 Gaojian Li
Gaojian Li Enoch Obeng1
Enoch Obeng1 Yulong He
Yulong He
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Immunol., 07 October 2020
Sec. Comparative Immunology
Volume 11 - 2020 | https://doi.org/10.3389/fimmu.2020.510943
Mycoplasma hyopneumoniae (M. hyopneumoniae, Mhp) is a geographically widespread and economically devastating pathogen that colonizes ciliated epithelium; the infection of Mhp can damnify the mucociliary functions as well as leading to Mycoplasma pneumonia of swine (MPS). MPS is a chronic respiratory infectious disease with high infectivity, and the mortality can be increased by secondary infections as the host immunity gets down-regulated during Mhp infection. The host immune responses are regarded as the main driving force for the disease development, while MPS is prone to attack repeatedly in farms even with vaccination or other treatments. As one of the smallest microorganisms with limited genome scale and metabolic pathways, Mhp can use several mechanisms to achieve immune evasion effect and derive enough nutrients from its host, indicating that there is a strong interaction between Mhp and porcine organism. In this review, we summarized the immune evasion mechanisms from genomic variability and post-translational protein processing. Besides, Mhp can induce the immune cells apoptosis by reactive oxygen species production, excessive nitric oxide (NO) release and caspase activation, and stimulate the release of cytokines to regulate inflammation. This article seeks to provide some new points to reveal the complicated interaction between the pathogen and host immune system with Mhp as a typical example, further providing some new strategies for the vaccine development against Mhp infection.
Mycoplasma hyopneumoniae (M. hyopneumoniae, Mhp) is one of the primary pathogens of Mycoplasma pneumonia of swine (MPS), also known as enzootic pneumonia (EP) (1). This chronic respiratory pathogen mainly leads to decreased daily gain weight and serious clinical symptoms in growing and finishing pigs. MPS is reported worldwide and causes significant economic losses, mainly from the treatment, immunization, control/eradication and increased mortality caused by secondary infections (viruses, bacteria or parasites) (2). The in vitro culture of Mhp is strenuous because of its fastidious requirement for culture medium (3, 4), which limits the large-scale preparation and industrial production of bacteria-based vaccines. At present, commercial genetically engineered vaccines and other types of novel vaccines are still under development; vaccination (inactivated/attenuated vaccines) is the main method to control MPS, but it provides partial protection, and it cannot eliminate Mhp from farms or bodies completely (5).
Mhp is one of the smallest self-replicating microorganisms with less cell walls and reduced genome scale; these characteristics make Mhp rely on its surroundings (in vivo/in vitro) to acquire adequate nutrients like nucleic acid, lipid, and amino acid for its proliferation (6). Also, the limited genome size and protein expression scale allow Mhp to complicate its proteomics by Post-translational protein processing (6). Mhp infection is widespread in farms and also difficult to eliminate, which indicates a certain degree of immune evasion. However, a clear picture of the immune evasion mechanisms of Mhp and its interaction with host immune system is still missing.
In this review, we summarized the mechanisms of Mhp immune evasion from genomic variability and Post-translational protein processing. In addition, Mhp can induce the apoptosis of immune cells by reactive oxygen species (ROS) production, excessive nitric oxide (NO) release and caspase activation, and stimulate the release of cytokines to regulate inflammation. This article aims to use Mhp as an example to outline the comprehensive interaction between pathogen and the host immune system, and new strategies for the next-generation of efficient vaccine development for the control of MPS are also proposed.
The genomes of 12 Mhp strains (J, 7448, 232, 168, 168-L, 7422, KM014, TB1, 11, NCTC10127, ES-2, and F7.2C) were sequenced and uploaded into the database of the National Center for Biotechnology Information (NCBI). In general, the average genomic size of Mhp is 0.9199 Mb, encoding 744 genes and 645 proteins, the mean GC content is 28.54%, which is a significantly low value compared with other species. Figure 1 shows the genomic GC content of Mhp and other species; Supplementary Table 1 provides the detailed genomic GC content information of these species. Studies had reported that the GC-rich regions had more conservative mutation rate compared with the GC-poor regions (7). On the other hand, GC content is an important parameter for genome organization and gene expression (8, 9), the low GC content gives Mhp complex transcriptional organization, unique intrinsic terminator stem-loop formation and individual ribonuclease P (RNase P) structure (10, 11). Earlier studies demonstrated that Mollicutes genomes had increased recombination and mutation rate as the environment changed (12), and the high genomic diversity between different Mhp strains was reported in follow-up experiments (13).

Figure 1. The comparison of genomic GC content between Mycoplasma hyopneumoniae and other species, indicating a low GC content in almost all Mycoplasma species except Mycoplasma pneumoniae.
Genomic diversity can be associated with discrepant pathogenicity of pathogens (14). A comprehensive genomic analysis was performed to evaluate the genomic variability between pathogenic strain Mhp 168 and its high-passaged attenuated strain Mhp 168-L. Some mutative coding sequences were found in Mhp 168-L including adhesins (P97, P102, P146, P159, P216, and LppT), cell envelope protein (P95), cell surface antigen (P36), secretory proteins, and chaperone protein (DnaK), which could be associated with their different pathogenicity (15). Results from other experimental methods also supported the genomic variability between Mhp strains, including multiple locus variable number tandem repeat analysis (MLVA) (13, 16–18), random amplified polymorphic DNA (RAPD) analysis, pulsed-field gel electrophoresis and nested polymerase chain reaction (nPCR) (19–22). This review (23) provides detailed information about the Mhp genomic classification and variability.
In brief, genomic variability of Mhp has been identified, and the effects for controlling MPS are summarized as follows. (1) Gene mutations in immunogens like P97 and P146 (both of P97 and P146 are important adhesins and commonly used as immunogens in genetically engineered vaccines) have a negative impact for the vaccine development (24, 25). (2) Antibiotic efficacy decreased with the mutations happened in topoisomerase enzymes (26–29) and 23S rRNA (30, 31). (3) Genomic variability increases the porcine susceptibility when infected by different Mhp strains.
It is generally accepted that bacteria can use protein hydrolysis processing to accommodate the changing environment and manipulate host immune responses (32). Mhp cell surface proteins can be processed to play roles in cell growth, biofilm formation, peptide secretion and pathogenesis (33, 34). For bacteria with reduced genomes, proteolytic regulation allows them to produce novel proteoforms and digest host cell proteins for their synthesis (35). It is reported that the lipid-associated membrane proteins (LAMPs) from Mhp are necessary for cell adhesion, infection, disease development, and immunosuppressive effect (36). For example, a multifunctional adhesin mhp390 can induce apoptosis in different immune cells and exacerbate inflammatory responses (37). Besides, two surface-exposed Mhp proteases putative Xaa-Pro aminopeptidase (MHJ_0659; PepP) and oligoendopeptidase F (MHJ_0522; PepF) share substrates and participate in the cleavage of many biologically active proteins (bradykinin, substance P, neurokinin A, and neuropeptide Y) for the regulation of lung homeostasis (38).
The Post-translational protein processing events in Mhp proteomics can expand protein functions and expose more epitopes (39), which may increase the host immune burden. Protein cleavage events were reported in adhesins (25, 33, 34, 40–46), lipoproteins (6) as well as the surface moonlighting proteins (6, 39, 47, 48) (Table 1). Besides, plasminogen is an available host extracellular component in the respiratory tract, which can repair damaged tissues and regulate inflammatory responses (50, 51). Studies had found that during the experimental Mhp infection, plasmin activity was enhanced as a part of the host immune responses, and vaccination could weaken this effect (52). In this process, plasminogen can be activated by P97/P102 adhesion family proteins (53), P116 (44), glutamyl aminopeptidase (GAP; MHJ_0125) (47), leucine aminopeptidase (LAP; MHJ_0461) (54), and fructose-1, 6-bisphosphate aldolase (FBA) (55). The plasminogen activation can improve the Mhp adhesion efficiency and stimulate cytokines release like tumor necrosis factor α (TNF-α), interleukin (IL)-1β, and IL-6 (52), these may be secreted by endothelial cells, monocytes, macrophages, and dendritic cells (DCs) (56).
