
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Mar. Sci. , 04 April 2019
Sec. Marine Megafauna
Volume 6 - 2019 | https://doi.org/10.3389/fmars.2019.00125
 Julius Nielsen1,2,3,4*
Julius Nielsen1,2,3,4* Jørgen Schou Christiansen4,5
Jørgen Schou Christiansen4,5 Peter Grønkjær6
Peter Grønkjær6 Peter Bushnell7
Peter Bushnell7 John Fleng Steffensen1
John Fleng Steffensen1 Helene Overgaard Kiilerich6
Helene Overgaard Kiilerich6 Kim Præbel8
Kim Præbel8 Rasmus Hedeholm2
Rasmus Hedeholm2Current knowledge on the feeding ecology of the Greenland shark (Somniosus microcephalus), a potential top predator in arctic marine ecosystems, is based on small sample sizes as well as narrow size ranges of sharks. Therefore, potential size-related feeding patterns remain poorly documented. Using stomach content data (N = 88) and stable isotope values of white muscle tissue (N = 40), this study evaluates the diet of sharks ranging in size from 81 to 474 cm (total length). The importance of prey categories (“Fish,” “Mammal,” “Squid,” “Crustacean,” and “Other”) was evaluated based on the reconstructed prey biomass of the stomach contents. Stable isotope values of δ13C and δ15N ranged between -14.4 to -19.9‰ and 11.8 to 17.2‰, respectively. The importance of each prey category was estimated by the Index of Relative Importance (IRI). Our findings suggest that the smallest Greenland sharks (<200 cm) feed on lower trophic level prey, predominantly squids. Larger sharks (>200 cm) mainly feed on higher trophic level prey such as seals, epibenthic and benthic fishes including gadoids (Gadidae), skates (Rajidae), righteye flounders (Pleuronectidae), lumpfish (Cyclopteridae), wolffish (Anarhichadidae), and redfish (Sebastidae). Redfish were, however, only found to be important in the largest sharks sampled (>400 cm). In addition to demonstrating ontogenetic shifts in their feeding preferences, this study supports that Greenland sharks are capable of active predation on fast swimming seals and large fishes.
During the past 10 years, the Greenland shark (Somniosus microcephalus) has been subject to increased scientific attention due to its role as a long-lived, highly migratory, and relatively abundant top predator in arctic marine food webs (MacNeil et al., 2012; Campana et al., 2015; Nielsen et al., 2016). The Greenland shark is distributed across the Arctic and in the cold temperate waters of the Atlantic and reaches more than 5 m in length. (Compagno, 1984; Mecklenburg et al., 2018). Although they are occasionally reported in shallow waters (<30 m, Harvey-Clark et al., 2005; Eriksen, 2011), Greenland sharks predominantly occur in deep waters (>200 m, Fisk et al., 2012; Campana et al., 2015), where they have been recorded as deep as 2,992 m near the mid-Atlantic Ridge (Porteiro et al., 2017). In Greenland waters, Greenland sharks are most often encountered at depths of 400–700 m (Nielsen et al., 2014).
Previous studies have investigated the diet and trophic interactions of Greenland sharks by analyzing stomach contents, stable isotopes, and fatty acids (Fisk et al., 2002; Yano et al., 2007; McMeans et al., 2010; Leclerc et al., 2012; McMeans et al., 2013; Nielsen et al., 2014). These studies are mostly based on relatively few individuals (N < 50) from 200 to 400 cm in length, with limited information on larger individuals. Ontogenetic dietary shifts have been documented for other large predatory sharks such as sevengill (Notorynchus cepedianus), white (Carcharodon carcharias), and tiger sharks (Galeocerdo cuvier) (Lowe et al., 1996; Ebert, 2002; Estrada et al., 2006), and preliminary data suggest that small Greenland sharks have a lower trophic position compared to larger sharks (Fisk et al., 2002). Dietary differences across Greenland shark size thus becomes relevant to investigate.
Size distribution of Greenland sharks appears to vary spatially. While sharks <400 cm seem to dominate in the Canadian Arctic, northern Greenland and the Svalbard Archipelago (Norway), larger sharks >400 cm seem to be more common off southern Greenland, Iceland, and Newfoundland (Nielsen et al., 2014; Campana et al., 2015; Lydersen et al., 2016; Devine et al., 2018).
This study documents the feeding ecology of the Greenland shark, investigated across a large size range, by analyzing stable isotopes and stomach contents. Specifically, we identified prey in stomach contents to the lowest possible taxonomical level, estimated their biomass, and calculated the relative importance of prey through ontogeny. Stable isotopes levels were used to determine whether prey composition was reflected in the 15N signal. Elaborating on the feeding ecology of different sized Greenland shark, currently listed as Near Threatened in the IUCN Red List of Threatened Species and as Data Deficient in the Norwegian Red List (Henriksen and Hilmo, 2015; IUCN, 2018), will allow for a more complete understanding of the species’ role as an important predator in arctic ecosystems.
Greenland sharks (N = 78) from Greenland waters were caught from 2013 to 2017 as part of the international collaboration project “Old and Cold – Biology of the Greenland shark.” Sampling was obtained by the Greenland Institute of Natural Resources (GINR) annual fish surveys (RV Pâmiut, RV Sanna), the TUNU IV and V Expeditions (RV Helmer Hanssen) (Christiansen, 2012), the commercial trawler FV Sisimiut, and targeted Greenland shark expeditions with RV Porsild and RV Sanna. All sampling was carried out in accordance with laws, regulations and authorization from the Government of Greenland (Ministry of Fisheries, Hunting and Agriculture, document number 565466, 935119, 20179208, C-17-129, C-15-17, and C-13-16). Overlap in both sampling area and season (West and East Greenland, inshore and offshore, May–September) allowed us to pool stomach content data with an additional 30 specimens caught in 2012 [reported separately in Nielsen et al. (2014)], increasing the total number of sharks included in this study to 108. All sharks were measured (total length, TL, cm) and the sex determined from the presence (males) or absence (females) of claspers. All shark lengths in this study are reported as TL. To evaluate possible size-related dietary differences, sharks were grouped into six “shark size groups” (TL bins); <200 cm, 201–250 cm, 251–300 cm, 301–350 cm, 351–400 cm, and >400 cm. As in previous studies, data for males and females were combined (see Yano et al., 2007; McMeans et al., 2010; Leclerc et al., 2012; Nielsen et al., 2014). Bin ranges were chosen as “juvenile” sharks of both sexes have previously been defined as those <200 cm (Hussey et al., 2015). In this study (and in general), sharks >400 cm are strictly females and further also potentially mature [cf. Yano et al. (2007) suggesting that males mature at ∼300 cm and females mature at >400 cm]. Shark size groups between 200 and 400 cm are thus composed by both immature males and females and potentially mature males.
Each stomach was removed and contents examined following the procedure described in Nielsen et al. (2014), with each prey item counted, measured, weighed, and identified to the lowest possible taxonomical level. All prey items were grouped into five prey categories: “Fish,” “Mammal,” “Squid,” “Crustacean,” and “Other.” The “Other” category included birds and all invertebrates except squids and crustaceans. The remaining stomach contents were designated “Non-prey items” and encompassed small rocks/stones, fishing gear, macro algae, metal pieces, fragments of Porifera, small bivalves (<1 cm), and scavenging lyssianassid amphipods. For all “Non-prey items” only frequency of occurrence was calculated.
