 Oriana Flores1
Oriana Flores1 Camila Prince1
Camila Prince1 Mauricio Nuñez1Alejandro Vallejos2Claudia Mardones2
Mauricio Nuñez1Alejandro Vallejos2Claudia Mardones2 Carolina Yañez1
Carolina Yañez1 Ximena Besoain3
Ximena Besoain3 Roberto Bastías1*
Roberto Bastías1*- 1Laboratorio de Microbiología, Instituto de Biología, Facultad de Ciencias, Pontificia Universidad Católica de Valparaíso, Valparaíso, Chile
- 2Departamento de Análisis Instrumental, Facultad de Farmacia, Universidad de Concepción, Concepción, Chile
- 3Laboratorio de Fitopatología, Escuela de Agronomía, Pontificia Universidad Católica de Valparaíso, Valparaíso, Chile
In recent years, Chilean kiwifruit production has been affected by the phytopathogen Pseudomonas syringae pv. actinidiae (Psa), which has caused losses to the industry. In this study, we report the genotypic and phenotypic characterization of 18 Psa isolates obtained from Chilean kiwifruits orchards between 2012 and 2016 from different geographic origins. Genetic analysis by multilocus sequence analysis (MLSA) using four housekeeping genes (gyrB, rpoD, gltA, and gapA) and the identification of type III effector genes suggest that the Chilean Psa isolates belong to the Psa Biovar 3 cluster. All of the isolates were highly homogenous in regard to their phenotypic characteristics. None of the isolates were able to form biofilms over solid plastic surfaces. However, all of the isolates formed cellular aggregates in the air–liquid interface. All of the isolates, except for Psa 889, demonstrated swimming motility, while only isolate Psa 510 demonstrated swarming motility. The biochemical profiles of the isolates revealed differences in 22% of the tests in at least one Psa isolate when analyzed with the BIOLOG system. Interestingly, all of the isolates were able to produce indole using a tryptophan-dependent pathway. PCR analysis revealed the presence of the genes aldA/aldB and iaaL/matE, which are associated with the production of indole-3-acetic acid (IAA) and indole-3-acetyl-3-L-lysine (IAA-Lys), respectively, in P. syringae. In addition, IAA was detected in the cell free supernatant of a representative Chilean Psa strain. This work represents the most extensive analysis in terms of the time and geographic origin of Chilean Psa isolates. To our knowledge, this is the first report of Psa being able to produce IAA. Further studies are needed to determine the potential role of IAA in the virulence of Psa during kiwifruit infections and whether this feature is observed in other Psa biovars.
Introduction
Pseudomonas syringae pv. actinidiae (Psa) is the causal agent of bacterial canker in Actinidia deliciosa and Actinidia chinensis that has caused severe losses in all of the major areas of kiwifruit cultivation, including Italy, China, New Zealand, and Chile (Scortichini et al., 2012; Ferrante and Scortichini, 2015). This bacterium infects host plants by entering natural openings and wounds, moving inside the plant, and promoting the appearance of necrotic leaf spots, red exudate production, and canker and necrosis in the trunk. In the late stages of the infection, the plants wilt and desiccate which leads to the death of the kiwifruit vine (Vanneste et al., 2012; Cellini et al., 2014). Since its identification in Japan in 1984, successive outbreaks of Psa have been observed worldwide, and therefore it is now considered to be a pandemic phytopathogen (Scortichini et al., 2012; McCann et al., 2017). Comparative analysis using multilocus sequence analysis (MLSA), the detection of type III secretion system effector genes and phytotoxins (phaseolotoxin or coronatine) in Psa isolates from different geographic origins have revealed the existence of five clusters of biovars (Marcelletti et al., 2011; Ciarroni et al., 2015; Ferrante and Scortichini, 2015; Fujikawa and Sawada, 2016; McCann et al., 2017): biovar 1, comprising Japanese strains which are able to produce phaseolotoxin; biovar 2, including only South Korean strains which produce coronatine; biovar 3 or Psa-V, which includes the most virulent strains that are characterized by not producing phytotoxins and were first isolated in Italy (2008–2009) and have been subsequently reported to cause outbreaks in different countries (Butler et al., 2013; Ciarroni et al., 2015; Cunty et al., 2015a); biovar 4, contain strains with low virulence and was recently proposed to be a new pathovar called P. syringae pv. actinidifoliorum (Psaf) (Abelleira et al., 2015; Cunty et al., 2015b); and finally, biovar 5 with Japanese strains isolated in 2012 which do not produce phytotoxins. Recently, a potential new biovar was described in Japan, which produces both phaseolotoxin and coronatine (Fujikawa and Sawada, 2016).
The genetic analysis of the Psa biovars described a set of genes that participate in distinct phases of kiwifruit infection and niche colonization, both outside and inside of the host plant. These genes are related to bacterial motility, biofilm formation, copper and antibiotic resistance, siderophore production, and the degradation of lignin (Marcelletti et al., 2011; Scortichini et al., 2012; Ghods et al., 2015; Gao et al., 2016; Colombi et al., 2017; Patel et al., 2017). However, the mechanisms that determine infection and the interactions between Psa with the kiwifruit plant remain unknown. The production of the phytohormone indole-3-acetic acid (IAA) is another virulence factor that has been described in Pseudomonas savastanoi and P. syringae pathovars. This compound can perturb the regulation of the hormone balance in the plant and increase its susceptibility to infection (Glickmann et al., 1998; Cerboneschi et al., 2016). IAA production using the indole-3-acetamide (IAM) pathway is the most common mechanism in phytopathogenic bacteria, including P. syringae, and has mostly been characterized in P. savastanoi pv. savastanoi (Psav) (Baltrus et al., 2011; Aragón et al., 2014) where IAA biosynthesis begins from L-tryptophan (Trp) and involves the activity of the enzymes tryptophan-2-monooxygenase (IaaM) and IAM hydrolase (IaaH) encoded by the iaaM and iaaH genes, respectively. However, in other P. syringae pathovars, the IAA production involves other genes that lack homology to iaaM and iaaH (Glickmann et al., 1998), and recently aldehyde dehydrogenase family proteins encoded by genes aldA and aldB, were associated with IAA synthesis in P. syringae pv. tomato (McClerklin et al., 2018). For instance, P. savastanoi pv. nerii can conjugate IAA to the amino acid lysine producing indole-3-acetyl-3-L-lysine (IAA-Lys) due to the action of the enzyme IAA-Lys ligase encoded by the iaaL gene (Cerboneschi et al., 2016). This gene has been found in several P. syringae pathovars where it is arranged in synteny with the gene matE, which encodes a putative MATE family transporter, and has been implicated in the fitness and virulence of P. syringae pv. tomato (Pst) in tomato plants (Glickmann et al., 1998; Castillo-Lizardo et al., 2015).
Pseudomonas syringae pv. actinidiae was first reported in Chile in 2010 following its isolation from kiwifruit orchards in the Maule Region, and since 2011, it has been considered to be a pest under the official control of the Agricultural and Livestock Service (SAG) of the Government of Chile (McCann et al., 2013). Previous studies included classifying the first Chilean Psa isolates in biovar 3 together with strains from China, Europe, and New Zealand (Butler et al., 2013; McCann et al., 2013; Cunty et al., 2015a). However, the scope of these studies was limited by the number of Chilean strains. In this study, we report the genotypic and phenotypic characterization of Chilean Psa isolates obtained between 2012 and 2016 from the regions that accumulate more than 80% of the Psa-positive orchards in Chile. In addition, we show the first evidence of Psa strains producing IAA.
Materials and Methods
Bacterial Strains and Culture Conditions
Chilean Psa isolates are listed in Table 1 and were obtained from the SAG from kiwi orchards of different geographic areas in the central-south of Chile in 2012, 2013, and 2016. P. syringae pv. tomato DC3000 was provided by Dr. Paula Salinas of the Universidad Santo Tomás (Santiago, Chile). Escherichia coli DH5α, E. coli K12, Pseudomonas aeruginosa PAO1, Azospirillum brasilense SP7, Salmonella bongori X9617, and Cupriavidus metallidurans CH34 were obtained from the bacterial collection of the Laboratory of Microbiology of the Pontificia Universidad Católica de Valparaíso (PUCV). Pseudomonas antarctica S63 (Vásquez-Ponce et al., 2018) was provided by Dr. Jorge Olivares from the PUCV. The bacteria were grown at 25°C in Luria-Bertani (LB) medium except when another medium is specified. Growth curve were performed in 96 multi-well plates at 25°C during 30 h in a microplate spectrophotometer Infinite® M200 NanoQuant (TECAN). Optical density (OD600 nm) was determined each 30 min. All curves were performed in biological triplicates.
