 Camelia Frantz
Camelia Frantz Cedric Auffray
Cedric Auffray Jerome Avouac
Jerome Avouac Yannick Allanore*
Yannick Allanore*- INSERM U1016, UMR8104, Cochin Institute, Paris Descartes University, Paris, France
In recent years, accumulating evidence suggest that regulatory T cells (Tregs) are of paramount importance for the maintenance of immunological self-tolerance and immune homeostasis, even though they represent only about 5–10% of the peripheral CD4+ T cells in humans. Their key role is indeed supported by the spontaneous development of autoimmune diseases after Tregs depletion in mice. Moreover, there is also a growing literature that investigates possible contribution of Tregs numbers and activity in various autoimmune diseases. The contribution of Tregs in autoimmune disease has opened up a new therapeutic avenue based on restoring a healthy balance between Tregs and effector T-cells, such as Treg-based cellular transfer or low-dose IL-2 modulation. These therapies hold the promise of modulating the immune system without immunosuppression, while several issues regarding efficacy and safety need to be addressed. Systemic sclerosis (SSc) is an orphan connective tissue disease characterized by extensive immune abnormalities but also microvascular injury and fibrosis. Recently, data about the presence and function of Tregs in the pathogenesis of SSc have emerged although they remain scarce so far. First, there is a general agreement in the medical literature with regard to the decreased functional ability of circulating Tregs in SSc. Second the quantification of Tregs in patients have led to contradictory results; although the majority of the studies report reduced frequencies, there are conversely some indications suggesting that in case of disease activity circulating Tregs may increase. This paradoxical situation could be the result of a compensatory, but inefficient, amplification of Tregs in the context of inflammation. Nevertheless, these results must be tempered with regards to the heterogeneity of the studies for the phenotyping of the patients and of the most importance for Tregs definition and activity markers. Therefore, taking into account the appealing developments of Tregs roles in autoimmune diseases, together with preliminary data published in SSc, there is growing interest in deciphering Tregs in SSc, both in humans and mice models, to clarify whether the promises obtained in other autoimmune diseases may also apply to SSc.
Regulatory T Cells: Gatekeepers of Immunological Tolerance
Human regulatory T cells (Tregs) expressing the transcription factor FoxP3 have a crucial role for the maintenance of immunological self-tolerance and immune homeostasis (1–3). The loss of dominant peripheral tolerance, exerted by Tregs, can lead to autoimmune diseases, immunopathology, allergy or metabolic disease (4, 5).
The pivotal role of Treg cells in the protection from autoimmunity is exemplified by spontaneous development of immunopathology in scurfy mice which are deficient for FoxP3+ Tregs (4, 6). Mutations in the human ortholog result in a similar X-linked lymphoproliferative disorder characterized by immune dysregulation, polyendocrinopathy, enteropathy, defined by the acronym IPEX (7, 8). Moreover, lack of Treg-mediated control has been shown to play a role in many animal models of autoimmunity (4) but also in numerous autoimmune disorders (9–12). In this review, we will focus on CD4+FoxP3+ Treg cell subset.
CD4+ Tregs: Cell Subsets and Identification
Human Tregs were first characterized as CD4+CD25+ T cells in 2001 (13–15) based on the findings by Sakaguchi et al. that mouse Tregs constitutively express CD25, the α-chain of the IL2 receptor (16). However, CD25 is also upregulated on responder T cells upon activation. Therefore, much research has been focused on the identification of further markers to precisely distinguish the Treg population from recently activated T cells. In 2003, the transcription factor FoxP3 was shown to regulate the generation and function of Tregs in mice (17–19). Subsequently, in humans, FoxP3 was shown to be expressed predominantly by CD4+CD25high T cells (20). However, whereas in mice FoxP3 expression seems to be restricted to Tregs (21, 22), in humans, FoxP3 is also expressed by non-regulatory CD4+CD25+ T cells (23, 24), restricting the usage of FoxP3 as a specific marker for human Tregs. Moreover, FoxP3 being an intracellular protein, it cannot be used for Tregs isolation with the goal to perform functional studies. Later, it was shown that low expression of CD127, the α-chain of the IL7 receptor, acts as an additional marker for the characterization of Treg cells among CD4+CD25high T cells (25, 26). Indeed, FoxP3 expression and suppressive capacity are enriched in CD4+ T cells that express low levels of CD127. However, CD127 expression tends to be downregulated also in activated conventional CD4+ T cells (27, 28). Moreover, Klein et al. demonstrated that a high percentage of CD127+ cells expressed FoxP3 and, reciprocally, that there was a high percentage of CD127low/− cells that did not express FoxP3. These results suggest that these markers did not represent the same population of Tregs (29). A number of additional cell markers for the identification of CD4+ Tregs have been proposed (30–32) but many of these are also induced upon activation of non-regulatory CD4+ T cells.
In this context, Miyara and coworkers further delineated the Treg cell compartment into three subpopulations using the combination of FoxP3 and CD45RA expression: (i) CD45RA+FoxP3low resting Tregs (rTregs); (ii) CD45RA−FoxP3high activated Tregs (aTregs), both of which are strongly suppressive in vitro; and (iii) non-suppressive cytokine-secreting CD45RA−FoxP3low non-Tregs (33). In vitro, CD45RA−FoxP3high aTregs were activated, highly suppressive and died by apoptosis after exertion of suppression, whereas rTregs were in a quiescent state, proliferated upon activation and converted into aTreg cells in vitro and in vivo. A major stake of this combination is based on the identification of a non-regulatory FoxP3+ T cell population, enabling to overcome the contamination by this cell population when studying Treg cells.
CD49d (α-chain of the integrin VLA-4) was also described as a marker that could discriminate contaminating effector cells from immune-suppressive Foxp3+ Treg cells. This marker is present on the majority of proinflammatory effector cells but absent from Foxp3+ Treg cells. Therefore, depletion with α-CD49d removes proinflammatory effector cells from CD25highCD4+ cells and, in combination with α-CD127, it provides access to hugely pure populations of Foxp3+ cells (34).
Mechanisms of Action and Functional Characterization of CD4+ Treg
The best common way to analyze Treg function is based on their capacity to suppress target cell proliferation, and consist of in vitro suppression assays. This method relies on isolation of effector and regulatory cell populations immunomagnetically or by fluorescence activated cell sorting (FACS). Effector cells are then activated in the presence or absence of the regulatory population. After a defined period of time, their proliferation, and/or cytokine production are examined. However, FoxP3 being an intracellular protein, live human Tregs cannot be isolated using FoxP3 as a marker, and the lack of specific Treg cell surface markers precludes the isolation of a pure Treg population to test in these in vitro suppression assays.
Numerous mechanisms have been described as to how Tregs exert their suppressive function, including cell-cell contact dependent suppression, inhibitory cytokine release (IL-10, TGFβ, IL-35, Granzymes A et B), IL-2 deprivation, modulation of antigen-presenting cell function via CTLA-4, cytolysis and metabolic disruption of the target cell. These mechanisms have been extensively reviewed (35–38) and will not be further discussed in this article.
Defects in the number and/or function of Treg cells could each lead to a suboptimal T cell regulation, and subsequently to the development of autoimmunity.
Systemic Sclerosis
Systemic sclerosis (SSc) is an orphan connective tissue disease characterized by extensive immune abnormalities, microvascular injury and fibrosis of skin and internal organs (39). It is the most severe connective tissue disease, associated with a high mortality risk (40). Patients with SSc are classified according to skin involvement extent: limited cutaneous SSc (LcSSc), with skin involvement restricted to the hands, arms, and face; and diffuse cutaneous SSc (DcSSc), with more extensive skin thickening (truncal and proximal) and more frequent visceral involvement (41).
Although the pathogenesis of SSc is complex and remains incompletely understood (42), research in the area has confirmed that immune dysfunction is one of the most important component of the pathogenesis. Innate and adaptive immune abnormalities can be observed, and culminate in auto-antibodies production and activation of cell-mediated autoimmunity. Moreover, immune cells may trigger the complex biochemical and molecular changes that promote vasculopathy and fibrosis. Indeed, there is increasing evidence that places immune activation as a cause and not a consequence of the vasculopathy and fibrosis. First, histological studies indicate that an inflammatory infiltrate is present in the very early stages, preceding the onset of fibrosis (43). This cellular infiltrates consist mostly of T cells which are predominantly CD4+ cells (44). Second, fibroblasts with increased expression of type I and III procollagen mRNA can often be identified in areas adjacent to the infiltrating mononuclear cells (45, 46). Third, T cells in the skin and in the peripheral blood of SSc patients express an oligoclonal T cell receptor (TCR) repertoire, strongly suggestive of a proliferation and clonal expansion of these cells in response to a specific Ag(s) (47, 48). Furthermore, several studies have demonstrated an association of particular HLA alleles with SSc (49–52), which supports the concept of an Ag-driven T cell response in SSc. It should be noted that the genotype varies particularly strongly according to the presence of different types of autoantibodies associated with SSc: anti-centromere antibodies was associated with DRB1*01:01, DRB1*01:04, DRB1*01:08, DQB1*05:01, DPB1*04:02 and anti-topoisomerase I with DRB1*11-*15:02, DPB1*13:01 and DPB1**09:01 (51, 52). In a large study of HLA class II genes carried out in 1,300 SSc cases and 1,000 controls, the DRB1*11:04, DQA1*05:01 and DQB1*03:01 haplotypes and the DQB1 allele were the strongest associations identified (49). The association of DRB1*11:04, DQA1*05:01 and DQB1*03:01 haplotype with SSc was confirmed in a similar study (944 Caucasian SSc patients and 1,320 unaffected controls) (50). Although not specific to SSc, these HLAs were not found in many other autoimmune diseases.
