 Johanna Schleifenbaum
Johanna Schleifenbaum
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Med. , 11 June 2019
Sec. Nephrology
Volume 6 - 2019 | https://doi.org/10.3389/fmed.2019.00107
This article is part of the Research Topic Renal Hypertension at the Crossroads: Theoretical, Experimental and Clinical Aspects View all 7 articles
Only a few years ago, alamandine was found to be a member of the protective arm of the renin-angiotensin system. It turned out to be an endogenous ligand of the G protein-coupled receptor MrgD. So far, MrgD had predominantly been studied in a neuronal context. The expression of the receptor in non-neuronal tissue showed hitherto unknown effects mediated by MrgD, most strikingly alamandine-induced vasodilation. Alamandine being a part of the non-classical renin-angiotensin system, a protective role of receptor activation seemed natural. This review summarizes the different effects of MrgD activation by alamandine in vasculature, in the central nervous system, and in organs as kidney and heart. Alamandine and MrgD are promising novel drug targets to protect the kidney and heart through anti-hypertensive actions.
When it comes to discussing G protein-coupled receptors (GPCRs) involvement in the development of hypertension, the first molecule that comes into everyone's mind is usually the angiotensin II receptor. The combination of the search terms “angiotensin receptor” and “hypertension” presents more than 7,000 publications on Pubmed. Many of those publications discuss the angiotensin II receptor subtype 1 (AT1R) as the “bad guy” promoting the development of hypertension, inflammation, remodeling, and more. AT1R inhibition reduces blood pressure and has anti-inflammatory and anti-proliferative effects. An increasing number of publications emphasize the role of the angiotensin II receptor subtype 2 (AT2R) in the so-called protective arm of the renin-angiotensin system (RAS). Following the development of hypertension research, one will soon come across another receptor belonging to the protective arm of the RAS: the Mas receptor. The search terms “Mas receptor” and “hypertension” still offer about 200 publications in Pubmed. The Mas receptor can be activated by its ligand angiotensin (1-7) [Ang (1-7)]. Derived from angiotensin II (Ang II) by Ang II-converting enzyme 2 (ACE2), it shows vasodilating (thus protective) properties. Ang (1-7) can be decarboxylated to a peptide called alamandine. Interestingly, alamandine was found to be an endogenous ligand for a GPCR related to Mas, the Mas-related G protein-coupled receptor member D (MrgD), also known as TGR7 (1) or hGPCR45 (2). The structural resemblance of the ligands Ang (1-7) and alamandine as well as of the receptors Mas and MrgD (3) suggests a possible role of MrgD in blood pressure regulation and hypertension, respectively (Figure 1). Very surprisingly, the search term “MrgD” provides only 42 publications in total on Pubmed. This can be explained by the fact that the investigation of the MrgD receptor in non-neuronal tissue started only a few years ago.
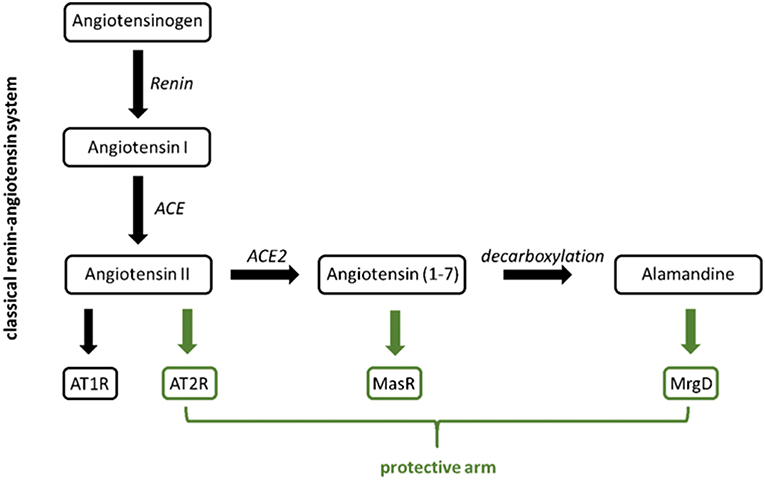
Figure 1. Classical (black) vs. protective arm (green) of the renin-angiotensin system.