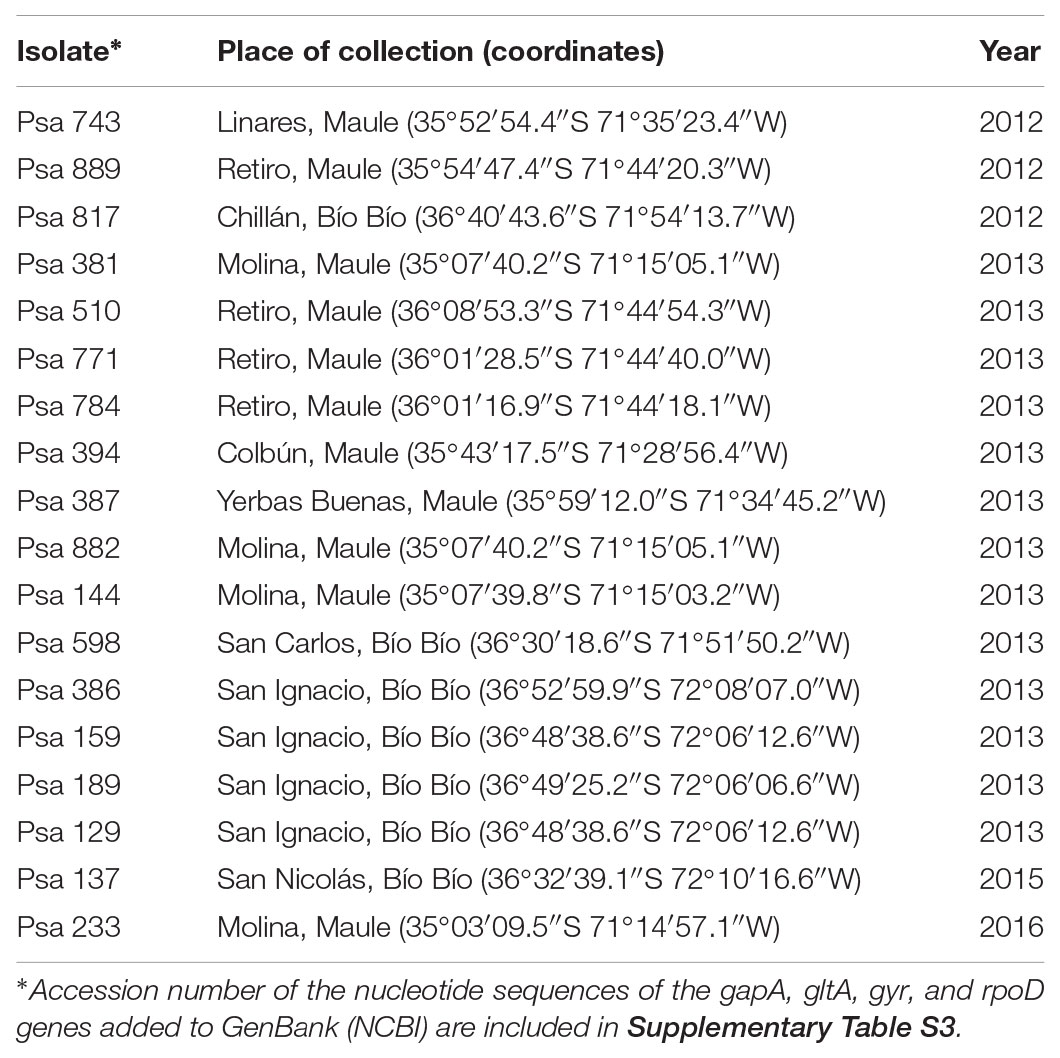
TABLE 1. Chilean Psa isolates used in this work.
Molecular Identification and Characterization of the Psa Isolates
Pseudomonas syringae pv. actinidiae strain molecular identification was performed using RG-PCR and duplex-PCR as previously described (Rees-George et al., 2010; Gallelli et al., 2011). For RG-PCR, specific primers were used to amplify the internal transcribed spacer (ITS) between the 16S and 23S rRNA sequences, and for duplex-PCR, specific primers against ompP1 (Outer Membrane Protein P1) and avrD1 (effector) genes were used. All 18 isolates amplified produced bands of the expected size (Supplementary Figure S1). In addition, the identity of these isolates was also confirmed by partial 16S rDNA sequences. For genomic DNA isolation, the bacteria were grown in LB media for 16 h until the stationary phase. Total genomic DNA was extracted using a Wizard® Genomic DNA Purification Kit (Promega) according to the manufacturer’s instructions. The DNA concentration was determined using MaestroNano MN-913 (Maestrogen, Inc.). For the molecular identification of the type III effector genes, the reference genome of Psa ICMP 18884 biovar 3 strain (GenBank accession number: NZ_CP011972.2) (Templeton et al., 2015) and contigs of the Chilean Psa genomes, ICMP 19439 (ANJM00000000.1) and ICMP 19455 (ANJK00000000.1), available in GenBank (NCBI) were used to design specific primers for the PCRs. Comparative sequence analysis was performed using the Geneious R11 software (Kearse et al., 2012). The amplicons of effector genes obtained from strain Psa 743 were purified using an E.Z.N.A.® Cycle Pure Kit (Omega Bio-Tek, Inc.) and sequenced using the Sanger method by Macrogen, Inc. (South Korea). The quality and assembly of the sequences were analyzed using Geneious R11 software, which were compared with the NCBI database using BLASTN and BLASTX to identify the genes. Primers and annealing temperatures used in the PCRs are listed in Supplementary Table S1. In all cases, PCR was performed on a SureCycler 8800 Thermal Cycler (Agilent Technologies) using SapphireAmp Fast PCR Master Mix (Takara Bio) according to the manufacturer’s instructions. PCR products were separated using electrophoresis in agarose gel (1.5% agarose in 1× buffer TAE) stained with GelRedTM (Biotium), and the bands were visualized under UV light. PCRs were performed in triplicate. The genomic DNA of P. syringae pv. tomato DC3000 and E. coli DH5α were used as the control reactions. The sequences of the effector genes of a selected strain (Psa 743) were deposited in GenBank (NCBI), and the accession numbers are listed in Supplementary Table S2.
Phylogenetic Analysis by MLSA
The gapA, gltA, gyrB, and rpoD genes, encoding glyceraldehyde-3-phosphate dehydrogenase, citrate synthase, DNA gyrase B, and sigma factor 70, respectively, were amplified from the genomic DNA of Psa isolates using the primers listed in Supplementary Table S1 as previously described (Ferrante and Scortichini, 2010). PCR was performed in triplicate using a SureCycler 8800 Thermal Cycler (Agilent Technologies) with GoTaq G2 Flexi polymerase (Fermentas) according to the manufacturer’s instructions. The PCR products were visualized using electrophoresis in agarose gels and purified using an E.Z.N.A.® Gel Extraction Kit (Omega Bio-Tek, Inc.). The automated sequencing of the amplicons was performed by Macrogen, Inc. (South Korea), and the sequences were analyzed using the Geneious R9 software package (Biomatters Limited) (Kearse et al., 2012). The nucleotide sequences of the gapA, gltA, gyr, and rpoD genes of Chilean Psa strains were added to GenBank (NCBI) and are listed in Supplementary Table S3. The sequences of other Psa biovars available in GenBank (NCBI) were included in the analysis and are listed in Supplementary Table S4. In addition, sequences of P. syringae pv. tomato strain DC3000 were included: gapA (AE016853.1:1415258-1416259), cts (AE016853.1:2414332-2415621), gyrB (AE016853.1:4147-6564), and rpoD (AE016853.1:588846-590696) (Buell et al., 2003). The sequences of each locus were aligned using the CLUSTALW included in the MEGA7 software (Kumar et al., 2016). A dendrogram from four-locus concatenated sequences was generated using neighbor-joining (UPGMA) and 1,000 bootstrap iterations.
Biochemical Characterization
The bacteria were streaked out from a -80°C stock onto LB plates and incubated at 25°C for 48 h. Biochemical patterns were determined using the Biolog GEN III MicroPlateTM system (BiologTM, United States) according to the manufacturer’s instructions. BIOLOG plates were read in an Infinite M200 PRO plate reader, TECAN. Reactions were considered positive if the OD590 nm was greater than 50% of the positive control (∼0.7). Reactions indistinguishable from the negative control and with an OD590 nm below 25% of the positive control (∼0.35) were considered to be negative. Reactions between these two parameters were considered borderline.
Determination of Streptomycin and Copper Susceptibility
The copper and streptomycin susceptibility was determined using the broth microdilution method (Biebl and Pfennig, 1978; Mergeay et al., 1985). Bacterial strains were grown in Tris minimal (for the copper assay) or Mueller–Hinton (for the streptomycin assay) media during 18 h, and the optical density at 600 nm (OD600 nm) was adjusted to 0.7. For the copper susceptibility assays, 10 μL of each bacterial culture were inoculated in Tris minimal agar media (1.5% agar) supplemented with the corresponding copper sulfate concentration (0, 75, 100, 125, 150, 175, 200, 225, 250, 275, and 300 μg/mL). To assess the streptomycin susceptibility, bacterial strains were inoculated in Mueller–Hinton agar media supplemented with the corresponding antibiotic concentration (0, 3.9, 7.8, 15.7, 31.25, 62.5, 125, 250, 500, 1,000, and 2,000 μg/mL). Plates were incubated for 5 days at 25°C, and the bacterial growth was observed. C. metallidurans CH34 and P. antarctica S63 were used as experimental controls (von Rozycki and Nies, 2009; Vásquez-Ponce et al., 2018). All experiments were performed in biological and technical triplicates.
Biofilm Production
Microtiter plate biofilm production was performed and adapted as previously described (Merritt et al., 2011; O’Toole, 2011; Ueda and Saneoka, 2015). Briefly, overnight bacterial cultures were adjusted to an optical density of 0.1 (OD600 nm) and diluted 10-fold. Aliquots (100 μL) of the dilution were added to each well (96-well microtiter plates), and the plates were incubated for 7 days at 25°C. After incubation, the liquid supernatant was removed and the plates were washed with distilled water. The wells were stained with 0.1% violet crystal solution, and the biofilm was solubilized with a 30% acetic acid solution. The biofilm production was quantified spectrophotometrically (550 nm) in a Tecan Infinite M200® microplate reader. For the air–liquid interface biofilm assay, 1 mL of the bacterial dilution was added to each well (12-well plates), and the plates were incubated at 25°C for 96 h. Surface biofilm formation was monitored and photo documented every 24 h. All of the experiments were performed in biological and technical triplicates, and P. aeruginosa PAO1 was used as the positive control (Ghafoor et al., 2011).
