 Jing Ouyang1,2,3†
Jing Ouyang1,2,3† Stéphane Isnard1,2†
Stéphane Isnard1,2† John Lin1,2
John Lin1,2 Brandon Fombuena1,2,4
Brandon Fombuena1,2,4 Xiaorong Peng1,2,5Seema Nair Parvathy6
Xiaorong Peng1,2,5Seema Nair Parvathy6 Yaokai Chen3Michael S. Silverman6
Yaokai Chen3Michael S. Silverman6 Jean-Pierre Routy1,2,7*
Jean-Pierre Routy1,2,7*- 1Infectious Diseases and Immunity in Global Health Program, McGill University Health Centre, Research Institute, Montreal, QC, Canada
- 2Chronic Viral Illness Service, McGill University Health Centre, Montreal, QC, Canada
- 3Chongqing Public Health Medical Center, Chongqing, China
- 4Department of Microbiology and Immunology, McGill University, Montreal, QC, Canada
- 5State Key Laboratory for Diagnosis and Treatment of Infectious Diseases, National Clinical Research Center for Infectious Diseases, Collaborative Innovation Center for Diagnosis and Treatment of Infectious Diseases, College of Medicine, The First Affiliated Hospital, Zhejiang University, Hangzhou, China
- 6Infectious Diseases Division, St. Joseph's Health Care, London, ON, Canada
- 7Division of Hematology, McGill University Health Centre, Montreal, QC, Canada
The gastrointestinal (GI) tract is a complex and well-balanced milieu of anatomic and immunological barriers. The epithelial surface of the GI tract is colonized by trillions of microorganisms, known as the gut microbiota, which is considered an “organ” with distinctive endocrine and immunoregulatory functions. Dysregulation of the gut microbiota composition, termed dysbiosis, has been associated with epithelial damage and translocation of microbial products into the circulating blood. Dysbiosis, increased gut permeability and chronic inflammation play a major role on the clinical outcome of inflammatory bowel diseases, graft-vs.-host disease (GVHD) and HIV infection. In this review, we focus on GVHD and HIV infection, conditions sharing gut immune damage leading to dysbiosis. The degree of dysbiosis and level of epithelial gut damage predict poor clinical outcome in both conditions. Emerging interventions are therefore warranted to promote gut microbiota homeostasis and improve intestinal barrier function. Interventions such as anti-inflammatory medications, and probiotics have toxicity and/or limited transitory effects, justifying innovative approaches. Fecal microbiota transplantation (FMT) is one such approach where fecal microorganisms are transferred from healthy donors into the GI tract of the recipient to restore microbiota composition in patients with Clostridium difficile-induced colitis or inflammatory bowel diseases. Preliminary findings point toward a beneficial effect of FMT to improve GVHD and HIV-related outcomes through the engraftment of beneficial donor bacteria, notably those producing anti-inflammatory metabolites. Herein, we critically review the potential for FMT in alleviating dysbiosis and gut damage in patients with GVHD or HIV-infection. Understanding the underlying mechanism by which FMT restores gut function will pave the way toward novel scalable and targeted interventions.
Introduction
Trillions of microorganisms reside in the human gut, encompassing not only bacteria but also fungi, archaea, viruses, and eukaryotic microbes, collectively termed microbiota. The gut microbiota was recently considered as an essential organ, playing a critical role in various host functions such as maintenance of the gut barrier and modulation of systemic immune response (1). Furthermore, the endocrine function of the gut microbiota was demonstrated through the production of vitamins and immunoregulatory short chain fatty acids (SCFA) (2). Dysregulation of gut microbiota composition, also known as dysbiosis, can lead to barrier dysfunction and translocation of microbial products leading to systemic inflammation (3). Recent evidence has shown that patients with diabetes, inflammatory bowel diseases (IBD), cancer, graft-vs.-host disease (GVHD) or HIV infection present with gut dysbiosis, gut damage, and microbial translocation (4–7).
Allo-hematopoietic stem cell transplantation (HSCT) is used in the treatment of hematological cancers where donor derived T-cells and natural killer cells target cancer cells in the recipient (4). Occurring after chemotherapy conditioning and HSCT, GVHD may develop as a serious complication when donor immune cells recognize the recipient as foreign and attack healthy cells in host's tissues. GVHD mostly occurs in the gut through the disruption of epithelial tight junctions, destruction of epithelial cells and inflammation in association with dysbiosis (5–8). A large multicenter study showed that gut microbiota composition independently predicted mortality in 1,362 HSCT patients with GVHD (9–11). Similarly, immune damage observed in the gut of people living with HIV (PLWH) was associated with gut dysbiosis, inflammation and clinical outcome (12–15). Despite long-term antiretroviral therapy (ART), damage to the gut mucosa and dysbiosis persist in PLWH, leading to systemic inflammation (8, 10, 11, 15). Like for people with GVHD, PLWH present with a disrupted gut epithelial barrier, immune-mediated intestinal damage, and increased gut permeability (15–22).
Given the association between microbiota composition and clinical outcome in both GVHD and HIV infection (5–11), strategies to modify the gut microbiota have come to light through dietary interventions, the antidiabetic drug metformin, selective antibiotics, probiotics, prebiotics, and fecal microbiota transplantation (FMT) (5, 23, 24). FMT refers to the transfer of fecal microorganisms from healthy donors into the GI tract of patients. It has shown to be effective in Clostridium difficile colitis (CDC), IBDs or obesity (25–28). As FMT has been recently shown to improve intestinal barrier function through promotion of gut microbiota homeostasis in GVHD and HIV infection, we discuss its relevance in both conditions in this review (29).
Dysbiosis and Increased Gut Permeability are Common Features in Patients With GVHD or HIV-Infection
In GVHD or HIV infection, a decrease of gut microbiota diversity is observed and associated with poor clinical outcome (30–33). Compared to patients undergoing allogeneic HSCT without GVHD, patients experiencing GVHD had decreased stool microbial diversity (32). Taur et al. reported that lower bacterial diversity was associated with increased transplant-related mortality in HSCT recipients (33). Nowak et al. also reported that the bacterial diversity of the gut microbiota was correlated with CD4 T-cell counts and inversely correlated with markers of microbial translocation and monocyte activation in PLWH (30).
The gut barrier is organized as a multi-layered and complex system which allows nutrient absorption while preventing the translocation of microbes and their products. Epithelial gut damage occurs in patients with GVHD and PLWH, with damaged enterocytes (basal barrier), non-functional Paneth cells (antimicrobial peptide production) and less mucosal-associated invariant T (MAIT) cells (5, 34–37). Several proteins have been used as gut damage markers. Plasma concentrations of regenerating islet-derived 3-alpha (REG3α), secreted by Paneth cells, were 3-fold higher in patients with gut GVHD at the onset of the disease compared to other HSCT patients (36, 37). Lower levels of REG3α at GVHD onset are correlated with higher 1 year survival (37). In PLWH, we observed that REG3α but not intestinal fatty acid binding protein (I-FABP) plasma levels were correlated with HIV disease progression, microbial translocation and immune activation (36). Similarly, soluble suppression of tumorigenicity (sST2) was also used to predict gut damage and clinical outcomes in patients with GVHD and PLWH (38–42).
