
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Immunol. , 11 February 2020
Sec. Molecular Innate Immunity
Volume 11 - 2020 | https://doi.org/10.3389/fimmu.2020.00111
 Tiago Lazzaretti Fernandes1,2,3*†
Tiago Lazzaretti Fernandes1,2,3*† Andreas H. Gomoll4Christian Lattermann3Arnaldo Jose Hernandez1,2
Andreas H. Gomoll4Christian Lattermann3Arnaldo Jose Hernandez1,2 Daniela Franco Bueno2Mariane Tami Amano2
Daniela Franco Bueno2Mariane Tami Amano2Cartilage lesions and osteoarthritis (OA) presents an ever-increasing clinical and socioeconomic burden. Synovial inflammation and articular inflammatory environment are the key factor for chondrocytes apoptosis and hypertrophy, ectopic bone formation and OA progression. To effectively treat OA, it is critical to develop a drug that skews inflammation toward a pro-chondrogenic microenvironment. In this narrative and critical review, we aim to see the potential use of immune cells modulation or cell therapy as therapeutic alternatives to OA patients. Macrophages are immune cells that are present in synovial lining, with different roles depending on their subtypes. These cells can polarize to pro-inflammatory (M1) and anti-inflammatory (M2) phenotypes, being the latter associated with wound-healing by the production of ARG-1 and pro-chondrogenic cytokines, such as IL-10, IL-1RA, and TGF-b. Emerging evidence reveals that macrophage shift can be determined by several stimuli, apart from the conventional in vitro IL-4, IL-13, and IL-10. Evidences show the potential of physical exercise to induce type 2 response, favoring M2 polarization. Moreover, macrophages in contact with oxLDL have effect on the production of anabolic mediators as TGF-b. In the same direction, type II collagen, that plays a critical role in development and maturation process of chondrocytes, can also induce M2 macrophages, increasing TGF-b. The mTOR pathway activation in macrophages was shown to be able to polarize macrophages in vitro, though further studies are required. The possibility to use mesenchymal stem cells (MSCs) in cartilage restoration have a more concrete literature, besides, MSCs also have the capability to induce M2 macrophages. In the other direction, M1 polarized macrophages inhibit the proliferation and viability of MSCs and impair their ability to immunosuppress the environment, preventing cartilage repair. Therefore, even though MSCs therapeutic researches advances, other sources of M2 polarization are attractive issues, and further studies will contribute to the possibility to manipulate this polarization and to use it as a therapeutic approach in OA patients.
Cartilage repair is the critical issue that patients with symptomatic cartilage lesions seek (1). Chondral lesion is a pathology with high prevalence, reaching as much as 63% of general population and 36% among athletes (2, 3). It has an impact in socioeconomic health system and the attempted treatment of these lesions is associated with a considerable economic burden (4). For instance, cartilage and osteoarthritis treatment can delay joint replacement and improve symptoms (5).
Articular cartilage tissue presents limited cellularity and lacks a vascular system, leading to restrained healing capability (6, 7). Actually, there is no available treatment to regenerate hyaline cartilage or modify disease progression (8). Consequently, cartilage injuries are often related to pain and joint instability that may diminish or even cease the tissue's functionality (6, 7). Thus, articular cartilage is at high risk of damage during initial trauma and, if left untreated, may results in lesions in the underlying subchondral bone, leading to biomechanics and homeostasis disturbances in the knee as a whole. This process may result in loss of mobility, wear and arthritis (9, 10).
Despite the numerous techniques available today, complete healing of damaged or defective cartilage and the consistent reproduction of normal hyaline cartilage is an elusive goal (5). For these reasons, continuous drug therapies and secondary surgeries are common, and new therapeutics for articular cartilage lesions is of elevated clinical relevance (11, 12).
Among cell therapeutics solutions, it is observed two main examples: Autologous Chondrocyte Implantation (ACI) and Mesenchymal Stem Cells (MSCs).
ACI is a two-step procedure that consists of healthy cartilage harvesting through arthroscopy followed by the expanded cell culture and, in a second step, cartilage defect filling (13–15). In spite of second and third ACI generations' versatility, those techniques use healthy cartilage tissue, “in vitro” related chondrocytes dedifferentiation and still fails to fully reproduce the hyaline characteristics of the original articular cartilage (6, 13, 15–17).
More recently, mesenchymal stem cell-based therapy has received considerable attention, because of the feasibility of handling the tissue harvest and ex vivo cell expansion and differentiation (12, 13). Moreover, these cells present minor immunological rejection due to the low surface expression of major histocompatibility complex (MHC) antigens, efficient engraftment and long-term coexistence in the host, which turns them an attractive therapeutic option (18, 19). According to the International Society of Cell Therapy (ISCT), MSCs are defined as plastic-adherent when maintained in standard culture conditions, specific surface antigen expression, and the cells must be able to differentiate to osteoblasts, adipocytes and chondroblasts under standard “in vitro” differentiating conditions (20).
The capacity of allogeneic MSCs to repair cartilage lesions has been reported in clinical trials (21) and in translational large animal models (12, 22). These cells migrate to damaged tissues and contribute to their repair by secretion of cytokines, chemokines, and extracellular matrix proteins (23). The chondrogenic potential and the known immunosuppressive characteristics of MSCs, point out these cells as a powerful tool in the therapy of osteoarthritis (OA) (18).
The estimated prevalence OA in the population is 22.7% and it is believed that by 2020, more than 50 million people will suffer from OA in the United States. This will be the major cause of morbidity and physical limitation among individuals aged over 40 years (24). Current therapy for OA is directed toward non-pharmacological treatments as physical activity through mechanical stimulation (25) and symptomatic treatment, focusing in pain management, and is not able to promote regeneration of degenerated cartilage or to attenuate joint inflammation (18).
A cartilage breakdown results in a release of molecular fragments into synovial fluid that starts macrophages fragment removal in the synovium and further signaling in a positive feedback loop triggering apoptosis in chondrocytes (26).
OA is a disease which affects all joint tissues, characterized by progressive degeneration of the articular cartilage, neovascular invasion of articular surface, subchondral bone remodeling, osteophyte formation, bone marrow lesions, meniscal damage and synovial inflammation (synovitis) (18, 27). Joint effusion is detected in half of patients with OA symptoms and no radiographic findings, indicating that synovitis is not restricted to severe OA only, but is associated with increased pain and dysfunction (28).
Accumulating evidence suggests that synovial inflammation is correlative with the pathogenesis and progression of OA (27). And articular inflammatory environment is the key factor for initiation and aggregation of cartilage lesion (29). Clinical symptoms of OA are attributed to synovial inflammation (26).
Synovial inflammation is characterized by synovial thickening with hypertrophy and hyperplasia (30). Increased vascular density and inflammatory cell infiltration (lymphocytes and macrophages) are common features of OA (31). Macrophage accumulation in the intimal lining, reflecting mostly proliferative synovial tissue, is the principal morphological characteristic of synovitis (27).
