 Andrés J. Cortés
Andrés J. Cortés Felipe López-Hernández
Felipe López-Hernández Daniela Osorio-Rodriguez
Daniela Osorio-Rodriguez- 1Corporación Colombiana de Investigación Agropecuaria AGROSAVIA, C.I. La Selva, Rionegro, Colombia
- 2Departamento de Ciencias Forestales, Facultad de Ciencias Agrarias, Universidad Nacional de Colombia – Sede Medellín, Medellín, Colombia
- 3Division of Geological and Planetary Sciences, California Institute of Technology (Caltech), Pasadena, CA, United States
Molecular evolution offers an insightful theory to interpret the genomic consequences of thermal adaptation to previous events of climate change beyond range shifts. However, disentangling often mixed footprints of selective and demographic processes from those due to lineage sorting, recombination rate variation, and genomic constrains is not trivial. Therefore, here we condense current and historical population genomic tools to study thermal adaptation and outline key developments (genomic prediction, machine learning) that might assist their utilization for improving forecasts of populations’ responses to thermal variation. We start by summarizing how recent thermal-driven selective and demographic responses can be inferred by coalescent methods and in turn how quantitative genetic theory offers suitable multi-trait predictions over a few generations via the breeder’s equation. We later assume that enough generations have passed as to display genomic signatures of divergent selection to thermal variation and describe how these footprints can be reconstructed using genome-wide association and selection scans or, alternatively, may be used for forward prediction over multiple generations under an infinitesimal genomic prediction model. Finally, we move deeper in time to comprehend the genomic consequences of thermal shifts at an evolutionary time scale by relying on phylogeographic approaches that allow for reticulate evolution and ecological parapatric speciation, and end by envisioning the potential of modern machine learning techniques to better inform long-term predictions. We conclude that foreseeing future thermal adaptive responses requires bridging the multiple spatial scales of historical and predictive environmental change research under modern cohesive approaches such as genomic prediction and machine learning frameworks.
On the Challenges of Studying Genomic Thermal Adaptation
Warming is imposing an unprecedented climate emergency on nature, food, energy supply, and economy around the world (Ripple et al., 2020). While evolutionary genomics may improve prediction of populations’ responses to thermal change (Waldvogel et al., 2020a), geologic records of temperature and carbon dioxide (CO2) variations (Supplementary Figure S1) are also insightful into the coupling of biodiversity, climate, and the carbon cycle and hence may help predicting the consequences of future carbon emissions (Zachos et al., 2008). For instance, several reports of fire activity (Whitlock and Bartlein, 2003; Bush et al., 2008) and hydroclimate changes (Wang et al., 2017) as records of thermal changes during the Holocene have taught us that extinction is a slow process and that many species may already be functionally extinct (Cronk, 2016). A key modern advance has precisely been to couple the extinction risk with the migratory potential under an ecological niche conservatism scenario (Steinbauer et al., 2018), and predictions of population-level genomic and phenotypic responses to thermal change (Hoffmann and Sgro, 2011). Although atmospheric CO2 has been found to be better correlated with richness of (plant) species (Supplementary Figure S1C) than temperature itself throughout the Cenozoic up until 20 Mya (Jaramillo et al., 2006; Royer and Chernoff, 2013), we need to improve our understanding on how thermal change vulnerability impacts current and historical adaptive genetic variation in order to enhance populations response projections (Razgour et al., 2019).
Genomes are diverse in signatures of the populations’ evolutionary past across timescales (Wolf and Ellegren, 2017) and therefore are informative on historical adaptive responses to ancient and more recent events of climate change (Figure 1 and Table 1). By revealing the nature of these signatures and learning from previous reactions to environmental change, genomics can truly assist modern predictions aimed at incorporating responses beyond migration. Yet, disentangling often confused selective and demographic signatures from those due to genetic drift and genomic constrains is challenging (Ellegren and Galtier, 2016), consequently delaying the factual utilization of genomics for forecasting. Therefore, in this mini-review we envision summarizing modern tools from the genomic era that are enriching our comprehension of the genetic consequences of past and recent climate change, while offering a perspective on how to improve predictive models that incorporate thermal adaptation. Specifically, we aim prospecting how genomic prediction (GP) and machine learning (ML) approaches may offer cohesive frameworks to (1) integrate more traditional, but heterogeneous, genomic, and ecological datasets across temporal scales, by (2) maximizing prediction accuracies, while (3) understating the relative contribution of the underlying genomic processes. This is still a future avenue of research, and so we close by offering perspectives. Different drivers of the genomic landscape to thermal adaption (Gompert et al., 2014; Ravinet et al., 2017; Cortés and Blair, 2018; López-Hernández and Cortés, 2019), such as disruptive and background selection, gene flow (Miller et al., 2020), shared ancestral polymorphism, and mutation/recombination rate variation (Feder et al., 2012; Ellegren and Wolf, 2017; Cortés et al., 2018b), have been identified. In order to discern among them, a first necessary step toward the evaluation of the adaptive potential involves typifying the genomic landscape by using summary statistics like nucleotide diversity, π (Nei, 1987), and relative, FST (Weir and Cockerham, 1984), and absolute, DXY (Nei, 1987), divergence. FST vs. DXY contrasts inform population divergence in the presence of gene flow (co-occurrence of peaks in both profiles), recurrent selection across subpopulations (FST peaks match shallow DXY valleys), and selective sweeps predating the subpopulations’ split (FST peaks match deep DXY valleys) (Nachman and Payseur, 2012; Cruickshank and Hahn, 2014; Irwin et al., 2016). Inferences are more robust if carried out across replicated samplings of contrasting populations (e.g., in terms of thermal variation) within a hierarchically nested framework of divergence (Cortés et al., 2018b). A second step refers to the detection of selection signatures, if any – i.e., hard vs. soft selection sweeps (Pritchard et al., 2010; Zahn and Purnell, 2016), which must be followed by a third validation step across replicated demographics (Roesti et al., 2014; Lotterhos and Whitlock, 2015) and temporal levels (Nosil and Feder, 2011; Matos et al., 2015; Fragata et al., 2018).

Figure 1. Potential approaches to assess populations’ thermal adaptation by looking into their genomic past. Genomic analyses allow reconstructing populations’ adaptive responses to previous events of climate change across various temporal scales (A,B,D,F), as a tool to improve forecasting (C,E,G,H). (A) Empirical approaches such as replicated “common garden” (provenance) tests and space-for-time (SFT) substitution allow studying in situ ongoing genomic thermal adaptation. The inset plot exemplifies a significant genotype-by-environment (GxE) interaction, as can be quantified using reciprocal transplant experiments between habitat types that differ in their thermal stress. (B) Coalescent and approximate Bayesian computation (ABC) analyses help infer recent thermal-driven selective and demographic responses. The inset diagram shows a typical coalescent genealogy depicting divergence with gene flow. (C) The breeder’s equation predicts responses of genetically correlated traits over one generation (vector R) given standardized selection gradients to thermal stress (vector β) by means of the variance–covariance matrix (G) of additive genetic parameter estimates. Alternatively, experimental evolution traces real-time changes in allele frequencies (Δp) across generations. (D) When genomic signatures of thermal selection are under divergent selection after several generations, genome-wide association (GWAS), and selection (GWSS) scans, as well as genome–environment associations (GEA), allow characterizing the genomic architecture of thermal adaptation. The inset Manhattan plot schematizes a hypothetical genomic scan between populations that contrast in their thermal adaptation. (E) Modern high-throughput genotyping may facilitate predictions of the thermal adaptive potential over multiple generations using infinitesimal models under a genomic prediction (GP) framework. (F) Phylogeographic approaches offer an understanding of the genomic consequences of deep-time thermal shifts at an evolutionary time scale. The inset tree represents an imaginary phylogeny. Finally, (G) machine learning (ML) approaches (H) trained using heterogeneous past responses to thermal variation may enhance long-term predictions of the thermal adaptive potential. ML’s modus operandi, as GPs, requires partitioning the calibrating historical dataset between training (TRN) and testing (TST) subsets that are iteratively imputed into a N-fold cross-validation scheme.
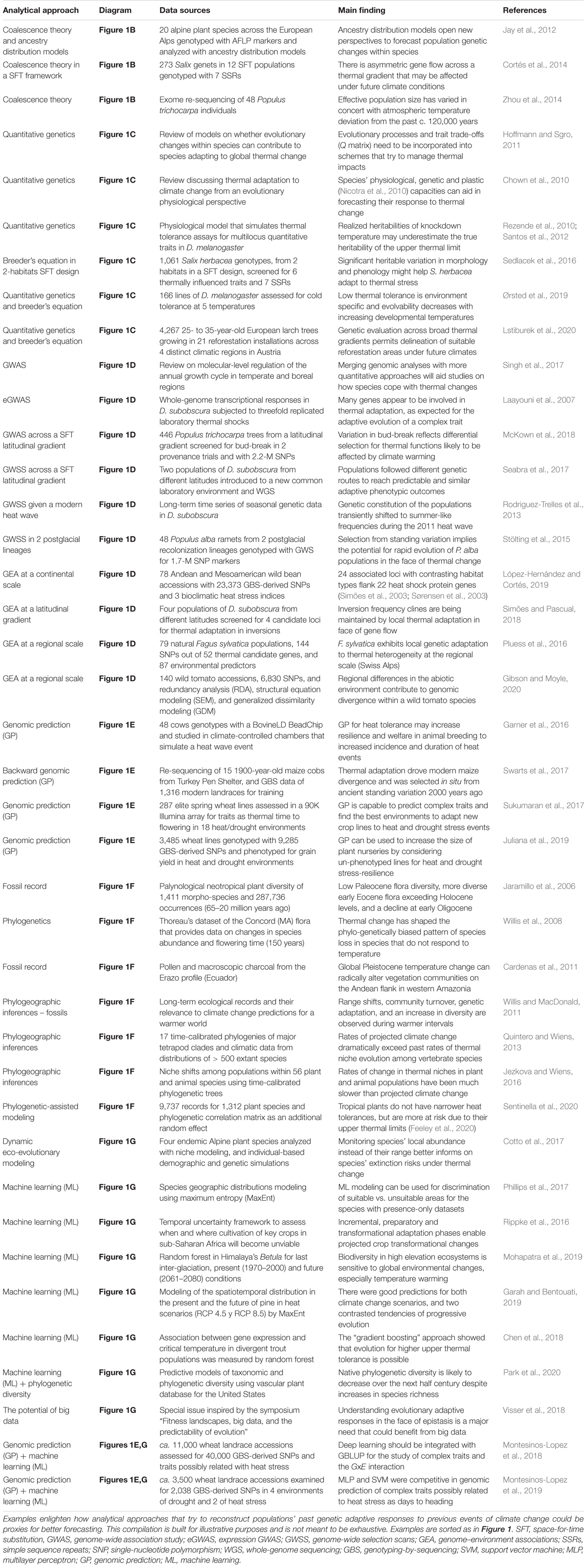
Table 1. Case studies that have addressed thermal adaptation at different temporal scales using diverse genetic analyses.