Epithelial gut damage allows microbial translocation of microbial products from the lumen to the bloodstream, inducing local and systemic inflammation (43). Circulating levels of lipopolysaccharide (LPS), a pro-inflammatory bacterial cell wall component, is a clinically significant marker to assess the level of microbial translocation (44). LPS leakage in the circulation could induce innate immune activation, in association with mortality in GVHD (45–47). In PLWH, we and others have shown that LPS translocation is correlated with immune dysfunction and increased risk of non-AIDS comorbidities (48–51). Additionally, cytomegalovirus (CMV) primarily replicates in mucosal epithelial cells, decreasing gut barrier integrity. In patients with GVHD and PLWH, CMV latent infection or reactivation is associated with poor clinical outcomes (52–57). These findings suggest that patients with GVHD and HIV infection share similar features in gut damage and related microbial translocation.
Moreover, the gut microbiota can influence host cell physiology via production of metabolites such as SCFAs and bile acids. SCFAs, especially butyrate, constitute the primary energy source for colon epithelial cells. SCFAs play an important role in protecting intestinal barrier function, preventing microbial translocation and reducing inflammation through regulation of host epigenetics (58–60). GVHD patients or PLWH present with a lower abundance of SCFA-producing bacteria and a lower level of SCFAs, compared to non-GVHD HSCT patients or HIV-negative individuals, respectively (61–64). In both conditions, lower levels of SCFAs have been associated with gut damage and inflammation (62, 64–66). Conflicting results exist on the role of butyrate in GVHD as one report shows that patients developing GVHD had higher butyrate production (67). Furthermore, microbiota modulation leading to poor bile acids reabsorption could also be associated with gut damage in both patients with GVHD or PLWH (68–72).
Globally, gut dysbiosis, increased gut permeability, inflammation and systemic immune activation are common features of patients with GVHD or PLWH.
FMT in Patients With Gut GVHD
Given the dysbiosis and gut permeability in patients with GVHD, and regarding the vital role of gut microbiota in intestinal barrier and homeostasis, strategies targeting the microbiota offer one promising avenue for preventing or treating this condition. In the 1990s, investigators attempted to prevent the development of acute GVHD by drastically reducing the gut microbiota mass with antibiotics, removing the triggers of inflammation (73–75). However, newer studies have proven that gut microbiota-depleted patients had a higher risk of developing acute GVHD following HSCT than non-depleted patients (76, 77). Therefore, strategies promoting a “healthy” microbiota including FMT have attracted recent attention. Kakihana et al. (78) conducted a pilot study on four patients with steroid-resistant or steroid-dependent gut GVHD to observe the effects of FMT from spouses or relatives via nasoduodenal tube. All patients responded to FMT with three complete responses, one partial response, all in absence of severe adverse events. Spindelboeck et al. (79) reported successful FMT in three patients with severe acute GVHD. After one to six FMTs delivered via colonoscopy, all three patients showed increased diversity of the gut microbiota, with two complete remissions of GVHD and one partial remission. Qi et al. (80) reported eight patients with steroid-refractory gut GVHD receiving FMT through a nasoduodenal tube, from a stool bank. After FMT, all patients' clinical symptoms were relieved, bacteria diversity was enriched, and the gut microbiota diversity was restored. Compared to those who did not receive FMT, these eight patients achieved a longer progression-free survival. These case studies suggest that FMT can serve as a promising therapeutic option for gut GVHD, however larger controlled studies are required to confirm these effects.
FMT in PLWH
In PLWH, the mucosal immune system is disturbed by HIV infection. Th17 and Th22 cells, important components of mucosal immunity, are rapidly depleted following HIV or simian immunodeficiency virus (SIV) infection, contributing to a reduced barrier integrity, microbial translocation, and systemic immune activation (81–83). In a pilot study, Hensley-McBain et al. (84) reported that FMT significantly increases the number of peripheral Th17 and Th22 cells and reduced CD4 T-cell activation in the gut in SIV-infected macaques receiving ART. Moreover, the transplant was well-tolerated and no side effects were observed (84).
A pilot study in ART-suppressed individuals who received one-time FMT from stool bank via colonoscopy reported no serious adverse effects during the 24 weeks of follow-up. Microbial engraftment occurred but was partial, and limited to specific bacterial taxa including an increase of Faecalibacterium (85), which has been shown to exert anti-inflammatory effects in murine experimental colitis (86, 87). The authors considered that the limited effects of FMT might be related to the single dose of FMT given and the absence of antibiotic pre-treatment to “provide space” before FMT (85). Serrano-Villar et al. reported that repeated oral capsular FMT was one way to safely introduce incremental compositional changes into the gut microbiota in ART-treated PLWH (88). Compared to placebo, FMT significantly decreased the gut damage marker I-FABP 4 weeks after initiating FMT. Furthermore, mild engraftment of the donor's microbiota persisted until week 36 after initiating FMT and greater engraftment was observed among the four subjects who had received antibiotics in the 12 week period before FMT (88) (Figure 1).
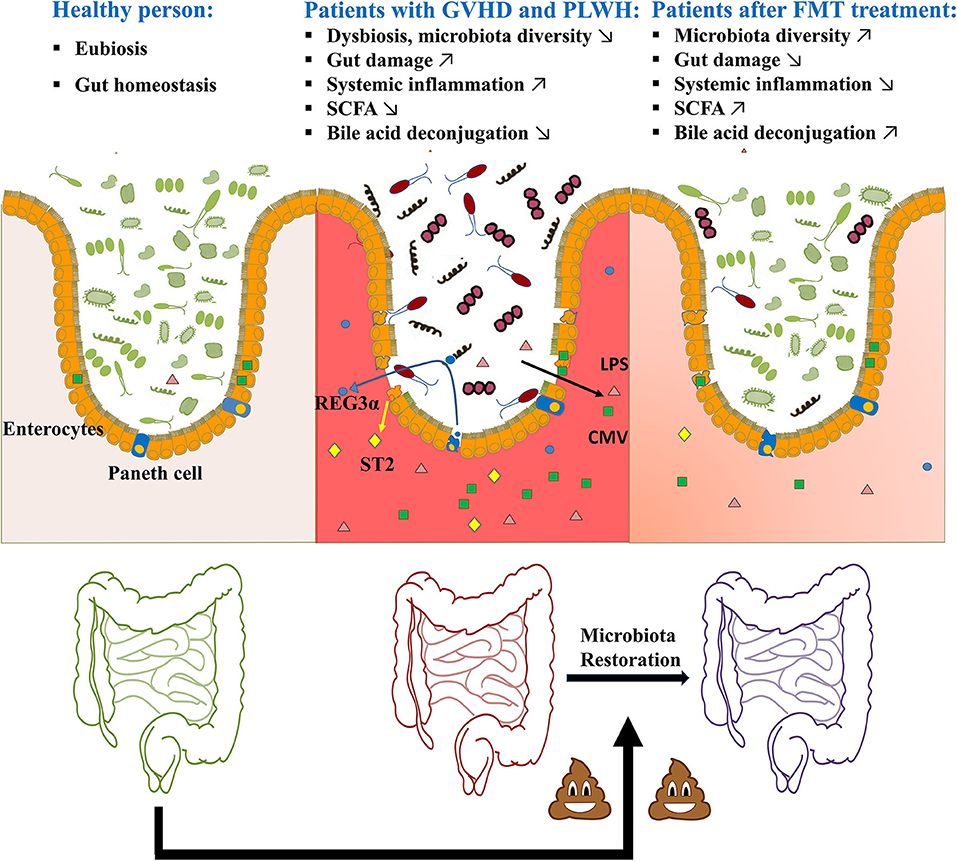
Figure 1. Potential of FMT in GVHD or HIV infection-related gut damage.