Bacterial Motility Assay
Motility assays were adapted for the Psa assays as described by Hosseinidoust et al. (2013). Swimming motility assays were performed by inoculating 2 μL of stationary-phase bacterial culture (OD600 nm∼1.3) into the center of 0.3% LB agar plates. Swarming motility assays were performed utilizing the same procedure except that 0.5% LB agar plates were used. The zone sizes were measured after incubation at 30°C for 72 h. The assays were performed in biological and technical triplicates. E. coli K12 was used as the experimental control (Swiecicki et al., 2013). Statistical analysis was performed using one-way ANOVA and Dunnett’s multiple comparison test with p ≤ 0.05.
Indole Production and Identification of IAA Pathway Genes
The indole production was determined using Salkowski’s method as previously described (Mazzola and White, 1994; Mohite, 2013). Briefly, each strain was grown in LB media supplemented with Trp (2 g/L) and incubated at 25°C for 24 h. After incubation, the bacterial density was measured (OD600 nm), and the cultures were centrifuged at 10,000 rpm for 10 min. Cell-free supernatants were mixed with 0.5 mL of Salkowski’s reagent (12 g of FeCl3 per liter in 7.9 M H2SO4). The mixture was incubated for 30 min at room temperature in the dark, and the absorbance at 530 nm was determined. The concentration of indole in each sample was determined using a standard curve of indoleacetic acid (Sigma) (0–30 μg/mL) (Supplementary Figure S3). IAA concentrations were normalized to the cell density. A. brasilense SP7 (Bar and Okon, 1993) and S. bongori X9617 (De La Rosa Fraile et al., 1980) strains were used as experimental positive and negative controls, respectively. All of the analyses were performed in biological and technical triplicates. Statistical analysis was performed using one-way ANOVA and Dunnett’s multiple comparison test with p ≤ 0.05. The detection of iaaL, matE, iaaH, iaaM, aldA, and aldB genes in the Chilean Psa isolates was performed using specific primers designed on the basis of conserved regions from the sequences of different P. syringae pathovars (Supplementary Table S5). The primers designed are listed in Supplementary Table S1. PCRs were performed on a SureCycler 8800 Thermal Cycler (Agilent Technologies) using a SapphireAmp Fast PCR Master Mix (Takara Bio) according to the manufacturer’s instructions. The PCR conditions were as follows: 5 min at 95°C, followed by 35 cycles of 30 s at 95°C, 30 s at the annealing temperature (Supplementary Table S1), 2 min at 72°C, and a final elongation step of 5 min at 72°. Sanger automated sequencing of the amplicons from Psa 743, Psa 598, and Psa 889 was performed by Macrogen, Inc. (South Korea). The sequences were compared with those in the NCBI database using BLASTN and BLASTX for gene identification. The sequences obtained were deposited in GenBank (NCBI), and the accession numbers are listed in Supplementary Table S2.
LC-ESI-MS/MS Analysis
To detect IAA, Psa strain 743 was grown in minimal media (4.5 g/L KH2PO4, 10.5 g/L K2HPO4, 1 g/L (NH4)2SO4, and 0.5 g/L sodium citrate) supplemented with Trp (2 g/L) and incubated at 25°C for 72 h. After incubation, the bacterial density was measured (OD600 nm), and the cultures were centrifuged at 10,000 rpm for 10 min. The supernatant was filtered (0.22 μm). Methanol and acetic acid were added to the cell-free supernatant at a final concentration of 10 and 0.05%, respectively, and then filtered through a PVDF filter (0.22 μm). At the end, the sample was subjected to LC-ESI-MS/MS analysis using indoleacetic acid and lysine (Sigma) as standards. The analysis was performed using a Shimadzu Nexera HPLC system coupled to a 3200Q TRAP mass spectrometer equipped with a turbo ion spray interface (Applied Biosystems/MDS Sciex, ON, Canada). A Kinetex C18 core shell column (150 mm × 4.6 mm i.d.; 2.6 μm particle size; Kinetex, Phenomenex) protected by a C18 UHPLC Ultra column guard (0.5 μm Porosity × 4, 6 mm. i.d., Phenomenex, United States) was used. The elution gradient was adapted from Matsuda et al. (2005) and consisted of a mixture of methanol:water containing 0.05% acetic acid (methanol gradient: 10–90% in 13 min; 95% from 13.1 to 28 min) at a flow rate of 0.4 mL/min and a column temperature of 30°C. MS was conducted in the positive ion mode during the following conditions: curtain gas (CUR), 10 psi; collision activated dissociation (CAD), medium; ion spray voltage (IS), 4500 V; nebulizer gas (Gas1), 60 psi; turbo gas (Gas2), 40 psi; temperature (TEM), 400°C. The detection was performed using multiple reaction monitoring (MRM). The data obtained were processed using Analyst 1.3 software (Applied Biosystems).
Results
Phylogenetic Analysis and Molecular Characterization of the Chilean Psa Isolates
The 18 Chilean Psa isolates used in this study were collected from kiwi plants with canker disease symptoms by the SAG. These isolates were obtained between 2012 and 2016 from orchards in central-south Chile (Bío Bío and Maule Regions) that is the site of the vast majority of kiwifruit production in the country (Oficina de Estudios y Políticas Agrarias [ODEPA], 2018) and accumulates more than 50% of the Psa-infected orchards in Chile (Figure 1 and Table 1). All of the isolates were confirmed as Psa strains by PCR using different sets of primers (see section “Materials and Methods”).
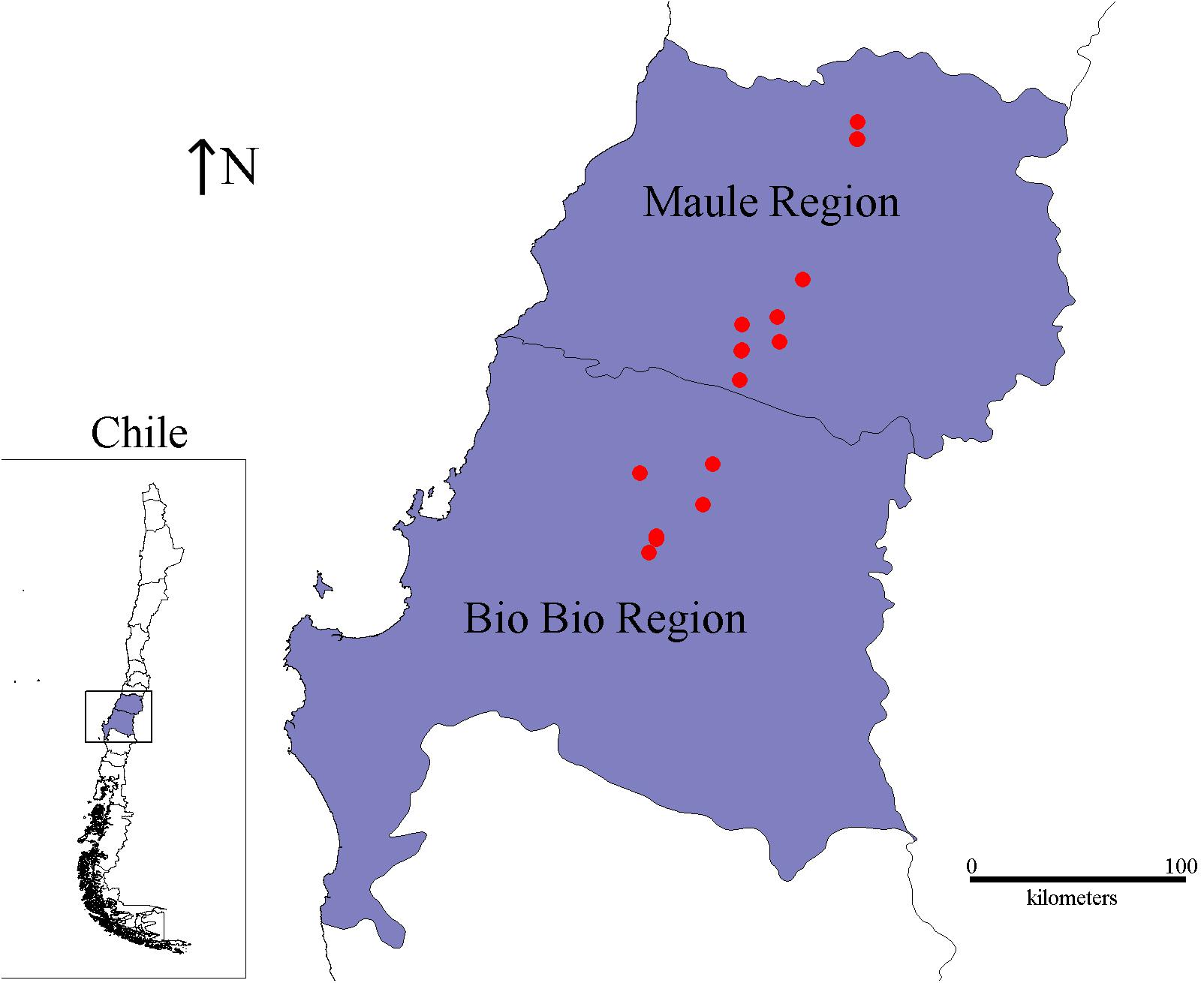
FIGURE 1. Geographic localization of the Chilean Psa strains. The map shows the localization of the Maule and Bío Bío regions. The red spots represent the exact location from where the Chilean Psa strains were isolated.
