 Marie-Laure Golinski1,2
Marie-Laure Golinski1,2 Mélanie Demeules1
Mélanie Demeules1 Céline Derambure3
Céline Derambure3 Gaetan Riou1
Gaetan Riou1 Maud Maho-Vaillant1,2
Maud Maho-Vaillant1,2 Olivier Boyer1,4
Olivier Boyer1,4 Pascal Joly1,2
Pascal Joly1,2 Sébastien Calbo1*
Sébastien Calbo1*- 1INSERM U1234, Normandy University, Rouen, France
- 2Department of Dermatology, Rouen University Hospital, Rouen, France
- 3INSERM U1245, Normandy University, Rouen, France
- 4Department of Immunology, Rouen University Hospital, Rouen, France
CD11c+ B cells have been reported to be increased in autoimmune diseases, but they are detected in the blood of healthy individuals as well. We aimed to characterize CD11c+ B cells from healthy donors by flow cytometry, microarray analysis, and in vitro functional assays. Here, we report that CD11c+ B cells are a distinct subpopulation of B cells, enriched in the memory subpopulation even if their phenotype is heterogeneous, with overexpression of genes involved in B-cell activation and differentiation as well as in antigen presentation. Upon activation, CD11c+ B cells can differentiate into antibody-secreting cells, and CD11c could be upregulated in CD11c− B cells by B-cell receptor activation. Finally, we show that patients with pemphigus, an autoimmune disease mediated by B cells, have a decreased frequency of CD11c+ B cell after treatment, relative to baseline. Our findings show that CD11c+ B cells are mainly memory B cells prone to differentiate into antibody secreting cells that accumulate with age, independently of gender.
Introduction
Integrins are a family of heterodimeric cell adhesion receptors with function in cell migration, survival, and proliferation (1). CD11c (ITGAX gene) is the integrin αX chain protein, which combines with the β2 chain (CD18; ITGB2 gene). The CD11c/CD18 complex binds complement iC3b-coated particles to induce phagocytosis (2), cell adhesion molecules [intercellular adhesion molecule (ICAM)-1, ICAM-2, ICAM-4, and vascular cell adhesion molecule 1] (3, 4), lipopolysaccharide (5), and collagen and fibrinogen (6, 7). CD11c is abundantly expressed on monocytes, granulocytes, on tissue macrophages, on a subset of dendritic cells, and at low level on neutrophil, but can be expressed as well on a subset of B cells, T cells, and NK cells, with expression level varying from dim to bright (8).
CD11c+ B cells are reported to be increased in autoimmune diseases as rheumatoid arthritis (RA) (9), Sjogren's syndrome (10), multiple sclerosis (11), and systemic lupus erythematosus (SLE) (12), or after malarial infection (13). CD11c+ B cells accumulate with age in mice and in women with RA and are thus known as “age-associated B cells” (ABC) (9). However, CD11c+ B cells are detected in young healthy donors (HD) as well (14). This broad range of conditions could reflect their state of activation depending on the disease or age and might explain why cell surface expression of CD11c+ B cells has been reported to be heterogeneous, corresponding to: CD27+ memory cell (9), double-negative (DN) IgD−CD27− (13), CD21low (10), and T-bet+ (12, 15) or T-bet−, at least in mice (16). In addition, we reported previously that following toll-like receptor 9 (TLR9) and B-cell receptor (BCR) stimulation, CD11c+ B cells from HD were unable to secrete interleukin (IL)-10 (14), while IL-10 is one of the highest messenger RNA (mRNA) expressed found in CD11c+ B cells from SLE patient (12).
In this study, we compared the phenotype, frequency depending on HD's age, gene expression profile, mRNA cytokine expression, and capacity of CD11c+ vs. CD11c− B cells to differentiate into plasma cells. Despite the fact that CD11c expression was detected in all B cell subpopulations, our findings demonstrate that CD11c+ B cells are enriched in memory B cells, which explains their strong ability to differentiate into antibody-secreting cells. Moreover, we show that CD11c upregulation is controlled by BCR stimulation. We report an increase in the frequency of CD11c+ B cells from blood in Pemphigus, an autoantibody-mediated disease targeting the skin and the mucous membranes, which return to HD's level following treatment.
Materials and Methods
Phenotyping of PBMCs
Peripheral blood mononuclear cells (PBMC) were isolated from buffy coats obtained via the French blood bank using Ficoll–Paque density gradient centrifugation (GE Healthcare) (authorization number: PLER-UPR/2018/014). In addition, PBMCs from pemphigus patients from the clinical trial number NCT00784589 were used. This study was approved by the Ethics Committee of the North West in France and conducted according to the Declaration of Helsinki principles. HD age was 20–35 years old, which is the age group that most often donates large volume of blood in our area, unless specified. Representative frequency's examples depicted in Figures 1–7 were obtained from donors in this age group, which is the group of age with the lowest frequency of CD11c+ B cells according to Figure 1E.
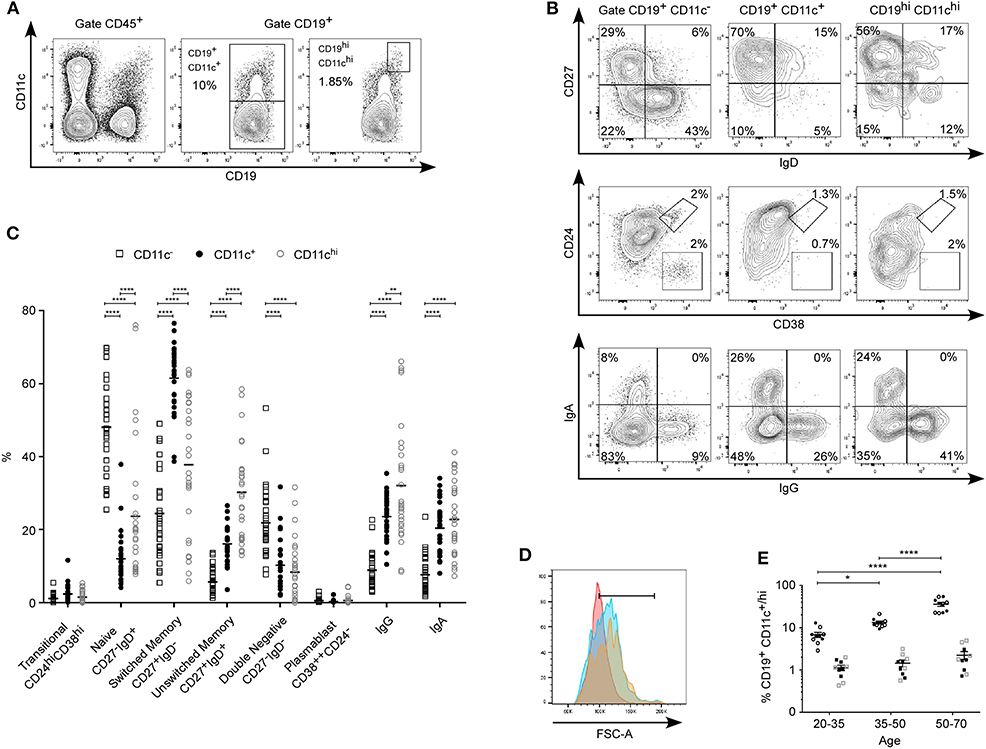
Figure 1. Phenotyping of human CD11c+ B cells. (A) Expression level of CD11c and CD19 and gating strategy to study CD19+CD11c−, CD19+CD11c+, or CD19+CD11chi with one representative frequency. (B) CD27, IgD, CD24, CD38, IgA, and IgG expression on CD19+CD11c−, CD19+CD11c+, or CD19+CD11chi with one representative frequency, and (C) proportion of transitional B cells, naive, switched memory, unswitched memory, double negative, plasmablast, IgG+, and IgA+ for n = 30 healthy donors. (D) Forward scatter histogram overlay for CD19+ CD11c− (pink), CD11c+ (blue), and CD11chi (yellow). The gate is used to determine the Geo mean fluorescence intensity, which is 0.98, 10.8, and 11.3 × 104, respectively. (E) Percentage of CD11c+ and CD11chi B cells for donors between age 20 and 35, 35 and 50, and 50 and 70 years old (n = 10 for each group; circle = CD11c+ B cells, square = CD11chi B cells, open symbol = woman, fill symbol = man). Significant difference is determined by two-way ANOVA with correction by Sidak's multiple comparison test in (C) and with correction by Tukey's multiple comparison test in (E). *p < 0.05, **p < 0.01, ****p < 0.0001. Dot plots from (A,B), and histogram from (D) were obtained from a donor age 32.
Phenotype analysis was performed with the cytometer FortessaTM (Becton Dickinson) using the following markers: LIVE/DEAD® Fixable Blue Dead Cell Stain (Invitrogen), Fc Receptor Blocking Solution (Human TruStain FcX, Biolegend), CD19-PE-Cy7 (clone Hib19, eBioscience), CD11c-PE or APC (clone Bu15, Biolegend), IgA-VioBright-FITC (clone IS11-8E10, Miltenyi), CD27-BV421 (clone M-T271, Becton Dickinson), IgD-AF700 (clone IA6-2, Becton Dickinson), CD38-PerCP-Cy5.5 (clone HIT2, Becton Dickinson), CD24-PE-CF594 (clone ML5, Becton Dickinson), IgG-BV510 (clone G18-145, Becton Dickinson), IgM-BV605 (clone G20-127, Becton Dickinson), CD138-BV711 (clone MI15, Becton Dickinson), CD45-BV785 (clone HI30, Sony), and CD20-APC (clone 2H7, Sony).
To confirm the microarray data, PBMC from five different HD were labeled with the following antibodies: LIVE/DEAD® Fixable Blue Dead Cell Stain (Invitrogen), Fc Receptor Blocking Solution (Human TruStain FcX, Biolegend), CD19-PeCy7, CD11c-PE or APC, CD1c-BV421 (clone L161, Biolegend), CD58-PeCy5 (clone TS2/9, Biolegend), CD84-PE (clone CD84.1.21, Biolegend), CD27-BV421, CD86-BV510 (clone IT2.2, Biolegend), CD95-FITC (clone DX2, Biolegend), CD6-FITC (clone BL-CD6, Biolegend), CD200-BV605 (clone OX104, Biolegend), CD80-BV650 (clone 2D10, Biolegend), CD21-PE (clone HB5, eBioscience) CD274-BV711 (clone 29E.2A3, Biolegend), CD68-PerCP-Cy5.5 (clone Y1/821, Biolegend), IL-27/IL-35 EBI3-PE (clone B032F6, Biolegend), IL-1β-PE (clone AS10, Becton Dickinson), IFN-γ PE (clone 45.15, Beckman Coulter), IL-10 PE (BT-10, Miltenyi), IL-6 PE (clone MQ2-13A5, eBioscience), and BCMA-APC (polyclonal, R&D). All cytometry data were analyzed using FlowJo software (TreeStar Inc). Fluorescence-minus-one controls were used to compensate all flow cytometry data (17).