The responsible gene Mrgprd is found in rodents as well as in humans (4), the coding sequence is located within a single exon (5). First publications described the receptor expression as constrained to a subset of pain-sensitive small-diameter neurons (1, 4–6). The MrgD receptor was associated with a role in the modulation of neuropathic pain (4, 5) and the perception of itching (7). Coexpression with MrgE, another member of the Mrg family, was found in macaque peripheral nociceptive neurons (8); both subtypes were shown to form heteromeres in HEK cells (9). Expression of several Mrg receptor subtypes was increased in mouse inflamed intestinal tissue (10, 11).
In a cardiovascular context, expression was detected in arterial smooth muscle cells, endothelial nitric oxide synthase (eNOS)-positive endothelial cells, and in atherosclerotic plaques (12). Oliveira et al. localized the receptor in blood vessels, cardiomyocytes (mainly in the membrane, perinuclear, and nuclear region), and the cardiovascular center of the mouse brain. They also studied MrgD-deficient mice and found left ventricular remodeling and a pronounced dilated cardiomyopathy, decreasing the systolic function of the mice (13). If treated with angiotensin II, hearts of spontaneously hypertensive rats (SHR) and cardiomyocytes showed an increased expression of MrgD. The MrgD ligand alamandine was able to attenuate hypertension and alleviate cardiac hypertrophy in this model (14).
The MrgD receptor is G protein-coupled. Most studies were performed in heterologous expression systems, interestingly suggesting a possible coupling to different heterotrimeric G protein subtypes (Table 1). Shinohara et al. were the first ones to show that ß-alanine, a neurotransmitter and a ligand of the MrgD receptor, initiates calcium influx into MrgD-expressing Chinese hamster ovary (CHO) cells (indicating Gq protein coupling), but also reduces forskolin-induced cAMP production (sensitive to pertussis toxin, thus indicating Gi protein coupling) (6). The ß-alanine-induced receptor activation also increased intracellular calcium concentration and stimulated ERK1/2 phosphorylation in Human embryonic kidney 293 (HEK293) cells (9). In cells with coexpression of MrgD and MrgE, ß-alanine-induced ERK1/2 phosphorylation was increased, while MrgD internalization was reduced followed by a prolonged calcium influx (9). Coexpression of MrgD and voltage-sensitive KCNQ2/3 potassium channels in HEK293 cells resulted in a strong inhibition of KCNQ2/3 currents (neuronal “M current”) upon ß-alanine-induced receptor activation. This effect was blocked completely by phospholipase C (PLC) inhibition (again indicating Gq protein coupling) and partially by pertussis toxin (indicating Gi protein coupling). Interestingly, results were partially confirmed when experiments have been performed in isolated DRG neurons. Here, KCNQ2/3 current activation was partially inhibited by PLC blockade and blocked completely by pertussis toxin (15).
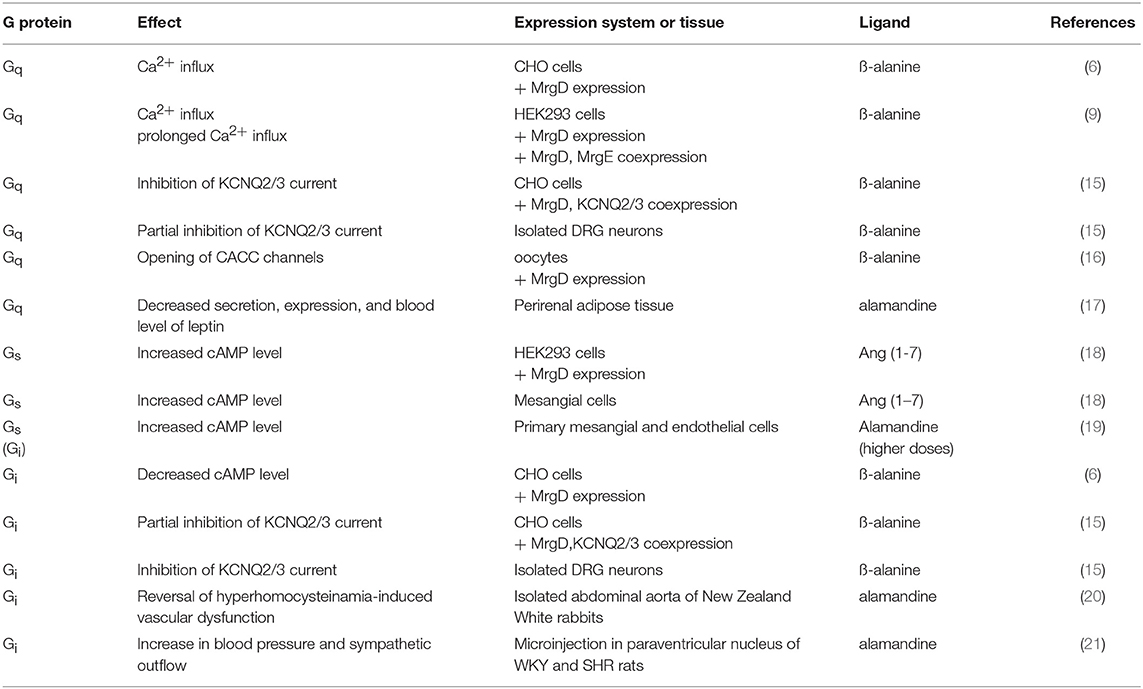
Table 1. Overview of MrgD receptor G protein-coupling under different conditions.
