 Elena Moreno-Grúa1†Sara Pérez-Fuentes1†Asunción Muñoz-Silvestre1
Elena Moreno-Grúa1†Sara Pérez-Fuentes1†Asunción Muñoz-Silvestre1 David Viana1
David Viana1 Ana B. Fernández-Ros2Celia Sanz-Tejero2Juan M. Corpa1*
Ana B. Fernández-Ros2Celia Sanz-Tejero2Juan M. Corpa1* Laura Selva1*
Laura Selva1*- 1Biomedical Research Institute (PASAPTA-Pathology Group), Facultad de Veterinaria, Universidad CEU Cardenal Herrera, CEU Universities, Valencia, Spain
- 2EXOPOL Autovacunas y Diagnóstico, Zaragoza, Spain
Infections caused by methicillin-resistant Staphylococcus aureus (MRSA) have been a growing problem in human medicine since the 1960s, and more recently in veterinary medicine with the appearance of livestock-associated MRSA (LA-MRSA). Nevertheless, information about the presence of MRSA in rabbits is quite scarce since only one LA-MRSA identification has been previously reported. The present study aimed to determine genotypic characterization by verifying the presence of resistance determinants, virulence, and toxin genes of different S. aureus strains that cause lesions in rabbits, and their phenotypic traits based on the antimicrobial susceptibility profile. The analysis of 240 S. aureus isolates obtained from different lesion types collected from 89 Spanish and Portuguese rabbit commercial farms in the last 4 years (2014–2017) was performed. The methicillin-resistant gene mecA was found in 11.25% of the studied isolates (27 of 240) from 19 farms (13 Spanish and 6 Portuguese). Staphylococcal cassette chromosome mec (SCCmec) typing predominantly revealed type III (n = 15). Additionally, three MRSA isolates carrying the mecC gen were detected in samples from three different farms (two Spanish and one Portuguese). None of the 30 MRSA isolates was PVL-positive or tst-positive. After the multilocus sequence typing (MLST) procedure, 16 belonged to ST2855, 6 to ST146, 6 to ST398, and 2 ST4774. No ST121 isolate was mec-positive. ST398 and ST4774 isolates lacked the immune-evasion-cluster (IEC) genes. ST2855 strains were associated with the presence only of the sak gene, and ST146 isolates were ascribed to IEC type E. Therefore, this is the first description of LA-MRSA from rabbits belonging to ST2855. Interestingly, one ST2855 and two ST4774 isolates were mecC-positive, which could act as a mecC-MRSA reservoir. More studies are needed to further characterize these isolates and their relationship with humans and other animal species.
Introduction
Staphylococcus aureus multi-resistant to antibiotics is a leading cause of bacterial infections in hospitals and communities. Specifically, methicillin-resistant S. aureus (MRSA) has spread worldwide in the second half of the 20th century and is now considered endemic in healthcare facilities in all industrialized countries (Kobayashi et al., 2015). In 2005 livestock-associated MRSA (LA-MRSA) emerged in pigs (Armand-Lefevre et al., 2005), where it plays an important role a reservoir of infection to humans (Frana et al., 2013). It was later reported in different farm animals, including horses, cattle or poultry worldwide (Graveland et al., 2011; Aires-de-Sousa, 2016). S. aureus frequently infects commercial rabbits (Corpa et al., 2009), but there is only one reported case in which LA-MRSA was identified in this animal species, and was also isolated from farmers and their relatives (Agnoletti et al., 2014).
The results of population genetics studies have shown that most S. aureus strains are host-specific, which indicates low frequency of cross-species transmission (Fitzgerald, 2012). However, more recent studies that employed the multilocus sequence typing (MLST) have identified several sequence types (ST) that are associated with multiple host species. This finding implies either zoonotic transmission or a recent common ancestor (Spoor et al., 2013).
Several S. aureus MLST lineages have been associated with animals, including clonal complexes (CC): CC1 (livestock), CC5 (avian), CC130 (multi-host), CC133 (ruminants), CC151 (ruminants), CC398 (livestock), and CC425 (ruminants and wild mammals) (Harrison et al., 2017). S. aureus CC121 has a multi-host tropism (Viana et al., 2015a), including humans, where it is considered a globally disseminated hypervirulent clone, although 90% of ST121 strains are methicillin-sensitive (Rao et al., 2015). In commercial rabbits, ST121 is by far the most frequently isolated clone, with prevalences close to, and even higher than, 90% (Viana et al., 2011; Guerrero et al., 2015). This causes important economic loss associated with several lesions, such as mastitis, multisystemic abscessation and pododermatitis (Corpa et al., 2009). However, no information in rabbits about the susceptibility of ST121 strains to methicillin is available.
Due to the increasing presence of LA-MRSA strains in different animal species and the scarcity of available information on commercial rabbits, the aims of this study were to: (i) evaluate the presence of MRSA in a S. aureus collection obtained from staphylococcal lesions on different Spanish and Portuguese rabbit farms; (ii) compare the isolated strains using MLST, SCCmec, and agr typing; (iii) determine antibiotic resistance profile and virulence factors.
Materials and Methods
Isolation and Characterization of S. aureus Isolates
Two hundred and forty S. aureus isolates from rabbit clinical samples obtained on 89 rabbitries located in Spain (n = 82) and Portugal (n = 7), were studied in our laboratories in the last 4 years (2014–2017). These selected samples came from rabbits with different lesion types, including mastitis (n = 86), subcutaneous abscesses (n = 33), pododermatitis (n = 31), dermatitis (n = 21), otitis (n = 13), metritis (n = 12), conjunctivitis (n = 11), pneumonia (n = 9), rhinitis (n = 3), hepatitis (n = 3), peritonitis (n = 2), pericarditis (n = 1), and osteomyelitis (n = 1). Fourteen S. aureus isolates from nasal carriers were also analyzed.