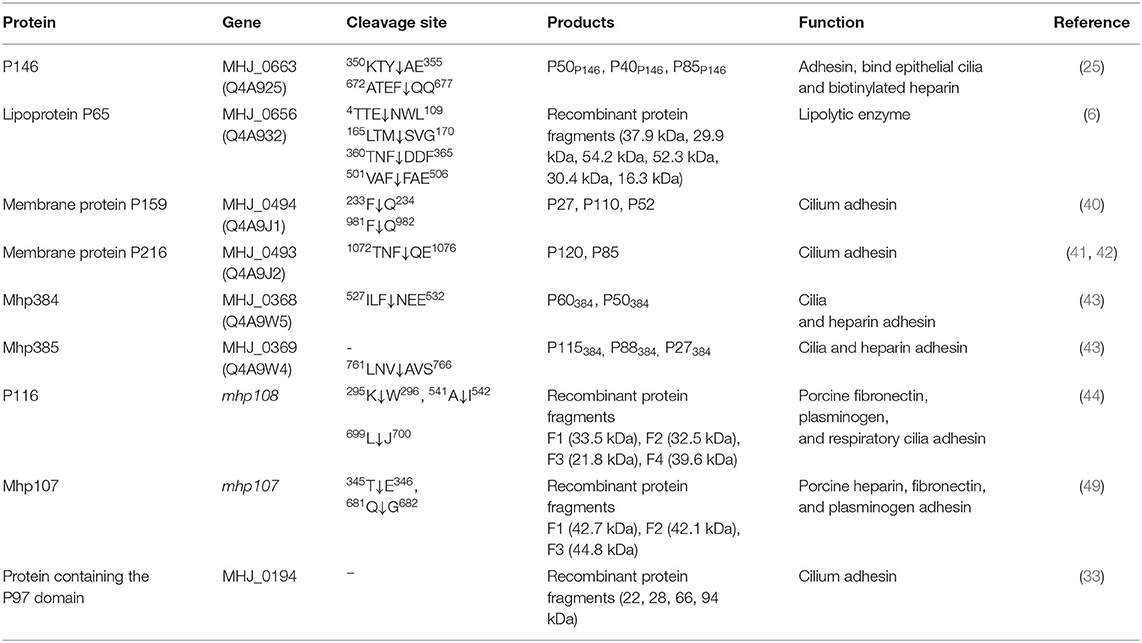
Table 1. Protein processing events reported in Mhp proteins.
MIB–MIP is a mycoplasmal system that captures and cleaves immunoglobulin (Ig) G, which is mediated by Mycoplasma immunoglobulin binding protein (MIB) and Mycoplasma immunoglobulin protease (MIP), respectively. The MIB-MIP system had been reported in Mycoplasma mycoides subspecies capri, and the multiple copies of MIB-MIP genes were also found in Mhp genome (57), while the role of the MIB-MIP system in the interaction between Mhp and host immune system still needs further studies.
In short, as a genome-reduced pathogen, Post-translational protein processing and genomic variability complicate the Mhp cell surface antigen appearance and expand protein functions, which may aggravate disease development and regulate lung homeostasis to promote the proliferation of Mhp. For the immune system, abundant exposed epitopes can affect the Pathogen-Associated Molecular Patterns (PAMPs) to reduce immune efficiency, and more functional virulence factors can contribute to the negative regulation of the immune system.
Different immune cells can form a complex and sophisticated network to recognize pathogens and provide direct defense against their invasion. In the respiratory tract, the mucosal immune system is constructed by epithelial barriers, mucosal dendritic cells, innate lymphoid cells, natural killer cells, natural killer T cells, mast cells, and eosinophils (58). Mhp can invade organisms by endotracheal, intranasal or aerosol, while the endotracheal is the most effective method to induce MPS experimentally (59). Thereafter, Mhp can adhere to the cilium surface and spread through the respiratory tract, destroying cilium structures and causing apoptosis to epithelial cells (60). Clinical symptoms of MPS can be summarized as dry coughing, dyspnea, inflammation response, and immunosuppression (61, 62), the accumulation of mononuclear leukocytes in bronchioles and perivascular was also reported (63). Partial pattern diagram of Mhp infection is shown in Figure 2, Mhp infection destroys cilium structures and induces the apoptosis of epithelial cells, which may increase the risk of secondary infections. Mhp can survive inside the epithelia cells and move to other tissues with disease development. The production of secretory IgA (SIgA), IgG, and phagocytic action of macrophage are involved in the immune responses.
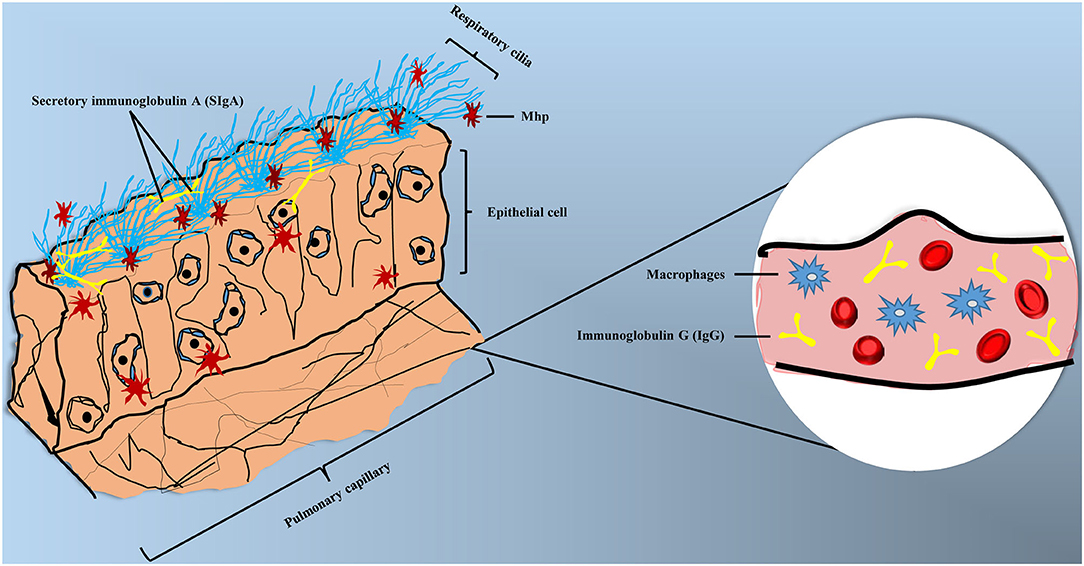
Figure 2. Partial pattern diagram of Mhp infection. Mhp infection destroys the cilium structures, leading to the apoptosis of epithelial cells, as well as increasing the secondary infection risk. Mhp can survive inside the epithelia cells and move to other tissues with disease development. The production of secretory IgA (SIgA) and IgG and phagocytic action of macrophage are involved in the immune responses.
Studies had reported that PAMs could recognize and provide immune defense during Mhp infection (64). Before and after Mhp infection, the high throughput cDNA microarray assays were used to evaluate the PAMs responses and found some cell behaviors were changed significantly, which could be clustered into inflammatory response, apoptosis, anti-apoptosis, programmed cell death, defense response, signal transduction, and pattern recognition receptors (PRRs) activation (65). PAMs can express Toll-like receptors (TLRs), and the TLR2-TLR6 dimeride can recognize the PAMPs from Mhp, thereby activating the classical Toll/IL-1 receptor signal pathway (66, 67). The up-regulation of TLR2 and TLR4 can also increase the level of IgA in mouse LDC/B cell coculture model (68). Besides, Mhp infection improves the level of proinflammatory cytokines like TNF-α, IL-1β, and IL-6 in PAMs (69, 70), which can be regulated by activator protein-1, nuclear factor κB (NF-κB) and mitogen-activated protein kinase (71, 72).
