 Woo Jae Shin
Woo Jae Shin Brian A. Zabel
Brian A. Zabel Russell K. Pachynski
Russell K. Pachynski- 1Division of Oncology, Washington University School of Medicine, St. Louis, MO, United States
- 2Palo Alto Veterans Institute for Research (PAVIR), VA Palo Alto Health Care Systems (VAPAHCS), Palo Alto, CA, United States
- 3The Andrew M. and Jane M. Bursky Center for Human Immunology & Immunotherapy Programs (CHiiPs), St. Louis, MO, United States
Chemerin [RARRES2 [retinoic acid receptor responder 2], TIG2 [tazarotene induced gene 2 (TIG2)]] is a multifunctional cytokine initially described in skin cultures upon exposure to the synthetic retinoid tazarotene. Its secreted pro-form, prochemerin, is widely expressed, found systemically, and is readily converted into active chemerin by various proteases. Subsequent studies elucidated major roles of chemerin as both a leukocyte chemoattractant as well as an adipokine. Chemerin's main chemotactic receptor, the G-protein coupled receptor CMKLR1, is expressed on macrophages, dendritic, and NK cells. With respect to its role in immunology, chemerin mediates trafficking of these cells to sites of inflammation along its concentration gradient, and likely helps coordinate early responses, as it has been shown to have antimicrobial and angiogenic properties, as well. Recently, there has been mounting evidence that chemerin is an important factor in various cancers. As with its role in immune responses—where it can act as both a pro- and anti-inflammatory mediator—the potential functions or correlations chemerin has in or with cancer appears to be context dependent. Most studies, however, suggest a downregulation or loss of chemerin/RARRES2 in malignancies compared to the normal tissue counterparts. Here, we perform a comprehensive review of the literature to date and summarize relevant findings in order to better define the roles of chemerin in the setting of the tumor microenvironment and tumor immune responses, with an ultimate focus on the potential for therapeutic intervention.
Introduction
Chemerin [also known as retinoic acid receptor responder 2 (RARRES2) or tazarotene induced gene 2 (TIG2)] is a multifunctional, chemoattractant protein known for its roles in adipogenesis, angiogenesis, skin function, metabolic activity, and, recently, tumorigenesis. Initially secreted by the liver and white adipose tissue as prochemerin, the 163-amino acid precursor protein is readily cleaved by a specific set of serine proteases to become a chemotactically-active protein isoform of chemerin (1). Depending on the site of cleavage and subsequent interaction with its cognate heptahelical receptors, CMKLR1 (chemokine-like receptor 1), CCRL2 (C-C chemokine receptor-like 2), and GPR1 (G protein-coupled receptor 1), chemerin may exhibit varying degrees of bioactivity and elicit pro- or anti-inflammatory effects in different biological environments (1–3). In the literature, CMKLR1 is usually discussed as the primary receptor for chemerin interaction, while CCRL2 is described to participate in various functions by binding and presenting chemerin in a non-signaling manner to establish concentration gradients (4, 5). On the other hand, GPR1 is less-well characterized and found in the central nervous system (5) and reproductive organs (6, 7) and may play a role in metabolism (8).
Since its initial discovery in skin cultures upon stimulation with anti-psoriatic synthetic retinoid tazarotene, chemerin has been further described in a number of biological settings (9). In human endothelial cells, chemerin has been found to mediate angiogenesis via interactions with CMKLR1 (10), while, in the epidermis, chemerin has been shown to engage in significant antimicrobial activity (11). Moreover, chemerin has been linked with conditions such as obesity and diabetes, where it may modulate metabolism and adipocyte development (12, 13). Most recently, chemerin has been shown to mediate the chemoattraction of various immunocytes in the tumor microenvironment, while expression of chemerin's receptors, CMKLR1 and CCRL2, has been identified on a number of leukocyte subsets, namely, dendritic cell subsets, natural killer cells, and macrophages (1, 2, 14, 15). With recent findings that chemerin's receptors are also expressed on malignant tumor cells and that chemerin's expression is often altered in different cancer types, newer studies have focused on chemerin's novel roles in immune surveillance and tumor progression.
As expected, these recent studies have confirmed the notion that chemerin's functions in cancer are context driven. In some cancer types [e.g., glioblastoma, mesothelioma, neuroblastoma, squamous cell carcinoma of the esophagus, and squamous cell carcinoma of the oral tongue (SCCOT)], chemerin is upregulated (16–22). In most cancer types [e.g., acute myeloid leukemia (AML), adrenocortical carcinoma (ACC), breast cancer, Ewing sarcoma, hepatocellular carcinoma, melanoma, non-small cell lung cancer (NSCLC), prostate cancer, and squamous cell carcinoma of the skin] chemerin is downregulated, likely via hypermethylation of RARRES2 (15, 19, 23–28) (Table 1). Adding to this complexity, the ultimate effects on tumor growth are also context-dependent; both tumor suppression and accelerated growth have been observed as a result of altered chemerin expression levels (Table 2). Thus, in this review, we attempt to catalog and analyze chemerin's specific functions in each tumor type, with a focus on identifying patterns in its mechanism of action and suggesting ways in which the chemerin system may be utilized or manipulated for clinical benefit.
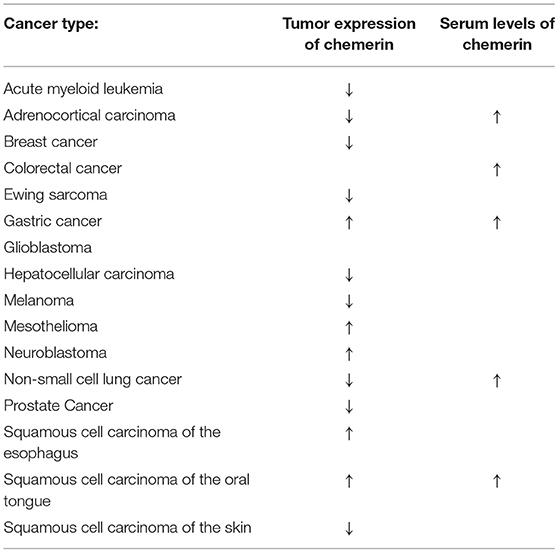
Table 1. Alterations of chemerin expression profiles by cancer type.
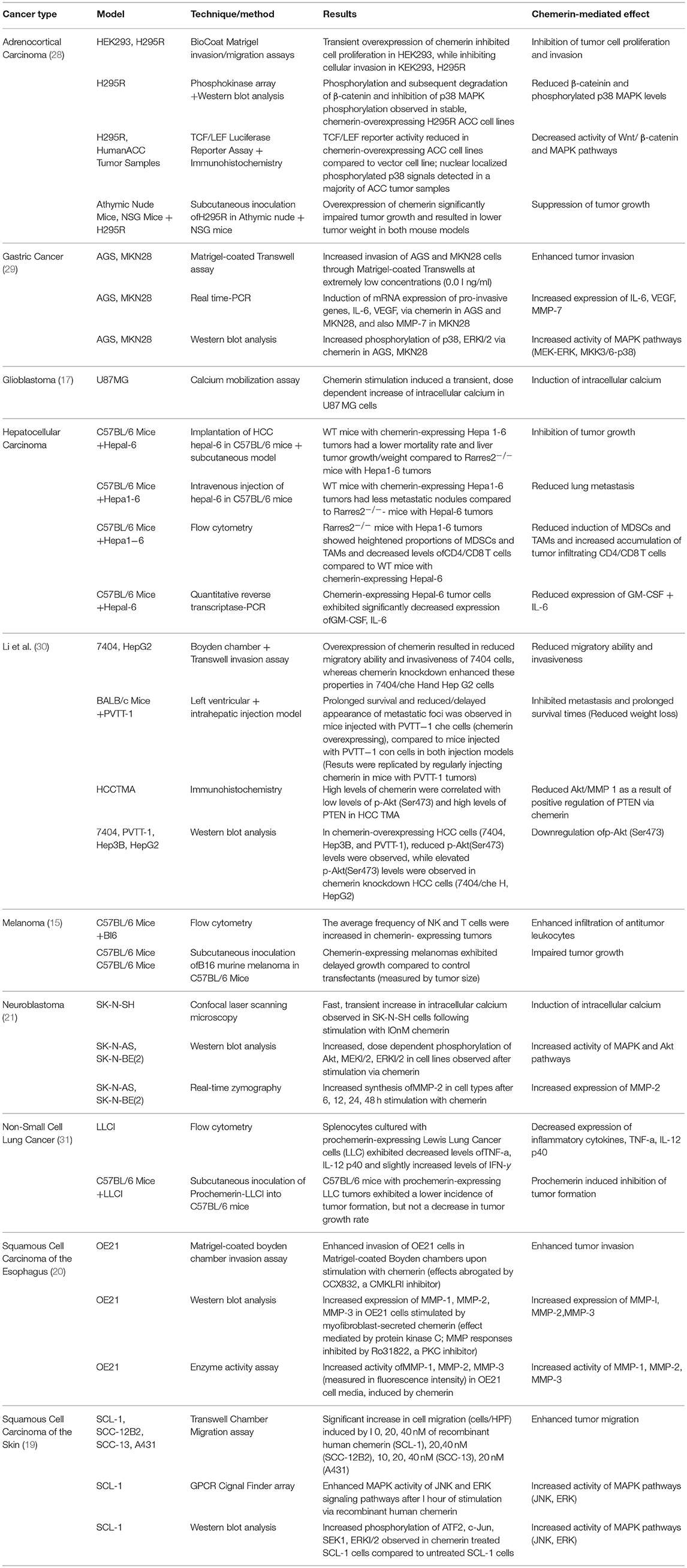
Table 2. Mechanisms of chemerin-mediated effects by cancer type.