Isolation of CD11c+ B Cells
B cells were isolated from PBMC using Dynabeads Untouched Human B cells kit (Life Technologies) following the manufacturer's instructions. More than 90% of the isolated cells were CD19+. B cells were suspended at 10.106 cells/ml in cold buffer and further stained with LIVE/DEAD Fixable Aqua Dead Cell Stain Kit (Life Technologies), Fc Receptor Blocking Solution (Human TruStain FcX, Biolegend), anti-CD19 Pe-Cy7 (clone HIB19, Biolegend), and anti-CD11c APC (clone BU15, Biolegend). CD11c+ and CD11c− CD19+ B cells were sorted using BD FACSAria III 4-Laser (BD Biosciences). The purity of CD11c+ and CD11c− B cells was checked after each sorting and was mean 95.3% (±3.4%) and mean 99.3% (±0.4%), respectively (Figure 2). In total, we have performed 26 cell sorting, from 26 different HD. Owing to the low frequency of CD11c+ B cells and the cell sorter's setting to purity (which decreased the yield), B cells were purified from buffy coat, for which the donors are predominantly between 20 and 35 years old in our area. On average, out of 722 ± 301 million PBMC (n = 26), we end up with 46 ± 30 million purified B cells that were used for sorting. CD11c+ B cells (80,000–1.4 million) were obtained (mean, 5.9 × 105 ± 4.3 × 105 cells, n = 26) depending on the donor. Owing to the low number of CD11chi B cells expected recovery, which represent, on average, 1.6 ± 1.1% of CD19+ B cells as shown in Figure 1E (n = 30), CD11chi B cells were included in the sorting of CD11c+ B cells.
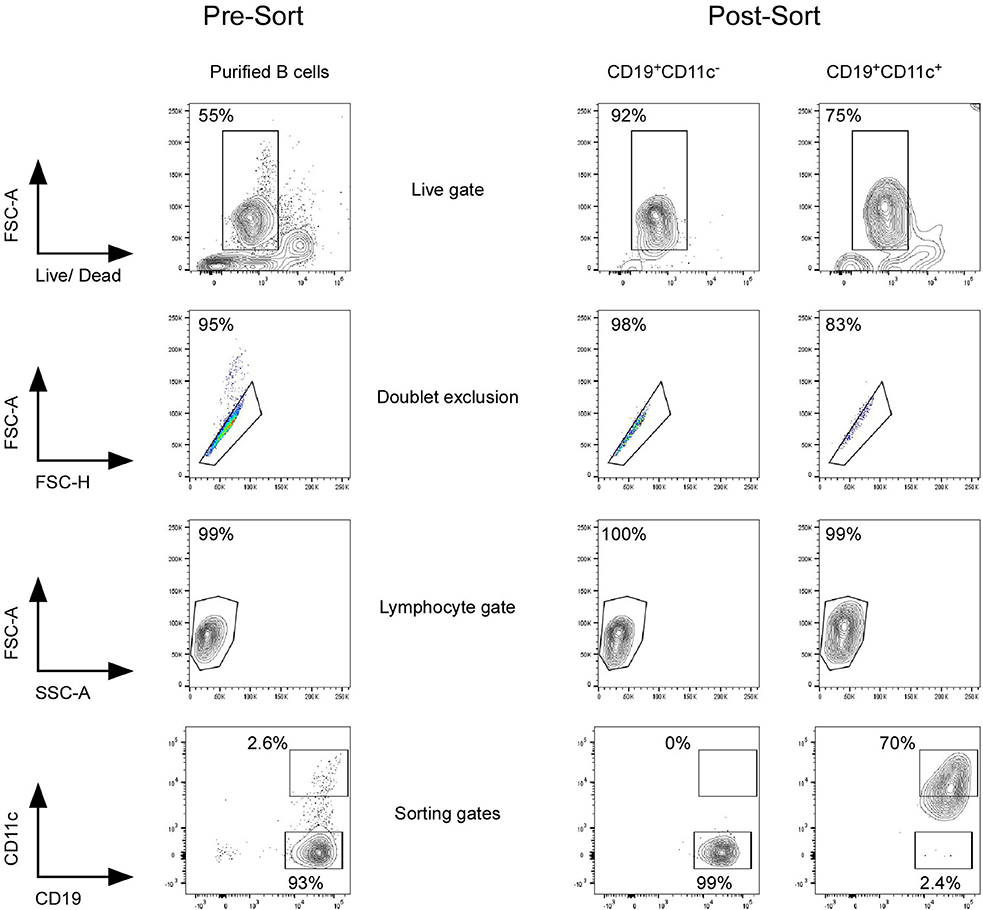
Figure 2. Sorting strategy of CD11c+ and CD11c− B cells. Purified B cells by negative selection using magnetic beads were stained with LIVE/DEAD Fixable Aqua Dead Cell Stain Kit, Fc Receptor Blocking Solution, and anti-CD19 and anti-CD11c antibodies. CD11c+, including CD11chi and CD11c− CD19+ B cells, were sorted using FACSAria III. Purity of CD11c+ and CD11c− B cells was checked after each sorting and was mean 95.3% (±3.4%) and mean 99.3% (±0.4%), respectively. Twenty-six different HD were sorted with age between 20 and 35 years old.
In vitro Culture of B Cells
CD11c+ and CD11c− B cells were cultured at a cell density of 106 cells/ml in Roswell Park Memorial Institute 1640 supplemented with 10% fetal bovine serum, 2 mM L-glutamine, 100 U/ml penicillin, and 100 μg/ml streptomycin (all reagents from Thermo Fisher Scientific). B cells were left unstimulated or stimulated with 3 μg/ml CpG-B 2006 (InvivoGen), 10 μg/ml goat anti-human immunoglobulin A (IgA) IgG IgM (H + L) (anti-Ig; Jackson ImmunoResearch Laboratories) or 1 μg/ml R848 for 48 h at 37°C with 5% CO2. For plasmocyte differentiation and antibody production assay, CD11c+ and CD11c− B cells were cultured at a cell density of 2.105 cells/ml in the above media and were stimulated with 6 μg/ml CpG-B 2006, 50 ng/ml IL-21, and Staphylococcus aureus Cowan I cells (SAC, 1×; Pansorbin cells, Calbiochem) for 7 days at 37°C with 5% CO2, unless otherwise stated. Quantity of IgM and IgG secreted were measured by ELISA (READY-SET-GO! Kit, eBioscience). In Figure 5E, PBMC from 10 HD were cultured in 96-well plate U bottom in full media in the presence of Brefeldin A for 4 h, at 2 × 106 cells/ml, without any stimulation. Cells were then stained with Live and Dead, anti-CD45, anti-CD19, and anti-CD11c, washed, fixed, and permeabilized, and stained with different cytokine-specific antibodies that are commercially available. Settings were done using PMA + ionomycin-stimulated cells and fmo.
RNA Preparation
Total RNAs from CD11c+ or CD11c− B cells of 15 HD [5 donors for transcriptome analysis and 10 donors for quantitative PCR (qPCR)] were extracted with RNeasy Mini kit according to the manufacturer's recommendations (Qiagen). The quality and quantity of isolated mRNAs were assessed using the 2100 Bioanalyzer (Agilent Technologies) and the Nanodrop device (Thermo Scientific). All RNA samples had an RNA integrity number of ≥8.2 ± 0.4.
Microarrays
Whole human genomic DNA microarrays were used to perform one-colored gene expression profiling (4 × 44K Whole Human Genome, Agilent Technologies). One hundred nanogram for each CD11c+ or CD11c− B cells sample from 5 HD were labeled by Cyanine-3 according to the manufacturer's instructions (Low Input QuickAmp Labeling Kit, Agilent Technologies). Hybridization was performed at 65°C for 17 h using a hybridization kit (Agilent Technologies). After wash steps, microarrays were scanned with a 5-μM pixel size using the DNA Microarray Scanner GB (Agilent Technologies). Image analysis and extraction of raw data were performed with Feature Extraction Software 10.5.1.1 (Agilent Technologies). Data were normalized using 75 percentile shift and baseline transformed by median of all samples. Only spots that passed these quality controls on 100% of arrays in at least one of two conditions (CD11c+ or CD11c−) were selected for further analysis. Hierarchical clustering was performed with Pearson coefficient metric and Wards linkage to build the transcripts and sample dendrograms. Data were in agreement with the guidelines for Minimum Information About a Microarray Experiment and were deposited in the database of the National Center for Biotechnology Information's Gene Expression Omnibus (https://www.ncbi.nlm.nih.gov/geo/). Data are accessible using the following accession number: GSE112515. Non-uniform and saturated spots or spots with intensities below the background were not taken into account.
Functional Analysis of Microarrays
Data from transcriptomic analysis were analyzed with GeneSpring GX V.13.0 (Agilent Technologies). Paired Student's t test (p < 0.05) with Benjamini–Hochberg correction to check the false discovery rate was used to determine the statistical significance in gene expression levels between CD11c+ and CD11c− B cells. The Gene Ontology (GO) analysis was used to investigate the biological processes, molecular function, or cellular localization enriched in the transcripts list showing a significant fluctuation in gene expression between CD11c+ and CD11c− B cells. p value was computed by standard hypergeometric distribution. The GeneSpring Single Experiment Analysis bioinformatics tool was used for computational analysis to identify potential curated canonical pathways with setting parameters (Reactome and GenMAPP for pathway source), which are enriched in the differentially expressed transcripts list, using WikiPathways database (http://www.wikipathways.org). The significance of the association between the genes and the pathways was measured by Fisher's exact test (minimal number of entities in pathways ≥6 and p ≤ 0.001). Text mining was performed with GeneSpring GX V.13.0 (direct interactions with relation score ≥9 and relation types chosen: binding, expression, member, metabolism, promoter binding, protein modification, regulation, and transport).
Quantitative Real-Time Polymerase Chain Reaction
Three nanograms of each RNA sample were retro-transcribed and preamplified in a 96-well plate containing 0.2 μl Platinum Taq polymerase and SuperScript III reverse transcriptase (Invitrogen), a mixture of Taqman primer probes specific for the transcripts of interest (18), and 5 μl CellsDirect quantitative real-time PCR buffer (Invitrogen). After centrifugation, samples were incubated at 50°C for 15 min for reverse transcription, followed by 20 cycles of 95°C, 15 s and 60°C, 4 min for amplification. Subsequent preamplified complementary DNA (cDNA) was stored at −20°C until analysis. After one-fourth dilution in Tris–EDTA buffer, each cDNA sample was then separated into 48 separate reactions for further qPCR using the BioMark 48.48 dynamic array nanofluidic chip (Fluidigm, Inc.) as described in (19). Wells with no RNA were used as negative controls. Data were analyzed using Real-Time PCR Analysis software with normalization of the Ct value for each gene using the mean of five housekeeping genes (GAPDH, β2-M, TUBB, GUSB, and HPRT1) as calibrator, using geNorm (20). We considered that gene was expressed if Ct value was <40, and expression curve was a sigmoid. To compare expression level between samples, a standard curve was first created with a pool of cDNA from all samples. From this pool, serial dilutions were performed, and qPCR values were assigned arbitrary values (6 points ranging from 100 to 0.4). The gene expression level of samples was obtained using this standard curve.
