial sciences by individual-based modelling. Nat. Rev. Microbiol. 14, 461–471. doi: 10.1038/nrmicro.2016.62
Hemes, S., Desbois, G., Urai, J. L., Schröppel, B., and Schwarz, J.-O. (2015). Multi-scale characterization of porosity in Boom clay (HADES-level, Mol, Belgium) using a combination of X-ray (-CT, 2D BIB-SEM and FIB-SEM tomography. Microporous Mesoporous Mater. 208, 1–20. doi: 10.1016/j.micromeso.2015.01.022
Herrmann, A. M., Ritz, K., Nunan, N., Clode, P. L., Pett-Ridge, J., Kilburn, M. R., et al. (2007). Nano-scale secondary ion mass spectrometry – A new analytical tool in biogeochemistry and soil ecology: a review article. Soil Biol. Biochem. 39, 1835–1850. doi: 10.1016/j.soilbio.2007.03.011
Hesterberg, D., Duff, M. C., Dixon, J. B., and Vepraskas, M. J. (2011). X-ray microspectroscopy and chemical reactions in soil microsites. J. Environ. Qual. 40, 667–678. doi: 10.2134/jeq2010.0140
Heße, F., Radu, F. A., Thullner, M., and Attinger, S. (2009). Upscaling of the advection-diffusion-reaction equation with Monod reaction. Adv. Water Resour. 32, 1336–1351. doi: 10.1016/j.advwatres.2009.05.009
Hitchcock, A., Morin, C., Li, L., Brash, J., Scholl, A., and Doran, A. (2004). Methods and examples of quantitative chemical mapping using synchrotron soft X-ray spectromicroscopy. Microsc. Microanal. 10, 1050–1051. doi: 10.1017/S1431927604886926
Holland, J. H. (1990). “Emergent models,” in Frontiers of Science, ed. A. Scott (Oxford: Blackwell), 107–125.
Hopkins, S., and Boswell, G. P. (2012). Mycelial response to spatiotemporal nutrient heterogeneity: a velocity-jump mathematical model. Fungal Ecol. 5, 124–136. doi: 10.1016/j.funeco.2011.06.006
Hopmans, J. W., and Dane, J. H. (1986). Calibration of a dual-energy gamma radiation system for multiple point measurements in a soil. Water Resour. Res. 22, 1109–1114. doi: 10.1029/WR022i007p01109
Houben, M. E., Desbois, G., and Urai, J. L. (2013). Pore morphology and distribution in the shaly facies of opalinus Clay (Mont Terri, Switzerland): Insights from representative 2D BIB–SEM investigations on mm to nm scale. Appl. Clay Sci. 71, 82–97. doi: 10.1016/j.clay.2012.11.006
Houston, A. N., Otten, W., Baveye, P. C., and Hapca, S. (2013a). Adaptive-window indicator kriging: a thresholding method for computed tomography images of porous media. Comput. Geosci. 54, 239–248. doi: 10.1016/j.cageo.2012.11.016
Houston, A. N., Schmidt, S., Tarquis, A. M., Otten, W., Baveye, P. C., and Hapca, S. M. (2013b). Effect of scanning and image reconstruction settings in X-ray computed microtomography on quality and segmentation of 3D soil images. Geoderma 207, 154–165. doi: 10.1016/j.geoderma.2013.05.017
Houston, A. N., Otten, W., Falconer, R., Monga, O., Baveye, P. C., and Hapca, S. M. (2017). Quantification of the pore size distribution of soils: assessment of existing software using tomographic and synthetic 3D images. Geoderma 299, 73–82. doi: 10.1016/j.geoderma.2017.03.025
Hron, P., Jost, D., Bastian, P., Gallert, C., Winter, J., and Ippisch, O. (2015). Application of reactive transport modeling to growth and transport of microorganisms in the capillary fringe. Vadose Zone J. 14:vzj2014.07.0092. doi: 10.2136/vzj2014.07.0092
Hu, H.-W., Chen, D., and He, J.-Z. (2015). Microbial regulation of terrestrial nitrous oxide formation: understanding the biological pathways for prediction of emission rates. FEMS Microbiol. Rev. 39, 729–749. doi: 10.1093/femsre/fuv021
Iassonov, P., Gebrenegus, T., and Tuller, M. (2009). Segmentation of X-ray computed tomography images of porous materials: a crucial step for characterization and quantitative analysis of pore structures. Water Resour. Res. 45:W09415. doi: 10.1029/2009wr008087
Iliev, O., Lakdawala, Z., Neßler, K. H. L., Prill, T., Vutov, Y., Yang, Y., et al. (2017). On the pore-scale modeling and simulation of reactive transport in 3D geometries. Mathemat. Modell. Anal. 22, 671–694. doi: 10.3846/13926292.2017.1356759
Iltis, G. C., Armstrong, R. T., Jansik, D. P., Wood, B. D., and Wildenschild, D. (2011). Imaging biofilm architecture within porous media using synchrotron-based X-ray computed microtomography. Water Resour. Res. 47:W02601. doi: 10.1029/2010wr009410
Inselsbacher, E., Oyewole, O. A., and Näsholm, T. (2014). Early season dynamics of soil nitrogen fluxes in fertilized and unfertilized boreal forests. Soil Biol. Biochem. 74, 167–176. doi: 10.1016/j.soilbio.2014.03.012
Ishii, S., Tago, K., and Senoo, K. (2010). Single-cell analysis and isolation for microbiology and biotechnology: methods and applications. Appl. Microbiol. Biotechnol. 86, 1281–1292. doi: 10.1007/s00253-010-2524-4
Ishoey, T., Woyke, T., Stepanauskas, R., Novotny, M., and Lasken, R. S. (2008). Genomic sequencing of single microbial cells from environmental samples. Curr. Opin. Microbiol. 11, 198–204. doi: 10.1016/j.mib.2008.05.006
Ishøy, T., Kvist, T., Westermann, P., and Ahring, B. K. (2006). An improved method for single cell isolation of prokaryotes from meso-, thermo- and hyperthermophilic environments using micromanipulation. Appl. Microbiol. Biotechnol. 69, 510–514. doi: 10.1007/s00253-005-0014-x
Jacobson, A. R., Dousset, S., Andreux, F., and Baveye, P. C. (2007). Electron microprobe and synchrotron X-ray fluorescence mapping of the heterogeneous distribution of copper in high-copper vineyard soils. Environ. Sci. Technol. 41, 6343–6349. doi: 10.1021/es070707m
Jasinska, E., Wetzel, H., Baumgartl, T., and Horn, R. (2006). Heterogeneity of physico-chemical properties in structured soils and its consequences. Pedosphere 16, 284–296. doi: 10.1016/S1002-0160(06)60054-4
Jassogne, L., Hettiarachchi, G., McNeill, A., and Chittleborough, D. (2012). Characterising the chemistry of micropores in a sodic soil with strong texture-contrast using synchrotron X-ray techniques and LA-ICP-MS. Soil Res. 50, 424–435. doi: 10.1071/sr11312
Jeger, M. J., Lamour, A., Gilligan, C. A., and Otten, W. (2008). A fungal growth model fitted to carbon-limited dynamics of Rhizoctonia solani. New Phytol. 178, 625–633. doi: 10.1111/j.1469-8137.2008.02394.x
Jenkinson, D. S. (1966). Studies on the decomposition of plant material in soil: II. Partial sterilization of soil and the soil biomass. J. Soil Sci. 17, 280–302. doi: 10.1111/j.1365-2389.1966.tb01474.x
Ji, N. (2017). Adaptive optical fluorescence microscopy. Nat. Methods 14, 374–380. doi: 10.1038/nmeth.4218
Jiang, Y. J., Sun, B., Li, H. X., Liu, M. Q., Chen, L. J., and Zhou, S. (2015). Aggregate-related changes in network patterns of nematodes and ammonia oxidizers in an acidic soil. Soil Biol. Biochem. 88, 101–109. doi: 10.1016/j.soilbio.2015.05.013
Joergensen, R. G., and Wichern, F. (2018). Alive and kicking: why dormant soil microorganisms matter. Soil Biol. Biochem. 116, 419–430. doi: 10.1016/j.soilbio.2017.10.022
Johnson, R. A., Ellsworth, T. E., Hudson, R. J. M., and Sims, G. K. (2013). Diffusion limitation for atrazine biodegradation in soil. Adv. Microbiol. 3, 412–420. doi: 10.4236/aim.2013.35056
Jones, B., and Feng, Y. T. (2011). “Permeability assessment of heterogeneous porous media using the Lattice Boltzmann method,” in Particle-Based Methods: Fundamentals and Applications, eds E. Oñate and R. Owen (Berlin: Springer), 865–881.
Jones, D., and Griffiths, E. (1964). The use of thin soil sections for the study of soil microorganisms. Plant Soil 20, 232–240. doi: 10.1007/bf01376452
Joschko, M., Graff, O., Muller, P. C., Kotzke, K., Lindner, P., Pretschner, D. P., et al. (1991). A non-destructive method for the morphological assessment of earthworm burrow systems in three dimensions by x-ray computed tomography. Biol. Fertil. Soils 11, 88–92. doi: 10.1007/BF00336369
Juyal, A., Eickhorst, T., Falconer, R., Baveye, P. C., Spiers, A., and Otten, W. (2018). Control of pore geometry in soil microcosms and its effect on the growth of Pseudomonas and Bacillus sp. Front. Environ. Sci. 6:73. doi: 10.3389/fenvs.2018.00073
Kaisermann, A., Maron, P. A., Beaumelle, L., and Lata, J. C. (2015). Fungal communities are more sensitive indicators to non-extreme soil moisture variations than bacterial communities. Appl. Soil Ecol. 86, 158–164. doi: 10.1016/j.apsoil.2014.10.009
Kalyuzhnaya, M. G., Lapidus, A., Ivanova, N., Copeland, A. C., McHardy, A. C., Szeto, E., et al. (2008). High-resolution metagenomics targets specific functional types in complex microbial communities. Nat. Biotechnol. 26, 1029–1034. doi: 10.1038/nbt.1488
Kang, Y., Norris, M. H., Zarzycki-Siek, J., Nierman, W. C., Donachie, S. P., and Hoang, T. T. (2011). Transcript amplification from single bacterium for transcriptome analysis. Genome Res. 21, 925–935. doi: 10.1101/gr.116103.110
Karadimitriou, N. K., and Hassanizadeh, S. M. (2012). A review of micromodels and their use in two-phase flow studies. Vadose Zone J. 11. doi: 10.2136/vzj2011.0072
Karsanina, M. V., Gerke, K. M., Skvortsova, E. B., and Mallants, D. (2015). Universal spatial correlation functions for describing and reconstructing soil microstructure. PLoS One 10:e0126515. doi: 10.1371/journal.pone.0126515
Keck, H., Strobel, B. W., Gustafsson, J. P., and Koestel, J. (2017). Quantitative imaging of the 3-D distribution of cation adsorption sites in undisturbed soil. Soil 3, 177–180. doi: 10.5194/soil-3-177-2017
Keiluweit, M., Gee, K., Denney, A., and Fendorf, S. (2018). Anoxic microsites in upland soils dominantly controlled by clay content. Soil Biol. Biochem. 118, 42–50. doi: 10.1016/j.soilbio.2017.12.002
Keiluweit, M., Wanzek, T., Kleber, M., Nico, P., and Fendorf, S. (2017). Anaerobic microsites have an unaccounted role in soil carbon stabilization. Nat. Commun. 8:1771. doi: 10.1038/s41467-017-01406-6
Kettridge, N., and Binley, A. (2011). Characterization of peat structure using X-ray computed tomography and its control on the ebullition of biogenic gas bubbles. J. Geophys. Res. 116:G01024. doi: 10.1029/2010JG001478
Killham, K., Amato, M., and Ladd, J. N. (1993). Effect of substrate location in soil and soil porewater regime on carbon turnover. Soil Biol. Biochem. 25, 57–62. doi: 10.1016/0038-0717(93)90241-3
Kim, M., and Or, D. (2016). Individual-based model of microbial life on hydrated rough soil surfaces. PLoS One 11:e0147394. doi: 10.1371/journal.pone.0147394
Kinyangi, J., Solomon, D., Liang, B., Lerotic, M., Wirick, S., and Lehmann, J. (2006). Nanoscale biogeocomplexity of the organomineral assemblage in soil: application of STXM microscopy and C 1s-NEXAFS spectroscopy. Soil Sci. Soc. Am. J. 70, 1708–1718. doi: 10.2136/sssaj2005.0351
Kirschbaum, M. U. F. (2006). The temperature dependence of organic matter decomposition - still a topic of debate. Soil Biol. Biochem. 38, 2510–2518. doi: 10.1016/j.soilbio.2006.01.030
Kitanidis, P. K. (2015). Persistent questions of heterogeneity, uncertainty, and scale in subsurface flow and transport. Water Resour. Res. 51, 5888–5904. doi: 10.1002/2015wr017639
Knauth, S., Schmidt, H., and Tippkotter, R. (2013). Comparison of commercial kits for the extraction of DNA from paddy soils. Lett. Appl. Microbiol. 56, 222–228. doi: 10.1111/lam.12038