Cancers
Acute Myeloid Leukemia
Acute myeloid leukemia (AML) is the most prevalent form of acute leukemia affecting adults, with a particularly high rate of incidence (12.2 cases per 100,000 people) for those above the age of 65 (32–34). Though recent advances in AML treatments have significantly improved prognosis for younger patients, those who are older have little chance of survival (35). In fact, a majority of AML patients over the age of 65 die within a year of diagnosis, underlying an urgent need for improved methods of detection that may enable earlier life-preserving treatment (36).
In light of this need, chemerin has recently been identified as a potential biomarker for diagnosis of AML. A study in 2017 revealed that chemerin was downregulated in the bone marrow mononuclear cells of AML patients compared to that of healthy controls, with ROC curve analysis showing a test specificity of 79% (true positive) and sensitivity of 54% (true negative) for chemerin expression as a marker for AML diagnosis. The same group performed a cohort study of 149 patients, 32 of whom had high chemerin expression and 117 of whom had low expression, with no significant variability in certain gene mutations, white blood cell count, platelets, and hemoglobin, and found that patients with low chemerin expression correlated with poorer overall survival. Moreover, multivariate analysis on parameters such as age, various gene mutations, chemerin expression, karyotypic classifications, and white blood cell count verified that chemerin was independently able to prognosticate AML patients, while univariate analysis of chemerin expression levels showed that high chemerin expression was associated with positive prognosis (22). In terms of chemerin receptor expression levels (independent of associations between chemerin expression and clinical outcome), a different group showed that CCRL2 was overexpressed in AML, identifying the non-classically signaling chemerin receptor as a potential therapeutic target, along with other GPCRs that were also overexpressed (37).
Adrenocortical Carcinoma
Adrenocortical carcinoma (ACC) is an extremely rare and aggressive tumor that is associated with poor prognosis in patients. The incidence rate for ACC is 0.7–2.0 cases per million people per year, and for patients that undergo first-line treatment (surgical resection), the 5-year median survival rate is 38.6%. For patients ineligible or unwilling to undergo adrenalectomy, there is little chance of remission (38–40). In fact, the potential risk of ACC is considered the standard for adrenalectomy in patients with adrenal incidentalomas (38).
Recent research suggests that serum chemerin levels have prognostic value in ACC and that manipulating chemerin levels in ACC tumors may prove efficacious in ACC patients. Chemerin's role as a chemoattractant, in recruiting immune cells to sites of inflammation, has already been well documented; for instance, chemerin has been shown to suppress neoplasia by eliciting natural killer cells to the tumor site in melanoma (15). In line with those findings, a study by Chittenden et al. (28) featuring a group of 20 ACC patients, 53 benign tumor patients, and 21 healthy individuals reported that serum chemerin levels were elevated in ACC patients as compared to those with benign adrenocortical tumors, and the difference further increased when tested against healthy controls. They also discovered that higher serum chemerin levels were strongly associated with better overall survival. Though seemingly paradoxical, it was proposed that two important, proven factors may explicate this phenomenon. First, a mouse xenograft model showed that increased serum chemerin levels were not a result of secretions from the tumor itself but from the host environment (28). Second, as mentioned previously, chemerin has been shown to be an effective recruiter of immune cells to sites of inflammation (1, 14, 15, 41). Thus, it was suggested that ACC may downregulate chemerin as a method of immune evasion and that the host environment may increase secretion of chemerin in serum as a counteractive response (28).
Interestingly, in addition to the potential for immune-dependent methods of tumor suppression, a new study has found evidence that chemerin may directly alter malignant cells in an immune-independent manner. A study by Liu-Chittenden et al. (42) found that the RARRES2 gene was transcriptionally downregulated in ACC, in line with similar findings in other cancer types. Specifically, the silenced RARRES2 gene in ACC tumors was characterized by hypermethylation at five CpG sites. This was confirmed in three human ACC cell lines and HEK293 cells via treatment with a DNA methyltransferase inhibitor, which showed that such treatment reversed the effects of methylation at all CpG sites in a dose dependent manner (42, 43). [As an aside, in Ewing sarcoma, a rare, malignant tumor that grows inside the bones and in nearby soft tissues, RARRES2 showed a high rate of methylation and was one of only eight genes to have a frequency of silencing >20% (25). Thus, hypermethylation of RARRES2 is likely a common method of gene silencing in tumors where chemerin is downregulated].
When ACC cell lines were transfected to express chemerin, significant inhibition of tumor growth was observed in a dose dependent manner, as the cell lines that had higher expression of chemerin correlated to a more compelling reduction in tumorigenesis (42). Supporting these results, exogenous addition of recombinant chemerin in vitro showed no meaningful effect in affecting cell proliferation among various ACC cell lines, implying that the mechanism of growth inhibition was not mediated by the binding of chemerin to its receptors, such as CMKLR1 (42). In fact, chemerin's function in reducing ACC tumor growth was found to be mediated by two different, immune-independent mechanisms. First, chemerin overexpression could induce β-catenin phosphorylation, and thus, proteasome mediated degradation. In phosphokinase arrays of a chemerin overexpressing ACC cell line, significant reduction in total β-catenin levels was observed, whereas treatment with a proteasome inhibitor prevented proteasome mediated degradation, allowing for detection of elevated phosphorylated β-catenin levels (42). The phosphorylated sites were identified as Ser33, Ser37, and Thr41. Consequently, a decrease in Wnt/β-catenin pathway activity was observed and confirmed via a TCF/LEF luciferase reporter assay (42). Second, chemerin overexpression could inhibit p38 MAPK phosphorylation. The phosphokinase array, which showed a decrease in total β-catenin levels, also showed a reduction in phospho-p38 MAPK levels (42). In vivo xenograft studies in athymic nude mice and NSG mice, both immunodeficient mouse models, confirmed that chemerin overexpression resulted in lower β-catenin and phosphorylated p38 MAPK levels (42). Together, these results indicated that chemerin could promote tumor suppression through immune-independent pathways, both in vitro and in vivo.
These findings may have positive implications for ACC patients and other cancer patients alike. β-catenin (CTNNB1), a proto-oncogene, is frequently mutated in ACC, resulting in constitutive activation of the Wnt/β-catenin pathway (44). The occurrence of activating mutations of CTNNB1 is generally known to be a significant pathway for ACC tumorigenesis. Indeed, a majority of adrenocortical tumors exhibit activation of the Wnt/β-catenin pathway, which is correlated with poor outcomes in ACC patients (45, 46). Aberrant activation of the Wnt/β-catenin pathway is common in many other cancer types, such as breast cancer, lung cancer, hepatocellular carcinoma, and squamous cell carcinoma (47). Additionally, elevated phospho-p38 MAPK levels are found in a majority of adrenocortical tumors (42). In other cancer types such as lung cancer and colon cancer, abnormal activation of p38 signals are detected (48, 49). As such, it is reasonable to suspect that chemerin may have similar, tumor-suppressive effects in other cancer types.
Finally, chemerin may also serve as a prognostic marker for ACC. Among patients, tissue chemerin levels did not correspond to prognosis, though chemerin expression was downregulated in ACC tissue samples (28). However, serum chemerin levels did correspond to prognosis. A clinical survey showed that serum chemerin levels were significantly elevated in ACC patients in comparison to patients with benign tumors and even more so in comparison to healthy controls. Survival analysis of median serum chemerin levels in ACC patients determined that higher serum chemerin levels correlated with longer survival. Moreover, prognosis for patients with recurrent ACC could be stratified according to varying serum chemerin levels, further verifying its value as a marker for ACC prognosis (28).
Breast Cancer
Breast cancer is the most common malignancy in women, accounting for a quarter of all cancer cases in females (50). It is estimated that one in every eight women will develop breast cancer at some point in their lives (51). In this context, chemerin has not been deeply investigated; though, there are some preliminary pieces of evidence showing that chemerin is significantly downregulated in breast adenocarcinomas, one of the most common forms of breast cancer (15). Additionally, studies have detected CMKLR1 in breast tissue (52). Taken together, these facts raise the suspicion that chemerin may exhibit antitumor effects in breast cancer.
However, pro-tumor effects are also possible. Given the increased risk of breast cancer due to post-menopausal obesity and chemerin's known associations with obesity and obesity related parameters, such as blood pressure and BMI, some suspect that chemerin may be correlated to higher risk of breast cancer, albeit indirectly (51, 53). Moreover, it has been experimentally shown that some adipose-derived angiogenic factors may promote breast cancer growth, though this remains to be tested in the context of chemerin (54, 55). It has also been speculated that chemerin may play a role in instigating metastasis through its angiogenic functions (51). For prognosis, it is unclear whether chemerin can be used as a biomarker for breast cancer. A cross sectional study involving 117 breast cancer patients by Akin et al. (51) showed that serum chemerin levels could not be used in staging of breast cancer, as there were no differences in serum chemerin levels of patients with metastatic and non-metastatic cancer.
Colorectal Cancer
Colorectal cancer has the third highest incidence rate and fourth highest mortality rate of all cancers types, with 1.4 million new cases and 700,000 related deaths per year (56). Current treatment methods range from chemotherapy (e.g., single-agent 5-fluorouracil, oxaliplatin, and/or irinotecan) to other regimens consisting of newer, targeted substances, such as anti-vascular endothelial growth factor-A antibodies or anti-epidermal growth factor receptor antibodies (57).
Among the early diagnostic tools that have become available in recent years, serum chemerin levels have been identified as an effective biomarker for colorectal cancer. A study in 2018 compared 32 colorectal cancer patients with sex, BMI, and age matched healthy volunteers and reported that chemerin was significantly upregulated in serum of colorectal cancer patients, increasing with higher tumor stage progression (58). The area under the receiver operating characteristic (ROC) curve for serum chemerin levels was 1, with a sensitivity and specificity of 100% (58). The results clearly indicated that serum chemerin could be an effective biomarker for colorectal cancer and stage progression.
