
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 04 September 2020
Sec. Biogeography and Macroecology
Volume 8 - 2020 | https://doi.org/10.3389/fevo.2020.00280
This article is part of the Research Topic Invaders on the Horizon! Scanning the Future of Invasion Science and Management View all 16 articles
 Bernd Lenzner1*†
Bernd Lenzner1*† Guillaume Latombe1
Guillaume Latombe1 César Capinha2
César Capinha2 Céline Bellard3†Franck Courchamp3†
Céline Bellard3†Franck Courchamp3† Christophe Diagne3†Stefan Dullinger1
Christophe Diagne3†Stefan Dullinger1 Marina Golivets4†Severin D. H. Irl5†
Marina Golivets4†Severin D. H. Irl5† Ingolf Kühn4,6,7†Brian Leung8†
Ingolf Kühn4,6,7†Brian Leung8† Chunlong Liu9†Dietmar Moser1Núria Roura-Pascual9†
Chunlong Liu9†Dietmar Moser1Núria Roura-Pascual9† Hanno Seebens10†
Hanno Seebens10† Anna Turbelin3Patrick Weigelt11Franz Essl1
Anna Turbelin3Patrick Weigelt11Franz Essl1Biological invasions are a major threat to global biodiversity with particularly strong implications for island biodiversity. Much research has been dedicated towards understanding historic and current changes in alien species distribution and impacts on islands and potential changes under future climate change. However, projections of how alien species richness and impacts on islands might develop in the future are still lacking. In the absence of reliable projections, expert-based assessments are a valuable tool to investigate the importance of different drivers and pathways and the distributions of potential impacts of future biological invasions. These insights can guide subsequent quantification efforts and inform invasive species management and policy. In this study, we performed a survey among 126 experts in invasion science ranging from scientists to managers and decision makers with a focus on island systems until the mid-21st century. The survey revealed that out of 15 drivers, six were considered important by almost all respondents (>90%). Of these, trade and transport was identified as most important at the introduction stage (99.2%) and land use/cover change as most important at the establishment (96.8%) and spread (95.2%) stage. Additionally, the experts considered that alien species were more likely to be introduced (93.7%) and spread (78.6%) as stowaways than through any other pathway. In general, respondents agreed that the impacts of alien species will increase on all types of islands, particularly on oceanic islands, followed by atolls and continental islands. Within islands, terrestrial ecosystems were assumed to be impacted more severely than marine ecosystems. Finally, the survey hints toward the potential for effective communication, scientific research and increased pro-active management of alien species on islands to reduce their future consequences. Given the major threat represented by invasive alien species on islands, these results provide crucial insights relevant for global and regional conservation efforts.
Biological invasions have been identified as one of the major threats to biodiversity worldwide and are an important facet of global environmental change (Maxwell et al., 2016; IPBES, 2018). Islands are hotspots of both endemic (Myers et al., 2000; Kier et al., 2009) and alien species richness (Bellard et al., 2017; Dawson et al., 2017; Essl et al., 2019a) with a subset of alien species – invasive alien species (IAS) – causing negative impacts on biodiversity and driving many recent extinctions (Tershy et al., 2015; Bellard et al., 2016). Current knowledge of the mechanisms driving biological invasions on islands, and of the threats IAS pose on island biota, largely relies on information from several well studied regions (e.g., Macaronesian Islands, Hawaii; e.g., Levine and D’Antonio, 1999; Daehler, 2006; Kueffer et al., 2014), while research in many other island regions is often less extensive or even lacking.
Two issues are crucial in understanding island invasions for conservation and scientific purposes: (i) a comparison of trends in alien species richness in mainland and island regions (Seebens et al., 2018); (ii) how environmental and socio-economic factors could change in relative importance over time for driving future invasions. Improved understanding of these issues should lead ultimately to better knowledge on how invasion impacts could change over time in magnitude and geographical distribution (Lenzner et al., 2019). Answers to these questions are complex, as drivers of biological invasions may change distinctly across taxa, habitats and island regions (Latombe et al., 2019b).
Assessments of potential future invasions and impacts of alien species require the use of scenarios of future trajectories for various facets of specific systems, such as invasion pathways or other drivers of alien species richness, composition, abundance and impact. For most other key drivers of biodiversity change such scenarios have already been established. These include climate change (Moss et al., 2010; IPCC, 2014), land-use change (Hurtt et al., 2009), sea-level rise (Wetzel et al., 2012; Hinkel et al., 2014) and human population change (Lutz et al., 2014). However, comprehensive long-term scenarios are still missing for biological invasions (Lenzner et al., 2019). One reason for the lack of scenarios for potential future states of biological invasions is their complex nature and until recently a lack of comprehensive global datasets (Courchamp et al., 2017). Moreover, biological invasions have arisen from a complex interplay of environmental, socio-economic and societal changes that are difficult to project using classical modeling techniques, like static habitat suitability models, population dynamic models or cellular automata (see Buchadas et al., 2017; Capinha et al., 2018; Lenzner et al., 2019). To overcome such multi-disciplinary challenges, combining classical forecasting techniques with expert-based assessments has proven to be a promising approach (e.g., through qualitative surveys; Berg et al., 2016; Symstad et al., 2017; Reside et al., 2018).
Here, we present the outcome of an expert survey with the aim to identify the importance of different drivers and pathways of biological invasions on islands in the 21st century. Further, based on the knowledge of experts on island ecology and conservation, we aim to gain a better understanding of potential future impacts of biological invasions on islands. The identification of these aspects of biological invasions in the future is crucial for subsequent quantification efforts. Specifically, we focus on three overarching questions: (1) Which drivers will contribute most strongly to alien species richness increase during the introduction, establishment and spread phases of the invasion process? (2) Which pathways will substantially increase alien species richness during the introduction and spread phases of the invasion process? (3) How strongly will different island contexts be affected by an increase in alien species richness?
A set of 15 major drivers of biological invasions was selected during a workshop on scenarios for biological invasions held in Vienna, Austria in October 2016. A comprehensive list of drivers of biological invasions based on an extensive literature search was compiled prior to the workshop. This list was then provided to a group of invasion scientists with complementary backgrounds in related fields (e.g., land-use change, systems analysis, global environmental change), who assessed the importance of each driver and identified a set of 15 most relevant ones for future biological invasions. For the selection procedure, each workshop participant identified three most important drivers for future invasions and the highest ranked drivers were selected. This set of drivers underlies complementary assessments of biological invasion scenarios that are currently under development (Essl et al., 2019b, 2020; Roura-Pascual et al., under review). We adopted this updated list of 15 drivers and classified them into three thematic groups: (i) environmental change drivers (climate change; ocean acidification; eutrophication and pollution; biodiversity loss and degradation), (ii) socio-economic activity drivers (trade and transport; land use/cover change; socio-economic change; demography and migration) and (iii) society and technology (awareness, values and lifestyle; recreation and tourism; communication and outreach; technology and innovation; cooperation, legislation and agreements; IAS science; IAS management). For the description of individual drivers and their rationale as provided in the questionnaire, see Table 1.
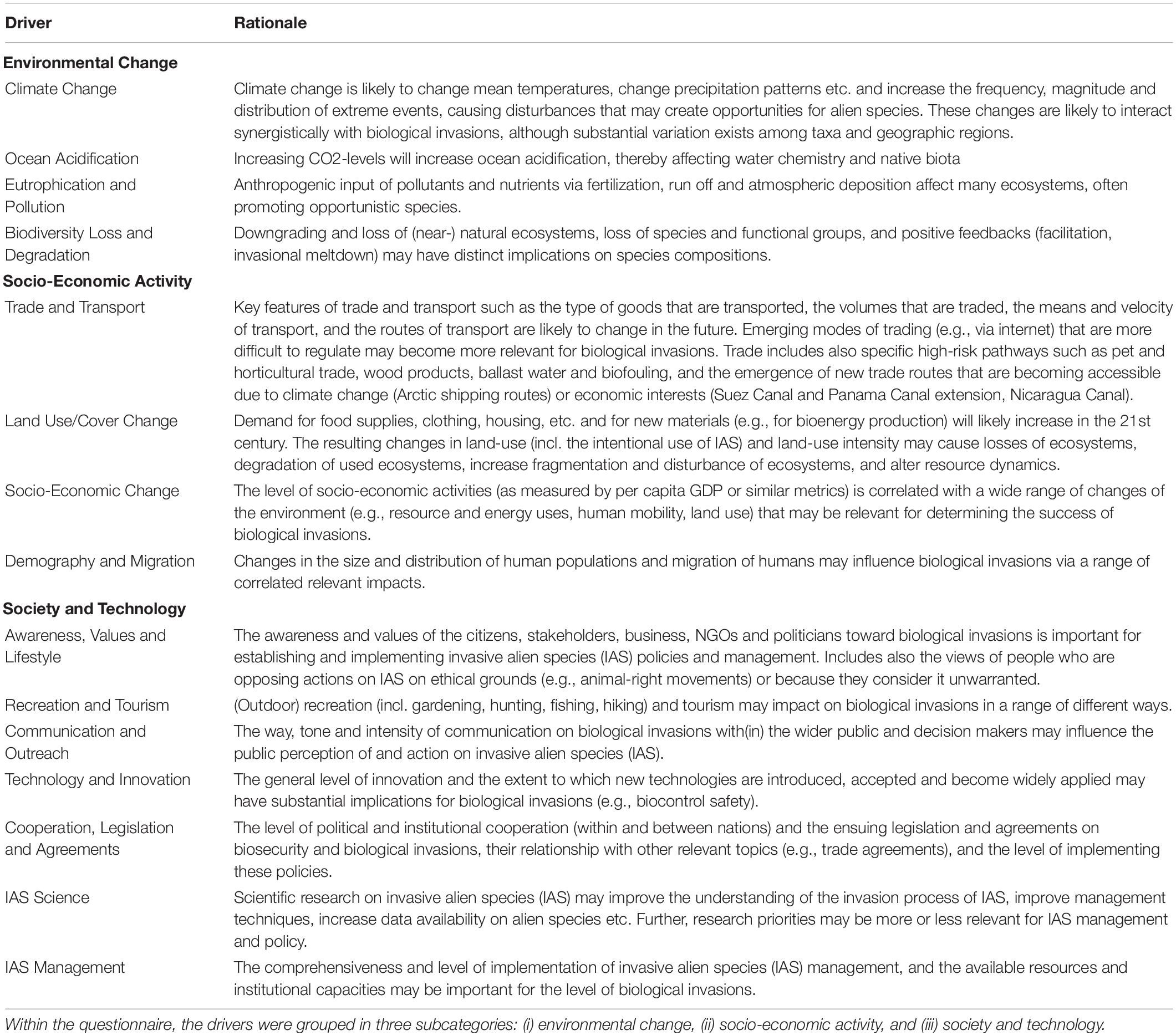
Table 1. Overview table of the 15 drivers used in the survey and their rationale on how they might affect biological invasions.
The questionnaire was divided into four sections: (1) drivers of biological invasions, (2) pathways of biological invasions, (3) effects of alien species richness increases related to three island contexts (i.e., types, systems and habitats), and (4) personal background and expertise of respondents. Section 1 contained questions related to three thematic groups of drivers (see above). For each thematic group, we asked the survey respondents to indicate which of the drivers will strongly increase alien species richness on islands within the 21st century in each of the three main stages of invasion (i.e., introduction, establishment and spread). In section 2, we asked which pathways will substantially increase alien species richness in the introduction and spread stages on islands within the 21st century. The pathways are based on Hulme et al. (2008) and include six categories: stowaway, escape, contaminant, release, corridor, unaided. Section 3 related to the respondents’ projection of how strongly an increase in invasive alien species richness in the 21st century will affect different island types (oceanic islands, continental islands and atolls), island systems (marine, freshwater, terrestrial) and habitats (e.g., wetlands, agricultural land; see the full list in Supplementary Material S2). Finally, section 4 aimed at collecting background information on the level and area of expertise of the respondents (focal taxonomic groups, geographic regions, island types, and socio-economic status of their study area) and personal information (gender, age, country of home institution).