The first Chilean Psa isolates had been previously assigned to the biovar 3 group (Butler et al., 2013; McCann et al., 2013; Cunty et al., 2015a). An MLSA using the housekeeping genes gyrB (DNA gyrase B), rpoD (sigma factor 70), gltA (citrate synthase), and gapA (glyceraldehyde-3-phosphate dehydrogenase) showed that the genes sequenced have 100% identity with the corresponding genes in different Psa strains belonging to biovar 3, including Chilean strains obtained in 2010. The phylogenetic analysis including other Psa strains shows a clear clustering of different biovars except for biovar 2 and 5 that are grouped together (Figure 2). The results show that all the Chilean Psa isolates group together with the other Psa biovar 3 isolates, confirming the findings of previous studies. These results were also confirmed by the PCR detection of the 16 type III effector genes that have been described in Psa biovar 3 strains (McCann et al., 2013; Ferrante and Scortichini, 2015). Type III effector genes were detected in all of the Chilean Psa strains, including those encoded in plasmid DNA in Psa biovar 3. The identity of these genes was confirmed by sequencing the amplicons of Psa strain 743 as a representative of the other Chilean Psa strains (Supplementary Table S2). These results also suggest that no new biovars have been introduced to Chile during this period.
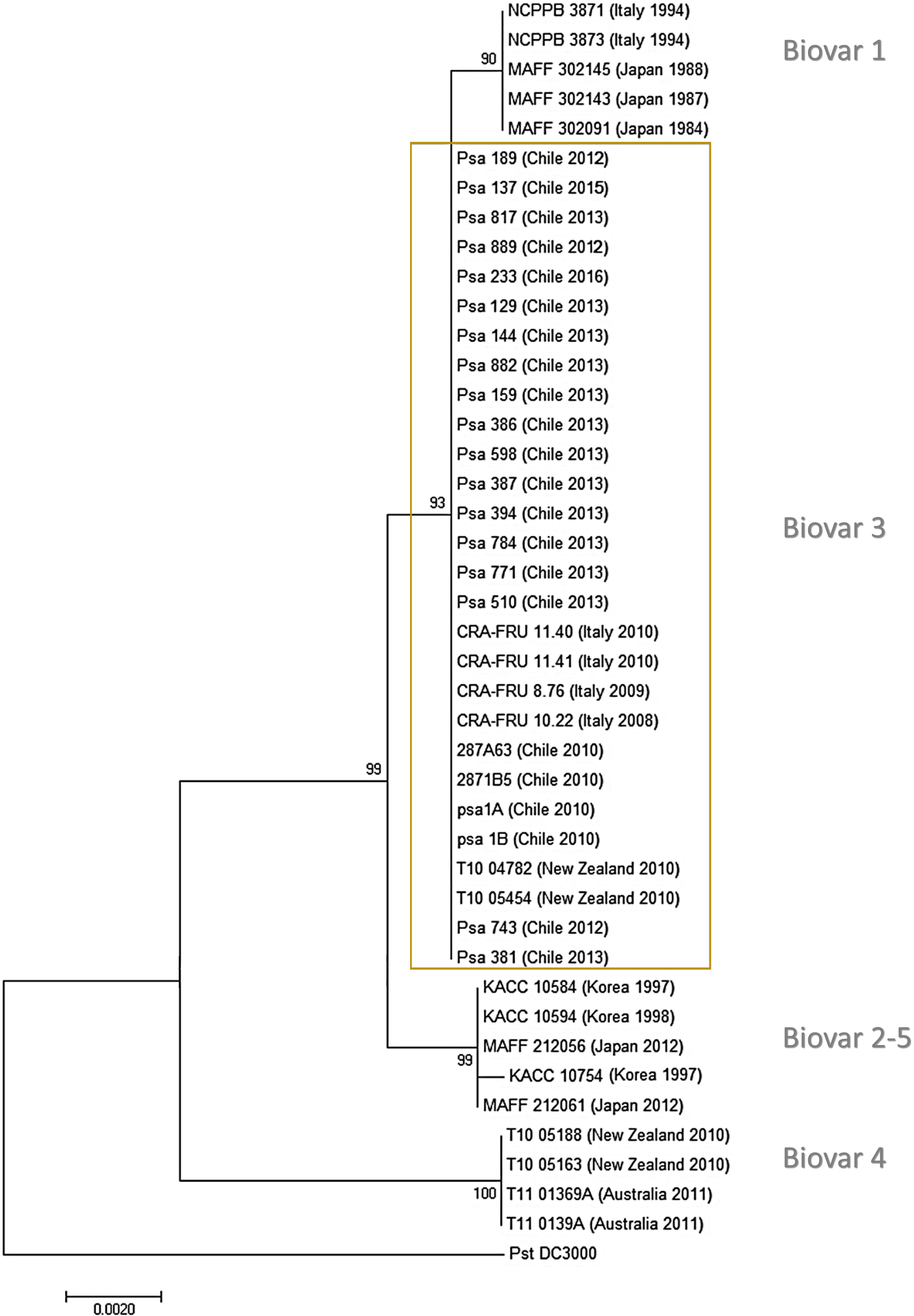
FIGURE 2. Phylogenetic tree of Pseudomonas syringae pv. actinidiae isolates derived from multilocus sequence analysis (MLSA). Phylogenetic tree using the neighbor-joining method (bootstrap: 1,000 replicates) and concatenated sequences of the genes gapA, gltA, gyrB, and rpoB for each isolate. Country, year of isolation, and biovar clade are indicated for each strain.
Phenotypic Characterization of the Psa Isolates
Different features implicated in the fitness and virulence of Psa were evaluated in the 18 Chilean isolates. None of the strains showed differences in their growth parameters (data not shown). However, their biochemical profile determined using the Biolog GEN III MicroPlate revealed differences in 22% of the different tests in at least one of the 18 strains (Supplementary Table S6). All of the strains were able to use different carbon sources such as D-glucose, D-mannose, D-galactose, glycerol, D-mannitol, L-arginine, L-serine, acetic acid, and citric acid. However, they varied in their ability to use sucrose, D-fructose, inosine, L-glutamic acid, and formic acid. Alternatively, all of the strains were resistant to antibiotics such as rifamycin SV, lincomycin, and vancomycin, while they were sensitive to minocycline and troleandomycin and showed variable sensitivity to aztreonam, nalidixic acid, and fusidic acid. Despite these differences, all of the strains were identified as P. syringae pathovars according to the Biolog GEN III database (version 2.8). Interestingly, all of the isolates were susceptible to copper (MIC 75 μg/mL Cu2+) and streptomycin (MIC 3.9 μg/mL), suggesting that no resistance has developed in these strains despite the use of copper compounds as antimicrobials in the Chilean kiwifruit industry.
Biofilm production has been proposed to be an important virulence factor in P. syringae (Ghods et al., 2015; Ueda and Saneoka, 2015). Therefore, the ability to produce biofilm was evaluated in the different Chilean Psa isolates. The results showed that none were able to produce biofilm over an abiotic surface. However, they do produce a thin layer of biofilm (pellicle) in the air–liquid interface. Initially a thin layer of cells was observed in the center of static cultures after 24 h of incubation, turning to a fully grown biofilm after 96 h (Supplementary Figure S2). Swimming and swarming motility was also evaluated among the different Psa isolates. The results show that all of the isolates exhibit swimming motility except for strain Psa 889 which shows a significant reduced displacement in comparison to the other strains (p < 0.05). In contrast, none of the strains except for Psa 510 demonstrated swarming motility under the experimental conditions (p < 0.05) (Figure 3). These results show that the Chilean Psa strains demonstrate a high phenotypic homogeneity with specific differences in particular strains.
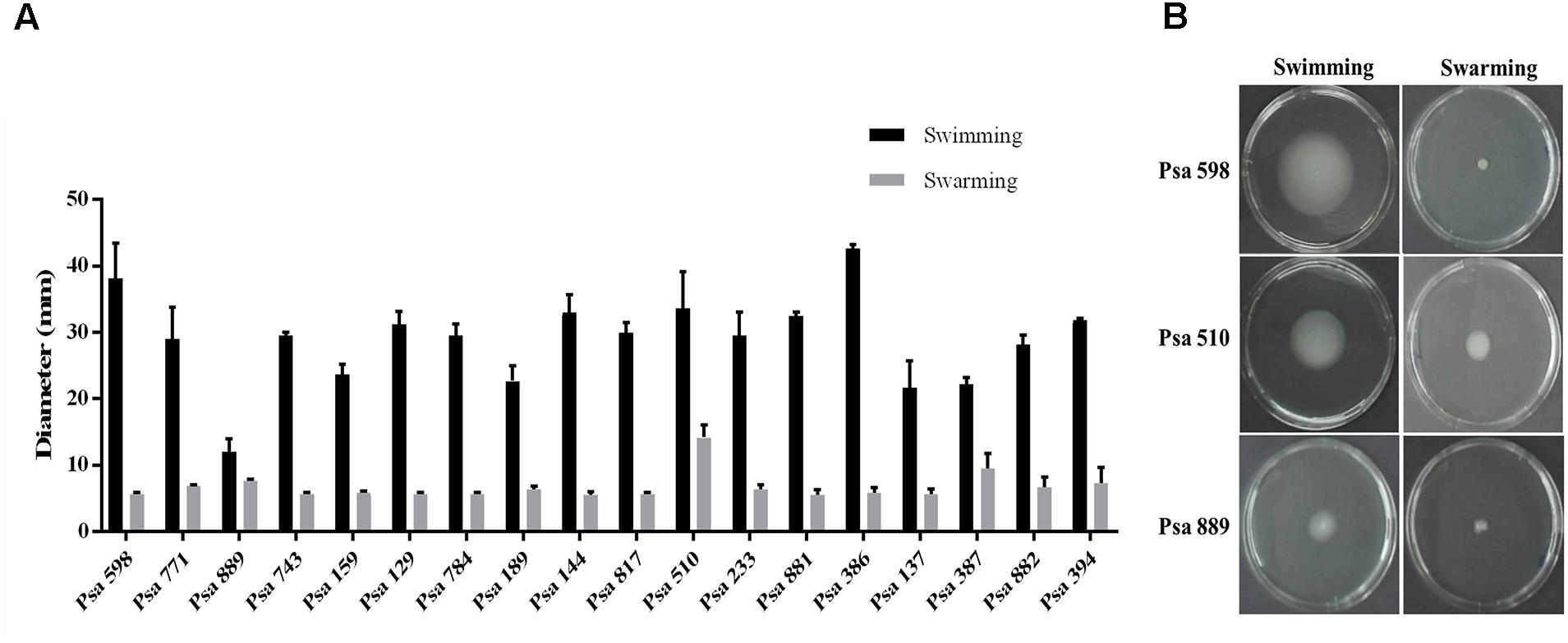
FIGURE 3. Motility of the Chilean Psa isolates. (A) Swimming or swarming motility of different Chilean Psa strains. (B) Representative image of selected strains. Swimming or swarming movement was determined at 72 h post-inoculation measuring the diameter of displacement. The names of each strain are shown.