Additionally, a pilot study of 110 colorectal cancer survivors in Korea showed that there was an inverse relationship between serum chemerin levels and colorectal cancer-related quality of life, defined in terms of Functional Assessment of Cancer Therapy (FACT)-General, -Colorectal, and -Fatigue scores (59). Given that chemerin has been shown to have antimicrobial properties, it was proposed that chemerin may play a role in modulating gut microbial activity, which may play a role in the various bowel symptoms affecting colorectal cancer patients (11, 60). Supporting this hypothesis, colorectal cancer patients who have undergone resection surgery have consistently reported altered gut microbial activity, which has been linked to numerous bowel symptoms in these patients (61–63). Interestingly, the gut microbiome is somewhat altered in chemerin KO mice compared to WT controls, with an increase in Desulfovibrionaceae (64). Increases in this pathobiont have been reported as a component of IBD-associated gut dysbiosis (65).
Gastric Cancer
Gastric cancer is the fourth most common cancer and the second most common cause of cancer-related death in the world (66). Treatment of gastric cancer generally consists of a combination of surgery and chemotherapy (66). Though current clinical parameters for diagnosis are incapable of accurately predicting patient outcomes, recent evidence has revealed chemerin's prognostic value in gastric cancer, as well as its functions in tumor growth (67).
A 2014 study comparing 196 gastric cancer patients with 196 age- and sex-matched healthy controls showed that preoperative plasma chemerin levels were significantly elevated in cancer patients (67). Multivariate analysis of clinical parameters currently used for diagnosis and serum chemerin levels showed that serum chemerin could be used as an independent predictor of 5-year mortality, 5-year adverse event, overall survival, and 5-year disease free survival, though further research is needed to adjust for the possibility of postoperative alterations (67).
In terms of functional roles, studies have shown that chemerin exhibits pro-tumor effects in gastric cancer, confirming the notion that chemerin's pro/anti-tumor functions are context dependent. Indeed, elevated serum chemerin levels were observed in gastric cancer patients and were shown to be correlated with increased tumor invasiveness (29). Specifically, chemerin contributed to tumor growth by inducing phosphorylation of p38 and ERK 1/2 MAPKs and upregulating IL-6, MMP-7, and VEGF (29). Chemerin treatment did not alter the in vitro proliferation of two human gastric cancer cell lines, AGS and MKN28. However, chemerin increased the invasiveness of both cell types through Matrigel-coated transwells with only miniscule concentrations of chemerin. In fact, the number of cells invading through the membrane increased fivefold or more for AGS and MKN28 cells exposed to chemerin. Moreover, chemerin increased mRNA expression of VEGF and IL-6 in AGS and MKN28 cells, while also increasing expression of MMP-7 exclusively in MKN28 (29). VEGF, IL-6, and MMP-7 have all been associated with enhanced tumor invasiveness in gastric cancer (68–70), while high expression of VEGF and IL-6 have been shown to stimulate metastasis of malignant cells and indicate poor clinical outcomes in gastric cancer patients, suggesting a potential impact of chemerin in this setting (69–72).
In addition to increased expression of various pro-invasion genes, stimulation of AGS and MKN28 via chemerin resulted in elevated phosphorylation of p38 and ERK1/2 in both cancer lines, suggesting that chemerin could activate MKK3/6-p38 and MEK-ERK MAPK signaling cascades (29). Importantly, these are known mechanisms by which tumor cells modulate growth and invasion in gastric cancer (73–76). Elevation of phosphorylated ERK 1/2 levels have been associated with poor survival in patients, and p38 activation has been found to incite peritoneal dissemination in gastric cancer (75, 76). Given that these effects were abrogated by treatment with inhibitors of MEK-ERK signaling, it was clear that the MEK-ERK MAPK pathway was primarily associated with increased gastric cancer invasiveness (29). Together, the results of the studies indicated that chemerin may engender pro-tumor effects in gastric cancer, while also serving as a marker for patient prognosis.
Glioblastoma
Glioblastoma is the most malignant form of brain tumors affecting adults (77). Recent advances in glioblastoma research have made new treatment methods available to patients, such as tumor-treating fields and immunotherapy. However, the classical approach to treating glioblastoma remains a multipronged effort, using a combination of surgical resection, radiation, and chemotherapy (77). The incidence rate of glioblastoma hovers around 3.19 cases per 100,000 people in the United States and is very uncommon in children (78).
Though chemerin's role in glioblastoma is still unclear, studies have reported an altered chemerin profile in glioblastoma cases (17). First, one study found that chem158 K, a bioactive isoform of chemerin, was elevated in the cerebrospinal fluid of patients with malignant glioblastoma (79). A later study found that chemerin mRNA expression was significantly increased in grade 3 and 4 glioma, equivalent to malignant glioblastoma, compared to epilepsy and grade 2 glioma (17). In contrast, CMKLR1 and CCRL2 mRNA expression was unaltered in those cases. To further test whether chemerin could function as a signaling molecule, in vitro experiments were conducted using U-87 MG cells, a human glioblastoma cell line. Experimental results determined that the addition of chem157S, another bioactive isoform of chemerin, to U-87 MG cells resulted in a transient, dose-dependent increase of intracellular calcium, indicating that chemerin could instigate intracellular signaling in U-87 MG cells (17). Whether these results translate to experiments in vivo remains to be seen.
Hepatocellular Carcinoma
Hepatocellular carcinoma (HCC) is the fifth most prevalent cancer in the world, with more than 500,000 new cases being diagnosed each year (80). It is mainly caused by chronic liver damage due to hepatitis B or hepatitis C and is associated with risk factors such as obesity and diabetes (80). Currently, a variety of treatment options are available to patients, including liver transplantation, curative resection, radiotherapy, radio/chemo-embolization, and systemic therapies (81).
In recent years, chemerin has been identified as a potential therapeutic agent for HCC. A study comparing chemerin expression levels in 124 HCC patient tumors and matched, normal tissues showed that chemerin was significantly downregulated in 72 of the patients' liver tissues and that tissue chemerin expression correlated with tumor size, grade, and infiltration of dendritic cells and natural killer cells. Additionally, chemerin was identified as an independent factor for prognosis via multivariate analysis, with lower chemerin expression corresponding to poorer overall survival (23).
A later study by Lin et al. (82) further reported that chemerin could inhibit HCC tumor growth. In an orthotopic mouse model of HCC, chemerin knockout mice showed aggressive tumor growth and metastasis. In contrast, overexpression of chemerin in mice resulted in delayed tumor growth, suggesting that chemerin may hamper tumor progression. Specifically, the inhibitory effect was mediated by suppressing pro-tumor inflammatory cytokines. When Hepa1-6 cells were treated with chemerin in vitro, cell survival and proliferation was unaffected. Moreover, chemerin knockout mice with accelerated tumor growth exhibited increased expression of granulocyte-macrophage colony-stimulating factor (GM-CSF) and IL-6 and accumulation of myeloid-derived suppressor cells (MDSCs) and tumor-associated macrophages (TAMs). Neutralization of GM-CSF and IL-6 reversed the effects in the chemerin knockout mice, showing that these two cytokines were mainly responsible for the accelerated growth. Combined with the fact that serum chemerin levels were inversely correlated to GM-CSF and IL-6 expression levels in patients with HCC, the study suggested that chemerin may have a negative regulatory role for these two cytokines (82).
Further verifying these claims, chemerin was shown to inhibit nuclear factor-κB activation, an important factor for GM-CSF and IL-6 expression (83, 84). The data consistently showed that chemerin targeted tumor cells and tumor-associated endothelial cells, the major source of GM-CSF and IL-6, via interaction with CMKLR1 and CCRL2, reducing expression of inflammatory cytokines and inhibiting NF-κB activation (82). Downregulation of CMKLR1 and CCRL2 via siRNA in Hepa1-6 cells reestablished GM-CSF and IL-6 expression, indicating that decreased expression of the cytokines was mediated by chemerin-CMKLR1/CCRL2 interactions. The observed consequences of reduced GM-CSF and IL-6 expression were impaired MDSC accumulation, reestablishment of anti-tumor IFN-γ+ T-cell activity, and hampered tumor angiogenesis (82).
The inhibitory function of chemerin in HCC was mediated by both T-cell dependent and independent mechanisms. Because chemerin showed less of an inhibitory effect in Rag1 knockout and CD8-T-cell depleted mice, it was determined that chemerin's function in HCC was not completely based on T-cell anti-tumor activity (82). It was postulated that chemerin's T-cell independent mechanism of inhibition was mediated by reduced accumulation of MDSCs, which are a major source of pro-angiogenic factors Bv8 and MMP-9. Supporting this hypothesis, overexpression of chemerin in mice resulted in a notable decrease in Bv8 and MMP-9 expression and, ultimately, tumor angiogenesis (82).
A recently published study showed a novel interaction of chemerin and the tumor suppressor PTEN in a mouse model of HCC (30). In vitro, overexpression of chemerin resulted in reduced migratory ability and invasiveness of human HCC line 7404. Chemerin knockdown, in turn, resulted in increased migratory ability and invasiveness. In vivo, overexpression of chemerin inhibited intra- and extrahepatic metastases of HCC cells in nude mice, lengthening survival times of HCC inoculated mice. Nude mice with chemerin overexpressing tumors showed markedly less distant metastases and delayed appearance of metastatic foci using control or chemerin-expressing PVTT-1 HCC cells. Consistent observations were made in an intrahepatic injection model. Moreover, regular intraperitoneal injections or intrahepatical injections of chemerin in mice with PVTT-1 tumors replicated these results, with the additional benefit of reduced weight loss, showing that chemerin could potentially be therapeutically administered to suppress HCC metastases.
Underlying these tumor suppressive effects, the mechanism of reduced HCC cell migration and invasion via chemerin was shown to be mediated by negative regulation of p-Akt (Ser473) by PTEN. P-Akt and MMP-1 levels, as well as ubiquitination of PTEN, were reduced in chemerin-overexpressing HCC cell lines and elevated in chemerin-knockdown HCC cell lines. PTEN levels were upregulated in chemerin-overexpressing HCC cell lines and downregulated in chemerin-knockdown HCC cell lines. Immunohistochemistry of HCC tissues showed that upregulation of chemerin correlated with low p-Akt and high PTEN levels. Supporting these findings in vivo, data from their tumor models showed that overexpression of chemerin was associated with reduced tumor MMP-1 and p-Akt levels, elevated PTEN levels, and reduced metastases. Taken in aggregate, these studies provided strong support for therapeutic applications of chemerin in HCC.
