 Jeanne Bigot1
Jeanne Bigot1 Loïc Guillot2
Loïc Guillot2 Juliette Guitard1
Juliette Guitard1 Manon Ruffin2
Manon Ruffin2 Harriet Corvol3
Harriet Corvol3 Viviane Balloy2*†
Viviane Balloy2*† Christophe Hennequin1*†
Christophe Hennequin1*†- 1Sorbonne Université, Inserm, Centre de Recherche Saint-Antoine, CRSA, AP-HP, Hôpital Saint-Antoine, Service de Parasitologie-Mycologie, Paris, France
- 2Sorbonne Université, Inserm, Centre de Recherche Saint-Antoine, Paris, France
- 3Sorbonne Université, Inserm, Centre de Recherche Saint-Antoine, CRSA, AP-HP, Hôpital Trousseau, Service de Pneumologie Pédiatrique, Paris, France
Aspergillus fumigatus is an environmental filamentous fungus that can be pathogenic for humans, wherein it is responsible for a large variety of clinical forms ranging from allergic diseases to life-threatening disseminated infections. The contamination occurs by inhalation of conidia present in the air, and the first encounter of this fungus in the human host is most likely with the bronchial epithelial cells. Although alveolar macrophages have been widely studied in the Aspergillus–lung interaction, increasing evidence suggests that bronchial epithelium plays a key role in responding to the fungus. This review focuses on the innate immune response of the bronchial epithelial cells against A. fumigatus, the predominant pathogenic species. We have also detailed the molecular interactants and the effects of the different modes of interaction between these cells and the fungus.
Introduction
Aspergillus spp. are saprophytic filamentous fungi capable of colonizing different ecological habitats. They are usually isolated from soils, decaying organic matters, and plants but are also present in the air and indoor environments (1). Aspergillus spores, or so-called conidia, represent the disseminating form of the fungus that spreads through the air. These conidia are produced through asexual reproduction by differentiated fungal cells called phialides, which are themselves carried on a conidiophore. Conidia remain “dormant” or metabolically inactive until they encounter favorable environmental conditions. In this case, the conidia swell, germinate to produce hyphae that grow into a mycelium that harbor conidiophores, and then form conidia (2).
Aspergillus genus encompasses several hundred of species (3). Aspergillus fumigatus is by far the most frequent pathogenic species, responsible for about 90% of the cases of Aspergillus diseases, followed by Aspergillus flavus, Aspergillus niger, Aspergillus terreus, and Aspergillus nidulans (3, 4). Indeed, A. fumigatus is the predominant fungal species isolated from the ambient air able to grow at 37°C, the human body temperature.
Humans inhale around a few hundred conidia daily (5). Due to their small size (2- to 3-μm diameter) they can reach the lower respiratory tract (4) but, in most of cases, this phenomenon does not lead to any symptoms thanks to their evacuation by the mucociliary machinery of the tracheobronchial epithelium. However, depending on the immune status of the host, this contamination can be followed by a wide spectrum of manifestations (1). Concisely, immunocompromised patients are at risk for invasive infection, so-called invasive pulmonary aspergillosis (IPA) and patients with pre-formed lung cavity (typically following previous pulmonary tuberculosis) are prone to chronic pulmonary aspergillosis, of which aspergilloma is one of the main presentations. Finally, patients with altered mucociliary clearance, such as cystic fibrosis (CF) patients, may be colonized which can turn, in patients with exacerbated immune response, into allergic bronchopulmonary aspergillosis (ABPA).
The essential role of neutrophils and monocytes in anti-Aspergillus immunity has been emphasized by the high rate of incidence of Aspergillus invasive infection in patients with quantitative (neutropenia) or qualitative (corticosteroid therapy, chronic granulomatous disease) deficiency of these cells (6–8). However, the role of the bronchial epithelium should not be underestimated as it represents the first physical and biological barrier preventing fungal implantation.
While studies looking at the interactions between Aspergillus and leukocytes (alveolar macrophages and recruited neutrophils) are numerous (6, 9–11), data on the role of bronchial epithelial cells (BECs) in anti-Aspergillus defense are still limited. Yet, BECs seem to play a crucial role in the innate immune response against Aspergillus particularly in preventing the bronchial colonization. The high prevalence of Aspergillus bronchial colonization in patients suffering from CF (12, 13), a disease characterized by the thickening of the bronchial mucus, highlights this phenomenon. Bronchial Aspergillus colonization, whose role in the subsequent development of IPA is still debated, may have deleterious consequences as it is the starting point of Aspergillus bronchitis and immuno-allergic forms (14, 15). In CF patients, while remaining superficial, bronchial colonization is associated with the occurrence of bronchial exacerbations, a decline in lung function, and ABPA with a prevalence ranging between 1 and 15% (16). Fungal sensitization to Aspergillus antigens may also occur in allergic patients (17) but the role of the bronchial epithelium in these diseases won't be analyzed in this review.
Thanks to experimental studies, there is increasing knowledge on the interactions between the different morphotypes of A. fumigatus and BECs. This review aims to decipher these interactions at the molecular level and their effect on anti-Aspergillus immunity.
Study Models of the Interaction Between Aspergillus fumigatus and Bronchial Epithelial Cells
The respiratory tract is lined by epithelial cells whose types vary according to the anatomic structure of the airways. Trachea, bronchi, and bronchioles are lined by the pseudostratified epithelium, while type I and II pneumocytes constitute the alveolar epithelium. At the bronchial level, the pseudostratified epithelium is mostly composed of ciliated, secretory, and basal cells from which the first two derive.
To understand the interactions between Aspergillus and BECs, different cell lines (immortalized or tumor) have been commonly used. Among the most popular bronchial cell lines used and commercially available, we can cite BEAS-2B and 16HBE, both isolated from normal human bronchial epithelium and secondarily immortalized through transfection of a replication-defective SV40 plasmid (18, 19). NCI-H292 cells derive from a lymph node metastasis sample of a pulmonary mucoepidermoid carcinoma. But other respiratory cell lines are occasionally used in some studies. All these cell lines have major advantages such as easy to maintain (cultured in simple and inexpensive culture media), capable of growing at high densities, and exhibiting an extended life span (20). However, these cells represent only one donor, and many cellular processes are deregulated due to immortalization. Refinement of the model consists of the use of commercially available primary bronchial cells that are free from any genetic modification and whose physiological functions are intact. However, before they are used, those cells must undergo antibiotic, antifungal, and growth factor treatment. Usually, these cells have a limited life span with limited proliferation capacity and are more difficult to culture than cell lines requiring more complex, specialized, and expensive cell culture media. Irrespective of the cell type, cells are usually cultured under submerged conditions, i.e., in flat-bottom plastic wells filled with culture medium, that hamper cells to differentiate. To better mimic physiological conditions, air-liquid interface cell culture (ALI) systems have been developed (21). In this case, primary BECs and also some cell lines such as 16HBE, differentiate until they develop the mucociliary phenotype characteristic of a pseudostratified epithelium and express mucins (20). Basal surface is therefore in contact with the liquid medium and the apical part of the cellular layer is exposed to the air. Obtaining this type of differentiated epithelium is time-consuming and requires specific technical skills but such cell culture systems mimic the required in vivo conditions in the best way.
Different approaches can be used to mimic an Aspergillus infection. Aspergillus, mostly in the form of dormant conidia can be inoculated in cell culture supernatants and then recovered after defined incubation intervals to measure the parameters of interest (such as cytokine level, cytotoxicity, etc.). In these conditions, A. fumigatus hyphae are usually obtained after 15 h of incubation. Killed (UV or irradiated for example) resting, swollen conidia and hyphae have also been used as inoculum. Differences between experimental protocols, especially the use of different multiplicity of infection, likely far from reality, could explain some discrepancies in the results obtained in different studies.
In addition to in vitro models, in vivo models of Aspergillus infection have already been used. Mice, rats or rabbits are the animals the most commonly used. They are immunosuppressed or not, and infected with A. fumigatus through inhalation of conidia administered either intranasal or via intratracheal route. In addition to the measurements of mortality rate and/or fungal load in the lungs, more precise descriptions of the immune response have also been reported, looking at the immune cells recruitment or inflammatory response (22). In vivo models have the considerable advantage of most closely imitate lung infection and immunity as a whole, however, there is a paucity of in vivo models (conditional and inducible transgenic mice targeting bronchial/airway epithelial cells) allowing the study of BECs against Aspergillus challenge specifically.
