
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Immunol. , 12 March 2019
Sec. Mucosal Immunity
Volume 10 - 2019 | https://doi.org/10.3389/fimmu.2019.00430
 Romain Marlin1,2,3
Romain Marlin1,2,3 Marie-Thérèse Nugeyre1,2,3
Marie-Thérèse Nugeyre1,2,3 Nicolas Tchitchek1
Nicolas Tchitchek1 Matteo Parenti1,3Cécile Lefebvre3,4Hakim Hocini3,4Fahd Benjelloun1,2Claude Cannou1,2
Matteo Parenti1,3Cécile Lefebvre3,4Hakim Hocini3,4Fahd Benjelloun1,2Claude Cannou1,2 Silvia Nozza5
Silvia Nozza5 Nathalie Dereuddre-Bosquet1
Nathalie Dereuddre-Bosquet1 Yves Levy3,4,6Françoise Barré-Sinoussi3,7Gabriella Scarlatti3,8
Yves Levy3,4,6Françoise Barré-Sinoussi3,7Gabriella Scarlatti3,8 Roger Le Grand1,3
Roger Le Grand1,3 Elisabeth Menu1,2,3*
Elisabeth Menu1,2,3*HIV-1 sexual transmission occurs mainly via mucosal semen exposures. In the female reproductive tract (FRT), seminal plasma (SP) induces physiological modifications, including inflammation. An effective HIV-1 vaccine should elicit mucosal immunity, however, modifications of vaccine responses by the local environment remain to be characterized. Using a modified vaccinia virus Ankara (MVA) as a vaccine model, we characterized the impact of HIV-1+ SP intravaginal exposure on the local immune responses of non-human primates. Multiple HIV-1+ SP exposures did not impact the anti-MVA antibody responses. However, SP exposures revealed an anti-MVA responses mediated by CD4+ T cells, which was not observed in the control group. Furthermore, the frequency and the quality of specific anti-MVA CD8+ T cell responses increased in the FRT exposed to SP. Multi-parameter approaches clearly identified the cervix as the most impacted compartment in the FRT. SP exposures induced a local cell recruitment of antigen presenting cells, especially CD11c+ cells, and CD8+ T cell recruitment in the FRT draining lymph nodes. CD11c+ cell recruitment was associated with upregulation of inflammation-related gene expression after SP exposures in the cervix. We thus highlight the fact that physiological conditions, such as SP exposures, should be taken into consideration to test and to improve vaccine efficacy against HIV-1 and other sexually transmitted infections.
Semen is a complex fluid composed of a cellular fraction containing spermatozoa and leukocytes, and a non-cellular fraction, the so-called seminal plasma (SP) including diverse set of components such as cytokines, chemokines, fibrils, immunoglobulins, complement factors, and bacteria (1–3). Semen deposition in the female reproductive tract (FRT) during intercourse induces physiological modifications (4), such as the induction of pro-inflammatory cytokine/chemokine production (5, 6), and upregulation of inflammatory upstream regulator expression, such as Cyclooxygenase-2 (COX-2) (7), and leukocyte infiltration (8, 9). These modifications of the local environment have been described to promote fertility and are not due to intercourse itself, but require SP exposure (10), as described in vitro and in vivo in mice (5) and pigs (11) as well as humans (10). SP exposure also induces the recruitment of regulatory T cells to the FRT, showing that SP does not have only a pro-inflammatory effect (12).
Semen is the main vector of HIV-1 transmission. The virus is present as both cell-free particles in the SP and infected mononuclear cells (13). Both have been shown to be infectious in animal models (14–16) and human cervical explants (17). SP is not a passive carrier of viral particles (18); it contributes to reduce the protective acidic pH of the vagina (19) and favors the attachment of virions to target cells (20). In contrast, several antiviral factors have been characterized in SP (21). The composition of semen varies according to the presence and stage of HIV-1 infection. In particular, the cytokine and chemokine network is different between the SP from HIV-1neg and HIV-1+ individuals (22), pro-inflammatory factors are higher in the SP of HIV-1+ individuals (23). The effect of HIV-1+ SP on the local environment of the FRT is still uncharacterized.
The FRT mucosae are the main portal of HIV-1 entry during heterosexual intercourse. As a consequence, the induction of efficient mucosal immune responses in the FRT is a strategy to prevent heterosexual transmission of HIV-1. However, many preclinical efficacy studies in animal models and non-human primates (NHP) in particular, testing vaccine candidates or microbicides, used a cell-free culture medium-derived virus inoculum for the challenge phase, which does not allow evaluation of the physiological effects of SP on local immunity. We have recently shown that systemic immunization with an MVA vector-based vaccine was able to induce vaccine-specific CD8+ T cells in all FRT compartments in macaques (24). However, the effect of SP on such vaccine-induced mucosal immune responses is completely unknown.
The aim of the present study was to determine the effect of vaginal HIV-1+ SP exposure on local immunity and vaccine-specific responses in the FRT. We used an MVA vector-based vaccine as a vaccine model and show that SP exposures increase the specific CD8+ T-cell response, myeloid dendritic cells (mDC) recruitment and inflammation-related gene expression. A local specific CD4+ T-cell response was also detected after vaginal SP exposures. Multi-parameter approaches clearly identified the cervix as the most affected compartment in the FRT.
Human semen was collected from 14 HIV-1 infected subjects (among them, 13 were naive for antiretroviral therapy) attending the Infectious Disease Dept., OSR, under the supervision of Dr. Sivia Nozza. Patients were informed of the study and signed an informed consent (HIVSPERM study, protocol number 5/INT/2014 of 2014/02/06). At collection, the median age was 37.5 years (min 28; max 55), the median infection time was 5.5 years (min 2; max 20), the median plasma viral load was 2,468 copies/ml (min 45; max 119,116) and their median CD4 count was 636.5 (min 371; max 10,027). SP was obtained after liquefaction of the semen at 37°C for 30 min and centrifugation at 1,000 × g for 10 min. Supernatant were collected, pooled and stored at −80°C.
Pro-inflammatory and anti-inflammatory cytokines/chemokines and TGFβ isoforms were measured in SP pool by Luminex assays (cytokine human magnetic 25-plex panel; Invitrogen, Courtaboeuf, France and TGFβ 1,2,3 Magnetic Bead Kit, MerckMillipore, Germany). HIV-1 RNA viral load was determined in SP pool by COBAS TaqMan HIV-1 test V2.0 (Roche).
On day zero (D0) and D58, the macaques received two subcutaneous injections per time point in the right and left side of the upper back, delivering 2 × 1 ml of inoculum containing a total of 4 × 108 plaque-forming units (PFU) of recombinant MVA-HIV-1 expressing the Gag, Pol, and Nef proteins from HIV-1 strain LAI (ANRS-MVA HIV-B, MVATG17401, Transgene Ltd., France). The animals were monitored daily for signs of disease, appetite loss and lethargy. A physical examination was performed at each blood sampling and each inoculation. All experimental procedures (handling, immunization, blood sampling) were conducted after sedation with ketamine hydrochloride (Rhône-Mérieux, Lyon, France, 10 mg/kg). An intramuscular injection of a synthetic variant of progesterone (Depoprovera, 30 mg, Pfeizer, France) was given 42 days after the first vaccine injection to synchronize their hormonal cycle. On D70, D72, D74, and D76, animals were exposed intra-vaginally to 800 μl 1X PBS (control group n = 6) or 800 μl SP-pool/PBS (v:v) (SP group, n = 6). The animals were sedated 77 days post-vaccination with ketamine hydrochloride (10 mg/kg) and euthanized by intravenous injection of 180 mg/kg sodium pentobarbital.
Blood, serum and vaginal fluid were collected before and after each vaccine inoculation and at the time of euthanasia. Collection of serum, cervicovaginal fluids, and PBMC was previously described (24), briefly serum was isolated by centrifugation, cervicovaginal fluid was collected with a Weck-Cel Spear and PMBCs were isolated in heparin CPT tubes. LNs and tissues were collected at necropsy. LN cells were obtained by mechanical dissociation. FRT tissues (vagina and cervix which included both ectocervix and endocervix, uterus, and fallopian tubes) were isolated and cut into small pieces. Each tissue was digested for 1 h at 37°C with agitation in digestion buffer, consisting of RPMI 1640 (Fisher Scientific, Illkirch, France), collagenase IV (0.3 mg/ml, Sigma Aldrich, St Quantin Fallavier, France), fetal calf serum (5%, Fisher Scientific), HEPES (0.025 M, Fisher Scientific), DNase (0.1 mg/ml, Roche, Mannheim, Germany), and antibiotics (Fisher Scientific). Undigested pieces were subjected to up to three more digestion steps. Cell suspensions from LNs and FRT tissues were filtered through 70-μm sterile nylon cell strainers (BD Biosciences).