Melanoma
Melanoma has one of the fastest growing incidence rates in the world, which in the US has risen from 8.2–9.4 cases per 100,000 people in 1975 to 24.2–35.4 cases per 100,000 people in 2010 (85). Unlike many malignancies, melanoma affects a diverse range of age groups, with a median age of diagnosis of 57 years (86). It is also widely recognized as the most aggressive form of skin cancer, accounting for the majority of skin cancer-related deaths vs. a fraction of total cases (86). Significantly, recent research has elucidated chemerin's ability to induce tumor suppression in melanoma, where chemerin was found to be downregulated (15).
Our group previously identified the mechanism of tumor suppression via chemerin in melanoma, showing that tumor expression of chemerin led to inhibited tumor growth in vivo. When chemerin-expressing B16F0 (hereafter, B16) cells were implanted into mice, the mouse melanoma showed significant delay in growth compared to control B16 cell lines. Importantly, the RARRES2-transfected B16 cells expressed bioactive chemerin, confirmed via chemotaxis, and both the transfected line and control B16 lines lacked expression of CMKLR1. When tested in vitro, chemerin treatment failed to alter B16 cell proliferation, suggesting that chemerin's inhibitory effects in melanoma was mediated by host immune responses (15). It was also noted that chemerin expressing tumors exhibited enhanced infiltration by NK and T cells; ratios of NK and T cells to MDSCs and/or plasmacytoid dendritic cells (pDC) were also increased (15, 87, 88). Given that certain subsets of pDCs and MDSCs have been reported to be tolerogenic, suppressing the body's antitumor immune responses, it was noted that increased ratios of antitumor immune cells to tolerogenic ones would translate to a more favorable environment for tumor suppression (89, 90). Moreover, chemerin's tumor suppressive effects in melanoma were mediated by NK cells, as their depletion abrogated the antitumor effects, while a lack of T cells and B cells did not alter the tumor growth-inhibition phenotype (15). Further experiments showed that host expression of CMKLR1 was necessary for inhibited tumor growth, since chemerin-expressing mouse melanoma grew faster than control B16 tumors in CMKLR1-negative mice. Finally, local administration of chemerin suppressed tumor growth in vivo (15).
Thus, in melanoma, chemerin was shown to inhibit tumor growth by eliciting antitumor responses and altering the tumor microenvironment in favor of growth inhibition. Whether by tumoral expression or local administration, these tumor-suppressive effects could be observed in vivo (15). Additionally, high expression of chemerin was shown to be associated with better outcomes for patients in two clinical studies, demonstrating chemerin's potential for therapeutic intervention in melanoma (15).
Mesothelioma
Mesothelioma is a tumor that grows in the linings of various organs, such as the lungs or the heart. Tumorigenesis is instigated by exposure to specific, carcinogenic mineral fibers, namely asbestos (63). Due to the commercial use of such materials, the incidence of mesothelioma has increased over the years, from near non-existence to several thousand cases per year (63). Though treatments are available, such as chemotherapy with cisplatin and pemetrexed, prognosis is still dismal and diagnosis very difficult, as it may take several decades for symptoms to appear (91). Regarding chemerin's role in mesothelioma, literature on the topic is scarce, though it was one of the first identified cancer types where RARRES2 expression was altered (significantly increased, compared to matched normal tissue) (16). It remains unclear if chemerin contributes in any way to mesothelioma progression.
Neuroblastoma
Neuroblastoma is a pediatric cancer affecting the sympathetic nervous system. With < 50% probability of cure for high-risk cases, the prognosis for children with advanced stage neuroblastoma is bleak (92). For those with high-risk neuroblastoma, treatment is intensive, consisting of several modalities, such as chemotherapy, surgery, immunotherapy, stem cell rescue, and differentiation therapy (93).
A recent study by Tümmler et al. (21) showed that expression of chemerin receptors could successfully prognosticate neuroblastoma. Based on data from public datasets, the study found that high expression of GPR1 and CMKLR1 was associated with low survival rates. Furthermore, expression of CMKLR1 and CCRL2 was found to be upregulated in neuroblastoma cohorts in comparison to benign counterparts, while increased CCRL2 expression was correlated to poor prognosis. Increased chemerin expression was also found in neuroblastoma cohorts in comparison to neural crest, though no differences in expression levels were found between neuroblastoma and benign neurofibroma. Overall, CMKLR1 and GPR1 expression was shown to be a potentially viable indicator of prognosis in neuroblastoma patients (21).
According to the same study, inhibition of the chemerin/CMKLR1 axis exhibited antitumor effects in vitro and in vivo, suggesting that the chemerin/CMKLR1 axis could be a potential therapeutic target for neuroblastoma. In vitro, expression of CMKLR1, chemerin mRNA/protein, and GPR1 was verified in neuroblastoma cell lines (as well as primary tumors) via real time-PCR and western blot analysis (21). Exposure to inflammatory cytokines TNF-α, IL-1β, and serum resulted in increased secretion of chemerin by neuroblastoma cells. Moreover, the mechanistic effects of added chemerin in neuroblastoma cell lines was the induction of intracellular calcium, activation of MAPK and Akt signaling, and synthesis of MMP-2. Specifically, in a human neuroblastoma cell line (SK-N-SH), chemerin induced a rapid increase in intracellular calcium. In SK-N-AS and SK-N-BE(2) cells, a dose-dependent increase in MEK 1/2, Akt, and ERK 1/2 phosphorylation was observed upon addition of chemerin, indicating the activation of Akt and MAPK pathways. In the same cell lines, a dose-dependent increase in MMP-2 synthesis was shown via real-time zymography, after cells had been stimulated via chemerin for various timepoints. Significantly, inhibition of CMKLR1 on four neuroblastoma cell lines via α-NETA, a recently described CMKLR1 inhibitor (94), dose-dependently reduced cell viability and clonogenicity (21). Similar results were achieved in vivo. In a SK-N-AS xenograft model, mice that were pretreated with α-NETA, before tumors reached a specified volume, showed longer survival and delayed tumor growth compared to control mice (21). Thus, the results of both in vitro and in vivo experiments indicated that targeting chemerin/CMKLR1 could potentially elicit antitumor effects in clinical settings.
Non-small Cell Lung Cancer
Lung cancer is the leading cause of cancer-related deaths in the world, having a 5-year survival rate of < 15% and resulting in ~1.4 million deaths per year (66, 95). This is despite the fact that various chemotherapy-based treatment methods are available to patients. For non-small cell lung cancer (NSCLC), which accounts for the majority (~85%) of lung cancer cases, multiple studies indicate that chemerin may be of great diagnostic and prognostic value (24, 27, 96). Two independent studies, one in 2011 and another in 2016, reported that chemerin was downregulated in NSCLC (24, 27). Zhao et al. (24) compared 108 NSCLC tumor samples with corresponding disease-free tissues and found that 56 of the NSCLC patient tumors showed a lower chemerin expression profile. They also found a positive correlation between chemerin expression levels and infiltration of NK cells into tumor, as revealed by CD56 IHC staining of NSCLC patient tissue samples. Moreover, multivariate analysis of various parameters, such as age, smoking history, tumor size, and differentiation grade, and univariate analysis, together, revealed that lower levels of chemerin expression in tissue were significantly associated with tumor-node metastasis stage, degree of differentiation, and poorer survival rates (24). In line with these findings, a 2016 study by Cai et al. (27) comparing 20 NSCLC tumor samples with their corresponding non-tumor tissues found that 16 of the 20 tumor samples exhibited a significant downregulation of chemerin. Chemerin mRNA expression was also tested in 26 NSCLC tumor samples along with their healthy tissue counterparts, and the RT-qPCR results of these specimen showed that chemerin expression was significantly reduced in most (19) of the samples. Furthermore, chi-squared analysis of chemerin expression levels in NSCLC specimen, distinguished between low and high based on a mean score, found that tissue chemerin expression levels were associated with tumor-node metastasis stage, differentiation, and lymph node metastasis (24, 27). Both studies identified tissue chemerin levels as an independent prognostic factor for NSCLC and reported that, overall, higher chemerin expression was associated with positive prognosis (24, 27).
Unlike tissue expression levels, elevated serum chemerin levels were observed in NSCLC patients, which correlated to poorer overall survival, and was identified as an independent risk factor for poorer prognosis in NSCLC patients (97). In a study by Xu et al. (97), analysis of serum samples from 189 NSCLC patients and 120 healthy individuals revealed that serum chemerin levels were upregulated in NSCLC patients. Furthermore, both early stage and advanced stage NSCLC patients showed elevated levels of chemerin in serum, with a further increase for patients with advanced stage NSCLC. For diagnosis of NSCLC, serum chemerin levels had a test sensitivity and specificity of 62.4% and 67.5%, respectively. When combined with carcinoembryonic antigen tests, the sensitivity and specificity of the test increased to 78.3 and 84.2%, respectively. Thus, serum chemerin expression was also established as a viable biomarker for diagnosis and prognosis of NSCLC (97).
To elucidate chemerin's mechanistic roles in lung cancer, Unver et al. (31) examined chemerin's functions in Lewis lung carcinoma (LLC), a mouse model of lung cancer. LLC cells were genetically altered to secrete prochemerin at low levels insufficient to induce chemotaxis. Media from the modified LLC cells was used to culture splenic leukocytes and suppress their expression of inflammatory cytokines, TNF-α, and IL-12 p40 (31). Results were replicated in vivo, as expression of TNF-α and IL-12 p40 was also reduced in the prochemerin expressing LLC tumor tissues. These observations are notable because inflammation, though sometimes necessary for antitumor activity, is often exploited by tumors to establish a pro-tumor environment, particularly in cases of NSCLC (98, 99). In syngeneic C57BL6 mice implanted with prochemerin-expressing LLC grafts, tumor formation was impeded by prochemerin expression (31). However, in mice with successful tumor formation, control tumors and prochemerin expressing tumors showed no difference in growth rate, indicating that prochemerin expression may play a role in modulating tumorigenesis through reduced inflammation but not in tumor suppression of LLC (31).
