
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Immunol. , 07 July 2020
Sec. Microbial Immunology
Volume 11 - 2020 | https://doi.org/10.3389/fimmu.2020.01386
This article is part of the Research Topic The Role of Gene Polymorphism in Modulating the Immune Responses against Tropical Infectious Diseases View all 14 articles
 Amanda Farage Frade-Barros1,2,3,4,5*
Amanda Farage Frade-Barros1,2,3,4,5* Barbara Maria Ianni1
Barbara Maria Ianni1 Sandrine Cabantous3
Sandrine Cabantous3 Cristina Wide Pissetti6Bruno Saba7Hui Tzu Lin-Wang7Paula Buck1José Antonio Marin-Neto8André Schmidt8
Cristina Wide Pissetti6Bruno Saba7Hui Tzu Lin-Wang7Paula Buck1José Antonio Marin-Neto8André Schmidt8 Fabrício Dias8
Fabrício Dias8 Mario Hiroyuki Hirata9
Mario Hiroyuki Hirata9 Marcelo Sampaio7Abílio Fragata7
Marcelo Sampaio7Abílio Fragata7 Alexandre Costa Pereira1
Alexandre Costa Pereira1 Eduardo Donadi8
Eduardo Donadi8 Virmondes Rodrigues6
Virmondes Rodrigues6 Jorge Kalil1,2,5
Jorge Kalil1,2,5 Christophe Chevillard10†
Christophe Chevillard10† Edecio Cunha-Neto1,2,4†
Edecio Cunha-Neto1,2,4†Background: Chagas disease, caused by the protozoan Trypanosoma cruzi, is endemic in Latin America. Thirty percent of infected individuals develop chronic Chagas cardiomyopathy (CCC), an inflammatory dilated cardiomyopathy that is the most important clinical consequence of T. cruzi infection, while the others remain asymptomatic (ASY). IFN-γ and IFN-γ-producing Th1-type T cells are increased in peripheral blood and CCC myocardium as compared to ASY patients, while the Th1-antagonizing cytokine IL-10 is more expressed in ASY patients. Importantly IFN-γ-producing Th1-type T cells are the most frequent cytokine-producing T cell subset in CCC myocardium, while expression of Th1-antagonizing cytokines IL-10 and IL-4 is unaltered. The control of IFN-γ production by Th1-type T cells may be a key event for progression toward CCC. A genetic component to disease progression was suggested by the familial aggregation of cases and the association of gene polymorphisms with CCC development. We here investigate the role of gene polymorphisms (SNPs) in several genes involved in the control of IFN-γ production and Th1 T cell differentiation in CCC development.
Methods: We studied a Brazilian population including 315 CCC cases and 118 ASY subjects. We assessed 35 Tag SNPs designed to represent all the genetic information contained in the IL12B, IL10, IFNG, and IL4 genes.
Results: We found 2 IL12 SNPs (rs2546893, rs919766) and a trend of association for a IL10 SNP (rs3024496) to be significantly associated with the ASY group. these associations were confirmed by multivariate analysis and allele tests. The rs919766C, 12rs2546893G, and rs3024496C alleles were associated to an increase risk to CCC development.
Conclusions: Our data show that novel polymorphisms affecting IL12B and IL10, but not IFNG or IL4 genes play a role in genetic susceptibility to CCC development. This might indicate that the increased Th1 differentiation and IFN-γ production associated with CCC is genetically controlled.
Chagas disease (American trypanosomiasis) is caused by the protozoan Trypanosoma cruzi (T. cruzi) and mainly transmitted by the reduviid arthropod vector. It is endemic in Latin America, where an estimated 8 million people are infected (https://www.cdc.gov/parasites/chagas/index.html). Imported disease is increasingly recognized as an emerging problem in the USA and Europe due to immigration from Latin America (https://www.who.int/chagas/disease/en/). Decades after acute infection, 20–30% of the patients develop chronic Chagas disease cardiomyopathy (CCC), where heart tissue inflammation and remodeling, fibrosis, electrical and structural abnormalities often lead to life-threatening heart failure and arrhythmia (1). Five to 10% of infected individuals develop digestive disease, with denervation and dilation of the digestive tract. The remaining two-thirds of infected individuals remain asymptomatic (ASY) and free from symptomatic disease for life (2). Heart failure due to CCC has a worse prognosis, with 50% shorter survival when compared to like ischemic, idiopathic and hypertensive cardiomyopathies (3, 4). The pathogenic determinants of differential disease progression have not yet been completely elucidated.
The finding that 30% of Chagas disease patients develop CCC suggested the participation of genetics in differential disease progression. In addition, familial aggregation of CCC cases has been described (5), and a significant number of genetic polymorphisms has been associated with CCC development. A recent literature search found ca. 150 case-control genetic association studies addressing polymorphisms in 76 genes, which disclosed 62 SNPs from 44 genes to be associated with CCC, mostly related to immunology and inflammation. From those, a much smaller number of SNPs were associated with CCC severity (ventricular dysfunction) (6–8).
Many of these SNPs are related to pathogen resistance and disease tolerance genes—those related to direct control of parasitism or to reduction of tissue damage, respectively (7). Chagas disease is associated with increased production of inflammatory cytokines; IFN-γ is a major pathogen resistance gene and plays a major role in the disease; T. cruzi infection of mice genetically deficient of IFNG leads to uncontrolled parasitism and 100% mortality (9, 10). While it is a key player in pathogen protection, excessive levels lead to increased inflammation and tissue damage during the acute phase as well as CCC (7). During acute infection, T. cruzi pathogen-associated molecular patterns (PAMPs) trigger innate immunity (3, 11, 12) leading to the release of proinflammatory cytokines and chemokines, including IL-12 and IL-18, the major drivers of differentiation of IFN-γ-producing T. cruzi–specific Th1 T cells which migrate to sites of T. cruzi-induced inflammation (13, 14). IFNγ is then produced at high levels, inducing production of microbicidal and potentially tissue-damaging reactive oxygen and nitrogen species that kill intracellular T. cruzi. This leads to the control—but not complete elimination—of parasitism, establishing a low-grade chronic persistent infection by T. cruzi. On the other hand, cytokines that are able to modulate excessive IFN-γ production/Th1 T cell differentiation can modulate tissue damage in the acute phase of infection. Mice genetically deficient in disease tolerance genes IL10 (15), IL4 (16) and IL27b/Ebi3 (17), factors known to limit Th1 T cell differentiation and IFNγ production (17–20) display increased inflammation and may die early. Animals who keep a balance between inflammatory Th1 and anti-inflammatory cytokines may survive and progress to the chronic phase. As a result of persistent infection, both CCC and ASY chronic Chagas disease patients produce higher levels of IFNγ than seronegative controls (21, 22), but those who develop Chagas cardiomyopathy display a particularly strong Th1-type immune response with increased numbers of IFN-γ-producing Th1 cells in peripheral blood mononuclear cells (PBMC) (19, 20, 22, 23) and decreased IFNγ -modulating factors IL-10, IL-27b/Ebi3, and Treg cells (17–20). Accordingly, CCC patients displayed increased plasma levels of IFNγ while levels of IL-10 are reduced when compared to ASY individuals (24–26). These data suggest that the cytokine balance can modulate progression to CCC development. Indeed, CCC hearts display a prominent IFNγ -producing T-cell dependent myocarditis. IFNγ is the most highly expressed cytokine in CCC myocardium (27), and mononuclear cells producing this cytokine are abundant in CCC heart (12, 14, 22). mRNA expression IL-10 and other Th1-antagonizing cytokines and cell types (Treg and Th2) was low or undetectable in CCC heart tissue. This indicates that the Th1 infiltrate in CCC myocardium is essentially unopposed by regulatory cells or cytokines, and subject to little regulation (27). The failure to regulate Th1 responses and IFNγ production may thus be one key immune defect of patients who progress to CCC. This creates a paradox, in that IFNγ is essential for control of parasitism in acute and chronic Chagas disease, but at the same time causes tissue damage (7).