Exclusively phenotypic empirical methods (Figure 1A), such as in situ monitoring, growth chamber experiments, and “common garden” (provenance) tests (Miller et al., 2020), constitute baseline evidence of thermal adaptation and should therefore inform more advanced genomic approaches. Naturally available environmental gradients (e.g., elevation or latitudinal clines) can also be used as proxies for climate change (Wheeler et al., 2016; Cortés and Wheeler, 2018), which is known as space-for-time (SFT) substitution. Replicated “common garden” tests (a.k.a. reciprocal transplants) carried out in an SFT framework are in turn useful to test whether populations can cope with changes through local adaptation (standing variation) or via phenotypic plasticity, especially in long-living species (Bridle and Vines, 2007; Sedlacek et al., 2015). Within an SFT framework, restricted gene flow can lead to small-scale genetic structures (Stanton et al., 1997) or distorted source/sink-like patterns (e.g., Cortés et al., 2014) driven by environmental factors (Nathan and Muller-Landau, 2000). Asymmetric migratory potential in a local scale may provide suitable habitats within only a few meters of the current locations (Yamagishi et al., 2005; Scherrer and Körner, 2011) but may also lead to narrowly adapted populations, even in the face of gene flow (Fitzpatrick et al., 2015), that may respond poorly to future conditions (North et al., 2011; Miller et al., 2020).
From Recent Genetic Responses to Short-Term Predictions
Coalescence Informs on Contemporary Thermal-Driven Selective and Demographic Changes
In order to trace back thermal-driven selective and demographic changes at recent temporal scales (Figure 1B), coalescent theory (Wakeley, 2008) helps in discriminating among authentic signatures of selection and those related to demography (e.g., bottlenecks and among populations reduced gene flow), from spurious covariates (Yeaman and Otto, 2011) such as lineage sorting (Wolf and Ellegren, 2017; Becher et al., 2020) and inversions (Dolgova et al., 2010; Fragata et al., 2014). Recursive simulation-based tools to incorporate the mutation/selection balance (Bustamante et al., 2001) across various scenarios of divergence and gene flow are approximate Bayesian computation – ABC (Csilléry et al., 2010; Cornuet et al., 2014), and pairwise sequentially Markovian coalescent – PSMC (Nadachowska-Brzyska et al., 2016). These approaches can inform how isolated populations that usually occupy climates with scarce habitat complexity (Flantua et al., 2019) may favor thermal generalists, while intricate local-scale heterogeneity at larger scales could trigger (Hughes, 2006; Cortés et al., 2018a) thermal specialists with limited migration potential (Cuesta et al., 2019). They can also model population sizes (Beerli, 2006) in concert with thermal changes (Zhou et al., 2014; Lehnert et al., 2019). Yet, these approaches may be limited by computational burden as they rely on simulation-based rejection sampling, while much effort is gone into the design of multiple scenarios, dimensionality reduction, and feature selection (Schrider and Kern, 2018).
The Breeder’s Equation Assists Multi-Trait Predictions Over a Few Generations
In order for thermal adaptation to happen, there must be heritable trait variation upon which selection, enforced by climate change, acts (Darwin, 1874). A simple deterministic model that condenses this evolutionary paradigm, aiding in the forecast of adaptive trait responses across few generations, comes from the quantitative genetic discipline and is known as the breeder’s equation (Figure 1C). Its multivariate form (Walsh, 2008) allows predicting responses of genetically correlated traits (vector R) to standardized thermal selection gradients (vector β) over one generation, so that R = Gβ, where G is the variance–covariance matrix of additive genetic parameter estimates – a proxy for traits’ heritabilities and trade-offs (Falconer and Mackay, 1996). The potential evolutionary response can therefore be computed using selection-gradient estimates derived from fitness proxies (i.e., fitness values regressed as a function of standardized trait values) and marker-based heritabilities (Lynch and Ritland, 1999). This approach by itself is not novel, but what makes it powerful is that it can be coupled with SFT (Wheeler et al., 2014), among other trials, to predict thermal responses to thermal change (Sedlacek et al., 2016). Yet, a major drawback is that selection gradients heavily depend on the nature of the fitness proxies (Sedlacek et al., 2016). Alternatively, experimental evolution studies (Exposito-Alonso et al., 2019) could test more explicitly how rapidly growing populations may respond to different thermal scenarios (Kawecki et al., 2012) that, together with evolve and re-sequence analyses (Turner and Miller, 2012), may contribute to understand the genetic basis of short-term thermal adaptation.
From Deeper Genomic Signatures of Selection to Mid-Term Predictions
Genome-Wide Scans Reveal Signatures of Divergent Selection to Past Thermal Adaptation
Assuming that enough generations have passed as to exhibit divergent selection to thermal changes, genome-wide association (GWAS) (Hirschhorn and Daly, 2005) and selection (GWSS) (Sabeti et al., 2007) scans (Figure 1D) are essential analytical tools to reconstruct the genomic architecture of adaptive trait divergence to thermal stress (Lecheta et al., 2020; Zwoinska et al., 2020). These methods assume that some allele variants are in linkage disequilibrium (LD) (Slatkin, 2008) with causal variants that influence the adaptive phenotype (Morris and Borevitz, 2011; Tam et al., 2019), a.k.a. genetic “hitchhiking” (Maynard Smith and Haigh, 1974; Feder and Nosil, 2010). An interface between GWAS and GWSS studies where loci are directly correlated with niche’s thermal variables is named genome–environment association (GEA) (Forester et al., 2016) and is insightful to infer past thermal adaptation, too (Hancock et al., 2011; Pluess et al., 2016; López-Hernández and Cortés, 2019). Yet, these approaches partly disregard non-additive and highly polygenic architectures (Stephan, 2016; Csillery et al., 2018; Barghi et al., 2020) and may be misleading (Maher, 2008; Pennisi, 2014) if standardized data (Waldvogel et al., 2020b) and statistical covariates (Lambert and Black, 2012), such as population stratification (Barton et al., 2019) and genomic constrains (Wray et al., 2013; Huber et al., 2016), are incorrectly accounted for.
Genomic Prediction May Assist Forecasting of Adaptive Traits Over Multiple Generations
A cutting-edge development that materialized after bringing genomics into quantitative genetics theory is genomic prediction (GP) (Desta and Ortiz, 2014; Crossa et al., 2017; Grattapaglia et al., 2018). GP uses historical phenotypic data to adjust marker-based infinitesimal (Figure 1E) models (Meuwissen et al., 2001; Gianola et al., 2006; de los Campos et al., 2013) that may overcome some of the restraints described in the previous section. GP may offer a more thoughtful picture of complex traits (e.g., thermal adaptation), presumably regulated by many low-effect loci (Pritchard et al., 2010). GP has so far informed predictions of single adaptive traits in populations with known pedigrees (Saint Pierre et al., 2012; Cros et al., 2019) and hybrid origins (Technow et al., 2014; Tan et al., 2017), as well as multi-trait inferences across diverse unrelated populations (Crossa et al., 2007, 2016; Resende et al., 2012; Suontama et al., 2019) under genotype by environment interactions (GxE) (Montesinos-Lopez et al., 2018; Crossa et al., 2019) facing polygenic climate adaptation (Isabel et al., 2020). GP of thermal adaptive traits across multiple generations and populations may be incipient (Table 1), yet it harbors a promising potential, as was demonstrated by reversely predicting unobserved thermal phenology in 1900-year-old ancient corn (Swarts et al., 2017), and as we prospect in the last section of this mini-review.
From Deep-Time Genomic Consequences of Thermal Shifts to Long-Term Predictions
Phylogeography Offers Insights Into Past Responses at an Evolutionary Scale
Phylogeographic inferences (Figure 1F) offer insights into how species (1) diversify (Quintero and Wiens, 2013) and (2) face the effects of past thermal variation (Jezkova and Wiens, 2016; Richardson et al., 2019) by boosting complex interactions such as species facilitation (Wheeler et al., 2015), adaptive introgression, and hybrid speciation (Coyne and Orr, 2004; Abbott et al., 2013; Payseur and Rieseberg, 2016; Marques et al., 2019). For instance, interspecific hybrids with intermediate niche requirements may rescue population’s gene pools in the face of climate change, while they can also display signals of heterosis for thermal adaption due to dominance on recessive alleles or overdominance via novel allele combinations (Abdelmula et al., 1999; Leinonen et al., 2011). Modern phylogeographic inferences currently rely on abundant and unlinked genetic markers (Bryant et al., 2012) that are capable of bypassing traditional assumptions of single gene mutation models (Caliebe, 2008) while accounting for scenarios of reticulate evolution (Vargas et al., 2017). Marker-based inferences also offer higher resolution to validate cases where adaptive radiation (Madriñán et al., 2013), and ecological parapatric speciation resulted from local patterns of environmental variation (Cortés et al., 2018a) that may resemble those expected by thermal change. Mosaics of local-habitat heterogeneity can ultimately enforce thermal pre-adaptation (Cortés and Wheeler, 2018). Distance-based phylogenic reconstruction without proper out-groups (Baum et al., 2005; Cortés, 2013) is yet a major risk of these approaches.
Machine Learning May Bridge Historical Genomics and Long-Term Predictions
A promising way to simultaneously make sense of multiple sources of historical genomic data that can be utilized to predict populations’ adaptive responses is by merging them into a machine learning (ML) framework (Figures 1G,H). ML bypasses the “curse of dimensionality” and benefits from high-dimensional inputs of heterogeneous dependent variables (“features”) without a priori knowledge of their joint probability distribution (Schrider and Kern, 2018). This improves predictions’ “recall” (true positive) rate among a set of possible responses, especially when the classification is iteratively trained using “labeled” data (i.e., historical thermal responses may offer novel calibration datasets, Table 1) via N-fold cross-validation. ML has been routinely used to make ecological niche modeling (Phillips et al., 2017; Valencia et al., 2020) and functional predictions across genomes (Libbrecht and Noble, 2015). Yet, ML may likely displace other tools useful to characterize the genomic consequences of thermal adaptation, already introduced in this mini-review, such as ABC modeling (Liu et al., 2019) and GWSS (Schrider and Kern, 2018).