Koestel, J. (2017). SoilJ: an ImageJ plugin for the semiautomatic processing of three-dimensional X-ray images of soils. Vadose Zone J. 17. doi: 10.2136/vzj2017.03.0062
Kögel-Knabner, I., Ekschmitt, K., Flessa, H., Guggenberger, G., Matzner, E., Marschner, B., et al. (2008). An integrative approach of organic matter stabilization in temperate soils: linking chemistry, physics, and biology. J. Plant Nutr. Soil Sci. 171, 5–13. doi: 10.1002/jpln.200700215
Kohlmeier, S., Smits, T. H. M., Ford, R. M., Keel, C., Harms, H., and Wick, L. Y. (2005). Taking the fungal highway: mobilization of pollutant-degrading bacteria by fungi. Environ. Sci. Technol. 39, 4640–4646. doi: 10.1021/es047979z
Kopittke, P. M., Wang, P., Lombi, E., and Donner, E. (2017). Synchrotron-based X-Ray approaches for examining toxic trace metal(loid)s in soil-plant systems. J. Environ. Qual. 46, 1175–1189. doi: 10.2134/jeq2016.09.0361
Kotani-Tanoi, T., Nishiyama, M., Otsuka, S., and Senoo, K. (2007). Single particle analysis reveals that bacterial community structures are semi-specific to the type of soil particle. Soil Sci. Plant Nutr. 53, 740–743. doi: 10.1111/j.1747-0765.2007.00200.x
Kowalchuk, G. A. (2012). Bad news for soil carbon sequestration? Science 337, 1049–1050. doi: 10.1126/science.1227303
Kravchenko, A., Chun, H. C., Mazer, M., Wang, W., Rose, J. B., Smucker, A., et al. (2013). Relationships between intra-aggregate pore structures and distributions of Escherichia coli within soil macro-aggregates. Appl. Ecol. 63, 134–142. doi: 10.1016/j.apsoil.2012.10.001
Kravchenko, A., Falconer, R. E., Grinev, D., and Otten, W. (2011). Fungal colonization in soils with different management histories: modeling growth in three-dimensional pore volumes. Ecol. Appl. 21, 1202–1210. doi: 10.1890/10-0525.1
Kravchenko, A. N., Wang, A. N. W., Smucker, A. J. M., and Rivers, M. L. (2011). Long-term differences in tillage and land use affect intra-aggregate pore heterogeneity. Soil Sci. Soc. Am. J. 75, 1658–1666. doi: 10.2136/sssaj2011.0096
Kravchenko, A. N., and Guber, A. K. (2017). Soil pores and their contributions to soil carbon processes. Geoderma 287, 31–39. doi: 10.1016/j.geoderma.2016.06.027
Kravchenko, A. N., Negassa, W., Guber, A. K., and Schmidt, S. (2014a). New approach to measure soil particulate organic matter in intact samples using X-Ray computed microtomography. Soil Sci. Soc. Am. J. 78, 1177–1185. doi: 10.2136/sssaj2014.01.0039
Kravchenko, A. N., Negassa, W. C., Guber, A. K., Hildebrandt, B., Marsh, T. L., and Rivers, M. L. (2014b). Intra-aggregate pore structure influences phylogenetic composition of bacterial community in macroaggregates. Soil Sci. Soc. Am. J. 78, 1924–1939. doi: 10.2136/sssaj2014.07.0308
Kravchenko, A. N., Negassa, W. C., Guber, A. K., and Rivers, M. L. (2015). Protection of soil carbon within macro-aggregates depends on intra-aggregate pore characteristics. Sci. Rep. 5:16261. doi: 10.1038/srep16261
Kravchenko, A. N., Toosi, E. R., Guber, A. K., Ostrom, N. E., Yu, J., Azeem, K., et al. (2017). Hotspots of soil N2O emission enhanced through water absorption by plant residue. Nat. Geosci. 10:496. doi: 10.1038/ngeo2963
Kühl, F.-C., Mueller, M., Schellhorn, M., Mann, K., Wieneke, S., and Eusterhues, K. (2016). Near-edge x-ray absorption fine structure spectroscopy at atmospheric pressure with a table-top laser-induced soft x-ray source. J. Vacuum Sci. Technol. A 34, 041302. doi: 10.1116/1.4950599
Kuka, K., Franko, U., and Ruhlmann, J. (2007). Modelling the impact of pore space distribution on carbon turnover. Ecol. Modell. 208, 295–306. doi: 10.1016/j.ecolmodel.2007.06.002
Kulkarni, R., Tuller, M., Fink, W., and Wildenschild, D. (2012). Three-dimensional multiphase segmentation of X-Ray CT data of porous materials using a bayesian markov random field framework. Vadose Zone J. 11. doi: 10.2136/vzj2011.0082
Kuzyakov, Y. (2010). Priming effects: interactions between living and dead organic matter. Soil Biol. Biochem. 42, 1363–1371. doi: 10.1016/j.soilbio.2010.04.003
Kuzyakov, Y., and Blagodatskaya, E. (2015). Microbial hotspots and hot moments in soil: concept and review. Soil Biol. Biochem. 83, 184–199. doi: 10.1016/j.soilbio.2015.01.025
Kvist, T., Ahring, B. K., Lasken, R. S., and Westermann, P. (2007). Specific single-cell isolation and genomic amplification of uncultured microorganisms. Appl. Microbiol. Biotechnol. 74, 926–935. doi: 10.1007/s00253-006-0725-7
Lafond, J. A., Han, L., Allaire, S. E., and Dutilleul, P. (2012). Multifractal properties of porosity as calculated from computed tomography (CT) images of a sandy soil, in relation to soil gas diffusion and linked soil physical properties. Eur. J. Soil Sci. 63, 861–873. doi: 10.1111/j.1365-2389.2012.01496.x
Lal, R., and Bruce, J. P. (1999). The potential of world cropland soils to sequester C and mitigate the greenhouse effect. Environ. Sci. Policy 2, 177–185. doi: 10.1016/S1462-9011(99)00012-X
Lanning, L. M., and Ford, R. M. (2002). Glass micromodel study of bacterial dispersion in spatially periodic porous networks. Biotechnol. Bioeng. 78, 556–566. doi: 10.1002/bit.10236
Lasken, R. S. (2012). Genomic sequencing of uncultured microorganisms from single cells. Nat. Rev. Microbiol. 10, 631–640. doi: 10.1038/nrmicro2857
Lasken, R. S. (2013). Single-cell sequencing in its prime. Nat. Biotechnol. 31, 211–212. doi: 10.1038/nbt.2523
Laughlin, R. J., and Stevens, R. J. (2002). Evidence for fungal dominance of denitrification and codenitrification in a grassland soil. Soil Sci. Soc. Am. J. 66, 1540–1548. doi: 10.2136/sssaj2002.1540
Lavelle, P., Barot, S., Blouin, M., Decaens, T., Jimenez, J. J., and Jouquet, P. (2007). “Earthworms as key actors in self-organized soil systems,” in Ecosystem Engineers: Plants to Protists, eds K. Cuddington, J. E. Byers, W. G. Wilson, and A. Hastings (Cambrdge, MA: Academic press), 77–106. doi: 10.1016/S1875-306X(07)80007-4
Lavelle, P., Spain, A., Blouin, M., Brown, G., Decaens, T., Grimaldi, M., et al. (2016). Ecosystem engineers in a self-organized soil: a review of concepts and future research Questions. Soil Sci. 181, 91–109. doi: 10.1097/ss.0000000000000155
LeCun, Y., Bengio, Y., and Hinton, G. (2015). Deep learning. Nature 521, 436–444. doi: 10.1038/nature14539
Ledbetter, M., and Budker, D. (2013). Zero-field nuclear magnetic resonance. Phys. Today 66, 44–49. doi: 10.1063/PT.3.1948
Lee, B. H., and Lee, S. K. (2017). Probing the water distribution in porous model sans with two immiscible fluids: a nuclear magentic resonance micro-imaging study. J. Hydrol. 553, 637–650. doi: 10.1016/j.jhydrol.2017.08.035
Lehmann, J., Solomon, D., Kinyangi, J., Dathe, L., Wirick, S., and Jacobsen, C. (2008). Spatial complexity of soil organic matter forms at nanometre scales. Nat. Geosci. 1, 238–242. doi: 10.1038/ngeo155
Lehmann, J., Wyss, P., Flisch Lehmann, E., Vontobel, P., Krafczyk, M., Kaestner, A., et al. (2006). Tomographical imaging and mathematical description of porous media used for the prediction of fluid distribution. Vadose Zone J. 5, 80–97. doi: 10.2136/vzj2004.0177
Lerch, T. Z., Nunan, N., Dignac, M. F., Chenu, C., and Mariotti, A. (2011). Variations in microbial isotopic fractionation during soil organic matter decomposition. Biogeochemistry 106, 5–21. doi: 10.1007/s10533-010-9432-7
Letey, J. (1991). The study of soil structure: science or art? Austr. J. Soil Res. 29, 699–707. doi: 10.1071/sr9910699
Li, J., Pignatello, J. J., Smets, B. F., Grasso, D., and Monserrate, E. (2005). Bench-scale evaluation of in situ bioremediation strategies for soil at a former manufactured gas plant site. Environ. Toxicol. Chem. 24, 741–749. doi: 10.1897/04-247r.1
Li, L., Peters, C. A., and Celia, M. A. (2006). Upscaling geochemical reaction rates using pore-scale network modeling. Adv. Water Resour. 29, 1351–1370. doi: 10.1016/j.advwatres.2005.10.011
Li, L., Peters, C. A., and Celia, M. A. (2007a). Applicability of averaged concentrations in determining geochemical reaction rates in heterogeneous porous media. Am. J. Sci. 307, 1146–1166. doi: 10.2475/10.2007.02
Li, L., Peters, C. A., and Celia, M. A. (2007b). Effects of mineral spatial distribution on reaction rates in porous media. Water Resour. Res. 43:W01419. doi: 10.1029/2005WR004848
Li, L., Peters, C. A., and Celia, M. A. (2007c). Reply to “Comment on upscaling geochemical reaction rates using pore-scale network modeling” by Peter C. Lichtner and Qinjun Kang. Adv. Water Resour. 30, 691–695. doi: 10.1016/j.advwatres.2006.05.002
Li, L., Wen, K., Li, C., and Amini, F. (2017). FIB/SEM imaging of microbial induced calcite precipitation in sandy soil. Microsc. Microanal. 23, 310–311. doi: 10.1017/S1431927617002239
Li, M., Xu, J., Romero-Gonzalez, M., Banwart, S. A., and Huang, W. E. (2012). Single cell Raman spectroscopy for cell sorting and imaging. Curr. Opin. Biotechnol. 23, 56–63. doi: 10.1016/j.copbio.2011.11.019
Li, R., Yang, Y. S., Pan, J., Pereira, G. G., Taylor, J. A., Clennell, B., et al. (2014). Lattice Boltzmann modeling of permeability in porous materials with partially percolating voxels. Phys. Rev. E 90:33301. doi: 10.1103/PhysRevE.90.033301
Li, Y., Dick, W. A., and Tuovinen, O. H. (2003). Evaluation of fluorochromes for imaging bacteria in soil. Soil Biol. Biochem. 35, 737–744. doi: 10.1016/s0038-0717(02)00196-7
Li, Y., Dick, W. A., and Tuovinen, O. H. (2004). Fluorescence microscopy for visualization of soil microorganisms – A review. Biol. Fertil. Soils 39, 301–311. doi: 10.1007/s00374-004-0722-x
Li, Z., Zhang, X., Wang, D., and Liu, Y. (2018). Direct methods to calculate the mass exchange between solutes inside and outside aggregates in macroscopic model for solute transport in aggregated soil. Geoderma 320, 126–135. doi: 10.1016/j.geoderma.2018.01.021
Lichtner, P. C., and Kang, Q. (2007b). Upscaling pore-scale reactive transport equations using a multiscale continuum formulation. Water Resour. Res. 43:W12s15.
Lichtner, P. C., and Kang, Q. J. (2007a). Comment on: “Upscaling geochemical reaction rates using pore-scale network modeling” by Li, Peters and Celia. Adv. Water Resour. 30, 686–690. doi: 10.1016/j.advwatres.2006.05.005
Lilje, O., Lilje, E., Marano, A. V., and Gleason, F. H. (2013). Three dimensional quantification of biological samples using micro-computer aided tomography (microCT). J. Microbiol. Methods 92, 33–41. doi: 10.1016/j.mimet.2012.10.006
Lin, H. (2012). “Understanding soil architecture and its functional manifestation across scales,” in Hydropedology, ed. H. Lin (Amsterdam: Elsevier),41–74.
Lin, H., Flühler, H., Otten, W., and Vogel, H. J. (2010). Soil architecture and preferential flow across scales. J. Hydrol. 393, 1–2. doi: 10.1016/j.jhydrol.2010.07.026
Liu, J., Pereira, G. G., Liu, Q., and Regenauer-Lieb, K. (2016). Computational challenges in the analyses of petrophysics using microtomography and upscaling: a review. Comput. Geosci. 89, 107–117. doi: 10.1016/j.cageo.2016.01.014
Liu, Y., King, H. E., van Huis, M. A., Drury, M. R., and Plümper, O. (2017). FIB-SEM tomography of porous geological materials: a state-of-the-art 3D characterization technique. GIT. Imag. Microsc. 2, 41–43.