Safety should be the primary focus of any intervention. Concern persists on the safety of FMT administration, even more so in immunosuppressed recipients. However, PLWH with low CD4 T-cell count were shown to have the most profound modification of their gut microbiota and therefore would benefit greatly from FMT (89). As reviewed by Shogbesan et al. (27), FMT is successful in the treatment of recurrent CDC in immunocompromised patients including organ transplant recipients and PLWH. Encouragingly, FMT showed similar rates of adverse events in immunocompromised participants compared to immunocompetent ones including PLWH with CD4 counts lower than 200 cells/mm3 (90–93). Additionally, Schunemann et al. showed that FMT increased CD4 counts in an individual with HIV (94). To better assess the efficacy and safety of FMT, well-designed RCT clinical trials are ongoing and presented in Table 1. However, large studies assessing the influence of FMT in PLWH are still needed.
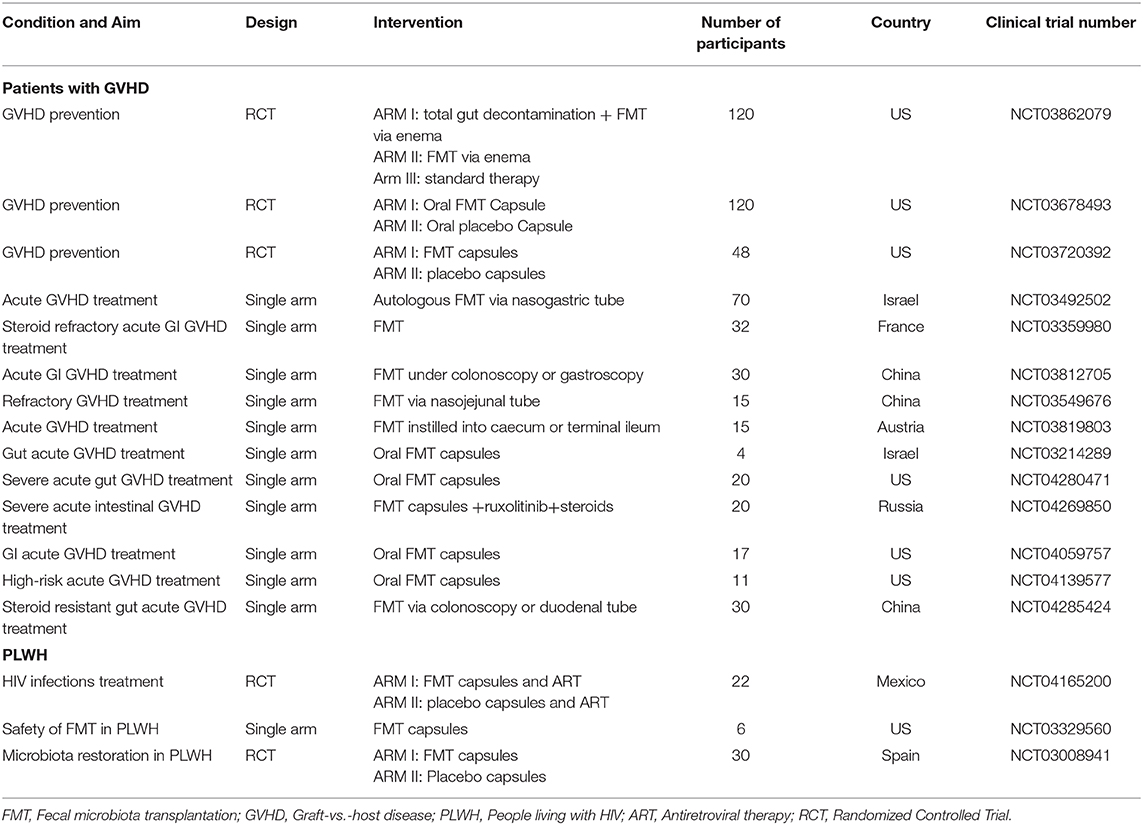
Table 1. Ongoing clinical trials using FMT as a treatment for GVHD and HIV.
Challenges of FMT for Patients With GVHD and PLWH
FMT needs further confirmation of its efficacy in decreasing gut damage in patient with GVHD or PLWH since studies assessing FMT with GVHD or PLWH involved a small number of participants. Moreover, safety needs to be validated as rare side effects may not have been observed in small studies. Therefore, challenges in designing formulations, preventing potential risks and implementing application in clinic for patients with GVHD and PLWH still remain.
Firstly, both healthy donors and patients have a microbiota composition with a high inter-person variability, and the key factors causing microbiota composition variation over time are not fully characterized. The precise influence of different microbiota composition and metabolites on epithelial barrier and clinical outcomes remain poorly understood and need further studies to define their distinctive role on the development of GVHD and HIV infection. Therefore, it remains difficult to select donors and special products for FMT formulation. Moreover, FMT treatment may carry pathogens for digestive and bloodstream infection, as DeFilipp et al. recently reported two cases of drug-resistant Escherichia coli bacteremia transmitted by FMT (95). Therefore, despite the absence of a uniform standard for “qualified” microbial communities, donors have to be thoroughly screened for transmissible diseases (e.g., HIV and hepatitis) and other non-infectious conditions (e.g., obesity and diabetes) that may be influenced by changes in the microbiome. In the light of Coronavirus Disease 2019, efforts to screen for novel infectious diseases should be implemented in the future. SARS-CoV-2, the virus that causes this disease, was found in stools even after diminution of respiratory symptoms and could be transmitted through a fecal-oral route (96, 97). Donors who may transfer undesirable agents (e.g., antibiotics, anti-acid proton pump inhibitors, systemic immunosuppressive agents, antineoplastic agents, and glucocorticoids) which can affect the safety or efficacy of FMT should also be excluded (98). Hence, screening for potential donors is costly and time consuming (99). Fortunately, new techniques allow freezing and storage of donor stools for extended periods of time, possibly facilitating FMT implementation (100).
As donor selection is a difficult process, and in order to favor clinical improvement, engraftment of the donor's microbiota should be optimal. Antibiotic conditioning given to the recipient just before FMT seems to improve microbiota engraftment (88). This procedure may destabilize the existing microbial community and promote engraftment of another community. By preventing niche competition in the mucosa between the xenomicrobiota and indigenous microbiota, preparing the gut with antibiotics was shown to facilitate xenomicrobiota colonization, thus enhancing the overall gut microbiota modification efficiency (101). Preliminary results by Serrano-villar et al. showed greater engraftment in four PLWH who had received antibiotics before FMT (88). Pre-therapy with antibiotics before FMT to alleviate GVHD is currently under study (NCT03862079, Table 1).