Prostate Cancer
Prostate cancer is one of the most frequently diagnosed cancers in the United States, with more than 164,000 new cases per year (100). It is also the second leading cause of cancer-related mortality in the country (101). Treatment of prostate cancer mainly consists of surgery and radiotherapy for localized cases and androgen deprivation therapy and chemotherapy for metastatic cases (102).
Studies have shown that chemerin is significantly downregulated in prostate cancer. Both analyses of public datasets and RT-qPCR of human prostate tumor samples in several studies demonstrated reduced expression of chemerin, indicating that prostate tumors may downregulate chemerin as a means of immune evasion (15, 26, 103, 104). Thus, it is possible that therapeutic application of chemerin in prostate cancer may enhance anti-tumor immunity and slow tumor progression.
Squamous Cell Carcinoma of the Esophagus
Esophageal cancer affects millions of people worldwide, with approximately half a million new cases being diagnosed each year (105). Of those cases, esophageal squamous cell carcinoma (ESCC) accounts for a dominant majority, 90% (105). The outlook for patients is bleak, as esophageal cancers are among the deadliest tumors in the world due to its fast progression and late diagnosis (106). Standard therapy for esophageal cancer patients consists of esophagectomy, radiotherapy, and/or chemotherapy (106).
Identifying chemerin's role in ESCC, kumar et al. (20) showed that chemerin was upregulated in ESCC myofibroblasts, which also expressed CMKLR1 (20). Moreover, chemerin was shown to increase invasiveness of ESCC in vitro, which was mediated by the accumulation of MMP-1, MMP-2, and MMP-3. In Boyden chambers, chemerin stimulated the migration and invasion of OE21 cells, a human ESCC cell line, and this effect was inhibited by antagonists of CMKLR1 and chemerin siRNA, supporting previous results. Analysis of the OE21 media showed that there was increased expression of MMP-1, MMP-2, and MMP-3, and this effect could be reduced by inhibiting p44/42 MAPK kinase and protein kinase C, showing that they were responsible for increased MMP expression levels (20). Combined with previous reports that MMP-1 expression is associated with poor prognosis in esophageal cancer, the study suggested that chemerin could elicit pro-tumor effects in ESCC (107–109). Thus, it was postulated that targeting chemerin/CMKLR1 may be a viable therapeutic approach for ESCC.
Squamous Cell Carcinoma of the Oral Tongue
Squamous cell carcinoma of the oral tongue (SCCOT) is the most common form of oral squamous cell carcinoma (OSCC), which has a worldwide incidence rate of ~6.6 cases per 100,000 males and 2.9 cases per 100,000 females (110). Current treatment for SCCOT ranges from surgery alone, for stage 1 and 2 cases, to a combination of surgery and radiotherapy for later stages of disease (111). Though early diagnosis and treatment methods are currently available, the primary causes of mortality in SCCOT patients are lymph node metastasis and recurrence (112). Thus, biomarkers for those parameters may be of great prognostic value for SCCOT patients.
A study by Wang et al. (18), looking at chemerin mRNA expression in 19 SCCOT tumor tissues and matched adjacent tissues via qRT-PCR and IHC stains of 147 SCCOT specimen and their associated peritumoral, healthy tissues, determined that both chemerin mRNA and protein was upregulated in primary SCCOT specimens (18). Significantly, overexpression of chemerin was correlated with poor differentiation, high clinical stage, and lymph node metastasis. Multivariate survival analysis further showed that chemerin was an independent prognostic factor for SCCOT and that patients with overexpressed chemerin had a shorter cancer-related survival time (18). Though chemerin's role in SCCOT tumor progression is still unclear, the results indicate that chemerin may be a therapeutic target for inhibiting tumor growth.
Another study found that serum and salivary levels of chemerin were also elevated in OSCC patients, indicating that chemerin may be used to diagnose patients (113). The study included serum and salivary samples from 15 patients with early stage OSCC, 15 patients with oral premalignant lesions (OPML), and 15 healthy individuals, and analysis of these specimen showed that serum and salivary levels of chemerin, and MMP-9, were upregulated in OSCC compared to OPML, and in OPML compared to healthy patients. Analysis of the ROC curve revealed that serum and salivary chemerin levels had a test sensitivity and specificity of 100% for detecting early stage OSCC (113).
Squamous Cell Carcinoma of the Skin
Cutaneous Squamous Cell Carcinoma (CSCC) is the second most prevalent form of skin cancer in the world (114). Affecting the elderly at disproportionately higher rates, CSCC is mainly caused by chronic exposure to ultraviolet radiation (114, 115). Fortunately, advances in treatment have made new, targeted molecular therapies available to patients, in addition to the standard options of surgery and chemotherapy (116).
A recent study indicated that chemerin may contribute to CSCC cell migration and tumor growth. The study by Farsam et al. (19) found that chemerin was downregulated in CSCC but upregulated in senescent fibroblasts and skin samples of elderly patients (19, 117). Transcriptional analysis of CCRL2 in CSCC cell lines further revealed that CCRL2 expression was significantly increased on CSCC tumor cells in vitro, and enhanced tumor cell migration was observed as a result of increased levels of senescence-associated chemerin, which was abrogated by inhibition of chemerin in senescent fibroblasts. Moreover, CMKLR1, or the combination of CCRL2 and GPR1, was necessary for CSCC cell migration via chemerin, given that suppression of either one of the receptors reversed these effects. Finally, chemerin was shown to activate MAPK signaling in SCL-1 cells, a human CSCC cell line, with high expression of CCRL2 and low expression of GPR1. Specifically, the JNK and ERK 1/2 pathways were indicated as the primary mediators of chemerin-mediated effects in CSCC tumor cells, as inhibition of these pathways neutralized the previously observed migratory response. Taken together, these results showed that chemerin enhanced CSCC cell migration and promoted tumor growth through chemerin/CMKLR1/CCRL2/GPR1 interactions and subsequent activation of MAPK pathway subtypes (19).
Conclusion
Chemerin is a versatile protein with significant functions in modulating tumor growth. With altered expression profiles in a variety of cancer types, it can have context dependent effects on tumorigenesis and tumor progression. In some cases, cancers may silence RARRES2 via hypermethylation to evade immune surveillance. In others, host systems may increase chemerin expression as a defensive measure, recruiting antitumor immunocytes, reducing secretion of inflammatory cytokines, and/or modifying signaling pathways. Through both immune-independent and immune-dependent mechanisms, chemerin has been shown to elicit tumor-suppressive effects in various cancers, and there is a strong possibility that the mechanisms of tumor inhibition seen in these tumor types may be replicated in others as well. Importantly, for cases in which silencing of RARRES2 has been reported, the restoration and/or forced overexpression of chemerin in the microtumor environment may incite compelling antitumor effects, indicating new avenues of research for chemerin in cancer.
Here, we have comprehensively reviewed the current data on chemerin's functions in cancer, along with its underlying mechanisms when available. Concentrating on aspects that may lead to clinical applications, chemerin's diagnostic and/or prognostic value have also been evaluated in a few cancer types. Given the efficacy of chemerin-mediated anti-tumor responses seen now in several tumor settings, future research investigating chemerin's roles in additional tumor types is warranted, with a particular focus on manifesting its therapeutic potential for cancer treatment. Importantly, it should be noted that chemerin-based treatments may add to the efficacy of approved checkpoint inhibitors, as they likely act through independent mechanisms to diminish tumor growth.
Author Contributions
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank Christina Mizerny and Kayla Corman for administrative support. RP is funded by American Cancer Society #125078-MRSG-13-244-01-LIB and a Kimmel Scholar Award. BZ was supported by 3R01AI079320, R01CA169354 (E.C. Butcher, PI), and the Emerson Collective Cancer Research Fund.
References
1. Zabel BA, Allen SJ, Kulig P, Allen JA, Cichy J, Handel TM, et al. Chemerin activation by serine proteases of the coagulation, fibrinolytic, and inflammatory cascades. J Biol Chem. (2005) 280:34661–6. doi: 10.1074/jbc.M504868200
2. Wittamer V, Franssen JD, Vulcano M, Mirjolet JF, Le Poul E, Migeotte I, et al. Specific recruitment of antigen-presenting cells by chemerin, a novel processed ligand from human inflammatory fluids. J Exp Med. (2003) 198:977–85. doi: 10.1084/jem.20030382
3. Yoshimura T, Oppenheim JJ. Chemokine-like receptor 1 (CMKLR1) and chemokine (C-C motif) receptor-like 2 (CCRL2); two multifunctional receptors with unusual properties. Exp Cell Res. (2011) 317:674–84. doi: 10.1016/j.yexcr.2010.10.023
4. Zabel BA, Nakae S, Zuniga L, Kim JY, Ohyama T, Alt C, et al. Mast cell-expressed orphan receptor CCRL2 binds chemerin and is required for optimal induction of IgE-mediated passive cutaneous anaphylaxis. J Exp Med. (2008) 205:2207–20. doi: 10.1084/jem.20080300
5. Bondue B, Wittamer V, Parmentier M. Chemerin and its receptors in leukocyte trafficking, inflammation and metabolism. Cytokine Growth Factor Rev. (2011) 22:331–8. doi: 10.1016/j.cytogfr.2011.11.004
6. Yang YL, Ren LR, Sun LF, Huang C, Xiao TX, Wang BB, et al. The role of GPR1 signaling in mice corpus luteum. J Endocrinol. (2016) 230:55–65. doi: 10.1530/JOE-15-0521
7. Li L, Ping M, Huang C, Liu Y, Zhang Y, Gao C, et al. Expression of chemerin and its receptors in rat testes and its action on testosterone secretion. J Endocrinol. (2014) 220:155–63. doi: 10.1530/JOE-13-0275
8. Rourke JL, Muruganandan S, Dranse HJ, McMullen NM, Sinal CJ. Gpr1 is an active chemerin receptor influencing glucose homeostasis in obese mice. J Endocrinol. (2014) 222:201–15. doi: 10.1530/JOE-14-0069
9. Nagpal S, Patel S, Jacobe H, DiSepio D, Ghosn C, Malhotra M, et al. Tazarotene-induced gene 2 (TIG2), a novel retinoid-responsive gene in skin. J Invest Dermatol. (1997) 109:91–5.