Concluding Remarks
Thermal adaptation is a complex polygenic trait well-described in terms of its genetic architecture and selection footprints across a wide range of phylogenetically diverse taxa (Way and Oren, 2010; Valladares et al., 2014; López-Hernández and Cortés, 2019). While genomics has enabled these achievements that rely on past events of thermal variation, forward predictions remain one step behind partly because (1) disentangling selective and demographic drivers of the genomic landscape from fortuitous genomic constrains (Logan and Cox, 2020) is puzzling (Ellegren and Galtier, 2016) and (2) merging these heterogeneous signatures and data sources into a cohesive predictive framework was unfeasible, until recently. In this mini-review, we advocated for novel approaches that may enhance our understanding of the genetic consequences of past climate change, while offering new avenues to calibrate more accurate predictive models of the thermal adaptive potential. For instance, ML advances are likely to now move beyond species distribution modeling (Phillips et al., 2017) and functional genomics (Libbrecht and Noble, 2015) to permeate the backward interpretation of recent genetic demographic responses and genomic signatures to historical thermal selection by updating popular but sometimes intractable methods such as ABC modeling and GWSS (Schrider and Kern, 2018). Meanwhile, GP and ML might boost forward predictions of the adaptive potential beyond a single generation by training multifactorial models that can try incorporating genomic heterogeneous evidence of historical thermal adaption across a wide spectrum of temporal scales. Ultimately, understanding how biotas formed in response to historical environmental change may improve our ability to predict and mitigate the threats to species posed by global warming (Ding et al., 2020).
Despite GP’s and ML’s being useful to comprehend and predict thermal adaptation, these new paradigms are not exempt of criticism. A reiterative misconception is that because these methodologies aim at strengthening predictions and classification boundaries, they do not offer a mechanistic understanding of the subjacent processes. However, even though GP and ML rely on algorithmically generated models, both are far from “black boxes” because they allow direct measurement of the contribution of each genetic marker (Resende et al., 2012; Spindel et al., 2016) and “feature” (Schrider and Kern, 2018), to the point that they can offer higher resolution than traditional genetic mapping (Hirschhorn and Daly, 2005) and deterministic model building (Otto and Day, 2007) techniques. A second misconception assumes computational burden. Although both GP and ML require a large number of simulations, they do not depend on rejection sampling, which means they may efficiently use all of the simulations to inform the mapping of historical thermal data to parameters (Schrider and Kern, 2018).
Future Directions
So far, GP and ML have been mostly utilized to address thermal adaptation individually (Table 1). For instance, GP has been used to project heat tolerance in diverse wheat lines (Sukumaran et al., 2017; Juliana et al., 2019), and bovine genotypes (Garner et al., 2016), in all cases more as a proof of concept. Similarly, ML approaches have not only deepened our understating on populations’ range shifts in the light of thermal variation (Rippke et al., 2016; Garah and Bentouati, 2019; Mohapatra et al., 2019) but also assisted eGWAS of critical temperature thresholds (Chen et al., 2018) and phylogenetic forecasting in plants (Park et al., 2020). However, since GP and ML are both cutting-edge tools, there is still room and need for new developments. For instance, merging more cohesively past adaptive responses to previous events of environmental change into cutting-edge analytical frameworks like GP and ML will ultimately allow predicting whether populations’ adaptive potential may keep up with the pace of current thermal increase (Franks and Hoffmann, 2012; Franks et al., 2014). Swarts et al. (2017) illustrates that across-temporal predictions may be useful not only to improve forecasting (Sweet et al., 2019) but also to better understand previous responses to thermal variation, since they used backward GP to demonstrate that thermal adaptation in maize was selected in situ from ancient standing variation 2000 years ago. By enlightening on the nature of these historical genetic signatures to past climate change, genomics can also enhance predictions that aim at incorporating adaptive responses beyond extirpation and range shifts (Chen et al., 2011).
Data sources incorporated into GP and ML can transcend those with a direct genomic connotation and involve others that can modulate or be informative of the thermal responses. For instance, from an abiotic point of view, nutrient availability (Little et al., 2016), absorption (Wu et al., 2020), and soil interactions (Sedlacek et al., 2014) could act as enhancers or limiting factors of the adaptive responses. From a biotic perspective, among-ecotype differentiation (Cortés et al., 2012a,b, 2013; Blair et al., 2016), intrapopulation divergence (Cortés et al., 2011; Blair et al., 2012, 2018; Kelleher et al., 2012), and within-family variation (Galeano et al., 2012; Blair et al., 2013) could encourage or coerce adaptation. Population’s functioning, abundance, distribution, and diversity, as predicted from controlled experiments (Way and Oren, 2010; Elmendorf et al., 2012; Wolkovich et al., 2012; Andresen et al., 2016; Becklin et al., 2017; Singh et al., 2017), experimental evolution (Tenaillon et al., 2012; Mallard et al., 2018; Pfenninger and Foucault, 2020), biological monitoring (Walther et al., 2002; Franks et al., 2013; Wipf et al., 2013; Reichstein et al., 2014; Hällfors et al., 2020), and shifts observed in the fossil record (Alsos et al., 2009; Willis and MacDonald, 2011; Lyons et al., 2016; Bruelheide et al., 2018), can feed back on climate change (Pearson et al., 2013) and so be considered as drivers themselves. Regardless of the exact nature and extent of the data type, both GP and ML may offer suitable scenarios to merge diverse, and even conflicting, data sources in order to pinpoint emergent properties (Street et al., 2011) out of a complex system, as is thermal genomic adaptation. Therefore, a key guideline for new developments concerns a better coupling of GP and ML approaches. Until now, only a few works have relied on both methodologies, in the context of thermal adaptation in wheat landraces (Montesinos-Lopez et al., 2018, 2019), but have not gone beyond technical comparisons/recommendations, nor have designed integrated pipelines. Also, reconciling modern genomics with last-generation predictive inferences of the thermal adaptive potential and stochastic demographic modeling (Jenouvrier et al., 2009) is necessary. Open-access resources and data sharing platforms are as crucial in this effort as new integrated analytical pipelines. We are looking forward to seeing more cohesive (Beyer et al., 2020) and systematic studies and predictions across the rich and informative temporal spectrum (Kristensen et al., 2018) of past and future environmental variation (Franks et al., 2013). These efforts should be carried out through a wide range of spatial scales (Parmesan and Hanley, 2015; Way et al., 2015; Gonzalez et al., 2020) spanning contrasting ecosystems (Lenoir et al., 2020), microhabitats (Zellweger et al., 2020), and unrelated taxa, which together may already be keeping heritable adaptive trait differentiation valuable for long-term thermal responses and informative for conservation prioritizations (Barnosky et al., 2017; Elsen et al., 2020).
Author Contributions
AC conceived this mini-review. FL-H collected the literature and prepared diagrams. DO-R compiled the historical climate data. AC wrote the first draft of the mini-review with further contributions from FL-H and DO-R. All authors contributed to the article and approved the submitted version.
Funding
AC was supported by grants 4.1-2016-00418 and BS2017-0036 from Vetenskapsrådet (VR) and Kungliga Vetenskapsakademien (KVA), respectively. The National Science Foundation (NSF) and the SIMONS Collaboration on the Origins of Life support DO-R. The editorial fund from the Colombian Corporation for Agricultural Research (AGROSAVIA) was thanked for subsidizing the mini-review B-type processing charge.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We acknowledge thoughtful discussions with M. W. Blair regarding the genetic basis of thermal adaptation that took place with AC and DO-R during the Erasmus funded workshop “Molecular Breeding for Abiotic Constraints in Plants” held in Montpellier (France) during the summer of 2012. Some of the ideas discussed here were also framed into perspective, thanks to suggestions from A. A. Hoffmann to AC as part of the “Climate Change and Evolution” symposium during the XIV Congress of the European Society for Evolutionary Biology (ESEB) held at Lisbon (Portugal) in August 2013. The Evolutionary Biology Centre (EBC) Graduate School on Genomes and Phenotypes from Uppsala University is recognized for promoting AC participation in this meeting. AGROSAVIA’s Department for Research Capacity Building is credited for granting time to AC to carry out synergistic discussions and progress meetings during 2016 and 2017 in order to pursue this mini-review, as well as for sponsoring FL-H’s internship during 2018. We thank D. Royer for the Cenozoic temperature, CO2, and species richness dataset. Special thanks are given to M. J. Torres-Urrego for support while drafting and revising this mini-review. The topic editor and the two reviewers are recognized for their thoughtful suggestions to improve the scope of the mini-review, as well as for making possible the insightful special issue on “Coping with Climate Change: A Genomic Perspective on Thermal Adaptation.”
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2020.564515/full#supplementary-material
FIGURE S1 | Past and future of thermal and CO2 variation, and their correlates with past biodiversity. (A) Temperature and richness of plant species (from pollen) for the Cenozoic Era (65 Mya – present). Temperature estimates (Supplementary Table S1) were computed by Hansen et al. (2013) using the original δ18O record from Zachos et al. (2008). Richness of plant species from pollen data (Supplementary Table S1) is based on 15 Neotropical stratigraphic sections inspected by Jaramillo et al. (2006). This profile goes from 65 to 20 Mya due to a lack of more recent suitable sampling records. (B) Projections of the near-surface temperature anomalies to 2,050 (Supplementary Table S2), which follow the CIMP5 RCP 8.5 scenario from the KNMI (http://climexp.knmi.nl/) repository averaged from an original 5-min resolution. Light gray shaded areas depict minimum and maximum estimates. (C) Atmospheric CO2 and richness of plant species (as in A) for the Cenozoic Era (65 Mya – present). CO2 records are an updated version (Supplementary Table S1) derived from Royer and Chernoff (2013), originally compiled by Beerling and Royer (2011). (D) Projected CO2 concentration (ppm) to 2,050 also follow the CIMP5 RCP 8.5 scenario, as in B (Supplementary Table S3).
TABLE S1 | Dataset of temperature, atmospheric CO2, and richness of plant species for the Cenozoic Era (65 Mya – present for temperature and CO2, and 65–20 Mya for richness of plant species). Temperature estimates were computed by Hansen et al. (2013) from five-point running means of the original temporal resolution of the δ18O record from Zachos et al. (2008), a profile of surface low-magnesium calcitic fossils (including planktonic foraminifera, belemintes, brachiopods, and bivalves) that was lower during periods with warmer seawater. Atmospheric CO2 corresponds to an updated version from Royer and Chernoff (2013), originally compiled by Beerling and Royer (2011). Richness of plant species is based on pollen data from Jaramillo et al. (2006), who analyzed 1,530 samples from 15 stratigraphic sections in Colombia and Venezuela (Neotropics).
TABLE S2 | Projections of thermal variation to 2,050. Simulation of Near-Surface Air Temperature Anomalies (°C) from 1,860 to 2,050 follow the CIMP5 RCP 8.5 scenario from the KNMI (http://climexp.knmi.nl/) database averaged from an original 5 min resolution. Minimum and maximum temperature estimates were generated by the coupled ACCESS v.1.0 model specifically designed for the CIMP5 project (Kowalczyk et al., 2013).
TABLE S3 | Projections of CO2 concentration (ppm) to 2,050. Simulations follow the CIMP5 RCP 8.5 scenario from 1,860 to 2,050 available at KNMI (http://climexp.knmi.nl/) database averaged from an original 5 min resolution.
References
Abbott, R., Albach, D., Ansell, S., Arntzen, J. W., Baird, S. J. E., Bierne, N., et al. (2013). Hybridization and speciation. J. Evol. Biol. 26, 229–246.