Samples were inoculated on blood-agar (BioMérieux, Marcy l’Etoile, France) and incubated aerobically at 37°C for 24–48 h. S. aureus strains were identified on the basis of morphological growth characteristics and hemolytic properties (Devriese et al., 1996). To perform PCR, genomic DNA was extracted from each isolate with a Genelute Bacterial Genomic DNA kit (Sigma), according to the manufacturer’s protocol, except for bacterial cells, which were lysed by lysostaphin (12.5 μg/ml, Sigma) at 37°C for 1 h before DNA purification. Isolates were genotyped by MLST (Enright et al., 2000).
All the strains were checked for the presence of the mecA/mecC genes by PCR, as previously described (Geha et al., 1994; Khairalla et al., 2017). Furthermore, SCC mec cassette element classification was carried out for all the mecA-positive isolates, as described elsewhere (Zhang et al., 2005; Milheirico et al., 2007).
Detection chp, sak, sea, sep, scn, tst, and PVL-Encoding Genes and agr Typing
The MRSA isolates were subjected to a PCR assay to detect the lukF/S-PV genes that encode the PVL toxin and the tst gene that encodes the TSST-1 toxin and agr typing, as previously described (Viana et al., 2015b). These strains were also checked for the presence of the immune-evasion cluster (IEC) genes (sea, sep, sak, chp, and scn) by PCR, as formerly described (van Wamel et al., 2006).
Antibiotic Susceptibility Testing
The antibiotic susceptibility of the MRSA isolates was determined by the disk diffusion method on Mueller-Hinton agar (MHA, CONDA, Spain), according to the recommendations of the Clinical and Laboratory Standards Institute (CLSI). The disk diffusion assay was done with 14 antibiotics: bacitracin (10 U), enrofloxacin (5 μg) (OXOID), streptomycin (10 μg), spiramycin (100 μg), sulfadiazine (25 μg), chloramphenicol (30 μg) (BD), doxycycline (30 μg), erythromycin (15 μg), gentamicin (10 μg), neomycin (30 μg), penicillin (10 U), tetracycline (30 μg), cefoxitin (30 μg), and trimethoprim/sulfamethoxazole (1.25 μg/23.75 μg, respectively) (BIO-RAD). Minimum inhibitory concentrations (MIC) for cefoxitin and vancomycin was determined by using MIC Test Strip (Liofilchem) on inoculated Mueller Hinton agar plates and the results were interpreted according EUCAST breakpoints. S. aureus strain ATCC 25923 and Enterococcus faecalis strain ATCC 29212 were used as controls in the susceptibility test.
Statistical Analysis
Categorical data were compared using Fisher exact test. All reported p-values are two-tailed and analyses were performed using GraphPad software.1 Variables with p < 0.01 were considered to be statistically significant.
Results
Identification of MRSA and Lesions
Of all the analyzed S. aureus isolates, 12.5% (30/240) were identified as methicillin-resistant. Of the 30 MRSA isolates, mecA was detected in 27 isolates. In 3 isolates mecC were amplified. The 30 isolates were cefoxitin-resistant, with inhibition zone diameters <22 mm (range 0–20 mm) using disk diffusion method and MIC values >4 μg/ml (range 8–256 μg/ml).
When considering lesion type, MRSA was most frequently present in hepatitis samples (100%, 3/3), followed by metritis (58.3%, 7/12), pneumonia (44.4%, 4/9), rhinitis (33.3%, 1/3), otitis (23.1%, 3/13), conjunctivitis (18.2%, 2/11), mastitis (9.3%, 8/86), dermatitis (4.8%, 1/21), and pododermatitis (3.2%, 1/31). Hepatitis and metritis were strongly associated with MRSA (p < 0.0001) (see Table 1). When only the commonest lesions (mastitis, abscesses, and pododermatitis) were taken into account, MRSA was identified in 6% (9/150) of the cases. None of the 14 isolates from nasal carriers were MRSA.
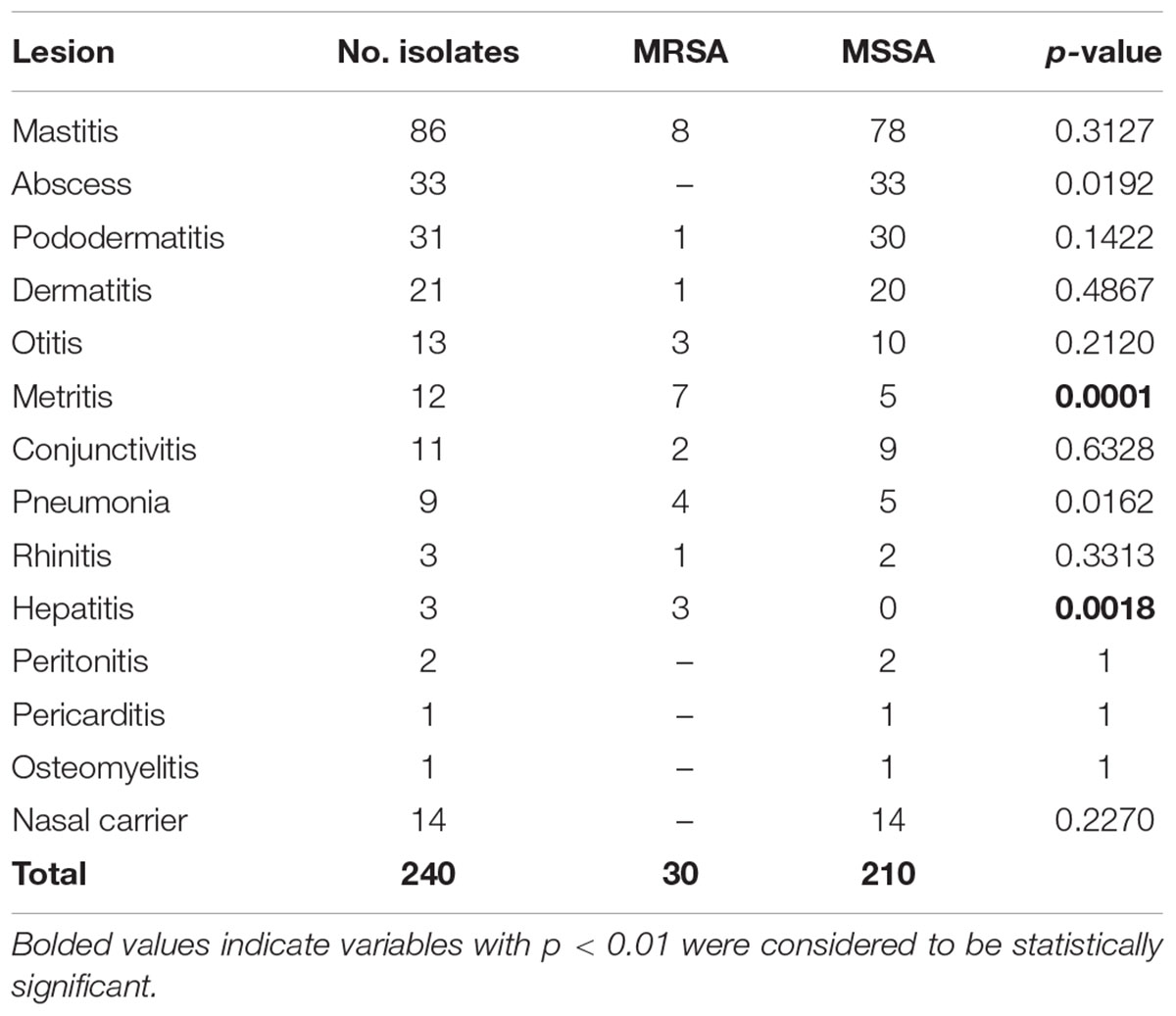
TABLE 1. Number of MRSA and MSSA isolates identified from different lesions.
