 Anna L. K. Nilsson1,2*
Anna L. K. Nilsson1,2* Trond Reitan1,3
Trond Reitan1,3 Thomas Skaugen3Jan Henning L’Abée-Lund3
Thomas Skaugen3Jan Henning L’Abée-Lund3 Marlène Gamelon4Kurt Jerstad5Ole Wiggo Røstad6Tore Slagsvold1
Marlène Gamelon4Kurt Jerstad5Ole Wiggo Røstad6Tore Slagsvold1 Nils C. Stenseth1
Nils C. Stenseth1 L. Asbjørn Vøllestad1Bjørn Walseng7
L. Asbjørn Vøllestad1Bjørn Walseng7- 1Centre for Ecological and Evolutionary Synthesis (CEES), Department of Biosciences, University of Oslo, Oslo, Norway
- 2Norwegian Institute for Nature Research, Bergen, Norway
- 3Norwegian Water Resources and Energy Directorate, Oslo, Norway
- 4Centre for Biodiversity Dynamics, Department of Biology, Norwegian University of Science and Technology, Trondheim, Norway
- 5Jerstad Viltforvaltning, Mandal, Norway
- 6Faculty of Environmental Sciences and Natural Resource Management, Norwegian University of Life Sciences, Ås, Norway
- 7Norwegian Institute for Nature Research, Oslo, Norway
Ecological and evolutionary effects of environmental variation on wild populations are of particular interest in a changing world. Large-scale environmental indices are classically used as environmental explanatory variables to study climate change effects on wild populations. However, these measures neglect the fact that individuals are affected by the local conditions in their home ranges. Freshwater ecosystems are particularly sensitive to climate change induced alterations in precipitation and discharge affecting lower trophic level prey and cascading up the food chain to impact keystone species. Here, we study how small-scale environmental variation at the level of each territory affects fitness-related traits and in particular reproductive success in the white-throated dipper Cinclus cinclus, a bird dependent on aquatic prey. We take advantage of a long-term study of breeding dippers in a natural river system in Norway (1978–2015). We relate daily river discharge and other important hydrological and territory location variables to clutch and brood size simultaneously, while accounting for male and female age, polygyny, and population density. We also address uncertainty in estimates of both clutch and brood size when modeling reproductive success. The most influential variable overall was the altitudinal gradient, where birds breeding at higher altitudes laid fewer eggs and fledged fewer young. Clutch size decreased with increased population density and high temperatures, while it was positively affected by female age. Brood size varied greatly among territories, where some territories had consistently larger broods than others. Increased minimum and maximum temperatures, river discharge above 10 l/s and increased variability in snow depth had a positive effect on brood size, whereas polygyny had a negative effect. In conclusion, territory-specific variables explained a large amount of the variation in the reproductive output in the dipper, emphasizing the importance of the local habitat, and not only climatic variation, in a keystone species in freshwater ecosystems.
Introduction
Understanding how environmental fluctuations affect individual fitness is central in ecology and evolution. Freshwater ecosystems are particularly sensitive to climate change-induced alterations in precipitation and river discharge (Hanssen-Bauer, 2005). Indeed, extreme fluctuations in river discharge cause disturbances in aquatic food webs, including lower trophic level-prey. These disturbances may also cascade up the food chain and negatively affect the reproductive success of higher trophic levels. Also, shifts in timing of droughts and river flooding events might directly reduce reproductive opportunities for species that strongly depend on water to reproduce, such as waterbirds (Chiu et al., 2008; Royan et al., 2013).
Large scale climate indices such as the North Atlantic Oscillation (NAO) are often assumed to represent an environment experienced by all population members (Sæther et al., 2000; Frederiksen et al., 2008; Nilsson et al., 2011a; Gamelon et al., 2017). However, even on a population scale, the environment is rarely uniform (Cole et al., 2015; Hinks et al., 2015), and such indices are unlikely to accurately capture the environmental conditions at the most biologically relevant scale (Mauck et al., 2018). Individuals in a population are rarely roaming the entire population space, particularly not territorial species. Thus, focusing on the scale at which the environment is interacting with the individual is key to understand the causes and consequences of ecological and evolutionary processes on individual fitness, and is an area where we commonly are hampered by a lack of small-scale environmental measurements (Nilsson et al., 2011b; Cole et al., 2015).
Understanding how environmental variation affect individual fitness requires accurate estimates of fitness-related traits in the studied population. For instance, in birds, recording the number of eggs in a clutch (clutch size) when the eggs have already hatched seems straightforward enough; there simply must have been at least as many eggs as nestlings (brood size). Thus, brood size obviously depends on clutch size. However, assuming that the brood size accurately represents the initial reproductive effort of a female may often be an underestimated source of uncertainty (eggs or nestlings may be lost before the nests are visited, eggs could be unfertilized or not hatched by other reasons). Also recording the number of nestlings when the brood rapidly is approaching fledging (leaving the nest) is also associated with uncertainty. For instance, there could be one or more offspring already having left the nest, or in worst case, the field observer might not encounter the nest until after fledging where there might be obvious signs of the brood having successfully fledged but the number of fledglings is unknown. Accounting for uncertainty when estimating fitness-related traits is fundamental in science (Fischhoff and Davis, 2014). If uncertainty is not accounted for, this can lead to serious errors during model selection in regression analyses and thus lead to misleading hypothesis testing. In this study, we acknowledge the fact that clutch and brood size are not independent variables, and we furthermore address the uncertainty in the estimates of both clutch and brood size when modeling breeding success in a study system where uncertainty has been recorded since the start of the study in 1978 (Nilsson et al., 2019).
Here, we focus on the effects of local environmental conditions at the territory scale (i.e., multiple hydrological variables at each territory, and model-input derived from a 1 × 1 km meteorological grid; Skaugen et al., 2015) on reproductive success in the white-throated dipper Cinclus cinclus (hereafter dipper), taking advantage of a long-term monitoring program (1978–2015; Nilsson et al., 2011a). The dipper is a passerine bird with an obligate aquatic lifestyle. Previous work has shown that the species population dynamics responds to variation in winter climate and spring phenology, responding to warmer springs but also to variation in territory quality (Nilsson et al., 2011a, 2019; Gamelon et al., 2017, 2018). However, the documented effects of regional temperatures and precipitation, ice conditions on the major lake in the study area, and NAO, are most likely extrapolations of microclimatic conditions, such as local river discharge and temperature, snow conditions within the territories, which we previously have had no information on. Based on the model, and the model input variables derived from a small-scale meteorological grid, we could use a number of accurate estimates of local environmental variables and relate these to reproductive success. Given the effect of future climate predictions on population dynamics (Gamelon et al., 2017), attributing reproductive success to variation in specific local variables is of particular interest. The lower parts of the river system have been limed to mitigate the effects of acidic precipitation, because such acidification has negative impacts on dipper reproductive performance (Ormerod et al., 1991; Nybø et al., 1997; Nilsson et al., 2018). We therefore also included information on liming status among the explanatory variables. In the present study, while assessing the effect of local environmental conditions on reproductive output, we have also taken male and female age, polygyny and population density into account. In addition to shedding light on the variables that are most important for breeding success, this exercise led us to draw some general conclusions about statistical modeling of biological systems, which deserves more attention.
Materials and Methods
Study Species and Study System
The dipper is a medium-sized passerine bird (50–70 g) distributed across mountainous areas in the Palearctic. It feeds on submerged aquatic invertebrates and fish in running clear freshwater (Ormerod et al., 1987; Tyler and Ormerod, 1994). Breeding is restricted to the immediate vicinity of fast-flowing rapids, where the nest is situated so that the opening is almost always placed immediately above fast-flowing water. Dippers lay approximately 5 eggs and start incubation after the clutch is completed. The female incubates for approximately 17 days and the young remain in the nest for about 22 days (Borgström, 1991). The Scandinavian dipper population is partially migratory, meaning that part of the population undertakes migration while the remaining birds winter on or close to the breeding grounds (Terrill and Able, 1988). Norwegian migratory dippers are recovered mainly in Denmark, Sweden, Northern Germany, and Poland (Bakken et al., 2006).