While opening of KCNQ channels hyperpolarizes the cell membrane, opening calcium-activated chloride channels leads to a depolarization. KCNQ channels can be inhibited by PIP2 depletion, thus by activation of Gq protein-coupled receptors. An increase in intracellular calcium, also resulting from Gq protein activation, opens calcium-activated chloride channels. Thus, it was not very surprising that in MrgD-expressing oocytes, a ß-alanine-induced current was found to be sensitive to calcium-activated chloride channel inhibitors and calcium chelators. The chloride current was reduced by phospholipase C (PLC) inhibition and by inositol 1,4,5-trisphosphate (IP3) receptor antisense nucleotides, while phosphokinase C (PKC) inhibition had no effect (16). This indicates a signal transduction from MrgD to CaCC via calcium as second messenger, while the transduction pathway diacylglycerol (DAG)/PKC is neglectable under those conditions.
Thus, ß-alanine is able to signal via MrgD receptor activation inducing Gq or Gi protein-coupled pathways. It remains unclear, which transduction is used under physiological conditions, though both pathways are involved in the regulation of neuronal excitability. This suggests a limitation of ß-alanine as a ligand of MrgD receptors expressed in neurons.
Other ligands of the MrgD receptor are more relevant in the cardiovascular context. Ang (1-7), known as a ligand for the Mas receptor, is also binding to the MrgD receptor and stimulating the Gs protein-coupled signaling pathway. Moreover, it increased cyclic adenosine monophosphate (cAMP) concentrations in MrgD-transfected HEK293 cells and in mesangial cells, but not in MrgD-deficient mesangial cells (18). Similar effects were found for alamandine, an endogenous MrgD ligand circulating in the human blood. It elevated cAMP concentration in primary endothelial and mesangial cells (18), also suggesting Gs coupling. Interestingly, higher concentrations of alamandine were also involved in the Gi-coupled pathway in primary mesangial and endothelial cells, showing specific pharmacodynamics properties of alamandine compared to Ang (1-7) (19). Reversal of hyperhomocysteinamia-induced vascular dysfunction by alamandine in isolated abdominal aorta of New Zealand White rabbits was dependent on Gi protein signaling (20). In blood vessels of New Zealand White rabbits on an atherogenic diet and control animals, alamandine enhanced acetylcholine-mediated vasodilation in thoracic aorta, and iliac artery, while vasodilation of renal arteries by acetylcholine was reduced. In total, the vasoreactivity of alamandine was reduced in atherogenic arteries compared to controls (12). In vivo, participation of the Gi signaling pathway in an alamandine effect could be shown after microinjection of alamandine in the paraventricular nucleus of normotensive Wistar Kyoto (WKY) rats and SHR rats. Blood pressure as well as sympathetic outflow increased in both groups upon activation of MrgD receptor dependent on cAMP and protein kinase A (PKA), while the increase in SHR rats was larger than in the control group (21).