Characterization of S. aureus Isolates
The MLST typing analysis revealed 13 different STs. The MLST type, including the largest number of isolates, was ST121 (n = 73). This was immediately followed by ST3764 (n = 57), ST96 (n = 36), ST2855 (n = 32), and ST1 (n = 13). Other less frequent STs were ST398 (n = 11), ST146 (n = 6), ST5 (n = 4), ST3761 (n = 3), ST4774 (n = 2), a novel tpi single locus variant of ST130, ST4473 (n = 1), a novel arc single locus variant of ST4470, ST407 (n = 1), and ST3759 (n = 1).
The 30 MRSA isolates from 22 farms (15 Spanish and 7 Portuguese) belonged to ST2855 (53.3%, 16/30), ST146 (20%, 6/30), ST398 (20%, 6/30), and ST4774 (6.6%, 2/30). For SCCmec typing, the multiplex PCR assay identified 15 of the 27 mecA MRSA isolates with SCCmec type III, 6 with SCCmec type IV, and 6 with SCCmec type V. The 3 mecC MRSA isolates belonged to a ST4774 and one ST2855.
All the MRSA strains belonging to the same lineage displayed an identical accessory gene regulator (agr) type and SCCmec-type: ST2855 harbored agrIII and SCCmec-type III; ST146 displayed agrII and SCCmec-type IV; ST398 contained agrI and SCCmec-type V; ST4774 harbored agrIII (Table 2). Methicillin-susceptible S. aureus (MSSA) strains with agr type I belonged to lineage ST398 and ST407; ST5 and ST3759 harbored agrII; ST1, ST96, ST2855, and ST4473 displayed agrIII; ST121, ST3764, and ST3761 contained agrIV.
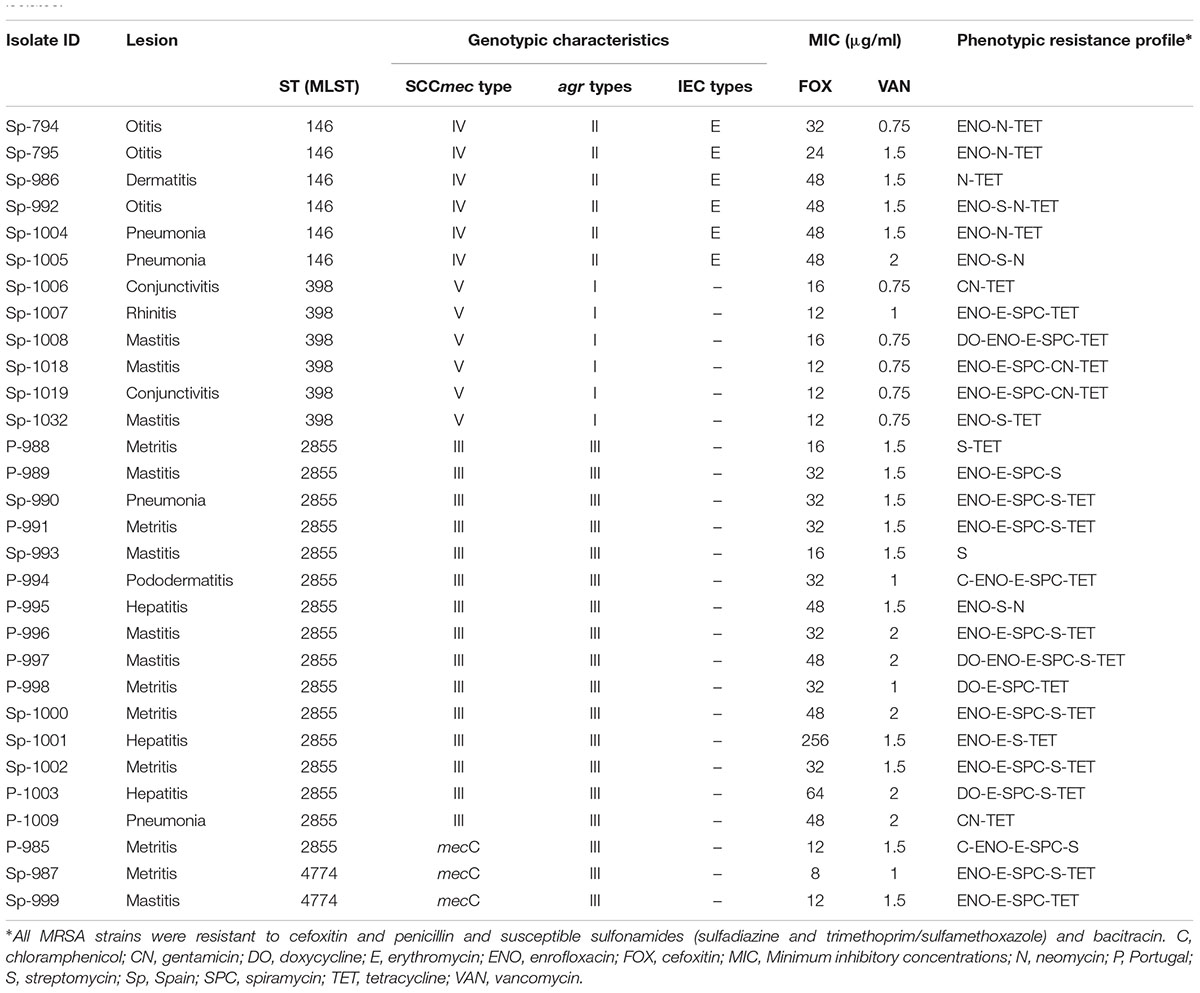
TABLE 2. Relationship among the type of lesions and genotypic (MLST, SCCmec, agr and IEC types) and phenotypic (antibiotic profile) characteristics in the MRSA isolates.