Abdelmula, A. A., Link, W., Von Kittlitz, E., and Stelling, D. (1999). Heterosis and inheritance of drought tolerance in Faba Bean, Vicia Faba L. Plant Breed. 118, 485–490. doi: 10.1046/j.1439-0523.1999.00411.x
Alsos, I. G., Alm, T., Normand, S., and Brochmann, C. (2009). Past and future range shifts and loss of diversity in Dwarf Willow (Salix Herbacea L.). Inferred from genetics, fossils and modelling. Glob. Ecol. Biogeogr. 18, 223–239. doi: 10.1111/j.1466-8238.2008.00439.x
Andresen, L. C., Müller, C., De Dato, G., Dukes, J. S., Emmett, B. A., Estiarte, M., et al. (2016). Shifting impacts of climate change. Adv. Ecol. Res. 55, 437–473.
Barghi, N., Hermisson, J., and Schlötterer, C. (2020). Polygenic adaptation: a unifying framework to understand positive selection. Nat. Rev. Genet. doi: 10.1038/s41576-020-0250-z
Barnosky, A. D., Hadly, E. A., Gonzalez, P., Head, J., Polly, P. D., Lawing, A. M., et al. (2017). Merging paleobiology with conservation biology to guide the future of terrestrial ecosystems. Science 355: eaah4787.
Baum, D. A., Smith, S. D., and Donovan, S. S. (2005). The tree-thinking challenge. Science 310, 979–970. doi: 10.1126/science.1117727
Becher, H., Jackson, B. C., and Charlesworth, B. (2020). Patterns of genetic variability in genomic regions with low rates of recombination. Curr. Biol. 30, 94.e3–100.e3.
Becklin, K. M., Walker, S. M. II, Way, D. A., and Ward, J. K. (2017). Co2 studies remain key to understanding a future world. New Phytol. 214, 34–40. doi: 10.1111/nph.14336
Beerli, P. (2006). Comparison of Bayesian and maximum-likelihood inference of population genetic parameters. Bioinformatics 22, 341–345. doi: 10.1093/bioinformatics/bti803
Beerling, D. J., and Royer, D. L. (2011). Convergent cenozoic Co2 history. Nat. Geosci. 4, 418–420. doi: 10.1038/ngeo1186
Beyer, R. M., Krapp, M., and Manica, A. (2020). High-resolution terrestrial climate, bioclimate and vegetation for the last 120,000 years. Sci. Data 7:236. doi: 10.1038/s41597-020-0552-1
Blair, M. W., Cortes, A. J., Farmer, A. D., Huang, W., Ambachew, D., Penmetsa, R. V., et al. (2018). Uneven recombination rate and linkage disequilibrium across a reference Snp map for common Bean (Phaseolus Vulgaris L.). PLoS One 13:e0189597. doi: 10.1371/journal.pone.0189597
Blair, M. W., Cortés, A. J., Penmetsa, R. V., Farmer, A., Carrasquilla-Garcia, N., and Cook, D. R. (2013). A high-throughput snp marker system for parental polymorphism screening, and diversity analysis in common Bean (Phaseolus Vulgaris L.). Theoret. Appl. Genet. 126, 535–548. doi: 10.1007/s00122-012-1999-z
Blair, M. W., Cortés, A. J., and This, D. (2016). Identification of an Erecta gene and its drought adaptation associations with wild and cultivated common Bean. Plant Sci. 242, 250–259. doi: 10.1016/j.plantsci.2015.08.004
Blair, M. W., Soler, A., and Cortés, A. J. (2012). Diversification and population structure in common Beans (Phaseolus Vulgaris L.). PLoS One 7:e49488. doi: 10.1371/journal.pone.0049488
Bridle, J. R., and Vines, T. H. (2007). Limits to evolution at range margins: when and why does adaptation fail? Trends Ecol. Evol. 22, 140–147. doi: 10.1016/j.tree.2006.11.002
Bruelheide, H., Dengler, J., Purschke, O., Lenoir, J., Jiménez-Alfaro, B., Hennekens, S. M., et al. (2018). Global trait–environment relationships of plant communities. Nat. Ecol. Evol. 2, 1906–1917.
Bryant, D., Bouckaert, R., Felsenstein, J., Rosenberg, N. A., and Roychoudhury, A. (2012). Inferring species trees directly from biallelic genetic markers: bypassing gene trees in a full coalescent analysis. Mol. Biol. Evol. 29, 1917–1932. doi: 10.1093/molbev/mss086
Bush, M. B., Silman, M. R., Mcmichael, C., and Saatchi, S. (2008). Fire, climate change and biodiversity in amazonia: a late-holocene perspective. Philos. Trans. R. Soc. Lond. B Biol. Sci. 363, 1795–1702. doi: 10.1098/rstb.2007.0014
Bustamante, C. D., Wakeley, J., Sawyer, S., and Hartl, D. L. (2001). Directional selection and the site-frequency spectrum. Genetics 159, 1779–1788.
Cardenas, M. L., Gosling, W. D., Sherlock, S. C., Poole, I., Pennington, R. T., and Mothes, P. (2011). The response of vegetation on the Andean Flank in Western Amazonia to pleistocene climate change. Science 331, 1055–1058. doi: 10.1126/science.1197947
Chen, I. C., Hill, J. K., Ohlemuller, R., Roy, D. B., and Thomas, C. D. (2011). Rapid range shifts of species associated with high levels of climate warming. Science 333, 1024–1026. doi: 10.1126/science.1206432
Chen, Z., Farrell, A. P., Matala, A., and Narum, S. R. (2018). Mechanisms of thermal adaptation and evolutionary potential of conspecific populations to changing environments. Mol. Ecol. 27, 659–674. doi: 10.1111/mec.14475
Chown, S. L., Hoffmann, A. A., Kristensen, T. N., Angilletta, M. J., Stenseth, N. C., and Pertoldi, C. (2010). Adapting to climate change: a perspective from evolutionary physiology. Clim. Res. 43, 3–15. doi: 10.3354/cr00879
Cornuet, J. M., Pudlo, P., Veyssier, J., Dehne-Garcia, A., Gautier, M., Leblois, R., et al. (2014). Diyabc V2.0: a software to make approximate bayesian computation inferences about population history using single nucleotide polymorphism, DNA sequence and microsatellite data. Bioinformatics 30, 1187–1189. doi: 10.1093/bioinformatics/btt763
Cortés, A. J. (2013). On the origin of the common Bean (Phaseolus Vulgaris L.). Am. J. Plant Sci. 4, 1998–2000.
Cortés, A. J., and Blair, M. W. (2018). Genotyping by sequencing and genome – environment associations in wild common bean predict widespread divergent adaptation to drought. Front. Plant Sci. 9:128. doi: 10.3389/fpls.2018.00128
Cortés, A. J., Chavarro, M. C., and Blair, M. W. (2011). Snp marker diversity in common Bean (Phaseolus Vulgaris L.). Theoret. Appl. Genet. 123, 827–845. doi: 10.1007/s00122-011-1630-8
Cortés, A. J., Chavarro, M. C., Madriñán, S., This, D., and Blair, M. W. (2012a). Molecular ecology and selection in the drought-related Asr gene polymorphisms in wild and cultivated common Bean (Phaseolus Vulgaris L.). BMC Genet. 13:58. doi: 10.1186/1471-2156-13-58
Cortés, A. J., This, D., Chavarro, C., Madriñán, S., and Blair, M. W. (2012b). Nucleotide diversity patterns at the drought-related Dreb2 encoding genes in wild and cultivated common Bean (Phaseolus Vulgaris L.). Theoret. Appl. Genet. 125, 1069–1085. doi: 10.1007/s00122-012-1896-5
Cortés, A. J., Garzón, L. N., Valencia, J. B., and Madriñán, S. (2018a). On the causes of rapid diversification in the páramos: isolation by ecology and genomic divergence in Espeletia. Front. Plant Sci. 9:1700. doi: 10.3389/fpls.2018.01700
Cortés, A. J., Skeen, P., Blair, M. W., and Chacón-Sánchez, M. I. (2018b). Does the genomic landscape of species divergence in Phaseolus Beans coerce parallel signatures of adaptation and domestication? Front. Plant Sci. 9:1816. doi: 10.3389/fpls.2018.01816
Cortés, A. J., Monserrate, F., Ramírez-Villegas, J., Madriñán, S., and Blair, M. W. (2013). Drought tolerance in wild plant populations: the case of common Beans (Phaseolus Vulgaris L.). PLoS One 8:e62898. doi: 10.1371/journal.pone.0062898
Cortés, A. J., Waeber, S., Lexer, C., Sedlacek, J., Wheeler, J. A., Van Kleunen, M., et al. (2014). Small-scale patterns in snowmelt timing affect gene flow and the distribution of genetic diversity in the Alpine Dwarf Shrub Salix Herbacea. Heredity 113, 233–239. doi: 10.1038/hdy.2014.19
Cortés, A. J., and Wheeler, J. A. (2018). “The environmental heterogeneity of mountains at a fine scale in a changing world,” in Mountains, Climate, and Biodiversity, eds C. Hoorn, A. Perrigo, and A. Antonelli (New York, NY: Wiley).
Cotto, O., Wessely, J., Georges, D., Klonner, G., Schmid, M., Dullinger, S., et al. (2017). A dynamic eco-evolutionary model predicts slow response of alpine plants to climate warming. Nat. Commun. 8:15399.
Cros, D., Mbo-Nkoulou, L., Bell, J. M., Oum, J., Masson, A., Soumahoro, M., et al. (2019). Within-family genomic selection in rubber tree (Hevea Brasiliensis) increases genetic gain for rubber production. Ind. Crops Prod. 138:111464. doi: 10.1016/j.indcrop.2019.111464
Crossa, J., Burgueno, J., Dreisigacker, S., Vargas, M., Herrera-Foessel, S. A., Lillemo, M., et al. (2007). Association analysis of historical bread wheat germplasm using additive genetic covariance of relatives and population structure. Genetics 177, 1889–1913. doi: 10.1534/genetics.107.078659
Crossa, J., Jarquin, D., Franco, J., Perez-Rodriguez, P., Burgueno, J., Saint-Pierre, C., et al. (2016). Genomic prediction of gene bank wheat landraces. G3 6, 1819–1834. doi: 10.1534/g3.116.029637
Crossa, J., Martini, J. W. R., Gianola, D., Perez-Rodriguez, P., Jarquin, D., Juliana, P., et al. (2019). Deep Kernel and deep learning for genome-based prediction of single traits in multienvironment breeding trials. Front. Genet. 10:1168. doi: 10.3389/fgene.2019.01168
Crossa, J., Perez-Rodriguez, P., Cuevas, J., Montesinos-Lopez, O., Jarquin, D., De Los Campos, G., et al. (2017). Genomic selection in plant breeding: methods, models, and perspectives. Trends Plant Sci. 22, 961–975.
