 Fernando Félix
Fernando Félix Santiago F. Burneo
Santiago F. Burneo- 1Museo de Zoología, Escuela de Ciencias Biológicas, Pontificia Universidad Católica del Ecuador, Quito, Ecuador
- 2Museo de Ballenas, Salinas, Ecuador
A long-term study of a common bottlenose dolphin (Tursiops truncatus) population inhabiting the Gulf of Guayaquil, Ecuador (2°33′ S, 79°20′W), has been carried out for almost 30 years. Similarly, as in other parts of the world, this population is structured socially and spatially in well-defined subunits or communities. Two of these communities, referred to as Posorja and El Morro, have been studied with major intensity in the last 10 years in the western inner estuary, among others to calculate population parameters that allow assessing their viability in time. Calculated parameters include annual abundance, age and sex composition, annual crude birth rate, calf survival, calf production interval, and average annual mortality/emigration. With these parameters and others derived from other better-studied populations, the trend of both subunits was modeled using the software Vortex. Results show that even under an optimistic scenery both communities will be extinct in the short (Posorja) and mid-term (El Morro), if current stressors continue. Most population parameters calculated in both communities show similar values as in populations elsewhere, but a very low calf survival in Posorja and high mortality/emigration ratios in adults, and probably in juveniles in both communities, contribute to this trend. Population deterioration seems to be the result of different human-induced threats such as fisheries, maritime traffic and others still not well assessed, as well as stochastic demographic events. We recommend taking actions in the short term to halt population decline addressing the major causes of mortality affecting these dolphin communities.
Introduction
The conservation status of several species of coastal small cetaceans is of worldwide concern because of the increasing evidence of population declines, in some cases to critical levels, particularly those species of discrete distribution (e.g., Slooten, 2007; Huang et al., 2014; Taylor et al., 2016; Minton et al., 2017). Mortality in fishing gear and collisions with vessels have been identified as the main causes of population decline (Van Waerebeek et al., 2007; Reeves et al., 2013; Food and Agriculture Organization [FAO], 2018), but other less-known causes such as environmental degradation and pollution predispose animals to disease, impaired reproduction or cause a variety of other metabolic disorders (Wells et al., 2005; Jepson et al., 2015; Murphy et al., 2015). In many cases, a population can decline very rapidly due to a low reproduction or calf survival rate, which complicates an early diagnosis (Wells and Scott, 1990; Thompson et al., 2000). The situation worsens when no baseline information is available to recognize changes occurring in the ecosystem and while actions are defined and management measures implemented, the situation may deteriorate until it becomes irreversible. The role of marine mammals in the ecosystem and how changes in the trophic structure could affect them is not known in depth (see Morissette et al., 2012), as changes could be exacerbated by alterations generated by both human activities and large-scale phenomena such as climate change acting synergistically (Wells, 2010; Wild et al., 2019). Thus, addressing the conservation problem of small cetaceans is complex, and requires financial and human resources to improve our understanding of the ecosystem functioning and how wild populations respond to both natural and human-induced pressures.
Estimating population parameters such as abundance, birth rate, mortality, and morbidity among others, are key issues in defining the viability of a population and is the first step to define appropriate management measures (Wells and Scott, 1990; Cantor et al., 2012; Manlik et al., 2016). Life histories of identifiable individuals in long-term studies have been used to estimate population and reproductive parameters of small cetaceans (e.g., Wells and Scott, 1990; Fruet et al., 2015; Martin and Da Silva, 2018). However, highly structured populations of coastal cetaceans living in small units require finer-scale assessments than less structured offshore populations, as it has been demonstrated in the inshore common bottlenose dolphin (Tursiops truncatus) (Sellas et al., 2005; Richards et al., 2013; Fruet et al., 2014), the Indo-Pacific bottlenose dolphin (T. aduncus) (Wiszniewski et al., 2009), coastal killer whales (Orcinus orca) (Parsons et al., 2009) and Guiana dolphins (Sotalia guianensis) (Cantor et al., 2012). Such an approach demands enormous efforts to follow- specific individuals for sufficient time to understand the role of these animals within their population units as their reproductive status and physical maturity changes to properly interpret the fluctuations in the face of environmental changes (Parsons et al., 2009; Wells, 2014).
A resident population of inshore bottlenose dolphins inhabits the inner estuary of the Gulf of Guayaquil in Ecuador. Intermittently studied since the early 1990s, this population has experienced a 50% decrease in abundance over the last three decades (Félix, 1994, 1997; Félix et al., 2017; Bayas-Rea et al., 2018). More detailed analysis of the population subunits that conform this metapopulation shows different biological and conservation conditions, they are likely being unevenly affected by human and non-human stressors (Félix et al., 2018, 2019c), it is not clear why some subunits are more resilient than others. Addressing the causes is key to designing tailored conservation actions (Manlik et al., 2016). For example, artisanal fishermen in the inner estuary of the Gulf of Guayaquil use a high diversity of gear with different intensity at different sites (see Herrera et al., 2013). Shipping and port activities could also have a different impact on those population units inhabiting nearby ports than those animals living in areas where such activities are less intense. The identification of management units in structured populations of small coastal cetaceans such as bottlenose dolphins is key to prevent habitat fractionating and ensuring their conservation (e.g., Sellas et al., 2005; Fruet et al., 2014). Therefore, the conservation of this metapopulation must take into account the variability of threats occurring in a relatively small area where well-differentiated population entities inhabit. While the concept of management units has been traditionally used for demographic and spatially isolated populations (Palsbøll et al., 2006; Sveegaard et al., 2015), in highly social species such as some matrilineal cetaceans their social and even cultural aspects could have an important evolutionary role and explain aspects such as low genetic diversity and habitat specificity (Whitehead, 2017), so they are determining features to define population units.
In this study, we evaluate population parameters of two neighboring population units of bottlenose dolphins inhabiting estuarine waters in the Gulf of Guayaquil, Ecuador, aiming at defining their viability in the short and medium-term. Being neighboring communities with a certain level of interaction, it would be expected that population trajectories are not significantly different. Based on our demographic analysis we propose management measures to be implemented immediately to reduce the impact of anthropogenic disturbances in these dolphin communities before the point of no return is surpassed.
Methodology
The Study Area
With around 12,000 km2, the Gulf of Guayaquil is the largest estuary on the west coast of South America (2°33′ S, 79°20′ W). The inner estuary extends into the continent approximately 100 km. This study focused on the western part of the inner estuary where one of its two main branches, the Estero Salado, is located (Figure 1). The other main branch, the Guayas River, runs some 20 kilometers eastward and parallel to the Estero Salado. The salinity in the estuary is seasonally influenced by the Guayas River runoff, ranging between 20 and 25 pus in the Estero Salado throughout the year. The Estero Salado extends for about 70 km in a northeasterly direction. Secondary channels divert from the main water body and form numerous mangrove channels and islands aside. The inner estuary is subject to a semi-diurnal tide regime ranging from 2.5 to 3.5 m, with currents up to 3 knots (Stevenson, 1981). Two well-differentiated seasons with different rainfall regimes characterize the coast of Ecuador; a short and rainy season between December and April and a longer and drier season between May and November, with relatively short transition periods. The surface temperature of the estuary varies between 22 and 28°C and does not affect the distribution of bottlenose dolphins in the study area but it does the annual variation in salinity during the rainy season in other sites of the inner estuary (Félix, 1994).
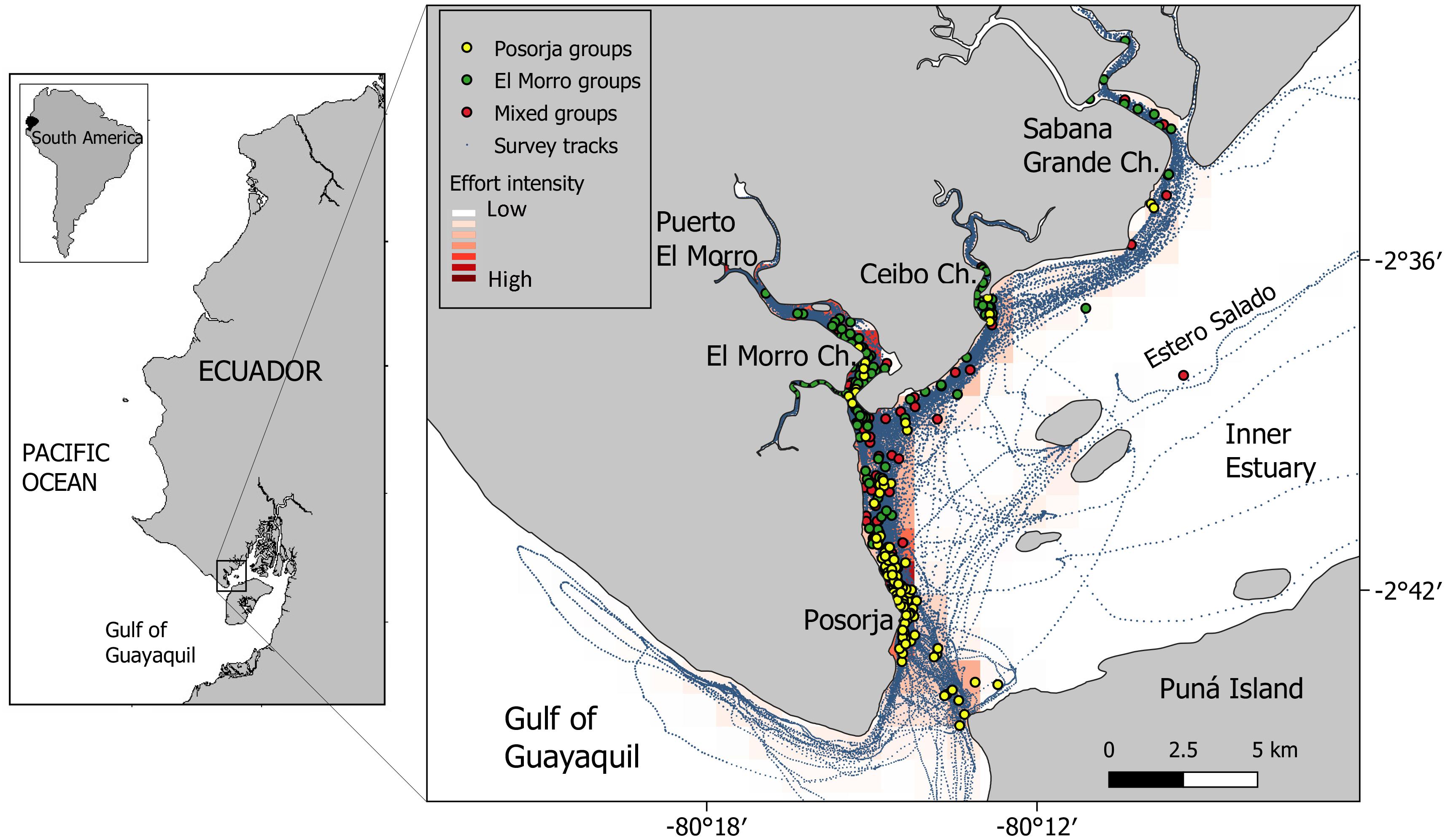
Figure 1. The study area in the western part of the inner estuary of the Gulf of Guayaquil. Blue lines correspond to the survey tracks conducted during the study period. Colored dots correspond to geo-referenced positions where dolphin groups were recorded, and the colors indicate their membership (Posorja, El Morro and mixed groups). The red-colored background layer shows the effort intensity.
The study area includes about 30 km along the east side of the Estero Salado between Posorja and Sabana Grande channels and the northwest area of Puná Island by the south (Figure 1). With 30,000 inhabitants, Posorja is one of the most important industrial fishing ports of Ecuador. A new port facility for post-panamax ships started operating in Posorja in August 2019 increasing maritime activities in the area. Other activities carried out along the Estero Salado are artisanal fishing, shrimp aquaculture, and boat-based tourism. Part of the study area is within the El Morro Mangrove Wildlife Refuge (REVISEMEM), a coastal marine reserve created in 2007 with an extent of 10,230 ha (Ministerio del Ambiente del Ecuador [MAE], 2010).
Population Units
Two socially and geographically well-defined bottlenose dolphin population units inhabit the study area, one in the southern part around the city of Posorja and another one further north along the west side of the Estero Salado (Félix et al., 2017). These population units are called communities (sensu Wells et al., 1987) because their members maintain a higher association degree with each other than with individuals from neighboring communities. Membership of individuals to a specific community was based on the analysis of the level of association among individuals (see Félix et al., 2017). For practical purposes, these two dolphin communities have been referred to as Posorja and El Morro. Mixed groups containing individuals of both communities represented between 13 and 16% of the total groups recorded (Félix, 1997; Félix et al., 2017). Bottlenose dolphins in the Gulf of Guayaquil are genetically distinct from other coastal populations and have low genetic diversity at mtDNA control region (Hd = 0.557 ± 0.086 and π = 0.022 ± 0.019) (Bayas-Rea et al., 2018). Mostly shared but also unique haplotypes were found in these two communities, confirming their genetic relatedness (Bayas-Rea et al., 2018).
Boat-Based Surveys
Boat-based surveys onboard open boats of 6–7 m in length powered by outboard engines (48–150 HP) have been made around the study area since May 2005. Trips were conducted opportunistically onboard tourist boats until 2009. Restarting in 2011, both opportunistic and dedicated research trips increased gradually with a maximum of 58 trips in 2018 (Table 1). Posorja was used as a base port until 2014 and after which Puerto El Morro, located 8 km upstream within El Morro channel. The area was surveyed randomly, beginning the search for dolphins near the departing port and continuing further until finding a group. Except when tourist boats were used, trips aimed to cover the greater distance possible. Survey extended between 20 and 100 km and lasted between 2 and 7 h. There was high variability in terms of area coverage during the surveys, therefore not every area was covered equally. A heatmap layer with the survey tracks was prepared with the software QGIS v3.4 (Qgis Development Team, 2018) using an interpolation algorithm to create a density raster from a vector layer with the kernel density estimate (Figure 1).