Whole blood, LN cells and cells from FRT compartments were analyzed by flow cytometry. The cells were incubated with the antibodies listed in Table S1, washed, and then fixed with FACS lysing buffer or BD Cell Fix solution. A Fortessa 2-UV 6-Violet 2-Blue 5-Yelgr 3-Red laser configuration was used (BD Biosciences), with Diva (BD) and FlowJo 9.8.3 (Tristar, USA) software. At least 500 events for rare cell populations (i.e., pDC) were recorded.
Specific cellular immune responses were evaluated with in vitro stimulation assays. The cells were incubated for 5 h at 37°C in DMEM medium (Fisher Scientific) supplemented with 10% FCS and antibiotics, alone or medium with 0.3 PFU/cell of live wild-type MVA, or PMA (5 ng/ml) and ionomycin (500 ng/ml) (Sigma Aldrich). Brefeldin A was then added (5 μg/ml, Sigma Aldrich) and the cells incubated for a further 10 h at 37°C. For HIV-1 stimulation, cells were incubated with 4 μg/ml overlapping Gag peptide pools (HIV-1 LAI strain) in DMEM medium supplemented with 10% FCS, antibiotics and costimulatory antibodies, for 1 h at 37°C, then for an additional 4 h with brefeldin A (5 μg/ml). The cells were stained with LIVE/DEAD® Fixable Blue Dead Cell Stain (Thermo Fisher) to assess viability, fixed, and then permeabilized with BD Fix&Perm reagent (BD Bioscience). The antibodies listed in Table S2 were used for intracellular staining. At least 5,000 events in the CD8+ T cell gate were recorded. The gating strategy was as described elsewhere (25). Briefly, cytokine and activation marker expression was evaluated in CD4+ and CD8+ T cells, and Boolean gate analyses were performed with FlowJo software. The percentages of cells positive for cytokines and activation markers between unstimulated and MVA-stimulated cells were then compared.
Specific antibodies were measured by EIA in sera and vaginal fluids, as described previously (24). First, 96-well MaxiSorp microplates (Nunc, Thermo Fisher) were coated overnight with 105 PFU/well wtMVA (Transgene, Illkirch, France) in NaHCO3/Na2CO3 buffer, or 1 μg/ml p24 antigen (kind gift from Bernard Verrier, LBTI UMR5305) in PBS. The plates were then blocked for 1 h with PBS containing 3% (w/v) bovine serum albumin (BSA, Sigma Aldrich) or PBS containing 10% skimmed milk. The plates were washed five times with PBS containing 0.1% Tween 20 and 10 mM EDTA, then incubated with two-fold serial dilutions of macaque fluids diluted in PBS containing 1% BSA for 1 h at room temperature (to detect anti-MVA IgG/IgA) or PBS containing 1% skimmed milk and 0.05% Tween 20 for 1 h at 37°C (to detect anti-HIV IgG), starting at 1:50 for serum and 1:20 for vaginal fluid. The plates were then washed five times and incubated for 1 h with a 1:20,000 dilution of horseradish peroxidase-conjugated goat anti-monkey H+L chain IgG (Bio-Rad, Marne-la-Coquette, France) or a 1:5,000 dilution of horseradish peroxidase-conjugated goat anti-monkey IgA (Alpha Diagnostic international, San Antonio, TX). The plates were washed five times, then 100 μL o-phenylenediamine dihydrochloride (OPD) (Sigma Aldrich) was added and the plates incubated for 30 min at room temperature in the dark. The reaction was stopped by adding 2N H2SO4. Absorbance was measured at 492 nm with a spectrophotometer (Tecan, Lyon, France), and the data analyzed using Magellan software (Tecan). Antibody titers were calculated by extrapolation from the OD as a function of a serum dilution curve and defined as the dilution of the test serum reaching 2 OD of the corresponding preimmune serum or vaginal fluid, tested at 1:50 and 1:30, respectively.
Tissue biopsies were immediately immersed in RLT-beta-mercaptoethanol 1/100 lysis buffer (Qiagen, Courtaboeuf, France), disrupted, and then homogenized with a TissueLyser LT (Qiagen). RNA was purified using Qiagen RNeasy microkits. Contaminating DNA was removed using the RNA Cleanup step of the RNeasy microkit. Purified RNA was quantified with a ND-8000 spectrophotometer (NanoDrop Technologies, Fisher Scientific, Illkirch, France) before being checked for integrity on a 2100 BioAnalyzer (Agilent Technologies, Massy, France). cDNA was synthesized and biotin-labeled using the Ambion Illumina TotaPrep RNA amplification kit (Applied Biosystem/Ambion, Saint-Aubin, France). Labeled cRNA was hybridized on Illumina Human HT-12V4 BeadChips, which target 47,323 probes corresponding to 34 694 genes. The manufacturers' protocols were followed.
Comparison between the control group and the SP-exposed group (antibody and T cell responses, and cell population abundance) were analyzed using the two-tailed Mann-Whitney test. The two-tailed Wilcoxon signed rank test was used to compare paired conditions (T cell responses within each animal group). Polyfunctional profiles of MVA-specific CD8+ T cells were compared between the two groups using the Chi-squared test. The statistical threshold of significance was fixed at p < 0.05 for these tests.
Microarray were analyzed using R/Bioconductor software. Gene expression values were quantile normalized. Differentially expressed genes were identified with a paired non-parametric t-test (q-value < 0.05), based on a 1.2-fold-change cutoff. Functional enrichment analysis was performed using QIAGEN's Ingenuity Pathway Analysis (IPA, QIAGEN, Redwood City, https://www.qiagenbioinformatics.com/products/ingenuity-pathway-analysis/). Hierarchical clustering presented in the heatmaps were generated with the Euclidian metric and complete linkage methods. The correlation between molecular and cellular data were computed using Spearman coefficients of correlation (R > 0.70 and p < 0.01), based on the abundance of cell populations and normalized gene expression values across differentially expressed genes in the vagina, cervix, and uterus. Significant correlations were restricted to those between cell population abundances and gene expression levels. Microarray raw data are available from the EBI-ArrayExpress database under accession number E-MTAB-7639.
Two groups of six cynomolgus macaques immunized by subcutaneous injection of 4 × 108 PFU of rMVA-HIV-1 at weeks 0 and 8, were exposed to PBS, as controls, or to a SP pool at days 70, 72, 74, and 76 following first vaccine injection. The seminal plasma pool was obtained from HIV-1 infected patients and contained 22,000 vRNA copies/ml, and inflammatory as well as anti-inflammatory cytokines/chemokines (Table 1). As expected, TGFβ was the most abundant.
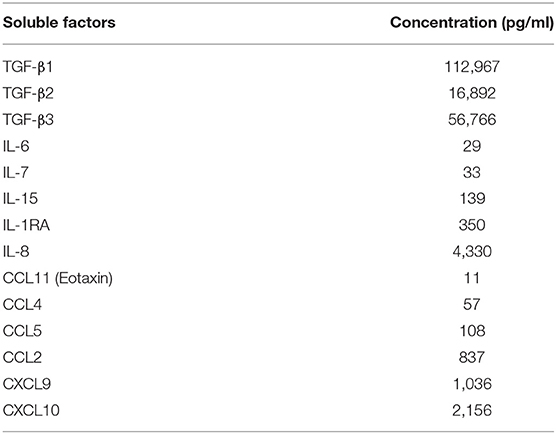
Table 1. Cytokine and chemokine composition of the SP pool.
All animals had been treated with synthetic progesterone on day 42 in order to ensure a similar cervico-vaginal mucosa structural organization and immune status of all animals at time of intravaginal exposure. In addition, progesterone treatment, mimicking luteal phase or contraceptive medication in women, has been shown to increase susceptibility to SIV transmission in macaques (26), therefore representing a “worse case” scenario for exploring local vaccine induced immunity.