In an attempt to reflect the initial (undigested) quantity of prey consumed, a “reconstructed biomass” was calculated for all prey items (except mammals, see explanation below). For each fish prey item, the reconstructed biomass was calculated from species or genus-specific length-weight relationship acquired from FishBase (Froese and Pauly, 2018). Squid size (pen length) and reconstructed biomass were calculated from beak size according to Zumholz and Frandsen (2006). No attempt was made to reconstruct the initial biomass of any mammal prey items (Nielsen et al., 2014) as it would be unrealistic to assume that an entire large marine mammal, potentially weighing hundreds or thousands of kilograms, was consumed by the shark. Therefore, for calculation purposes, the reconstructed biomass for each mammal prey item was set to be the actual wet mass as this would conservatively reflect the initial quantity consumed. Whenever possible, body morphometrics and wet mass of intact cnidarians (sea anemones), molluscs (gastropods), crustaceans (amphipods and decapods), echinoderms (sea urchins, sea stars, brittle stars, and sea cucumbers), and birds were used to estimate initial wet mass of digested specimens. The proportion of reconstructed biomass for each prey category (i.e., “Fish,” “Mammal,” “Squid,” “Crustacean,” and “Other”) was plotted as mean values (± SE) for each of the six shark size groups.
The index of relative importance (IRI) for prey items was calculated from the number of prey items (N), the reconstructed biomass of prey items (B) and the frequency of occurrence of prey items (F) expressed as proportions (%), as described in Nielsen et al. (2014). For shark size groups with significantly different 15N levels (cf. size group <200 cm, see section “Results”) the %IRI was calculated for each prey type. %IRI was also calculated separately for each of the six shark size groups and presented as a summarized %IRI-score for prey categories (“Fish,” “Mammal,” “Squid,” “Crustacean,” and “Other”) as well as at family level within these categories.
Stable isotope analysis of Greenland shark muscle tissue poses two challenges. Firstly, the muscle tissue is lipid rich which demands either chemical lipid extraction or mathematical correction to avoid bias from lipid carbon (Shipley et al., 2017). Secondly, shark muscle contains urea and trimethylamine N-oxide (TMAO), which tend to be depleted in 15N relative to protein N (Carlisle et al., 2017; Shipley et al., 2017). In order to compare data with the three main studies that present Greenland shark isotope values (Fisk et al., 2002; Hansen et al., 2012; and McMeans et al., 2013), we did not extract the TMAO, but performed lipid extraction. Hence, the δ13C values presented are from lipid extracted samples, whereas the δ15N values are from non-lipid extracted samples, thereby circumventing both the effect of δ13C depleted lipid on δ13C values and the potential effects of lipid extraction on δ15N values.
White muscle tissue (5–10 g) sampled dorsal to the vertebrae and anterior to the first dorsal fin, was dried at 60°C for at least 48 h and split in two aliquots. One sample was lipid extracted using 5 ml of a 2:1 chloroform/methanol mixture which was put into a sealed vial. The sample was allowed to extract for 24 h after which the supernatant was removed. This procedure was repeated three times. The vial and sample was then rinsed with another 5 ml of the chloroform/methanol mixture, and the rinsed sample was allowed to dry for 24 h in a fume hood. Both the lipid extracted and non-lipid extracted samples were pulverized in an agate mortar and 0.4–1 mg replicate samples of both aliquots were packed in tin capsules. The δ15N and δ13C values were standardized using a Gelatine A (Gel-A) standard with known isotopic values of δ15N = 5.4‰ and δ13C = -21.8‰. In order to correct for daily offsets and drift, two or three internal 0.2–0.7 mg Gel-A standards were assayed every nine or ten tissue samples.
All the samples were analyzed at Department of Bioscience, Center for Geomicrobiology, University of Aarhus, Aarhus, Denmark using comparable techniques to previous stable isotope analysis on Greenland sharks (e.g., Fisk et al., 2002). The samples were measured by means of Isotope Ratio Mass Spectrometry in combination with an Element Analyzer and an operational interface (Thermo Electon Corporation Flash EA 1112 series and Thermo Scientific Delta V Plus Isotope Ratio MS). Results are expressed in a δ notation as the deviation from international standards in parts per thousand (‰) according to the formula:
where X is either 13C or 15N, Rsample is 13C/12C or 15N/14N in the sample, and Rstandard is the ratio for the given standards. Standards for δ15N were calibrated against atmospheric air. Statistical analysis was carried out in the statistical computing program R (R Development Core Team, 2017). Analysis of variance and post hoc Tukey HSD test were used to evaluate the variation in δ15N and δ13C values between shark size groups. The level of significance was P < 0.05.
A total of 108 Greenland sharks were sampled from May to September 2012–2017 in inshore and offshore waters (Figure 1). Males measured from 104 to 372 cm (N = 27) and females from 81 to 474 cm (N = 79) (Supplementary Table S1). Due to wounds inflicted by conspecifics during capture, the sex of two sharks could not be determined. Stomachs were available from 102 specimens. Since fourteen stomachs were empty, 88 stomachs from sharks ranging in size from 81 to 474 cm were included in the stomach content analysis (Table 1 and Supplementary Table S1). The total wet biomass of stomach contents was 462.3 kg and ranged between 0.029 and 52.0 kg for individual stomachs (mean ± SD:5.3 ± 9.1 kg). In total, 3.5 kg was categorized as “Non-prey items” and of the remaining 458.8 kg of stomach contents, 96.9 % was assigned to one of the following prey categories: “Fish,” “Mammal,” “Squid,” “Crustacean,” or “Other.” The remaining 3.1% was made up by non-identifiable digested biological material. Of the 88 stomachs analyzed, a total of 697 prey items were identified representing 57 different prey types (Table 2).
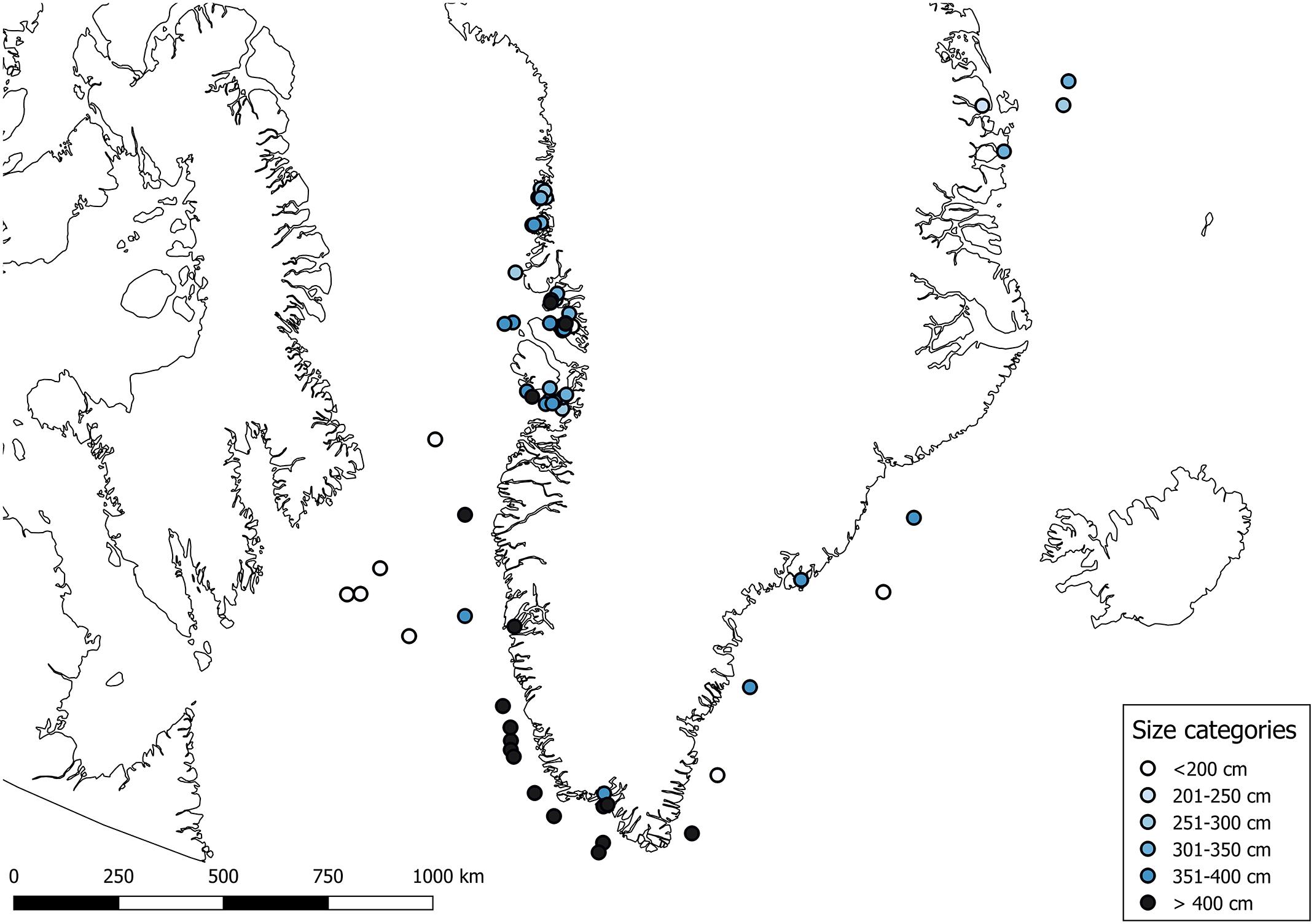
Figure 1. Capture locations for Greenland sharks (N = 108). Color indicates shark size (TL).