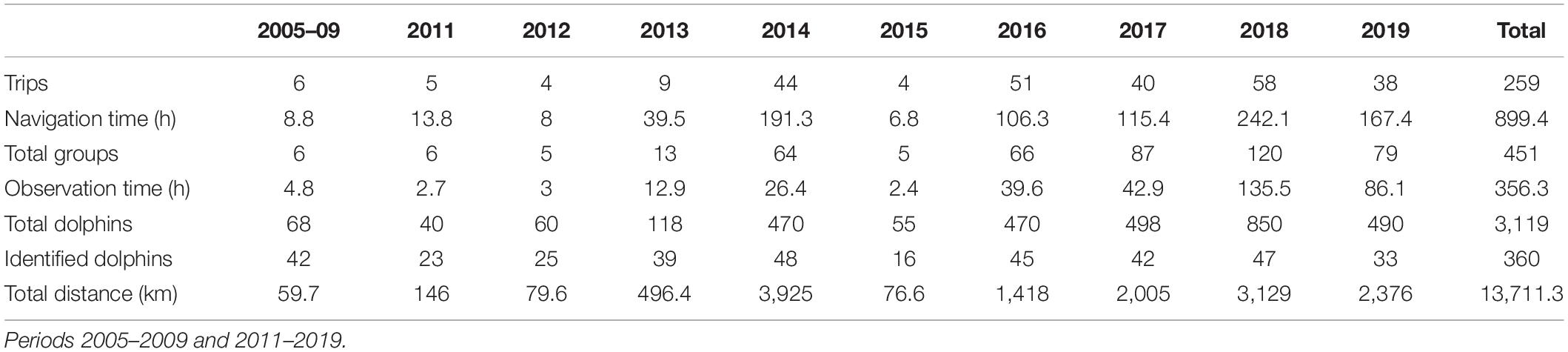
Table 1. The yearly effort made during the study period and the number of groups and dolphins recorded.
Sightings
Once a group of dolphins was found, a cautious approach was made to a safe distance to take information on behavior, size and group composition. Digital cameras (18–24 megapixels) with 70–300 mm and 100–400 mm zoom lenses were used to take photographs of dorsal fins and other characteristic body parts. Both the shape and the notches on the posterior edge of the dorsal fins were used for individual identification, but also temporal scars product of teeth rakes and sharp objects (see previous work Félix, 1997; Félix et al., 2017). More than 150,000 photographs were taken during the study and around 30% were retained for photo-ID analysis. The best quality photographs regarding focus and perpendicular angle were used to create a catalog that was updated regularly as new particular features appeared in the fins. Each individual was assigned an alphanumeric identification code. The photographs were also useful for recording behaviors and additional specific individual characteristics such as skin diseases and scars (Van Bressem et al., 2015; Félix et al., 2018). The sightings were georeferenced using a Garmin® 64 GPS and the route information subsequently used to estimate the sampling effort and build distribution maps using QGIS. Positions were also used to estimate 95% home ranges of both communities using the package adehabitatHR v. 0.4.18 for R (Calengue, 2006). The model uses the ‘utilization distribution’ and considers that animals use of the space can be described by a bivariate probability density function (UD). The percent value corresponds to the smallest areas on which the probability to relocate the animals is equal to 0.95.
Definitions and Population Parameters Evaluated
Group
A group of dolphins refers to all animals that were recorded during the sighting period, usually performing similar activities and moving as a unit with similar speed and same direction (Félix et al., 2017). The number of animals in a group was determined by direct counts of identified animals or as an average of the range estimated in the field when all the animals in the group could not be identified. Size and group composition were adjusted once photographs were processed.
Encounter Rate
The number of dolphins and groups recorded per 10 kilometers of survey.
Occurrence Index
Calculated as the frequency in which identified individuals were seen for the total possible recapture occasions in a given period (Morteo et al., 2012), considering the home range spatial coverage of each community. A value of 1 indicates that an animal was always seen on each sampling occasion. OI described the residence patterns of individuals within the study area.
Site Fidelity
Is the tendency to return to a previously occupied location. Animals were classified as residents and non-resident according to their sighting frequency along the study period based on an annual recording basis. Individuals seen at least once in three different years were considered as resident, otherwise, they were classified as non-residents. This criterion was not applied to calves.
Age and Sex Class Determination
Individuals were assigned to one of three age categories (adults, subadults, or calves) according to their relative size and their life histories (Félix, 1997; Félix et al., 2017). As some individuals were recorded since they were calves or juveniles and changed their class with the time, the age class was assigned on an annual basis. The sex of the animals was determined by different methods: 1) using photographs of the dolphin’s anogenital area; 2) molecular methods (Domínguez, 2019); and 3) using indirect information based on breeding condition and behavior: adult females were closely and regularly associated with a calf at some point in their lives and males never associated with a calf during the study period (at least 5 years) but regularly associated with another adult of similar characteristics, presumably another male (Félix et al., 2019a).
Annual Crude Birth Rate
The annual average rate of newborn animals as a proportion of the total number of animals.
Calf Survival
The proportion of offspring that survived the first 12 months of life.
Calving Period
The period a calf remained with its mother until weaning. It was considered that a calf was weaned from the first time the mother and her calf were not part of the same group and verified in subsequent sightings. The calving period was accounted since the 1st day a newborn calf was recorded with its mother, so the estimated nursing period is the minimum.
Calf Production Interval
The period between the birth of two offspring of the same mother.
Average Annual Mortality/Emigration
The annual average number of resident adults found dead or considered emigrated as a ratio of the total number of animals. A resident adult animal that ceased to be observed for three consecutive years was considered as dead/emigrated. Because it was not possible to know if animals that disappeared for more than 3 years from the study area died or emigrated, both parameters were calculated together. Conversely, adult survival was derived by subtracting the animals that die or emigrated from the remaining animals.
Abundance and Apparent Survival
Given the small size of the dolphin communities studied and the fact that all resident animals have been identified by photographs, including small calves, annual population estimates were calculated using direct counts. In the case of calves, individuals were size either due to small nicks on the trailing edge of the fin, fin shape, permanent pigmentations, or due to temporal marks on the surface produced by teeth or sharp objects. If a previously identified animal was not seen in a given year but seen in subsequent years, it was considered alive in the year it was not recorded.
Annual abundance estimates were also obtained with the Lincoln-Petersen estimator for closed populations modified by Chapman (Seber, 2002). Closed population models assume that the population is closed demographically and geographically (i.e., N is constant), that all animals have the same probability to be caught in the first sample, that the second sample is a simple random sample, that marks does not affect catchability, that marks are permanent and all marks are reported (Seber, 2002). We used two consecutive years for samples 1 and 2, starting in 2011 for Posorja and 2013 for El Morro. The last sampling year was 2019. Individual capture histories obtained during the 12 months of a year were collapsed to have an annual sample with all individuals sighted in that year. Calves were excluded from the dataset to reduce bias related to population closeness.
The Cormack-Jolly-Seber (CJS) formulation implemented in Program MARK (White and Burnham, 1999) was used to explore additional open population models to complement estimates on apparent survival. CJS is based on a conditional product multinominal likelihood where the first capture of the animals is conditioned and only recaptures are modeled (Pledger et al., 2003). The model estimate capture probabilities (p) and apparent survival probabilities (phi). Datasets for Posorja and El Morro were assessed separately organizing the dataset collapsing the capture histories by 6-months periods. In the dataset, individuals were marked as females and males. Finally, the minimum real mortality was also included in Mark indicating which individuals die and when using the life story format LDLD (live recoveries L and dead animals D between sampling periods). Model selection was based on the lowest AICc (Akaike Information Criterion) among 16 pre-defined models in each case. The goodness of fit test was accomplished on the Posorja and the El Morro datasets to estimate the magnitude of overdispersion of the data or variance inflation factor ĉ (c-hat). The difference in fit (deviance) between the saturated model and other candidate models is asymptotically X2 (Chi-square) distributed. Calculated ĉ was used to adjust the fit of candidate models in both datasets.
Population Trend
Population viability analyses were conducted with the software VORTEX (version 10.3.7.0), well suited for low fecundity groups such as dolphins (Lacy and Pollak, 2019). VORTEX simulates the effects of deterministic forces as well as demographic, environmental and genetic stochastic events on wildlife populations. VORTEX models demographic stochasticity by determining the occurrence of probabilistic events such as reproduction, litter size, sex determination, and death with a pseudo-random number generator. Predictions should be considered about what would be most likely happens to the populations if various hypotheses about the status of the populations and threats are true.
Most demographic parameters used in the modeling analysis were estimated during this study and were complemented with information from other better studied coastal populations of T. truncatus (Table 2). Dolphins have a long-term polygyny reproductive system with sexual maturity reached between 5 – 13 years for females and 9 – 14 years for males, having produced offspring until 48 yrs for females and 40 years for males (Wells, 2003; Wells and Scott, 2009). Life span varied by sex, up to 48 years for males and 57 years for females (Wells and Scott, 2009; Wells et al., 2014). One offspring is produced in each reproductive event, which is dependent on its mother for about 3 – 6 years (Wells and Scott, 2009), the calving production period for this study was used to infer the percentage of reproductive adult females.
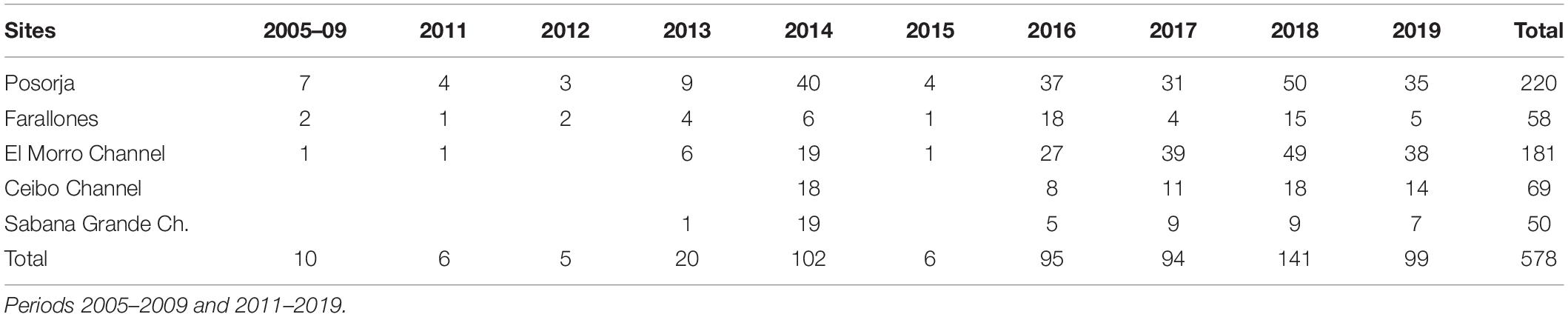
Table 2. The number of times sites and channels surveyed within the study area.
The models were run with 100 iterations and 150 years into the future; extinction was defined as the moment when only one sex remains in the populations. Models were run both with Posorja and El Morro as discrete populations and with them as a metapopulation. For each case, two models were run, optimistic and pessimistic, based on high and low carrying capacity estimates: a high value of 115 individuals and the lowest value of 56 individuals according to the estimates of the dolphins’ communities’ average size in the Gulf of Guayaquil in the early 1990s (Félix, 1997).
Results
Effort
During the study period, 259 trips were conducted totaling 13,711.3 km (Table 1). Navigation time was 899.4 h, of which 356.3 h were of direct observation on the animals. A total of 451 groups and 3,119 dolphins were recorded, most of them resighted animals. The number of different animals identified per year ranged between 16 and 48. Although trips were carried out throughout the year, the effort was concentrated in the period June-October (Figure 2). December and January were the months with the least effort. Dolphins were recorded every month demonstrating year-round residence in both dolphin communities. No significant difference in the group encounter rate throughout the year was found (X211 = 0.9, p > 0.05), although dolphins were significantly more abundant in February and March (X211 = 61.6, p < 0.01), indicating that groups were larger in this part of the year.