Statistical Analyses
All experiments were performed using at least three different cell cultures or blood donors in independent experiments. GraphPad Prism 7 was used to create the figures. Student's t test and the Wilcoxon paired t test were used to assess normally distributed data and non-normally distributed data, respectively. One-way ANOVA with correction by Bonferroni's posttest, two-way ANOVA with correction by Sidak's posttest, and one-way ANOVA with Dunnett's posttest were used. p < 0.05 was considered significant.
Results
Phenotyping of Human CD11c+ B Cell in Blood of HD
We used HD's blood from the French blood bank, in an age group between 20 and 70 years and with an equivalent number of men and women. Using flow cytometry, expression level of CD11c on blood monocytes and B cells from HD was equivalent, fluctuating from dim to bright (Figure 1A) with percentages of CD19+CD11c+ varying from 3 to 55% (mean, 19 ± 3, n = 30). Among CD11c+ B cells, a small subpopulation, bright for CD19 and CD11c, was detected (1.6 ± 0.2%; range, 0.4–5%, n = 30). First, CD11c+ B cells were compared with CD11c− B cells. We found that CD11c+ B cells were significantly enriched in the switched memory population (CD27+IgD−: 62 ± 2% for CD11c+ vs. 24 ± 2% for CD11c−, p < 0.0001), as in the unswitched memory population (CD27+IgD+: 16 ± 1 vs. 6 ± 1%, p < 0.0001) (Figures 1B,C). In addition, CD11c+ B cells had a significantly higher proportion of IgG+ and IgA+ (IgG+, 24 ± 1 vs. 9 ± 1%, p < 0.0001; IgA+, 20 ± 1 vs. 8 ± 1%, p < 0.0001). CD11c+ B-cell frequency was reduced in the naive (CD27−IgD+, 12 ± 1 vs. 48 ± 2%, p < 0.0001) and in the DN population (CD27−IgD−, 10 ± 1 vs. 22 ± 2%, p < 0.0001). No significant difference in frequency was observed between CD11c+ vs. CD11c− in plasmablast (CD38++CD24−: 0.3 ± 0.1 vs. 0.5 ± 0.1%) nor in transitional B cell population (CD24hiCD38hi, 2.3 ± 0.4 vs. 1.1 ± 0.2%; p = NS). Second, CD11c+ B cells were compared to CD11chi B cells. Interestingly, the CD11chi population was enriched in the naive (24 ± 3%, p < 0.0001) and the unswitched memory population (30 ± 2%, p < 0.0001), and decreased in the switched memory population (38 ± 3%, p < 0.0001) (Figures 1B,C), suggesting a retrograde control of CD11c expression during B-cell ontogeny and differentiation. No significant difference was observed between CD11chi and CD11c+ frequencies in the transitional, DN, and plasmablast populations (respectively for CD11chi: 1.5 ± 0.3, 8.4 ± 1.5, and 0.6 ± 0.2%). However, CD11chi B cells had a significantly higher proportion of IgG+ (IgG+, 32 ± 2.9%, p < 0.01). Of note, CD11chi B cells had a higher proportion of larger cells than CD11c+ and the CD11c− B cells (Figure 1D), suggesting a different stage of differentiation or activation (forward scatter-A geometric mean fluorescence intensity = 111.57, 104.31, 95 × 103 for CD11chi, CD11c+, and CD11c− B cells, respectively. p < 0.0001, n = 30).
Since CD11c+CD27+ B cells have been described to accumulate in aged women with RA (9), we assessed CD11c+ B-cell frequency depending on age and gender. Figure 1E shows that CD11c+ B-cell frequency increased with age from 7 ± 1% in HD aged 20–35 years, up to 35 ± 5% in HD aged 50–70 years (p < 0.0001), with no difference depending on gender. However, no difference was observed for CD11chi B-cell frequency between age groups.
Overall, CD11c was expressed in all B cell subpopulations, suggesting a role in all B-cell development steps, with an increased frequency in antigen-driven B cells, which accumulate with age of HD, independently of gender.
Gene Expression Profile
To better define the cellular characteristics of circulating CD11c+ B cells, we then performed a microarray analysis of CD11c+ vs. CD11c− B cells from five HD's blood. After negative selection using magnetic bead, purified B cells from buffy coat were sorted based on CD11c expression using BD FACSAria III. Purity of CD11c+ and CD11c− B cells was checked after each sorting and was 95.3% (±3.4%) and 99.3% (±0.4%), respectively (Figure 2).
To identify the differentially expressed genes (DEG) between paired CD11c+ and CD11c− B cells, total RNA were hybridized on whole human genome microarrays by one-color technology and analyzed with GeneSpring GX V.13.0 (Agilent Technologies). Out of 19,905 transcripts that passed the quality filters after exclusion of spikes and flagged probes, a fold change cut-off of 2 between CD11c+ and CD11c− samples identified 1,894 transcripts. To highlight the most discriminant transcripts, we applied a fold change cut-off of 3. After paired t test with Benjamini–Hochberg correction for multiple testing (p < 0.05), removal of duplicate probes, pseudogenes, and uncharacterized loci, 307 DEG were found to be upregulated and 205 downregulated in CD11c+ compared with CD11c− B cells (Supplemental Table 1).
GO analysis was performed with the 512 DEG and identified 145 biological processes, which mainly corresponded to regulation of cell activation (GO:0050865; p = 7.44 × 10−12), regulation of multicellular organismal process (GO:0051239; p = 5.41 × 10−11), regulation of immune system process (GO:0002682; p = 1.83 × 10−9), regulation of cell adhesion (GO:0030155; p = 1.49 × 10−8), and response to stimulus (GO:0050896; p = 2.25 × 10−9). To find molecular functions enriched in these DEG set, Wiki and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analyses were performed using the Single Experiment Analysis tool from GeneSpring software and DAVID Bioinformatics Resources (21). Pathways related to TLR signaling (WP1449-45049, p = 4.2 × 10−4; WP75-46210, p = 0.001; KEGG 04620, p = 0.029), inflammatory response (WP453-41201, p = 1.02 × 10−5), cell adhesion molecules (KEGG 04514, p = 0.021), FoxO signaling pathway (KEGG 04068, p = 0.034), and cytokine signaling pathway (KEGG 04060, p = 0.04) were listed. These findings indicate that CD11c+ B cells have a clear distinctive gene expression profile related to active immune response.
Differential Expression of CD1c, CD58, CD68, CD84, CD86, CD27, CD274, Fas, and CD200 on Human CD11c+ B Cells
Out of 512 DEG, we selected 12 transcripts coding for cell surface receptors. CD6 and CD200 were underexpressed, and CD11c (ITGAX), CD1c, CD84, CD68, CD27, Fas (CD95), CD80, CD58, CD274, and CD86 were overexpressed in CD11c+ B cells vs. CD11c− B cells. Principal component analysis (Figure 3A) and hierarchical clustering (Figure 3B) performed with this minimal combination of 12 transcripts were sufficient to accurately discriminate CD11c+ from CD11c− B cells.
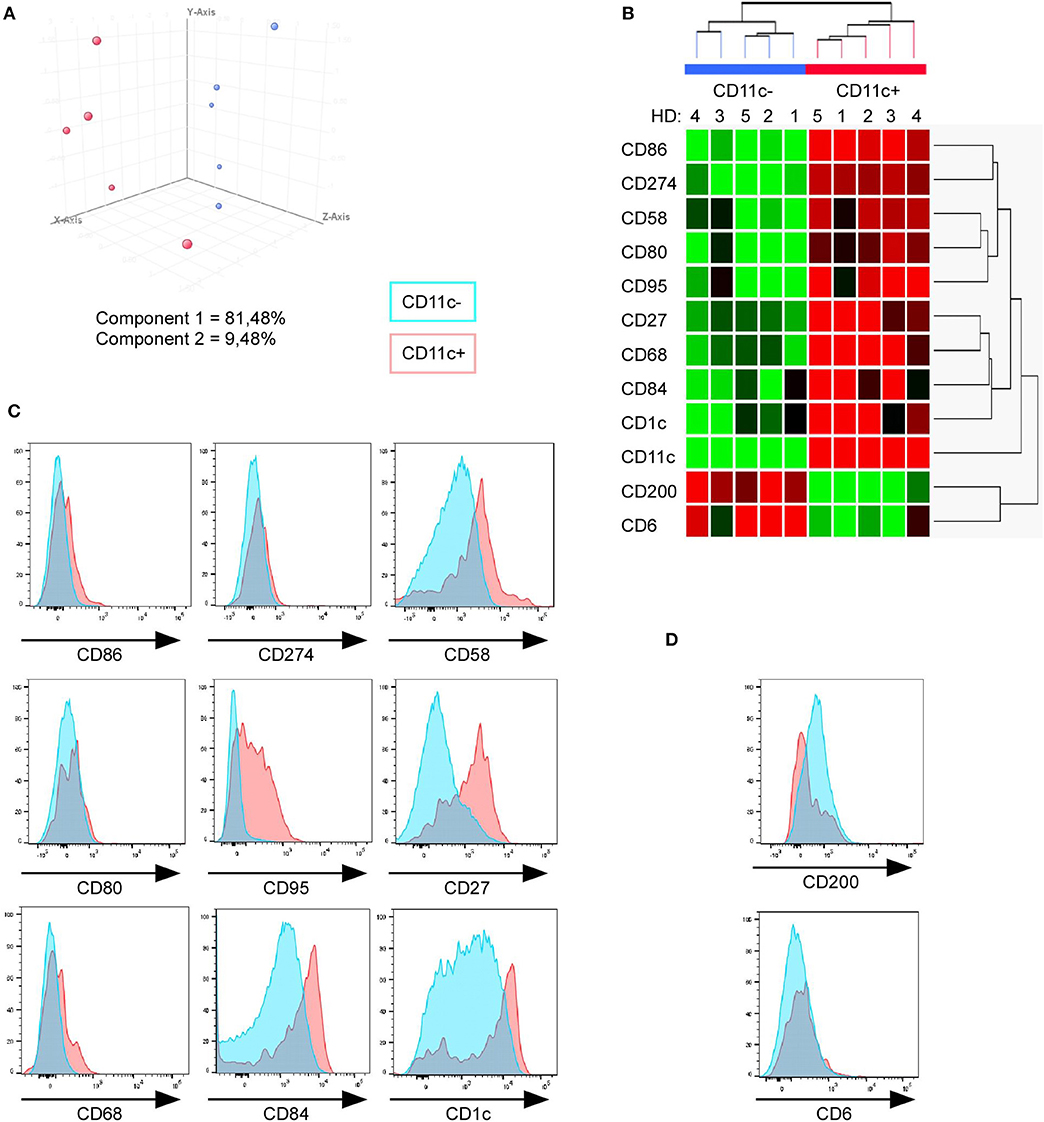
Figure 3. Minimal combination of 12 cell surface markers perfectly discriminate CD11c+ from CD11c− B cells. (A) Principal component analysis of 10 samples (paired CD11c+ and CD11c− B cells from five HD) performed with gene expression level of the 12 mRNA corresponding to cell surface markers among the 512 DEG between CD11c+ (red dots) and CD11c− (blue dots) B cells. (B) Hierarchical clustering shows that a minimal combination of 12 transcripts is sufficient to perfectly discriminate both populations. (C,D) FACS analysis of CD11c+, including CD11chi (red) and CD11c− (blue), was performed to validate this combination of markers at the protein level (one representative result of three independent experiments).