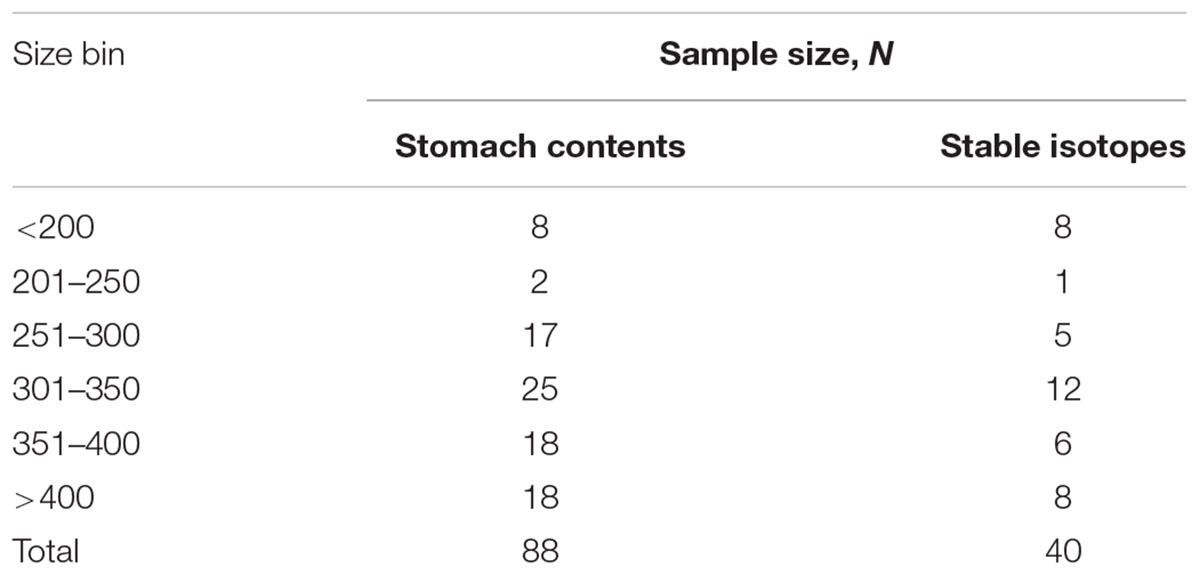
Table 1. Overview of number of sharks for each analysis (stomach contents and stable isotopes) in each of the six size bins.
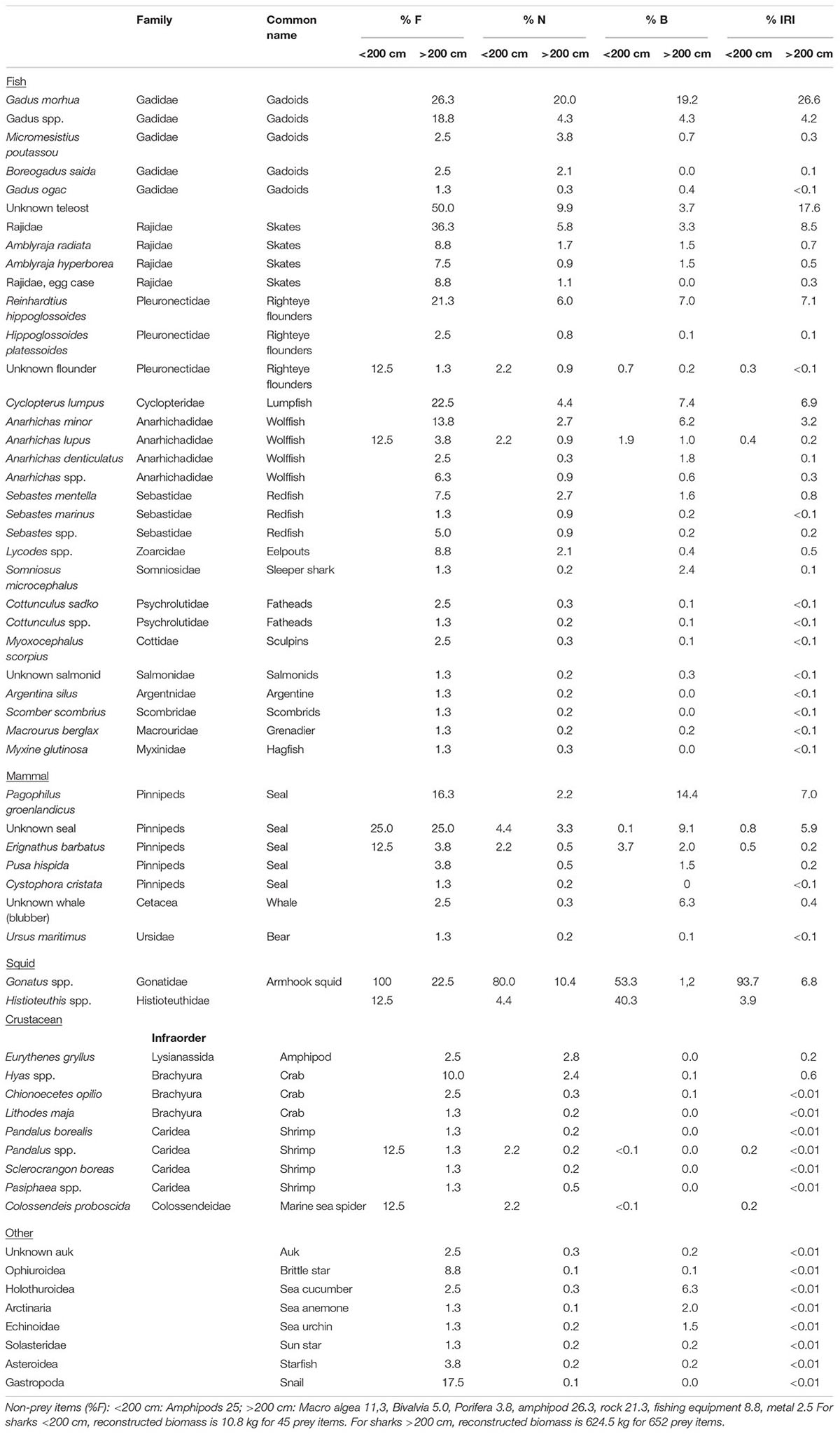
Table 2. %F = frequency of occurrence, %N = number of prey items, %B = reconstructed biomass, and %IRI = Index of relative importance, for sharks <200 cm (N = 8) and >200 cm (N = 80).