Encouragingly, multiple clinical trials studying the potential of FMT as a treatment for GVHD or HIV-related gut damage are ongoing (Table 1). In these trials, several routes of administration for FMT are under investigations, including oral capsules, nasal tube, colonoscopy, or enema. The optimal administration route may depend on the characteristics of the disease, and general condition of the patient. Compared with enema, colonoscopy could deliver the FMT to deep cecum, and increase engraftment while the donor stools are expelled less rapidly. However, colonoscopy remains a relatively invasive procedure (102): Kelly et al. reported one case of death from lung-aspiration injury during sedation for FMT administered via colonoscopy (103). Furthermore, nasal administration is considered inconvenient as some cases of intestinal bleeding and rare peritonitis have been reported (104). However, oral capsules have been developed to pass through the acidic environment of the stomach and ensure a delayed delivery of live microbial communities into the intestine (105). By using questionnaires, this route is considered to be most convenient for patients. Kao et al. compared oral capsule and colonoscopy delivered FMT on recurrent CDC showed similar efficacy, with less adverse events (106). Further studies should analyze the preferential route of FMT to alleviate gut damage patients in GVHD and PLWH.
Conclusion
Both gut GVHD and HIV infection have been associated with dysbiosis and increased gut permeability, contributing to microbial translocation, inflammation, and poor clinical outcomes. Progress has been made in discerning the role of the microbiota in GVHD patients and PLWH. Manipulating the gut microbiota with FMT has been successfully used to treat CDC through microbiota restoration and has paved the way as a novel strategy to improve the outcomes of GVHD patients and PLWH. Several clinical trials are ongoing to assess the efficacy and safety of treating GVHD and HIV-induced gut damage with FMT. However, most trials and published studies are pilot or case series, thus making it difficult to confirm its efficacy and safety. Only large multicentre RCT studies will address the merit of such intervention. Moreover, a standard FMT procedure needs to be implemented and described, including pre-treatment with antibiotics and delivery with oral capsules to favor engraftment. Overall, collaborative efforts encompassing microbiology, clinical care, and pharmacy will define the optimal procedure and number of FMT to obtain a significant and lasting benefit from FMT for individuals with GVHD and HIV. In the future, FMT will pave avenues toward the characterization of important species and their metabolites in modulation of gut damage in patient with GVHD or PLWH, leading to more effective interventions.
Author Contributions
JO and SI wrote the first draft of the manuscript. JL, BF, XP, SN, YC, and MS provided critical revision of the manuscript. J-PR conceived and designed the manuscript. All authors approved it for publication.
Funding
Our research is funded by the Fonds de la Recherche Québec-Santé (FRQ-S): Réseau SIDA/Maladies infectieuses and Thérapie cellulaire; the Canadian Institutes of Health Research (CIHR; grants MOP 103230 and PTJ 166049); the Vaccines & Immunotherapies Core of the CIHR Canadian HIV Trials Network (CTN; CTN PT038); the Canadian Foundation for AIDS Research (CANFAR; grant 02-512); CIHR- funded Canadian HIV Cure Enterprise (CanCURE) Team Grant HB2-164064; and Chongqing Basic and Frontier Research project (No. cstc2018jcyjAX0652).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We are highly grateful to Angie Massicotte, Josée Girouard, and Cezar Iovi for coordination and assistance.
References
1. Sircana A, Framarin L, Leone N, Berrutti M, Castellino F, Parente R, et al. Altered gut microbiota in type 2 diabetes: just a coincidence? Curr Diab Rep. (2018) 18:98. doi: 10.1007/s11892-018-1057-6
2. Lynch SV, Pedersen O. The human intestinal microbiome in health and disease. N Engl J Med. (2016) 375:2369–79. doi: 10.1056/NEJMra1600266
3. Vinolo MA, Rodrigues HG, Nachbar RT, Curi R. Regulation of inflammation by short chain fatty acids. Nutrients. (2011) 3:858–76. doi: 10.3390/nu3100858
4. Ingham AC, Kielsen K, Cilieborg MS, Lund O, Holmes S, Aarestrup FM, et al. Specific gut microbiome members are associated with distinct immune markers in pediatric allogeneic hematopoietic stem cell transplantation. Microbiome. (2019) 7:131. doi: 10.1186/s40168-019-0745-z
5. Fredricks DN. The gut microbiota and graft-versus-host disease. J Clini Invest. (2019) 129:1808–17. doi: 10.1172/JCI125797
6. Ghimire S, Weber D, Mavin E, Wang XN, Dickinson AM, Holler E. Pathophysiology of GvHD and other HSCT-related major complications. Front Immunol. (2017) 8:79. doi: 10.3389/fimmu.2017.00079
7. Nalle SC, Zuo L, Ong M, Singh G, Worthylake AM, Choi W, et al. Graft-versus-host disease propagation depends on increased intestinal epithelial tight junction permeability. J Clin Invest. (2019) 129:902–14. doi: 10.1172/JCI98554
8. Gilliam AC, Whitaker-Menezes D, Korngold R, Murphy GF. Apoptosis is the predominant form of epithelial target cell injury in acute experimental graft-versus-host disease. J Invest Dermatol. (1996) 107:377–83. doi: 10.1111/1523-1747.ep12363361
9. Peled JU, Gomes ALC, Devlin SM, Littmann ER, Taur Y, Sung AD, et al. Microbiota as predictor of mortality in allogeneic hematopoietic-cell transplantation. N Engl J Med. (2020) 382:822–34. doi: 10.1056/NEJMoa1900623
10. Zitvogel L, Kroemer G. Immunostimulatory gut bacteria. Science. (2019) 366:1077–8. doi: 10.1126/science.aaz7595
11. Stein-Thoeringer CK, Nichols KB, Lazrak A, Docampo MD, Slingerland AE, Slingerland JB, et al. Lactose drives Enterococcus expansion to promote graft-versus-host disease. Science. (2019) 366:1143–9. doi: 10.1126/science.aax3760
12. Gianella S, Letendre SL, Iudicello J, Franklin D, Gaufin T, Zhang Y, et al. Plasma (1 –> 3)-beta-D-glucan and suPAR levels correlate with neurocognitive performance in people living with HIV on antiretroviral therapy: a CHARTER analysis. J Neurovirol. (2019) 25:837–43. doi: 10.1007/s13365-019-00775-6
13. Hoenigl M, de Oliveira MF, Perez-Santiago J, Zhang Y, Morris S, McCutchan AJ, et al. (1–>3)-beta-D-Glucan levels correlate with neurocognitive functioning in HIV-infected persons on suppressive antiretroviral therapy: a cohort study. Medicine. (2016) 95:e3162. doi: 10.1097/MD.0000000000003162
14. Gelpi M, Vestad B, Hansen SH, Holm K, Drivsholm N, Goetz A, et al. Impact of HIV-related gut microbiota alterations on metabolic comorbidities. Clin Infect Dis. (2020). doi: 10.1093/cid/ciz1235. [Epub ahead of print].