We then evaluated by fluorescence-activated cell sorting (FACS) if these 11 markers (excepting CD11c) could discriminate CD11c+ from CD11c− B cells as suggested by the microarray data. We found that CD11c+ B cells overexpressed CD86, CD58, CD95, CD27, CD68, CD84, and CD1c in comparison with CD11c− B cells (Figure 3C), even if the expression level for these markers was heterogeneous, which might suggest a specific level of expression by the different CD11c+ B cell subpopulations. On the other hand, CD200 expression was lower in a fraction of CD11c+ B cells compared with CD11c− B cells (Figure 3D). No difference was observed for the CD274, CD80, and CD6 markers, which were weakly expressed by B cells (Figures 3C,D). Overall, the differences in the mRNA expression level between CD11c+ and CD11c− B cells were confirmed at the protein level for 8 out of 11 markers, although the expression of these markers was not strictly associated with the CD11c+ B cell population. However, text mining from bibliographic data performed with natural language processing algorithm showed that 9 out of 12 cell surface receptors (CD11c, CD1c, CD27, CD58, CD68, CD80, CD86, CD95, and CD274) were related and constituted a functional network associated with CD11c (Figure 4).
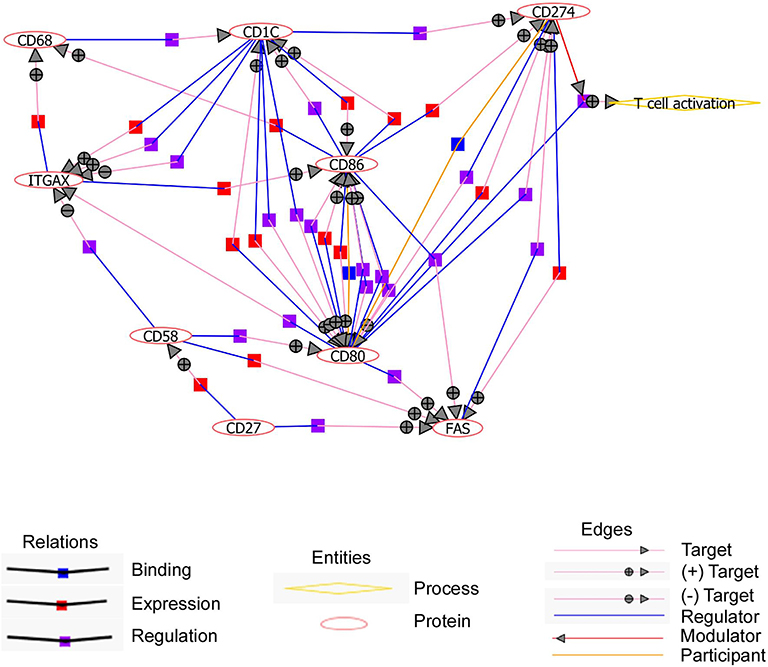
Figure 4. Upregulated cell surface markers on CD11c+ B cells interplay to favor T cell activation. Text mining performed with natural language processing (NLP) algorithm shows that 9 out of 12 cell surface receptors are connected all together. Pictograms representing relations, entities, and edges are shown. Circles with + and – symbols represent positive and negative regulations, respectively. Symbols (blue, red, and purple squares) show different modes of relations like binding, expression, and regulation.
Cytokine Gene Expression of CD11c+ B Cells
To characterize the cytokines expressed by CD11c+ and CD11c− B cells, we analyzed the expression of 25 cytokines known to be secreted by B cells, in addition to 5 housekeeping genes and 1 transcription factor (19), using Biomark HD real-time PCR. Sorted B cells from 10 HD were analyzed. EBI3, IL7, IL1β, IL12p40, IFNγ, Baff, and IL10 were found to be significantly upregulated, while IL6, IL15, IL23p19, and BCMA were found significantly downregulated in ex vivo purified CD11c+ B cells (Figures 5A,B). qPCR analyses confirmed the significantly different expression of EBI3 and IL6 observed using microarray, with a fold change > 3 (Supplemental Table 1) and that of IL7, IL15, and IL23p19 with a fold change > 2 (data not shown). No significant difference in expression of the genes, April, TNFα, IL1RA, IL12p35, TNFβ, Trail, TGFβ2, CD19, Taci, and BAFF-R, was observed between CD11c+ and CD11c− B cells (data not shown). IL2, IL5, IL9, IL13, IL17A, IL17F, IL27p28, and IL21 mRNAs were not detected by qPCR. To confirm cytokine preferential expression at the protein level, PBMC from 10 HD were cultured in the presence of Brefeldin A for 4 h, without any stimulation, and then stained intracellularly with commercially available antibodies specific for: EBI3, IL-1β, IFN-γ, IL-10, IL-6, and BCMA. We confirmed a significant preferential expression for EBI3, IL-1β, IFN-γ, and IL-10 at the protein level for CD11c+ B cells (n = 10) and a tendency for a lower expression of BCMA, even if the percentage of stained cells was very low (Figure 5E). However, we observed a higher percentage of IL-6+ cells among CD11c+ B cells than CD11c− B cells, which is the opposite of what we observed at the RNA level. This discrepancy could be due to the instability of IL6 mRNA (22) or by a regulatory IL-1β feedback loop as IL-6 is induced by IL1-β (23).
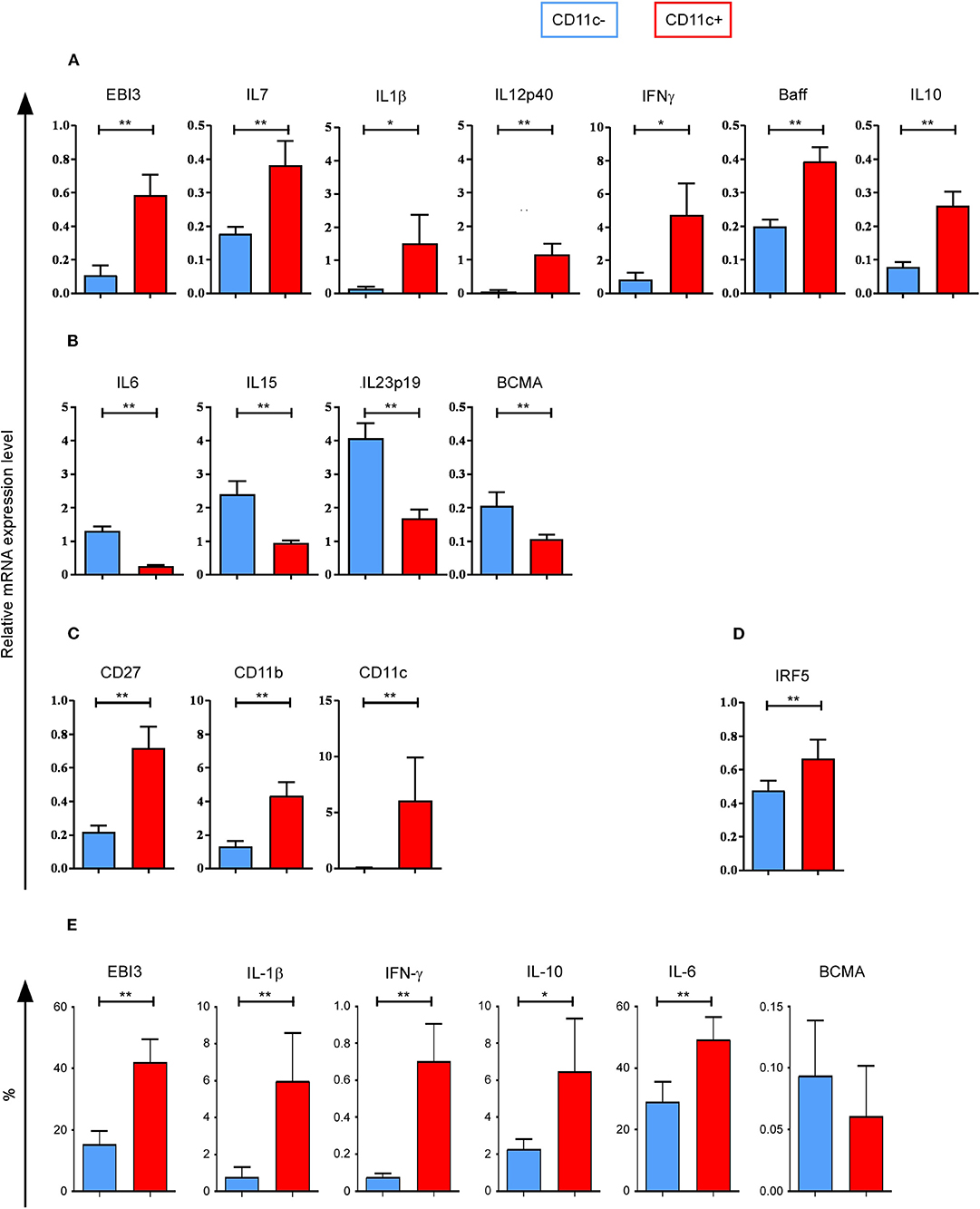
Figure 5. Cytokine profile of CD11c+ B cells as well as CD27, CD11b, CD11c, and IRF5 messenger RNA (mRNA) expression. Quantitative PCR (qPCR) analysis of cytokine genes for CD11c+ and CD11c− B cells from 10 healthy donors (HD) shows (A) upregulated genes and (B) downregulated gene expression levels for cytokine genes. (C) CD11c+ B cells upregulated CD27, CD11b, and CD11c gene coding for cell surface markers compared to CD11c− B cells. (D) IRF5 gene expression level is shown. (E) Peripheral blood mononuclear cells (PBMC) from 10 HD were cultured in the presence of Brefeldin A for 4 h, without any stimulation, and then stained intracellularly with antibodies specific for: Epstein–Barr virus-induced 3 (EBI3), interleukin (IL)-1β, interferon gamma (IFN-γ), IL-10, IL-6, and B-cell maturation antigen (BCMA). Percentage of cytokine expressing cells for CD11c+ and CD11c− B cells are shown. Bar graphs show mean ± SEM of relative expression, compared using the Wilcoxon paired t test: *p < 0.05; **p < 0.01.