The regulation of cytokines can also be mediated through other signaling pathways. The cytochrome P450 1A1 (CYP1A1) was down-regulated in lung tissues during Mhp infection; further studies found that the overexpression of CYP1A1 could activate the PPAR-γ signaling pathway and up-regulated the IL-1β, IL-6, IL-8, and TNF-α levels in Mhp-treated PAMs (70). Moreover, sonicated protein fractions from Mhp could stimulate PAMs to produce higher level of IL-1β, IL-6, TNF-α, COX-2, and inducible NO synthase. Besides, differential genes expression analysis indicated that Mus musculus chemokine (C-C motif) ligand3 (Ccl3) and M. musculus serum amyloid A 3 (Saa3) were associated with the inflammatory responses (69). In general, PAMs are involved in inflammatory responses during Mhp infection with the excessive production of proinflammatory cytokines both in vitro (64, 73) and in vivo (52, 74, 75).
NO is a multifunctional biological molecule produced by NO synthase, studies have reported that some Mycoplasma species can induce immunosuppressive effect by excess NO production (69, 76, 77). The LAMPs from Mhp can induce a time-dependent apoptosis to PAMs by NO production, oxidative stress response, and caspase-3 activation (36). In addition, research in Mhp-treated murine alveolar macrophage found that the up-regulation of NO level and proinflammatory cytokines were associated with the activation of NF-κB and three MAPK signaling pathways (64). On the other hand, the increased synthesis level of phosphoribosyl pyrophosphate (PRPP), nicotinamide adenine dinucleotide (NAD), and nucleotide in pathogenic strain Mhp 168 can be regarded as an effective protection method for potential ROS damage to Mhp cells (78).
Mhp cannot synthesize purines or pyrimidines de novo while it is able to assimilate exogenous nucleobases and nucleosides and then synthesizes nucleotides by salvage pathways and interconversions (79). Studies had shown that Mhp can digest extracellular DNA to form biofilm in the respiratory tract and abiotic surfaces, which could be inhibited by nuclease treatment (80). Besides, the multifunctional nuclease Mhp597 had been reported (81), and Mhp597 was able to digest extracellular nucleic acid such as induced macrophage extracellular traps (METs) to synthesize nucleotides (80, 82, 83). Another study found Mhp could assimilate nucleotides more effective from METs than free nucleotides in medium, which suggested a close connection between nuclease degradation and nucleotide transportation (84). In general, PAMs can provide nucleotides and protein synthesis materials for Mhp proliferation, and they may serve as a shelter for Mhp to escape immune attacks (85).
DCs are the most powerful antigen presenting cells that can activate effector CD4+ T cells, cross-present antigens to CD8+ T cells, and stimulate B cells to produce antibodies (86). DCs are widely distributed among respiratory tracts and have a high density above and below the basement membrane of the tracheal epithelium, which allows DCs to efficiently ingest, process, and present antigens (87). Studies showed that the quantity of SIgA positive cells, T cells, SLA-II-DR+CD11b+ DCs, and SLA-II-DR+SWC3a+ DCs decreased obviously during Mhp infection (88). In addition, the level of IL-12 and interferon γ (IFN-γ) was also down-regulated, which indicated that DCs preferred Th2 immune response (89). Another study evaluated the cytokines production of bone marrow–derived DCs (BMDCs) in a coinfection model, and reported the lower IL-12 production of Mhp infection and higher IL-10 level with the coinfection of M. flocculare and Mhp (90). Vaccine strain Mhp 168-L treated BMDCs can activate T-cell proliferation effectively, while the up-regulation of IL-10 and down-regulation of IFN-γ may limit its effect (91).
Previous studies evaluated the capability of antibody production by peripheral blood mononuclear cells (PBMCs) and found that older pigs possessed a higher antibody level during Mhp infection (92). Follow-up studies reported Mhp infection could induce apoptosis to PBMCs by ROS and NO production, which were related to the Bax/Bcl-2, p38 MAPK signaling pathway and caspase activation (93). When PBMCs were treated with heat-killed Mhp cells, the proinflammatory cytokines (TNF-α, IL-1β, IL-8, and IL-18) and anti-inflammatory cytokine (IL-10) were up-regulated; at the same time, the level of antigen-specific IFN-γ and IL-10 were gradually decreased during infection (74). These results indicated that Mhp could regulate immune responses by inducing several cytokines secretion and cause immunosuppression with disease development.
The mucosal surface of the respiratory tract is the first defense line to prevent pathogens infection and eliminate microorganisms (bacteria, virus, parasites, and fungi) or foreign bodies by producing active components like SIgA (94, 95). On the other hand, mucosal immunity is an important sign of Mhp infection and could be improved by cathepsin L, which should be considered into the medication development (96).
The deviation of IgG subclass during Mhp infection indicates a Th1/Th2 response bias, in general, the IL-12 is related to the Th1 immune response, the IL-4 and IL-6 are related to the Th2 immune response, while the specific immune type that Mhp induced remains ambiguous (97). Studies found a lower production of IL-12 during Mhp infection, which indicated that the immune system preferred the Th2 immune response (90). Some studies also reported that Mhp could induce a higher level of IgG1 in local and systemic immune responses, which could stimulate the differentiation of naive Th1 cells to Th2 cells (74). On the contrary, some experiments reported the secretion of IgG2 dominated the humoral immune responses, and indicated Mhp could firstly induce the cell-mediated immunity (97–99). In addition, Mhp was thought to exist only extracellularly and adhere to the surface of respiratory tract (60), while recent studies have found that Mhp can also survive inside the epithelial cells (100). Meanwhile, the Th1 immune response can resist the invasion of intracellular pathogens, indicating that Mhp can induce cellular immune response firstly (98).
At present, vaccination (inactivated/attenuated vaccines) is a commonly used and effective method to prevent Mhp infection, while these vaccines provide partial protection and cannot cure MPS or eliminate Mhp from farms or bodies completely. Vaccines preparation need excellent strains with high immunogenicity and adequate supply of bacteria, besides, the product price, immunization efficiency and vaccination method also influence the popularization of such vaccines. In order to improve the immunogenicity and biological characteristics of Mhp strains, the self-replicating plasmids with the origin of replication (oriC) sequences from Mhp could be useful genetic engineering vectors for strain improvement, which had expressed EGFP in Mhp cells successfully (85, 101). On the other hand, genome-scale metabolic engineering techniques provided more information for the optimization of fermentation process, studies had reported that the addition of pyruvate could increase Mhp growth rate in special fermentation experiment (102).
The genetically engineered vaccines are still under development, which mainly includes adhesion factor-based single antigen vaccines, multiple antigens combined vaccines, and multiple disease combined vaccines (5). Feasibility phase of vaccine development depends on the effective antigens selection, expression system, and experimental batch proof of concept, which not only wastes time but also extends the research and development cycle (103). On the other hand, genomic variability and Post-translational protein processing will increase the risk of these vaccine development, and may further reduce the general applicability.
To improve the feasibility of novel vaccine development, the computer-aided vaccinology can provide some new strategies and methods. For example, bacterial pan genome analysis tool (BPGA) can evaluate the genomic diversity and classify the core (conserved), accessory (dispensable), and unique (strain-specific) genes of different strains, which can be used for the choice of conserved and dominant immunogens (104). On the other hand, reverse vaccinology can also provide novel framework for the development of multi-epitope vaccines, which had been used to prevent the infection of bacteria (105), virus (106), and parasite (107).
Mhp is an important respiratory pathogen that can cause MPS, and brings enormous economic losses worldwide. MPS can develop into a recessive disease with the immune defense and other treatments; the isolation of Mhp from other tissues has been reported, which indicates that Mhp can turn into an internal organ and exist within its host without causing disease (27, 108–110). Mhp infection decreases host immunity and induces significant apoptosis or dysfunction to immune cells, as well as increasing the secondary viral or bacterial infection risk.