Cytokine gene polymorphisms may contribute to the control of Th1 T cell differentiation/IFN-γ production. Previous studies have assessed the association of SNPs in cytokines important for regulation of IFN-γ responses with CCC. For IFNG, no SNP was found to be associated with CCC when compared to ASY after comparison for multiple testing (28). Likewise, IL4 SNP case-control studies between CCC and ASY failed to show associations, but may have been underpowered, with only110 and 260 total Chagasic patients, respectively (29, 30). A promoter polymorphism in IL12B +1188 (rs3212227) was found to be associated with CCC in an Colombian population (31). The SNP IL10-1082 (rs1800896) was shown to be functional (32) and was associated to CCC in a Brazilian population comparing ASY individuals vs. CCC (33). On a Brazilian population, the frequency of the polymorphic rs1800896A allele (associated with lower expression of IL-10) was higher in the asymptomatic group than in the cardiac group. On the other hand, Florez et al. compared CCC (n = 130) to ASY (n = 130) cases in a Colombian population. Florez et al. failed to show associations in the genotypes and allele frequencies neither in IL10 SNPs IL10-1082 (rs1800896) nor in the additional SNPs IL10-819 (rs1800871) and IL10-592 (rs1800872). It is possible that this lack of association in our study is was due to a small sample size or to epistasis between IL-10 and MHC.
Since the previous studies had some limitations, i.e., focused on selected polymorphisms, failed to be replicated in other populations, or lacked appropriate statistical power, we here comprehensively assessed the role of polymorphisms in these genes on CCC progression. For that matter we here performed a thorough genetic study focusing on 35 polymorphisms of IL4, IL12B, IL10, and IFNG using the Tag SNP approach, which represent all the genetic information contained in the mentioned genes, in a larger cohort (n = 433) of Brazilian Chagas disease patients, including CCC patients with or without ventricular dysfunction as well as asymptomatic patients. With the tag SNP approach, we could both replicate the study of previously investigated SNPs as well as novel polymorphisms. This way, we could perform a more sensitive assessment of the contribution of genetic variants in prognosis to CCC either confirming or finding additional associated SNPs in the mentioned genes. In addition, our experimental design allowed the study of possible interactions between polymorphisms in different cytokines.
Written informed consent was obtained from all the patients, in accordance with the guidelines of the various internal review boards of all the involved institutions. The protocol was also approved by the INSERM Internal Review Board and the Brazilian National Ethics in Research Commission (CONEP). All the patients enrolled in this study were over 21 years old. Investigations were conformed to the principles outlined in the declaration of Helsinki.
The diagnostic criteria for Chagas disease included the detection of antibodies against T. cruzi in at least two of three independent serological tests (EIA [Hemobio Chagas; Embrabio São Paulo], indirect immunofluorescence assays [IFA-immunocruzi; Biolab Merieux], and indirect hemagglutination tests [Biolab Merieux]) (34). All Chagas disease patients underwent standard electrocardiography and echocardiography. Echocardiography was performed at the hospital, with a Sequoia model 512 echocardiograph with a broad-band transducer. Left ventricular dimensions and regional and global function, including the recording of left ventricular ejection fraction (LVEF), were evaluated with a two-dimensional, M-mode approach, in accordance with the recommendations of the American Society of Echocardiography. ASY subjects had no electrocardiography and echocardiography changes. CCC patients presented typical conduction abnormalities (right bundle branch block and/or left anterior division hemiblock) (35). CCC patients with significant left ventricular systolic dysfunction (LVEF <40%) were classified as having severe CCC, whereas those with no significant ventricular dysfunction (LVEF ≥ 40%) were classified as having moderate CCC. We selected 40% as arbitrary cutoff value that has been previously used to define significant ventricular dysfunction by our group and others (36–38).
The CCC and ASY patients were born and raised in rural areas of Sao Paulo, Minas Gerais and Bahia states and enrolled in one of the study centers (Incor, FMUSP, FMRP, UFTM, IDPC). Patients with digestive forms were excluded of this study. Patients were classified as ASY (n = 118) or as having CCC (n = 315). ASY individuals were used as the control subjects for this study because they were from the same areas of endemicity as the patients with CCC, had encountered the parasite and had tested seropositive for T. cruzi infection, but the infection had not progressed to CCC. Here, the Case/Control ratio, which is 0.375, illustrate the difficulty to enroll real ASY subjects. For our association study, if we are fixing the significant level to 0.05, the disease prevalence to 30%, the disease allele Frequency to 0.2 and the genotype relative risk to 1.5, the expected power of our association study reaches 0.895.
Of 315 patients with CCC, 106 (42 men [39.6%] and 64 women [60.4%]) showed no significant ventricular dysfunction (LVEF ≥ 40%) and were thus classified as having moderate CCC, whereas 199 (144 men [72.4%] and 55 women [27.6%]) had severe ventricular dysfunction (LVEF <40%) and were classified as having severe CCC. Data for left ventricular ejection fraction were missing for 10 patients with CCC. So, when we compared moderate patients to severe patients, these 10 individuals were excluded from the analysis. Regarding progression of the ASY cases to CCC, the yearly progression rate –regardless of age group- is ca. 1–2%/year. The average age of Subjects with asymptomatic form was above 55 years. Taking into account that they were all born in endemic areas before vector transmission was interrupted, it is likely that in most if not all cases vector-borne infection occurred in early childhood. The odds that a significant number of such mature patients convert to CCC, and that this thwarts our statistical calculation is rather low; however, this is a pitfall of all cross sectional studies on diseases that display progression.