Anti-Aspergillus Physicochemical Activity Exhibited by the Bronchial Epithelium
Inhaled conidia first face the physical barriers of the upper airways that include the mouth, nose, larynx, and pharynx. Mucociliary clearance from the nasal walls and mechanical defenses such as coughing and sneezing help eliminate most of the inhaled particles. If the conidia pass these first barriers, they then arrive in the lower airways consisting of the trachea that divided into two-stem bronchial tubes, which in turn are subdivided into several smaller bronchial tubes, followed by bronchioles that end with the alveoli. The bronchial epithelium participates in the clearance of inhaled conidia to prevent their germination and growth locally. Secretory cells, including serous and goblet cells, together with submucosal glands, participate in the formation of mucus, which protects the epithelium from the inhaled particles. Basically, the mucus traps the inhaled particles, which are then actively transported by the beating of the cilia to the oropharynx where they are swallowed or expectorated. Under healthy conditions, mucus is composed of 97% water and 3% of mucins, non-mucin proteins, salts, lipids, and cellular debris (23). Mucins, namely MUC5AC and MUC5B, are the major macromolecular constituents of the mucus. They are large glycoproteins with serine-/threonine-rich domains linked by their hydroxyl side groups to sugar chains forming a polymeric gel that ensure the properties of the mucus (24). Ciliated cells also play a fundamental role in the elimination of particles engulfed in the mucus because they mechanize the movement of the mucus blanket (25). The role of these physicochemical barriers associated with the bronchial epithelium can be better understood in patients suffering from CF or primary ciliary dyskinesia (PCD) in whom mucus properties and/or ciliary beating are impaired. Hence, CF patients are frequently colonized by A. fumigatus (14), and a similar trend has been noticed in patients with PCD (26). This machinery can also be altered by pathogens. Indeed, mycotoxins secreted by A. fumigatus, damage epithelial cells and inhibit ciliary beating (27). Among those toxins, gliotoxin has been extensively studied because it is the most abundant one produced by A. fumigatus and it exhibits immunosuppressive properties that have been described extensively in a previously reported review (28).
Recognition of Aspergillus fumigatus by Bronchial Epithelial Cells
Schematically, the interaction between Aspergillus fumigatus and the BECs leads to three main types of cellular response: internalization, synthesis of cytokines/chemokines and release of bioactive molecules potentially active against Aspergillus. Specific interactions between A. fumigatus and BECs require close contact between the fungus and the cell-surface ligands. Fungal cell-wall polysaccharides and, to a lesser extent, some proteins or the genetic material (ADN or ARN) act as pathogen-associated molecular patterns (PAMPs). They are sensed through pathogen recognition receptors (PRRs), several of which but not all, have been identified in BECs (5) (Table 1).
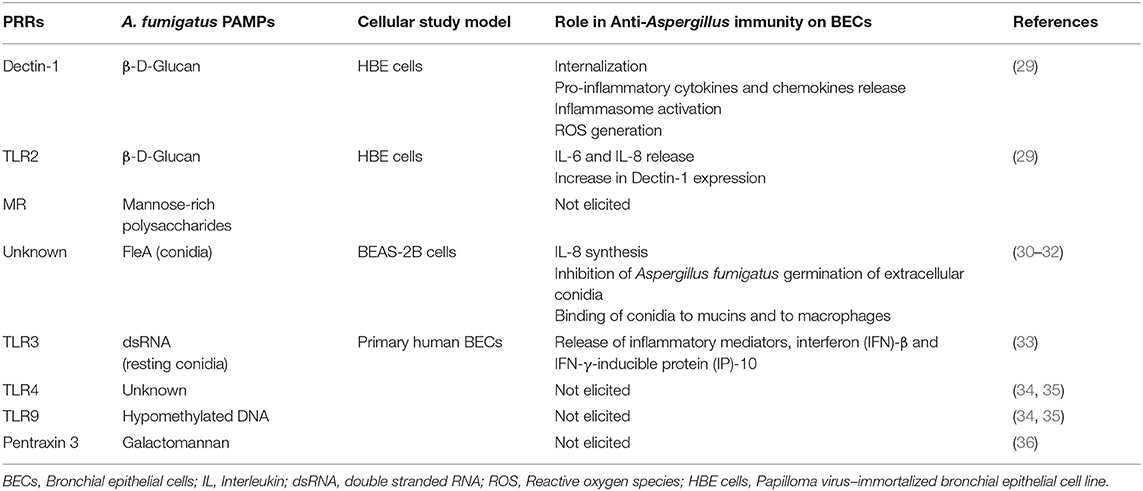
Table 1. Pattern recognition receptors (PRRs) and pathogen-associated molecular patterns (PAMPs) involved in the recognition of Aspergillus fumigatus by bronchial epithelial cells and the consequences of their activation.
A. fumigatus cell wall encompasses an inner and outer layer whose composition varies along with the fungus' life cycle (Figure 1). The outer part of the conidial surface is composed of hydrophobic RodA proteins that conceal an underlying fungal pigment, dihydroxynaphthalene (DHN) melanin (37). This outer layer plays a key role in conidial dispersion, their protection against external stress factors such as desiccation, physical damage, drugs, and UV radiation. They also mask the epitopes present in the underlying layer by inhibiting their recognition by the host's innate immune system (38–40). Whereas, dormant conidia are described as immunologically inert, the FleA lectin, a fucose-binding lectin expressed on their surface (30, 41), has been shown to mediate their binding both to the airway mucins produced by the epithelial cells and also to macrophages (31). Moreover, stimulation of BEAS-2B cells by FleA has been reported to lead to an increase in interleukin (IL)-8 synthesis and contributed to the inflammatory response (30). Thus, FleA acts as a PAMP-like molecule whose cellular ligands remain to be identified.
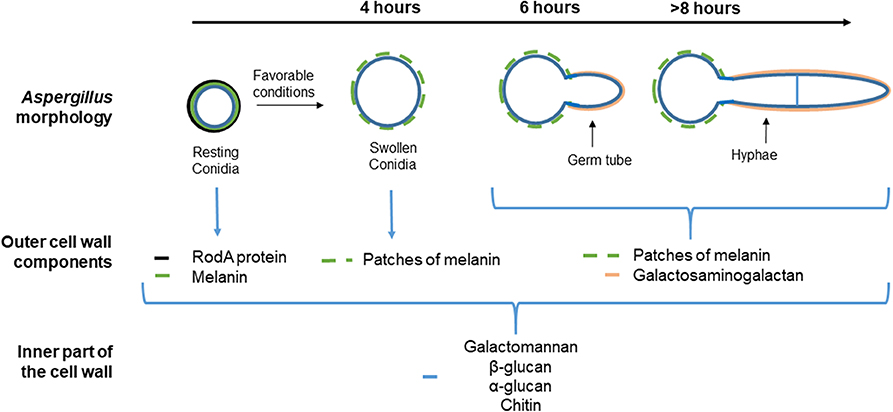
Figure 1. Different morphological stages and compositions of the cell wall during the vegetative cycle of Aspergillus fumigatus. The cell wall of the resting conidia consists of (the most external part to the inner) (i) the rodlet layer (black), composed of the hydrophobin RodA, (ii) the melanin layer below (green), and (iii) polysaccharides (blue). Under favorable conditions, the resting conidia begin to swell after 4 h. The rodlet layer is lost by proteolytic degradation and conidium swelling is due to an increase in the internal osmotic pressure. The melanin layer is then disorganized and the inner layer of the swollen conidium forms the mycelium cell wall. After 8 h, hyphae are apparent. Then, an extracellular matrix mainly composed of galactosaminogalactan (GAG) (pink) covers up the hyphae. Specific and universal components of each morphotype are shown below each image.
Under favorable conditions, the dormant conidia become metabolically active and ensue an increased intracellular osmotic pressure followed by water uptake and isodiametric growth (42). The resulting swollen conidia lose their rodlet layer by proteolytic degradation, and consequently, their hydrophobicity. This phenomenon is associated with a rupture of the melanin layer by a still-unknown mechanism (43). Then, the swollen conidia grow in a polarized way that leads to the formation of a germ tube. At this stage, the melanin layer is disrupted but the inner layer of the cell wall remains unchanged and participates in hyphal formation. The hyphae are mainly composed of galactosaminogalactan (GAG) that allows adhesion of the filaments to various biotic or abiotic surfaces (44, 45). Recent studies have also shown that GAG, expressed during conidial germination, exhibits a possible anti-inflammatory effect. Indeed, GAG induces the release of the IL-1 receptor antagonist (IL-1Ra), a potent anti-inflammatory cytokine that blocks IL-1 signaling (46), by macrophages and neutrophils. Furthermore, in a mouse model of aspergillosis, treatment by GAG before and during intranasally induced A. fumigatus infection inhibits neutrophil infiltration in the lung at the site of infection (6, 44, 46).