The study population is located in the river Lyngdalselva in southernmost Norway (Figure 1; 58°08′–58°40′N, 6°56′–7°20′E). The dipper population has been studied since 1978 following a standardized monitoring program (see Nilsson et al., 2011a for details). There is a strong altitudinal gradient in the study river system, from the outlet into the sea up to 700 m above sea level, in addition to a coastal-inland gradient reaching 60 km inland (Figure 1). The population size fluctuates between years with a range of 20–117 breeding pairs, typically declining after cold winters and increasing after mild ones (Nilsson et al., 2011a). The total number of recorded breeding events are 2585 by 2165 breeding females.
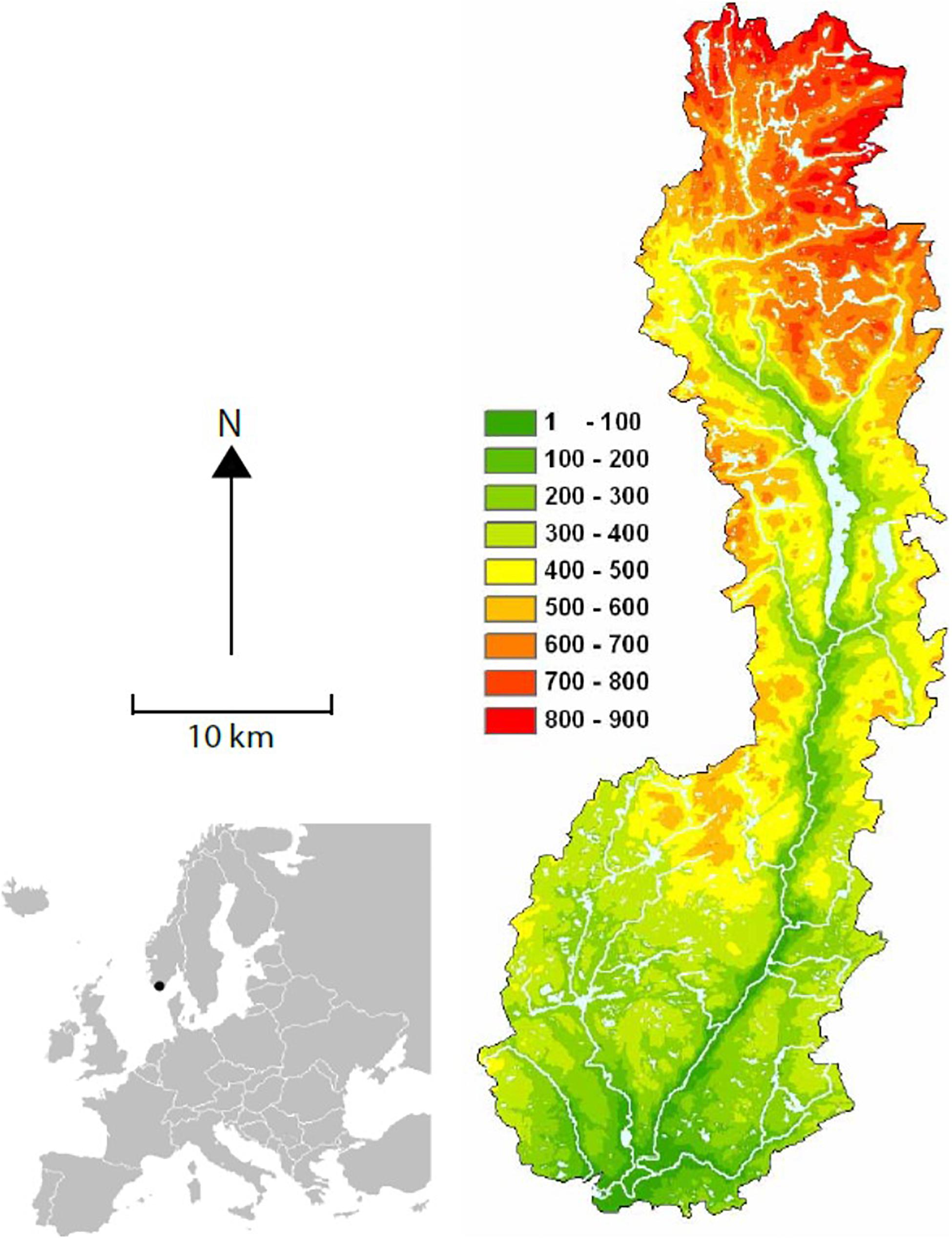
Figure 1. Map showing the altitudinal gradient (m) in the Lygna watershed.
Dippers defend territories, which contain one or more appropriate nest sites. Because the dipper has such particular nest site requirements (see above), the number of territories in our study system is limited to 158. Some territories are almost always occupied, while others have only been in use for a single year. The downstream and upstream boundaries of a territory might vary slightly between years, particularly depending on whether the neighboring territories are occupied or not, but the major part of a territory remains constant between years (Nilsson et al., 2019).
All known territories in the river system were visited in the early morning hours during the nest building phase and scanned for dipper activity. Breeding was defined as positive when inner nest building was completed. Dippers build an outer and an inner nest, where the outer nest can be used year after year. The inner nest is rebuilt each year and both sexes participate. At first encounter, adults were caught and ringed with an aluminum ring and given an individual color code in the form of two plastic color rings. Adult birds were accurately aged and sexed according to Svensson (1992). The clutch and brood sizes of almost all occupied nests are thus known and nearly all young were ringed with an aluminum ring. Clutch size was recorded during incubation, and brood size was recorded when the young were ringed in the nest. Uncertainty in the exact clutch and brood size was recorded when appropriate. Uncertainty can be caused by the disappearance of eggs, re-laid clutches, variation in chick survival and food provisioning, or when the nest is inaccessible and requires aid from mountain climbers, etc.
The mating system in the dipper is primarily monogamous, but occasionally polygynous. The prevalence of polygyny varies between populations (Wilson, 1996). Polygynous males occupy two or more territories with one female in each territory. In our area, assessing the mating system is complicated by the fact that occasionally females might sequentially make several breeding attempts with different males, often when the first breeding attempt fail. In some instances, females have even been observed to build nests at two different territories with two different males. However, the female only lays eggs in one of the built nests. We termed this mating strategy sequential monogamy. Polyandry is defined as multiple mating where the female’s clutch is fertilized by more than one male (Parker and Birkhead, 2013), but we have no observations of females actually mating with both males. The incidence of extra-pair young seems generally very low in the dipper, only 2% in another river in Norway (Øigarden and Linløkken, 2010).
Hydrology and Small-Scale Data
Hydrology data are usually obtained from gauged sites, but in Lyngdalselva only two sites are gauged. Therefore, hydrology data in this study is based on predictions in ungauged basins, at each dipper breeding territory (145 of 158 territories; the remaining territories had too small catchments to allow predictions) using the rainfall-runoff model (the Distance Distribution Dynamics (DDD) model; Skaugen et al., 2015). Digitized maps of terrain and river networks form the basis for estimations of many of the DDD model parameters. For the study period 1978–2015, daily river discharge data have been estimated, in addition to snow covered area (SCA), snow water equivalent (SWE) and groundwater levels. The model for Lyngdalselva is extremely good when comparing model predictions and gauged sites; the Kling Gupta Efficiency criterion (KGE; Gupta et al., 2009; Kling et al., 2012) for both gauged sites in Lyngdalselva is 0.94. Thiemig et al. (2013) regard values KGE 0.5–0.75 as intermediate, and 0.75–0.9 as good; a KGE of 0.94 for Lyngdalselva is thus very good.