Alamandine decreases secretion, expression, and blood levels of leptin. When leptin levels are low, expenditure is reduced and hunger is increased (22). Genetic deficiency of leptin or its receptor, in both mice and humans, “fools” the brain into thinking that fat stores are absent, resulting in extreme hunger and obesity (22). However, the role of leptin in maintaining energy balance is asymmetric; low levels strongly promote restoration of fat stores, whereas high levels weakly resist obesity. Another feedback signal probably serves this latter function (i.e., to resist obesity), but the identity of the signal (or signals) is unknown (22). Remarkably, leptin plasma levels are elevated under hypertensive conditions (23). This suggests a potentially protective role of the protein. This effect in perirenal adipose tissue of Wistar rats was mediated by Gq protein signaling and also involved c-Src, p38 mitogen-activated protein, and IκB activation (17). Additionally, alamandine induced expression of iNOS and plasminogen activator inhibitor-1 (PAI-1) in adipose tissue and isolated adipocytes (17).
Reduction in leptin expression by alamandine was reversed by the NOS inhibitor L-NG-nitroarginine methyl ester (L-NAME), while leptin expression was inhibited by S-nitroso-L-glutathione, an donor of nitric oxide (NO) (17). The vasodilating molecule NO is produced by the enzymes iNOS, eNOS, and nNOS. Indeed, observed vasoreactive effects of alamandine are mediated by NO release. Leao et al. compared the vasorelaxing effects of alamandine in spontaneously hypertensive stroke prone (SHRSP) rats. They found an enhanced vasorelaxation in SHRSP rats mediated by release of NO and prostaglandins1. At least in aortic rings, the vasodilating effects of alamandine are sensitive to inhibition of endothelial NO release by L-NAME. Also, they can be blocked by ß-alanine, probably through competitive inhibition (24).
The literature about vasodilating effects of alamandine mediated through MrgD receptor activation is very rare; some studies are available only as abstracts. Local vasodilation, particularly in arterial beds with high peripheral resistance, could be a powerful tool in fighting hypertension. Thus, further studies are needed to investigate the involvement of the MrgD receptor and its ligand alamandine. Both molecules, i.e., MrgD and alamandine, could be of interest for the development of new therapies against cardiovascular diseases. The fact that the new RAS hormone alamandine can be easily administered orally as a HPβCD inclusion compound and produced antihypertensive effects in spontaneously hypertensive rats and pronounced protective effects in cardiac fibrosis, opens new perspectives for exploring the therapeutic potential of angiotensin-(1–7)–related peptides. The identification of 2 novel components of the RAS, alamandine and its receptor, will be important for improving our understanding of the physiological and pathophysiological role of the this key regulatory system. It is possible that these 2 novel compounds of the RAS play a pathophysiological role in kidney failure since hemodialysis patients seem to exhibit increased serum concentrations of alamandine (as measured by alamandine /Ang II ratios in 5 patients) (24).
Several studies have been published showing an effect of alamandine-induced MrgD activation on blood pressure. In renovascular hypertensive rats, alamandine showed a biphasic hemodynamic effect after infusion: AT1R-mediated signaling by alamandine under normal conditions (normotensive rats) is superimposed by activation of a PD123319-sensitive receptor under pathophysiological conditions (renovascular hypertensive 2K1C rats) (25). PD123319 has been regarded as an specific AT2R inhibitor, but also inhibits MrgD receptors, as found in experiments using knock-out animals (26). This study is the only one hypothesizing so far that alamandine is also signaling via AT1R activation. The authors suggest an agonistic effect of alamandine based on the knowledge of the structural related Ang (1-7) being an AT1R agonist (27). When injected into the caudal ventrolateral medulla, alamandine showed an MrgD-dependent hypotensive effect in normotensive control rats, while the blood pressure curve was U-shaped in 2K1C animals (28). In vivo, oral doses of alamandine lead to a long-lasting antihypertensive effect in SHR rats, as well as to an anti-fibrotic affect in rats treated with isoprotenerol (24). Effects specifically in the heart were seen upon treatment of cultured neonatal rat cardiomyocytes with alamandine. The treatment prevented angiotensin II-induced hypertrophy via MrgD activation involving a 5′ AMP-activated protein kinase (AMPK)/NO pathway (29). In the same cell type, and additionally in mice treated with lipopolysaccharide (LPS), alamandine attenuated sepsis-induced cardiac dysfunction via inhibition of mitogen-activated protein kinase (MAPK) signaling pathways. Alamandine prevented myocardial inflammation, apopotosis, and autophagy induced by lipopolysaccharides (LPS). It also reversed decreases in cardiac ejection fraction and NOS expression, leading to reduced blood pressure (30). As previously mentioned, administration of alamandine attenuated hypertension and alleviated cardiac hypertrophy in angiotensin II-treated SHR hearts (14). In normotensive SD rats, alamandine administration in vivo increased the plasma level of atrial natriuretic peptide (ANP) via MrgD activation. It was also improving postischemic left ventricular pressure and decreasing the infarct size, while decreasing apoptotic protein and increasing antioxidative protein expression (31).