Therefore, among this aberrant immune response, T lymphocytes seem to be of particular importance in the pathogenesis of SSc. These cells are predominantly CD4+ cells, display markers of activation, with a predominant Th2 cytokine profile characterized by high levels of IL-4, IL-5, and IL-13 in skin, lung and peripheral blood (53–56). This key role of T cell proliferation and cytokine secretion in SSc suggests that this condition could be associated with a defective control of T cell activation.
Circulating Treg in SSc
Evidence for numerical and functional changes of Treg population in SSc has been obtained in several studies (Table 1). The majority of the studies reported decreased frequencies and/or impaired function of circulating Tregs in SSc patients compared to controls (12, 57–65, 74). Banica et al. investigated Treg cells in peripheral blood of patients with different connective tissue diseases, as compared with blood from healthy controls. They found a reduced percentage of CD4+CD25hi T cells in SSc compared to controls but also to other connective tissue diseases (12). Antiga et al. also reported fewer CD4+CD25brightFoxP3+ cells in SSc patients naïve to any systemic treatment compared with healthy controls and with patients having other common immune-mediate dermatoses (psoriasis, atopic dermatitis) (57). This decrease was associated with reduced total TGFβ1 and IL-10 serum levels. The authors concluded that this reduced frequency of Tregs, together with that of total TGFβ1 and IL-10, may be responsible for the loss of tolerance observed in SSc. Papp and coworkers observed decreased CD4+CD25+FoxP3+ T cells percentages in peripheral blood of patients with SSc associated with increased Th17 cell percentages and decreased circulating IL-10 levels (59). In addition, both CD4+ and CD8+ central memory T-cell percentages were increased representing an immunologically active state. Similarly, the results of Fenoglio et al. supported an imbalanced ratio between Th17 and Treg cell subsets in SSc patients, with increased proportion of circulating Th17 cells, and decreased proportion of both CD4+CD25+CD127− and CD8+CD28− Treg cells (60). Lower frequencies of CD4+CD25+FoxP3+ T cells in SSc patients were reproducibly reported by other groups (62–64). Kataoka et al. found reduced frequencies of CD4+CD25+FoxP3+ T cells in treatment-free SSc patients compared to healthy controls, particularly in patients with late-stage disease (65).
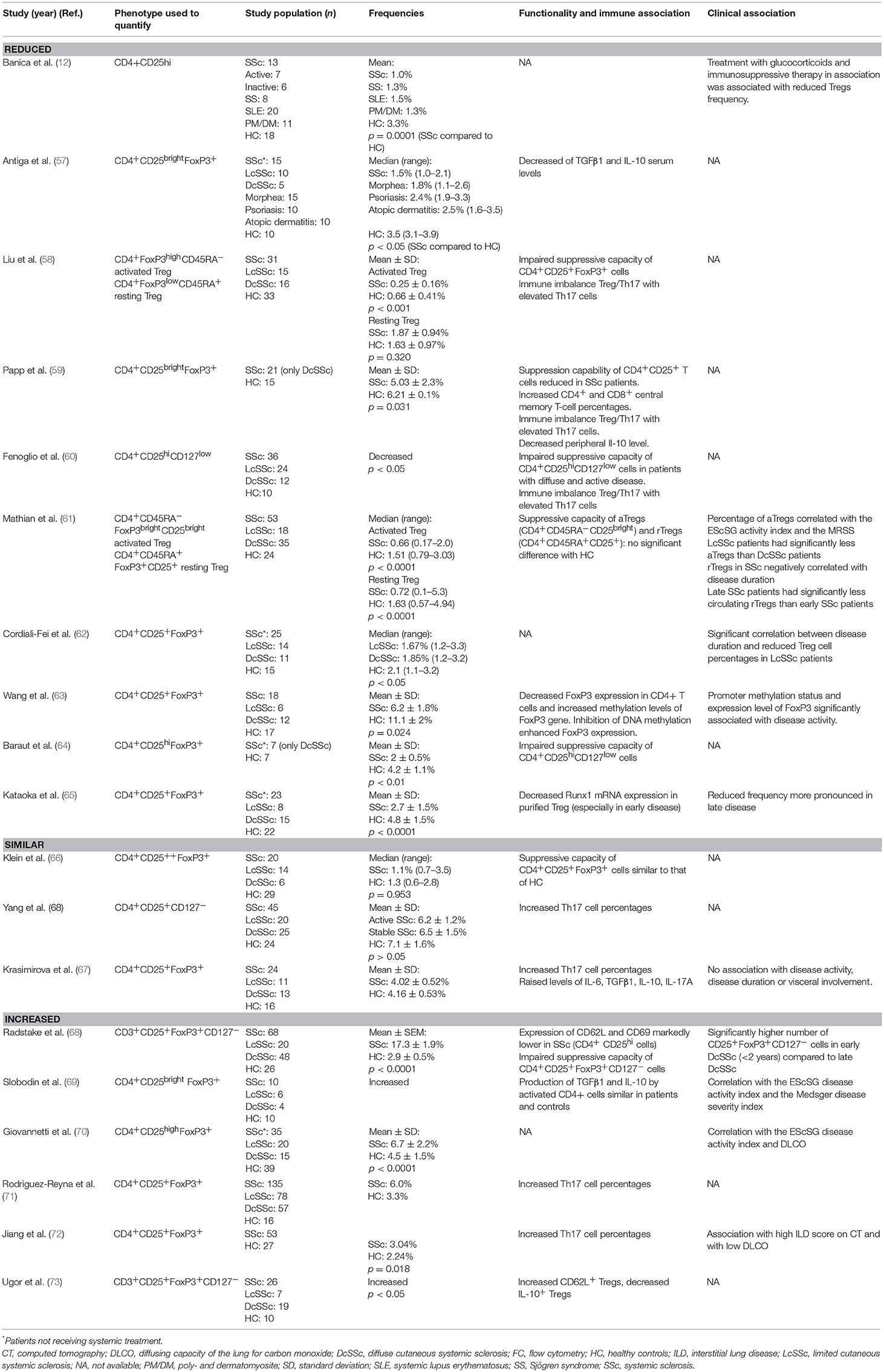
Table 1. Circulating regulatory T cells in systemic sclerosis.
Among other reports, some studies have reported decreased frequency of circulating Tregs but not reaching statistical significance (66, 67, 75).
Some studies have reported an increase in circulating Tregs (68–73), particularly in early phase and active disease (68–70). Radstake et al. reported an increase in the frequency of circulating CD25+FoxP3+CD127− T cells in SSc patients, especially in early phase of the disease. However, despite this increase, Tregs from SSc patients harbored a defective suppressive capacity correlated with a dramatic reduction in CD62L and CD69 expression. Interestingly, co-incubation of Treg cells from healthy donors with plasma from SSc patients abrogated suppressive activity, suggesting the presence of specific soluble factors inhibiting Treg function in SSc patients (68). Two studies reported higher number of CD25+FoxP3+ T cells in SSc patients, correlated with disease activity and severity (70, 69). Three more studies reported a significantly higher frequency of circulating Tregs in SSc compared to controls, although no functional studies were performed (71–73).
These discrepancies reflect the challenge of the phenotypic characterization of Treg cells and possible contamination by activated CD25+ T cells. In this context, two studies have used the combination of markers described by Miyara et al. (33) discriminating CD4+FoxP3lowCD45RA+ resting Treg (rTreg), CD4+FoxP3highCD45RA− activated Treg (aTreg) from non-regulatory Foxp3+ cells. Mathian and coworkers found that the percentages and absolute counts for both aTreg and rTreg were decreased in SSc compared to controls, but not those for non-regulatory FoxP3+ CD4+ T cells (61). Interestingly, aTreg were decreased at any disease stage while rTreg frequency declined in late phases of SSc. Moreover, the quantitative Treg defect was less pronounced in diffuse cutaneous and/or active disease. Similarly, Liu et al. reported lower proportions of aTreg and higher proportions of non-regulatory Foxp3+ cells in SSc patients compared to healthy controls (58). In the latter study, the frequency of CD4+CD25+FoxP3+ Treg cells was significantly increased in patients with SSc, suggesting that the increase in this cell population was mainly due to elevated CD4+CD25+FoxP3lowCD45RA− non-Treg cells. These results within the same patient population support the notion that the complexity of the phenotypic characterization of this cell population explains conflicting results in the literature.
However, the discrepancies among these studies cannot be solely explained by the use of different Treg markers. Indeed, it should be emphasized that flow cytometry gating is rather subjective and depends partly on researcher selection. Therefore, studies using same Treg markers are not necessarily comparable. Moreover, the patients' characteristics, such as disease duration, disease severity and activity, concomitant treatments, might also contribute to the discrepancy among different studies as previously stated.