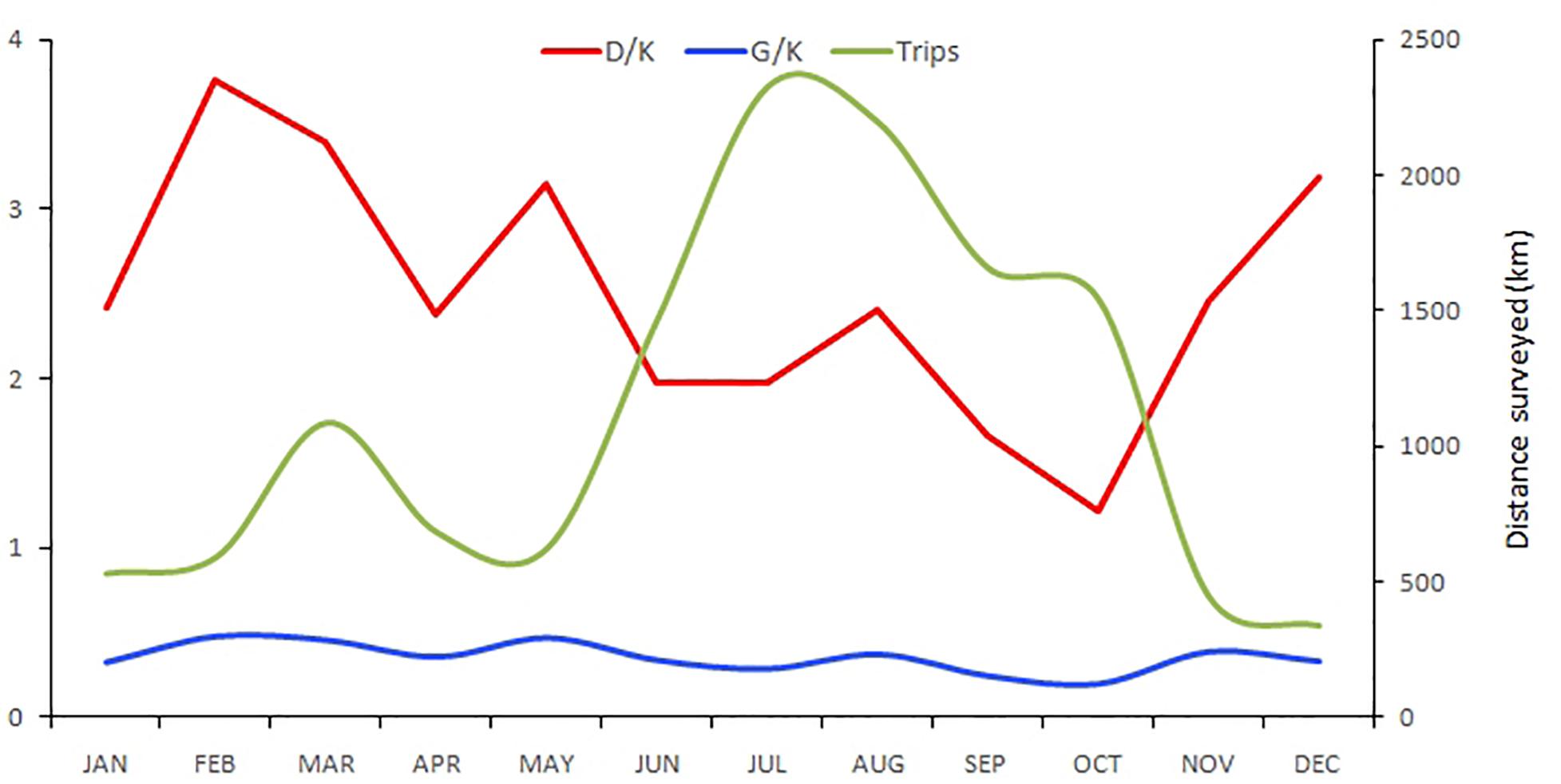
Figure 2. Cumulative monthly effort. Number of trips (green line), group encounter rate per 10 km of survey (G/K) (blue line) and dolphin encounter rate per 10 km of survey (D/K) (red line).
The effort was not uniformly distributed along the study area but concentrated within the core distribution sites of both dolphin communities (Posorja and El Morro Channel) (Table 2). Surveys extended beyond the core areas of both dolphin communities along the Estero Salado, northwestern Puná Island, and west Posorja, where a group density was considerably lower (see Figure 1).
Distribution
Groups containing only Posorja community individuals were recorded clumped around the city (Figure 1). The 95% home range of the Posorja dolphin community covers 24.5 km2, extending 8 km along the continent border and around 3 km on the northeast coast of Puná Island, where small rocky islets called Farallones are located (Figure 1). Some scattered groups were also seen along 20 km of the Estero Salado as far north up to Sabana Grande Channel mouth. A few groups were also seen within the el Morro channel. Most groups observed within El Morro were pairs of males that joined El Morro females with calves. No adult female from Posorja was seen within El Morro Channel. Posorja dolphins were generally found within 500 m off the city coastline where maritime activities concentrate.
The 95% home range of the El Morro dolphin community extends 105 km2 along 17 km of the western side of the Estero Salado and several channels branching from it. Most groups were recorded within the El Morro channel and to a lower extent within Ceibo and Sabana Grande channels, using the Estero Salado mostly as a transit corridor (Figure 1). The core area of El Morro community consists of 100–200 m wide channels fringed by mangrove trees. Some groups were also recorded south near Posorja.
Although core areas were well defined, home ranges of both communities overlap to some extent. There were 116 groups (25.7% of the total recorded) in which at least one animal belonging to another community was present. Most mixed groups contained one or two individuals of the other community, generally adult or subadult males, were found along the entire study area. The largest mixed groups were located between Posorja and El Morro Channel where groups of both communities join and socialize up to several hours before splitting and return to their respective core areas.
Group Size
Average group size in Posorja community was significantly larger than in El Morro, 5.35 dolphins/group (SD = 3.6, n = 172, range 1–20) and 4.1 dolphins/group (SD = 2.82, n = 139, range 1–12), respectively (t = −2.96, P = 0.003) Modal size in both communities was 2 (Figure 3). Group size average increased to 13.9 dolphins/group (SD = 6.1, n = 116, range 3–36) in mixed groups. Overall group size was 7.2 dolphins/group (SD = 5.88, n = 427).
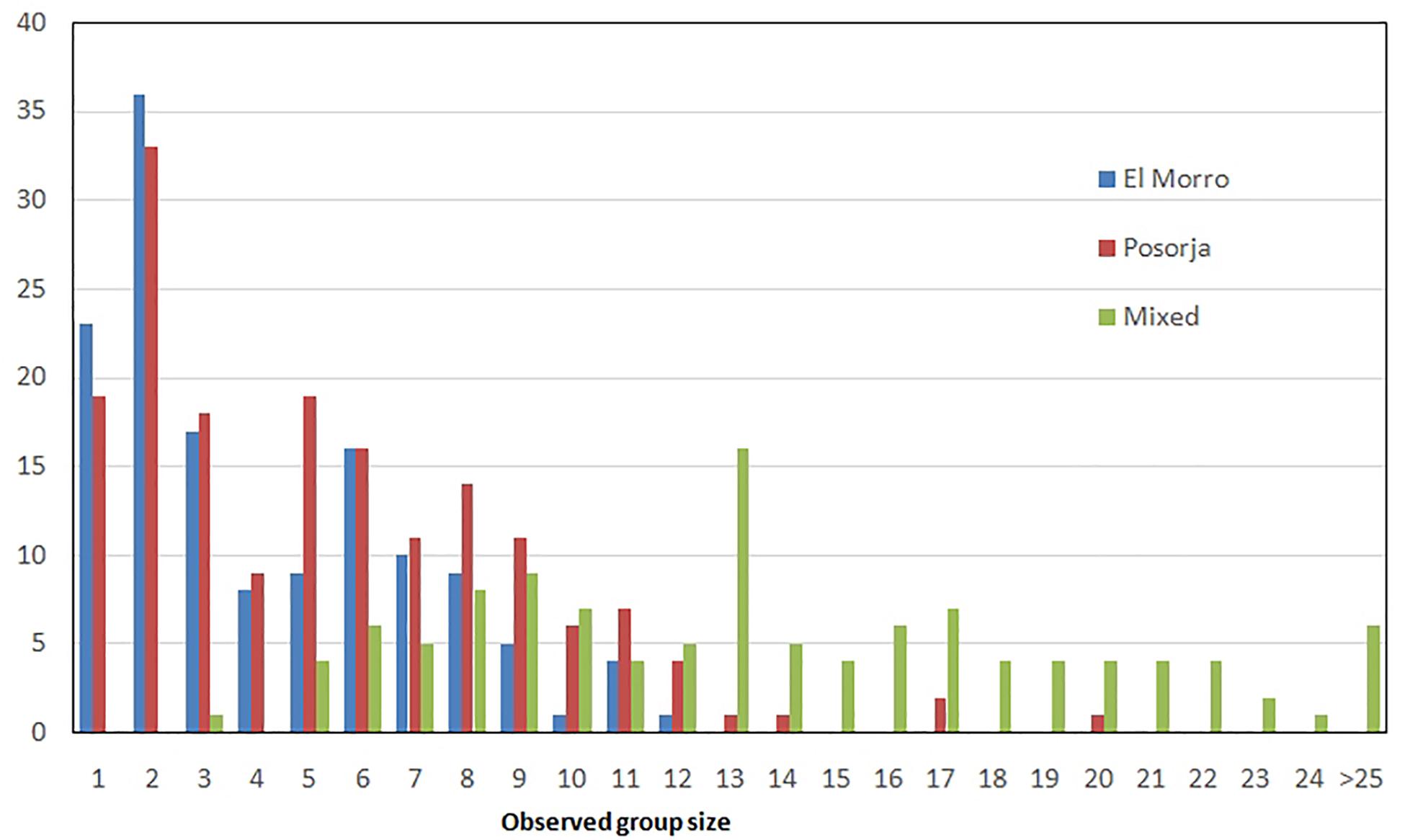
Figure 3. Group size frequency distribution in each dolphin community and mixed groups.
Abundance and Apparent Survival
Posorja
Twenty-six residents and 46 non-resident animals, not belonging to the El Morro community, were recorded during the study period at Posorja. A clear negative population trend of both resident and non-resident animals was found in this dolphin community (Figure 4 and Table 3). The highest number of resident animals (n = 23) was recorded in the year 2014. Ten of the 21 resident animals recorded in the period 2005–2009 were recorded until 2014 and only four of them until 2019. Abundance estimates in the period 2011–2014 were more consistent ranging between 20 and 23 resident animals, then started a steady decrease until 2019 when only 14 resident animals were recorded. The number of non-resident animals also decreased in this period from six in 2011 to two in 2019. Annual abundance obtained with the Chapman estimator resulted in higher values than direct counts (Table 3). Results show a decreasing trend from 36 animals in 2012 to 23 in 2029, but also a stabilization in the number of dolphins in the last 5 years. The most precise estimates were obtained for years 2017 and 2018 with values between 5 and 9% higher than direct counts. The estimate in 2019, however, was around 30% higher than direct counts.
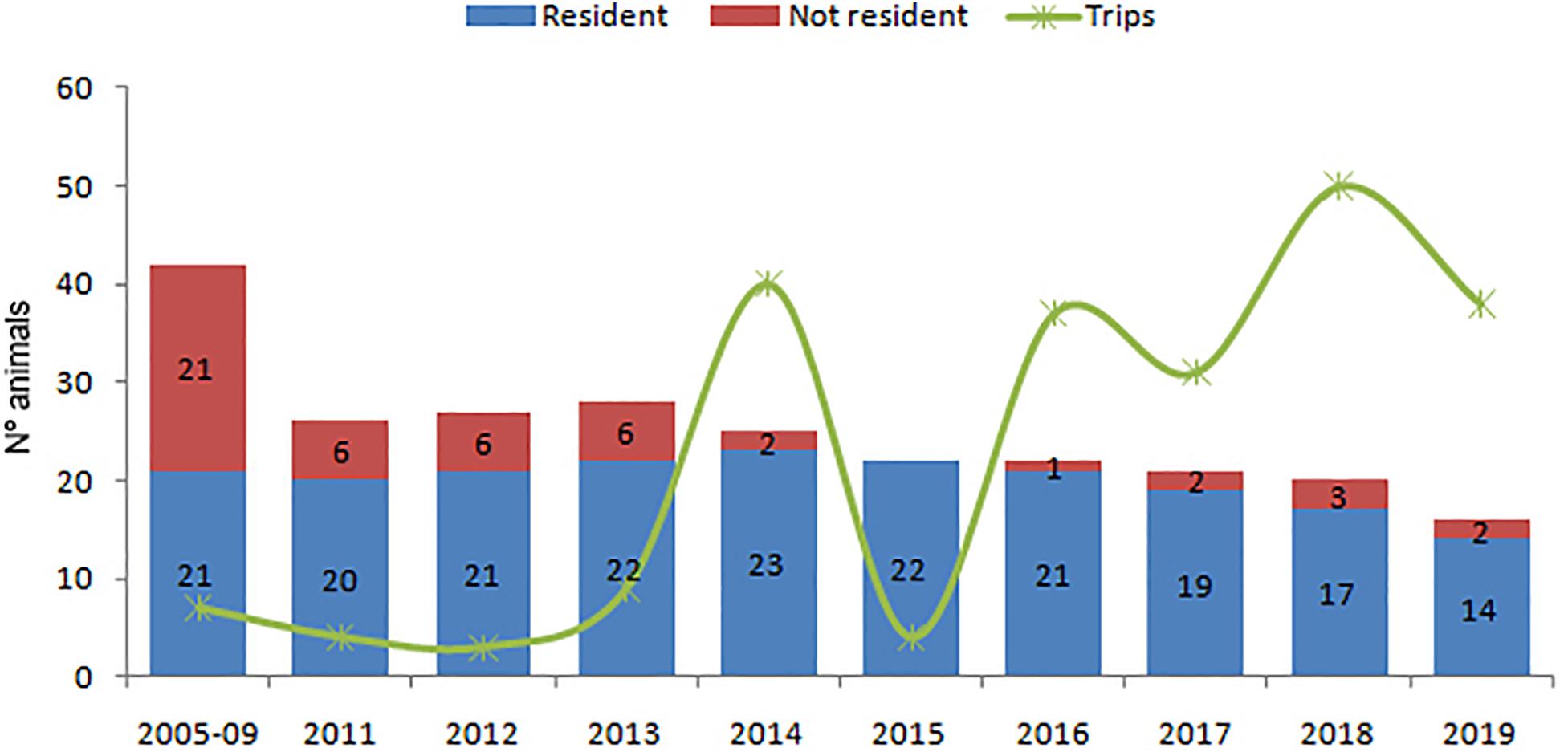
Figure 4. Annual population estimates of the Posorja dolphin community. Resident dolphins in blue and non-residents dolphins in red. The green line indicates the number of trips per year conducted within the distribution area of the Posorja dolphin community. Periods 2005–2009 and 2011–2019.