Cruickshank, T. E., and Hahn, M. W. (2014). Reanalysis suggests that genomic Islands of speciation are due to reduced diversity, not reduced gene flow. Mol. Ecol. 23, 3133–3157. doi: 10.1111/mec.12796
Csilléry, K., Blum, M. G. B., Gaggiotti, O. E., and François, O. (2010). Approximate Bayesian computation (Abc) in practice. Trends Ecol. Evol. 25, 410–418. doi: 10.1016/j.tree.2010.04.001
Csillery, K., Rodriguez-Verdugo, A., Rellstab, C., and Guillaume, F. (2018). Detecting the genomic signal of polygenic adaptation and the role of epistasis in evolution. Mol. Ecol. 27, 606–612. doi: 10.1111/mec.14499
Cuesta, F., Tovar, C., Llambí, L. D., Gosling, W. D., Halloy, S., Carilla, J., et al. (2019). Thermal Niche traits of high alpine plant species and communities across the tropical andes and their vulnerability to global warming. J. Biogeogr. 47, 408–420. doi: 10.1111/jbi.13759
Darwin, C. R. (1874). The Descent of Man and Selection in Relation to Sex. New York, NY: Hurst and Company.
de los Campos, G., Hickey, J. M., Pong-Wong, R., Daetwyler, H. D., and Calus, M. P. (2013). Whole-genome regression and prediction methods applied to plant and animal breeding. Genetics 193, 327–345. doi: 10.1534/genetics.112.143313
Desta, Z. A., and Ortiz, R. (2014). Genomic selection: genome-wide prediction in plant improvement. Trends Plant Sci. 19, 592–601. doi: 10.1016/j.tplants.2014.05.006
Ding, W. N., Ree, R. H., Spicer, R. A., and Xing, Y. W. (2020). Ancient orogenic and monsoon-driven assembly of the world’s richest temperate alpine flora. Science 369, 578–581. doi: 10.1126/science.abb4484
Dolgova, O., Rego, C., Calabria, G., Balanya, J., Pascual, M., Rezende, E. L., et al. (2010). Genetic constraints for thermal coadaptation in Drosophila Subobscura. BMC Evol. Biol. 10:363. doi: 10.1186/1471-2148-10-363
Ellegren, H., and Galtier, N. (2016). Determinants of genetic diversity. Nat. Rev. Genet. 17, 422–433.
Ellegren, H., and Wolf, J. B. W. (2017). Parallelism in genomic landscapes of differentiation, conserved genomic features and the role of linked selection. J. Evol. Biol. 30, 1516–1518. doi: 10.1111/jeb.13113
Elmendorf, S. C., Henry, G. H. R., Hollister, R. D., Björk, R. G., Bjorkman, A. D., Callaghan, T. V., et al. (2012). Global assessment of experimental climate warming on tundra vegetation: heterogeneity over space and time. Ecol. Lett. 15, 164–175. doi: 10.1111/j.1461-0248.2011.01716.x
Elsen, P. R., Monahan, W. B., Dougherty, E. R., and Merenlender, A. M. (2020). Keeping pace with climate change in global terrestrial protected areas. Sci. Adv. 6:eaay0814. doi: 10.1126/sciadv.aay0814
Exposito-Alonso, M., Burbano, H. A., Bossdorf, O., Nielsen, R., and Weigel, D. (2019). Natural selection on the Arabidopsis thaliana genome in present and future climates. Nature 573, 126–129. doi: 10.1038/s41586-019-1520-9
Falconer, D. S., and Mackay, T. F. C. (1996). Introduction to Quantitative Genetics. Essex: Longman.
Feder, J. L., Gejji, R., Yeaman, S., and Nosil, P. (2012). Establishment of new mutations under divergence and genome hitchhiking. Philos. Trans. R. Soc. Lond. B Biol. Sci. 367, 461–474. doi: 10.1098/rstb.2011.0256
Feder, J. L., and Nosil, P. (2010). The efficacy of divergence hitchhiking in generating genomic islands during ecological speciation. Evolution 64, 1729–1747. doi: 10.1111/j.1558-5646.2009.00943.x
Feeley, K., Martinez-Villa, J., Perez, T., Silva Duque, A., Triviño Gonzalez, D., and Duque, A. (2020). The thermal tolerances, distributions, and performances of tropical montane tree species. Front. For. Glob. Change 3:25. doi: 10.3389/ffgc.2020.00025
Fitzpatrick, S. W., Gerberich, J. C., Kronenberger, J. A., Angeloni, L. M., and Funk, W. C. (2015). Locally adapted traits maintained in the face of high gene flow. Ecol. Lett. 18, 37–47. doi: 10.1111/ele.12388
Flantua, S. G. A., O’dea, A., Onstein, R. E., Giraldo, C., and Hooghiemstra, H. (2019). The flickering connectivity system of the North Andean Páramos. J. Biogeogr. 46, 1808–1825. doi: 10.1111/jbi.13607
Forester, B. R., Jones, M. R., Joost, S., Landguth, E. L., and Lasky, J. R. (2016). Detecting spatial genetic signatures of local adaptation in heterogeneous landscapes. Mol. Ecol. 25, 104–120. doi: 10.1111/mec.13476
Fragata, I., Lopes-Cunha, M., Barbaro, M., Kellen, B., Lima, M., Santos, M. A., et al. (2014). How much can history constrain adaptive evolution? A real-time evolutionary approach of inversion polymorphisms in Drosophila Subobscura. J. Evol. Biol. 27, 2727–2738. doi: 10.1111/jeb.12533
Fragata, I., Simões, P., Matos, M., Szathmáry, E., and Santos, M. (2018). playing evolution in the laboratory: from the first major evolutionary transition to global warming. Europhys. Lett. 122:38001. doi: 10.1209/0295-5075/122/38001
Franks, P. J., Adams, M. A., Amthor, J. S., Barbour, M. M., Berry, J. A., Ellsworth, D. S., et al. (2013). Sensitivity of plants to changing atmospheric Co2 concentration: from the geological past to the next century. New Phytol. 197, 1077–1094. doi: 10.1111/nph.12104
Franks, S. J., and Hoffmann, A. A. (2012). Genetics of climate change adaptation. Annu. Rev. Genet. 46, 185–208.
Franks, S. J., Weber, J. J., and Aitken, S. N. (2014). Evolutionary and plastic responses to climate change in terrestrial plant populations. Evol. Appl. 7, 123–139. doi: 10.1111/eva.12112
Galeano, C. H., Cortés, A. J., Fernandez, A. C., Soler, A., Franco-Herrera, N., Makunde, G., et al. (2012). Gene-based single nucleotide polymorphism markers for genetic and association mapping in common Bean. BMC Genet. 13:48. doi: 10.1186/1471-2156-13-48
Garah, K., and Bentouati, A. (2019). Using the maxent model for assessing the impact of climate change on the Aurasian Aleppo pine distribution in algeria. Afr. J. Ecol. 57, 500–511. doi: 10.1111/aje.12630
Garner, J. B., Douglas, M. L., Williams, S. R., Wales, W. J., Marett, L. C., Nguyen, T. T., et al. (2016). Genomic selection improves heat tolerance in dairy cattle. Sci. Rep. 6:34114.
Gianola, D., Fernando, R. L., and Stella, A. (2006). Genomic-assisted prediction of genetic value with semiparametric procedures. Genetics 173, 1761–1776. doi: 10.1534/genetics.105.049510
Gibson, M. J. S., and Moyle, L. C. (2020). Regional differences in the abiotic environment contribute to genomic divergence within a wild tomato species. Mol. Ecol. 29, 2204–2217. doi: 10.1111/mec.15477
Gompert, Z., Comeault, A. A., Farkas, T. E., Feder, J. L., Parchman, T. L., Buerkle, C. A., et al. (2014). Experimental evidence for ecological selection on genome variation in the wild. Ecol. Lett. 17, 369–379. doi: 10.1111/ele.12238
Gonzalez, A., Germain, R. M., Srivastava, D. S., Filotas, E., Dee, L. E., Gravel, D., et al. (2020). Scaling-up biodiversity-ecosystem functioning research. Ecol. Lett. 23, 757–776.
Grattapaglia, D., Silva-Junior, O. B., Resende, R. T., Cappa, E. P., Muller, B. S. F., Tan, B., et al. (2018). Quantitative genetics and genomics converge to accelerate forest tree breeding. Front. Plant Sci. 9:1693. doi: 10.3389/fpls.2018.01693
Hällfors, M. H., Antao, L. H., Itter, M., Lehikoinen, A., Lindholm, T., Roslin, T., et al. (2020). Shifts in timing and duration of breeding for 73 boreal bird species over four decades. PNAS 117, 18557–18565. doi: 10.1073/pnas.1913579117
Hancock, A. M., Brachi, B., Faure, N., Horton, M. W., Jarymowycz, L. B., Sperone, F. G., et al. (2011). Adaptation to climate across the Arabidopsis Thaliana genome. Science 334, 83–86. doi: 10.1126/science.1209244
Hansen, J., Sato, M., Russell, G., and Kharecha, P. (2013). Climate sensitivity, sea level and atmospheric carbon dioxide. Philos. Trans. A Math. Phys. Eng. Sci. 371:20120294. doi: 10.1098/rsta.2012.0294
Hirschhorn, J. N., and Daly, M. J. (2005). Genome-wide association studies for common diseases and complex traits. Nat. Rev. Genet. 6, 95–108. doi: 10.1038/nrg1521
Hoffmann, A. A., and Sgro, C. M. (2011). Climate change and evolutionary adaptation. Nature 470, 479–485.
Huber, C. D., Degiorgio, M., Hellmann, I., and Nielsen, R. (2016). Detecting recent selective sweeps while controlling for mutation rate and background selection. Mol. Ecol. 25, 142–156. doi: 10.1111/mec.13351
Hughes, C. (2006). From the cover: Island radiation on a continental scale: exceptional rates of plant diversification after uplift of the andes. Proc. Natl. Acad. Sci. U.S.A. 103, 10334–10339. doi: 10.1073/pnas.0601928103
Irwin, D. E., Alcaide, M., Delmore, K. E., Irwin, J. H., and Owens, G. L. (2016). Recurrent selection explains parallel evolution of genomic regions of high relative but low absolute differentiation in a ring species. Mol. Ecol. 25, 4488–4507. doi: 10.1111/mec.13792
Isabel, N., Holliday, J. A., and Aitken, S. N. (2020). Forest genomics: advancing climate adaptation, forest health, productivity, and conservation. Evol. Appl. 13, 3–10. doi: 10.1111/eva.12902
Jaramillo, C., Rueda, M. J., and Mora, G. (2006). Cenozoic plant diversity in the neotropics. Science 311, 1893–1896. doi: 10.1126/science.1121380
Jay, F., Manel, S., Alvarez, N., Durand, E. Y., Thuiller, W., Holderegger, R., et al. (2012). Forecasting changes in population genetic structure of alpine plants in response to global warming. Mol. Ecol. 21, 2354–2368. doi: 10.1111/j.1365-294x.2012.05541.x
Jenouvrier, S., Caswell, H., Barbraud, C., Holland, M., Stroeve, J., and Weimerskirch, H. (2009). Demographic models and Ipcc climate projections predict the decline of an emperor penguin population. PNAS 106, 1844–1847. doi: 10.1073/pnas.0806638106
Jezkova, T., and Wiens, J. J. (2016). Rates of change in climatic niches in plant and animal populations are much slower than projected climate change. Proc. Biol. Sci. 283:20162104. doi: 10.1098/rspb.2016.2104
Juliana, P., Montesinos-López, O. A., Crossa, J., Mondal, S., González Pérez, L., Poland, J., et al. (2019). Integrating genomic-enabled prediction and high-throughput phenotyping in breeding for climate-resilient bread wheat. Theoret. Appl. Genet. 132, 177–194. doi: 10.1007/s00122-018-3206-3
Kawecki, T. J., Lenski, R. E., Ebert, D., Hollis, B., Olivieri, I., and Whitlock, M. C. (2012). Experimental evolution. Trends Ecol. Evol. 27, 547–560.