As regards clinical association, most of the studies reporting increased frequency of circulating Tregs have demonstrated a correlation with disease activity (69, 70) and severity (69, 72), and with early disease (68), whereas reduced frequency of Tregs seemed to be associated with late disease (61, 62, 65). Elevated Tregs were also reported in patients with a high interstitial lung disease (ILD) score on computed tomography (72) and with low DLCO (70, 72). No other clinical association was found, in particular, no difference was observed between the two subsets of the disease.
Effect of medications on Tregs frequency have been raised by some authors. It is of note that most of the patients in studies reporting increased or similar Tregs frequency received systemic treatment which may have bias the results, since some authors reported that immunosuppressive therapy is able to increase the pool of circulating Treg (76, 77). On the other hand, immunosuppressive therapy and bosentan showed no significant effect on the frequency of Tregs in some reports (66, 73). In contrast, treatment with glucocorticoids and immunosuppressive therapy in association was associated with reduced Tregs frequency in the study of Banica et al. (12). Thus, more investigations are needed to evaluate the impact of therapies on Treg cells.
Regarding the functional capacity of circulating Tregs, almost all studies agree that Tregs fail to produce inhibitory cytokines or suppress the effector T cells in SSc (58–60, 64, 68).
Pulmonary arterial hypertension (PAH) is one of the most severe complication of SSc. Several studies have investigated the role of Tregs in PAH. Two studies reported elevated CD4+CD25+FoxP3+ T cells in the peripheral blood of idiopathic PAH patients (78, 79), but not in the lungs (78). Huertas et al. investigated the functional status of CD4+CD25+FoxP3+ Treg function by measuring Treg STAT3 phosphorylation in patients with idiopathic PAH, heritable PAH or SSc-PAH, compared to controls (80). Although Treg cell numbers were similar between patients and controls, they found that Tregs were dysfunctional in all these PAH subgroups, including SSc-PAH, with reduced proportion of Treg-pSTAT3+ cells compared to controls.
Mechanisms of Tregs Dysfunction in SSc
Mechanisms of Tregs dysfunction in SSc have been investigated by several studies. Aberrant epigenetic modifications, such as microRNA, DNA methylation, histone modifications, affecting FoxP3 and other key genes in Tregs have been shown to contribute to disease activity and tissue inflammation in autoimmune diseases (81). In SSc, Wang et al. reported elevated methylation levels of the FoxP3 promoter, inversely correlated with FoxP3 mRNA expression, and accompanied by reduced proportion of CD4+CD25+FoxP3+ Tregs (63). Furthermore, treatment of SSc CD4+ T cells with 5-azacytidine, a DNA methylation inhibitor, reduced the mean methylation levels, increased FoxP3 expression and induced Treg generation. Interestingly, the promoter methylation status and expression level of Foxp3 were significantly associated with disease activity. D'Amico et al. provide evidence of the association between rs2294020 FoxP3 polymorphism and disease progression in a female Italian population (82). Otherwise, it has been reported a significantly higher frequency of skewed X chromosomal inactivation in patients with SSc compared with controls, correlated with lower FoxP3 expression in CD4+CD25+ cells and less efficient suppressive activity (83). Kataoka and al. reported reduced expression of the transcription factor Runx1 mRNA correlated with decreased proportion of Tregs even in early stages of the disease (65). Semaphorin 3A serum levels along with cell expression on Tregs were reported to be low in SSc patients (84).
In addition, recent evidence indicate that Tregs could contribute to SSc pathogenesis by conversion into pathogenic effector T cells in the presence of appropriate environment. Thus, because accumulating evidence suggest that Th17 cells could be responsible for prominent features of SSc (53, 71, 85), it has been hypothesized that a Treg/Th17 imbalance could be a pivotal component of SSc pathogenesis. Indeed, Fenoglio et al. found a significant correlation between increased circulating Th17 cells and alteration of the Treg compartment (60). One could argue that the observed decrease in Treg cells could be the result of conversion to Th17. Several groups have reported the conversion of Tregs to Th17 cells in both mouse and human (86–88), supporting this hypothesis. Moreover, IL-6 and IL-1β, that are highly expressed in inflammatory conditions, have been shown to convert Tregs to Th17 cells (89, 90). Liu et al. found that CD4+CD25+FoxP3lowCD45RA− non-regulatory T cells produced high levels of IL-17 (58). They hypothesized that this population of FoxP3+ non-regulatory T cells expressing IL-17 could represent a transitional phase in the conversion process from Treg to Th17 cells. Consistent with this result, T cells that co-express IL-17 and FoxP3 have been identified by other groups (91, 92). In contrast, although they reported decreased Tregs proportions, no IL-17 amplification was observed in blood and skin of SSc patients in the study of Mathian et al. (61).
More recently, MacDonald et al., using flow cytometry, analyzed FoxP3 and cytokine expression among skin-resident T cells isolated from cultured explants (93). They found that Tregs from SSc-skin produced significant amount of Th2 cell-associated cytokines IL-4 and IL-13 compared to controls. On the other hand, circulating Tregs of SSc patients did not produce Th2 cytokines, but they contained a significantly higher proportion of skin-homing cells expressing Th2 cell-associated chemokine receptors. The authors also found evidence that IL-33 might be an important stimulator of tissue-localized loss of normal Tregs function and polarization into Th2-like cells. Altogether, these results further support the hypothesis that the skin of SSc patients provides the appropriate environment for transdifferentiation of Tregs toward a Th2-like phenotype, that might contribute to fibrosis in patients with SSc.
Tregs at the Site of Inflammation in SSc
When studying Treg cells in such diseases, one needs to consider potential differences between Treg cells derived from the peripheral blood vs. the inflamed organs (skin, lungs) in terms of function and frequency. In contrast to the lung, skin-resident Tregs are being actively investigated, probably due to relative ease of access for tissue samples. Therefore, data from mice and human subjects have revealed the importance of correct Treg cell positioning in the skin for the maintenance of immune homeostasis and prevention of spontaneous autoimmune and inflammatory disease (94). Contradictory results have been reported by the few studies investigating the presence of Tregs in the skin of patients with SSc (Table 2). Earlier studies have found fewer FoxP3+ cells by immunohistochemistry as compared to healthy controls or control diseases (psoriasis and atopic dermatitis) (57, 66). Interestingly, no significant difference was revealed when comparing lesional and non-lesional skin of SSc patients (66). This decrease was associated with reduced TGFβ and IL-10, which are regulatory cytokines involved in Treg suppressive function, both in skin and blood of SSc patients in the study of Antiga and coworkers (57). By contrast, in the study of Yang et al. FoxP3+ Treg cells was reported to be enriched in both the dermis and epidermis of patients with early SSc compared with patients with late SSc and healthy controls (75). The authors hypothesized that this expansion of FoxP3+ cells in early SSc skin may reflect a regulatory feedback mechanism to restore cellular tolerance and ameliorate harmful autoimmune responses. It should be noted that disease duration was not reported in the two studies that have found fewer FoxP3+ cells. More recently, MacDonald et al., found that FoxP3+ cells with high IL-4 and IL-13 production could be detected more frequently in the skin of SSc patients compared to normal controls (93). This study provides the first evidence for the differentiation of human Treg cells into Th2 cytokine-producing cells that might contribute to fibrosis in patients with SSc.
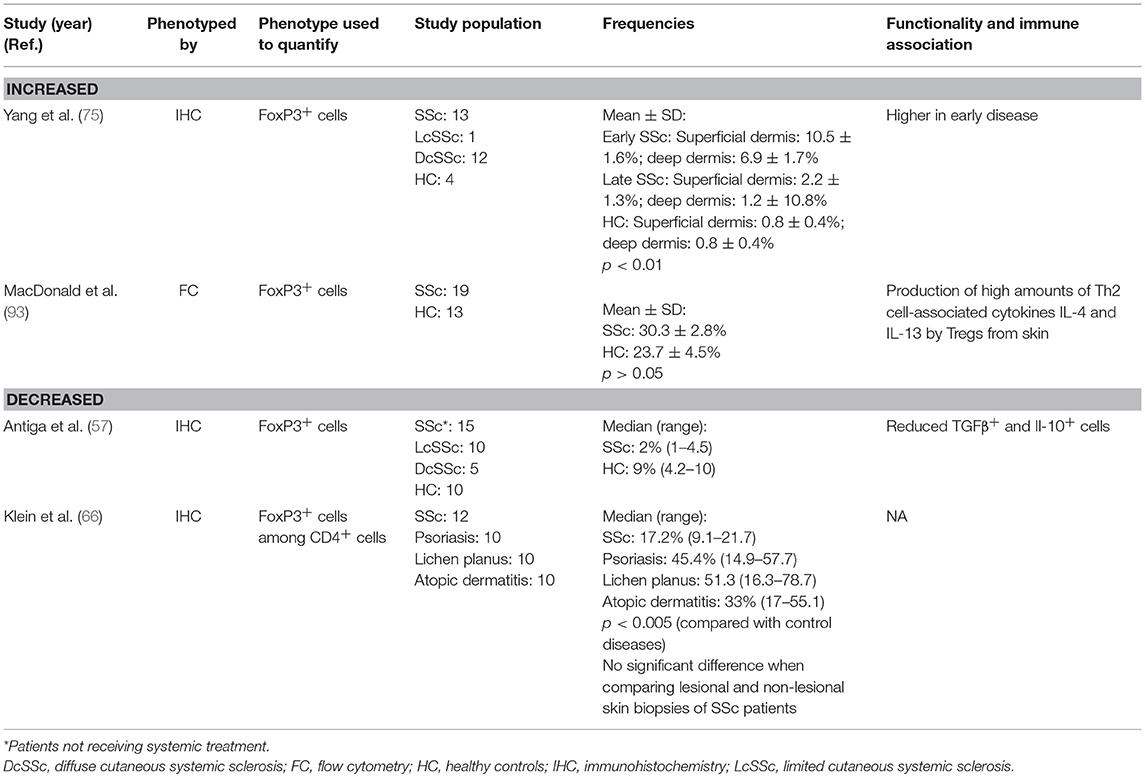
Table 2. Regulatory T cells in systemic sclerosis skin.