Regardless of the A. fumigatus morphotype, the inner layer, the so-called fibrillary core, is continuously composed of branched β-(1, 3)-glucan/β-(1, 4)-glucan, chitin, galactomannan, and α-(1, 3)-glucan (43). A few studies have shown that, as already demonstrated for macrophages and digestive epithelial cells (47), Dectin-1 is a major but not the unique receptor for β-glucan on a papilloma virus-immortalized BEC cell line (HBE cells) (29). Dectin-1 is a transmembrane receptor and member of the C-type lectin receptor family (11). The role of Dectin-1 in the immune response against A. fumigatus has been highlighted in different in vivo studies. For example, immunocompetent mice lacking Dectin-1 are more sensitive to intratracheal challenge with A. fumigatus than control mice (48). In humans, mutations in Dectin-1 are associated with increased susceptibility to IPA (49). It has also been shown that immunocompromised mice, transfected to upregulate Dectin-1 expression in airway epithelial cells, have a lower fungal burden, an increase in the recruitment of neutrophils into the lungs and a greater survival rate in response to intratracheal injection of A. fumigatus conidia compared to the controls (50). After ligation of β-glucan to Dectin-1, two distinct signaling pathways are activated through the spleen tyrosine kinase (SYK)-caspase recruitment domain-containing protein 9 (CARD9) or through RAF-1 (51). These pathways act synergistically to induce nuclear factor-κB activation and pro-inflammatory gene expression (52). The SYK–CARD9 pathway also activates the NOD-, LRR-, and pyrin domain-containing 3 (NLRP3) inflammasome, which results in the proteolytic activation of the pro-inflammatory cytokines IL-1β and IL-18 by caspase 1. The role of A. fumigatus antigens in this activation cascade has been demonstrated by Jeong et al. They showed that sensitized mice intratracheally challenged with A. fumigatus crude antigens displayed an increased in immunofluorescence intensities of NLRP3 and caspase-1 in lung tissue, particularly in epithelial cell layers, leading to an increase in IL1-β concentration in the lung tissue (53). Similar results were obtained by using an in vitro model utilizing primary human BECs stimulated with the same A. fumigatus antigens (53). The mannose receptor (MR) is another C-type lectin receptor involved in fungal, and notably Aspergillus conidia, recognition and is expressed by 16HBE cells and primary BECs (54). The MR recognizes carbohydrates rich in mannose typically produced by many microorganisms including fungi (55) but its immune-specific role against A. fumigatus in association with BECs has not been investigated.
Toll-like receptors (TLRs) are another family of conserved PRRs (56). So far 10 different TLRs have been identified and described. TLR2, TLR4, and TLR9 are the main molecules involved in sensing fungal components (11, 57, 58). These receptors possess extracellular leucine-rich repeat ligand-binding domains and a conserved intracellular toll/IL-1R (TIR) signaling domain that induces specific signaling cascades through intracellular TIR containing adaptors such as MyD88. Interestingly, the expression of the 10 TLRs has been detected by reverse transcription–quantitative real-time polymerase chain reaction in two independent studies using human primary BECs (34, 35). Using a silencing method, it has been demonstrated that TLR2, in a heterodimer form with TLR1 or TLR6, is required for the expression of Dectin-1 by HBE cells in response to A. fumigatus infection (29). Moreover, blocking TLR2 with antibodies results in the inhibition of the release of IL-6 and IL-8 by BEAS-2B cells infected with A. fumigatus hyphal fragments (59). Certain polymorphisms in TLR4, receptor for the bacterial lipopolysaccharide, are clearly associated with increased susceptibility to invasive aspergillosis (60). TLR4 expressed by mouse macrophages, recognizes A. fumigatus conidia and hyphae and induces the release of pro-inflammatory molecules. However, after blocking TLR4 with a specific monoclonal antibody, Øya et al. failed to detect any significant change in the levels of IL-6 and IL-8 released by BEAS-2B cells infected with X-ray-treated hyphal fragments of A. fumigatus (59). Hypomethylated DNA, the natural ligand of TLR9, has been extracted in A. fumigatus hyphae (61). In murine model of invasive aspergillosis, Leiva-Juarez et al. revealed that therapeutic stimulation of lung epithelial defenses by inhalation of a synergistic combination of TLR 2/6 and TLR9 agonists robustly protects against the development of A. fumigatus lung infection despite the profound immune dysfunction (62). TLR9 is expressed by BECs (16HBE cells) (63) but its role in response to Aspergillus challenge remains to be investigated in the context of these cells. TLR3 is localized onto endosomal membranes and recognizes double-stranded (ds)RNA. It is primarily involved in the recognition of viruses but has also been implicated in the recognition of A. fumigatus dsRNA of resting conidia (33). In this study, the infection by Aspergillus of human primary BECs cultivated either in submerged or in ALI culture system, induced the release of inflammatory mediators, notably interferon-β and interferon-γ-inducible protein-10, through TLR3 signaling. Interestingly, this induction is internalization dependent, as demonstrated by the use of an actin-polymerization inhibitor and observed only with killed resting conidia (heat- or UV light-inactivated).
Finally, pentraxin 3, a soluble PRR, plays a key role in the recognition, uptake, and killing of Aspergillus conidia by macrophages and dendritic cells through binding to galactomannan (64). This molecule can act as an opsonizing factor for activating the complement system and subsequent phagocytosis by macrophages (65). Pentraxin 3 is secreted by both human primary bronchial and BEAS-2B cells but, again, its precise role in these cell types has yet to be determined (36).
Internalization of Aspergillus fumigatus Conidia by Bronchial Epithelial Cells
The alveolar epithelial cell line A549 has been extensively used to study conidia internalization by non-professional phagocytic cells (66–68). Wasylnka and Moore described that internalized conidia fused with lysosomes and colocalized with lysosomal protein (Lysosomal-associated membrane protein 1 and CD63). Nonetheless, a significant percentage of internalized conidia persist and germinate in A549 epithelial cells. Little is known about the internalization by BECs and conflicting results have been published (Figure 2A). First, Paris et al., showed by microscopic observations that rabbit tracheal epithelial cells were able to internalize A. fumigatus conidia after 6 h of incubation and that conidia were enclosed in membrane-bound vacuoles (69). Then, other studies reported that the conidia of A. fumigatus were taken up in vitro by a human bronchial epithelial cell line cultured in monolayers (70, 71). Indeed, Clark et al. observed the internalization of 10 to 20% of the conidia in contact with BEAS-2B cells at 6 and 9 h after challenge. In comparison, 70% of conidia in contact with macrophages were internalized after 1 h incubation (71). A 41% internalization rate has also been reported with 16HBE cells after 6 h incubation (70). Overall, these results contrast with those from other studies that have considered the internalization by BECs a very minor phenomenon. By using a model of primary BECs grown in ALI culture system, Toor et al. showed that only 1% of the bound conidia were internalized 6 h after exposure (72). Similarly, Fernandes et al. were unable to demonstrate any case of internalization using primary BECs cultured in ALI (73). However, these authors described the formation, within the cell, of an actin tunnel not altering the viability of the penetrated cells (Figure 2A). The authors suggested that this event could explain the penetration of the hyphae into the underlying parenchyma leading to the development of invasive infection in immunocompromised patients (73). It is noteworthy that the internalization of conidia by BECs was not seen 18 h after an intratracheal challenge in an immunosuppressed mouse model (74). The molecular mechanism(s) involved in the internalization of Aspergillus conidia by BECs remains incompletely understood. Adhesion of conidia onto BECs induces actin polymerization (75), a phenomenon dependent on the activity of the human actin reorganization complex 2 and 3, regulated by Wiskott-Aldrich syndrome protein-interacting proteins (76). Using the BEAS-2B cell line, Clark et al. identified 7 host markers—caveolin, flotillin-2, RAB5C, RAB8B, RAB7A, 2xFYVE, and FAPP1—that consistently localized around the internalized conidia (71).