A substantial part of OA patients develops synovial activation (32). Synovial lining macrophages play a crucial role in driving joint pathology, such as cartilage damage and ectopic bone formation (32). Macrophage-derived inflammatory cytokines [tumor necrosis factor alpha (TNF-a) and interleukin (IL)-1b] shift synovial tissue homeostasis toward catabolism by promoting production of matrix degrading enzymes that results with an increased bone and cartilage resorption (18). In a rat model of OA induced by anterior cruciate ligament transection, it was also observed increased of inflammatory OA-related cytokines, such as TNF-a, IL-1b, and matrix metallopeptidase 13 (MMP13). While moderate physical activity decreased the expression of these cytokines and increased the level of anti-inflammatory and chondroprotective proteins, such as IL-4 and IL-10 (25) corroborating the idea of immune response contributing to the synovial dynamic.
The synovial membrane is an area of high functional importance within the joint, responsible for the production of synovial fluid, which lubricates and nourishes chondrocytes (31). The membrane is composed of two cell types, synovial macrophages and fibroblasts (31). Normal synovium consists of two distinct tissue layers. One is the intimal lining layer with two to three layers of macrophages (type A cells) with phagocytic function and fibroblast-like synoviocytes (type B cells) with secretory function that produces hyaluronan and lubricin (25, 26); the other is the synovial sublining layer, composed of fibrous connective tissue and blood vessels, with few lymphocytes or macrophages (27).
OA development involves multiple pathological changes, including synoviocytes dysfunction, chondrocytes apoptosis and hypertrophy and immune cells activation (25, 33). These changes have an impact in the articular microenvironment, being crucial for the cartilage repair (33). Labinsky et al. (34) stated that OA inflammation is not homogeneous, different inflammatory phenotypes exists and it may influence each patient selection criteria and treatment (28). Thus, the OA treatment requires a multifactorial approach that restrain inflammatory response and provide a pro-chondrogenic microenvironment. Macrophages are protagonists in the balance of inflammation and regeneration, which catches the attention for the possibility of their use as therapeutic tools.
Macrophages, together with fibroblasts, are present in the synovial lining of joints. They are involved in synovial inflammation, and have been shown to play a prominent role in the progression of OA (27). Macrophages are innate immune cells that express MHC class II, which gives them the ability to initiate adaptive immune response through T cell activation. Macrophage from synovial tissue expresses CD14, CD68, CSF1R, HLA-DRA, and MARCO (35).
After initial stimulation, macrophages acquire a phenotype, ranging from pro-inflammatory (M1) to anti-inflammatory (M2) (36). In the joint, CD14 and CD163 are associated to pro-inflammatory and anti-inflammatory phenotypes, respectively (28). Synovial macrophages can have different origins, being embryonic- or bone marrow-derived, and depending on the source they might present different roles in arthritis (37). Culemann et al. demonstrated a dynamic membrane-like structure formed by resident synovial CX3CR1+ macrophages that physically isolate the joint and restrict inflammation. These CX3CR1+ macrophages are derived from mononuclear cells embedded in the synovial tissue, and present opposite function from infiltrating monocyte-derived macrophages, which are responsible to inflammatory response in the joint (38).
These data suggest a therapeutic potential of the modulation of macrophages in OA.
Macrophages M1 were seen to be associated to Th1 response. Mice strains that were known to favor Th1 (C57Bl6) or Th2 (BALB/c) responses, presented preferentially induction to M1 or M2 differentiation, respectively (39). M1 macrophages are the “classically activated” and are associated with high production of pro-inflammatory cytokines and chemokines as shown in Table 1 (27, 40). M1 macrophages can be induced by interferon-g (IFN-g) and lipopolysaccharide (LPS) increasing CD80 expression (41). Lepage et al. also stated that monocytes subjected to TNF-a can polarize to M1 phenotype (36). Once in contact with these stimuli, transcription factors, such as IRF5 activates transcription of genes encoding IL-12, IL-23 and represses IL-10 (42). M1 stimulation also leads to nitric oxide (NO) production and cell surface expression of the co-stimulatory molecules CD86, CD80, and MHC, which are required for T cell activation (43). In general, M1 macrophages have high microbicidal activity and secrete large amounts of pro-inflammatory cytokines (Table 1) (36, 44).
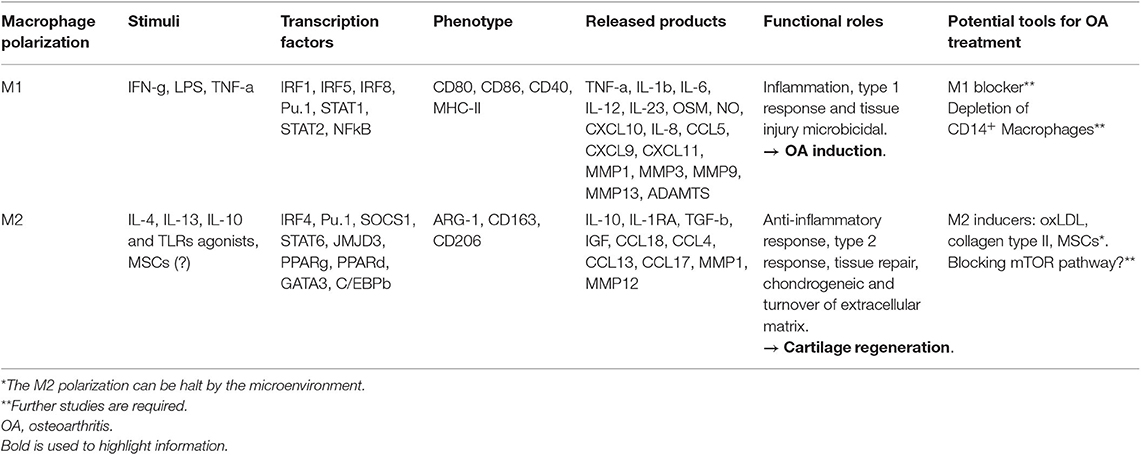
Table 1. Macrophage polarization in OA and cartilage regeneration.
The “alternatively activated” macrophages were denominated M2, and they are known as wound-healing macrophages. The M2 macrophages have been further divided into specific subtypes: M2a (induced by IL-4 and IL-13), M2b (induced by TLRs agonists), and M2c (induced by IL-10). All subtypes promote anti-inflammatory responses (44). After M2 induction, the transcription factor IRF4 among others are activated favoring the polarization (45) (Table 1). M2 macrophages are characterized by the expression of CD163 and CD206 markers and production of arginase (ARG)-1 (Table 1). These cells present an anti-inflammatory function producing, IL-10, IL-1RA, chemokine (CeC motif) ligand 18 (CCL18), and TGF-b (31, 46) as well as pro-chondrogenic factors: TGF-b1, TGF-b2, TGF-b3, insulin-like growth factor 1 (IGF1), and 2 (IGF2) (33) (Table 1). The IL-10-induced subtype M2c plays a role in tissue remodeling (36). Some of the known M2-related genes are ARG-1, resistin like alpha (FIZZ1), mannose receptor, C type 1 (MRC), human macrophage galactose-type C-type lectin (CLEC10A) (47). Arginine metabolism into nitric oxide (NO) and citrulline (M1 macrophages) or ornithine and urea (M2 macrophages) may be used to distinguish phenotypes. Their relative proportion of NO/urea is useful for functional readout since it reflects the ratio of M1/M2 polarization (8).