Tregs in SSc Mouse Models
Numerous inducible and genetic mouse models of SSc have been developed and characterized in the last years (95). Conversely to other autoimmune diseases, very scarce data about Tregs in SSc mouse models have been produced. In the topoisomerase mouse model, treatment with topoisomerase I and Freund's complete adjuvant (CFA) induces SSc-like skin, lung fibrosis and autoimmune abnormalities with anti-topoisomerase I auto-antibody production (96). This was associated with increased IL-6, TGFβ1, and IL-17 production and decreased IL-10 production. In this model, mice treated with topoisomerase I and CFA exhibited significantly increased frequencies of Th1 cells, Th2 cells, Th17 cells and Treg cells in bronchoalveolar lavage fluid compared with mice treated with saline or with topoisomerase I. Functional characteristics of Tregs was not assessed in this study.
In the mouse model of bleomycin-induced pulmonary fibrosis, contradictory results have been produced. Birjandi et al. found that treatment by IL-2 complex, used to expand CD4+CD25highFoxP3+ cells in the lung, leads to immune deviation that is dominated by type 2 immune response within the lung, and associated with exacerbate lung fibrosis (97). Moreover, they showed that bleomycin had a modifying and profibrotic effect on the CD4+CD25highFoxp3+ cells. This was corroborated by adoptive transfer experiments in Rag−/− mice. The authors concluded that a therapeutic strategy of expanding CD4+CD25highFoxP3+ in humans may be harmful via the augmentation of Th2 immune responses in patients with idiopathic pulmonary fibrosis and other fibroproliferative lung diseases. On the other hand, adoptive transfer of Tregs on day 14 after a bleomycin challenge significantly reduced pulmonary fibrosis in another report (98). Moreover, although splenocytes significantly improved bleomycin-induced pulmonary fibrosis when they were administered on day 14, this effect was abolished by depleting Tregs with an anti-CD25 monoclonal antibody. Finally, another group found that early depletion of Tregs with an anti-CD25 antibody led to favorable outcomes whereas late depletion of Tregs led to increased fibrosis, suggesting that Tregs play a detrimental role in early stages but protective role in late stages of pulmonary fibrosis in mice (99).
Although these results have to be taken into account in future therapeutic strategy using Treg cells in such diseases, it should be noted that extrapolation of these data from a mouse model to human is challenging since bleomycin may not accurately recapitulate human SSc ILD or other human fibrotic lung diseases. Moreover, there remains much controversy in the field about the actual role of Tregs.
Therapeutics Application
Taking into account that Tregs are immunodominant suppressors, there is a huge interest in the therapeutic potential of Tregs in several immune-mediated diseases. Indeed, adoptive cellular therapies may offer fewer risks and better efficacy than traditional pharmacological strategies. So far, clinical research has involved mostly hematopoietic stem cell transplantations, solid organ transplantations, and autoimmunity. Mechanisms of actions are incompletely understood but Tregs protect from auto-aggression and damage to tissues; the effect is executed mainly via cell-to-cell contacts, and also “control by starvation/theft” of IL-2. In general, what is critical is the balance between Tregs and effector T cells (Teff). Therefore, changing the balance between Tregs and Teff is a promising avenue to restore immune homeostasis and to treat autoimmune diseases. Moreover, since the recognition of antigen is a central part in Treg function and their therapeutic use, the modulation of T cell receptor specificity may be offer very stimulating perspectives (100).
Early reports demonstrated that adoptive transfer of Treg cells associated with hematopoietic stem cell transplantation (HSCT) in mice promoted the graft vs. leukemia effect (GvL) and protected from graft vs. host disease (GvHD). Unfortunately, this simple strategy cannot be translated to humans, such as the clinical efficacy requires the administration of a high number of cells whereas Treg cells represent a very low percentage of leukocytes in the blood. Thus, manufacturing procedures to expand Tregs in vitro before administration was developed. Multiple sclerosis (MS) is a well-defined autoimmune disease with solid evidence of Treg involvement. Remarkably, remission or prevention of experimental autoimmune encephalomyelitis (EAE), was associated with the induction of CD4+CD25+ Tregs. The adoptive transfer of Tregs further confirmed this hypothesis: transfer before EAE induction prevented EAE, and transfer to mice that already had EAE alleviated symptoms (101).
In humans, infusion of Tregs that is the direct approach to increase Tregs have been used in several phase 1 clinical trials for the prevention of GvHD or type 1 diabetes. Currently, several trials are registered in clinicaltrials.gov website for various conditions, such as liver transplantation (NCT01624077), autoimmune hepatitis (NCT02704338), chronic graft vs. host disease (NCT02385019 and NCT01937468), kidney transplantation (NCT02088931), type 1 diabetes (NCT01210664), systemic lupus erythematosus (NCT02428309). Although it is manageable to produce large numbers of alloantigen-reactive Tregs through selective stimulation by allogeneic antigen presenting cells, producing a high number of tissue antigen-specific Tregs for autoimmune diseases is far more challenging. Indeed, the low frequency of Treg precursor and the tendency of Tregs to destabilize after repeated in vitro stimulation impair easy large quantity production (102, 103).
Manipulating Tregs is another avenue that may even be complementary to adoptive cell transfer. In order to restore a safe equilibrium between Treg and Teff, anti-CD3 strategies mainly using antibodies was first developed (104, 105). However, efficacy was restricted to some patients and only at early stages. Co-stimulation may also be used and low-dose CTLA4-Ig therapy can enhance Treg and prevent immune activation (105). Finally, the most stimulating findings have been reported with IL2 and relate to the harboring by Tregs of high affinity IL-2 receptor promoting preferential expansion in conditions of low amounts of IL-2. Very stimulating data have been reported in humans in type 1 diabetes (106), GvHD (107), and regrowth of the scalp and/or body hair could be seen in alopecia areata (108). Patients with hepatitis C virus-induced vasculitis have a set of symptoms including fatigue, skin purpura, arthralgia, neuropathy and kidney involvement. In eight out of ten patients treated with low-dose IL-2, these symptoms progressively disappeared. It must be pointed out that in most cases, clinical improvements started to be observed after the second or third course of IL-2 therapy (109). Systemic lupus erythematosus (SLE) shares with SSc several immune disturbances. A dysbalance between Treg and Teff was recently confirmed in SLE with correlations between these findings and disease activity. In vitro experiments showed that lack of IL-2 production by CD4+ T cells accounted for the loss of CD25 expression in SLE Treg (110). Preliminary data in few patients receiving low-dose IL2 showed effectiveness to expand Treg (111). Clinical effects are now under investigations. Thus, IL-2 therapy is a promising avenue for expanding Treg cells and improving clinical outcomes for patients with autoimmune disease and trials are ongoing in connective tissues diseases including systemic sclerosis (TRANSREG study NCT01988506).
Similarly, anti-CD25 therapy results in prevention of activation and proliferation of T cells and inhibition of T cell responses. It is indicated for the prevention of acute organ rejection in adult and pediatric renal transplant recipients in combination with other immunosuppressive agents and has been studied in some immune diseases. Basiliximab is a chimeric (human/murine) anti-CD25 monoclonal antibody. It was administered to 10 SSc patients with severe skin involvement in addition to concomitant immunosuppressive and vasoactive treatments (112). Outcomes showed a reduction in skin fibrosis at week 68 and improvement in lung function at week 44. Treatment with basiliximab was well-tolerated. Although erythema, transient nausea, fatigue and weakness were common, severe reactions with significant dyspnea occurred in only one case. No patient had a documented severe infection and only one patient needed antibacterial therapy because of suspected respiratory infection. However, the application of this therapy in auto-immune diseases could be questionable. Indeed, since CD25 is also expressed on regulatory T cells, elimination or inhibition of the functional capacity of this subset using CD25 antibody might be counterproductive.