Indole Production in the Psa Isolates
Indole-3-acetic acid production has been described in different P. syringae pathovars and P. savastanoi (Glickmann et al., 1998; Cerboneschi et al., 2016) but not in Psa. It is produced mostly from Trp via IAM by enzymes encoded in the genes iaaM and iaaH. Therefore, all 18 isolates were evaluated for their ability to produce IAA (Glickmann and Dessaux, 1995). The results show that all of the Chilean Psa isolates can produce indole at different concentrations (Figure 4A). In addition, some of the Chilean Psa isolates (Psa 882 and Psa 394) produce indole concentrations similar to those of A. brasilense (63 μg/mL IAA) that produces exceptionally large amounts of IAA (Bar and Okon, 1993). In all cases, indole was produced only in the presence of Trp, suggesting that, as observed in other P. syringae, this amino acid is the precursor of IAA synthesis in Psa. IAA production was also confirmed in the Chilean Psa strain 743 using LC-ESI-MS/MS analysis, showing a strong signal for IAA in the supernatant of the Psa 743 cell-free cultures (Supplementary Figure S4). The iaaM and iaaH genes were not detected in the Chilean Psa isolates using PCR and specific primers, suggesting an alternative route of synthesis exists in these strains. Recently, a novel IAA synthesis pathway was reported in P. syringae pv. tomato DC3000 (Pst), which involves the participation of an indole-3-acetaldehyde dehydrogenase encoded by the gene aldA and its homolog, aldB (McClerklin et al., 2018). Comparative analysis by BLASTN showed 95 and 97% identity between the aldA and aldB genes, and an aldehyde dehydrogenase sequence (GenBank accession number: CP011972.2: 149109–150602) and a carnitine dehydratase/3-oxoadipate enol-lactonase sequences (GenBank accession number: CP011972.2: 3182732–3184213) were encoded in the Biovar 3 Psa strain ICMP 18884. PCR with specific primers revealed that the aldA and aldB genes were also detected in all of the Chilean Psa strains, suggesting that they are likely to be responsible for the synthetic route of IAA. The identity of genes aldA and aldB was confirmed in strains Psa 889, Psa 743, and Psa 598 using Sanger sequencing (Supplementary Table S2).
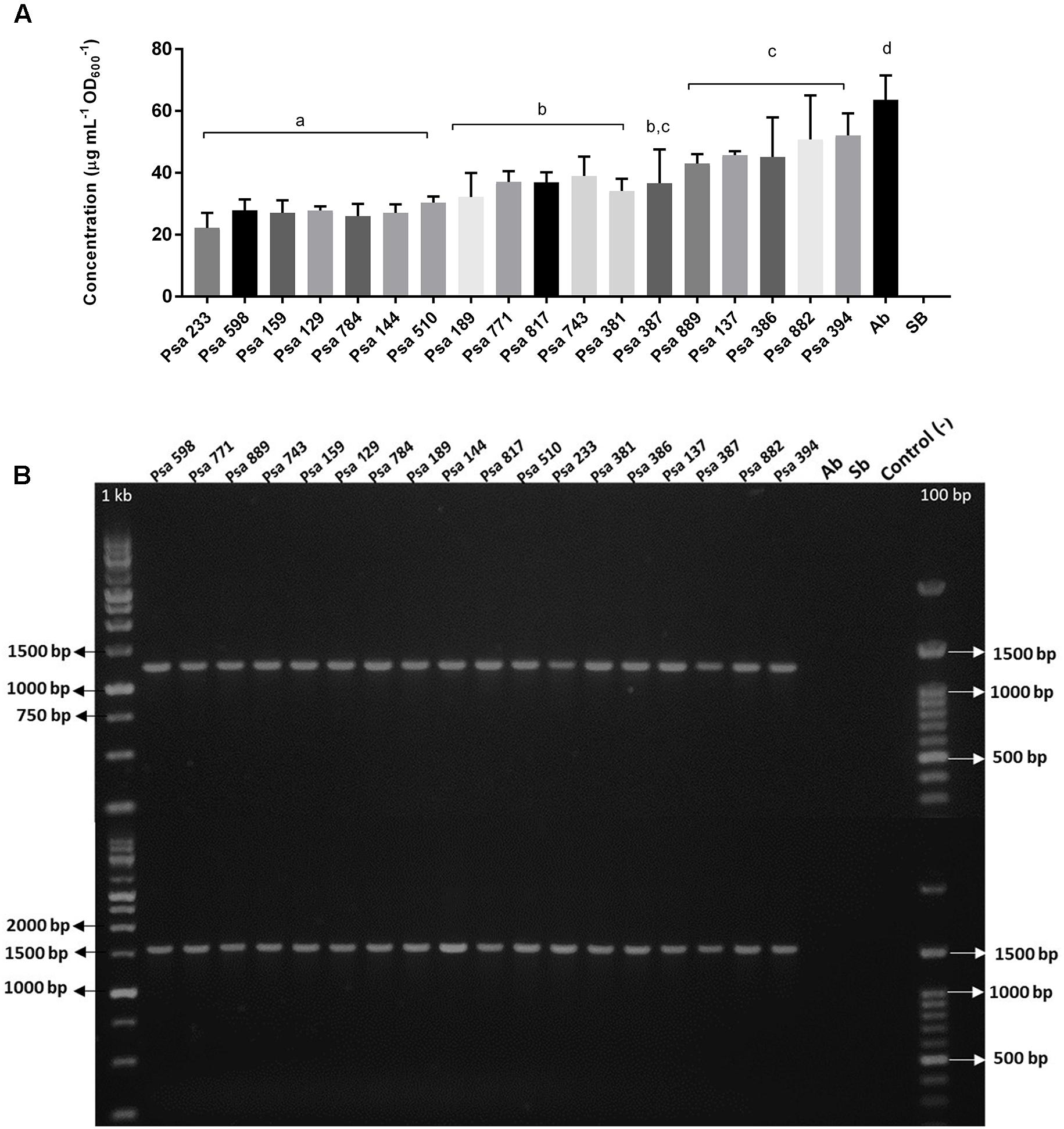
FIGURE 4. Indole production and detection of iaaL/matE genes in the Chilean Psa isolates. (A) Indole produced by the Chilean Psa isolates. Bacterial strains are organized according their level of indole production, and letters (a–d) show significant differences (p ≤ 0.05). The calibration curve with IAA (Sigma) is shown in Supplementary Figure S3. (B) PCR detection of genes iaaL (top) and matE (bottom). Molecular markers: 1 kb and 100 bp. Control (–): Reaction without DNA. A. brasilense (Ab) and S. bongori (Sb) were used for the positive and negative controls, respectively.
It has also been reported that IAA can be conjugated to the amino acid lysine to produce IAA-Lys by the enzymatic activity of the iaaL gene product (Glickmann et al., 1998; Castillo-Lizardo et al., 2015; Cerboneschi et al., 2016). Our analysis detected the presence of the genes iaaL and matE in all of the Chilean Psa isolates (Figure 4B), which are in tandem in the Hrp regulon and are associated with IAA-Lys production. However, IAA-Lys production was not detected using the LC-ESI-MS/MS analysis. The identity of the iaaL and matE genes was also confirmed using Sanger sequencing in strains Psa 889, Psa 743, and Psa 598 (Supplementary Table S3). Taken together, these results strongly suggest that the Chilean Psa isolates produce IAA using a Trp-dependent pathway.
Discussion
Genetic Analysis of the Chilean Psa Isolates
Pseudomonas syringae pv. actinidiae was first isolated in Chile in 2010, and since then, it has been considered to be a quarantine pest under the official control of the SAG of Chile. The 18 Chilean Psa isolates included in this study were obtained as part of the monitoring program established by the SAG. They were isolated from the central south region of Chile, which is the zone that accumulates the majority of Psa infections reported in the country (Servicio Agrícola y Ganadero [SAG], 2018). These strains were obtained between the years 2012 and 2016, representing the most extended study performed on Psa in Chile. All of these strains were identified by the SAG and then confirmed by the standard molecular techniques used with this pathovar (Rees-George et al., 2010; Gallelli et al., 2011). As reported previously, the use of specific primers for the ITS amplification was not specific to Psa and also amplified a fragment from P. syringae (Vanneste, 2013). Therefore, a duplex-PCR analysis was necessary to positively identify the Psa isolates.