There are only a few studies about the MrgD receptor and its endogenous ligand alamandine, but many of them show a clear relevance of both molecules for the cardiovascular system. Though the molecular details of the signaling pathways are not completely evolved yet, some physiological effects of receptor and ligand are proven.
Most striking is an increased expression of NOS enzymes upon alamandine-induced activation of MrgD, leading to NO-mediated vasodilation. The induced vasodilation is sufficient for a reversal of vascular endothelial dysfunction as well as an alleviation of the venous return to the heart, thus decreasing the preload of the heart. The vasodilation is subsidized by an alamandine-induced increase in ANP plasma levels. The depressor effect of MrgD activation is masking a preceding pressure effect in healthy animals, while the mechanism seems dysfunctional in rat models of hypertension. Also relevant is an anti-fibrotic effect, preventing hypertrophy of cardiomyocytes. The protective impact of alamandine-induced MrgD activation is strengthened by changes in expression of proteins improving the outcome of cardiac infarcts.
Though studies investigating alamandine and/or MrgD receptor are done in different models, the findings are consistent and fit into a bigger picture. All alamandine-induced MrgD receptor actions are affecting the cardiovascular system, so targeting both molecules can be a worthwhile goal for the development of future antihypertensive and cardioprotective drugs.
JS sifted all publications regarding the topic, drafted, and finalized the manuscript.
The author acknowledges support from the German Research Foundation (DFG) and the Open Access Publication Fund of Charité - Universitätsmedizin Berlin.
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The handling Editor declared a shared affiliation, though no other collaboration on this topic or techniques, with the author.
1. Parmentier M, Libert F, Maenhaut C, Lefort A, Gerard C, Perret J, et al. Molecular cloning of the thyrotropin receptor. Science. (1989) 246:1620–2. doi: 10.1126/science.2556796
2. Takeda S, Kadowaki S, Haga T, Takaesu H, Mitaku S. Identification of G protein-coupled receptor genes from the human genome sequence. FEBS Lett. (2002) 520:97–101. doi: 10.1016/S0014-5793(02)02775-8
3. Young D, Waitches G, Birchmeier C, Fasano O, Wigler M. Isolation and characterization of a new cellular oncogene encoding a protein with multiple potential transmembrane domains. Cell. (1986) 45:711–9. doi: 10.1016/0092-8674(86)90785-3
4. Zylka MJ, Dong X, Southwell AL, Anderson DJ. A typical expansion in mice of the sensory neuron-specific Mrg G protein-coupled receptor family. Proc Natl Acad Sci USA. (2003) 100:10043–8. doi: 10.1073/pnas.1732949100
5. Dong X, Han S, Zylka MJ, Simon MI, Anderson DJ. A diverse family of GPCRs expressed in specific subsets of nociceptive sensory neurons. Cell. (2001) 106:619–32. doi: 10.1016/S0092-8674(01)00483-4
6. Shinohara T, Harada M, Ogi K, Maruyama M, Fujii R, Tanaka H, et al. Identification of a G protein-coupled receptor specifically responsive to beta-alanine. J Biol Chem. (2004) 279:23559–64. doi: 10.1074/jbc.M314240200