Use of autologous hematopoietic stem cell transplantation SCT (aHSCT) has recently gained interest in systemic sclerosis (64, 113–115) although the right regimen and best patient profile remain a matter of debate. It must be pointed out that sustained regression of skin and lung fibrosis has been reported for some patients. After aHSCT, reappearance of functional B cells, T-cell development, reconstitution of effector cells and efficient antigen presentation to reconstitute the pre-transplantation immune repertoire has been described. In SSc, there is scarce data about Treg restoration after aHSCT and contradictory results have been reported. A preliminary report about 7 patients focused on Treg showing a decrease number of Treg at baseline and altered suppressive capacity contrasting with restoration of Treg numbers and suppressive activity 24 months after aHSCT although a high variability was observed (64). On the other hand, in another report about 11 patients, both CD4+ activated effector T cells and Tregs did not reconstitute well after aHSCT with high dose cyclophosphamide conditioning, although the patients displayed resolution of clinical SSc (115). Therefore, the authors concluded that a complete reconstitution of the immune system, including Tregs, is not necessary for a treatment effect. Besides reconstitution of Treg numbers and suppressive capacity, increase in diversity of the TCR repertoire of Treg could be a crucial mechanism for the re-establishment of immune tolerance after aHSCT (116). Thus, future research into the effect of aHSCT on Treg cell compartment in SSc is required to clarify the underlying mechanisms of Treg cell pool renewal and the potential link with clinical outcome.
Conclusion
Evidence for numerical and functional changes of Tregs in SSc has been obtained in several studies. While the majority of the studies reported reduced frequencies of circulating Tregs in SSc patients compared to controls, it seems that some patients, especially in early phase and active disease have increased number of circulating Tregs. This paradoxical situation could be the result of a compensatory, but inefficient, amplification of Tregs in the context of active inflammation. In addition to diminished suppressive capacity, recent evidence indicate that Tregs could contribute to SSc pathogenesis by conversion into pathogenic effector T cells in the presence of appropriate environment, such as Th17 cells and Th2 cytokine-producing cells.
Nevertheless, these results should be tempered with regards to the heterogeneity of the studies in terms of patient's phenotype, and of the most importance regarding Tregs definition and activity markers. Moreover, although most previous studies analyzed peripheral blood of patients, the studies on Treg cells investigating phenotype and function in the site of inflammation are still sparse. Furthermore, conversely to other auto-immune diseases, very scarce data about Tregs in SSc mouse models have been produced. Thus, since the available data points toward a central role of Treg cells in SSc, future research is definitively needed to clarify the role of this cell population in SSc pathogenesis.
Finally, since the effect of existing treatment modalities on Tregs in SSc has not been elaborated sufficiently, gaining a better understanding of the natural history of Treg function and the affected mechanisms in SSc will certainly lead to new avenues in therapy, and will help to clarify whether the promises obtained in other autoimmune diseases may also apply to SSc.
Author Contributions
CF wrote the first draft of the manuscript; YA wrote sections of the manuscript. All authors contributed to manuscript revision, read and approved the submitted version.
Funding
This work was supported by a grant from the Association des Sclerodermiques de France.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer SS and handling Editor declared their shared affiliation.
References
1. Sakaguchi S, Miyara M, Costantino CM, Hafler DA. FOXP3+ regulatory T cells in the human immune system. Nat Rev Immunol. (2010) 10:490–500. doi: 10.1038/nri2785
2. Sakaguchi S, Yamaguchi T, Nomura T, Ono M. Regulatory T cells and immune tolerance. Cell (2008) 133:775–87. doi: 10.1016/j.cell.2008.05.009
3. Wing JB, Sakaguchi S. Multiple treg suppressive modules and their adaptability. Front Immunol. (2012) 3:178. doi: 10.3389/fimmu.2012.00178
4. Sakaguchi S, Ono M, Setoguchi R, Yagi H, Hori S, Fehervari Z, et al. Foxp3+ CD25+ CD4+ natural regulatory T cells in dominant self-tolerance and autoimmune disease. Immunol Rev. (2006) 212:8–27. doi: 10.1111/j.0105-2896.2006.00427.x
5. Calzada D, Baos S, Cremades-Jimeno L, Cárdaba B. Immunological mechanisms in allergic diseases and allergen tolerance: the role of Treg cells. J Immunol Res. (2018) 2018:6012053. doi: 10.1155/2018/6012053
6. Brunkow ME, Jeffery EW, Hjerrild KA, Paeper B, Clark LB, Yasayko S-A, et al. Disruption of a new forkhead/winged-helix protein, scurfin, results in the fatal lymphoproliferative disorder of the scurfy mouse. Nat Genet. (2001) 27:68–73. doi: 10.1038/83784
7. Bennett CL, Christie J, Ramsdell F, Brunkow ME, Ferguson PJ, Whitesell L, et al. The immune dysregulation, polyendocrinopathy, enteropathy, X-linked syndrome (IPEX) is caused by mutations of FOXP3. Nat Genet. (2001) 27:20–1. doi: 10.1038/83713
8. Wildin RS, Ramsdell F, Peake J, Faravelli F, Casanova JL, Buist N, et al. X-linked neonatal diabetes mellitus, enteropathy and endocrinopathy syndrome is the human equivalent of mouse scurfy. Nat Genet. (2001) 27:18–20. doi: 10.1038/83707
9. Hull CM, Peakman M, Tree TIM. Regulatory T cell dysfunction in type 1 diabetes: what's broken and how can we fix it? Diabetologia (2017) 60:1839–50. doi: 10.1007/s00125-017-4377-1
10. Rother N, van der Vlag J. Disturbed T cell signaling and altered Th17 and regulatory t cell subsets in the pathogenesis of systemic lupus erythematosus. Front Immunol. (2015) 6:610. doi: 10.3389/fimmu.2015.00610
11. Kleinewietfeld M, Hafler DA. Regulatory T cells in autoimmune neuroinflammation. Immunol Rev. (2014) 259:231–44. doi: 10.1111/imr.12169
12. Banica L, Besliu A, Pistol G, Stavaru C, Ionescu R, Forsea A-M, et al. Quantification and molecular characterization of regulatory T cells in connective tissue diseases. Autoimmunity (2009) 42:41–9. doi: 10.1080/08916930802282651
13. Baecher-Allan C, Brown JA, Freeman GJ, Hafler DA. CD4+CD25high regulatory cells in human peripheral blood. J Immunol. (2001) 167:1245–53. doi: 10.4049/jimmunol.167.3.1245
14. Ng WF, Duggan PJ, Ponchel F, Matarese G, Lombardi G, Edwards AD, et al. Human CD4(+)CD25(+) cells: a naturally occurring population of regulatory T cells. Blood (2001) 98:2736–44. doi: 10.1182/blood.V98.9.2736
15. Jonuleit H, Schmitt E, Stassen M, Tuettenberg A, Knop J, Enk AH. Identification and functional characterization of human CD4(+)CD25(+) T cells with regulatory properties isolated from peripheral blood. J Exp Med. (2001) 193:1285–94. doi: 10.1084/jem.193.11.1285
16. Sakaguchi S, Sakaguchi N, Asano M, Itoh M, Toda M. Immunologic self-tolerance maintained by activated T cells expressing IL-2 receptor alpha-chains (CD25). Breakdown of a single mechanism of self-tolerance causes various autoimmune diseases. J Immunol. (1995) 155:1151–64.