The total reconstructed biomass was 635.3 kg, adding an additional 176.5 kg to the observed biomass. The reconstructed biomasses for prey categories showed that “Squid” dominated the diet for Greenland sharks <200 cm, whereas “Fish” and “Mammal” were of major importance for all larger size groups (Figure 2). For these (i.e., sharks >200 cm), “Fish” constituted approximately 70% of the reconstructed biomass, and “Mammal” became gradually more prevalent with body length, increasing from 10 to 20% (Figure 2). For sharks <200 cm, armhook squid (Gonatus spp.) was present in all stomachs as the most dominant prey item (%IRI = 93.7, Table 2). For sharks >200 cm, Atlantic cod (Gadus morhua) (%IRI = 26.6) and unknown teleost (%IRI = 17.6) were the main prey followed by Greenland halibut (Reinhardtius hippoglossoides), skates (Rajidae), lumpfish (Cyclopterus lumpus), harp seal (Pagophilus groenlandicus), armhook squid, and spotted wolffish (Anarhichas minor) with %IRIs ranging from 3.2 to 7.9 (Table 2).
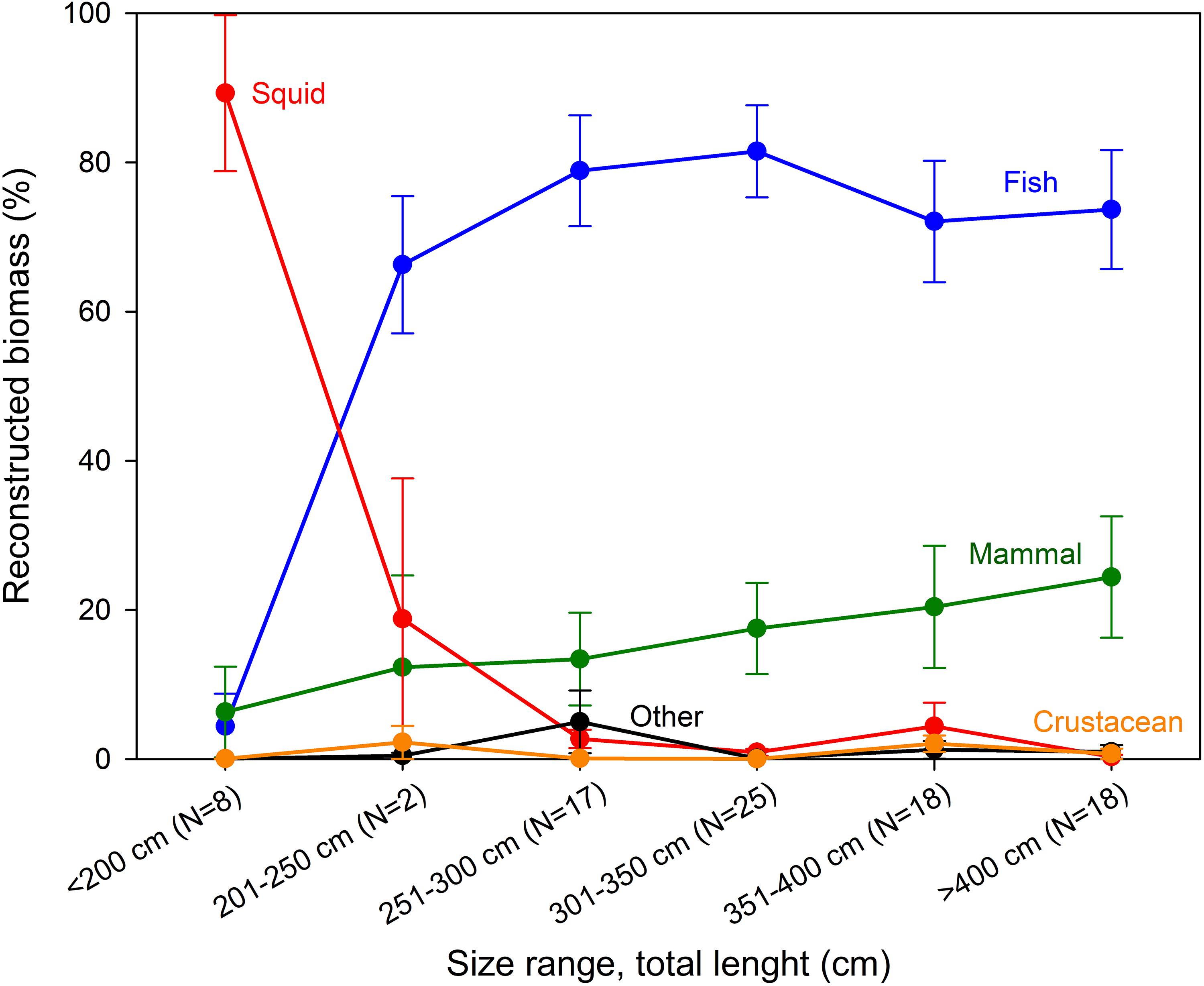
Figure 2. Proportional composition of reconstructed biomass of prey categories (“Fish,” “Mammal,” “Squid,” “Crustacean,” and “Other”) in the different shark size categories, for the 88 sharks with non-empty stomachs. Reconstructed biomass of each prey category is plotted as mean ± SE and the sample size (N) is given for each shark size category.
The summarized %IRI for each prey category is presented in Table 3 for the six shark size groups. Common to the four largest shark size groups (from 251 to 300 cm and larger), was that gadoids (dominated by Atlantic cod), Unknown teleost, and seal (dominated by harp seal) had a %IRI > 5 (Table 3). In contrast, righteye flounders (dominated by Greenland halibut) only had a %IRI > 5 for the 251–300 cm and 301–351 cm shark size groups (%IRI of 29.8 and 10.7, respectively), whereas righteye flounders had a %IRI < 2.5 for the two largest sizes classes (Table 3). Redfish (Sebastidae) were only important in the largest sharks >400 cm (%IRI > 5). Skates were of least importance in the largest sharks >400 cm (%IRI = 2.0), compared to sharks between 201 and 400 cm (%IRI range between 10.3 to 19.8, Table 3).
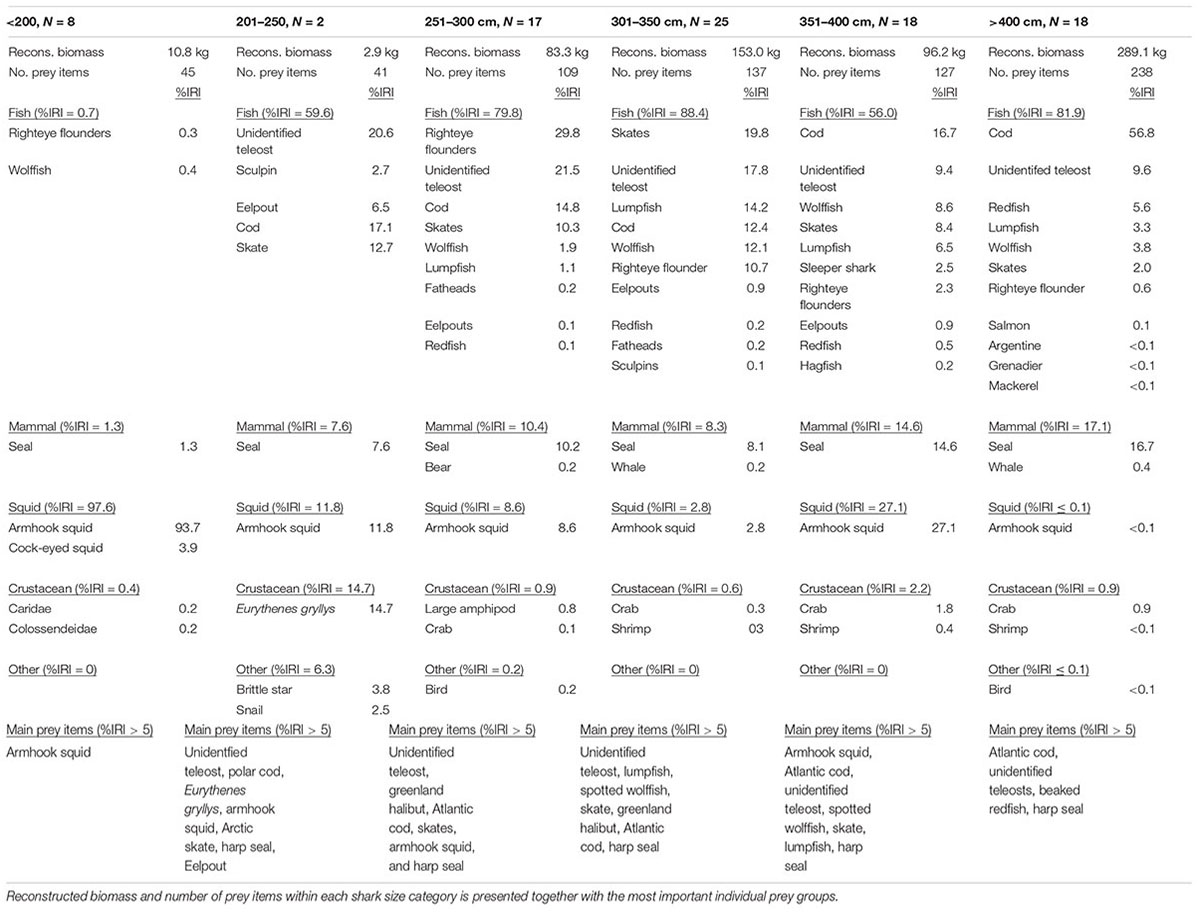
Table 3. %IRI calculated for each prey category and for prey families for each shark size category.