Kelleher, C. T., Wilkin, J., Zhuang, J., Cortés, A. J., Quintero, ÁL. P., Gallagher, T. F., et al. (2012). Snp discovery, gene diversity, and linkage disequilibrium in wild populations of Populus Tremuloides. Tree Genet. Genomes 8, 821–829. doi: 10.1007/s11295-012-0467-x
Kowalczyk, E. A., Stevens, L., Law, R. M., Dix, M., Wang, Y. P., Harman, I. N., et al. (2013). The land surface model component of access: description and impact on the simulated surface climatology. Aust. Meteorol. Oceanogr. 63, 65–82. doi: 10.22499/2.6301.005
Kristensen, T. N., Ketola, T., and Kronholm, I. (2018). Adaptation to environmental stress at different timescales. Ann. N. Y. Acad. Sci. [Epub ahead of print] doi: 10.1111/nyas.13974
Laayouni, H., Garcia-Franco, F., Chavez-Sandoval, B. E., Trotta, V., Beltran, S., Corominas, M., et al. (2007). Thermal evolution of gene expression profiles in Drosophila Subobscura. BMC Evol. Biol. 7:42. doi: 10.1186/1471-2148-7-42
Lambert, C. G., and Black, L. J. (2012). Learning from our gwas mistakes: from experimental design to scientific method. Biostatistics 13, 195–203. doi: 10.1093/biostatistics/kxr055
Lecheta, M. C., Awde, D. N., O’leary, T. S., Unfried, L. N., Jacobs, N. A., Whitlock, M. H., et al. (2020). Integrating Gwas and transcriptomics to identify the molecular underpinnings of thermal stress responses in Drosophila Melanogaster. Front. Genet. 11:658. doi: 10.3389/fgene.2020.00658
Lehnert, S. J., Kess, T., Bentzen, P., Kent, M. P., Lien, S., Gilbey, J., et al. (2019). Genomic signatures and correlates of widespread population declines in salmon. Nat. Commun. 10:2996.
Leinonen, P. H., Remington, D. L., and Savolainen, O. (2011). Local adaptation, phenotypic differentiation, and hybrid fitness in diverged natural populations of Arabidopsis Lyrata. Evolution 65, 90–107. doi: 10.1111/j.1558-5646.2010.01119.x
Lenoir, J., Bertrand, R., Comte, L., Bourgeaud, L., Hattab, T., Murienne, J., et al. (2020). Species better track climate warming in the oceans than on land. Nat. Ecol. Evol. 4, 1044–1059. doi: 10.1038/s41559-020-1198-2
Libbrecht, M. W., and Noble, W. S. (2015). Machine learning applications in genetics and genomics. Nat. Rev. Genet. 16, 321–332. doi: 10.1038/nrg3920
Little, C. J., Wheeler, J. A., Sedlacek, J., Cortés, A. J., and Rixen, C. (2016). Small-scale drivers: the importance of nutrient availability and snowmelt timing on performance of the Alpine Shrub Salix Herbacea. Oecologia 180, 1015–1024. doi: 10.1007/s00442-015-3394-3
Liu, S., Cornille, A., Decroocq, S., Tricon, D., Chague, A., Eyquard, J. P., et al. (2019). The complex evolutionary history of apricots: species divergence, gene flow and multiple domestication events. Mol. Ecol. 28, 5299–5314. doi: 10.1111/mec.15296
Logan, M. L., and Cox, C. L. (2020). Genetic constraints, transcriptome plasticity, and the evolutionary response to climate change. Front. Genet. doi: 10.3389/fgene.2020.538226
López-Hernández, F., and Cortés, A. J. (2019). Last-generation genome–environment associations reveal the genetic basis of heat tolerance in common Bean (Phaseolus Vulgaris L.). Front. Genet. 10:22. doi: 10.3389/fgene.2019.00954
Lotterhos, K. E., and Whitlock, M. C. (2015). The relative power of genome scans to detect local adaptation depends on sampling design and statistical method. Mol. Ecol. 24, 1031–1046. doi: 10.1111/mec.13100
Lstiburek, M., Schueler, S., El-Kassaby, Y. A., Hodge, G. R., Stejskal, J., Korecky, J., et al. (2020). In Situ genetic evaluation of european larch across climatic regions using marker-based pedigree reconstruction. Front. Genet. 11:28. doi: 10.3389/fgene.2020.00028
Lynch, M., and Ritland, K. (1999). Estimation of pairwise relatedness with molecular markers. Genetics 152, 1753–1766.
Lyons, S. K., Amatangelo, K. L., Behrensmeyer, A. K., Bercovici, A., Blois, J. L., Davis, M., et al. (2016). Holocene shifts in the assembly of plant and animal communities implicate human impacts. Nature 529, 80–83. doi: 10.1038/nature16447
Madriñán, S., Cortés, A. J., and Richardson, J. E. (2013). Páramo is the world’s fastest evolving and coolest biodiversity hotspot. Front. Genet. 4:192. doi: 10.3389/fgene.2013.00192
Mallard, F., Nolte, V., Tobler, R., Kapun, M., and Schlötterer, C. (2018). A simple geneticbasis of adaptation to a novel thermal environment results in complex metabolic rewiring in Drosophila. A simple genetic basis of adaptation to a novel thermal environment results in complex metabolic rewiring in Drosophila. Genome Biol. 19:119.
Marques, D. A., Meier, J. I., and Seehausen, O. (2019). A combinatorial view on speciation and adaptive radiation. Trends Ecol. Evol. 34, 531–544. doi: 10.1016/j.tree.2019.02.008
Matos, M., Simoes, P., Santos, M. A., Seabra, S. G., Faria, G. S., Vala, F., et al. (2015). History, chance and selection during phenotypic and genomic experimental evolution: replaying the tape of life at different levels. Front. Genet. 6:71. doi: 10.3389/fgene.2015.00071
Maynard Smith, J., and Haigh, J. (1974). The hitch-hiking effect of a favourable gene. Genet. Res. 23, 23–35. doi: 10.1017/s0016672300014634
McKown, A. D., Klapste, J., Guy, R. D., El-Kassaby, Y. A., and Mansfield, S. D. (2018). Ecological genomics of variation in bud-break phenology and mechanisms of response to climate warming in Populus Trichocarpa. New Phytol. 220, 300–316. doi: 10.1111/nph.15273
Meuwissen, T. H. E., Hayes, B. J., and Goddard, M. E. (2001). Prediction of total genetic value using genome-wide dense marker maps. Genetics 157, 1819–1829.
Miller, A. D., Coleman, M. A., Clark, J., Cook, R., Naga, Z., Doblin, M. A., et al. (2020). Local thermal adaptation and limited gene flow constrain future climate responses of a marine ecosystem engineer. Evol. Appl. 13, 918–934. doi: 10.1111/eva.12909
Mohapatra, J., Singh, C., Hamid, M., Verma, A., Semwal, S. C., Gajmer, B., et al. (2019). Modelling Betula Utilis distribution in response to climate-warming scenarios in Hindu-Kush Himalaya using random forest. Biodiv. Conserv. 28, 2295–2317. doi: 10.1007/s10531-019-01731-w
Montesinos-Lopez, A., Montesinos-Lopez, O. A., Gianola, D., Crossa, J., and Hernandez-Suarez, C. M. (2018). Multi-environment genomic prediction of plant traits using deep learners with dense architecture. G3 8, 3813–3828.
Montesinos-Lopez, O. A., Martin-Vallejo, J., Crossa, J., Gianola, D., Hernandez-Suarez, C. M., Montesinos-Lopez, A., et al. (2019). A benchmarking between deep learning, support vector machine and bayesian threshold best linear unbiased prediction for predicting ordinal traits in plant breeding. G3 9, 601–618.
Morris, B. B. G. P., and Borevitz, J. O. (2011). Genome-wide association studies in plants: the missing heritability is in the field. Genome Biol. 12:232. doi: 10.1186/gb-2011-12-10-232
Nachman, M. W., and Payseur, B. A. (2012). Recombination rate variation and speciation: theoretical predictions and empirical results from rabbits and mice. Philos. Trans. R. Soc. Lond. B Biol. Sci. 367, 409–421. doi: 10.1098/rstb.2011.0249
Nadachowska-Brzyska, K., Burri, R., Smeds, L., and Ellegren, H. (2016). Psmc analysis of effective population sizes in molecular ecology and its application to black-and-white Ficedula Flycatchers. Mol. Ecol. 25, 1058–1072. doi: 10.1111/mec.13540
Nathan, R., and Muller-Landau, H. C. (2000). Spatial patterns of seed dispersal, their determinants and consequences for recruitment. Trends Ecol. Evol. 15, 278–285. doi: 10.1016/s0169-5347(00)01874-7
Nicotra, A. B., Atkin, O. K., Bonser, S. P., Davidson, A. M., Finnegan, E. J., Mathesius, U., et al. (2010). Plant phenotypic plasticity in a changing climate. Trends Plant Sci. 15, 684–692. doi: 10.1016/j.tplants.2010.09.008
North, A., Pennanen, J., Ovaskainen, O., and Laine, A.-L. (2011). Local adaptation in a changing world: the roles of gene-flow, mutation, and sexual reproduction. Evolution 65, 79–89. doi: 10.1111/j.1558-5646.2010.01107.x
Nosil, P., and Feder, J. L. (2011). Genomic divergence during speciation: causes and consequences. Philos. Trans. R. Soc. B Biol. Sci. 367, 332–342. doi: 10.1098/rstb.2011.0263
Ørsted, M., Hoffmann, A. A., Rohde, P. D., Sørensen, P., and Kristensen, T. N. (2019). Strong impact of thermal environment on the quantitative genetic basis of a key stress tolerance trait. Heredity 122, 315–325. doi: 10.1038/s41437-018-0117-7
Otto, S. P., and Day, T. (2007). A Biologist’s Guide to Mathematical Modeling in Ecology and Evolution. Princeton, NJ: Princeton University Press.