Loehle, C., and Johnson, P. (1994). A framework for modeling microbial transport and dynamics in the subsurface. Ecol. Modell. 73, 31–49. doi: 10.1016/0304-3800(94)90096-5
Lombard, N., Prestat, E., van Elsas, J. D., and Simonet, P. (2011). Soil-specific limitations for access and analysis of soil microbial communities by metagenomics. FEMS Microbiol. Ecol. 78, 31–49. doi: 10.1111/j.1574-6941.2011.01140.x
Lugo-Mendez, H. D., Valdes-Parada, F. J., Porter, M. L., Wood, B. D., and Ochoa-Tapia, J. A. (2015). Upscaling diffusion and nonlinear reactive mass transport in homogeneous porous media. Transp. Porous Media 107, 683–716. doi: 10.1007/s11242-015-0462-4
Luo, L., Lin, H., and Li, S. (2010). Quantification of 3-D soil macropore networks in different soil types and land uses using computed tomography. J. Hydrol. 393, 53–64. doi: 10.1016/j.jhydrol.2010.03.031
Lühl, L., Mantouvalou, I., Schaumann, I., Vogt, C., and Kanngießer, B. (2013). Three-dimensional chemical mapping with a confocal XRF setup. Anal. Chem. 85, 3682–3689. doi: 10.1021/ac303749b
Macura, J., Szolnoki, J., Kunc, F., Vancura, V., and Babicky, A. (1965). Decomposition of glucose continuously added to soil. Folia Microbiol. 10, 44–54. doi: 10.1007/bf02869840
Malka, V., Faure, J., Gauduel, Y. A., Lefebvre, E., Rousse, A., and Phuoc, K. T. (2008). Principles and applications of compact laser-plasma accelerators. Nat. Phys. 4, 447–453. doi: 10.1038/nphys966
Männik, J., Driessen, R., Galajda, P., Keymer, J. E., and Dekker, C. (2009). Bacterial growth and motility in sub-micron constrictions. Proc. Natl. Acad. Sci. U.S.A. 106, 14861–14866. doi: 10.1073/pnas.0907542106
Manson, S. M. (2001). Simplifying complexity: a review of complexity theory. Geoforum 32, 405–414. doi: 10.1016/s0016-7185(00)00035-x
Marcy, Y., Ouverney, C., Bik, E. M., Lösekann, T., Ivanova, N., Martin, H. G., et al. (2007). Dissecting biological “dark matter” with single-cell genetic analysis of rare and uncultivated TM7 microbes from the human mouth. Proc. Natl. Acad. Sci. U.S.A. 104, 11889–11894. doi: 10.1073/pnas.0704662104
Maron, P. A., Mougel, C., and Ranjard, L. (2011). Soil microbial diversity: methodological strategy, spatial overview and functional interest. CR Biol. 334, 403–411. doi: 10.1016/j.crvi.2010.12.003
Martin-Sotoca, J. J., Saa-Requejo, A., Grau, J. B., and Tarquis, A. M. (2017). New segmentation method based on fractal properties using singularity maps. Geoderma 287, 40–53. doi: 10.1016/j.geoderma.2016.09.005
Martys, N. S., and Chen, H. (1996). Simulation of multicomponent fluids in complex three-dimensional geometries by the lattice Boltzmann method. Phys. Rev. E53, 743–750. doi: 10.1103/PhysRevE.53.743
Masue-Slowey, Y., Kocar, B. D., Jofre, S. A. B., Mayer, K. U., and Fendorf, S. (2011). Transport implications resulting from internal redistribution of arsenic and iron within constructed soil aggregates. Environ. Sci. Technol. 45, 582–588. doi: 10.1021/es1027663
Masue-Slowey, Y., Ying, S. C., Kocar, B. D., Pallud, C. E., and Fendorf, S. (2013). Dependence of arsenic fate and transport on biogeochemical heterogeneity arising from the physical structure of soils and sediments. J. Environ. Qual. 42, 1119–1129. doi: 10.2134/jeq2012.0253
McDonald, P. J., and Turner, M. N. (2016). Combining effective media and multi-phase methods of Lattice Boltzmann modelling for the characterisation of liquid-vapour dynamics in multi-length scale heterogeneous structural materials. Modell. Simul. Mater. Sci. Eng. 2. doi: 10.1088/0965-0393/24/1/015010
McIntosh, K. G., Cordes, N. L., Patterson, B. M., and Havrilla, G. J. (2015). Laboratory-based characterization of plutonium in soil particles using micro-XRF and 3D confocal XRF. J. Anal. Atomic Spectros. 30, 1511–1517. doi: 10.1039/C5JA00068H
Mcnaughton, S. J., Booth, T., Embley, T. M., and O’Donnell, A. G. (1996). Physical stabilization and confocal microscopy of bacteria on roots using 16S rRNA targeted, fluorescent-lavebeled oligonucleotide probes. J. Microb. Methods 26, 279–285. doi: 10.1016/0167-7012(96)00923-2
Meckenstock, R. U., Elsner, M., Griebler, C., Lueders, T., Stumpp, C., Aamand, J., et al. (2015). Biodegradation: updating the concepts of control for microbial cleanup in contaminated aquifers. Environ. Sci. Technol. 49, 7073–7081. doi: 10.1021/acs.est.5b00715
Meira Cassaro, F. A., Posadas Durand, A. N., Gimenez, D., and Pedro Vaz, C. M. (2017). Pore-size distributions of soils derived using a geometrical approach and multiple resolution microCT images. Soil Sci. Soc. Am. J. 81, 468–476. doi: 10.2136/sssaj2016.09.0291
Mendes, L. W., Tsai, S. M., Navarrete, A. A., de Hollander, M., van Veen, J. A., and Kuramae, E. E. (2015). Soil-borne microbiome: linking diversity to function. Microb. Ecol. 70, 255–265. doi: 10.1007/s00248-014-0559-2
Michelson, K., Sanford, R. A., Valocchi, A. J., and Werth, C. J. (2017). Nanowires of geobacter sulfurreducens require redox cofactors to reduce metals in pore spaces too small for cell passage. Environ. Sci. Technol. 51, 11660–11668. doi: 10.1021/acs.est.7b02531
Milne, A. E., Lehmann, J., Solomon, D., and Lark, R. M. (2011). Wavelet analysis of soil variation at nanometre- to micrometre-scales: an example of organic carbon content in a micro-aggregate. Eur. J. Soil Sci. 62, 617–628. doi: 10.1111/j.1365-2389.2011.01352.x
Min, W., Freudiger, C. W., Lu, S., and Xie, X. S. (2011). “Coherent nonlinear optical imaging: beyond fluorescence microscopy,” in Annual Review of Physical Chemistry, Vol. 62, eds S. R. Leone, P. S. Cremer, J. T. Groves, and M. A. Johnson (Palo Alto, CA: Annual Reviews), 507–530.
Minasny, B., Arrouays, D., McBratney, A. B., Angers, D. A., Chambers, A., Chaplot, V., et al. (2018). Rejoinder to comments on minasny et al., 2017 soil carbon 4 per mille geoderma 292, 59-86. Geoderma 309, 124–129. doi: 10.1016/j.geoderma.2017.05.026
Minasny, B., Malone, B. P., McBratney, A. B., Angers, D. A., Arrouays, D., Chambers, A., et al. (2017). Soil carbon 4 per mille. Geoderma 292, 59–86. doi: 10.1016/j.geoderma2017.01.002
Monga, O., Bousso, M., Garnier, P., and Pot, V. (2009). Using pore space 3D geometrical modelling to simulate biological activity: impact of soil structure. Comput. Geosci. 35, 1789–1801. doi: 10.1016/j.cageo.2009.02.007
Monga, O., Ngom, N. F., and Delerue, J. F. (2007). Representing geometric structures in 3D tomography soil images: aplication to pore space modelling. Comput. Geosci. 33, 1140–1161. doi: 10.1016/j.cageo.2006.11.015
Mooney, S. J. (2002). Three-dimensional visualization and quantification of soil macroporosity and water flow patterns using computed tomography. Soil Use Manage. 18, 142–151. doi: 10.1111/j.1475-2743.2002.tb00232.x
Mourou, G., Brocklesby, B., Tajima, T., and Limpert, J. (2013). The future is fibre accelerators. Nat. Photon. 7, 258–261. doi: 10.1038/nphoton.2013.75
Muci, A. L., Jorquera, M. A., Avila, A. I., Rengel, Z., Crowley, D. E., and de la Luz Mora, M. (2012). A combination of cellular automata and agent-based models for simulating the root surface colonization by bacteria. Ecol. Modell., 247, 1–10. doi: 10.1016/j.ecolmodel.2012.07.035
Mueller, C. W., Köbl, A., Hoeschen, C., Hillion, F., Hiester, K., Herrmann, A. M., et al. (2012). Submicron scale imaging of soil organic matter dynamics using NanoSIMS – From single particles to intact aggregates. Organ. Geochem. 42, 1476–1488. doi: 10.1016/j.orggeochem.2011.06.003
Mueller, C. W., Remusat, L., and Rumpel, C. (2017). “Characterization of biogeochemical processes at the microscale,” in Terrestrial Ecosystem Research Infrastructures, eds A. Chabbi and H. Loescher (Boca Raton, FL: CRC Press), 193–212. doi: 10.1201/9781315368252-9
Mueller, C. W., Weber, P. K., Kilburn, M. R., Hoeschen, C., Kleber, M., and Pett-Ridge, J. (2013). Advances in the analysis of biogeochemical interfaces: nanoSIMS to investigate soil microenvironments. Adv. Agron. 121, 1–46. doi: 10.1016/b978-0-12-407685-3.00001-3
Müller, M., Mey, T., Niemeyer, J., and Mann, K. (2014). Table-top soft x-ray microscope using laser-induced plasma from a pulsed gas jet. Optics Express 22, 23489–23495. doi: 10.1364/oe.22.023489
Nannipieri, P., Ascher, J., Ceccherini, M. T., Landi, L., Pietramellara, G., and Renella, G. (2003). Microbial diversity and soil functions. Eur. J. Soil Sci. 54, 655–670. doi: 10.1046/j.1351-0754.2003.0556.x
Nazir, R., Shen, J.-P., Wang, J.-T., Hu, H.-W., and He, J.-Z. (2017). Fungal networks serve as novel ecological routes for enrichment and dissemination of antibiotic resistance genes as exhibited by microcosm experiments. Sci. Rep. 7:1547. doi: 10.1038/s41598-017-15660-7
Negassa, W. C., Guber, A. K., Kravchenko, A. N., Marsh, T. L., Hildebrandt, B., and Rivers, M. L. (2015). Properties of soil pore space regulate pathways of plant residue decomposition and community structure of associated bacteria. PLoS One 10:e0123999. doi: 10.1371/journal.pone.0123999
Ngom, N. F., Monga, O., Mohamed, M. M. O., and Garnier, P. (2012). 3D shape extraction segmentation and representation of soil microstructures using generalized cylinders. Comput. Geosci. 39, 50–63. doi: 10.1016/j.cageo.2011.06.010
Nishizawa, T., Tago, K., Uei, Y., Ishii, S., Isobe, K., Otsuka, S., et al. (2012). Advantages of functional single-cell isolation method over standard agar plate dilution method as a tool for studying denitrifying bacteria in rice paddy soil. AMB Express 2, 50–50. doi: 10.1186/2191-0855-2-50
Nunan, N., Lerch, T. Z., Pouteau, V., Mora, P., Changey, E., Katterer, T., et al. (2015). Metabolising old soil carbon: simply a matter of simple organic matter? Soil Biol. Biochem. 88, 128–136. doi: 10.1016/j.soilbio.2015.05.018
Nunan, N., Ritz, K., Crabb, D., Harris, K., Wu, K. J., Crawford, J. W., et al. (2001). Quantification of the in situ distribution of soil bacteria by large-scale imaging of thin sections of undisturbed soil. FEMS Microbiol. Ecol. 37, 67–77. doi: 10.1111/j.1574-6941.2001.tb00854.x
Nunan, N., Ritz, K., Rivers, M., Feeney, D. S., and Young, I. M. (2006). Investigating microbial micro-habitat structure using X-ray computed tomography. Geoderma 133, 398–407. doi: 10.1016/j.geoderma.2005.08.004
Nunan, N., Wu, K., Young, I. M., Crawford, J. W., and Ritz, K. (2002). In situ spatial patterns of soil bacterial populations, mapped at multiple scales, in an arable soil. Microb. Ecol. 44, 296–305. doi: 10.1007/s00248-002-2021-0
Nunan, N., Wu, K. J., Young, I. M., Crawford, J. W., and Ritz, K. (2003). Spatial distribution of bacterial communities and their relationships with the micro-architecture of soil. FEMS Microbiol. Ecol. 44, 203–215. doi: 10.1016/S0168-6496(03)00027-8
Oburger, E., and Schmidt, H. (2016). New methods to unravel rhizosphere processes. Trends Plant Sci. 21, 243–255. doi: 10.1016/j.tplants.2015.12.005
O’Donnell, A. G., Young, I. M., Rushton, S. P., Shirley, M. D., and Crawford, J. W. (2007). Visualization, modelling and prediction in soil microbiology. Nat. Rev. Microbiol. 5, 689–699. doi: 10.1038/nrmicro1714
Ojeda-Magana, B., Quintanilla-Dominguez, J., Ruelas, R., Tarquis, A. M., Gomez-Barba, L., and Andina, D. (2014). Identification of pore spaces in 3D CT soil images using PFCM partitional clustering. Geoderma 217, 90–101. doi: 10.1016/j.geoderma.2013.11.005
Olson, M. S., Ford, R. M., Smith, J. A., and Fernandez, E. J. (2004). Quantification of bacterial chemotaxis in porous media using magnetic resonance imaging. Environ. Sci. Technol. 38, 3864–3870. doi: 10.1021/es035236s
Or, D. (2002). Physical processes affecting microbial habitats and activity in unsaturated poous media. Agric. Sci. 7, 39–45.