7. Liu Q, Sikand P, Ma C, Tang Z, Han L, Li Z, et al. Mechanisms of itch evoked by beta-alanine. J Neurosci. (2012) 32:14532–7. doi: 10.1523/JNEUROSCI.3509-12.2012
8. Zhang L, Taylor N, Xie Y, Ford R, Johnson J, Paulsen JE, et al. Cloning and expression of MRG receptors in macaque, mouse, and human. Brain Res. (2005) 133:187–97. doi: 10.1016/j.molbrainres.2004.10.007
9. Milasta S, Pediani J, Appelbe S, Trim S, Wyatt M, Cox P, et al. Interactions between the Mas-related receptors MrgD and MrgE alter signalling and trafficking of MrgD. Mol Pharmacol. (2006) 69:479–91. doi: 10.1124/mol.105.018788
10. Avula LR, Buckinx R, Alpaerts K, Costagliola A, Adriaensen D, Van Nassauw L, et al. The effect of inflammation on the expression and distribution of the MAS-related gene receptors MrgE and MrgF in the murine ileum. Histochem Cell Biol. (2011) 136:569–85. doi: 10.1007/s00418-011-0862-7
11. Avula LR, Buckinx R, Favoreel H, Cox E, Adriaensen D, Van Nassauw L, et al. Expression and distribution patterns of Mas-related gene receptor subtypes A-H in the mouse intestine: inflammation-induced changes. Histochem Cell Biol. (2013) 139:639–58. doi: 10.1007/s00418-013-1086-9
12. Habiyakare B, Alsaadon H, Mathai ML, Hayes A, Zulli A. Reduction of angiotensin A and alamandine vasoactivity in the rabbit model of atherogenesis: differential effects of alamandine and Ang(1-7). Int J Exp Pathol. (2014) 95:290–5. doi: 10.1111/iep.12087
13. Oliveira AC, Melo MB, Motta-Santos D, Peluso AA, Souza-Neto F, da Silva RF, et al. Genetic deletion of the alamandine receptor mrgd leads to dilated cardiomyopathy in mice. Am J Physiol. (2018) 316:H123–33. doi: 10.1152/ajpheart.00075.2018
14. Liu C, Yang CX, Chen XR, Liu BX, Li Y, Wang XZ, et al. Alamandine attenuates hypertension and cardiac hypertrophy in hypertensive rats. Amino Acids. (2018) 50:1071–81. doi: 10.1007/s00726-018-2583-x
15. Crozier RA, Ajit SK, Kaftan EJ, Pausch MH. MrgD activation inhibits KCNQ/M-currents and contributes to enhanced neuronal excitability. J Neurosci. (2007) 27:4492–6. doi: 10.1523/JNEUROSCI.4932-06.2007
16. Zhuo RG, Ma XY, Zhou PL, Liu XY, Zhang K, Wei XL, et al. Mas-related G protein-coupled receptor D is coupled to endogenous calcium-activated chloride channel in Xenopus oocytes. J Physiol Biochem. (2014) 70:185–91. doi: 10.1007/s13105-013-0292-1
17. Uchiyama T, Okajima F, Mogi C, Tobo A, Tomono S, Sato K. Alamandine reduces leptin expression through the c-Src/p38 MAP kinase pathway in adipose tissue. PLoS ONE. (2017) 12:e0178769. doi: 10.1371/journal.pone.0178769
18. Tetzner A, Gebolys K, Meinert C, Klein S, Uhlich A, Trebicka J, et al. G-protein-coupled receptor MrgD is a receptor for angiotensin-(1-7) involving adenylyl cyclase, cAMP, and phosphokinase A. Hypertension. (2016) 68:185–94. doi: 10.1161/HYPERTENSIONAHA.116.07572
19. Tetzner A, Naughton M, Gebolys K, Eichhorst J, Sala E, Villacanas O, et al. Decarboxylation of Ang-(1-7) to Ala(1)-Ang-(1-7) leads to significant changes in pharmacodynamics. Eur J Pharmacol. (2018) 833:116–23. doi: 10.1016/j.ejphar.2018.05.031