15. Vujkovic-Cvijin I, Sortino O, Verheij E, Sklar J, Wit FW, Kootstra NA, et al. HIV-associated gut dysbiosis is independent of sexual practice and correlates with noncommunicable diseases. Nat Commun. (2020) 11:2448. doi: 10.1038/s41467-020-16222-8
16. Mehandru S, Tenner-Racz K, Racz P, Markowitz M. The gastrointestinal tract is critical to the pathogenesis of acute HIV-1 infection. J Allergy Clin Immunol. (2005) 116:419–22. doi: 10.1016/j.jaci.2005.05.040
17. Brenchley JM, Douek DC. HIV infection and the gastrointestinal immune system. Mucosal Immunol. (2008) 1:23–30. doi: 10.1038/mi.2007.1
18. Estes JD, Kityo C, Ssali F, Swainson L, Makamdop KN, Del Prete GQ, et al. Defining total-body AIDS-virus burden with implications for curative strategies. Nat Med. (2017) 23:1271–6. doi: 10.1038/nm.4411
19. Hensley-McBain T, Berard AR, Manuzak JA, Miller CJ, Zevin AS, Polacino P, et al. Intestinal damage precedes mucosal immune dysfunction in SIV infection. Mucosal Immunol. (2018) 11:1429–40. doi: 10.1038/s41385-018-0032-5
20. Batman PA, Miller AR, Forster SM, Harris JR, Pinching AJ, Griffin GE. Jejunal enteropathy associated with human immunodeficiency virus infection: quantitative histology. J Clin Pathol. (1989) 42:275–81. doi: 10.1136/jcp.42.3.275
21. Batman PA, Kotler DP, Kapembwa MS, Booth D, Potten CS, Orenstein JM, et al. HIV enteropathy: crypt stem and transit cell hyperproliferation induces villous atrophy in HIV/Microsporidia-infected jejunal mucosa. AIDS. (2007) 21:433–9. doi: 10.1097/QAD.0b013e3280142ee8
22. Heise C, Dandekar S, Kumar P, Duplantier R, Donovan RM, Halsted CH. Human immunodeficiency virus infection of enterocytes and mononuclear cells in human jejunal mucosa. Gastroenterology. (1991) 100:1521–7. doi: 10.1016/0016-5085(91)90648-5
23. Li J, Riaz Rajoka MS, Shao D, Jiang C, Jin M, Huang Q, et al. Strategies to increase the efficacy of using gut microbiota for the modulation of obesity. Obes Rev. (2017) 18:1260–71. doi: 10.1111/obr.12590
24. Vujkovic-Cvijin I, Somsouk M. HIV and the gut microbiota: composition, consequences, and avenues for amelioration. Curr HIV/AIDS Rep. (2019) 16:204–13. doi: 10.1007/s11904-019-00441-w
25. Cammarota G, Ianiro G, Tilg H, Rajilic-Stojanovic M, Kump P, Satokari R, et al. European consensus conference on faecal microbiota transplantation in clinical practice. Gut. (2017) 66:569–80. doi: 10.1136/gutjnl-2016-313017
26. Mullish BH, Quraishi MN, Segal JP, McCune VL, Baxter M, Marsden GL, et al. The use of faecal microbiota transplant as treatment for recurrent or refractory Clostridium difficile infection and other potential indications: joint British society of gastroenterology (BSG) and healthcare infection society (HIS) guidelines. Gut. (2018) 67:1920–41. doi: 10.1136/gutjnl-2018-316818
27. Shogbesan O, Poudel DR, Victor S, Jehangir A, Fadahunsi O, Shogbesan G, et al. a systematic review of the efficacy and safety of fecal microbiota transplant for Clostridium difficile infection in immunocompromised patients. Can J Gastroenterol Hepatol. (2018) 2018:1394379. doi: 10.1155/2018/1394379
28. Konturek PC, Koziel J, Dieterich W, Haziri D, Wirtz S, Glowczyk I, et al. Successful therapy of Clostridium difficile infection with fecal microbiota transplantation. J Physiol Pharmacol. (2016) 67:859–66. doi: 10.1016/S0016-5085(17)31403-8
29. Kang Y, Cai Y. Altered gut microbiota in hiv infection: future perspective of fecal microbiota transplantation therapy. AIDS Res Hum Retroviruses. (2019) 35:229–35. doi: 10.1089/aid.2017.0268
30. Nowak P, Troseid M, Avershina E, Barqasho B, Neogi U, Holm K, et al. Gut microbiota diversity predicts immune status in HIV-1 infection. AIDS. (2015) 29:2409–18. doi: 10.1097/QAD.0000000000000869
31. Tuddenham SA, Koay WLA, Zhao N, White JR, Ghanem KG, Sears CL. The impact of human immunodeficiency virus infection on gut microbiota alpha-diversity: an individual-level meta-analysis. Clin Infect Dis. (2020) 70:615–27. doi: 10.1093/cid/ciz258
32. Jenq RR, Ubeda C, Taur Y, Menezes CC, Khanin R, Dudakov JA, et al. Regulation of intestinal inflammation by microbiota following allogeneic bone marrow transplantation. J Exp Med. (2012) 209:903–11. doi: 10.1084/jem.20112408
33. Taur Y, Jenq RR, Perales MA, Littmann ER, Morjaria S, Ling L, et al. The effects of intestinal tract bacterial diversity on mortality following allogeneic hematopoietic stem cell transplantation. Blood. (2014) 124:1174–82. doi: 10.1182/blood-2014-02-554725
34. Troeger H, Hering NA, Bojarski C, Fromm A, Barmeyer C, Uharek L, et al. Epithelial barrier dysfunction as permissive pathomechanism in human intestinal graft-versus-host disease. Bone Marrow Transplant. (2018) 53:1083–6. doi: 10.1038/s41409-018-0144-2
35. Nalle SC, Turner JR. Endothelial and epithelial barriers in graft-versus-host disease. Adv Exp Med Biol. (2012) 763:105–31. doi: 10.1007/978-1-4614-4711-5_5
36. Isnard S, Ramendra R, Dupuy FP, Lin J, Fombuena B, Kokinov N, et al. Plasma levels of C-type lectin REG3alpha and gut damage in people with human immunodeficiency virus. J Infect Dis. (2020) 221:110–21. doi: 10.1093/infdis/jiz423
37. Ferrara JL, Harris AC, Greenson JK, Braun TM, Holler E, Teshima T, et al. Regenerating islet-derived 3-alpha is a biomarker of gastrointestinal graft-versus-host disease. Blood. (2011) 118:6702–8. doi: 10.1182/blood-2011-08-375006
38. Vander Lugt MT, Braun TM, Hanash S, Ritz J, Ho VT, Antin JH, et al. ST2 as a marker for risk of therapy-resistant graft-versus-host disease and death. N Engl J Med. (2013) 369:529–39. doi: 10.1056/NEJMoa1213299
39. Rowan C, Pike F, Cooke KR, Krance R, Carpenter PA, Duncan C, et al. Assessment of ST2 for risk of death following graft-versus-host disease in the pediatric and adult age groups. Blood. (2020) 135:1428–37. doi: 10.2139/ssrn.3409311
40. Younas M, Psomas C, Mehraj V, Cezar R, Portales P, Tuaillon E, et al. Plasma level of soluble ST2 in chronically infected HIV-1 patients with suppressed viremia. Open AIDS J. (2017) 11:32–5. doi: 10.2174/1874613601711010032