Oremland, R. S., Capone, D. G., Stolz, J. F., and Fuhrman, J. A. (2005). Whither or wither geomicrobiology in the era of ‘community metagenomics’. Nat. Rev. Microbiol. 3, 572–578. doi: 10.1038/nrmicro1182
Ostadi, H., Rama, P., Liu, Y., Chen, R., Zhang, X. X., and Jiang, K. (2010). Threshold fine-tuning and 3D characterisation of porous media using X-ray nanotomography. Curr. Nanosci. 6, 226–231. doi: 10.2174/157341310790945623
Otten, W., and Gilligan, C. A. (2006). Soil structure and soil-borne diseases: using epidemiological concepts to scale from fungal spread to plant epidemics. Eur. J. Soil Sci. 57, 26–37. doi: 10.1111/j.1365-2389.2006.00766.x
Otten, W., Gilligan, C. A., Watts, C. W., Dexter, A. R., and Hall, D. (1999). Continuity of air-filled pores and invasion thresholds for a soilborne fungal plant pathogen, Rhizoctonia solani. Soil Biol. Biochem. 31, 1803–1810. doi: 10.1016/s0038-0717(99)00099-1
Otten, W., Hall, D., Harris, K., Ritz, K., Young, I. M., and Gilligan, C. A. (2001). Soil physics, fungal epidemiology and the spread of Rhizoctonia solani. New Phytol. 151, 459–468. doi: 10.1046/j.0028-646x.2001.00190.x
Pajor, R., Falconer, R., Hapca, S., and Otten, W. (2010). Modelling and quantifying the effect of heterogeneity in soil physical conditions on fungal growth. Biogeosciences 7, 3731–3740. doi: 10.5194/bg-7-3731-2010
Pallud, C., Masue-Slowey, Y., and Fendorf, S. (2010). Aggregate-scale spatial heterogeneity in reductive transformation of ferrihydrite resulting from coupled biogeochemical and physical processes. Geochim. Cosmochim. Acta 74, 2811–2825. doi: 10.1016/j.gca.2010.01.032
Pamp, S. J., Harrington, E. D., Quake, S. R., Relman, D. A., and Blainey, P. C. (2012). Single-cell sequencing provides clues about the host interactions of segmented filamentous bacteria (SFB). Genome Res. 22, 1107–1119. doi: 10.1101/gr.131482.111
Pan, X., Durrett, R. E., Zhu, H., Tanaka, Y., Li, Y., Zi, X., et al. (2013). Two methods for full-length RNA sequencing for low quantities of cells and single cells. Proc. Natl. Acad. Sci. U.S.A. 110, 594–599. doi: 10.1073/pnas.1217322109
Papadopoulos, A., Whitmore, A. P., White, R. P., Mooney, S. J., and Bird, N. R. A. (2009). Combining spatial resolutions in the multiscale analysis of soil pore size distributions. Vadose Zone J. 8, 227–232. doi: 10.2136/vzj2008.0042
Parkin, T. B., Starr, J. L., and Meisinger, J. J. (1987). Influence of sample size on measurement of soil denitrification. Soil Sci. Soc. Am. J. 51, 1492–1501. doi: 10.2136/sssaj1987.03615995005100060017x
Patankar, S. V. (1980). Numerical Heat Transfer and Fluid Flow. London: Taylor and Francis. doi: 10.1201/9781482234213
Patterson, B. M., Campbell, J., and Havrilla, G. J. (2010). Integrating 3D images using laboratory-based micro X-ray computed tomography and confocal X-ray fluorescence techniques. X-Ray Spectrom. 39, 184–190. doi: 10.1002/xrs.1245
Paustian, K., Andren, O., Janzen, H. H., Lal, R., Smith, P., Tian, G., et al. (1997). Agricultural soils as a sink to mitigate CO2 emissions. Soil Use Manage. 13, 230–244. doi: 10.1111/j.1475-2743.1997.tb00594.x
Pedersen, L. L., Smets, B. F., and Dechesne, A. (2015). Measuring biogeochemical heterogeneity at the micro scale in soils and sediments. Soil Biol. Biochem. 90, 122–138. doi: 10.1016/j.soilbio.2015.08.003
Pendleton, D. E., Dathe, A., and Baveye, P. (2005). Influence of image resolution and evaluation algorithm on estimates of the lacunarity of porous media. Phys. Rev. E72:041306, doi: 10.1103/PhysRevE.72.041306
Peng, S., Marone, F., and Dultz, S. (2014). Resolution effect in X-ray microcomputed tomography imaging and small pore’s contribution to permeability for a Berea sandstone. J. Hydrol. 510, 403–411. doi: 10.1016/j.jhydrol.2013.12.028
Pennell, K. D. (2016). “Specific surface area,” in Reference Module in Earth Systems and Environmental Sciences, ed. S. A. Elias (Oxford: Elsevier), 1–8.
Pereira, G. G. (2016). Grayscale lattice Boltzmann model for multiphase heterogeneous flow through porous media. Phys. Rev. E93:063301. doi: 10.1103/PhysRevE.93.063301
Perez-Reche, F. J., Taraskin, S. N., Otten, W., Viana, M. P., Costa, L. d. F., and Gilligan, C. A. (2012). Prominent effect of soil network heterogeneity on microbial invasion. Phys. Rev. Lett. 109:098102. doi: 10.1103/PhysRevLett.109.098102
Perfect, E., Cheng, C.-L., Kang, M., Bilheux, H. Z., Lamanna, J. M., Gragg, M. J., et al. (2014). Neutron imaging of hydrogen-rich fluids in geomaterials and engineered porous media: a review. Earth-Sci. Rev. 129, 120–135. doi: 10.1016/j.earscirev.2013.11.012
Persson, M., Huber, B., Karlsson, S., Liu, X., Chen, H., Xu, C., et al. (2014). Energy-resolved CT imaging with a photon-counting silicon-strip detector. Phys. Med. Biol. 59, 6709–6727. doi: 10.1088/1361-6560/59/22/6709
Peszynska, M., Trykozko, A., Iltis, G., Schlueter, S., and Wildenschild, D. (2016). Biofilm growth in porous media: experiments, computational modeling at the porescale, and upscaling. Adv. Water Resour. 95, 288–301. doi: 10.1016/j.advwatres.2015.07.008
Peth, C., Barkusky, F., and Mann, K. (2008). Near-edge x-ray absorption fine structure measurements using a laboratory-scale XUV source. J. Phys. D-Appl. Phys. 41:10. doi: 10.1088/0022-3727/41/10/105202
Peth, S., Chenu, C., Leblond, N., Mordhorst, A., Garnier, P., Nunan, N., et al. (2014). Localization of soil organic matter in soil aggregates using synchrotron-based X-ray microtomography. Soil Biol. Biochem. 78, 189–194. doi: 10.1016/j.soilbio.2014.07.024
Petrovic, A. M., Siebert, J. E., and Rieke, P. E. (1982). Soil bulk density analysis in 3 dimensions by computed tomographic scanning. Soil Sci. Soc. Am. J. 46, 445–450. doi: 10.2136/sssaj1982.03615995004600030001x
Pham, V. H. T., and Kim, J. (2012). Cultivation of unculturable soil bacteria. Trends Biotechnol. 30, 475–484. doi: 10.1016/j.tibtech.2012.05.007
Philip, J. R. (1991). Soils, natural science, and models. Soil Sci. 151, 91–98. doi: 10.1097/00010694-199101000-00011
Phillips, J. D. (1995). Self-organization and landscape evolution. Prog. Phys. Geogr. 19, 309–321. doi: 10.1177/030913339501900301
Phillips, J. D. (2000). Signatures of divergence and self-organization in soils and weathering profiles. J. Geol. 108, 91–102. doi: 10.1086/314386
Piccoli, I., Camarotto, C., Lazzaro, B., Furlan, L., and Morari, F. (2017). Conservation agriculture had a poor impact on the soil porosity of Veneto low-lying plain silty soils after a 5-year transition period. Land Degrad. Dev. 28, 2039–2050. doi: 10.1002/ldr.2726
Pickup, R. W. (1995). “Sampling and detecting bacterial populations in natural environments,” in Population Genetics of Bacteria, eds S. Baumerg, J. P. W. Young, E. M. H. Wellington, and J. R. Saunders (Cambridge: Cambridge University Press), 295–315.
Pini, R., and Madonna, C. (2016). Moving across scales: a quantitative assessment of X-ray CT to measure the porosity of rocks. J. Porous Mater. 23, 325–338. doi: 10.1007/s10934-015-0085-8
Pires, L. F., Borges, J., Bacchi, O., and Reichardt, K. (2010). Twenty-five years of computed tomography in soil physics: a literature review of the Brazilian contribution. Soil Tillage Res. 110, 197–210. doi: 10.1016/j.still.2010.07.013
Podar, M., Abulencia, C. B., Walcher, M., Hutchison, D., Zengler, K., Garcia, J. A., et al. (2007). Targeted access to the genomes of low-abundance organisms in complex microbial communities. Appl. Environ. Microbiol. 73, 3205–3214. doi: 10.1128/aem.02985-06
Popescu, L., and Cao, Z.-P. (2018). From microscopy to genomic approach in soil biodiversity assessment. Curr. Issues Mol. Biol. 27, 195–198. doi: 10.21775/cimb.027.195
Porta, G. M., Ceriotti, G., and Thovert, J. F. (2016). Comparative assessment of continuum-scale models of bimolecular reactive transport in porous media under pre-asymptotic conditions. J. Contam. Hydrol. 185, 1–13. doi: 10.1016/j.jconhyd.2015.12.003
Postma, J., and Altemuller, H. J. (1990). Bacteria in thin soil sections stained with the fluorescent brightener calcofluor white M2R. Soil Biol. Biochem. 22, 89–96. doi: 10.1016/0038-0717(90)90065-8
Postma, J., and van Veen, J. A. (1990). Habitable pore space and survival of Rhizobium leguminosarum biovartrifolii introduced into soil. Microb. Ecol. 19, 149–161. doi: 10.1007/BF02012096
Pot, V., Peth, S., Monga, O., Vogel, L. E., Genty, A., Garnier, P., et al. (2015). Three-dimensional distribution of water and air in soil pores: comparison of two-phase two-relaxation-times lattice-Boltzmann and morphological model outputs with synchrotron X-ray computed tomography data. Adv. Water Resour. 84, 87–102. doi: 10.1016/j.advwatres.2015.08.006
Power, R. M., and Huisken, J. (2017). A guide to light-sheet fluorescence microscopy for multiscale imaging. Nat. Methods 14, 360–373. doi: 10.1038/nmeth.4224
Powlson, D., Xu, J., and Brookes, P. (2017). “Through the eye of the needle: the story of the soil microbial biomass,” in Microbial Biomass: A Paradigm Shift in Terrestrial Biogeochemistry, ed. K. R. Tate (London: World Scientific),1–40.