Macrophages play pivotal roles in innate immunity and exhibit a high degree of plasticity. Synovial macrophages have similar phenotype than others resident macrophages, including CD11b, CD14, CD16, and CD68 (30). O'Brien et al. (26) found that there were more macrophages in the early stages of synovial OA than compared to the late stages. They also demonstrated that synovial macrophages are decreased in pre-OA joints in comparison to normal knees, and that MSCs and macrophages are spatially closer to each other in normal and pre-OA than in OA cases (26). M1 affects OA cartilage by inhibiting genes associated with matrix production, upregulation of matrix degenerating genes and induction of inflammation. Fahy et al. (31) stated that the M1-associated cytokines IL-6, IL1b, TNF-a, and Oncostatin M (OSM) induce destructive processes in chondrocytes including down regulation of collagen type II and aggrecan synthesis. Synovial M1 macrophages were also shown to up regulate the production of proteolytic enzymes, such as matrix metalloproteinase (MMP)-1, MMP3, MMP13, MMP9 aggrecanases (ADAMTS), and cyclooxygenase-2, which contribute to articular degeneration (8, 30, 31). It was demonstrated that synovial macrophages and monocyte-derived pro-inflammatory macrophages negatively affected chondrogenesis of MSCs (36).
M2 macrophages have a major role in tissue repair (44), and the shift from M0 toward M2 macrophages in the lesion might contribute to repair the damaged articular cartilage. Dai et al. showed that under the stimulation of certain biomaterials M2 macrophages could be induced, releasing certain regulatory cytokines and exerting an immunomodulatory effect on tissue healing (33) (Figure 1).
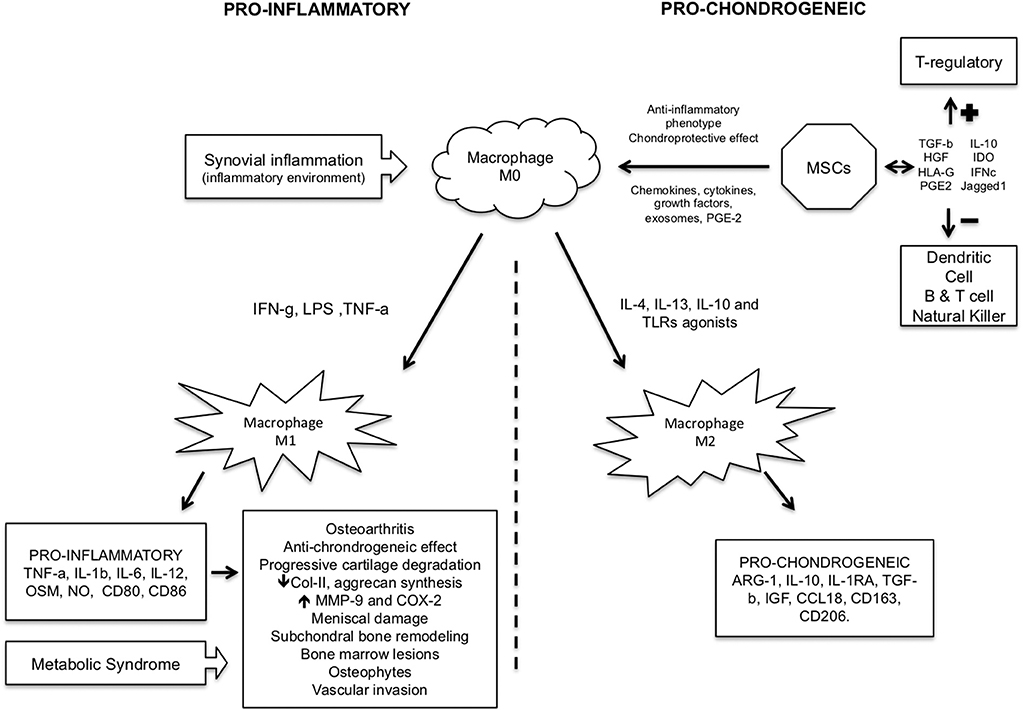
Figure 1. Pro-inflammatory and pro-chondrogeneic macrophage pathways in cartilage injury/repair.
It seems clear that pro-inflammatory macrophage M1 contributes to OA, while M2 could reverse it or favor the chondrogenesis process. There are evidences showing that moderate physical activity could change the synovial microenvironment, shifting it from a type 1 to type 2 immune response, which is associated to M2 macrophages with the presence of IL-4 and IL-10 cytokines. These changes could provide a protective environment in the joints of OA patients (25). The use of gold nanoparticles were shown to present anti-inflammatory macrophage response (48), and a pilot study was opened as clinical trial (NCT03389906) in order to see impact of these particles in macrophages from OA knees, though no further analysis concerning M1 and M2 was mentioned.
Lepage et al. showed that M1 and M2a macrophages did not affect OA cartilage, while M2c seemed to induce inflammation to some extent, although less intense than M1. Both M2a and M2c were unable to counteract the pro-inflammatory effects of M1 (36). This was somewhat unexpected since it was previously demonstrated that M2 macrophages induced by macrophage colony-stimulating factor (M-CSF) enhanced chondrogenesis in vitro (36). The analogous M1 polarization to M2 M-CSF-induced is the stimulation with granulocyte macrophage colony-stimulating fator (GM-CSF) (49). The use of anti-GM-CSF was applied in a clinical trial with patients with OA. Although some positive effect was seen in diminishing the pain, MRI images showed no change in synovitis after anti-GM-CSF treatment (50). Wood et al. (35) declared that sub-groups of synovial macrophages functions as a spectrum with different remodeling signatures and Tardito et al. (51) emphasized the need of a not classical sub-division into M1 and M2 macrophages, but a M1 and M2 coexistence and equilibrium.
To date, there are few clinical studies in humans considering macrophage polarization and osteoarthritis (27, 34, 50, 52). Most of them do not access macrophage polarization and when they access, they are in vitro studies with human cells. It points out the novelty and clinical relevance of this topic, and the possibility for new clinical perspectives and treatments for OA.
Therefore, the M2 induction is crucial for chondrogenesis development, and inhibiting M1 macrophage appears to be a good target for treatments. However, further investigations are required before translating these approaches to the patients.