20. Qaradakhi T, Matsoukas MT, Hayes A, Rybalka E, Caprnda M, Rimarova K, et al. Alamandine reverses hyperhomocysteinemia-induced vascular dysfunction via PKA-dependent mechanisms. Cardiovasc Ther. (2017) 35:e12306. doi: 10.1111/1755-5922.12306
21. Shen YH, Chen XR, Yang CX, Liu BX, Li P. Alamandine injected into the paraventricular nucleus increases blood pressure and sympathetic activation in spontaneously hypertensive rats. Peptides. (2018) 103:98–102. doi: 10.1016/j.peptides.2018.03.014
22. Lowell BB. New neuroscience of homeostasis and drives for food, water, and salt. N Engl J Med. (2019) 380:459–71. doi: 10.1056/NEJMra1812053
23. Shankar A, Xiao J. Positive relationship between plasma leptin level and hypertension. Hypertension. (2010) 56:623–8. doi: 10.1161/HYPERTENSIONAHA.109.148213
24. Lautner RQ, Villela DC, Fraga-Silva RA, Silva N, Verano-Braga T, Costa-Fraga F, et al. Discovery and characterization of alamandine: a novel component of the renin-angiotensin system. Circ Res. (2013) 112:1104–11. doi: 10.1161/CIRCRESAHA.113.301077
25. Soltani Hekmat A, Javanmardi K, Kouhpayeh A, Baharamali E, Farjam M. Differences in cardiovascular responses to alamandine in two-kidney, one clip hypertensive and normotensive rats. Circ J. (2017) 81:405–12. doi: 10.1253/circj.CJ-16-0958
26. Blankley CJ, Hodges JC, Klutchko SR, Himmelsbach RJ, Chucholowski A, Connolly CJ, et al. Synthesis and structure-activity relationships of a novel series of non-peptide angiotensin II receptor binding inhibitors specific for the AT2 subtype. J Med Chem. (1991) 34:3248–60. doi: 10.1021/jm00115a014
27. Galandrin S, Denis C, Boularan C, Marie J, M'Kadmi C, Pilette C, et al. Cardioprotective angiotensin-(1-7) peptide acts as a natural-biased ligand at the angiotensin II type 1 receptor. Hypertension. (2016) 68:1365–74. doi: 10.1161/HYPERTENSIONAHA.116.08118
28. Soares ER, Barbosa CM, Campagnole-Santos MJ, Santos RAS, Alzamora AC. Hypotensive effect induced by microinjection of Alamandine, a derivative of angiotensin-(1-7), into caudal ventrolateral medulla of 2K1C hypertensive rats. Peptides. (2017) 96:67–75. doi: 10.1016/j.peptides.2017.09.005
29. Jesus ICG, Scalzo S, Alves F, Marques K, Rocha-Resende C, Bader M, et al. Alamandine acts via MrgD to induce AMPK/NO activation against ANG II hypertrophy in cardiomyocytes. Am J Physiol Cell Physiol. (2018) 314:C702–11. doi: 10.1152/ajpcell.00153.2017
30. Li P, Chen XR, Xu F, Liu C, Li C, Liu H, et al. Alamandine attenuates sepsis-associated cardiac dysfunction via inhibiting MAPKs signaling pathways. Life Sci. (2018) 206:106–16. doi: 10.1016/j.lfs.2018.04.010
Keywords: alamandine, MrgD, GPCR, renin-angiotensin system, hypertension
Citation: Schleifenbaum J (2019) Alamandine and Its Receptor MrgD Pair Up to Join the Protective Arm of the Renin-Angiotensin System. Front. Med. 6:107. doi: 10.3389/fmed.2019.00107
Received: 01 February 2019; Accepted: 30 April 2019;
Published: 11 June 2019.
Edited by:
Maik Gollasch, Charité Medical University of Berlin, GermanyReviewed by:
Dmitry Tsvetkov, University of Tübingen, GermanyCopyright © 2019 Schleifenbaum. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Johanna Schleifenbaum, am9oYW5uYS5zY2hsZWlmZW5iYXVtQGNoYXJpdGUuZGU=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.