The MLSA confirmed that the Chilean Psa isolates belong to biovar 3. In this case, four housekeeping genes were used (gyrB, rpoD, gltA, and gapA), which seems to be sufficient to discriminate between biovar 3 and the other biovars; however, it is not sufficient to distinguish between biovars 2 and 5, which according to previous research, are very closely related (Fujikawa and Sawada, 2016). This phylogenetic analysis included sequences from several Psa strains with different biovars and origins, including some older Chilean strains that were also grouped in biovar 3. This suggests that this “hypervirulent” group (Ciarroni et al., 2015) is the only found in Chile, and no other biovar has entered or emerged. The conclusions of this study are consistent with previous research in which the Chilean Psa isolates were classified in the Psa Biovar 3 cluster using different approaches: REP-PCR fingerprinting, MLVA (multiple locus variable number of tandem repeats analysis) assay and MLST (Ferrante and Scortichini, 2010, 2015; Vanneste et al., 2010; Ciarroni et al., 2015; Biondi et al., 2017). Genomic analyses of the Chilean Psa strains suggest that they originated from China forming a sub-group in biovar 3 (Butler et al., 2013; Ciarroni et al., 2015).
Nearly 50 putative effector genes have been identified in Psa and are found in most of the biovars (McCann et al., 2013; Ferrante and Scortichini, 2015; Fujikawa and Sawada, 2016). Sixteen type III effector genes, among others, were identified in all of the Chilean Psa isolates, including genes that were reported in conjugative DNA plasmids in other biovar 3 Psa strains (hopAV1 and hopAU1). The emergence of resistant strains as an evolutionary response to the use of antimicrobial compounds was observed in countries affected by recent outbreaks of Psa biovar 3 strains (Han et al., 2004; Vanneste, 2013; Colombi et al., 2017).
Phenotypic Features of the Chilean Psa Isolates
The results of this study show a high phenotypic homogeneity. However, it is still possible to observe differences between specific features and specific strains. For instance, the biochemical profile shows differences between the various Chilean Psa strains (Supplementary Table S6). These differences are related to carbon source utilization and chemical susceptibility assays. Moura et al. (2015) reported similar results with different Psa isolates from Portugal. Using the BIOLOG system, they observed differences in the ability to use at least 12 different carbon sources among the Portuguese strains. Interestingly, both the Chilean and Portuguese strains varied in their ability to use methyl pyruvate, bromo-succinic acid, and acetoacetic acid as carbon sources showing that variations in the biochemical repertory are not exclusive to the Chilean strains. Both groups of strains are susceptible to minocycline, lithium chloride, and sodium butyrate. The Chilean Psa strains are also resistant to antibiotics not used in agriculture such as rifamycin SV or vancomycin. However, curiously they were susceptible to streptomycin (MIC 3.9 μg/mL) that, in the past, has been authorized for use to control Psa infections in Chile. This suggests that no resistance has evolved among the Chilean Psa strains, in contrast to what has been reported by others where Psa strains can have a MIC for streptomycin greater than 2,000 μg/mL (Cameron and Sarojini, 2014). A similar situation has been observed for copper resistance in which other studies have reported Psa strains with a MIC from 100 μg/mL to more than 1,000 μg/mL (Cameron and Sarojini, 2014), while the Chilean strains have a MIC of 75 μg/mL. The absence of resistance among the Chilean Psa strains could be due to multiple factors such as low selective pressures from the environment or low plasticity in the Psa genome of these strains. However, is not possible to disregard the existence of resistant Chilean Psa strains in the environment. Our results do not show a clear correlation between these differences in the biochemical profiles and the origin or isolation year of the strains, but it would be interesting to determine if these differences have any relevance for fitness or niche colonization in the natural environment of Psa.
All of the Chilean isolates demonstrate a similar range of swimming motility (Figure 3) with strain Psa 889 being the only exception that lacks motility. In contrast, none of the Chilean Psa strains show swarming motility, except for strain Psa 510 that demonstrates a slightly but significantly greater amount of displacement than the other strains. The differences observed between strains Psa 889 and Psa 510 are probably related to alterations in their flagella, since no differences were observed in the growth of any of the strains according to our analysis (Supplementary Figure S5). Flagellar motility is an important virulence factor that allows the infection of plants through natural openings on their tissue surfaces (Ichinose et al., 2013). Therefore, it remains to be determined if these differences in strains Psa 889 and Psa 510 are correlated with alterations in their virulence.
Psa infections are very persistent, and once they are detected in a region, it is very difficult or even impossible to eradicate the bacteria (Vanneste, 2017). This persistence could be related to the ability to endure environmental conditions through biofilm formation (Danhorn and Fuqua, 2007; Renzi et al., 2012). It has been reported that Psa can form biofilm (Ghods et al., 2015). However, our analysis showed that the Chilean Psa strains are not able to form biofilms over abiotic solid surfaces. This and other differences observed between the Chilean Psa strains and the other Psa are probably related to the unique clonal origin of the Psa strains present in Chile (Butler et al., 2013). However, the low affinity to form biofilms over solid surfaces has been observed in the P. syringae pathovars (Ueda and Saneoka, 2015). Therefore, it seems that biofilm formation is not a hallmark of this species. Interestingly the Chilean strains do form a thin layer of cells at the air–liquid interface in liquid cultures. This phenomenon has been described for other Pseudomonas species where an air–liquid interface would represent a favorable environment due to the oxygen access enabling a more rapid rate of growth (Constantin, 2009; Ueda and Saneoka, 2015). All of these results confirm the high degree of homogeneity among the different Chilean Psa strains. Further studies are needed to determine if the differences between the Chilean strains affect the colonization and infection of the kiwifruit plants.
Indole Production in Psa Isolates
Several phytopathogens, including P. syringae pathovars, produce auxins that can alter the host’s physiology and promote plant susceptibility to infection (Glickmann et al., 1998; Cerboneschi et al., 2016). To our knowledge, this is the first report showing that Psa can produce indole using a Trp-dependent pathway. All of the Chilean Psa strains evaluated produce indole, some of them at levels similar to A. brasilense, which is a plant growth promoting bacterium (Masciarelli et al., 2013). The common route for IAA production in P. syringae pathovars is via the IAM pathway using the enzymes IaaM and IaaH. This pathway has been studied in P. syringae pv. syringae (Pss) and Psav (Glickmann et al., 1998; Baltrus et al., 2011; Aragón et al., 2014; Cerboneschi et al., 2016), and the only related report in Psa is from a strain isolated in 1984 belonging to biovar 1 which has putative ORFs of an IAM pathway (Baltrus et al., 2011). The Chilean Psa strains have the genes aldA and aldB which are associated with an alternative synthesis route of IAA recently found in P. syringae pv. tomato (McClerklin et al., 2018). Therefore, this is the most probable pathway in the Chilean Psa strains. Interestingly, bioinformatics analysis revealed that the genes iaaH and iaaM, associated with the common synthesis route of IAA, are only found in the Psa strains from biovar 4, which are now considered to be a new pathovar designated P. syringae pv. actinidifoliorum that is characterized by low virulence in kiwifruit plants (Abelleira et al., 2015). In this regard, the presence of the IAM pathway represents another distinctive feature differencing the former biovar 4 from the other Psa biovars.
Pseudomonas syringae pv. tomato and other species, such as P. savastanoi pv. nerii, also produce the enzyme IAA-lysine ligase, encoded by the iaaL gene, which is responsible for IAA-Lys production (Glickmann et al., 1998; Castillo-Lizardo et al., 2015; Cerboneschi et al., 2016). In the P. syringae pv. tomato (Pst) genome, iaaL is found in synteny with the matE gene that encodes a multidrug transporter of the MatE family. The analysis of the Chilean Psa strains revealed that all of the strains contain the genes iaaL and matE. A bioinformatic analysis showed that the iaaL gene was first annotated as a pre-protein translocase subunit Tim44 in several P. syringae pathovars; however, later it was annotated as an indoleacetate-lysine ligase gene in P. syringae pv. tomato (Castillo-Lizardo et al., 2015). According to this analysis, the matE and iaaL genes are conserved in Psa Biovar 1, 2, 3, and 5 strains with near 100% identity in their amino acid sequences (Supplementary Figure S6). There are reports on the importance of IAA production, and IAA-Lys in particular, in the virulence of P. syringae. For instance, mutations in the IAM pathway of Pss affect its growth in Phaseolus vulgaris (Mazzola and White, 1994), and the deletion of the aldA, aldB, iaaL, or matE genes in P. syringae pv. tomato result in a reduction in fitness, colonization, and virulence in infected tomato plants (Castillo-Lizardo et al., 2015; McClerklin et al., 2018). In addition, studies on the IAA-Lys effect on plants suggest that IAA conjugation can modulate hormone action and suppress the immune response (Romano et al., 1991). Our results show that the Chilean Psa strains produce IAA. However, we were not able to demonstrate IAA-Lys production. Despite this, the presence of the genes iaaL and matE in the Chilean and other Psa strains, including different biovars, raise the possibility that this compound could be produced in conditions other than those evaluated in this study. To date, the exact mechanism of action of IAA and IAA-Lys in the virulence of P. syringae species is not totally understood. The results presented here show that the Chilean Psa strains produce IAA, but it is unknown if this feature is shared with other Psa strains of biovar 3 and other biovars. The results represent the starting point to determine the mechanisms and regulation of IAA production (and possibly IAA-Lys) in Psa and its participation during infection in kiwifruits plants.
Conclusion
The results of this study confirm that the Chilean Psa isolates belong to biovar 3. The isolates exhibit high homogeneity with phenotypic differences in specific isolates. This study is also the first report of Psa strains producing IAA using a Trp-dependent pathway. Several reports suggest that this compound may be related to virulence in P. syringae pathovars. Therefore, it would be interesting to determine whether this feature plays a role during bacterial canker in kiwi plants and to evaluate whether this is a common characteristic in different biovars of this pathovar.
Author Contributions
OF, CY, XB, and RB conceived and designed the study, and analyzed the results. OF, CP, and MN performed the experiments. AV and CM performed the LC-ESI-MS/MS analysis. OF and RB wrote the manuscript. All authors reviewed and approved the final manuscript.
