 Irene Maeve Rea1,2,3*
Irene Maeve Rea1,2,3* David S. Gibson2
David S. Gibson2 Victoria McGilligan2
Victoria McGilligan2 Susan E. McNerlan4
Susan E. McNerlan4 H. Denis Alexander2
H. Denis Alexander2 Owen A. Ross5,6,7
Owen A. Ross5,6,7
- 1School of Medicine, Dentistry and Biomedical Science, Queens University Belfast, Belfast, United Kingdom
- 2Northern Ireland Centre for Stratified Medicine, Biomedical Sciences Research Institute, University of Ulster, C-TRIC Building, Altnagelvin Area Hospital, Londonderry, United Kingdom
- 3Care of Elderly Medicine, Belfast Health and Social Care Trust, Belfast, United Kingdom
- 4Regional Genetics Service, Belfast Health and Social Care Trust, Belfast, United Kingdom
- 5Department of Neuroscience, Mayo Clinic, Jacksonville, FL, United States
- 6Department of Clinical Genomics, Mayo Clinic, Jacksonville, FL, United States
- 7School of Medicine and Medical Science, University College Dublin, Dublin, Ireland
Cytokine dysregulation is believed to play a key role in the remodeling of the immune system at older age, with evidence pointing to an inability to fine-control systemic inflammation, which seems to be a marker of unsuccessful aging. This reshaping of cytokine expression pattern, with a progressive tendency toward a pro-inflammatory phenotype has been called “inflamm-aging.” Despite research there is no clear understanding about the causes of “inflamm-aging” that underpin most major age-related diseases, including atherosclerosis, diabetes, Alzheimer’s disease, rheumatoid arthritis, cancer, and aging itself. While inflammation is part of the normal repair response for healing, and essential in keeping us safe from bacterial and viral infections and noxious environmental agents, not all inflammation is good. When inflammation becomes prolonged and persists, it can become damaging and destructive. Several common molecular pathways have been identified that are associated with both aging and low-grade inflammation. The age-related change in redox balance, the increase in age-related senescent cells, the senescence-associated secretory phenotype (SASP) and the decline in effective autophagy that can trigger the inflammasome, suggest that it may be possible to delay age-related diseases and aging itself by suppressing pro-inflammatory molecular mechanisms or improving the timely resolution of inflammation. Conversely there may be learning from molecular or genetic pathways from long-lived cohorts who exemplify good quality aging. Here, we will discuss some of the current ideas and highlight molecular pathways that appear to contribute to the immune imbalance and the cytokine dysregulation, which is associated with “inflammageing” or parainflammation. Evidence of these findings will be drawn from research in cardiovascular disease, cancer, neurological inflammation and rheumatoid arthritis.
Introduction
The inflammatory response must be tightly regulated to ensure effective immune protection. It is a dynamic network that is continuously remodeling throughout each person’s life as a result of the interaction between our genes, lifestyles, and environments (1–3). Infections and tissue damage from the external environment and our personal internal response to stress can act as triggers to initiate the inflammatory defense response. While inflammation is part of the normal repair response for healing, and essential in keeping us safe from bacterial and viral infections and noxious environmental agents, not all inflammation is good. When inflammation becomes prolonged and persists, it can become damaging and destructive (4). It is essential that inflammation is tailored to the initiating stress and resolves in a timely and controlled way, to avoid pathology associated with chronicity.
The cytokine network is a highly complex system of immune molecular messengers, with multiple layers of activation and control mediated through soluble receptors, receptor antagonists, diverse serum mediators, as well as gene polymorphisms (5). Proteomic methods measuring cytokine production and expression have demonstrated further layers of complexity and control in cytokine production and expression involving long coding RNAs, siRNAs, and miRNAs, which make for challenging interpretation of cytokine production and control in the inflammatory process (6). Many cytokines are able to act in more than one-way or paradoxically at different times and many act in feedback loops with the ability to auto-control their own production (7). Cytokine expression is also influenced by local cellular microenvironments, suggesting that multiple pathways exist to achieve homeostatic immunologic control and effectiveness, or to conversely accentuate chronic immune activation. However, what seems clear is that mirroring other body systems, the homeostatic control, titration, and modulation of immune responsiveness becomes more fragile and less tightly focused with increasing age. This loosening of the cytokine balance between the pro-inflammatory and anti-inflammatory control or resolving mechanisms, or inflamm-aging (8, 9), is a characteristic feature of both aging and aging-related diseases. This kind of inflammation is similar to that originally described as “parainflammation” by Medzhitov (10).
Today there is increasing recognition that inflammation is a common molecular pathway that underlies in part, the pathogenesis of diverse human diseases ranging from infection, to immune-mediated disorders, cardiovascular pathology, diabetes, metabolic syndrome, neurodegeneration, and cancer, to aging itself (4, 11, 12). Although there is no exact understanding about the causes of “inflamm-aging”, a common finding seems to involve a dysregulation of the cytokine network and its homeostasis. Several common molecular pathways have been identified that seem to be associated with both aging and low-grade inflammation. Excess oxidative stress and DNA damage trigger the inflammasome, stimulating NF-κB and the IL-1β-mediated inflammatory cascade. Autophagy, the cell machinery process that removes damaged proteins and large aggregates, is also slowed up at older age and in age-related disease, causing damaged material to accumulate and reduce cellular efficiency. Senescent cells increase with age and in age-related diseases, and the associated secretome or senescence-associated secretory phenotype (SASP) produces a self-perpetuating intracellular signaling loop and inflammatory cascade involving the NF-κB, IL-1α, TGF-β, IL-6 pathway that participates in the pro-inflammatory milieu. The molecular processes that damp down inflammation include the resolvin family of bioactive molecules, which have been much less evaluated in aging or age-related disease, but are important participants in effective and timely inflammation resolution.
Here, we will discuss some of the current ideas and highlight molecular pathways that appear to contribute to the immune imbalance and the cytokine dysregulation, which is associated with “inflamm-aging” or parainflammation. Evidence of these findings will be drawn from research in several age-related diseases, including cardiovascular and neurodegenerative disease, rheumatoid arthritis (RA), and cancers.
The Inflammation Pathway to Resolution
Inflammation is classically induced when innate cells detect infection or tissue injury. The pattern-recognition receptors (PRRs) on immune cells sense “danger” from protein-associated molecular patterns (PAMPs) associated with pathogens, or from danger-associated molecular patterns (DAMPs) triggered by a wide range of host-derived endogenous stress signals. DAMPs are molecules, such as ATP, the cytokine IL-1α, uric acid, and some cytoplasmic and nuclear proteins, which are released from damaged cells during necrosis and contribute to sterile inflammation (Figure 1). There have been suggestions that the extended IL-1 cytokine family (IL-1α, IL-1β, IL-18, IL-33, IL-36α, IL-36β, and IL-36γ) might also act as DAMPs and stimulate necrosis-initiated sterile inflammation, as well as amplify inflammation in response to infection-associated tissue injury (13).
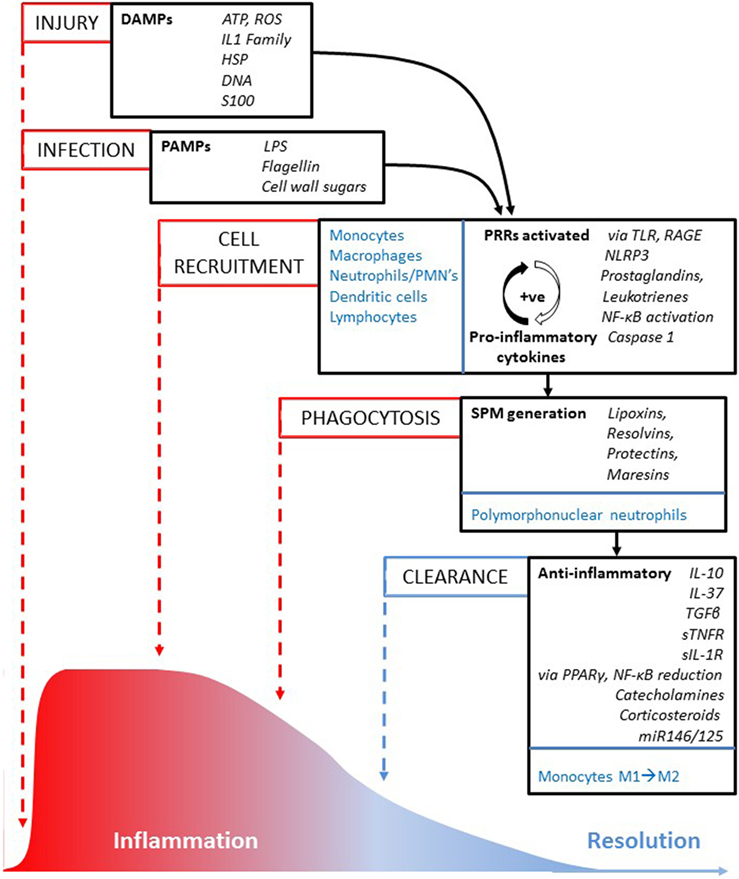
Figure 1. Inflammation pathway to resolution. An illustration of the sequence of key processes (in capitalized text), cells and molecules involved in reaction to injury or infection, and how the inflammatory episode is resolved over time (from left to right). Cells from the innate and adaptive immune system that are involved in cell recruitment, phagocytosis, and clearance processes are highlighted in blue text; key molecules are in italic text.
Members of the toll-like receptor (TLR) family are the major PRRs. They are expressed on monocytes, macrophages, neutrophils, and dendritic cells, and on some lymphocytes and they respond rapidly to the “danger” response. The cyclooxygenase (COX) and 5-lipoxygenase (5-LOX) pathways of arachidonic acid (AA) metabolism (14, 15) produce highly pro-inflammatory lipid mediators responsible for the classical signs of inflammation—redness, heat, pain, swelling, and loss of function, with the aim of removing the injurious and noxious stimuli. A third pathway involves the cytochrome 450 pathway of AA metabolism and P450 epoxygenases and hydroxylases that produce both vasoconstrictor and vasodilatory effects in blood vessels and other tissues (Figure 2). The reactive biolipid molecules synthesized from AA are; the prostanoids—prostaglandins (PGs), prostacyclins, and thromboxanes produced by the action of COX 1 and 2 (COX 1 and 2); the leukotrienes (LTs), hydroxyeicosatetraenoids (HETEs), and lipoxins (LXs) produced by the action of the 5-, 12-, and 15-lypooxygenase (5/12/15-LOX) enzymes and; the P450 epoxygenase generates HETEs and depoxyeicosatrienoids (epoxides) (16). PGs act to amplify the inflammatory response through enhancing the inflammatory cytokine cascade, upregulating the innate response to DAMPs and PAMPs, activating subsets of T helper cells, recruiting macrophages associated with chronic inflammation, and increasing cytokine expression from cytokine inflammatory genes. Additional factors, such as histamine, pro-inflammatory cytokines, and chemokines amplify the response further and make the vascular endothelium increasingly leaky. The increase in vascular permeability combined with the expression of cellular adhesion molecules (i.e., selectins and integrins) allows neutrophils, the first responders, to transmigrate across post-capillary venules to the sites of injury or microbial invasion. Together this increases polymorphonuclear (PMN) neutrophil chemotaxis and allows PMNs to transmigrate along chemotactic gradients in order to maximize phagocytosis and killing of pathogens, and deal with the “danger” signal effectively.
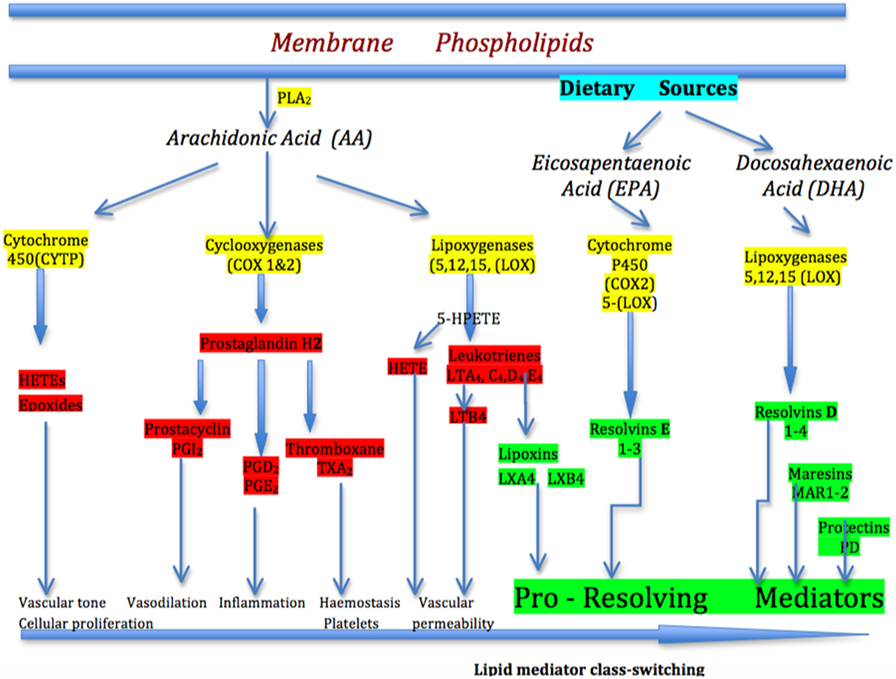
Figure 2. The arachidonic acid (AA) pathway of inflammation mediators. In the simplified pathway for the eicosanoid metabolic pathway, AA is released from membrane stores by phospholipase 2 (PLA2). AA is metabolized to biological mediators by three enzymatic pathways: cyclooxygenase, lipoxygenase, and cytochrome P450. Each pathway contains enzyme-specific steps that result in a wide variety of bioactive compounds that drive the pro-inflammatory (prostaglandins) response. After lipid mediator class-switching at the height of inflammation, the pro-resolving mediators-lipoxins begin to drive inflammation resolution. Eicosapentaenoic acid and docosahexaenoic acid-derived from dietary sources produce the E-series of resolvins and D-series of resolvins, maresins, and protectins, respectively, which are important pro-resolving mediators in progressing the resolution of inflammation.
As the acute inflammatory cascade develops to manage the “danger” signal, it is essential that a controlled resolution commences, so that immune homeostasis returns in an organized manner. If the inflammatory response does not shut down in a timely way, the inflammation cascade becomes chronic and smoldering. Lipid mediators derived from polyunsaturated fatty acids are now recognized to orchestrate the resolution of inflammation (17). At the peak of inflammation, the eicosanoids that initiated the inflammation undergo a class-switch so that they become the molecules that activate resolution, demonstrable through the clinical signs of removal of symptoms, relief of pain, restoration of function, regeneration of damaged tissues, and return to health. The so-called specialized pro-resolving mediators (SPMs) are key to resolving inflammation and include lipoxins derived from the 5-LOX arm of the AA pathway; the E-group of resolvins derived from dietary-derived eicosapentaenoic acid (EPA); the D-group of resolvins from dietary–derived docosahexaenoic acid (DHA); and protectins (PD), and maresins (MaR) (17–19) (Figure 2). The lipid class-switch starts early in inflammation and is initiated by lipoxins LXA4 and LXB4, and considered to be produced by platelets when they begin to aggregate with PMNs at the sites of inflammation (18).
After class-switching of the lipid molecules has occurred, SPMs are produced. Pro-resolving monocyte-derived macrophages begin to clear PMNs from the site of injury by a process called efferocytosis that removes apoptotic neutrophils, microbes, and necrotic debris. As resolution progresses, monocytes and macrophages, change from a pro-inflammatory (M1) to a pro-resolving phenotype (M2) by genetic and epigenetic reprogramming (20–22). Recent investigations suggest that SPMs, particularly the D-series resolvins (resolving D1 and resolving D2) and MaR 1 modulate adaptive immune responses in human peripheral blood lymphocytes. These lipid mediators reduce cytokine production by activated CD8+ T cells and CD4+ T helper 1 (TH1) and TH 17 cells, but do no modulate T cell inhibitory receptors or reduce their ability to proliferate (23, 24). Other reports show an increase in plasma cell differentiation and antibody production that supports the involvement of SPMs in the humoral response during late stages of inflammation and pathogen clearance (25). The anti-inflammatory cytokines interleukin 10 (IL-10), and IL-37 a member of the IL-1 family, together with TGF-β that is released from monocytes and platelets, are important contributors to damping down the inflammation. The soluble receptors, TNFR and IL-1 receptor (IL-1R) also limit inflammation in acting as decoy receptors, by binding to and neutralizing their respective cytokines, and inhibiting the biological activity. Additional anti-inflammatory mechanisms, include stress hormones, particularly corticosteroids and catecholamines and negative regulators, such as microRNAs—MiR-146 and MiR-125 (26).
The local environment and context also play an important role in the production and function of SPMs, which have both autocrine and paracrine actions. Inflammation resolution is likely to depend on prompt class-switching to pro-resolving lipid mediators, effective apoptosis, and efferocytic clearance of inflammatory cells and debris, timely damping down of pro-inflammatory signals and integrated repair of collateral damage. An imbalance between pro-inflammatory and pro-resolving mediators has been linked to a number of chronic inflammatory diseases (27).
In normal inflammation SPMs do not compromise host immune competence with examples of pro-resolving mediators increasing survival from infections in mouse models (28, 29). The common mechanism by which this occurs appears to be through suppression of the NF-κB activation in a partly PPAR-γ-dependent manner, with associated downstream signaling and alteration in transcriptomics pathways (30, 31). A maresin mediator has been shown to have potent anti-inflammatory and pro-resolving actions in a model of colitis, and attenuated inflammation in vascular smooth muscle and endothelial cells (32, 33). In human studies, the role of SPMs are being explored in chronic inflammatory diseases, such as RA (34), atherosclerosis (27), and cancer (35). In Alzheimer’s disease, several SPMs promoted neuronal survival and β-amyloid uptake by microglia in “in vitro” models in Alzheimer’s disease (36, 37). However, little is known about the pro-resolving mediators in aging itself. Studies are needed to assess whether pro-resolving molecules, such as E and D-resolvins, and maresins decrease or are less effective in damping down inflammation with increasing age and whether they could contribute to the pro-inflammatory phenotype associated with aging. Already synthetic analogs are in process of development, and so the design of pharmacological mimetics of naturally occurring pro-resolving mediators and their receptors offers new potential targets for drug design and the opportunity to investigate the underpinning molecular mechanisms of inflammation resolution.
Could life-style factors play a role in the epidemic of non-communicable and age-related diseases and the associated pro-inflammatory phenotype? Evidence exists that suggests that the Mediterranean diet which includes olive oil and some omega-3 lipids, can ameliorate RA (38), may give some protection from atrial fibrillation and myocardial infarction (MI) (39), and improves diabetic control (40). Research has also demonstrated a protective role of the Mediterranean diet in gene/Mediterranean diet interactions for the risk TT allele of the TCF7L2-rs7903146 gene in stroke risk and mortality (41, 42). Improving knowledge about how inflammation shuts down in a timely way is crucial to the understanding of how chronic inflammation contributes to aging and age-related diseases. Further studies are likely to be needed to advise if dietary modifications with omega-3 lipids or whether synthetic resolving mimetics are part of the answer.
Triggers of the Inflammation Pathway
Several common molecular pathways have been identified that seem to be associated with both aging and low-grade inflammation. These pathways trigger the inflammasome, stimulating NF-κB, and the IL-1β-mediated inflammatory cascade.
Age-Related Redox Imbalance
A redox imbalance has long been associated with aging and led to the development of the redox stress hypothesis of aging (43). Redox stress is caused by an imbalance between unregulated and overproduced reactive oxygen species (ROS) that are produced secondary to mitochondrial energy production, active immunological phagocytic processes, and the prostaglandin pathway through COX enzyme production. While ROS are important molecules regulating numerous physiological and pathological processes in the cell, there is now clear evidence that overproduction of ROS is involved in the development of a number of diseases, such as Alzheimer’s disease, rheumatoid, and cardiovascular diseases. Increasing evidence supports the notion that low concentrations of ROS or “primary ROS” are involved in well controlled processes (44), where their effect on reactive target molecules can be reversible, suggesting that “primary” ROS acts as an important intracellular signaling molecule (45). In contrast, the very active OH ROS is less effectively controlled and forms the main damaging type of ROS that is able to react with many macromolecules, such as lipids, proteins, and nucleic acids. This results in DNA oxidation and cell membrane damage, which contributes to the burden of damaged molecules related to aging and age-related diseases.
Mitochondrial ROS
Mitochondria are highly efficient producers of energy, but in doing so they produce ROS. It is estimated that about 90% of intracellular ROS is generated in the mitochondria through the mitochondrial transport chain. The chain of electron flow is considered to leak prematurely between complexes 1, 11, and 111 leading to the formation of damaging oxidants like . This ROS has been considered to cause damaging mutations in the mitochondrial genes with increasing age (43). With increasing age, mitochondrial function becomes sluggish and this compromises energy production, which in turn further contributes to mitochondrial dysfunction (46). A vicious cycle develops with age-reduced physical activity producing muscles that become weaker, are infiltrated with fat cells, and show less efficient mitochondria energy production (47). Ischemia and apoptosis can trigger , and mitochondria themselves can be damaged by ROS production. Mitophagy, the removal of damaged mitochondria is also reduced as age increases (48). A reduced age-related capacity of the body’s anti-oxidative defense systems to mop up free radicals also plays an important role in maintaining the inflammatory background of chronic inflammation (49).
The Nicotamide Adenine Dinucleotide Phosphate (NADPH) Pathway of ROS
One of the other main producers of ROS is the specialized enzyme group of the NADPH oxidases of the NOX family—(NOX1, NOX2, NOX3 NOX4, NOX5, DUOX1, and DUOX2). The NOX family or NADPH oxidases’ generate or H2O2 radicals by transferring electrons from cytoplasmic NADPH or the “NOX” catalytic subunit to molecular oxygen (50). The ROS produced by these enzymes has an essential function in neutrophils and macrophages as a mechanism for effective bacterial killing and host defense (51, 52). When the phagocytes sense an endogenous or exogenous danger signal, the NADPH-oxidase unit translocates to fuse with the plasma membrane to form the phagosome. This generates large amounts of highly reactive ROS called the phagocytic burst that is very effective in killing microbes, though phagosomal pH and ion concentration are also likely to be contributors.
Although NOX family of isoenzymes was initially associated with the ROS produced in phagocytes, other members of the NOX family are now known to be involved in a wide range of regulatory functions in many tissues and seem likely to play a role in aging and age-related diseases. Studies in the human vascular system suggest that NOX1, NOX2, and NOX5 promote endothelial dysfunction, inflammation, and apoptosis in the vessel walls, whereas NOX4 by contrast is vasoprotective, by increasing nitric oxide bioavailability (53). NOX enzymes, therefore, appear to play a role in vascular pathology as well as in the maintenance of normal physiological vascular function. Activation of NOX2 and NOX4 occurs in humans with atrial fibrillation and inhibition of NOX by angiotension converting enzyme inhibitor drugs or statins has proved helpful in preventing post-operative atrial fibrillation (54).
COX Pathways of ROS
The biolipids are highly reactive substances that contribute to both inflammation and healing and their pathways produce and use ROS signaling. The reaction that converts AA through COX2 into prostaglandin H2 (PGH2) by a two-stage free radical mechanism (55) involves superoxide and can contribute to cellular oxidative stress as well as signaling. Other enzymes that generate ROS during AA metabolism include the arachidonic 12-lipoxygenase (LOX-12 or ALOX12) and arachidonic 5-lipoxygenase (LOX5 or ALOX5), both of which also activate and induce NADPH-oxidases (56).
While mitochondrial ROS are traditionally seen as the main source of intracellular ROS and, therefore, major mediators of ROS-induced damage, the relative contribution of mitochondrial and non-mitochondrial sources of ROS to induction of cellular senescence remain unclear. Both mitochondrial ROS and NADPH-produced ROS appear to be able to cross signal between each other and mitochondria have significant antioxidant capacity, which may act as a cellular redox buffer for NADPH-produced ROS, suggesting there is tight control and integration of ROS signaling within the cell.
The cellular systems that protect against ROS, include the anti-oxidative defense enzymes, superoxidase dismutase, glutathione peroxidase, and catalase (57), oxidant scavengers (vitamin E, vitamin C, carotenoids, uric acid, and polyphenols), and mechanisms to repair oxidant damage to lipids, proteins, or DNA. Despite these protective mechanisms, uncontrolled ROS can overwhelm the antioxidant capacity of the cell causing mitochondrial dysfunction (49). Increased ROS production from the various cellular sources stimulates intracellular danger-sensing multi-protein platforms called inflammasomes (58–60). Through the inflammasome, the ROS activates NF-κB which sets in motion the transcription of a cascade of pro-inflammatory cytokines—tumor necrosis factor-alpha (TNF-α), IL-1β, IL-2, and IL-6, chemokines—IL-8 and RANTES, and adhesion molecules, such as ICAM-1, VCAM, and E-selectin, that are central mediators in the inflammatory response.
Autophagy Slowing and Aging
Approximately a third of all newly synthesized proteins are formed in the endoplasmic reticulum (ER), where they are folded, modified, sorted, and transported to sites where they perform specialized roles. Stressors, such as low glucose as in fasting, alterations in calcium levels, low oxygen states, viruses, cytokines, and nutrient excess or deficiency can trigger the autophagy pathway with the aim of returning normal homeostasis to the cell.
Autophagy is a cellular process whereby cellular waste, such as modified proteins, protein aggregates, and damaged organelles are removed from the cell. It is a tightly controlled process that plays a role in growth and development and maintains a balance between the synthesis, degradation, and subsequent recycling of cellular products. Autophagy can be considered a protein and organelle quality control mechanism that maintains normal cellular homeostasis.