Antibiotic Resistance Profile
The full resistance rates among the 30 MRSA isolates tested in the present study were as follows: 100% (n = 30) for cefoxitin and penicillin, 83.3% (n = 25) for tetracycline, 76.6% (n = 23) for enrofloxacin, 60% (n = 18) for erythromycin, 56.6% (n = 17) for streptomycin and spiramycin, 23.3% (n = 7) for neomycin, 13.3% (n = 4) for doxycycline and gentamicin and 6.6% (n = 2) for chloramphenicol. Conversely, all the tested isolates were susceptible to sulfonamides (sulfadiazine and trimethoprim/sulfamethoxazole) and bacitracin.
The majority of the tested MRSA isolates (n = 29, 96.6%) were multidrug-resistant (resistant to three antimicrobial classes or more). A comparison of occurrence of antimicrobial resistance among the investigated MRSA isolates in relation to different STs is presented in Table 2. The ST2855 strains showed resistance to a larger number of antibiotic groups than other MRSA strains; specifically to tetracycline, macrolides (erythromycin and spiramycin) and enrofloxacin. The highest percentage of tetracyclines (100%) resistance was recorded among the ST398 strains. ST146 isolates showed resistance to a smaller number of antibiotics, of which neomycin (100%), enrofloxacin (83.3%), and tetracycline (83.3%) stood out. The two ST4774 isolates showed resistance to enrofloxacin, macrolides, and tetracycline (100%) and in one case also to streptomycin. For the remaining antimicrobial groups, no differences in resistance were observed among the STs. MIC Test Strip for cefoxitin detect a strain ST2855 with high level resistance (MIC, 256 μg/ml). None of the MRSA isolates showed resistance to vancomycin, with MICs ranged from 0.5 to 2 μg/ml.
Detection of IEC Cluster (scn, chp, sak, and sea or sep), tst, and PVL Genes Among MRSA Isolates
The PCR detection of the IEC genes, tst and PVL-encoding genes was carried out for the MRSA isolates. None of the MRSA isolates were positive for sea or sep, chp, tst, and PVL genes. The sea/sep gene along with the sak (73.3%), chp (0%), and scn (20%) genes, modulators of different parts of the innate immune system, forming an immune evasion cluster (IEC) (van Wamel et al., 2006). Depending on the presence or absence of these genes and their different combinations, S. aureus isolates were classified into 7 different IEC types according to patters previously described. The scn gene is mandatory for the consideration of the IEC types (Benito et al., 2016). All ST146 isolates contained IEC type E (comprised of sak and scn genes). However, isolates belonging to ST2855 were associated with the presence only of the sak gene, while the strains belonging to ST398 and ST4774 did not contain IEC genes.
Discussion
In the present study, 240 S. aureus isolates obtained from rabbits suffering different lesions, located on 89 farms of Spain and Portugal, were analyzed between 2014 and 2017. Isolates were obtained mainly from mastitis (n = 86), abscesses (n = 33) and pododermatitis (n = 31), which were the most frequently observed lesions associated with S. aureus infections in commercial rabbits (Segura et al., 2007; Viana et al., 2007).
Screening for methicillin-resistant isolates allowed the identification of 30 strains as MRSA. The lesion types from which they were isolated differed compared with those upper previously indicated as more frequent (mastitis, abscesses, and pododermatitis). MRSA was detected in 100% (3 out of 3) of hepatitis cases, metritis (58.3%, 7/12), pneumonia (44.4%, 4/9), rhinitis (33.3%, 1/3), otitis (23.1%, 3/13), conjunctivitis (18.2%, 2/11), mastitis (9.3%, 8/86), dermatitis (4.8%, 1/21), and pododermatitis (3.2%, 1/31). Therefore, while the percentage of the most frequent lesions (mastitis, abscesses, and pododermatitis) caused by S. aureus was 62.5% of all lesions (150/240), MRSA was involved only in 30% (9/30) of these same lesions. They were not isolated from abscesses and there was only one pododermatitis case. Hepatitis and metritis were found to be strongly associated with MRSA (hepatitis p < 0.0018; metritis p < 0.0001). This unusual lesion pattern occasioned with MRSA strains versus habitual S. aureus infections could indicate a different pathogenesis of MRSA infections in rabbits.
In order to understand the pathogenesis of staphylococcal infections, the correct identification of the involved strain is vital. Thus the development of high discriminatory typing techniques, such as MLST, is very important (Enright et al., 2000). In the present study, the most frequently detected MLST type was ST121 (73/240). It has been reported that the majority of chronic staphylococcal infections in rabbits are caused by high virulence strains that belong mainly to the ST121 lineage (Vancraeynest et al., 2006; Viana et al., 2011; Guerrero et al., 2015). The ST121 lineage was also the predominant one in this study, followed immediately by ST3764 (57/240). This last lineage, together with strains ST3761, a minority in our study (n = 3), belong to clonal complex CC121. S. aureus CC121 has a multi-host tropism, which is a common cause of human skin and soft-tissue infections (Viana et al., 2015a). In humans, the ST121 lineage is also considered a hypervirulent clone, although approximately 90% of the ST121 strains were methicillin-sensitive (Rao et al., 2015). The present study agrees with these results since all the ST121 strains from rabbit isolates were methicillin-sensitive.