Metabolic syndrome. is a combination of pathological processes that increases cardiovascular risks associating hypertension, diabetes, obesity and high levels of low-density lipoproteins (LDL). And it was reported to be associated to OA, though the mechanisms involved remain unclear (32, 53). Oxidized LDL (oxLDL) is a modified LDL and it interacts with macrophages that accumulate in the subendothelial space and transform into foam cells (54). This accumulation of cells leads to chronic inflammation in the arterial wall culminating in atherosclerosis initiation (54, 55). Despite the inflammation observed by foam cells, macrophages treated with oxLDL polarize to M2 phenotype, presenting increased production of IL-10 and TGFb (56).
de Munter et al. and Griffin et al. had postulated that low grade inflammation during OA could induce local oxidation of LDL aggravating OA pathology (57). Synovial fluid (SF) contains LDL (28, 58). Either serum- or serum free (SF)-derived LDL could be oxidized under inflammatory conditions and taken up by synovial cells (59). However, the same group demonstrated later that injections of oxLDL in knee joints in mouse model significantly increased TGF-b without inducing catabolic response and inflammatory response (32). Curiously, injection of oxLDL in macrophage-depleted animals increased inflammation, with higher expression of CCL2 and CCL3, attracting more monocytes and neutrophils and increasing synovial thickening (32). These data suggest that oxLDL injections could contribute to OA protection by inducing M2 macrophages.
Type II collagen has been classically recognized as the indispensable collagenous component in articular cartilage, and plays a critical role in the development and maturation process of chondrocytes (33). In vitro studies reported that type II collagen increases secretion of cartilage matrix by chondrocytes (60).
M2 macrophages express mannose receptors MRC1 (CD206) and together with MRC2 can recognize several types of collagen, promoting internalization and lysosomal degradation (61, 62). M2 macrophages were shown to contribute to collagen turnover to keep the extracellular matrix homeostasis (63). On the other hand, type II collagen was shown to induced M2 polarization, by increasing the expression of M2-related genes (MR, Arg1, Fizz1, and Ym1) as well as the pro-chondrogenic cytokines (TGF-b and IGF). In an OA rat model, one of the groups was treated with collagen type II injections and it was observed increased M2 macrophages, higher production of TGF-b in synovial fluid, diminished chondrocyte apoptosis and decreased MMP13 production (33). MMP13 degrades cartilage matrix (type II collagen and proteoglycan) (64).
Since M2-associated cytokines participate in tissue repair (65), we could infer that collagen type II could favor M2 polarization, turning the microenvironment prone to chondrogenesis, contributing to cartilage repair and keeping the extracellular matrix stability.
Emerging evidence reveals that the mammalian target of rapamycin (mTOR) pathway plays key roles in macrophage polarization (27). The canonical activation of this pathway is by the phosphorylation of PI3K, followed by AKT that phosphorylate the tuberous sclerosis complex (TSC1-TSC2-TBC1D7) (66). The TSC2 phosphorylated inhibits the Ras homolog enriched in brain (Rheb), which in turn activates mTORC1. The downstream activation of this pathway leads to changes in metabolism that favor cellular growth signals, modulating innate and adaptive immune response (67). Genetically modified animals with TSC specifically depleted from myeloid cells (TSC−/−), which therefore leads to constitutive mTOR complex 1 (mTORC1) activation, were shown to favor M1 and fail to polarize to M2 phenotype (68, 69). In a low synovial activation OA model, the TSC−/− mice presented increased M1 accumulation in synovial and exacerbated experimental OA (27). Conversely, deletion of Rheb1 in the myeloid lineage (Rheb1−/−) enhanced synovial macrophage M2 polarization and attenuated OA (27).
The production of IL-12, iNOS, and TNF-a (M1-like macrophage markers) are upregulated in TSC1−/− macrophages in comparison to control macrophages, indicating the M1 inflammatory phenotype with mTORC1 activation (68). Taken together, these findings demonstrate that M1-polarized macrophages induced by mTORC1 promote hypertrophic chondrocyte differentiation and maturation, suggesting that it plays a critical role in cartilage degeneration during OA. The inhibition of mTOR pathway has long been used in transplanted patients (with drugs, such as rapamycin, sirolimus, and everolimus), because of its property of blocking T cell proliferation (70), one could consider the use of these inhibitors to treat OA. However, Zhu et al. demonstrated that inhibition of mTOR did not reverse the M1 response in TSC−/− cells (69). Moreover, PI3K knockout (71) or mTORC2 deleted macrophages (72), both presenting impaired mTOR pathway activation, also favored M1 polarization, showing a complex scenario before considering a therapeutic approach by manipulating this pathway.
It is well-known that MSCs exhibit immune-tolerance capacity by downregulating effector immune cells response and favoring an immunosuppressed environment. MSCs can influence innate immune cells, such as macrophages, dendritic cells, and natural killer cells, as well as adaptive immune cells, such as T and B lymphocytes (23, 73, 74). One of the mechanisms that MSCs can influence immune cells is through secretion of immune regulating molecules, such as TGFb, hepatocyte growth factor (HGF), HLA-G, prostaglandin (PGE2), IL-10, and indoleamine 2,3-dioxygenase (IDO) (23, 73). Besides the paracrine secretion of cytokines, MSC can also modulate inflammatory response by cell to cell (46).
MSCs inhibit activation of inflammatory M1 macrophages and promote anti-inflammatory M2 polarization in vitro (18). MSCs is associated to the conversion of TNF-a and IL-1 inflammatory cytokines into immunosuppressive IL-10 production by macrophages, which attenuate joint inflammation and promote cartilage regeneration (18).
CD14 is a membrane antigen (glycoprotein) expressed on the surface on macrophages and monocytes (52). In the other sense of immune-modulation, Han et al. stated that depletion of human CD14+ synovial macrophages allows osteoarthritic synovial MSCs for chondrogenic potential (52). NF-kB represents a family of inducible dimeric transcription factors that stimulates osteoarthritis and it is a pivotal factor that induces suppression of the chondrogenic potential of human osteoarthritic synovial MSCs (52). Chahal et al. (75) demonstrated a reduced level of monocytes/macrophages pro-inflammatory IL-12 cytokine in synovial fluid levels and likely improved clinical efficacy in patient-reported outcomes after 3 months of high doses of MSCs injection in the knee (75).
It was demonstrated that intra-articular injection of adipose derived stromal cells (ASCs) in OA animal models exert anti-inflammatory and chondroprotective effects (76, 77). In cell culture, MSCs secrete a large number of chemokines, cytokines, and growth factors that pushes macrophages to polarize toward an anti-inflammatory phenotype (46).
Hamilton et al. demonstrated a decrease in proportion of iNOS and reduction of pro-inflammatory macrophages after MSCs injection in a murine OA model (78). It was reported that MSCs decreased synovial inflammation and fibrosis (78).