41. Mehraj V, Jenabian MA, Ponte R, Lebouche B, Costiniuk C, Thomas R, et al. The plasma levels of soluble ST2 as a marker of gut mucosal damage in early HIV infection. AIDS. (2016) 30:1617–27. doi: 10.1097/QAD.0000000000001105
42. Mehraj V, Ponte R, Routy JP. The dynamic role of the IL-33/ST2 axis in chronic viral-infections: alarming and adjuvanting the immune response. EBioMedicine. (2016) 9:37–44. doi: 10.1016/j.ebiom.2016.06.047
43. Mu J, Chen Q, Zhu L, Wu Y, Liu S, Zhao Y, et al. Influence of gut microbiota and intestinal barrier on enterogenic infection after liver transplantation. Curr Med Res Opin. (2019) 35:241–8. doi: 10.1080/03007995.2018.1470085
44. Brenchley JM, Price DA, Schacker TW, Asher TE, Silvestri G, Rao S, et al. Microbial translocation is a cause of systemic immune activation in chronic HIV infection. Nat Med. (2006) 12:1365–71. doi: 10.1038/nm1511
45. Price KS, Nestel FP, Lapp WS. Progressive accumulation of bacterial lipopolysaccharide in vivo during murine acute graft-versus-host disease. Scand J Immunol. (1997) 45:294–300. doi: 10.1046/j.1365-3083.1997.d01-404.x
46. Daguindau E, Gautier T, Chague C, Pais de Barros JP, Deckert V, Lagrost L, et al. Is it time to reconsider the lipopolysaccharide paradigm in acute graft-versus-host disease? Front Immunol. (2017) 8:952. doi: 10.3389/fimmu.2017.00952
47. Hill GR, Crawford JM, Cooke KR, Brinson YS, Pan L, Ferrara JL. Total body irradiation and acute graft-versus-host disease: the role of gastrointestinal damage and inflammatory cytokines. Blood. (1997) 90:3204–13. doi: 10.1182/blood.V90.8.3204
48. Jenabian MA, El-Far M, Vyboh K, Kema I, Costiniuk CT, Thomas R, et al. Immunosuppressive tryptophan catabolism and gut mucosal dysfunction following early HIV infection. J Infect Dis. (2015) 212:355–66. doi: 10.1093/infdis/jiv037
49. Ramendra R, Isnard S, Mehraj V, Chen J, Zhang Y, Finkelman M, et al. Circulating LPS and (1–>3)-beta-D-Glucan: a folie a deux contributing to HIV-associated immune activation. Front Immunol. (2019) 10:465. doi: 10.3389/fimmu.2019.00465
50. Morris A, Hillenbrand M, Finkelman M, George MP, Singh V, Kessinger C, et al. Serum (1–>3)-beta-D-glucan levels in HIV-infected individuals are associated with immunosuppression, inflammation, and cardiopulmonary function. J Acquir Immune Defic Syndr. (2012) 61:462–8. doi: 10.1097/QAI.0b013e318271799b
51. Xu W, Luo Z, Alekseyenko AV, Martin L, Wan Z, Ling B, et al. Distinct systemic microbiome and microbial translocation are associated with plasma level of anti-CD4 autoantibody in HIV infection. Sci Rep. (2018) 8:12863. doi: 10.1038/s41598-018-31116-y
52. Maidji E, Somsouk M, Rivera JM, Hunt PW, Stoddart CA. Replication of CMV in the gut of HIV-infected individuals and epithelial barrier dysfunction. PLoS Pathog. (2017) 13:e1006202. doi: 10.1371/journal.ppat.1006202
53. Ramendra R, Isnard S, Lin J, Fombuena B, Ouyang J, Mehraj V, et al. CMV seropositivity is associated with increased microbial translocation in people living with HIV and uninfected controls. Clin Infect Dis. (2019). doi: 10.1093/cid/ciz1001. [Epub ahead of print].
54. Collins-McMillen D, Buehler J, Peppenelli M, Goodrum F. Molecular determinants and the regulation of human cytomegalovirus latency and reactivation. Viruses. (2018) 10:444. doi: 10.3390/v10080444
55. Camargo JF, Wieder ED, Kimble E, Benjamin CL, Kolonias DS, Kwon D, et al. Deep functional immunophenotyping predicts risk of cytomegalovirus reactivation after hematopoietic cell transplantation. Blood. (2019) 133:867–77. doi: 10.1182/blood-2018-10-878918
56. Cantoni N, Hirsch HH, Khanna N, Gerull S, Buser A, Bucher C, et al. Evidence for a bidirectional relationship between cytomegalovirus replication and acute graft-versus-host disease. Biol Blood Marrow Transplant. (2010) 16:1309–14. doi: 10.1016/j.bbmt.2010.03.020
57. Broers AE, van Der Holt R, van Esser JW, Gratama JW, Henzen-Logmans S, Kuenen-Boumeester V, et al. Increased transplant-related morbidity and mortality in CMV-seropositive patients despite highly effective prevention of CMV disease after allogeneic T-cell-depleted stem cell transplantation. Blood. (2000) 95:2240–5. doi: 10.1182/blood.V95.7.2240.007k08_2240_2245
58. Woo V, Alenghat T. Host-microbiota interactions: epigenomic regulation. Curr Opin Immunol. (2017) 44:52–60. doi: 10.1016/j.coi.2016.12.001
59. Wang HB, Wang PY, Wang X, Wan YL, Liu YC. Butyrate enhances intestinal epithelial barrier function via up-regulation of tight junction protein Claudin-1 transcription. Dig Dis Sci. (2012) 57:3126–35. doi: 10.1007/s10620-012-2259-4
60. Tan J, McKenzie C, Potamitis M, Thorburn AN, Mackay CR, Macia L. The role of short-chain fatty acids in health and disease. Adv Immunol. (2014) 121:91–119. doi: 10.1016/B978-0-12-800100-4.00003-9
61. Gonzalez-Hernandez LA, Ruiz-Briseno MDR, Sanchez-Reyes K, Alvarez-Zavala M, Vega-Magana N, Lopez-Iniguez A, et al. Alterations in bacterial communities, SCFA and biomarkers in an elderly HIV-positive and HIV-negative population in western Mexico. BMC Infect Dis. (2019) 19:234. doi: 10.1186/s12879-019-3867-9
62. Dillon SM, Kibbie J, Lee EJ, Guo K, Santiago ML, Austin GL, et al. Low abundance of colonic butyrate-producing bacteria in HIV infection is associated with microbial translocation and immune activation. AIDS. (2017) 31:511–21. doi: 10.1097/QAD.0000000000001366
63. Biagi E, Zama D, Nastasi C, Consolandi C, Fiori J, Rampelli S, et al. Gut microbiota trajectory in pediatric patients undergoing hematopoietic SCT. Bone Marrow Transplant. (2015) 50:992–8. doi: 10.1038/bmt.2015.16
64. Mathewson ND, Jenq R, Mathew AV, Koenigsknecht M, Hanash A, Toubai T, et al. Gut microbiome-derived metabolites modulate intestinal epithelial cell damage and mitigate graft-versus-host disease. Nat Immunol. (2016) 17:505–13. doi: 10.1038/ni.3400
65. Kohler N, Zeiser R. Intestinal microbiota influence immune tolerance post allogeneic hematopoietic cell transplantation and intestinal GVHD. Front Immunol. (2018) 9:3179. doi: 10.3389/fimmu.2018.03179