For each subject, 5–15 ml of blood were collected in EDTA tubes. Genomic DNA was isolated on a silica-membrane according to the manufacturer's protocol (QIAamp DNA Blood Max Kit, Qiagen, Hilden, Germany).
Tag single nucleotide polymorphisms (SNPs) were selected on the basis on Ensembl Release GRCh38 for the Caucasian and Yoruba reference populations. Tag SNPs were selected within a region extending 5 kb on either side of the candidate gene. The minor allele frequency (MAF) cut off value was arbitrarily set at 20% (so the markers characterized by a MAF <20% were excluded from the analysis by lack of power). In each reference population, the markers with MAF > 20% are included in different blocks of correlation (based on the r2 values). For each correlation block in the gene, one marker was selected and considered as a Tag SNP. Indeed, markers located in the same block of correlation yielded the same genetic information in association studies. Tag SNPs characterized by a MAF over 20% on at least one reference population were selected. These Tag SNPs were thus defined to gather all the genetic information from each candidate gene. we selected a total of 35 Tag SNPs: sixteen tag SNPs for IL12B, six tag SNPs for IL10, six for IFNG and seven tag SNPs for IL4.
Most of the genotyping was done with the Golden Gate genotyping assay (Illumina, San Diego, USA). In some cases, genotyping assays were performed with the TaqMan system (Applied Biosystems, Foster City, USA) according to the manufacturer's instructions. Genotyes are available on request.
SPSS Statistics software v. 17.0 (IBM, Armonk, USA) was used for statistical analyses. We performed stepwise binary logistic regression analysis on the whole population, to analyse the relationship between the probability of an individual to develop chronic Chagas cardiomyopathy and the main covariates (sex and polymorphisms). Sex was considered as a binary covariate. In our stepwise binary logistic regression analysis, genotypes were considered as binary covariates. Indeed, for each polymorphism we had two alleles (A frequent one; a rare one). So, we obtained three genotypes (AA, Aa, and aa). In our stepwise binary logistic regression analysis, genotypes were considered as binary covariates. So, we performed three different analyses (Analysis 1: AA vs. Aa + aa (we supposed that the a allele is dominant); Analysis 2: AA + aa vs. Aa (we supposed that the heterozygote carriers are different from the homozygote ones); Analysis 3: AA + Aa vs. aa (we supposed that the A allele is dominant).
In multivariate analyses, several polymorphisms and gender were included as covariates. All the covariates are analyzed in the same time. In a stepwise approach, the worse associated covariate (non-significant) is removed and the analysis is run again up to keep only significant associated covariates.
An investigation of a candidate gene should test many SNPs individually for association. However, such multiple testing will increase the false-positive (type I error) rate under nominal significance thresholds (α = 0.05). So, we applied the Holm-Bonferroni sequential correction for multiple testing (39).
We conducted a study focusing on the IL12B, IL10, IFNG, and IL4 genes. 35 Tag SNPs were selected (Supplementary Table 1). 97.8% markers were genotyped successfully on our cohort including ASY subjects (n = 118) and CCC patients (n = 315). The genotype distribution of each SNP is summarized in Supplementary Table 2. All the SNPs were in Hardy-Weinberg equilibrium on the ASY individuals considered as control subjects. Of the 118 ASY subjects, 45.3% were male, whereas in the CCC patients group, this percentage reaches 61.3%. The difference in sex distribution between the groups was significant (p = 1.21E-4; OR = 2.126; 95%CI: 1.450–3.12). It is well-known that male patients infected with T. cruzi have a higher risk of progression to CCC than female patients. Of 315 patients with CCC, 106 (42 men [39.6%] and 64 women [60.4%]) showed no significant ventricular dysfunction and were thus classified as having moderate CCC, whereas 199 (144 men [72.4%] and 55 women [27.6%]) had severe ventricular dysfunction and were classified as having severe CCC.
Fourteen on sixteen tag SNPs for the IL12B gene were genotyped successfully. IL-12B genotype data are shown in Tables 1, 2. All these markers are located in intronic region of IL12B gene. In the CCC subjects group, 269 (86.8%) subjects carried the rs2546893G/G or G/A genotypes whereas 79 (73.8%) of the ASY carried these genotypes (p = 0.002; OR = 1.548; 95%CI: 1.176–2.037 (see Table 1). These data suggest that rs2546893G/G or G/A genotypes are associated to an increased susceptibility to CCC.