PGE2 is a lipid mediator derived from the conversion of arachidonic acid to the prostaglandin through COX1 and COX2 enzymes that has an important role in MSC immunosuppression. PGE2 production by MSCs promote conversion of M1 to M2 phenotype (79). Manferdini et al. demonstrated that PGE2, mainly produced by ASC, was directly responsible for inhibition of the inflammatory cytokines TNF-a and IL6 (46) and blocking PGE2 by EP4 receptor antagonist showed the opposite effect corroborating these data (80).
Nevertheless, osteoarthritic-conditioned medium and synovial fluid were shown to inhibit the chondrogenic differentiation of MSCs (81, 82), which indicates that the presence of a destructive inflammatory environment, as found in OA, may halt the MSCs properties to cartilage repair. Fahy et al. showed that M1 macrophages inhibit chondrogenic differentiation of MSCs. These findings suggests that synovial macrophages are key regulators of the chondrogenesis of OA synovium (31).
Even though MSCs are clearly a potent M2 inducer, if the environment is exacerbated inflamed, MSCs by themselves may not be able to conduct the cells to type 2 response. Other stimuli, such as oxLDL, collagen type II, and signaling pathways manipulation could help to control the OA damage or cartilage repair.
Osteoarthritis is a disabling and very incapacitant disease with no definitive treatment that concerns patients and physicians. International societies are working hard on osteoarthritis treatment. Currently, no final or effective modifying-disease treatment is available and clinicians can only prescribe physical activity, alleviate symptoms or post-pone surgeries.
The clinical relevance of this study was to perform a critical review of literature and analysis of the current knowledge to open new roads for innovative and translational clinical trials of macrophage polarization in humans to treat osteoarthritis.
Macrophages plasticity can provide interesting therapeutic approaches in OA. There are several evidences that macrophage polarization could contribute to cartilage repair, either by inducing M2 or blocking M1 macrophages. M2 macrophages increase TGF-b, a well-known anti-inflammatory and pro-chondrogenic cytokine that contributes to MSCs differentiation and cartilage formation.
Since MSCs in an already inflamed tissue have their properties impaired, in addition to the attempts of the use of MSCs and their products (vesicles, micro-vesicles, and exosomes) in cartilage restoration, M2 macrophages induction could improve even more the MSCs therapeutic effects. Thus, further studies on M2 induction in combination with MSCs in cartilage repair could enormously contribute to OA treatments perspectives.
TF and MA took responsibility for the integrity of the study from conception and design to completion. Drafting of the manuscript and critical revision was performed by TF, MA, CL, AG, AH, and DB.
This study was supported by Fundação de Amparo a Pesquisa do Estado de São Paulo (FAPESP–Process 2017/05774-5), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES–Process 88881.171651/2018-01), grant from the International Society of Arthroscopy, Knee Surgery and Orthopaedic Sports Medicine and the Orthopaedic Research and Education Foundation (ISAKOS Osteoarthritis Grant−2018), Sírio-Libanês Hospital and IOT HC-FMUSP Hospital das Clínicas, Faculdade de Medicina da USP.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
1. Farr J, Gomoll AH. 2016 barriers to cartilage restoration. J Clin Orthop Trauma. (2016) 7:183–6. doi: 10.1016/j.jcot.2016.05.001
2. Perera JR, Gikas PD, Bentley G. The present state of treatments for articular cartilage defects in the knee. Ann R Coll Surg Engl. (2012) 94:381–7. doi: 10.1308/003588412X13171221592573
3. Flanigan DC, Harris JD, Trinh TQ, Siston RA, Brophy RH. Prevalence of chondral defects in athletes' knees: a systematic review. Med Sci Sports Exerc. (2010) 42:1795–801. doi: 10.1249/MSS.0b013e3181d9eea0
4. Samuelson EM, Brown DE. Cost-effectiveness analysis of autologous chondrocyte implantation: a comparison of periosteal patch versus type I/III collagen membrane. Am J Sports Med. (2012) 40:1252–8. doi: 10.1177/0363546512441586
5. Fernandes TL, SantAnna JP, Frisene I, Gazarini JP, Pinheiro CCG, Gomoll AH, et al. Systematic review of human dental pulp stem cells for cartilage regeneration. Tissue Eng Part B Rev. (2019). doi: 10.1089/ten.TEB.2019.0140. [Epub ahead of print].
6. Kubosch EJ, Lang G, Furst D, Kubosch D, Izadpanah K, Rolauffs B, et al. The potential for synovium-derived stem cells in cartilage repair. Curr Stem Cell Res Ther. (2018) 13:174–84. doi: 10.2174/1574888X12666171002111026
7. Zainal Ariffin SH, Kermani S, Megat Abdul Wahab R, Senafi S, Zainal Ariffin Z, Abdul Razak M. In vitro chondrogenesis transformation study of mouse dental pulp stem cells. Sci World J. (2012) 2012:827149. doi: 10.1100/2012/827149
8. Haltmayer E, Ribitsch I, Gabner S, Rosser J, Gueltekin S, Peham J, et al. Co-culture of osteochondral explants and synovial membrane as in vitro model for osteoarthritis. PLoS ONE. (2019) 14:e0214709. doi: 10.1371/journal.pone.0214709
9. Gomoll AH, Madry H, Knutsen G, van Dijk N, Seil R, Brittberg M, et al. The subchondral bone in articular cartilage repair: current problems in the surgical management. Knee Surg Sports Traumatol Arthrosc. (2010) 18:434–47. doi: 10.1007/s00167-010-1072-x
10. Showery JE, Kusnezov NA, Dunn JC, Bader JO, Belmont PJ Jr, Waterman BR. The rising incidence of degenerative and posttraumatic osteoarthritis of the knee in the United States military. J Arthroplasty. (2016) 31:2108–14. doi: 10.1016/j.arth.2016.03.026
11. Tuan RS, Chen AF, Klatt BA. Cartilage regeneration. J Am Acad Orthop Surg. (2013) 21:303–11. doi: 10.5435/JAAOS-21-05-303