With the development of modern swine production goes to larger-scale and intensive, coinfections of Mhp and other pathogens are becoming common and serious. Porcine circovirus type 2 (PCV2) infection is immunosuppressive by damaging lymphoid tissues; the coinfection of Mhp and PCV2 contributes to a range of polymicrobial disease syndromes (111). What is more, the coinfection of Mhp and reproductive and respiratory syndrome virus (PRRSV) is generally considered to be universal. The quantitative microbial ecology analysis of microbiota was performed in bronchoalveolar lavage fluid (BALF) from PRRSV-infected pigs, and found the dominant bacterial groups were Haemophilus parasuis and Mycoplasma hyorhinis, which indicated that the pathological importance of Mycoplasma hyorhinis had been underestimated (112). Earlier research pointed out that Mhp infection could increase the lung susceptibility to Pasteurella multocida (113). Particularly, Mhp plays a leading role during the adhesion of Pasteurella multocida type A, which can provide binding sites on the epithelial cell surface (114). In addition, Actinobacillus pleuropneumoniae infection can cause Mhp-infected pigs to show more severe respiratory symptoms (115). In general, coinfection of Mhp and other pathogens aggravates the clinical symptoms and increases the mortality. Therefore, the development of appropriate therapeutic plans and multiple pathogens combined vaccines are crucial in the actual disease treatment.
The main contents of this article are briefly summarized in Figure 3. Genomic variability and Post-translational protein processing complicate the Mhp cell surface antigens and virulence factors appearance, which enhance the immune evasion of Mhp and make the treatment of MPS more difficult. Protein cleavage can produce more functional motifs and make excess epitopes exposed, which can promote disease development and increase the immune burden. Mhp can invade organisms by endotracheal, intranasal or aerosol. In the interaction between Mhp and the host immune system, Mhp can regulate the inflammatory responses by stimulating the release of cytokines. Mhp can also induce immune cells apoptosis by ROS production, excessive NO release and caspase activation, and increases the risk of coinfection with other pathogens. For vaccine development, the self-replicating plasmid system and metabolic engineering techniques can be used for strain improvement, and the computer-aided reverse vaccinology is effective to increase the feasibility of novel vaccine development.

Figure 3. The interaction between Mhp and pig immune system, and new strategies for next-generation vaccine development. Genomic variability and Post-translational protein processing events increase the immune evasion capability of Mhp. Mhp can invade organisms by endotracheal, intranasal, or aerosol. In the interaction between Mhp and the host immune system, Mhp can regulate the inflammatory responses by stimulating the release of cytokines. Mhp can also induce apoptosis to immune cells by ROS production, excessive NO release, and caspase activation. For vaccine development, self-replicating plasmid system and metabolic engineering techniques can be used for strain improvement, and the computer-aided reverse vaccinology is effective to increase the feasibility of novel vaccine development.
Mhp is an important pathogen that leads to MPS and causes major economic losses to pig industry, some commercial vaccines are already available, while the limited immune protective effect is unable to eliminate Mhp from farms or bodies. The balance between immune evasion of the pathogen and immune defense of the host determines the disease development, and based on the current research status, we proposed the genomic variability and Post-translational protein processing are important immune evasion mechanisms of Mhp, but it still needs direct concept proof and deeper research on the mechanisms of immune evasion. In order to reduce the adverse effects of immune evasion on vaccine development and increase the general applicability of vaccines, some novel research methods, such as metabolic engineering techniques and computer-aided reverse vaccinology, can improve the feasibility of novel vaccines as well as reducing the research and development cycle. Vaccine development based on reverse vaccinology has been applied to pathogens with immune evasion ability like COVID-19 (116), while the practicality of these technologies still needs further exploration. On the other hand, Mhp infection can impair host immunity by ROS production, excessive NO release, and caspase activation. In order to strengthen immune responses, certain adjuvants can be used as a component in the vaccine formula. What's more, the use of appropriate pharmacotherapy to improve the host immunity, especially mucosal immunity, is another attractive treatment method (96), which can be regarded as an assistant method for vaccine development. Reducing the adverse effects of immune evasion and enhancing host immunity are critical directions for vaccine development and medicine research and unraveling the immune evasion mechanisms and pathogenesis of Mhp will pave the way for the development of new therapeutics for MPS.
GL and YH conceived the idea. GL wrote the manuscript. EO drew the Figures 2, 3 as well as the grammar and spell checked the manuscript. JinS collected the data and plotted Figure 1. JianS, JC, YW, and YH provided suggestions for the outline and modified the article. All authors read and approved the final version of the manuscript.
This work was funded by Key Research and Development Program of Zhejiang Province [No. 2019C02043]; Public Welfare Technology Application Research Project of Zhejiang Province [No. 2017C32049]; and The Science Foundation of Zhejiang Sci-Tech University [No. 16042062-Y].
JS was employed by Zhejiang Hom-Sun Biosciences Co., Ltd.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We warmly thank all study participants of the Department of Biopharmacy in College of Life Sciences and Medicine, Zhejiang Sci-Tech University, who helped in the realization of this study. In addition, we are grateful to Yu Tao, Rui Yang, Chenyu Wang, and Ke Lu for their valuable suggestions to this manuscript.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2020.510943/full#supplementary-material
1. Ross RF, Young TF. The nature and detection of mycoplasmal immunogens. Vet Microbiol. (1993) 37:369–80. doi: 10.1016/0378-1135(93)90035-6
2. Maes D, Segales J, Meyns T, Sibila M, Pieters M, Haesebrouck F. Control of Mycoplasma hyopneumoniae infections in pigs. Vet Microbiol. (2008) 126:297–309. doi: 10.1016/j.vetmic.2007.09.008
4. Cook BS, Beddow JG, Manso-Silvan L, Maglennon GA, Rycroft AN. Selective medium for culture of Mycoplasma hyopneumoniae. Vet Microbiol. (2016) 195:158–64. doi: 10.1016/j.vetmic.2016.09.022
5. Tao Y, Shu J, Chen J, Wu Y, He Y. A concise review of vaccines against Mycoplasma hyopneumoniae. Res Vet Sci. (2019) 123:144–52. doi: 10.1016/j.rvsc.2019.01.007
6. Tacchi JL, Raymond BB, Haynes PA, Berry IJ, Widjaja M, Bogema DR, et al. Post-translational processing targets functionally diverse proteins in Mycoplasma hyopneumoniae. Open Biol. (2016) 6:150210–27. doi: 10.1098/rsob.150210
7. Niu Z, Xue Q, Wang H, Xie X, Zhu S, Liu W, et al. Mutational biases and GC-biased gene conversion affect GC content in the plastomes of Dendrobium genus. Int J Mol Sci. (2017) 18:E2307. doi: 10.3390/ijms18112307
8. Eyre-Walker A, Hurst LD. The evolution of isochores. Nat Rev Genet. (2001) 2:549–55. doi: 10.1038/35080577
9. Mukhopadhyay P, Basak S, Ghosh TC. Nature of selective constraints on synonymous codon usage of rice differs in GC-poor and GC-rich genes. Gene. (2007) 400:71–81. doi: 10.1016/j.gene.2007.05.027
10. Svard SG, Mattsson JG, Johansson KE, Kirsebom LA. Cloning and characterization of the RNase P RNA genes from two porcine mycoplasmas. Mol Microbiol. (1994) 11:849–59. doi: 10.1111/j.1365-2958.1994.tb00363.x
11. Fritsch TE Siqueira FM Schrank IS. Intrinsic terminators in Mycoplasma hyopneumoniae transcription. BMC Genomics. (2015) 16:273–84. doi: 10.1186/s12864-015-1468-6
12. Razin S, Yogev D, Naot Y. Molecular biology and pathogenicity of mycoplasmas. Microbiol Mol Biol Rev. (1998) 62:1094–156.