Samples for stable isotope analysis were available from 40 sharks measuring between 81 and 474 cm (Supplementary Table S1, Table 1). White muscle δ13C values ranged from -14.4 to -19.9‰ and δ15N between 11.8 and 17.2‰ (Supplementary Table S1, Figure 3). While there was no significant difference in δ13C values among size groups (ANOVA, F4,34 = 0.86, P = 0.5), δ15N values differed significantly (ANOVA, F4,34 = 5.8, P < 0.05, Figure 4). Sharks <200 cm had significantly lower δ15N values compared to the three largest size groups (post hoc Tukey HSD test P < 0.05) whereas the four largest size groups (251–300 cm, 301–350 cm, 351–400 cm, >400 cm) were not statistically different (Figure 4). As the δ15N value for the 201–250 cm size group was based on only one individual, it was not included in the ANOVA.
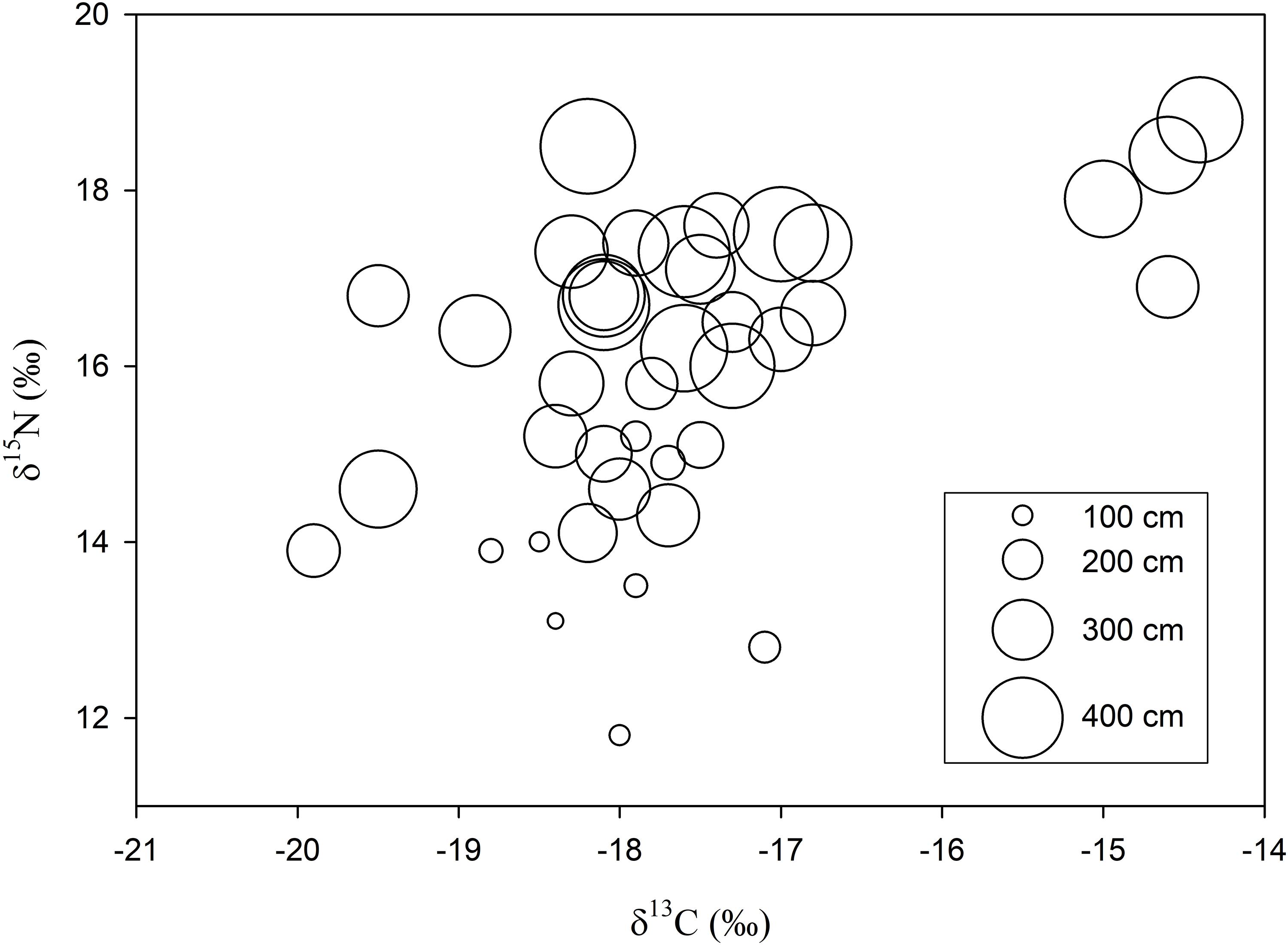
Figure 3. Bi-plot of δ13C and δ15N values for the 40 sharks ranging in size from 81 to 474 cm. The diameter of circles are proportional to the TL (cm) of the individual shark.