Two major pathways degrade cellular proteins. The ubiquitin-proteasome system (UPS) degrades 80–90% of denatured and damaged proteins. In the ATP-dependent UPS, damaged or misfolded proteins are tagged with a small protein called ubiquitin. Three different sets of enzymes—E1, E2, and E3, identify and categorize proteins in order to link ubiquitin or ubiquitin complexes to the damaged proteins. The ubiquitin-protein complexes pass through the proteasome, where they are degraded and discharged as free amino acids into the cytoplasm (Figure 3A).
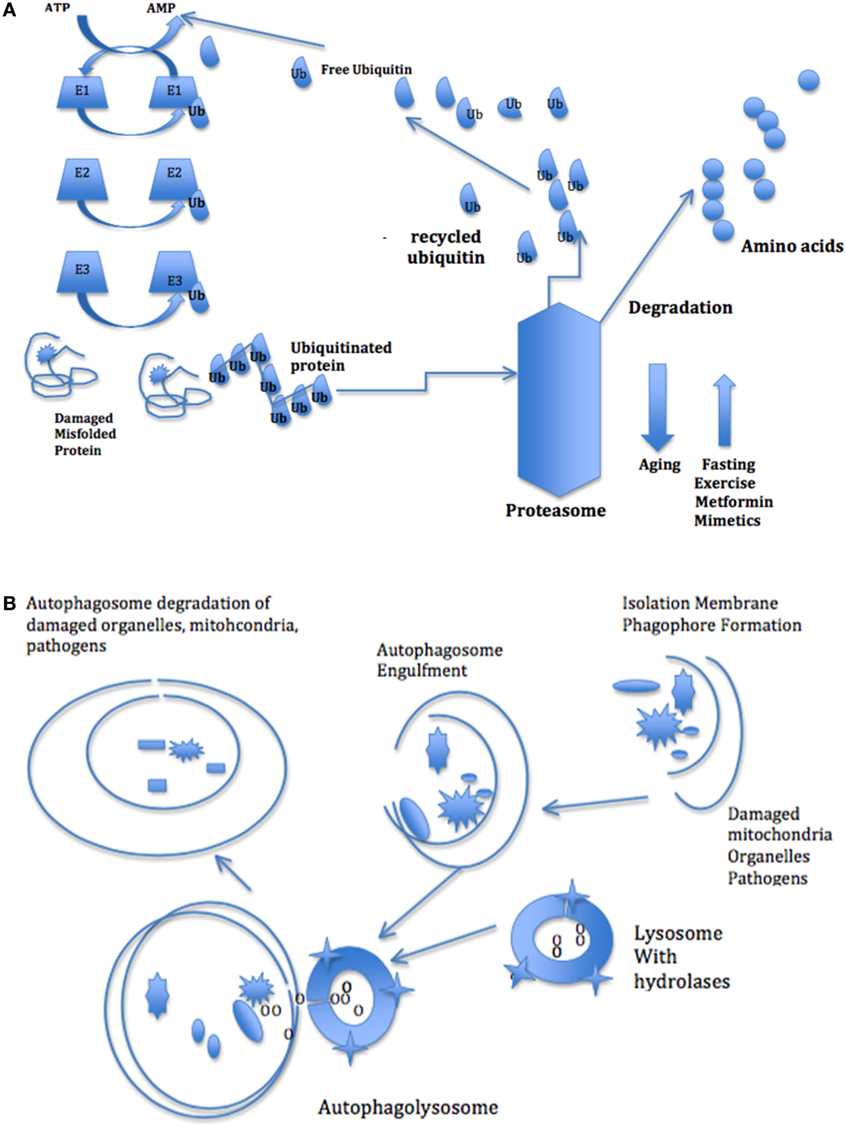
Figure 3. (A) The ubiquitin–proteasome pathway of protein degradation. Three different sets of enzymes—E1, E2, and E3, identify and categorize proteins in order to link ubiquitin or ubiquitin complexes to the damaged proteins. The ubiquitin-protein complexes pass through the proteasome where they are degraded and discharged as free amino acids into the cytoplasm. (B) The autophagy pathway of degradation of damaged organelles and pathogens. The autophagy system degrades larger aggregated proteins and cellular organelles, such as mitochondria, peroxisomes, and infectious organisms. The process involves membrane formation, fusion, and degradation. A small separate membrane called a phagophore forms and then forms the autophagosome that fuses with the lysosome. The autophagosome contents are degraded by lysosomal hydrolases.
The other main pathway is the autophagy system that degrades cystolic components, including larger aggregated proteins and cellular organelles, such as mitochondria, peroxisomes, and infectious organisms (61). This process involves membrane formation, fusion, and degradation (Figure 3B). When autophagy is induced, a small separate membrane structure called a phagophore arises in the cytoplasm, which gradually expands to form the autophagosome. The outer membrane of the autophagosome fuses with the lysosome and the autophagosome contents are degraded by lysosomal hydrolases (62). Like the proteasome, the macroautophagy system is stimulated by intracellular and extracellular stress-related signals, including oxidative stress. Both proteasome and autophagy produce small polypeptides that help maintain a pool of amino acids and control energy balance in starvation, since recycling amino acids is more energy efficient than de novo amino acid synthesis.
In aging and age-related disease there are gradual reductions of cellular repair mechanisms that lead to the accumulation of damaged molecules, proteins, DNA, and lipids leading to loss of efficient cellular function. The cell’s capacity for autophagic degradation also declines with age and this in itself may contribute to the aging process (63). While both major systems for intracellular protein degradation are slowed up with increasing age, a physical reduction of autophagy-related proteins also contributes to the accumulation of misfolded proteins and damaged macromolecules in the cell. Diseases associated with increased oxidative stress, such as cardiovascular and Crohn’s disease and obesity also slow up cellular clearing and reduce autophagy, further contributing to disease (64–66).
The lysosome–autophagy system carries out a wide range of non-specific intracellular degradation and cleaning processes, which include managing pathogens, damaged intra-cellular macromolecules, and surface receptors (67–69). Lysosomal dysfunction is associated with age-related pathology that reduce lifespan, such as Parkinson’s and Alzheimer’s diseases (70, 71). Senescent cells accumulate abnormal protein aggregates in the cytoplasm, and contribute to neurodegenerative disease (72).
The dysregulation in autophagy has important effects in the innate immune response, in aging and age-related diseases by influencing inflammasome activity, cytokine secretion, antigen presentation, and lymphocyte function (73, 74). Under normal circumstances the nod-like receptor 3 (NLRP3) inflammasome fine-tunes the progression of the innate immune response that it has initiated, by upregulating autophagy activity so that the removal of immune mediators is expedited (74). In aging and age-related diseases, the autophagy response becomes blunted, the immune mediators remain active and prolong the inflammatory response (75).
The UPS and autophagy act synergistically and cooperatively to maintain cellular homeostasis (76). Effective autophagic uptake of dysfunctional mitochondria and efficient lysosomal degradation of damaged aggregated proteins and macromolecules are crucial elements in maintaining tissue homeostasis and good health (77). The decline in the autophagy capacity, that impairs cellular housekeeping in aging, seems to be an attractive molecular pathway to target to improve the quality of aging.
Two groups of drugs, the mammalian target of rapamycin (mTOR) inhibitors and AMP-activated protein kinase (AMPK) activators are promising pharmacological agents which stimulate autophagic degradation (78–80). Other drugs, such as the diabetic drug metformin and the oncology agent 5-aminimidazole-4-carboxamide ribonucleoside are pharmacological activators of AMPK, which are soon planned for clinical studies in relation to aging (81–83). A number of substances, such as curcumin, berberine, and quercetin, regularly available in normal diets, appear able to mimic the action of AMPK and upregulate autophagy. The action of AMPK has important anti-inflammatory and immunosuppressive effects (83). By upregulating autophagic activity, AMPK promotes effective clearing of DAMPs and by preventing the activation of the inflammasome, it reduces the triggering of the inflammatory cascade. Further evidence of the anti-inflammatory role comes from research with the AMPK agonist A-769662 that mimics AMPK activity (84). This AMPK mimetic has been shown to suppress inflammatory arthritis in mice and reduce IL-6 expression in serum and arthritic joints, suggesting that targeted AMPK activation could be an effective therapeutic strategy for IL-6-dependent inflammatory arthritis (85).
Non-pharmacological life-style changes also upregulate autophagy. One of the best researched is the effect of exercise which improves mitochondrial mitogenesis and stimulates mitogeny, so improving the quality of muscle function and exercise performance, with improvement in the quality of aging (86, 87). Furthermore in animal model studies, both modulated caloric restriction and exercise increase autophagy, downregulate endotoxin-induced IL-1β production, improve the aging-related pro-inflammatory profile, and reduce disease symptoms (78, 88).
Further understanding of molecular pathways of the signaling networks underpinning autophagy should help to identify other novel drug targets. Important research areas include those that could improve the sensitivity of degradation inhibitors useful to improve anticancer treatment, or new drugs to upregulate autophagy to maintain good cellular housekeeping, with the potential for improving the quality of aging and the management of age-related degenerative diseases.
Senescent Cells
Senescent cells increase with age and are considered important contributors to the pro-inflammatory phenotype (89). The two major hallmarks of cellular senescence are an irreversible arrest of cell proliferation and production of the pro-inflammatory secretome, called the SASP. When replicative senescence was first identified in serial cell passage studies (90), telomere attrition was considered to cause the cellular growth arrest that acted as a mechanism to stop damaged or transformed cells from proliferation and transiting to tumor initiation. Today senescence is considered to have much broader role as both a contributor to damage protection and in the control of cellular growth, or as both a “friend and foe” depending on the cellular context. Senescence together with apoptosis is recognized to play an important physiological role in normal embryonic development, in ongoing tissue homeostasis throughout life (91, 92), but is increasingly considered to have a role in causing or exacerbating aging and age-related diseases (91, 93–95).
Senescence is a stress response triggered not only by telomere attrition as originally described (90, 96), but also by stress insults, such as genomic instability, DNA damage, protein misfolding and/or aggregation, and ROS. There is also an association between senescent cells and the dysregulated mitochondrial network and associated metabolic dysfunction that is seen with increasing age (97). Through the SASP, the senescent cell has an important influence on the extrinsic microenvironment, which suggests a link between senescence and alterations in intracellular and intercellular communications (93).
Cells that express senescence markers accumulate with age in some tissues in studies in mice and man (98–100). Senescent cells are found in association with age-related diseases, such as atherosclerosis, RA, neurodegenerative diseases, and cancer (101–104). In RA patients T-cells are described as showing a pre-aged phenotype with apparent loss of CD28 expression that reduces T-cell activation and this in association with reduced RA-related NK surveillance, could allow senescent cells and the associated SASP to persist. In cancer, SASP factors promote angiogenesis, cell proliferation, and cancer invasiveness. Cells attracted by SASP influence the local microenvironment with the potential to promote tumor invasion and cancer progression (105). Senescent cells have been seen in atherosclerotic plaques (101). Recent data from several laboratories has suggested that both aging and age-related neurodegenerative diseases show an increase in SASP-expressing senescent cells of non-neuronal origin in the brain, which correlated with changes in neurodegeneration (103).
The SASP consists of a complex combination of growth factors, proteases, chemokines, matrix metalloproteinases, and is particularly enriched in pro-inflammatory cytokines, especially IL-6 (106–108). The SASP-secreting cells respond by switching on a self-perpetuating intracellular pro-inflammatory signaling loop, centered around the NF-κB, TGF-β, IL-1α, IL-6 pathway (109–111), with suggested mechanisms related to higher basal phosphorylation and altered threshold signaling (112) or alternative splicing (113). Senescent cells influence other cells by paracrine and bye-stander effects (114). There appears to be multi-level control of senescence and the SASP secretome, which includes the tumor suppressor pathways involved in the cell cycle arrest and the NF-κB and persistent damage response (DAMP) pathway, involved in triggering transcription of the SASP-related factors (115). Several pathways of investigation suggest that senescent primary human CD8+ T cells use anaerobic glycolysis to generate energy for effector functions and that p38 mitogen-activated protein kinase (p38 MAPK) blockade may reverse senescence via the mTOR-independent pathway (116). Low doses of glucocorticoid suppress elements of the SASP in patients with RA and improve clinical symptoms (117). Senescent cells effectively recruit the immune system to organize their removal, but with increasing age, removal becomes sluggish or otherwise impaired (118, 119).
It can be argued that the increase in senescent cells with aging reflects either an increase in their rate of generation or a decrease in their rate of clearance because the immune response is attenuated or weakened with aging and less capable of clearing senescent cells (120–122). Senescent cells express ligands for cytotoxic immune cells, such as natural killer (NK) cells, and have been shown to be able to be specifically eliminated by the immune system (123, 124). Through a proteomics analysis of senescent cell chromatin, the NF-κB pathway appeared to act as a master regulator of the SASP, with NF-κB suppression causing escape from immune recognition by NK cells (125). Other studies show that processes which eliminate senescent cells with p16(Ink4a)-positive markers, delay age-related pathologies in the mouse model of aging though side-effects can be problematical (126, 127). Therapies that specifically recognize and trigger the elimination of senescent cells would seem important to enhance the immune system in older people. New methods are in the process of being developed to enhance the immune clearance and autophagy of the increased senescent cell burden in aging and age-related disease (128).
Inflammasome NLRP3
The inflammasomes, intra-cellular multiprotein sensors that recognize danger signals, are likely key players in initiating and maintaining the pro-inflammatory phenotype found associated with aging. The NLRP3 is a major inflammasome sensor for intracellular stress molecules called DAMPs, which together with damaged aggregated proteins that are released from destabilized lysosomes and damaged mitochondria contribute to the cellular stress (ROS) and trigger NLRP3 activation (129). Once activated, the NLRP3 inflammasome initiates the inflammatory response cascade by stimulating caspase-1 (casp-1) that acts to induce the active precursors of pro-inflammatory cytokines, such as IL-1β, IL-1α, and IL-18, and on-going interaction with NF-κB (130, 131) (Figure 4). Although the baseline activity of NLRP3 is low, the initiation process of the inflammatory cascade requires a complex oligomerization-priming phase that includes association with NF-κB and so contributes several layers of regulatory control.
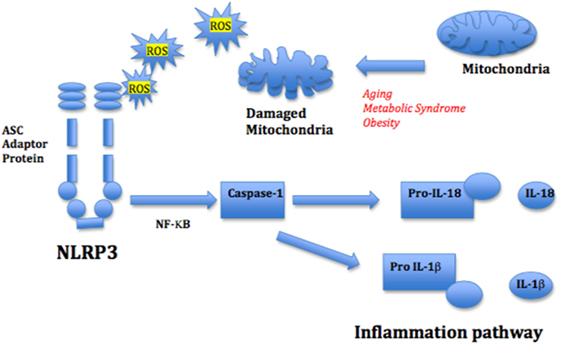
Figure 4. Mitochondrial reactive oxygen species (ROS) and nod-like receptor 3 (NLRP3) activation of inflammation pathway. Mitochondrial ROS from damaged mitochondria triggers the inflammasome NLRP3, stimulating NF-κB and the IL-1β and IL-18-mediated inflammatory cascade. The adapter protein ASC mediates innate signaling by bridging the interaction between the damage recognition receptor and the NF-κB caspase-1 inflammasome complex.
Nod-like receptor 3 has been shown to be able to activate NF-κB and induce cytokines in response to sterile signals, such as monosodium urate crystals and aluminum adjuvant, suggesting that NLRP3 could initiate NF-κB activation to both pathogen-induced and sterile inflammation (132). Conversely NF-κB, which primes the NLPR3 inflammasome for activation also prevents excessive inflammation and restrains NLRP3 activation by enhancing the NF-κB-p62 mitophagy pathway. By self-limiting the host response, the NF-κB-p62 mitophagy pathway maintains homeostasis which under normal conditions leads to tissue repair (75). It is, however, unclear if this layer of control of NF-κB function remains as tightly controlled in aging and age-related disease.
The NLRP3 inflammasome is a key component of the innate inflammatory response to pathogenic infection and tissue damage. It responds to a wide range of cellular stress and is considered to contribute to the aging process and to age-related diseases (133). Zhou and colleagues identified that mitochondrial ROS was involved in the activation of NLRP3 (58). This study emphasized the important role of mitochondria in maintaining a correct balance between cellular energy production and ROS production and that effective clearance of damaged mitochondria through autophagy was an important regulatory activity. Damaged mitochondria increase with aging and age-related diseases (134). Mitochondrial dysfunction drives mitochondrial mutagenesis, affecting respiratory chain genes, and compromising the efficiency of oxidative phosphorylation, which may lead to further mt-DNA mutations and more cell damage. The subsequent mitochondrial impairment leads to more ROS that further reduce ATP generation and increases the chance of cell death. Mitochondria have been identified as a key source of DAMPs, the so-called mito-DAMPs, which have been considered to play a role in DAMPS-modulated inflammation in diseases, such as RA, cancer, and heart disease (135–138) as well as in the aging process (139). Degraded mt-DNA has also been reported in neuroinflammation (140). Dysfunctional mitochondria seem to be able to initiate an auto-feedback loop to increase autophagy, so that damaged mitochondria or misfolded proteins are degraded which reduces inflammasome activation and risk of further tissue injury, though this system is less efficient in aging (141).
Lyosomal destabilization is also associated with NLRP3 activation and can be induced by a number of molecules, including cholesterol crystals in macrophages linking atherosclerosis progression with inflammation (142). There is deposition of other harmful intra- and extracellular material in several age-related diseases. The aggregates compromise cellular homeostasis and can provoke the activation of the NLRP3 inflammasome. Research has shown that amyloid fibrils and Alzheimer’s amyloid-β can trigger NLRP3 inflammasomes and in that way stimulate inflammation and enhance pathogenesis and association between type 2 diabetes and Alzheimer’s disease, respectively (143). Palmitate, a saturated fatty acid has been shown to activate NLPR3, whereas oleic acid did not initiate the same inflammatory response (144). The inflammasome has been implicated in the development of the metabolic syndrome through impairment of adipose tissue sensitivity. Evidence showed that obesity triggered NLRP3 activation, and that the secreted IL-1β impaired insulin signaling which promoted insulin resistance in mice (145). Other research has shown that obesity was associated with the activation of the NLRP3 in adipose tissues (146, 147).
A number of intracellular processes seem likely to work together to stimulate and augment the inflammasome pathway and contribute to pro-inflammatory cytokine upregulation associated with increased age and age-related diseases. Both the redox-sensitive inflammatory pathway and the senescent cell-related SASP activate the inflammasome through the NF-κB and IL-α cascade, causing persistence of the inflammatory response that delays resolution and healing (125, 138). Similarly, reduced autophagy processes allow the accumulation of damaged intracellular proteins and senescent cells that further perpetuate and amplify the pro-inflammatory milieu that is found with increased age and is associated with age-related diseases.
Pro-Inflammatory and Anti-Inflammatory Cytokine Dysregulation
Pro-Inflammatory Cytokines in Aging and Age-Related Disease
Various biomarkers and biochemical indices are used in medicine and age-related diseases as a way of improving diagnosis, beyond the well-recognized clinical signs. Modest increases in concentration of C-reactive protein, a circulating marker of inflammation, have been widely reported to be associated with a large number of age-related conditions and lifestyles felt to be associated with poor health; these conditions represent or reflect minor metabolic stresses. Alongside C-reactive proteins, cytokines have come under investigation as the molecular processes and pathways underpinning inflammation have become better identified. A common finding in aging and age-related diseases is “inflamm-aging,” a dysregulation of the cytokine network and its homeostasis. Downstream from NF-κB signaling, the pro-inflammatory cytokines play a central role in the remodeling of the immune system with age (Figure 5).
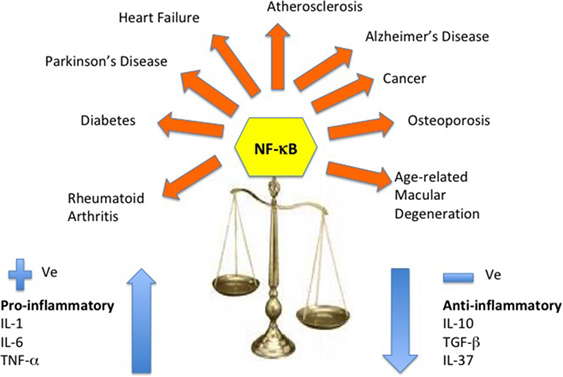
Figure 5. Cytokine dysregulation and NF-κB inflammation pathway. This reshaping of cytokine expression pattern with a progressive tendency toward a pro-inflammatory phenotype has been called “inflamm-aging” and is found associated with age-related diseases. Several molecular pathways have been identified that trigger the inflammasome and stimulate the NF-κB and the IL-1β-mediated inflammatory cascade of cytokines.
The major pro-inflammatory cytokines, such as IL-6, TNF-α, and IL-1α contribute significantly to the phenomenon of inflamm-aging in healthy elderly individuals (8), while also playing a major role in many age-related diseases (11, 27, 148–151). The key to healthy aging must lie in the ability to maintain a balanced response to these immune messengers and a prompt and integrated return to inflammation resolution and immune homeostasis (17). A summary of the changes that have been described in pro-inflammatory and anti-inflammatory cytokines in aging and some age-related diseases are outlined in this section.
Interleukin-1 (IL-1) Family
IL-1α and IL-1β, known as IL-1, and IL-18 are important cytokine initiators of the stress-induced inflammatory cascade (152). IL-1β and IL-18 are cleaved to active forms by Casp-1, whereas IL-1α is activated by calpain protease. All bind to and activate the IL-1R that is downregulated by the receptor anatagonist IL-1Rα, which blocks IL-1-mediated signal transduction.
Studies in elderly people, including centenarians have reported an age-related rise in the IL-1R antagonist, (IL-1Rα), whereas IL-1β showed no detectable age-related trend. The age-related rise is associated with increased co-morbidity, age-related disease, and mortality (153–156).
Certain IL-1 haplotype-carriers produce increased IL-1β, and IL-1 gene variations associate with earlier onset or more severe progression of cardiovascular and Alzheimer’s disease, but not with osteoporosis (157–161). In centenarians, no single IL-1 gene polymorphism showed a survival advantage, but in Swedish elderly males an IL-1 gene polymorphism shortened life expectancy (153, 162, 163). IL-1 gene variants appear to increase the risk of age-related diseases and recombinant drugs, such as IL-1Rα-blockers may have a role in the clinical control of inflammation (164).
Interleukin-18 (IL-18)
Interleukin-18, a linked IL-1 pro-inflammatory cytokine, signals in a complex with IL-18 receptors α (Rα) and β (Rβ) chains and induces IFN-γ that is essential for defense against infections (165). IL-18’s multiple pro-inflammatory effects are modulated through IL-18 binding protein (166).
Higher levels of IL-18 have been found in centenarians, associated with heart failure, ischemic heart disease, and type 1 diabetes in patients, and in the Alzheimer’s disease brain (167–172). IL-18 levels associate with physical functioning and with a frailty index in the English longitudinal study of aging, where carriers of IL-18 gene polymorphism that reduced IL-18 levels, showed improved walking speed (173–175). Evidence consistently shows that IL-1 and IL-18 are mediators of inflammation and associated with the aging process (168). Drugs blocking binding between IL-18 and the receptors are currently in development and may provide benefit in the treatment in diabetes, macular degeneration, and autoimmune disease (176).
Interleukin-6 (IL-6)
Interleukin-6 has been long recognized as important in aging and age-related disease and has been called the “gerontologist’s cytokine” (177, 178). IL-6 plays a key role in the acute phase response, in the transition from innate to acquired immunity, in metabolic control, and in the pathogenesis many chronic diseases (11, 148–151, 179). It has both pro- and anti-inflammatory activities, and modulates the acute inflammatory response by producing IL-1 Rα and soluble tumor necrosis factor receptor p55 (sTNF-R55), which suppresses TNF-α and IL-1.
Interleukin-6 is normally present in low levels in the blood, but is increased in aging and in subjects with markers of frailty and chronic disease, where it tracks with mortality (180–183). IL-6 is a risk factor associated with cardiovascular disease and is associated with sarcopenia and muscle loss (184, 185).
The G allele of IL-6-174C/G polymorphism shows higher IL-6 levels and associates with cognitive decline and mortality in age-related vascular disease, whereas CC allele carriers show decreased Alzheimer’s risk (186–191). In a meta-analysis of longevity in a large cohort of European nonagenarians and centenarians there was longevity benefit for carriers of the lower cytokine producing IL-6 allele, with similar supporting findings for this IL-6 allele in a case control study (192, 193). IL-6 or IL-6 receptor blockers are already used successfully in the treatment of RA, and are proof of concept that damping down IL-6, a product of the NF-κB pro-inflammatory cascade, can improve clinical symptoms. Studies are either in progress or planned to assess the outcome of blocking IL-6-related inflammation in other age-related diseases with the potential for contributing to more successful aging (194, 195).
Tumor Necrosis Factor Alpha
Another major player in the immune response is the pro-inflammatory cytokine TNF-α, which increases with age and is associated with age-related disease (196). It is a pro-inflammatory mediator that can be beneficial when it acts locally in the tissues, but can be highly harmful when released systemically.
Tumor necrosis factor-α has been reported to be increased in intracellular aging studies in elderly people, in centenarians and octogenarians with atherosclerosis, and associated with mortality (197–202). In post-MI patients, a rise in TNF-α increased risk of recurrent cardiac events and in renal patients TNF-α receptors predicted cardiovascular disease (203–205). In genetic studies, the A allele of TNF-α 308 G/A gene associated with risk for MI, whereas TNF-α polymorphisms and TNF-α itself, have been variably associated with increased Alzheimer’s disease risk (206–210). TNF-α mediates metabolic changes and increased TNF-α was found in type 2 diabetes mellitus and was associated with lower muscle mass and strength in older groups (211).