Powlson, D. S., Whitmore, A. P., and Goulding, K. W. T. (2011). Soil carbon sequestration to mitigate climate change: a critical re-examination to identify the true and the false. Eur. J. Soil Sci. 62, 42–55. doi: 10.1111/j.1365-2389.2010.01342.x
Prakash, O., Shouche, Y., Jangid, K., and Kostka, J. E. (2013). Microbial cultivation and the role of microbial resource centers in the omics era. Appl. Microbiol. Biotechnol. 97, 51–62. doi: 10.1007/s00253-012-4533-y
Pratama, A. A., and van Elsas, J. D. (2018). The “neglected” soil virome: potential role and impact. Trends Microbiol. 26, 649–662. doi: 10.1016/j.tim.2017.12.004
Preston, A. R., Bird, N. R. A., Kinchesh, P., Randall, E. W., and Whalley, W. R. (2001). STRAFI-NMR studies of water transport in soil. Magn. Reson. Imaging 19, 561–563. doi: 10.1016/S0730-725X(01)00305-8
Prietzel, J., Thieme, J., Neuhausler, U., Susini, J., and Kögel-Knabner, I. (2003). Speciation of sulphur in soils and soil particles by X-ray spectromicroscopy. Eur. J. Soil Sci. 54, 423–433. doi: 10.1046/j.1365-2389.2003.00543.x
Prosser, J. I. (2015). Dispersing misconceptions and identifying opportunities for the use of ‘omics’ in soil microbial ecology. Nat. Rev. Microbiol. 13, 439–446. doi: 10.1038/nrmicro3468
Prosser, J. I., Bohannan, B. J. M., Curtis, T. P., Ellis, R. J., Firestone, M. K., Freckleton, R. P., et al. (2007). The role of ecological theory in microbial ecology. Nat. Rev. Microbiol. 5, 384–392. doi: 10.1038/nrmicro1643
Puspita, I. D., Kamagata, Y., Tanaka, M., Asano, K., and Nakatsu, C. H. (2012). Are uncultivated bacteria really uncultivable? Microbes Environ. 27, 356–366. doi: 10.1264/jsme2.ME12092
Rab, M. A., Haling, R. E., Aarons, S. R., Hannah, M., Young, I. M., and Gibson, D. (2014). Evaluation of X-ray computed tomography for quantifying macroporosity of loamy pasture soils. Geoderma 213, 460–470. doi: 10.1016/j.geoderma.2013.08.037
Rabbi, S. M. F., Daniel, H., Lockwood, P. V., Macdonald, C., Pereg, L., Tighe, M., et al. (2016). Physical soil architectural traits are functionally linked to carbon decomposition and bacterial diversity. Sci. Rep. 6:33012. doi: 10.1038/srep33012
Rabbi, S. M. F., Wilson, B. R., Lockwood, P. V., Daniel, H., and Young, I. M. (2014). Soil organic carbon mineralization rates in aggregates under contrasting land uses. Geoderma 216, 10–18. doi: 10.1016/j.geoderma.2013.10.023
Rabot, E., Lacoste, M., Henault, C., and Cousin, I. (2015). Using X-ray computed tomography to describe the dynamics of nitrous oxide emissions during soil drying. Vadose Zone J. 14, 1–34. doi: 10.2136/vzj2014.12.0177
Rabot, E., Wiesmeier, M., Schlüter, S., and Vogel, H. J. (2018). Soil structure as an indicator of soil functions: a review. Geoderma 314, 122–137. doi: 10.1016/j.geoderma.2017.11.009
Radulovich, R., Sollins, P., Baveye, P., and Solorzano, E. (1992). Bypass water flow through unsaturated microaggregated tropical soils. Soil Sci. Soc. Am. J. 56, 721–726. doi: 10.2136/sssaj1992.03615995005600030008x
Raghunathan, A., Ferguson, H. R., Bornath, C. J., Song, W., Driscoll, M., and Lasken, R. S. (2005). Genomic DNA amplification from a single bacterium. Appl. Environ. Microbiol. 71, 3342–3347. doi: 10.1128/AEM.71.6.3342-3347.2005
Ranjard, L., and Richaume, A. S. (2001). Quantitative and qualitative microscale distribution of bacteria in soil. Res. Microbiol. 152, 707–716. doi: 10.1016/S0923-2508(01)01251-7
Rawlins, B. G., Wragg, J., Reinhard, C., Atwood, R. C., Houston, A. N., Lark, R. M., et al. (2016). Three-dimensional soil organic matter distribution, accessibility and microbial respiration in macroaggregates using osmium staining and synchrotron X-ray computed tomography. Soil 2, 659–671. doi: 10.5194/soil-2-659-2016
Raynaud, X., and Nunan, N. (2014). Spatial ecology of bacteria at the microscale in soil. PLoS One 9:287217. doi: 10.1371/journal.pone.0087217
Razafimbelo, T., Chevallier, T., Albrecht, A., Chapuis-Lardy, L., Rakotondrasolo, F. N., Michellon, R., et al. (2013). Texture and organic carbon contents do not impact amount of carbon protected in Malagasy soils. Sci. Agricola 70, 204–208. doi: 10.1590/S0103-90162013000300009
Razafimbelo, T. M., Albrecht, A., Oliver, R., Chevallier, T., Chapuis-Lardy, L., and Feller, C. (2008). Aggregate associated-C and physical protection in a tropical clayey soil under Malagasy conventional and no-tillage systems. Soil Tillage Res. 98, 140–149. doi: 10.1016/j.still.2007.10.012
Razavi, B. S., Zarebanadkouki, M., Blagodatskaya, E., and Kuzyakov, Y. (2016). Rhizosphere shape of lentil and maize: spatial distribution of enzyme activities. Soil Biol. Biochem. 96, 229–237. doi: 10.1016/j.soilbio.2016.02.020
Reardon, S. (2017). Diamond offers MRI for molecules. Nature 543, 162–162. doi: 10.1038/nature.2017.21573
Remusat, L., Hatton, P. J., Nico, P. S., Zeller, B., Kleber, M., and Derrien, D. (2012). NanoSIMS study of organic matter associated with soil aggregates: advantages, limitations, and combination with STXM. Environ. Sci. Technol. 46, 3943–3949. doi: 10.1021/es203745k
Renault, P., and Stengel, P. (1993). Modeling oxygen diffusion in aggregated soils: I. Anaerobiosis inside the aggregates. Soil Sci. Soc. Am. J. 58, 1017–1023. doi: 10.2136/sssaj1994.03615995005800040004x
Rennert, T., Totsche, K. U., Heister, K., Kersten, M., and Thieme, J. (2012). Advanced spectroscopic, microscopic, and tomographic characterization techniques to study biogeochemical interfaces in soil. J. Soils Sediments 12, 3–23. doi: 10.1007/s11368-011-0417-5
Resat, H., Bailey, V., McCue, L. A., and Konopka, A. (2012). Modeling microbial dynamics in heterogeneous environments: growth on soil carbon sources. Microb. Ecol. 63, 883–897. doi: 10.1007/s00248-011-9965-x
Rillig, M. C., Muller, L. A. H., and Lehmann, A. (2017). Soil aggregates as massively concurrent evolutionary incubators. ISME J. 11, 1943–1948. doi: 10.1038/ismej.2017.56
Ringeisen, B. R., Rincon, K., Fitzgerald, L. A., Fulmer, P. A., and Wu, P. K. (2015). Printing soil: a single-step, high-throughput method to isolate micro-organisms and near-neighbour microbial consortia from a complex environmental sample. Methods Ecol. Evol. 6, 209–217. doi: 10.1111/2041-210x.12303
Rogasik, H., Crawford, J. W., Wendroth, O., Young, I. M., Joschko, M., and Ritz, K. (1999). Discrimination of soil phases by dual energy x-ray tomography. Soil Sci. Soc. Am. J. 63, 741–751. doi: 10.2136/sssaj1999.634741x
Rogasik, H., Onasch, I., Brunotte, J., Jegou, D., and Wendroth, O. (2003). “Assessment of soil structure using X-ray computed tomography,” in Applications of X-ray Computed Tomography in the geosciences, eds F. Mees, R. Swennen, M. Van Geet, and P. Jacobs (London: Geological Society, Special Publications), 151–165.
Ronn, R., Vestergard, M., and Ekelund, F. (2012). Interactions between bacteria, protozoa and nematodes in soil. Acta Protozool. 51, 223–235. doi: 10.4467/16890027ap.12.018.0764
Roose, T., Keyes, S. D., Daly, K. R., Carminati, A., Otten, W., Vetterlein, D., et al. (2016). Challenges in imaging and predictive modeling of rhizosphere processes. Plant Soil 407, 9–38. doi: 10.1007/s11104-016-2872-7
Rousk, J., and Jones, D. L. (2010). Loss of low molecular weight dissolved organic carbon (DOC) and nitrogen (DON) in H2O and 0.5M K2SO4 soil extracts. Soil Biol. Biochem. 42, 2331–2335. doi: 10.1016/j.soilbio.2010.08.017
Rovira, A. D., and Greacen, E. L. (1957). The effect of aggregate disruption on the activity of microorganisms in the soil. Austr. J. Agric. Res. 8, 659–673. doi: 10.1071/ar9570659
Rozenbaum, O., Bruand, A., and Le Trong, E. (2012). Soil porosity resulting from the assemblage of silt grains with a clay phase: new perspectives related to utilization of X-ray synchrotron computed microtomography. Comptes Rendus Geosci. 344, 516–525. doi: 10.1016/j.crte.2012.09.004
Ruamps, L. S., Nunan, N., and Chenu, C. (2011). Microbial biogeography at the soil pore scale. Soil Biol. Biochem. 43, 280–286. doi: 10.1016/j.soilbio.2010.10.010
Ruamps, L. S., Nunan, N., Pouteau, V., Leloup, J., Raynaud, X., Roy, V., et al. (2013). Regulation of soil organic C mineralisation at the pore scale. FEMS Microbiol. Ecol. 86, 26–35. doi: 10.1111/1574-6941.12078
Rubinstein, R. L., Kadilak, A. L., Cousens, V. C., Gage, D. J., and Shor, L. M. (2015). Protist-facilitated particle transport using emulated soil micromodels. Environ. Sci. Technol. 49, 1384–1391. doi: 10.1021/es503424z
Rubol, S., Dutta, T., and Rocchini, D. (2016). 2D visualization captures the local heterogeneity of oxidative metabolism across soils from diverse land-use. Sci. Total Environ. 572, 713–723. doi: 10.1016/j.scitotenv.2016.06.252
Salzer, M., Prill, T., Spettl, A., Jeulin, D., Schladitz, K., and Schmidt, V. (2015). Quantitative comparison of segmentation algorithms for FIB-SEM images of porous media. J. Microsc. 257, 23–30. doi: 10.1111/jmi.12182
San José Martínez, F., Caniego, F. J., and García-Gutiérrez, C. (2017). Lacunarity of soil macropore space arrangement of CT images: effect of soil management and depth. Geoderma 287, 80–89. doi: 10.1016/j.geoderma.2016.09.007
San José Martínez, F., Muñoz Ortega, F. J., Caniego Monreal, F. J., Kravchenko, A. N., and Wang, W. (2015). Soil aggregate geometry: measurements and morphology. Geoderma 23, 36–48. doi: 10.1016/j.geoderma.2014.08.003
Sanderlin, A. B., Vogt, S. J., Grunewald, E., Bergin, B. A., and Codd, S. L. (2013). Biofilm detection in natural unconsolidated porous media using a low-field magnetic resonance system. Environ. Sci. Technol. 47, 987–992. doi: 10.1021/es3040686
Santner, J., Larsen, M., Kreuzeder, A., and Glud, R. N. (2015). Two decades of chemical imaging of solutes in sediments and soils – a review. Anal. Chim. Acta 878, 9–42. doi: 10.1016/j.aca.2015.02.006
Schaap, M. G., Porter, M. L., Christensen, B. S. B., and Wildenschild, D. (2007). Comparison of pressure-saturation characteristics derived from computed tomography and lattice Boltzmann simulations. Water Resour. Res. 43:W12S06. doi: 10.1029/2006wr005730
Scheffer, M., Baveco, J., DeAngelis, D., Rose, K., and VanNes, E. (1995). Super-individuals, a simple solution for modeling large populations on an individual basis. Ecol. Modell. 80, 161–170. doi: 10.1016/0304-3800(94)00055-M
Scheibe, T. D., Perkins, W. A., Richmond, M. C., McKinley, M. I., Romero-Gomez, P. D. J., Oostrom, M., et al. (2015). Pore-scale and multiscale numerical simulation of flow and transport in a laboratory-scale column. Water Resour. Res. 51, 1023–1035. doi: 10.1002/2014wr015959
Schlüter, S., Leuther, F., Vogler, S., and Vogel, H. J. (2016). X-ray microtomography analysis of soil structure deformation caused by centrifugation. Solid Earth 7, 129–140. doi: 10.5194/se-7-129-2016
Schlüter, S., Sheppard, A., Brown, K., and Wildenschild, D. (2014). Image processing of multiphase images obtained via X- ray microtomography: a review. Water Resour. Res. 50, 3615–3639. doi: 10.1002/2014wr015256
Schlüter, S., and Vogel, H. J. (2016). Analysis of soil structure turnover with garnet particles and X-ray microtomography. PLoS One 11:e0159948. doi: 10.1371/journal.pone.0159948
Schlüter, S., Weller, U., and Vogel, H.-J. (2010). Segmentation of X-ray microtomography images of soil using gradient masks. Comput. Geosci. 36, 1246–1251. doi: 10.1016/j.cageo.2010.02.007
Schmidt, H., and Eickhorst, T. (2014). Detection and quantification of native microbial populations on soil-grown rice roots by catalyzed reporter deposition-fluorescence in situ hybridization. FEMS Microbiol. Ecol. 87, 390–402. doi: 10.1111/1574-6941.12232
Schmidt, H., Eickhorst, T., and Mussmann, M. (2012a). Gold-FISH: a new approach for the in situ detection of single microbial cells combining fluorescence and scanning electron microscopy. Syst. Appl. Microbiol. 35, 518–525. doi: 10.1016/j.syapm.2012.04.006
Schmidt, H., Eickhorst, T., and Tippkoetter, R. (2012b). Evaluation of tyramide solutions for an improved detection and enumeration of single microbial cells in soil by CARD-FISH. J. Microbiol. Methods 91, 399–405. doi: 10.1016/j.mimet.2012.09.021
Schmidt, H., Vetterlein, D., Koehne, J. M., and Eickhorst, T. (2015). Negligible effect of X-ray mu-CT scanning on archaea and bacteria in an agricultural soil. Soil Biol. Biochem. 84, 21–27. doi: 10.1016/j.soilbio.2015.02.010
Schumacher, M., Christl, I., Scheinost, A. C., Jacobsen, C., and Kretzschmar, R. (2005). Chemical heterogeneity of organic soil colloids investigated by scanning transmission X-ray microscopy and C-1s NEXAFS microspectroscopy. Environ. Sci. Technol. 39, 9094–9100. doi: 10.1021/es050099f
Schweizer, S. A., Hoeschen, C., Schlüter, S., Kögel-Knabner, I., and Mueller, C. W. (2018). Rapid soil formation after glacial retreat shaped by spatial patterns of organic matter accrual in microaggregates. Global Change Biol. 24, 1637–1650. doi: 10.1111/gcb.14014
Sedlmair, J., Gleber, S.-C., Peth, C., Mann, K., Niemeyer, J., and Thieme, J. (2012). Characterization of refractory organic substances by NEXAFS using a compact X-ray source. J. Soils Sediments 12, 24–34. doi: 10.1007/s11368-011-0385-9
Shah, S. M., Gray, F., Crawshaw, J. P., and Boek, E. S. (2016). Micro-computed tomography pore-scale study of flow in porous media: effect of voxel resolution. Adv. Water Resour. 95, 276–287. doi: 10.1016/j.advwatres.2015.07.012
Sheehy, J., Regina, K., Alakukku, L., and Six, J. (2015). Impact of no-till and reduced tillage on aggregation and aggregate-associated carbon in Northern European agroecosystems. Soil Tillage Res. 150, 107–113. doi: 10.1016/j.still.2015.01.015
Shi, X., Gao, W., Wang, J., Chao, S.-H., Zhang, W., and Meldrum, D. R. (2015). Measuring gene expression in single bacterial cells: recent advances in methods and micro-devices. Crit. Rev. Biotechnol. 35, 448–460. doi: 10.3109/07388551.2014.899556
Shokri, N. (2014). Pore-scale dynamics of salt transport and distribution in drying porous media. Phys. Fluids 26:012106. doi: 10.1063/1.4861755
Silversmit, G., Vekemans, B., Nikitenko, S., Schmitz, S., Schoonians, T., Brenker, F. E., et al. (2010). Spatially resolved 3D micro-XANES by a confocal detection scheme. Phys. Chem. Chem. Phys. 12, 5653–5659. doi: 10.1039/C004103N
Smagin, A. V. (1989). Soil as a product of biogeocenosis self-organization. Doklady Akademii Nauk Sssr 308, 729–731.