The questionnaire was implemented using Google Forms©. The respondents had to answer all questions (i.e., they were not able to skip a questions) in the provided order and could only switch to the next page once all questions had been answered. Introductory text and guidance were provided for each question where necessary (see survey layout in Supplementary Material S2). Survey responses were in the form of a Likert scale (e.g., strongly disagree < disagree < neutral < agree < strongly agree, or low < medium < strong). The link to the online survey was distributed among the 262 participants of the 3rd International Conference of Island Biology 2019 on La Réunion Island.1 The survey was sent out twice during the conference and once after the conference (July 2019). Additionally, to reach an even wider audience, the survey was circulated once through relevant mailing lists (August 2019), namely the Island Conservation Network mailing list (Island-L;aXNsYW5kcy1sQGxpc3RzZXJ2LmJnY2kub3Jn) which has 382 subscribers (as of February 2020) and the ALIENS-L mailing list (aliens-l@list.auckland.ac.nz) of the Invasive Species Specialist Group (ISSG) of the IUCN Species Survival Commission with 1449 subscribers (as of February 2020).
For two of the questions concerning their personal background and study system, the respondents were given the option to provide free text additions to their answers. Where possible, we integrated the additional information on personal background into the default categories, whereas the answers stating a role in policy or government-related positions were assigned to a new additional category named “policy.” For the question on study systems, we likewise added an additional post-survey category named “island-like system”; this category encompassed respondents who stated they work with island-like systems (e.g., fragmented landscapes) but not “true” islands. We provide the original data and their category assignment in the Supplementary Table S1.
To assess the agreement between respondents on the importance of the different drivers, pathways and island characteristics for each stage of invasion, we fitted proportional odds models with a logit-link function using the ‘clm’-function in the package ‘ordinal’ (Christensen, 2019) in the statistical programming software R version 3.6.1 (R Core Team, 2019). Individual models were run for each invasion stage for section 1 (three models: introduction, establishment and spread) and 2 (two models: introduction and spread) of the survey. Survey responses were modeled as a function of the individual categorical drivers of invasion. Similarly, three different models were run for section 3 to assess how island types, systems and habitats might be affected in the future. Again, survey responses were modeled as a function of the individual categorical drivers of invasion. Model fit was assessed visually and none of the models violated the proportional odds assumption.
To assess the agreement or disagreement of respondents with the postulated contribution of drivers to future invasions, we evaluated whether given answers differed statistically from a neutral assumption of random answers. To this end, we introduced an additional factor level (i.e., formally a new driver) which was assumed to be assessed at random by all respondents, i.e., we assigned an equal number (n = 25) of responses to the response options “strongly disagree,” “disagree,” “agree” and “strongly agree” and 26 responses to the neutral response option to resemble the 126 respondents of the survey. We then used this dummy category as a reference level and all the other drivers were tested on whether they differed significantly from it. A significantly higher odds ratio was interpreted as agreement with the postulated effect, a non-significantly different one as neutral and a significantly lower odds ratio as disagreement.
In total, 126 responses were obtained. If fully attributed to either of the channels through which we distributed the survey, this would equate to response rates of 48% of the conference participants, 32% of the members of the Island-L mailing list, or 9% of the Aliens-L mailing list. The gender ratio among the respondents was 74 men to 50 women with two respondents not providing this information (option “prefer not to say”). The age ranged from <25 (n = 3) to >65 (n = 10) years of age with most people in the age class of “36–45” (n = 51). The responses regarding the location of home institution (which was an open-text question) sometimes included only a country or an island group, which might have somewhat distorted the true picture as many islands are administrative units of certain countries; meanwhile, four participants did not provide a clear answer that could be assigned to a country or island group. Overall, home institutions were distributed across the world with participants situated on islands within all major ocean realms, as well as on all continents, but most participants had their home institutions in Europe (n = 42) followed by mainland United States (n = 28). However, most researchers worked on North Pacific Islands (n = 37), North Atlantic Islands (n = 37), Indian Ocean Islands (n = 33) and islands in the South Pacific Ocean (n = 26). Fewest worked on South Pacific Islands (n = 14), Southern Ocean Islands (n = 6), and Arctic Ocean Islands (n = 1). Finally, 19 respondents indicated that they worked at a global scale. Within their study regions, respondents mainly worked with oceanic islands (n = 108), followed by continental islands (n = 39) and atolls (n = 16). Most islands studied by the respondents are situated in developed countries (n = 85), followed by developing (n = 40) and emerging (n = 39) ones (see the questionnaire in Supplementary Material S2 for definitions of the socio-economic background of the study regions). Most respondents indicated that their professional background was in conservation management (n = 80), followed by applied research (n = 55), basic research (n = 52), policy (n = 21), other stakeholders (n = 4) and interested citizens (n = 3). Finally, respondents working in the marine realm had highest expertise with vertebrates, followed by invertebrates, plants and microorganisms. In the terrestrial realm, highest expertise was indicated for plants, followed by vertebrates, invertebrates and microorganisms. All data on personal information and scientific background and expertise are summarized in Supplementary Figure S1.
For the introduction stage, respondents almost uniformly pinpointed trade and transport (99.2% agreement) as an important driver of future alien species richness, followed by recreation and tourism (92.9% agreement) and demography and migration (92.8% agreement), and with some respondents indicating distance by land use/cover change (80.2% agreement), socio-economy (76.2% agreement) climate change (68.3% agreement) and biodiversity loss and degradation (68.2% agreement). The remaining drivers were not considered to contribute to the increase in alien species richness at the introduction stage, i.e., the agreement for these drivers was <50% and the odds ratio of agreement to their contribution was not significantly higher as for the dummy category (see Figures 1, 2A and Table 2A).
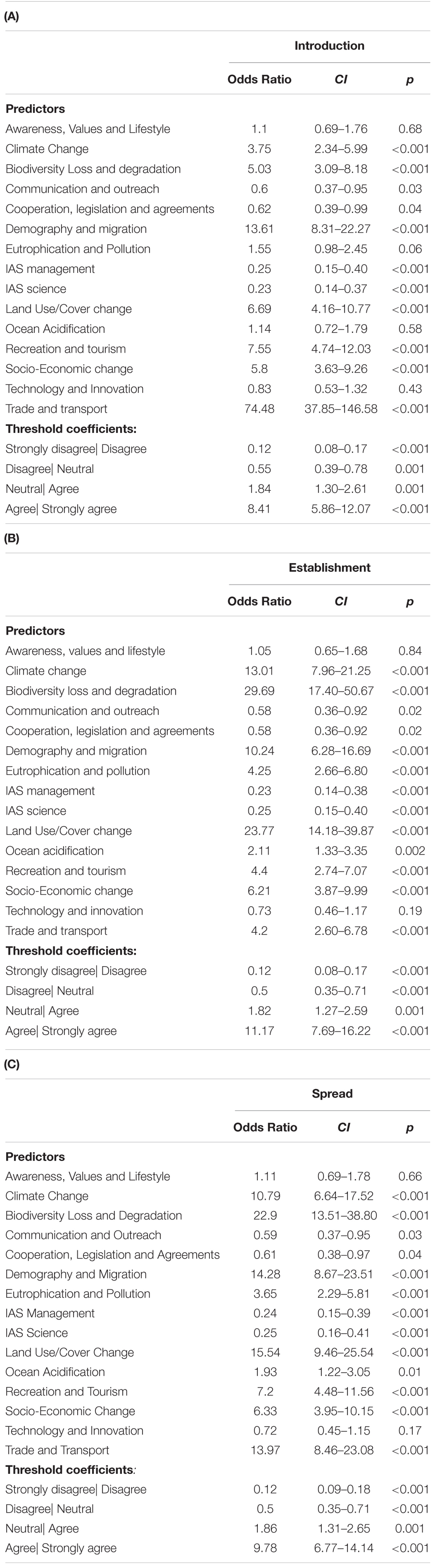
Table 2. Proportional odds models analyzing if the 15 drivers of biological invasions addressed in the survey significantly increase alien species (A) introduction, (B) establishment, and (C) spread on islands in the 21st century.
At the establishment stage, the main drivers of the increase in alien species richness were considered to be land use/cover change (96.8% agreement), biodiversity loss and degradation (96.0% agreement), climate change (93.7% agreement), and demography and migration (85.7% agreement) as well as socio-economy (80.2% agreement), recreation and tourism (74.6% agreement), eutrophication and pollution (73.8% agreement) and trade and transport (65.9% agreement). The remaining drivers were considered to not contribute substantially to alien species richness increase at the establishment stage, with <50% agreement and odd ratios that did not deviate significantly from the dummy category (see Figures 1, 2B and Table 2B).
At the spread stage, the drivers of alien species richness were considered to be land use/cover change (95.2% agreement), climate change (91.3% agreement), biodiversity loss and degradation (91.3% agreement), demography and migration (88.9% agreement), recreation and tourism (86.5% agreement), trade and transport (86.5% agreement), socio-economy (81.0% agreement) and eutrophication and pollution (68.2% agreement). The remaining drivers were considered not to contribute substantially to alien species richness increase at the spread stage, with <50% agreement and non-significantly deviating odd ratios from the dummy category (see Figures 1, 2C and Table 2C).
Across invasion stages, two drivers – technology and innovation and awareness, values and lifestyle – consistently showed no clear trend, i.e., not deviating significantly from the dummy category. For technology & innovation respondents disagreed slightly more with their importance as a driver of alien species richness (introduction: 30.2% agreement and 38.9% disagreement; establishment: 28.6% agreement and 41.3% disagreement; spread: 27.0% agreement and 41.3% disagreement; see Figure 2 and Table 2), whereas for awareness, values and lifestyle, the results were more balanced (introduction: 38.9% agreement and 35.7% disagreement; establishment: 35.7% agreement and 33.3% disagreement; spread: 34.9% agreement and 32.5% disagreement; see Figure 2 and Table 2).
For the introduction stage, respondents consistently agreed that all six pathways will substantially contribute to an increase in alien species richness, i.e., > 50% agreement and significantly higher odd ratios than for the dummy category for all of them. The responses identified strongest agreement for species introductions as stowaway (93.7% agreement), followed by escapes (89.7% agreement), contaminants (86.5% agreement), release (74.6% agreement), corridor (67.5% agreement) and unaided (65.9% agreement).
For the spread stage, respondents agreed again to all introduction pathways increasing alien species richness. Strongest agreement was provided for stowaway (78.6% agreement), escape (74.6% agreement) and corridor (71.4% agreement), followed by contaminant (69.8% agreement), unaided (65.9% agreement) and release (64.3% agreement). The overall agreement on pathway contributions to alien species richness was stronger at the introduction rather than the spread stage. See Figure 3 and Table 3 for the answer structure and model outputs.