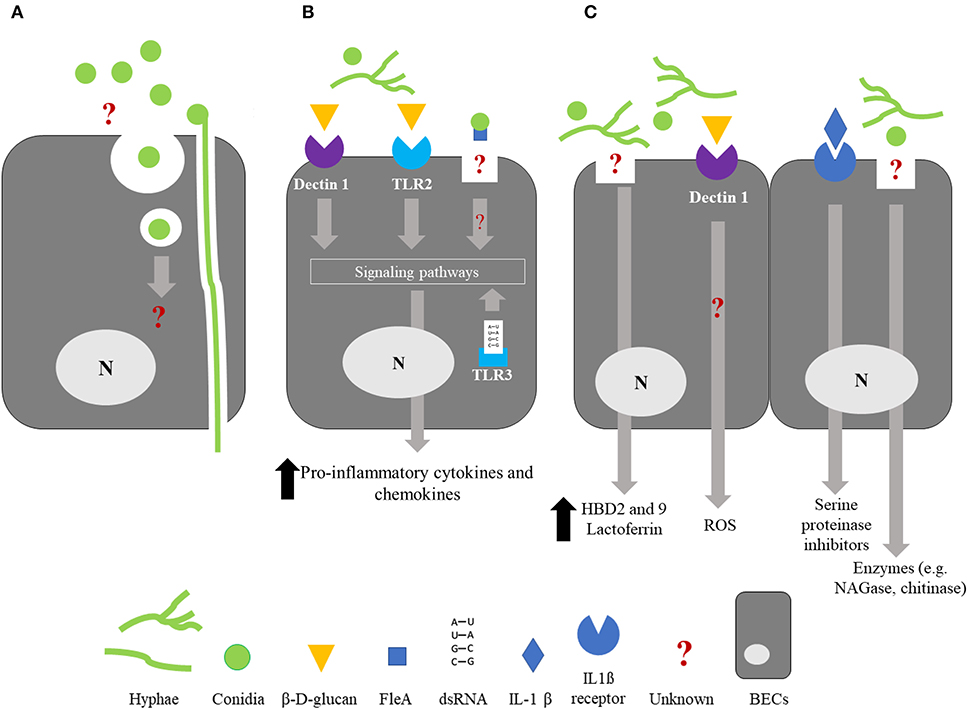
Figure 2. Summary of the interactions between bronchial epithelial cells and Aspergillus fumigatus. For clarity, it should be noted that the schematized epithelial cells are not polarized. (A) Internalization; BECs are able to internalized A. fumigatus conidia but the molecular mechanisms involved in this phenomenon and the fate of the internalized conidia are still unknown. An alternative way of penetration of the hyphae through an actin channel within the cell has been proposed. (B) Cytokine/chemokine release; Recognition of β-D-glucan by BECs through the membrane receptors Dectin-1 and TLR2, and recognition of A. fumigatus dsRNA by the endosomal receptor TLR3 result in the activation of intracellular signaling pathways and a significant increase in the release of proinflammatory cytokines and chemokines, leading to phagocyte recruitment. FleA lectin, expressed on the surface of A. fumigatus conidia, induces IL-8 synthesis by BECs and binding of the conidia to airway mucins. Recognition of FleA by BEC induces the inhibition of conidium germination. (C) Bioactive molecules potentially active against A. fumigatus; BECs synthetize antimicrobials molecules such as defensins HBD2 and HBD9, lactoferrin, reactive oxygen species (ROS), SLPI, ESI, and chitinase in response to A. fumigatus infection or inflammatory challenge. Until now, an antifungal activity has only been described for lactoferrin, ROS, SLPI, ESI, and chitinase. BECs, Bronchial epithelial cells; SLPI, Secretory leukocyte proteinase inhibitor; ESI, Elastase-specific inhibitor; NAGase, N-Acetyl-β-d-glucosaminidase; ROS, Reactive oxygen species; N, nucleus.
Even if the internalization of Aspergillus fumigatus conidia by BECs is a rare event, however, it could represent a starting point of invasive forms in immunosuppressed patients. The fate of the internalized conidia is still unknown and one can postulate that some of them remain quiescent within the cells until they reactivate, thanks to iatrogenic immunosuppression (77). This could explain the breakthrough of some invasive aspergillosis cases occurring in immunocompromised patients otherwise protected from contamination from the ambient air using a high-efficiency particulate air filter chamber (78).
Synthesis and Release of Bioactive Molecules by Bronchial Epithelial Cells Infected With Aspergillus fumigatus
Cytokine/Chemokine Synthesis
One of the most studied consequences of PAMP/PRR interaction is the induction of cytokine/chemokine synthesis. Several studies have shown that infection of BECs with Aspergillus fumigatus leads to the release of pro-inflammatory cytokines, mostly IL-6, IL-8, or tumor necrosis factor-α provided that the infection time is more than 6 h and allows conidia germination (79–81) (Figure 2B). Hence, after 6 h of incubation, expression (mRNA) of not only tumor necrosis factor-α and IL-8 but also granulocyte macrophage-colony stimulating factor (GM-CSF) in HBE cells exposed to A. fumigatus conidia, is significantly increased by 8 to 14 times in regards to non-infected cells (29). Similarly, BEAS-2B cells release an increased amount of IL-8, 8 h after having been infected with A. fumigatus conidia, at a time corresponding to the hyphal formation (79) (Figure 1). In contrast, IL-8 synthesis was not triggered 15 h post-infection in BEAS-2B cells infected with a mutant strain of A. fumigatus unable to germinate (79). Considering the differences in the cell-wall content between the conidial and hyphal stages, these observations strongly suggest a role for some parietal molecules in masking resting conidia, avoiding their recognition by the BECs. RodA hydrophobin, whose masking role toward immune cells has been mentioned above, could be involved but, to the best of our knowledge, this hiding role toward BECs has never been checked.
Inflammatory mediators act synergistically to establish an organized and regulated host response against Aspergillus fumigatus. For example, IL-8, also known as CXCL-8, is a chemokine exhibiting a pleiotropic effect on neutrophils: strong chemotactic influence, degranulation of lysosomes with release of enzymes within the phagosome, production of reactive oxygen species (ROS), and increased expression of adhesion molecules (82). GM-CSF acts both as a hematopoietic growth factor favoring the proliferation and differentiation of myeloid cells into mature cells such as neutrophils and macrophages, and as an enhancer of the antimicrobial functions of those cells. Very recently, our group demonstrated that the inflammatory response of BEAS-2B cells and human primary BECs against A. fumigatus could be reprogrammed after the first contact with a microbial ligand, in this case Pseudomonas aeruginosa flagellin (81). Pre-stimulation with this TLR5 ligand led to a significantly enhanced release of two proinflammatory cytokines, IL-6 and IL-8, after an A. fumigatus challenge. This is comparable to the phenomenon called trained immunity or innate immune memory that has been largely studied using monocytes/macrophages (83–85).
Molecules Potentially Active Against Aspergillus fumigatus
Antimicrobial peptides (AMPs) are cationic small-peptide chains that exhibit antimicrobial activity against a variety of pathogens including fungi (86). Although membrane permeabilization is the main mechanism of action of AMPs against pathogens, additional mechanisms have been described including inhibition of macromolecular synthesis (87). Under basal conditions, BECs release a number of AMPs or proteins, some of which exhibit potential antifungal activity (Figure 2C).
The defensin family, divided into three classes (α-, β-, and θ-defensins), includes broad-spectrum antimicrobial peptides that are evolutionarily conserved across the living world (88). Human β-defensins are a characteristic of epithelial tissues and present a constitutive expression in primary human BECs (89). Human β-defensin 2 and human β-defensin 9 are reported to be highly expressed by BECs (HBE and 16HBE cells) exposed to different morphotypes of A. fumigatus (29, 90). Lactoferrin is a protein synthesized and released by 16HBE cells (91). Interestingly, according to Lupetti et al., a synthetic peptide based on the human lactoferrin sequence but containing only the first cationic domain is one of the most potent antimicrobial peptides against A. fumigatus hyphae and conidia in vitro (92). Lactoferrin can also act by reducing the toxic effect on host cells (cytotoxicity, oxidation level, and DNA damage) of aflatoxin, a mycotoxin synthesized by Aspergillus (93). Secretory leukocyte proteinase inhibitor (SLPI), also called anti-leukoprotease or mucus proteinase inhibitor, and elastase-specific inhibitor (or elafin and trappin-2) are two serine proteinase inhibitors constitutively secreted from the airway epithelium (94). Both are secreted by BECs in response to pro-inflammatory cytokines such as IL-1β (95, 96). Different biological functions have been reported for these molecules: protection of the lungs against the damage induced by neutrophil serine proteases and also antimicrobial activity notably against fungi like A. fumigatus (97, 98). However, these proteins are not considered as AMPs because their size, 11.7 and 9.9 kDa for SPLI and elastase-specific inhibitor respectively, are too large to be classified as peptides.