In studies in nonagenarian/centenarian groups from three European countries, there was no attrition of the TNF-α-308 A/G polymorphism in centenarians (162, 212, 213). With increasing evidence of an association between increases in TNF-α and age-related diseases, research re-purposing anti-inflammatory drugs are under development. Research has demonstrated that TNF-α inhibitors may have possible prophylactic or ameliorating roles in cardiovascular and Alzheimer’s disease in animal models (214, 215).
Other Pro-Inflammatory Cytokines
Other pro-inflammatory cytokines are increasingly being recognized as dysregulated in association with aging and age-related disease.
Interleukin-2
Interleukin-2 plays a pivotal role in the immune response. It is a growth factor that promotes NK cell activity and the differentiation of naïve T cells into Th1 and Th2 cells (216). Conversely, IL-2, acting via STAT5 pathway negatively regulates interleukin 17 (IL-17) production (217). Most studies show that lymphocytes in elderly people produce significantly less IL-2, compared to young people (218–220). Intracellular cytokine studies have shown variable results for IL-2, whereas mitogen-induced stimulation of mononuclear cells from elderly subjects showed significant decreases in IL-2 and IFNγ production (197, 221).
The IL-7/IL-7R
The IL-7/IL-7R network is essential at various stages in T-cell development and survival (222). It has an important role in the maintenance of a vigorous health span and higher IL7R gene expression is associated with long life (223–225). Serum IL-7 is increased in some age-related diseases, including osteoarthritis and genetic variation in the IL7RA/IL7 pathway increased susceptibility to multiple sclerosis (226, 227). Research has suggested that silencing of the IL-7R gene may be an important mechanism underpinning an aging-related loss of binding to NK-κB (228), linking IL-7R gene to the NF-κB pathway and inflammation control.
Interleukin-12
Interleukin-12, a pro-inflammatory member of the IL-6 family has an active role in the development of cardiovascular diseases, such as atherosclerosis, MI, and stroke (229). Patients with cardiovascular disease show increased levels of IL-12, 23, and 27 with higher IL-12 predicting poorer long-term outcome after acute MI (230). Other research shows variable results for IL-12 and its receptor antagonist, with increased IL-12 (total) and IL-12p40 in apparently healthy nonagenarians, lower IL-12p70 and IL-23 production in association with frailty and IL-12/23p40 ameliorating Alzheimer’s disease in animal models (231–233).
Interleukin 17
Interleukin 17 is a key pro-inflammatory cytokine that belongs to a family of six cytokine members (A–F). IL-17A (referred to as IL-17) plays a central role in host defense against invading pathogens and is produced by a subset of CD4+ cells (234, 235). Elderly people (age ≥65) have shown a decreased frequency of IL-17-producing cells in memory subset of CD4+ T cells compared to healthy younger people (236). IL-17 enhances production of IL-6, TNF-α, the acute phase reactants, C-reactive protein, and serum amyloid A and activates the induction of IL-6, IL-8, and G-CSF in non-immune cells, such as fibroblasts and epithelial cells, in part through activation of the NF-κB transcription factor (237). IL-17 promotes inflammation and is overexpressed in many autoimmune diseases, such as RA, systemic lupus erythematosus, inflammatory bowel disease, and psoriasis and its effects are stabilized by IL-23 (238–241). An IL-17 expressing CD8+ T subset of cells has also been reported to be involved in psoriatic arthritis and some other autoimmune diseases (242, 243).
Interleukin-8
Interleukin-8 (or CXCL8) is a chemokine secreted by monocyte/macrophages whose key role in the inflammation process is the recruitment and activation of neutrophils. IL-8 has been implicated in a number of inflammatory conditions, such as cystic fibrosis, asthma, chronic pulmonary disease, inflammatory bowel disease, and some autoimmune diseases, including RA and psoriasis.
Increased levels of IL-8 have been detected after LPS-stimulation of leukocytes from elderly individuals (244). In one small study of centenarians, IL-8 was proposed as a possible longevity factor (245). A single study of IL-8 polymorphisms found no significant difference in IL-8 -251 A/T polymorphisms in nonagenarians compared to young controls (212). IL-8 signaling occurs via the MAPK and PI3K pathways, by binding to the IL-8 receptors-CXCR1/2. Several agents that block IL-8-CXCR1/2 signaling have been developed in an attempt to target inflammatory pathways in cancer, asthma, chronic obstructive pulmonary disease, psoriasis, and RA (246).
Anti-Inflammatory Cytokines in Aging and Age-Related Disease
The anti-inflammatory cytokines play a key role in balancing the immune response, and in preventing the tipping of the steady state of immune homeostasis across into inflamm-aging and a disease-inducing state. Anti-inflammatory cytokines are an important arm of inflammation resolution. They block or modulate the synthesis of IL-1α, TNF, and other major pro-inflammatory cytokines and damp down the inflammatory response, so that inflammation resolution can begin. Specific cytokine receptors for IL-1, TNF-α, and IL-18, together with soluble receptor antagonists, chemokines, microRNA, siRNAs, also function as inhibitors for pro-inflammatory cytokines. The anti-inflammatory cytokines and families of soluble receptor antagonists work within a complex network of control of immune regulation. They are critical for balancing the inflammatory outcome and together with pro-resolving lipoxins are critical to resolving inflammation in an integrated and organized manner.
As age increases and in age-related diseases, a chronic inflammatory state predominates, which is not properly contained or resolved and the anti-inflammatory side of the immune system seems to be similarly dysregulated, and unable to damp down the inflammatory episode in a timely effective manner. The following cytokines are the major players in the anti-inflammatory pathway of the control of inflammation and changes in their production and expression have been quite widely reported in aging and age-related disease. Where increases in anti-inflammatory cytokines have been reported, one interpretation would be that increases might reflect the immune system’s attempt to suppress the persistent pro-inflammatory response and support a return to immune homeostasis.
IL-10 Family
Interleukin 10 is one of the key anti-inflammatory cytokines, which suppresses the actions of IL-6, TNF-α, and IL-8 (247, 248). Higher IL-10 serum levels and production by both lymphocytes and monocytes have been reported in elderly people (155, 244, 249). Conversely an age and gender-related decline in cellular stimulation studies has been reported (250).
In age-related disease, IL-10 has been reported to be associated with vascular protection in atherosclerosis and improved endothelial dysfunction (251–253). However, at variance, the authors from the ERA (254) and PROSPER (255) studies, concluded that elevated IL-10 increased cardiovascular risk among elderly groups, and suggested that IL-10 blockers merited investigation. In male Sicilian centenarians, male carriers of the high producing GG 1,082 allele of the IL-10 promoter polymorphism showed a survival advantage, suggesting that IL-10 anti-inflammatory activities might be a marker for male longevity (213). This result was not replicated in Sardinian, Irish, or Finnish nonagenarian/centenarians (162, 212, 256). It has been argued that an enhanced anti-inflammatory phenotype could be beneficial and contribute to longevity by controlling the pro-inflammatory milieu that predominates in later life and contributes to increased morbidity and mortality (9, 11, 257).
TGF-β
TGF-β, another important anti-inflammatory cytokine limits both the acute phase response, and is involved in tissue repair post-damage or infection (258). Several authors have reported that TGF-β was increased in octogenarians and centenarians (148, 259). It is also involved in aging-related disease, such as in obesity, in vascular wall integrity, in muscle loss and sarcopenia, in osteoarthritis, and with frailty in the Newcastle longitudinal study (260–264). In stroke, TGF-β signaling was increased in microglia and macrophages suggesting that increased TGF-β likely regulated glial scar formation (265). Reports have linked TGF-β or its polymorphisms with atherosclerosis and Alzheimer’s disease (266–268). Other research found TGF-β genotypes associated with longevity in Italian centenarians, a finding not replicated in BELFAST nonagenarians (212, 269). Context-specific environmental factors, epigenetic regulation, and non-coding RNAs are suggested to play a role in TGF-β’s paradoxical pro-and anti-inflammatory functions (7, 270, 271), but important uses have been found for TGF-β in fibrosis management and oncology (272).
Interleukin-37
Interleukin-37, formerly an IL-1 cytokine, limits innate inflammation via suppression of pro-inflammatory cytokine production (273). Carriage of an IL-37 haplotype that decreases IL-37 levels contributes to increased inflammation. Research demonstrates that IL-37 reduces TNF-α and IL-1β cytokine production from human macrophages, is increased in chronic heart failure patients and attenuated the production of inflammatory cytokines in serum or synovial joints in RA, suggesting IL-37 may have a role in clinical disease (274–276).
Age-Related Diseases
Cancer
Cancer increases with aging, with one in two people likely to develop malignant tumors in their lifetime. Probable reasons for this age-related increase include exposure to environmental toxins, declining immune surveillance, and increasingly ineffective DNA repair mechanisms. Inflammation is involved at different stages of tumor development, at initiation, promotion, malignant conversion, invasion, and metastasis, has a paracrine bystander role and is an essential part of the tumor micro-environment. Inflammation also affects immune surveillance and responses to therapy (277). Thus, malignancy is a major threat to successful aging.
While inflammatory pathways are vital to promote immune homeostasis, over-activation or dysregulation can be pathological and lead to malignant progression. Prolonged inflammation, either as a result of chronic infections, or reduced homeostasis in the inflammatory response, plays a role through the production of pro-inflammatory cytokines that may be directly or indirectly implicated in the oncogenesis (278, 279). More recent investigations have focused on the role of the inflammasone pathway, whose biochemical function is to activate casp-1, which leads to the activation of the IL-1β and IL-18 pathways and induction of pyroptosis, a form of cell death. Although inflammasomes have an important role in inhibiting cancer, through the triggering of the programmed-death pathway, they both initiate and maintain carcinogenesis, dependent on tumor type and the tumor environment (280, 281).
Bacterial and viral infections are associated with malignancies. For example, Helicobacter pylori (H. pylori) infection of the gut is associated with both gastric cancer and mucosa-associated lymphoid tissue (MALT) lymphoma (282). Epstein–Barr virus (EBV) is a causative agent in Hodgkin’s disease (HD), where chronic inflammation is considered a major contributory factor (283), human papilloma virus is implicated in most cases of cervical cancer (284), while human T-lymphotrophic virus 1 (HTLV-1) is a causative agent in adult T-cell leukemia lymphoma (285). A common factor is the association of infection with oncogenesis, with chronic inflammation a contributory factor.
In H. pylori chronic infection, elevated levels of IL-1β are detected and recognized as important in the development of gastric carcinoma. Normally gastric acid in the stomach does not permit bacterial survival, but in circumstances of low stomach acidity, H. pylori grow vigorously in the mucosa and induces caspase-mediated cleavage of pro-IL-1β and pro-IL-18 in association with the NLRP3 inflammasome. The overexpression of IL-1β induces NF-κB activation and the transcription and expression of IL-6, TNF-α, and IL-10. The proinflammatory cytokine milieu increases the risk for developing both gastric carcinoma and MALT lymphoma (286). Persistently high levels of IL-1β and IL-18 suppress acid secretion, allow hypoacidity in the stomach, loss of parietal cells, gastric atrophy, metaplasia, and eventually gastric cancer. In addition, IL-1β inhibits gastric acid secretion and carriers of IL-1β polymorphisms producing higher IL-1β carry increased gastric cancer risk (287, 288). H. pylori infection of gastric mucosa can cause a monoclonal B cell proliferation, with a histological diagnosis of MALT lymphoma. This tumor-like proliferation of gastric mucosal cells and clonal B cells can regress after eradication of the H. pylori infection with combined antibiotic therapy and proton pump inhibitor treatment (289).
Viral infections strongly stimulate inflammatory responses and may lead to malignant transformation of the host cell (290). Although the activation of the inflammasome benefits the clearance of viruses and the regression of cancer, there are several examples of viruses, such as EBV and HTLV-1 developing strategies to evade detection, triggering the inflammasome, and high-jacking the inflammatory cascade to induce, and amplify the cancer spread. For example, when EBV infects B-lymphocytes and nasopharyngeal cells through its receptor CD21 (291), this leads to a proliferation of infected B cells, followed by an increase in CD8+ T cells, that controls the infected cells by lysis. However, where the normal infection-limiting response is “exhausted” or dysregulated, B cell proliferation continues unabated leading to chromosomal damage, which drives cell proliferation outside normal control mechanisms and may result in an aggressive non-Hodgkin’s or Burkitt’s lymphoma (292). NLRP3 activation has been demonstrated in EBV-associated cancerous tissues (293). Furthermore, EBV has been shown to be able to overcome the immune response by means of EBV miRNA binding to the 3′-untranslated region of NLRP3 (294), so preventing effective immune activation and control mechanisms.
Retro-viruses stimulate inflammatory responses and are associated with malignant transformation of host cells. They reverse transcribe their RNA into the host cell’s DNA, leading to dysregulation of cellular proliferation and programmed cell death responses, and elicit a pro-inflammatory response. HTLV-1 causes adult T-cell leukemia by targeting CD4+ T cells that express CD25 (IL-2Rα) and FoxP3, similar to Tregs (295, 296). The persistent activation of the NF-κB pathway in HTLV-1-infected T cells and the associated NF-κB oncoprotein Tax contribute to the oncogenic transformation (297). The resulting hijacking of the NF-κB pathway, allows uncontrolled upregulation of cellular genes that govern growth-signal transduction, amplify the pro-inflammatory cytokines (IL-2, IL-6, IL-15, TNF), together with increasing expression of proto-oncogenes (c-Myc), and antiapoptotic proteins (bcl-xl) Hiscott Rayet (298, 299). Inter-individual susceptibility to HTLV-1 infection has been associated with allele carrier status of the NLRP3 gene (300).
In summary, the interaction of infective agents, host cells, adaptive immune cells, cytokine production, and the inflammasome response is complex and incompletely understood. Many cancers arise from sites of infection, chronic irritation, and inflammation, which although sometimes reversible in the pre-malignant phase by eradicating the causative virus or bacterium, often treatments are too delayed to prevent the cancer development. There needs to be improved understanding about the roles of inflammation, the inflammatory cells, and the paracrine effects that allow tumor cell proliferation, survival, and migration. Does the pro-inflammatory environment found in aging enhance and facilitate cancer cell proliferation or does it alternatively represent an upregulated immune surveillance mechanism to deal with increased damaged and dangerous cancer cells? Improved understanding of the pathways involved should begin to provide insights that could contribute to new anticancer and anti-inflammatory therapeutic approaches through manipulation of autophagy for cancer treatment regimes or conversely tagging cancer cells for destruction through proteasome or autophagy upregulation (301).
Rheumatoid Arthritis
Chronic tissue inflammation has an important role in the etiology and immunopathogenesis of RA (302), with genetic and environmental factors contributing to a predilection to develop the disease. In the pre-clinical asymptomatic phase of RA disease, the immune system is characterized by reduced self-tolerance and production of autoantibodies, whereas in the clinical phase (303) innate and adaptive immune cells infiltrate the synovial joints and produce symptoms of joint pain and stiffness (304, 305). As RA progresses, immune cells and synovial fibroblasts produce a pro-inflammatory environment in the joint (306, 307) leading to joint destruction (302). Cell-specific cytokines, include TNF-α, IL-1, and IL-6 from macrophages, IL-6, IL-7, and IL-15 from memory T-cells, IL-1 and IL-17 from helper T-cells, and IL-1, IL-6, IL-18, GM-CSF, and TGF-β from synovial fibroblasts (303, 308). This complex cytokine milieu attracts further immune cells, promotes abnormal angiogenesis and osteoclastogenesis, poorly formed leaky vasculature and leads to systemic effects (309).
There is evidence to suggest that activation of the NLRP3-inflammasome contributes to the inflammatory processes in RA. Active RA subjects have increased expression of NLRP3 and NLRP3-mediated IL-18 secretion in whole blood upon stimulation via TLR3 and TLR4, but not TLR2 receptors (310, 311). Functional polymorphisms in the genes coding for NLRP3 and its component parts, including CARD8 has been shown to contribute to higher disease activity at diagnosis and for response in the early months of treatment (312, 313).
Patients with RA show premature immune aging and accumulation of CD28− pre-aged effector T cells that associate with disease activation and prognosis (314, 315). A novel T-cell subset CD28− Treg-like cell has been described that produce pro-inflammatory cytokines, mirroring the SASP associated with senescent cells (316). RA patients who show CD28− senescent Treg-like cells in blood seem to demonstrate earlier and more severe osteoporosis (317).
Limiting inflammation before damage occurs is central to successful RA management and the use of specific monoclonal antibodies has been a key therapeutic strategy. The central roles of TNF and IL-6 in RA have been corroborated by clinical trials of biologic drugs, which can specifically target and neutralize these cytokines. Evidence from RA clinical subgroups stratified by responses to specific biologic drugs strongly suggest that for a particular individual, inflammation is coordinated by a predominant cytokine pathway, such as TNF or IL-6 (318).
Anti-TNF biologics, such as adalimumab, etanercept, and infliximab reduce inflammation, pain, neovascularization, lymphocyte infiltration, and increase macrophage apoptosis (318–321). Anti-IL-6R biologics, such as tociluzimab and anti-IL-6, such as sirukumab, strongly reduce disease activity and erosive progression (322, 323). Evidence suggests that the predominant cell cytokines seen in synovial histopathology may act as prognostic biomarkers for stratification of RA patients (324–326).
Studies of TNF and IL-6 gene polymorphisms further support their role in RA risk and severity. SNPs in IL-6 and IL-6R genes associate with increased RA risk and joint damage (327–329), and the TNF 308 G gene polymorphism with RA disease severity and poor response to anti-TNF treatment (330–334). In the elderly person with RA, there is difficulty in distinguishing whether chronic inflammation or genetic “predisposition” initiates disease or if late-onset RA is hastened by the pro-inflammatory phenotype associated with aging. TNF-α inhibitors used as disease-modifying agents in RA improve not only the clinical symptoms of RA, but also decrease the associated vascular risk (335), suggesting that a stratified biologic approach may be of use to therapeutically dampen chronic systemic inflammation related to aging and other age-related diseases.
Like other age-related diseases and aging itself, there is evidence for dysregulation in both the autophagy–lysosomal and the ubiquitin–proteasomal systems in RA (102). Autophagy seems to be activated in RA in a TNFα-dependent manner and regulates osteoclast differentiation and bone resorption, emphasizing a central role for autophagy in joint destruction (336). Gene and allele frequency population differences seem also to contribute to how effectively cellular autophagy processes work within the cell in removing damaged proteins and other necrotic cellular debris. Polymorphisms of the ubiquitn E3 ligase gene that directly influence autophagy have also been identified and have been associated with the etiology and response to drug treatment in RA (337, 338). Both are likely important contributors to the action and effectiveness of disease modifying and monoclonal biological drugs used in RA treatment. The role of the NLPR3 inflammasome may give opportunities for developing other disease-modifying drugs by targeting upstream triggers of the NLPR3 pathway.
Atherosclerosis
Atherosclerosis is recognized as a chronic inflammatory condition (339) and atherosclerotic plaques show cellular senescence (340, 341). Cytokines are involved in all stages of the pathogenesis of atherosclerosis, having both pro- or anti-atherogenic effects (342, 343). In response to increased low-density lipoprotein (LDL), hypertension, and subsequent shear stress, cytokines modulate endothelial cell permeability and recruit monocytes and T-lymphocytes (344, 345). The continuous monocyte recruitment, foam cell and fatty steak formation eventually result in unstable plaque development, thrombosis, and a cardiac event (345, 346).
Chronic unresolved inflammation is a key feature in atherosclerosis and the levels of SPMs, particularly resolvin D1, and the ratio of SPMs to pro-inflammatory leukotriene B4 (LTB4), are significantly decreased in the vulnerable plaque regions (27). Vulnerable atherosclerotic plaques are recognized as having distinct features; increased inflammation; oxidative stress; areas of necrosis overlain by a thin protective layer of collagen (fibrous cap). In advanced atherosclerotic plaques, macrophages have more abundant nuclear 5-LOX, which is suggested to lead to conversion of AA to proinflammatory LTs, with the potential to contribute to plaque rupture (27).
The NLRP3 inflammasome, a central regulator of inflammation (58), is activated by cholesterol crystals and oxidized LDL (347, 348) that drives the IL-1β inflammation pathway. Recent research targeting IL-1β inflammation in atherosclerosis using cannakinumab, a therapeutic monoclonal antibody, has shown up to 15% lower rates of recurrent cardiovascular events, which was independent of lipid lowering (349). As well as playing a major role in chronic inflammation, NLRP3 is also upregulated during endothelial cell senescence (350) via ROS, and is negatively regulated by autophagy (351, 352). The NLRP3 inflammasome, therefore, appears to warrant further investigation as a potential target for inflamm-aging related to atherosclerosis given that such mechanisms are now of well known importance in atherosclerosis (353).
The gut microbiome has been implicated in age-related inflammation (354) with numerous studies reporting bacterial organisms in arterial plaque (355–357). Emerging research reports bacterial DNA in blood associated with a personal microbiota fingerprint as a predictor of cardiovascular events and stool microbiome as a signature of cardiovascular disease (358, 359). Similarly, bacterial DNA has been noted in cell-free plasma in cardiovascular and chronic renal disease patients (360, 361). Altered gut microbiota composition or dysbiosis is also seen in elderly people, and is associated with inflammatory markers (354). Aging leads to changes in intestinal permeability in gut bacterial milieu (362), and the increased circulatory bacterial DNA observed associated with atherosclerosis support further investigation of the microbiome as a contributory factor to age-related inflammation and atherosclerosis.
Neuroinflammation and Neurodegenerative Disease
Inflammation has been well established as a major component of neurodegenerative disorders, but it has never been clear if this was a direct cause of the disease or a consequence of the progressive degenerative process that was occurring (363, 364). The central role of cytokines in regulating the immune response has been implicated in neurodegeneration, but over the past decade, there has been a revolution in our understanding of how cytokines contribute to the etiology of the leading neurodegenerative disorders, including Alzheimer’s (AD) and Parkinson’s disease (PD).
In AD, central events seem to include the inflammasome, the NF-κB pathway, and the activation of microglia by a variety of factors, including beta amyloid and pro-inflammatory cytokines (172). Microglia, the primary components of the CNS innate immune system (365), produce cytokines and monitor the integrity of CNS. Together with astrocytes, microglia are the primary effectors of neuroinflammation and express PPRs that allow early recognition of PAMPs and DAMPs. When the NLRP3 inflammasome is activated, the inflammation cascade begins with casp-1 that facilitates the processing of IL-1β and IL18. These proinflammatory cytokines drive the inflammatory cascade through downstream signaling pathways and lead to neuronal damage and death (366). The activated microglia release proinflammatory cytokines, such as IL-1β, IL-6, TNF-α, and IL-18, that contributes to neuronal death and dysfunction.
There is interest in the role of sphingolipid metabolites, such as ceramide and sphingosine-1-phosphate, which regulate a diverse range of cellular processes that are important in immunity, inflammation, and inflammatory diseases (367). Growing evidence suggests that ceramide may play a critical role in NLRP3 inflammasome assembly in neuroinflammation. Research has shown that microglia treated with sodium palmitate (PA) induce de novo ceramide synthesis, triggering the expression of NLRP3 inflammasome assembly and resulting in release of IL-1β (368), linking neuroinflammation with dietary lipids. Recent insights into the molecular mechanisms of action of sphingolipid metabolites suggest roles in altering membrane composition, with effects on cellular interactions and signaling pathways with potential causal relationships to neuroinflammatory disease.
Dysregulated autophagy has been considered to play a role in neurodegenerative diseases, particularly AD, and is felt to be a key regulator of Aβ abnormal protein generation and clearance (369). In AD the maturation of autophagolysosomes (i.e., autophagosomes that have undergone fusion with lysosomes) and their clearance are hindered. Evidence suggests that Aβ peptides are released from neurons in an autophagy-dependent manner and that the accumulation of intracellular Aβ plaques is toxic to brain cells leading to AD pathology (370). Furthermore, lysosomal and autophagocytic dysfunction has been associated with both Alzheimer’s and Parkinson’s diseases (71, 72). Senescent cells too, accumulate abnormal protein aggregates in the cytoplasm that contribute to neurodegenerative disease (72). Cellular senescence has been reported in the aging brain with an increase in SASP-expressing senescent cells of non-neurological origin that are likely to contribute to the pro-inflammatory background (103, 371).
In AD and PD, the application of genome-wide association studies (GWAS) has demonstrated a number of key genes, relating to immunity, including the human leukocyte antigen (HLA) complex on chromosome six that regulates the immune and inflammatory response (372, 373). In the most recent Parkinson’s disease GWAS a locus containing the IL-1R2 gene was identified as significantly associated with disease risk and awaits further investigation (372). There is some evidence that carriage of certain pro-inflammatory cytokine gene alleles may confer increased Alzheimer’s disease risk. Single studies have reported that carriers of the A allele of the TNF-α 308 G/A gene were variably associated with increased risk of Alzheimer’s disease (207–210) and that carriage the higher IL-6 producing allele of IL-6 (174 G/C) may confer increased risk (186, 190, 191). Animal studies have provided some clearer understanding of the role of TNF-α in Alzheimer’s disease with evidence of disease modulation with the use of anti-TNF agents (215). Three studies, published in 2013, confirmed a role for the immune response in AD identifying the microglia-related gene TREM2 as harboring an intermediate effect size variant in risk of AD that has also been implicated in other related neurodegenerative diseases (374–376). A recent study of rare variants has also implicated a role for microglial-mediated innate immunity in AD (377).