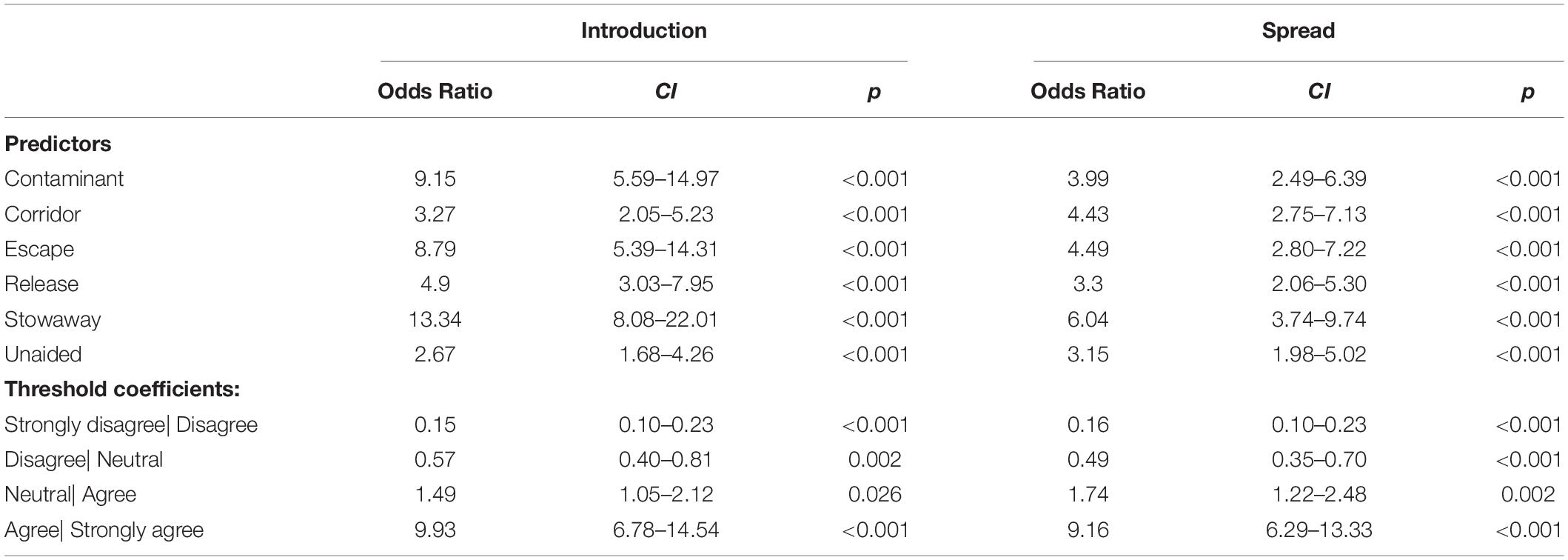
Table 3. Proportional odds models, analyzing which pathway will significantly increase alien species (A) introductions and (B) spread to/on islands in the 21st century.
All respondents agreed that each island type would be affected by a future increase in alien species richness, with oceanic islands being affected more strongly (85.7% strong effects) than atolls (57.1% strong effects) and continental islands (52.4% strong effects).
The respondents were also consistently convinced that an increase in invasive alien species richness will have ecological implications across different types of island systems. Strongest effects were expected for terrestrial (81.8% strong effects), freshwater (69.0% strong effects) and marine systems (65.9% strong effects).
Across habitats, strongest effects were suggested for terrestrial coastal habitats (70.6% strong effects), wetlands (67.5% strong effects), dry forests (60.3% strong effects), marine habitats (57.1% strong effects), and settlements (52.4% strong effects). Least strong effects were anticipated for mountain and alpine habitats (43.6% strong effects; odds ratio = 3.6; p-value = 0.004) and (semi-)deserts (25.4% strong effects; odds ratio = 0.8; p-value = 0.346). For information on the answer structure and model outputs for all impact related analyses see Figure 4 and Table 4.
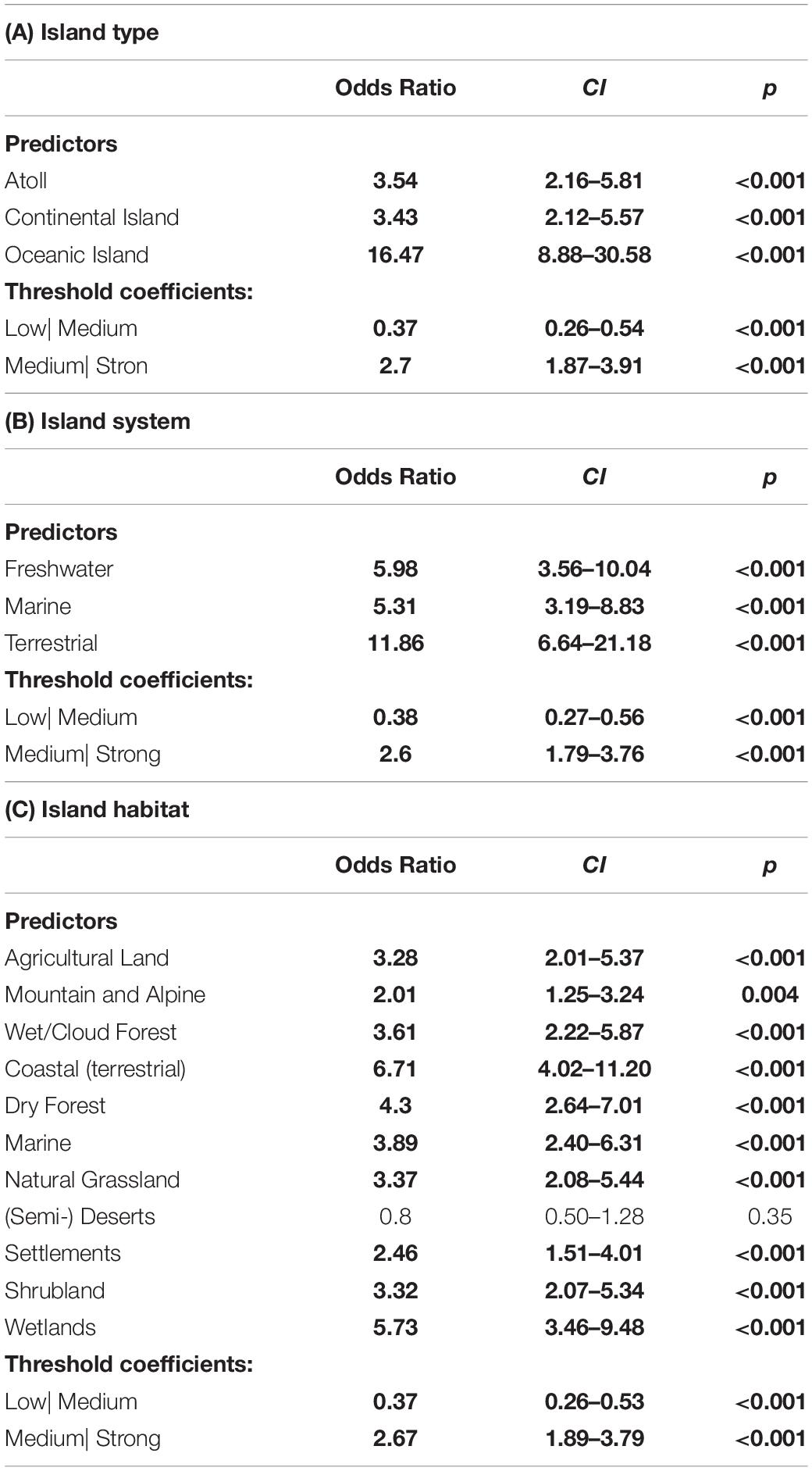
Table 4. Proportional odds models, analyzing how strongly an increase in alien species richness in the 21st century will affect different (A) island types, (B) island systems, and (C) island habitats.
Our study identifies six drivers that, according to experts, should substantially contribute to alien species richness increase on islands during the 21st century. At the introduction stage these drivers are mainly related to socio-economic activities like trade and transport, recreation and tourism and demography and migrations. At the establishment and spread stage the drivers change and mainly include environmental and anthropogenic change drivers, namely land use/cover change, biodiversity loss and degradation and climate change (Figure 1). Major pathways assumed to substantially increase future alien species richness at the introduction stage are stowaways, escapes and contaminants. At the spread stage the first two pathways remain similar, with contaminants being third in importance (Figure 1). All results are discussed in more detail in the following sections.
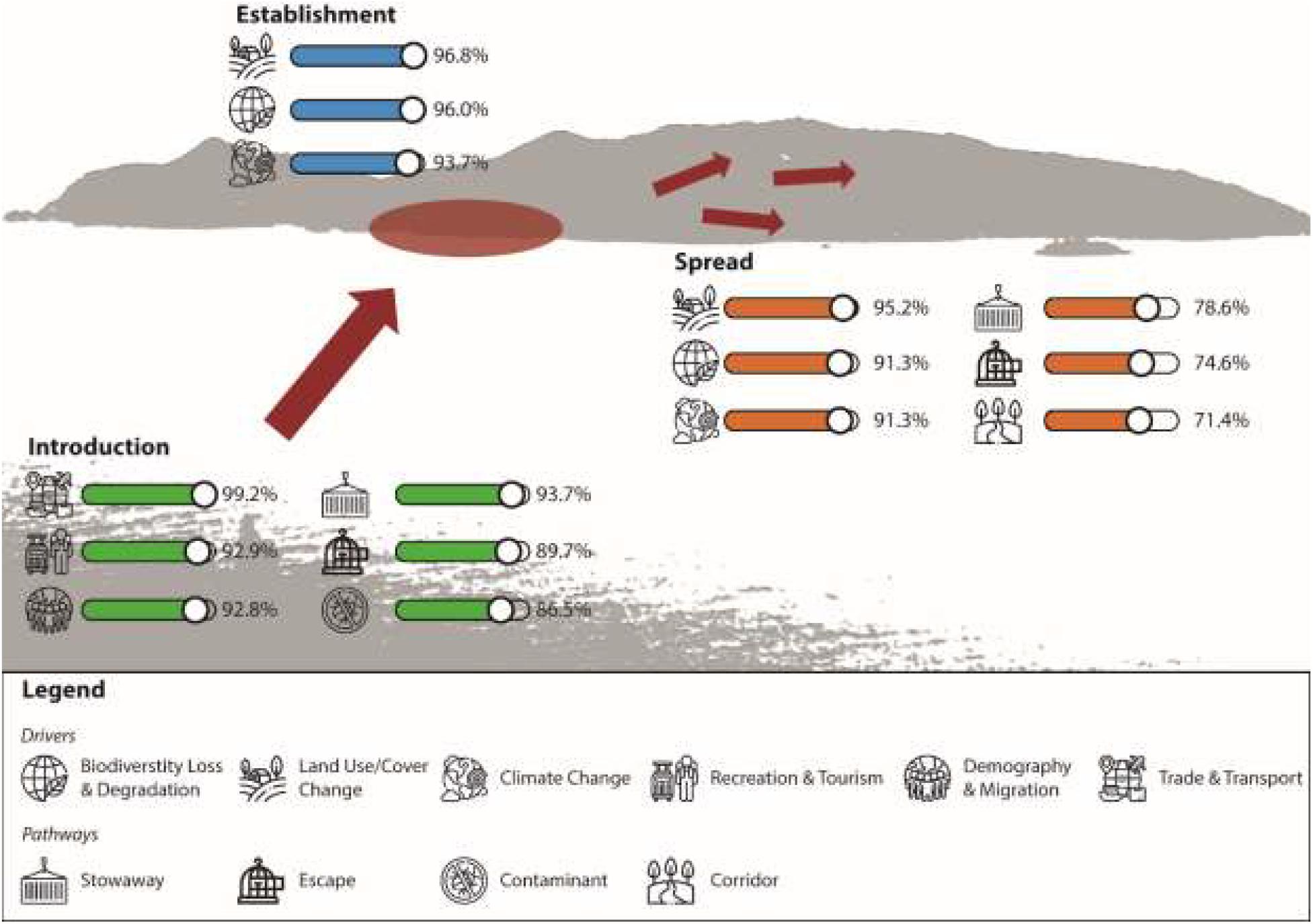
Figure 1. Summary of the three main drivers of alien species richness for all invasion stages and the three main pathways of alien species introduction and spread contributing to substantially to an increase of alien species richness. Values are given for the percent of combined answers in the categories “agree” and “strongly agree.” Icons are made by “Freepik” from “www.flaticon.com.”