Interestingly, BEAS-2B and HBE cells are also able to produce ROS (29, 99). In their study on BECs challenged with A. fumigatus, Sun et al. showed that conidia induced ROS generation in a Dectin-1-dependent manner after 6 h of infection (29). ROS are known to be produced by neutrophils thanks to nicotinamide adenine dinucleotide phosphate oxidase produced in response to germinating A. fumigatus challenge (100, 101). They are released from granules either into phagosomes or into the extracellular environment, inducing damage to Aspergillus (6). But until now, the precise role of ROS produced by BECs in antifungal activity remains undetermined.
The study of the secretome of BEAS-2B cells infected with A. fumigatus also gave more insight into the role of some molecules that may possibly act against the fungal infection (102). Among the most significant results, Fekkar et al. found the release of lysosomal enzymes such as N-Acetyl-β-d-glucosaminidase, cathepsin B, and cathepsin D. N-Acetyl-β-d-glucosaminidase is responsible for the hydrolysis of glycosidic bonds. Cathepsin B and D, members of the lysosomal cysteine protease family, are known to acidify the phagosome of macrophages but their role in BECs has not been determined yet. Chitinase, a member of N-Acetyl-β-d-glucosaminidase, degrades the major fungal-wall component chitin and as such could play a role in the control of Aspergillus infection (103, 104).
At this time, not all the active molecules may have been described. Recently, Richard et al. reported that BEAS-2B cells prevent the germination of conidia without internalization but failed to demonstrate the role of any soluble compound present in the supernatants of infected cells (32). This antifungal activity of BECs is fungistatic and occurs via a mechanism that is phosphoinositide 3-kinase dependent. The same kind of observation was made by Clark et al. who found that after infecting BEAS-2B cells with A. fumigatus conidia, a large subset of conidia is rendered metabolically inactive, as measured with the metabolic marker FUN-1, while not being internalized by the cells (71).
Conclusion
The evidence of BECs playing an important role in the innate immunity-based defense mechanism against Aspergillus fumigatus is now growing. Recent studies have been able to demonstrate that, in addition to the production of mucus and ciliary beating that allow the clearance of the conidia, BECs are directly involved in an immune response against A. fumigatus through the recognition of fungal cell-wall components, mainly polysaccharides, by cellular ligands such as Dectin-1 or TLRs. There is a consensus for a pro-inflammatory response by BECs stimulated by A. fumigatus. However, additional studies are needed to better decipher BECs response. Studies focused on the internalization of the conidia by BECs reported divergent results according to the study model. This warrants further studies to clarify this point such as the investigation of the fate of the internalized conidia that could play a role in the future development of invasive aspergillosis. Different types of AMPs are also produced by BECs, some of them being active directly or indirectly against Aspergillus, making these molecules appealing for new therapeutic approaches. Overall, the bronchial epithelium appears as a suitable target for novel therapeutic strategies aiming to restore barrier integrity and to enhance defenses against inhaled pathogens.
Author Contributions
JB, VB, and CH: drafting of the manuscript. JB, VB, CH, LG, JG, MR, and HC: revision of the manuscript.
Funding
VB received grants from Vaincre la mucoviscidose RF20170501940 and RF20190502450.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors would like to dedicate this paper in the honor of their deceased colleague and friend, Michel Chignard.
Abbreviations
BECs, Bronchial epithelial cells; IPA, Invasive pulmonary aspergillosis; CF, Cystic fibrosis; PCD, Primary ciliary dyskinesia; ABPA, Allergic bronchopulmonary aspergillosis; ALI, Air-liquid interface; DHN, Dihydroxynaphthalene; HBE cells, papilloma virus–immortalized bronchial epithelial cell line; PAMPs, Pathogen-associated molecular patterns; PRRs, Pathogen recognition receptors; IL-1Ra, Interleukin-1 receptor antagonist; NLRP3, NOD-, LRR-, and pyrin domain-containing 3; ROS, Reactive oxygen species; IL, Interleukin; AMPs, Antimicrobial peptides; hBD, Human β-defensins; NAGase, N-Acetyl-β-d-glucosaminidase.
References
1. Latgé JP, Chamilos G. Aspergillus fumigatus and aspergillosis in 2019. Clin Microbiol Rev. (2019) 33:e00140-18. doi: 10.1128/CMR.00140-18
2. Samson RA, Visagie CM, Houbraken J, Hong SB, Hubka V, Klaassen CHW, et al. Phylogeny, identification and nomenclature of the genus Aspergillus. Stud Mycol. (2014) 78:141–73. doi: 10.1016/j.simyco.2014.07.004
3. Paulussen C, Hallsworth JE, Álvarez-Pérez S, Nierman WC, Hamill PG, Blain D, et al. Ecology of aspergillosis: insights into the pathogenic potency of Aspergillus fumigatus and some other Aspergillus species. Microb Biotechnol. (2017) 10:296–322. doi: 10.1111/1751-7915.12367
4. Latgé J-P. Aspergillus fumigatus and aspergillosis. Clin Microbiol Rev. (1999) 12:310–50. doi: 10.1128/CMR.12.2.310
5. van de Veerdonk FL, Gresnigt MS, Romani L, Netea MG, Latgé JP. Aspergillus fumigatus morphology and dynamic host interactions. Nat Rev Microbiol. (2017) 15:661–74. doi: 10.1038/nrmicro.2017.90
6. Cunha C, Kurzai O, Löffler J, Aversa F, Romani L, Carvalho A. Neutrophil responses to aspergillosis: new roles for old players. Mycopathologia. (2014) 178:387–93. doi: 10.1007/s11046-014-9796-7
7. Balloy V, Huerre M, Latgé JP, Chignard M. Differences in patterns of infection and inflammation for corticosteroid treatment and chemotherapy in experimental invasive pulmonary aspergillosis. Infect Immun. (2005) 73:494–503. doi: 10.1128/IAI.73.1.494-503.2005
8. Nawada R, Amitani R, Tanaka E, Niimi A, Suzuki K, Murayama T, et al. Murine model of invasive pulmonary aspergillosis following an earlier stage, noninvasive Aspergillus infection. J Clin Microbiol. (1996) 34:1433–9. doi: 10.1128/JCM.34.6.1433-1439.1996
9. Ibrahim-Granet O, Philippe B, Boleti H, Boisvieux-Ulrich E, Grenet D, Stern M, et al. Phagocytosis and intracellular fate of Aspergillus fumigatus conidia in alveolar macrophages. Infect Immun. (2003) 71:891–903. doi: 10.1128/IAI.71.2.891-903.2003
10. Philippe B, Ibrahim-Granet O, Prévost MC, Gougerot-Pocidalo MA, Sanchez Perez M, Van der Meeren A, et al. Killing of Aspergillus fumigatus by alveolar macrophages is mediated by reactive oxidant intermediates. Infect Immun. (2003) 71:3034–42. doi: 10.1128/IAI.71.6.3034-3042.2003
11. Romani L. Immunity to fungal infections. Nat Rev Immunol. (2011) 11:275–88. doi: 10.1038/nri2939
12. Brandt C, Roehmel J, Rickerts V, Melichar V, Niemann N, Schwarz C. Aspergillus bronchitis in patients with cystic fibrosis. Mycopathologia. (2018) 183:61–9. doi: 10.1007/s11046-017-0190-0
13. Zhao J, Cheng W, He X, Liu Y. The Co-colonization prevalence of Pseudomonas aeruginosa and Aspergillus fumigatus in cystic fibrosis: a systematic review and meta-analysis. Microbial Pathog. (2018) 125:122–28. doi: 10.1016/j.micpath.2018.09.010
14. Armstead J, Morris J, Denning DW. Multi-country estimate of different manifestations of aspergillosis in cystic fibrosis. PLoS ONE. (2014) 9:e98502. doi: 10.1371/journal.pone.0098502
15. Gago S, Denning DW, Bowyer P. Pathophysiological aspects of Aspergillus colonization in disease. Med Mycol. (2019) 57(Suppl. 2):S219–27. doi: 10.1093/mmy/myy076
16. Stevens DA, Moss RB, Kurup VP, Knutsen AP, Greenberger P, Judson MA, et al. Allergic bronchopulmonary aspergillosis in cystic fibrosis—state of the art: cystic fibrosis foundation consensus conference. Clin Infect Dis. (2003) 37(Suppl. 3):S225–64. doi: 10.1086/376525
17. Agarwal R. Allergic bronchopulmonary aspergillosis. Chest. (2009) 135:805–26. doi: 10.1378/chest.08-2586
18. Reddel RR, Ke Y, Gerwin BI, McMenamin MG, Lechner JF, Su RT, et al. Transformation of human bronchial epithelial cells by infection with SV40 or adenovirus-12 SV40 hybrid virus, or transfection via strontium phosphate coprecipitation with a plasmid containing SV40 early region genes. Cancer Res. (1988) 48:1904–9.