We confirmed that CD27 was significantly upregulated in CD11c+ B cells, as well as CD11b and CD11c (Figure 5C). The transcription factor IRF5 was also significantly upregulated in CD11c+ B cells (Figure 5D), which was confirmed by microarray data with a fold change > 2 (data not show).
In short, cytokine genes overexpressed in CD11c+ B cells corresponded to both pro-inflammatory (IFN-γ, EBI3, IL-1β, IL-12p40) and anti-inflammatory (EBI3, IL-10) cytokines, and cytokines involved in B-cell survival (IL-7, Baff, IL-10), activation, and differentiation (IL-1β, IFN-γ) as well as in isotype switch (EBI3, IFN-γ, and IL-10) (24–26).
Differentiation of CD11c+ B Cells Into Plasma Cells
Since CD11c+ B cells were enriched in the memory subpopulation and upregulated IRF5, which is involved in the differentiation into plasma cells (27, 28), we assessed the ability of CD11c+ B cells to differentiate into antibody-secreting cells. CD11c+ and CD11c− were cultured without or with stimulation (CPG + SAC + IL-21) for 7 days. Plasmablast and plasmocyte differentiation was evaluated by FACS using CD27+CD38+CD138− and CD27+CD38+CD138+ expression, respectively (Figure 6A) (27). A higher proportion of CD11c+ than CD11c− B cells was able to differentiate into plasmablasts (31 ± 10 vs. 7 ± 3%; n = 6, p < 0.05) and plasmocytes (8 ± 3 vs. 1 ± 0.2%; p < 0.04) (Figure 6B). Interestingly, the number of CD11c− B cells upregulating the plasmocyte-specific marker CD138 did not increase, even after a longer stimulation time of 9 days (data not shown). On the opposite, for a shorter stimulation time of 5 days, 29% (±17.1%) CD11c+ B cells differentiate into plasmablasts vs. 6.3% (±2.4%) for CD11c− B cells, and 3.1% (±2.7%) CD11c+ B cells differentiate into plasmocytes vs. 0% (±0%) for CD11c− B cells (n = 3; p > 0.3, data not shown).
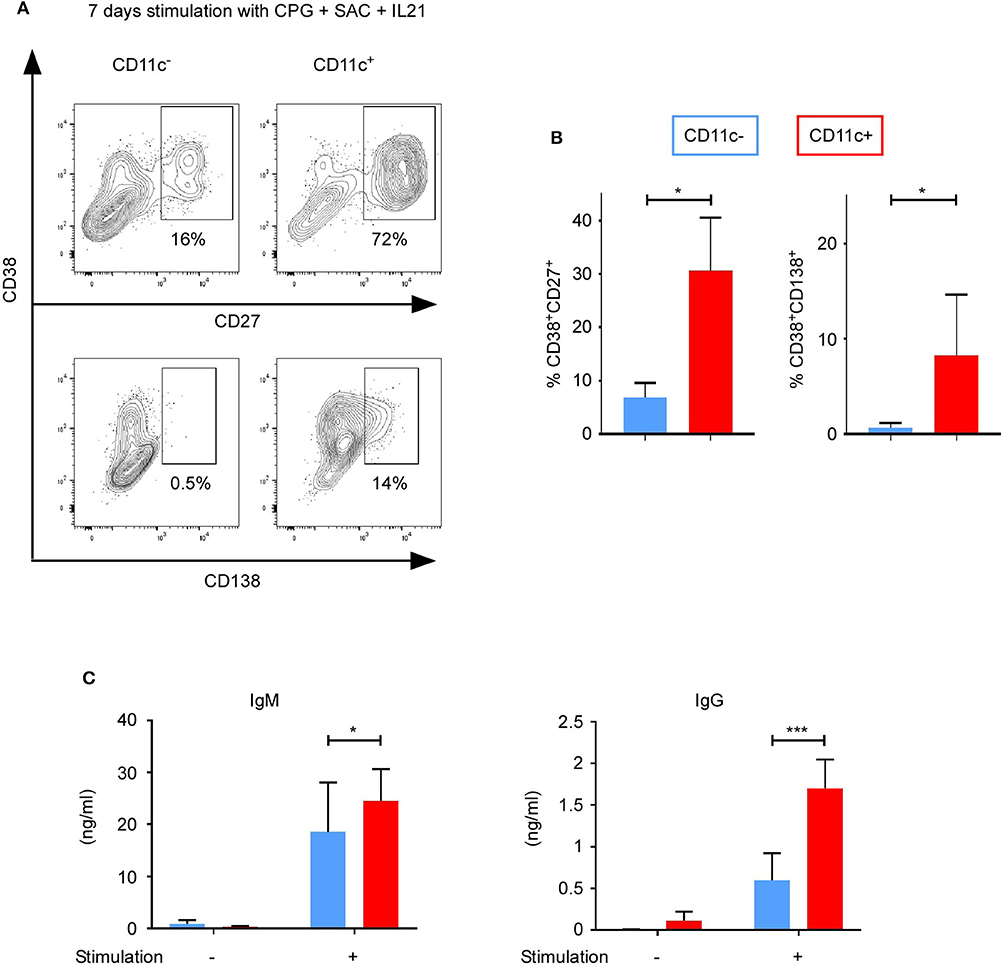
Figure 6. CD11c+ B cells differentiate into antibody-secreting cells. Purified CD11c+ and CD11c− B cells cultured with CPG + SAC + IL-21. Cells and supernatants were analyzed at day 7. (A) Gating strategy to study plasma cells (CD38+CD27+) and plasmocytes (CD38+CD138+) (one representative result of three independent experiments is presented). (B) Bar graphs show mean ± SEM of % CD38+CD27+ and % CD38+CD138+. (C) IgG and IgM secretion were measured by ELISA (n = 7). Out of seven sorting performed, only three gave us a sufficient number of CD11c+ B cells to perform flow cytometry and ELISA analysis. Bar graphs show mean ± SEM of quantity of antibodies (ng/ml). Significant differences are determined using Student's t test for (B) (n = 3; *p < 0.05) and two-way ANOVA with correction by Sidak's multiple comparison test for C) *p < 0.05; ***p < 0.001.
We then measured the IgM and IgG secretion using ELISA. After 7 days of stimulation, CD11c+ B cells secreted 1.3-fold more IgM and 2.8-fold more IgG than CD11c− B cells (Figure 6C) (for IgM from CD11c+ B cells, 25 ± 6 ng/ml, and for CD11c− B cells, 19 ± 10 ng/ml, n = 7, p = 0.011; for IgG from CD11c+ B cells, 1.7 ± 0.3 ng/ml, and for CD11c− B cells, 0.6 ± 0.3 ng/ml; n = 7, p = 0.0002). A 2.7-fold higher secretion of IgM and 1.4-fold higher secretion of IgG by CD11c+ B cells than CD11c− B cells was also observed after a shorter stimulation duration of 5 days (IgM, 27.1 ± 10.3 vs. 10 ± 5.8 ng/ml; p = 0.014; IgG, 1.4 ± 0.3 vs. 1 ± 0.5 ng/ml; p = 0.0093). Without stimulation, neither CD11c+ nor CD11c− B cells upregulated CD138 (data not shown) nor produced IgM or significant quantity of IgG (Figure 6C). Therefore, upon stimulation, CD11c+ B cells were found prone to differentiate into antibody-secreting cells, especially plasmocytes, with a higher frequency of immunoglobulin class switched, which is probably due to the higher frequency of memory B cells in the CD11c+ B cells subpopulation.
Upregulation of CD11c Upon BCR Stimulation
We next addressed whether CD11c− B cells could upregulate CD11c upon activation. Purified CD11c− B cells were cultured without stimulation or with different stimuli: CpG (TLR9 agonist), anti-Ig, R848 (TLR7/8 agonist) for 48 h. CD11c expression was measured by qPCR and flow cytometry. TLR9 or TLR7 stimulation did not upregulate CD11c, while BCR stimulation upregulated CD11c at the protein and mRNA levels (Figure 7), demonstrating that CD11c upregulation is controlled by BCR stimulation. Interestingly, the combine stimulation TLR + anti-Ig inhibit CD11c upregulation induced by BCR stimulation alone, at the protein and mRNA level for TLR9, while only at the protein level for TLR7. Therefore, CD11c upregulation is controlled by BCR stimulation but can be counteracted by TLR7 or TLR9 costimulation.
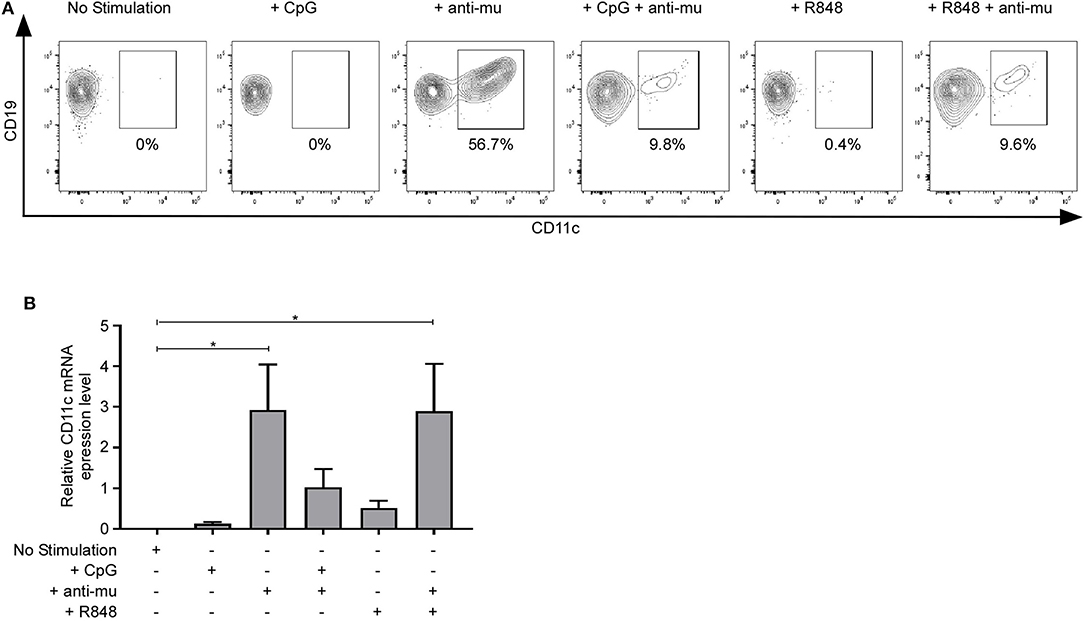
Figure 7. Upregulation of CD11c upon B-cell receptor (BCR) stimulation. CD11c expression was measured after defined stimulation on purified CD11c− B cells by (A) flow cytometry (one representative result of three independent experiments is presented) or (B) by qPCR (n = 4). Bar graphs show mean ± SEM of relative CD11c expression. Means were compared using one-way analysis of variance followed by Dunnett post hoc test: *p < 0.05.