Park, D. S., Willis, C. G., Xi, Z., Kartesz, J. T., Davis, C. C., and Worthington, S. (2020). Machine learning predicts large scale declines in native plant phylogenetic diversity. New Phytol. 227, 1544–1556. doi: 10.1111/nph.16621
Parmesan, C., and Hanley, M. E. (2015). Plants and climate change: complexities and surprises. Ann. Bot. 116, 849–864. doi: 10.1093/aob/mcv169
Payseur, B. A., and Rieseberg, L. H. (2016). A genomic perspective on hybridization and speciation. Mol. Ecol. 25, 2337–2360. doi: 10.1111/mec.13557
Pearson, R. G., Phillips, S. J., Loranty, M. M., Beck, P. S. A., Damoulas, T., Knight, S. J., et al. (2013). Shifts in arctic vegetation and associated feedbacks under climate change. Nat. Clim. Change 3, 673–677. doi: 10.1038/nclimate1858
Pfenninger, M., and Foucault, Q. (2020). Genomic processes underlying rapid adaptation of a natural Chironomus Riparius population to unintendedly applied experimental selection pressures. Mol. Ecol. 29, 536–548. doi: 10.1111/mec.15347
Phillips, S. J., Anderson, R. P., Dudiík, M., Schapire, R. E., and Blair, M. E. (2017). Opening the black box: An open-source release of maxent. Ecography 40, 887–893. doi: 10.1111/ecog.03049
Pluess, A. R., Frank, A., Heiri, C., Lalague, H., Vendramin, G. G., and Oddou-Muratorio, S. (2016). Genome-environment association study suggests local adaptation to climate at the regional scale in Fagus Sylvatica. New Phytol. 210, 589–601. doi: 10.1111/nph.13809
Pritchard, J. K., Pickrell, J. K., and Coop, G. (2010). The genetics of human adaptation: hard sweeps, soft sweeps, and polygenic adaptation. Curr. Biol. 20, R208–R215.
Quintero, I., and Wiens, J. J. (2013). Rates of projected climate change dramatically exceed past rates of climatic niche evolution among vertebrate species. Ecol. Lett. 16, 1095–1103. doi: 10.1111/ele.12144
Ravinet, M., Faria, R., Butlin, R. K., Galindo, J., Bierne, N., Rafajlovic, M., et al. (2017). Interpreting the genomic landscape of speciation: a road map for finding barriers to gene flow. J. Evol. Biol. 30, 1450–1477. doi: 10.1111/jeb.13047
Razgour, O., Forester, B., Taggart, J. B., Bekaert, M., Juste, J., Ibanez, C., et al. (2019). Considering adaptive genetic variation in climate change vulnerability assessment reduces species range loss projections. PNAS 116, 10418–10423. doi: 10.1073/pnas.1820663116
Reichstein, M., Bahn, M., Mahecha, M. D., Kattge, J., and Baldocchi, D. D. (2014). Linking plant and ecosystem functional biogeography. Proc. Natl. Acad. Sci. U.S.A. 111, 13697–13702. doi: 10.1073/pnas.1216065111
Resende, M. D. V. Jr., Resende, M. F. R., Sansaloni, C. P., Petroli, C. D., Missiaggia, A. A., Aguiar, A. M., et al. (2012). Genomic selection for growth and wood quality in Eucalyptus: capturing the missing heritability and accelerating breeding for complex traits in forest trees. New Phytol. 194, 116–128. doi: 10.1111/j.1469-8137.2011.04038.x
Rezende, E. L., Tejedo, M., and Santos, M. (2010). Estimating the adaptive potential of critical thermal limits: methodological problems and evolutionary implications. Funct. Ecol. 25, 111–121. doi: 10.1111/j.1365-2435.2010.01778.x
Richardson, J. E., Madriñán, S., Gómez-Gutiérrez, M. C., Valderrama, E., Luna, J., Banda-R, K., et al. (2019). Using dated molecular phylogenies to help reconstruct geological, climatic, and biological history: Examples from Colombia. Geol. J. 53, 2935–2943. doi: 10.1002/gj.3133
Rippke, U., Ramirez-Villegas, J., Jarvis, A., Vermeulen, S. J., Parker, L., Mer, F., et al. (2016). Timescales of transformational climate change adaptation in Sub-Saharan African agriculture. Nat. Clim. Change 6, 605–609. doi: 10.1038/nclimate2947
Ripple, W. J., Wolf, C., Newsome, T. M., Barnard, P., and Moomaw, W. R. (2020). World Scientists’ warning of a climate emergency. BioScience 70, 8–12.
Rodriguez-Trelles, F., Tarrio, R., and Santos, M. (2013). Genome-wide evolutionary response to a heat wave in Drosophila. Biol. Lett. 9:20130228. doi: 10.1098/rsbl.2013.0228
Roesti, M., Gavrilets, S., Hendry, A. P., Salzburger, W., and Berner, D. (2014). The genomic signature of parallel adaptation from shared genetic variation. Mol. Ecol. 23, 3944–3956. doi: 10.1111/mec.12720
Royer, D. L., and Chernoff, B. (2013). Diversity in neotropical wet forests during the cenozoic is linked more to atmospheric Co2 than temperature. Proc. Biol. Sci. 280:20131024. doi: 10.1098/rspb.2013.1024
Sabeti, P. C., Varilly, P., Fry, B., Lohmueller, J., Hostetter, E., Cotsapas, C., et al. (2007). Genome-wide detection and characterization of positive selection in human populations. Nature 449, 913–918. doi: 10.1038/nature06250
Saint Pierre, C., Crossa, J. L., Bonnett, D., Yamaguchi-Shinozaki, K., and Reynolds, M. P. (2012). Phenotyping transgenic wheat for drought resistance. J. Exp. Bot. 63, 1799–1808. doi: 10.1093/jxb/err385
Santos, M., Castaneda, L. E., and Rezende, E. L. (2012). Keeping pace with climate change: what is wrong with the evolutionary potential of upper thermal limits? Ecol. Evol. 2, 2866–2880. doi: 10.1002/ece3.385
Scherrer, D., and Körner, C. (2011). Topogaphically controlled thermal-habitat differentiation buffers Alpine plant diversity against climate warming. J. Biogeogr. 38, 406–416. doi: 10.1111/j.1365-2699.2010.02407.x
Schrider, D. R., and Kern, A. D. (2018). Supervised machine learning for population genetics: a new paradigm. Trends Genet. 34, 301–312. doi: 10.1016/j.tig.2017.12.005
Seabra, S. G., Fragata, I., Antunes, M. A., Faria, G. S., Santos, M. A., Sousa, V. C., et al. (2017). Different genomic changes underlie adaptive evolution in populations of contrasting history. Mol. Biol. Evol. 35, 549–563. doi: 10.1093/molbev/msx247
Sedlacek, J., Bossdorf, O., Cortés, A. J., Wheeler, J. A., and Van-Kleunen, M. (2014). What role do plant-soil interactions play in the habitat suitability and potential range expansion of the Alpine Dwarf Shrub Salix Herbacea? Basic Appl. Ecol. 15, 305–315. doi: 10.1016/j.baae.2014.05.006
Sedlacek, J., Cortés, A. J., Wheeler, J. A., Bossdorf, O., Hoch, G., Klapste, J., et al. (2016). Evolutionary potential in the Alpine: trait heritabilities and performance variation of the Dwarf Willow Salix Herbacea from different elevations and microhabitats. Ecol. Evol. 6, 3940–3952. doi: 10.1002/ece3.2171
Sedlacek, J., Wheeler, J. A., Cortés, A. J., Bossdorf, O., Hoch, G., Lexer, C., et al. (2015). The response of the Alpine Dwarf Shrub Salix Herbacea to altered snowmelt timing: lessons from a multi-site transplant experiment. PLoS One 10:e0122395. doi: 10.1371/journal.pone.0122395
Sentinella, A. T., Warton, D. I., Sherwin, W. B., Offord, C. A., Moles, A. T., and Wang, Z. (2020). Tropical plants do not have narrower temperature tolerances, but are more at risk from warming because they are close to their upper thermal limits. Glob. Ecol. Biogeogr. 29, 1387–1398. doi: 10.1111/geb.13117
Simões, J. L., Gouvêa, N., and Margis, M. (2003). Small heat shock proteins genes are differentially expressed in distinct varieties of common Bean. Braz. J. Plant Physiol. 15, 33–41. doi: 10.1590/s1677-04202003000100005
Simões, P., and Pascual, M. (2018). Patterns of geographic variation of thermal adapted candidate genes in Drosophila Subobscura sex chromosome arrangements. BMC Evol. Biol. 18:60. doi: 10.1186/s12862-018-1178-1
Singh, R. K., Svystun, T., Aldahmash, B., Jonsson, A. M., and Bhalerao, R. P. (2017). Photoperiod- and temperature-mediated control of phenology in trees - a molecular perspective. New Phytol. 213, 511–524. doi: 10.1111/nph.14346
Slatkin, M. (2008). Linkage disequilibrium — understanding the evolutionary past and mapping the medical future. Nat. Rev. Genet. 9, 477–485. doi: 10.1038/nrg2361
Sørensen, J. G., Kristensen, T. N., and Loeschcke, V. (2003). The evolutionary and ecological role of heat shock proteins. Ecol. Lett. 6, 1025–1037. doi: 10.1046/j.1461-0248.2003.00528.x
Spindel, J. E., Begum, H., Akdemir, D., Collard, B., Redona, E., Jannink, J. L., et al. (2016). Genome-wide prediction models that incorporate de novo gwas are a powerful new tool for tropical rice improvement. Heredity 116, 395–408. doi: 10.1038/hdy.2015.113
Stanton, M. L., Galen, C., and Shore, J. (1997). Population structure along a steep environmental gradient: consequences of flowering time and habitat variation in the snow buttercup, Ranunculus Adoneus. Evolution 51, 79–94. doi: 10.2307/2410962
Steinbauer, M. J., Grytnes, J. A., Jurasinski, G., Kulonen, A., Lenoir, J., Pauli, H., et al. (2018). Accelerated increase in plant species richness on mountain summits is linked to warming. Nature 556, 231–234.