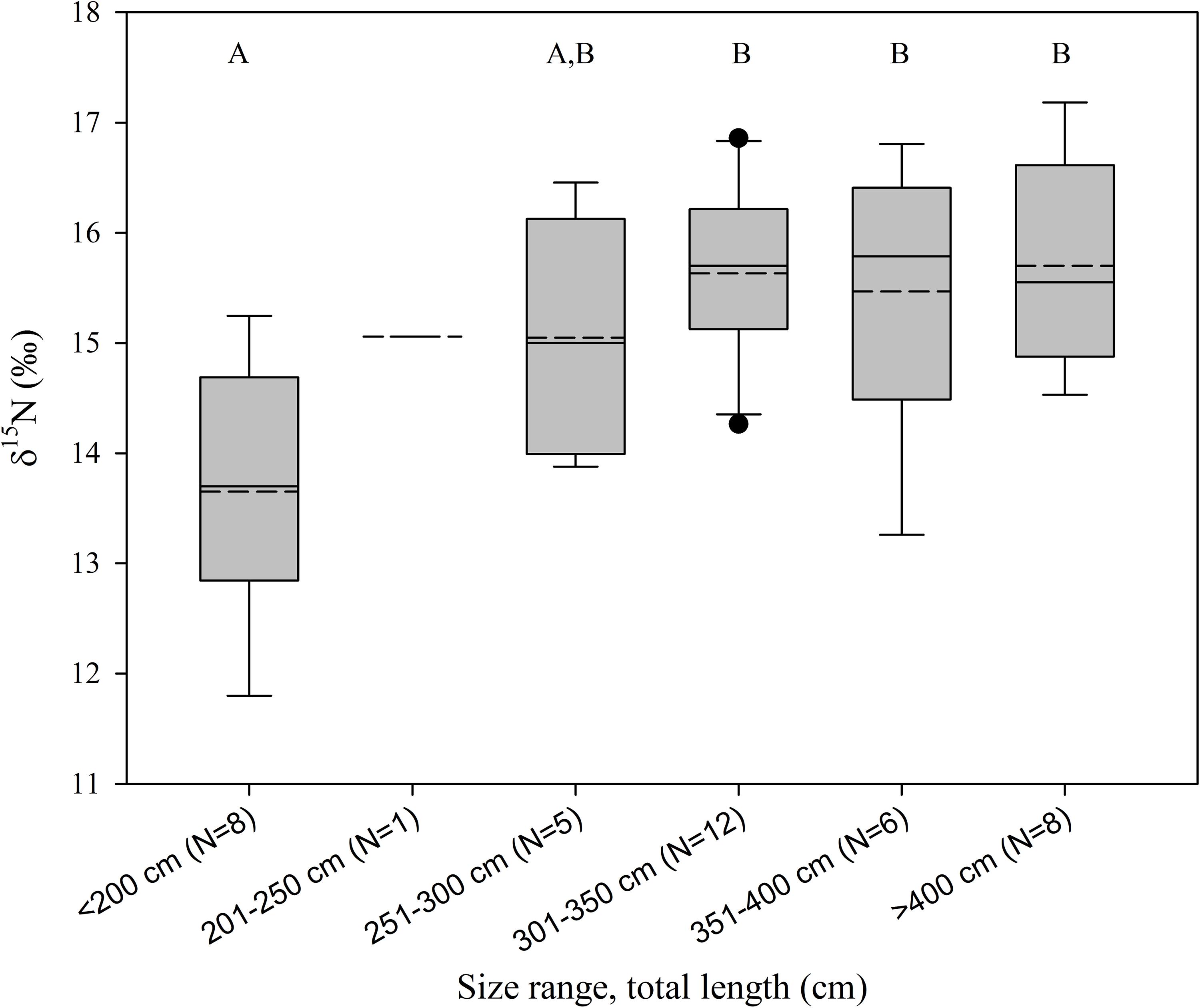
Figure 4. Boxplot [10th percentile, first quartile, median (dashed line), mean (full line) third quartile, and 90th percentile, dots represent outliers] of δ15N plotted against shark size categories. Different letters show significant differences between size categories.
This study investigated the ontogenetic shift of the diet of Greenland sharks and found that the smallest sharks (<200 cm) generally fed at a lower trophic level compared to larger sharks, both in terms of prey items and isotopic δ15N levels. This finding mirrors observations by Fisk et al. (2002), who reported lower δ15N values in two 135 cm sharks compared to fifteen specimens >250 cm. We also observed that the stomach contents of the <200 cm sharks were mainly armhook squid and the δ15N values were similar to that of other squid-feeding predators of the North Atlantic (Hooker et al., 2001; Mendes et al., 2007). The δ15N values of larger sharks >200 cm indicated a diet consisting of higher trophic level prey such as large piscivorous fishes and seals corresponding with stomach content results for these size classes, which primarily consisted of Atlantic cod, Greenland halibut and seals (see stable isotope values for Greenlandic marine food webs in Hansen et al., 2012). Taken together, the stomach contents and δ15N analyses suggests an ontogenetic shift from small (<200 cm), primarily squid-eating sharks, to larger (>200 cm), fish and mammal-eating sharks. However, sample size was limited for sharks from 201 to 250 cm, making it challenging to determine at which exact size the ontogenetic shift occurs.
When investigating the feeding ecology of a species, it is important to combine data of stomach contents and stable isotopes because stable isotopes provide an integrated and long-term estimate of trophic position that cannot be inferred from stomach content (Hobson and Welch, 1992; Christiansen et al., 2012). Although, stomach content analysis enables prey species identification and provides a detailed snapshot of recently ingested prey, regurgitation during capture and varying rates of digestion can skew the stomach content results leading to spurious conclusions, especially when sample size is small (Wetherbee et al., 2012). In this study, armhook squid was the single most important prey for the smallest sharks (<200 cm, %IRI = 97.6, Table 2), and was also relatively important in sharks >200 cm (%IRI = 6.8, Table 2), and more specifically, for 351–400 cm sharks (%IRI = 27.1, Table 3). For the reasons described below, we are not convinced that IRI-levels accurately reflect the importance of squids for larger sharks >200 cm. Firstly, one Greenland shark of 351 cm (GS17, Table 1) contained 32 squid beaks making this single observation accountable for 29% of all armhook squid observations in sharks >200 cm. Secondly, the contribution of the reconstructed biomass to the squid IRI-score is minimal but driven by a high frequency of occurrence (F) and numerical abundance (N) which is different from sharks <200 cm where all IRI parameters (F, N, and B) were high for squids. Thirdly, squids were much smaller (pen length mean ± SD: 16.8 ± 6.88 cm, N = 88) than prey items of >200 cm Greenland sharks typically measuring 21.3–68.8 cm in length (Nielsen et al., 2014). Lastly, the vast majority of squid observations were based on hard-to-digest chitin beaks, which for large sharks, especially, are prone to be derived from secondary stomach contents of large fully swallowed and digested prey like seal, Greenland halibut and Atlantic cod (a “matrijoska effect”). Based on the arguments outlined above, we believe that the importance of squids for sharks >200 cm is overestimated by our IRI calculations. In contrast, however, we believe that squids are very important for <200 cm sharks as indicated by their numerical abundance, frequency of occurrence, reconstructed biomass, and by δ15N values. The prominence of Atlantic cod (%IRI = 26.6) in the diet of sharks >200 cm was to some extent, also driven by observations from only few sharks, with 69% of all cod observations (87 of 127 specimens) being present in only two stomachs. However, Atlantic cod were recorded in 26% of all stomachs and are (in contrast to squids) very unlikely to be consumed secondarily. Thus the generally high importance of Atlantic cod seems a reasonable conclusion, which together with skates, seals, Greenland halibut, lumpfish, and wolffish are the most important prey for sharks >200 cm.
Our findings also show that prey fishes differ between shark size groups. For example, righteye flounder (mainly Greenland halibut, Table 3) were among the most important prey for sharks between 251 and 350 cm, but of limited importance to larger sharks (i.e., 351–400 and >400 cm). Furthermore, despite redfish being abundant across the Greenland continental shelf (ICES, 2017), they were only an important part of the diet among the largest sharks (>400 cm). Females this size from Iceland shelf waters have also been reported to feed predominantly on redfish (McMeans et al., 2010) suggesting some degree of prey selectivity. For sharks >400 cm, the scarcity or complete lack of deep sea fishes such as Greenland halibut, grenadiers and slickheads (Alepocephalidae) in the stomachs is noteworthy and is supported by depth records from a recent tagging study in Greenland waters (Nielsen, 2018), which found that >400 cm females do not spend much time at depths >800 m. Instead, females this size seem to mainly occupy and forage on the continental shelf and the upper part of the continental slope.
The apparent shift in diet between small and large sharks was corroborated by significantly lower δ15N values in sharks of <200 cm compared to sharks >200 cm. As noted in the Section “Materials and Methods” we did not attempt to extract TMAO and urea from the samples, though these compounds have been shown to lower the δ15N values to a variable extent (Carlisle et al., 2017). As we primarily used the isotopic values to compare between shark size groups, we believe any TMAO and urea produced bias in δ15N would be expected to have a similar effect across size groups. Hansen et al. (2012) analyzed Greenland shark samples from the same areas around Greenland (lengths 270 to 473 cm) and found average δ15N values of 16.7‰ which is similar to our values. Greenland sharks from Svalbard (lengths 245 to 404 cm) were reported by McMeans et al. (2013) to have average values of 15.9 (δ15N) and -18.6 (δ13C), which are also very similar to the ones reported here. In contrast, the δ13C values found by Hansen et al. (2012) were approximately 2‰ higher (-15.6‰ compared to -17.6‰ in our study). While the reason for the discrepancy between the two values is not clear, it could be due to varying isotopic baselines in different areas of Greenland (Hansen et al., 2012), as well as differences in the average size of the sharks examined in the different studies. Fisk et al. (2002) found δ15N values similar to this study, but had more depleted δ13C values, which may be attributed to their use of toluene as solvent to remove lipids.