A better understanding of the molecular pathways involved in the use of established drugs, such as non-steroidal anti-inflammatory or statin drugs in risk and progression of neurological disorders may provide further opportunities to treat earlier or prevent disease onset (378–380). It has been considered that downregulation of the type and magnitude of the pro-inflammatory immune response in neurodegeneration might be a key to earlier and more successful targeting of these pathways. However results, to date, have been disappointing and anti-TNF-α therapies and targeted treatment of TNF-α levels that are elevated in cerebrospinal fluid and in patients’ serum, have produced, at best, modest results (381). Multiple sclerosis patients have benefited from treatment with fingolimod (FTY720) that has been reported to attenuate neuroinflammation, by regulating the activation and neuroprotective effects of microglia, by modulating the sphingosine-1-phosphate receptor (S1P receptor) (382). Given the success of FTY720 for treatment of multiple sclerosis, it is hoped that next-generation S1PR1 modulators will find wider therapeutic uses in other inflammatory disorders. Fingolimod is now under a phase 2 clinical trials for acute stroke and phase 4 for neurodegeneration (383).
Future Considerations
Aging is heterogeneous among people and highly variable between different organs and tissues. Our genes, our lifestyles, and our response to stress are infinitely individual and variable, so that the immunobiography of each life tells a different story of how each will respond to the internal and external environmental stressors (1–3, 384). But evidence is accumulating that the aging process may be malleable.
Because aging is the major risk factor for age-related diseases, understanding age better and maintaining the health of older people and societies is highly important personally and for societies and governments. Knowledge about the underlying molecular pathways and the genetic and life-style processes associated with age-related disease and aging itself is increasing. Evidence from centenarian and nonagenarian studies suggests that these oldest members of populations have had the ability to delay aging and age-related disease (385, 386). Other studies suggest that centenarians may demonstrate optimized cardiovascular risk factors (387, 388), or have either intuitively or through social example, adopted lifestyles which have interacted with their genes to facilitate a successful aging phenotype (3, 389, 390).
Population studies across the world show that the age-specific incidence of cardiovascular disease, stroke, and dementia is decreasing (391–395). This suggests that better blood pressure and diabetic control and statin use may directly or indirectly link into and downregulate molecular pathways associated with inflammation (396–399). Research into how carriage of certain gene alleles, such as TCF7L2 or IL-6 can increase inflammation or stroke risk, respectively, and can be ameliorated by following a Mediterranean-type diet (42, 400, 401), or how gene splicing and features of senescence may be modulated by resveratrol in food (402), herald research into how gene, diet, and lifestyles can interact, with positive or negative effects on the immune system and health. Increased knowledge is emerging as to how epigenetic modulation can affect cytokine genes with reports linking cytokine epigenetic change to neuroinflammation (403–405). Obesity, smoking, and malnutrition have been shown to have next generational epigenetic effects, and seem likely to contribute to the predilection of offspring developing age-related disease or conversely the longevity phenotype (406–409).
Other strategies should be adopted which link with public health messages and encourage people to adopt behavioral changes in lifestyles. Modifications should include: changes in diets to include more omega-3 containing foods or fruits and vegetables as in the Mediterranean diet (410–413); engagement in regular moderated exercise routines (414–417); continued engagement with social connections and intellectual activities in daily lives (418–420); or best of all a combination of life-style factors (3, 421, 422), all of which have been shown to reduce the inflammatory profile and improve the quality of aging. Although the role of diet on human health and connections through nutrition, inflammation, and cancer are not as linear as those between tobacco, smoking, and lung cancer, obesity is linked to chronic inflammation through several mechanisms, including the dysregulation of autophagy, whereas fasting has anti-inflammatory effects, similar to the effect of exercise (423–426), and may downregulate inflammatory biomarkers (427–429). There is, therefore, considerable interest in the role of the intestinal microbiota and health and the so-called immune-relevant microbiome (324, 354), with important correlations between inflammation and neurodegenerative disease (430), bacterial β-hydroxybutyrate metabolites (431), and the role of vagal stimulation (432).
Increasing evidence shows that many signaling pathways are activated in a stress-type-dependent fashion, and all appear to converge with nuclear factor (NF)-κB signaling, which is a central controller of the immune response, and inflammatory cascade (110, 433–436). With increasing age, immune homeostasis loosens, NF-κB signaling becomes less tightly controlled or is more readily triggered, cytokine dysregulation occurs, and a pro-inflammatory phenotype predominates that underpins most major age-related diseases from atherosclerosis to cancer, and aging itself (Figure 5). Understanding how different factors trigger the NF-κB cascade is an important pathway of research (434). In animal models, miRNA-based regulatory networks involving miR-155 and miR-146a, finely regulate NF-κB activity, with miR-146a downregulating and miR-155 upregulating NF-κB expression (435). There is an important temporal separation of miR-155 and miR-146a cellular expression that allows finely controlled NK-κB signaling and enables a precise macrophage inflammatory response, which merits further research.
Therapeutic opportunities may arise through better understanding of the molecular mechanisms that induce senescent cells and SASP in the cellular environments of chronic disease or whether senescent cells can be removed by upregulating autophagy and using sophisticated tagging mechanisms (110). There will be increased opportunities to use the knowledge gained from clinical studies in autoimmune disease, about the roles and actions of monoclonal antibodies in modulating inflammation, which may be able to be utilized in treatments for other age-related diseases involving inflammation (436). The formulations of new and more specific drugs are likely to become available as the modes of action of kinases, such a AMPK and mTOR which control the senescence and inflammation pathways, become better understood (81, 84, 437). Old drugs, such as metformin, still used in diabetes control, are being repurposed and have been shown to have exciting new uses through their ability to modify epigenetic gene expression. Clinical studies are underway to assess any modulating effect of metformin in aging and age-related diseases (81). The use of histone deacetylating drugs is likely to increase as the clinical use of deacetylation and methylation agents is evaluated in cancer with improved knowledge of their effects and safety criteria (438). The current interest in diet and modified diets will encourage further studies assessing how nutrachemicals modify gene expression, for example, through the regulation of intracellular receptors that bind the promoters of certain genes, and may help to design more specific drugs to modify metabolism and benefit health (439).
Turning research to focus on improved understanding of the mechanisms of inflammation resolution in aging and age-related disease, should also be prioritized, since it is an under researched area. Developing synthetic resolvins for use in inflammation resolution may have advantages over the use of single biological anti-inflammatory blockers in autoimmune disease clinical management, since cytokine networks are highly interactive and complex (440), with many auto-regulatory feedback loops. All these molecular pathways are, or have the potential for being developed as drug targets toward clinical interventions useful in damping down and modulating inflammation (441, 442) and may have a role in delaying the onset or treatment of age-related diseases.
Evidence from on-going global studies of the oldest members of our societies, such as centenarians and nonagenarians (443–454) suggests that it may be possible to delay age-related diseases and that aging may be a potentially modifiable risk factor (455). Further investigation has shown that centenarians and super-centenarians also have an enhanced pro-inflammatory background (9, 456, 457), which at first seems surprising, given their long lives. However, studies have demonstrated that the pro-inflammatory background is accompanied and perhaps modulated, by an enhanced anti-inflammatory status in some centenarians. Some have argued that an enhanced anti-inflammatory phenotype could be beneficial as a contributor to longevity by effectively controlling the pro-inflammatory background (9, 11, 257). Others suggest that some inflammation is good, in the same way as hormetic stress triggers systems, and upgrades them but does not overwhelm them (458). Regular exposure to pro-inflammatory stressors could train the immune system to upregulate and fine-tune its cellular processes, so that it responds better and provides better outcomes, when faced with real life-threatening pathogenic threats.
Genome-wide association studies have proved a powerful methodology to assess the influence of common variation in AD and PD disease susceptibility, but by their nature have reflected low effect size variants that likely have a cumulative effect on risk (459). As next-generation sequencing technology becomes more cost-effective, the ability to identify variants that are less common (<1% minor allele frequency) will become more achievable. These unbiased approaches should aid the identification of key players in the inflamm-aging pathway and will play a critical role in the development of therapeutic intervention strategies in neurodegenerative and age-related diseases.
There is the increasing opportunity to link large global datasets with the technologies of genomics, transcriptomics, and proteomics through bioinformatics and artificial intelligence methods to unlock the physiological, genetic, and molecular pathways that underpin the pro-inflammatory aging-phenotype. Using systems biology methods has the potential to lead to the generation of novel therapeutic approaches for old diseases and modern health challenges. Improving knowledge about how to delay or modify the pro-inflammatory aging-phenotype, the hallmark of aging and age-related disease, will give hope of a better quality aging and the longevity dividend for all.
Author Contributions
IR conceived and designed the outline of the manuscript. All authors IR, DG, VM, SM, DA, and OR contributed to the manuscript draft. All authors contributed to the drafting and revising of the manuscript and approved the manuscript prior to submission.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
VM, DG, and DA were supported by £11.5M grant awarded to Professor Tony Bjourson from European Union Regional Development Fund EU Sustainable Competitiveness Programme for N. Ireland; Northern Ireland Public Health Agency (HSC R&D) & Ulster University and a project supported by the European Union’s INTERREG VA Programme, managed by the Special EU Programmes Body. Interreg grant number is IVA 5306. OR receives support from the Mayo Clinic Center of Individualized Medicine. IMR was funded in part by EU Socrates Erasmus Programme for Thematic Network, Interfacing Science, Literature and Humanities ACUME2 (227942-CP-1-2006-1-IT-ERASMUS-TN2006-2371/001 SO2-23RETH), LSH-2002-2.1.4-1—Genetic factors of longevity and healthy ageing, Atlantic Philanthropies, Changing Ageing Partnership Grant, Queens Foundation Trust (R9158PHM) (IMR), Wellcome Trust Project Grant (045519/Z/95/Z) (IMR), Eastern Health and Social Care Board Research Fellowship Grant (IMR) and Belfast Trust Fund (Research and Education into Ageing (0-132) (IMR). IR thanks the nonagenarians from the BELFAST study who enthusiastically engaged in the Super Vivere and Beyond 90 Together projects.
References
1. Ter Horst R, Jaeger M, Smeekens SP, Oosting M, Swertz MA, Li Y, et al. Host and environmental factors influencing individual human cytokine responses. Cell (2016) 167(4):1111e–24e. doi:10.1016/j.cell.2016.10.018
2. Govindaraju D, Atzmon G, Barzilai N. Genetics, lifestyle and longevity: lessons from centenarians. Appl Transl Genom (2015) 4:23–32. doi:10.1016/j.atg.2015.01.001
3. Rea JNM, Carvalho A, McNerlan SE, Alexander HD, Rea IM. Genes and life-style factors in BELFAST nonagenarians: nature, nurture and narrative. Biogerontology (2015) 16(5):587–97. doi:10.1007/s10522-015-9567-y
4. Liu Y-Z, Wang Y-X, Jiang C-L. Inflammation: the common pathway of stress-related diseases. Front Hum Neurosci (2017) 11:316. doi:10.3389/fnhum.2017.00316
5. Abe K, Hashimoto Y, Yatsushiro S, Yamamura S, Bando M, Hiroshima Y, et al. Simultaneous immunoassay analysis of plasma IL-6 and TNF-α on a microchip. PLoS One (2013) 8(1):e53620. doi:10.1371/journal.pone.0053620
6. Battle A, Khan Z, Wang SH, Mitrano A, Ford MJ, Pritchard JK, et al. Genomic variation. Impact of regulatory variation from RNA to protein. Science (2015) 347(6222):664–647. doi:10.1126/science.1260793
7. Kubiczkova L, Sedlarikova L, Hajek R, Sevcikova S. TGF-beta – an excellent servant but a bad master. J Transl Med (2012) 10:183. doi:10.1186/1479-5876-10-183
8. Franceschi C, Bonafè M, Valensin S, Olivieri F, De Luca M, Ottaviani E, et al. Inflamm-aging. An evolutionary perspective on immunosenescence. Ann N Y Acad Sci (2000) 908:244–54. doi:10.1111/j.1749-6632.2000.tb06651.x
9. Franceschi C, Capri M, Monti D, Giunta S, Olivieri F, Sevini F, et al. Inflammaging and antiinflammaging: a systemic perspective on aging and longevity emerged from studies in humans. Mech Ageing Dev (2007) 128:92–105. doi:10.1016/j.mad.2006.11.016
10. Medzhitov R. Origin and physiological role of inflammation. Nature (2008) 454:428–35. doi:10.1038/nature07201
11. Franceschi C, Campisi J. Chronic inflammation (inflammaging) and its potential contribution to age-associated diseases. J Gerontol A Biol Sci Med Sci (2014) 69(Suppl 1):S4–9. doi:10.1093/gerona/glu057
12. Chung HY, Cesari M, Anton S, Marzetti E, Giovannini S, Seo AY, et al. Molecular inflammation: underpinnings of aging and age-related diseases. Ageing Res Rev (2009) 8(1):18–30. doi:10.1016/j.arr.2008.07.002
13. Martin SJ. Cell death and inflammation: the case for IL-1 family cytokines as the canonical DAMPs of the immune system. FEBS J (2016) 283(14):2599–615. doi:10.1111/febs.13775
14. Kumar V, Abbas A, Aster J. Robbins & Cotran Pathologic Basis of Disease. IX ed. Amsterdam: Elsevier (2014).
15. Nathan C. Points of control in inflammation. Nature (2002) 420(6917):846–52. doi:10.1038/nature01320
16. Chiurchiù V, Leuti A, Maccarrone M. Bioactive lipids and chronic inflammation: managing the fire within. Front Immunol (2018) 9:38. doi:10.3389/fimmu.2018.00038
17. Serhan CN, Chiang N, Dalli J, Levy BD. Lipid mediators in the resolution of inflammation. Cold Spring Harb Perspect Biol (2015) 7(2):a016311. doi:10.1101/cshperspect.a016311
18. Serhan CN, Chiang N, Dalli J. New pro-resolving n-3 mediators bridge resolution of infectious inflammation to tissue regeneration. Mol Aspects Med (2017). doi:10.1016/j.mam.2017.08.002
19. Serhan CN. Pro-resolving lipid mediators are leads for resolution physiology. Nature (2014) 510(7503):92–101. doi:10.1038/nature134
20. Awad F, Assrawi E, Jumeau C, Georgin-Lavialle S, Cobret L, Duquesnoy P, et al. Impact of human monocyte and macrophage polarization on NLR expression and NLRP3 inflammasome activation. PLoS One (2017) 12(4):e0175336. doi:10.1371/journal.pone.0175336
21. Mantovani A, Biswas SK, Galdiero MR, Sica A, Locati M. Macrophage plasticity and polarization in tissue repair and remodelling. J Pathol (2013) 229:176–85. doi:10.1002/path.4133
22. Kittan NA, Allen RM, Dhaliwal A, Cavassani KA, Schaller M, Gallagher KA, et al. Cytokine induced phenotypic and epigenetic signatures are key to establishing specific macrophage phenotypes. PLoS One (2013) 8(10):e78045. doi:10.1371/journal.pone.0078045
23. Chiurchiù V, Leuti A, Dalli J, Jacobsson A, Battistini L, Maccarrone M, et al. Proresolving lipid mediators resolvin D1, resolvin D2, and maresin 1 are critical in modulating T cell responses. Sci Transl Med (2016) 8:353ra111.1. doi:10.1126/scitranslmed.aaf7483
24. Krishnamoorthy N, Burkett PR, Dalli J, Abdulnour RE, Colas R, Ramon S, et al. Cutting edge: maresin-1 engages regulatory T cells to limit type 2 innate lymphoid cell activation and promote resolution of lung inflammation. J Immunol (2015) 194(3):863–7. doi:10.4049/jimmunol.1402534
25. Ramon S, Gao F, Serhan CN, Phipps RP. Specialized proresolving mediators enhance human B cell differentiation to antibody-secreting cells. J Immunol (2012) 189:1036–42. doi:10.4049/jimmunol.1103483
26. Recchiuti A, Serhan CN. Pro-resolving lipid mediators (SPMs) and their actions in regulating miRNA in novel resolution circuits in inflammation. Front Immunol (2012) 3:298. doi:10.3389/fimmu.2012.00298
27. Fredman G, Hellmann J, Proto JD, Kuriakose G, Colas RA, Dorweiler B, et al. An imbalance between specialized pro-resolving lipid mediators and pro-inflammatory leukotrienes promotes instability of atherosclerotic plaques. Nat Commun (2016) 7:2859. doi:10.1038/ncomms1285
28. Spite M, Norling LV, Summers L, Spite M, Norling LV, Summers L, et al. Resolvin D2 is a potent regulator of leukocytes and controls microbial sepsis. Nature (2009) 461(7268):1287–91. doi:10.1038/nature08541
29. Buckley CD, Gilroy DW, Serhan CN. Pro-Resolving lipid mediators and mechanisms in the resolution of acute inflammation. Immunity (2014) 40(3):315–27. doi:10.1016/j.immuni.2014.02.009
30. Arita M, Ohira T, Sun YP, Elangovan S, Chiang N, Serhan CN. Resolvin E1 selectively interacts with leukotriene B4 receptor BLT1 and ChemR23 to regulate inflammation. J Immunol (2007) 178:3912–7. doi:10.4049/jimmunol.178.6.3912
31. Liao Z, Dong J, Wu W, Yang T, Wang T, Guo L, et al. Resolvin D1 attenuates inflammation in lipopolysaccharide-induced acute lung injury through a process involving the PPARgamma/NF-kappaB pathway. Respir Res (2012) 13:110. doi:10.1186/1465-9921-13-110
32. Chatterjee A, Sharma A, Chen M, Toy R, Mottola G, Conte MS. The pro-resolving lipid mediator maresin 1 (MaR1) attenuates inflammatory signaling pathways in vascular smooth muscle and endothelial cells. PLoS One (2014) 9(11):e113480. doi:10.1371/journal.pone.0113480
33. Marcon R, Bento AF, Dutra RC, Bicca MA, Leite DF, Calixto JB. Maresin 1, a proresolving lipid mediator derived from omega-3 polyunsaturated fatty acids, exerts protective actions in murine models of colitis. J Immunol (2013) 191:4288–98. doi:10.4049/jimmunol.1202743
34. Perretti M, Norling LV. Actions of SPM in regulating host responses in arthritis. Mol Aspects Med (2017) 58:57–64.1. doi:10.1016/j.mam.2017.04.005
35. Sulciner ML, Serhan CN, Gilligan MM, Mudge DK, Chang J, Gartung A, et al. Resolvins suppress tumor growth and enhance cancer therapy. J Exp Med (2018) 215(1):115–40. doi:10.1084/jem.20170681
36. Fiala M, Terrando N, Dalli J. Specialized pro-resolving mediators from omega-3 fatty acids improve amyloid-β phagocytosis and regulate inflammation in patients with minor cognitive impairment. J Alzheimers Dis (2015) 48:293–301.1. doi:10.3233/JAD-150367
37. Zhu M, Wang X, Hjorth E, Colas RA, Schroeder L, Granholm A-C, et al. Pro-resolving lipid mediators improve neuronal survival and increase Aβ42 phagocytosis. Mol Neurobiol (2016) 53:2733–49. doi:10.1007/s12035-015-9544-0
38. Skoldstam L, Hagfors L, Johansson G. An experimental study of a Mediterranean diet intervention for patients with rheumatoid arthritis. Ann Rheum Dis (2003) 62(3):208–14. doi:10.1136/ard.62.3.208
39. Fung TT, Rexrode KM, Mantzoros CS, Manson JE, Willett WC, Hu FB. Mediterranean diet and incidence of and mortality from coronary heart disease and stroke in women. Circulation (2009) 119(8):1093–100. doi:10.1161/CIRCULATIONAHA.108.816736
40. Esposito K, Maiorino MI, Bellastella G, Chiodini P, Panagiotakos D, Giugliano D. A journey into a Mediterranean diet and type 2 diabetes: a systematic review with meta-analyses. BMJ Open (2015) 5:e008222. doi:10.1136/bmjopen-2015-008222
41. Demarin V, Lisak M, Morović S. Mediterranean diet in healthy lifestyle and prevention of stroke. Acta Clin Croat (2011) 50(1):67–77.
42. Corella D, Carrasco P, Sorlí JV, Estruch R, Rico-Sanz J, Martínez-González MÁ, et al. Mediterranean diet reduces the adverse effect of the TCF7L2-rs7903146 polymorphism on cardiovascular risk factors and stroke incidence: a randomized controlled trial in a high-cardiovascular-risk population. Diabetes Care (2013) 36:3803–11. doi:10.2337/dc13-0955
43. Harman D. Aging: a theory based on free radical and radiation chemistry. J Gerontol (1956) 11:298–300. doi:10.1093/geronj/11.3.298
44. Bedard K, Krause KH. The NOX family of ROS-generating NADPH oxidases: physiology and pathophysiology. Physiol Rev (2007) 87:245–313. doi:10.1152/physrev.00044.2005
45. Daiber A. Redox signaling (cross-talk) from and to mitochondria involves mitochondrial pores and reactive oxygen species. Biochim Biophys Acta (2010) 1797(6–7):897–906. doi:10.1016/j.bbabio.2010.01.032
46. Marcus RL, Addison O, Kidde JP, Dibble LE, Lastayo PC. Skeletal muscle fat infiltration: impact of age, inactivity, and exercise. J Nut Health Aging (2010) 14:362–6. doi:10.1007/s12603-010-0081-2
47. Rea IM. Towards ageing well: use it or lose it: exercise, epigenetics and cognition. Biogerontology (2017) 18(4):679–91. doi:10.1007/s10522-017-9719-3
48. Diot A, Morten K, Poulton J. Mitophagy plays a central role in mitochondrial ageing. Mamm Genome (2016) 27:381–95. doi:10.1007/s00335-016-9651-x
49. Sohal RS, Orr WC. The redox stress hypothesis of aging. Free Radic Biol Med (2012) 52(3):539–55. doi:10.1016/j.freeradbiomed.2011.10.445
50. Takac I, Schroder K, Brandes RP. The Nox family of NADPH oxidases: friend or foe of the vascular system? Curr Hypertens Rep (2012) 14:70–8. doi:10.1007/s11906-011-0238-3
51. Babior BM, Lambeth JD, Nauseef W. The neutrophil NADPH oxidase. Arch Biochem Biophys (2002) 397(2):342–324. doi:10.1006/abbi.2001.2642
52. Nguyen GT, Green ER, Mecsas J. Neutrophils to the ROScue: mechanisms of NADPH oxidase activation and bacterial resistance. Front Cell Infect Microbiol (2017) 7:373. doi:10.3389/fcimb.2017.00373
53. Drummond GR, Sobey CG. Endothelial NADPH oxidases: which NOX to target in vascular disease? Trends Endocrinol Metab (2014) 25(9):452–63. doi:10.1016/j.tem.2014.06.012
54. Youn J-Y, Zhang J, Zhang Y, Chen H, Liu D, Ping P, et al. Oxidative stress in atrial fibrillation: an emerging role of NADPH oxidase. J Mol Cell Cardiol (2013) 62:72–9. doi:10.1016/j.yjmcc.2013.04.019
55. Marnett LJ, Rowlinson SW, Goodwin DC, Kalgutkar AS, Lanzo CA. Arachidonic acid oxygenation by COX-1 and COX-2. Mechanisms of catalysis and inhibition. J Biol Chem (1999) 274:22903–6. doi:10.1074/jbc.274.33.22903
56. Cho K-J, Seo J-M, Kim J-H. Bioactive lipoxygenase metabolites stimulation of NADPH oxidases and reactive oxygen species. Mol Cells (2011) 32(1):1–5. doi:10.1007/s10059-011-1021-7
57. Kienhofer J, Haussler DJF, Ruckelshausen F, Muessig E, Weber K, Pimentel D, et al. Association of mitochondrial antioxidant enzymes with mitochondrial DNA as integral nucleoid constituents. FASEB J (2009) 23(7):2034–44. doi:10.1096/fj.08-113571
58. Zhou R, Yazdi AS, Menu P, Tschopp J. A role for mitochondria in NLRP3 inflammasome activation. Nature (2011) 469:221–5. doi:10.1038/nature09663
59. Tschopp J. Mitochondria: sovereign of inflammation? Eur J Immunol (2011) 41:1196–202. doi:10.1002/eji.201141436
60. Abais JM, Xia M, Zhang Y, Boini KM, Li P-L. Redox regulation of NLRP3 inflammasomes: ROS as trigger or effector? Antioxid Redox Signal (2015) 22(13):1111–29. doi:10.1089/ars.2014.5994
61. Ciechanover A. Intracellular protein degradation: from a vague idea thru the lysosome and the ubiquitin-proteasome system and onto human diseases and drug targeting. Biochim Biophys Acta (2012) 1824:3–13. doi:10.1038/sj.cdd.4401692
62. Tanida I. Autophagosome formation and molecular mechanism of autophagy. Antioxid Redox Signal (2011) 14(11):2201–14. doi:10.1089/ars.2010.3482