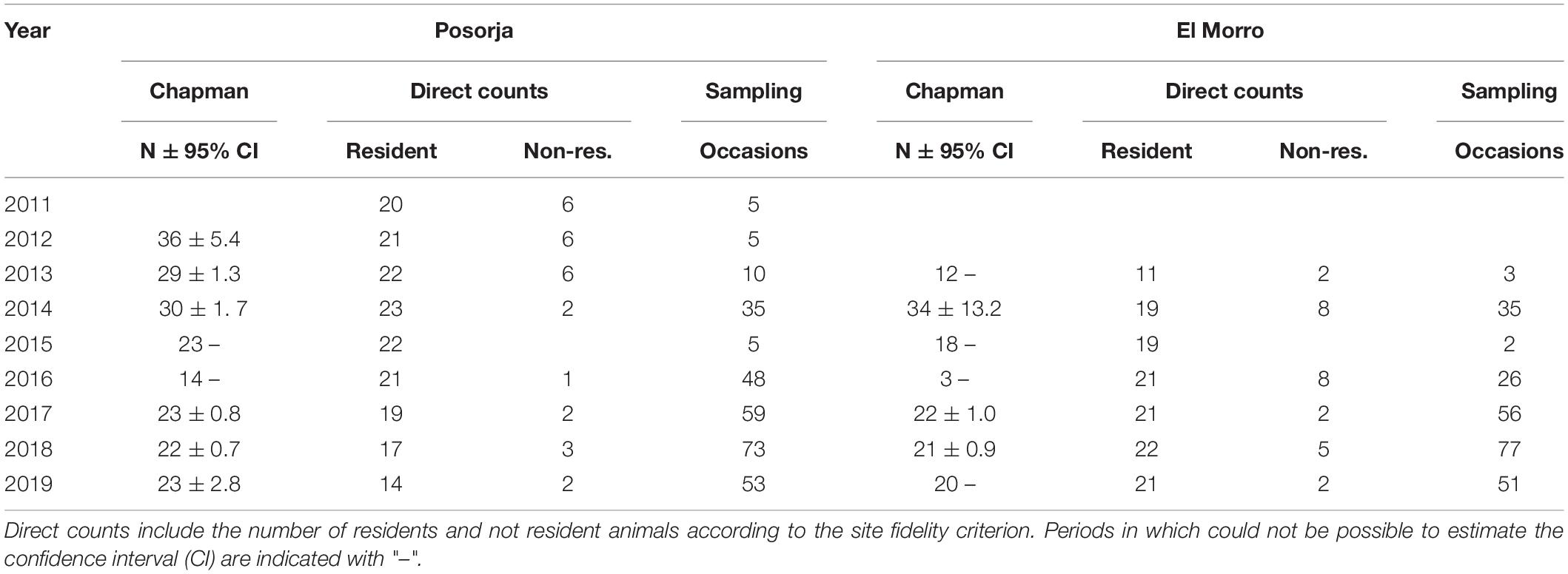
Table 3. Annual population estimates with the Chapman estimator for closed populations and direct counts for each dolphin community.
Regarding survival, two adult dolphins were found dead in the period 2011–2019 (P33F and P72M) which allows calculating the maximum annual survival (0.98%, SD = 0.035) and five other adult animals disappeared for more than 3 years (P4F, P16M, P21M, P27M, and P61M). The average annual survival rate of females in Posorja was 0.93 (SD = 0.13) and in males (0.96, SD = 0.50). Annual average survival in Posorja was 0.95 (SD = 0.06). Apparent survival with the CJS modeling in Mark produced two models that fitted the data with similar probability, constant survival for both sexes and capture probability time dependant {Phi(.) p(t)} and constant survival but sex dependant and capture probability time dependant {Phi(g) p(t)} (Supplementary Table 1). In the first case, survival was 0.94. In the case of the second CJS model, the apparent survival of females was 0.94 and for males 0.97, which is less than 1% higher than obtained with direct counts.
El Morro
Twenty-seven residents and 23 non-resident animals, not belonging to the El Morro community, were recorded in the period 2013–2019 (Figure 5). Contrarily to what occurs in Posorja, the number of resident dolphins in El Morro experienced a slight increase from 19 in 2014 to 21 animals in 2019, mainly due to the production of offspring (Figure 7). Non-resident animals showed a decrease over time from eight in 2014 and 2016 to two in 2019. Annual abundance estimates with the Petersen estimator tended to underestimate the number of animals respect to direct counts (Table 3). The most precise estimates were obtained in the years 2017 and 2018, were estimates were between 5 and 28% lower than direct counts. This model also shows stability in the last 3 years in this community. Subestimation would be caused because calves were not included in the analysis, but they were included in direct counts.
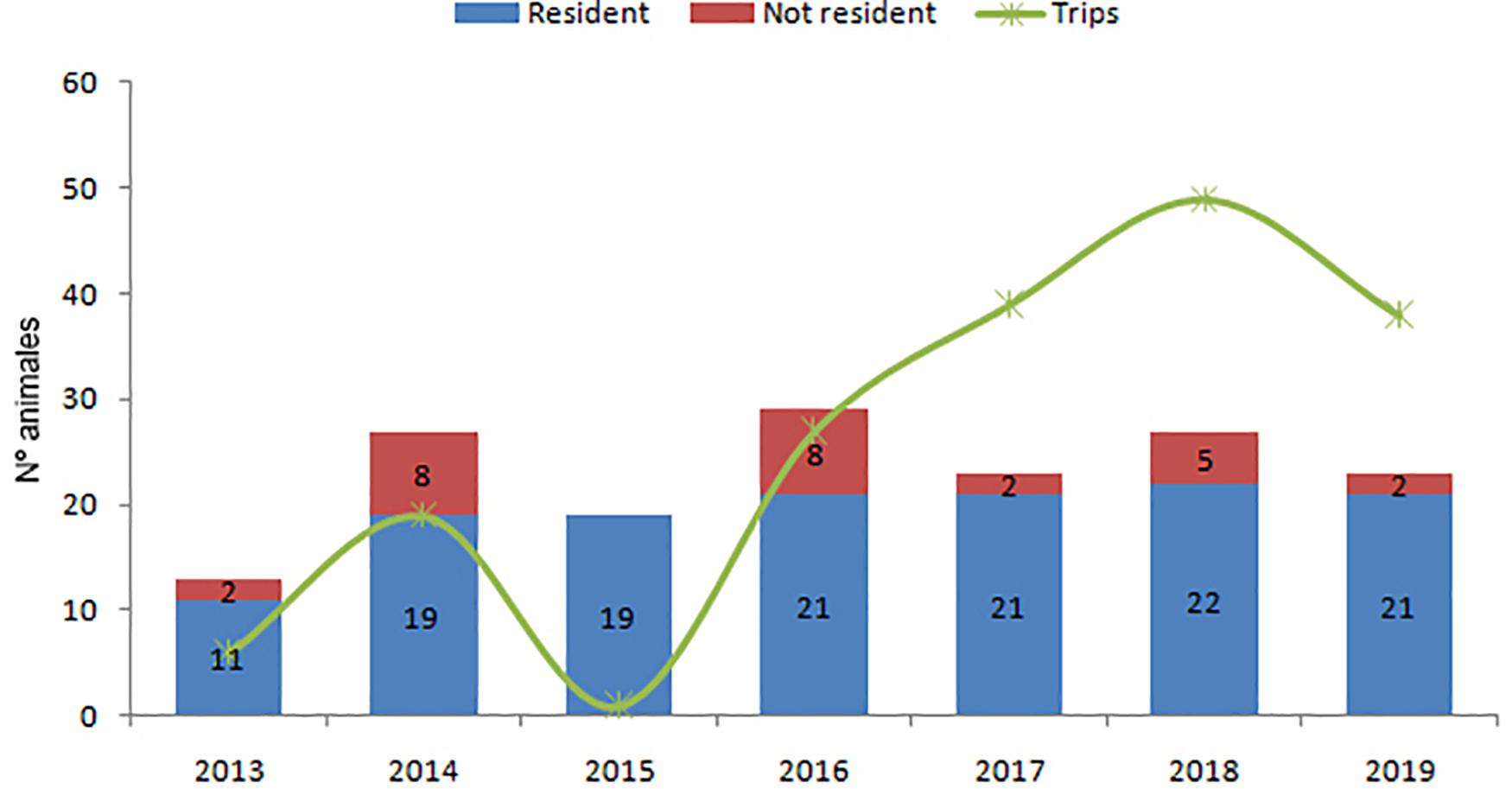
Figure 5. Annual population estimates of the El Morro dolphin community. Resident dolphins in blue and non-resident dolphins in red. The green line indicates the number of trips per year conducted within the distribution area of this dolphin community. Period 2013–2019.
Regarding survival, no resident animal was found dead in El Morro but two animals disappeared for more than 3 years (P85M and P93F). The average annual survival rate in females calculated with direct counts in El Morro was 0.96 (SD = 0.08) and 0.98 (SD = 0.36) in males. Annual average survival in El Morro was 0.98 (SD = 0.53). Apparent survival with the CJS modeling with Mark produced one model that fitted our data, constant survival for both sexes and capture probability time dependant {Phi(.) p(t)} (Supplementary Table 2). The survival estimated for adults in El Morro was 0.96 (SE 0.012), which is 1.9% lower than that obtained with direct counts.
Age and Class Structure
The Posorja community shows a marked bias towards males in the adult population that decreased from 3:1 (15 males and 5 females) in 2011 to 2.3:1 (7 males and 3 females) in 2019 apparently because of the emigration of adult males (Figure 6). Only two calves survived to become subadults in Posorja between 2011 and 2018. Subadults represented on the average 5.5% and calves 4.8% of the total dolphins in Posorja, respectively. In the El Morro community, there was also a bias towards males in the adult population that has maintained constant over time in 2.25:1 in the period 2014–2019 (Figure 7). However, the number of adult males reduced from 12 to nine and adult females from five to four in this period. Contrarily, subadults and calves increased in El Morro three and five times and were observed regularly throughout the study period. On average subadults and calves represented 10.2 and 11.2% of the total animals, respectively.
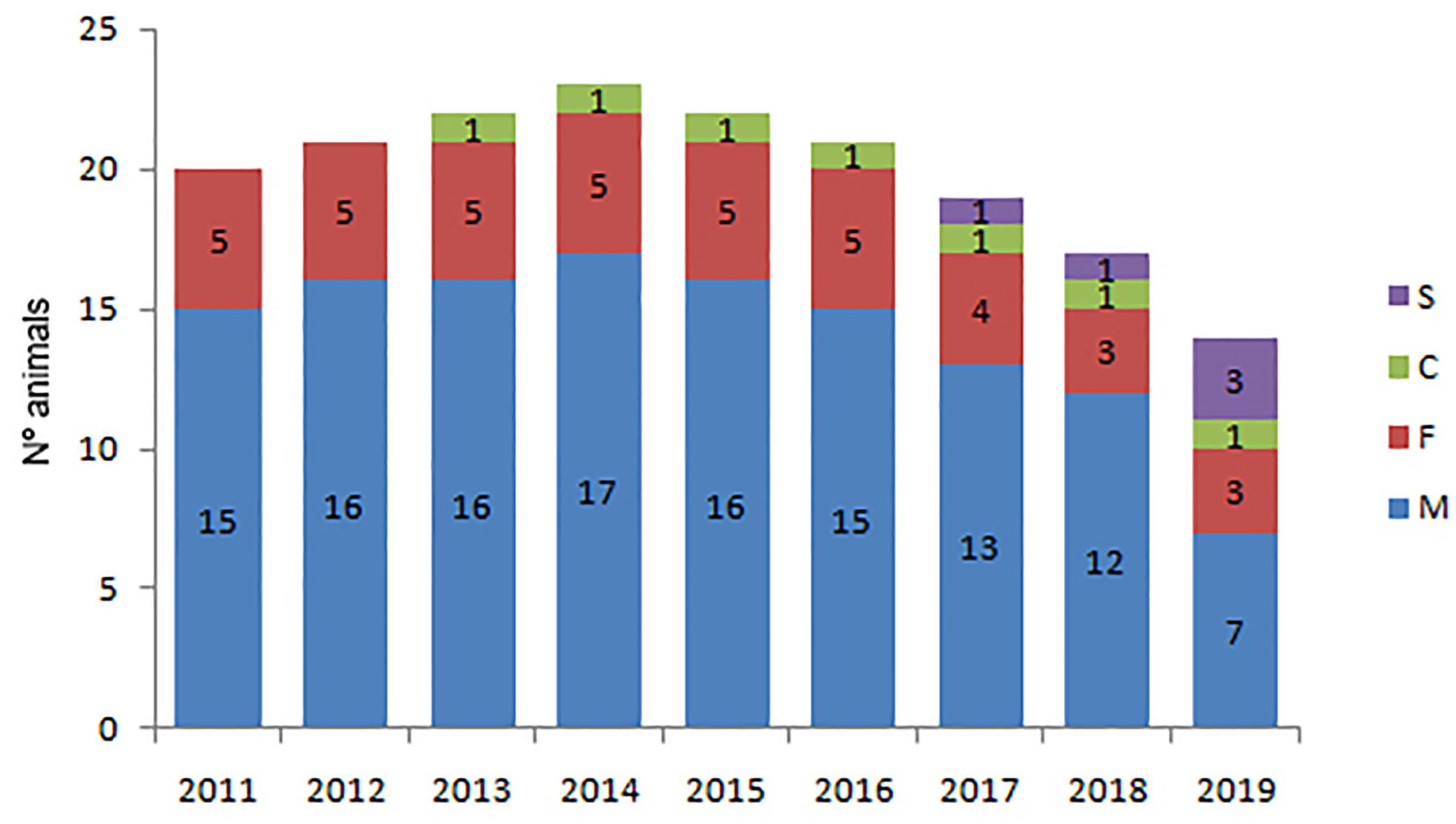
Figure 6. Number of individuals per age and sex class in the Posorja community during the period 2011–2019 M = adult males, F = adult females, S = subadults, C = calves.