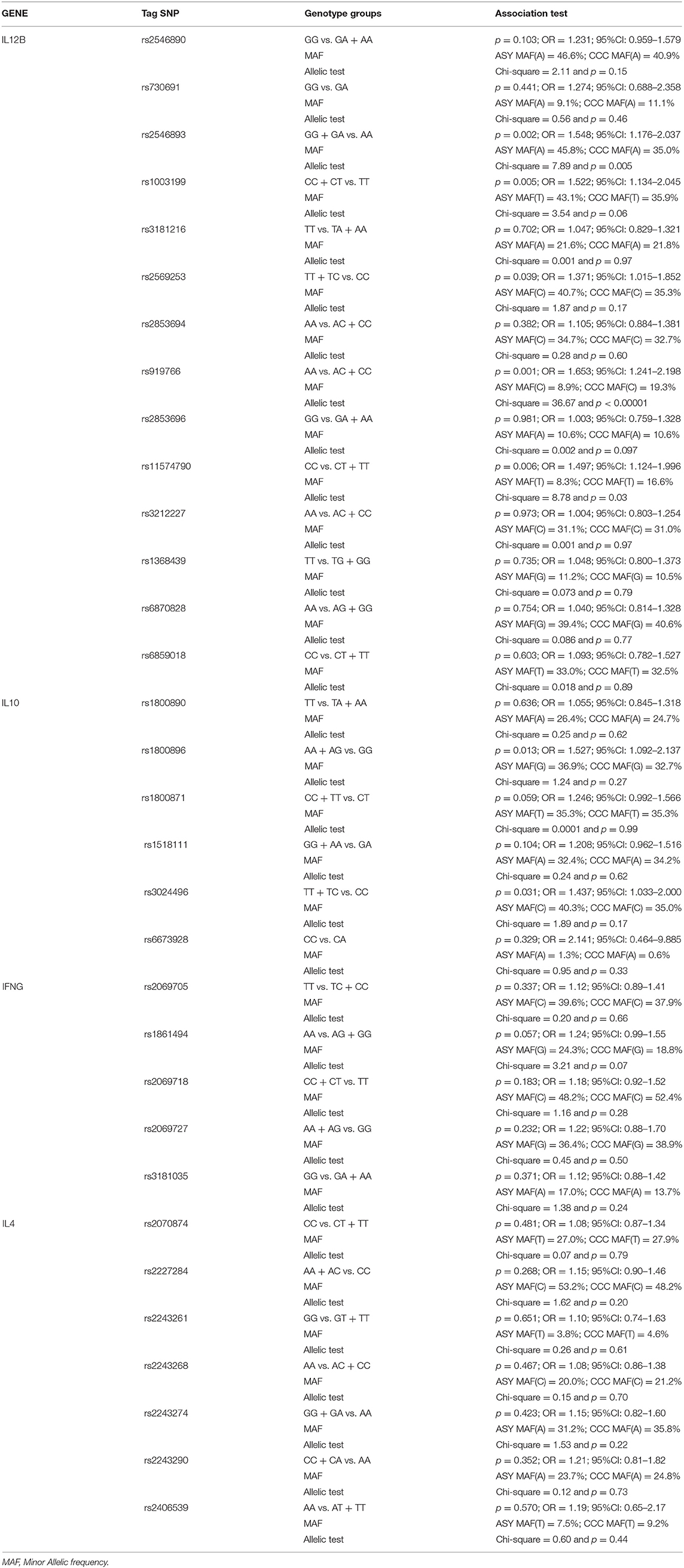
Table 1. Association studies between CCC and ASY including as covariates the gender and the polymorphisms (genotype associations and allelic tests).
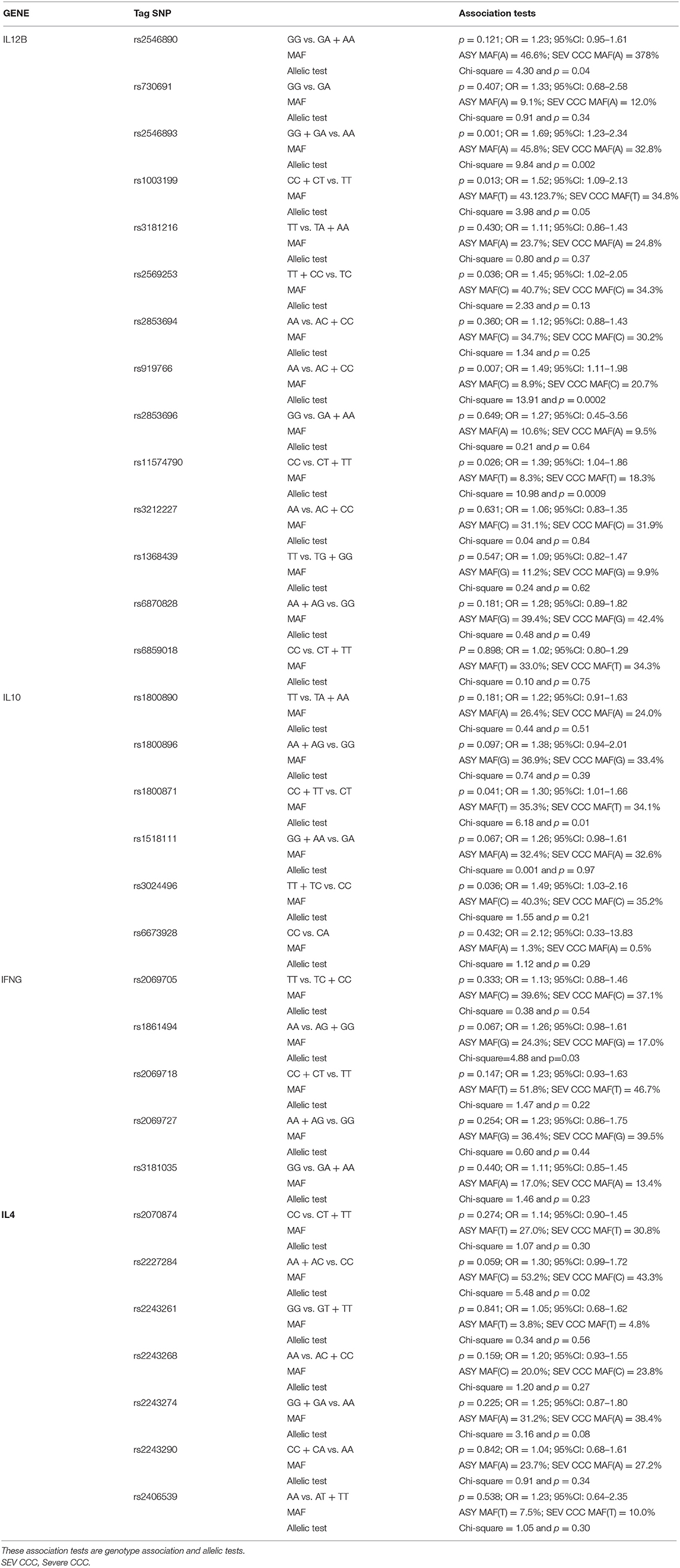
Table 2. Association studies between CCC with a left ventricular ejection fraction value under 0.4% and ASY including as covariates the gender and the polymorphism one by one.
For the rs919766A/C polymorphism, 107 (34.7%) CCC subjects carried the C/C or A/C genotypes vs. 17 (15.9%) for the ASY group (p = 0.001; OR = 1.653; 95%CI: 1.241–2.198). For the rs1003199C/T polymorphism, 276 (88.7%) CCC subjects carried the C/C or C/T genotypes vs. 285 (78.7%) for the ASY group. This difference was significant (p = 0.005; OR = 1.522; 95%CI: 1.134–2.045). For the rs11574790C/T polymorphism, 95 (30.5%) CCC subjects carried the C/T or T/T genotypes vs. 17 (15.7%) for the ASY group (p = 0.006; OR = 1.497; 95%CI: 1.124–1.996). For the rs2569253T/C polymorphism, 256 (87.46%) CCC subjects carried the T/T or T/C genotypes vs. 81 (79.4%) for the ASY (p = 0.039; OR = 1.371; 95%CI: 1.015–1.852). After performing a correction for multiple testing using the Bonferroni methods, the two main polymorphisms (rs919766 and rs25466893) remain significant (corrected p-values were 0.014 and 0.026, respectively).