12. Fernandes TL, Shimomura K, Asperti A, Pinheiro CCG, Caetano HVA, Oliveira C, et al. Development of a novel large animal model to evaluate human dental pulp stem cells for articular cartilage treatment. Stem Cell Rev. (2018) 14:734–43. doi: 10.1007/s12015-018-9820-2
13. Shimomura K, Ando W, Moriguchi Y, Sugita N, Yasui Y, Koizumi K, et al. Next generation mesenchymal stem cell (MSC)-based cartilage repair using scaffold-free tissue engineered constructs generated with synovial mesenchymal stem cells. Cartilage. (2015) 6:13S−29S. doi: 10.1177/1947603515571002
14. Ochi M, Adachi N, Nobuto H, Yanada S, Ito Y, Agung M. Articular cartilage repair using tissue engineering technique–novel approach with minimally invasive procedure. Artif Organ. (2004) 28:28–32. doi: 10.1111/j.1525-1594.2004.07317.x
15. Fernandes TL, Kimura HA, Pinheiro CCG, Shimomura K, Nakamura N, Ferreira JRM, et al. Human synovial MSC good manufacturing practices for articular cartilage regeneration. Tissue Eng Part C Methods. (2018) 24:709–16. doi: 10.1089/ten.TEC.2018.0219
16. Niemeyer P, Salzmann G, Feucht M, Pestka J, Porichis S, Ogon P, et al. First-generation versus second-generation autologous chondrocyte implantation for treatment of cartilage defects of the knee: a matched-pair analysis on long-term clinical outcome. Int Orthop. (2014) 38:2065–70. doi: 10.1007/s00264-014-2368-0
17. Hickery MS, Bayliss MT, Dudhia J, Lewthwaite JC, Edwards JC, Pitsillides AA. Age-related changes in the response of human articular cartilage to IL-1alpha and transforming growth factor-beta (TGF-beta): chondrocytes exhibit a diminished sensitivity to TGF-beta. J Biol Chem. (2003) 278:53063–71. doi: 10.1074/jbc.M209632200
18. Harrell CR, Markovic BS, Fellabaum C, Arsenijevic A, Volarevic V. Mesenchymal stem cell-based therapy of osteoarthritis: current knowledge and future perspectives. Biomed Pharmacother. (2019) 109:2318–26. doi: 10.1016/j.biopha.2018.11.099
19. Prockop DJ. Repair of tissues by adult stem/progenitor cells (MSCs): controversies, myths, and changing paradigms. Mol Ther. (2009) 17:939–46. doi: 10.1038/mt.2009.62
20. Dominici M, Le Blanc K, Mueller I, Slaper-Cortenbach I, Marini F, Krause D, et al. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy. (2006) 8:315–7. doi: 10.1080/14653240600855905
21. Park YB, Ha CW, Lee CH, Yoon YC, Park YG. Cartilage regeneration in osteoarthritic patients by a composite of allogeneic umbilical cord blood-derived mesenchymal stem cells and hyaluronate hydrogel: results from a clinical trial for safety and proof-of-concept with 7 years of extended follow-up. Stem Cells Transl Med. (2017) 6:613–21. doi: 10.5966/sctm.2016-0157
22. Shimomura K, Ando W, Tateishi K, Nansai R, Fujie H, Hart DA, et al. The influence of skeletal maturity on allogenic synovial mesenchymal stem cell-based repair of cartilage in a large animal model. Biomaterials. (2010) 31:8004–11. doi: 10.1016/j.biomaterials.2010.07.017
23. Waterman RS, Tomchuck SL, Henkle SL, Betancourt AM. A new mesenchymal stem cell (MSC) paradigm: polarization into a pro-inflammatory MSC1 or an Immunosuppressive MSC2 phenotype. PLoS ONE. (2010) 5:e10088. doi: 10.1371/journal.pone.0010088
24. Shen J, Chen D. Recent progress in osteoarthritis research. J Am Acad Orthop Surg. (2014) 22:467–8. doi: 10.5435/JAAOS-22-07-467
25. Castrogiovanni P, Di Rosa M, Ravalli S, Castorina A, Guglielmino C, Imbesi R, et al. Moderate physical activity as a prevention method for knee osteoarthritis and the role of synoviocytes as biological key. Int J Mol Sci. (2019) 20:E511. doi: 10.3390/ijms20030511
26. O'Brien K, Tailor P, Leonard C, DiFrancesco LM, Hart DA, Matyas JR, et al. Enumeration and localization of mesenchymal progenitor cells and macrophages in synovium from normal individuals and patients with pre-osteoarthritis or clinically diagnosed osteoarthritis. Int J Mol Sci. (2017) 18:E774. doi: 10.3390/ijms18040774
27. Zhang H, Lin C, Zeng C, Wang Z, Wang H, Lu J, et al. Synovial macrophage M1 polarisation exacerbates experimental osteoarthritis partially through R-spondin-2. Ann Rheum Dis. (2018) 77:1524–34. doi: 10.1136/annrheumdis-2018-213450
28. Griffin TM, Scanzello CR. Innate inflammation and synovial macrophages in osteoarthritis pathophysiology. Clin Exp Rheumatol. (2019) 37:57–63.
29. Bondeson J, Blom AB, Wainwright S, Hughes C, Caterson B, van den Berg WB. The role of synovial macrophages and macrophage-produced mediators in driving inflammatory and destructive responses in osteoarthritis. Arthritis Rheum. (2010) 62:647–57. doi: 10.1002/art.27290
30. Manferdini C, Paolella F, Gabusi E, Silvestri Y, Gambari L, Cattini L, et al. From osteoarthritic synovium to synovial-derived cells characterization: synovial macrophages are key effector cells. Arthritis Res Ther. (2016) 18:83. doi: 10.1186/s13075-016-0983-4
31. Fahy N, de Vries-van Melle ML, Lehmann J, Wei W, Grotenhuis N, Farrell E, et al. Human osteoarthritic synovium impacts chondrogenic differentiation of mesenchymal stem cells via macrophage polarisation state. Osteoarthritis Cartilage. (2014) 22:1167–75. doi: 10.1016/j.joca.2014.05.021
32. de Munter W, Geven EJ, Blom AB, Walgreen B, Helsen MM, Joosten LA, et al. Synovial macrophages promote TGF-beta signaling and protect against influx of S100A8/S100A9-producing cells after intra-articular injections of oxidized low-density lipoproteins. Osteoarthritis Cartilage. (2017) 25:118–27. doi: 10.1016/j.joca.2016.07.020
33. Dai M, Sui B, Xue Y, Liu X, Sun J. Cartilage repair in degenerative osteoarthritis mediated by squid type II collagen via immunomodulating activation of M2 macrophages, inhibiting apoptosis and hypertrophy of chondrocytes. Biomaterials. (2018) 180:91–103. doi: 10.1016/j.biomaterials.2018.07.011
34. Labinsky H, Panipinto PM, Ly KA, Khuat DK, Madarampalli B, Mahajan V, et al. Multiparameter analysis identifies heterogeneity in knee osteoarthritis synovial responses. Arthritis Rheumatol. (2019). doi: 10.1002/art.41161. [Epub ahead of print].