66. Serrano-Villar S, Vazquez-Castellanos JF, Vallejo A, Latorre A, Sainz T, Ferrando-Martinez S, et al. The effects of prebiotics on microbial dysbiosis, butyrate production and immunity in HIV-infected subjects. Mucosal Immunol. (2017) 10:1279–93. doi: 10.1038/mi.2016.122
67. Golob JL, DeMeules MM, Loeffelholz T, Quinn ZZ, Dame MK, Silvestri SS, et al. Butyrogenic bacteria after acute graft-versus-host disease (GVHD) are associated with the development of steroid-refractory GVHD. Blood Adv. (2019) 3:2866–9. doi: 10.1182/bloodadvances.2019000362
68. Sayin SI, Wahlstrom A, Felin J, Jantti S, Marschall HU, Bamberg K, et al. Gut microbiota regulates bile acid metabolism by reducing the levels of tauro-beta-muricholic acid, a naturally occurring FXR antagonist. Cell Metab. (2013) 17:225–35. doi: 10.1016/j.cmet.2013.01.003
69. Inagaki T, Moschetta A, Lee YK, Peng L, Zhao G, Downes M, et al. Regulation of antibacterial defense in the small intestine by the nuclear bile acid receptor. Proc Natl Acad Sci USA. (2006) 103:3920–5. doi: 10.1073/pnas.0509592103
70. Cramp ME, Hing MC, Marriott DJ, Freund J, Cooper DA. Bile acid malabsorption in HIV infected patients with chronic diarrhoea. Aust N Z J Med. (1996) 26:368–71. doi: 10.1111/j.1445-5994.1996.tb01924.x
71. Steuerwald M, Bucher HC, Muller-Brand J, Gotze M, Roser HW, Gyr K. HIV-enteropathy and bile acid malabsorption: response to cholestyramine. Am J Gastroenterol. (1995) 90:2051–3.
72. Joshi NM, Hassan S, Jasani P, Dixon S, Cavenagh JD, Oakervee HE, et al. Bile acid malabsorption in patients with graft-versus-host disease of the gastrointestinal tract. Br J Haematol. (2012) 157:403–7. doi: 10.1111/j.1365-2141.2011.09014.x
73. Vossen JM, Heidt PJ, van den Berg H, Gerritsen EJ, Hermans J, Dooren LJ. Prevention of infection and graft-versus-host disease by suppression of intestinal microflora in children treated with allogeneic bone marrow transplantation. Eur J Clin Microbiol Infect Dis. (1990) 9:14–23. doi: 10.1007/BF01969527
74. Beelen DW, Haralambie E, Brandt H, Linzenmeier G, Muller KD, Quabeck K, et al. Evidence that sustained growth suppression of intestinal anaerobic bacteria reduces the risk of acute graft-versus-host disease after sibling marrow transplantation. Blood. (1992) 80:2668–76. doi: 10.1182/blood.V80.10.2668.bloodjournal80102668
75. Beelen DW, Elmaagacli A, Muller KD, Hirche H, Schaefer UW. Influence of intestinal bacterial decontamination using metronidazole and ciprofloxacin or ciprofloxacin alone on the development of acute graft-versus-host disease after marrow transplantation in patients with hematologic malignancies: final results and long-term follow-up of an open-label prospective randomized trial. Blood. (1999) 93:3267–75. doi: 10.1182/blood.V93.10.3267.410k22_3267_3275
76. Routy B, Letendre C, Enot D, Chenard-Poirier M, Mehraj V, Seguin NC, et al. The influence of gut-decontamination prophylactic antibiotics on acute graft-versus-host disease and survival following allogeneic hematopoietic stem cell transplantation. Oncoimmunology. (2017) 6:e1258506. doi: 10.1080/2162402X.2016.1258506
77. Gjaerde LK, Schmidt M, Sengelov H. Gut decontamination during allogeneic hematopoietic stem cell transplantation and the risk of acute graft-versus-host disease. Bone Marrow Transplant. (2018) 53:1061–4. doi: 10.1038/s41409-018-0131-7
78. Kakihana K, Fujioka Y, Suda W, Najima Y, Kuwata G, Sasajima S, et al. Fecal microbiota transplantation for patients with steroid-resistant acute graft-versus-host disease of the gut. Blood. (2016) 128:2083–8. doi: 10.1182/blood-2016-05-717652
79. Spindelboeck W, Schulz E, Uhl B, Kashofer K, Aigelsreiter A, Zinke-Cerwenka W, et al. Repeated fecal microbiota transplantations attenuate diarrhea and lead to sustained changes in the fecal microbiota in acute, refractory gastrointestinal graft-versus-host-disease. Haematologica. (2017) 102:e210–3. doi: 10.3324/haematol.2016.154351
80. Qi X, Li X, Zhao Y, Wu X, Chen F, Ma X, et al. Treating steroid refractory intestinal acute graft-vs.-host disease with fecal microbiota transplantation: a pilot study. Front Immunol. (2018) 9:2195. doi: 10.3389/fimmu.2018.02195
81. Klatt NR, Estes JD, Sun X, Ortiz AM, Barber JS, Harris LD, et al. Loss of mucosal CD103+ DCs and IL-17+ and IL-22+ lymphocytes is associated with mucosal damage in SIV infection. Mucosal Immunol. (2012) 5:646–57. doi: 10.1038/mi.2012.38
82. Klatt NR, Brenchley JM. Th17 cell dynamics in HIV infection. Curr Opin HIV AIDS. (2010) 5:135–40. doi: 10.1097/COH.0b013e3283364846
83. Brenchley JM, Paiardini M, Knox KS, Asher AI, Cervasi B, Asher TE, et al. Differential Th17 CD4 T-cell depletion in pathogenic and nonpathogenic lentiviral infections. Blood. (2008) 112:2826–35. doi: 10.1182/blood-2008-05-159301
84. Hensley-McBain T, Zevin AS, Manuzak J, Smith E, Gile J, Miller C, et al. Effects of fecal microbial transplantation on microbiome and immunity in simian immunodeficiency virus-infected macaques. J Virol. (2016) 90:4981–9. doi: 10.1128/JVI.00099-16
85. Paquin-Proulx D, Ching C, Vujkovic-Cvijin I, Fadrosh D, Loh L, Huang Y, et al. Bacteroides are associated with GALT iNKT cell function and reduction of microbial translocation in HIV-1 infection. Mucosal Immunol. (2017) 10:69–78. doi: 10.1038/mi.2016.34
86. Sokol H, Pigneur B, Watterlot L, Lakhdari O, Bermudez-Humaran LG, Gratadoux JJ, et al. Faecalibacterium prausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota analysis of Crohn disease patients. Proc Natl Acad Sci USA. (2008) 105:16731–6. doi: 10.1073/pnas.0804812105
87. Frank DN, St. Amand AL, Feldman RA, Boedeker EC, Harpaz N, Pace NR. Molecular-phylogenetic characterization of microbial community imbalances in human inflammatory bowel diseases. Proc Natl Acad Sci USA. (2007) 104:13780–5. doi: 10.1073/pnas.0706625104
88. Serrano-Villar S, Talavera A, Madrid-Elena N, Pérez-Molina JA, Gosalbes MJ, Budree S, et al. Rational donor fecal microbiota transplantation in HIV (refresh study). In: Conference on Retroviruses and Opportunistic Infections (CROI). Boston, MA (2020).