In a multivariate analysis, including these five polymorphisms and gender as covariates, the gender (p = 0.004; OR = 2.036; 95% CI: 1.249–3.318) and two markers (rs2546893: p = 0.005; OR = 1.536; 95% CI: 1.139–2.070; rs919766: p = 0.014; OR = 1.477; 95% CI: 1.081–2.020) remained significantly associated. The three other markers were excluded from the best model. Allelic tests confirmed the association of these two IL12 markers. The rs919766C allele (p < 0.00001) and the rs2546893G allele (p = 0.005) were associated to an increase risk to CCC development.
The same analysis was performed between the ASY subjects and severe CCC patients. The same polymorphisms were associated, in univariate analysis, to an increased risk to develop severe chronic cardiomyopathy (rs2546893G/A: p = 0.001; OR = 1.706; 95%CI: 1.236–2.353; rs919766A/C: p = 0.001; OR = 1.672; 95%CI: 1.231–2.267; rs1003199C/T: p = 0.013; OR = 1.531; 95%CI: 1.236–2.353; rs2569253T/C: p = 0.037; OR = 1.445; 95%CI: 1.022–2.041; rs11574790C/T: p = 0.004; OR = 1.562; 95%CI: 1.151–2.123) (see Table 2). After correction for multiple testing the polymorphism rs2546893 remains significant (corrected p-value = 0.014) and the polymorphism rs919766 is borderline (corrected p-value = 0.091). In a multivariate analysis, the gender and the two polymorphisms remained significantly associated (rs2546893G/A: p = 0.005; OR = 1.667; 95% CI: 1.170–2.375; rs919766A/C: p = 0.024; OR = 1.471; 95%CI: 1.051–2.058; gender: p = 2.25 × 10−5; OR = 3.261; 95% CI: 1.888–5.632). the allelic tests confirmed that the rs2546893G allele (p < 0.002) and the rs919766C allele (p = 0.0002) were associated to an increase risk to severe CCC. None of the studied IL12B markers discriminated moderate from severe CCC patient groups.
Six tag SNPs were genotyped for the IL10 gene. Table 1 shows comparison between CCC and ASY, while Table 2 shows comparison of severe CCC and ASY. For the rs1800896A/G polymorphism, 282 (91.9%) CCC subjects carried the A/A or A/G genotypes vs. 90 (84.1%) for the ASY (p = 0.013; OR = 1.527; 95%CI: 1.092–2.137). The same result was obtained when the analysis was restricted to severe CCC vs. ASY (p = 0.028; OR = 1.536; 95%CI: 1.047–2.252) (see Table 2). For the rs3024496T/C polymorphism, 284 (91.3%) CCC subjects carried the T/T or T/C genotypes vs. 92 (84.3%) for the ASY (p = 0.031; OR = 1.437; 95%CI: 1.033–2.000) (see Table 1). The same result was obtained when the analysis was restricted to severe CCC vs. ASY (p = 0.049; OR = 1.462; 95%CI: 1.002–2.132) (see Table 2). For the rs1800871 marker, we found a significant association when severe CCC patients were compared the ASY subjects (p = 0.035; OR = 1.311; 95%CI: 1.019–1.686). No association was found for this last marker in the comparison between ASY and the whole CCC group (see Table 1). After correction for multiple testing, the polymorphism rs1800896A/G and rs3024496T/C has shown only trends of association (rs1800896A/G: corrected p-value = 0.078). The allelic tests reach the same trends of association (rs3024496C p = 0.17 and rs1800896G p = 0.27).
Based on the results of the multivariate analysis, we had an association between CCC and ASY with two SNPs located in the IL12B gene (rs2546893 and rs919766) and one polymorphism in the IL10 gene (rs3024496). Subjects carrying the three susceptibility genotypes [rs2546893(GG or GA) + rs919766(CC or AC) + rs3024496(TT or TC)] have a higher risk (OR = 2.68) to develop CCC. In contrast, subjects carrying the resistant genotypes (rs2546893AA + r919766AA + rs3024496CC) have a lower risk [OR = 4.48 (1/0.223)] to develop CCC (Table 3).
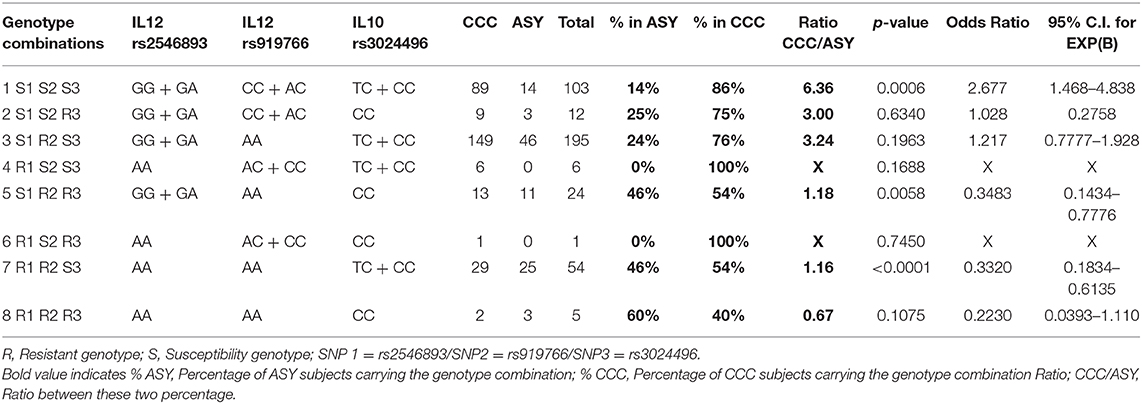
Table 3. Genotype combination analysis based on the following polymorphisms (rs2546893/rs919766/rs3024496).
Six tag SNPs were genotyped for the IFNG gene. For the rs1861494 A/G polymorphism, a trend of association was detected. Indeed, 202 (64.7%) CCC subjects carried the A/A genotype vs. 62 (54.9%) for the ASY (p = 0.057; OR = 1.24; 95%CI: 0.99–1.55) (see Supplementary Table 2). The same trend appeared on severe cases (p = 0.067; OR = 1.26; 95%CI: 0.98–1.61) (see Table 2). Seven tag SNPs into the IL4 gene were genotyped on our whole study population (see Supplementary Table 2). Comparisons between ASY and CCC patients disclosed no significant associations (see Tables 1, 2).