35. Wood MJ, Leckenby A, Reynolds G, Spiering R, Pratt AG, Rankin KS, et al. Macrophage proliferation distinguishes 2 subgroups of knee osteoarthritis patients. JCI Insight. (2019) 4:125325. doi: 10.1172/jci.insight.125325
36. Lepage SIM, Robson N, Gilmore H, Davis O, Hooper A, St John S, et al. Beyond cartilage repair: the role of the osteochondral unit in joint health and disease. Tissue Eng Part B Rev. (2019) 25:114–25. doi: 10.1089/ten.teb.2018.0122
37. Tu J, Hong W, Guo Y, Zhang P, Fang Y, Wang X, et al. Ontogeny of synovial macrophages and the roles of synovial macrophages from different origins in arthritis. Front Immunol. (2019) 10:1146. doi: 10.3389/fimmu.2019.01146
38. Culemann S, Gruneboom A, Nicolas-Avila JA, Weidner D, Lammle KF, Rothe T, et al. Locally renewing resident synovial macrophages provide a protective barrier for the joint. Nature. (2019) 572:670–5. doi: 10.1038/s41586-019-1471-1
39. Mills CD, Kincaid K, Alt JM, Heilman MJ, Hill AM. M-1/M-2 macrophages and the Th1/Th2 paradigm. J Immunol. (2000) 164:6166–73. doi: 10.4049/jimmunol.164.12.6166
40. Murray PJ, Allen JE, Biswas SK, Fisher EA, Gilroy DW, Goerdt S, et al. Macrophage activation and polarization: nomenclature and experimental guidelines. Immunity. (2014) 41:14–20. doi: 10.1016/j.immuni.2014.06.008
41. Tarique AA, Logan J, Thomas E, Holt PG, Sly PD, Fantino E. Phenotypic, functional, and plasticity features of classical and alternatively activated human macrophages. Am J Respir Cell Mol Biol. (2015) 53:676–88. doi: 10.1165/rcmb.2015-0012OC
42. Krausgruber T, Blazek K, Smallie T, Alzabin S, Lockstone H, Sahgal N, et al. IRF5 promotes inflammatory macrophage polarization and TH1-TH17 responses. Nat Immunol. (2011) 12:231–8. doi: 10.1038/ni.1990
43. Shapouri-Moghaddam A, Mohammadian S, Vazini H, Taghadosi M, Esmaeili SA, Mardani F, et al. Macrophage plasticity, polarization, and function in health and disease. J Cell Physiol. (2018) 233:6425–40. doi: 10.1002/jcp.26429
44. Benoit M, Desnues B, Mege JL. Macrophage polarization in bacterial infections. J Immunol. (2008) 181:3733–9. doi: 10.4049/jimmunol.181.6.3733
45. Platanitis E, Decker T. Regulatory networks involving STATs, IRFs, and NFkappaB in inflammation. Front Immunol. (2018) 9:2542. doi: 10.3389/fimmu.2018.02542
46. Manferdini C, Paolella F, Gabusi E, Gambari L, Piacentini A, Filardo G, et al. Adipose stromal cells mediated switching of the pro-inflammatory profile of M1-like macrophages is facilitated by PGE2: in vitro evaluation. Osteoarthritis Cartilage. (2017) 25:1161–71. doi: 10.1016/j.joca.2017.01.011
47. Raes G, Van den Bergh R, De Baetselier P, Ghassabeh GH, Scotton C, Locati M, et al. Arginase-1 and Ym1 are markers for murine, but not human, alternatively activated myeloid cells. J Immunol. (2005) 174:6561; author reply 6561–2. doi: 10.4049/jimmunol.174.11.6561
48. Seifert O, Matussek A, Sjogren F, Geffers R, Anderson CD. Gene expression profiling of macrophages: implications for an immunosuppressive effect of dissolucytotic gold ions. J Inflamm (Lond). (2012) 9:43. doi: 10.1186/1476-9255-9-43
49. Lacey DC, Achuthan A, Fleetwood AJ, Dinh H, Roiniotis J, Scholz GM, et al. Defining GM-CSF- and macrophage-CSF-dependent macrophage responses by in vitro models. J Immunol. (2012) 188:5752–65. doi: 10.4049/jimmunol.1103426
50. Conaghan PG, Cook AD, Hamilton JA, Tak PP. Therapeutic options for targeting inflammatory osteoarthritis pain. Nat Rev Rheumatol. (2019) 15:355–63. doi: 10.1038/s41584-019-0221-y
51. Tardito S, Martinelli G, Soldano S, Paolino S, Pacini G, Patane M, et al. Macrophage M1/M2 polarization and rheumatoid arthritis: a systematic review. Autoimmun Rev. (2019) 18:102397. doi: 10.1016/j.autrev.2019.102397
52. Han SA, Lee S, Seong SC, Lee MC. Effects of CD14 macrophages and proinflammatory cytokines on chondrogenesis in osteoarthritic synovium-derived stem cells. Tissue Eng Part A. (2014) 20:2680–91. doi: 10.1089/ten.tea.2013.0656
53. Zhuo Q, Yang W, Chen J, Wang Y. Metabolic syndrome meets osteoarthritis. Nat Rev Rheumatol. (2012) 8:729–37. doi: 10.1038/nrrheum.2012.135
54. Hansson GK, Robertson AK, Soderberg-Naucler C. Inflammation and atherosclerosis. Annu Rev Pathol. (2006) 1:297–329. doi: 10.1146/annurev.pathol.1.110304.100100
55. Steinberg D, Parthasarathy S, Carew TE, Khoo JC, Witztum JL. Beyond cholesterol. Modifications of low-density lipoprotein that increase its atherogenicity. N Engl J Med. (1989) 320:915–24. doi: 10.1056/NEJM198904063201407
56. Rios FJ, Koga MM, Pecenin M, Ferracini M, Gidlund M, Jancar S. Oxidized LDL induces alternative macrophage phenotype through activation of CD36 and PAFR. Mediators Inflamm. (2013) 2013:198193. doi: 10.1155/2013/198193
57. de Munter W, van der Kraan PM, van den Berg WB, van Lent PL. High systemic levels of low-density lipoprotein cholesterol: fuel to the flames in inflammatory osteoarthritis? Rheumatology (Oxford). (2016) 55:16–24. doi: 10.1093/rheumatology/kev270
58. Oliviero F, Lo Nigro A, Bernardi D, Giunco S, Baldo G, Scanu A, et al. A comparative study of serum and synovial fluid lipoprotein levels in patients with various arthritides. Clin Chim Acta. (2012) 413:303–7. doi: 10.1016/j.cca.2011.10.019
59. James MJ, van Reyk D, Rye KA, Dean RT, Cleland LG, Barter PJ, et al. Low density lipoprotein of synovial fluid in inflammatory joint disease is mildly oxidized. Lipids. (1998) 33:1115–21. doi: 10.1007/s11745-998-0313-8
60. Rutgers M, Saris DB, Vonk LA, van Rijen MH, Akrum V, Langeveld D, et al. Effect of collagen type I or type II on chondrogenesis by cultured human articular chondrocytes. Tissue Eng Part A. (2013) 19:59–65. doi: 10.1089/ten.tea.2011.0416