89. Lu W, Feng Y, Jing F, Han Y, Lyu N, Liu F, et al. Association between gut microbiota and CD4 recovery in HIV-1 infected patients. Front Microbiol. (2018) 9:1451. doi: 10.3389/fmicb.2018.01451
90. Mandalia A, Ward A, Tauxe W, Kraft CS, Dhere T. Fecal transplant is as effective and safe in immunocompromised as non-immunocompromised patients for Clostridium difficile. Int J Colorectal Dis. (2016) 31:1059–60. doi: 10.1007/s00384-015-2396-2
91. Ott SJ, Waetzig GH, Rehman A, Moltzau-Anderson J, Bharti R, Grasis JA, et al. Efficacy of sterile fecal filtrate transfer for treating patients with Clostridium difficile infection. Gastroenterology. (2017) 152:799–811.e7. doi: 10.1053/j.gastro.2016.11.010
92. Elopre L, Rodriguez M. Fecal microbiota therapy for recurrent Clostridium difficile infection in HIV-infected persons. Ann Intern Med. (2013) 158:779–80. doi: 10.7326/0003-4819-158-10-201305210-00021
93. Gathe JC Jr, Diejomaoh EM, Mayberry CC, Clemmons JB. Fecal transplantation for Clostridium difficile-“all stool may not be created equal”. J Int Assoc Provid AIDS Care. (2016) 15:107–8. doi: 10.1177/2325957415627695
94. Schunemann M, Oette M. Fecal microbiota transplantation for Clostridium difficile-associated colitis in a severely immunocompromized critically ill AIDS patient: a case report. AIDS. (2014) 28:798–9. doi: 10.1097/QAD.0000000000000148
95. DeFilipp Z, Bloom PP, Torres Soto M, Mansour MK, Sater MRA, Huntley MH, et al. Drug-resistant E. coli bacteremia transmitted by fecal microbiota transplant. N Engl J Med. (2019) 381:2043–50. doi: 10.1056/NEJMoa1910437
96. Song Y, Liu P, Shi XL, Chu YL, Zhang J, Xia J, et al. SARS-CoV-2 induced diarrhoea as onset symptom in patient with COVID-19. Gut. (2020) 69:1143–4. doi: 10.1136/gutjnl-2020-320891
97. Wu Y, Guo C, Tang L, Hong Z, Zhou J, Dong X, et al. Prolonged presence of SARS-CoV-2 viral RNA in faecal samples. Lancet Gastroenterol Hepatol. (2020) 5:434–35. doi: 10.1016/S2468-1253(20)30083-2
98. Canada H. Guidance Document: Fecal Microbiota Therapy Used in the Treatment of Clostridium difficile Infection Not Responsive to Conventional Therapies: Health Canada. (2019). Available online at: https://www.canada.ca/en/health-canada/services/drugs-health-products/biologics-radiopharmaceuticals-genetic-therapies/applications-submissions/guidance-documents/regulation-fecal-microbiota-therapy-treatment-difficile-infections.html (accessed February 21, 2020).
99. Craven LJ, Nair Parvathy S, Tat-Ko J, Burton JP, Silverman MS. Extended screening costs associated with selecting donors for fecal microbiota transplantation for treatment of metabolic syndrome-associated diseases. Open Forum Infect Dis. (2017) 4:ofx243. doi: 10.1093/ofid/ofx243
100. Hamilton MJ, Weingarden AR, Sadowsky MJ, Khoruts A. Standardized frozen preparation for transplantation of fecal microbiota for recurrent Clostridium difficile infection. Am J Gastroenterol. (2012) 107:761–7. doi: 10.1038/ajg.2011.482
101. Ji SK, Yan H, Jiang T, Guo CY, Liu JJ, Dong SZ, et al. Preparing the gut with antibiotics enhances gut microbiota reprogramming efficiency by promoting xenomicrobiota colonization. Front Microbiol. (2017) 8:1208. doi: 10.3389/fmicb.2017.01208
102. Konig J, Siebenhaar A, Hogenauer C, Arkkila P, Nieuwdorp M, Noren T, et al. Consensus report: faecal microbiota transfer - clinical applications and procedures. Aliment Pharmacol Ther. (2017) 45:222–39. doi: 10.1111/apt.13868
103. Kelly CR, Ihunnah C, Fischer M, Khoruts A, Surawicz C, Afzali A, et al. Fecal microbiota transplant for treatment of Clostridium difficile infection in immunocompromised patients. Am J Gastroenterol. (2014) 109:1065–71. doi: 10.1038/ajg.2014.133
104. Kassam Z, Lee CH, Yuan Y, Hunt RH. Fecal microbiota transplantation for Clostridium difficile infection: systematic review and meta-analysis. Am J Gastroenterol. (2013) 108:500–8. doi: 10.1038/ajg.2013.59
105. Youngster I, Russell GH, Pindar C, Ziv-Baran T, Sauk J, Hohmann EL. Oral, capsulized, frozen fecal microbiota transplantation for relapsing Clostridium difficile infection. JAMA. (2014) 312:1772–8. doi: 10.1001/jama.2014.13875
Keywords: fecal microbiota transplantation, graft-vs.-host disease, HIV infection, gut epithelial damage, dysbiosis
Citation: Ouyang J, Isnard S, Lin J, Fombuena B, Peng X, Nair Parvathy S, Chen Y, Silverman MS and Routy J-P (2020) Treating From the Inside Out: Relevance of Fecal Microbiota Transplantation to Counteract Gut Damage in GVHD and HIV Infection. Front. Med. 7:421. doi: 10.3389/fmed.2020.00421
Received: 15 April 2020; Accepted: 30 June 2020;
Published: 06 August 2020.
Edited by:
Levinus Albert Dieleman, University of Alberta, CanadaReviewed by:
Rosica S. Valcheva, University of Alberta, CanadaAngel Lanas, University of Zaragoza, Spain
Copyright © 2020 Ouyang, Isnard, Lin, Fombuena, Peng, Nair Parvathy, Chen, Silverman and Routy. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jean-Pierre Routy, amVhbi1waWVycmUucm91dHlAbWNnaWxsLmNh
†These authors have contributed equally to this work