Funding
This work was financially supported by CONICYT grants FONDEF/II Concurso IDeA en Dos Etapas ID15I10032 and FONDECYT Postdoctorado 2017 No. 3170567.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors wish to acknowledge the Agricultural and Livestock Service (SAG) for facilitating the Psa isolate collection, the Chilean Kiwifruit Committee for support assistance, and Dr. Paula Salinas and Dr. Jorge Olivares for providing bacterial strains.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.01907/full#supplementary-material
References
Abelleira, A., Ares, A., Aguin, O., Peñalver, J., Morente, M. C., López, M. M., et al. (2015). Detection and characterization of Pseudomonas syringae pv. actinidifoliorum in kiwifruit in Spain. J. Appl. Microbiol. 119, 1659–1671. doi: 10.1111/jam.12968
Aragón, I. M., Pérez-Martínez, I., Moreno-Pérez, A., Cerezo, M., and Ramos, C. (2014). New insights into the role of indole-3-acetic acid in the virulence of Pseudomonas savastanoi pv. savastanoi. FEMS Microbiol. Lett. 356, 184–192. doi: 10.1111/1574-6968.12413
Baltrus, D. A., Nishimura, M. T., Romanchuk, A., Chang, J. H., Mukhtar, M. S., Cherkis, K., et al. (2011). Dynamic evolution of pathogenicity revealed by sequencing and comparative genomics of 19 Pseudomonas syringae isolates. PLoS Pathog. 7:e1002132. doi: 10.1371/journal.ppat.1002132
Bar, T., and Okon, Y. (1993). Tryptophan conversion to indole-3-acetic acid via indole-3-acetamide in Azospirillum brasilense Sp7. Can. J. Microbiol. 39, 81–86. doi: 10.1139/m93-011
Biebl, H., and Pfennig, N. (1978). Growth yields of green sulfur bacteria in mixed cultures with sulfur and sulfate reducing bacteria. Arch. Microbiol. 117, 9–16. doi: 10.1007/BF00689344
Biondi, E., Zamorano, A., Vega, E., Ardizzi, S., Sitta, D., De Salvador, R., et al. (2017). Draft whole genome sequence analyses on Pseudomonas syringae pv. actinidiae HR negative strains detected from kiwifruit bleeding sap samples. Phytopathology 108, 552–560. doi: 10.1094/PHYTO-08-17-0278-R
Buell, C. R., Joardar, V., Lindeberg, M., Selengut, J., Paulsen, I. T., Gwinn, M. L., et al. (2003). The complete genome sequence of the Arabidopsis and tomato pathogen Pseudomonas syringae pv. tomato DC3000. Proc. Natl. Acad. Sci. U.S.A. 100, 10181–10186. doi: 10.1073/pnas.1731982100
Butler, M. I., Stockwell, P. A., Black, M. A., Day, R. C., Lamont, I. L., and Poulter, R. T. M. (2013). Pseudomonas syringae pv. actinidiae from recent outbreaks of kiwifruit bacterial canker belong to different clones that originated in China. PLoS One 8:e57464. doi: 10.1371/journal.pone.0057464
Cameron, A., and Sarojini, V. (2014). Pseudomonas syringae pv. actinidiae: chemical control, resistance mechanisms and possible alternatives. Plant Pathol. 63, 1–11. doi: 10.1111/ppa.12066
Castillo-Lizardo, M. G., Aragón, I. M., Carvajal, V., Matas, I. M., Pérez-Bueno, M. L., Gallegos, M.-T., et al. (2015). Contribution of the non-effector members of the HrpL regulon, iaaL and matE, to the virulence of Pseudomonas syringae pv. tomato DC3000 in tomato plants. BMC Microbiol. 15:165. doi: 10.1186/s12866-015-0503-8
Cellini, A., Fiorentini, L., Buriani, G., Yu, J., Donati, I., Cornish, D. A., et al. (2014). Elicitors of the salicylic acid pathway reduce incidence of bacterial canker of kiwifruit caused by Pseudomonas syringae pv. actinidae. Ann. Appl. Biol. 165, 441–453. doi: 10.1111/aab.12150
Cerboneschi, M., Decorosi, F., Biancalani, C., Ortenzi, M. V., Macconi, S., Giovannetti, L., et al. (2016). Indole-3-acetic acid in plant–pathogen interactions: a key molecule for in planta bacterial virulence and fitness. Res. Microbiol. 167, 774–787. doi: 10.1016/j.resmic.2016.09.002
Ciarroni, S., Gallipoli, L., Taratufolo, M. C., Butler, M. I., Poulter, R. T. M., Pourcel, C., et al. (2015). Development of a multiple loci variable number of tandem repeats analysis (MLVA) to unravel the intra-pathovar structure of Pseudomonas syringae pv. actinidiae populations worldwide. PLoS One 10:e0135310. doi: 10.1371/journal.pone.0135310
Colombi, E., Straub, C., Künzel, S., Templeton, M. D., McCann, H. C., and Rainey, P. B. (2017). Evolution of copper resistance in the kiwifruit pathogen Pseudomonas syringae pv. actinidiae through acquisition of integrative conjugative elements and plasmids. Environ. Microbiol. 19, 819–832. doi: 10.1111/1462-2920.13662
Constantin, O. (2009). Bacterial biofilms formation at air liquid interfaces. Innov. Rom. Food Biotechnol. 5, 18–22.
Cunty, A., Cesbron, S., Poliakoff, F., Jacques, M. A., and Manceau, C. (2015a). Origin of the outbreak in France of Pseudomonas syringae pv. actinidiae biovar 3, the causal agent of bacterial canker of kiwifruit, revealed by a multilocus variable-number tandem-repeat analysis. Appl. Environ. Microbiol. 81,6773–6789. doi: 10.1128/AEM.01688-15
Cunty, A., Poliakoff, F., Rivoal, C., Cesbron, S., Fischer-Le Saux, M., Lemaire, C., et al. (2015b). Characterization of Pseudomonas syringae pv. actinidiae (Psa) isolated from France and assignment of Psa biovar 4 to a de novo pathovar: Pseudomonas syringae pv. actinidifoliorum pv. nov. Plant Pathol. 64, 582–596. doi: 10.1111/ppa.12297
Danhorn, T., and Fuqua, C. (2007). Biofilm formation by plant-associated bacteria. Annu. Rev. Microbiol. 61, 401–422. doi: 10.1146/annurev.micro.61.080706.093316
De La Rosa Fraile, M., Vega Aleman, D., and Fernandez Gutierrez, C. (1980). Evaluation of urea-motility-indole medium for recognition and differentiation of salmonella and shigella species in stool cultures. J. Clin. Microbiol. 12, 310–313.
Ferrante, P., and Scortichini, M. (2010). Molecular and phenotypic features of Pseudomonas syringae pv. actinidiae isolated during recent epidemics of bacterial canker on yellow kiwifruit (Actinidia chinensis) in central Italy. Plant Pathol. 59, 954–962. doi: 10.1111/j.1365-3059.2010.02304.x
Ferrante, P., and Scortichini, M. (2015). Redefining the global populations of Pseudomonas syringae pv. actinidiae based on pathogenic, molecular and phenotypic characteristics. Plant Pathol. 64, 51–62. doi: 10.1111/ppa.12236
Fujikawa, T., and Sawada, H. (2016). Genome analysis of the kiwifruit canker pathogen Pseudomonas syringae pv. actinidiae biovar 5. Sci. Rep. 6, 1–11. doi: 10.1038/srep21399
Gallelli, A., Talocci, S., L’urora, A., and Loreti, S. (2011). canker of kiwifruit, from symptomless fruits and twigs, and from pollen. Phytopathol. Mediterr. 50, 462–472. doi: 10.14601/Phytopathol_Mediterr-10039
Gao, X., Huang, Q., Zhao, Z., Han, Q., Ke, X., Qin, H., et al. (2016). Studies on the infection, colonization, and movement of Pseudomonas syringae pv. actinidiae in kiwifruit tissues using a GFPuv-labeled strain. PLoS One 11:e0151169. doi: 10.1371/journal.pone.0151169
Ghafoor, A., Hay, I. D., and Rehm, B. H. A. (2011). Role of exopolysaccharides in Pseudomonas aeruginosa biofilm formation and architecture. Appl. Environ. Microbiol. 77, 5238–5246. doi: 10.1128/AEM.00637-11
Ghods, S., Sims, I. M., Moradali, M. F., and Rehma, B. H. A. (2015). Bactericidal compounds controlling growth of the plant pathogen Pseudomonas syringae pv. actinidiae, which forms biofilms composed of a novel exopolysaccharide. Appl. Environ. Microbiol. 81, 4026–4036. doi: 10.1128/AEM.00194-15
Glickmann, E., and Dessaux, Y. (1995). A critical examination of the specificity of the Salkowski reagent for indolic compounds produced by phytopathogenic bacteria. Appl. Environ. Microbiol. 61, 793–796.
Glickmann, E., Gardan, L., Jacquet, S., Hussain, S., Elasri, M., Petit, A., et al. (1998). Auxin production is a common feature of most pathovars of Pseudomonas syringae. Mol. Plant Microbe Interact. 11, 156–162. doi: 10.1094/MPMI.1998.11.2.156
Han, H. S., Koh, Y. J., Hur, J., and Jung, J. S. (2004). Occurrence of the strA-strB streptomycin resistance genes in Pseudomonas species isolated from kiwifruit plants. J. Microbiol. 42, 365–368.