63. Gelino S, Hansen M. Autophagy – an emerging anti-aging mechanism. J Clin Exp Pathol (2012) Suppl 4:006.
64. Brunk UT, Terman A. The mitochondrial-lysosomal axis theory of aging. Accumulation of damaged mitochondria as a result of imperfect autophagocytosis. Eur J Biochem (2002) 269:1996–2002. doi:10.1046/j.1432-1033.2002.02869.x
65. Carrard G, Bulteau A-L, Petropoulos I, Friguet B. Impairment of proteasome structure and function in aging. Int J Biochem Cell Biol (2002) 34:1461–74. doi:10.1016/S1357-2725(02)00085-7
66. Rubinsztein DC, Marino G, Kroemer G. Autophagy and aging. Cell (2011) 146:682–95. doi:10.1016/j.cell.2011.07.030
68. Orenstein SJ, Cuervo AM. Chaperone-mediated autophagy: molecular mechanisms and physiological relevance. Semin Cell Dev Biol (2010) 21(7):719–26. doi:10.1016/j.semcdb.2010.02.005
69. Green DR, Galluzzi L, Kroemer G. Mitochondria and the autophagy-inflammation-cell death axis in organismal aging. Science (2011) 333:1109–12. doi:10.1126/science.1201940
70. Jiang P, Mizushima N. Autophagy and human diseases. Cell Res (2014) 24:69–79. doi:10.1038/cr.2013.161
71. Wolfe DM, Lee J-H, Kumar A, Lee S, Orenstein SJ, Nixon RA. Autophagy failure in Alzheimer’s disease and the role of defective lysosomal acidification. Eur J Neurosci (2013) 37:1949–61. doi:10.1111/ejn.12169
72. Nah J, Yuan J, Jung Y-K. Autophagy in neurodegenerative diseases: from mechanism to therapeutic approach. Mol Cells (2015) 38(5):381–9. doi:10.14348/molcells.2015.0034
73. Cuervo AM, Macian F. Autophagy and the immune function in aging. Curr Opin Immunol (2014) 29:97–104. doi:10.1016/j.coi.2014.05.006
74. Shi C-S, Shenderov K, Huang N-N, Kabat J, Abu-Asab M, Fitzgerald KA, et al. Activation of autophagy by inflammatory signals limits IL-1a production by targeting ubiquitinated inflammasomes for destruction. Nat Immunol (2012) 13(3):255–63. doi:10.1038/ni.2215
75. Zhong Z, Umemura A, Sanchez-Lopez E, Liang S, Shalapour S, Wong J, et al. NF-κB restricts inflammasome activation via elimination of damaged mitochondria. Cell (2016) 164(5):896–910. doi:10.1016/j.cell.2015.12.057
76. Nam T, Han JH, Sushil Devkota S, Lee H-W. Emerging paradigm of crosstalk between autophagy and the ubiquitin-proteasome system. Mol Cells (2017) 40(12):897–905. doi:10.14348/molcells.2017.0226
77. Mizushima N, Levine B, Cuervo AM, Klionsky DJ. Autophagy fights disease through cellular self-digestion. Nature (2008) 451:1069–75. doi:10.1038/nature06639
78. Blagosklonny MV. Linking calorie restriction to longevity through sirtuins and autophagy: any role for TOR. Cell Death Dis (2010) 1:e12. doi:10.1038/cddis.2009.17
79. Blagosklonny MV. Rapamycin extends life- and health span because it slows aging. Aging (Albany NY) (2013) 5(8):592–8. doi:10.18632/aging.100591
80. Zoncu R, Efeyan A, Sabatini DM. mTOR: from growth signal integration to cancer, diabetes and ageing. Nat Rev Mol Cell Biol (2011) 12:21–35. doi:10.1038/nrm3025
81. Barzilai NR. Targeting aging with metformin (TAME). Innov Aging (2017) doi:10.1093/geroni/igx004.2682
82. Vaiserman AM, Marotta F. Longevity-promoting pharmaceuticals: is it a time for implementation? Trends Pharmacol Sci (2016) 37(5):331–3. doi:10.1016/j.tips.2016.02.003
83. Fogarty S, Hardie DG. Development of protein kinase activators: AMPK as a target in metabolic disorders and cancer. Biochim Biophys Acta (2010) 1804:581–91. doi:10.1016/j.bbapap.2009.09.012
84. O’Neill LA, Hardie DG. Metabolism of inflammation limited by AMPK and pseudo-starvation. Nature (2013) 493:346–55. doi:10.1038/nature11862
85. Guma M, Wang Y, Viollet B, Liu-Bryan R. AMPK Activation by A-769662 controls IL-6 expression in inflammatory arthritis. PLoS One (2015) 10(10):e0140452. doi:10.1371/journal.pone.0140452
86. Grumati P, Coletto L, Schiavinato A, Castagnaro S, Bertaggia E, Sandri M, et al. Physical exercise stimulates autophagy in normal skeletal muscles but is detrimental for collagen VI-deficient muscles. Autophagy (2011) 7:1415–23. doi:10.4161/auto.7.12.17877
87. Yan Z, Lira VA, Greene NP. Exercise training-induced regulation of mitochondrial quality. Exerc Sport Sci Rev (2012) 40(3):159–64. doi:10.1097/JES.0b013e3182575599
88. Ferreira-Marques M, Aveleira CA, Carmo-Silva S, Botelho M, de Almeida LP, Cavadas C. Caloric restriction stimulates autophagy in rat cortical neurons through neuropeptide Y and ghrelin receptors activation. Aging (Albany NY) (2016) 8(7):1470–84. doi:10.18632/aging.100996
89. Campisi J. Aging, cellular senescence, and cancer. Annu Rev Physiol (2013) 75:685–705. doi:10.1146/annurev-physiol-030212-183653
90. Hayflick L, Moorhead PS. The serial cultivation of human diploid cell strains. Exp Cell Res (1961) 25:585–621. doi:10.1016/0014-4827(61)90192-6
91. Muñoz-Espín D, Cañamero M, Maraver A, Gómez-López G, Contreras J, Murillo-Cuesta S, et al. Programmed cell senescence during mammalian embryonic development. Cell (2013) 155:1104–11. doi:10.1016/j.cell.2013.10.019
92. Storer M, Mas A, Robert-Moreno A, Pecoraro M, Ortells MC, Di Giacomo V, et al. Senescence is a developmental mechanism that contributes to embryonic growth and patterning. Cell (2013) 155(5):1119–30. doi:10.1016/j.cell.2013.10.041
93. McHugh D, Gil J. Senescence and aging: causes, consequences, and therapeutic avenues. J Cell Biol (2018) 217(1):65–77. doi:10.1083/jcb.201708092
94. Wiley CD, Campisi J. From ancient pathways to aging cells—connecting metabolism and cellular senescence. Cell Metab (2016) 23(6):1013–21. doi:10.1016/j.cmet.2016.05.010
95. Serrano M. Senescence helps regeneration. Dev Cell (2014) 31(6):671–2. doi:10.1016/j.devcel.2014.12.007
96. Bodnar AG, Ouellette M, Frolkis M, Holt SE, Chiu CP, Morin GB, et al. Extension of life-span by introduction of telomerase into normal human cells. Science (1998) 279(5349):349–52. doi:10.1126/science.279.5349.349
97. Sun N, Youle RJ, Finkel T. The mitochondrial basis of aging. Mol Cell (2016) 61(5):654–66. doi:10.1016/j.molcel.2016.01.028
98. Dimri GP, Lee X, Basile G, Acosta M, Scott G, Roskelley C, et al. A biomarker that identifies senescent human cells in culture and in aging skin in vivo. Proc Natl Acad Sci U S A (1995) 92(20):9363–7. doi:10.1073/pnas.92.20.9363
99. Jeyapalan JC, Sedivy JM. Cellular senescence and organismal aging. Mech Ageing Dev (2008) 129(7–8):467–74. doi:10.1016/j.mad.2008.04.001
100. Krizhanovsky V, Yon M, Dickins RA, Hearn S, Simon J, Miething C, et al. Senescence of activated stellate cells limits liver fibrosis. Cell (2008) 134(4):657–67. doi:10.1016/j.cell.2008.06.049
101. Erusalimsky JD, Kurz DJ. Cellular senescence in vivo: its relevance in ageing and cardiovascular disease. Exp Gerontol (2005) 40(8–9):634–42. doi:10.1016/j.exger.2005.04.010
102. Chalan P, van den Berg A, Kroesen B-J, Brouwer L, Boots A. Rheumatoid arthritis, immunosenescence and the hallmarks of aging. Curr Aging Sci (2015) 8(2):131–46. doi:10.2174/1874609808666150727110744
103. Chinta SJ, Woods G, Rane A, Demaria M, Campisi J, Andersen JK. Cellular senescence and the aging brain. Exp Gerontol (2015) 68:3–7. doi:10.1016/j.exger.2014.09.018
104. Campisi J, Andersen J, Kapahi P, Melov S. Cellular senescence: a link between cancer and age-related degenerative disease? Semin Cancer Biol (2011) 21(6):354–9. doi:10.1016/j.semcancer.2011.09.001
105. Ghosh K, Capell BC. The senescence-associated secretory phenotype: critical effector in skin cancer and aging. J Invest Dermatol (2016) 136(11):2133–9. doi:10.1016/j.jid.2016.06.621
106. Coppé JP, Desprez PY, Krtolica A, Campisi J. The senescence-associated secretory phenotype: the dark side of tumor suppression. Annu Rev Pathol (2010) 5:99–118. doi:10.1146/annurev-pathol-121808-102144
107. Kuilman T, Michaloglo C, Mooi WJ, Peeper DS. The essence of senescence. Genes Dev (2010) 24:2463–79. doi:10.1101/gad.1971610
108. Rodier F, Campisi J. Four faces of cellular senescence. J Cell Biol (2011) 192(4):547–56. doi:10.1083/jcb.201009094
109. Ovadya Y, Krizhanovsky V. Senescent cells: SASPected drivers of age-related pathologies. Biogerontology (2014) 15:627–42. doi:10.1007/s10522-014-9529-9
110. Freund A, Orjalo AV, Desprez PY, Campisi J. Inflammatory networks during cellular senescence: causes and consequences. Trends Mol Med (2010) 16(5):238–46. doi:10.1016/j.molmed.2010.03.003
111. Salminen A, Kauppinen A, Kaarniranta K. Emerging role of NF-κB signaling in the induction of senescence-associated secretory phenotype (SASP). Cell Signal (2012) 24(4):835–45. doi:10.1016/j.cellsig.2011.12.006
112. Fulop T, Witkowski JM, Le Page A, Fortin C, Pawelec G, Larbi A. Intracellular signalling pathways: targets to reverse immunosenescence. Clin Exp Immunol (2017) 187:35–43. doi:10.1111/cei.12836
113. Deschênes M, Chabot B. The emerging role of alternative splicing in senescence and aging. Aging Cell (2017) 16:918–33. doi:10.1111/acel.12646
114. Nelson G, Wordsworth J, Wang C, Jurk D, Lawless C, Martin-Ruiz C, et al. A senescent cell bystander effect: senescence-induced senescence. Aging Cell (2012) 11:345–9. doi:10.1111/j.1474-9726.2012.00795.x
115. Watanabe S, Kawamoto S, Ohtani N, Hara E. Impact of senescence-associated secretory phenotype and its potential as a therapeutic target for senescence-associated diseases. Cancer Sci (2017) 108(4):563–9. doi:10.1111/cas.13184
116. Hongo A, Okumura N, Nakahara M, Kay EP, Koizumi N. The Effect of a p38 mitogen-activated protein kinase inhibitor on cellular senescence of cultivated human corneal endothelial cells. Invest Ophthalmol Vis Sci (2017) 58(9):3325–34. doi:10.1167/iovs.16-21170
117. Laberge RM, Zhou L, Sarantos MR, Rodier F, Freund A, de Keizer PL, et al. Glucocorticoids suppress selected components of the senescence-associated secretory phenotype. Aging Cell (2012) 11:569–78. doi:10.1111/j.1474-9726.2012.00818.x
118. Akbar AN. The convergence of senescence and nutrient sensing during lymphocyte ageing. Clin Exp Immunol (2016) 187:4–5. doi:10.1111/cei.12876
119. Henson SM, Lanna A, Riddell NE, Franzese O, Macaulay R, Griffiths SJ, et al. p38 signaling inhibits mTORC1-independent autophagy in senescent human CD8? T cells. J Clin Invest (2014) 124(9):4004–16. doi:10.1172/JCI75051
120. Hoenicke L, Zender L. Immune surveillance of senescent cells – biological significance in cancer- and non-cancer pathologies. Carcinogenesis (2012) 33(6):1123–6. doi:10.1093/carcin/bgs124
121. Kang TW, Yevsa T, Woller N, Hoenicke L, Wuestefeld T, Dauch D, et al. Senescence surveillance of pre-malignant hepatocytes limits liver cancer development. Nature (2011) 479(7374):547–51. doi:10.1038/nature10599
122. McElhaney JE, Effros RB. Immunosenescence: what does it mean to health outcomes in older adults? Curr Opin Immunol (2009) 21:418–24. doi:10.1016/j.coi.2009.05.023
123. Krishnamurthy J, Torrice C, Ramsey MR, Kovalev GI, Al-Regaiey K, Su L, et al. Ink4a/Arf expression is a biomarker of aging. J Clin Invest (2004) 114(9):1299–307. doi:10.1172/JCI22475
124. Xue W, Zender L, Miething C, Dickins RA, Hernando E, Krizhanovsky V, et al. Senescence and tumour clearance is triggered by p53 restoration in murine liver carcinomas. Nature (2007) 445(7128):656–60. doi:10.1038/nature05529
125. Chien Y, Scuoppo C, Wang X, Fang X, Baigley B, Bolden JE, et al. Control of the senescence-associated secretory phenotype by NF-κB promotes senescence and enhances chemosensitivity. Genes Dev (2011) 25:2125–36. doi:10.1101/gad.17276711
126. Baker DJ, Wijshake T, Tchkonia T, LeBrasseur NK, Childs BG, van de Sluis B, et al. Clearance of p16Ink4a-positive senescent cells delays ageing-associated disorders. Nature (2011) 479(7372):232–6. doi:10.1038/nature10600
127. Baker DJ, Childs BG, Durik M, Wijers ME, Sieben CJ, Zhong J, et al. Naturally occurring p16Ink4a-positive cells shorten healthy lifespan. Nature (2016) 530(7589):184–9. doi:10.1038/nature16932
128. Velarde MC, Demaria M, Campisi J. Senescent cells and their secretory phenotype as targets for cancer therapy. Interdiscip Top Gerontol (2013) 38:17–27. doi:10.1159/000343572
129. Cruz CM, Rinna A, Forman HF, Ventura ALM, Persechini PM, Ojcius DM. ATP activates a reactive oxygen species-dependent oxidative stress response and secretion of proinflammatory cytokines in macrophages. J Biol Chem (2007) 282:2871–9. doi:10.1074/jbc.M608083200
130. Chen G, Shaw MH, Kim YG, Nunez G. NOD-like receptors; role in innate immunity and inflammatory disease. Annu Rev Pathol (2009) 4:365–98. doi:10.1146/annurev.pathol.4.110807.092239
131. Schroder K, Tschopp J. The inflammasomes. Cell (2010) 140:821–32. doi:10.1016/j.cell.2010.01.040
132. Kinoshita T, Imamura R, Kushiyama H, Suda T. NLRP3 mediates NF-κB activation and cytokine induction in microbially induced and sterile inflammation. PLoS One (2015) 10(3):e0119179. doi:10.1371/journal.pone.0119179
133. Youm YH, Grant RW, McCabe LR, Albarado DC, Nguyen KY, Ravussin A, et al. Canonical Nlrp3 inflammasome links systemic low-grade inflammation to functional decline in aging. Cell Metab (2013) 18:519–32. doi:10.1016/j.cmet.2013.09.015
134. Payne BAI, Chinnery PF. Mitochondrial dysfunction in aging: much progress but many unresolved questions. Biochim Biophys Acta (2015) 1847(11):1347–53. doi:10.1016/j.bbabio.2015.05.022
135. Fang C, Wei X, Wei Y. Mitochondrial DNA in the regulation of innate immune responses. Protein Cell (2016) 7(1):11–6. doi:10.1007/s13238-015-0222-9
136. Collins LV, Hajizadeh S, Holme E, Jonsson IM, Tarkowski A. Endogenously oxidized mitochondrial DNA induces in vivo and in vitro inflammatory responses. J Leukoc Biol (2004) 7(5):995–1000. doi:10.1189/jlb.0703328
137. Ding Z, Liu S, Wang X, Khaidakov M, Dai Y, Mehta JL. Oxidant stress in mitochondrial DNA damage, autophagy and inflammation in atherosclerosis. Sci Rep (2013) 3:1077. doi:10.1038/srep01077
138. Escames G, López LC, Garcıa JA, Garcıa-Corzo L, Ortiz F, Acun ~a-Castroviejo D. Mitochondrial DNA and inflammatory diseases. Hum Genet (2012) 131:161–73. doi:10.1007/s00439-011-1057-y
139. Salminen A, Ojala J, Kaarniranta K, Kauppinen A. Mitochondrial dysfunction and oxidative stress activate inflammasomes: impact on the aging process and age-related diseases. Cell Mol Life Sci (2012) 69(18):2999–3013. doi:10.1007/s00018-012-0962-0
140. Mathew A, Lindsley TA, Sheridan A, Bhoiwala DL, Hushmendy SF, Yager EJ, et al. Degraded mitochondrial DNA is a newly identified subtype of the damage associated molecular pattern (DAMP) family and possible trigger of neurodegeneration. J Alzheimers Dis (2012) 30:617–27. doi:10.3233/JAD-2012-120145
141. Scherz-Shouval R, Elazar Z. Regulation of autophagy by ROS: physiology and pathology. Trends Biochem Sci (2011) 36:30–8. doi:10.1016/j.tibs.2010.07.007
142. Rajamaki L, Lappalainen J, Oorni K, Vallimaki E, Matikanien S, Kovanen PT, et al. Cholesterol crystals activate the NLRP3 inflammasome in human macrophages: a novel link between cholesterol metabolism and inflammation. PLoS One (2010) 5:e11765. doi:10.1371/journal.pone.0011765
143. Masters SL, O’Nelll LA. Disease-associated amyloid and misfolded protein aggregrates activate the inflammasome. Trends Mol Med (2011) 17:276–82. doi:10.1016/j.molmed.2011.01.005
144. Wen H, Fris D, Lei Y, Jha S, Zhang L, Huang MTH, et al. Fatty acid induced NLRP-3AS inflammasome activation interferes with insulin signaling. Nat Immunol (2011) 12:408–15. doi:10.1038/ni.2022
145. Mori MA, Bezy O, Kahn CR. Metabolic Syndrome: is Nlrp3 inflammasome a trigger or a target of insulin resistance? Circ Res (2011) 108(10):1160–2. doi:10.1161/RES.0b013e318220b57b
146. Vandanmagsar B, Youm YH, Ravussin A, Galgani JE, Stadler K, Mynatt RL, et al. The NLRP3 inflammasome instigates obesity-induced inflammation and insulin resistance. Nat Med (2011) 17(2):179–88. doi:10.1038/nm.2279
147. Schroder K, Zhou R, Tschopp J. The NLRP3 inflammasome: a sensor for metabolic danger? Science (2010) 327:296–300. doi:10.1126/science.1184003
148. Forsey RJ, Thompson JM, Ernerudh J, Hurst TL, Strindhall J, Johnsson B, et al. Plasma cytokine profiles in elderly humans. Mech Ageing Devel (2003) 124:487–93. doi:10.1016/S0047-6374(03)00025-3
149. Ferrucci L, Harris TB, Guralnik JM, Tracy RP, Corti MC, Cohen HJ, et al. Serum IL-6 level and the development of disability in older persons. J Am Geriatr Soc (1999) 47(6):639–46. doi:10.1111/j.1532-5415.1999.tb01583.x
150. Harris TB, Ferrucci L, Tracy RP, Corti MR, Wacholder S, Ettinger WH Jr, et al. Associations of elevated interleukin-6 and C-reactive protein levels with mortality in the elderly. Am J Med (1999) 106(5):506–12. doi:10.1016/S0002-9343(99)00066-2
151. Wei J, Xu H, Davies JL, Hemmings JP. Increase of plasma IL-6 concentration with age in healthy subjects. Life Sci (1992) 51(25):1953–6. doi:10.1016/0024-3205(92)90112-3
152. Sims JE, Smith DE. The IL-1 family: regulators of immunity. Nat Rev Immunol (2010) 10:89–102. doi:10.1038/nri2691
153. Cavallone L, Bonafe M, Olivieri F, Cardelli M, Marchegiani F, Giovagnetti S, et al. The role of IL-1 gene cluster in longevity: a study in Italian population. Mech Ageing Dev (2003) 124:533–8. doi:10.1016/S0047-6374(03)00033-2
154. Di Iorio A, Ferrucci L, Sparvieri E, Cherubini A, Volpato S, Corsi A, et al. Serum IL-1beta levels in health and disease: a population based study. “The InCHIANTI Study”. Cytokine (2003) 22:198–205. doi:10.1016/S1043-4666(03)00152-2
155. Sansoni P, Vescovini R, Fagnoni F, Biasini C, Zanni F, Zanlari L, et al. The immune system in extreme longevity. Exp Gerontol (2008) 43(2):61–5. doi:10.1016/j.exger.2007.06.008128-
156. Jylha M, Paavilainen P, Lehtimäki T, Goebeler S, Karhunen PJ, Hervonen A, et al. Interleukin-1 receptor antagonist, interleukin-6, and C-reactive protein as predictors of mortality in nonagenarians: the vitality 90+ study. J Gerontol A Biol Sci Med Sci (2007) 62(9):1016–21. doi:10.1093/gerona/62.9.1016
157. Zhou L, Cai J, Liu G, Wei Y, Tang H. Associations between interleukin-1 gene polymorphisms and coronary heart disease risk: a meta-analysis. PLoS One (2012) 7(9):e45641. doi:10.1371/journal.pone.0045641
158. Mun M-J, Kim J-H, Choi J-Y, Jang W-C. Genetic polymorphisms of interleukin genes and the risk of Alzheimer’s disease: an update meta-analysis. Meta Gene (2016) 8:1–10. doi:10.1016/j.mgene.2016.01.001
159. Trompet S, de Craen AJ, Slagboom P, Shepherd J, Blauw GJ, Murphy MB, et al. Genetic variation in the interleukin-1 beta-converting enzyme associates with cognitive function. The PROSPER study. Brain (2008) 131(4):1069–77. doi:10.1093/brain/awn023
160. Mrak RE, Griffin WS. Interleukin-1 and the immunogenetics of Alzheimer disease. J Neuropathol Exp Neurol (2000) 59(6):471–6. doi:10.1093/jnen/59.6.471
161. Langdahl BL, Lokke E, Carstens M, Stenkjaer LL, Eriksen EF. Osteoporotic fractures are associated with an 86-base pair repeat polymorphism in the interleukin-1 receptor antagonist gene but not with polymorphisms in the interleukin-1beta gene. J Bone Miner Res (2000) 15(3):402–14. doi:10.1359/jbmr.2000.15.3.402
162. Wang XY, Hurme M, Jylha M, Hervonen A. Lack of association between human longevity and polymorphisms of IL-1 cluster, IL-6, IL-10 and TNF-alpha genes in Finnish nonagenarians. Mech Ageing Dev (2001) 123:29–38. doi:10.1016/S0047-6374(01)00338-4
163. Cederholm T, Persson M, Andersson P, Stenvinkel P, Nordfors L, Madden J, et al. Polymorphisms in cytokine genes influence long-term survival differently in elderly male and female patients. J Intern Med (2007) 262:215–23. doi:10.1111/j.1365-2796.2007.01803.x
164. Dinarello CA, Simon A, van der Meer JWM. Treating inflammation by blocking interleukin-1 in a broad spectrum of diseases. Nat Rev Drug Discov (2012) 11:633–52. doi:10.1038/nrd3800
165. Smith DE. The biological paths of IL-1 family members IL-18 and IL-33. J Leukoc Biol (2011) 89(3):383–92. doi:10.1189/jlb.0810470
166. Dinarello CA, Novick D, Kim S, Kaplanski G. Interleukin-18 and IL-18 binding protein. Front Immunol (2013) 4:289. doi:10.3389/fimmu.2013.00289
167. Gangemi S, Basile G, Merendino RA, Minciullo PL, Novick D, Rubinstein M, et al. Increased circulating interleukin-18 levels in centenarians with no signs of vascular disease: another paradox of longevity? Exp Gerontol (2003) 38:669–72. doi:10.1016/S0531-5565(03)00061-5
168. Dinarello CA. Interleukin 1 and interleukin 18 as mediators of inflammation and the aging process. Am J Clin Nutr (2006) 83(2):447S–55S. doi:10.1093/ajcn/83.2.447S
169. Mallat Z, Heymes C, Corbaz A, Logeart D, Alouani S, Cohen-Solal A, et al. Evidence of altered interleukin (IL)-18 pathway in human heart failure. FASET J (2004) 18(14):1752–4. doi:10.1096/fj.04-2426fje
170. Jefferis BJ, Papacosta O, Owen CG, Wannamethee SG, Humphries SE, Woodward M, et al. Interleukin 18 and coronary heart disease: prospective study and systematic review. Atherosclerosis (2011) 217(1):227–33. doi:10.1016/j.atherosclerosis.2011.03.015
171. Harms RZ, Yarde DN, Guinn Z, Lorenzo-Arteaga KM, Corley KP, Cabrera MS, et al. Increased expression of IL-18 in the serum and islets of type 1 diabetics. Mol Immunol (2015) 64(2):306–12. doi:10.1016/j.molimm.2014.12.012
172. Liu L, Chan C. The role of inflammasome in Alzheimer’s disease. Ageing Res Rev (2014) 15:6–15. doi:10.1016/j.arr.2013.12.007
173. Thomas K, Radiq S, Frayling T, Ebrahim S, Kumari M, Gallacher J, et al. Interleukin-18 polymorphism and physical functioning in older people: a replication study and meta-analysis. J Gerontol A Biol Sci Med Sci (2009) 64(11):1177–82. doi:10.1093/gerona/glp092
174. Mekli K, Marshall A, Nazroon J, Vanhoutte B, Pendleton N. Genetic variant of Interleukin-18 gene is associated with the frailty index in the English longitudinal study of ageing. Age Ageing (2015) 44(6):938–42. doi:10.1093/ageing/afv122
175. Frayling TM, Rafiq S, Murray A, Hurst AJ, Weedon MN, Henley W, et al. Aninterleukin-18 polymorphism is associated with reduced serum concentrations and better functioning in older people. J Gerontol A Biol Sci Med Sci (2007) 62:73–8. doi:10.1093/gerona/62.1.73
176. Doyle SL, Ozaki E, Brennan K, Humphries MM, Mulfaul K, Keaney J, et al. IL-18 attenuates experimental choroidal neovascularization as a potential therapy for wet age-related macular degeneration. Sci Transl Med (2014) 6(230):230ra44. doi:10.1126/scitranslmed.3007616
177. Ershler WB. Interleukin-6: a cytokine for gerontologists. J Am Geriatr Soc (1993) 41:176–81. doi:10.1111/j.1532-5415.1993.tb02054.x
178. Ershler WB, Keller ET. Age-associated increased interleukin-6 gene expression, late-life diseases, and frailty. Annu Rev Med (2000) 51:245–70. doi:10.1146/annurev.med.51.1.245
179. Weiss TW, Arnesen H, Seljeflot I. Components of the interleukin-6 trans-signalling system are associated with the metabolic syndrome, endothelial dysfunction and arterial stiffness. Metabolism (2013) 62(7):1008–13. doi:10.1016/j.metabol.2013.01.019