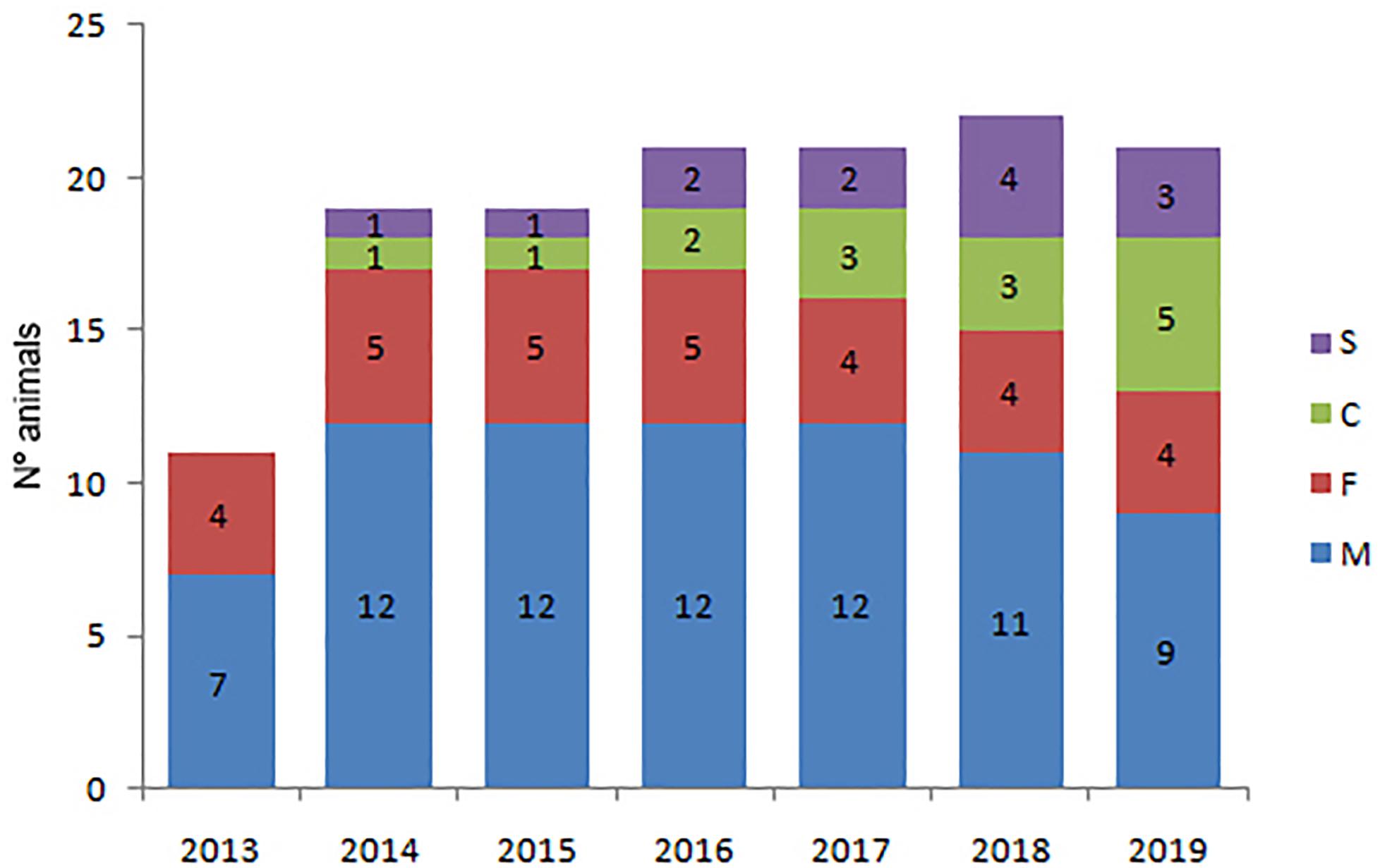
Figure 7. Number of individuals per age and sex class in the El Morro community during the period 2013–2019. M = adult males, F = adult females, S = subadults, C = calves.
Occurrence Index
Adult females showed the highest OI values in both communities (Posorja: P48, P7, and P67, and El Morro: P78, P91, P76, and P77) over time (Figures 8, 9). Although in general terms the pattern of residence by sex was similar in both communities, some differences were found in adult males. In the case of Posorja, there were five adult males with high levels of residence in the last 3 years (P89, P12, P69, P25 and P51), while in Morro there were only two males with similar high level of residence, particularly in the last 3 years (ES117 and ES118).
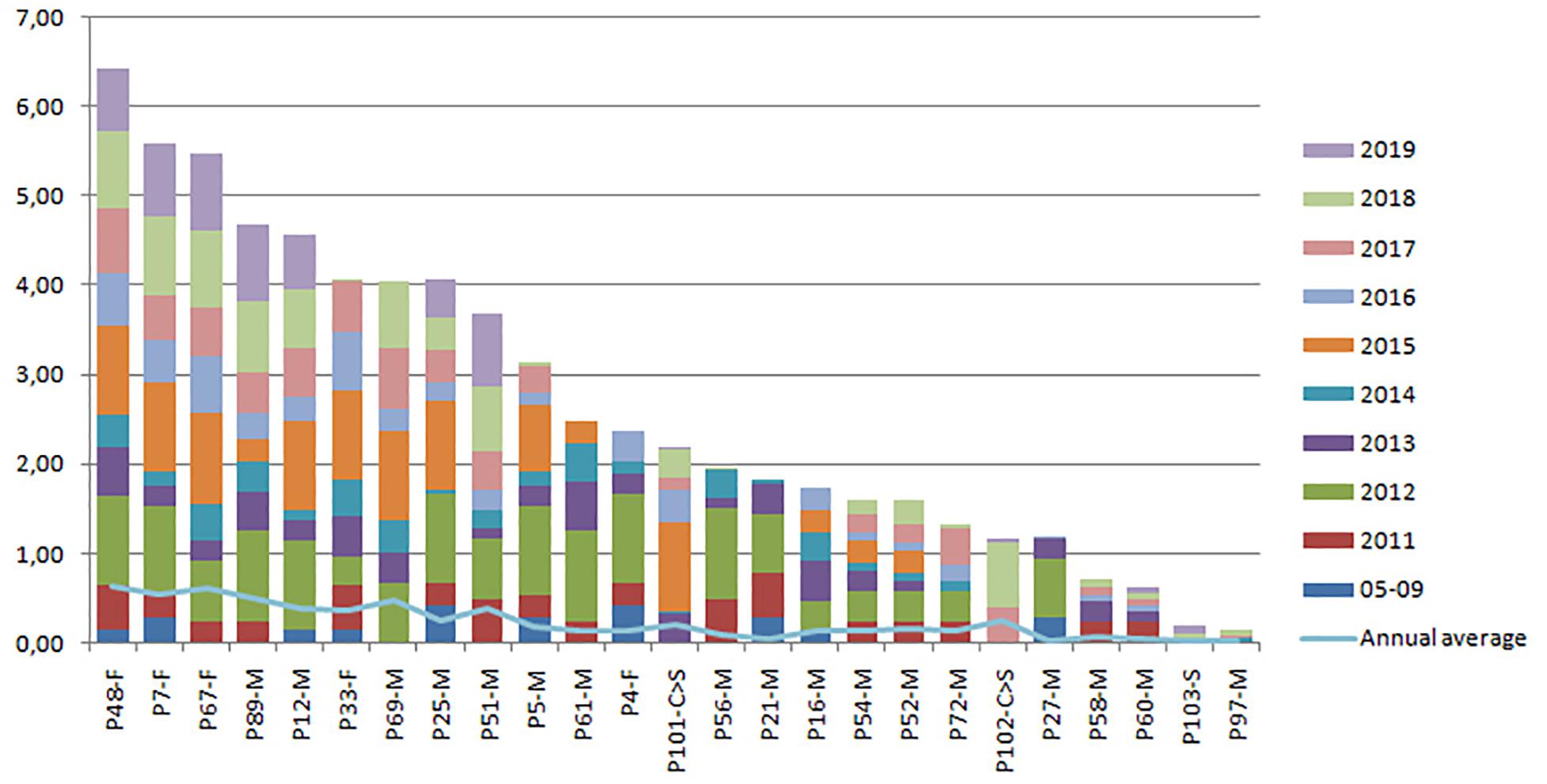
Figure 8. Cumulative annual residency level of resident individuals in the Posorja community during the period 2011–2019. Each color represents 1 year. F = adult female, M = adult male, S = subadult, C = calf. The light blue line indicates the annual average of the residence level of each individual.
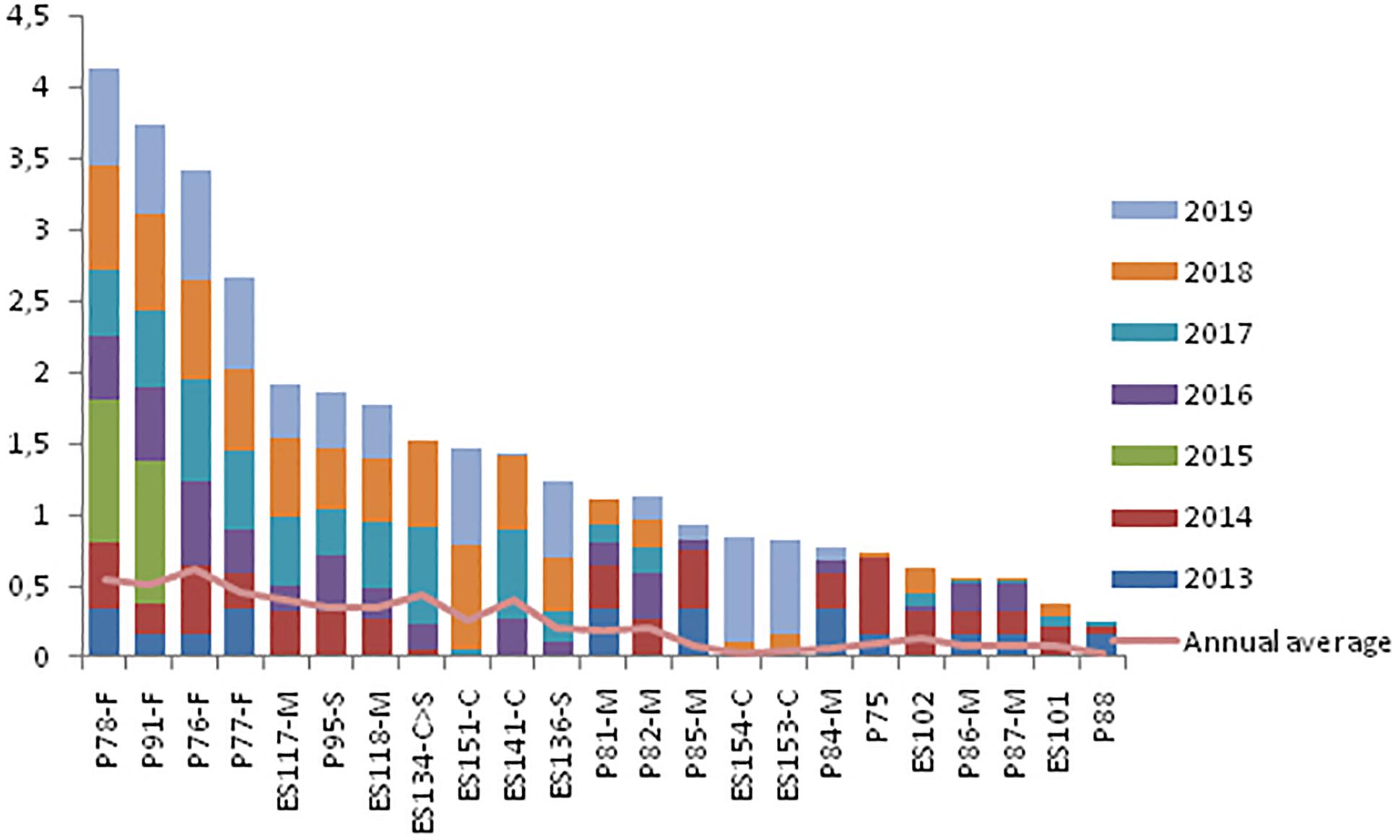
Figure 9. Cumulative annual residency levels of resident individuals in the El Morro community during the period 2013–2019. Each color represents 1 year. F = adult female, M = adult male, S = subadult, C = calf. The light red line indicates the annual average of the level of residence of each individual.
Population Parameters
Crude Birth Rate
In Posorja there were ten births between 2011 and 2019. One female has one calve, three had two calves and one had three. The average annual crude birth rate was 0.055 (SD = 0.08). In El Morro, there were 10 births between 2013 and 2019. Two females had one calf, two have three calves, and one had two. The average annual crude birth rate in the El Morro was 23% higher (0.068, SD = 0.05) (Table 4).