One day (D77 of the study) following the last SP/PBS exposure, animals were euthanized and immune responses explored in tissues collected at necropsy. The MVA-specific IgG titers in the sera of control and SP-exposed animals were similar (30,307 ± 6,884 and 31,315 ± 8,390, respectively; mean ± SEM of six animals) (Figure S1A). In addition, we detected MVA-specific IgA in the sera of both, control (1,314 ± 328), and SP-exposed animals (1,955 ± 1,162) without any statistical difference (Figure S1B). In the vaginal fluids, the MVA-specific IgG and IgA titers of the control group (389 ± 111 and 151 ± 54, respectively) were not significantly different from those of the SP-exposed group (219 ± 52 and 205 ± 62, respectively) (Figures S1D,E). The MVA we used also encodes HIV-1 Gag, Pol and Nef proteins, as a consequence, animals may raise anti-HIV responses which may be boosted by exposure to SP obtained from HIV-1 infected donors. However, anti-HIV Gag IgG titers in the SP-exposed group (847 ± 714) were not significantly different from those of the control group (155 ± 54) (Figure S1C). HIV Gag-specific IgG were detected in sera but not in vaginal fluids (Figure S1F). Overall, these results suggest that SP exposures do not significantly modify the vaccine-specific humoral responses.
We evaluated the MVA-specific CD4+ T cell response in the blood, lymph nodes (LNs) and FRT following ex-vivo re-stimulation of isolated leukocytes. HIV-Gag-specific CD4+ T-cell responses were very weak and mainly detected in the blood. No differences between the two groups were detected (data not shown). MVA-specific CD4+ T-cells were mainly found in the blood and axillary LNs draining the vaccine injection site (Figure S2A), as previously described (24). No differences between the two groups were detected in these compartments. Within the FRT of the control animals, expression of CD154, used as a marker of activated cells, was significantly increased only in CD4+ T cells of the cervix (from 0.06 to 0.50% of total CD4+ T cells) following stimulation with MVA and relative to unstimulated cells (Figure 1A, left panel). Also not statistically significant, MVA-specific CD4+ T cells producing TNF-α or IFN-γ were found in the uterus of three of six control animals (Figures 1B,C, left panel). In contrast, MVA-specific CD4+ T-cells from the cervix and the uterus of all animals exposed to SP showed significantly increased CD154 expression (from 0.23 to 0.69%) (Figure 1A, right panel) associated with significantly increased production of TNF-α (from 0.03 to 0.30%) (Figure 1B, right panel), IFN-γ (from 0.04 to 0.22%) (Figure 1C, right panel), and IL-2 (from 0.02 to 0.21%) (Figure 1D, right panel). Percentage of IFN-γ producing CD4+ T cells was also increased in vagina of SP exposed animals. Also not statistically significant, unstimulated CD4+ T cells in the vagina of SP-exposed animals expressed a higher percentage of CD154 (from 0.21 to 1.00%) (Figure 1A, right panel) than those of control animals (from 0.06 to 0.32%) (Figure 1A, left panel), suggesting an effect of SP exposures on the basal activation state of vaginal CD4+ T cells.
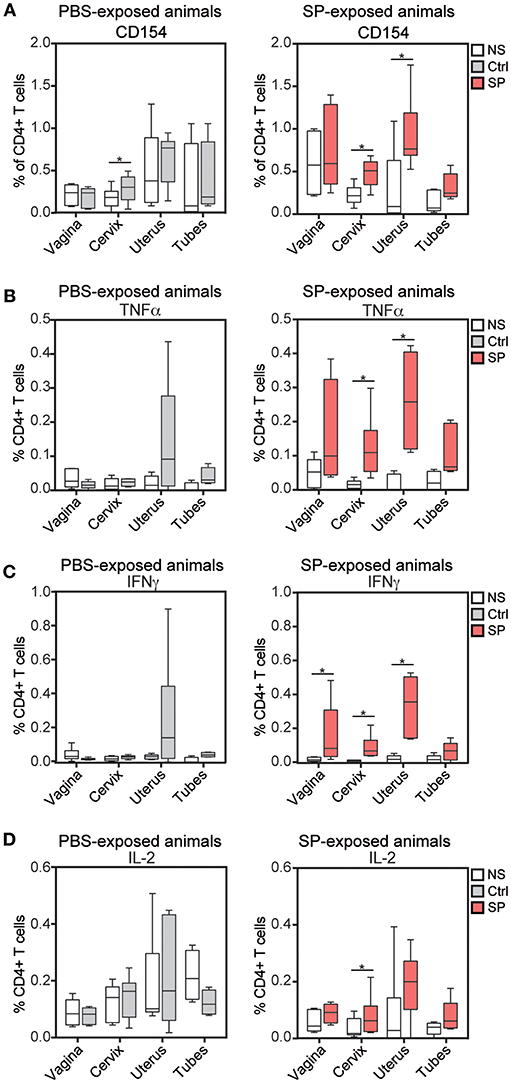
Figure 1. SP exposure enhances vaccine-specific mucosal CD4+ T cell responses. Percentage of CD154+ (A), TNF-α+ (B), IFN-γ+ (C), and IL-2+ (D) cells among CD4+ T cells after in vitro stimulation with medium (white) or wt MVA (gray for control animals, left panel and red for SP exposed animals, right panel) in FRT tissues. The Wilcoxon signed rank test (*) was used to compare NS vs. wt MVA stimulation antigens (*p < 0.05).
Overall, SP exposures appeared to enhance anti-MVA response mediated by CD4+ T cells.
We monitored the vaccine response mediated by CD8+ T cells in the blood, LNs and FRT of the control and SP-exposed animals using ex vivo re-stimulation assay. HIV-Gag-specific CD8+ T-cell responses were very weak and detected in the blood of few animals. No differences between the two groups were detected (data not shown). MVA-specific CD8+ T cell responses in the blood and LNs were detectable, but not different, between the two groups (Figure S2B). By contrast, percentages of IFN-γ (from 0.27 to 1.09% of total CD8+ T cells), MIP-1β (from 0.80 to 1.55%), TNF-α (from 0.11 to 0.53%) and IL-2-producing (from 0.06 to 0.24%) CD8+ T cells were statistically higher in the cervix of SP exposed animals compared to controls (Figure 2A). The percentage of MVA-specific uterine IL-2+ CD8+ T cells were also statistically higher in SP-exposed animals (Figure 2A). Higher proportion of polyfunctional MVA specific CD8+ T-cells, measured as the number of cytokines produced by a single cell, were detected in vaginal, cervical, and uterine of the SP-exposed animals compared to controls (Figure 2B), including those cells producing all four cytokines (IFN-γ, TNF-α, MIP-1β, and IL-2) (Figure 2C) and three cytokines (Figures 2C,D). Overall, these results provide evidence that SP exposure increase both the strength and the quality of the response against vaccine vector mediated by FRT CD8+ T cells.