19. Gruenert DC, Basbaum CB, Welsh MJ, Li M, Finkbeiner WE, Nadel JA. Characterization of human tracheal epithelial cells transformed by an origin-defective Simian virus 40. Proceedings Natl Acad Sci USA. (1988) 85:5951–5. doi: 10.1073/pnas.85.16.5951
20. Hiemstra PS, Grootaers G, van der Does AM, Krul CAM, Kooter IM. Human lung epithelial cell cultures for analysis of inhaled toxicants: lessons learned and future directions. Toxicology In Vitro. (2018) 47:137–46. doi: 10.1016/j.tiv.2017.11.005
21. Gray TE, Guzman K, Davis CW, Abdullah LH, Nettesheim P. Mucociliary differentiation of serially passaged normal human tracheobronchial epithelial cells. Am J Respir Cell Mol Biol. (1996) 14:104–12. doi: 10.1165/ajrcmb.14.1.8534481
22. Desoubeaux G, Cray C. Rodent models of invasive aspergillosis due to Aspergillus fumigatus: still a long path toward standardization. Front Microbiol. (2017) 8:841. doi: 10.3389/fmicb.2017.00841
23. Fahy JV, Dickey BF. Airway mucus function and dysfunction. N Engl J Med. (2010) 363:2233–47. doi: 10.1056/NEJMra0910061
24. Cowley AC, Thornton DJ, Denning DW, Horsley A. Aspergillosis and the role of mucins in cystic fibrosis. Pediatr Pulmonol. (2017) 52:548–55. doi: 10.1002/ppul.23618
25. Yaghi A, Dolovich MB. Airway epithelial cell cilia and obstructive lung disease. Cells. (2016) 5:40. doi: 10.3390/cells5040040
26. Sehgal IS, Dhooria S, Bal A, Agarwal R. Allergic bronchopulmonary aspergillosis in an adult with Kartagener syndrome. BMJ Case Rep. (2015) 2015:bcr2015211493. doi: 10.1136/bcr-2015-211493
27. Chotirmall SH, Mirkovic B, Lavelle GM, McElvaney NG. Immunoevasive Aspergillus virulence factors. Mycopathologia. (2014) 178:363–70. doi: 10.1007/s11046-014-9768-y
28. Arias M, Santiago L, Vidal-García M, Redrado S, Lanuza P, Comas L, et al. Preparations for invasion: modulation of host lung immunity during pulmonary aspergillosis by gliotoxin and other fungal secondary metabolites. Front Immunol. (2018) 9:2549. doi: 10.3389/fimmu.2018.02549
29. Sun WK, Lu X, Li X, Sun QY, Su X, Song Y, et al. Dectin-1 is inducible and plays a crucial role in Aspergillus-induced innate immune responses in human bronchial epithelial cells. Eur J Clin Microbiol Infect Dis. (2012) 31:2755–64. doi: 10.1007/s10096-012-1624-8
30. Houser J, Komarek J, Kostlanova N, Cioci G, Varrot A, Kerr SC, et al. A soluble fucose-specific lectin from Aspergillus fumigatus conidia - structure, specificity and possible role in fungal pathogenicity. PLoS ONE. (2013) 8:e83077. doi: 10.1371/journal.pone.0083077
31. Kerr SC, Fischer GJ, Sinha M, McCabe O, Palmer JM, Choera T, et al. FleA expression in Aspergillus fumigatus is recognized by fucosylated structures on mucins and macrophages to prevent lung infection. PLoS Pathog. (2016) 12:e1005555. doi: 10.1371/journal.ppat.1005555
32. Richard N, Marti L, Varrot A, Guillot L, Guitard J, Hennequin C, et al. Human bronchial epithelial cells inhibit Aspergillus fumigatus germination of extracellular conidia via FleA recognition. Sci Rep. (2018) 8:15699. doi: 10.1038/s41598-018-33902-0
33. Beisswenger C, Hess C, Bals R. Aspergillus fumigatus conidia induce interferon-β signalling in respiratory epithelial cells. Eur Respir J. (2012) 39:411–8. doi: 10.1183/09031936.00096110
34. Sha Q, Truong-Tran AQ, Plitt JR, Beck LA, Schleimer RP. Activation of airway epithelial cells by toll-like receptor agonists. Am J Respir Cell Mol Biol. (2004) 31:358–64. doi: 10.1165/rcmb.2003-0388OC
35. Mayer AK, Muehmer M, Mages J, Gueinzius K, Hess C, Heeg K, et al. Differential recognition of TLR-dependent microbial ligands in human bronchial epithelial cells. J Immunol. (2007) 178:3134–42. doi: 10.4049/jimmunol.178.5.3134
36. Han B, Mura M, Andrade CF, Okutani D, Lodyga M, dos Santos CC, et al. TNFα-induced long pentraxin PTX3 expression in human lung epithelial cells via JNK. J Immunol. (2005) 175:8303–11. doi: 10.4049/jimmunol.175.12.8303
37. Baltussen TJH, Zoll J, Verweij PE, Melchers WJG. Molecular mechanisms of conidial germination in Aspergillus spp. Microbiol Mol Biol Rev. (2020) 84:e00049-19. doi: 10.1128/MMBR.00049-19
38. Bayry J, Beaussart A, Dufrêne YF, Sharma M, Bansal K, Kniemeyer O, et al. Surface structure characterization of Aspergillus fumigatus conidia mutated in the melanin synthesis pathway and their human cellular immune response. Infect Immun. (2014) 82:3141–53. doi: 10.1128/IAI.01726-14
39. Valsecchi I, Dupres V, Stephen-Victor E, Guijarro JI, Gibbons J, Beau R, et al. Role of hydrophobins in Aspergillus fumigatus. J Fungi. (2018) 4:2. doi: 10.3390/jof4010002
40. Aimanianda V, Bayry J, Bozza S, Kniemeyer O, Perruccio K, Elluru SR, et al. Surface hydrophobin prevents immune recognition of airborne fungal spores. Nature. (2009) 460:1117–21. doi: 10.1038/nature08264
41. Kuboi S, Ishimaru T, Tamada S, Bernard EM, Perlin DS, Armstrong D. Molecular characterization of AfuFleA, an l-fucose-specific lectin from Aspergillus fumigatus. J Infect Chemother. (2013) 19:1021–8. doi: 10.1007/s10156-013-0614-9
42. Taubitz A, Bauer B, Heesemann J, Ebel F. Role of respiration in the germination process of the pathogenic mold Aspergillus fumigatus. Curr Microbiol. (2007) 54:354. doi: 10.1007/s00284-006-0413-y
43. Latgé JP, Beauvais A, Chamilos G. The cell wall of the human fungal pathogen Aspergillus fumigatus: biosynthesis, organization, immune response, and virulence. Annu Rev Microbiol. (2017) 71:99–116. doi: 10.1146/annurev-micro-030117-020406
44. Fontaine T, Delangle A, Simenel C, Coddeville B, van Vliet SJ, van Kooyk Y, et al. Galactosaminogalactan, a new immunosuppressive polysaccharide of Aspergillus fumigatus. PLoS Pathog. (2011) 7:e1002372. doi: 10.1371/journal.ppat.1002372
45. Gravelat FN, Beauvais A, Liu H, Lee MJ, Snarr BD, Chen D, et al. Aspergillus galactosaminogalactan mediates adherence to host constituents and conceals hyphal β-glucan from the immune system. PLoS Pathog. (2013) 9:e1003575. doi: 10.1371/journal.ppat.1003575
46. Gresnigt MS, Bozza S, Becker KL, Joosten LA, Abdollahi-Roodsaz S, van der Berg WB, et al. A polysaccharide virulence factor from Aspergillus fumigatus elicits anti-inflammatory effects through induction of interleukin-1 receptor antagonist. PLoS Pathog. (2014) 10:e1003936. doi: 10.1371/journal.ppat.1003936
47. Goyal S, Castrillón-Betancur JC, Klaile E, Slevogt H. The interaction of human pathogenic fungi with C-type lectin receptors. Front Immunol. (2018) 9:1261. doi: 10.3389/fimmu.2018.01261
48. Werner JL, Metz AE, Horn D, Schoeb TR, Hewitt MM, Schwiebert LM, et al. Requisite role for the Dectin-1 beta-glucan receptor in pulmonary defense against Aspergillus fumigatus. J Immunol. (2009) 182:4938–46. doi: 10.4049/jimmunol.0804250
49. Sainz J, Lupiáñez CB, Segura-Catena J, Vazquez L, Ríos R, Oyonarte S, et al. Dectin-1 and DC-SIGN polymorphisms associated with invasive pulmonary aspergillosis infection. PLoS ONE. (2012) 7:e32273. doi: 10.1371/journal.pone.0032273
50. Liu ZC, Wang M, Sun WK, Xia D, Tan MM, Ding Y, et al. Up-regulation of Dectin-1 in airway epithelial cells promotes mice defense against invasive pulmonary aspergillosis. Int J Clin Exp Med. (2015) 8:17489–97.