River discharge can be of vastly different magnitudes in different territories. Mean discharge in territories in mountainous brooks can be as small as 0.008 m3/s, while river discharge at the last territory before the outlet into the sea is 32 m3/s. To allow comparison between territories with large differences in discharge, we standardized river discharge with two methods: (1) as the relative river discharge (defined as discharge divided by the territory mean discharge for the study period), and (2) as specific discharge, that is discharge per area, measured in l/s/km2, in each breeding territory.
To determine during which time periods different environmental variables would affect the dippers’ reproductive success, we defined an annual trigger date for each territory (Supplementary Material 1). Whether the river is ice-covered or not, is of vital importance for the dipper and initiation of breeding, because dippers find all of their food under water. We therefore defined the annual trigger date as the first date when the daily air temperature exceeded 0°C for five consecutive days after 1st of February. Based on the trigger date we defined a number of different time periods, defined as trigger periods, with different starting dates and of different durations, in addition to a number of statistically defined periods. For a full description of trigger dates and trigger periods, see Supplementary Material 1.
Statistical Analyses
The data included 2585 breeding events, where 1177 (45.4%) had a definitive recorded clutch size (no uncertainty). Among the recorded brood sizes, 2474 (95.7%) were definite brood sizes (no uncertainty).
Breeding Success Model
If the exact number of eggs (clutch size) and chicks (brood size) were known, the probability of an egg hatching, i.e., the probability of a chick from an egg given the fixed effects could be modeling using standard General Linear Model (GLM) regression. However, for some of the observed breeding events, there was considerable uncertainty in the exact clutch and/or brood sizes and this uncertainty must be taken into account when analyzing the effects of the explanatory variables. We addressed this by using the likelihood expression for the mentioned GLM model but sum over the set of possible eggs and chicks. This likelihood expression mixes the regression coefficients for the number of eggs and chicks, thus the task of doing regression on eggs and chicks is no longer two separate tasks but must be made in one larger analysis. Typical binomial GLM models use logit(probability) = linear combinations of explanatory variables, and logit(probability) is equivalent to log(odds), where odds = probability/(1−probability); Thus the effect of a change in each explanatory variable can most readily be interpreted as a change in the odds, and we will thus discuss the results in this manner. For more details on the statistical modeling, such as how we handle random factors and per measurement variation, see the Supplementary Material 2.
Selection of Explanatory Variables
In addition to the hydrology and other deducted variables such as local temperature, precipitation, and catchment area size, from the work by Skaugen et al. (2015), we included altitude, distance from the coast, male and female age and identity, and polygyny status, as possible explanatory variables of breeding success in the dipper. A full list of the examined possible explanatory variables is available in Supplementary Table S2.1. The total number of possible explanatory variables examined were 109, among which 101 were continuous fixed effects, two were categorical fixed effects, and six were random effects.
The set of possible explanatory variables was explored in a step-wise-up manner, by starting with a model without any explanatory variables and for each iteration examining all models where each explanatory variable not already included is added to the explanation variables already included. Bayesian model likelihood (BML) was used to evaluate each model. The procedure stopped at the iteration where none of the new models examined was (statistically) significantly better than the best model from the previous iteration, see the Supplementary Material 2 for more information on how we explore the model space as well as for a description of the Markov Chain Monte Carlo (MCMC) algorithm used and the BML.
Sequential Monogamy
In the initial stages of modeling breeding success, sequential monogamy was a major contributor to explain variation in brood size. However, in sequential monogamy, the first breeding attempts usually failed (62.5%). After correcting for the natural sequence of events in sequential monogamy, sequential monogamy was excluded from among the variables contributing to explaining variation in breeding success. However, because of this instance, we decided to investigate this behavior further. Consequently, we first summarized the individual number of total breeding events and the number of sequentially monogamous breeding events, for each female. This enabled us to test for over-dispersion (binomial vs. beta-binomial model), in order to test whether there are females with a penchant for the behavior or whether it occurs randomly. After that, we examined sequential monogamy per breeding event as the response in a General Linear Model (GLM) model (binomial response), using a Bayesian Information Criterion (BIC)-based stepwise search. Lastly, we assessed whether the behavior could be an indicator of inherent, unmeasured female quality and how this would affect reproductive success. Thus, we created a new indicator variable for whether the female at any point in her history had had a sequentially monogamous breeding event. We added this new indicator variable to our resulting model for breeding success (egg and/or chick probability). If one of these extra models had a better Bayesian model probability, i.e., if sequential monogamous history could explain some of the reproductive success even when correcting for whether or not each particular breeding event was sequentially monogamous, then the history of that behavior as an indicator of inherent female quality would hold true.
Results
Breeding Success Model
The breeding success model resulted in the following effects: altitude (eggs and chicks), a temporal trend (eggs), territory id (chicks), population size (eggs), female age (eggs), temperature maximums from the trigger date and until 9 days after (eggs) and temperature minimums 10 days centered on the trigger date (chicks), polygyny (chicks), temperature maximums last autumn (chicks), the quadratic distance to the coast (eggs), when the discharge decreased below 10 l/s 35–50 days before the trigger date (chicks), the variation in snow depth (SWE) 20–5 days before the trigger date (chicks), the variation in the discharge from the trigger date and until 9 days after (eggs), etc. (for a full list, see Tables 1, 2), where the effects are ordered as they were found by the step-wise search (Tables 1,2; for interpretation of regression coefficients, see Eqs. (2) and (4) in the Supplementary Material 2).
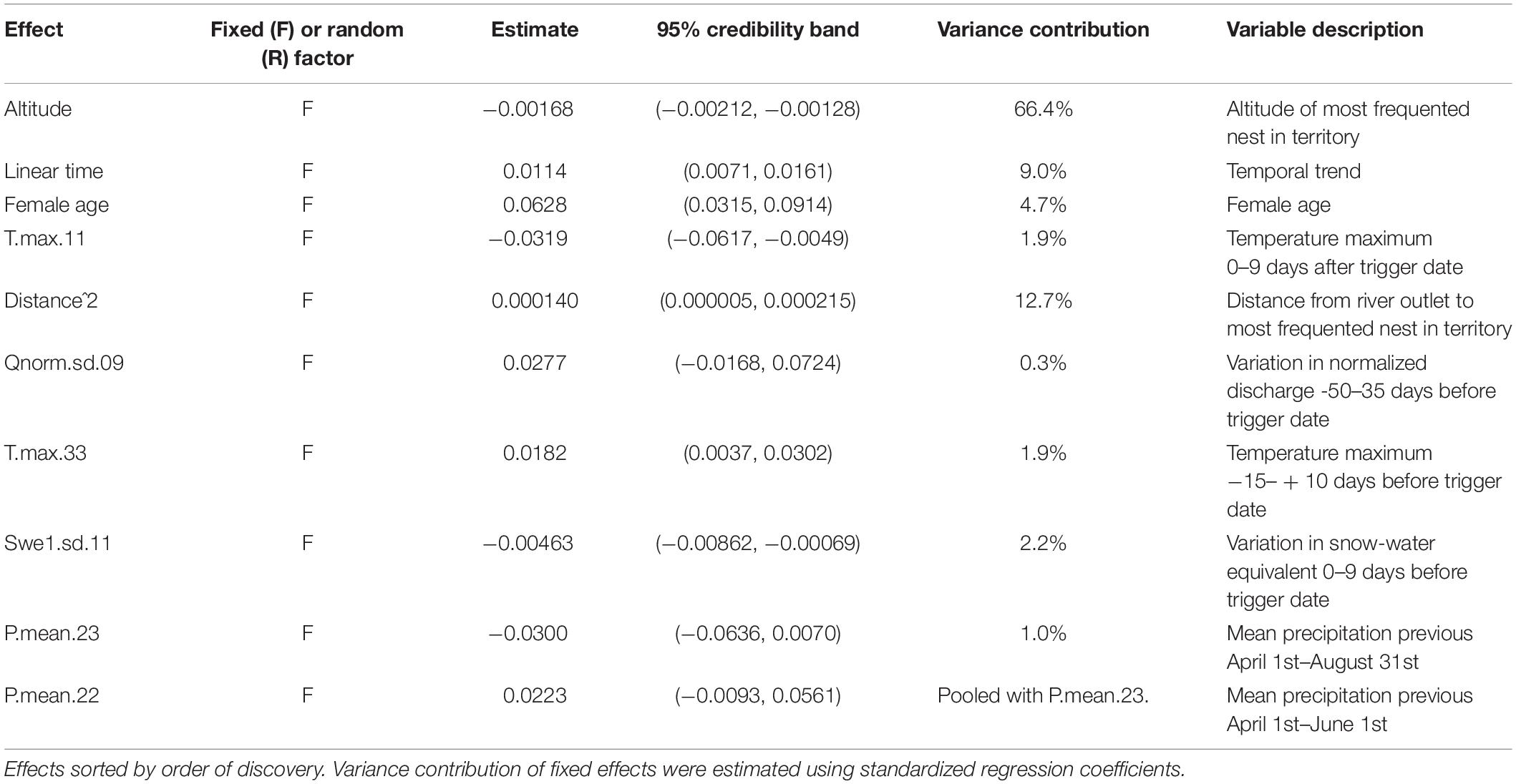
Table 1. Parameter estimates for a model explaining white-throated dipper clutch size in River Lyngdalselva 1978–2015 (see Eqs. (2) and (4) for interpretation, Supplementary Material 2).