A further ten different ST types among the tested isolates were identified: ST96 (n = 36), ST2855 (n = 32), ST1 (n = 13), ST398 (n = 11), ST146 (n = 6), ST5 (n = 4), ST4774 (n = 2), ST4473 (n = 1), ST407 (n = 1), and ST3759 (n = 1). However, MRSA isolates showed limited genetic diversity (ST2855, ST146, ST398, and ST4774), being the ST2855 the most predominant clone (53.3%; 16/30). It is noteworthy that 50% (16/32) of strains ST2855 were methicillin-sensitive. The only isolate described to date to belong to this lineage is an MSSA isolate of mastitis, described in a rabbit from Italy in 2012.2 All the ST146 strains isolated herein were methicillin-resistant. No data exists in the literature about S. aureus isolates from rabbits that belong to this lineage. ST146 belongs to clonal complex CC5. This clonal complex has been previously found in S. aureus isolates from rabbit carcasses, but none were methicillin-resistant (Merz et al., 2016). S. aureus associated with CC5 is commonly detected in humans or animal hosts, including poultry (Krupa et al., 2018). On the other hand, six of the 11 ST398 isolates were MRSA. The only LA-MRSA case to have occurred in rabbits for meat production belongs to this lineage (Agnoletti et al., 2014). Finally, the 3 mecC MRSA isolates belonged to ST4774, a novel tpi single locus variant of ST130, and one ST2855. Isolates reported to date and carrying mecC belonged mainly to lineages common in cattle, namely CC130, CC1943, and CC425, suggesting a zoonotic reservoir (Aires-de-Sousa, 2016). Besides cattle, mecC has also been found among other farm animals such isolates ST130 in sheep (Ariza-Miguel et al., 2014; Giacinti et al., 2017) and an isolate ST425 that caused a highly virulent infection in a rabbit (Paterson et al., 2012).
Other typing techniques used to characterize MRSA strains include the identification of the agr and staphylococcal cassette chromosome mec (SCCmec). The mecA-positive MRSA strains that belong to the same lineage displayed identical agr and mec types: ST2855 harbored agrIII and mec type III; ST146 displayed agrII and mec type IV and ST398 contained agrI and mec type V. The mecC-positive isolates harbored agrIII. The agr locus plays a critical role in MRSA pathogenesis and has been assumed to play a key role in human staphylococcal infections (El-Baz et al., 2017). Other lineages with agrIII include strains type ST96 or ST1. In rabbits, differences have been detected in the virulence between strains ST121 and ST96 (Guerrero et al., 2015; Viana et al., 2015b), which could be related with differences with this regulator type. Regarding the SCCmec type, different combinations of genes in several SCCmec types also leads to the strains displaying distinct antibiotic susceptibilities. Although the majority of the tested MRSA isolates were multidrug-resistant, the ST2855 strains showed resistance to a larger number of antibiotic groups than other strains, as well as higher frequency of resistance to macrolides. This multiresistance could justify its more extended presence compared to other MRSA strains. Types II and III SCCmec segments have been reported to be longer and to possess multidrug resistances to the strains that carry these elements. These types have frequently been demonstrated in HA-MRSA strains (Taherirad et al., 2016). Similarly to other studies, the ST398 strains identified in this study belonged to V SCCmec type (Witte et al., 2007; Bardiau et al., 2013). The highest percentage of tetracyclines (100%) resistance were recorded among the ST398 strains. Resistance to tetracycline is also associated with LA ST398 (Harrison et al., 2017), and acquisition of tetK as part of SCCmec type Vc by tetM-positive LA ST398 has been demonstrated (Larsen et al., 2016). On the other hand, ST146 isolates showed resistance to fewer antibiotics, of which neomycin (100%), enrofloxacin (83.3%) and tetracycline (83.3%) stand out. In Spain, ST146 MRSA has been detected in nasal carriage in Primary Healthcare Center patients. This isolate was typed as ST146-CC5 SCCmec IVc, and it presented a multiresistance phenotype, similarly to our ST146 rabbit strains. Strains with similar characteristics are considered hospital-acquired HA-MRSA (Lozano et al., 2015). Finally, mecC is part of a novel SCCmec assigned type XI (García-Álvarez et al., 2011). Aires-de-Sousa (2016) describes that isolates of this new genetic element are not resistant to antibiotics other than beta-lactams, in contrast to our results where mecC strains also showed resistance to enrofloxacin, tetracycline and macrolides.
Finally, all the MRSA strains described in this study were negative for the genes that code toxins PVL and TSST-1. A comparative analysis of the accessory genomes of the ST121 strains showed that the majority of human strains contained mobile genetic elements, which encode the potent toxins involved in human disease pathogenesis, such as PVL and TSST-1. According to our study, no rabbit strains carried lukS/F-PV or tst genes, which indicates that they are dispensable for the S. aureus infection of rabbits (Viana et al., 2015a). On the other hand, the presence of the immune-evasion-cluster (IEC) genes was determined for MRSA isolates to determine whether they may have human or animal origin. The presence of IEC type E in all ST146 isolates points to the possible human origin of this clone. However, ST398 and ST4774 (a single locus variant of ST130) isolates belonging to LA-lineages since were IEC negative. There are numerous reports of the presence of LA-MRSA in livestock caused by CC398 and several MRSA lineages as CC130 (Agnoletti et al., 2014; Paterson et al., 2014; Aires-de-Sousa, 2016; Harrison et al., 2017). However, isolates belonging to ST2855 were associated with the presence only of the sak gene. Stegger et al. (2013) described the presence of IEC in a porcine S. aureus CC398 isolate within the livestock clade, which supports that reacquisition of IEC enables LA-MRSA CC398 to spread in human populations. More recently Kraushaar et al. (2017) demonstrated that lysogenic conversion of LA-CC398 strains by virulence-associated phages may occur and that new pathotypes may emerge by this mechanism.