10. Kaur J, Adya R, Tan BK, Chen J, Randeva HS. Identification of chemerin receptor (ChemR23) in human endothelial cells: chemerin-induced endothelial angiogenesis. Biochem Biophys Res Commun. (2010) 391:1762–8. doi: 10.1016/j.bbrc.2009.12.150
11. Banas M, Zabieglo K, Kasetty G, Kapinska-Mrowiecka M, Borowczyk J, Drukala J, et al. Chemerin is an antimicrobial agent in human epidermis. PLoS ONE (2013) 8:e58709. doi: 10.1371/journal.pone.0058709
12. Ernst MC, Issa M, Goralski KB, Sinal CJ. Chemerin exacerbates glucose intolerance in mouse models of obesity and diabetes. Endocrinology (2010) 151:1998–2007. doi: 10.1210/en.2009-1098
13. Ernst MC, Sinal CJ. Chemerin: at the crossroads of inflammation and obesity. Trends Endocrinol Metab. (2010) 21:660–7. doi: 10.1016/j.tem.2010.08.001
14. Parolini S, Santoro A, Marcenaro E, Luini W, Massardi L, Facchetti F, et al. The role of chemerin in the colocalization of NK and dendritic cell subsets into inflamed tissues. Blood (2007) 109:3625–32. doi: 10.1182/blood-2006-08-038844
15. Pachynski RK, Zabel BA, Kohrt HE, Tejeda NM, Monnier J, Swanson CD, et al. The chemoattractant chemerin suppresses melanoma by recruiting natural killer cell antitumor defenses. J Exp Med. (2012) 209:1427–35. doi: 10.1084/jem.20112124
16. Mohr S, Bottin MC, Lannes B, Neuville A, Bellocq JP, Keith G, et al. Microdissection, mRNA amplification and microarray: a study of pleural mesothelial and malignant mesothelioma cells. Biochimie (2004) 86:13–9. doi: 10.1016/j.biochi.2003.11.008
17. Yamaguchi Y, Du XY, Zhao L, Morser J, Leung LL. Proteolytic cleavage of chemerin protein is necessary for activation to the active form, Chem157S, which functions as a signaling molecule in glioblastoma. J Biol Chem. (2011) 286:39510–9. doi: 10.1074/jbc.M111.258921
18. Wang N, Wang QJ, Feng YY, Shang W, Cai M. Overexpression of chemerin was associated with tumor angiogenesis and poor clinical outcome in squamous cell carcinoma of the oral tongue. Clin Oral Investig. (2014) 18:997–1004. doi: 10.1007/s00784-013-1046-8
19. Farsam V, Basu A, Gatzka M, Treiber N, Schneider LA, Mulaw MA, et al. Senescent fibroblast-derived Chemerin promotes squamous cell carcinoma migration. Oncotarget (2016) 7:83554–69. doi: 10.18632/oncotarget.13446
20. Kumar JD, Kandola S, Tiszlavicz L, Reisz Z, Dockray GJ, Varro A. The role of chemerin and ChemR23 in stimulating the invasion of squamous oesophageal cancer cells. Br J Cancer (2016) 114:1152–9. doi: 10.1038/bjc.2016.93
21. Tümmler C, Snapkov I, Wickström M, Moens U, Ljungblad L, Maria Elfman LH, et al. Inhibition of chemerin/CMKLR1 axis in neuroblastoma cells reduces clonogenicity and cell viability in vitro and impairs tumor growth in vivo. Oncotarget (2017) 8:95135–51. doi: 10.18632/oncotarget.19619
22. Zhang J, Zhou J, Tang X, Zhou LY, Zhai LL, Vanessa ME, et al. Reduced expression of chemerin is associated with poor clinical outcome in acute myeloid leukemia. Oncotarget (2017) 8:92536–44. doi: 10.18632/oncotarget.21440
23. Lin W, Chen YL, Jiang L, Chen JK. Reduced expression of chemerin is associated with a poor prognosis and a lowed infiltration of both dendritic cells and natural killer cells in human hepatocellular carcinoma. Clin Lab. (2011) 57:879–85.
24. Zhao S, Li C, Ye Y-B, Peng F, Chen Q. Expression of chemerin correlates with a favorable prognosis in patients with non-small cell lung cancer. Lab Med. (2011) 42:553–7. doi: 10.1309/LMWW79NITS6ZADPT
25. Alholle A, Brini AT, Gharanei S, Vaiyapuri S, Arrigoni E, Dallol A, et al. Functional epigenetic approach identifies frequently methylated genes in Ewing sarcoma. Epigenetics (2013) 8:1198–204. doi: 10.4161/epi.26266
26. Pachynski RK, Zabel B, Leong W.-I, Crowder R, Peehl D, Butcher E. Expression of the leukocyte chemoattractant chemerin in human prostate tumors. J Clin Oncol. (2015) 33(7_suppl.):81. doi: 10.1200/jco.2015.33.7_suppl.81
27. Cai Q, Huang Z, Qi L, Wang T, Shen Y, Huang J. Tazarotene-induced gene 2 is associated with poor survival in non-small cell lung cancer. Oncol Lett. (2016) 12:2680–5. doi: 10.3892/ol.2016.5025
28. Liu-Chittenden Y, Patel D, Gaskins K, Giordano TJ, Assie G, Bertherat J, et al. Serum RARRES2 is a prognostic marker in patients with Adrenocortical Carcinoma. J Clin Endocrinol Metab. (2016) 101:3345–52. doi: 10.1210/jc.2016-1781
29. Wang C, Wu WK, Liu X, To KF, Chen GG, Yu J, et al. Increased serum chemerin level promotes cellular invasiveness in gastric cancer: a clinical and experimental study. Peptides (2014) 51:131–8. doi: 10.1016/j.peptides.2013.10.009
30. Li JJ, Yin HK, Guan DX, Zhao JS, Feng YX, Deng YZ, et al. Chemerin suppresses hepatocellular carcinoma metastasis through CMKLR1-PTEN-Akt axis. Br J Cancer (2018) 118:1337–48. doi: 10.1038/s41416-018-0077-y
31. Unver N, Yilmaz G, Canpinar H, Guc D, Esendagli G. Regulatory role of chemerin in lung tumorigenesis and cancer-associated inflammation. Acta Medica (2018) 49:16–25.