Association studies of cytokine gene polymorphisms in Chagas disease frequently include a small number of patients, and are often not replicated in different studies or ethnical groups. The present study was designed to determine whether SNPs in IL12B, IL10, IFNG and IL4, key genes involved in the promotion and control of Th1 differentiation and IFN-g production to T. cruzi, are associated with Chagas disease outcome. For that, we used a larger population and performed a comprehensive assessment of 35 Tag SNPs covering all the genetic information of the mentioned genes. We found significant associations of genotypes and alleles of IL12B and IL10 with increased risk to CCC progression.
For the IL12B gene region, in a multivariate analysis, two SNPs were significantly associated to disease (rs2546893G/A, rs919766A/C). The rs919766C and rs2546893G alleles were associated to an increase risk to CCC development. Interestingly, these two markers are located 5 to 13 kb away from the rs3212227 polymorphism (IL12B 3′UTR region), that was previously associated to Chagas cardiomyopathy (31). Our results are consistent as these 3 markers are in linkage disequilibrium (D′ = 0.99) on the African (Yoruba) reference population and the markers rs919766 and rs3212227 are in strong linkage disequilibrium (D′ = 0.99) on the European reference population.
For the IL10 gene, the rs3024496 polymorphism (located in the 3′UTR region), and rs1800896 polymorphism (located in the promoter region) have shown trends of association to susceptibility to cardiomyopathy. The rs3024496C and rs1800896G alleles were more frequent in patients. Our data are consistent with the previous study that showed that the marker (IL10-1082 known as rs1800896 too) was previously associated with development of CCC (33). Costa et al. have shown that the A allele has been associated with lower IL-10 expression in disease and lower left ventricular ejection fraction values in CCC (33). The two markers (rs3024496 and rs1800896) are in strong linkage disequilibrium on the European reference population (D′ = 1) and on the Yoruba population (D′ = 0.83).
Three SNPs in the IL10 gene (the two rs3024496, rs1800896 mentioned above, plus rs1800871) detected trends of association. A study performed on a Brazilian population infected by Leishmania braziliensis has shown that these markers are correlated (r2 value >0.8) (40). So, from one Brazilian sub population to another one, the associated marker may change (due to light allelic frequency variation) or population stratification. However, the associated marker is coming from the same correlation block.
We did not find any significant association with SNPs from the IFNG and IL4 genes. This is in agreement with Arnez et al. who studied some of these same SNPs (rs2070874, rs2227284, rs2243268, rs2243274, rs2243290 in an Bolivian population (29), and Flórez et al. who studied one SNP not tested by our group (rs2243250) in a Colombian population, failing to find any association (30). Taken together, these results suggest that we do not have evidence that polymorphisms in or within the IFN gene control the expression of this key cytokine for the pathogenic process.
During T. cruzi infection, once the inflammatory process starts, IFN-γ will be produced by Th1 cells and act as a prime inflammatory cytokine in different pathways of the immune system, such as upregulating MHC class I and class II molecules, suppressing Th2 immune responses by antagonism of IL-4 production, inducing high levels of antigen presentation and activating macrophages (41). Moreover, CCC patients have an increased peripheral production of IFN-γ and TNF-α when compared to patients with the asymptomatic/indeterminate form. On the other hand, IFN-γ has direct effects on cardiomyocytes and perhaps other cells of the myocardium (34). At this level we can speculate that the existence of functional or pathogenic variants in or within the IFN gene will have a too strong deleterious effect that will be lethal. So, the genetic control of the human susceptibility to cardiomyopathy mainly target genes that are involved in the regulation of this TH1 response, such as IL12B and IL10 genes.
On the other hand, our data confirmed the involvement of IL12B and IL10 in the control of susceptibility to human Chagas cardiomyopathy. The IL10 polymorphism has been described as a functional polymorphism. It remains to be tested whether the IL12B polymorphism and linked SNPs are functional as well. Taking into account the effects of both cytokines on Th1 T cell differentiation and IFNγ production, it can be hypothesized that this is linked to the increased number of IFNγ producing Th1 T cells in the peripheral blood and heart tissue of CCC patients. Our study and others have identified several genes in the control of human susceptibility to chronic disease. However, each gene is characterized by a low odds ratio, implying a small contribution in Chagas disease infection. However, the combination of the several genetic markers increases the protection from disease development, with individual odds ratio around 1.5, to ca. 4.5 in patients carrying all the protection genotypes in all three loci. The identification of more genetic markers for CCC will provide information for pathogenesis as well as therapeutic targets. Identifying early or causal predictive genetic factors for the clinical progression of the disease is essential for clarifying the pathogenesis and defining appropriate treatment modalities.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
The studies involving human participants were reviewed and approved by The INSERM Internal Review Board, The Brazilian National Ethics in Research Commission (CONEP). The patients/participants provided their written informed consent to participate in this study.
JK, EC-N, and CC: study design. AF-B and SC: experiments. AF-B, BI, CP, BS, HL-W, PB, JM-N, AS, FD, MH, MS, AF, AP, ED, VR, JK, CC, and EC-N: manuscript preparation. All authors contributed to the article and approved the submitted version.
This work was supported by the Institut National de la Santé et de la Recherche Médicale (INSERM), Aix-Marseille University (Direction des Relations Internationales), the ARCUS II PACA Brésil program, CNPq (the Brazilian National Research Council), FAPESP (São Paulo State Research Funding Agency-Brazil), and the National Institute of Health (P50 AI098461-02 and U19AI098461-06; to EC-N). EC-N and CC were recipient for an international program funded either by the French ANR and the Brazilian FAPESP agencies (Br-Fr-chagas). AF-B hold fellowships from the São Paulo State Research Funding Agency, FAPESP. EC-N and JK have received a Council for Scientific and Technological Development—CNPq productivity award. CC was a recipient of a temporary professor position supported by the French consulate in Brazil and the University of São Paulo (USP).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We are thankful for the access to the genotyping platform (Denis Milan, Cecile Donnadieu, and Frederic Martins).