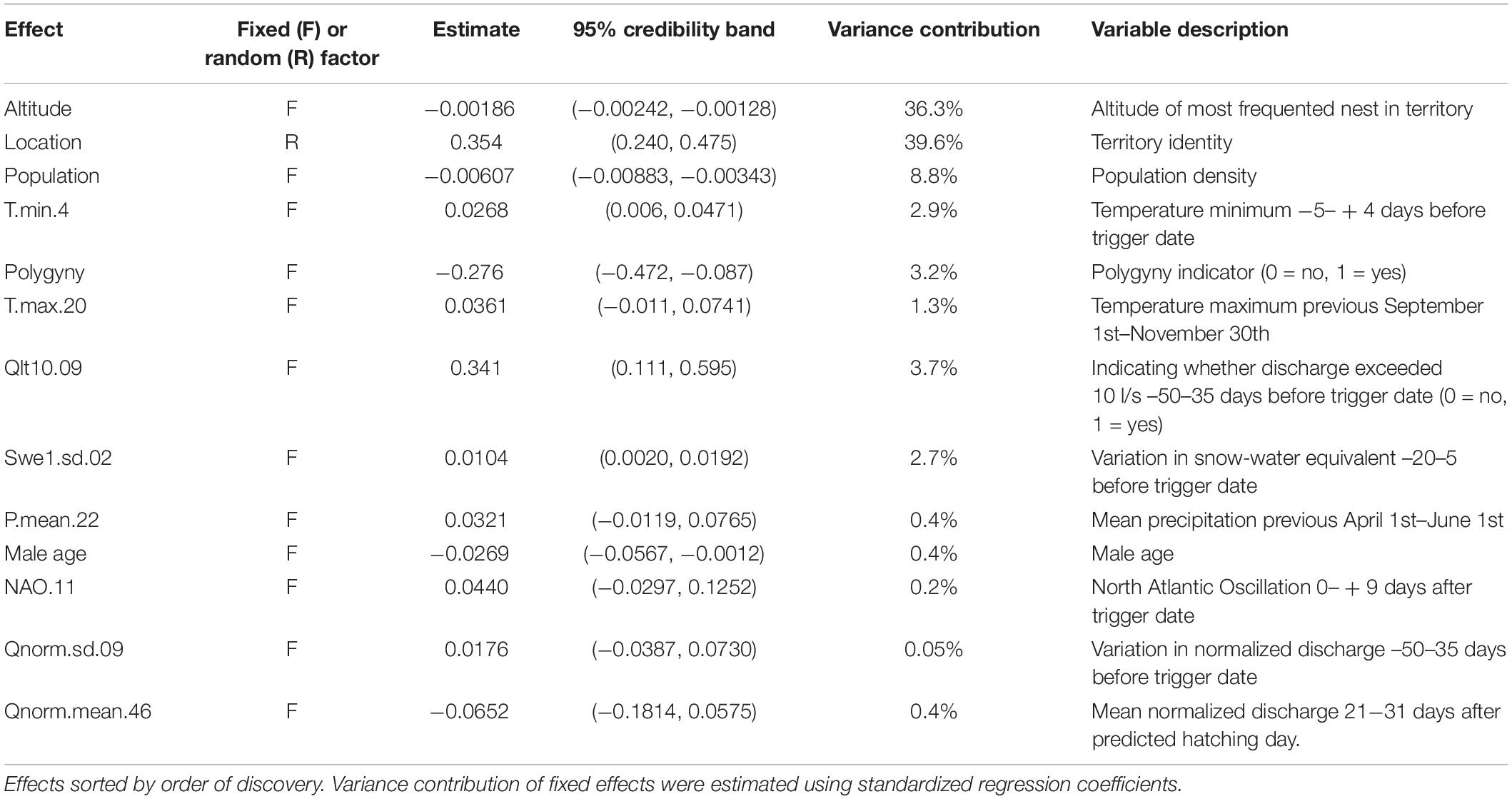
Table 2. Parameter estimates for a model explaining white-throated dipper brood size in River Lyngdalselva 1978–2015 (see Eqs. (2) and (4) for interpretation, Supplementary Material 2).
The strongest fixed effect was altitude for both the number of eggs and chicks (Tables 1, 2). The odds for producing an egg went down by a multiplicative factor of 0.84 for each 100 m higher up the territory was, that means that the odds were reduced by 16% for each extra 100 m of altitude. Similarly, the odds for producing a chick from each egg were reduced by a multiplicative factor of 0.83, i.e., a reduction of 17% per 100 m of extra altitude (Figure 2A). The odds for hatching success (effect on number of eggs times effect on number of chicks) was overall more than 12 times as high at sea level as it was at the territories at 700 m asl (Figure 2B). These were huge effects, highlighting the importance of the altitudinal gradient for breeding success.