For decades, scientists have noted the discrepancy between the lethargic appearance of the Greenland shark and the potentially faster moving prey species found in their stomachs (Jensen, 1914; Bigelow and Schroeder, 1948; Hansen, 1963; Watanabe et al., 2012) which has raised the question as to whether Greenland sharks are capable of active hunting or mainly feed as scavengers (Leclerc et al., 2012; Nielsen et al., 2014; Edwards et al., in press). Greenland shark scavenging events have been documented (Leclerc et al., 2011) but observations made in our study provide supporting evidence that Greenland sharks also are capable of active predation on fast swimming seals and large epibenthic fishes such as Atlantic cod and Greenland halibut. For example, we have observed circular bite wounds on large prey (Figures 5a,d) matching wounds observed on free swimming marine mammals (Figure 5c, Idrobo and Berkes, 2012 and MacNeil et al., 2012). Furthermore, two intact and freshly ingested seals were found without any scavenging fauna common for carcasses (Figure 5b) suggesting that the seals had been swallowed during a recent hunting event (Leclerc et al., 2012). How Greenland shark catch fast swimming prey such as seals remains to be resolved (Edwards et al., in press), but is has been suggested that predation occurs on seals sleeping in the water column (Leclerc et al., 2012).

Figure 5. (a) The characteristic circular gape shape of a Greenland shark (Photo: J. Nielsen). (b) Harp seals of 14 kg (top) and 19 kg (bottom) found in stomach of a 470 cm Greenland shark (Photo: J. Nielsen). (c) Circular bite marks on a freely swimming beluga whale (from MacNeil et al., 2012). (d) Seal chunks of skin, blubber, meat and ribs from shark stomach (Photo: J. Nielsen).
In conclusion, Greenland shark display a marked ontogenetic shift in trophic relationships from a squid diet in small sharks (<200 cm) to a diet dominated by fishes and seals in larger sharks (>200 cm). As the diet reflects available prey for given geographic areas and depths, how may we define Greenland shark in terms of feeding behavior? The squid diet for juvenile Greenland sharks is interesting and suggests a specialized feeding behavior where younger animals actively select squid, whereas larger animals appear generalists mainly feeding on demersal fishes and seals in addition to a minor proportion of benthic invertebrates. Clearly better information on the distribution, behavior and environments occupied by Greenland shark of different size and age is needed to clarify the understanding of the trophic relationships for this migratory and long-lived species.
JN coordinated this work. JN, HK, PB, KP, and JS collected the stomachs. JN and RH analyzed the stomach contents. HK, PG, and RH analyzed the stable isotopes. JN, JC, and RH designed the project and wrote the manuscript. All authors revised, reviewed, and finally approved the work.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We are grateful for the contribution of M. B. Backe and D. Bernal throughout the manuscript and for the great effort in the field by crew and student workers onboard research vessels RV Pâmiut, RV Sanna, RV Porsild, and RV Helmer Hanssen. We also thank the Greenland Institute of Natural Resources, the University of Copenhagen and the TUNU Programme (UiT The Arctic University of Norway) for ship time. Lastly, thanks to Thomas K. Kristensen assisting with identification of squid species from beak characteristics. This study is a part of the Greenland shark project (“Old and Cold - Biology of the Greenland shark”) which was financially supported by the Danish Council for Independent Research, GINR, Den Blå Planet-National Aquarium of Denmark, The Carlsberg Foundation, the Commission of Scientific Investigations in Greenland (KVUG), Danish Centre for Marine Research, Save Our Seas Foundation and National Geographic Foundation. The publication charges for this article have been funded by a grant from the publication fund of UiT The Arctic University of Norway.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2019.00125/full#supplementary-material
Bigelow, H. B., and Schroeder, W. C. (1948). “Sharks,” in Fishes of the Western North Atlantic, Part 1, eds J. Tee-Van, C. M. Breder, S. F. Hildebrand, A. E. Parr, and W. C. Schroeder (Yale, CT: Yale University, Sears Foundation for Marine Research), 59–546.
Campana, S. E., Fisk, A. T., and Klimley, A. P. (2015). Movements of Arctic and northwest Atlantic Greenland sharks (Somniosus microcephalus) monitored with archival satellite pop-up tags suggest long-range migrations. Deep Sea. Res. PT. II. 115, 109–115. doi: 10.1016/j.dsr2.2013.11.001
Carlisle, A. B., Litvin, S. Y., Madigan, D. J., Lyons, K., Bigman, J. S., Ibarra, M., et al. (2017). Interactive effects of urea and lipid content confound stable isotope analysis in elasmobranch fishes. Can. J. Fish. Aquat. Sci. 74, 419–428. doi: 10.1139/cjfas-2015-0584
Christiansen, J. S. (2012). “The TUNU-programme: Euro-Arctic marine fishes—diversity and adaptations,” in Adaptation and Evolution in Marine Environments, Volume 1, From Pole to Pole, eds G. Prisco and C. di Verde (Berlin: Springer-Verlag), 35–50. doi: 10.1007/978-3-642-27352-0_3
Christiansen, J. S., Hop, H., Nilssen, E. M., and Joensen, J. (2012). Trophic ecology of sympatric Arctic gadoids, Arctogadus glacialis (Peters, 1872) and Boreogadus saida (Lepechin, 1774), in NE Greenland. Polar Biol. 35, 1247–1257. doi: 10.1007/s00300-012-1170-y
Compagno, L. J. V. (1984). FAO Species Catalogue, Sharks of the World, An Annotated and Illustrated Catalogue of the Shark Species Known to Date, Part 1, Hexanchiformes to Lamniformes. Rome: FAO FisheriesSynopsis.
Devine, B. M., Wheeland, L. J., and Fisher, J. A. D. (2018). First estimates of Greenland shark (Somniosus microcephalus) local abundances in Arctic waters. Sci. Rep. 8:974. doi: 10.1038/s41598-017-19115-x
Ebert, D. A. (2002). Ontogenetic changes in the diet of the sevengill shark (Notorynchus cepedianus). Mar. Freshwater Res. 53, 517–523. doi: 10.1071/MF01143
Edwards, J. E., Broell, F., Bushnell, P. G., Campana, S. E., Christiansen, J. S., Devine, B. M., et al. (in press). Advancing our understanding of long-lived species: a case study on the Greenland shark. Front. Mar. Sci.
Estrada, J. A., Rice, A. N., Natanson, L. J., and Skomal, G. B. (2006). Use of isotopic analysis of vertebrae in reconstruction ontogenetic feeding ecology in white sharks. Ecology 87, 829–834. doi: 10.1890/0012-9658(2006)87[829:UOIAOV]2.0.CO;2
Fisk, A. T., Lydersen, C., and Kovacs, K. M. (2012). Archival pop-off tag tracking of Greenland sharks Somniosus microcephalus in the High Arctic waters of Svalbard, Norway. Mar. Ecol. Prog. Ser. 468, 255–265. doi: 10.3354/meps09962
Fisk, A. T., Tittlemier, S. A., Pranscke, J. L., and Norstrom, R. J. (2002). Using anthropogenic contaminants and stable isotopes to assess the feeding ecology of Greenland sharks. Ecology 83, 2162–2172. doi: 10.2307/3072048
Froese, R., and Pauly, D. (eds) (2018). FishBase. www.fishbase.org version (09/2018).
Hansen, J. H., Hedeholm, R. B., Sünksen, K., Christensen, J. T., and Grønkjær, P. (2012). Spatial variability of carbon (δ13C) and nitrogen (δ15N) stable isotope ratios in an Arctic marine food web. Mar. Ecol. Prog. Ser. 467, 47–59. doi: 10.3354/meps09945
Hansen, P. M. (1963). Hajer i Grønlandske Farvande. Charlottenlund: Skrivelse Danmarks Fiskeri-og Havundersøgelse, 83–95.