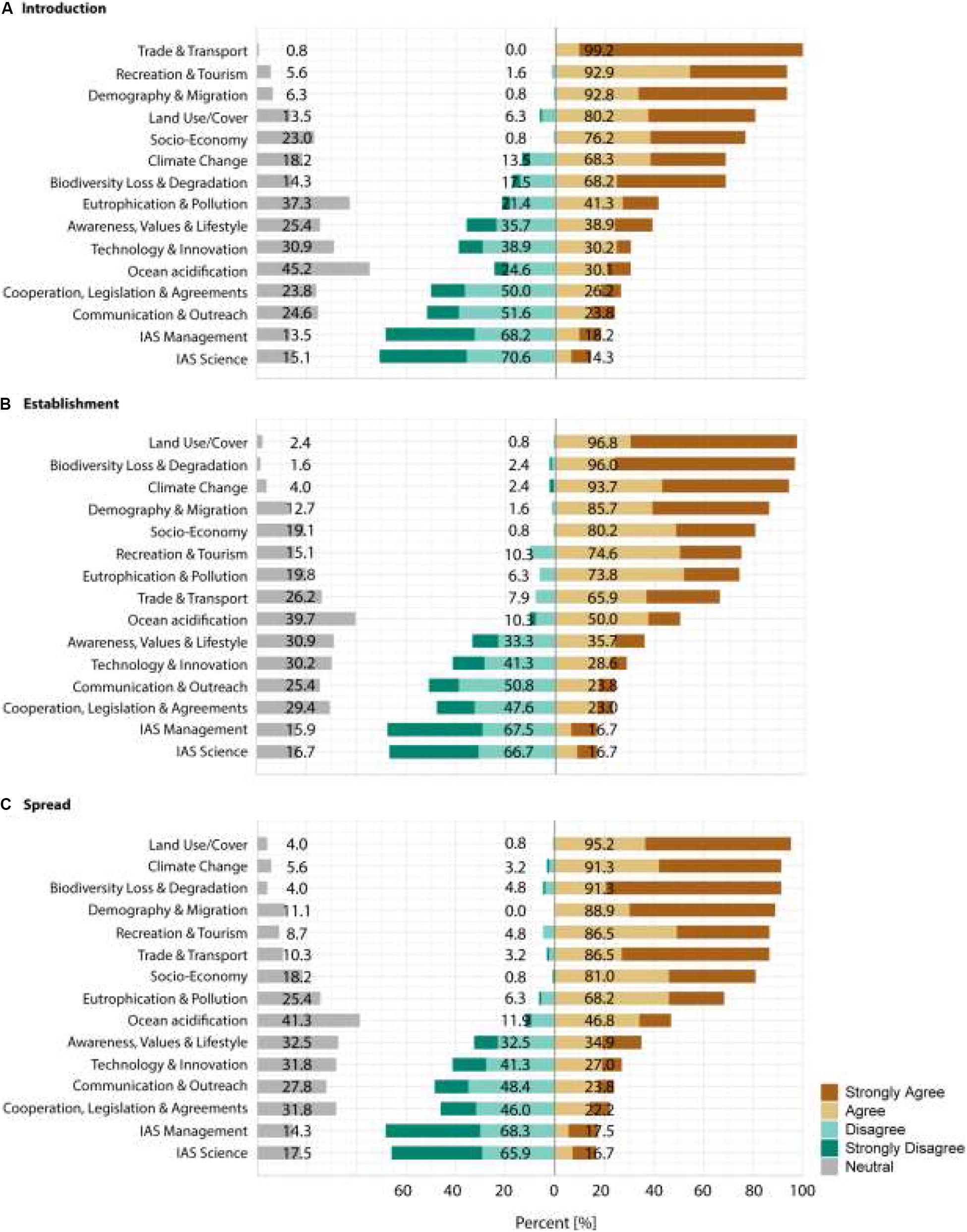
Figure 2. Summary of the answers provided by the 126 respondents for the first section of the survey. Respondents were asked which of the drivers will significantly increase alien species (A) introductions, (B) establishment and (C) spread to/on islands in the 21st century. Answers were provided on a 5-point Likert scale with the categories: strongly disagree, disagree, neutral, agree and strongly agree. Shown is the percentage of agreement with each of the categories. Values are given for the percent of answers in the neutral category and for the disagreement and agreement categories grouping the respective two answer possibilities.
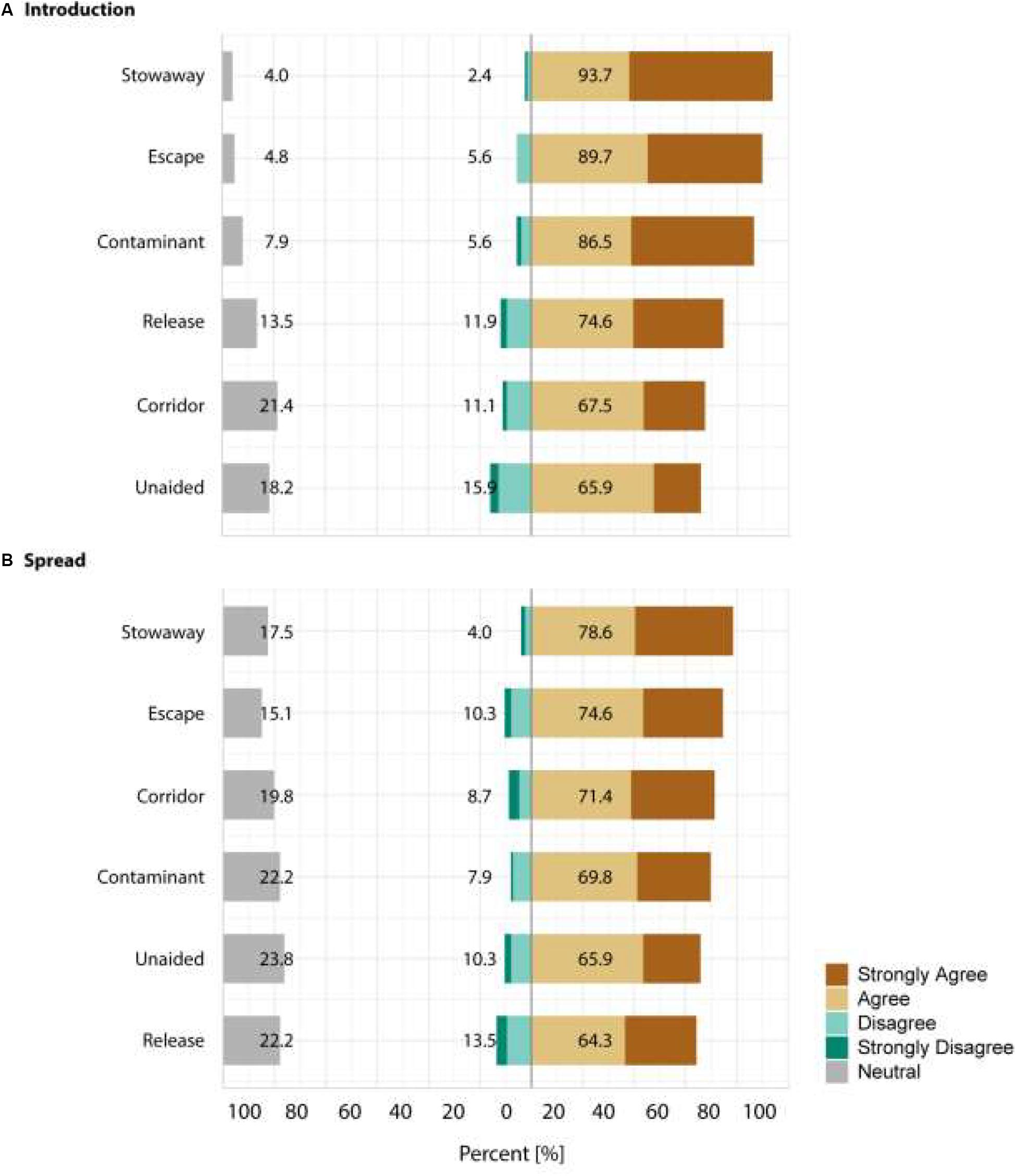
Figure 3. Summary of the answers provided by the 126 respondents for the second section of the survey. Respondents were asked which pathway will significantly increase alien species (A) introductions and (B) spread to/on islands in the 21st century. Answers were provided on a 5-point Likert scale with the categories: strongly disagree, disagree, neutral, agree, and strongly agree. Shown is the percentage of agreement with each of the categories. Values are given for the percent of answers in the neutral category and for the disagreement and agreement categories grouping the respective two answer possibilities.
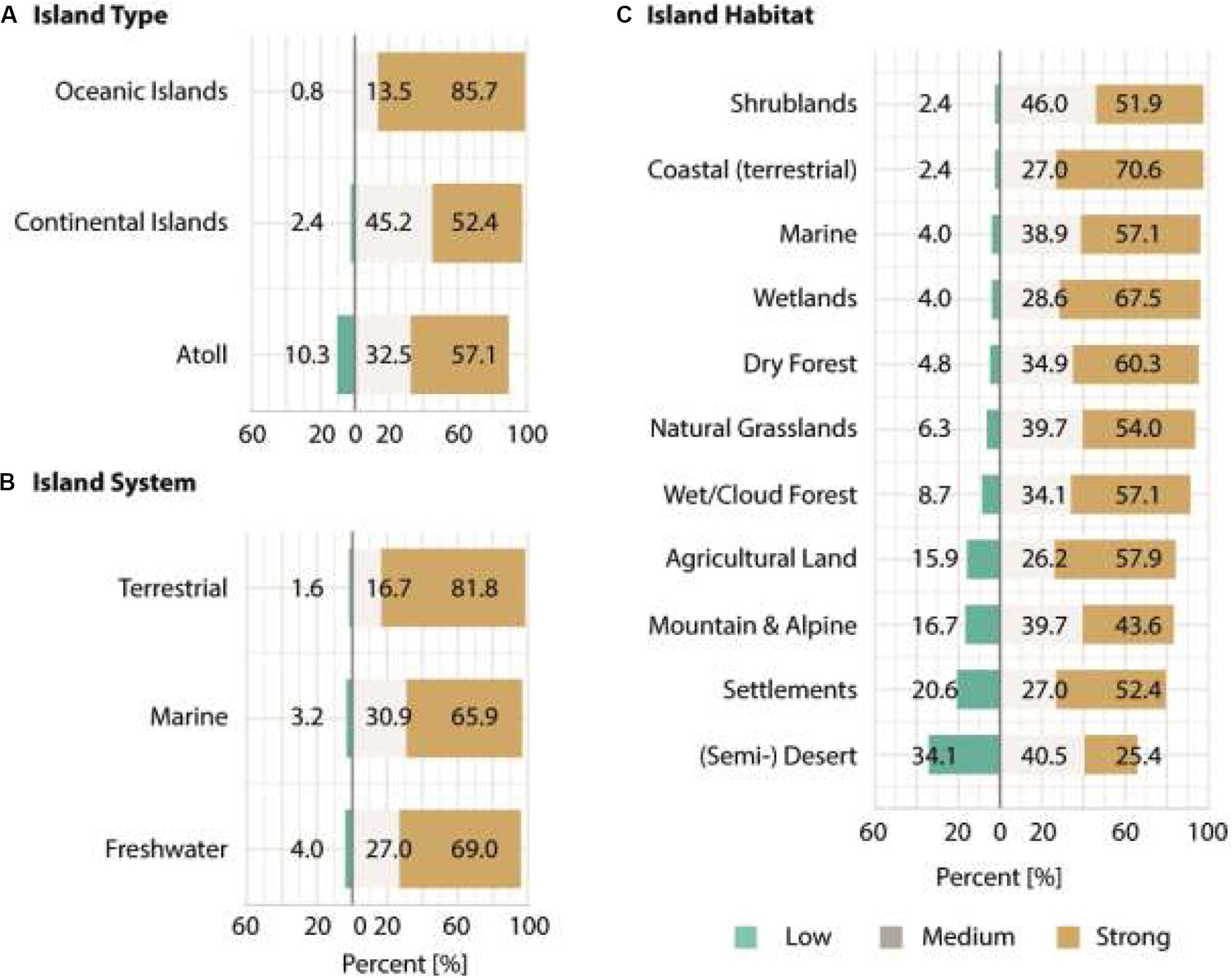
Figure 4. Summary of the answers provided by the 126 respondents for the third section of the survey. Respondents were asked how strongly an increase in alien species richness in the 21st century will affect different (A) island types, (B) island systems, and (C) island habitats. Answers had to be provided on a 3-point Likert scale with the categories: low, medium, strong. Shown are the percentage results for each of the categories.