180. Puzianowska-Kuźnicka M, Owczarz M, Wieczorowska-Tobis K, Nadrowski P, Chudek J, Slusarczyk P, et al. Interleukin-6 and C-reactive protein, successful aging, and mortality: the PolSenior study. Immun Ageing (2016) 13:21. doi:10.1186/s12979-016-0076-x
181. Van Epps P, Oswald D, Higgins PA, Hornick TR, Aung H, Banks RE, et al. Frailty has a stronger association with inflammation than age in older veterans. Immun Ageing (2016) 13:27. doi:10.1186/s12979-016-0082-z
182. Varadhan R, Yao W, Matteini A, Beamer BA, Xue QL, Yang H, et al. Simple biologically informed inflammatory index of two serum cytokines predicts 10 year all-cause mortality in older adults. J Gerontol A Biol Sci Med Sci (2014) 69:165–73. doi:10.1093/gerona/glt023
183. Hubbard RE, O’Mahony MS, Savva GM, Calver BL, Woodhouse KW. Inflammation and frailty measures in older people. J Cell Mol Med (2009) 13:3103–9. doi:10.1111/j.1582-4934.2009.00733
184. Alemán H, Esparza J, Ramirez FA, Astiazaran H, Payette H. Longitudinal evidence on the association between interleukin-6 and C-reactive protein with the loss of total appendicular skeletal muscle in free-living older men and women. Age Ageing (2011) 40(4):469–75. doi:10.1093/ageing/afr040
185. Ridker PM, Rifai N, Stampfer MJ, Hennekens CH. Plasma concentration of interleukin-6 and the risk of future myocardial infarction among apparently healthy men. Circulation (2000) 101:1767–72. doi:10.1161/01.CIR.101.15.1767
186. Fishman D, Faulds G, Jeffery R, Mohamed-Ali V, Yudkin JS, Humphries S, et al. The effect of novel polymorphisms in the interleukin-6 (IL- 6) gene on IL-6 transcription and plasma IL-6 levels, and an association with systemic-onset juvenile chronic arthritis. J Clin Invest (1998) 102:1369–76. doi:10.1172/JCI2629
187. Mooijaart SP, Sattar N, Trompet S, Lucke J, Stott DJ, Ford I, et al. Circulating interleukin-6 concentration and cognitive decline in old age: the PROSPER study. J Intern Med (2013) 274:77–85. doi:10.1111/joim.12052
188. Miwa K, Okazaki S, Sakaguchi M, Mochizuki H, Kitagawa K. Interleukin-6, interleukin-6 receptor gene variant, small-vessel disease and incident dementia. Eur J Neurol (2016) 23(3):656–63. doi:10.1111/ene.12921
189. Spoto B, Mattace-Raso F, Sijbrands E, Leonardis D, Testa A, Pisano A, et al. Association of IL-6 and a functional polymorphism in the IL-6 Gene with cardiovascular events in patients with CKD. Clin J Am Soc Nephrol (2014) 10:232–40. doi:10.2215/CJN.07000714
190. Dai L, Liu D, Guo H, Wang Y, Bai Y. Association between polymorphism in the promoter region of Interleukin 6 (174 G/C) and risk of Alzheimers disease: a meta-analysis. J Neurol (2012) 259(3):414–9. doi:10.1007/s00415-011-6164-0
191. Qi H-P, Qu Z-Y, Duan S-R, Wei S-Q, Wen S-R, Bi S. IL-6-174 G/C and 572 C/G polymorphisms and risk of Alzheimer’s disease. PLoS One (2012) 7(6):e37858. doi:10.1371/journal.pone.0037858
192. Di Bona D, Vasto S, Capurso C, Christiansen L, Deiana L, Franceschi C, et al. Effect of interleukin-6 polymorphisms on human longevity: a systematic review and meta-analysis. Ageing Res Rev (2009) 8:36–42. doi:10.1016/j.arr.2008.09.001
193. Soerensen M, Dato S, Tan Q, Thinggaard M, Kleindorp R, Beekman M, et al. Evidence from case-control and longitudinal studies supports associations of genetic variation in APOE, CETP, and IL6 with human longevity. Age (2013) 35(2):487–500. doi:10.1007/s11357-011-9373-7
194. Interleukin-6 Receptor Mendelian Randomisation Analysis (IL6R MR) Consortium, Swerdlow DI, Holmes MV, Kuchenbaecker KB, Engmann JEL, Shah T, et al. The interleukin-6 receptor as a target for prevention of coronary heart disease: a Mendelian randomisation analysis. Lancet (2012) 379:1214–24. doi:10.1016/S0140-6736(12)60110-X
195. Davies R, Choy E. Clinical experience of IL-6 blockade in rheumatic diseases—implications on IL-6 biology and disease pathogenesis. Semin Immunol (2014) 26:97–104. doi:10.1016/j.smim.2013.12.002
196. Ferruci L, Corsi A, Lauretani F, Bandinelli S, Bartali B, Taub DD, et al. The origins of age-related proinflammatory state. Blood (2005) 105(6):2294–9. doi:10.1182/blood-2004-07-2599
197. McNerlan SE, Rea IM, Alexander HD. A whole blood method for measurement of intracellular TNFα, IFNγ an IL-2 expression in stimulated CD3+ lymphocytes: differences between young and elderly subjects. Exp Gerontol (2002) 37:227–37. doi:10.1016/S0531-5565(01)00188-7
198. O’Mahoney L, Holland J, Jackson J, Feighery C, Hennessy TP, Mealy K. Quantitative intracellular cytokine measurement: age-related changes in proinflammatory cytokine production. Clin Exp Immunol (1998) 113(2):213–9. doi:10.1046/j.1365-2249.1998.00641.x
199. Armstrong ME, Alexander HD, Ritchie JL, McMillan SA, Rea IM. Age-related alterations in basal expression and in vitro, tumour necrosis factor alpha mediated, upregulation of CD11b. Gerontology (2001) 47:180–5. doi:10.1159/000052795
200. Bruunsgaard H, Pedersen AN, Schroll M, Skinhoj P, Pedersen BK. Ageing, TNF-α and atherosclerosis. Clin Exp Immunol (2000) 121:255–60. doi:10.1046/j.1365-2249.2000.01281.x
201. Bruunsgaard H, Ladelund S, Pedersen AN, Schroll M, Jørgensen T, Pedersen BK. Predicting death from tumour necrosis factor-alpha and interleukin-6 in 80 year old people. Clin Exp Immunol (2003) 132:24–31. doi:10.1046/j.1365-2249.2003.02137.x
202. Bruunsgaard H, Andersen-Ranberg K, Hjelmborg JV, Pedersen BK, Jeune B. Elevated levels of tumour necrosis factor alpha and mortality in centenarians. Am J Med (2003) 115:278–83. doi:10.1016/S0002-9343(03)00329-2
203. Ridker PM, Rifai N, Pfeffer M, Sacks F, Lepage S, Braunwald E. Elevation of tumor necrosis factor-alpha and increased risk of recurrent coronary events after myocardial infarction. Circulation (2000) 101(18):2149–53. doi:10.1161/01.CIR.101.18.2149
204. Nilsson L, Szymanowski A, Swahn E, Jonasson L. Soluble TNF receptors are associated with infarct size and ventricular dysfunction in ST-elevation myocardial infarction. PLoS One (2013) 8:e55477. doi:10.1371/journal.pone.0055477
205. Bae E, Cha R-H, Kim YC, An JN, Kim DK, Yoo KD, et al. Circulating TNF receptors predict cardiovascular disease in patients with chronic kidney disease. Medicine (2017) 96(19):e6666. doi:10.1097/MD.0000000000006666
206. Zhang P, Wu X, Li G, He Q, Dai H, Ai C, et al. Tumor necrosis factor-alpha gene polymorphisms and susceptibility to ischemic heart disease: a systematic review and meta-analysis. Medicine (2017) 96(14):e6569. doi:10.1097/MD.0000000000006569
207. Wang T. TNF-alpha G308A polymorphism and the susceptibility to Alzheimer’s disease: an updated meta-analysis. Arch Med Res (2015) 46(1):24.e–30.e. doi:10.1016/j.arcmed.2014.12.006
208. McCusker SM, Curran MD, Dynan KB, McCullagh CD, Urquhart DD, Middleton D, et al. Association between polymorphism in regulatory region of gene encoding tumour necrosis factor alpha and risk of Alzheimer’s disease and vascular dementia: a case-control study. Lancet (2001) 357:436–9. doi:10.1016/S0140-6736(00)04008-3
209. Collins JS, Perry RT, Watson B Jr, Harrell LE, Acton RT, Blacker D, et al. Association of a haplotype for tumour necrosis factor in siblings with late-onset Alzheimer disease. The NIMH Alzheimer disease genetics initiative. Am J Med Genet (2000) 96:823–30. doi:10.1002/1096-8628(20001204)96:6<823::AID-AJMG26>3.0.CO;2-I
210. Zheng C, Zhou Z-W, Wang J-Z. The dual roles of cytokines in Alzheimer’s disease: update on interleukins, TNF-α, TGF-β and IFN-γ. Transl Neurodegener (2016) 5(1):7. doi:10.1186/s40035-016-0054-4
211. Nilsson J, Jovinge S, Niemann A, Reneland R, Lithell H. Relation between plasma tumour necrosis factor-α and insulin sensitivity in elderly men with non-insulin-dependent diabetes mellitus. Arterioscler Thromb Vasc Biol (1998) 18(8):1199. doi:10.1161/01.ATV.18.8.1199
212. Ross OA, Curran MD, Meenagh A, Williams F, Barnett YA, Middleton D, et al. Study of age-association with cytokine gene polymorphisms in an aged Irish population. Mech Ageing Dev (2003) 124(2):199–206. doi:10.1016/S0047-6374(02)00132-X
213. Lio D, Scola L, Crivello A, Colonna-Romano G, Candore G, Bonafè M, et al. Inflammation, genetics and longevity: further studies on the prospective effects in men of IL-10-1082 promoter SNP and its interaction with TNF-alpha-308 promoter SNP. J Med Genet (2003) 40:296–9. doi:10.1136/jmg.40.4.296
214. Ruparelia N, Chai JT, Fisher EA, Choudhury RP. Inflammatory processes in cardiovascular disease: a route to targeted therapies. Nat Rev Cardiol (2017) 14:133–44. doi:10.1038/nrcardio.2017.33
215. Shamim D, Laskowski M. Inhibition of inflammation mediated through the tumor necrosis factor α biochemical pathway can lead to favorable outcomes in Alzheimer disease. J Cent Nerv Syst Dis (2017) 9:1179573517722512. doi:10.1177/1179573517722512
216. Mosmann TR, Sad S. The expanding universe of T cell subsets: Th1, Th2 and more. Immunol Today (1996) 17:138–46. doi:10.1016/0167-5699(96)80606-2
217. Laurence A, Tato CM, Davidson S, Kano Y, Chen Z, Yao Z, et al. Interleukin-2 signaling via STAT5 constrains T helper 17 cell generation. Immunity (2009) 26(3):371–81. doi:10.1016/j.immuni.2007.02.009
218. Gillis S, Watson J. Interleukin-2 dependent culture of cytolytic T cells. Immunol Rev (1981) 54:81–109. doi:10.1111/j.1600-065X.1981.tb00435.x
219. Caruso C, Candore G, Cigna D, DiLorenzo G, Sireci G, Dieli F, et al. Cytokine production pathway in the elderly. Immunol Res (1996) 15:84–90. doi:10.1007/BF02918286
220. Rea IM, Stewart M, Campbell P, Alexander HD, Crockard AD, Morris TCM. Changes in lymphocyte subsets, interleukin 2 and soluble interleukin 2 receptor in old and very old age. Gerontology (1996) 42:69–78. doi:10.1159/000213775
221. Pietschmann P, Gollob E, Brosch S, Hahn P, Kudlacek S, Willheim M, et al. The effect of age and gender on cytokine production by human peripheral blood mononuclear cells and markers of bone metabolism. Exp Gerontol (2003) 38:1119–27. doi:10.1016/S0531-5565(03)00189-X
222. Aspinall R. T cell development, ageing and interleukin-7. Mech Ageing Dev (2006) 127:572–8. doi:10.1016/j.mad.2006.01.016
223. Nguyen V, Mendelsohn A, LarricK JW. Interleukin-7 and Immunosenescence. J Immunol Res (2017) 2017:4807853. doi:10.1155/2017/4807853
224. Passtoors WM, Boer JM, Goeman JJ, van den Akker EB, Deelen J, Zwaan BJ. Transcriptional prefiling of human familial longevity indicates a role for ASF1A and IL-7R. PLoS One (2012) 7(1):e27759. doi:10.1371/journal.pone.0027759
225. Passtoors WM, van den Akker EB, Deelen J, Maier AB, van der Breggen R, Jansen R, et al. IL7R gene expression network associates with human healthy ageing. Immun Ageing (2015) 12:21. doi:10.1186/s12979-015-0048-6
226. Rübenhagen R, Schüttrumpf JP, Stürmer KM, Frosch KH. Interleukin-7 levels in synovial fluid increase with age and MMP-1 levels decrease with progression of osteoarthritis. Acta Orthop (2012) 83(1):59–64. doi:10.3109/17453674.2011.645195
227. Zuvich RL, McCauley JL, Oksenberg JR, Sawcer SJ, De Jager PL. Consortium international multiple sclerosis genetics, haines JL. genetic variation in the IL7RA/IL7 pathway increases multiple sclerosis susceptibility. Hum Genet (2010) 127(5):525–35. doi:10.1007/s00439-010-0789-4
228. Ucar D, Márquez EJ, Chung C-H, Marches R, Rossi RJ, Uyar A, et al. The chromatin accessibility signature of human immune aging stems from CD8+ T cells. J Exp Med (2017) 214(10):3123–44. doi:10.1084/jem.20170416
229. van der Heijden T, Bot I, Kuiper J. IL-12 cytokine family in cardiovascular disease. Cytokine (2017) 816:1–2. doi:10.1016/j.cyto.2017.10.010
230. Zykov MV, Barbarash OL, Kashtalap W, Kutikhin AG, Barbarash LS. Interleukin 12 serum level has prognostic value in patients with ST-segment elevation myocardial infarction. Heart Lung (2016) 45:336–40. doi:10.1016/j.hrtlng.2016.03.007
231. Rea IM, McNerlan SE, Alexander HD. Serum IL-12 and IL12p40 but not IL-12p70 are increased in the serum of older subjects: relationship to CD3+ and NK subsets. Cytokine (2000) 12(2):156–9. doi:10.1006/cyto.1999.0537
232. Compte N, Zouaoui Boudjeltia K, Vanhaeverbeek M, De Bruecker S, Tassignon J, Trelcat A, et al. Frailty in old age is associated with decreased interleukin 12/23 production in response to toll-like receptor ligation. PLoS One (2013) 8:e65325. doi:10.1371/journal.pone.0065325
233. Tan MS, Yu JT, Jiang T, Zhu XC, Guan HS, Tan L. Il-12/23 p40 inhibition ameliorates alzheimers disease-associated neuropathology and spatial memory in SAMP8 mice. J Alzheimers Dis (2014) 38(3):633–46. doi:10.1371/journal.pone.0176760
234. O’Quinn D, Palmer M, Lee Y, Weaver C. Emergence of the Th17 pathway and its role in host defense. Adv Immunol (2008) 99:115–63. doi:10.1016/S0065-2776(08)00605-6
235. Korn T, Bettelli E, Oukka M, Kuchroo VK. IL-17 and Th17 cells. Annu Rev Immunol (2009) 27:485–517. doi:10.1146/annurev.immunol.021908.132710
236. Lee JS, Lee WW, Kim SH, Kang Y, Lee N, Shin MS, et al. Age-associated alteration in naive and memory Th17 cell response in humans. Clin Immunol (2011) 140(1):84–91. doi:10.1016/j.clim.2011.03.018
237. Gaffen SL. The role of interleukin-17 in the pathogenesis of rheumatoid arthritis. Curr Rhematol Rep (2009) 11:365–70. doi:10.1007/s11926-009-0052-y
238. Zambrano-Zaragoza JF, Romo-Martínez EJ, Durán-Avelar Mde J, García-Magallanes N, Vibanco-Pérez N. Th17 cells in autoimmune and infectious diseases. Int J Inflam (2014) 2014:651503. doi:10.1155/2014/651503
239. McKensie BS, Kastelein RA, Cua DJ. Understanding the IL-23-IL-17 immune pathway. Trends Immunol (2006) 27:17–23. doi:10.1016/j.it.2005.10.003
240. Garrett-Sinha LA, John S, Gaffen SL. IL-17 and the Th17 lineage in systemic lupus erythematosus. Curr Opin Rheumatol (2008) 20:519–25. doi:10.1007/s10067-014-2656-5
241. Dong C. Regulation and pre-inflammatory function of interleukin-17 family cytokines. Immunol Rev (2008) 226:80–6. doi:10.1111/j.1600-065X.2008.00709.x
242. Srenathan U, Steel K, Taams LS. IL-17+ CD8+ T cells: differentiation, phenotype and role in inflammatory disease. Immunol Lett (2016) 178:20–6. doi:10.1016/j.imlet.2016.05.001
243. Menon B, Gullick NJ, Walter GJ, Rajasekhar M, Garrood T, Evans HG, et al. IL-17+ CD8+ T cells are enriched in the joints of patients with psoriatic arthritis and correlate with disease activity and joint damage progression. Arthritis Rheum (2014) 66:1272–81. doi:10.1002/art.38376
244. Rink L, Cakman I, Kirchner H. Altered cytokine production in the elderly. Mech Ageing Dev (1998) 102:199–209. doi:10.1016/S0047-6374(97)00153-X
245. Wieczorowska-Tobis K, Niemir ZI, Podkówka R, Korybalska K, Mossakowska M, Bręborowicz A. Can an increased level of circulating IL-8 be a predictor of human longevity? Med Sci Monit (2006) 12:CR118–21.
246. Campbell LM, Maxwell PJ, Waugh DJ. Rationale and means to target pro-inflammatory interleukin-8 (CXCL8) signaling in cancer. Pharmaceuticals (Basel) (2013) 6(8):929–59. doi:10.3390/ph6080929
247. Ouyang W, Rutz S, Crellin NK, Valdez PA, Hymowitz SG. Regulation and functions of the IL-10 family of cytokines in inflammation and disease. Annu Rev Immunol (2011) 29:71–109. doi:10.1146/annurev-immunol-031210-101312
248. Commins S, Steinke JW, Borish L. The extended IL-10 superfamily: IL-10, IL-19, IL-20, IL-22, IL-24, IL-26, IL-28, and IL-29. J Allergy Clin Immunol (2008) 121(5):1108–11. doi:10.1016/j.jaci.2008.02.026
249. Rea IM. IL-10 production from monocyte monolayers in very old age. Immunology (1996) 89(Suppl 1):68.
250. Hirokawa K, Utsuyama M, Hayashi Y, Kitagawa M, Makinodan T, Fulop T. Slower immune system aging in women versus men in the Japanese population. Immun Ageing (2013) 10:19. doi:10.1186/1742-4933-10-19
251. Didion SP, Kinzenbaw DA, Schrader LI, Chu Y, Faraci FM. Endogenous interleukin-10 inhibits angiotensin II-induced vascular dysfunction. Hypertension (2009) 54(3):619–24. doi:10.1161/HYPERTENSIONAHA.109.137158
252. Kinzenbaw DA, Chu Y, Peña Silva RA, Didion SP, Faraci FM. Interleukin-10 protects against aging-induced endothelial dysfunction. Physiol Rep (2013) 1(6):e00149. doi:10.1002/phy2.149
253. Fichtlscherer S, Breuer S, Heeschen C, Dimmeler S, Zeiher AM. Interleukin-10 serum levels and systemic endothelial vasoreactivity in patients with coronary artery disease. J Am Coll Cardiol (2004) 44(1):44–9. doi:10.1016/j.jacc.2004.02.054
254. Lakoski SG, Liu Y, Brosnihan KB, Herrington DM. Interleukin-10 concentration and coronary heart disease (CHD) event risk in the estrogen replacement and atherosclerosis (ERA) study. Atherosclerosis (2008) 197(1):443–7. doi:10.1016/j.atherosclerosis.2007.06.033
255. Welsh P, Murray HM, Ford I, Trompet S, de Craen AJ, Jukema JW, et al. Circulating interleukin-10 and risk of cardiovascular events a prospective study in the elderly at risk. Arterioscler Thromb Vasc Biol (2011) 31(10):2338–44. doi:10.1161/ATVBAHA.111.231795
256. Pes GM, Lio D, Carru C, Deiana L, Baggio G, Franceschi C, et al. Association between longevity and cytokine gene polymorphisms. A study in Sardinian centenarians. Aging Clin Exp Res (2004) 16(3):244–8. doi:10.1007/BF03327391
257. Westendorp RG, Langermans JAM, Huizinga TWJ, Elouali AH, Verweij CL, Boomsma DI, et al. Genetic influence on cytokine production in meningococcal disease. Lancet (1997) 349:170–3. doi:10.1016/S0140-6736(96)06413-6
258. Yoshimura A, Wakabayashi Y, Mori T. Cellular and molecular basis for the regulation of inflammation by TGF-beta. J Biochem (2010) 147(6):781–92. doi:10.1093/jb/mvq043
259. Rea IM, Maxwell LD, McNerlan SE, Alexander HD, Curran MD, Middleton D, et al. Killer immunoglobulin-like receptors (KIR) haplogroups A and B track with natural killer cells and cytokine profile in aged subjects: observations from octo/nonagenarians in the belfast elderly longitudinal free-living aging study (BELFAST). Immun Ageing (2013) 10(1):35. doi:10.1186/1742-4933-10-35
260. Krieglstein K, Miyazono K, Ten Dijke P, Unsicker K. TGF-beta in aging and disease. Cell Tissue Res (2012) 347:5–9. doi:10.1007/s00441-011-1278-1273
261. Pastrana JL, Sha X, Virtue A, Mai J, Cueto R, Lee IA, et al. Regulatory T cells and atherosclerosis. J Clin Exp Cardiolog (2012) (Suppl 12):002. doi:10.4172/2155-9880.S12-002
262. Burks TN, Cohn RD. Role of TGF-β signaling in inherited and acquired myopathies. Skelet Muscle (2011) 1:19. doi:10.1186/2044-5040-1-19
263. Baugé C, Girard N, Lhuissier E, Bazille C, Boumediene K. Regulation and role of TGFβ signaling pathway in aging and osteoarthritis joints. Aging Disease (2014) 5(6):394–405. doi:10.14336/AD.2014.0500394
264. Mitnitski A, Collerton J, Martin-Ruiz C, Jagger C, von Zglinicki T, Rockwood K, et al. Age-related frailty and its association with biological markers of ageing. BMC Med (2015) 13:161. doi:10.1186/s12916-015-0400-x
265. Doyle KP, Cekanaviciute E, Mamer LE, Buckwalter MS. TGFβ signaling in the brain increases with aging and signals to astrocytes and innate immune cells in the weeks after stroke. J Neuroinflammation (2010) 7:62. doi:10.1186/1742-2094-7-62
266. Yang Q, Wang E-Y, Jia H-W, Wang Y-P. Association between polymorphisms in transforming growth factor-β1 and sporadic Alzheimer’s disease in a Chinese population. Int J Neurosci (2016) 126:979–84. doi:10.3109/00207454.2015.1088849
267. Bosco P, Ferri R, Salluzzo MG, Castellano S, Signorelli M, Nicoletti F, et al. Role of the transforming-growth-factor-β1 gene in late-onset Alzheimer’s disease: implications for the treatment. Curr Genomics (2013) 14(2):147–56. doi:10.2174/1389202911314020007
268. Mallat Z, Gojova A, Marchiol-Fournigault C, Esposito B, Kamate C, Merval R, et al. Inhibition of transforming growth factor-beta signaling accelerates atherosclerosis and induces an unstable plaque phenotype in mice. Circ Res (2001) 89(10):930–4. doi:10.1161/hh2201.099415
269. Carrieri G, Marzi E, Olivieri F, Marchegiani F, Cavallone L, Cardelli M, et al. The G/C 915 polymorphism of transforming growth factor β1 is associated with human longevity: a study in Italian centenarians. Aging Cell (2004) 3:443–8. doi:10.1111/j.1474-9728.2004.00129.x
270. Tran Dat Q. TGF-β: the sword, the wand, and the shield of FOXP3+ regulatory T cells. J Mol Cell Biol (2012) 4(1):29–37. doi:10.1093/jmcb/mjr033
271. Han G, Li F, Singh TP, Wolf P, Wan X-J. The Pro-inflammatory Role of TGFβ1: a Paradox? Int J Biol Sci (2012) 8(2):228–35. doi:10.7150/ijbs.8.228
272. Akhurst RJ, Hata A. Targeting the TGFb signaling pathway in disease. Nat Rev Drug Discov (2012) 11:790–811. doi:10.1038/nrd3810
273. Nold MF, Nold-Petry CA, Zepp JA, Palmer BE, Bufler P, Dinarello CA. IL-37 is a fundamental inhibitor of innate immunity. Nat Immunol (2010) 11(11):1014–22. doi:10.1038/ni.1944
274. Shou X, Lin J, Xie C, Wang Y, Sun C. Plasma IL-37 elevated in patients with chronic heart failure and predicted major adverse cardiac events: a 1-year follow-up study. Dis Markers (2017) 2017:9134079. doi:10.1155/2017/913407
275. Dinarello CA, Nold-Petry C, Nold M, Fujita M, Li S, Kim S, et al. Suppression of innate inflammation and immunity by interleukin-37. Eur J Immunol (2016) 46(5):1067–81. doi:10.1002/eji.20154582854
276. Xu WD, Zhao Y, Liu Y. Insights into IL-37, the role in autoimmune diseases. Autoimmun Rev (2015) 14(12):1170–5. doi:10.1016/j.autrev.2015.08.006
277. Balkwill F, Charles KA, Mantovani A. Smoldering and polarized inflammation in the initiation and promotion of malignant disease. Cancer Cell (2005) 7:211–7. doi:10.1016/j.ccr.2005.02.013
278. Coussens LM, Werb Z. Inflammation and cancer. Nature (2002) 420(6917):860–7. doi:10.1038/nature01322
279. Grivennikov SI, Greten FR, Karin M. Immunity, inflammation, and cancer. Cell (2010) 140(6):883–99. doi:10.1016/j.cell.2010.01.025
280. Kantono M, Guo B. Inflammasomes and cancer: the dynamic role of the inflammasome in tumor development. Front Immunol (2017) 8:1132. doi:10.3389/fimmu.2017.01132
281. Lin C, Zhang J. Inflammasomes in inflammation- induced cancer. Front Immunol (2017) 8:271. doi:10.3389/fimmu.2017.00271
282. Venerito M, Vasapolli R, Rokkas T, Delchier JC, Malfertheiner P. Helicobacter pylori, gastric cancer and other gastrointestinal malignancies. Helicobacter (2017) 22(Suppl):1. doi:10.1111/hel.12413
283. Vockerodt M, Cader FZ, Shannon-Lowe C, Murray P. Epstein-Barr virus and the origin of Hodgkin lymphoma. Chin J Cancer (2014) 33(12):591–7. doi:10.5732/cjc.014.10193
284. de Sanjosé S, Brotons M, Pavón MA. The natural history of human papillomavirus infection. Best Pract Res Clin Obstet Gynaecol (2017). doi:10.1016/j.bpobgyn.2017.08.015
285. Greaves MF, Verbi W, Tilley R, Lister TA, Habeshaw J, Guo HG, et al. Human T-cell leukemia virus (HTLV) in the United Kingdom. Int J Cancer (1984) 33(6):795–806. doi:10.1002/ijc.2910330614
286. Diaconu S, Predescu A, Moldoveanu A, Pop CS, Fierbinteanu-Braticevici C. Helicobacter pylori infection: old and new. J Med Life (2017) 10(2):112–7.