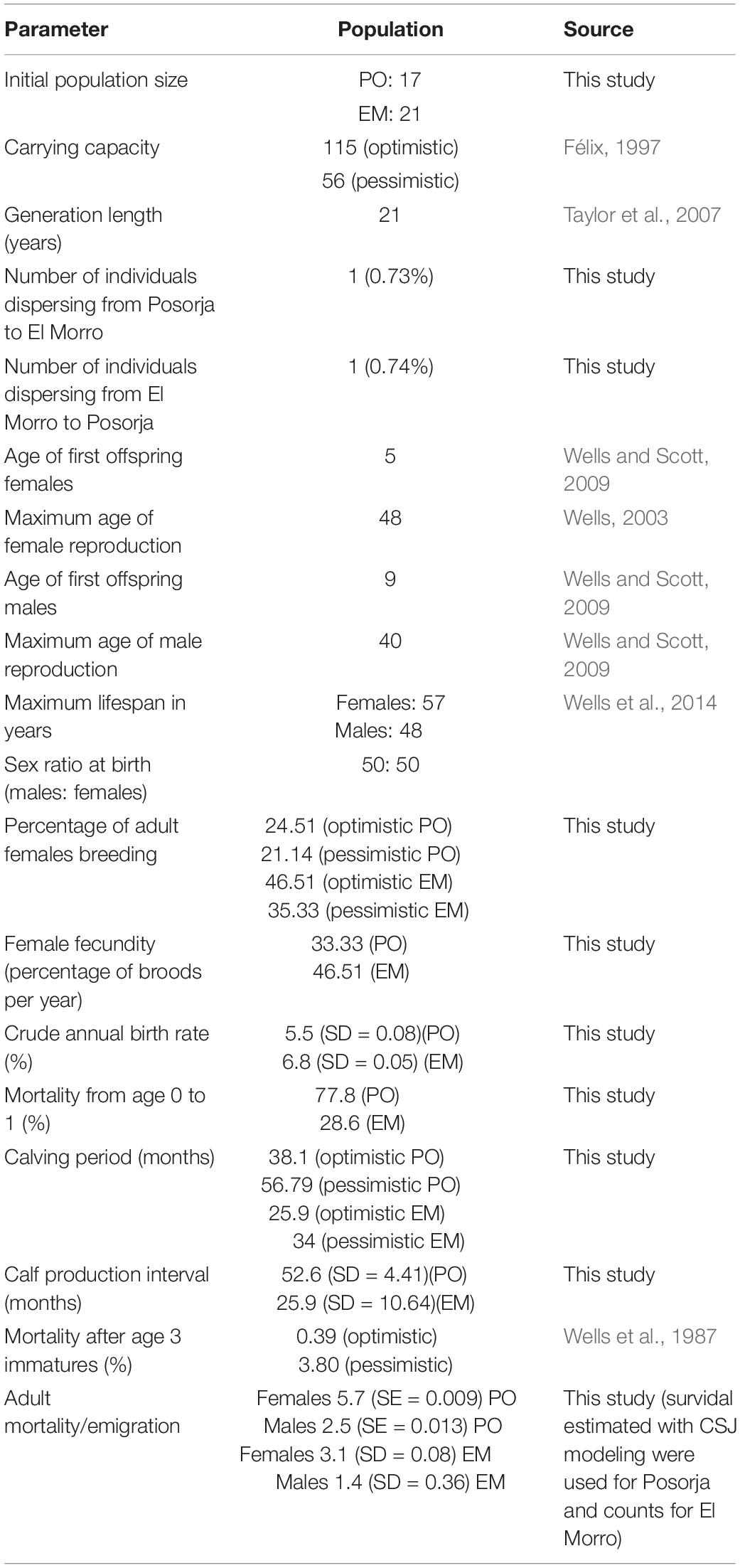
Table 4. Input data for the population viability analysis.
Calf Survival
In Posorja, two of nine offspring (22%) born between 2011 and 2018 survived more than 12 months. Interestingly, the two offspring that survived in Posorja were from the same female (P48). In El Morro, five of seven offspring born between 2013 and 2018 survived more than 12 months (71%) (Table 4). Three of the six were weaned during the study period and three were still with their mothers at the end of the study.
Calving Period
The calving period in Posorja was estimated based on two weaned calves of female P48 because the rest of the calves died a few months after birthing for unknown reasons. These cases showed the most extreme values of the two communities 52.2, the largest calving period, and 24 months the shortest calving period (mean 38.1). The calving period in El Morro was calculated based on three calves that were weaned on average at 32.6 months (Table 4).
Calf Production Interval
The average interval between offspring produced in Posorja was 52.6 months based in three cases and was twice longer than in the Morro (25.9 months) based on five cases. The average value was used in the optimistic model and the largest period reported in each community in the pessimistic model (Table 4). By the end of the study period, only one of three extant females in Posorja remained fertile, while in El Morro the four adult females were nursing calves, one of them (P91) raising simultaneously two calves with 14 months of difference. In El Morro, both females that lost calves gave birth to new calves in 2019. Female P77 lost a calf in May 2018 and the new calf was born 15 months later (August 2019). Female P76 lost her 11-month calf in September 2019 and by the end of December 2019 had another calf, which means this female was pregnant when she lost her previous calf. It is remarkable that two females P91 and P76 were nursing and pregnant simultaneously.
Population Trend
The modeled population trends varied slightly between the optimistic and the pessimistic scenarios for both dolphin communities (Figure 10). In both scenarios, the two communities tend to extinction, Posorja in the short-term and El Morro in the mid-term, if current conditions are not improved. Models predict that the Posorja community size will be reduced by 50% in 10 years and become extinct in a matter of four decades, although functionally will be lost in one generation due to the sex imbalance. In the case of El Morro, the community size will be reduced by 50% in 30 years and become extinct in about 100 years or five generations. If the two communities are modeled as a metapopulation of a single community, they eventually will behave as the El Morro community, which confirms the poor situation of the Posorja community (Figure 10). Additional information with the analysis output is provided as Supplementary material (SM2).
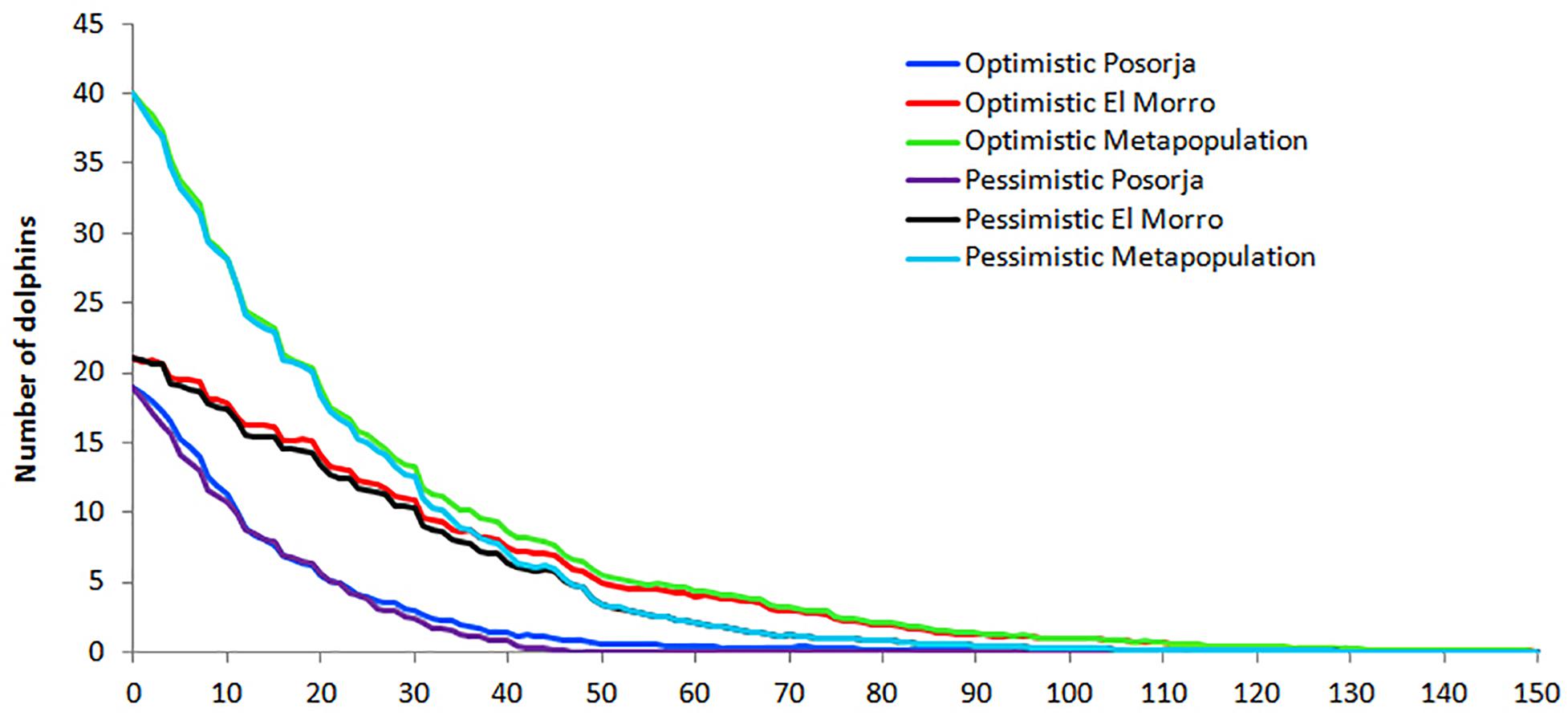
Figure 10. Population trajectories modeled under optimistic and pessimistic sceneries for the Posorja, El Morro and both communities as a metapopulation. Models extended to 150 years into the future, consensus for 100 iterations.
Discussion
The two bottlenose dolphin communities assessed in this study show negative trends that may be irreversible if the causes of such condition are not addressed in the short term, especially for the Posorja community is experiencing a decreasing trend for at least 15 years (Jiménez and Alava, 2014; Félix et al., 2017). As a consequence, the most plausible scenario is the extirpation of both communities, Posorja in the short-term and El Morro in the mid-term. The El Morro dolphin community shows improved population parameters and seems to have a better chance to recover and survive if appropriately managed. Although most population parameters included in the model were estimated in this study, a few of them were derived from other populations that not necessarily may reflect the real condition of bottlenose dolphins from the Gulf of Guayaquil. Therefore, adjustments must be done in future exercises once those parameters can be calculated, in particular, the mortality of immature animals and adult mortality and emigration. It is also important to take into account that no catastrophes were included in the models since no quantifiable data is available. But activities with high potential of impact as the new port facility at Posorja need to be also considered as a potential source of population deterioration.
Two key reproductive parameters illustrate well the differences between the two dolphin communities studied: calf survival and calf interval. In the case of the El Morro community, both parameters are in ranges reported in other bottlenose dolphin populations (e.g., Wells and Scott, 1990; Mann et al., 2000; Wells, 2000; Robinson et al., 2017; Baker et al., 2018), but in Posorja both parameters are 2.27 and 2 times lower than in El Morro, respectively. The high infant mortality in Posorja seems to be due to anthropogenic disturbances including maritime traffic around the port where dolphins concentrate nowadays their activities (Félix et al., 2018). Low calf survival in other bottlenose dolphin populations has also been attributed to anthropogenic sources including tourism boat traffic, fishing gear entanglement and pollution (Wells et al., 2005; Tezanos-Pinto et al., 2015). The second aspect, the calf interval, would be a consequence of aging, with El Morro females being younger and more fertile, which is consistent with reproductive senescence reported in bottlenose dolphins (Wells, 2000; Robinson et al., 2017; Karniski et al., 2018). Reproduction is probably the most important factor for population viability in animals with slow population growth as bottlenose dolphins (Manlik et al., 2016), and this is seriously compromised in the Posorja community. The few reproductive females at Posorja would make the site less attractive to adult males, which would explain why males started a process of emigration from this community since 2014. As a result of this, there is a population contraction that, if not reversed, will lead to the extinction of Posorja community in around two generations (see Figure 10).
Another aspect that characterizes both studied dolphin communities is the sex bias towards males. Stable populations of bottlenose dolphins maintain a sexual parity in the adult population or even with a slight bias towards females (Stanton and Mann, 2012; Wells, 2014; Manlik et al., 2016), as occurs in most mammals (Clutton-Brock et al., 1985). At the other extreme, an estuarine and stable bottlenose dolphin population in Southern Brazil has a bias towards females as high as 2:1 (Fruet et al., 2015). So, what happened to the adult females in these two communities in the inner estuary of the Gulf of Guayaquil? It is not clear whether this is a consequence of a differentiated impact of anthropogenic pressures or is part of a demographic process of selective emigration. A higher mortality rate of males in gillnets has been suggested by Fruet et al. (2015) to explain the bias towards females in Brazil. Such a trend has also been noticed in Ecuador with twice more males entangled than females (F. Félix, unpublished data). Thus, fisheries would not seem to be responsible for the male bias found in our case. On the other hand, selective emigration is consistent with the abandonment of two reproductive females in 2016, one from Posorja (ID N° P4) and another from El Morro (ID N° P93). These two animals represented a great loss for these small dolphin communities because they comprised 20% of the reproductive females. The emigration of individuals to other communities is an uncommon event in bottlenose dolphins and would occur mainly with males (Wells and Scott, 1990). Adult females in the inner estuary of the Gulf of Guayaquil have higher residence level and smaller home ranges than males (Félix et al., 2019a), which has also been observed in other bottlenose populations elsewhere (Wells et al., 1987; Möller and Beheregaray, 2004; Sprogis et al., 2015). However, higher dispersal rate by females has been reported in bottlenose dolphins in Scotland (Natoli et al., 2005) and in the Indo-Pacific bottlenose dolphin from Western Australia (Manlik et al., 2019), apparently driven by habitat quality, hence selective emigration cannot be ruled out as a potential explanation to what occurred with these two dolphin communities in our study area.
The way each dolphin community has responded to natural and anthropogenic disturbances could explain current demographic trends. Although both communities have similar anthropogenic disturbances such as urban development, fisheries, boat tourism, pollution and aquaculture because of their closeness (Jiménez and Alava, 2014; Félix et al., 2017, 2018), the intensity of some pressures over each dolphin community is different. In the case of Posorja, the main pressure is the intense maritime activity around the port, while in El Morro the main stressor is fisheries. The information obtained opportunistically in the first decade of the 21st century allows somehow to reconstruct the near past of the Posorja dolphin community. A first population crash occurred between 2005 and 2010 as only 24% of the population recorded at that time remained in the period 2011–2019. The most plausible explanations for this sudden decrease are a massive mortality event or population fragmentation. Die-off events caused by toxic algae or pathogens could explain not only the population collapse of Posorja but also in the El Morro and in other bottlenose dolphin communities in the inner estuary which show important decreases in abundance in the last decades (Félix et al., 2017). Harmful algal blooms (HABs) have caused massive mortalities of bottlenose dolphins in Florida (e.g., Flewelling et al., 2005; Fire et al., 2015), as well as outbreaks of morbillivirus around the world (Taubenberger et al., 1996; Van Bressem et al., 2014). Red tides are not rare events in the Gulf of Guayaquil and some of them were confirmed to have been toxic (Borbor-Cordova et al., 2019), so it is not ruled out that a mortality event could have happened at that time. Outbreaks of mass cetacean mortality have not been reported in Ecuador but in the north of Peru in 2012 (200 km south the Gulf of Guayaquil) large-scale mortality involving more than 800 small cetaceans of six different species including bottlenose dolphins were reported (Instituto del Mar del Perú [IMARPE], 2012). However, the precise cause remained unknown. A mass mortality event could explain the sudden population reduction but does not explain the sex imbalance in both dolphin communities.