13. Charlebois A, Marois-Crehan C, Helie P, Gagnon CA, Gottschalk M, Archambault M. Genetic diversity of Mycoplasma hyopneumoniae isolates of abattoir pigs. Vet Microbiol. (2014) 168:348–56. doi: 10.1016/j.vetmic.2013.11.006
14. Michiels A, Vranckx K, Piepers S, Del Pozo Sacristan R, Arsenakis I, Boyen F, et al. Impact of diversity of Mycoplasma hyopneumoniae strains on lung lesions in slaughter pigs. Vet Res. (2017) 48:2–15. doi: 10.1186/s13567-016-0408-z
15. Liu W, Xiao S, Li M, Guo S, Li S, Luo R, et al. Comparative genomic analyses of Mycoplasma hyopneumoniae pathogenic 168 strain and its high-passaged attenuated strain. BMC Genomics. (2013) 14:80–92. doi: 10.1186/1471-2164-14-80
16. Dos Santos LF, Sreevatsan S, Torremorell M, Moreira MA, Sibila M, Pieters M. Genotype distribution of Mycoplasma hyopneumoniae in swine herds from different geographical regions. Vet Microbiol. (2015) 175:374–81. doi: 10.1016/j.vetmic.2014.11.018
17. Felde O, Kreizinger Z, Sulyok KM, Marton S, Banyai K, Korbuly K, et al. Genotyping Mycoplasma hyopneumoniae isolates based on multi-locus sequence typing, multiple-locus variable-number tandem repeat analysis and analysing gene p146. Vet Microbiol. (2018) 222:85–90. doi: 10.1016/j.vetmic.2018.07.004
18. Rebaque F, Camacho P, Parada J, Lucchesi P, Ambrogi A, Tamiozzo P. Persistence of the same genetic type of Mycoplasma hyopneumoniae in a closed herd for at least two years. Rev Argent Microbiol. (2018) 50:147–50. doi: 10.1016/j.ram.2017.05.002
19. Stakenborg T, Vicca J, Butaye P, Maes D, Peeters J, de Kruif A, et al. The diversity of Mycoplasma hyopneumoniae within and between herds using pulsed-field gel electrophoresis. Vet Microbiol. (2005) 109:29–36. doi: 10.1016/j.vetmic.2005.05.005
20. Mayor D, Zeeh F, Frey J, Kuhnert P. Diversity of Mycoplasma hyopneumoniae in pig farms revealed by direct molecular typing of clinical material. Vet Res. (2007) 38:391–8. doi: 10.1051/vetres:2007006
21. Nathues H, Grosse Beilage E, Kreienbrock L, Rosengarten R, Spergser J. RAPD and VNTR analyses demonstrate genotypic heterogeneity of Mycoplasma hyopneumoniae isolates from pigs housed in a region with high pig density. Vet Microbiol. (2011) 152:338–45. doi: 10.1016/j.vetmic.2011.05.029
22. Tamiozzo P, Lucchesi P, Ambrogi A. Monitoring for Mycoplasma hyopneumoniae before and after a partial depopulation program using a typing scheme based on the polyserine repeat motif of p146. J Swine Health Prod. (2013) 21:309–12. Available online at: https://www.aasv.org/shap/issues/v21n6/v21n6p309.html
23. Betlach AM, Maes D, Garza-Moreno L, Tamiozzo P, Sibila M, Haesebrouck F, et al. Mycoplasma hyopneumoniae variability: current trends and proposed terminology for genomic classification. Transbound Emerg Dis. (2019) 66:1840–54. doi: 10.1111/tbed.13233
24. Minion FC, Adams C, Hsu T. R1 region of P97 mediates adherence of Mycoplasma hyopneumoniae to swine cilia. Infect Immun. (2000) 68:3056–60. doi: 10.1128/iai.68.5.3056-3060.2000
25. Bogema DR, Deutscher AT, Woolley LK, Seymour LM, Raymond BB, Tacchi JL, et al. Characterization of cleavage events in the multifunctional cilium adhesin Mhp684 (P146) reveals a mechanism by which Mycoplasma hyopneumoniae regulates surface topography. MBio. (2012) 3:e00282–11. doi: 10.1128/mBio.00282-11
26. Hooper DC. Mechanisms of action and resistance of older and newer fluoroquinolones. Clin Infect Dis. (2000) 31(Suppl. 2):S24–28. doi: 10.1086/314056
27. Le Carrou J, Laurentie M, Kobisch M, Gautier-Bouchardon AV. Persistence of Mycoplasma hyopneumoniae in experimentally infected pigs after marbofloxacin treatment and detection of mutations in the parC gene. Antimicrob Agents Chemother. (2006) 50:1959–66. doi: 10.1128/AAC.01527-05
28. Vicca J, Maes D, Stakenborg T, Butaye P, Minion F, Peeters J, et al. Resistance mechanism against fluoroquinolones in Mycoplasma hyopneumoniae field isolates. Microb Drug Resist. (2007) 13:166–70. doi: 10.1089/mdr.2007.716
29. Gautier-Bouchardon AV. Antimicrobial resistance in Mycoplasma spp. Microbiol Spectr. (2018) 6:ARBA-0030-2018. doi: 10.1128/microbiolspec.ARBA-0030-2018
30. Vester B, Douthwaite S. Macrolide resistance conferred by base substitutions in 23S rRNA. Antimicrob Agents Chemother. (2001) 45:1–12. doi: 10.1128/aac.45.1.1-12.2001
31. Stakenborg T, Vicca J, Butaye P, Maes D, Minion FC, Peeters J, et al. Characterization of in vivo acquired resistance of Mycoplasma hyopneumoniae to macrolides and lincosamides. Microb Drug Resist. (2005) 11:290–4. doi: 10.1089/mdr.2005.11.290
32. Marshall NC, Finlay BB, Overall CM. Sharpening host defenses during infection: proteases cut to the chase. Mol Cell Proteomics. (2017) 16(4 Suppl 1):S161–71. doi: 10.1074/mcp.O116.066456
33. Djordjevic SP, Cordwell SJ, Djordjevic MA, Wilton J, Minion FC. Proteolytic processing of the Mycoplasma hyopneumoniae cilium adhesin. Infect Immun. (2004) 72:2791–802. doi: 10.1128/iai.72.5.2791-2802.2004
34. Raymond BB, Jenkins C, Seymour LM, Tacchi JL, Widjaja M, Jarocki VM, et al. Proteolytic processing of the cilium adhesin MHJ_0194 (P123J) in Mycoplasma hyopneumoniae generates a functionally diverse array of cleavage fragments that bind multiple host molecules. Cell Microbiol. (2015) 17:425–44. doi: 10.1111/cmi.12377
35. Konovalova A, Sogaard-Andersen L, Kroos L. Regulated proteolysis in bacterial development. FEMS Microbiol Rev. (2014) 38:493–522. doi: 10.1111/1574-6976.12050
36. Bai F, Ni B, Liu M, Feng Z, Xiong Q, Xiao S, et al. Mycoplasma hyopneumoniae-derived lipid-associated membrane proteins induce apoptosis in porcine alveolar macrophage via increasing nitric oxide production, oxidative stress, and caspase-3 activation. Vet Immunol Immunopathol. (2013) 155:155–61. doi: 10.1016/j.vetimm.2013.07.004
37. Liu W, Zhou D, Yuan F, Liu Z, Duan Z, Yang K, et al. Surface proteins mhp390 (P68) contributes to cilium adherence and mediates inflammation and apoptosis in Mycoplasma hyopneumoniae. Microb Pathog. (2019) 126:92–100. doi: 10.1016/j.micpath.2018.10.035
38. Jarocki VM, Raymond BBA, Tacchi JL, Padula MP, Djordjevic SP. Mycoplasma hyopneumoniae surface-associated proteases cleave bradykinin, substance P, neurokinin A and neuropeptide Y. Sci Rep. (2019) 9:14585. doi: 10.1038/s41598-019-51116-w