Higher Frequency of CD11c+ B Cells in Pemphigus Patients Before Treatment
CD11c+ B cells have been associated with autoimmune diseases (9–12). Pemphigus is an autoimmune disease affecting skin and mucous membranes, with pathogenic autoantibodies directed against desmogleins 1 and 3 (Dsg1 and Dsg3). We therefore evaluated the frequency of CD11c+ B cells in patients with active pemphigus vulgaris (PV) in comparison to HD. Patients with active PV have not a significant higher frequency of CD11c+ B cells (p > 0.05) than HDs (Figure 8A). However, patients' age ranges from 19 to 79 years old, and Figure 1E shows a significant difference in CD11c+ B cells between age groups. We therefore compared CD11c+ B cells per age groups and found that patients with active PV have a significant higher frequency of CD11c+ B cells than HD in age groups 20–35 and 50–70, but not for the age group 35–50 (Figure 8B). We recently showed that first-line Rituximab treatment allowed achieving an 89% rate of complete remission off therapy at month 24 in pemphigus patients (29). Patients were then evaluated at month 36 when B cells were detectable in blood. PV patients who relapsed have a significant higher frequency of CD11c+ B cells than patients in complete remission at month 36 (Figure 8C). Interestingly, the frequency of CD11c+ B cells was 1.6 times higher in the relapse groups at month 36 when compare to baseline at day 0 (mean, 25 ± 6% at day 0 vs. 40 ± 6% at month 36, n = 5), while in the group of PV patients with no relapse, the frequency of CD11c+ B decreased at month 36 when compare to baseline at day 0 (mean, 22 ± 5% at day 0 vs. 16 ± 4% at month 36, n = 9). Therefore, CD11c+ B-cell frequency increases during active disease in PV patients but decreases after healing. We did not observed a correlation between the percentage of CD11c+ B cells and the autoantibodies, anti-Dsg1 and anti-Dsg3 titer (Figure 8D).
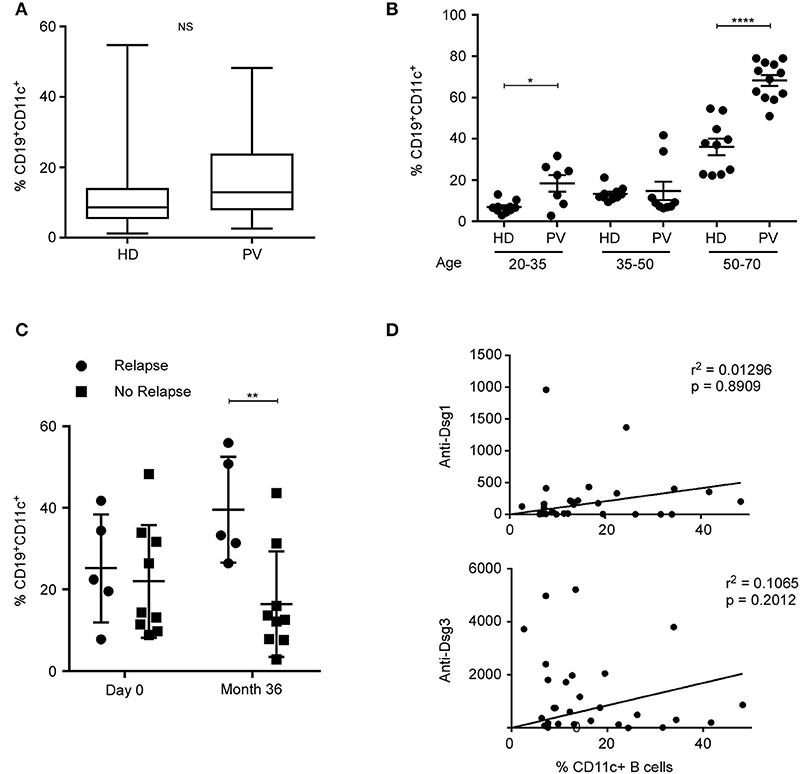
Figure 8. Higher frequency of CD11c+ B cells in pemphigus patients with active disease. (A) Box plot of CD11c+CD19+ frequency determined by cytometry in blood of healthy donors (HD) and pemphigus vulgaris (PV) patients. NS, non-significant. CD11c+ B cells' frequencies were measured for 28 patients with PV and 52 HD, independently of their age. There was no significant difference between patients with an active autoimmune disease and a large group of HD. (B) Percentage of CD11c+ B cells for HD and PV donors between age 20 and 35, 35 and 50, and 50 and 70 years old were compared. Significant difference is determined by one-way ANOVA with correction by Bonferroni's multiple comparison test. *p < 0.05, ****p < 0.0001. (C) Percentage of CD11c+ CD19+ B cells was measured by flow cytometry in PV patients at day 0 and 36 months after treatment. Patients were grouped according to whether or not they relapsed after treatment. Means were compared by two-way ANOVA with correction by Sidak's multiple comparison test: **p < 0.01. (D) Increase in CD11c B cells does not correlate to anti-DSG titer in PV patient blood (r, Pearson correlation coefficient; n = 28).
Discussion
An unusual B-cell subset characterized by CD11c expression has been described to be enriched in patients with different autoimmune diseases as RA (9), Sjogren's syndrome (10), multiple sclerosis (11), and SLE (12), as well as during infections or in aged individual (9, 13). However, these B cells can be found in HD as well. This broad range of condition could explain why markers used to characterize this population are diverse. Indeed, a B cell population that expressed Fc receptor-like 4 and CD11c was described in tissue and tonsil of HD, as memory B cells but CD27− (30). Later, Huifang Li described these so called tissue-like-memory B cells in HD blood as FcRL5+, CD21low, CD27−, and CD11c+ B cells (15). In our study, we focus our analysis on CD19+CD11c+ B cells, which include subpopulation expressing FcRL5 or CD21low or CD27− CD11c+ B cells, already described. Immunophenotyping, molecular, and functional analyses demonstrated in a consistent manner that CD11c+ B cells correspond to a subset of B cells, which mainly but not exclusively belong to memory B cells, and have a strong ability to differentiate into antibody-secreting cells.
We first confirmed that the percentage of CD11c+ B cells increased with age in HD (Figure 1E), as reported in mice in which CD11c+ B cells accumulate and are known as ABC (9). Despite the fact that CD11c+ B cells have been reported to be increased in women with RA, we did not evidence any relationship with gender in HD. No significant differences were observed between age groups for CD11chi B cells.
Phenotypic analyses showed that the CD11c+ B subpopulation was significantly enriched in switched and unswitched memory B cells, while CD11c− B cells were principally naive or DN B cells (Figure 1C), thus confirming Rubtsov's but not Wang's data (9, 12). However, by following Wang's gating strategy on CD19hiCD11chi cells, we observed a significant decrease in switched memory and a significant increase in unswitched memory and naive B cells in comparison with CD11c+ B cells. However, the DN B cells were not increased as Wang reported (12). The facts that [1] frequencies of CD11chi B cells did not change with age while CD11c+ B cells increase with age, [2] CD11chi B cells have a bigger size, and [3] are enriched in the unswitched memory phenotype in comparison with CD11c+ B cells strongly suggest that CD11chi and CD11c+ B cells are two distinct populations.
DN cells (CD27−IgD−) are atypical memory cells, which are reported to be exhausted, anergic, or preplasma cells. The role of DN cells remains unclear, but it has been observed that DN frequency increases during infection (HIV, malaria, Hantavirus), autoimmune disease (SLE, multiple sclerosis), as well as in non-small cell lung cancer (13, 31–35). In addition, DN cells are reported to express CD11c in HIV, malaria, Hantavirus infection, SLE, and multiple sclerosis (11, 36, 37). Based on those observations, one could expect to have a higher frequency of DN cells among CD19+CD11c+/hi. However, we analyzed HD blood in Figure 1C, and frequency of DN cells was lower in CD19+CD11c+/hi than in CD19+CD11c− B cells. Our interpretation is that DN CD11c+ B cells increase during ongoing immune response, associated with a strong inflammatory response. In addition, SC Jenks shows that DN CD11c+ B cells in SLE are poised to generate plasmablasts (33), and J. Fraussen shows that DN cells contain less somatic mutation than switched memory B cells in multiple sclerosis (32). Therefore, we hypothesize that DN CD11c+ B cells could be an intermediate step of differentiation toward plasma cells during B-cell activation, while in normal condition, DN CD11c− B cells could be anergic or exhausted B cells.
Despite the preferential expression of CD11c among memory B cells, our phenotypic analyses showed that CD11c was expressed by all B cell subpopulations that we analyzed. To identify a unique surface marker and transcription signature, we performed a microarray analysis on CD11c+ and CD11c− B cells purified from five HD.
CD11c+ B cells were extremely different from CD11c− B cells with 512 DEG with a fold change above 3. However, none of the cell surface markers identified by microarray was able to strictly discriminate CD11c+ from CD11c− B cells by FACS because of their heterogeneous expression (Figures 3C,D). Moreover, CD11c+ B cells are often characterized by low or negative expression of CD21 (9, 11). However, gene expression of CD21 was found downregulated in CD11c+ B cells with a fold change <3, and CD21 expression analyzed by FACS revealed that less than 10% of CD19+CD11c+ were CD21low/− in HD (data not shown). Similarly, IL-21R was reported to be upregulated in SLE patients (12). However, in our study, IL21R and IL21 are not a DEG, and IL21 was not detected by qPCR, which suggests that these markers are specific of CD11c+ B cells in SLE.
Wikipathways and GO analyses identified, in CD11c+ B cells, an enrichment of cell activation and cell adhesion molecules, TLR signaling, and inflammatory response. Indeed, upregulation of genes coding for the CD80 and CD86 costimulatory molecules observed in CD11c+ B cells was confirmed by FACS for CD86 (Figure 3C). This result is in accordance with studies showing the overexpression of costimulatory molecules in ABCs (9, 11, 12). Interestingly, CD200 was downregulated in CD11c+ B cells (Figure 3D), which can inhibit monocyte stimulation through binding to CD200-R (38), while the upregulation of CD58 lead to increased adhesion and activation of T cells through binding to CD2 (39).
TLR7 signaling is crucial for CD11c+ B-cell development in mice (40). In addition, TLR9 has been shown to be upregulated in CD11c+ B cells from SLE patients (12). Here, we demonstrated that TLR7 or TLR9 stimulation of purified CD11c− B cells was not sufficient to upregulate CD11c expression (Figure 7), whereas BCR stimulation was sufficient. Upregulation of CD11c was reduced as well by almost 2-fold (36.7% vs. 19.6%) by adding CpG on B cells stimulated by anti-IgM + anti-CD40 (12). Therefore, while TLR signaling is an important signaling pathway highlighted by the microarray analysis, TLR signaling does not seem to be crucial for CD11c upregulation in human peripheral B cells.