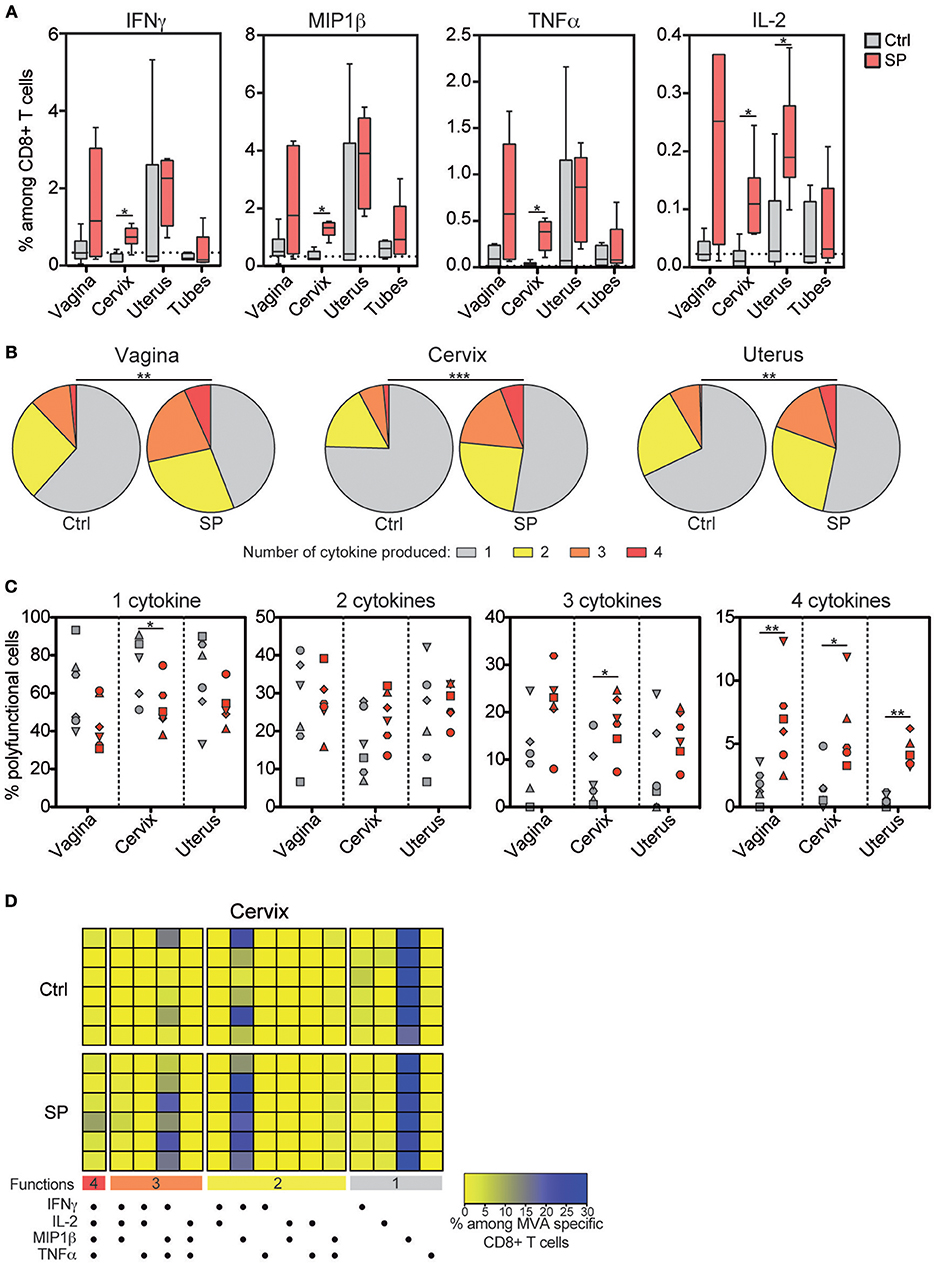
Figure 2. Frequency and quality of vaccine-specific mucosal CD8+ T cell responses increase after SP exposures. (A) Percentage of IFN-γ+, MIP-1β +, TNF-α+, and IL-2+ cells among MVA specific CD8+ T cells in FRT tissues after in vitro stimulation. Control animals (Ctrl) are indicated in gray and SP exposed animals (SP) in red. The horizontal dotted line indicates the mean background signal measured in unstimulated cells. (B) Pie chart graphs indicating the number of cytokines produced by the MVA-specific CD8+ T cells in the mucosal compartments. The means of the six animals of each group are represented in the pie charts. (C) Number of cytokines produced among the MVA specific CD8+ T cells in control (gray) and SP exposed (red) animals. Each symbol represents an animal. (D) Polyfunctional CD8+ T-cell profiles analyzed by Boolean gating in cervical tissue are represented as a heat map. Each colored horizontal line indicates one animal [n = 6 for the control group (Ctrl) and n = 6 for the SP exposed group (SP)]. The Mann-Whitney (*) was used to compare animal groups (*p < 0.05) and the Chi-squared test to compare the pie charts (**p < 0.005 and ***p < 0.001).
Our results show that SP exposures affect cellular immunity in the FRT. We therefore hypothesize that this could be the result of higher stimulation and/or recruitment of FRT immune cells, including lymphocytes and antigen presenting cells (APC). We thus characterized the immune cell composition in the various FRT compartments of the animals of both groups. The percentage of leukocytes (CD45+ cells) and CD4+ T cells were not affected by SP, except in the uterus (Figures 3A,B). However, the frequency of CD8+ T cells were significantly increased in the iliac LN of the SP-exposed animals (23.7 ± 2.7%; mean ± SEM) compared to controls (15.6 ± 2.7%) (Figure 3C). Analysis of the innate immunity components showed the percentage of NK cells and neutrophils to be similar in both groups (Figures 3D,E), except in the uterus, for which the percentage of neutrophils was significantly lower in the SP-exposed group (1.8 ± 0.3%) than in control group (4.0 ± 0.8%). The frequency of dendritic cells was higher in the vaginal, cervical, and uterine tissues of the SP-exposed group (8.4 ± 0.8, 7.2 ± 0.6, and 25 ± 3.5%, respectively) compared to controls (respectively 4.3 ± 0.6%, 4.6 ± 0.6% and 13.8 ± 3.8%) (Figure 3F). Phenotypic characterization of the dendritic subpopulations showed increased recruitment of mDC (CD11c+) after SP exposures (Figure 3G). In situ observations showed that CD11c+ cells were mainly localized to the subepithelial area (Figure 3H). Altogether, these results show that SP exposures induce the recruitment of lymphoid and myeloid dendritic cells in all FRT compartments.
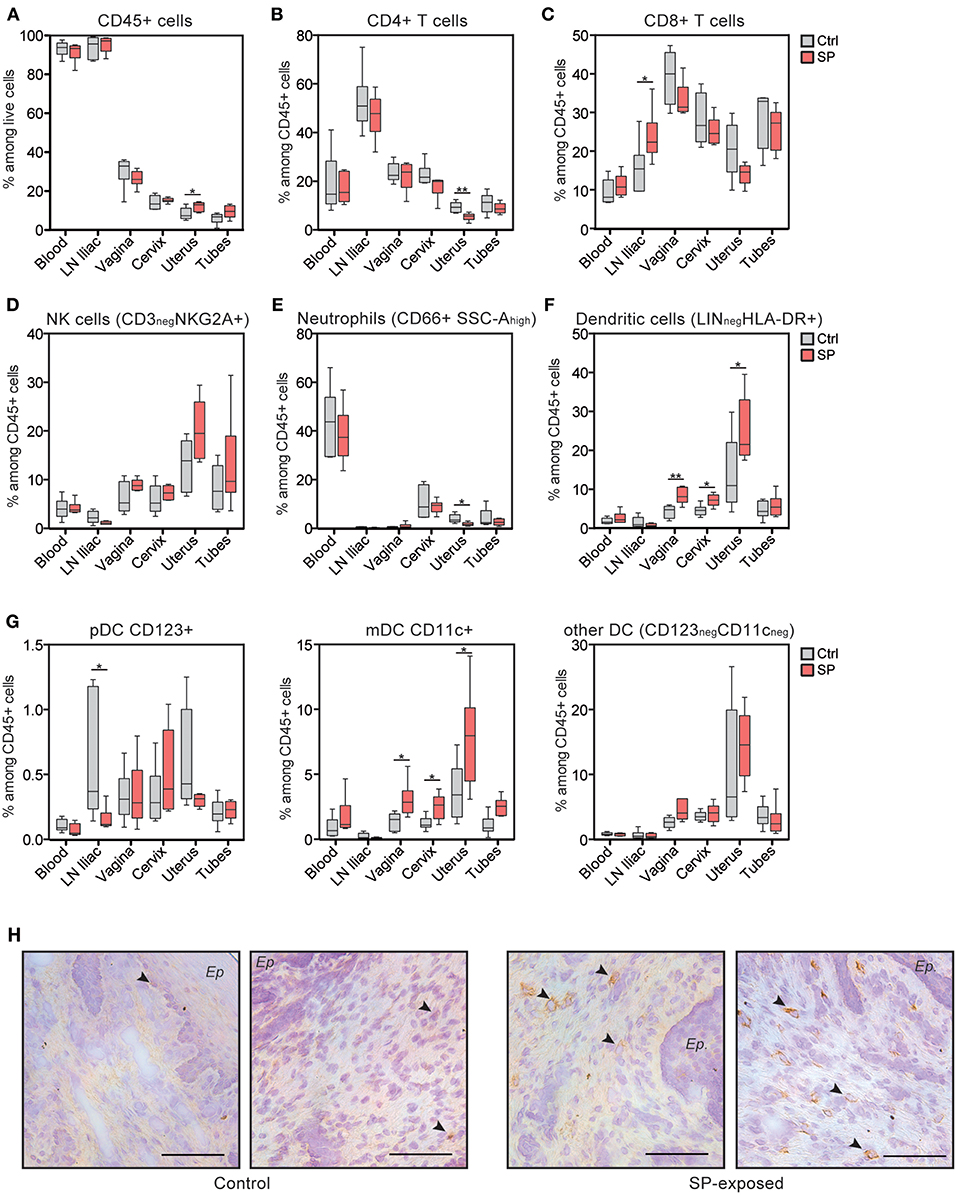
Figure 3. CD11c+ mDC are recruited to the FRT of SP exposed animals. (A) Distribution of leukocytes among living cells. Percentage of CD4+ T cells (B), CD8+ T cells (C), NK cells (D), neutrophils (E), and total dendritic cells (F) among leukocytes in the various compartments. (G) Percentage of dendritic cell subtypes [i.e., plasmacytoid DC (pDC), mDC, and other DC]. Control animals (Ctrl) are indicated in gray and SP exposed animals (SP) in red. The Mann-Whitney test (*) was used to compare animal groups (*p < 0.05). (H) Localization of mDC with anti-CD11c antibody in vaginal tissue sections from control (left panel) and SP-exposed animals (right panel). Black arrows indicate brown CD11c+ cells and the black bar indicates 100 μm. Ep, epithelium.