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2020.01386/full#supplementary-material
1. Bocchi EA, Bestetti RB, Scanavacca MI, Cunha Neto E, Issa VS. Chronic chagas heart disease management: from etiology to cardiomyopathy treatment. J Am Coll Cardiol. (2017) 70:1510–24. doi: 10.1016/j.jacc.2017.08.004
2. Coura JR, Borges-Pereira J. Chagas disease. What is known and what should be improved: a systemic review. Rev Soc Bras Med Trop. (2012) 45:286–96. doi: 10.1590/S0037-86822012000300002
3. Barbosa AP, Cardinalli Neto A, Otaviano AP, Rocha BF, Bestetti RB. Comparison of outcome between Chagas cardiomyopathy and idiopathic dilated cardiomyopathy. Arq Bras Cardiol. (2011) 97:517–25. doi: 10.1590/S0066-782X2011005000112
4. Bestetti RB, Muccillo G. Clinical course of Chagas' heart disease: a comparison with dilated cardiomyopathy. Int J Cardiol. (1997) 60:187–93. doi: 10.1016/S0167-5273(97)00083-1
5. Zicker F, Smith PG, Netto JC, Oliveira RM, Zicker EM. Physical activity, opportunity for reinfection, and sibling history of heart disease as risk factors for Chagas' cardiopathy. Am J Trop Med Hyg. (1990) 43:498–505. doi: 10.4269/ajtmh.1990.43.498
6. Acosta-Herrera M, Strauss M, Casares-Marfil D, Martin J. Genomic medicine in Chagas disease. Acta Trop. (2019) 197:105062. doi: 10.1016/j.actatropica.2019.105062
7. Chevillard C, Nunes JPS, Frade AF, Almeida RR, Pandey RP, Nascimento MS, et al. Disease tolerance and pathogen resistance genes may underlie Trypanosoma cruzi persistence and differential progression to chagas disease cardiomsyopathy. Front Immunol. (2018) 9:2791. doi: 10.3389/fimmu.2018.02791
8. Strauss M, Acosta-Herrera M, Alcaraz A, Casares-Marfil D, Bosch-Nicolau P, Lo Presti MS, et al. Association of IL18 genetic polymorphisms with Chagas disease in Latin American populations. PLoS Negl Trop Dis. (2019) 13:e0007859. doi: 10.1371/journal.pntd.0007859
9. Cunha-Neto E, Chevillard C. Chagas disease cardiomyopathy: immunopathology and genetics. Mediators Inflamm. (2014) 2014:683230. doi: 10.1155/2014/683230
10. Michailowsky V, Silva NM, Rocha CD, Vieira LQ, Lannes-Vieira J, Gazzinelli RT. Pivotal role of interleukin-12 and interferon-gamma axis in controlling tissue parasitism and inflammation in the heart and central nervous system during Trypanosoma cruzi infection. Am J Pathol. (2001) 159:1723–33. doi: 10.1016/S0002-9440(10)63019-2
11. Bilate AM, Cunha-Neto E. Chagas disease cardiomyopathy: current concepts of an old disease. Rev Inst Med Trop Sao Paulo. (2008) 50:67–74. doi: 10.1590/S0036-46652008000200001
12. Reis MM, Higuchi Mde L, Benvenuti LA, Aiello VD, Gutierrez PS, Bellotti G, et al. An in situ quantitative immunohistochemical study of cytokines and IL-2R+ in chronic human chagasic myocarditis: correlation with the presence of myocardial Trypanosoma cruzi antigens. Clin Immunol Immunopathol. (1997) 83:165–72. doi: 10.1006/clin.1997.4335
13. Muller U, Kohler G, Mossmann H, Schaub GA, Alber G, Di Santo JP, et al. IL-12-independent IFN-gamma production by T cells in experimental Chagas' disease is mediated by IL-18. J Immunol. (2001) 167:3346–53. doi: 10.4049/jimmunol.167.6.3346
14. Rocha Rodrigues DB, dos Reis MA, Romano A, Pereira SA, Teixeira Vde P, Tostes S, et al. In situ expression of regulatory cytokines by heart inflammatory cells in Chagas' disease patients with heart failure. Clin Dev Immunol. (2012) 2012:361730. doi: 10.1155/2012/361730
15. Holscher C, Mohrs M, Dai WJ, Kohler G, Ryffel B, Schaub GA, et al. Tumor necrosis factor alpha-mediated toxic shock in Trypanosoma cruzi-infected interleukin 10-deficient mice. Infect Immun. (2000) 68:4075–83. doi: 10.1128/IAI.68.7.4075-4083.2000
16. Soares MP, Gozzelino R, Weis S. Tissue damage control in disease tolerance. Trends Immunol. (2014) 35:483–94. doi: 10.1016/j.it.2014.08.001
17. Medina TS, Oliveira GG, Silva MC, David BA, Silva GK, Fonseca DM, et al. Ebi3 prevents Trypanosoma cruzi-induced myocarditis by dampening IFN-gamma-driven inflammation. Front Immunol. (2017) 8:1213. doi: 10.3389/fimmu.2017.01213
18. Araujo FF, Gomes JA, Rocha MO, Williams-Blangero S, Pinheiro VM, Morato MJ, et al. Potential role of CD4+CD25HIGH regulatory T cells in morbidity in Chagas disease. Front Biosci. (2007) 12:2797–806. doi: 10.2741/2273
19. Cunha-Neto E, Kalil J. Heart-infiltrating and peripheral T cells in the pathogenesis of human Chagas' disease cardiomyopathy. Autoimmunity. (2001) 34:187–92. doi: 10.3109/08916930109007383
20. Gomes JA, Bahia-Oliveira LM, Rocha MO, Martins-Filho OA, Gazzinelli G, Correa-Oliveira R. Evidence that development of severe cardiomyopathy in human Chagas' disease is due to a Th1-specific immune response. Infect Immun. (2003) 71:1185–93. doi: 10.1128/IAI.71.3.1185-1193.2003
21. Ribeirao M, Pereira-Chioccola VL, Renia L, Augusto Fragata Filho A, Schenkman S, Rodrigues MM. Chagasic patients develop a type 1 immune response to Trypanosoma cruzi trans-sialidase. Parasite Immunol. (2000) 22:49–53. doi: 10.1046/j.1365-3024.2000.00260.x
22. Abel LC, Rizzo LV, Ianni B, Albuquerque F, Bacal F, Carrara D, et al. Chronic Chagas' disease cardiomyopathy patients display an increased IFN-gamma response to Trypanosoma cruzi infection. J Autoimmun. (2001) 17:99–107. doi: 10.1006/jaut.2001.0523