17. Fontenot JD, Gavin MA, Rudensky AY. Foxp3 programs the development and function of CD4+CD25+ regulatory T cells. Nat Immunol. (2003) 4:330–6. doi: 10.1038/ni904
18. Hori S, Nomura T, Sakaguchi S. Control of regulatory T cell development by the transcription factor Foxp3. Science (2003) 299:1057–61. doi: 10.1126/science.1079490
19. Khattri R, Cox T, Yasayko S-A, Ramsdell F. An essential role for Scurfin in CD4+CD25+ T regulatory cells. Nat Immunol. (2003) 4:337–42. doi: 10.1038/ni909
20. Roncador G, Brown PJ, Maestre L, Hue S, Martínez-Torrecuadrada JL, Ling K-L, et al. Analysis of FOXP3 protein expression in human CD4+CD25+ regulatory T cells at the single-cell level. Eur J Immunol. (2005) 35:1681–91. doi: 10.1002/eji.200526189
21. Zheng Y, Rudensky AY. Foxp3 in control of the regulatory T cell lineage. Nat Immunol. (2007) 8:457–62. doi: 10.1038/ni1455
22. Stephens GL, Andersson J, Shevach EM. Distinct subsets of FoxP3+ regulatory T cells participate in the control of immune responses. J Immunol. (2007) 178:6901–11. doi: 10.4049/jimmunol.178.11.6901
23. Gavin MA, Torgerson TR, Houston E, DeRoos P, Ho WY, Stray-Pedersen A, et al. Single-cell analysis of normal and FOXP3-mutant human T cells: FOXP3 expression without regulatory T cell development. Proc Natl Acad Sci USA. (2006) 103:6659–64. doi: 10.1073/pnas.0509484103
24. Morgan ME, van Bilsen JHM, Bakker AM, Heemskerk B, Schilham MW, Hartgers FC, et al. Expression of FOXP3 mRNA is not confined to CD4+CD25+ T regulatory cells in humans. Hum Immunol. (2005) 66:13–20. doi: 10.1016/j.humimm.2004.05.016
25. Liu W, Putnam AL, Xu-Yu Z, Szot GL, Lee MR, Zhu S, et al. CD127 expression inversely correlates with FoxP3 and suppressive function of human CD4+ T reg cells. J Exp Med. (2006) 203:1701–11. doi: 10.1084/jem.20060772
26. Seddiki N, Santner-Nanan B, Martinson J, Zaunders J, Sasson S, Landay A, et al. Expression of interleukin (IL)-2 and IL-7 receptors discriminates between human regulatory and activated T cells. J Exp Med. (2006) 203:1693–700. doi: 10.1084/jem.20060468
27. Franchimont D, Galon J, Vacchio MS, Fan S, Visconti R, Frucht DM, et al. Positive effects of glucocorticoids on T cell function by up-regulation of IL-7 receptor alpha. J Immunol. (2002) 168:2212–8. doi: 10.4049/jimmunol.168.5.2212
28. Schluns KS, Kieper WC, Jameson SC, Lefrançois L. Interleukin-7 mediates the homeostasis of naïve and memory CD8 T cells in vivo. Nat Immunol. (2000) 1:426–32. doi: 10.1038/80868
29. Klein S, Kretz CC, Krammer PH, Kuhn A. CD127(low/–) and FoxP3(+) expression levels characterize different regulatory T-cell populations in human peripheral blood. J Invest Dermatol. (2010) 130:492–9. doi: 10.1038/jid.2009.313
30. Wing K, Onishi Y, Prieto-Martin P, Yamaguchi T, Miyara M, Fehervari Z, et al. CTLA-4 control over Foxp3+ regulatory T cell function. Science (2008) 322:271–5. doi: 10.1126/science.1160062
31. Shimizu J, Yamazaki S, Takahashi T, Ishida Y, Sakaguchi S. Stimulation of CD25(+)CD4(+) regulatory T cells through GITR breaks immunological self-tolerance. Nat Immunol. (2002) 3:135–42. doi: 10.1038/ni759
32. Piconese S, Valzasina B, Colombo MP. OX40 triggering blocks suppression by regulatory T cells and facilitates tumor rejection. J Exp Med. (2008) 205:825–39. doi: 10.1084/jem.20071341
33. Miyara M, Yoshioka Y, Kitoh A, Shima T, Wing K, Niwa A, et al. Functional delineation and differentiation dynamics of human CD4+ T cells expressing the FoxP3 transcription factor. Immunity (2009) 30:899–911. doi: 10.1016/j.immuni.2009.03.019
34. Kleinewietfeld M, Starke M, Mitri DD, Borsellino G, Battistini L, Rötzschke O, et al. CD49d provides access to “untouched” human Foxp3+ Treg free of contaminating effector cells. Blood (2009) 113:827–36. doi: 10.1182/blood-2008-04-150524
35. Vignali DAA, Collison LW, Workman CJ. How regulatory T cells work. Nat Rev Immunol. (2008) 8:523–32. doi: 10.1038/nri2343
36. Miyara M, Sakaguchi S. Natural regulatory T cells: mechanisms of suppression. Trends Mol Med. (2007) 13:108–16. doi: 10.1016/j.molmed.2007.01.003
37. Shevach EM. Mechanisms of foxp3+ T regulatory cell-mediated suppression. Immunity (2009) 30:636–45. doi: 10.1016/j.immuni.2009.04.010
38. Grant CR, Liberal R, Mieli-Vergani G, Vergani D, Longhi MS. Regulatory T-cells in autoimmune diseases: challenges, controversies and–yet–unanswered questions. Autoimmun Rev. (2015) 14:105–16. doi: 10.1016/j.autrev.2014.10.012
39. Elhai M, Avouac J, Kahan A, Allanore Y. Systemic sclerosis: recent insights. Joint Bone Spine (2015) 82:148–53. doi: 10.1016/j.jbspin.2014.10.010
40. Elhai M, Meune C, Avouac J, Kahan A, Allanore Y. Trends in mortality in patients with systemic sclerosis over 40 years: a systematic review and meta-analysis of cohort studies. Rheumatol Oxf. (2012) 51:1017–26. doi: 10.1093/rheumatology/ker269
41. van den Hoogen F, Khanna D, Fransen J, Johnson SR, Baron M, Tyndall A, et al. 2013 classification criteria for systemic sclerosis: an American College of Rheumatology/European League against Rheumatism collaborative initiative. Arthritis Rheum. (2013) 65:2737–47. doi: 10.1002/art.38098
42. Varga J, Trojanowska M, Kuwana M. Pathogenesis of systemic sclerosis: recent insights of molecular and cellular mechanisms and therapeutic opportunities. J Scleroderma Relat Disord. (2017) 2:137–52. doi: 10.5301/jsrd.5000249
43. Prescott RJ, Freemont AJ, Jones CJ, Hoyland J, Fielding P. Sequential dermal microvascular and perivascular changes in the development of scleroderma. J Pathol. (1992) 166:255–63. doi: 10.1002/path.1711660307
44. Roumm AD, Whiteside TL, Medsger TA, Rodnan GP. Lymphocytes in the skin of patients with progressive systemic sclerosis. Quantification, subtyping, and clinical correlations. Arthritis Rheum. (1984) 27:645–53. doi: 10.1002/art.1780270607
45. Scharffetter K, Lankat-Buttgereit B, Krieg T. Localization of collagen mRNA in normal and scleroderma skin by in-situ hybridization. Eur J Clin Invest. (1988) 18:9–17. doi: 10.1111/j.1365-2362.1988.tb01158.x
46. Kähäri VM, Sandberg M, Kalimo H, Vuorio T, Vuorio E. Identification of fibroblasts responsible for increased collagen production in localized scleroderma by in situ hybridization. J Invest Dermatol. (1988) 90:664–70. doi: 10.1111/1523-1747.ep12560826
47. Sakkas LI, Xu B, Artlett CM, Lu S, Jimenez SA, Platsoucas CD. Oligoclonal T cell expansion in the skin of patients with systemic sclerosis. J Immunol. (2002) 168:3649–59. doi: 10.4049/jimmunol.168.7.3649
48. Kreuter A, Höxtermann S, Tigges C, Hahn SA, Altmeyer P, Gambichler T. Clonal T-cell populations are frequent in the skin and blood of patients with systemic sclerosis. Br J Dermatol. (2009) 161:785–90. doi: 10.1111/j.1365-2133.2009.09213.x
49. Arnett FC, Gourh P, Shete S, Ahn CW, Honey RE, Agarwal SK, et al. Major histocompatibility complex (MHC) class II alleles, haplotypes and epitopes which confer susceptibility or protection in systemic sclerosis: analyses in 1300 Caucasian, African-American and Hispanic cases and 1000 controls. Ann Rheum Dis. (2010) 69:822–7. doi: 10.1136/ard.2009.111906
50. Beretta L, Rueda B, Marchini M, Santaniello A, Simeón CP, Fonollosa V, et al. Analysis of Class II human leucocyte antigens in Italian and Spanish systemic sclerosis. Rheumatol Oxf Engl. (2012) 51:52–9. doi: 10.1093/rheumatology/ker335
51. Kuwana M, Inoko H, Kameda H, Nojima T, Sato S, Nakamura K, et al. Association of human leukocyte antigen class II genes with autoantibody profiles, but not with disease susceptibility in Japanese patients with systemic sclerosis. Intern Med. (1999) 38:336–44. doi: 10.2169/internalmedicine.38.336
52. Gilchrist FC, Bunn C, Foley PJ, Lympany PA, Black CM, Welsh KI, et al. Class II HLA associations with autoantibodies in scleroderma: a highly significant role for HLA-DP. Genes Immun. (2001) 2:76–81. doi: 10.1038/sj.gene.6363734