We compared the transcriptomic profiles of axillary and iliac LNs, vaginal, cervical, and uterine tissues from SP-exposed and control animals. SP exposures modified the expression of 316 genes in the iliac LNs, 134 in the vagina, 80 in the axillary LNs, 339 in the cervix and 270 in the uterus (Figure 4A). Multidimensional scaling (MDS) representation, constructed based on the set of genes found to be differentially expressed (DEG) in a least comparison, showed that the samples from the FRT are segregated and distinct from LNs genes (Figure 4B). We focused in a first step, our analyses on the cervix, uterus and iliac LNs since the number of DEG in the vagina and axillary LNs were lower relative to other tissues. The Venn diagram underscores the specificity of the response of the iliac LNs, cervix, and uterus to SP exposures, as the number of common DEG was low (Figure 4C). The relative expression of DEG in the iliac LNs, cervix, and uterus are represented by a heatmap in Figures 4D,E and Figure S3, respectively. We separately analyzed downregulated and up-regulated gene sets resulting from SP exposures using Ingenuity Pathway Analysis (IPA). Functional enrichment of the most significant canonical pathways and upstream regulators (p < 0.003) are detailed under each heatmap. In the iliac LNs, SP exposures downregulated genes associated with stress responses and cell activation (PI3K/AKT signaling, oxidative stress response, TNF, PMA, estrogen, etc.), as well as genes associated with immunomodulation (LPS/IL-1 mediated inhibition of RXR function, tretinoin, TGFB1, dexamethasone, etc.) (Figure 4D). In contrast, up-regulated genes were associated with TCR signaling. The same analyses performed on the cervix showed that SP exposures down-regulated genes associated with cell-cell interactions (ILK, integrin, and tight junction signaling) and those associated with endocytosis and macropinocytosis (clathrin or caveolar endocytosis and Rho-associated pathways) (Figure 4E). In contrast, SP exposures of the cervix upregulated genes associated with antigen presentation (antigen presentation pathway and protein ubiquitination), and upstream regulators involved in immune function and inflammation (IRF1 and IRF2, IFNG, and IFNA2) (Figure 4E). The upstream regulators of these DEG are associated with immune cell functions and activation and inflammatory disease (i.e., rheumatic disease) (Figure 4F). Analyses performed on the uterus showed an effect on upstream regulators associated with the PIK3/AKT pathway, hormones (progesterone), and angiogenic factors (Vascular endothelial growth factor) (Figure S3); however, no significant canonical pathway was found except that of calcium signaling. Similarly, only one pathway related to glycolysis was significantly upregulated in the vagina and found to be statistically over-represented (p < 0.0001). Overall, these results show that SP exposures of the FRT mostly affect the expression of genes of the cervix.
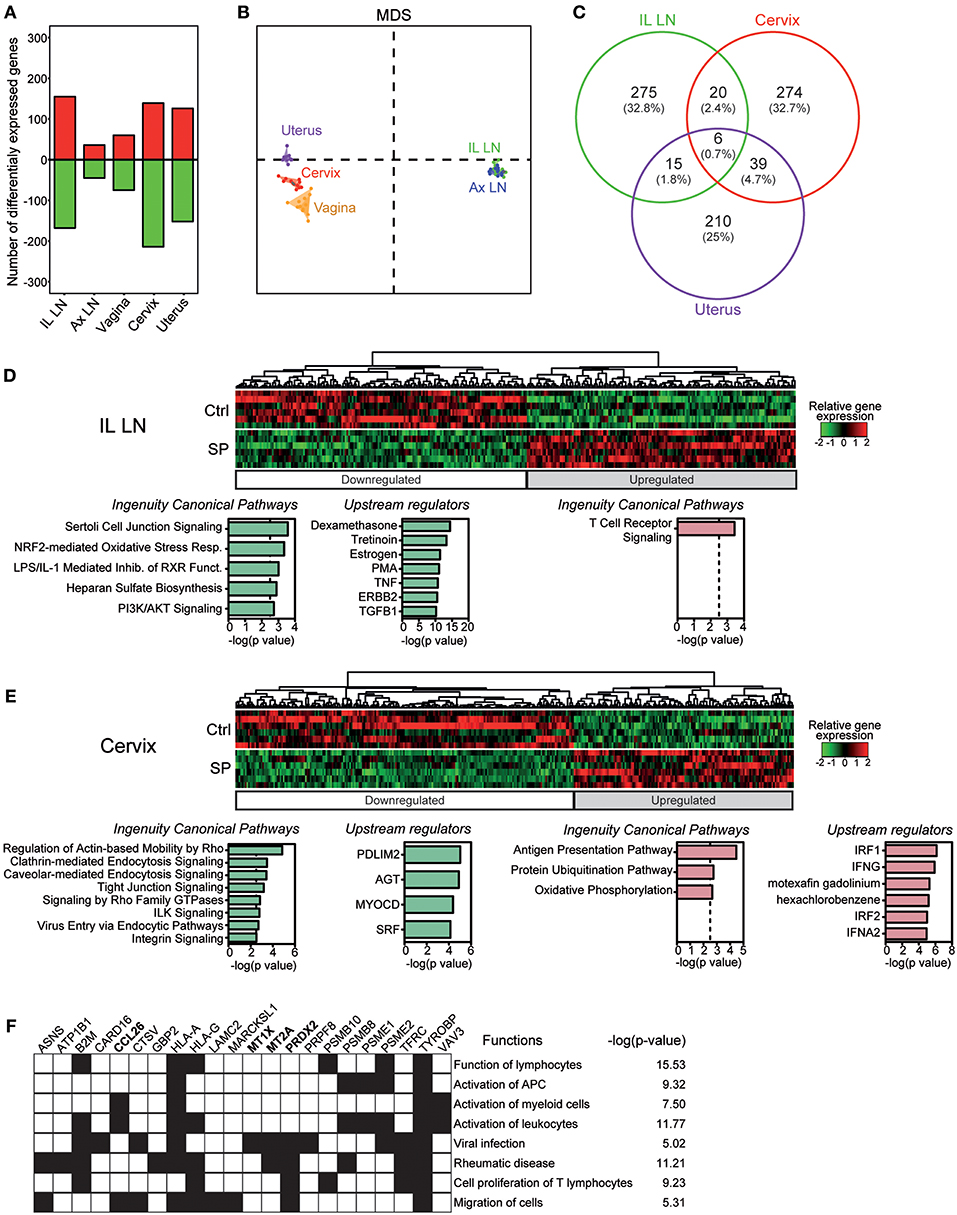
Figure 4. SP exposures impact mainly immune-related gene expression in the cervix. (A) Number of down-regulated (green) and upregulated (red) genes affected by SP exposures in the various compartments. (B) MDS representation of the whole dataset based on the DEG list for at least one condition. (C) Venn diagram showing the overlap between the list of DEG in the iliac LN, cervix and uterus. Heatmaps of relative gene expression in the iliac LN (D) and cervix (E). Associated canonical pathways and upstream regulators for down-regulated (green) and up-regulated (red) genes are detailed. (F) Biological processes found to be significantly over-represented (p < 0.05) in the lists of up-regulated genes targeted by upstream regulators in cervix. The main associated functions are shown.
We generated a correlation analysis to integrate flow cytometry and transcriptomic data. We restricted the analysis to positive correlations between DEG found in the FRT compartments (vagina, cervix and uterus) and mDCs, which were the cells recruited to the FRT after SP exposures (Figure 5A). Seventy-two genes had an expression profile correlated with the mDC abundance. Among these genes, 16 are known to be involved in immune responses or oxidative stress (Figure 5A in yellow). The expression of some of the same genes increased following SP exposures. For example, the expression of PRDX2 and the metallothioneins (MTE and MT1X), which are involved in protection against oxidative stress (27), positively correlated with the presence of mDCs and increased in cervical tissue following SP exposures (Figure 5B). In addition, these genes are also controlled by upstream regulators associated with cervical upregulated DEG (Figure 4F). The expression of PRDX2 and ACVR2A, a receptor involved in the activin/follistatin pathway (28), also increased in vaginal tissue following SP exposures (Figure 5B).