Per definition, alien species are distributed by human agency to regions outside their native range and hence their introduction is inherently tied to the movement of commodities and people around the world (Essl et al., 2018). Thus, respondents unanimously identified socio-economic activity drivers such as trade and transport, demography and migration and recreation and tourism as major drivers of future alien species richness on islands. As current scenarios of global trade assume that traded commodities will double or increase more than 4-fold between 2015 and 2050 (ITF, 2017), the numbers of species introductions are likely to increase for islands in the future. Additionally, the global trade network is projected to change over the course of the 21st century, including a stronger integration of hitherto less well integrated regions, which would change the frequencies, volumes and travel times of traded goods (World Trade Organisation, 2013). With changing frequencies and volumes of traded goods, propagule pressure and colonization will increase, and shorter travel times will increase the survival probability of transported propagules, all contributing to higher alien species richness in respective regions (Seebens et al., 2015; Sardain et al., 2019). Furthermore, the respondents identified stowaways and contaminants as the leading pathways, which is in line with what has been observed in other studies (Hulme, 2009; Pergl et al., 2017). The outlined changes in global trade are not exclusive to islands but apply to all regions worldwide. However, islands typically have high volumes of imports as most commodities are not produced on the islands themselves, which together with higher invasibility compared to mainland regions increases their invasion risk (Hulme, 2009; Moser et al., 2018; OECD, 2018). Hence, on islands the proportion of unintentionally introduced alien species will likely increase. Given the isolated nature of islands, biosecurity measures are more easily implemented compared to mainland regions, and prevention measures are preferred and most cost-efficient in the context of islands (Leung et al., 2002; Russell et al., 2017).
The identified main drivers of future invasions differ between the introduction and the establishment stages. The drivers associated with the environment (e.g., climate change and biodiversity loss and degradation) and human activity (e.g., land use/cover change, socio-economy or demography and migration) become prevalent at the establishment stage. Anthropogenic habitat destruction has indeed been shown to strongly increase alien species richness and abundance across habitats, at the expense of native species richness (Sanchez-Ortiz et al., 2019). Other anthropogenic disturbances like infrastructure development additionally increase alien species establishment (Alexander et al., 2016; Haider et al., 2018), and urban and artificial environments already hold a high diversity of alien species, often buffering them from adverse environmental conditions (Strubbe and Matthysen, 2009). Finally, anthropogenic habitat transformation (IPBES, 2018), future climate change (Bellard et al., 2013; Harter et al., 2015), and socio-economic activity (Hulme, 2009; Seebens et al., 2015) also indirectly promote the establishment of new alien species through the loss of native biodiversity.
Overall, the participants assessed alien species spread to be promoted by a combination of drivers relevant at the introduction and establishment stages. The respondents particularly agree that land use/cover change, biodiversity loss and degradation and climate change will strongly contribute to an increase of alien species richness in the future. These drivers are closely followed by the same drivers related to the movement of people (e.g., recreation and tourism) and goods (e.g., trade and transport) that are most important at the introduction stage. This assumption appears intuitive, as with ongoing anthropogenic pressure and disturbance more suitable habitat for alien species opens up, and increased movement of people and goods then facilitates secondary spread of species within and between island groups. Similar to the introduction phase, unintentional dispersal pathways (i.e., stowaways and contaminants) appear to be most relevant for alien species richness increases at the spread stage.
The strong support for demography & migration across invasion stages as one of the main drivers for alien species on islands is indicative of complex dynamics with other drivers, especially tourism. This appears to contradict future projections of human population trends on islands, which indicate an earlier saturation or even a decrease of human population sizes in island regions like the Caribbean, Oceania or the group of small island-developing countries compared to continental regions (United Nations et al., 2019). Additionally, many small island states face increasing threats of climate change driven sea-level rise, leading to forced migration (Kelman, 2010; Bellard et al., 2014). Emigration often leads to the abandonment of agricultural and highly modified regions and ecosystems. The subsequent succession in these areas often favors the establishment of alien species that are better adapted to human-modified landscapes compared to native species (Rey Benayas et al., 2007; Plieninger et al., 2014). However, many islands are tourist destinations with more people projected to arrive in the future. This emerging economic incentive might result in more people remaining or returning to islands, which would slow down expected demographic trends. Cumulative effects of demography and migration and recreation and tourism with the subsequent development of infrastructure (e.g., roads and hiking trails; Haider et al., 2018; Liedtke et al., 2020) might foster future establishment of alien species as these structures have been shown to be introduction pathways (Toral-Granda et al., 2017). Additionally, cultivation of alien species near tourist accommodations and in public green spaces further increases the probability that these species might jump the fence and establish new populations outside gardens (Cowie et al., 2008; van Kleunen et al., 2018). In several instances, charismatic alien species have as well adopted an economic value for the local community because they attract tourists (Jarić et al., 2020). The relevance of both drivers is in agreement with our study results.
The respondents have different perceptions on the role of technology and innovation for different invasion stages in the future. While there is a tendency toward technology optimism, meaning that technological development will not result in an increase of alien species richness, about one third of respondents have the opposite opinion. This ambivalence in responses might suggest that technological development can have strongly diverging effects on biological invasions. On the one hand, environmental DNA (eDNA) is already used for early detection and rapid response to alien species introductions (Thomsen and Willerslev, 2015; Dougherty et al., 2016). Especially in aquatic systems, eDNA is a highly effective tool to detect alien species at low population densities (Dejean et al., 2012). On islands, the use of eDNA methods will very likely result in better biosecurity effectiveness due to increased early detection rates and improved IAS surveillance (Herder et al., 2014; Tingley et al., 2019). Smart applications for reporting nature observations are used in citizen science projects to detect, monitor and manage alien species (Mannino and Balistreri, 2018; Roy et al., 2018; Johnson et al., 2020) and satellite data are used via remote sensing for alien species mapping and management (Henderson and Dawson, 2009; Robin et al., 2011; Rocchini et al., 2015; Rivas-Torres et al., 2018). On the other hand, e-commerce has resulted in increased alien species introductions and has emerged as a new pathway of introductions that is difficult to manage (Lenda et al., 2014; Humair et al., 2015), which likely explains why the divergence in opinion was higher for the introduction than for the two other stages of invasion. Furthermore, upon the initial introduction of goods to major transportation hubs, secondary spread of alien species might be facilitated by more localized trade with higher transportation frequencies resulting from increased automation using, for example, block chain techniques, reduced transit times, and new consumer good distribution techniques (McKinsey Global Institute, 2019).
A similarly ambiguous assessment was provided for the effects of awareness, values and lifestyle on alien species richness across invasion stages. Prevention of introduction and post-introduction early detection and rapid response are the most effective management options against alien species introduction (Reaser et al., 2020), which is reflected in the responses, showing that this driver has the lowest contribution toward the increase of alien species richness at the introduction stage. At later stages, building awareness might be more difficult, as many alien species that were introduced a long time ago, may have been incorporated culturally or economically into local communities and are now being perceived as “native.” For example, the prickly pear (Opuntia ficus-indica) was introduced to the Macaronesian islands as an economically important fodder and to obtain red and purple pigment (Prance and Nesbit, 2005). Nowadays prickly pear occurs across all islands and is even used to advertise the beauty of the islands to tourists. Another aspect related to peoples’ values toward alien species might lie in their perception of a species based on its charisma (Jarić et al., 2020) and in several instances, alien species management plans have failed due to strong opposition of the general public and activist groups (e.g., Bertolino and Genovesi, 2003; Verbrugge et al., 2013). Finally, there can be important discrepancies in the perception of alien species between archipelagos and even between islands within an archipelago, with people in more remote islands tending to see more benefits in alien species (Meyer and Fourdrigniez, 2019). It is hence important to include all relevant stakeholders in the decision-making process for managing alien species as well as transparently communicating such actions and their relevance to the general public (Novoa et al., 2018; Shackleton et al., 2019). Especially on islands, where native biodiversity is a major economic pillar, such societal transformations with respect to awareness, values and lifestyle are crucial and feasible. However, it is likely that efforts toward societal transformation will not show their result within the near future, a delay, which probably motivated the ambivalent responses in our study.
Finally, the survey shows consensus in that knowledge generation (i.e., IAS science), dissemination (i.e., communication and outreach) and pro-active actions (i.e., IAS management) does not substantially increase future alien species richness. For a long time, islands have been at the forefront of biological research, dating back to Charles Darwin and Alfred Russel Wallace. They have provided valuable testing grounds for theory development and conservation planning (Whittaker and Fernandez-Palacios, 2007). Given their isolated nature and limited spatial extent, biosecurity measures are substantially easier to implement compared to mainland regions. For example, on the Galapagos Islands, where nature-based tourism is a vital part of the local economy, IAS management is a top priority, with the highest allocated budget of 2.5 million US$/yr in a study of 21 protected areas globally (González et al., 2008; Self et al., 2010; Shackleton et al., 2020). Similarly, New Zealand has some of the strictest biosecurity protocols regarding IAS and an ambitious governmental program (“Predator-Free NZ”) aiming to eradicate a selection of invasive predators by 2050 (Russell et al., 2015). This strong history of research communication and acknowledgment of the risk of alien species might contribute to a slowdown of future alien species richness increases on islands even though this conclusion cannot be drawn directly from our survey (see limitations section below for a short discussion).
Most respondents agree that effects of increases in alien species richness will occur mainly on oceanic islands, followed by atolls and continental islands. Oceanic islands show a higher degree of endemism and more disharmonic floras and faunas than the other island types (König et al., 2019; Taylor et al., 2019). This disharmony and proportion of endemism increases with island isolation (Kier et al., 2009; König et al., 2017) and at the same time isolation has been shown to increase island invasibility (Moser et al., 2018). Continental islands, on the other hand, have a more diverse set of native biota including functional guilds usually underrepresented or absent on oceanic islands and thus might be less affected by alien species introductions (Atkinson, 1989; Apanius et al., 2000). However, their proximity to the mainland generally leads to higher anthropogenic use (e.g., higher population sizes) and socio-economic exchange with the mainland (e.g., trade) and higher propagule pressure due to close proximity compared to more distant (usually oceanic) islands, facilitating species introductions (Ficetola and Padoa-Schioppa, 2009).