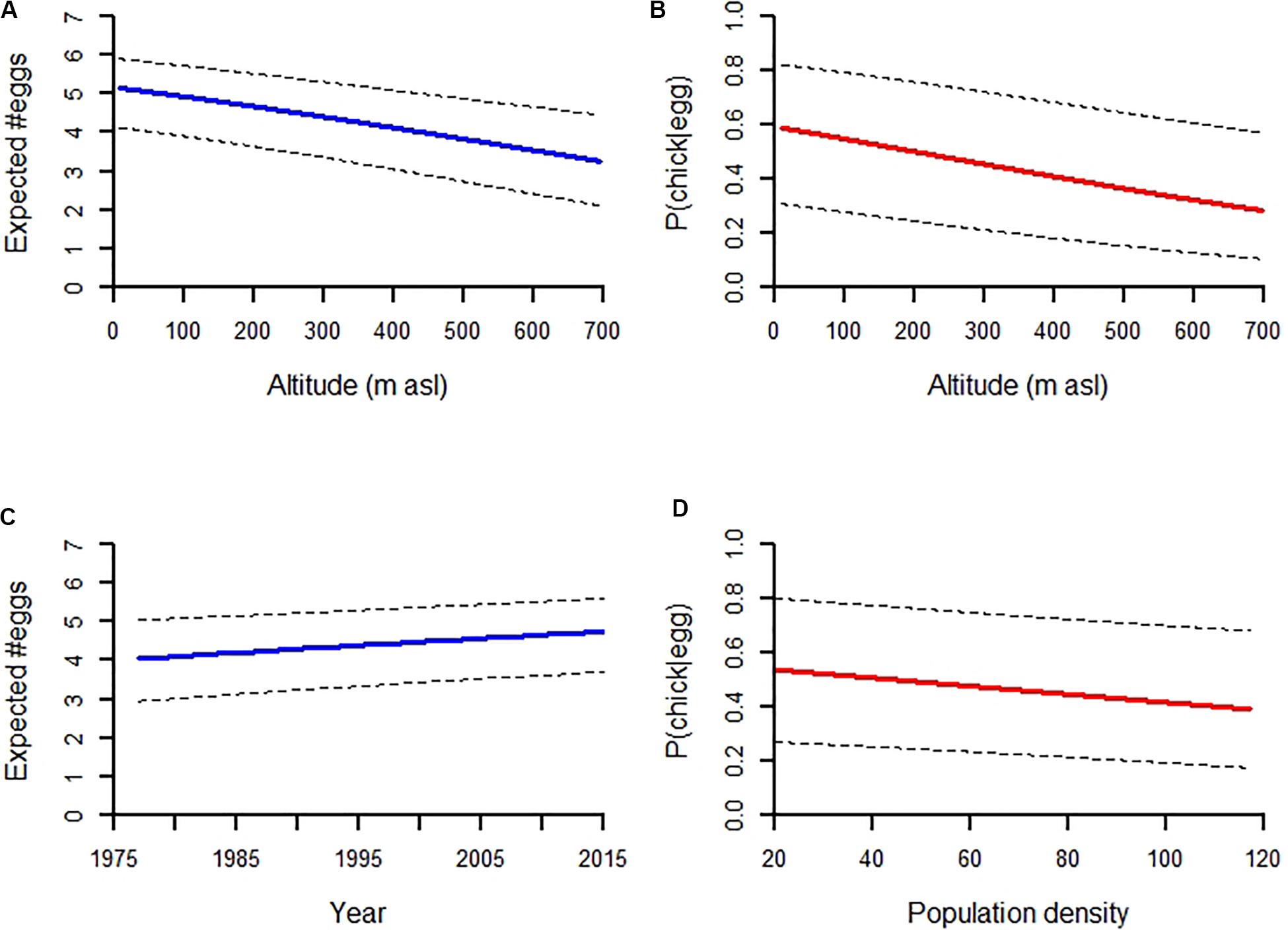
Figure 2. The causes of variation in breeding success in the white-throated dipper population in River Lyngdalselva 1978–2015. Effects are based on model predictions: Influence of altitude on the (A) clutch and (B) brood size, the (C) temporal increase in clutch size, and the (D) density effect on brood size. N = 2585.
The regression estimate for territory identity as a random factor for the number of chicks means that the odds can vary with a multiplicative factor ranging with 95% probability between 0.50 and 2.00. In essence, territory can halve or double the odds for chicks. This implies that some territories consistently produced larger clutches than expected, while other territories produced much smaller clutches. The odds for producing an egg increased by 12% per decade (Figure 2C), by 6.4% for each extra year the female ages, and by 7.9% when the distance to the coast went from 0 to the median value of 23.3 km (however, this was a quadratic effect). The odds for producing chicks decreased by 5.9% for an increase in population size of 10 new individuals (Figure 2D), decreased by 24% if the male was polygynous and increased 41% if the indicator for low discharge was set (which it is if the discharge becomes lower than 10 l/s in the period going from 50 to 35 days before the temperature-driven trigger date).
In summary, the most important model effects on the number of eggs laid were altitude (66.4%), distance from the coast (12.7%), the temporal trend (9.0%), female age (4.7%), temperature (3.8%) and snow (2.2%), while the most important model effects on number of chicks were altitude (36.3%) and territory identity (39.6%), followed by population size (8.8%), temperature (4.2%), discharge (4.2%), polygyny (3.2%) and snow (2.7%) (Tables 1, 2).
Sequential Monogamy
Before correcting for causality in the breeding success analysis, sequential monogamy was a major contributor to the model. Note, however, that our indicator for sequential monogamy indicated whether the female at any point in the season changed partner, and would thus be set before the change of partner actually took place. Females that later changed partner (during the same breeding season) usually failed at their first breeding attempt. There would therefore be a clear circularity in the analyses, overemphasizing the first failed breeding attempt of sequentially monogamous females, which could then trigger new breeding attempts and thus make partner change possible. This serves as an example demonstrating how potential errors in the analyses might arise due to faulty causality.
Yet, the rarity of sequential monogamy among females warrants further investigation. The first statistically strong but wrong result originated from curiously few field observations, where 40 breeding events (1.8%) by 38 females (3.2%) had a seemingly overwhelming effect when compared to the other 2125. We found no indication of over-dispersion in the number of sequentially monogamous breeding events for all females. Thus, we found no support for the notion that some females had an enhanced probability of sequential monogamy. We found mean temperature for the previous summer and autumn to be reliable explanatory variables for sequential monogamy. This climatic driver suggested that if the previous year was cold, there would be an increased probability of sequential monogamy the following breeding season.
Over-Dispersion
Over-dispersion suggested extra sources of variation not caught in the explanatory variables, i.e., that there were extra effects and phenomena not accounted for in the model. It was estimated to be negligible for clutch size. However, the over-dispersion for brood size given clutch size was estimated to be substantial (κ = 0.59, see Supplementary Material 2). When examining a model where the distributional modeling of over-dispersion was replaced with a per-measurement random factor, this factor stood for 88% of the overall variance contribution for number of chicks given the number of eggs. However, a look at the distribution of number of chicks for different number of eggs (see Supplementary Figure S2.1 in the Supplementary Material 2), suggested that zero-inflation may have been the reason for the over-dispersion, further suggesting that failure at the batch-level as well as individual level may have reduced the hatching success.
Discussion
Overall, territory location had a strong effect on the breeding success of the dippers, although local climatic variation also contributed. Birds breeding at higher altitudes further from the coast laid fewer eggs and fledged fewer young than dippers at lower altitudes closer to the coast. The altitudinal and coastal-inland gradients were most likely reflecting microclimatic variation; the microclimate would thus become progressively cooler while moving up the altitudinal gradient, by −0.65° per 100 m increase in altitude, and moving away from the coast to breeding locations further inland. The main river spread into smaller tributaries upstream, and reduced breeding success at higher altitudes could be a result of a generally lower food availability in smaller tributaries and perhaps reduced environmental buffering against flooding and drought events (Chiu et al., 2008). Reduced breeding success at higher altitudes could potentially also be a consequence of delayed breeding, because later breeding phenology is associated with lower fitness (Gamelon et al., 2018; Nilsson et al., 2019), although dippers are generally thought to breed multiple times during one breeding season (Fransson and Hall-Karlsson, 2008).
Earlier breeding might be the most commonly reported response to climate warming (Parmesan and Yohe, 2003; Dunn and Winkler, 2010), the dipper being no exception (Gamelon et al., 2018; Nilsson et al., 2019). In general, birds breeding earlier also produce larger clutches and more fledglings, although breeding time was not a significant contributor in this study. Accompanying earlier breeding in the dipper, the number of eggs in the clutch has increased during the study period. The increased clutch size might be a consequence of climate warming resulting in territories also at higher altitudes becoming available earlier, and increasing food availability prior to the breeding season in early spring. This would improve female body condition and enable females to lay more eggs. Thus, here we report a case of climate warming resulting in increased breeding effort, in addition to previous reports of earlier breeding (Parmesan and Yohe, 2003; Dunn and Winkler, 2010; Gamelon et al., 2018; Nilsson et al., 2019). Despite more eggs laid, there has not been a subsequent increase in the number of chicks fledged in the river system. The probability of fledging has not increased, probably because climate change has not only led to increased mean temperatures but also greater variability in the weather conditions (Easterling et al., 2000). Thus, greater climatic variation might reduce the dippers ability to forage efficiently enough to raise all chicks from hatching to fledging, due to droughts or sudden flooding events. An additional complicating factor is that the dippers in Lyngdalselva are partial migrants with the additional time constraint of arriving back on the breeding grounds ahead of breeding; the responses of partial migrants to climate change have proven difficult to predict (Berthold, 2001; Nilsson et al., 2006).