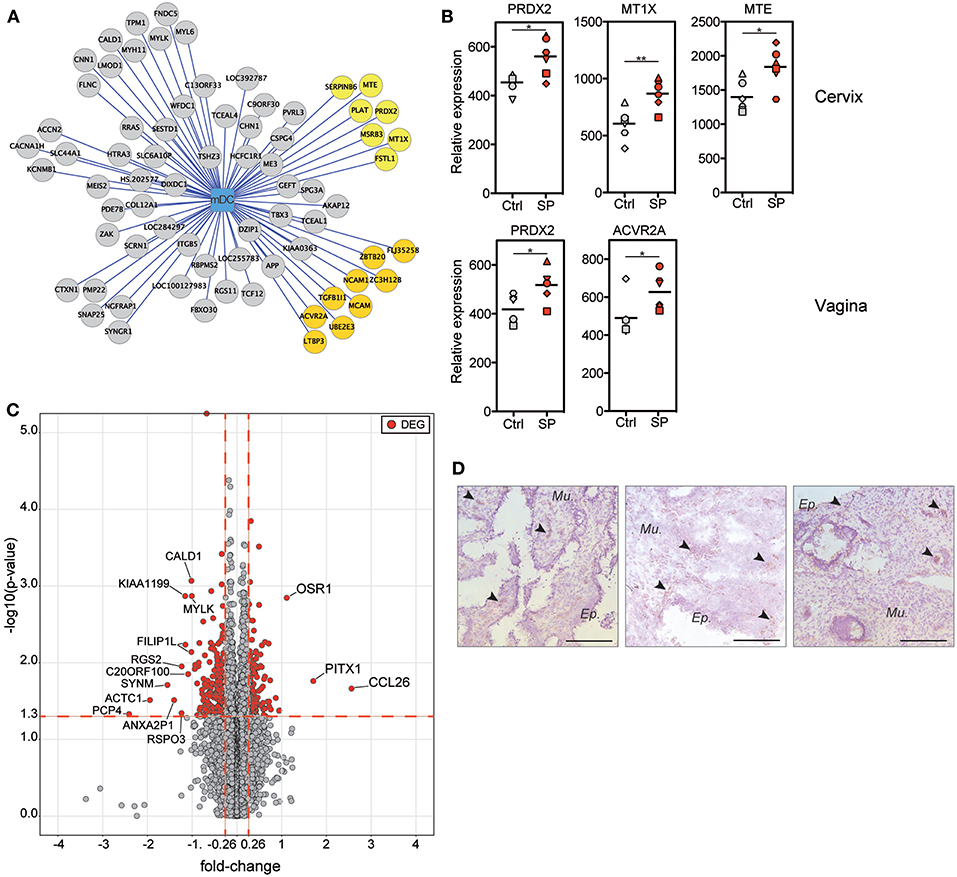
Figure 5. Upregulation of inflammation-related gene expression is associated with mDC recruitment after SP exposures. (A) Graph showing the correlations between the mDC abundance and DEG expression in the FRT (vagina, cervix, and uterus). Each node of the graph corresponds to a biological variable and links between the nodes correspond to significant positive correlations (Spearman correlation coefficient). Genes are represented by circles and the mDC cell population by the square. Gene circles colored in yellow are known to be involved in immune-related process. (B) Relative expression of inflammation-related genes identified in (A) that are upregulated by SP exposures. Each symbol represents one animal from control (gray) or SP group (red). The Mann-Whitney test (*) was used to compare gene expression between the two groups (*p < 0.05). (C) Volcano plots showing fold-change (FC, x axis) and statistical significance distribution (–log(p-value), y axis) for DEG between to PBS or SP exposure in the cervix. (D) Location of CCL26-producing cells in cervical tissue sections from SP exposed animals (n = 3). Black arrows indicate brown CCL26+ cells and the black bar indicates 100 μm.
Our results show that the effects of SP exposures on immunity are mainly localized in cervical tissue. We thus represented the DEG by volcano plots to identify potential biomarkers associated with SP exposures in the cervix (Figure 5C). Three genes were strongly upregulated by SP exposures: OSR-1, PITX-1, and CCL26. The OSR-1 and PITX-1 genes code for transcriptional regulators (29, 30). The CCL26 gene is controlled by upstream regulators associated with cervical upregulated DEG (Figure 4F). CCL26 is a chemokine involved in the regulation of inflammatory processes (31). Immunohistochemistry staining of SP-exposed cervical tissue showed CCL26 producing cells localized both within the epithelium and the mucosa (Figure 5D).
Overall, our transcriptomic analyses provide evidence that the effect of SP exposures in the cervix is predominantly on inflammatory responses that are associated with mDC recruitment.
We set up a model of repeated vaginal exposures to HIV-1 SP in MVA-HIV-B vaccinated NHPs to demonstrate the effect of semen during HIV-1 prevention studies, such as vaccination and challenge studies.
Our study characterized the impact of SP on local immunity in the FRT. Here, we showed recruitment of both CD8+ T cells to the LNs draining the FRT and CD11c+ dendritic cells to all compartments of the FRT. In addition to altering the proportion of mucosal immune cells, SP exposures also affected antigen-specific cellular responses. Indeed, HIV-1+ SP exposures increased responses against vaccine vector mediated by CD4+ and CD8+ T cells. SP clearly influenced the activation of immune cells as there was an increase in CD154 marker expression in vaginal tissue exposed to SP, even before in vitro restimulation with MVA. Moreover, SP exposures showed anti-MVA responses mediated by CD4+ T-cells mainly in the cervix. This CD4+ T-cell response was not observed in unexposed mucosae.
The anti-MVA CD8+ T cell response was also affected by SP exposures, once again mainly in the cervix. Indeed, the frequency of anti-MVA CD8+ T cells producing IFN-γ, MIP-1β, TNF-α, or IL-2 increased after SP exposures. Antigen-specific CD8+ T cells were specifically recruited following SP exposures, as the percentage of total CD8+ T cells was not modified. Such recruitment could be due to the pro-inflammatory chemokines found in HIV-1+ SP (direct effect) (21) or factors produced by SP-stimulated FRT epithelial cells (indirect effect) (32). In addition to affecting the strength, SP exposures also affected the quality of the response mediated by anti-MVA CD8+ T cells in the FRT. Thus, MVA-specific CD8+ T cells producing three or four cytokines per cell were more frequent in the cervix after SP exposures. This increase may be related to the inflammatory environment induced by SP. Indeed, pro-inflammatory cytokines in HIV-1+ SP could attract cells carrying a co-stimulatory signal for CD8+ T cells. Co-stimulatory signals for T cells are classically provided by antigen presenting cells (33). HIV-1+ SP exposures induced specific recruitment of CD11c+ DC, which could affect MVA-specific CD8+ T cell stimulation. Our enrichment analysis of the transcriptional profile from cervical tissue showed that endocytosis-related pathways were downregulated by SP exposures, whereas the antigen presentation pathway was significantly upregulated. This pattern suggests a dendritic cell maturation profile, as mature dendritic cells have a lower endocytic capacity than immature dendritic cells associated with antigen presentation specialization (34). Thus, the association of pro-inflammatory cytokines of HIV-1+ SP and the recruitment of mature CD11c+ APC could be responsible for the higher quality of the CD8+ T cell response after SP exposure. However, APC recruited after HIV-1+ SP exposures could locally constitute new target cells for HIV-1. Moreover, antigen-specific CD4+ T cells detected after SP exposures could also constitute target cells for the virus. Thus, a challenge study following HIV-1+ SP exposures may help to determine whether the effects that we characterized increase protection against or susceptibility to HIV-1 infection.
Although we detected MVA-specific humoral responses in body fluids, we did not observe an effect of SP exposures on levels of anti-MVA antibodies in vaginal secretion. Further analyses to determine whether SP exposures affect the functions of secreted anti-MVA immunoglobulin, such as its neutralization capacity, may be informative.
Transcriptomic analyses showed that the expression of inflammation-related chemokine CCL26 and factors involved in protection against oxidative stress and inflammation, such as PRDX2 and metallothioneins (MTE, MT1X, and MT2A) significantly increased after SP exposures. Upregulation of these factors suggests that our analyses were performed after the peak of inflammation. Indeed, our analyses were performed 24 h after the last SP inoculation, and thus we may have missed the upregulation of early inflammatory genes such as COX-2 (10), and rapidly recruited immune cells, such as neutrophils (35). Our results are consistent with those of previous studies performed in women, which showed a local effect of HIV-1neg SP on cervical immune cells and inflammation (10).