51. Drummond RA, Brown GD. The role of dectin-1 in the host defence against fungal infections. Curr Opin Microbiol. (2011) 14:392–9. doi: 10.1016/j.mib.2011.07.001
52. Geijtenbeek TB, Gringhuis SI. Signalling through C-type lectin receptors: shaping immune responses. Nat Rev Immunol. (2009) 9:465–79. doi: 10.1038/nri2569
53. Jeong JS, Lee KB, Kim SR, Kim DI, Park HJ, Lee HK, et al. Airway epithelial phosphoinositide 3-kinase-δ contributes to the modulation of fungi-induced innate immune response. Thorax. (2018) 73:758–68. doi: 10.1136/thoraxjnl-2017-210326
54. Michael CF, Waters CM, LeMessurier KS, Samarasinghe AE, Song CY, Malik KU, et al. Airway epithelial repair by a prebiotic mannan derived from Saccharomyces cerevisiae. J Immunol Res. (2017) 2017:8903982. doi: 10.1155/2017/8903982
55. Gazi U, Martinez-Pomares L. Influence of the mannose receptor in host immune responses. Immunobiology. (2009) 214:554–61. doi: 10.1016/j.imbio.2008.11.004
56. De Nardo D. Toll-like receptors: activation, signalling and transcriptional modulation. Cytokine. (2015) 74:181–9. doi: 10.1016/j.cyto.2015.02.025
57. Balloy V, Si-Tahar M, Takeuchi O, Philippe B, Nahori MA, Tanguy M, et al. Involvement of toll-like receptor 2 in experimental invasive pulmonary aspergillosis. Infect Immun. (2005) 73:5420–5. doi: 10.1128/IAI.73.9.5420-5425.2005
58. Chignard M, Balloy V, Sallenave JM, Si-Tahar M. Role of toll-like receptors in lung innate defense against invasive aspergillosis. Distinct impact in immunocompetent and immunocompromized hosts. Clin Immunol. (2007) 124:238–43. doi: 10.1016/j.clim.2007.05.004
59. Øya E, Becher R, Ekeren L, Afanou AKJ, Øvrevik J, Holme JA. Pro-Inflammatory responses in human bronchial epithelial cells induced by spores and hyphal fragments of common damp indoor molds. Int J Environ Res Public Health. (2019) 16:1085. doi: 10.3390/ijerph16061085
60. Bochud PY, Chien JW, Marr KA, Leisenring WM, Upton A, Janer M, et al. Toll-like receptor 4 polymorphisms and aspergillosis in stem-cell transplantation. N Engl J Med. (2008) 359:1766–77. doi: 10.1056/NEJMoa0802629
61. Ramirez-Ortiz ZG, Specht CA, Wang JP, Lee CK, Bartholomeu DC, Gazzinelli RT, et al. Toll-like receptor 9-dependent immune activation by unmethylated CpG motifs in Aspergillus fumigatus DNA. Infect Immun. (2008) 76:2123–9. doi: 10.1128/IAI.00047-08
62. Leiva-Juárez MM, Ware HH, Kulkarni VV, Zweidler-McKay PA, Tuvim MJ, Evans SE. Inducible epithelial resistance protects mice against leukemia-associated pneumonia. Blood. (2016) 128:982–92. doi: 10.1182/blood-2016-03-708511
63. Greene CM, Carroll TP, Smith SG, Taggart CC, Devaney J, Griffin S, et al. TLR-induced inflammation in cystic fibrosis and non-cystic fibrosis airway epithelial cells. J Immunol. (2005) 174:1638–46. doi: 10.4049/jimmunol.174.3.1638
64. Garlanda C, Hirsch E, Bozza S, Salustri A, De Acetis M, Nota R, et al. Non-redundant role of the long pentraxin PTX3 in anti-fungal innate immune response. Nature. (2002) 420:182–6. doi: 10.1038/nature01195
65. Bozza S, Campo S, Arseni B, Inforzato A, Ragnar L, Bottazzi B, et al. PTX3 binds MD-2 and promotes TRIF-dependent immune protection in aspergillosis. J Immunol. (2014) 193:2340–8. doi: 10.4049/jimmunol.1400814
66. Wasylnka JA, Moore MM. Aspergillus fumigatus conidia survive and germinate in acidic organelles of A549 epithelial cells. J Cell Sci. (2003) 116:1579–87. doi: 10.1242/jcs.00329
67. DeHart DJ, Agwu DE, Julian NC, Washburn RG. Binding and germination of Aspergillus fumigatus conidia on cultured A549 pneumocytes. J Infect Dis. (1997) 175:146–50. doi: 10.1093/infdis/175.1.146
68. Zhang Z, Liu R, Noordhoek JA, Kauffman HF. Interaction of airway epithelial cells (A549) with spores and mycelium of Aspergillus fumigatus. J Infect. (2005) 51:375–82. doi: 10.1016/j.jinf.2004.12.012
69. Paris S, Boisvieux-Ulrich E, Crestani B, Houcine O, Taramelli D, Lombardi L, et al. Internalization of Aspergillus fumigatus conidia by epithelial and endothelial cells. Infect Immunity. (1997) 65:1510–14. doi: 10.1128/IAI.65.4.1510-1514.1997
70. Gomez P, Hackett TL, Moore MM, Knight DA, Tebbutt SJ. Functional genomics of human bronchial epithelial cells directly interacting with conidia of Aspergillus fumigatus. BMC Genomics. (2010) 11:358. doi: 10.1186/1471-2164-11-358
71. Clark HR, Powell AB, Simmons KA, Ayubi T, Kale SD. Endocytic markers associated with the internalization and processing of Aspergillus fumigatus conidia by BEAS-2B cells. mSphere. (2019) 4:e00663-18. doi: 10.1128/mSphere.00663-18
72. Toor A, Culibrk L, Singhera GK, Moon KM, Prudova A, Foster LJ, et al. Transcriptomic and proteomic host response to Aspergillus fumigatus conidia in an air-liquid interface model of human bronchial epithelium. PLoS ONE. (2018) 13:e0209652. doi: 10.1371/journal.pone.0209652
73. Fernandes J, Hamidi F, Leborgne R, Beau R, Castier Y, Mordant P, et al. Penetration of the human pulmonary epithelium by Aspergillus fumigatus hyphae. J Infect Dis. (2018) 218:1306–13. doi: 10.1093/infdis/jiy298
74. Rammaert B, Jouvion G, de Chaumont F, Garcia-Hermoso D, Szczepaniak C, Renaudat C, et al. Absence of fungal spore internalization by bronchial epithelium in mouse models evidenced by a new bioimaging approach and transmission electronic microscopy. Am J Pathol. (2015) 185:2421–30. doi: 10.1016/j.ajpath.2015.04.027
75. Croft CA, Culibrk L, Moore MM, Tebbutt SJ. Interactions of Aspergillus fumigatus conidia with airway epithelial cells: a critical review. Front Microbiol. (2016) 7:472. doi: 10.3389/fmicb.2016.00472
76. Culibrk L, Croft CA, Toor A, Yang SJ, Singhera GK, Dorscheid DR, et al. Phagocytosis of Aspergillus fumigatus by human bronchial epithelial cells is mediated by the Arp2/3 complex and WIPF2. Front Cell Infect Microbiol. (2019) 9:16. doi: 10.3389/fcimb.2019.00016
77. Amitani R, Kawanami R. Interaction of Aspergillus with human respiratory mucosa: a study with organ culture model. Med Mycol. (2009) 47(Suppl. 1):S127–31. doi: 10.1080/13693780802558959
78. Roilides E, Simitsopoulou M. Local innate host response and filamentous fungi in patients with cystic fibrosis. Med Mycol. (2010) 48:S22–31. doi: 10.3109/13693786.2010.511286
79. Balloy V, Sallenave JM, Wu Y, Touqui L, Latgé JP, Si-Tahar M, et al. Aspergillus fumigatus-induced interleukin-8 synthesis by respiratory epithelial cells is controlled by the phosphatidylinositol 3-kinase, P38 MAPK, and ERK1/2 pathways and not by the toll-like receptor-MyD88 pathway. J Biol Chem. (2008) 283:30513–21. doi: 10.1074/jbc.M803149200