Vancomycin is an important antibiotic to treat MRSA isolates, so the emergence of vancomycin-resistant S. aureus (VRSA) strains poses a serious global threat to public health. Two mechanisms, including cell wall changes and acquired van genes were involved in vancomycin resistance. None of the MRSA isolates showed resistance to vancomycin.
A cefoxitin disk diffusion test is used to predict the presence of mecA in S. aureus (Cauwelier et al., 2004; Swenson et al., 2005). All MRSA isolates were cefoxitin-resistant, with inhibition zone diameters <22 mm using disk diffusion method. Therefore, this method is a good test for routine detection of all classes of MRSA. On the other hand, isolates harboring mecC typically yield negative results for conventional mecA PCR, which can translate in detection errors in a subset of MRSA isolates (Paterson et al., 2012). Therefore, detecting mecA/mecC genes proves to be more reliable test for detecting methicillin resistance among staphylococci.
Methicillin-resistant S. aureus prevalence varies according to sampling type and the studied animal species. The European Food Safety Authority [EFSA] (2009) has reported a 26.9% MRSA prevalence, which was detected in the dust of pigs’ production holdings of the European Union. Recent research into small ruminants has indicated an overall estimated prevalence of 0.70% (2/286) in milk samples in herds (Giacinti et al., 2017). In cattle a 4.4% prevalence has been reported in isolates collected from bovine mastitis cases (Bardiau et al., 2013). In the only MRSA case published in rabbits, only in 1 of 40 (2.5%) farms with clinical staphylococcosis has detected MRSA in skin samples (Agnoletti et al., 2014). Although the present study was not designed according to an epidemiological point of view, an unexpectedly large number of positive samples (12.5%; 30 of 240 isolates) from 22 different farms (22 of 89 farms) in Spain (n = 15) and Portugal (n = 7) was detected. Therefore, it can be stated that the spread of MRSA strains on Spanish and Portuguese rabbitries in the last 4 years is worrisome.
Conclusion
An unexpected large number of MRSA strains obtained from numerous rabbit farms, isolated from infrequent lesions in S. aureus infections, is herein reported. A new lineage of MRSA, ST2855 associated with livestock, has not yet been described. This is the first description of LA-MRSA in rabbits belonging to ST2855 and the first report of mecC MRSA in rabbit samples from Iberian Peninsula. More studies are needed to further characterize these isolates and their relationship with humans and other species.
Ethics Statement
No approval from the Animal Welfare Ethics Committee was required, since all isolates were analyzed as part of microbiological diagnostics in accordance with the Spanish law RD53/2013.
Author Contributions
JC and LS conceived the presented idea. EM-G, SP-F, and AM-S carried out the experiments. AF-R and CS-T contributed to samples preparations. LS wrote the manuscript with support from DV and JC. All authors discussed the results and contributed to the final manuscript.
Funding
This study has been supported by grants from the Universidad CEU Cardenal Herrera (INDI 17/07) and the Comisión Interministerial de Ciencia y Tecnología (AGL2014-53405-C2-2-P). Grants for SP-F from the Generalitat Valenciana (ACIF/2016/085), and for AM-S and EM-G from the Universidad CEU Cardenal Herrera, are also gratefully acknowledged.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Footnotes
References
Agnoletti, F., Mazzolini, E., Bacchin, C., Bano, L., Berto, G., Rigoli, R., et al. (2014). First reporting of methicillin-resistant Staphylococcus aureus ( MRSA ) ST398 in an industrial rabbit holding and in farm-related people. Vet. Microbiol. 170, 172–177. doi: 10.1016/j.vetmic.2014.01.035
Aires-de-Sousa, M. (2016). MRSA among animals: current overview. Clin. Microbiol. Infect. 23, 373–380. doi: 10.1016/j.cmi.2016.11.002
Ariza-Miguel, J., Hernández, M., Fernández-Natal, I., and Rodríguez-Lázaro, D. (2014). Methicillin-resistant Staphylococcus aureus harboring mecC in livestock in Spain. J. Clin. Microbiol. 52, 4067–4069. doi: 10.1128/JCM.01815-14
Armand-Lefevre, L., Ruimy, R., and Andremont, A. (2005). Clonal comparison of Staphylococcus aureus isolates from healthy pig farmers, human controls, and pigs. Emerg. Infect. Dis. 11, 711–714. doi: 10.3201/eid1105.040866
Bardiau, M., Yamazaki, K., Duprez, J. N., Taminiau, B., Mainil, J. G., and Ote, I. (2013). Genotypic and phenotypic characterization of methicillin- resistant Staphylococcus aureus ( MRSA ) isolated from milk of bovine mastitis. Lett. Appl. Microbiol. 57, 181–186. doi: 10.1111/lam.12099
Benito, D., Gómez, P., Aspiroz, C., Zarazaga, M., Lozano, C., and Torres, C. (2016). Molecular characterization of Staphylococcus aureus isolated from humans related to a livestock farm in Spain, with detection of MRSA-CC130 carrying mecC gene: A zoonotic case? Enferm. Infecc. Microbiol. Clín. 34, 280–285. doi: 10.1016/j.eimc.2015.03.008
Cauwelier, B., Gordts, B., Descheemaecker, P., and Van Landuyt, H. (2004). Evaluation of a disk diffusion method with cefoxitin (30 microg) for detection of methicillin-resistant Staphylococcus aureus. Eur. J. Clin. Microbiol. Infect. Dis. 23, 389–392. doi: 10.1007/s10096-004-1130-8
Corpa, J. M., Hermans, K., and Haesebrouck, F. (2009). Main pathologies associated with Staphylococcus aureus infections in rabbits: a review. World Rabbit Sci. 17, 115–125.