Smith, A. P., Bond-Lamberty, B., Benscoter, B. W., Tfaily, M.-M., Hinkle, C. R., Liu, C., et al. (2017). Shifts in pore connectivity from precipitation versus groundwater rewetting increases soil carbon loss after drought. Nat. Commun. 8:1335. doi: 10.1038/s41467-017-01320-x
Smith, R., Wright, K. L., and Ashton, L. (2016). Raman spectroscopy: an evolving technique for live cell studies. Analyst 141, 3590–3600. doi: 10.1039/C6AN00152A
Smolek, S., Pemmer, B., Folser, M., Streli, C., and Wobrauschek, P. (2012). Confocal micro-X-ray fluorescence spectrometer for light element analysis. Rev. Sci. Instrum. 83:083703. doi: 10.1063/1.4744934
Soane, B. D. (1967). Dual energy gamma-ray transmission for coincident measurement of water content and dry bulk density of soil. Nature 214, 1273–1274. doi: 10.1038/2141273a0
Söderström, B., and Erland, S. (1986). Isolation of fluorescein diacetate stained hyphae from soil by micromanipulation. Trans. Br. Mycol. Soc. 86, 465–468. doi: 10.1016/S0007-1536(86)80190-5
Sok, R. M., Varslot, T., Ghous, A., Latham, S., Sheppard, A. P., and Knackstedt, M. A. (2010). Pore scale characterization of carbonates at multiple scales: integration of micro-CT, BSEM, and FIBSEM. Petrophysics 51, 379–387.
Sollins, P., and Radulovich, R. (1988). Effects of soil physical structure on solute transport in a weathered tropical soil. Soil Sci. Soc. Am. J. 52, 1168–1173. doi: 10.2136/sssaj1988.03615995005200040050x
Solomon, D., Lehmann, J., Harden, J., Wang, J., Kinyangi, J., Heymann, K., et al. (2012). Micro- and nano-environments of carbon sequestration: multi-element STXM-NEXAFS spectromicroscopy assessment of microbial carbon and mineral associations. Chem. Geol. 329, 53–73. doi: 10.1016/j.chemgeo.2012.02.002
Solomon, D., Lehmann, J., Kinyangi, J., Liang, B. Q., and Schafer, T. (2005). Carbon K-edge NEXAFS and FTIR-ATR spectroscopic investigation of organic carbon speciation in soils. Soil Sci. Soc. Am. J. 69, 107–119. doi: 10.2136/sssaj2005.0107dup
Son, K., Brumley, D. R., and Stocker, R. (2015). Live from under the lens: exploring microbial motility with dynamic imaging and microfluidics. Nat. Rev. Microbiol. 13, 761–775. doi: 10.1038/nrmicro3567
Spanne, P., Thovert, J., Jacquin, J., Lindquist, W. B., Jones, K., and Coker, D. (1994). Synchotron computed microtomography of porous media: topology and transports. Phys. Rev. Lett. 73, 2001–2004. doi: 10.1103/PhysRevLett.73.2001
Spohn, M., Carminati, A., and Kuzyakov, Y. (2013). Soil zymography – A novel in situ method for mapping distribution of enzyme activity in soil. Soil Biol. Biochem. 58, 275–280. doi: 10.1016/j.soilbio.2012.12.004
Sposito, G. (2013). Green water and global food security. Vadose Zone J. 12:vzj2013.02.0041. doi: 10.2136/vzj2013.02.0041
Stanley, C. E., Grossmann, G., iSolvas, X., and deMello, A. J. (2016). Soil-on-a-Chip: microfluidic platforms for environmental organismal studies. Lab Chip 16, 228–241. doi: 10.1039/C5LC01285F
Stanley, C. E., Stockli, M., van Swaay, D., Sabotic, J., Kallio, P. T., Kunzler, M., et al. (2014). Probing bacterial-fungal interactions at the single cell level. Integr. Biol. (Camb.) 6, 935–945. doi: 10.1039/c4ib00154k
Stanley, C. E., and van der Heijden, M. G. A. (2017). Microbiome-on-a-chip: new frontiers in plant–microbiota research. Trends Microbiol. 25, 610–613. doi: 10.1016/j.tim.2017.05.001
Starkloff, T., Larsbo, M., Stolte, J., Hessel, R., and Ritsema, C. (2017). Quantifying the impact of a succession of freezing-thawing cycles on the pore network of a silty clay loam and a loamy sand topsoil using X-ray tomography. Catena 156, 365–374. doi: 10.1016/j.catena.2017.04.026
Steefel, C. I., Molins, S., and Trebotich, D. (2013). Pore scale processes associated with subsurface CO2 injection and sequestration. Rev. Mineral. Geochem. 77, 259–303. doi: 10.2138/rmg.2013.77.8
Stefana, G., Cornelia, B., Jorg, R., and Michael, B. (2014). Soil water availability strongly alters the community composition of soil protists. Pedobiologia 57, 205–213. doi: 10.1016/j.pedobi.2014.10.001
Steffens, M., Rogge, D. M., Mueller, C. W., Höschen, C., Lugmeier, J., Kölbl, A., et al. (2017). Identification of distinct functional microstructural domains controlling C storage in soil. Environ. Sci. Technol. 51, 12182–12189. doi: 10.1021/acs.est.7b03715
Stepanauskas, R. (2012). Single cell genomics: an individual look at microbes. Curr. Opin. Microbiol. 15, 613–620. doi: 10.1016/j.mib.2012.09.001
Stepanauskas, R. (2015). Wiretapping into microbial interactions by single cell genomics. Front. Microbiol. 6:258. doi: 10.3389/fmicb.2015.00258
Stewart, T. L., and Fogler, H. S. (2001). Biomass plug development and propagation in porous media. Biotechnol. Bioeng. 72, 353–363. doi: 10.1002/1097-0290(20010205)72:3<353::AID-BIT13>3.0.CO;2-U
Stotzky, G. (1986). “Influence of soil mineral colloids on metabolic processes, growth, adhesion, and ecology of microbes and viruses,” in Interactions of Soil Minerals with Natural Organics and Microbes, eds P. M. Huang and M. Schnitzer (Madison, WI: Soil Science Society of America), 305–428.
Strawn, D. G., and Baker, L. L. (2009). Molecular characterization of copper in soils using X-ray absorption spectroscopy. Environ. Pollut. 157, 2813–2821. doi: 10.1016/j.envpol.2009.04.018
Strong, D. T., DeWever, H., Merckx, R., and Recous, S. (2004). Spatial location of carbon decomposition in the soil pore system. Eur. J. Soil Sci. 55, 739–750. doi: 10.1111/j.1365-2389.2004.00639.x
Strong, D. T., Sale, P. W. G., and Helyar, K. R. (1997). Initial soil pH affects the pH at which nitrification ceases due to self-induced acidification of microbial microsites. Aust. J. Soil Res. 35, 565–570. doi: 10.1071/S96055
Su, C., Lei, L., Duan, Y., Zhang, K.-Q., and Yang, J. (2012). Culture-independent methods for studying environmental microorganisms: methods, application, and perspective. Appl. Microbiol. Biotechnol. 93, 993–1003. doi: 10.1007/s00253-011-3800-7
Sukop, M. C., and Thorne, D. T. (2006). Lattice Boltzmann Modeling: An Introduction for Geoscientists and Engineers. Berlin: Springer Verlag.
Svensson, V., Natarajan, K. N., Ly, L.-H., Miragaia, R. J., Labalette, C., Macaulay, I. C., et al. (2017). Power analysis of single-cell RNA-sequencing experiments. Nat. Methods 14, 381–387. doi: 10.1038/nmeth.4220
Taina, I. A., Heck, R. J., and Elliot, T. R. (2008). Application of X-ray computed tomography to soil science: a literature review. Can. J. Soil Sci. 88, 1–20. doi: 10.4141/CJSS06027
Tartakovsky, A. M., Meakin, P., Scheibe, R. D., and Wood, B. D. (2007). A smoothed particle hydrodynamics model for reactive transport and mineral precipitation in porous, and fractured porous media. Water Resour. Res. 43:W05437. doi: 10.1029/2005WR004770
Tecon, R., and Or, D. (2017a). Biophysical processes supporting the diversity of microbial life in soil. FEMS Microbiol. Rev. 41, 599–623. doi: 10.1093/femsre/fux039
Tecon, R., and Or, D. (2017b). Cooperation in carbon source degradation shapes spatial self-organization of microbial consortia on hydrated surfaces. Sci. Rep. 7:43726. doi: 10.1038/srep43726
Terrat, S., Christen, R., Dequiedt, S., Lelievre, M., Nowak, V., Regnier, T., et al. (2012). Molecular biomass and MetaTaxogenomic assessment of soil microbial communities as influenced by soil DNA extraction procedure. Microb. Biotechnol. 5, 135–141. doi: 10.1111/j.1751-7915.2011.00307.x
Terrat, S., Horrigue, W., Dequietd, S., Saby, N. P. A., Lelievre, M., Nowak, V., et al. (2017). Mapping and predictive variations of soil bacterial richness across France. PLoS One 12:e0186766. doi: 10.1371/journal.pone.0186766
Thomas, G. W. (1992). In defense of observations and measurements. Soil Sci. Soc. Am. J. 56, 1979. doi: 10.2136/sssaj1992.03615995005600060054x
Thoreau, H. D. (1854). Walden: Life in the Woods. Boston, MA: Ticknor and Fields. doi: 10.5962/bhl.title.146169
Thullner, M., and Baveye, P. (2008). Computational pore network modeling of the influence of biofilm permeability on bioclogging in porous media. Biotechnol. Bioeng. 99, 1337–1351. doi: 10.1002/bit.21708
Tian, J., Pausch, J., Yu, G., Blagodatskaya, E., Gao, Y., and Kuzyakov, Y. (2015). Aggregate size and their disruption affect C-14-labeled glucose mineralization and priming effect. Appl. Soil Ecol. 90, 1–10. doi: 10.1016/j.apsoil.2015.01.014
Tiedje, J. M., Cho, J. C., Murray, A., Treves, D., Xia, B., and Zhou, J. (2001). “Soil teeming with life: new frontiers for soil science,” in Sustainable Management of Soil Organic Matter, eds R. M. Rees, B. C. Ball, and C. A. Watson (Wallingford: CAB International), 393–412. doi: 10.1079/9780851994659.0393
Tiedje, J. M., Sexstone, A. J., Parkin, T. B., Revsbech, N. P., and Shelton, D. R. (1984). Anaerobic processes in soils. Plant Soil 76, 197–212. doi: 10.1007/BF02205580
Tippkötter, R., Ritz, K., and Darbyshire, J. F. (1986). The preparation of soil thin sections for biological studies. J. Soil Sci. 37, 681–690. doi: 10.1111/j.1365-2389.1986.tb00396.x
Tippkötter, R., Eickhorst, T., Taubner, H., Gredner, B., and Rademaker, G. (2009). Detection of soil water in macropores of undisturbed soil using microfocus X- ray tube computerized tomography (CT). Soil Tillage Res. 105, 12–20. doi: 10.1016/j.still.2009.05.001
Tisdall, J. M., and Oades, J. M. (1982). Organic matter and water-stable aggregates in soils. J. Soil Sci. 33, 141–163. doi: 10.1111/j.1365-2389.1982.tb01755.x
Topp, G. C. (1971). Soil-water hysteresis: domain theory extended to pore interaction conditions. Soil Sci. Soc. Am. Proc. 35, 219–225. doi: 10.2136/sssaj1971.03615995003500020017x
Topp, G. C., and Miller, E. E. (1966). Hysteretic moisture characteristics and hydraulic conductivities for glass-bead media. Soil Sci. Soc. Am. Proc. 30, 156–162. doi: 10.2136/sssaj1966.03615995003000020008x
Torrance, J., Elliot, T. R., Martin, R., and Heck, R. (2008). X-ray computed tomography of frozen soil. Cold Regions Sci. Technol. 53, 75–82. doi: 10.1016/j.coldregions.2007.04.010
Torre, I. G., Losada, J. C., Heck, R. J., and Tarquis, A. M. (2018). Multifractal analysis of 3D images of tillage soil. Geoderma 311, 167–174. doi: 10.1016/j.geoderma.2017.02.013
Totsche, K. U., Amelung, W., Gerzabek Martin, H., Guggenberger, G., Klumpp, E., Knief, C., et al. (2017). Microaggregates in soils. J. Plant Nutr. Soil Sci. 181, 104–136. doi: 10.1002/jpln.201600451
Tötzke, C., Kardjilov, N., Manke, I., and Oswald, S. E. (2017). Capturing 3D water flow in rooted soil by ultra-fast neutron tomography. Sci. Rep. 7:6192. doi: 10.1038/s41598-017-06046-w
Tracy, S. R., Daly, K. R., Sturrock, C. J., Crout, N. M. J., Mooney, S. J., and Roose, T. (2015). Three-dimensional quantification of soil hydraulic properties using X-ray computed tomography and image-based modeling. Water Resour. Res. 51, 1006–1022. doi: 10.1002/2014WR016020
Treves, D. S., Xia, B., Zhou, J., and Tiedje, J. M. (2003). A two-species test of the hypothesis that spatial isolation influences microbial diversity in soil. Microb. Ecol. 45, 20–28. doi: 10.1007/s00248-002-1044-x
Tuller, M., Kulkarni, R., and Fink, W. (2013). “Segmentation of X-Ray CT data of porous materials: a review of global and locally adaptive algorithms,” in Soil-Water-Root Processes: Advances in Tomography and Imaging, eds S. H. Anderson and J. W. Hopmans, Vol. 61 (Madison, WI: American Society of Agronomy), 157–182.