39. Berry IJ, Jarocki VM, Tacchi JL, Raymond BBA, Widjaja M, Padula MP, et al. N-terminomics identifies widespread endoproteolysis and novel methionine excision in a genome-reduced bacterial pathogen. Sci Rep. (2017) 7:11063–79. doi: 10.1038/s41598-017-11296-9
40. Raymond BB, Tacchi JL, Jarocki VM, Minion FC, Padula MP, Djordjevic SP. P159 from Mycoplasma hyopneumoniae binds porcine cilia and heparin and is cleaved in a manner akin to ectodomain shedding. J Proteome Res. (2013) 12:5891–903. doi: 10.1021/pr400903s
41. Tacchi JL, Raymond BB, Jarocki VM, Berry IJ, Padula MP, Djordjevic SP. Cilium adhesin P216 (MHJ_0493) is a target of ectodomain shedding and aminopeptidase activity on the surface of Mycoplasma hyopneumoniae. J Proteome Res. (2014) 13:2920–30. doi: 10.1021/pr500087c
42. Wilton J, Jenkins C, Cordwell SJ, Falconer L, Minion FC, Oneal DC, et al. Mhp493 (P216) is a proteolytically processed, cilium and heparin binding protein of Mycoplasma hyopneumoniae. Mol Microbiol. (2009) 71:566–82. doi: 10.1111/j.1365-2958.2008.06546.x
43. Deutscher AT, Tacchi JL, Minion FC, Padula MP, Crossett B, Bogema DR, et al. Mycoplasma hyopneumoniae surface proteins Mhp385 and Mhp384 bind host cilia and glycosaminoglycans and are endoproteolytically processed by proteases that recognize different cleavage motifs. J Proteome Res. (2012) 11:1924–36. doi: 10.1021/pr201115v
44. Seymour LM, Deutscher AT, Jenkins C, Kuit TA, Falconer L, Minion FC, et al. A processed multidomain Mycoplasma hyopneumoniae adhesin binds fibronectin, plasminogen, and swine respiratory cilia. J Biol Chem. (2010) 285:33971–8. doi: 10.1074/jbc.M110.104463
45. Bogema DR, Scott NE, Padula MP, Tacchi JL, Raymond BB, Jenkins C, et al. Sequence TTKF↓QE defines the site of proteolytic cleavage in Mhp683 protein, a novel glycosaminoglycan and cilium adhesin of Mycoplasma hyopneumoniae. J Biol Chem. (2011) 286:41217–29. doi: 10.1074/jbc.M111.226084
46. Deutscher AT, Jenkins C, Minion FC, Seymour LM, Padula MP, Dixon NE, et al. Repeat regions R1 and R2 in the P97 paralogue Mhp271 of Mycoplasma hyopneumoniae bind heparin, fibronectin and porcine cilia. Mol Microbiol. (2010) 78:444–58. doi: 10.1111/j.1365-2958.2010.07345.x
47. Robinson MW, Buchtmann KA, Jenkins C, Tacchi JL, Raymond BB, To J, et al. MHJ_0125 is an M42 glutamyl aminopeptidase that moonlights as a multifunctional adhesin on the surface of Mycoplasma hyopneumoniae. Open Biol. (2013) 3:130017. doi: 10.1098/rsob.130017
48. Widjaja M, Harvey KL, Hagemann L, Berry IJ, Jarocki VM, Raymond BBA, et al. Elongation factor Tu is a multifunctional and processed moonlighting protein. Sci Rep. (2017) 7:11227. doi: 10.1038/s41598-017-10644-z
49. Seymour LM, Falconer L, Deutscher AT, Minion FC, Padula MP, Dixon NE, et al. Mhp107 is a member of the multifunctional adhesin family of Mycoplasma hyopneumoniae. J Biol Chem. (2011) 286:10097–104. doi: 10.1074/jbc.M110.208140
50. Matsuoka H, Sisson TH, Nishiuma T, Simon RH. Plasminogen-mediated activation and release of hepatocyte growth factor from extracellular matrix. Am J Respir Cell Mol Biol. (2006) 35:705–13. doi: 10.1165/rcmb.2006-0006OC
51. Degen JL, Bugge TH, Goguen JD. Fibrin and fibrinolysis in infection and host defense. J Thromb Haemost. (2007) 5(Suppl. 1):24–31. doi: 10.1111/j.1538-7836.2007.02519.x
52. Woolley LK, Fell SA, Djordjevic SP, Eamens GJ, Jenkins C. Plasmin activity in the porcine airways is enhanced during experimental infection with Mycoplasma hyopneumoniae, is positively correlated with proinflammatory cytokine levels and is ameliorated by vaccination. Vet Microbiol. (2013) 164:60–6. doi: 10.1016/j.vetmic.2013.02.003
53. Seymour LM, Jenkins C, Deutscher AT, Raymond BB, Padula MP, Tacchi JL, et al. Mhp182 (P102) binds fibronectin and contributes to the recruitment of plasmin(ogen) to the Mycoplasma hyopneumoniae cell surface. Cell Microbiol. (2012) 14:81–94. doi: 10.1111/j.1462-5822.2011.01702.x
54. Jarocki VM, Santos J, Tacchi JL, Raymond BB, Deutscher AT, Jenkins C, et al. MHJ_0461 is a multifunctional leucine aminopeptidase on the surface of Mycoplasma hyopneumoniae. Open Biol. (2015) 5:140175–87. doi: 10.1098/rsob.140175
55. Yu Y, Liu M, Hua L, Qiu M, Zhang W, Wei Y, et al. Fructose-1,6-bisphosphate aldolase encoded by a core gene of Mycoplasma hyopneumoniae contributes to host cell adhesion. Vet Res. (2018) 49:114–26. doi: 10.1186/s13567-018-0610-2
56. Syrovets T, Lunov O, Simmet T. Plasmin as a proinflammatory cell activator. J Leukoc Biol. (2012) 92:509–19. doi: 10.1189/jlb.0212056
57. Arfi Y, Minder L, Di Primo C, Le Roy A, Ebel C, Coquet L, et al. MIB-MIP is a mycoplasma system that captures and cleaves immunoglobulin G. Proc Natl Acad Sci U S A. (2016) 113:5406–11. doi: 10.1073/pnas.1600546113
58. Kiyono H, Izuhara K. New trends in mucosal immunology and allergy. Allergol Int. (2019) 68:1–3. doi: 10.1016/j.alit.2018.12.002
59. Garcia-Morante B, Segales J, Lopez-Soria S, de Rozas AP, Maiti H, Coll T, et al. Induction of mycoplasmal pneumonia in experimentally infected pigs by means of different inoculation routes. Vet Res. (2016) 47:54–63. doi: 10.1186/s13567-016-0340-2
60. Blanchard B, Vena MM, Cavalier A, Le Lannic J, Gouranton J, Kobisch M. Electron microscopic observation of the respiratory tract of SPF piglets inoculated with Mycoplasma hyopneumoniae. Vet Microbiol. (1992) 30:329–41. doi: 10.1016/0378-1135(92)90020-t
61. Pointon AM, Byrt D, Heap P. Effect of enzootic pneumonia of pigs on growth performance. Aust Vet J. (1985) 62:13–8. doi: 10.1111/j.1751-0813.1985.tb06032.x
62. Thacker EL. Diagnosis of Mycoplasma hyopneumoniae. Anim Health Res Rev. (2004) 5:317–20. doi: 10.1079/ahr200491
63. Okada M, Asai T, Ono M, Sakano T, Sato S. Cytological and immunological changes in bronchoalveolar lavage fluid and histological observation of lung lesions in pigs immunized with Mycoplasma hyopneumoniae inactivated vaccine prepared from broth culture supernate. Vaccine. (2000) 18:2825–31. doi: 10.1016/s0264-410x(00)00077-3
64. Damte D, Lee SJ, Hwang MH, Gebru E, Choi MJ, Lee JS, et al. Inflammatory responses to Mycoplasma hyopneumoniae in murine alveolar macrophage cell lines. N Z Vet J. (2011) 59:185–90. doi: 10.1080/00480169.2011.579553
65. Bin L, Luping D, Bing S, Zhengyu Y, Maojun L, Zhixin F, et al. Transcription analysis of the porcine alveolar macrophage response to Mycoplasma hyopneumoniae. PLoS ONE. (2014) 9:e101968. doi: 10.1371/journal.pone.0101968