Respondents assumed strong impacts across all island systems (marine, freshwater and terrestrial) with the latter experiencing most dramatic impacts. While most anthropogenic activity is undoubtedly directed toward terrestrial ecosystems, all systems are tightly interconnected with strong cascading effects across all island ecosystems (Graham et al., 2018). Lower assumed impacts in marine regions might result from the fact that marine regions are less isolated from similar regions than terrestrial island regions and thus are less prone to biological invasions (results only including 17 participants out of 126 with high expertise for at least one marine taxonomic group provide a similar ranking; see Supplementary Figure S2). However, marine alien species introductions via the pet trade like the lionfish in the Caribbean (Pterois volitans and P. miles; Ricardo et al., 2011) have been shown to dramatically affect local environments and species communities (Ballew et al., 2016), highlighting the need to monitor and manage alien species in marine systems. It is noteworthy, that data availability on IAS and their management is particularly scarce in the marine realm (Ojaveer et al., 2015; Latombe et al., 2019a) and that therefore, our results may be influenced by this lack of knowledge and the lower taxonomic expertise of the survey participants as compared to other systems.
Lastly, island habitats that will likely experience the strongest assumed impacts in the future are coastal regions, wetlands and dry forests. Coastal habitats on islands are exposed to highest anthropogenic pressures, with major cities and transportation hubs and highest population densities found there (Russell et al., 2017; Andrew et al., 2019). Dry forest ecosystems tend to be situated close to coastal regions, especially on tropical and subtropical islands (Janzen, 1988). During the initial human colonization, many dry forest ecosystems were degraded and thus became highly vulnerable to species invasions (Janzen, 1988; Barbé et al., 2015), a trend that will likely persist in the future. Wetlands are probably even more vulnerable to alien species introduction and establishment than other habitats, as native wetland biodiversity (especially amphibians) is highly endemic (Duellman and Trueb, 1994; Inger, 2001). The introduction of invasive species to island wetlands, such as the cane toad (Bufo marinus) in the Pacific and Hawaiian Islands (Eldredge, 2000; Ellison, 2009), are a serious threat to local biodiversity. For all other habitats, the respondents agree that at least medium but mainly strong impacts are likely to occur in the future. The only exceptions are agricultural land and settlements, which are probably assumed to be already highly invaded with strong impacts and consequently impacts are assumed to be low in the future, as well as mountain and alpine and (semi-) desert habitats. For the latter two, the survey did not provide a clear trend in impact severity. While both habitats provide environmental conditions that might not suit many alien species, mountains are an attractive destination for tourists who can introduce species to these habitats (Seipel et al., 2012; Alexander et al., 2016).
Our study aims to provide a global picture of future drivers of alien species richness on islands by targeting a large panel of diverse respondents. Necessarily, such generality imposes trade-offs with respect to the level of detail in the questions asked and the framing of the drivers, to make them applicable across regions and systems. In section 1, the drivers are defined in a broad sense (see Table 1) and it is thus impossible to disentangle their effects at fine scales. For example, climate change encompasses many relevant facets like temperature changes, changes in precipitation patterns or the shifts in extreme event severity or frequency or alterations of oceanic currents and ocean chemistry. All these aspects are very important at the local and regional level, however will very likely vary dramatically between different regions of the world. Follow-up studies that target specific island regions with a more differentiated and context specific set of drivers are necessary to draw such conclusions. An interesting angle would for example be to discriminate between different taxonomic groups, which was not addressed in this study. Our study nonetheless provides a valuable synthesis on which drivers to look at more specifically.
In section 1, we asked the participants to provide an assessment of the degree of their (dis)agreement that certain drivers would increase alien species richness in the future. Disagreement does not necessarily imply the opposite effect, i.e., that the driver leads to a decrease in alien species richness. This is especially relevant for a set of drivers related to communication, management and research, and technology and innovation, for which the participants disagreed with a positive effect of the drivers on alien species richness. We interpreted this result as the expression of a negative relationship between these drivers and species richness, rather than the absence of a relationship. Although this assumption cannot be directly inferred from the survey results, it is based on the similarity in expertise between the co-authors of this study and the respondents and is supported by the existing body of literature.
Expert surveys are only as representative as the sample of respondents that take part in it and the same survey among a different group of experts might produce diverging results. We acknowledge that the participants’ background is skewed toward European and Northern American institutions, expertise in terrestrial systems and plants and vertebrates, a reoccurring bias in ecological research (e.g., Troudet et al., 2017; Nuñez et al., 2019). For example, the results regarding alien species richness effects on island types or systems mirror to some degree the expertise of the respondents. We thus cannot exclude any bias in the answers given but are confident of the validity of the results following our discussion and the link to existing literature. Overall, our sample size of 126 participants is substantial and the participants represent all important island regions, major taxonomic groups and relevant scientific fields. We additionally include the answer structure to the survey including only respondents with specific expertise (e.g., for taxonomic groups or realms; Supplementary Figure S2).
Our survey provides a comprehensive expert-based assessment of the future importance of drivers, pathways of biological invasions on islands and their effects in different contexts. Experts across different fields of expertise and with varying backgrounds have high confidence that the movements of goods and people and related activities like tourism and recreation will continue to be the major drivers of alien species introduction and subsequent spread in the future. Additionally, biotic and abiotic factors, such as land use/cover change, biodiversity loss and climate change, are acknowledged to play crucial roles in the increase of alien species richness on islands after introduction.
In the meantime, the responses from our survey suggest that experts do not expect a silver bullet that will provide a mid-term solution to alien species-related issues. Technological innovation might play out either way, supporting or restricting alien species introduction and spread. Societal processes like changes in awareness, lifestyles and values are considered to have little to no substantial effects within the next few decades. However, transformation of societal norms is crucial in the long term, and this is supported by a consensus among the respondents. The adoption of strict biosecurity measures and pro-active communication regarding the threats from alien species is believed to lead toward effective prevention and management of biological invasions.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
BL and FE designed the research. BL performed the analysis with input from GL, CC, and FE. BL led the writing of the initial manuscript with input by GL, CC, and FE. BL designed the questionnaire with input by SDHI, HS, PW, GL, CC, and FE. All other authors contributed to writing and commented on the manuscript. All authors contributed to the article and approved the submitted version.
BL, FE, DM, SD, GL, FC, CD, AT, MG, IK, HS, and NR-P appreciate funding by the BiodivERsA-Belmont Forum Project “Alien Scenarios” (BL, FE, DM, SD, and GL: FWF project no: I 4011-B32; NR-P: AEI PCI2018-092966; MG and IK: BMBF/PT DLR 01LC1807C; HS: BMBF/PT DLR 01LC1807A). FC, CD, and AT were also supported by the AXA Chair in Invasion Biology.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The handling editor declared a past co-authorship with several of the authors, BL, FC, and FE.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2020.00280/full#supplementary-material
Alexander, J. M., Lembrechts, J. J., Cavieres, L. A., Daehler, C., Haider, S., Kueffer, C., et al. (2016). Plant invasions into mountains and alpine ecosystems: current status and future challenges. Alp. Bot. 126, 89–103. doi: 10.1007/s00035-016-0172-178
Andrew, N. L., Bright, P., de la Rua, L., Teoh, S. J., and Vickers, M. (2019). Coastal proximity of populations in 22 Pacific Island Countries and Territories. PLoS One 14:e223249. doi: 10.1371/journal.pone.0223249
Apanius, V., Yorinks, N., Bermingham, E., and Ricklefs, R. E. (2000). Island and taxon effects in parasitism and resistance of Lesser Antillean birds. Ecology 81, 1959–1969. doi: 10.1890/0012-96582000
Atkinson, I. A. E. (1989). “Introduced animals and extinctions,” in Conservation for the Twenty-First Century, eds D. Western and M. Pearl (New York, NY: Oxford University Press), 54–69.
Ballew, N. G., Bacheler, N. M., Kellison, G. T., and Schueller, A. M. (2016). Invasive lionfish reduce native fish abundance on a regional scale. Sci. Rep. 6, 1–7. doi: 10.1038/srep32169
Barbé, M., Fenton, N. J., Lavergne, C., Le Péchon, T., Baider, C., and Gigord, L. D. B. (2015). Changes in lowland dry-forest native and alien plant communities on Réunion Island (Indian Ocean) over 16 years. Botany 93, 843–857. doi: 10.1139/cjb-2015-2112
Bellard, C., Cassey, P., and Blackburn, T. M. (2016). Alien species as a driver of recent extinctions. Biol. Lett. 12:20150623. doi: 10.1098/rsbl.2015.0623
Bellard, C., Leclerc, C., and Courchamp, F. (2014). Impact of sea level rise on the 10 insular biodiversity hotspots. Glob. Ecol. Biogeogr. 23, 203–212. doi: 10.1111/geb.12093
Bellard, C., Rysman, J. F., Leroy, B., Claud, C., and Mace, G. M. (2017). A global picture of biological invasion threat on islands. Nat. Ecol. Evol. 1, 1862–1869. doi: 10.1038/s41559-017-0365-366
Bellard, C., Thuiller, W., Leroy, B., Genovesi, P., Bakkenes, M., and Courchamp, F. (2013). Will climate change promote future invasions? Glob. Chang. Biol. 19, 3740–3748. doi: 10.1111/gcb.12344
Berg, C., Rogers, S., and Mineau, M. (2016). Building scenarios for ecosystem services tools: Developing a methodology for efficient engagement with expert stakeholders. Futures 81, 68–80. doi: 10.1016/j.futures.2015.10.014
Bertolino, S., and Genovesi, P. (2003). Spread and attempted eradication of the grey squirrel (Sciurus carolinensis) in Italy, and consequences for the red squirrel (Sciurus vulgaris) in Eurasia. Biol. Conserv. 109, 351–358. doi: 10.1016/S0006-3207(02)00161-161
Buchadas, A., Zav, A. S., Honrado, J. P., Alagador, D., Bastos, R., Cabras, J. A., et al. (2017). Dynamic models in research and management of biological invasions. J. Environ. Manag. 196, 594–606.
Capinha, C., Essl, F., Seebens, H., Pereira, H. M., and Kühn, I. (2018). Models of alien species richness show moderate predictive accuracy and poor transferability. NeoBiota 38, 77. doi: 10.3897/neobiota.38.23518
Christensen, R. (2019). ordinal - Regression Models for Ordinal Data. Available online at: http://www.cran.r-project.org/package=ordinal
Courchamp, F., Fournier, A., Bellard, C., Bertelsmeier, C., Bonnaud, E., Jeschke, J. M., et al. (2017). Invasion Biology: specific problems and possible solutions. Trends Ecol. Evol. 32, 13–22. doi: 10.1016/j.tree.2016.11.001
Cowie, R. H., Hayes, K. A., Tran, C. T., and Meyer, W. M. (2008). The horticultural industry as a vector of alien snails and slugs: widespread invasions in Hawaii. Int. J. Pest Manag. 54, 267–276. doi: 10.1080/09670870802403986
Daehler, C. (2006). Invasibility of tropical islands by introduced plants: Partitioning the influence of isolation and propagule pressure. Preslia 78, 389–404.
Dawson, W., Moser, D., Van Kleunen, M., Kreft, H., Pergl, J., Pyšek, P., et al. (2017). Global hotspots and correlates of alien species richness across taxonomic groups. Nat. Ecol. Evol. 1:0186. doi: 10.1038/s41559-017-0186
Dejean, T., Valentini, A., Miquel, C., Taberlet, P., Bellemain, E., and Miaud, C. (2012). Improved detection of an alien invasive species through environmental DNA barcoding: the example of the American bullfrog Lithobates catesbeianus. J. Appl. Ecol. 49, 953–959. doi: 10.1111/j.1365-2664.2012.02171.x
Dougherty, M. M., Larson, E. R., Renshaw, M. A., Gantz, C. A., Egan, S. P., Erickson, D. M., et al. (2016). Environmental DNA (eDNA) detects the invasive rusty crayfish Orconectes rusticus at low abundances. J. Appl. Ecol. 53, 722–732. doi: 10.1111/1365-2664.12621
Duellman, W. E., and Trueb, L. (1994). Biology of Amphibians. Baltimore, MD: John Hopkins University Press.