Location, or territory identity, had a substantial influence on the size of the brood, where some territories were consistently associated with a large number of nestlings, while others generally produced fewer nestlings. Specifically, location is a variable quantifying unknown and unmeasured but obviously important characteristics of the territory that might include habitat quality, local food abundance, nest site suitability, etc. Habitat quality obviously influences fitness (Newton, 1998), for example, in black kites Milvus migrans, high occupancy territories had higher food abundance and low predation risk, and produced most of the recruits (Sergio and Newton, 2003). Occupancy differed strongly between territories also in the present population (Nilsson et al., 2019).
We found that high population density had a negative effect on clutch size. Presumably, when population density was high, more of the low-quality territories were occupied, reducing the population average reproductive output. This is called density-dependent fecundity, the habitat heterogeneity hypothesis or the buffer effect, and explains how variation in breeding territory quality contributes to population dynamics (Brown, 1969; Dhondt et al., 1992; Mcpeek et al., 2001). Despite high population density favoring earlier breeding due to the competitive advantage of young fledging early (Nilsson et al., 2019), contradictorily high population density was associated with smaller clutches. Note that there were no density effects on brood size. From a population dynamical context, the contribution by strong fluctuations in winter mortality seemed to be greater than the relatively constant contribution by reproduction. In a long-term perspective, although successively warmer winters favor positive population growth, density-dependence can cause populations to become less buffered against occasional weather extremes (Gamelon et al., 2017).
Female and male age had an effect on clutch size, where the female effect was considerably stronger. Older females laid larger clutches than younger females. The female age effect on clutch size was not quadratic, contrary to earlier findings demonstrating a clear peak for female age in breeding phenology (Nilsson et al., 2019). Usually, age-specific reproductive output improves with age until leveling off (Forslund and Pärt, 1995). Presumably, reproduction improves with age and experience, while individuals with poor breeding performance might also suffer lower survival, which might contribute to explain the observed pattern (selective disappearance; Reid et al., 2003; McCleery et al., 2008). Thus, we did not find any evidence of senescence in female reproductive parameters (Monaghan et al., 2008). This might lend some support to the terminal investment hypothesis (Clutton-Brock, 1984; Duffield et al., 2017), which states that when an individual’s chance of future reproduction grows smaller, the investment in current reproduction should increase. Although the effect of male age on clutch size was marginal, older males were associated with smaller clutches than younger males. This is in contrast to results from other study systems, where male age had no effect on clutch size (Sanz, 2002; Reid et al., 2003) or a positive effect (Mills, 1973; Hasselquist, 1998). However, Kindsvater and Alonzo (2014) found that when mortality is low, young females might reduce current clutch size and prioritize the next breeding opportunity. Male polygyny status had a negative effect on brood size where secondary females produced fewer young, possibly because polygynous males did not provide as much parental care as monogamous males (Slagsvold and Lifjeld, 1994).
Surprisingly, the climatic influence was of lesser consequence compared to other effects in explaining variation in clutch and brood sizes. Considering the importance of winter weather for population dynamics in this species (Nilsson et al., 2011a; Gamelon et al., 2017), the minor influence of weather in the following months is remarkable. Temperature regimes around the trigger date, meaning when the minimum temperatures had exceeded 0°C for five consecutive days, from 5 days before and until 9 days after, were most prominent. High maximum temperatures had a diminishing effect on clutch size while high minimum temperatures increased brood size. High minimum temperatures most likely improves feeding success and chick survival, because then there will most likely be less risk of temporary setbacks with freezing over of the river rendering prey inaccessible. In birds, spring temperature is a key determinant of timing of breeding (Parmesan and Yohe, 2003; Charmantier et al., 2008; Nilsson et al., 2019), and the negative effect of high maximum temperatures on clutch size might indicate that birds have failed to time-optimize laying date, leading to a reduced clutch. Interestingly, high maximum temperatures the previous autumn enabled dippers to raise more chicks. Presumably, this is an effect of improved adult body condition in early winter, enabling dippers to invest more energy into parental care, despite similar clutch sizes. Higher winter river discharge in a 2-week period 1 month before the trigger date had a positive effect on both clutch and brood size. Given that one of the discharge variables were a threshold variable for when discharge dropped below 10 l/s at each nest site, we conclude that higher river discharge in late winter naturally is positive for dipper reproductive success. Water birds are well known to be affected by floods and droughts (Royan et al., 2013), and dipper prey might become more inaccessible during flooding events (Chiu et al., 2008, 2013), because dippers are visual predators and floods lead to high water turbidity. Droughts during breeding might reduce the available river area used for foraging, but more importantly, it may render the nest visible to predators because dipper nest sites are selected for their ability to flush away chick excrements (Tyler and Ormerod, 1994). Because dippers depend entirely on submerged prey and rapidly running water to hide the nest, the absence of discharge variables of high importance for breeding success is rather unexpected. Presumably, the dipper has adapted to the natural variation in river discharge by choosing nest sites where droughts rarely occur and foraging at alternative sites with shallow water, such as swells and lakes, during floods.
Important Reflections From Analyzing Sequential Monogamy
Investigating sequential monogamy, the only variable able to explain some of the variation was the mean temperature during the previous summer and autumn. The probability of sequential monogamy increased with decreasing temperatures, indicating that sequential monogamy might be related to poor body condition. Body condition during molt and establishment of wintering territories in late summer and autumn, prior to the critical winter months, might carry over to the breeding season (Harrison et al., 2011), where females in poor condition make the best of a bad situation. Neither did we find support for the notion that some females have an enhanced probability of sequential monogamous breeding events; two females showed this behavior twice, which is not implying that these females had a penchant for sequential monogamy. Worth noting, the observed number of sequentially monogamous breeding attempts is a conservative measure, because to detect sequentially monogamous breeding events a large field effort is required during the early breeding season, and some events might have passed under the radar. Incidentally, extra-pair paternity seems to be low in the dipper, approximately 2% (Øigarden and Linløkken, 2010), but it would be interesting to know whether the frequency is higher in sequentially monogamous females. For a discussion about circularity in statistical analyses of biological systems, see the Supplementary Material 2.
Conclusion
Low occupancy, and presumably low quality, territories in years of high population density lead to reduced fledging success, also called density-dependent fecundity (Brown, 1969; Dhondt et al., 1992; Mcpeek et al., 2001). Breeding success in the dipper was heavily dependent on the territory the breeding pair defended; there is therefore fierce competition for the best territories in the river system. The differential rates of occupancy between the territories in the population has thus received its natural explanation (Nilsson et al., 2019). In addition to the territory effect, females laid larger clutches at the end of the study period than at the start of it. Considering the climate warming in the study area, female body condition might have improved due to milder pre-breeding weather, and territories at higher altitudes might have become available earlier than at the start of the study. Increased clutch size is thus presumably explained by climatic factors, and it is for that reason surprising that climatic conditions explained comparatively little of the variation in fledging success. Hitherto, dipper breeding success might have been buffered against negative effects of climate change. Whether this can continue under further warming trends and increasing incidences of weather extremes is presently unknown, but long-term studies remain important in predicting future climate responses (Gamelon et al., 2017).