53. Truchetet M-E, Brembilla NC, Montanari E, Allanore Y, Chizzolini C. Increased frequency of circulating Th22 in addition to Th17 and Th2 lymphocytes in systemic sclerosis: association with interstitial lung disease. Arthritis Res Ther. (2011) 13:R166. doi: 10.1186/ar3486
54. Boin F, De Fanis U, Bartlett SJ, Wigley FM, Rosen A, Casolaro V. T cell polarization identifies distinct clinical phenotypes in scleroderma lung disease. Arthritis Rheum. (2008) 58:1165–74. doi: 10.1002/art.23406
55. Salmon-Ehr V, Serpier H, Nawrocki B, Gillery P, Clavel C, Kalis B, et al. Expression of interleukin-4 in scleroderma skin specimens and scleroderma fibroblast cultures. Potential role in fibrosis. Arch Dermatol. (1996) 132:802–6. doi: 10.1001/archderm.1996.03890310088013
56. Wynn TA. Fibrotic disease and the T(H)1/T(H)2 paradigm. Nat Rev Immunol. (2004) 4:583–94. doi: 10.1038/nri1412
57. Antiga E, Quaglino P, Bellandi S, Volpi W, Del Bianco E, Comessatti A, et al. Regulatory T cells in the skin lesions and blood of patients with systemic sclerosis and morphoea. Br J Dermatol. (2010) 162:1056–63. doi: 10.1111/j.1365-2133.2010.09633.x
58. Liu X, Gao N, Li M, Xu D, Hou Y, Wang Q, et al. Elevated levels of CD4(+)CD25(+)FoxP3(+) T cells in systemic sclerosis patients contribute to the secretion of IL-17 and immunosuppression dysfunction. PloS ONE (2013) 8:e64531. doi: 10.1371/journal.pone.0064531
59. Papp G, Horvath IF, Barath S, Gyimesi E, Sipka S, Szodoray P, et al. Altered T-cell and regulatory cell repertoire in patients with diffuse cutaneous systemic sclerosis. Scand J Rheumatol. (2011) 40:205–10. doi: 10.3109/03009742.2010.528021
60. Fenoglio D, Battaglia F, Parodi A, Stringara S, Negrini S, Panico N, et al. Alteration of Th17 and Treg cell subpopulations co-exist in patients affected with systemic sclerosis. Clin Immunol. (2011) 139:249–57. doi: 10.1016/j.clim.2011.01.013
61. Mathian A, Parizot C, Dorgham K, Trad S, Arnaud L, Larsen M, et al. Activated and resting regulatory T cell exhaustion concurs with high levels of interleukin-22 expression in systemic sclerosis lesions. Ann Rheum Dis. (2012) 71:1227–34. doi: 10.1136/annrheumdis-2011-200709
62. Cordiali-Fei P, Mussi A, D'Agosto G, Trento E, Bordignon V, Trincone S, et al. Assessment of T regulatory cells and expanded profiling of autoantibodies may offer novel biomarkers for the clinical management of systemic sclerosis and undifferentiated connective tissue disease. Clin Dev Immunol. (2013) 2013:390563. doi: 10.1155/2013/390563
63. Wang YY, Wang Q, Sun XH, Liu RZ, Shu Y, Kanekura T, et al. DNA hypermethylation of the forkhead box protein 3 (FOXP3) promoter in CD4+ T cells of patients with systemic sclerosis. Br J Dermatol. (2014) 171:39–47. doi: 10.1111/bjd.12913
64. Baraut J, Grigore EI, Jean-Louis F, Khelifa SH, Durand C, Verrecchia F, et al. Peripheral blood regulatory T cells in patients with diffuse systemic sclerosis (SSc) before and after autologous hematopoietic SCT: a pilot study. Bone Marrow Transplant. (2014) 49:349–54. doi: 10.1038/bmt.2013.202
65. Kataoka H, Yasuda S, Fukaya S, Oku K, Horita T, Atsumi T, et al. Decreased expression of Runx1 and lowered proportion of Foxp3+ CD25+ CD4+ regulatory T cells in systemic sclerosis. Mod Rheumatol. (2015) 25:90–5. doi: 10.3109/14397595.2014.899736
66. Klein S, Kretz CC, Ruland V, Stumpf C, Haust M, Hartschuh W, et al. Reduction of regulatory T cells in skin lesions but not in peripheral blood of patients with systemic scleroderma. Ann Rheum Dis. (2011) 70:1475–81. doi: 10.1136/ard.2009.116525
67. Krasimirova E, Velikova T, Ivanova-Todorova E, Tumangelova-Yuzeir K, Kalinova D, Boyadzhieva V, et al. Treg/Th17 cell balance and phytohaemagglutinin activation of T lymphocytes in peripheral blood of systemic sclerosis patients. World J Exp Med. (2017) 7:84–96. doi: 10.5493/wjem.v7.i3.84
68. Radstake TRDJ, van Bon L, Broen J, Wenink M, Santegoets K, Deng Y, et al. Increased frequency and compromised function of T regulatory cells in systemic sclerosis (SSc) is related to a diminished CD69 and TGFbeta expression. PloS ONE (2009) 4:e5981. doi: 10.1371/journal.pone.0005981
69. Slobodin G, Ahmad MS, Rosner I, Peri R, Rozenbaum M, Kessel A, et al. Regulatory T cells (CD4(+)CD25(bright)FoxP3(+)) expansion in systemic sclerosis correlates with disease activity and severity. Cell Immunol. (2010) 261:77–80. doi: 10.1016/j.cellimm.2009.12.009
70. Giovannetti A, Rosato E, Renzi C, Maselli A, Gambardella L, Giammarioli AM, et al. Analyses of T cell phenotype and function reveal an altered T cell homeostasis in systemic sclerosis. Correlations with disease severity and phenotypes. Clin Immunol. (2010) 137:122–33. doi: 10.1016/j.clim.2010.06.004
71. Rodríguez-Reyna TS, Furuzawa-Carballeda J, Cabiedes J, Fajardo-Hermosillo LD, Martínez-Reyes C, Díaz-Zamudio M, et al. Th17 peripheral cells are increased in diffuse cutaneous systemic sclerosis compared with limited illness: a cross-sectional study. Rheumatol Int. (2012) 32:2653–60. doi: 10.1007/s00296-011-2056-y
72. Jiang N, Li M, Zeng X. Correlation of Th17 cells and CD4+CD25+ regulatory T cells with clinical parameters in patients with systemic sclerosis. Chin Med J. (2014) 127:3557–61. doi: 10.3760/cma.j.issn.0366-6999.20141395
73. Ugor E, Simon D, Almanzar G, Pap R, Najbauer J, Németh P, et al. Increased proportions of functionally impaired regulatory T cell subsets in systemic sclerosis. Clin Immunol. (2017) 184:54–62. doi: 10.1016/j.clim.2017.05.013
74. Slobodin G, Rimar D. Regulatory T cells in systemic sclerosis: a comprehensive review. Clin Rev Allergy Immunol. (2017) 52:194–201. doi: 10.1007/s12016-016-8563-6
75. Yang X, Yang J, Xing X, Wan L, Li M. Increased frequency of Th17 cells in systemic sclerosis is related to disease activity and collagen overproduction. Arthritis Res Ther. (2014) 16:R4. doi: 10.1186/ar4430
76. Braitch M, Harikrishnan S, Robins RA, Nichols C, Fahey AJ, Showe L, et al. Glucocorticoids increase CD4CD25 cell percentage and Foxp3 expression in patients with multiple sclerosis. Acta Neurol Scand. (2009) 119:239–45. doi: 10.1111/j.1600-0404.2008.01090.x
77. Antiga E, Fabbri P, Caproni M. Immunosuppressive therapy may affect the number of circulating regulatory cells in systemic sclerosis: pay attention to the patient selection criteria. Cell Immunol. (2010) 264:186. doi: 10.1016/j.cellimm.2010.06.007
78. Austin ED, Rock MT, Mosse CA, Vnencak-Jones CL, Yoder SM, Robbins IM, et al. T lymphocyte subset abnormalities in the blood and lung in pulmonary arterial hypertension. Respir Med. (2010) 104:454–62. doi: 10.1016/j.rmed.2009.10.004
79. Ulrich S, Nicolls MR, Taraseviciene L, Speich R, Voelkel N. Increased regulatory and decreased CD8+ cytotoxic T cells in the blood of patients with idiopathic pulmonary arterial hypertension. Respir Int Rev Thorac Dis. (2008) 75:272–80. doi: 10.1159/000111548
80. Huertas A, Phan C, Bordenave J, Tu L, Thuillet R, Le Hiress M, et al. Regulatory T cell dysfunction in idiopathic, heritable and connective tissue-associated pulmonary arterial hypertension. Chest (2016) 149:1482–93. doi: 10.1016/j.chest.2016.01.004
81. Shu Y, Hu Q, Long H, Chang C, Lu Q, Xiao R. Epigenetic variability of CD4+CD25+ Tregs contributes to the pathogenesis of autoimmune diseases. Clin Rev Allergy Immunol. (2017) 52:260–72. doi: 10.1007/s12016-016-8590-3
82. D'Amico F, Fiorito G, Skarmoutsou E, Granata M, Rossi GA, Trovato C, et al. FOXP3, ICOS and ICOSL gene polymorphisms in systemic sclerosis: FOXP3 rs2294020 is associated with disease progression in a female Italian population. Immunobiology (2018) 223:112–7. doi: 10.1016/j.imbio.2017.10.004
83. Broen JCA, Wolvers-Tettero ILM, Geurts-van Bon L, Vonk MC, Coenen MJH, Lafyatis R, et al. Skewed X chromosomal inactivation impacts T regulatory cell function in systemic sclerosis. Ann Rheum Dis. (2010) 69:2213–6. doi: 10.1136/ard.2010.129999