61. Engelholm LH, List K, Netzel-Arnett S, Cukierman E, Mitola DJ, Aaronson H, et al. uPARAP/Endo180 is essential for cellular uptake of collagen and promotes fibroblast collagen adhesion. J Cell Biol. (2003) 160:1009–15. doi: 10.1083/jcb.200211091
62. Martinez-Pomares L, Wienke D, Stillion R, McKenzie EJ, Arnold JN, Harris J, et al. Carbohydrate-independent recognition of collagens by the macrophage mannose receptor. Eur J Immunol. (2006) 36:1074–82. doi: 10.1002/eji.200535685
63. Madsen DH, Leonard D, Masedunskas A, Moyer A, Jurgensen HJ, Peters DE, et al. M2-like macrophages are responsible for collagen degradation through a mannose receptor-mediated pathway. J Cell Biol. (2013) 202:951–66. doi: 10.1083/jcb.201301081
64. Pap T, Korb-Pap A. Cartilage damage in osteoarthritis and rheumatoid arthritis–two unequal siblings. Nat Rev Rheumatol. (2015) 11:606–15. doi: 10.1038/nrrheum.2015.95
65. Mantovani A, Biswas SK, Galdiero MR, Sica A, Locati M. Macrophage plasticity and polarization in tissue repair and remodelling. J Pathol. (2013) 229:176–85. doi: 10.1002/path.4133
66. Inoki K, Li Y, Zhu T, Wu J, Guan KL. TSC2 is phosphorylated and inhibited by Akt and suppresses mTOR signalling. Nat Cell Biol. (2002) 4:648–57. doi: 10.1038/ncb839
67. Jones RG, Pearce EJ. MenTORing immunity: mTOR signaling in the development and function of tissue-resident immune cells. Immunity. (2017) 46:730–42. doi: 10.1016/j.immuni.2017.04.028
68. Byles V, Covarrubias AJ, Ben-Sahra I, Lamming DW, Sabatini DM, Manning BD, et al. The TSC-mTOR pathway regulates macrophage polarization. Nat Commun. (2013) 4:2834. doi: 10.1038/ncomms3834
69. Zhu L, Yang T, Li L, Sun L, Hou Y, Hu X, et al. TSC1 controls macrophage polarization to prevent inflammatory disease. Nat Commun. (2014) 5:4696. doi: 10.1038/ncomms5696
70. Baroja-Mazo A, Revilla-Nuin B, Ramirez P, Pons JA. Immunosuppressive potency of mechanistic target of rapamycin inhibitors in solid-organ transplantation. World J Transplant. (2016) 6:183–92. doi: 10.5500/wjt.v6.i1.183
71. Amano MT, Castoldi A, Andrade-Oliveira V, Latancia MT, Terra FF, Correa-Costa M, et al. The lack of PI3Kgamma favors M1 macrophage polarization and does not prevent kidney diseases progression. Int Immunopharmacol. (2018) 64:151–61. doi: 10.1016/j.intimp.2018.08.020
72. Festuccia WT, Pouliot P, Bakan I, Sabatini DM, Laplante M. Myeloid-specific Rictor deletion induces M1 macrophage polarization and potentiates in vivo pro-inflammatory response to lipopolysaccharide. PLoS ONE. (2014) 9:e95432. doi: 10.1371/journal.pone.0095432
73. Leyendecker A Jr, Pinheiro CCG, Amano MT, Bueno DF. The use of human mesenchymal stem cells as therapeutic agents for the in vivo treatment of immune-related diseases: a systematic review. Front Immunol. (2018) 9:2056. doi: 10.3389/fimmu.2018.02056
74. Fibbe WE, Nauta AJ, Roelofs H. Modulation of immune responses by mesenchymal stem cells. Ann N Y Acad Sci. (2007) 1106:272–8. doi: 10.1196/annals.1392.025
75. Chahal J, Gomez-Aristizabal A, Shestopaloff K, Bhatt S, Chaboureau A, Fazio A, et al. Bone marrow mesenchymal stromal cell treatment in patients with osteoarthritis results in overall improvement in pain and symptoms and reduces synovial inflammation. Stem Cells Transl Med. (2019) 8:746–57. doi: 10.1002/sctm.18-0183
76. Desando G, Cavallo C, Sartoni F, Martini L, Parrilli A, Veronesi F, et al. Intra-articular delivery of adipose derived stromal cells attenuates osteoarthritis progression in an experimental rabbit model. Arthritis Res Ther. (2013) 15:R22. doi: 10.1186/ar4156
77. Schelbergen RF, van Dalen S, ter Huurne M, Roth J, Vogl T, Noel D, et al. Treatment efficacy of adipose-derived stem cells in experimental osteoarthritis is driven by high synovial activation and reflected by S100A8/A9 serum levels. Osteoarthritis Cartilage. (2014) 22:1158–66. doi: 10.1016/j.joca.2014.05.022
78. Hamilton AM, Cheung WY, Gomez-Aristizabal A, Sharma A, Nakamura S, Chaboureau A, et al. Iron nanoparticle-labeled murine mesenchymal stromal cells in an osteoarthritic model persists and suggests anti-inflammatory mechanism of action. PLoS ONE. (2019) 14:e0214107. doi: 10.1371/journal.pone.0214107
79. Ylostalo JH, Bartosh TJ, Coble K, Prockop DJ. Human mesenchymal stem/stromal cells cultured as spheroids are self-activated to produce prostaglandin E2 that directs stimulated macrophages into an anti-inflammatory phenotype. Stem Cells. (2012) 30:2283–96. doi: 10.1002/stem.1191
80. Barminko JA, Nativ NI, Schloss R, Yarmush ML. Fractional factorial design to investigate stromal cell regulation of macrophage plasticity. Biotechnol Bioeng. (2014) 111:2239–51. doi: 10.1002/bit.25282
81. Heldens GT, Blaney Davidson EN, Vitters EL, Schreurs BW, Piek E, van den Berg WB, et al. Catabolic factors and osteoarthritis-conditioned medium inhibit chondrogenesis of human mesenchymal stem cells. Tissue Eng Part A. (2012) 18:45–54. doi: 10.1089/ten.tea.2011.0083
Keywords: M1/M2 macrophages, cartilage regeneration, synovial inflammation, mesenchymal stem cells, osteoarthritis, articular cartilage, cell therapy
Citation: Fernandes TL, Gomoll AH, Lattermann C, Hernandez AJ, Bueno DF and Amano MT (2020) Macrophage: A Potential Target on Cartilage Regeneration. Front. Immunol. 11:111. doi: 10.3389/fimmu.2020.00111
Received: 30 September 2019; Accepted: 15 January 2020;
Published: 11 February 2020.
Edited by:
Jagadeesh Bayry, Institut National de la Santé et de la Recherche Médicale (INSERM), FranceReviewed by:
Xiaoxuan Zhuang, National Institute of Allergy and Infectious Diseases (NIAID), United StatesCopyright © 2020 Fernandes, Gomoll, Lattermann, Hernandez, Bueno and Amano. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tiago Lazzaretti Fernandes, dGlhZ28ubGF6emFyZXR0aUB1c3AuYnI=
†ORCID: Tiago Lazzaretti Fernandes orcid.org/0000-0002-6665-3608
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.