Harvey-Clark, C. J., Gallant, J. J., and Batt, J. H. (2005). Vision and its relationship to novel behaviour in St. Lawrence River Greenland Sharks, Somniosus microcephalus. Can. Field Nat. 119, 355–359. doi: 10.22621/cfn.v119i3.145
Hobson, K. A., and Welch, H. E. (1992). Determination of trophic relationships within a high Arctic marine food web using δ13C and δ15N analysis. Mar. Ecol. Prog. Ser. 84, 9–18. doi: 10.3354/meps084009
Hooker, S. K., Iverson, S. J., Ostrom, P., and Smith, S. C. (2001). Diet of northern bottlenose whales inferred from fatty-acid and stable-isotope analyses of biopsy samples. Can. J. Zool. 79, 1442–1454. doi: 10.1139/z01-096
Hussey, N. E., Cosandey-Godin, A., Walter, R. P., Hedges, K. J., VanGerwen, T., Barkley, A. N., et al. (2015). Juvenile Greenland sharks Somniosus microcephalus (Bloch & Schneider, 1801) in the Canadian Arctic. Polar Biol. 38, 493–504. doi: 10.1007/s00300-014-1610-y
Idrobo, C. J., and Berkes, F. (2012). Pangnirtung inuit and the Greenland shark: co-producing knowledge of a little discussed species. Hum. Ecol. 40, 405–414. doi: 10.1007/s10745-012-9490-7
Jensen, A. S. (1914). The Selachians of Greenland. Copenhagen: Zoological Museum of the University Copenhagen. doi: 10.5962/bhl.title.11662
Leclerc, L. M., Lydersen, C., Haug, T., Bachmann, L., Fisk, A. T., and Kovacs, K. M. (2012). A missing piece in the Arctic food web puzzle? Stomach contents of Greenland sharks sampled in Svalbard, Norway. Polar Biol. 35, 1197–1208. doi: 10.1007/s00300-012-1166-7
Leclerc, L. M., Lydersen, C., Haug, T., Glover, K. A., Fisk, A. T., and Kovacs, K. M. (2011). Greenland shark (Somniosus microcephalus) scavenge offal from minke (Balaenoptera acutorostrata) whaling operations in Svalbard (Norway). Polar Res. 30:7342. doi: 10.3402/polar.v30i0.7342
Lowe, C. G., Wetherbee, B. M., Crow, G. L., and Tester, A. L. (1996). Ontogenetic dietary shifts and feeding behavior of the tiger shark, Galeocerdo cuvier, in Hawaiian waters. Environ. Biol. Fish. 47, 203–211. doi: 10.1007/BF00005044
Lydersen, C., Fisk, A. T., and Kovacs, K. M. (2016). A review of Greenland shark (Somniosus microcephalus) studies in the Kongsfjorden area, Svalbard Norway. Polar Biol. 39, 2169–2178. doi: 10.1007/s00300-016-1949-3
MacNeil, M. A., McMeans, B. C., Hussey, N. E., Vecsei, P., Svavarsson, J., Kovacs, K. M., et al. (2012). Biology of the Greenland shark Somniosus microcephalus. J. Fish Biol. 80, 991–1018. doi: 10.1111/j.1095-8649.2012.03257.x
McMeans, B. C., Arts, M. T., Lydersen, C., Kovacs, K. M., Hop, H., Falk-Petersen, S., et al. (2013). The role of Greenland sharks (Somniosus microcephalus) in an Arctic ecosyste assessed via stable isotopes and fatty acids. Mar. Biol. 160, 1223–1238. doi: 10.1007/s00227-013-2174-z
McMeans, B. C., Svarvarsson, J., Dennard, S., and Fisk, A. T. (2010). Diet and resource use among Greenland sharks (Somniosus microcephalus) and teleosts sampled in Iceland waters, using δ13C, δ15N, and mercury. Can. J. Fish. Aquat. Sci. 67, 1428–1438. doi: 10.1139/F10-072
Mecklenburg, C. W., Lynghammar, A., Johannesen, E., Byrkjedal, I., Christiansen, J. S., Dolgov, A. V., et al. (2018). Marine Fishes of the Arctic Region, Vol I. Available at: http://hdl.handle.net/11374/2116
Mendes, S., Newton, J., Reid, R. J., Zuur, A. F., and Pierce, G. J. (2007). Stable carbon and nitrogen isotope ratio profiling of sperm whale teeth reveals ontogenetic movements and trophic ecology. Oecologia 151, 605–615. doi: 10.1007/s00442-006-0612-z
Nielsen, J. (2018). The Greenland Shark (Somniosus Microcephalus) – Diet, Tracking and Radiocarbon Age Estimates Reveal the World’s Oldest Vertebrate. Doctoral dissertation, Department of Biology, Copenhagen.
Nielsen, J., Hedeholm, R. B., Heinemeier, J., Bushnell, P. G., Christiansen, J. S., Olsen, J., et al. (2016). Eye lens radiocarbon reveal centuries of longevity in the Greenland shark (Somniosus microcephalus). Science 353, 702–704. doi: 10.1126/science.aaf1703
Nielsen, J., Hedeholm, R. B., Simon, M., and Steffensen, J. F. (2014). Distribution and feeding ecology of the Greenland shark (Somniosus microcephalus) in Greenland waters. Polar Biol. 37, 37–46. doi: 10.1007/s00300-013-1408-3
Porteiro, F. M., Sutton, T. T., Byrkjedal, I., Orlov, A. M., Heino, M., Menezes, G., et al. (2017). Fishes of the northern Mid-Atlantic Ridge collected during the MAR-ECO cruise in June–July 2004: an annotated checklist. Arquipelago Life Mar. Sci. 10, 1–125.
R Development Core Team (2017). R: A Language and Environment for Statistical Computing. Available at: https://www.R-project.org
Shipley, O. N., Brooks, E. J., Madigan, D. J., Sweeting, C. J., and Grubbs, R. D. (2017). Stable isotope analysis in deep-sea chondrichthyans: recent challenges, ecological insights, and future directions. Rev. Fish. Biol. Fisher. 27, 481–497. doi: 10.1007/s11160-017-9466-1
Watanabe, Y., Lydersen, C., Fisk, A. T., and Kovacs, K. M. (2012). The slowest fish: swim speed and tail-beat frequency of Greenland sharks. J. Exp. Mar. Bio. Ecol. 426-427, 5–11. doi: 10.1016/j.jembe.2012.04.021
Wetherbee, B. M., Cortés, E., and Bizzarro, J. J. (2012). “Food consumption and feeding habits,” in Biology of Sharks and Their Relatives, eds J. C. Carrier, J. A. Musick, and M. R. Heithaus (Boca Raton, FL: CRC Press),239–264.
Yano, K., Stevens, J. D., and Compagno, L. J. V. (2007). Distribution, reproduction and feeding of the Greenland shark Somniosus (Somniosus) microcephalus, with notes on two other sleeper sharks, Somniosus (Somniosus) pacificus and Somniosus (Somniosus) antarcticus. J. Fish Biol. 70, 374–390. doi: 10.1111/j.1095-8649.2007.01308.x
Keywords: Greenland shark, diet, feeding ecology, stomach content, isotopes
Citation: Nielsen J, Christiansen JS, Grønkjær P, Bushnell P, Steffensen JF, Kiilerich HO, Præbel K and Hedeholm R (2019) Greenland Shark (Somniosus microcephalus) Stomach Contents and Stable Isotope Values Reveal an Ontogenetic Dietary Shift. Front. Mar. Sci. 6:125. doi: 10.3389/fmars.2019.00125
Received: 28 September 2018; Accepted: 28 February 2019;
Published: 04 April 2019.
Edited by:
Jeremy Kiszka, Florida International University, United StatesReviewed by:
Charlie Huveneers, Flinders University, AustraliaCopyright © 2019 Nielsen, Christiansen, Grønkjær, Bushnell, Steffensen, Kiilerich, Præbel and Hedeholm. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Julius Nielsen, anVsaXVzbmllbHNlbjg4QGdtYWlsLmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.