Data Availability Statement
The code used in this analysis is available at https://github.com/trondreitan/egg_chick_intervals. The datasets analyzed during the current study are not publicly available, due to it being a private, unique long-term collection of individual data on a breeding dipper population, which has cost an enormous effort in the field 1973–present. However, the data are available from the corresponding author upon reasonable request. The complete dipper data are stored at the Norwegian University of Life Sciences, and managed by OWR (b2xlLnJvc3RhZEBubWJ1Lm5v).
Ethics Statement
The field work was conducted with respect for the animals’ well-being and adheres to the Guidelines for the Use of Animals in Research (Anonymous, 2006). It complies with the laws and regulations for animals used in research in Norway (FOR-2003-03-18-349, LOV-2009-06-19-100, LOV-1981-05-29-38§ 26). Ethical approval is not required for bird ringing (FOR-2003-03-18-349§§ 21-24), but Ringing licenses were issued and approved by the Norwegian Bird Ringing Centre on behalf of the Norwegian Environment Agency.
Author Contributions
ALKN and JHLL conceived the study. KJ collected the field data. TSk ran the hydrological simulations and also provided DDD model input for the analyses. OWR organized the dipper data collection. TR conducted the analyses. ALKN and TR wrote the paper with input from MG, TSl, LAV, TSk, OWR, BW, KJ, and NCS. All authors contributed to the article and approved of the submitted version.
Funding
This study was supported by the Norwegian Research Council (nr. 221393), the Research Council of Norway through its Centres of Excellence funding scheme (nr. 223257 to CBD), and the Norwegian Water Resources and Energy Directorate.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We are grateful to the many people involved in the field work during the last 40 years. Thanks also to Øystein Holen for helpful discussions about sequential monogamy. We are also grateful to two reviewers and Mark Mainwaring for constructive comments on previous versions of the paper.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2020.542846/full#supplementary-material
References
Anonymous (2006). Guidelines for the treatment of animals in behavioural research and teaching. Anim. Behav. 71, 245–253. doi: 10.1016/j.anbehav.2005.10.001
Bakken, V., Runde, O., and Tjørve, E. (2006). Norsk Ringmerkningsatlas. Stavanger: Stavanger Museum.
Borgström, E. (1991). Strömstarens Cinclus c. cinclus utbredning och häckningsbiologi i Värmland. Ornis Svec. 1, 93–101.
Brown, J. L. (1969). Buffer effect and productivity in tit populations. Am. Nat. 103, 347–354. doi: 10.1086/282607
Charmantier, A., McCleery, R. H., Cole, L. R., Perrins, C., Kruuk, L. E. B., and Sheldon, B. C. (2008). Adaptive phenotypic plasticity in response to climate change in a wild bird population. Science 320, 800–803. doi: 10.1126/science.1157174
Chiu, M.-C., Kuo, M.-H., Hong, S.-Y., and Sun, Y.-H. (2013). Impact of extreme flooding on the annual survival of a riparian predator, the brown dipper Cinclus pallasii. Ibis 155, 377–383. doi: 10.1111/ibi.12035
Chiu, M.-C., Kuo, M.-H., Sun, Y.-H., Hong, S.-Y., and Kuo, H.-C. (2008). Effects of flooding on avian top-predators and their invertebrate prey in a monsoonal Taiwan stream. Freshw. Biol. 53, 1335–1344. doi: 10.1111/j.1365-2427.2008.01968.x
Clutton-Brock, T. H. (1984). Reproductive effort and terminal investment in iteroparous animals. Am. Nat. 123, 212–229. doi: 10.1086/284198
Cole, E. F., Long, P. R., Zelazowski, P., Szulkin, M., and Sheldon, B. C. (2015). Predicting bird phenology from space: satellite-derived vegetation green-up signal uncovers spatial variation in phenological synchrony between birds and their environment. Ecol. Evol. 5, 5057–5074. doi: 10.1002/ece3.1745
Dhondt, A. A., Kempenaers, B., and Adriaensen, F. (1992). Density-dependent clutch size caused by habitat heterogeneity. J. Anim. Ecol. 61, 643–648. doi: 10.2307/5619
Duffield, K. R., Bowers, E. K., Sakaluk, S. K., and Sadd, B. M. (2017). A dynamic threshold model for terminal investment. Behav. Ecol. Sociobiol. 71:185.
Dunn, P. O., and Winkler, D. W. (2010). “Effects of climate change on timing of breeding and reproductive success in birds,” in Effects of Climate Change on Birds, eds A. P. Møller, W. Fiedler, and P. Berthold (Oxford: Oxford University Press), 113–128.
Easterling, D. R., Meehl, G. A., Parmesan, C., Changnon, S. A., Karl, T. R., and Mearns, L. O. (2000). Climate extremes: observations, modeling, and impacts. Science 289, 2068–2074. doi: 10.1126/science.289.5487.2068
Fischhoff, B., and Davis, A. L. (2014). Communicating scientific uncertainty. Proc. Natl. Acad. Sci. U.S.A. 111, 13664–13671.
Forslund, P., and Pärt, T. (1995). Age and reproduction in birds - hypotheses and tests. Tree 10, 374–378. doi: 10.1016/s0169-5347(00)89141-7
Fransson, T., and Hall-Karlsson, S. (2008). Strömstare Cinclus cinclus, Swedish Bird Ringing Atlas. Stockholm: Swedish Museum of Natural History & Swedish Ornithological Society.
Frederiksen, M., Daunt, F., Harris, M. P., and Wanless, S. (2008). The demographic impact of extreme events: stochastic weather drives survival and population dynamics in a long-lived seabird. J. Anim. Ecol. 77, 1020–1029. doi: 10.1111/j.1365-2656.2008.01422.x
Gamelon, M., Grøtan, V., Nilsson, A. L. K., Engen, S., Hurrell, J. W., Jerstad, K., et al. (2017). Interactions between demography and environmental effects are important determinants of population dynamics. Sci. Adv. 3:e1602298. doi: 10.1126/sciadv.1602298
Gamelon, M., Tufto, J., Nilsson, A. L. K., Jerstad, K., Røstad, O. W., Stenseth, N. C., et al. (2018). Environmental drivers of varying selective optima in a small passerine: a multivariate, multiepisocid approach. Evolution 72, 2325–2342. doi: 10.1111/evo.13610
Gupta, H. V., Kling, H., Yilmaz, K. K., and Martinez, G. F. (2009). Decomposition of the mean squared error and NSE performance criteria: implications for improving hydrological modelling. J. Hydrol. 377, 80–91. doi: 10.1016/j.jhydrol.2009.08.003
Hanssen-Bauer, I. (2005). Regional Temperature and Precipitation Series for Norway: Analyses of Time-Series Updated to 2004. Climate Report no. 15. Norway: Norwegian Meteorological Institute.