32. Yamamoto JF, Goodman MT. Patterns of leukemia incidence in the United States by subtype and demographic characteristics, 1997–2002. Cancer Causes Control. (2008) 19:379–90. doi: 10.1007/s10552-007-9097-2
33. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2015. CA Cancer J Clin. (2015) 65:5-29. doi: 10.3322/caac.21254
34. De Kouchkovsky I, Abdul-Hay M. Acute myeloid leukemia: a comprehensive review and 2016 update. Blood Cancer J. (2016) 6:e441. doi: 10.1038/bcj.2016.50
35. Shah A, Andersson TM, Rachet B, Bjorkholm M, Lambert PC. Survival and cure of acute myeloid leukaemia in England, 1971-2006: a population-based study. Br J Haematol. (2013) 162:509–16. doi: 10.1111/bjh.12425
36. Meyers J, Yu Y, Kaye JA, Davis KL. Medicare fee-for-service enrollees with primary acute myeloid leukemia: an analysis of treatment patterns, survival, and healthcare resource utilization and costs. Appl Health Econ Health Policy (2013) 11:275–86. doi: 10.1007/s40258-013-0032-2
37. Maiga A, Lemieux S, Pabst C, Lavallee VP, Bouvier M, Sauvageau G, et al. Transcriptome analysis of G protein-coupled receptors in distinct genetic subgroups of acute myeloid leukemia: identification of potential disease-specific targets. Blood Cancer J. (2016) 6:e431. doi: 10.1038/bcj.2016.36
38. Kebebew E, Reiff E, Duh QY, Clark OH, McMillan A. Extent of disease at presentation and outcome for adrenocortical carcinoma: have we made progress? World J Surg. (2006) 30:872–8. doi: 10.1007/s00268-005-0329-x
39. Bilimoria KY, Shen WT, Elaraj D, Bentrem DJ, Winchester DJ, Kebebew E, et al. Adrenocortical carcinoma in the United States: treatment utilization and prognostic factors. Cancer (2008) 113:3130–6. doi: 10.1002/cncr.23886
40. Kerkhofs TM, Verhoeven RH, Van der Zwan JM, Dieleman J, Kerstens MN, Links TP, et al. Adrenocortical carcinoma: a population-based study on incidence and survival in the Netherlands since 1993. Eur J Cancer (2013) 49:2579–86. doi: 10.1016/j.ejca.2013.02.034
41. Vermi W, Riboldi E, Wittamer V, Gentili F, Luini W, Marrelli S, et al. Role of ChemR23 in directing the migration of myeloid and plasmacytoid dendritic cells to lymphoid organs and inflamed skin. J Exp Med. (2005) 201:509–15. doi: 10.1084/jem.20041310
42. Liu-Chittenden Y, Jain M, Gaskins K, Wang S, Merino MJ, Kotian S, et al. RARRES2 functions as a tumor suppressor by promoting beta-catenin phosphorylation/degradation and inhibiting p38 phosphorylation in adrenocortical carcinoma. Oncogene (2017) 36:3541–52. doi: 10.1038/onc.2016.497
43. Rechache NS, Wang Y, Stevenson HS, Killian JK, Edelman DC, Merino M, et al. DNA methylation profiling identifies global methylation differences and markers of adrenocortical tumors. J Clin Endocrinol Metab. (2012) 97:e1004–1013. doi: 10.1210/jc.2011-3298
44. Assie G, Letouze E, Fassnacht M, Jouinot A, Luscap W, Barreau O, et al. Integrated genomic characterization of adrenocortical carcinoma. Nat Genet. (2014) 46:607–12. doi: 10.1038/ng.2953
45. Tissier F, Cavard C, Groussin L, Perlemoine K, Fumey G, Hagnere AM, et al. Mutations of beta-catenin in adrenocortical tumors: activation of the Wnt signaling pathway is a frequent event in both benign and malignant adrenocortical tumors. Cancer Res. (2005) 65:7622–7. doi: 10.1158/0008-5472.CAN-05-0593
46. Gaujoux S, Grabar S, Fassnacht M, Ragazzon B, Launay P, Libe R, et al. beta-catenin activation is associated with specific clinical and pathologic characteristics and a poor outcome in adrenocortical carcinoma. Clin Cancer Res. (2011) 17:328–36. doi: 10.1158/1078-0432.CCR-10-2006
47. Polakis P. Wnt signaling in cancer. Cold Spring Harb Perspect Biol. (2012) 4:a008052. doi: 10.1101/cshperspect.a008052
48. Wagner EF, Nebreda AR. Signal integration by JNK and p38 MAPK pathways in cancer development. Nat Rev Cancer (2009) 9:537–49. doi: 10.1038/nrc2694
49. Chiacchiera F, Grossi V, Cappellari M, Peserico A, Simonatto M, Germani A, et al. Blocking p38/ERK crosstalk affects colorectal cancer growth by inducing apoptosis in vitro and in preclinical mouse models. Cancer Lett. (2012) 324:98–108. doi: 10.1016/j.canlet.2012.05.006
50. Ghoncheh M, Pournamdar Z, Salehiniya H. Incidence and mortality and epidemiology of breast cancer in the World. Asian Pac J Cancer Prev. (2016) 17:43–6. doi: 10.7314/APJCP.2016.17.S3.43
51. Akin S, Akin S, Gedik E, Haznedaroglu E, Dogan AL, Altundag MK. Serum chemerin level in breast cancer. Int J Hematol Oncol. (2017) 27:127–32. doi: 10.4999/uhod.171847
52. Cash JL, Hart R, Russ A, Dixon JP, Colledge WH, Doran J, et al. Synthetic chemerin-derived peptides suppress inflammation through ChemR23. J Exp Med. (2008) 205:767–75. doi: 10.1084/jem.20071601
53. Bozaoglu K, Bolton K, McMillan J, Zimmet P, Jowett J, Collier G, et al. Chemerin is a novel adipokine associated with obesity and metabolic syndrome. Endocrinology (2007) 148:4687–94. doi: 10.1210/en.2007-0175
54. Lorincz AM, Sukumar S. Molecular links between obesity and breast cancer. Endocr Relat Cancer (2006) 13:279–92. doi: 10.1677/erc.1.00729
55. Perrier S, Caldefie-Chezet F, Vasson MP. IL-1 family in breast cancer: potential interplay with leptin and other adipocytokines. FEBS Lett. (2009) 583:259–65. doi: 10.1016/j.febslet.2008.12.030
56. Ferlay J, Soerjomataram I, Dikshit R, Eser S, Mathers C, Rebelo M, et al. Cancer incidence and mortality worldwide: sources, methods and major patterns in GLOBOCAN 2012. Int J Cancer (2015) 136:e359–386. doi: 10.1002/ijc.29210
57. Stintzing S. Management of colorectal cancer. F1000Prime Rep. (2014) 6:108. doi: 10.12703/P6-108
58. Alkady MM, Abdel-Messeih PL, Nosseir NM. Assessment of serum levels of the adipocytokine chemerin in colorectal cancer patients. J Med Biochem. (2017) 37:313–9. doi: 10.1515/jomb-2017-0062
59. Lee JY, Lee MK, Kim NK, Chu SH, Lee DC, Lee HS, et al. Serum chemerin levels are independently associated with quality of life in colorectal cancer survivors: a pilot study. PLoS ONE (2017) 12:e0176929. doi: 10.1371/journal.pone.0176929
60. Kulig P, Kantyka T, Zabel BA, Banas M, Chyra A, Stefanska A, et al. Regulation of chemerin chemoattractant and antibacterial activity by human cysteine cathepsins. J Immunol. (2011) 187:1403–10. doi: 10.4049/jimmunol.1002352
61. Vironen JH, Kairaluoma M, Aalto AM, Kellokumpu IH. Impact of functional results on quality of life after rectal cancer surgery. Dis Colon Rectum. (2006) 49:568–78. doi: 10.1007/s10350-006-0513-6
62. Phipps E, Braitman LE, Stites S, Leighton JC. Quality of life and symptom attribution in long-term colon cancer survivors. J Eval Clin Pract. (2008) 14:254–8. doi: 10.1111/j.1365-2753.2007.00842.x
63. Yang H, Testa JR, Carbone M. Mesothelioma epidemiology, carcinogenesis, and pathogenesis. Curr Treat Options Oncol. (2008) 9:147–57. doi: 10.1007/s11864-008-0067-z
64. Dranse HJ, Zheng A, Comeau AM, Langille MGI, Zabel BA, Sinal CJ. The impact of chemerin or chemokine-like receptor 1 loss on the mouse gut microbiome. PeerJ. (2018) 6:e5494. doi: 10.7717/peerj.5494
65. Nagao-Kitamoto H, Kamada N. Host-microbial cross-talk in inflammatory bowel disease. Immune Netw. (2017) 17:1–12. doi: 10.4110/in.2017.17.1.1
66. Jemal A, Center MM, DeSantis C, Ward EM. Global patterns of cancer incidence and mortality rates and trends. Cancer Epidemiol Biomarkers Prev. (2010) 19:1893–907. doi: 10.1158/1055-9965.EPI-10-0437
67. Zhang J, Jin HC, Zhu AK, Ying RC, Wei W, Zhang FJ. Prognostic significance of plasma chemerin levels in patients with gastric cancer. Peptides (2014) 61:7–11. doi: 10.1016/j.peptides.2014.08.007
68. Yonemura Y, Endou Y, Fujita H, Fushida S, Bandou E, Taniguchi K, et al. Role of MMP-7 in the formation of peritoneal dissemination in gastric cancer. Gastric Cancer (2000) 3:63–70. doi: 10.1007/PL00011698
69. Ashizawa T, Okada R, Suzuki Y, Takagi M, Yamazaki T, Sumi T, et al. Clinical significance of interleukin-6 (IL-6) in the spread of gastric cancer: role of IL-6 as a prognostic factor. Gastric Cancer (2005) 8:124–31. doi: 10.1007/s10120-005-0315-x
70. Yang LP, Fu LC, Guo H, Xie LX. Expression of vascular endothelial growth factor C correlates with lymphatic vessel density and prognosis in human gastroesophageal junction carcinoma. Onkologie (2012) 35:88–93. doi: 10.1159/000336807
71. Takahashi Y, Kitadai Y, Bucana CD, Cleary KR, Ellis LM. Expression of vascular endothelial growth factor and its receptor, KDR, correlates with vascularity, metastasis, and proliferation of human colon cancer. Cancer Res. (1995) 55:3964–8.