Previous in vitro studies reported an opposite effect of SP exposure on lymphocyte immunity (36, 37). Indeed, it has been shown that SP downregulated T cell and NK cell functions, such as cytotoxicity and cytokine production. The origins of the cells may explain this discrepancy with our results. The inhibitory effect of SP on T cells was observed after direct contact of SP with cells from peripheral blood (36). In the lower FRT, SP contact with T cells could be facilitated due to microtrauma of the epithelium during intercourse. However, the intraepithelial T cells are not abundant in the lower tract (38) so SP interacts mainly with epithelial cells (10). Thus, SP stimulates epithelial cells, which then produced soluble factors, inducing cell recruitment and T-cell activation (32, 39, 40). SP could also directly stimulate the DC found in FRT epithelium. In vitro studies have demonstrated that SP induces the production of soluble factors by epithelial cells, including pro-inflammatory cytokines (41). Among these, follistatin, also highlighted by our study is of great interest. Indeed, the function of this glycoprotein is to bind and inhibit members of the TGFβ family, in particular, activin (28). The loop of activin/follistatin process appeared in our transcriptional analysis on FRT as expression of the activin A receptor type A2 (ACVR2A gene) that was upregulated after SP exposures and correlated with CD11c+ mDC recruitment (Figures 5A,B). Furthermore, high concentrations of follistatin are found in SP (42) and SP regulates the synthesis of activin and follistatin (43). Moreover, follistatin has been shown to increase the function of antigen-specific CD8+ T cells (44). Future studies to investigate the role of follistatin in the upregulation of MVA specific CD8+ T cell function after SP exposures in cervical tissue may be informative.
In conclusion, our combined analysis of molecular and cellular events shows that HIV-1+ SP affects mainly on cervical immune cells and MVA-specific T cell responses. Further studies are required to characterize the role of each protagonist (i.e. factors in SP, epithelial cells, and HIV-1+ particles) in the increase of the strength and quality of vaccine-CD8+ T cell response after SP exposures.
Twelve sexually mature adult female cynomolgus macaques (Macaca fascicularis) imported from Mauritius were housed in the Infectious Disease Models and Innovative Therapies (IDMIT) facilities at the Commissariat à l'Energie Atomique et aux Energies Alternatives (CEA, Fontenay-aux-Roses, France). The treatment of NHPs at the CEA complies with French national regulations (CEA authorization A 92-032-02), the Standards for Human Care and Use of Laboratory Animals (OLAW Assurance number #A5826-01), and European Directive 2010/63 (recommendation #9). Experiments were supervised by veterinarians in charge of the animal facility. This study was approved and accredited by the Ethics Committee on Animal Experimentation (Comité d'Ethique en Expérimentation Animale) of the CEA (A14-080) and the French Research Ministry. Animals were housed in pairs under controlled humidity, temperature and light (12-h light/dark cycles). Water was available ad libitum. The animals were monitored and fed once or twice a day with commercial monkey chow and fruits, by trained personnel, and were provided with environmental enrichment including toys, novel foodstuffs and music, under the supervision of the CEA Animal Welfare Officer.
RM, YL, FB-S, GS, RL, and EM: study conception and design. SN and ND-B: resources. RM, M-TN, MP, CL, HH, CC, and ND-B: acquisition of data. RM, NT, GS, RL, and EM: analysis and interpretation of data. RM, M-TN, NT, RL, and EM: drafting of manuscript. M-TN, NT, FB, YL, RL, and EM: critical revision.
NT held a fellowship from the ANRS (France Recherche Nord & Sud Sida-HIV Hépatites). FB held fellowships from the ANRS, Sidaction and Institut Carnot-Pasteur, Microbes & Santé. This work was supported by the Programme Investissement d'Avenir (PIA) managed by the ANR under reference ANR-11-INBS-0008, funding the Infectious Disease Models and Innovative Therapies (IDMIT, Fontenay-aux-Roses, France) infrastructure, the ANR-10-EQPX-02-01, funding the FlowCyTech facility (IDMIT, Fontenay-aux-Roses, France) and the ANR-10-LABX-77-01 funding the Vaccine Research Institute (VRI). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank Drs. Anne-Sophie Beignon, Mireille Centlivre, and the Bmuc division members for scientific discussions. Alex Edelman & Associates for critical editing of the manuscript. Transgene for providing the wild type MVA strain and ANRS/INSERM and the Vaccine Research Institute the MVA-HIV-B vaccine. Andrea Galli for preparation of SP pool. Rahima Yousfi for technical assistance. Dr. Véronique Avettand-Fenoel for HIV-1 viral load quantification. Dr. Bernard Verrier for providing the p24 HIV-1 antigen and all members of the IDMIT infrastructure for their excellent expertise and outstanding contribution.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2019.00430/full#supplementary-material
1. Owen DH, Katz DF. A review of the physical and chemical properties of human semen and the formulation of a semen simulant. J Androl. (2005) 26:459–69. doi: 10.2164/jandrol.04104
2. Rodríguez-Martínez H, Kvist U, Ernerudh J, Sanz L, Calvete JJ. Seminal plasma proteins: what role do they play? Am J Reprod Immunol. (2011) 66:11–22. doi: 10.1111/j.1600-0897.2011.01033.x
3. Rolland AD, Lavigne R, Dauly C, Calvel P, Kervarrec C, Freour T, et al. Identification of genital tract markers in the human seminal plasma using an integrative genomics approach. Hum Reprod. (2013) 28:199–209. doi: 10.1093/humrep/des360
4. Southern PJ. Missing out on the biology of heterosexual HIV-1 transmission. Trends Microbiol. (2013) 21:245–52. doi: 10.1016/j.tim.2013.02.002
5. Robertson SA, Mau VJ, Tremellen KP, Seamark RF. Role of high molecular weight seminal vesicle proteins in eliciting the uterine inflammatory response to semen in mice. J Reprod Fertil. (1996) 107:265–77.
6. Robertson SA, Allanson M, Mau VJ. Molecular regulation of uterine leukocyte recruitment during early pregnancy in the mouse. Placenta. (1998) 19:101–19. doi: 10.1016/S0143-4004(98)80009-X
7. Joseph T, Zalenskaya IA, Sawyer LC, Chandra N, Doncel GF. Seminal plasma induces prostaglandin-endoperoxide synthase (PTGS) 2 expression in immortalized human vaginal cells: involvement of semen prostaglandin E2 in PTGS2 upregulation. Biol Reprod. (2013) 88:13. doi: 10.1095/biolreprod.112.101956
8. Pandya IJ, Cohen J. The leukocytic reaction of the human uterine cervix to spermatozoa. Fertil Steril. (1985) 43:417–21.
9. Johansson M, Bromfield JJ, Jasper MJ, Robertson SA. Semen activates the female immune response during early pregnancy in mice. Immunology. (2004) 112:290–300. doi: 10.1111/j.1365-2567.2004.01876.x
10. Sharkey DJ, Tremellen KP, Jasper MJ, Gemzell-Danielsson K, Robertson SA. Seminal fluid induces leukocyte recruitment and cytokine and chemokine mRNA expression in the human cervix after coitus. J Immunol. (2012) 188:2445–54. doi: 10.4049/jimmunol.1102736
11. O'Leary S, Jasper MJ, Warnes GM, Armstrong DT, Robertson SA. Seminal plasma regulates endometrial cytokine expression, leukocyte recruitment and embryo development in the pig. Reproduction. (2004) 128:237–47. doi: 10.1530/rep.1.00160
12. Robertson SA, Guerin LR, Bromfield JJ, Branson KM, Ahlström AC, Care AS. Seminal fluid drives expansion of the CD4+CD25+ T regulatory cell pool and induces tolerance to paternal alloantigens in mice. Biol Reprod. (2009) 80:1036–45. doi: 10.1095/biolreprod.108.074658
13. Anderson DJ, Politch JA, Nadolski AM, Blaskewicz CD, Pudney J, Mayer KH. Targeting trojan horse leukocytes for HIV prevention. AIDS. (2010) 24:163–87. doi: 10.1097/QAD.0b013e32833424c8
14. Kaizu M, Weiler AM, Weisgrau KL, Vielhuber KA, May G, Piaskowski SM, et al. Repeated intravaginal inoculation with cell-associated simian immunodeficiency virus results in persistent infection of nonhuman primates. J Infect Dis. (2006) 194:912–6. doi: 10.1086/507308
15. Khanna KV, Whaley KJ, Zeitlin L, Moench TR, Mehrazar K, Cone RA, et al. Vaginal transmission of cell-associated HIV-1 in the mouse is blocked by a topical, membrane-modifying agent. J Clin Invest. (2002) 109:205–11. doi: 10.1172/JCI13236
16. Sallé B, Brochard P, Bourry O, Mannioui A, Andrieu T, Prevot S, et al. Infection of macaques after vaginal exposure to cell-associated simian immunodeficiency virus. J Infect Dis. (2010) 202:337–44. doi: 10.1086/653619
17. Gupta P, Collins KB, Ratner D, Watkins S, Naus GJ, Landers DV, et al. Memory CD4+ T cells are the earliest detectable human immunodeficiency virus type 1 (HIV-1)-infected cells in the female genital mucosal tissue during HIV-1 transmission in an organ culture system. J Virol. (2002) 76:9868–76. doi: 10.1128/JVI.76.19.9868
18. Hladik F, Doncel GF. Preventing mucosal HIV transmission with topical microbicides: challenges and opportunities. Antiviral Res. (2010) 88:S3–9. doi: 10.1016/j.antiviral.2010.09.011
19. Bouvet J-P, Grésenguet G, Bélec L. Vaginal pH neutralization by semen as a cofactor of HIV transmission. Clin Microbiol Infect. (1997) 3:19–23.