Stephan, W. (2016). Signatures of positive selection: from selective sweeps at individual loci to subtle allele frequency changes in polygenic adaptation. Mol. Ecol. 25, 79–88. doi: 10.1111/mec.13288
Stölting, K. N., Paris, M., Meier, C., Heinze, B., Castiglione, S., Bartha, D., et al. (2015). Genome-wide patterns of differentiation and spatially varying selection between postglacial recolonization lineages of Populus Alba (Salicaceae), a widespread forest tree. New Phytol. 207, 723–734. doi: 10.1111/nph.13392
Street, N. R., Jansson, S., and Hvidsten, T. (2011). A systems biology model of the regulatory network in Populus leaves reveals interacting regulators and conserved regulation. BMC Plant Biol. 11:13. doi: 10.1186/1471-2229-11-13
Sukumaran, S., Crossa, J., Jarquin, D., Lopes, M., and Reynolds, M. P. (2017). Genomic prediction with pedigree and genotype x environment interaction in spring wheat grown in South and West Asia, North Africa, and Mexico. G3 7, 481–495. doi: 10.1534/g3.116.036251
Suontama, M., KláPšTě, J., Telfer, E., Graham, N., Stovold, T., Low, C., et al. (2019). Efficiency of genomic prediction across two Eucalyptus Nitens seed orchards with different selection histories. Heredity 122, 370–379. doi: 10.1038/s41437-018-0119-5
Swarts, K., Gutaker, R. M., Benz, B., Blake, M., Bukowski, R., Holland, J., et al. (2017). Genomic estimation of complex traits reveals ancient maize adaptation to temperate North America. Science 357, 512–515. doi: 10.1126/science.aam9425
Sweet, L. C., Green, T., Heintz, J. G. C., Frakes, N., Graver, N., Rangitsch, J. S., et al. (2019). Congruence between future distribution models and empirical data for an iconic species at Joshua Tree National Park. Ecosphere 10:e0276. doi: 10.1002/ecs2.2763
Tam, V., Patel, N., Turcotte, M., Bosseì, Y., Pareì, G., and Meyre, D. (2019). Benefits and limitations of genome-wide association studies. Nat. Rev. Genet. 20, 467–484. doi: 10.1038/s41576-019-0127-1
Tan, B., Grattapaglia, D., Martins, G. S., Ferreira, K. Z., Sundberg, B. R., and Ingvarsson, P. R. K. (2017). Evaluating the accuracy of genomic prediction of growth and wood traits in two eucalyptus species and their F1 hybrids. BMC Plant Biol. 17:110. doi: 10.1186/s12870-017-1059-6
Technow, F., Schrag, T. A., Schipprack, W., Bauer, E., Simianer, H., and Melchinger, A. E. (2014). Genome properties and prospects of genomic prediction of hybrid performance in a breeding program of maize. Genetics 197, 1343–1355. doi: 10.1534/genetics.114.165860
Tenaillon, O., Rodriguez-Verdugo, A., Gaut, R. L., Mcdonald, P., Bennett, A. F., Long, A. D., et al. (2012). The molecular diversity of adaptive convergence. Science 335, 457–461. doi: 10.1126/science.1212986
Turner, T. L., and Miller, P. M. (2012). Investigating natural variation in drosophila courtship song by the evolve and resequence approach. Genetics 191, 633–642. doi: 10.1534/genetics.112.139337
Valencia, J. B., Mesa, J., León, J. G., Madriñán, S., and Cortés, A. J. (2020). Climate vulnerability assessment of the Espeletia complex in Páramo sky islands of the northern Andes. Front. Ecol. Evol. doi: 10.3389/fevo.2020.565708
Valladares, F., Matesanz, S., Guilhaumon, F., Araujo, M. B., Balaguer, L., Benito-Garzon, M., et al. (2014). The effects of phenotypic plasticity and local adaptation on forecasts of species range shifts under climate change. Ecol. Lett. 17, 1351–1364. doi: 10.1111/ele.12348
Vargas, O. M., Ortiz, E. M., and Simpson, B. B. (2017). Conflicting phylogenomic signals reveal a pattern of reticulate evolution in a recent high-andean diversification (Asteraceae: Astereae: Diplostephium). New Phytol. 214, 1736–1750. doi: 10.1111/nph.14530
Visser, J. A. G. M. D., Elena, S. F., Fragata, I. S., and Matuszewski, S. (2018). The utility of fitness landscapes and big data for predicting evolution. Heredity 121, 401–405. doi: 10.1038/s41437-018-0128-4
Waldvogel, A. M., Feldmeyer, B., Rolshausen, G., Exposito-Alonso, M., Rellstab, C., Kofler, R., et al. (2020a). Evolutionary genomics can improve prediction of species’ responses to climate change. Evol. Lett. 4, 4–18. doi: 10.1002/evl3.154
Waldvogel, A. M., Schreiber, D., Pfenninger, M., and Feldmeyer, B. (2020b). Climate change genomics calls for standardised data reporting. Front. Ecol. Evol. 8:242. doi: 10.3389/fevo.2020.00242
Walther, G. R., Post, E., Convey, P., Menzel, A., Parmesan, C., Beebee, T. J. C., et al. (2002). Ecological responses to recent climate change. Nature 416, 389–395.
Wang, X., Edwards, R. L., Auler, A. S., Cheng, H., Kong, X., Wang, Y., et al. (2017). Hydroclimate changes across the amazon lowlands over the past 45,000 years. Nature 541, 204–207. doi: 10.1038/nature20787
Way, D. A., and Oren, R. (2010). Differential responses to changes in growth temperature between trees from different functional groups and biomes: a review and synthesis of data. Tree Physiol. 30, 669–688. doi: 10.1093/treephys/tpq015
Way, D. A., Oren, R., and Kroner, Y. (2015). The space-time continuum: the effects of elevated Co2 and temperature on trees and the importance of scaling. Plant Cell Environ. 38, 991–1007. doi: 10.1111/pce.12527
Weir, B. S., and Cockerham, C. (1984). Estimating F-Statistics for the analysis of population structure. Evolution 38, 1358–1370. doi: 10.2307/2408641
Wheeler, J. A., Cortés, A. J., Sedlacek, J., Karrenberg, S., Van Kleunen, M., Wipf, S., et al. (2016). The snow and the willows: accelerated spring snowmelt reduces performance in the low-lying Alpine Shrub Salix Herbacea. J. Ecol. 104, 1041–1050. doi: 10.1111/1365-2745.12579
Wheeler, J. A., Hoch, G., Cortés, A. J., Sedlacek, J., Wipf, S., and Rixen, C. (2014). Increased Spring freezing vulnerability for Alpine Shrubs under early snowmelt. Oecologia 175, 219–229. doi: 10.1007/s00442-013-2872-8
Wheeler, J. A., Schnider, F., Sedlacek, J., Cortés, A. J., Wipf, S., Hoch, G., et al. (2015). With a little help from my friends: community facilitation increases performance in the Dwarf Shrub Salix Herbacea. Basic Appl. Ecol. 16, 202–209. doi: 10.1016/j.baae.2015.02.004
Whitlock, C., and Bartlein, P. J. (2003). holocene fire activity as a record of past environmental change. Dev. Quatern. Sci. 1, 479–490. doi: 10.1016/s1571-0866(03)01022-4
Willis, C. G., Ruhfel, B., Primack, R. B., Miller-Rushing, A. J., and Davis, C. C. (2008). Phylogenetic patterns of species loss in Thoreau’s woods are driven by climate change. PNAS 105, 17029–17033. doi: 10.1073/pnas.0806446105
Willis, K. J., and MacDonald, G. M. (2011). Long-term ecological records and their relevance to climate change predictions for a warmer world. Annu. Rev. Ecol. Evol. Syst. 42, 267–287. doi: 10.1146/annurev-ecolsys-102209-144704
Wipf, S., Stöckli, V., Herz, K., and Rixen, C. (2013). The oldest monitoring site of the alps revisited: accelerated increase in plant species Richness on Piz Linard summit since 1835. Plant Ecol. Divers. 6, 447–455. doi: 10.1080/17550874.2013.764943
Wolf, J. B., and Ellegren, H. (2017). Making sense of genomic islands of differentiation in light of speciation. Nat. Rev. Genet. 18, 87–100. doi: 10.1038/nrg.2016.133
Wolkovich, E. M., Cook, B. I., Allen, J. M., Crimmins, T. M., Betancourt, J. L., Travers, S. E., et al. (2012). Warming experiments underpredict plant phenological responses to climate change. Nature 485, 494–497. doi: 10.1038/nature11014
Wray, N. R., Yang, J., Hayes, B. J., Price, A. L., Goddard, M. E., and Visscher, P. M. (2013). Pitfalls of predicting complex traits from Snps. Nat. Rev. Genet. 14, 507–515. doi: 10.1038/nrg3457
Wu, X., Islam, A. S. M. F., Limpot, N., Mackasmiel, L., Mierzwa, J., Cortés, A. J., et al. (2020). Genome-wide Snp identification and association mapping for seed mineral concentration in Mung Bean (Vigna Radiata L.). Front. Genet. 11:656. doi: 10.3389/fgene.2020.00656
Yamagishi, H., Allison, T. D., and Ohara, M. (2005). Effect of snowmelt timing on the genetic structure of an Erythronium Grandiflorum population in an Alpine environment. Ecol. Res. 20, 199–204. doi: 10.1007/s11284-004-0032-7
Yeaman, S., and Otto, S. P. (2011). Establishment and maintenance of adaptive genetic divergence under migration, selection, and drift. Evolution 65, 2123–2129. doi: 10.1111/j.1558-5646.2011.01277.x
Zachos, J. C., Dickens, G. R., and Zeebe, R. E. (2008). An early cenozoic perspective on greenhouse warming and carbon-cycle dynamics. Nature 451, 279–283. doi: 10.1038/nature06588
Zahn, L. M., and Purnell, B. A. (2016). Genes under pressure. Science 354:52. doi: 10.1126/science.354.6308.52
Zellweger, F., De Frenne, P., Lenoir, J., Vangansbeke, P., Verheyen, K., Bernhardt-Römermann, M., et al. (2020). Forest microclimate dynamics drive plant responses to warming. Science 368, 772–775.
Zhou, L., Bawa, R., and Holliday, J. A. (2014). Exome resequencing reveals signatures of demographic and adaptive processes across the genome and range of Black Cottonwood (Populus Trichocarpa). Mol. Ecol. 23, 2486–2499. doi: 10.1111/mec.12752
Keywords: coalescent theory, genome-wide association studies, genome-wide selection scans, genome–environment associations, phylogeography, breeder’s equation, genomic prediction, machine learning
Citation: Cortés AJ, López-Hernández F and Osorio-Rodriguez D (2020) Predicting Thermal Adaptation by Looking Into Populations’ Genomic Past. Front. Genet. 11:564515. doi: 10.3389/fgene.2020.564515
Received: 21 May 2020; Accepted: 24 August 2020;
Published: 25 September 2020.
Edited by:
Margarida Matos, University of Lisbon, PortugalReviewed by:
Anti Vasemägi, Swedish University of Agricultural Sciences, SwedenTakeshi Kawakami, Independent Researcher, Boston, United States
Copyright © 2020 Cortés, López-Hernández and Osorio-Rodriguez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Andrés J. Cortés, YWNvcnRlc0BhZ3Jvc2F2aWEuY28=