Eldredge, L. G. (2000). “Non-indigenous freshwater fishes, amphibians, and crustaceans of the Pacific and HawaiianIslands,” in Invasive Species in the Pacific:A Technical Review and Draft Regional Strategy, ed. G. Sherley (Oakland, CA: SREP), 173–190. doi: 10.1016/S0378-777X(82)80016-4
Ellison, J. C. (2009). Wetlands of the pacific island region. Wetl. Ecol. Manag. 17, 169–206. doi: 10.1007/s11273-008-9097-9093
Essl, F., Bacher, S., Genovesi, P., Hulme, P. E., Jeschke, J. M., Katsanevakis, S., et al. (2018). Which taxa are alien? criteria, applications, and uncertainties. Bioscience 68, 496–509. doi: 10.1093/biosci/biy057
Essl, F., Dawson, W., Kreft, H., Pergl, J., Pyšek, P., Van Kleunen, M., et al. (2019a). Drivers of the relative richness of naturalized and invasive plant species on Earth. AoB Plants 11, 1–13. doi: 10.1093/aobpla/plz051
Essl, F., Lenzner, B., Courchamp, F., Dullinger, S., Jeschke, J. M., Kühn, I., et al. (2019b). Introducing AlienScenarios: A project to develop scenarios and models of biological invasions for the 21st century. NeoBiota 45, 1–17. doi: 10.3897/neobiota.45.33366
Essl, F., Lenzner, B., Bacher, S., Bailey, S., Capinha, C., Daehler, C. C., et al. (2020). Drivers of future alien species impacts: an expert-based assessment. Glob. Change Biol. doi: 10.1111/gcb.15199
CrossRef Full Text [Online ahead of print]
Ficetola, G. F., and Padoa-Schioppa, E. (2009). Human activities alter biogeographical patterns of reptiles on Mediterranean islands. Glob. Ecol. Biogeogr. 18, 214–222. doi: 10.1111/j.1466-8238.2008.00433.x
González, J. A., Montes, C., Rodriguez, J., and Tapia, W. (2008). Rethinking the Galapagos Islands as a complex social-ecological system: Implications for conservation and management. Ecol. Soc. 13:13. doi: 10.5751/ES-02557-130213
Graham, N. A. J., Wilson, S. K., Carr, P., Hoey, A. S., Jennings, S., and MacNeil, M. A. (2018). Seabirds enhance coral reef productivity and functioning in the absence of invasive rats. Nature 559, 250–253. doi: 10.1038/s41586-018-0202-203
Haider, S., Kueffer, C., Bruelheide, H., Seipel, T., Alexander, J. M., Rew, L. J., et al. (2018). Mountain roads and non-native species modify elevational patterns of plant diversity. Glob. Ecol. Biogeogr. 00, 1–12. doi: 10.1111/geb.12727
Harter, D. E. V., Irl, S. D. H., Bumsuk, S., Steinbauer, M. J., Gillespie, R., Triantis, K. A., et al. (2015). Impacts of global climate change on the floras of oceanic islands – Projections, implications and current knowledge. Perspect. Plant Ecol. Evol. Syst. 17, 160–183. doi: 10.1016/j.ppees.2015.01.003
Henderson, S., and Dawson, T. P. (2009). Alien invasions from space observations: Detecting feral goat impacts on Isla Isabela, Galapagos Islands with the AVHRR. Int. J. Remote Sens. 30, 423–433. doi: 10.1080/01431160802339472
Herder, J. E., Valentini, A., Bellemain, E., Dejean, T., van Delft, J. J. C. W., Thomsen, P. F., et al. (2014). Environmental DNA - A Review of the Possible Applications for the Detection of (invasive) Species. Nijmegen: Stichting RAVON, 2013–2104.
Hinkel, J., Lincke, D., Vafeidis, A. T., Perrette, M., Nicholls, R. J., Tol, R. S. J., et al. (2014). Coastal flood damage and adaptation costs under 21st century sea-level rise. Proc. Natl. Acad. Sci. U.S.A. 111, 3292–3297. doi: 10.1073/pnas.1222469111
Hulme, P. E. (2009). Trade, transport and trouble: Managing invasive species pathways in an era of globalization. J. Appl. Ecol. 46, 10–18. doi: 10.1111/j.1365-2664.2008.01600.x
Hulme, P. E., Bacher, S., Kenis, M., Klotz, S., Kaahn, I., Minchin, D., et al. (2008). Grasping at the routes of biological invasions: A framework for integrating pathways into policy. J. Appl. Ecol. 45, 403–414. doi: 10.1111/j.1365-2664.2007.01442.x
Humair, F., Humair, L., Kuhn, F., and Kueffer, C. (2015). E-commerce trade in invasive plants. Conserv. Biol. 29, 1658–1665. doi: 10.1111/cobi.12579
Hurtt, G. C., Chini, L. P., Frolking, S., Betts, R. A., Feddema, J., Gc, H., et al. (2009). Harmonisation of global land-use scenarios for the period 1500–2100 for IPCC-AR5 Recommended Citation. ILEAPS Newsl. 7, 6–8.
Inger, R. F. (2001). The biogeographical relations and snakes of Sundaland. J. Biogeogr. 28, 863–891. doi: 10.1046/j.1365-2699.2001.00580.x
IPBES (2018). The IPBES Assessment on Land Degradation and Restoration. Bonn: IPBES. doi: 10.4324/9781315640051-105
IPCC (2014). Climate Change 2013 - The Physical Science Basis, ed. R. K. Pachauri (Geneva: IPCC). doi: 10.1017/CBO9781107415324
Janzen, D. H. (1988). Management of Habitat Fragments in a Tropical Dry Forest Growth. Ann. Missouri Bot. Gard. 75, 105–116. doi: 10.2307/2399468
Jarić, I., Courchamp, F., Correia, R. A., Crowley, S. L., Essl, F., Fischer, A., et al. (2020). The role of species charisma in biological invasions. Front. Ecol. Environ. 18, 345–353. doi: 10.1002/fee.2195
Johnson, B. A., Mader, A. D., Dasgupta, R., and Kumar, P. (2020). Citizen science and invasive alien species: An analysis of citizen science initiatives using information and communications technology (ICT) to collect invasive alien species observations. Glob. Ecol. Conserv. 21:e00812. doi: 10.1016/j.gecco.2019.e00812
Kelman, I. (2010). Earth’s Future Special Section: Difficult decisions: Migration from Small Island Developing States under climate change. Future 3, 133–142. doi: 10.1002/2014EF000278.Received
Kier, G., Kreft, H., Lee, T. M., Jetz, W., Ibisch, P. L., Nowicki, C., et al. (2009). A global assessment of endemism and species richness across island and mainland regions. Proc. Natl. Acad. Sci. U.S.A. 106, 9322–9327. doi: 10.1073/pnas.0810306106
König, C., Weigelt, P., and Kreft, H. (2017). Dissecting global turnover in vascular plants. Glob. Ecol. Biogeogr. 26, 228–242. doi: 10.1111/geb.12536
König, C., Weigelt, P., Taylor, A., Stein, A., Dawson, W., Essl, F., et al. (2019). Disharmony of the world’ s island floras. bioRxiv [Preprint] doi: 10.1101/523464
Kueffer, C., Drake, D. R., and Fernández-Palacios, J. M. (2014). Island biology: looking towards the future. Biol. Lett. 10, 1–4. doi: 10.1098/rsbl.2014.0719
Latombe, G., Canavan, S., Hirsch, H., Hui, C., Kumschick, S., Nsikani, M. M., et al. (2019a). A four-component classification of uncertainties in biological invasions: implications for management. Ecosphere 10:e02669. doi: 10.1002/ecs2.2669
Latombe, G., Roura-Pascual, N., and Hui, C. (2019b). Similar compositional turnover but distinct insular environmental and geographical drivers of native and exotic ants in two oceans. J. Biogeogr. 46, 2299–2310. doi: 10.1111/jbi.13671
Lenda, M., Skórka, P., Knops, J. M. H., Moroń, D., Sutherland, W. J., Kuszewska, K., et al. (2014). Effect of the internet commerce on dispersal modes of invasive alien species. PLoS One 9:e99786. doi: 10.1371/journal.pone.0099786
Lenzner, B., Leclère, D., Franklin, O., Seebens, H., Roura-Pascual, N., Obersteiner, M., et al. (2019). A Framework for Global Twenty-First Century Scenarios and Models of Biological Invasions. Bioscience 69, 697–710. doi: 10.1093/biosci/biz070
Leung, B., Lodge, D. M., Finnoff, D., Shogren, J. F., Lewis, M. A., and Lamberti, G. (2002). An ounce of prevention or a pound of cure: bioeconomic risk analysis of invasive species. Proc. R. Soc. B Biol. Sci. 269, 2407–2413. doi: 10.1098/rspb.2002.2179
Levine, J. M., and D’Antonio, C. M. (1999). Elton revisited: a review of evidence linking diversity and invasibility. Oikos 87, 15–26. doi: 10.2307/3546992
Liedtke, R., Barros, A., Essl, F., Lembrechts, J. J., Wedegärtner, R. M., Pauchard, A., et al. (2020). Hiking trails as conduits for the spread of non-native species into mountain areas. Biol. Invasions 22, 1121–1134. doi: 10.1007/s10530-019-02165-9
Lutz, W., Butz, W. P., and Samir, K. (2014). World Population and the Human Capital in the Twenty-First Century. Oxford: Oxford University Press.
Mannino, A. M., and Balistreri, P. (2018). Citizen science: a successful tool for monitoring invasive alien species (IAS) in Marine Protected Areas. The case study of the Egadi Islands MPA (Tyrrhenian Sea, Italy). Biodiversity 19, 42–48. doi: 10.1080/14888386.2018.1468280
Maxwell, S. L., Fuller, R. A., Brooks, T. M., and Watson, J. E. M. (2016). Biodiversity: The ravages of guns, nets and bulldozers. Nature 536, 143–145. doi: 10.1038/536143a
McKinsey Global Institute (2019). Globalization in Transition: The Future of Trade and Value Chains. San Francisco, CA: McKinsey Global Institute.
Meyer, J. Y., and Fourdrigniez, M. (2019). Islander perceptions of invasive alien species: the role of socio-economy and culture in small isolated islands of French Polynesia (South Pacific). Island invasives: scaling up to meet the challenge 62, 510.