Devriese, L. A., Hendrickx, W., Godard, C., Okerman, L., and Haesebrouck, F. (1996). A new pathogenic Staphylococcus aureus type in commercial rabbits. J. Vet. Med. B 43, 313–315. doi: 10.1111/j.1439-0450.1996.tb00319.x
El-Baz, R., Rizk, D. E., Barwa, R., and Hassan, R. (2017). Virulence characteristics and molecular relatedness of methicillin resistant Staphylococcus aureus harboring different staphylococcal cassette chromosome mec. Microb. Pathog. 113, 385–395. doi: 10.1016/j.micpath.2017.11.021
Enright, M. C., Day, N. P., Davies, C. E., Peacock, S. J., and Spratt, B. G. (2000). Multilocus sequence typing for characterization of methicillin- resistant and methicillin-susceptible clones of Staphylococcus aureus. J. Clin. Microbiol. 38, 1008–1015.
European Food Safety Authority [EFSA] (2009). Analysis of the baseline survey on the prevalence of methicillin-resistant Staphylococcus aureus (MRSA) in holdings with breeding pigs, in the EU, 2008, part A: MRSA prevalence estimates. EFSA J. 7:1376. doi: 10.2903/j.efsa.2009.1376
Fitzgerald, J. R. (2012). Livestock-associated Staphylococcus aureus: origin, evolution and public health threat. Trends Microbiol. 20, 192–198. doi: 10.1016/j.tim.2012.01.006
Frana, T. S., Beahm, A. R., Hanson, B. M., Kinyon, J. M., Layman, L. L., Karriker, L. A., et al. (2013). Isolation and characterization of methicillin-resistant Staphylococcus aureus from pork farms and visiting veterinary students. PLoS One 8:e53738. doi: 10.1371/journal.pone.0053738
García-Álvarez, L., Holden, M. T., Lindsay, H., Webb, C. R., Brown, D. F., Curran, M. D., et al. (2011). Methicillin-resistant Staphylococcus aureus with a novel mecA homologue in human and bovine populations in the UK and Denmark: a descriptive study. Lancet Infect. Dis. 11, 595–603. doi: 10.1016/S1473-3099(11)70126-8
Geha, D. J., Uhl, J. R., Gustaferro, C. A., and Persing, D. H. (1994). Multiplex PCR for identification of methicillin-resistant staphylococci in the clinical laboratory. J. Clin. Microbiol. 32, 1768–1772.
Giacinti, G., Carfora, V., Caprioli, A., Sagrafoli, D., Marri, N., Giangolini, G., et al. (2017). Prevalence and characterization of methicillin-resistant Staphylococcus aureus carrying mecA or mecC and methicillin-susceptible Staphylococcus aureus in dairy sheep farms in Central Italy. J. Dairy Sci. 100, 7857–7863. doi: 10.3168/jds.2017-12940
Graveland, H., Duim, B., van Duijkeren, E., Heederik, D., and Wagenaar, J. A. (2011). Livestock-associated methicillin-resistant Staphylococcus aureus in animals and humans. Int. J. Med. Microbiol. 301, 630–634. doi: 10.1016/j.ijmm.2011.09.004
Guerrero, I., Ferrian, S., Penadés, M., García-Quirós, A., Pascual, J. J., Selva, L., et al. (2015). Host responses associated with chronic staphylococcal mastitis in rabbits. Vet. J. 204, 338–344. doi: 10.1016/j.tvjl.2015.03.020
Harrison, E. M., Coll, F., Toleman, M. S., Blane, B., Brown, N. M., Török, M. E., et al. (2017). Genomic surveillance reveals low prevalence of livestock- associated methicillin-resistant Staphylococcus aureus in the East of England. Sci. Rep. 7:7406. doi: 10.1038/s41598-017-07662-2
Khairalla, A. S., Wasfi, R., and Ashour, H. M. (2017). Carriage frequency, phenotypic, and genotypic characteristics of methicillin-resistant Staphylococcus aureus isolated from dental health-care personnel, patients, and environment. Sci. Rep. 7:7390. doi: 10.1038/s41598-017-07713-8
Kobayashi, S. D., Malachowa, N., and DeLeo, F. R. (2015). Pathogenesis of Staphylococcus aureus abscesses. Am. J. Pathol. 185, 1518–1527. doi: 10.1016/j.ajpath.2014.11.030
Kraushaar, B., Hammerl, J. A., Kienöl, M., Heinig, M. L., Sperling, N., Thanh, M. D., et al. (2017). Acquisition of virulence factors in livestock-associated MRSA: lysogenic conversion of CC398 strains by virulence gene containing phages. Sci. Rep. 7:2004. doi: 10.1038/s41598-017-02175-4
Krupa, P., Bystro, J., Bania, J., Podkowik, M., Empel, J., and Mroczkowska, A. (2018). Genotypes and oxacillin resistance of Staphylococcus aureus from chicken and chicken meat in Poland. Poult. Sci. 93, 3179–3186. doi: 10.3382/ps.2014-04321
Larsen, J., Clasen, J., Hansen, J. E., Paulander, W., Petersen, A., Larsen, A. R., et al. (2016). Copresence of tet(K) and tet(M) in livestock-associated methicillin- resistant Staphylococcus aureus clonal complex 398 is associated with increased fitness during exposure to sublethal concentrations of tetracycline. Antimicrob. Agents Chemother. 60, 4401–4403. doi: 10.1128/AAC.00426-16
Lozano, C., Marí, A., Aspiroz, C., Gómez-Sanz, E., Ceballos, S., Fortuño, B., et al. (2015). Nasal carriage of coagulase positive staphylococci in patients of a Primary-Healthcare-Center: genetic lineages and resistance and virulence genes. Enferm. Infecc. Microbiol. Clín. 33, 391–396. doi: 10.1016/j.eimc.2014.09.007
Merz, A., Stephan, R., and Johler, S. (2016). Genotyping and DNA microarray based characterization of Staphylococcus aureus isolates from rabbit carcasses. Meat Sci. 112, 86–89. doi: 10.1016/j.meatsci.2015.11.002