Tumlinson, L. G., Liu, H., Silk, W. K., and Hopmans, J. W. (2008). Thermal neutron computed tomography of soil water and plant roots. Soil Sci. Soc. Am. J. 72, 1234–1242. doi: 10.2136/sssaj2007.0302
Uroz, S., Kelly, L. C., Turpault, M. P., Lepleux, C., and Frey-Klett, P. (2015). The mineralosphere concept: mineralogical control of the distribution and function of mineral-associated bacterial communities. Trends Microbiol. 23, 751–762. doi: 10.1016/j.tim.2015.10.004
Valdes-Parada, F. J., Porter, M. L., Narayanaswamy, K., Ford, R. M., and Wood, B. D. (2009). Upscaling microbial chemotaxis in porous media. Adv. Water Resour. 32, 1413–1428. doi: 10.1016/j.advwatres.2009.06.010
Valocchi, A. J. (2012). “Hydrogeochemical models,” in Delivery and Mixing in the Subsurface: Processes and Design Principles for In Situ Remediation, eds P. K. Kitanidis and P. L. McCarty (New York, NY: Springer), 77–116. doi: 10.1007/978-1-4614-2239-6_4
Van den Bulcke, J., Boone, M., Van Acker, J., and Van Hoorebeke, L. (2009). Three-dimensional x-ray imaging and analysis of fungi on and in wood. Microsc. Microanal. 15, 395–402. doi: 10.1017/s1431927609990419
van der Wal, A., and de Boer, W. (2017). Dinner in the dark: illuminating drivers of soil organic matter decomposition. Soil Biol. Biochem. 105, 45–48. doi: 10.1016/j.soilbio.2016.11.006
van Elsas, J. D., Costal, R., Jansson, J., Sjoling, S., Bailey, M., Nalin, R., et al. (2008). The metagenomics of disease-suppressive soils – Experiences from the METACONTROL project. Trends Biotechnol. 26, 591–601. doi: 10.1016/j.tibtech.2008.07.004
van Groenigen, J. W., van Kessel, C., Hungate, B. A., Oenema, O., Powlson, D. S., and van Groenigen, K. J. (2017). Sequestering soil organic carbon: a nitrogen dilemma. Environ. Sci. Technol. 51, 4738–4739. doi: 10.1021/acs.est.7b01427
Van Loo, D., Bouckaert, L., Leroux, O., Pauwels, E., Dierick, M., and Van Hoorebeke, L. (2014). Contrast agents for soil investigation with X-ray computed tomography. Geoderma 213, 485–491. doi: 10.1016/j.geoderma.2013.08.036
Vandevivere, P., and Baveye, P. (1992a). Effect of bacterial extracellular polymers on the saturated hydraulic conductivity of sand columns. Appl. Environ. Microbiol. 58, 1690–1698.
Vandevivere, P., and Baveye, P. (1992b). Improved preservation of bacterial exoplolymers for scanning elecrton microscopy. J. Microsc. Oxford 167, 323–330. doi: 10.1111/j.1365-2818.1992.tb03242.x
Vandevivere, P., and Baveye, P. (1992c). Sampling method for the observation of microorganisms in unconsolidated porous media via scanning electron microscopy. Soil Sci. 153, 482–485. doi: 10.1097/00010694-199206000-00007
Vandevivere, P., and Baveye, P. (1992d). Saturated hydraulic conductivity reduction caused by aerobic bacteria in sand columns. Soil Sci. Soc. Am. J. 56, 1–13. doi: 10.2136/sssaj1992.03615995005600010001x
Vandevivere, P., Baveye, P., Delozada, D. S., and Deleo, P. (1995). Microbial clogging of saturated soils and aquifer materials: evaluation of mathematical models. Water Resour. Res. 31, 2173–2180. doi: 10.1029/95wr01568
Vaz, C. M. P., de Maria, I. C., Lasso, P. O., and Tuller, M. (2011). Evaluation of an advanced benchtop micro-computed tomography system for quantifying porosities and pore-size distributions of two brazilian oxisols. Soil Sci. Soc. Am. J. 75, 832–841. doi: 10.2136/sssaj2010.0245
Verbruggen, E., Veresoglou, S. D., Anderson, I. C., Caruso, T., Hammer, E. C., Kohler, J., et al. (2012). Arbuscular mycorrhizal fungi – Short-term liability but long-term benefits for soil carbon storage? New Phytol. 197, 366–368. doi: 10.1111/nph.12079
Veres, M., Lacey, G., and Taylor, G. W. (2015). Deep learning architectures for soil property prediction. Paper Presented at the 12th Conference on Computer and Robot Vision, Halifax, NS, Canada. doi: 10.1109/CRV.2015.15
Vidal, A., Hirte, J., Bender, S. F., Mayer, J., Gattinger, A., Höschen, C., et al. (2018). Linking 3D structure and plant-microbe-soil carbon transfer in the rhizosphere. Front. Environ. Sci. 6:9. doi: 10.3389/fenvs.2018.00009
Villemin, G., Mansot, J. L., Watteau, F., Ghanbaja, J., and Toutain, F. (1995). Study of the biodegradation and humification of soil organic matter of plant origin by Electron Energy Loss Spectroscopy (EELS): distribution of carbon, nitrogen and C/N evaluation at ultrastructural level in situ. Comptes Ren. Acad. Sci. Série 2. Sci. Terre Planèt. 321, 861–868.
Vogel, H. J. (1997). Morphological determination of pore connectivity as a function of pore size using serial sections. Eur. J. Soil Sci. 48, 365–377. doi: 10.1046/j.1365-2389.1997.00096.x
Vogel, H. J., Bartke, S., Daedlow, K., Helming, K., Kögel-Knabner, I., Lang, B., et al. (2018). A systemic approach for modeling soil functions. Soil 4, 83–92. doi: 10.5194/soil-4-83-2018
Vogel, H.-J., Cousin, I., and Roth, K. (2002). Quantification of pore structure and gas diffusion as a function of scale. Eur. J. Soil Sci. 53, 465–473. doi: 10.1046/j.1365-2389.2002.00457.x
Vogel, H. J., and Ippisch, O. (2008). Estimation of a critical spatial discretization limit for solving Richards’ equation at large scales. Vadose Zone J. 7, 112–114. doi: 10.2136/vzj2006.0182
Vogel, H. J., and Roth, K. (2001). Quantitative morphology and network representation of soil pore structure. Adv. Water Resour. 24, 233–242. doi: 10.1016/S0309-1708(00)00055-5
Vogel, H. J., Weller, U., and Schlüter, S. (2010). Quantification of soil structure based on Minkowski functions. Comput. Geosci. 36, 1236–1245. doi: 10.1016/j.cageo.2010.03.007
Vogel, L. E., Makowski, D., Garnier, P., Vieuble-Gonod, L., Coquet, Y., Raynaud, X., et al. (2015). Modeling the effect of soil meso- and macropores topology on the biodegradation of a soluble carbon substrate. Adv. Water Resour. 83, 123–136. doi: 10.1016/j.advwatres.2015.05.020
Vogel, L. E., Pot, V., Makowski, D., Garnier, P., and Baveye, P. C. (2018). To what extent do uncertainty and sensitivity analyses help unravel the influence of microscale physical and biological drivers in soil carbon dynamics models? Ecol. Modell. 383, 10–22. doi: 10.1016/j.ecolmodel.2018.05.007
Vogel, T. M., Simonet, P., Jansson, J. K., Hirsch, P. R., Tiedje, J. M., van Elsas, J. D., et al. (2009). TerraGenome: a consortium for the sequencing of a soil metagenome. Nat. Rev. Microbiol. 7, 252–252. doi: 10.1038/nrmicro2119
Vogt, S. J., Sanderlin, A. B., Seymour, J. D., and Codd, S. L. (2013). Permeability of a growing biofilm in a porous media fluid flow analyzed by magnetic resonance displacement-relaxation correlations. Biotechnol. Bioeng. 110, 1366–1375. doi: 10.1002/bit.24803
Voltolini, M., Tas, N., Wang, S., Brodie, E. L., and Ajo-Franklin, J. B. (2017). Quantitative characterization of soil micro-aggregates: new opportunities from sub-micron resolution synchrotron X-ray microtomography. Geoderma 305, 382–393. doi: 10.1016/j.geoderma.2017.06.005
Vos, M., Wolf, A. B., Jennings, S. J., and Kowalchuk, G. A. (2013). Micro-scale determinants of bacterial diversity in soil. FEMS Microbiol. Rev. 37, 936–954. doi: 10.1111/1574-6976.12023
Votrubová, J., Císlerová, M., Amin, M. H. G., and Hall, L. D. (2003). Recurrent ponded infiltration into structured soil: a magnetic resonance imaging study. Water Resour. Res. 39:1371. doi: 10.1029/2003WR002222
Wachinger, G., Fiedler, S., Zepp, K., Gattinger, A., Sommer, M., and Roth, K. (2000). Variability of soil methane production on the micro-scale: spatial association with hot spots of organic material and archaeal populations. Soil Biol. Biochem. 32, 1121–1130. doi: 10.1016/s0038-0717(00)00024-9
Wagner, A. O., Praeg, N., Reitschuler, C., and Illmer, P. (2015). Effect of DNA extraction procedure, repeated extraction and ethidium monoazide (EMA)/propidium monoazide (PMA) treatment on overall DNA yield and impact on microbial fingerprints for bacteria, fungi and archaea in a reference soil. Appl. Soil Ecol. 93, 56–64. doi: 10.1016/j.apsoil.2015.04.005
Walsh, S. D. C., Burwinkle, H., and Saar, M. O. (2009). A new partial-bounceback lattice-Boltzmann method for fluid flow through heterogeneous media. Comput. Geosci. 35, 1186–1193. doi: 10.1016/j.cageo.2008.05.004
Walsh, S. D. C., and Saar, M. O. (2013). Developing extensible Lattice-Boltzmann simulators for general-purpose graphics-processing units. Commun. Comput Phys. 13, 867–879. doi: 10.4208/cicp.351011.260112s
Wan, J., Tyliszczak, T., and Tokunaga, T. K. (2007). Organic carbon distribution, speciation, and elemental correlations within soil micro aggregates: applications of STXM and NEXAFS spectroscopy. Geochim. Cosmochim. Acta 71, 5439–5449. doi: 10.1016/j.gca.2007.07.030
Wang, D., and Bodovitz, S. (2010). Single cell analysis: the new frontier in ‘omics’. Trends Biotechnol. 28, 281–290. doi: 10.1016/j.tibtech.2010.03.002
Wang, G., and Or, D. (2010). Aqueous films limit bacterial cell motility and colony expansion on partially saturated rough surfaces. Environ. Microbiol. 12, 1363–1373. doi: 10.1111/j.1462-2920.2010.02180.x
Wang, J., Guo, L., Bai, Z., and Yang, L. (2016). Using computed tomography (CT) images and multi-fractal theory to quantify the pore distribution of reconstructed soils during ecological restoration in opencast coal-mine. Ecol. Eng. 92, 148–157. doi: 10.1016/j.ecoleng.2016.03.029
Wanzek, T., Keiluweit, M., Baham, J., Dragila, M. I., Fendorf, S., Fiedler, S., et al. (2018). Quantifying biogeochemical heterogeneity in soil systems. Geoderma 324, 89–97. doi: 10.1016/j.geoderma.2018.03.003
Warmink, J. A., Nazir, R., Corten, B., and van Elsas, J. D. (2011). Hitchhikers on the fungal highway: the helper effect for bacterial migration via fungal hyphae. Soil Biol. Biochem. 43, 760–765. doi: 10.1016/j.soilbio.2010.12.009
Warner, G. S., Nieber, J. L., Moore, I. D., and Geise, R. A. (1989). Characterizing macropores in soil by computed tomography. Soil Sci. Soc. Am. J. 53, 653–660. doi: 10.2136/sssaj1989.03615995005300030001x
Watrous, J. D., and Dorrestein, P. C. (2011). Imaging mass spectrometry in microbiology. Nat. Rev. Microbiol. 9, 683–694. doi: 10.1038/nrmicro2634
Wenz, J., Schleede, S., Khrennikov, K., Bech, M., Thibault, P., Heigoldt, M., et al. (2015). Quantitative X-ray phase-contrast microtomography from a compact laser-driven betatron source. Nat. Commun. 6:7568. doi: 10.1038/ncomms8568
Wertz, S., Degrange, V., Prosser, J. I., Poly, F., Commeaux, C., Freitag, T., et al. (2006). Maintenance of soil functioning following erosion of microbial diversity. Environ. Microbiol. 8, 2162–2169. doi: 10.1111/j.1462-2920.2006.01098.x
Wertz, S., Degrange, V., Prosser, J. I., Poly, F., Commeaux, C., Guillaumaud, N., et al. (2007). Decline of soil microbial diversity does not influence the resistance and resilience of key soil microbial functional groups following a model disturbance. Environ. Microbiol. 9, 2211–2219. doi: 10.1111/j.1462-2920.2007.01335.x
West, B. A., Hodgdon, T. S., Parno, M. D., and Song, A. J. (2018). Improved workflow for unguided multiphase image segmentation. Comput. Geosci. 118, 91–99. doi: 10.1016/j.cageo.2018.05.013
Weyens, N., van der Lelie, D., Taghavi, S., Newman, L., and Vangronsveld, J. (2009). Exploiting plant–microbe partnerships to improve biomass production and remediation. Trends Biotechnol. 27, 591–598. doi: 10.1016/j.tibtech.2009.07.006
White, D., FitzPatrick, E. A., and Kilham, K. (1994). Use of stained bacterial inocula to assess spatial distribution after introduction into soil. Geoderma 63, 245–254. doi: 10.1016/0016-7061(94)90066-3
White, R. E., Davidson, B., Lam, S. K., and Chen, D. L. (2018). A critique of the paper ‘Soil carbon 4 per mille’ by Minasny et al. (2017). Geoderma 309, 115–117. doi: 10.1016/j.geoderma.2017.05.025
Whitley, K. D., Comstock, M. J., and Chemla, Y. R. (2017). “High-resolution “fleezers”: dual-trap optical tweezers combined with single-molecule fluorescence detection,” in Optical Tweezers: Methods and Protocols, ed. A. Gennerich (New York, NY: Springer), 183–256.