66. Muneta Y, Uenishi H, Kikuma R, Yoshihara K, Shimoji Y, Yamamoto R, et al. Porcine TLR2 and TLR6: identification and their involvement in Mycoplasma hyopneumoniae infection. J Interferon Cytokine Res. (2003) 23:583–90. doi: 10.1089/107999003322485080
67. Heine H, Ulmer AJ. Recognition of bacterial products by toll-like receptors. Chem Immunol Allergy. (2005) 86:99–119. doi: 10.1159/000086654
68. Li X, Zhang YK, Yin B, Liang JB, Jiang F, Wu WX. Toll-Like Receptor 2 (TLR2) and TLR4 mediate the IgA immune response induced by Mycoplasma hyopneumoniae. Infect Immun. (2019) 88:e00697–19. doi: 10.1128/IAI.00697-19
69. Damte D, Lee SJ, Birhanu BT, Suh JW, Park SC. Sonicated protein fractions of Mycoplasma hyopneumoniae induce inflammatory responses and differential gene expression in a murine alveolar macrophage cell line. J Microbiol Biotechnol. (2015) 25:2153–9. doi: 10.4014/jmb.1506.06049
70. Fang X, Zhao W, Xu J, Tu F, Wang X, Li B, et al. CYP1A1 mediates the suppression of major inflammatory cytokines in pulmonary alveolar macrophage (PAM) cell lines caused by Mycoplasma hyponeumoniae. Dev Comp Immunol. (2016) 65:132–8. doi: 10.1016/j.dci.2016.06.023
71. Grimm S, Baeuerle PA. The inducible transcription factor NF-kappa B: structure-function relationship of its protein subunits. Biochem J. (1993) 290:297–308. doi: 10.1042/bj2900297
72. Su B, Karin M. Mitogen-activated protein kinase cascades and regulation of gene expression. Curr Opin Immunol. (1996) 8:402–11. doi: 10.1016/s0952-7915(96)80131-2
73. Hwang MH, Chang ZQ, Kang EH, Lim JH, Yun HI, Rhee MH, et al. Surfactin C inhibits Mycoplasma hyopneumoniae-induced transcription of proinflammatory cytokines and nitric oxide production in murine RAW 264.7 cells. Biotechnol Lett. (2008) 30:229–33. doi: 10.1007/s10529-007-9552-x
74. Muneta Y, Minagawa Y, Shimoji Y, Ogawa Y, Hikono H, Mori Y. Immune response of gnotobiotic piglets against Mycoplasma hyopneumoniae. J Vet Med Sci. (2008) 70:1065–70. doi: 10.1292/jvms.70.1065
75. Woolley LK, Fell S, Gonsalves JR, Walker MJ, Djordjevic SP, Jenkins C, et al. Evaluation of clinical, histological and immunological changes and qPCR detection of Mycoplasma hyopneumoniae in tissues during the early stages of mycoplasmal pneumonia in pigs after experimental challenge with two field isolates. Vet Microbiol. (2012) 161:186–95. doi: 10.1016/j.vetmic.2012.07.025
76. Obara H, Harasawa R. Nitric oxide causes anoikis through attenuation of E-cadherin and activation of caspase-3 in human gastric carcinoma AZ-521 cells infected with Mycoplasma hyorhinis. J Vet Med Sci. (2010) 72:869–74. doi: 10.1292/jvms.09-0573
77. Dusanic D, Bencina D, Oven I, Cizelj I, Bencina M, Narat M. Mycoplasma synoviae induces upregulation of apoptotic genes, secretion of nitric oxide and appearance of an apoptotic phenotype in infected chicken chondrocytes. Vet Res. (2012) 43:7–20. doi: 10.1186/1297-9716-43-7
78. Li S, Fang L, Liu W, Song T, Zhao F, Zhang R, et al. Quantitative proteomic analyses of a pathogenic strain and its highly passaged attenuated strain of Mycoplasma hyopneumoniae. Biomed Res Int. (2019) 2019:4165735–53. doi: 10.1155/2019/4165735
79. Bizarro CV, Schuck DC. Purine and pyrimidine nucleotide metabolism in mollicutes. Genet Mol Biol. (2007) 30:190–201. doi: 10.1590/S1415-47572007000200005
80. Raymond BBA, Jenkins C, Turnbull L, Whitchurch CB, Djordjevic SP. Extracellular DNA release from the genome-reduced pathogen Mycoplasma hyopneumoniae is essential for biofilm formation on abiotic surfaces. Sci Rep. (2018) 8:10373. doi: 10.1038/s41598-018-28678-2
81. Li P, Zhang Y, Li X, Zhou W, Li X, Jiang F, et al. Mycoplasma hyopneumoniae Mhp597 is a cytotoxicity, inflammation and immunosuppression associated nuclease. Vet Microbiol. (2019) 235:53–62. doi: 10.1016/j.vetmic.2019.05.011
82. Minion FC, Jarvill-Taylor KJ, Billings DE, Tigges E. Membrane-associated nuclease activities in mycoplasmas. J Bacteriol. (1993) 175:7842–7. doi: 10.1128/jb.175.24.7842-7847.1993
83. Jarvill-Taylor KJ, VanDyk C, Minion FC. Cloning of mnuA, a membrane nuclease gene of Mycoplasma pulmonis, and analysis of its expression in Escherichia coli. J Bacteriol. (1999) 181:1853–60.
84. Henthorn CR, Chris Minion F, Sahin O. Utilization of macrophage extracellular trap nucleotides by Mycoplasma hyopneumoniae. Microbiology. (2018) 164:1394–404. doi: 10.1099/mic.0.000717
85. Deeney AS, Maglennon GA, Chapat L, Crussard S, Jolivet E, Rycroft AN. Mycoplasma hyopneumoniae evades phagocytic uptake by porcine alveolar macrophages in vitro. Vet Res. (2019) 50:51–65. doi: 10.1186/s13567-019-0667-6
86. Merad M, Sathe P, Helft J, Miller J, Mortha A. The dendritic cell lineage: ontogeny and function of dendritic cells and their subsets in the steady state and the inflamed setting. Annu Rev Immunol. (2013) 31:563–604. doi: 10.1146/annurev-immunol-020711-074950
87. Bimczok D, Post A, Tschernig T, Rothkotter HJ. Phenotype and distribution of dendritic cells in the porcine small intestinal and tracheal mucosa and their spatial relationship to epithelial cells. Cell Tissue Res. (2006) 325:461–8. doi: 10.1007/s00441-006-0195-3
88. Shen Y, Hu W, Wei Y, Feng Z, Yang Q. Effects of Mycoplasma hyopneumoniae on porcine nasal cavity dendritic cells. Vet Microbiol. (2017) 198:1–8. doi: 10.1016/j.vetmic.2016.11.018
89. Fourour S, Tocqueville V, Paboeuf F, Lediguerher G, Morin N, Kempf I, et al. Pathogenicity study of Mycoplasma hyorhinis and M. flocculare in specific-pathogen-free pigs pre-infected with M. hyopneumoniae. Vet Microbiol. (2019) 232:50–7. doi: 10.1016/j.vetmic.2019.04.010
90. Fourour S, Marois-Crehan C, Martelet L, Fablet C, Kempf I, Gottschalk M, et al. Intra-species and inter-species differences in cytokine production by porcine antigen-presenting cells stimulated by Mycoplasma hyopneumoniae, M. hyorhinis, and M flocculare. Pathogens. (2019) 8:34–45. doi: 10.3390/pathogens8010034
91. Shen Y, Hu W, Wei Y, Feng Z, Yang Q. The immune mechanism of Mycoplasma hyopneumoniae 168 vaccine strain through dendritic cells. BMC Vet Res. (2017) 13:285–91. doi: 10.1186/s12917-017-1194-1
92. Wallgren P, Bolske G, Gustafsson S, Mattsson S, Fossum C. Humoral immune responses to Mycoplasma hyopneumoniae in sows and offspring following an outbreak of mycoplasmosis. Vet Microbiol. (1998) 60:193–205. doi: 10.1016/s0378-1135(98)00155-2