287. El-Omar EM, Rabkin CS, Gammon MD, Vaughan TL, Risch HA, Schoenberg JB, et al. Increased risk of noncardia gastric cancer associated with proinflammatory cytokine gene polymorphisms. Gastroenterology (2003) 124:1193–201.
288. El-Omar EM, Carrington M, Chow WH, McColl KE, Bream JH, Young HA, et al. Interleukin-1 polymorphisms associated with increased risk of gastric cancer. Nature (2000) 404:398–402. doi:10.1038/35006081
289. Iwamuro M, Takenaka R, Nakagawa M, Moritou Y, Saito S, Hori S, et al. Management of gastric mucosa-associated lymphoid tissue lymphoma in patients with extra copies of the MALT1 gene. World J Gastroenterol (2017) 23(33):6155–63. doi:10.3748/wjg.v23.i33.6155
290. Lupfer C, Malik A, Kanneganti TD. Inflammasome control of viral infection. Curr Opin Virol (2015) 12:38–46. doi:10.1016/j.coviro.2015.02.007
291. Fingeroth JD, Clabby ML, Strominger JD. Characterization of a T-lymphocyte Epstein-Barr virus/C3d receptor (CD21). J Virol (1988) 62(4):1442–7.
292. Bernheim A, Berger R, Lenoir G. Cytogenetic studies on African Burkitt’s lymphoma cell lines: t(8;14), t(2;8) and t(8;22) translocations. Cancer Genet Cytogenet (1981) 3(4):307–15. doi:10.1016/0165-4608(81)90039-X
293. Chen LC, Wang LJ, Tsang NM, Ojcius DM, Chen CC, Ouyang CN, et al. Tumour inflammasome-derived IL-1beta recruits neutrophils and improves local recurrence-free survival in EBV-induced nasopharyngeal carcinoma. EMBO Mol Med (2012) 4(12):1276–93. doi:10.1002/emmm.201201569
294. Haneklaus M, Gerlic M, Kurowska-Stolarska M, Rainey AA, Pich D, McInnes IB, et al. Cutting edge: miR-223 and EBV miR-BART15 regulate the NLRP3 inflammasome and IL-1beta production. J Immunol (2012) 189(8):3795–9. doi:10.4049/jimmunol.1200312
295. Longo DL, Gelmann EP, Cossman J, Young RA, Gallo RC, O’Brien SJ, et al. Isolation of HTLV-transformed B-lymphocyte clone from a patient with HTLV-associated adult T-cell leukaemia. Nature (1984) 310(5977):505–6. doi:10.1038/310505a0
296. Bangham CR, Toulza F. Adult T cell leukemia/lymphoma: FoxP3(+) cells and the cell-mediated immune response to HTLV-1. Adv Cancer Res (2011) 111:163–82. doi:10.1016/B978-0-12-385524-4.00004-0
297. Oliere S, Douville R, Sze A, Belgnaoui SM, Hiscott J. Modulation of innate immune responses during human T-cell leukemia virus (HTLV-1) pathogenesis. Cytokine Growth Factor Rev (2011) 22(4):197–210. doi:10.1016/j.cytogfr.2011.08.002
298. Rayet B, Gélinas C. Aberrant rel/nfkb genes and activity in human cancer. Oncogene (1999) 18(49):6938–47. doi:10.1038/sj.onc.1203221
299. Hiscott J, Kwon H, Génin P. Hostile takeovers: viral appropriation of the NF-kB pathway. J Clin Invest (2001) 107(2):143–51. doi:10.1172/JCI11918
300. Kamada AJ, Pontillo A, Guimarães RL, Loureiro P, Crovella S, Brandão LAC. NLRP3 polymorphism is associated with protection against human T-lymphotropicvirus 1 infection. Mem Inst Oswaldo Cruz (2014) 109(7):960–3. doi:10.1590/0074-0276140154
301. Wang D, DuBois RN. The role of COX-2 in intestinal inflammation and colorectal cancer. Oncogene (2010) 29(6):781–8. doi:10.1038/onc.2009.421
302. Smolen JS, Aletaha D, McInnes IB. Rheumatoid arthritis. Lancet (2016) 388:2023–38. doi:10.1016/S0140-6736(16)30173-8
303. Firestein GS, McInnes IB. Immunopathogenesis of rheumatoid arthritis. Immunity (2017) 46:183–96. doi:10.1016/j.immuni.2017.02.006
304. van de Sande MG, de Hair MJ, Schuller Y, van de Sande GP, Wijbrandts CA, Dinant HJ, et al. The features of the synovium in early rheumatoid arthritis according to the 2010 ACR/EULAR classification criteria. PLoS One (2012) 7:e36668. doi:10.1371/journal.pone.0036668
305. van der Ven M, van der Veer-Meerkerk M, Ten Cate DF, Rasappu N, Kok MR, Csakvari D, et al. Absence of ultrasound inflammation in patients presenting with arthralgia rules out the development of arthritis. Arthritis Res Ther (2017) 19:202. doi:10.1186/s13075-017-1405-y
306. Noack M, Miossec P. Selected cytokine pathways in rheumatoid arthritis. Semin Immunopathol (2017) 39:365–83. doi:10.1007/s00281-017-0619-z
307. van de Sande MG, Baeten DL. Immunopathology of synovitis: from histology to molecular pathways. Rheumatology (Oxford) (2016) 55:599–606. doi:10.1093/rheumatology/kev330
308. Bottini N, Firestein GS. Duality of fibroblast-like synoviocytes in RA: passive responders and imprinted aggressors. Nat Rev Rheumatol (2013) 9:24–33. doi:10.1038/nrrheum.2012.190
309. McInnes IB, Schett G. The pathogenesis of rheumatoid arthritis. N Engl J Med (2011) 365:2205–19. doi:10.1056/NEJMra1004965
310. Ruscitti P, Cipriani P, Di Benedetto P, Liakouli V, Berardicurti O, Carubbi F, et al. Monocytes from patients with rheumatoid arthritis and type 2 diabetes mellitus display an increased production of interleukin (IL)-1β via the nucleotide-binding domain and leucine-rich repeat containing family pyrin 3(NLRP3)-inflammasome activation: a possible implication for therapeutic decision in these patients. Clin Exp Immunol (2015) 182:35–44. doi:10.1111/cei.12667
311. Choulaki C, Papadaki G, Repa A, Kampouraki E, Kambas K, Ritis K, et al. Enhanced activity of NLRP3 inflammasome in peripheral blood cells of patients with active rheumatoid arthritis. Arthritis Res Ther (2015) 17:257. doi:10.1186/s13075-015-0775-2
312. Mathews RJ, Robinson JI, Battellino M, Wong C, Taylor JC; Biologics in Rheumatoid Arthritis Genetics and Genomics Study Syndicate (BRAGGSS), et al. Evidence of NLRP3-inflammasome activation in rheumatoid arthritis (RA); genetic variants within the NLRP3-inflammasome complex in relation to susceptibility to RA and response to anti-TNF treatment. Ann Rheum Dis (2014) 73(6):1202–10. doi:10.1136/annrheumdis-2013-203276
313. Jenko B, Praprotnik S, Tomšic M, Dolžan V. NLRP3 and CARD8 polymorphisms influence higher disease activity in rheumatoid arthritis. J Med Biochem (2016) 35:319–23. doi:10.1515/jomb-2016-0008
314. Weyand CM, Yang Z, Goronzy JJ. T cell aging in rheumatoid arthritis. Curr Opin Rheumatol (2014) 26(1):93–100. doi:10.1097/BOR.0000000000000011
315. Boots AM, Maier AB, Stinissen P, Masson P, Lories RJ, De Keyser F. The influence of ageing on the development and management of rheumatoid arthritis. Nat Rev Rheumatol (2013) 9(10):604–13. doi:10.1038/nrrheum.2013.92
316. Fessler J, Raicht A, Husic R, Ficjan A, Schwarz C, Duftner C, et al. Novel senescent regulatory T-cell subset with impaired suppressive function in rheumatoid arthritis. Front Immunol (2017) 20(8):300. doi:10.3389/fimmu.2017.00300
317. Fessler J, Husic R, Schwetz V, Lerchbaum E, Aberer F, Fasching P, et al. Senescent T-cells promote bone loss in rheumatoid arthritis. Front Immunol (2018) 1(9):95. doi:10.3389/fimmu.2018.00095
318. Catrina AI, Trollmo C, af Klint E, Engstrom M, Lampa J, Hermansson Y, et al. Evidence that anti-tumor necrosis factor therapy with both etanercept and infliximab induces apoptosis in macrophages, but not lymphocytes, in rheumatoid arthritis joints: extended report. Arthritis Rheum (2005) 52:61–72. doi:10.1002/art.20764
319. Hess A, Axmann R, Rech J, Finzel S, Heindl C, Kreitz S, et al. Blockade of TNF-alpha rapidly inhibits pain responses in the central nervous system. Proc Natl Acad Sci U S A (2011) 108:3731–6. doi:10.1073/pnas.1011774108
320. Izquierdo E, Canete JD, Celis R, Santiago B, Usategui A, Sanmarti R, et al. Immature blood vessels in rheumatoid synovium are selectively depleted in response to anti-TNF therapy. PLoS One (2009) 4:e8131. doi:10.1371/journal.pone.0008131
321. Nadkarni S, Mauri C, Ehrenstein MR. Anti-TNF-alpha therapy induces a distinct regulatory T cell population in patients with rheumatoid arthritis via TGF-beta. J Exp Med (2007) 204:33–9. doi:10.1084/jem.20061531
322. Smolen JS, Weinblatt ME, Sheng S, Zhuang Y, Hsu B. Sirukumab, a human anti-interleukin-6 monoclonal antibody: a randomised, 2-part (proof-of-concept and dose-finding), phase II study in patients with active rheumatoid arthritis despite methotrexate therapy. Ann Rheum Dis (2014) 73:1616–25. doi:10.1136/annrheumdis-2013-205137
323. Nishimoto N, Hashimoto J, Miyasaka N, Yamamoto K, Kawai S, Takeuchi T, et al. Study of active controlled monotherapy used for rheumatoid arthritis, an IL-6 inhibitor (SAMURAI): evidence of clinical and radiographic benefit from an x ray reader-blinded randomised controlled trial of tocilizumab. Ann Rheum Dis (2007) 66:1162–7. doi:10.1136/ard.2006.068064
324. Astorri E, Nerviani A, Bombardieri M, Pitzalis C. Towards a stratified targeted approach with biologic treatments in rheumatoid arthritis: role of synovial pathobiology. Curr Pharm Des (2015) 21:2216–24. doi:10.2174/1381612821666150310145758
325. Dennis G Jr, Holweg CT, Kummerfeld SK, Choy DF, Setiadi AF, Hackney JA, et al. Synovial phenotypes in rheumatoid arthritis correlate with response to biologic therapeutics. Arthritis Res Ther (2014) 16:R90. doi:10.1186/ar4555
326. Robinson WH, Lindstrom TM, Cheung RK, Sokolove J. Mechanistic biomarkers for clinical decision making in rheumatic diseases. Nat Rev Rheumatol (2013) 9:267–76. doi:10.1038/nrrheum.2013.14
327. Dar SA, Haque S, Mandal RK, Singh T, Wahid M, Jawed A, et al. Interleukin-6-174G > C (rs1800795) polymorphism distribution and its association with rheumatoid arthritis: a case-control study and meta-analysis. Autoimmunity (2017) 50:158–69. doi:10.1080/08916934.2016.1261833
328. Ferreira RC, Freitag DF, Cutler AJ, Howson JM, Rainbow DB, Smyth DJ, et al. Functional IL6R 358Ala allele impairs classical IL-6 receptor signaling and influences risk of diverse inflammatory diseases. PLoS Genet (2013) 9:e1003444. doi:10.1371/journal.pgen.1003444
329. Lopez-Lasanta M, Julia A, Maymo J, Fernandez-Gutierrez B, Urena-Garnica I, Blanco FJ, et al. Variation at interleukin-6 receptor gene is associated to joint damage in rheumatoid arthritis. Arthritis Res Ther (2015) 17:242–215. doi:10.1186/s13075-015-0737-8
330. Hussein YM, Mohamed RH, Pasha HF, El-Shahawy EE, Alzahrani SS. Association of tumor necrosis factor alpha and its receptor polymorphisms with rheumatoid arthritis in female patients. Cell Immunol (2011) 271:192–6. doi:10.1016/j.cellimm.2011.06.023
331. O’Rielly DD, Roslin NM, Beyene J, Pope A, Rahman P. TNF-alpha-308 G/A polymorphism and responsiveness to TNF-alpha blockade therapy in moderate to severe rheumatoid arthritis: a systematic review and meta-analysis. Pharmacogenomics J (2009) 9:161–7. doi:10.1038/tpj.2009.7
332. Stojanovic S, Bojana S, Stoimenov TJ, Nedovic J, Zivkovic V, Despotovic M, et al. Association of tumor necrosis factor-alpha (G-308A) genetic variant with matrix metalloproteinase-9 activity and joint destruction in early rheumatoid arthritis. Clin Rheumatol (2017) 36:1479–85. doi:10.1007/s10067-017-3699-1
333. Suarez-Gestal M, Perez-Pampin E, Calaza M, Gomez-Reino JJ, Gonzalez A. Lack of replication of genetic predictors for the rheumatoid arthritis response to anti-TNF treatments: a prospective case-only study. Arthritis Res Ther (2010) 12:R72. doi:10.1186/ar2990
334. Zeng Z, Duan Z, Zhang T, Wang S, Li G, Gao J, et al. Association between tumor necrosis factor-alpha (TNF-alpha) promoter -308 G/A and response to TNF-alpha blockers in rheumatoid arthritis: a meta-analysis. Mod Rheumatol (2013) 23:489–95. doi:10.1007/s10165-012-0699-5
335. Barnabe C, Martin BJ, Ghali WA. Systematic review and meta-analysis: anti–tumor necrosis factor α therapy and cardiovascular events in rheumatoid arthritis. Arthritis Care Res (2011) 63:522–9. doi:10.1002/acr.20371
336. Lin NY, Beyer C, Giessl A, Kireva T, Scholtysek C, Uderhardt S, et al. Autophagy regulates TNFα-mediated joint destruction in experimental arthritis. Ann Rheum Dis (2013) 72(5):761–8. doi:10.1136/annrheumdis-2012-201671
337. Kawaida R, Yamada R, Kobayashi K, Tokuhiro S, Suzuki A, Kochi Y, et al. CUL1, a component of E3 ubiquitin ligase, alters lymphocyte signal transduction with possible effect on rheumatoid arthritis. Genes Immun (2005) 6(3):194–202. doi:10.1038/sj.gene.6364177
338. Negi S, Kumar A, Thelma BK, Juyal RC. Association of cullin1 haplotype variants with rheumatoid arthritis and response to methotrexate. Pharmacogenet Genomics (2011) 21(9):590–3. doi:10.1097/FPC.0b013e3283492af7
339. Kannel WB, Vasan RS. Is age really a non-modifiable cardiovascular risk factor? Am J Cardiol (2009) 104(9):1307–10. doi:10.1016/j.amjcard.2009.06.051
340. Bolton E, Rajkumar C. The ageing cardiovascular system. Rev Clin Gerontol (2011) 21:99–109. doi:10.1017/S0959259810000389
341. Lv L, Ye M, Duan R, Yuan K, Chen J, Liang W, et al. Downregulation of Pin1 in human atherosclerosis and its association with vascular smooth muscle cell senescence. J Vasc Surg (2017). doi:10.1016/j.jvs.2017.09.006
342. Ramji DP, Davies TS. Cytokines in atherosclerosis: key players in all stages of disease and promising therapeutic targets. Cytokine Growth Factor Rev (2015) 26(6):673–85. doi:10.1016/j.cytogfr.2015.04.003
343. Moss JW, Ramji DP. Cytokines: roles in atherosclerosis disease progression and potential therapeutic targets. Future Med Chem (2016) 8(11):1317–30. doi:10.4155/fmc-2016-0072
344. McLaren JE, Michael DR, Ashlin TG, Ramji DP. Cytokines, macrophage lipid metabolism and foam cells: implications for cardiovascular disease therapy. Prog Lipid Res (2011) 50:331–47. doi:10.1016/j.plipres.2011.04.002
345. Ait-Oufella H, Taleb S, Mallat Z, Tedgui A. Recent advances on the role of cytokines in atherosclerosis. Arterioscler Thromb Vasc Biol (2011) 31:969–79. doi:10.1161/ATVBAHA.110.207415
346. Moore KJ, Sheedy FJ, Fisher EA. Macrophages in atherosclerosis: a dynamic balance. Nat Rev Immunol (2013) 13:709–21. doi:10.1038/nri3520
347. Karasawa T, Takahashi M. The crystal-induced activation of NLRP3 inflammasomes in atherosclerosis. Inflamm Regen (2017) 37:18. doi:10.1186/s41232-017-0050-9
348. Sheedy FJ, Grebe A, Rayner KJ, Kalantari P, Ramkhelawon B, Carpenter SB, et al. CD36 coordinates NLRP3 inflammasome activation by facilitating intracellular nucleation of soluble ligands into particulate ligands in sterile inflammation. Nat Immunol (2013) 14:812–20. doi:10.1038/ni.2639
349. Ridker PM, Everett BM, Thuren T, MacFadyen JG, Chang WH, Ballantyne C, et al. Anti inflammatory therapy with canakinumab for atherosclerotic disease. N Engl J Med (2017) 377(12):1119–31. doi:10.1056/NEJMoa1707914
350. Yin Y, Zhou Z, Liu W, Chang Q, Sun G, Dai Y. Vascular endothelial cells senescence is associated with NOD-like receptor family pyrin domain-containing 3 (NLRP3) inflammasome activation via reactive oxygen species (ROS)/thioredoxin-interacting protein (TXNIP) pathway. Int J Biochem Cell Biol (2017) 84:22–34. doi:10.1016/j.biocel.2017.01.001
351. Karasawa T, Takahashi M. Role of NLRP3 inflammasomes in atherosclerosis. J Atheroscler Thromb (2017) 24(5):443–51. doi:10.5551/jat.RV17001
352. Qian S, Fan J, Billiar TR, Scott MJ. Inflammasome and autophagy regulation: a two-way street. Mol Med (2017) 23:188–95. doi:10.2119/molmed.2017.00077
353. Sergin I, Evans TD, Zhang X, Bhattacharya S, Stokes CJ, Song E, et al. Exploiting macrophage autophagy-lysosomal biogenesis as a therapy for atherosclerosis. Nat Commun (2017) 8:15750. doi:10.1038/ncomms15750
354. Buford TW. (Dis)Trust your gut: the gut microbiome in age-related inflammation, health, and disease. Microbiome (2017) 5:80. doi:10.1186/s40168-017-0296-0
355. Håheim LL. The infection hypothesis revisited: oral infection and cardiovascular disease. Epidemiol Res Int (2014) 2014:735378. doi:10.1155/2014/735378
356. Tang WH, Hazen SL. The contributory role of gut microbiota in cardiovascular disease. J Clin Invest (2014) 124(10):4204–11. doi:10.1172/JCI72331
357. Renko J, Koskela KA, Lepp PW, Oksala N, Levula M, Lehtimäki T, et al. Bacterial DNA signatures in carotid atherosclerosis represent both commensals and pathogens of skin origin. Eur J Dermatol (2013) 23:53–8. doi:10.1684/ejd.2012.1908
358. Amar J, Lange C, Payros G, Garret C, Chabo C, Lantieri O, et al. Blood microbiota dysbiosis is associated with the onset of cardiovascular events in a large general population: the D.E.S.I.R. study. PLoS One (2013) 8(1):e54461. doi:10.1371/journal.pone.0054461
359. Jie Z, Xia H, Zhong S, Feng Q, Li S, Liang S, et al. The gut microbiome in atherosclerotic cardiovascular disease. Nat Commun (2017) 8(1):845. doi:10.1038/s41467-017-00900-1
360. Szeto C-C, Kwan BC-H, Chow K-M, Kwok JS-S, Lai K-B, Cheng PM-S, et al. Circulating bacterial-derived DNA fragment level is a strong predictor of cardiovascular disease in peritoneal dialysis patients. PLoS One (2015) 10(5):e0125162. doi:10.1371/journal.pone.0125162
361. Dinakaran V, Rathinavel A, Pushpanathan M, Sivakumar R, Gunasekaran P, Rajendhran J. Elevated levels of circulating dna in cardiovascular disease patients: metagenomic profiling of microbiome in the circulation. PLoS One (2014) 9(8):e105221. doi:10.1371/journal.pone.0105221
362. Thevaranjan N, Puchta A, Schulz C, Naidoo A, Szamosi JC, Verschoor CP, et al. Age-associated microbial dysbiosis promotes intestinal permeability, systemic inflammation, and macrophage dysfunction. Cell Host Microbe (2017) 21(4):455.e–66.e. doi:10.1016/j.chom.2017.03.002
363. McGeer PL, Rogers J, McGeer EG. Inflammation, antiinflammatory agents, and Alzheimer’s disease: the last 22 years. J Alzheimers Dis (2016) 54(3):853–7. doi:10.3233/JAD-160488
364. McGeer PL, McGeer EG. Inflammation and neurodegeneration in Parkinson’s disease. Parkinsonism Relat Disord (2004) 10(Suppl 1):S3–7. doi:10.1016/j.parkreldis.2004.01.005
365. Sousa C, Biber K, Michelucci A. Cellular and molecular characterization of microglia: a unique immune cell population. Front Immunol (2017) 8:198. doi:10.3389/fimmu.2017.00198
366. Song L, Pei L, Yao S, Wu Y, Shang Y. NLRP3 inflammasome in neurological diseases, from functions to therapies. Front Cell Neurosci (2017) 11:63. doi:10.3389/fncel.2017.00063
367. Maceyka M, Spiegel S. Sphingolipid metabolites in inflammatory disease. Nature (2014) 510(7503):58–67. doi:10.1038/nature13475
368. Scheiblich H, Schlütter A, Golenbock DT, Latz E, Martinez-Martinez P, Heneka MT. Activation of the NLRP3 inflammasome in microglia: the role of ceramide. J Neurochem (2017) 143(5):534–50. doi:10.1111/jnc.14225
369. Nilsson P, Saido TC. Dual roles for autophagy: degradation and secretion of Alzheimer’s disease abeta peptide. Bioessays (2014) 36:570–8. doi:10.1002/bies.201400002
370. Nilsson P, Loganathan K, Sekiguchi M, Matsuba Y, Hui K, Tsubuki S, et al. A beta secretion and plaque formation depend on autophagy. Cell Rep (2013) 5:61–9. doi:10.1016/j.celrep.2013.08.042
371. Boccardi V, Pelini L, Ercolani S, Ruggiero C, Mecocci P. From cellular senescence to Alzheimer’s disease: the role of telomere shortening. Ageing Res Rev (2015) 22:1–8. doi:10.1016/j.arr.2015.04.003
372. Chang D, Nalls MA, Hallgrímsdóttir IB, Hunkapiller J, van der Brug M, Cai F, et al. A meta-analysis of genome-wide association studies identifies 17 new Parkinson’s disease risk loci. Nat Genet (2017) 49(10):1511–6. doi:10.1038/ng.3955
373. Lambert JC, Ibrahim-Verbaas CA, Harold D, Naj AC, Sims R, Bellenguez C, et al. Meta-analysis of 74,046 individuals identifies 11 new susceptibility loci for Alzheimer’s disease. Nat Genet (2013) 45(12):1452–8. doi:10.1038/ng.2802
374. Rayaprolu S, Mullen B, Baker M, Lynch T, Finger E, Seeley WW, et al. TREM2 in neurodegeneration: evidence for association of the p.R47H variant with frontotemporal dementia and Parkinson’s disease. Mol Neurodegener (2013) 8:19. doi:10.1186/1750-1326-8-19
375. Jonsson T, Stefansson H, Steinberg S, Jonsdottir I, Jonsson PV, Snaedal J, et al. Variant of TREM2 associated with the risk of Alzheimer’s disease. N Engl J Med (2013) 368(2):107–16. doi:10.1056/NEJMoa1211103
376. Guerreiro R, Wojtas A, Bras J, Carrasquillo M, Rogaeva E, Majounie E, et al. TREM2 variants in Alzheimer’s disease. N Engl J Med (2013) 368(2):117–27. doi:10.1056/NEJMoa1211851
377. Sims R, van der Lee SJ, Naj AC, Bellenguez C, Badarinarayan N, Jakobsdottir J, et al. Rare coding variants in PLCG2, ABI3, and TREM2 implicate microglial-mediated innate immunity in Alzheimer’s disease. Nat Genet (2017) 49(9):1373–84. doi:10.1038/ng.3916
378. Carroll CB, Wyse RHK. Simvastatin as a potential disease-modifying therapy for patients with Parkinson’s disease: rationale for clinical trial, and current progress. J Parkinsons Dis (2017) 7(4):545–68. doi:10.3233/JPD-171203
379. Klegeris A, McGeer EG, McGeer PL. Therapeutic approaches to inflammation in neurodegenerative disease. Curr Opin Neurol (2007) 20(3):351–7. doi:10.1097/WCO.0b013e3280adc943
380. McGeer PL, Guo JP, Lee M, Kennedy K, McGeer EG. Alzheimer’s disease can be spared by non steroidal anti-inflammatory drugs. J Alzheimers Dis (2017) 30:1–4. doi:10.3233/JAD-170706
381. Decourt B, Lahiri DK, Sabbagh MN. Targeting tumor necrosis factor alpha for Alzheimer’s disease. Curr Alzheimer Res (2017) 4(4):412–25. doi:10.2174/1567205013666160930110551
382. Kolahdooz Z, Nasoohi S, Asle-Rousta M, Ahmadiani A, Dargahi L. Sphingosin-1-phosphate receptor 1: a potential target to inhibit neuroinflammation and restore the sphingosin-1-phosphate metabolism. Can J Neurol Sci (2015) 42(3):195–202. doi:10.1017/cjn.2015.19
383. Park S-J, Im D-S. Sphingosine 1-phosphate receptor modulators and drug discovery. Biomol Ther (seoul) (2017) 25(1):80–90. doi:10.4062/biomolther.2016.160
384. Franceschi C, Salvioli S, Garagnani P, de Equileor M, Monti D, Capri M. Immunobiography and the heterogeneity of immune responses in the elderly: a focus on inflammaging and trained immunity. Front Immunol (2017) 8:982. doi:10.3389/fimmu.2017.00982
385. Garagnani P, Giuliani C, Pirazzini C, Olivieri F, Bacalini MG, Ostan R, et al. Centenarians as super-controls to assess the biological relevance of genetic risk factors for common age-related diseases: a proof of principle on type 2 diabetes. Aging (Albany NY) (2013) 5:373–85. doi:10.18632/aging.100562
386. Evert J, Lawler E, Bogan H, Perls T. Morbidity Profiles of centenarians: survivors, delayers and escapers. J Gerontol A Biol Sci Med Sci (2003) 58:232–7. doi:10.1093/gerona/58.3.M232
387. Bennati E, Murphy A, Cambien F, Whitehead AS, Archbold GPR, Young IS, et al. BELFAST centenarians: a case of optimised cardiovascular risk? Curr Pharm Des (2010) 16:789–95. doi:10.2174/138161210790883697
388. Terry DF, Wilcox M, McCormick MA, Lawler E, Perls TT. Cardiovascular advantages among the offspring of centenarians. J Gerontol A Biol Sci Med Sci (2004) 59:M385–9. doi:10.1093/gerona/59.4.M385
389. Rea M. Living long and ageing well: insights from nonagenarians. In: Davidson S, Goodwin J, Rossall P editors. Improving Later Life: Understanding The Oldest Old Age. London (2013). p. 74–8.