Widdowson, M. A., Molz Fred, J., and Benefield Larry, D. (1988). A numerical transport model for oxygen-and nitrate-based respiration linked to substrate and nutrient availability in porous media. Water Resour. Res. 24, 1553–1565. doi: 10.1029/WR024i009p01553
Wildenschild, D., Hopmans, J. W., Vaz, C. M. P., Rivers, M. L., Rikard, D., and Christensen, B. S. B. (2002). Using X-ray computed tomography in hydrology: systems, resolutions, and limitations. J. Hydrol. 267, 285–297. doi: 10.1016/S0022-1694(02)00157-9
Wildenschild, D., and Sheppard, A. P. (2013). X-ray imaging and analysis techniques for quantifying pore-scale structure and processes in subsurface porous medium systems. Adv. Water Resour. 51, 217–246. doi: 10.1016/j.advwatres.2012.07.018
Willcock, S., Martínez-López, J., Hooftman, D. A. P., Bagstad, K. J., Balbi, S., Marzo, A., et al. (2018). Machine learning for ecosystem services. Ecosyst. Ser. (in press). doi: 10.1016/j.ecoser.2018.04.004
Williamson, K. E., Fuhrmann, J. J., Wommack, K. E., and Radosevich, M. (2017). Viruses in soil ecosystems: an unknown quantity within an unexplored territory. Ann. Rev. Virol. 4, 201–219. doi: 10.1146/annurev-virology-101416-041639
Willson, C. S., Lu, N., and Likos, W. J. (2012). Quantification of grain, pore, and fluid microstructure of unsaturatedsand from X-Ray computed tomography images. Geotech. Test J. 35, 1–13. doi: 10.1520/GTJ20120075
Wilmoth, J. L., Doak, P. W., Timm, A., Halsted, M., Anderson, J. D., Ginovart, M., et al. (2018). A microfluidics and agent -based modeling framework for investigating spatial organization in bacterial colonies: the case of Pseudomonas aeruginosa and H1-Type VI secretion interactions. Front. Microbiol. 9:33. doi: 10.3389/fmicb.2018.00033
Woese, C. R. (2004). A new biology for a new century. Microbiol. Mol. Biol. Rev. 68, 173–186. doi: 10.1128/mmbr.00002-09
Wolf, A. B., Vos, M., de Boer, W., and Kowalchuk, G. A. (2013). Impact of matric potential and pore size distribution on growth dynamics of filamentous and non-filamentous soil bacteria. PLoS One 8:e083661. doi: 10.1371/journal.pone.0083661
Wood, B. D. (2009). The role of scaling laws in upscaling. Adv. Water Resour. 32, 723–736. doi: 10.1016/j.advwatres.2008.08.015
Wood, B. D. (2010). Reply to comment by Philippe Baveye on “the role of scaling laws in upscaling.” Adv. Water Resour. 33, 125–127. doi: 10.1016/j.advwatres.2009.11.002
Worrich, A., Stryhanyuk, H., Musat, N., König, S., Banitz, T., Centler, F., et al. (2017). Mycelium-mediated transfer of water and nutrients stimulates bacterial activity in dry and oligotrophic environments. Nat. Commun. 8:15472. doi: 10.1038/ncomms15472
Woyke, T., Doud, D. F. R., and Schulz, F. (2017). The trajectory of microbial single-cell sequencing. Nat. Methods 14, 1045–1054. doi: 10.1038/nmeth.4469
Woyke, T., Tighe, D., Mavromatis, K., Clum, A., Copeland, A., Schackwitz, W., et al. (2010). One bacterial bell, one complete genome. PLoS One 5:e10314. doi: 10.1371/journal.pone.0010314
Wright, D. A., Kilham, K., Glover, L. A., and Prosser, J. L. (1995). Role of pore size location in determining bacterial activity during predation by protozoa in soil. Appl. Environ. Microbiol. 61, 3537–3543.
Xie, C., Wang, J., Wang, D., Pan, N., and Wang, M. (2015). Lattice Boltzmann modeling of thermal conduction in composites with thermal contact resistance. Commun. Comput. Phys. 17, 1037–1055. doi: 10.4208/cicp.2014.m360
Xiong, Q., Baychev, T. G., and Jivkov, A. P. (2016). Review of pore network modelling of porous media: experimental characterisations, network constructions and applications to reactive transport. J. Contam. Hydrol. 192, 101–117. doi: 10.1016/j.jconhyd.2016.07.002
Xun, W. B., Huang, T., Zhao, J., Ran, W., Wang, B. R., Shen, Q. R., et al. (2015). Environmental conditions rather than microbial inoculum composition determine the bacterial composition, microbial biomass and enzymatic activity of reconstructed soil microbial communities. Soil Biol. Biochem. 90, 10–18. doi: 10.1016/j.soilbio.2015.07.018
Yang, W., and Yuste, R. (2017). In vivo imaging of neural activity. Nature Methods 14, 349–359. doi: 10.1038/nmeth.4230
Yang, X., Mehmani, Y., Perkins, W. A., Pasquali, A., Schoenherr, M., Kim, K., et al. (2016). Intercomparison of 3D pore-scale flow and solute transport simulation methods. Adv. Water Resour. 95, 176–189. doi: 10.1016/j.advwatres.2015.09.015
Yehya, A., Naji, H., and Sukop, M. C. (2015). Simulating flows in multi-layered and spatially-variable permeability media via a new Gray Lattice Boltzmann model. Comput. Geotech. 70, 150–158. doi: 10.1016/j.compgeo.2015.07.017
Yilmaz, S., and Singh, A. K. (2012). Single cell genome sequencing. Curr. Opin. Biotechnol. 23, 437–443. doi: 10.1016/j.copbio.2011.11.018
Yoon, H., Kang, Q., and Valocchi, A. J. (2015). Lattice-Boltzmann-based approaches for pore-scale reactive transport. Rev. Mineral. Geochem. 80, 393–431. doi: 10.2138/rmg.2015.80.12
Yoon, H., Valocchi Albert, J., Werth Charles, J., and Dewers, T. (2012). Pore-scale simulation of mixing-induced calcium carbonate precipitation and dissolution in a microfluidic pore network. Water Resour. Res. 48:W02524. doi: 10.1029/2011WR011192
Yoshida, H., and Hayashi, H. (2014). Transmission-reflection coefficient in the lattice boltzmann method. J. Statist. Phys. 155, 277–299. doi: 10.1007/s10955-014-0953-7
Young, I. M., and Crawford, J. W. (2004). Interactions and self-organization in the soil-microbe complex. Science 304, 1634–1637. doi: 10.1126/science.1097394
Young, I. M., Crawford, J. W., Nunan, N., Otten, W., and Spiers, A. (2008). “Microbial distribution in soils: physics and scaling,” in Advances in Agronomy, Vol. 100, ed. D. L. Sparks (Amsterdam: Elsevier), 81–121.
Young, I. M., Crawford, J. W., and Rappoldt, C. (2001). New methods and models for characterising structural heterogeneity of soil. Soil Tillage Res. 61, 33–45. doi: 10.1016/s0167-1987(01)00188-x
Yu, X., Fu, Y., and Lu, S. (2017). Characterization of the pore structure and cementing substances of soil aggregates by a combination of synchrotron radiation X-ray micro-computed tomography and scanning electron microscopy. Eur. J. Soil Sci. 68, 66–79. doi: 10.1111/ejss.12399
Zalzale, M., McDonald, P. J., and Scrivener, K. L. (2013). A 3D lattice Boltzmann effective media study: understanding the role of C-S-H and water saturation on the permeability of cement paste. Modell. Simulat. Mater. Sci. Eng. 21, doi: 10.1088/0965-0393/21/8/085016
Zamir, A., Hagen, C., Diemoz, P. C., Endrizzi, M., Vittoria, F., Chen, Y., et al. (2017). Recent advances in edge illumination x-ray phase-contrast tomography. J. Med. imAging (Bellingham, Wash.) 4:040901. doi: 10.1117/1.jmi.4.4.040901
Zhang, X., Crawford, J. W., and Young, I. M. (2016). A Lattice Boltzmann model for simulating water flow at pore scale in unsaturated soils. J. Hydrol. 538, 152–160. doi: 10.1016/j.jhydrol.2016.04.013
Zhao, D., Xu, M., Liu, G., Yao, X., Tuo, D., Zhang, R., et al. (2017). Quantification of soil aggregate microstructure on abandoned cropland during vegetative succession using synchrotron radiation-based micro-computed tomography. Soil Tillage Res. 165, 239–246. doi: 10.1016/j.still.2016.08.007
Keywords: soil microbiology, biodiversity, upscaling, tomography, X-ray computed, NanoSIMS imaging, single-cell genomics
Citation: Baveye PC, Otten W, Kravchenko A, Balseiro-Romero M, Beckers É, Chalhoub M, Darnault C, Eickhorst T, Garnier P, Hapca S, Kiranyaz S, Monga O, Mueller CW, Nunan N, Pot V, Schlüter S, Schmidt H and Vogel H-J (2018) Emergent Properties of Microbial Activity in Heterogeneous Soil Microenvironments: Different Research Approaches Are Slowly Converging, Yet Major Challenges Remain. Front. Microbiol. 9:1929. doi: 10.3389/fmicb.2018.01929
Received: 27 March 2018; Accepted: 30 July 2018;
Published: 27 August 2018.
Edited by:
Eoin L. Brodie, Lawrence Berkeley National Laboratory (LBNL), United StatesReviewed by:
James J. Moran, Pacific Northwest National Laboratory (DOE), United StatesCopyright © 2018 Baveye, Otten, Kravchenko, Balseiro-Romero, Beckers, Chalhoub, Darnault, Eickhorst, Garnier, Hapca, Kiranyaz, Monga, Mueller, Nunan, Pot, Schlüter, Schmidt and Vogel. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Philippe C. Baveye, YmF2ZXllLnJwaUBnbWFpbC5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.