Hosseinidoust, Z., van de Ven, T. G. M., and Tufenkji, N. (2013). Evolution of Pseudomonas aeruginosa virulence as a result of phage predation. Appl. Environ. Microbiol. 79, 6110–6116. doi: 10.1128/AEM.01421-13
Ichinose, Y., Taguchi, F., and Mukaihara, T. (2013). Pathogenicity and virulence factors of Pseudomonas syringae. J. Gen. Plant Pathol. 79, 285–296. doi: 10.1007/s10327-013-0452-8
Kearse, M., Moir, R., Wilson, A., Stones-Havas, S., Cheung, M., Sturrock, S., et al. (2012). Geneious basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 28, 1647–1649. doi: 10.1093/bioinformatics/bts199
Kumar, S., Stecher, G., and Tamura, K. (2016). MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 33, 1870–1874. doi: 10.1093/molbev/msw054
Marcelletti, S., Ferrante, P., Petriccione, M., Firrao, G., and Scortichini, M. (2011). Pseudomonas syringae pv. actinidiae draft genomes comparison reveal strain-specific features involved in adaptation and virulence to Actinidia species. PLoS One 6:e27297. doi: 10.1371/journal.pone.0027297
Masciarelli, O., Urbani, L., Reinoso, H., and Luna, V. (2013). Alternative mechanism for the evaluation of indole-3-acetic acid (IAA) production by Azospirillum brasilense strains and its effects on the germination and growth of maize seedlings. J. Microbiol. 51, 590–597. doi: 10.1007/s12275-013-3136-3
Matsuda, F., Miyazawa, H., Wakasa, K., and Miyagawa, H. (2005). Quantification of indole-3- acetic acid and amino acid conjugates in rice by liquid chromatography-electrospray ionization-tandem mass spectrometry. Biosci. Biotechnol. Biochem. 69, 778–783. doi: 10.1271/bbb.69.778
Mazzola, M., and White, F. F. (1994). A mutation in the indole-3-acetic acid biosynthesis pathway of Pseudomonas syringae pv. syringae affects growth in Phaseolus vulgaris and syringomycin production. J. Bacteriol. 176, 1374–1382. doi: 10.1128/jb.176.5.1374-1382.1994
McCann, H. C., Li, L., Liu, Y., Li, D., Pan, H., Zhong, C., et al. (2017). Origin and evolution of the kiwifruit canker pandemic. Genome Biol. Evol. 9, 932–944. doi: 10.1093/gbe/evx055
McCann, H. C., Rikkerink, E. H. A., Bertels, F., Fiers, M., Lu, A., Rees-George, J., et al. (2013). Genomic analysis of the kiwifruit pathogen Pseudomonas syringae pv. actinidiae provides insight into the origins of an emergent plant disease. PLoS Pathog. 9:e1003503. doi: 10.1371/journal.ppat.1003503
McClerklin, S. A., Lee, S. G., Harper, C. P., Nwumeh, R., Jez, J. M., and Kunkel, B. N. (2018). Indole-3-acetaldehyde dehydrogenase-dependent auxin synthesis contributes to virulence of Pseudomonas syringae strain DC3000. PLoS Pathog. 14:e1006811. doi: 10.1371/journal.ppat.1006811
Mergeay, M., Nies, D. H., Schlegel, H. G., Gerits, J., Charles, P., and Gijsegem, F. V. (1985). Alcaligenes eutrophus CH34 is a facultative Chemolithotrophy with plasmid-bound resistance to heavy metals. J. Bacteriol. 162, 328–334.
Merritt, J. H., Kadouri, D. E., and O’Toole, G. A. (2011). Growing and analyzing static biofilms. Curr. Protoc. Microbiol. 22, 1B.1.1–1B.1.18. doi: 10.1002/9780471729259.mc01b01s22
Mohite, B. (2013). Isolation and characterization of indole acetic acid (IAA) producing bacteria from rhizospheric soil and its effect on plant growth. J. Soil Sci. Plant Nutr. 13, 638–649. doi: 10.4067/S0718-95162013005000051
Moura, L., García, E., Aguín, O., Ares, A., Abelleira, A., and Mansilla, P. (2015). Identificação e caracterização de Pseudomonas syringae pv. actinidiae (Psa) na Região do Entre Douro e Minho (Portugal). Soc. Ciencias Agrárias Port. 38, 196–205.
Oficina de Estudios y Políticas Agrarias [ODEPA] (2018). Ficha Nacional 2018. Available at: https://www.odepa.gob.cl/wp-content/uploads/2017/07/Ficha-Nacional-2018.pdf
O’Toole, G. A. (2011). Microtiter dish biofilm formation assay. J. Vis. Exp 47:e2437. doi: 10.3791/2437
Patel, H. K., Ferrante, P., Xianfa, M., Javvadi, S. G., Subramoni, S., Scortichini, M., et al. (2017). Identification of loci of Pseudomonas syringae pv. actinidiae involved in lipolytic activity and their role in colonization of kiwifruit leaves. Phytopathology 107, 645–653. doi: 10.1094/PHYTO-10-16-0360-R
Rees-George, J., Vanneste, J. L., Cornish, D. A., Pushparajah, I. P. S., Yu, J., Templeton, M. D., et al. (2010). Detection of Pseudomonas syringae pv. actinidiae using polymerase chain reaction (PCR) primers based on the 16S-23S rDNA intertranscribed spacer region and comparison with PCR primers based on other gene regions. Plant Pathol. 59, 453–464. doi: 10.1111/j.1365-3059.2010.02259.x
Renzi, M., Copini, P., Taddei, A. R., Rossetti, A., Gallipoli, L., Mazzaglia, A., et al. (2012). Bacterial canker on kiwifruit in Italy: anatomical changes in the wood and in the primary infection sites. Phytopathology 102, 827–840. doi: 10.1094/PHYTO-02-12-0019-R
Romano, C. P., Hein, M. B., and Klee, H. J. (1991). Inactivation of auxin in tobacco transformed with the indoleacetic acid-lysine synthetase gene of Pseudomonas savastanoi. Genes Dev. 5, 438–446. doi: 10.1101/gad.5.3.438
Scortichini, M., Marcelletti, S., Ferrante, P., Petriccione, M., and Firrao, G. (2012). Pseudomonas syringae pv. actinidiae: a re-emerging, multi-faceted, pandemic pathogen. Mol. Plant Pathol. 13, 631–640. doi: 10.1111/j.1364-3703.2012.00788.x
Servicio Agrícola y Ganadero [SAG] (2018). Estadísticas de Prospección de Psa en Kiwi 2018. Available at: http://www.sag.gob.cl/content/estadisticas-de-prospeccion-huertos-de-kiwi-positivos-huertos-y-muestras-negativas-0
Swiecicki, J.-M., Sliusarenko, O., and Weibel, D. B. (2013). From swimming to swarming: Escherichia coli cell motility in two-dimensions. Integr. Biol. 5, 1490–1494. doi: 10.1039/c3ib40130h
Templeton, M. D., Warren, B. A., Andersen, M. T., Rikkerink, E. H., and Fineran, P. C. (2015). Complete DNA sequence of Pseudomonas syringae pv. actinidiae, the causal agent of kiwifruit canker disease. Genome Announc. 3:e01054-15. doi: 10.1128/genomeA.01054-15
Ueda, A., and Saneoka, H. (2015). Characterization of the ability to form biofilms by plant-associated Pseudomonas species. Curr. Microbiol. 70, 506–513. doi: 10.1007/s00284-014-0749-7
Vanneste, J. L. (2013). Recent progress on detecting, understanding and controlling Pseudomonas syringae pv. actinidiae: a short review. N. Z. Plant Prot. 66, 170–177.
Vanneste, J. L. (2017). The scientific, economic, and social impacts of the new Zealand outbreak of bacterial canker of kiwifruit (Pseudomonas syringae pv. actinidiae). Annu. Rev. Phytopathol. 55, 377–399. doi: 10.1146/annurev-phyto-080516-035530
Vanneste, J. L., Yu, J., and Cornish, D. A. (2010). Molecular characterisations of Pseudomonas syringae pv. actinidiae strains isolated from the recent outbreak of bacterial canker on kiwifruit in Italy. N. Z. Plant Prot. 63, 7–14.
Vanneste, J. L., Yu, J., Cornish, D. A., Tanner, D. J., Windner, R., Chapman, J. R., et al. (2012). Identification, virulence and distribution of two biovars of Pseudomonas syringae pv. actinidiae in New Zealand. Plant Dis. 97, 708–719. doi: 10.1094/PDIS-07-12-0700-RE
Vásquez-Ponce, F., Higuera-Llantén, S., Pavlov, M. S., Marshall, S. H., and Olivares-Pacheco, J. (2018). Phylogenetic MLSA and phenotypic analysis identification of three probable novel Pseudomonas species isolated on King George Island, South Shetland, Antarctica. Braz. J. Microbiol. doi: 10.1016/j.bjm.2018.02.005 [Epub ahead of print].
Keywords: Pseudomonas syringae pv. actinidiae, Psa Biovar 3 (Psa-V), MLSA, kiwifruit, IAA, IAA production, indoleacetic acid lysine, IAA-L
Citation: Flores O, Prince C, Nuñez M, Vallejos A, Mardones C, Yañez C, Besoain X and Bastías R (2018) Genetic and Phenotypic Characterization of Indole-Producing Isolates of Pseudomonas syringae pv. actinidiae Obtained From Chilean Kiwifruit Orchards. Front. Microbiol. 9:1907. doi: 10.3389/fmicb.2018.01907
Received: 02 April 2018; Accepted: 30 July 2018;
Published: 22 August 2018.
Edited by:
Marco Scortichini, Consiglio per la Ricerca in Agricoltura e l’Analisi dell’Economia Agraria (CREA), ItalyReviewed by:
Stefania Tegli, Università degli Studi di Firenze, ItalyDavid John Studholme, University of Exeter, United Kingdom
Copyright © 2018 Flores, Prince, Nuñez, Vallejos, Mardones, Yañez, Besoain and Bastías. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Roberto Bastías, cm9iZXJ0by5iYXN0aWFzQHB1Y3YuY2w=