20. Kim K-A, Yolamanova M, Zirafi O, Roan NR, Staendker L, Forssmann W-G, et al. Semen-mediated enhancement of HIV infection is donor-dependent and correlates with the levels of SEVI. Retrovirology. (2010) 7:55. doi: 10.1186/1742-4690-7-55
21. Doncel GF, Anderson S, Zalenskaya I. Role of semen in modulating the female genital tract microenvironment—implications for HIV transmission. Am J Reprod Immunol. (2014) 71:564–574. doi: 10.1111/aji.12231
22. Lisco A, Introini A, Munawwar A, Vanpouille C, Grivel JC, Blank P, et al. HIV-1 imposes rigidity on blood and semen cytokine networks. Am J Reprod Immunol. (2012) 68:515–21. doi: 10.1111/aji.12015
23. Kafka JK, Sheth PM, Nazli A, Osborne BJ, Kovacs C, Kaul R, et al. Endometrial epithelial cell response to semen from HIV-infected men during different stages of infection is distinct and can drive HIV-1-long terminal repeat. AIDS. (2012) 26:27–36. doi: 10.1097/QAD.0b013e32834e57b2
24. Marlin R, Nugeyre M-T, Tchitchek N, Parenti M, Hocini H, Benjelloun F, et al. Modified vaccinia virus ankara vector induces specific cellular and humoral responses in the female reproductive tract, the main HIV portal of entry. J Immunol. (2017) 199:1923–32. doi: 10.4049/jimmunol.1700320
25. Guenounou S, Bosquet N, Dembek CJ, Le Grand R, Cosma A. OMIP-016: characterization of antigen-responsive macaque and human T-cells. Cytom Part A. (2013) 83A:182–4. doi: 10.1002/cyto.a.22233
26. Marx PA, Spira AI, Gettie A, Dailey PJ, Veazey RS, Lackner AA, et al. Progesterone implants enhance SIV vaginal transmission and early virus load. Nat Med. (1996) 2:1084–9.
27. Schulkens IA, Castricum KCM, Weijers EM, Koolwijk P, Griffioen AW, Thijssen VL. Expression, regulation and function of human metallothioneins in endothelial cells. J Vasc Res. (2014) 51:231–8. doi: 10.1159/000365550
28. Wijayarathna R, de Kretser DM. Activins in reproductive biology and beyond. Hum Reprod Update. (2016) 22:342–57. doi: 10.1093/humupd/dmv058
29. Katoh M. Molecular cloning and characterization of OSR1 on human chromosome 2p24. Int J Mol Med. (2002) 10:221–5. doi: 10.3892/ijmm.10.2.221
30. Mudie S, Bandarra D, Batie M, Biddlestone J, Moniz S, Ortmann B, et al. PITX1, a specificity determinant in the HIF-1α-mediated transcriptional response to hypoxia. Cell Cycle. (2014) 13:3878–91. doi: 10.4161/15384101.2014.972889
31. Petkovic V, Moghini C, Paoletti S, Uguccioni M, Gerber B. Eotaxin-3/CCL26 is a natural antagonist for CC chemokine receptors 1 and 5. A human chemokine with a regulatory role. J Biol Chem. (2004) 279:23357–63. doi: 10.1074/jbc.M309283200
32. Sharkey DJ, Macpherson AM, Tremellen KP, Robertson SA. Seminal plasma differentially regulates inflammatory cytokine gene expression in human cervical and vaginal epithelial cells. Mol Hum Reprod. (2007) 13:491–501. doi: 10.1093/molehr/gam028
33. Chen L, Flies DB. Molecular mechanisms of T cell co-stimulation and co-inhibition. Nat Rev Immunol. (2013) 13:227–42. doi: 10.1038/nri3405
34. Platt CD, Ma JK, Chalouni C, Ebersold M, Bou-Reslan H, Carano RAD, et al. Mature dendritic cells use endocytic receptors to capture and present antigens. Proc Natl Acad Sci USA. (2010) 107:4287–92. doi: 10.1073/pnas.0910609107
35. Thompson LA, Barratt CL, Bolton AE, Cooke ID. The leukocytic reaction of the human uterine cervix. Am J Reprod Immunol. (1992) 28:85–9.
36. Selva KJ, Kent SJ, Parsons MS. Modulation of innate and adaptive cellular immunity relevant to HIV-1 vaccine design by seminal plasma. AIDS. (2017) 31:333–42. doi: 10.1097/QAD.0000000000001319
37. Meuleman T, Snaterse G, van Beelen E, Anholts JDH, Pilgram GSK, van der Westerlaken LAJ, et al. The immunomodulating effect of seminal plasma on T cells. J Reprod Immunol. (2015) 110:109–16. doi: 10.1016/j.jri.2015.01.012
38. Pudney J, Quayle AJ, Anderson DJ. Immunological microenvironments in the human vagina and cervix: mediators of cellular immunity are concentrated in the cervical transformation zone. Biol Reprod. (2005) 73:1253–63. doi: 10.1095/biolreprod.105.043133
39. Moldenhauer LM, Keenihan SN, Hayball JD, Robertson SA. GM-CSF is an essential regulator of T cell activation competence in uterine dendritic cells during early pregnancy in mice. J Immunol. (2010) 185:7085–96. doi: 10.4049/jimmunol.1001374
40. Chen JC, Johnson BA, Erikson DW, Piltonen TT, Barragan F, Chu S, et al. Seminal plasma induces global transcriptomic changes associated with cell migration, proliferation and viability in endometrial epithelial cells and stromal fibroblasts. Hum Reprod. (2014) 29:1255–70. doi: 10.1093/humrep/deu047
41. Sharkey DJ, Macpherson AM, Tremellen KP, Mottershead DG, Gilchrist RB, Robertson SA. TGF- mediates proinflammatory seminal fluid signaling in human cervical epithelial cells. J Immunol. (2012) 189:1024–35. doi: 10.4049/jimmunol.1200005
42. Anderson RA, Evans LW, Irvine DS, McIntyre MA, Groome NP, Riley SC. Follistatin and activin A production by the male reproductive tract. Hum Reprod. (1998) 13:3319–25. doi: 10.1093/humrep/13.12.3319
43. Sharkey DJ, Robertson SA. 421. Seminal fluid TGFβ regulates follistatin mRNA expression human Ect1 cervical epithelial cells. Reprod Fertil Dev. (2008) 20:101. doi: 10.1071/SRB08Abs421
Keywords: female reproductive tract (FRT), seminal plasma, vaccine, mucosa, HIV-1
Citation: Marlin R, Nugeyre M-T, Tchitchek N, Parenti M, Lefebvre C, Hocini H, Benjelloun F, Cannou C, Nozza S, Dereuddre-Bosquet N, Levy Y, Barré-Sinoussi F, Scarlatti G, Le Grand R and Menu E (2019) Seminal Plasma Exposures Strengthen Vaccine Responses in the Female Reproductive Tract Mucosae. Front. Immunol. 10:430. doi: 10.3389/fimmu.2019.00430
Received: 23 November 2018; Accepted: 18 February 2019;
Published: 12 March 2019.
Edited by:
Cecil Czerkinsky, Institut National de la Santé et de la Recherche Médicale (INSERM), FranceReviewed by:
Pam Kozlowski, LSU Health Sciences Center New Orleans, United StatesCopyright © 2019 Marlin, Nugeyre, Tchitchek, Parenti, Lefebvre, Hocini, Benjelloun, Cannou, Nozza, Dereuddre-Bosquet, Levy, Barré-Sinoussi, Scarlatti, Le Grand and Menu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Elisabeth Menu, ZWxpc2FiZXRoLm1lbnVAcGFzdGV1ci5mcg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.