72. Huang SP, Wu MS, Wang HP, Yang CS, Kuo ML, Lin JT. Correlation between serum levels of interleukin-6 and vascular endothelial growth factor in gastric carcinoma. J Gastroenterol Hepatol. (2002) 17:1165–9. doi: 10.1046/j.1440-1746.2002.02873.x
73. Fu H, Hu Z, Wen J, Wang K, Liu Y. TGF-beta promotes invasion and metastasis of gastric cancer cells by increasing fascin1 expression via ERK and JNK signal pathways. Acta Biochim Biophys Sin. (2009) 41:648–56. doi: 10.1093/abbs/gmp053
74. Ho CC, Lai KC, Hsu SC, Kuo CL, Ma CY, Lin ML, et al. Benzyl isothiocyanate (BITC) inhibits migration and invasion of human gastric cancer AGS cells via suppressing ERK signal pathways. Hum Exp Toxicol. (2011) 30:296–306. doi: 10.1177/0960327110371991
75. Fujimori Y, Inokuchi M, Takagi Y, Kato K, Kojima K, Sugihara K. Prognostic value of RKIP and p-ERK in gastric cancer. J Exp Clin Cancer Res. (2012) 31:30. doi: 10.1186/1756-9966-31-30
76. Graziosi L, Mencarelli A, Santorelli C, Renga B, Cipriani S, Cavazzoni E, et al. Mechanistic role of p38 MAPK in gastric cancer dissemination in a rodent model peritoneal metastasis. Eur J Pharmacol. (2012) 674:143–52. doi: 10.1016/j.ejphar.2011.11.015
77. Davis ME. Glioblastoma: overview of disease and treatment. Clin J Oncol Nurs. (2016) 20(5 Suppl.):S2–8. doi: 10.1188/16.CJON.S1.2-8
78. Tamimi AF, Juweid M. Epidemiology and Outcome of Glioblastoma. In: De Vleeschouwer S, editor. Glioblastoma. Brisbane, QLD: Codon Publications (2017). doi: 10.15586/codon.glioblastoma.2017
79. Zhao L, Yamaguchi Y, Sharif S, Du XY, Song JJ, Lee DM, et al. Chemerin158K protein is the dominant chemerin isoform in synovial and cerebrospinal fluids but not in plasma. J Biol Chem. (2011) 286:39520–7. doi: 10.1074/jbc.M111.258954
80. Mittal S, El-Serag HB. Epidemiology of hepatocellular carcinoma: consider the population. J Clin Gastroenterol. (2013) 47(Suppl.):S2–6. doi: 10.1097/MCG.0b013e3182872f29
81. Raza A, Sood GK. Hepatocellular carcinoma review: current treatment, and evidence-based medicine. World J Gastroenterol. (2014) 20:4115–27. doi: 10.3748/wjg.v20.i15.4115
82. Lin Y, Yang X, Liu W, Li B, Yin W, Shi Y, et al. Chemerin has a protective role in hepatocellular carcinoma by inhibiting the expression of IL-6 and GM-CSF and MDSC accumulation. Oncogene (2017) 36:3599–608. doi: 10.1038/onc.2016.516
83. Park BK, Zhang H, Zeng Q, Dai J, Keller ET, Giordano T, et al. NF-kappaB in breast cancer cells promotes osteolytic bone metastasis by inducing osteoclastogenesis via GM-CSF. Nat Med. (2007) 13:62–9. doi: 10.1038/nm1519
84. He G, Karin M. NF-kappaB and STAT3 - key players in liver inflammation and cancer. Cell Res. (2011) 21:159–68. doi: 10.1038/cr.2010.183
85. Berwick M, Buller DB, Cust A, Gallagher R, Lee TK, Meyskens F, et al. Melanoma epidemiology and prevention. Cancer Treat Res. (2016) 167:17–49. doi: 10.1007/978-3-319-22539-5_2
86. Ali Z, Yousaf N, Larkin J. Melanoma epidemiology, biology and prognosis. EJC Suppl. (2013) 11:81–91. doi: 10.1016/j.ejcsup.2013.07.012
87. Sato E, Olson SH, Ahn J, Bundy B, Nishikawa H, Qian F, et al. Intraepithelial CD8+ tumor-infiltrating lymphocytes and a high CD8+/regulatory T cell ratio are associated with favorable prognosis in ovarian cancer. Proc Natl Acad Sci USA. (2005) 102:18538–43. doi: 10.1073/pnas.0509182102
88. Curran MA, Montalvo W, Yagita H, Allison JP. PD-1 and CTLA-4 combination blockade expands infiltrating T cells and reduces regulatory T and myeloid cells within B16 melanoma tumors. Proc Natl Acad Sci USA. (2010) 107:4275–80. doi: 10.1073/pnas.0915174107
89. Hadeiba H, Sato T, Habtezion A, Oderup C, Pan J, Butcher EC. CCR9 expression defines tolerogenic plasmacytoid dendritic cells able to suppress acute graft-versus-host disease. Nat Immunol. (2008) 9:1253–60. doi: 10.1038/ni.1658
90. Ostrand-Rosenberg S. Myeloid-derived suppressor cells: more mechanisms for inhibiting antitumor immunity. Cancer Immunol Immunother. (2010) 59:1593–600. doi: 10.1007/s00262-010-0855-8
92. Whittle SB, Smith V, Doherty E, Zhao S, McCarty S, Zage PE. Overview and recent advances in the treatment of neuroblastoma. Expert Rev Anticancer Ther. (2017) 17:369–86. doi: 10.1080/14737140.2017.1285230
93. Esposito MR, Aveic S, Seydel A, Tonini GP. Neuroblastoma treatment in the post-genomic era. J Biomed Sci. (2017) 24:14. doi: 10.1186/s12929-017-0319-y
94. Graham KL, Zhang JV, Lewen S, Burke TM, Dang T, Zoudilova M, et al. A novel CMKLR1 small molecule antagonist suppresses CNS autoimmune inflammatory disease. PLoS ONE (2014) 9:e112925. doi: 10.1371/journal.pone.0112925
95. Reck M, Heigener DF, Mok T, Soria JC, Rabe KF. Management of non-small-cell lung cancer: recent developments. Lancet (2013) 382:709–19. doi: 10.1016/S0140-6736(13)61502-0
96. Ramalingam SS, Owonikoko TK, Khuri FR. Lung cancer: new biological insights and recent therapeutic advances. CA Cancer J Clin. (2011) 61:91–112. doi: 10.3322/caac.20102
97. Xu CH, Yang Y, Wang YC, Yan J, Qian LH. Prognostic significance of serum chemerin levels in patients with non-small cell lung cancer. Oncotarget (2017) 8:22483–9. doi: 10.18632/oncotarget.14956
98. Dunn GP, Old LJ, Schreiber RD. The three Es of cancer immunoediting. Annu Rev Immunol. (2004) 22:329–60. doi: 10.1146/annurev.immunol.22.012703.104803
99. Esendagli G, Bruderek K, Goldmann T, Busche A, Branscheid D, Vollmer E, et al. Malignant and non-malignant lung tissue areas are differentially populated by natural killer cells and regulatory T cells in non-small cell lung cancer. Lung Cancer (2008) 59:32–40. doi: 10.1016/j.lungcan.2007.07.022
100. Noone AM, Howlader N, Krapcho M, Miller D, Brest A, Yu M, et al. SEER Cancer Statistics Review. Bethesda, MD: National Cancer Institute (1975–2015). Available online at: https://seer.cancer.gov/csr/1975_2015/
101. Brawley OW. Trends in prostate cancer in the United States. J Natl Cancer Inst Monogr. (2012) 2012:152–6. doi: 10.1093/jncimonographs/lgs035
102. Weiner AB, Kundu SD. Prostate cancer: a contemporary approach to treatment and outcomes. Med Clin North Am. (2018) 102:215–29. doi: 10.1016/j.mcna.2017.10.001
103. Stamey TA, Warrington JA, Caldwell MC, Chen Z, Fan Z, Mahadevappa M, et al. Molecular genetic profiling of Gleason grade 4/5 prostate cancers compared to benign prostatic hyperplasia. J Urol. (2001) 166:2171–7. doi: 10.1016/S0022-5347(05)65528-0
104. Lin PC, Giannopoulou EG, Park K, Mosquera JM, Sboner A, Tewari AK, et al. Epigenomic alterations in localized and advanced prostate cancer. Neoplasia (2013) 15:373–83. doi: 10.1593/neo.122146
105. Abnet CC, Arnold M, Wei W-Q. Epidemiology of esophageal squamous cell carcinoma. Gastroenterology (2018) 154:360–73. doi: 10.1053/j.gastro.2017.08.023
106. Liang H, Fan JH, Qiao YL. Epidemiology, etiology, and prevention of esophageal squamous cell carcinoma in China. Cancer Biol Med. (2017) 14:33–41. doi: 10.20892/j.issn.2095-3941.2016.0093
107. Murray GI, Duncan ME, O'Neil P, McKay JA, Melvin WT, Fothergill JE. Matrix metalloproteinase-1 is associated with poor prognosis in oesophageal cancer. J Pathol. (1998) 185:256–61.
108. Groblewska M, Siewko M, Mroczko B, Szmitkowski M. The role of matrix metalloproteinases (MMPs) and their inhibitors (TIMPs) in the development of esophageal cancer. Folia Histochem Cytobiol. (2012) 50:12–9. doi: 10.2478/18691
109. Tao YS, Ma XY, Chai DM, Ma L, Feng ZZ, Cheng ZN, et al. Overexpression of MMP-1 and VEGF-C is associated with a less favorable prognosis in esophageal squamous cell carcinoma. Oncol Res Treat. (2012) 35:651–6. doi: 10.1159/000343637
110. Markopoulos AK. Current aspects on oral squamous cell carcinoma. Open Dent J. (2012) 6:126–30. doi: 10.2174/1874210601206010126
111. Gujrathi D, Kerr P, Anderson B, Nason R. Treatment outcome of squamous cell carcinoma of the oral tongue. J Otolaryngol. (1996) 25:145–9.
112. Sano D, Myers JN. Metastasis of squamous cell carcinoma of the oral tongue. Cancer Metastasis Rev. (2007) 26:645–62. doi: 10.1007/s10555-007-9082-y
113. Ghallab NA, Shaker OG. Serum and salivary levels of chemerin and MMP-9 in oral squamous cell carcinoma and oral premalignant lesions. Clin Oral Investig. (2017) 21:937–47. doi: 10.1007/s00784-016-1846-8
114. Lomas A, Leonardi-Bee J, Bath-Hextall F. A systematic review of worldwide incidence of nonmelanoma skin cancer. Br J Dermatol. (2012) 166:1069–80. doi: 10.1111/j.1365-2133.2012.10830.x
115. Wischermann K, Popp S, Moshir S, Scharfetter-Kochanek K, Wlaschek M, de Gruijl F, et al. UVA radiation causes DNA strand breaks, chromosomal aberrations and tumorigenic transformation in HaCaT skin keratinocytes. Oncogene (2008) 27:4269–80. doi: 10.1038/onc.2008.70
116. Parikh SA, Patel VA, Ratner D. Advances in the management of cutaneous squamous cell carcinoma. F1000Prime Rep. (2014) 6:70. doi: 10.12703/P6-70
Keywords: chemerin, RARRES2, CMKLR1, CCRL2, GPR1, cytokine, chemoattractant, cancer
Citation: Shin WJ, Zabel BA and Pachynski RK (2018) Mechanisms and Functions of Chemerin in Cancer: Potential Roles in Therapeutic Intervention. Front. Immunol. 9:2772. doi: 10.3389/fimmu.2018.02772
Received: 13 August 2018; Accepted: 12 November 2018;
Published: 30 November 2018.
Edited by:
Annalisa Del Prete, Università degli Studi di Brescia, ItalyReviewed by:
Marcus Thelen, Institute for Research in Biomedicine (IRB), SwitzerlandPaul Proost, KU Leuven, Belgium
Gunes Esendagli, Hacettepe University, Turkey
Copyright © 2018 Shin, Zabel and Pachynski. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Russell K. Pachynski, rkpachynski@wustl.edu