93. Bai F, Ni B, Liu M, Feng Z, Xiong Q, Shao G. Mycoplasma hyopneumoniae-derived lipid-associated membrane proteins induce inflammation and apoptosis in porcine peripheral blood mononuclear cells in vitro. Vet Microbiol. (2015) 175:58–67. doi: 10.1016/j.vetmic.2014.11.013
94. Strugnell RA, Wijburg OL. The role of secretory antibodies in infection immunity. Nat Rev Microbiol. (2010) 8:656–67. doi: 10.1038/nrmicro2384
95. Brandtzaeg P. Secretory IgA: designed for anti-microbial defense. Front Immunol. (2013) 4:222. doi: 10.3389/fimmu.2013.00222
96. Zhang N, Gao P, Yin B, Li J, Wu T, Kuang Y, et al. Cathepsin L promotes secretory IgA response by participating in antigen presentation pathways during Mycoplasma hyopneumoniae infection. PLoS ONE. (2019) 14:e0215408. doi: 10.1371/journal.pone.0215408
97. Crawley A, Wilkie BN. Porcine Ig isotypes: function and molecular characteristics. Vaccine. (2003) 21:2911–22. doi: 10.1016/s0264-410x(03)00142-7
98. Raymond CR, Wilkie BN. Th-1/Th-2 type cytokine profiles of pig T-cells cultured with antigen-treated monocyte-derived dendritic cells. Vaccine. (2004) 22:1016–23. doi: 10.1016/j.vaccine.2003.08.026
99. Garcia-Morante B, Segales J, Fraile L, Llarden G, Coll T, Sibila M. Potential use of local and systemic humoral immune response parameters to forecast Mycoplasma hyopneumoniae associated lung lesions. PLoS ONE. (2017) 12:e0175034. doi: 10.1371/journal.pone.0175034
100. Raymond BBA, Turnbull L, Jenkins C, Madhkoor R, Schleicher I, Uphoff CC, et al. Mycoplasma hyopneumoniae resides intracellularly within porcine epithelial cells. Sci Rep. (2018) 8:17697. doi: 10.1038/s41598-018-36054-3
101. Maglennon GA, Cook BS, Matthews D, Deeney AS, Bosse JT, Langford PR, et al. Development of a self-replicating plasmid system for Mycoplasma hyopneumoniae. Vet Res. (2013) 44:63–72. doi: 10.1186/1297-9716-44-63
102. Kamminga T, Slagman SJ, Bijlsma JJE, Martins Dos Santos VAP, Suarez-Diez M, Schaap PJ. Metabolic modeling of energy balances in Mycoplasma hyopneumoniae shows that pyruvate addition increases growth rate. Biotechnol Bioeng. (2017) 114:2339–47. doi: 10.1002/bit.26347
103. Kamminga T, Slagman SJ, Martins Dos Santos VAP, Bijlsma JJE, Schaap PJ. Risk-based bioengineering strategies for reliable bacterial vaccine production. Trends Biotechnol. (2019) 37:805–16. doi: 10.1016/j.tibtech.2019.03.005
104. Chaudhari NM, Gupta VK, Dutta C. BPGA- an ultra-fast pan-genome analysis pipeline. Sci Rep. (2016) 6:24373. doi: 10.1038/srep24373
105. Dar HA, Zaheer T, Shehroz M, Ullah N, Naz K, Muhammad SA, et al. Immunoinformatics-aided design and evaluation of a potential multi-epitope vaccine against Klebsiella Pneumoniae. Vaccines. (2019) 7:88–105. doi: 10.3390/vaccines7030088
106. Khan A, Junaid M, Kaushik AC, Ali A, Ali SS, Mehmood A, et al. Computational identification, characterization and validation of potential antigenic peptide vaccines from hrHPVs E6 proteins using immunoinformatics and computational systems biology approaches. PLoS ONE. (2018) 13:e0196484. doi: 10.1371/journal.pone.0196484
107. Dhal AK, Pani A, Mahapatra RK, Yun SI. An immunoinformatics approach for design and validation of multi-subunit vaccine against Cryptosporidium parvum. Immunobiology. (2019) 224:747–57. doi: 10.1016/j.imbio.2019.09.001
108. Marois C, Le Carrou J, Kobisch M, Gautier-Bouchardon AV. Isolation of Mycoplasma hyopneumoniae from different sampling sites in experimentally infected and contact SPF piglets. Vet Microbiol. (2007) 120:96–104. doi: 10.1016/j.vetmic.2006.10.015
109. Marchioro SB, Maes D, Flahou B, Pasmans F, Del Pozo Sacristan R, Vranckx K, et al. Local and systemic immune responses in pigs intramuscularly injected with an inactivated Mycoplasma hyopneumoniae vaccine. Vaccine. (2013) 31:1305–11. doi: 10.1016/j.vaccine.2012.12.068
110. Vilalta C, Sanhueza JM, Murray D, Johnson L, Pieters M. Detection of Mycoplasma hyopneumoniae in piglet processing fluids. Vet Rec. (2019) 185:510–4. doi: 10.1136/vr.105475
111. Opriessnig T, Castro A, Karuppanan AK, Gauger PC, Halbur PG, Matzinger SR, et al. A porcine circovirus type 2b (PCV2b)-based experimental vaccine is effective in the PCV2b-Mycoplasma hyopneumoniae coinfection pig model. Vaccine. (2019) 37:6688–95. doi: 10.1016/j.vaccine.2019.09.029
112. Jiang N, Liu H, Wang P, Huang J, Han H, Wang Q. Illumina MiSeq sequencing investigation of microbiota in bronchoalveolar lavage fluid and cecum of the swine infected with PRRSV. Curr Microbiol. (2019) 76:222–30. doi: 10.1007/s00284-018-1613-y
113. Amass SF, Clark LK, van Alstine WG, Bowersock TL, Murphy DA, Knox KE, et al. Interaction of Mycoplasma hyopneumoniae and Pasteurella multocida infections in swine. J Am Vet Med Assoc. (1994) 204:102–7.
114. Park C, Jeong J, Kang I, Choi K, Park SJ, Chae C. Increased fucosyl glycoconjugate by Mycoplasma hyopneumoniae enhances adherences of Pasteurella multocida type A in the ciliated epithelial cells of the respiratory tract. BMC Vet Res. (2016) 12:25–30. doi: 10.1186/s12917-016-0650-7
115. Marois C, Gottschalk M, Morvan H, Fablet C, Madec F, Kobisch M. Experimental infection of SPF pigs with Actinobacillus pleuropneumoniae serotype 9 alone or in association with Mycoplasma hyopneumoniae. Vet Microbiol. (2009) 135:283–91. doi: 10.1016/j.vetmic.2008.09.061
Keywords: Mycoplasma hyopneumoniae, genomic variability, immune evasion, immune interaction, post-translational protein processing
Citation: Li G, Obeng E, Shu J, Shu J, Chen J, Wu Y and He Y (2020) Genomic Variability and Post-translational Protein Processing Enhance the Immune Evasion of Mycoplasma hyopneumoniae and Its Interaction With the Porcine Immune System. Front. Immunol. 11:510943. doi: 10.3389/fimmu.2020.510943
Received: 14 November 2019; Accepted: 27 August 2020;
Published: 07 October 2020.
Edited by:
Laurel J. Gershwin, University of California, Davis, United StatesReviewed by:
Sabine Hammer, University of Veterinary Medicine Vienna, AustriaCopyright © 2020 Li, Obeng, Shu, Shu, Chen, Wu and He. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yuehong Wu, d3V5dWVob25nMjAwM0AxNjMuY29t; Yulong He, aGV5dWxvbmcyMDAzQDE2My5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.