Milheirico, C., Oliveira, D. C., and de Lencastre, H. (2007). Update to the multiplex PCR strategy for assignment of mec element types in Staphylococcus aureus. Antimicrob. Agents Chemother. 51, 3374–3377. doi: 10.1128/AAC.00275-07
Paterson, G. K., Harrison, E. M., and Holmes, M. A. (2014). The emergence of mecC methicillin-resistant Staphylococcus aureus. Trends Microbiol. 22, 42–47. doi: 10.1016/j.tim.2013.11.003
Paterson, G. K., Larsen, A., Robb, A., Edwards, G., Penycott, T., Foster, G., et al. (2012). The newly described mecA homologue, mecALGA251, is present in methicillin-resistant Staphylococcus aureus isolates from a diverse range of host species. J. Antimicrob. Chemother. 67, 2809–2813. doi: 10.1093/jac/dks329
Rao, Q., Shang, W., Hu, X., and Rao, X. (2015). Staphylococcus aureus ST121: a globally disseminated hypervirulent clone. J. Med. Microbiol. 64, 1462–1473. doi: 10.1099/jmm.0.000185
Segura, P., Martinez, J., Peris, B., Selva, L., Viana, D., Penades, J. R., et al. (2007). Staphylococcal infections in rabbit does on two industrial farms. Vet. Rec. 160, 869–872. doi: 10.1136/vr.160.25.869
Spoor, L. E., McAdam, P. R., Weinert, L. A., Rambaut, A., Hasman, H., Aarestrup, F. M., et al. (2013). Livestock origin for a human pandemic clone of community- associated methicillin-resistant Staphylococcus aureus. mBio 4:e00356-13. doi: 10.1128/mBio.00356-13
Stegger, M., Liu, C. M., Larsen, J., Soldanova, K., Aziz, M., Contentecuomo, T., et al. (2013). Rapid differentiation between livestock-associated and livestock-independent Staphylococcus aureus CC398 clades. PLoS One 8:e79645. doi: 10.1371/journal.pone.0079645
Swenson, J. M., Tenover, F. C., and Cefoxitin Disk Study Group (2005). Results of disk diffusion testing with cefoxitin correlate with presence of mecA in Staphylococcus spp. J. Clin. Microbiol. 43, 3818–3823. doi: 10.1128/JCM.43.8.3818
Taherirad, A., Jahanbakhsh, R., Shakeri, F., Anvary, S., and Ghaemi, E. A. (2016). Staphylococcal cassette chromosome mec types among methicillin-resistant Staphylococcus aureus in Northern Iran. Jundishapur J. Microbiol. 9:e33933. doi: 10.5812/jjm.33933
van Wamel, W. J. B., Rooijakkers, S. H., Ruyken, M., van Kessel, K. P., and van Strijp, J. A. (2006). The innate immune modulators staphylococcal complement inhibitor and chemotaxis inhibitory protein of Staphylococcus aureus are located on beta hemolysin converting bacteriophages. J. Bacteriol. 188, 1310–1315. doi: 10.1128/JB.188.4.1310-1315.2006
Vancraeynest, D., Haesebrouck, F., Deplano, A., Denis, O., Godard, C., Wildemauwe, C., et al. (2006). International dissemination of a high virulence rabbit Staphylococcus aureus clone. J. Vet. Med. B Infect. Dis. Vet. Public Health 53, 418–422. doi: 10.1111/j.1439-0450.2006.00977.x
Viana, D., Comos, M., McAdam, P. R., Ward, M. J., Selva, L., Guinane, C. M., et al. (2015a). A single natural nucleotide mutation alters bacterial pathogen host tropism. Nat. Genet. 47, 361–366. doi: 10.1038/ng.3219
Viana, D., Selva, L., Penadés, M., and Corpa, J. M. (2015b). Screening of virulence genes in Staphylococcus aureus isolates from rabbits. World Rabbit Sci. 23, 185–195. doi: 10.4995/wrs.2015.3961
Viana, D., Selva, L., Callanan, J. J., Guerrero, I., Ferrian, J., and Corpa, J. M. (2011). Strains of Staphylococcus aureus and pathology associated with chronic suppurative mastitis in rabbits. Vet. J. 190, 403–407. doi: 10.1016/j.tvjl.2010.11.022
Viana, D., Selva, L., Segura, P., Penadés, J. R., and Corpa, J. M. (2007). Genotypic characterization of Staphylococcus aureus strains isolated from rabbit lesions. Vet. Microbiol. 121, 288–298. doi: 10.1016/j.vetmic.2006.12.003
Witte, W., Strommenger, B., Stanek, C., and Cuny, C. (2007). Staphylococcus aureus ST398 in humans and animals, Central Europe. Emerg. Infect. Dis. 13, 255–258. doi: 10.3201/eid1302.060924
Zhang, K., McClure, J., Elsayed, S., Louie, T., and Conly, J. M. (2005). Novel multiplex PCR assay for characterization and concomitant subtyping of staphylococcal cassette chromosome mec Types I to V in methicillin-resistant Staphylococcus aureus. J. Clin. Microbiol. 43, 5026–5033. doi: 10.1128/JCM.43.10.5026-5033.2005
Keywords: Staphylococcus aureus, MRSA, LA-MRSA, rabbit, lesions
Citation: Moreno-Grúa E, Pérez-Fuentes S, Muñoz-Silvestre A, Viana D, Fernández-Ros AB, Sanz-Tejero C, Corpa JM and Selva L (2018) Characterization of Livestock-Associated Methicillin-Resistant Staphylococcus aureus Isolates Obtained From Commercial Rabbitries Located in the Iberian Peninsula. Front. Microbiol. 9:1812. doi: 10.3389/fmicb.2018.01812
Received: 12 March 2018; Accepted: 19 July 2018;
Published: 14 August 2018.
Edited by:
Muna Anjum, Animal and Plant Health Agency, United KingdomReviewed by:
Ákos Tóth, National Public Health Institute (OKI), HungaryPierluigi Di Ciccio, Università degli Studi di Torino, Italy
Copyright © 2018 Moreno-Grúa, Pérez-Fuentes, Muñoz-Silvestre, Viana, Fernández-Ros, Sanz-Tejero, Corpa and Selva. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Juan M. Corpa, am1jb3JwYUB1Y2hjZXUuZXM= Laura Selva, bHNlbHZhQHVjaGNldS5lcw==
†These authors have contributed equally to this work