In the case of population fragmentation, this could occur after some extremely traumatic events able to break the strong social bonds that characterize the bottlenose dolphin society (Wells et al., 1987). Besides, the dolphins that emigrated must have found a new appropriate and productive territory otherwise they had returned to Posorja. The emigration of a complete bottlenose dolphin community was reported in Central Chile, from Chañaral Island to Choros Island, 27 km apart, due to long-term harassment by unregulated boat-based dolphin-watching tourism, leading to the death of at least one calf in the mid-1990s (Sanino and Yáñez, 2001). Dolphin watching is an additional stressor in the study area and although regulated in holidays it could be very intensive. In another case, 41 Indo-Pacific bottlenose dolphins emigrated from Mikura Island as far as 390 km away for undetermined causes (Tsuji et al., 2017). There is no evidence that something traumatic happened in Posorja although temporarily emigrated dolphins were recorded in other sites of the estuary. The development of a gillnet fishery targeting the stone crab (Menippe frontalis) in Farallones, a core area for Posorja bottlenose dolphin in the first decade of the century, could have caused dolphin mortality triggering the community to split up. By 2011, the remaining dolphins began to use more frequently the area around Posorja port where they concentrate activities nowadays. In support of the emigration hypothesis is the fact that in April 2016 female P3, an individual that had not been seen for 11 years was recorded once in the area, demonstrating this dolphin was alive and successfully emigrated. Wells and Scott (1990) also resighted an individual after 8 years and suggested that some animals may shift their core areas for extended periods. Population fractioning would explain the reduction of dolphin numbers in Posorja but does not explain the sex imbalance because females should emigrate with their offspring and adult males as well. Thus, both population size and sexual bias in Posorja and El Morro communities seem to be the result of stochastic demographic events aggravated by very small population size and that are not fully understood, but with implications for the metapopulation dynamics of the Gulf of Guayaquil.
Management Options
The condition of the Posorja dolphin communities, and to a lower extent the El Morro, is highly vulnerable not only because of their small population size but also because anthropogenic disturbances continue compromising their resilience. Understanding what led these communities to this critical level is key to seeking remediation measures and to prevent it to happen also to other bottlenose dolphin communities within the Gulf of Guayaquil. In this context, the management of this population demands urgent actions to reduce major threats already identified, as well as continuous monitoring to ensure measures taken are working (Félix et al., 2017, 2018), particularly regarding improving survival. The baseline has been established, but actions and control demand joined efforts by different local stakeholders including environmental and tourism authorities, and private sectors such as fishing, tourism, and transportation.
In the case of the Posorja, it seems difficult that the community could recover its members lost during the past decades, even if anthropogenic pressures could be stopped completely. Recently this dolphin community has been a population source for neighboring communities. To survive this community requires to reverse this trend and increase the recruitment of young animals from other communities. Thus, the most likely scenario is extinction in the short term. However, there would be a remote possibility of recovering an area used by this community but lost in the period 2000–2010 when the stone crab fishery developed at Farallones. At that time, this was the core zone for this dolphin community, so recovering the area for dolphins could be a potential solution. We propose the environmental authorities to promote the shift from bottom gillnets towards the use of traps in this area as traps are a highly effective method to catch crabs (Slack-Smith, 2001). The use of alternative fishing gear to gillnets, either longline or traps, has been effective in reducing the incidental mortality of small cetaceans such as harbor porpoise (Phocoena phocoena), Franciscana dolphin (Pontoporia blainvillei) and gray seals (Halichoerus grypus), achieving similar catch levels of target species (Ovegård et al., 2011; Food and Agriculture Organization [FAO], 2018).
Unlike Posorja where the area to be intervened is restricted to about 5 km2 around Farallones, fishery management at El Morro implies greater challenges. Not only the area is more extensive, but also the number of fishermen is higher and gear diverse, therefore surveillance and enforcement are more difficult. The substitution of gear seems a less viable option. From the information collected in these years, it is clear that both gillnets and longlines caused dolphin entanglement in this area (Félix et al., 2018, 2019b). Entanglements have been more frequent in El Morro than in Posorja, four vs. one case. The consequences of these entanglements remained in survivors as scars and appendage partial or whole mutilate, with a prevalence recently estimated at 13.2% (Félix et al., 2018). Management measures based on exclusion areas would appear to be more appropriate as well as to eradicate bottom gillnets in the whole area. Detailed information on distribution and habitat use is available to define the areas to be closed for fishing (e.g., Félix et al., 2017, 2018; Paladines, 2019). Specific measures such as banning the use of gillnets at the mouth of the main channels along the Estero Salado and leaving a corridor of at least of 300 m wide along the zones used for transiting between concentration areas at El Morro, Ceibo and Sabana Grande channels have been proposed (Félix and Burneo, 2019).
The existence of a marine protected area (REVISEMEM) in this part of the Gulf of Guayaquil gives the possibility to environment authorities to improve fishing management, as this is a specific provision stated in the new Ecuadorian Organic Environment Law issued in 2017 and regulated in 2019. We acknowledge the difficulties and challenges at taking such drastic measures, likewise, their implementation is complex and requires the commitment and agreements between fishermen and authorities. Surveillance will also demand additional costs that must be assumed by local authorities. But if measures are not taken in the short term, local extirpation of bottlenose dolphin communities from the Gulf of Guayaquil is not a distant scenario.
Data Availability Statement
The datasets generated for this study are available on request to the corresponding author.
Ethics Statement
Research activity for this animal study was granted authorization by the Ecuador Ministry of Environment (authorization numbers: N°004-IC-FAU-DPG/MAE, N°008-IC-FLO/FAU-DPG/MAE, N°006-2018-IC-FLO/FAU-DPAG/MAE, N°018-2019-IC-FLO/FAU-DPAG/MAE).
Author Contributions
FF designed the study, carried out fieldwork, organized databases, and wrote the first draft of the manuscript. SB performed the VPA modeling work. Both authors contributed to manuscript revision and approved the submitted version.
Funding
This research was financed by the Research Grants Program of the Pontifical Catholic University of Ecuador (Pontificia Universidad Católica del Ecuador, PUCE). Financial support was received from different sources along the time, including from Museo de Ballenas of Salinas and the Pontific University of Ecuador (PUCE) through its research grants program.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank several volunteers and researchers that participated during surveys collecting data including Ruby Centeno, Andrea Calderón, Michelle Vintimilla, Rosa Bayas, Anaid Paladines, and Gonzalo Domínguez. The Director of REVISMEM Óscar Vásconez for his unconditional support, as well as Juan Romero and other rangers who participated in almost every trip. Koen Van Waerebeek and the two reviewers made valuable comments to improve this article. Research authorizations were issued by the Ministry of the Environment of Ecuador (N°004-IC-FAU-DPG/MAE, N°008-IC-FLO/FAU-DPG/MAE, N°006-2018-IC-FLO/FAU-DPAG/MAE, N°018- 2019-IC-FLO/FAU-DPAG/MAE).
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2020.537010/full#supplementary-material
References
Baker, I., O’Brien, J., McHugh, K., and Berrow, S. (2018). Female reproductive parameters and population demographics of bottlenose dolphins (Tursiops truncatus) in the Shannon Estuary, Ireland. Mar. Biol. 165:15. doi: 10.1007/s00227-017-3265-z
Bayas-Rea, R., Félix, F., and Montúfar, R. (2018). Genetic divergence and fine scale population structure of the common bottlenose dolphin (Tursiops truncatus, Montagu) found in the Gulf of Guayaquil, Ecuador. PeerJ 6:e4589. doi: 10.7717/peerj.4589
Borbor-Cordova, M. J., Torres, G., Mantilla-Saltos, G., Casierra-Tomala, A., Bermúdez, J. R., Renteria, W., et al. (2019). Oceanography of harmful algal blooms on the ecuadorian coast (1997–2017): integrating remote sensing and biological data. Front. Mar. Sci. 6:13. doi: 10.3389/fmars.2019.00013
Calengue, C. (2006). The package adehabitat for the R software: tool for the analysis of space and habitat use by animals. Ecol. Model. 197:1035.
Cantor, M., Wedekin, L. L., Daura-Jorge, F. G., Rossi-Santos, M., and Simoes-Lopes, P. (2012). Assessing population parameters and trends of Guiana dolphins (Sotalia guianensis): an eight-year mark-recapture study. Mar. Mamm. Sci. 28, 63–83. doi: 10.1111/j.1748-7692.2010.00456.x
Clutton-Brock, T. H., Albon, S. D., and Guinness, F. E. (1985). Parental investment and sex differences in juvenile mortality in birds and mammals. Nature 313, 131–133. doi: 10.1038/313131a0
Domínguez, G. (2019). Estructura social y propuesta metodológica para evaluar el éxito reproductivo, parentesco y endogamia en delfín nariz de botella Tursiops truncatus, Montagu 1821 (Cetacea: Delphinidae). BS dissertation, Pontificia Universidad Católica del Ecuador (PUCE), Quito.
Félix, F. (1994). Ecology of the coastal bottlenose dolphin Tursiops truncatus in the Gulf of Guayaquil, Ecuador. Invest. Cetacea 25, 235–256.
Félix, F. (1997). Organization and social structure of the bottlenose dolphin Tursiops truncatus in the Gulf of Guayaquil, Ecuador. Aquat. Mamm. 23, 1–16.
Félix, F., and Burneo, S. (2019). Uso de Hábitat y Estructura Poblacional del Bufeo Costero (Tursiops truncatus) en el golfo de Guayaquil. Madrid: Ministry of Environment of Ecuador (MAE).
Félix, F., Calderón, A., Vintimilla, M., and Bayas-Rea Rosa, A. (2017). Decreasing population trend in coastal bottlenose dolphin (Tursiops truncatus) from the Gulf of Guayaquil, Ecuador. Aquat. Conserv. 27, 856–866. doi: 10.1002/aqc.2763
Félix, F., Centeno, R., Romero, J., Zavala, M., and Vásconez, Ó (2018). Prevalence of scars of anthropogenic origin in coastal bottlenose dolphin in Ecuador. J. Mar. Biol. Assoc. U.K. 98, 1177–1186. doi: 10.1017/S0025315417000686
Félix, F., Van Bressem, M. F., and Van Waerebeek, K. (2019a). Role of social behaviour in the epidemiology of lobomycosis-like disease in estuarine common bottlenose dolphins from Ecuador. Dis. Aquat. Organ. 134, 75–87. doi: 10.3354/dao03356
Félix, F., Vásconez, O., Centeno, R., and Romero, J. (2019b). Tough life: the case of a young coastal common bottlenose dolphin repeatedly entangled. Lat. Am. J. Aquat. Mamm. 13, 19–14. doi: 10.5597/lajam00243
Félix, F., Zavala, M., and Centeno, R. (2019c). Distribución espacial, estructura social y amenazas de conservación de una pequeña comunidad de delfines nariz de botella, Tursiops truncatus (Odontoceti: Delphinidae) en Ecuador. Rev. Bio. Trop. 67, 1059–1076.
Fire, S. E., Flewelling, L. J., Stolen, M., Durden, W. N., de Wit, M., Spellman, A. C., et al. (2015). Brevetoxin-associated mass mortality event of bottlenose dolphins and manatees along the east coast of Florida, USA. Mar. Ecol. Prog. Ser. 526, 241–251. doi: 10.3354/meps11225
Flewelling, J. L., Baden, D. G., Barros, N. B., Bossart, G. D., Bottein, M. D., Hammond, D. G., et al. (2005). Red Tides and marine mammal mortalities. Nature 435, 755–756. doi: 10.1038/nature435755a
Food, and Agriculture Organization [FAO] (2018). Report of the Expert Workshop on Means and Methods for Reducing Marine Mammal Mortality in Fishing and Aquaculture Operations, Rome, 20-23 March 2018. FAO Fish. Aquacult. Rep. 1231. Rome: FAO.
Fruet, P. F., Daura-Jorge, F., Vermeulen, E., Flores, P. A. C., Simoes-Lopes, P. C., Genoves, R. G., et al. (2014). Remarkably low genetic diversity and strong population structure in common bottlenose dolphins (Tursiops truncatus) from coastal waters of the Southwestern Atlantic Ocean. Conserv. Genet. 15, 879–895.
Fruet, P. F., Daura-Jorge, F. G., Moller, L. M., Genoves, R. G., and Secchi, E. (2015). Abundance and demography of bottlenose dolphins inhabiting a subtropical estuary in the Southwestern Atlantic Ocean. J. Mamm. 96, 332–343. doi: 10.1093/jmammal/gyv035
Herrera, M., Castro, R., Coello, D., Saa, I., and Elıas, E. (2013). Ports, coves and artisanal fishing settlements on mainland Ecuador. Ecuador: Instituto Nacional de Pesca.
Huang, S.-H., Chang, W.-L., and Karczmarski, L. (2014). Population trends and vulnerability of humpback dolphins Sousa chinensis off the west coast of Taiwan. Endanger. Species Res. 26, 147–159. doi: 10.3354/esr00619
Instituto del Mar del Perú [IMARPE] (2012). Mortandad de delfines en el litoral de la costa norte, febrero a abril de 2012. Informe Final. Lima: IMARPE.
Jepson, P. D., Deaville, R., Barber, J., Aguilar, A., Borrell, A., Simon, B., et al. (2015). PCB pollution continues to impact populations of orcas and other dolphins in European waters. Sci. Rep. 6:18573. doi: 10.1038/srep18573
Jiménez, P., and Alava, J. (2014). “Population ecology and anthropogenic stressors of the coastal bottlenose dolphin (Tursiops truncatus) in the El Morro Mangrove and Wildlife Refuge, Guayaquil Gulf, Ecuador: towards conservation and management actions,” in Dolphins: Ecology, behavior and conservation strategies, ed. J. Samuels (New York, NY: Nova Science Publisher), 129–163.