390. Laland KN, Odling-Smee J, Myles S. How culture shaped the human genome: bridging genetics and the human sciences together. Nat Rev Gen (2010) 11:137–48. doi:10.1038/nrg2734
391. Herrington W, Lacey B, Sherliker P, Armitage J, Lewington S. Epidemiology of atherosclerosis and the potential to reduce the global burden of atherothrombotic disease. Circ Res (2016) 118(4):535–46. doi:10.1161/CIRCRESAHA.115.307611
392. Roth GA, Forouzanfar MH, Moran AE, Barber R, Nguyen G, Feigin VL, et al. Demographic and epidemiologic drivers of global cardiovascular mortality. N Engl J Med (2015) 372:1333–41. doi:10.1056/NEJMoa1406656
393. Koton S, Schneider AL, Rosamond WD, Shahar E, Sang Y, Gottesman RF, et al. Stroke incidence and mortality trends in US communities, 1987 to 2011. JAMA (2014) 312(3):259–68. doi:10.1001/jama.2014.7692
394. Prince M. The global prevalence of dementia: a systemic review and meta-analysis. Alzheimers Dement (2013) 9(1):63–75. doi:10.1016/j.jalz.2012.11.007
395. Lee S, Shafe ACE, Cowie MR. UK stroke incidence, mortality and cardiovascular risk management 1999e2008: time-trend analysis from the general practice research database. BMJ Open (2011) 1:e000269. doi:10.1136/bmjopen-2011-000269
396. Xie X, Atkins E, Lv J, Bennett A, Neal B, Ninomiya T, et al. Effects of intensive blood pressure lowering on cardiovascular and renal outcomes: updated systematic review and meta-analysis. Lancet (2016) 387(10017):435–43. doi:10.1016/S0140-6736(15)00805-3
397. Ali MK, Bullard KM, Saaddine JB, Cowie CC, Imperatore G, Gregg EW. Achievement of goals in U.S. diabetes care, 1999-2010. N Engl J Med (2013) 368:1613–24. doi:10.1056/NEJMsa1213829
398. Satoh M, Tabuchi T, Itoh T, Nakamura M. NLRP3 inflammasome activation in coronary artery disease: results from prospective and randomized study of treatment with atorvastatin or rosuvastatin. Clin Sci (Lond) (2014) 126:233–41. doi:10.1042/CS20130043
399. Baigent C, Blackwell L, Emberson J, Holland LE, Reith C, Bhala N, et al. Efficacy and safety of more intensive lowering of LDL cholesterol: a meta-analysis of data from 170,000 participants in 26 randomised trials. Lancet (2010) 376:1670–81. doi:10.1016/S0140-6736(10)61350-5
400. Razquin C, Martinez JA, Martinez-Gonzalez MA, Fernández-Crehuet J, Santos JM, Marti A. A Mediterranean diet rich in virgin olive oil may reverse the effects of the -174G/C IL6 gene variant on 3-year body weight change. Mol Nutr Food Res (2010) 54(Suppl 1):S75–82. doi:10.1002/mnfr.200900257
401. Corella D, González JI, Bulló M, Carrasco P, Portolés O, Díez-Espino J, et al. The IL6 Gene Promoter SNP and Plasma IL-6 in Response to Diet Intervention. J Nutr (2009) 139(1):128–34. doi:10.3945/jn.108.093054
402. Latorre E, Birar VC, Sheerin AN, Jeynes CC, Hooper A, Dawe HR, et al. Small molecule modulation of splicing factor expression is associated with rescue from cellular senescence. BMC Cell Biol (2017) 18:31. doi:10.1186/s12860-017-0147-7
403. Falvo JV, Jasenosky LD, Kruidenier L, Goldfeld AE. Epigenetic control of cytokine gene expression: regulation of the TNF/LT Locus and T helper cell differentiation. Adv Immunol (2013) 118:37–128. doi:10.1016/B978-0-12-407708-9.00002-9
404. Freytag V, Carrillo-Roa T, Milnik A, Samann PG, Vukojevic V, Coynek D, et al. A peripheral epigenetic signature of immune system genes is linked to neocortical thickness and memory. Nat Commun (2017) 8:15193. doi:10.1038/ncomms15193
405. Kaminska B, Mot M, Pizzi M. Signal transduction and epigenetic mechanisms in the control of microglia activation during neuroinflammation. Biochim Biophys Acta (2016) 1862(3):339–51. doi:10.1016/j.bbadis.2015.10.026
406. Bauer M, Fink B, Thürmann L, Eszlinger M, Herberth G, Lehmann I. Tobacco smoking differently influences cell types of the innate and adaptive immune system—indications from CpG site methylation. Clin Epigenet (2016) 8:83. doi:10.1186/s13148-016-0249-7
407. Bell JT, Tsai PC, Yang TP, Pidsley R, Nisbet J, Glass D, et al. Epigenome-wide scans identify differentially methylated regions for age and age-related phenotypes in a healthy ageing population. PLoS Genet (2012) 8:e1002629. doi:10.1371/journal.pgen.1002629
408. Moskalev AA, Aliper AM, Smit-McBride Z, Buzdin A, Zhavoronkov A. Genetics and epigenetics of aging and longevity. Cell Cycle (2014) 13(7):1063–77. doi:10.4161/cc.28433
409. Rea IM, Dellet M, Mills KI; ACUME2 Project. Living long and ageing well: is epigenomics the missing link between nature and nurture? Biogerontology (2016) 17(1):33–54. doi:10.1007/s10522-015-9589-5
410. Martínez-González MA, Salas-Salvadó J, Estruch R, Corella D, Fitó M, Ros E, et al. Benefits of the Mediterranean diet: insights from the PREDIMED study. Prog Cardiovasc Dis (2015) 58(1):50–60. doi:10.1016/j.pcad.2015.04.003
411. Estruch R, Ros E, Salas-Salvadó J, Covas MI, Corella D, Martínez-González MA, et al. Primary prevention of cardiovascular disease with a Mediterranean diet. N Engl J Med (2013) 369:676–7. doi:10.1056/NEJMoa1200303
412. Camargo A, Delgado-Lista J, Garcia-Rios A, Cruz-Teno C, Yubero-Serrano EM, Perez-Martinez P, et al. Expression of pro-in- flammatory, pro-atherogenic genes is reduced by the Mediterranean diet in elderly people. Br J Nutr (2012) 108(3):500–8. doi:10.1017/S0007114511005812
413. de Lorgeril M, Salen P. The mediterranean-style diet for the prevention of cardiovascular diseases. Public Health Nutr (2006) 9(1):118–23. doi:10.1079/PHN2005933
414. Almeida OP, Khan KM, Hankey GJ, Yeap BB, Golledge J, Flicker L. 150 minutes of vigorous physical activity per week predicts survival and successful ageing: a population-based 11-year longitudinal study of 12,201 older Australian men. Br J Sports Med (2014) 48(3):220–5. doi:10.1136/bjsports-2013-092814
415. Carvalho A, Rea IM, Parimon T, Cusack BJ. Physical activity and cognitive function in individuals over 60 years of age: a systematic review. Clin Interv Aging (2014) 9:661–82. doi:10.2147/CIA.S55520
416. Hamer M, Lavoie KL, Bacon SL. Taking up physical activity in later life and healthy ageing: the English longitudinal study of ageing. Br J Sports Med (2014) 48(3):239–43. doi:10.1136/bjsports-2013-092993
417. Elosua R, Bartali B, Ordovas JM, Corsi AM, Lauretani F, Ferrucci L, et al. Association between physical activity, physical performance, and inflammatory biomarkers in an elderly population: the InCHIANTI study. J Gerontol A Biol Sci Med Sci (2005) 60(6):760–7. doi:10.1093/gerona/60.6.760
418. Yang YC, Boen C, Gerken K, Li T, Schorrp K, Harris KM. Social relationships and physiological determinants of longevity across the human life span. Proc Natl Acad Sci U S A (2016) 113(3):578–83. doi:10.1073/pnas.1511085112
419. Vemuri P, Lesnick TG, Przybelsk SA, Machulda M, Knopman DS, Mielke MM, et al. Association of lifetime intellectual enrichment with cognitive decline in the older population. JAMA Neurol (2014) 71(8):1017–24. doi:10.1001/jamaneurol.2014.963
420. Rea M, Rea S. Super Vivere: Reflections on Long Life and Ageing Well. Belfast: Blackstaff Press (2011). 187 p.
421. Yates LB, Djousse L, Kurth T, Buring JE, Gaziano M. Exceptional longevity in men: modificable factors associated with survival and function to age 90 years. Arch Intern Med (2008) 168:284–90. doi:10.1001/archinternmed.2007.77
422. Knoops KT, de Groot LC, Kromhout D, Perrin AE, Moreiras-Varela O, Menotti A, et al. Mediterranean diet, lifestyle factors, and 10-year mortality in elderly European men and women: the HALE project. JAMA (2004) 292(12):1433–9. doi:10.1001/jama.292.12.1433
423. Tam CS, Redman LM. Adipose tissue inflammation and metabolic dysfunction: a clinical perspective. Horm Mol Biol Clin Investig (2013) 15(1):19–24. doi:10.1515/hmbci-2013-0032
424. Holloszy JO, Fontana L. Caloric restriction in humans. Exp Gerontol (2007) 42(8):709–12. doi:10.1016/j.exger.2007.03.009
425. Jung SH, Park HS, Kim KS, Choi WH, Ahn CW, Kim BT, et al. Effect of weight loss on some serum cytokines in human obesity: increase in IL-10 after weight loss. J Nutr Biochem (2008) 19:371–5. doi:10.1016/j.jnutbio.2007.05.007
426. Picca A, Pesce V, Lezza AM. Does eating less make you live longer and better? An update on calorie restriction. Clin Interv Aging (2017) 12:1887–902. doi:10.2147/CIA.S126458
427. Jung KJ, Lee EK, Kim JY, Zou Y, Sung B, Heo HS, et al. Effect of short term calorie restriction on pro-inflammatory NF-kB and AP-1 in aged rat kidney. Inflamm Res (2009) 58:143–50. doi:10.1007/s00011-008-7227-2
428. Cronin O, Keohane DM, Molloy MG, Shanahan F. The effects of exercise interventions on inflammatory biomarkers in healthy, physically inactive subjects: a systemati review. QJM Int J Med (2017) 110:629–37. doi:10.1093/qjmed/hcx091
429. Wohlgemuth SE, Seo AY, Marzetti E, Lees HA, Leeuwenburgh C. Skeletal muscle autophagy and apoptosis during aging: effects of calorie restriction and life-long exercise. Exp Gerontol (2010) 45:138–48. doi:10.1016/j.exger.2009.11.002
430. Sampson TR, Debelius JW, Thron T, Janssen S, Shastri GG, Ilhan ZE, et al. Gut microbiota regulate motor deficits and neuroinflammation in a model of Parkinson’s disease. Cell (2016) 167(6):1469.e–80.e. doi:10.1016/j.cell.2016.11.018
431. Youm Y-H, Nguyen KY, Grant RW, Goldberg EL, Bodogai M, Kim D, et al. The ketone metabolite β-hydroxybutyrate blocks NLRP3 inflammasome-mediated inflammatory disease. Nat Med (2015) 21:263–9. doi:10.1038/nm.3804
432. Forsythe P, Bienenstock J, Kunze WA. Vagal pathways for microbiome-brain-gut axis communication. Adv Exp Med Biol (2014) 817:115–33. doi:10.1007/978-1-4939-0897-4_5
433. Salminen A, Huuskonen K, Ojala J, Kauppinen A, Kaarniranta K, Suuronen T. Activation of innate immunity system during ageing.: NF-kB signaling is the molecular culprit of inflamm-aging. Ageing Res Rev (2008) 7(2):83–105. doi:10.1016/j.arr.2007.09.002
434. Goldberg EL, Dixit VD. Drivers of age-related inflammation and strategies for healthspan extension. Immunol Rev (2015) 265(1):63–74. doi:10.1111/imr.12295
435. Mann M, Mehta A, Zhao JL, Lee K, Mariov GK, Garcia-Flores Y, et al. An NF-κB-microRNA regulatory network tunes macrophage inflammatory responses. Nat Commun (2017) 8:851. doi:10.1038/s41467-017-00972-z
436. Xu X, Ning Y-C, Wang W, Liu J, Bai X, Sun X, et al. Anti-inflamm-aging effects of long-term caloric restriction via overexpression of SIGIRR to inhibit NF-κB signaling pathway. Cell Physiol Biochem (2015) 37:1257–70. doi:10.1159/000430248
437. Dinarello CA. Anti-inflammatory agents: present and future. Cell (2010) 140(6):935–50. doi:10.1016/j.cell.2010.02.043
438. Falantes J, Pleyer L, Thépot S, Almeida AM, Maurillo L, Martínez-Robles V, et al. European ALMA + investigators. Real life experience with frontline azacitidine in a large series of older adults with acute myeloid leukemia stratified by MRC/LRF score: results from the expanded international E-ALMA series (E-ALMA+). Leuk Lymphoma (2018) 59(5):1113–20. doi:10.1080/10428194.2017.1365854
439. Joven J, Micol V, Segura-Carretero A, Alonso-Villaverde C, Menéndez JA, Bioactive Food Components Platform. Polyphenols and the modulation of gene expression pathways: can we eat our way out of the danger of chronic disease? Crit Rev Food Sci Nutr (2014) 54(8):985–1001. doi:10.1080/10408398.2011.621772
440. Uguccioni M, Teixeira MM, Locati M, Mantovani A. Editorial: regulation of inflammation, its resolution and therapeutic targeting. Front Immunol (2017) 8:415. doi:10.3389/fimmu.2017.00415
441. Figueira I, Fernandes A, Miadenovi DA, Lopez-Contreras A, Henriques CM, Selman C, et al. Interventions for age-related diseases: shifting the paradigm. Mech Ageing Develop (2016) 160:69–92. doi:10.1016/j.mad.2016.09.009
442. De Cabo R, Carmona-Gutierrez D, Bernier M, Hall MN, Madeo F. The search for antiaging interventions: from elixirs to fasting regimens. Cell (2014) 157:1515–26. doi:10.1016/j.cell.2014.05.031
443. Broer L, Buchman AS, Deelen J, Evans DS, Faul JD, Lunetta KL, et al. GWAS of longevity in CHARGE consortium confirms APOE and FOXO3 candidacy. J Gerontol A Biol Sci Med Sci (2015) 70(1):110–8. doi:10.1093/gerona/glu166
444. Deelen J, Beekman M, Uh H-W, Broer L, Ayers KL, Tan Q, et al. Genome-wide association meta-analysis of human longevity identifies a novel locus conferring survival beyond 90 years of age. Hum Mol Genet (2014) 23:4420–32. doi:10.1093/hmg/ddu139
445. Lu F, Guan H, Gong B, Liu X, Zhu R, Wang Y, et al. Genetic variants in PVRL2-TOMM40-APOE region are associated with human longevity in a Han Chinese population. PLoS One (2014) 9(6):e99580. doi:10.1371/journal.pone.0099580
446. Beekman M, Blanché H, Perola M, Hervonen A, Bezrukov V, Sikora E, et al. Genome-wide linkage analysis for human longevity: genetics of healthy ageing study. Aging Cell (2013) 12(2):184–93. doi:10.1111/acel.12039
447. Brooks-Wilson A. Genetics of healthy aging and longevity. Hum Genet (2013) 132:1323–38. doi:10.1007/s00439-013-1342-z
448. Murabito JM, Yuan R, Lunetta KL. The search for longevity and healthy aging genes: insights from epidemiological studies and samples of long-lived individuals. J Gerontol A Biol Sci Med Sci (2012) 67(5):470–9. doi:10.1093/gerona/gls089
449. Sebastiani P, Solovieff N, DeWan AT, Walsh KM, Puca A, Hartley SW, et al. Genetic signatures of exceptional longevity in humans. PLoS One (2012) 7(1):e29848. doi:10.1371/journal.pone.0029848
450. Deelan J, Beekman M, Hae-Won UH, Helmer Q, Kuningas M, Christiansen L, et al. Genome-wide association study identifies a single major locus contributing to survival into old age: the APOE locus revisited. Aging Cell (2011) 10:686–98. doi:10.1111/j.1474-9726.2011.00705.x
451. Soerensen M, Dato S, Christensen K, McGue M, Stevnsner T, Bohr VA, et al. Replication of an association of variation in the FOXO3A gene with human longevity using both case-control and longitudinal data. Aging Cell (2010) 9:1010–7. doi:10.1111/j.1474-9726.2010.00627.x
452. Willcox DC, Willcox BJ, Poon LW. Centenarian studies: important contributors to our understanding of the aging process and longevity. Curr Gerontol Geriatr Res (2010) 2010:6. doi:10.1155/2010/484529
453. Willcox BJ, Willcox DC, Ferrucci L. Secrets of healthy aging and longevity from exceptional survivors around the globe: lessons from octogenarians to supercentenarians. J Gerontol A Biol Sci Med Sci (2008) 63(11):1181–5. doi:10.1093/gerona/63.11.1181
454. Franceschi C, Bezrukov V, Blanche H, Bolund L, Christensen K, De Benedictis G, et al. Genetics of healthy aging in Europe: the EU-integrated project GEHA (GEnetics of healthy aging). Ann N Y Acad Sci (2007) 1100:21–45. doi:10.1196/annals.1395.003
455. Franceschi C. Inflammaging as a major characteristic of old people: can it be prevented or cured? Nutr Rev (2007) 65(12):S173–6. doi:10.1111/j.1753-4887.2007.tb00358.x
456. Arai Y, Martin-Ruiz C, Takayama M, Abe Y, Takbayashi T, Koyasu S, et al. Inflammation, but not telomere length, predicts successful ageing at extreme old age: a longitudinal study of semi-supercentenarians. EBioMedicine (2015) 2(10):1316–7. doi:10.1016/j.ebiom.2015.07.029
457. Baggio G, Donazzan S, Monti D, Mari D, Martini S, Gabelli C, et al. Lipoprotein(a) and lipoprotein profile in healthy centenarians: a reappraisal of vascular risk factors. FASEB J (1998) 12(6):433–7. doi:10.1096/fasebj.12.6.433
458. Calabrese EJ. Hormetic dose-response relationships in immunology: occurrence, quantitative features of the dose response, mechanistic foundations, and clinical implications. Crit Rev Toxicol (2005) 35(2–3):89–295. doi:10.1080/10408440590917044
459. Holmans P, Moskvina V, Jones L, Sharma M; International Parkinson’s Disease Genomics Consortium, Vedernikov A, et al. A pathway-based analysis provides additional support for an immune-related genetic susceptibility to Parkinson’s disease. Hum Mol Genet (2013) 22(5):1039–49. doi:10.1093/hmg/dds492
Keywords: aging, age-related diseases, inflamm-aging, redox, SASP, autophagy, cytokine dysregulation, inflammation resolution
Citation: Rea IM, Gibson DS, McGilligan V, McNerlan SE, Alexander HD and Ross OA (2018) Age and Age-Related Diseases: Role of Inflammation Triggers and Cytokines. Front. Immunol. 9:586. doi: 10.3389/fimmu.2018.00586
Received: 25 November 2017; Accepted: 08 March 2018;
Published: 09 April 2018
Edited by:
Rafael Solana, Universidad de Córdoba, SpainReviewed by:
Armando Luna López, Instituto Nacional de Geriatría, MexicoValerio Chiurchiù, Università Campus Bio-Medico, Italy
Beatriz Sánchez Correa, Universidad de Extremadura, Spain
Copyright: © 2018 Rea, Gibson, McGilligan, McNerlan, Alexander and Ross. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Irene Maeve Rea, i.rea@qub.ac.uk