80. Tomee JF, Wierenga AT, Hiemstra PS, Kauffman HK. Proteases from Aspergillus fumigatus induce release of proinflammatory cytokines and cell detachment in airway epithelial cell lines. J Infect Dis. (1997) 176:300–3. doi: 10.1086/517272
81. Bigot J, Guillot L, Guitard J, Ruffin M, Corvol H, Chignard M, et al. Respiratory epithelial cells can remember infection: a proof of concept study. J Infect Dis. (2019) 221:1000–5. doi: 10.1093/infdis/jiz569
82. Balloy V, Chignard M. The innate immune response to Aspergillus fumigatus. Microbes Infect. (2009) 11:919–27. doi: 10.1016/j.micinf.2009.07.002
83. Ifrim DC, Quintin J, Joosten LAB, Jacobs C, Jansen T, Jacobs L, et al. Trained immunity or tolerance: opposing functional programs induced in human monocytes after engagement of various pattern recognition receptors. Clin Vaccine Immunol. (2014) 21:534–45. doi: 10.1128/CVI.00688-13
84. Netea MG, Quintin J, van der Meer JW. Trained immunity: a memory for innate host defense. Cell Host Microbe. (2011) 9:355–61. doi: 10.1016/j.chom.2011.04.006
85. Netea MG, Joosten LAB, Latz E, Mills KHG, Natoli G, Stunnenberg HG, et al. Trained immunity: a program of innate immune memory in health and disease. Science. (2016) 352:aaf1098. doi: 10.1126/science.aaf1098
86. Kang HK, Kim C, Seo CH, Park Y. The therapeutic applications of antimicrobial peptides (AMPs): a patent review. J Microbiol. (2017) 55:1–12. doi: 10.1007/s12275-017-6452-1
87. Sierra JM, Fusté E, Rabanal F, Vinuesa T, Viñas M. An overview of antimicrobial peptides and the latest advances in their development. Expert Opin Biol Ther. (2017) 17:663–76. doi: 10.1080/14712598.2017.1315402
88. Ganz T, Weiss J. Antimicrobial peptides of phagocytes and epithelia. Semin Hematol. (1997) 34:343–54.
89. Arnason JW, Murphy JC, Kooi C, Wiehler S, Traves SL, Shelfoon C, et al. Human β-defensin-2 production upon viral and bacterial co-infection is attenuated in COPD. PLoS ONE. (2017) 12:e0175963. doi: 10.1371/journal.pone.0175963
90. Alekseeva L, Huet D, Féménia F, Mouyna I, Abdelouahab M, Cagna A, et al. Inducible expression of beta defensins by human respiratory epithelial cells exposed to Aspergillus fumigatus organisms. BMC Microbiol. (2009) 9:33. doi: 10.1186/1471-2180-9-33
91. van den Berge M, Jonker MR, Miller-Larsson A, Postma DS, Heijink I. H. Effects of fluticasone propionate and budesonide on the expression of immune defense genes in bronchial epithelial cells. Pulmon Pharmacol Ther. (2018) 50:47–56. doi: 10.1016/j.pupt.2018.04.002
92. Lupetti A, van Dissel JT, Brouwer CP, Nibbering PH. Human antimicrobial peptides' antifungal activity against Aspergillus fumigatus. Eur J Clin Microbiol Infect Dis. (2008) 27:1125–9. doi: 10.1007/s10096-008-0553-z
93. Zheng N, Zhang H, Li S, Wang J, Liu J, Ren H, et al. Lactoferrin inhibits aflatoxin B1- and aflatoxin M1-induced cytotoxicity and DNA damage in Caco-2, HEK, Hep-G2, SK-N-SH cells. Toxicon. (2018) 150:77–85. doi: 10.1016/j.toxicon.2018.04.017
94. Williams SE, Brown TI, Roghanian A, Sallenave JM. SLPI and Elafin: one glove, many fingers. Clin Sci. (2006) 110:21–35. doi: 10.1042/CS20050115
95. Sallenave JM, Shulmann J, Crossley J, Jordana M, Gauldie J. Regulation of secretory leukocyte proteinase inhibitor (SLPI) and elastase-specific inhibitor (ESI/elafin) in human airway epithelial cells by cytokines and neutrophilic enzymes. Am J Respir Cell Mol Biol. (1994) 11:733–41. doi: 10.1165/ajrcmb.11.6.7946401
96. Sallenave JM. Secretory leukocyte protease inhibitor and elafin/trappin-2. Am J Respir Cell Mol Biol. (2010) 42:635–43. doi: 10.1165/rcmb.2010-0095RT
97. Baranger K, Zani ML, Chandenier J, Dallet-Choisy S, Moreau T. The antibacterial and antifungal properties of trappin-2 (Pre-Elafin) do not depend on its protease inhibitory function. FEBS J. (2008) 275:2008–20. doi: 10.1111/j.1742-4658.2008.06355.x
98. Tomee JF, Hiemstra PS, Heinzel-Wieland R, Kauffman HF. Antileukoprotease: an endogenous protein in the innate mucosal defense against fungi. J Infect Dis. (1997) 176:740–7. doi: 10.1086/514098
99. Chan TK, Tan WSD, Peh HY, Wong WSF. Aeroallergens induce reactive oxygen species production and DNA damage and dampen antioxidant responses in bronchial epithelial cells. J Immunol. (2017) 199:39–47. doi: 10.4049/jimmunol.1600657
100. Imbert S, Bresler P, Boissonnas A, Gauthier L, Souchet L, Uzunov M, et al. Calcineurin inhibitors impair neutrophil activity against Aspergillus fumigatus in allogeneic hematopoietic stem cell transplant recipients. J Allergy Clin Immunol. (2016) 138:860–8. doi: 10.1016/j.jaci.2016.02.026
101. Chotirmall SH, Al-Alawi M, Mirkovic B, Lavelle G, Logan PM, Greene CM, et al. Aspergillus-associated airway disease, inflammation, and the innate immune response. Biomed Res Int. (2013) 2013:723129. doi: 10.1155/2013/723129
102. Fekkar A, Balloy V, Pionneau C, Marinach-Patrice C, Chignard M, Mazier D. Secretome of human bronchial epithelial cells in response to the fungal pathogen aspergillus fumigatus analyzed by differential in-gel electrophoresis. J Infect Dis. (2012) 205:1163–72. doi: 10.1093/infdis/jis031
103. Karamanos Y. Endo-N-Acetyl-β-D-glucosaminidases and their potential substrates : structure/function relationships. Res Microbiol. (1997) 148:661–71. doi: 10.1016/S0923-2508(99)80065-5
Keywords: bronchial epithelial cells, Aspergillus fumigatus, innate immunity, lung infection, mucociliary machinery
Citation: Bigot J, Guillot L, Guitard J, Ruffin M, Corvol H, Balloy V and Hennequin C (2020) Bronchial Epithelial Cells on the Front Line to Fight Lung Infection-Causing Aspergillus fumigatus. Front. Immunol. 11:1041. doi: 10.3389/fimmu.2020.01041
Received: 25 February 2020; Accepted: 30 April 2020;
Published: 22 May 2020.
Edited by:
Paul W. Bland, University of Gothenburg, SwedenReviewed by:
Paul King, Monash University, AustraliaZhengxiang He, Icahn School of Medicine at Mount Sinai, United States
Copyright © 2020 Bigot, Guillot, Guitard, Ruffin, Corvol, Balloy and Hennequin. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Viviane Balloy, dml2aWFuZS5iYWxsb3lAaW5zZXJtLmZy; Christophe Hennequin, Y2hyaXN0b3BoZS5oZW5uZXF1aW4tc2F0QGFwaHAuZnI=
†These authors have contributed equally to this work