23. Gomes JA, Bahia-Oliveira LM, Rocha MO, Busek SC, Teixeira MM, Silva JS, et al. Type 1 chemokine receptor expression in Chagas' disease correlates with morbidity in cardiac patients. Infect Immun. (2005) 73:7960–6. doi: 10.1128/IAI.73.12.7960-7966.2005
24. Ferreira RC, Ianni BM, Abel LC, Buck P, Mady C, Kalil J, et al. Increased plasma levels of tumor necrosis factor-alpha in asymptomatic/“indeterminate” and Chagas disease cardiomyopathy patients. Mem Inst Oswaldo Cruz. (2003) 98:407–11. doi: 10.1590/S0074-02762003000300021
25. Sousa GR, Gomes JA, Fares RC, Damasio MP, Chaves AT, Ferreira KS, et al. Plasma cytokine expression is associated with cardiac morbidity in chagas disease. PLoS ONE. (2014) 9:e87082. doi: 10.1371/journal.pone.0087082
26. Talvani A, Rocha MO, Ribeiro AL, Correa-Oliveira R, Teixeira MM. Chemokine receptor expression on the surface of peripheral blood mononuclear cells in Chagas disease. J Infect Dis. (2004) 189:214–20. doi: 10.1086/380803
27. Nogueira LG, Santos RH, Fiorelli AI, Mairena EC, Benvenuti LA, Bocchi EA, et al. Myocardial gene expression of T-bet, GATA-3, Ror-gammat, FoxP3, and hallmark cytokines in chronic Chagas disease cardiomyopathy: an essentially unopposed TH1-type response. Mediators Inflamm. (2014) 2014:914326. doi: 10.1155/2014/914326
28. de Alba-Alvarado M, Salazar-Schettino PM, Jimenez-Alvarez L, Cabrera-Bravo M, Garcia-Sancho C, Zenteno E, et al. Th-17 cytokines are associated with severity of Trypanosoma cruzi chronic infection in pediatric patients from endemic areas of Mexico. Acta Trop. (2018) 178:134–41. doi: 10.1016/j.actatropica.2017.11.009
29. Alvarado Arnez LE, Venegas EN, Ober C, Thompson EE. Sequence variation in the IL4 gene and resistance to Trypanosoma cruzi infection in Bolivians. J Allergy Clin Immunol. (2011) 127:279–82, 282 e1–3. doi: 10.1016/j.jaci.2010.10.026
30. Florez O, Martin J, Gonzalez CI. Interleukin 4, interleukin 4 receptor-alpha and interleukin 10 gene polymorphisms in Chagas disease. Parasite Immunol. (2011) 33:506–11. doi: 10.1111/j.1365-3024.2011.01314.x
31. Zafra G, Morillo C, Martin J, Gonzalez A, Gonzalez CI. Polymorphism in the 3′ UTR of the IL12B gene is associated with Chagas' disease cardiomyopathy. Microbes Infect. (2007) 9:1049–52. doi: 10.1016/j.micinf.2007.04.010
32. Turner DM, Williams DM, Sankaran D, Lazarus M, Sinnott PJ, Hutchinson IV. An investigation of polymorphism in the interleukin-10 gene promoter. Eur J Immunogenet. (1997) 24:1–8. doi: 10.1111/j.1365-2370.1997.tb00001.x
33. Costa GC, da Costa Rocha MO, Moreira PR, Menezes CA, Silva MR, Gollob KJ, et al. Functional IL-10 gene polymorphism is associated with Chagas disease cardiomyopathy. J Infect Dis. (2009) 199:451–4. doi: 10.1086/596061
34. Cunha-Neto E, Dzau VJ, Allen PD, Stamatiou D, Benvenutti L, Higuchi ML, et al. Cardiac gene expression profiling provides evidence for cytokinopathy as a molecular mechanism in Chagas' disease cardiomyopathy. Am J Pathol. (2005) 167:305–13. doi: 10.1016/S0002-9440(10)62976-8
35. Jorge MT, Macedo TA, Janones RS, Carizzi DP, Heredia RA, Acha RE. Types of arrhythmia among cases of American trypanosomiasis, compared with those in other cardiology patients. Ann Trop Med Parasitol. (2003) 97:139–48. doi: 10.1179/000349803235001561
36. Januzzi JL Jr. Natriuretic peptides, ejection fraction, and prognosis: parsing the phenotypes of heart failure. J Am Coll Cardiol. (2013) 61:1507–9. doi: 10.1016/j.jacc.2013.01.039
37. van Veldhuisen DJ, Linssen GC, Jaarsma T, van Gilst WH, Hoes AW, Tijssen JG, et al. B-type natriuretic peptide and prognosis in heart failure patients with preserved and reduced ejection fraction. J Am Coll Cardiol. (2013) 61:1498–506. doi: 10.1016/j.jacc.2012.12.044
38. Nogueira LG, Santos RH, Ianni BM, Fiorelli AI, Mairena EC, Benvenuti LA, et al. Myocardial chemokine expression and intensity of myocarditis in Chagas cardiomyopathy are controlled by polymorphisms in CXCL9 and CXCL10. PLoS Negl Trop Dis. (2012) 6:e1867. doi: 10.1371/journal.pntd.0001867
40. Salhi A, Rodrigues V Jr, Santoro F, Dessein H, Romano A, Roberto Castellano L, et al. Immunological and genetic evidence for a crucial role of IL-10 in cutaneous lesions in humans infected with Leishmania braziliensis. J Immunol. (2008) 180:6139–48. doi: 10.4049/jimmunol.180.9.6139
Keywords: Chagas disease, cardiomyopathy, susceptibility, IL12, IL 10, IFN, IL4
Citation: Farage Frade-Barros A, Ianni BM, Cabantous S, Pissetti CW, Saba B, Lin-Wang HT, Buck P, Marin-Neto JA, Schmidt A, Dias F, Hirata MH, Sampaio M, Fragata A, Pereira AC, Donadi E, Rodrigues V, Kalil J, Chevillard C and Cunha-Neto E (2020) Polymorphisms in Genes Affecting Interferon-γ Production and Th1 T Cell Differentiation Are Associated With Progression to Chagas Disease Cardiomyopathy. Front. Immunol. 11:1386. doi: 10.3389/fimmu.2020.01386
Received: 21 March 2020; Accepted: 29 May 2020;
Published: 07 July 2020.
Edited by:
Paul Laszlo Bollyky, Stanford University, United StatesReviewed by:
Javier Martin, Instituto de Parasitología y Biomedicina López-Neyra (IPBLN), SpainCopyright © 2020 Frade-Barros, Ianni, Cabantous, Pissetti, Saba, Lin-Wang, Buck, Marin-Neto, Schmidt, Dias, Hirata, Sampaio, Fragata, Pereira, Donadi, Rodrigues, Kalil, Chevillard and Cunha-Neto. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Amanda Farage Frade-Barros, YW1hbmRhLmZyYWRlQHVuaXZlcnNpZGFkZWJyYXNpbC5lZHUuYnI=
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.