IRF5 is a transcription factor that can induce IFNγ mRNA transcription (41) and regulate IgG class switching in B cells (27, 42). Our qPCR analysis showed an overexpression of IRF5 by CD11c+ B cells (Figure 5D), which is in agreement with a previous report showing that IRF5 participates in the development of ABCs in systemic autoimmunity in mice (43). Activation of TLR7 and TLR9 induces IRF5 translocation in the nucleus, leading to the transcription of IFN and other pro-inflammatory cytokine genes (43). Indeed, we observed a significant increase in IFNγ gene expression as well as IL1β, IL7, and IL12p40 (Figure 5A). On the other hand, transcriptomic and qPCR analyses showed that the gene expression of EBI3, which is a subunit of the cytokine IL-27, was increased in CD11c+ B cells (Figure 5A) (44). Interestingly, IL-27 and IFN-γ can induce B-cell intrinsic T-bet expression, which regulates Ig class switching and is required for the production of long-lived antibody-secreting cells (25, 45).
As previously described in SLE patients, gene expression level of IL6 was decreased and IL10 was increased in CD11c+ compared with CD11c− B cells (Figure 5) (12). Although IL-6 is a pro-inflammatory cytokine (46, 47) and IL-10 has anti-inflammatory effects, both help in B-cell differentiation and survival as well. Moreover, we previously reported that IL-10 secretion following TLR9 and BCR stimulation was not detected by ELISA in CD11c+ B cells from HD (14), suggesting that IL-10 might be completely consumed by B-activated cells differentiating into antibody-secreting cells.
Furthermore, we observed an overexpression of TBX21 (T-bet) in CD11c+ B cells, confirming studies that demonstrated that ABC express high levels of CD11c and transcription factor T-bet (48–52). Although the transcription factor T-bet is not usually expressed by B cells (53, 54), several studies demonstrated its role in Ig class switching (55, 56). Moreover, Rubtsov et al. showed that CD11c+ T-bet+ B cells were more effective in presenting Ag than other B cell subpopulations (57). Consequently, we observed that, upon stimulation, CD11c+ B cells were found prone to differentiate into antibody-secreting cells, especially plasmocytes, with a higher frequency of immunoglobulin class switched than CD11c− B cells subpopulation (Figure 6). However, not all CD11c+ B cells are T-bet+ in SLE patients or HD, and some T-bet+ B cells do not express CD11c (12). Moreover, B-cell-intrinsic T-bet deletion in a murine lupus model exerted no impact of CD11c+ B cells generation in vivo (16).
Finally, we observed that the frequency of CD11c+ B cells was higher in PV patients than HD defined per age groups, as previously reported in RA. Moreover, the frequency of CD11c+ B cells increased in a group of PV patients with active disease in comparison with a group in complete remission, 36 months after treatment (Figure 8C), suggesting that CD11c+ B-cell frequencies could be an indicator of disease activity. During active disease, CD11c+ B cells were enriched in the memory compartment (switched, unswitched, and DN), and we did not observed a preferential expansion of DN cells (data not shown). In addition, we did not observed a correlation between the frequency of CD11c+ B cells in the blood and the autoantibodies titer as previously reported in SLE (12), which could be due to the skin localization of B cells in the skin lesions of PV patients (58).
In conclusion, our study has characterized CD11c+ B cells in the blood of HD. CD11c is expressed in many B cell subpopulations but is enriched in memory B cells, which have a strong ability to differentiate into plasmocytes and secrete Ig. Accumulation of CD11c+ B cells, which naturally occurs with age, could thus promote the emergence of autoimmunity.
Data Availability Statement
The datasets generated for this study can be found in the National Center for Biotechnology Information's Gene Expression Omnibus (https://www.ncbi.nlm.nih.gov/geo/). Data are accessible using the following accession number: GSE112515.
Ethics Statement
The studies involving human participants were reviewed and approved by Ethics Committee of the North West in France. The patients/participants provided their written informed consent to participate in this study.
Author Contributions
M-LG and SC wrote and conceived the manuscript. M-LG, SC, MD, CD, GR, and MM-V generated and analyzed data. OB and PJ read and revised the manuscript.
Funding
This study was supported by INSERM, Normandy University, and Rouen University Hospital, Dermatology Department, France.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank the Center for Human Immunology at Institut Pasteur for providing access to Biomark Fluidigm technology. We are grateful to Nikki Sabourin-Gibbs, Rouen University Hospital, for editing the manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2020.00032/full#supplementary-material
Supplemental Table 1. List of DEG.
Abbreviations
ABC, age-associated B cells; DEG, differentially expressed genes; HD, healthy donors; PV, pemphigus vulgaris; RA, rheumatoid arthritis; SLE, systemic lupus erythematosus.
References
1. Schittenhelm L, Hilkens CM, Morrison VL. beta2 integrins as regulators of dendritic cell, monocyte, and macrophage function. Front Immunol. (2017) 8:1866. doi: 10.3389/fimmu.2017.01866
2. Rosen H, Law SK. The leukocyte cell surface receptor(s) for the iC3b product of complement. Curr Top Microbiol Immunol. (1990) 153:99–122. doi: 10.1007/978-3-642-74977-3_6
3. Blackford J, Reid HW, Pappin DJ, Bowers FS, Wilkinson JM. A monoclonal antibody, 3/22, to rabbit CD11c which induces homotypic T cell aggregation: evidence that ICAM-1 is a ligand for CD11c/CD18. Eur J Immunol. (1996) 26:525–31. doi: 10.1002/eji.1830260304
4. Ihanus E, Uotila LM, Toivanen A, Varis M, Gahmberg CG. Red-cell ICAM-4 is a ligand for the monocyte/macrophage integrin CD11c/CD18: characterization of the binding sites on ICAM-4. Blood. (2007) 109:802–10. doi: 10.1182/blood-2006-04-014878
5. Ingalls RR, Golenbock DT. CD11c/CD18, a transmembrane signaling receptor for lipopolysaccharide. J Exp Med. (1995) 181:1473–9. doi: 10.1084/jem.181.4.1473
6. Garnotel R, Rittie L, Poitevin S, Monboisse JC, Nguyen P, Gillery P. Human blood monocytes interact with type I collagen through alpha x beta 2 integrin (CD11c-CD18, gp150-95). J Immunol. (2000) 164:5928–34. doi: 10.4049/jimmunol.164.11.5928
7. Loike JD, Sodeik B, Cao L, Leucona S, Weitz JI, Silverstein SC. CD11c/CD18 on neutrophils recognizes a domain at the N terminus of the A alpha chain of fibrinogen. Proc Natl Acad Sci USA. (1991) 88:1044–48. doi: 10.1073/pnas.88.3.1044
8. Kamp VM, Pillay J, Lammers JW, Pickkers P, Ulfman LH, Koenderman L. Human suppressive neutrophils CD16bright/CD62Ldim exhibit decreased adhesion. J Leukoc Biol. (2012) 92:1011–20. doi: 10.1189/jlb.0612273
9. Rubtsov AV, Rubtsova K, Fischer A, Meehan RT, Gillis JZ, Marrack P. Toll-like receptor 7 (TLR7)-driven accumulation of a novel CD11c(+) B-cell population is important for the development of autoimmunity. Blood. (2011) 118:1305–15. doi: 10.1182/blood-2011-01-331462
10. Saadoun D, Terrier B, Bannock J, Vazquez T, Massad C, Kang I, et al. Expansion of autoreactive unresponsive CD21-/low B cells in Sjogren's syndrome-associated lymphoproliferation. Arthritis Rheum. (2013) 65:1085–96. doi: 10.1002/art.37828
11. Claes N, Fraussen J, Vanheusden M, Hellings N, Stinissen P, Van Wijmeersch B, et al. Age-associated B cells with proinflammatory characteristics are expanded in a proportion of multiple sclerosis patients. J Immunol. (2016) 197:4576–83. doi: 10.4049/jimmunol.1502448
12. Wang S, Wang J, Kumar V, Karnell JL, Naiman B, Ettinger R. IL-21 drives expansion and plasma cell differentiation of autoreactive CD11c(hi)T-bet(+) B cells in SLE. Nat Commun. (2018) 9:1758. doi: 10.1038/s41467-018-03750-7
13. Weiss GE, Crompton PD, Li S, Walsh LA, Moir S, Pierce SK. Atypical memory B cells are greatly expanded in individuals living in a malaria-endemic area. J Immunol. (2009) 183:2176–82. doi: 10.4049/jimmunol.0901297
14. Lin W, Cerny D, Chua E, Duan K, Yi JT, Calbo S. Human regulatory B cells combine phenotypic and genetic hallmarks with a distinct differentiation fate. J Immunol. (2014) 193:2258–66. doi: 10.4049/jimmunol.1303214
15. Li H, Borrego F, Nagata S, Tolnay M. Fc receptor-like 5 expression distinguishes two distinct subsets of human circulating tissue-like memory B cells. J Immunol. (2016) 196:4064–74. doi: 10.4049/jimmunol.1501027
16. Du SW, Arkatkar T, Jacobs HM, Rawlings DJ, Jackson SW. Generation of functional murine CD11c(+) age-associated B cells in the absence of B cell T-bet expression. Eur J Immunol. (2019) 49:170–8. doi: 10.1002/eji.201847641
17. Tung JW, Heydari K, Tirouvanziam R, Sahaf B, Parks DR, Herzenberg LA. Modern flow cytometry: a practical approach. Clin Lab Med. (2007) 27:453–68. doi: 10.1016/j.cll.2007.05.001
18. Berkani N, Joly P, Golinski ML, Colliou N, Lim A, Larbi A, et al. B-cell depletion induces a shift in self antigen specific B-cell repertoire and cytokine pattern in patients with bullous pemphigoid. Sci Rep. (2019) 9:3525. doi: 10.1038/s41598-019-40203-7. Erratum in: Sci Rep. (2019) 9:18991.