Karniski, C., Krzyszczyk, E., and Mann, J. (2018). Senescence impacts reproduction and maternal investment in bottlenose dolphins. Proc. R. Soc. B 285:20181123. doi: 10.1098/rspb.2018.1123
Lacy, R. C., and Pollak, J. P. (2019). VORTEX: A stochastic simulation of the extinction process. Version 10.3.7.0.
Manlik, O., Chabanne, D., Caniel, C., Bejder, L., Allen, S. J., and Sherwim, W. B. (2019). Demography and genetics suggest reversal of dolphin source-sink dynamics, with implications for conservation. Mar. Mamm. Sci. 35, 732–759. doi: 10.1111/mms.12555
Manlik, O., McDonald, J. A., Mann, J., Raudino, H. C., Bejder, L., Krutzen, M., et al. (2016). The relative importance of reproduction and survival for the conservation of two dolphin populations. Ecol. Evol. 6, 3496–3612. doi: 10.1002/ece3.2130
Mann, J., Connor, R. C., Barre, L. M., and Heithaus, M. R. (2000). Female reproductive success in bottlenose dolphins (Tursiops sp.): life history, habitat provisioning, and group-size effects. Behav. Ecol. 11, 210–219. doi: 10.1093/beheco/11.2.210
Martin, A. R., and Da Silva, V. M. (2018). Reproductive parameters of the Amazon river dolphin or boto, Inia geoffrensis (Cetacea: Iniidae); an evolutionary outlier bucks no trends. Biol. J. Linn. Soc. 123, 666–676. doi: 10.1093/biolinnean/bly005
Ministerio del Ambiente del Ecuador [MAE] (2010). Plan de Manejo del Refugio de Vida Silvestre Manglares El Morro. Playas: Fundación Ecuatoriana para el Estudio de Mamíferos Marinos (FEMM).
Minton, G., Smith, B. D., Braulik, G. T., Kreb, D., Sutaria, D., and Reeves, R. (2017). Orcaella brevirostris. The IUCN Red List of Threatened Species 2017: eT15419A123790805. Gland: IUCN.
Möller, L. M., and Beheregaray, L. B. (2004). Genetic evidence for sex-biased dispersal in resident bottlenose dolphins (Tursiops aduncus). Mol. Ecol. 13, 1607–1612. doi: 10.1111/j.1365-294x.2004.02137.x
Morissette, L., Christensen, V., and Pauly, D. (2012). Marine mammal impacts in exploited ecosystems: would large scale culling benefit fisheries? PLoS One 7:e43966. doi: 10.1371/journal.pone.0043966
Morteo, E., Rocha-Olivares, A., and Morteo, R. (2012). Sensitivity analysis of residency and site fidelity estimations to variations in sampling effort and individual catchability. Rev. Mex. Biodivers. 83, 487–495.
Murphy, S., Barber, J. L., Learmonth, J. A., Read, F. L., Deaville, R., Perkins, M. W., et al. (2015). Reproductive failure in UK harbour porpoises Phocoena phocoena: legacy of pollutant exposure? PLoS One 10:e0131085. doi: 10.1371/journal.pone.0131085
Natoli, A., Birkun, A., Aguilar, A., Lopes, A., and Hoelzel, A. R. (2005). Habitat structure and the dispersal of male and female bottlenose dolphins (Tursiops truncatus). P. Roy. Soc. B Biol. Sci. 272, 1217–1226. doi: 10.1098/rspb.2005.3076
Ovegård, M., Königson, S., Persson, A., and Lunneryd, S. G. (2011). Size selective capture of Atlantic cod (Gadus morhua) in floating pots. Fish. Res. 107, 239–244. doi: 10.1016/j.fishres.2010.10.023
Paladines, A. S. (2019). Evaluación del ámbito hogareño del delfín nariz de botella (Tursiops truncatus) en el Golfo de Guayaquil y potenciales impactos de la construcción del Puerto de Aguas Profundas. BS disertation, Pontificia Universidad Católica del Ecuador PUCE, Quito.
Palsbøll, P. J., Bérubé, M., and Allendorf, F. W. (2006). Identification of management units using population genetic data. Trends Ecol. Evol. 22, 11–16. doi: 10.1016/j.tree.2006.09.003
Parsons, K. M., Balcomb, K. C., Ford, J. K. B., and Durban, J. W. (2009). The social dynamics of southern resident killer whales and conservation implications for this endangered population. Anim. Behav. 77, 963–971. doi: 10.1016/j.anbehav.2009.01.018
Pledger, S., Pollock, K. H., and Norris, J. L. (2003). Open capture-recapture models with heterogeneity: I. Cormack-jolly-seber model. Biometrics 59, 786–794. doi: 10.1111/j.0006-341x.2003.00092.x
Qgis Development Team (2018). QGIS Geographic Information System. Open Source Geospatial Foundation Project. Chicago, IL: Open Source Geospatial Foundation.
Reeves, R. R., McClellan, K., and Werner, T. (2013). Marine mammal bycatch in gillnet and other entangling net fisheries, 1990 to 2011. Endanger. Species Res. 20, 71–97. doi: 10.3354/esr00481
Richards, V. P., McCulloch, S. D., Politz, C., Natoli, A., Driscoll, C. A., Hoelzel, R., et al. (2013). Patterns of population structure for inshore bottlenose dolphins along the eastern United States. J. Hered. 104, 765–778. doi: 10.1093/jhered/est070
Robinson, K. P., Sim, T. M. C., Culloch, R. M., Bean, T. S., Cordoba Aguilar, I., Eisfeld, S. M., et al. (2017). Female reproductive success and calf survival in a North Sea coastal bottlenose dolphin (Tursiops truncatus) population. PLoS One 12:e0185000. doi: 10.1371/journal.pone.0185000
Sanino, G. P., and Yáñez, J. L. (2001). Nueva técnica de video identificación y estimación de tamaño poblacional en cetáceos, aplicada en delfines nariz de botella, Tursiops truncatus, de isla Choros, IV Región de Chile. Bol. Mus. Nac. Hist. Nat. 50, 37–63.
Seber, G. A. F. (2002). The Estimation of Animal Abundance and Related Parameters. Blackburn: The Blackburn Press.
Sellas, A. B., Wells, R. S., and Rosel, P. E. (2005). Mitochondrial and nuclear DNA analyses reveal fine scale geographic structure in bottlenose dolphins (Tursiops truncatus) in the Gulf of Mexico. Conserv. Genet. 6, 715–728. doi: 10.1007/s10592-005-9031-7
Slooten, E. (2007). Conservation management in the face of uncertainty: effectiveness of four options for management Hectors’s dolphin bycatch. Enanger. Species Res. 3, 169–179.
Sprogis, K. R., Raudino, H. C., Rankin, R., MacLeod, C. D., and Bejder, L. (2015). Home range size of adult Indo-Pacific bottlenose dolphins (Tursiops aduncus) in a coastal and estuarine system is habitat and sex-specific. Mar. Mamm. Sci. 32, 287–308. doi: 10.1111/mms.12260
Stanton, M. A., and Mann, J. (2012). Early social networks predict survival in wild bottlenose dolphins. PLoS One 7:e47508. doi: 10.1371/journal.pone.0047508
Stevenson, M. (1981). Seasonal variations in the Gulf of Guayaquil, a tropical estuary. Instituto Nacional de Pesca de Ecuador. Boletín Científico y Técnico 4, 1–131.
Sveegaard, S., Galatius, A., Dietz, R., Kyhn, L., Koblitz, J. C., Amundin, M., et al. (2015). Defining management units for cetaceans by combining genetics, morphology, acoustic and satellite tracking. Glob. Ecol. Conserv. 3, 839–850.
Taubenberger, J. K., Tsai, M., Krafft, A. E., Lichy, J. H., Reid, A. H., Schulman, F. Y., et al. (1996). Two Morbilliviruses implicated in bottlenose dolphin epizootics. Emerg. Infect. Dis. 2, 213–216.
Taylor, B. L., Chivers, S. J., Larese, J., and Perrin, W. F. (2007). Generation Length and Percent Mature Estimates for IUCN Assessments of Cetaceans. Administrative Report LJ-07-01. La Jolla, CA: National Marine Fisheries Service.
Taylor, B. L., Rojas-Bracho, L., Moore, J., Jaramillo-Legorreta, A., Ver Hoef, J. M., Cardenas-Hinojosa, G., et al. (2016). Extinction is imminent for Mexico’s endemic porpoise unless fishery bycatch is eliminated. Conserv. Lett. 10, 588–595. doi: 10.1111/conl.12331
Tezanos-Pinto, G., Constantine, R., Berghan, J., and Baker, C. S. (2015). High calf mortality in bottlenose dolphins in the Bay of Islands, New Zealand—a local unit in decline. Mar. Mamm. Sci. 31, 540–559. doi: 10.1111/mms.12174
Thompson, P. M., Wilson, B., Grellier, K., and Hammond, P. S. (2000). Combining power analysis and population viability analysis to compare traditional and precautionary approaches to conservation of coastal cetaceans. Conserv. Biol. 14, 1253–1263. doi: 10.1046/j.1523-1739.2000.00099-410x
Tsuji, K., Kogi, K., Sakai, M., and Morisaka, T. (2017). Emigration of Indo-Pacific Bottlenose Dolphins (Tursiops aduncus) from Mikura Island, Japan. Aquat. Mamm. 43, 585–593. doi: 10.1578/AM.43.6.2017.585
Van Bressem, M. F., Duignan, P. J., Banyard, A., Barbieri, M., Kathleen, M. C., Andrew, D., et al. (2014). Cetacean morbillivirus: current knowledge and future directions. Viruses 6, 5145–5181. doi: 10.3390/v6125145
Van Bressem, M. F., Simoes-Lopes, P., Félix, F., Kiszka, J., Daura-Jorge, F. G., Avila, I. C., et al. (2015). Epidemiology of lobomycosis-like disease in bottlenose dolphins (Tursiops spp.) from South America and southern Africa. Dis. Aquat. Organ. 117, 59–75. doi: 10.3354/dao02932
Van Waerebeek, K., Baker, A. N., Félix, F., Gedamke, J., Iñiguez, M., Sanino, G. P., et al. (2007). Vessel collisions with small cetaceans worldwide and with large whales in the Southern Hemisphere, and initial assessment. Lat. Am. J. Aquat. Mamm. 6, 43–69.
Wells, R. S. (2000). “Reproduction in wild bottlenose dolphins: overview of patterns observed during a long-term study,” in Bottlenose Dolphin Reproduction Workshop Report, eds D. Duffield and T. Robeck (Silver Spring, MD: AZA Marine Mammal Taxon Advisory Group), 57–74.
Wells, R. S. (2003). “Dolphin social complexity: lessons from long-term study and life history,” in Animal Social Complexity: Intelligence, Culture, and Individualized Societies, eds F. B. M. Waal and P. L. Tyack (Cambridge: Harvard University Press), 32–56.
Wells, R. S. (2010). Feeling the heat, potential climate change impacts on bottlenose dolphins. Whalewatcher 39, 12–17.
Wells, R. S. (2014). “Social structure and life history of bottlenose dolphins near Sarasota Bay, Florida: insights from four decades and five generations,” in Primates and Cetaceans: Field Research and Conservation of Complex Mammalian Societies, eds J. Yamagiwa and L. Karczmarski (Tokio: Springer), 149–172.
Wells, R. S., and Scott, M. D. (1990). Estimating bottlenose dolphin population parameters from individual identification and capture-release techniques. Rep. Int. Whal. Comm. 12, 407–415.
Wells, R. S., and Scott, M. D. (2009). “Common bottlenose dolphin: Tursiops truncatus,” in Encyclopedia of Marine Mammals, 2nd Edn, eds W. F. Perrin, B. Wursig, and J. G. M. Thewissen (San Diego: Elsevier), 249–255.
Wells, R. S., Scott, M. D., and Irvine, A. B. (1987). “The social structure of free-ranging bottlenose dolphins,” in Current Mammalogy, ed. H. H. Genoways (New York, NY: Plenum Press), 247–305.
Wells, R. S., Smith, C. R., Sweeney, J. C., Townsend, F. I., Fauquier, D. A., and Stone, R. (2014). Fetal survival of common bottlenose dolphins (Tursiops truncatus) in Sarasota Bay, Florida. Aquat. Mamm. 40, 252–259.
Wells, R. S., Tornero, V., Borrell, A., Aguilar, A., Rowles, T. K., Rhinehart, H. L., et al. (2005). Integrating life-history and reproductive success data to examine potential relationships with organochlorine compounds for bottlenose dolphins (Tursiops truncatus) in Sarasota Bay, Florida. Sci. Total Environ. 349, 106–119.
White, G. C., and Burnham, K. P. (1999). Program MARK: survival estimation from populations of marked animals. Bird Study 46(Suppl.), 120–138.
Wild, S., Krutzen, M., Rankin, R. W., Hoppitt, W. J. E., Gerber, L., and Allen, S. J. (2019). Long-term decline in survival and reproduction of dolphins following a marine heatwave. Curr. Biol. 29, R225–R240. doi: 10.1016/j.cub.2019.02.047
Keywords: bottlenose dolphin, Ecuador, management, reproductive parameters, population trend
Citation: Félix F and Burneo SF (2020) Imminent Risk of Extirpation for Two Bottlenose Dolphin Communities in the Gulf of Guayaquil, Ecuador. Front. Mar. Sci. 7:537010. doi: 10.3389/fmars.2020.537010
Received: 21 February 2020; Accepted: 12 August 2020;
Published: 03 September 2020.
Edited by:
Jeremy Kiszka, Florida International University, United StatesReviewed by:
Katherine McHugh, Chicago Zoological Society, United StatesPedro Friedrich Fruet, Federal University of Rio Grande, Brazil
Copyright © 2020 Félix and Burneo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Fernando Félix, ZmVmZWxpeDkwQGhvdG1haWwuY29t