Harrison, X. A., Blount, J. D., Inger, R., Norris, D. R., and Bearhop, S. (2011). Carry-over effects as drivers of fitness differences in animals. J. Anim. Ecol. 80, 4–18. doi: 10.1111/j.1365-2656.2010.01740.x
Hasselquist, D. (1998). Polygyny in great reed warblers: a long-term study of factors contributing to male fitness. Ecology 79, 2376–2390. doi: 10.1890/0012-9658(1998)079[2376:PIGRWA]2.0.CO;2
Hinks, A. E., Cole, E. F., Daniels, K. J., Nakagawa, S., and Sheldon, B. C. (2015). Scale-dependent phenological synchrony between songbirds and their caterpillar food source. Am. Nat. 186, 84–97. doi: 10.1086/681572
Kindsvater, H. K., and Alonzo, S. H. (2014). Females allocate differentially to offspring size and number in response to male effects on female and offspring fitness. Proc. Roy. Soc. B 281:20131981. doi: 10.1098/rspb.2013.1981
Kling, H., Fuchs, M., and Paulin, M. (2012). Runoff conditions in the upper Danube basin under an ensemble of climate change scenarios. J. Hydrol. 424–425, 264–277. doi: 10.1016/j.jhydrol.2012.01.011
Mauck, R. A., Dearborn, D. C., and Huntington, C. E. (2018). Annual global mean temperature explains reproductive success in a marine vertebrate from 1955 to 2010. Glob. Change Biol. 24, 1599–1613. doi: 10.1111/gcb.13982
McCleery, R. H., Perrins, C. M., Sheldon, B. C., and Charmantier, A. (2008). Age-specific reproduction in a long-lived species: the combined effects of senescence and individual quality. Proc. Roy. Soc. B 275, 963–970. doi: 10.1098/rspb.2007.1418
Mcpeek, M. A., Rodenhouse, N. L., Holmes, R. T., and Sherry, T. W. (2001). A general model of site-dependent population regulation: population-level regulation without individual-level interactions. Oikos 94, 417–424. doi: 10.1034/j.1600-0706.2001.940304.x
Mills, J. A. (1973). Influence of age and pair-bond on breeding biology of red-billed Gull Larus novaehollandiae scopulinus. J. Anim. Ecol. 42, 147–162. doi: 10.2307/3409
Monaghan, P., Charmantier, A., Nussey, D. H., and Ricklefs, R. E. (2008). The evolutionary ecology of senescence. Funct. Ecol. 22, 371–378. doi: 10.1111/j.1365-2435.2008.01418.x
Nilsson, A. L. K., Knudsen, E., Jerstad, K., Røstad, O. W., Walseng, B., Slagsvold, T., et al. (2011a). Climate effects on population fluctuations of the white-throated dipper Cinclus cinclus. J. Anim. Ecol. 80, 235–243. doi: 10.1111/j.1365-2656.2010.01755.x
Nilsson, A. L. K., Stige, L. C., Jerstad, K., Røstad, O. W., Slagsvold, T., Knudsen, E., et al. (2011b). To make the most of what we have: extracting phenological data from nestling measurements. Int. J. Biometeorol. 55, 797–804. doi: 10.1007/s00484-011-0461-2
Nilsson, A. L. K., L’Abee-Lund, J. H., Vøllestad, L. A., Jerstad, K., Larsen, B. M., Røstad, O. W., et al. (2018). The potential influence of Atlantic salmon Salmo salar and brown trout Salmo trutta on density and breeding of the white-throated dipper Cinclus cinclus. Ecol. Evol. 8, 4065–4073. doi: 10.1002/ece3.3958
Nilsson, A. L. K., Lindström, Å, Jonzen, N., Nilsson, S. G., and Karlsson, L. (2006). The effect of climate change on partial migration - the blue tit paradox. Glob. Change Biol. 12, 2014–2022. doi: 10.1111/j.1365-2486.2006.01237.x
Nilsson, A. L. K., Slagsvold, T., Røstad, O. W., Knudsen, E., Jerstad, K., Cadahia, L., et al. (2019). Territory location and quality, together with climate, affect the timing of breeding in the white-throated dipper. Sci. Rep. 9:7671. doi: 10.1038/s41598-019-43792-5
Nybø, S., Staurnes, M., and Jerstad, K. (1997). Thinner eggshells of dipper (Cinclus cinclus) eggs from an acidified area compared to an non-acidified area in Norway. Water Air Soil Pollut. 93, 255–266. doi: 10.1007/BF02404760
Øigarden, T., and Linløkken, A. (2010). Is the breeding success of the white-throated dipper Cinclus cinclus in Hedmark, Norway, influenced by acid rain. Ornis Norveg. 33, 118–129. doi: 10.15845/on.v33i0.151
Ormerod, S. J., Efteland, S., and Gabrielsen, L. E. (1987). The diet of breeding dippers Cinclus cinclus cinclus and their nestlings in southwestern Norway. Holarctic Ecol. 10, 201–205. doi: 10.1111/j.1600-0587.1987.tb00760.x
Ormerod, S. J., O’Halloran, J., Gribbin, S. D., and Tyler, S. J. (1991). The ecology of dippers Cinclus cinclus in relation to stream acidity in upland Wales: breeding performance, calcium physiology and nestling growth. J. Appl. Ecol. 28, 419–433. doi: 10.2307/2404559
Parker, G. A., and Birkhead, T. R. (2013). Polyandry: the history of a revolution. Philos. T. R. Soc. B 368:20120335. doi: 10.1098/rstb.2012.0335
Parmesan, C., and Yohe, G. (2003). A globally coherent fingerprint of climate change impacts across natural systems. Nature 421, 37–42. doi: 10.1038/nature01286
Reid, J. M., Bignal, E. M., Bignal, S., McCracken, D. I., and Monaghan, P. (2003). Age-specific reproductive performance in red-billed choughs Pyrrhocorax pyrrhocorax: patterns and processes in a natural population. J. Anim. Ecol. 72, 765–776. doi: 10.1046/j.1365-2656.2003.00750.x
Royan, A., Hannah, D. M., Reynolds, S. J., Noble, D. G., and Sadler, J. P. (2013). Avian community responses to variability in river hydrology. PLoS One 8:e83221. doi: 10.1371/journal.pone.0083221
Sanz, J. J. (2002). Climate change and breeding parameters of great and blue tits throughout the western Palaearctic. Glob. Change Biol. 8, 409–422. doi: 10.1046/j.1365-2486.2002.00496.x
Sergio, F., and Newton, I. (2003). Occupancy as a measure of territory quality. J. Anim. Ecol. 72, 857–865. doi: 10.1046/j.1365-2656.2003.00758.x
Skaugen, T., Peerebom, I. O., and Nilsson, A. (2015). Use of a parsimonious rainfall-run-off model for predicting hydrological response in ungauged basins. Hydrol. Process. 29, 1999–2013. doi: 10.1002/hyp.10315
Slagsvold, T., and Lifjeld, J. T. (1994). Polygyny in birds: the role of competition between females for male parental care. Am. Nat. 143, 59–94. doi: 10.1086/285596
Sæther, B. E., Tufto, J., Engen, S., Jerstad, K., Røstad, O. W., and Skåtan, J. E. (2000). Population dynamical consequences of climate change for a small temperate songbird. Science 287, 854–856. doi: 10.1126/science.287.5454.854
Terrill, S. B., and Able, K. P. (1988). Bird migration terminology. Auk 105, 205–206. doi: 10.1093/auk/105.1.205
Thiemig, V., Rojas, R., Zambrano-Bigiarini, M., and De Roo, A. (2013). Hydrological evaluation of satellite-based rainfall estimates over the Volta and Baro-Akobo basin. J. Hydrol. 499, 324–338. doi: 10.1016/j.jhydrol.2013.07.012
Keywords: brood size, Cinclus cinclus, clutch size, habitat quality, hydrology, polygamy, treating uncertainty
Citation: Nilsson ALK, Reitan T, Skaugen T, L’Abée-Lund JH, Gamelon M, Jerstad K, Røstad OW, Slagsvold T, Stenseth NC, Vøllestad LA and Walseng B (2020) Location Is Everything, but Climate Gets a Share: Analyzing Small-Scale Environmental Influences on Breeding Success in the White-Throated Dipper. Front. Ecol. Evol. 8:542846. doi: 10.3389/fevo.2020.542846
Received: 14 March 2020; Accepted: 25 August 2020;
Published: 14 October 2020.
Edited by:
Mark C. Mainwaring, University of Montana, United StatesReviewed by:
Stanley Mastrantonis, The University of Western Australia, AustraliaJonathan Bennie, University of Exeter, United Kingdom
Copyright © 2020 Nilsson, Reitan, Skaugen, L’Abée-Lund, Gamelon, Jerstad, Røstad, Slagsvold, Stenseth, Vøllestad and Walseng. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Anna L. K. Nilsson, YW5uYS5uaWxzc29uQG5pbmEubm8=