84. Rimar D, Nov Y, Rosner I, Slobodin G, Rozenbaum M, Halasz K, et al. Semaphorin 3A: an immunoregulator in systemic sclerosis. Rheumatol Int. (2015) 35:1625–30. doi: 10.1007/s00296-015-3269-2
85. Kurasawa K, Hirose K, Sano H, Endo H, Shinkai H, Nawata Y, et al. Increased interleukin-17 production in patients with systemic sclerosis. Arthritis Rheum. (2000) 43:2455–63. doi: 10.1002/1529-0131(200011)43:11 < 2455::AID-ANR12>3.0.CO;2-K
86. Koenen HJPM, Smeets RL, Vink PM, van Rijssen E, Boots AMH, Joosten I. Human CD25highFoxp3pos regulatory T cells differentiate into IL-17-producing cells. Blood (2008) 112:2340–52. doi: 10.1182/blood-2008-01-133967
87. Osorio F, LeibundGut-Landmann S, Lochner M, Lahl K, Sparwasser T, Eberl G, et al. DC activated via dectin-1 convert Treg into IL-17 producers. Eur J Immunol. (2008) 38:3274–81. doi: 10.1002/eji.200838950
88. Yang XO, Nurieva R, Martinez GJ, Kang HS, Chung Y, Pappu BP, et al. Molecular antagonism and plasticity of regulatory and inflammatory T cell programs. Immunity (2008) 29:44–56. doi: 10.1016/j.immuni.2008.05.007
89. Beriou G, Costantino CM, Ashley CW, Yang L, Kuchroo VK, Baecher-Allan C, et al. IL-17-producing human peripheral regulatory T cells retain suppressive function. Blood (2009) 113:4240–9. doi: 10.1182/blood-2008-10-183251
90. Bending D, De la Peña H, Veldhoen M, Phillips JM, Uyttenhove C, Stockinger B, et al. Highly purified Th17 cells from BDC2.5NOD mice convert into Th1-like cells in NOD/SCID recipient mice. J Clin Invest. (2009) 119:565–72. doi: 10.1172/JCI37865
91. Weaver CT, Hatton RD. Interplay between the TH17 and TReg cell lineages: a (co-)evolutionary perspective. Nat Rev Immunol. (2009) 9:883–9. doi: 10.1038/nri2660
92. Voo KS, Wang Y-H, Santori FR, Boggiano C, Wang Y-H, Arima K, et al. Identification of IL-17-producing FOXP3+ regulatory T cells in humans. Proc Natl Acad Sci USA. (2009) 106:4793–8. doi: 10.1073/pnas.0900408106
93. MacDonald KG, Dawson NAJ, Huang Q, Dunne JV, Levings MK, Broady R. Regulatory T cells produce profibrotic cytokines in the skin of patients with systemic sclerosis. J Allergy Clin Immunol. (2015) 135:946–.e9. doi: 10.1016/j.jaci.2014.12.1932
94. Dudda JC, Perdue N, Bachtanian E, Campbell DJ. Foxp3+ regulatory T cells maintain immune homeostasis in the skin. J Exp Med. (2008) 205:1559–65. doi: 10.1084/jem.20072594
95. Soare A, Ramming A, Avouac J, Distler JHW. Updates on animal models of systemic sclerosis. J Scleroderma Relat Disord. (2016) 1:266–76. doi: 10.5301/jsrd.5000220
96. Yoshizaki A, Yanaba K, Ogawa A, Asano Y, Kadono T, Sato S. Immunization with DNA topoisomerase I and Freund's complete adjuvant induces skin and lung fibrosis and autoimmunity via interleukin-6 signaling. Arthritis Rheum. (2011) 63:3575–85. doi: 10.1002/art.30539
97. Birjandi SZ, Palchevskiy V, Xue YY, Nunez S, Kern R, Weigt SS, et al. CD4(+)CD25(hi)Foxp3(+) cells exacerbate bleomycin-induced pulmonary fibrosis. Am J Pathol. (2016) 186:2008–20. doi: 10.1016/j.ajpath.2016.03.020
98. Kamio K, Azuma A, Matsuda K, Usuki J, Inomata M, Morinaga A, et al. Resolution of bleomycin-induced murine pulmonary fibrosis via a splenic lymphocyte subpopulation. Respir Res. (2018) 19:71. doi: 10.1186/s12931-018-0783-2
99. Boveda-Ruiz D, D'Alessandro-Gabazza CN, Toda M, Takagi T, Naito M, Matsushima Y, et al. Differential role of regulatory T cells in early and late stages of pulmonary fibrosis. Immunobiology (2013) 218:245–54. doi: 10.1016/j.imbio.2012.05.020
100. Chae W-J, Bothwell ALM. Therapeutic potential of gene-modified regulatory T cells: from bench to bedside. Front Immunol. (2018) 9:303. doi: 10.3389/fimmu.2018.00303
101. Danikowski KM, Jayaraman S, Prabhakar BS. Regulatory T cells in multiple sclerosis and myasthenia gravis. J Neuroinflamm. (2017) 14:117. doi: 10.1186/s12974-017-0892-8
102. Jeffery HC, Braitch MK, Brown S, Oo YH. Clinical potential of regulatory T cell therapy in liver diseases: an overview and current perspectives. Front Immunol. (2016) 7:334. doi: 10.3389/fimmu.2016.00334
103. Giang S, La Cava A. Regulatory T cells in SLE: biology and use in treatment. Curr Rheumatol Rep. (2016) 18:67. doi: 10.1007/s11926-016-0616-6
104. Kuhn C, Weiner HL. Therapeutic anti-CD3 monoclonal antibodies: from bench to bedside. Immunotherapy (2016) 8:889–906. doi: 10.2217/imt-2016-0049
105. Spence A, Klementowicz JE, Bluestone JA, Tang Q. Targeting Treg signaling for the treatment of autoimmune diseases. Curr Opin Immunol. (2015) 37:11–20. doi: 10.1016/j.coi.2015.09.002
106. Spence A, Tang Q. Restoring regulatory T cells in type 1 diabetes. Curr Diab Rep. (2016) 16:110. doi: 10.1007/s11892-016-0807-6
107. Matsuoka K, Koreth J, Kim HT, Bascug G, McDonough S, Kawano Y, et al. Low-dose interleukin-2 therapy restores regulatory T cell homeostasis in patients with chronic graft-versus-host disease. Sci Transl Med. (2013) 5:179ra43. doi: 10.1126/scitranslmed.3005265
108. Castela E, Le Duff F, Butori C, Ticchioni M, Hofman P, Bahadoran P, et al. Effects of low-dose recombinant interleukin 2 to promote T-regulatory cells in alopecia areata. JAMA Dermatol. (2014) 150:748–51. doi: 10.1001/jamadermatol.2014.504
109. Saadoun D, Rosenzwajg M, Joly F, Six A, Carrat F, Thibault V, et al. Regulatory T-cell responses to low-dose interleukin-2 in HCV-induced vasculitis. N Engl J Med. (2011) 365:2067–77. doi: 10.1056/NEJMoa1105143
110. Heiler S, Lötscher J, Kreuzaler M, Rolink J, Rolink A. Prophylactic and therapeutic effects of interleukin-2 (IL-2)/Anti-IL-2 complexes in systemic lupus erythematosus-like chronic graft-versus-host disease. Front Immunol. (2018) 9:656. doi: 10.3389/fimmu.2018.00656
111. von Spee-Mayer C, Siegert E, Abdirama D, Rose A, Klaus A, Alexander T, et al. Low-dose interleukin-2 selectively corrects regulatory T cell defects in patients with systemic lupus erythematosus. Ann Rheum Dis. (2016) 75:1407–15. doi: 10.1136/annrheumdis-2015-207776
112. Becker MO, Brückner C, Scherer HU, Wassermann N, Humrich JY, Hanitsch LG, et al. The monoclonal anti-CD25 antibody basiliximab for the treatment of progressive systemic sclerosis: an open-label study. Ann Rheum Dis. (2011) 70:1340–1. doi: 10.1136/ard.2010.137935
113. van Laar JM, Farge D, Sont JK, Naraghi K, Marjanovic Z, Larghero J, et al. Autologous hematopoietic stem cell transplantation vs intravenous pulse cyclophosphamide in diffuse cutaneous systemic sclerosis: a randomized clinical trial. JAMA (2014) 311:2490–8. doi: 10.1001/jama.2014.6368
114. Sullivan KM, Goldmuntz EA, Keyes-Elstein L, McSweeney PA, Pinckney A, Welch B, et al. Myeloablative autologous stem-cell transplantation for severe scleroderma. N Engl J Med. (2018) 378:35–47. doi: 10.1056/NEJMoa1703327
115. Tsukamoto H, Nagafuji K, Horiuchi T, Mitoma H, Niiro H, Arinobu Y, et al. Analysis of immune reconstitution after autologous CD34+ stem/progenitor cell transplantation for systemic sclerosis: predominant reconstitution of Th1 CD4+ T cells. Rheumatol Oxf Engl. (2011) 50:944–52. doi: 10.1093/rheumatology/keq414
Keywords: systemic sclerosis, regulatory T cells, immune tolerance, auto-immunity, thymus
Citation: Frantz C, Auffray C, Avouac J and Allanore Y (2018) Regulatory T Cells in Systemic Sclerosis. Front. Immunol. 9:2356. doi: 10.3389/fimmu.2018.02356
Received: 18 June 2018; Accepted: 24 September 2018;
Published: 15 October 2018.
Edited by:
Qizhi Tang, University of California, San Francisco, United StatesReviewed by:
Dennis O. Adeegbe, Moffitt Cancer Center, United StatesShomyseh Sanjabi, University of California, San Francisco, United States
Kathryn S. Torok, University of Pittsburgh, United States
Tamiko Robin Katsumoto, Stanford University, United States
Copyright © 2018 Frantz, Auffray, Avouac and Allanore. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yannick Allanore, yannick.allanore@aphp.fr