19. Hebert V, Petit M, Maho-Vaillant M, Golinski ML, Riou G, Calbo S. Modifications of the transcriptomic profile of autoreactive B cells from pemphigus patients after treatment with rituximab or a standard corticosteroid regimen. Front Immunol. (2019) 10:1794. doi: 10.3389/fimmu.2019.01794
20. Vandesompele J, De Preter K, Pattyn F, Poppe B, Van Roy N, De Paepe A, et al. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. (2002) 3:RESEARCH0034. doi: 10.1186/gb-2002-3-7-research0034
21. Huang da W, Sherman BT, Lempicki RA. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat Protoc. (2009) 4:44–57. doi: 10.1038/nprot.2008.211
22. Paschoud S, Dogar AM, Kuntz C, Grisoni-Neupert B, Richman L, Kuhn LC. Destabilization of interleukin-6 mRNA requires a putative RNA stem-loop structure, an AU-rich element, and the RNA-binding protein AUF1. Mol Cell Biol. (2006) 26:8228–41. doi: 10.1128/MCB.01155-06
23. Cahill CM, Rogers JT. Interleukin (IL) 1beta induction of IL-6 is mediated by a novel phosphatidylinositol 3-kinase-dependent AKT/IkappaB kinase alpha pathway targeting activator protein-1. J Biol Chem. (2008) 283:25900–912. doi: 10.1074/jbc.M707692200
24. Vazquez MI, Catalan-Dibene J, Zlotnik A. B cells responses and cytokine production are regulated by their immune microenvironment. Cytokine. (2015) 74:318–26. doi: 10.1016/j.cyto.2015.02.007
25. Stone SL, Peel JN, Scharer CD, Risley CA, Chisolm DA, Lund FE. T-bet transcription factor promotes antibody-secreting cell differentiation by limiting the inflammatory effects of IFN-gamma on B cells. Immunity. (2019) 50:1172–87.e1177. doi: 10.1016/j.immuni.2019.04.004
26. Lund FE. Cytokine-producing B lymphocytes-key regulators of immunity. Curr Opin Immunol. (2008) 20:332–8. doi: 10.1016/j.coi.2008.03.003
27. De S, Zhang B, Shih T, Singh S, Winkler A, Donnelly RJ, et al. B cell-intrinsic role for IRF5 in TLR9/BCR-induced human B cell activation, proliferation, and plasmablast differentiation. Front Immunol. (2017) 8:1938. doi: 10.3389/fimmu.2017.01938
28. Lien C, Fang CM, Huso D, Livak F, Lu R, Pitha PM. Critical role of IRF-5 in regulation of B-cell differentiation. Proc Natl Acad Sci USA. (2010) 107:4664–8. doi: 10.1073/pnas.0911193107
29. Joly P, Maho-Vaillant M, Prost-Squarcioni C, Hebert V, Houivet E, Calbo S, et al. First-line rituximab combined with short-term prednisone versus prednisone alone for the treatment of pemphigus (Ritux 3): a prospective, multicentre, parallel-group, open-label randomised trial. Lancet. (2017) 389:2031–40. doi: 10.1016/S0140-6736(17)30070-3
30. Ehrhardt GR, Hsu JT, Gartland L, Leu CM, Zhang S, Cooper MD. Expression of the immunoregulatory molecule FcRH4 defines a distinctive tissue-based population of memory B cells. J Exp Med. (2005) 202:783–91. doi: 10.1084/jem.20050879
31. Centuori SM, Gomes CJ, Kim SS, Putnam CW, Larsen BT, Martinez JD. Double-negative (CD27(-)IgD(-)) B cells are expanded in NSCLC and inversely correlate with affinity-matured B cell populations. J Transl Med. (2018) 16:30. doi: 10.1186/s12967-018-1404-z
32. Fraussen J, Marquez S, Takata K, Beckers L, Montes Diaz G, Zografou C, et al. Phenotypic and Ig repertoire analyses indicate a common origin of IgD(-)CD27(-) double negative B cells in healthy individuals and multiple sclerosis patients. J Immunol. (2019) 203:1650–64. doi: 10.4049/jimmunol.1801236
33. Jenks SA, Cashman KS, Zumaquero E, Marigorta UM, Patel AV, Sanz I. Distinct effector B cells induced by unregulated toll-like receptor 7 contribute to pathogenic responses in systemic lupus erythematosus. Immunity. (2018) 49:725–739.e726. doi: 10.1016/j.immuni.2018.08.015
34. Kerkman PF, Dernstedt A, Tadala L, Mittler E, Dannborg M, Sundling C, et al. The generation of plasma cells and CD27–IgD– B cells during Hantavirus infection are associated with distinct pathological findings. bioRXiv. (2019). doi: 10.1101/723585
35. Moir S, Ho J, Malaspina A, Wang W, DiPoto AC, Fauci AS. Evidence for HIV-associated B cell exhaustion in a dysfunctional memory B cell compartment in HIV-infected viremic individuals. J Exp Med. (2008) 205:1797–805. doi: 10.1084/jem.20072683
36. Knox JJ, Buggert M, Kardava L, Seaton KE, Eller MA, Betts MR. T-bet+ B cells are induced by human viral infections and dominate the HIV gp140 response. JCI Insight. (2017) 2:e92943. doi: 10.1172/jci.insight.92943
37. Sullivan RT, Kim CC, Fontana MF, Feeney ME, Jagannathan P, Greenhouse B. FCRL5 delineates functionally impaired memory B cells associated with Plasmodium falciparum exposure. PLoS Pathog. (2015) 11:e1004894. doi: 10.1371/journal.ppat.1004894
38. Jenmalm MC, Cherwinski H, Bowman EP, Phillips JH, Sedgwick JD. Regulation of myeloid cell function through the CD200 receptor. J Immunol. (2006) 176:191–9. doi: 10.4049/jimmunol.176.1.191
39. Leitner J, Herndler-Brandstetter D, Zlabinger GJ, Grubeck-Loebenstein B, Steinberger P. CD58/CD2 is the primary costimulatory pathway in human CD28-CD8+ T cells. J Immunol. (2015) 195:477–87. doi: 10.4049/jimmunol.1401917
40. Rubtsov AV, Rubtsova K, Kappler JW, Marrack P. TLR7 drives accumulation of ABCs and autoantibody production in autoimmune-prone mice. Immunol Res. (2013) 55:210–6. doi: 10.1007/s12026-012-8365-8
41. Barnes BJ, Moore PA, Pitha PM. Virus-specific activation of a novel interferon regulatory factor, IRF-5, results in the induction of distinct interferon alpha genes. J Biol Chem. (2001) 276:23382–90. doi: 10.1074/jbc.M101216200
42. Savitsky DA, Yanai H, Tamura T, Taniguchi T, Honda K. Contribution of IRF5 in B cells to the development of murine SLE-like disease through its transcriptional control of the IgG2a locus. Proc Natl Acad Sci USA. (2010) 107:10154–9. doi: 10.1073/pnas.1005599107
43. Manni M, Gupta S, Ricker E, Chinenov Y, Park SH, Pernis AB. Regulation of age-associated B cells by IRF5 in systemic autoimmunity. Nat Immunol. (2018) 19:407–19. doi: 10.1038/s41590-018-0056-8
44. Pflanz S, Timans JC, Cheung J, Rosales R, Kanzler HA. IL-27, a heterodimeric cytokine composed of EBI3 and p28 protein, induces proliferation of naive CD4+ T cells. Immunity. (2002) 16:779–90. doi: 10.1016/S1074-7613(02)00324-2
45. Yoshimoto T, Okada K, Morishima N, Kamiya S, Owaki T, Asakawa M, et al. Induction of IgG2a class switching in B cells by IL-27. J Immunol. (2004) 173:2479–85. doi: 10.4049/jimmunol.173.4.2479
46. Bao Y, Cao X. The immune potential and immunopathology of cytokine-producing B cell subsets: a comprehensive review. J Autoimmun. (2014) 55:10–23. doi: 10.1016/j.jaut.2014.04.001
47. Samoilova EB, Horton JL, Hilliard B, Liu TS, Chen Y. IL-6-deficient mice are resistant to experimental autoimmune encephalomyelitis: roles of IL-6 in the activation and differentiation of autoreactive T cells. J Immunol. (1998) 161:6480–6.
48. Barnett BE, Staupe RP, Odorizzi PM, Palko O, Tomov VT, Wherry EJ. Cutting edge: B cell-intrinsic T-bet expression is required to control chronic viral infection. J Immunol. (2016) 197:1017–22. doi: 10.4049/jimmunol.1500368
49. Hao Y, O'Neill P, Naradikian MS, Scholz JL, Cancro MP. A B-cell subset uniquely responsive to innate stimuli accumulates in aged mice. Blood. (2011) 118:1294–304. doi: 10.1182/blood-2011-01-330530
50. Isnardi I, Ng YS, Menard L, Meyers G, Saadoun D, Meffre E. Complement receptor 2/CD21- human naive B cells contain mostly autoreactive unresponsive clones. Blood. (2010) 115:5026–36. doi: 10.1182/blood-2009-09-243071
51. Rubtsova K, Rubtsov AV, Thurman JM, Mennona JM, Kappler JW, Marrack P. B cells expressing the transcription factor T-bet drive lupus-like autoimmunity. J Clin Invest. (2017) 127:1392–404. doi: 10.1172/JCI91250
52. Rubtsova K, Rubtsov AV, van Dyk LF, Kappler JW, Marrack P. T-box transcription factor T-bet, a key player in a unique type of B-cell activation essential for effective viral clearance. Proc Natl Acad Sci USA. (2013) 110:E3216–24. doi: 10.1073/pnas.1312348110
53. Cruz-Guilloty F, Pipkin ME, Djuretic IM, Levanon D, Lotem J, Rao A. Runx3 and T-box proteins cooperate to establish the transcriptional program of effector CTLs. J Exp Med. (2009) 206:51–9. doi: 10.1084/jem.20081242
54. Townsend MJ, Weinmann AS, Matsuda JL, Salomon R, Farnham PJ, Glimcher LH. T-bet regulates the terminal maturation and homeostasis of NK and Valpha14i NKT cells. Immunity. (2004) 20:477–94. doi: 10.1016/S1074-7613(04)00076-7
55. Liu N, Ohnishi N, Ni L, Akira S, Bacon KB. CpG directly induces T-bet expression and inhibits IgG1 and IgE switching in B cells. Nat Immunol. (2003) 4:687–93. doi: 10.1038/ni941
56. Peng SL, Szabo SJ, Glimcher LH. T-bet regulates IgG class switching and pathogenic autoantibody production. Proc Natl Acad Sci USA. (2002) 99:5545–50. doi: 10.1073/pnas.082114899
57. Rubtsov AV, Rubtsova K, Kappler JW, Jacobelli J, Friedman RS, Marrack P. CD11c-expressing B cells are located at the T cell/B cell border in spleen and are potent APCs. J Immunol. (2015) 195:71–9. doi: 10.4049/jimmunol.1500055
Keywords: human, CD11c, B cells, microarray, plasma cells, pemphigus
Citation: Golinski M-L, Demeules M, Derambure C, Riou G, Maho-Vaillant M, Boyer O, Joly P and Calbo S (2020) CD11c+ B Cells Are Mainly Memory Cells, Precursors of Antibody Secreting Cells in Healthy Donors. Front. Immunol. 11:32. doi: 10.3389/fimmu.2020.00032
Received: 20 October 2019; Accepted: 08 January 2020;
Published: 25 February 2020.
Edited by:
Deborah K. Dunn-Walters, University of Surrey, United KingdomReviewed by:
Daniela Frasca, University of Miami, United StatesYoshiya Tanaka, University of Occupational and Environmental Health, Japan
Copyright © 2020 Golinski, Demeules, Derambure, Riou, Maho-Vaillant, Boyer, Joly and Calbo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sébastien Calbo, c2ViYXN0aWVuLmNhbGJvJiN4MDAwNDA7dW5pdi1yb3Vlbi5mcg==