Moser, D., Lenzner, B., Weigelt, P., Dawson, W., Kreft, H., Pergl, J., et al. (2018). Remoteness promotes biological invasions on islands worldwide. Proc. Natl. Acad. Sci. U.S.A. 115, 9270–9275. doi: 10.1073/pnas.1804179115
Moss, R., Edmonds, J., Hibbard, K., Manning, M., Rose, S., van Vuuren, D., et al. (2010). The next generation of scenarios for climate change research and assessment. Nature 463, 747–756. doi: 10.1038/nature08823
Myers, N., Mittermeier, R. A., Mittermeier, C. G., da Fonseca, G. A., and Kent, J. (2000). Biodiversity hotspots for conservation priorities. Nature 403, 853–858. doi: 10.1038/35002501
Novoa, A., Shackleton, R., Canavan, S., Cybèle, C., Davies, S. J., Dehnen-Schmutz, K., et al. (2018). A framework for engaging stakeholders on the management of alien species. J. Environ. Manag. 205, 286–297. doi: 10.1016/j.jenvman.2017.09.059
Nuñez, M. A., Barlow, J., Cadotte, M., Lucas, K., Newton, E., Pettorelli, N., et al. (2019). Assessing the uneven global distribution of readership, submissions and publications in applied ecology: Obvious problems without obvious solutions. J. Appl. Ecol. 56, 4–9. doi: 10.1111/1365-2664.13319
OECD (2018). Making Development Co-operation Work for Small Island Developing States. Paris: OECD, doi: 10.1787/9789264287648-en
Ojaveer, H., Galil, B. S., Campbell, M. L., Carlton, J. T., Canning-Clode, J., Cook, E. J., et al. (2015). Classification of non-indigenous species based on their impacts: considerations for application in marine management. PLoS Biol. 13:e1002130. doi: 10.1371/journal.pbio.1002130
Pergl, J., Pyšek, P., Bacher, S., Essl, F., Genovesi, P., Harrower, C. A., et al. (2017). Troubling travellers: Are ecologically harmful alien species associated with particular introduction pathways? NeoBiota 32, 1–20. doi: 10.3897/neobiota.32.10199
Plieninger, T., Hui, C., Gaertner, M., and Huntsinger, L. (2014). The Impact of Land Abandonment on Species Richness and Abundance in the Mediterranean Basin: A Meta-Analysis. PLoS One 9:e98355. doi: 10.1371/journal.pone.0098355
Reaser, J. K., Burgiel, S. W., Kirkey, J., Brantley, K. A., Veatch, S. D., and Burgos-Rodríguez, J. (2020). The early detection of and rapid response (EDRR) to invasive species: a conceptual framework and federal capacities assessment. Biol. Invasions 22, 1–19. doi: 10.1007/s10530-019-02156-w
Reside, A. E., Critchell, K., Crayn, D. M., Goosem, M., Goosem, S., Hoskin, C. J., et al. (2018). Beyond the model: expert knowledge improves predictions of species’ fates under climate change. Ecol. Appl. 29, 1–15. doi: 10.1002/eap.1824
Rey Benayas, J. M., Martins, A., Nicolau, J. M., and Schulz, J. (2007). Abandonment of agricultural land: an overview of drivers and consequences. CAB Rev. 2:057.
Ricardo, B. R., Hines, A., Arturo, A. P., Ortí, G., Wilbur, A. E., and Freshwater, D. W. (2011). Reconstructing the lionfish invasion: Insights into Greater Caribbean biogeography. J. Biogeogr. 38, 1281–1293. doi: 10.1111/j.1365-2699.2011.02496.x
Rivas-Torres, G. F., Benítez, F. L., Rueda, D., Sevilla, C., and Mena, C. F. (2018). A methodology for mapping native and invasive vegetation coverage in archipelagos: An example from the Galápagos Islands. Prog. Phys. Geogr. 42, 83–111. doi: 10.1177/0309133317752278
Robin, M., Chapuis, J. L., and Lebouvier, M. (2011). Remote sensing of vegetation cover change in islands of the Kerguelen archipelago. Polar Biol. 34, 1689–1700. doi: 10.1007/s00300-011-1069-z
Rocchini, D., Andreo, V., Forster, M., Garzon-Lopez, C. X., Gutierrez, A. P., Gillespie, T. W., et al. (2015). Potential of remote sensing to predict species invasions: a modelling perspective. Prog. Phys. Geogr. 39, 283–309. doi: 10.1177/0309133315574659
Roy, H., Groom, Q., Adriaens, T., Agnello, G., Antic, M., Archambeau, A.-S., et al. (2018). Increasing understanding of alien species through citizen science (Alien-CSI). Res. Ideas Outcomes 4:31412. doi: 10.3897/rio.4.e31412
Russell, J. C., Innes, J. G., Brown, P. H., and Byrom, A. E. (2015). Predator-free New Zealand: Conservation country. Bioscience 65, 520–525. doi: 10.1093/biosci/biv012
Russell, J. C., Meyer, J. Y., Holmes, N. D., and Pagad, S. (2017). Invasive alien species on islands: Impacts, distribution, interactions and management. Environ. Conserv. 44, 359–370. doi: 10.1017/S0376892917000297
Sanchez-Ortiz, K., Taylor, K. J. M., Palma, A., De König, C., Pyšek, P., Weigelt, P., et al. (2019). Effects of land — use change and related pressures on alien and native subsets of island communities. bioRxiv [Preprint] doi: 10.1101/2019.12.16.878041
Sardain, A., Sardain, E., and Leung, B. (2019). Global forecasts of shipping traffic and biological invasions to 2050. Nat. Sustain. 2, 274–282. doi: 10.1038/s41893-019-0245-y
Seebens, H., Blackburn, T. M., Dyer, E. E., Genovesi, P., Hulme, P. E., Jeschke, J. M., et al. (2018). Global rise in emerging alien species results from increased accessibility of new source pools. Proc. Natl. Acad. Sci. U.S.A. 115, E2264–E2273. doi: 10.1073/pnas.1719429115
Seebens, H., Essl, F., Dawson, W., Fuentes, N., Moser, D., Pergl, J., et al. (2015). Global trade will accelerate plant invasions in emerging economies under climate change. Glob. Chang. Biol. 21, 4128–4140. doi: 10.1111/gcb.13021
Seipel, T., Kueffer, C., Rew, L. J., Daehler, C. C., Pauchard, A., Naylor, B. J., et al. (2012). Processes at multiple scales affect richness and similarity of non-native plant species in mountains around the world. Glob. Ecol. Biogeogr. 21, 236–246. doi: 10.1111/j.1466-8238.2011.00664.x
Self, R. M., Self, D. R., and Bell-Haynes, J. (2010). Marketing Tourism In The Galapagos Islands: Ecotourism Or Greenwashing? Int. Bus. Econ. Res. J. 9, 111–126. doi: 10.19030/iber.v9i6.590
Shackleton, R. T., Foxcroft, L. C., Pyšek, P., Wood, L. A., and Richardson, D. M. (2020). Assessing biological invasions in protected areas after 30 years: Revisiting nature reserves targeted by the 1980s SCOPE programme. Biol. Cons. 243:108424. doi: 10.1016/j.biocon.2020.108424
Shackleton, R. T., Richardson, D. M., Shackleton, C. M., Bennett, B., Crowley, S. L., Dehnen-Schmutz, K., et al. (2019). Explaining people’s perceptions of invasive alien species: A conceptual framework. J. Environ. Manag. 229, 10–26. doi: 10.1016/j.jenvman.2018.04.045
Strubbe, D., and Matthysen, E. (2009). Establishment success of invasive ring-necked and monk parakeets in Europe. J. Biogeogr. 36, 2264–2278. doi: 10.1111/j.1365-2699.2009.02177.x
Symstad, A. J., Fisichelli, N. A., Miller, B. W., Rowland, E., and Schuurman, G. W. (2017). Multiple methods for multiple futures: Integrating qualitative scenario planning and quantitative simulation modeling for natural resource decision making. Clim. Risk Manag. 17, 78–91. doi: 10.1016/j.crm.2017.07.002
Taylor, A., Weigelt, P., König, C., Zotz, G., and Kreft, H. (2019). Island disharmony revisited using orchids as a model group. New Phytol. 223, 597–606. doi: 10.1111/nph.15776
Tershy, B. R., Shen, K. W., Newton, K. M., Holmes, N. D., and Croll, D. A. (2015). The importance of islands for the protection of biological and linguistic diversity. Bioscience 65, 592–597. doi: 10.1093/biosci/biv031
Thomsen, P. F., and Willerslev, E. (2015). Environmental DNA - An emerging tool in conservation for monitoring past and present biodiversity. Biol. Conserv. 183, 4–18. doi: 10.1016/j.biocon.2014.11.019
Tingley, R., Greenless, M., van Rooyen, A. R., and Weeks, A. R. (2019). Environmental DNA sampling as a surveillance tool for cane toad Rhinella marina introductions on offshore islands. Biol. Inv. 21, 1–6. doi: 10.1007/s10530-018-1810-4
Toral-Granda, M. V., Causton, C. E., Jäger, H., Trueman, M., Izurieta, J. C., Araujo, E., et al. (2017). Alien species pathways to the Galapagos Islands. Ecuador. PLoS One 12:e0184379. doi: 10.1371/journal.pone.0184379
Troudet, J., Grandcolas, P., Blin, A., Vignes-Lebbe, R., and Legendre, F. (2017). Taxonomic bias in biodiversity data and societal preferences. Sci. Rep. 7, 1–14. doi: 10.1038/s41598-017-09084-6
United Nations, Department of Economic and Social Affairs and Population Division (2019). World Population Prospects 2019. (ST/ESA/SER.A/423). Available online at: http://www.ncbi.nlm.nih.gov/pubmed/12283219 (accessed May, 2020).
van Kleunen, M., Essl, F., Pergl, J., Brundu, G., Carboni, M., Dullinger, S., et al. (2018). The changing role of ornamental horticulture in alien plant invasions. Biol. Rev. 93, 1421–1437. doi: 10.1111/brv.12402
Verbrugge, L. N. H., Van Den Born, R. J. G., and Lenders, H. J. R. (2013). Exploring public perception of non-native species from a visions of nature perspective. Environ. Manag. 52, 1562–1573. doi: 10.1007/s00267-013-0170-1
Wetzel, F. T., Kissling, W. D., Beissmann, H., and Penn, D. J. (2012). Future climate change driven sea-level rise: secondary consequences from human displacement for island biodiversity. Glob. Chang. Biol. 18, 2707–2719. doi: 10.1111/j.1365-2486.2012.02736.x
Whittaker, R. J., and Fernandez-Palacios, J. M. (2007). Island Biogeography: Ecology, Evolution and Conservation. Oxford: Oxford University Press.
Keywords: biodiversity change, biological invasions, drivers, islands, expert-based approach, plausible futures, scenarios, survey
Citation: Lenzner B, Latombe G, Capinha C, Bellard C, Courchamp F, Diagne C, Dullinger S, Golivets M, Irl SDH, Kühn I, Leung B, Liu C, Moser D, Roura-Pascual N, Seebens H, Turbelin A, Weigelt P and Essl F (2020) What Will the Future Bring for Biological Invasions on Islands? An Expert-Based Assessment. Front. Ecol. Evol. 8:280. doi: 10.3389/fevo.2020.00280
Received: 20 May 2020; Accepted: 10 August 2020;
Published: 04 September 2020.
Edited by:
Ross Taylor Shackleton, University of Lausanne, SwitzerlandReviewed by:
Louisa Wood, Centre for Environment, Fisheries and Aquaculture Science (CEFAS), United KingdomCopyright © 2020 Lenzner, Latombe, Capinha, Bellard, Courchamp, Diagne, Dullinger, Golivets, Irl, Kühn, Leung, Liu, Moser, Roura-Pascual, Seebens, Turbelin, Weigelt and Essl. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Bernd Lenzner, YmVybmQubGVuem5lckB1bml2aWUuYWMuYXQ=
†ORCID ID: Bernd Lenzner orcid.org/0000-0002-2616-3479 Céline Bellard orcid.org/0000-0003-2012-1160 Franck Courchamp orcid.org/0000-0001-7605-4548 Christophe Diagne orcid.org/0000-0002-6406-1270 Marina Golivets orcid.org/0000-0003-1278-2663 Severin D. H. Irl orcid.org/0000-0002-1734-8607 Ingolf Kühn orcid.org/0000-0003-1691-8249 Brian Leung orcid.org/0000-0002-8323-9628 Chunlong Liu orcid.org/0000-0002-3376-4555 Núria Roura-Pascual orcid.org/0000-0003-0025-2972 Hanno Seebens orcid.org/0000-0001-8993-6419
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.